
Submitted:
06 September 2024
Posted:
10 September 2024
You are already at the latest version
Abstract
Worldwide the severity of abiotic stress like heat, drought, salinity, cold, waterlogging, etc. are being increased because of global warming. Wheat crops must adapt to harsh environments through physiological, molecular, and genetic manipulation. Because of abiotic stress, reactive oxygen species (ROS) and reactive nitrogen species (RNS) produced in wheat plants break down DNA and RNA bases, protein, lipid, and other macro- and micromolecules, resulting in stunted growth and yield decline. Abiotic stress also induces the crop's molecular functions, physiological, biological, and cellular processes, and their compositions. Different stressors like high light intensity, heat, drought, and salinity also impair photosynthesis damaging various pigments. Antioxidant enzymes, such as catalase (CAT), superoxide dismutase (SOD), peroxidase (POD), and ascorbate peroxidase (APX), etc. synthesized in harsh environments can ameliorate ROS/RNS produced in oxidative stress. Under abiotic stress, heat shock proteins (HSPs) heat shock transcription factors (HSFs) in heat stress, and dehydration-responsive element binding proteins (DREBs) in drought stress are activated to synthesize proteins to mitigate the deleterious effects of those stressors. Na+ exclusive genes (NAX1, NAX2) and Na+/H+ (K+) antiporter genes (NHX1, NHX2) hinder those ions from uptake into leaf sheaths and blades of wheat resulting in the declination of salinity in plants. Exogenous hormones like gibberellic acid, salicylic acid, and melatonin, etc., and osmoprotectants like proline, and gene manipulation in plants through genetic engineering are being employed to mitigate the toxic effects of abiotic stress on plants, as portrayed in many literatures. The review depicts how abiotic stress affects phenotypic, physiological, and molecular structures and functions of wheat crops, and their mitigation strategies to lessen their noxious effects on wheat.
Keywords:
Drought
; Heat
; Genes
; Antioxidants
; Salinity
; Oxidative stress
1. Introduction
Abiotic stress largely reduces wheat production globally. Extreme abiotic stressors, such as drought, high temperatures, salinity, cold, waterlogging, etc., and biotic stress like insects-pests, and diseases decrease the yield of wheat in the least developed and developing countries by 20-30%, according to recent projections by the ICARDA, CIMMYT, OECD, and FAO (Islam et al. 2016). According to Zhao et al. (2017), if crops aren’t genetically enhanced for heat, drought, salinity, cold, etc., resilience, here’s the predicted yield of 6% loss of wheat for each 1°C temperature increase. In addition, if anthropogenic activities continue to cause global warming, the Intergovernmental Panel on Climate Change (IPCC) predicts a 6.4°C temperature will rise by the end of the century, and the sea level will rise by 59 cm as a result of glacier melting same time (IPCC 2014). Climate change can increase floods, droughts, storms, and precipitation patterns, etc. Since agriculture is climate-sensitive, temperature, humidity, and rainfall affect crop yield (Atanga and Tankpa 2021). A major cereal crop known as wheat (Triticum aestivum L.) is grown around the world in various agro-ecologies (Abhinandan et al. 2018). According to forecasts, there will be 60% rise in demand for wheat in 2050 to feed the expected 9.7 billion people on the planet (Yadav et al. 2020). It is widely grown in almost all continents, such as Asia, Africa, Europe, Australia, and America. Drought stress affects crops’ growth and development. Dryness during early growth reduces wheat seedling stand establishment and tiller production. Mid-growth droughts reduce dry matter output, productive tillers, and grains per plant (Dudziak et al. 2019; Gull et al. 2019). Drought during terminal growth reduces grain weight, fertility, and absorption, killing wheat. Drought stress disrupts antioxidant defenses and ROS generation. During prolonged water stress, chloroplast ROS production degrades the wheat photosynthetic system (Dudziak et al. 2019). Malondialdehyde (MDA) generation may indicate ROS-induced oxidative damage. Wheat cells’ high H2O2 and MDA levels inhibit drought-induced oxidative stress tolerance (Djanaguiraman et al. 2020; Rath et al. 2022). Wheat ripens best around 14-15°C, although grain weight decreases at 25°C. Diurnal temperature variation under a changing environment results in yield loss (Alam et al. 2013c; Raza et al. 2019; Hossain et al. 2020). Grain filling, formation, yield loss, and spike yield decrease with temperature. Extreme weather occurrences like frost (0°C) and heat shock (brief periods of very high temperatures (>33°C) pose a significant risk to crop productivity (Alam et al. 2014; Mukherjee et al. 2019). Salinity affects 20% of the world’s arable land, and anthropogenic and environmental changes are growing daily. 33% of irrigated agricultural land and 20% of peripheral areas have excessive salinity, as estimated by Chele et al. (2021). Saline soil is one with an electrical conductivity of the saturation extract at the root zone greater than 4 dS/m (approximately 40 mM NaCl) and an exchangeable salt content greater than 15% (Shrivastava and Kumar 2015). High soil salt concentration reduces leaf water potential, decreases turgor pressure due to osmotic imbalance, closes stomata, decreases CO2 conductivity, affects photosynthetic rate, chlorophyll (Chl) content, carotenoids, membrane variability, cell wall integrity, oxidative stress due to increased ROS production, and increases toxic metabolites. This may damage DNA, RNA, proteins, and lipids, impede plant function, and kill the plants (Turkan 2018; Alam et al. 2021). Stress activates enzymatic, structural, and regulatory genes. Transgenic investigations started with single-action genes. The first targets were water channel proteins, osmolyte biosynthesis (proline, betaine, sugars like trehalose, and polyamines), detoxifying enzymes, and transport proteins. However, stress tolerance is regulated by multiple genes simultaneously, thus single-gene tolerance is implausible. Plants have stress-induced regulatory genes. One gene can regulate many stress-related proteins. In all species, temperature boosts HSP transcription (Chen et al. 2023). Stress converts a non-DNA-binding monomeric form of HSF into a DNA-binding trimeric form (Table 1). High temperatures create HSPs (ul Haq et al. 2019). Transgenic plants with higher plastid Elongation factor-thermal unstable (EF-Tu) reduced thylakoid membrane damage and boosted photosynthetic rate and grain yield during high-temperature stress. EF-Tu may protect photosynthetic membranes and photosynthesis-related enzymes from high-temperature stress, boosting CO2 fixation. Detox gene overexpression of CBF1/DREB1B genes increased rice, wheat, and canola’s temperature tolerance (Table 1) (Djanaguiraman et al. 2020). Co-expression of Escherichia coli P5C biosynthetic enzymes GK74 and GPR in Arabidopsis and tobacco increased proline biosynthesis. Transgenic plants have increased high-temperature stress tolerance due to cell wall proline-rich proteins. Simultaneous co-expression of DHAR, GR or GST, and GR in tobacco plants boosted temperature stress tolerance (Table 1) (Reguera et al. 2012). ROS are poisonous molecules that can cause oxidative damage to lipids, proteins, and DNA. Overexpression of Mn-superoxide dismutase in wheat increased temperature tolerance and field yields. APX and CAT detoxify H2O2. The cAPX gene improves tomato’s heat tolerance (Chen et al. 2023). Combining antioxidant enzyme expression may improve high-temperature stress resistance. Constitutive expression of MBF1c in Arabidopsis thaliana increases transgenic plants’ tolerance to bacterial infection, high temperature, and osmotic stress. Enhanced transgenic plant resistance to osmotic and high-temperature stress was maintained when combined (Tian et al. 2013). Transcriptomic profiling and inhibitor experiments reveal that MBF1c expression increases transgenic plant tolerance to high temperature and osmotic stress by partially activating the ethylene-response signal transduction system (Ma et al. 2024). Katiyar-Agarwal et al. (2003) improved a high-temperature-tolerant transgenic rice line (Pusa basmati). The findings showed that almost all transgenic plants recovered after 45-50°C heat stress and grew vigorously at 28°C, while untransformed plants did not. Overexpressing sHSP17.7 produces high-temperature-tolerant rice plants. mtHSP70 overexpression reduces programmed cell death in rice protoplasts via preserving mitochondrial membrane potential and limiting ROS signal amplification (Murakami et al. 2004). Abiotic stress induces ROS/RNS, MDA, electrolyte leakage, and metabolite production resulting in an imbalance of radicles found in plant cells, called oxidative stress. During oxidative stress carbohydrates, proteins, fatty acids, DNA, and RNA bases of plants are broken down, ultimately uneven plant growth producing the reduced yield and contributing characteristics of wheat, and resulting in poor grain yield (Alam et al. 2013c; Hossain et al. 2020). This review provokes how abiotic stresses affect the morphological, physiological, biochemical, and genetic levels of wheat crops, as well as their mitigation strategies.
2. Detrimental effects of abiotic stress on wheat crop
2.1. Wheat seed germination and growth under abiotic stress
Originally wheat was a cool-loving plant. Winter wheat germinates at 6°C, while spring wheat germinates from 6.0 to 26°C. Genetically, winter wheat seedlings can survive <10°C. Non- acclimated winter wheat seedlings can survive at - 5°C, while acclimated ones can live up to - 8°C (Barlow et al. 2015). Drought stress reduced seedling growth and development, turning leaves into pale to yellow colors lacking moisture in the soil. Heat stress for cold-loving wheat resulted in burned and curled leaves of seedlings, and uneven growth. The rates of seed germination and seedling survivability were also reduced under salinity stress (Fig. 1) (Yadav et al. 2022; Hossain et al. 2020; Rane et al. 2007; Munns and Tester 2008).
2.2. ROS production under abiotic stress and their effects on wheat
Generally, abiotic stress provokes heat shock, high temperature, drought, salinity, waterlogging, chilling stress, etc. Moderately reduced forms of oxygen (O2) are called ROS. Excitation of molecular O2 in the presence of abiotic stress like high temperature, drought, salinity, cold and waterlogging stress, etc. produces singlet oxygen (1O2) or transfers electrons to O2 to form hydrogen peroxide (H2O2), hydroxyl ion (OH−) and superoxide ion (O2•–). During abiotic stress, the structure of molecules of plants or living organisms, such as amino acid, DNA, RNA, fatty acid, etc. is broken down by donating or accepting electron(s) (Miura and Furumoto 2013). Result, radicals like OH¯, H2O2, and O2•–, and signaling molecules like nitric oxide (NO) and nitrogen dioxide (NO2) are formed resulting in a disturbance of cell homeostasis (Fig 1). Because of the disorganization of cells, plant growth and development, metabolisms, and physiological and biochemical activities were hindered (Alam et al. 2018a, Alam et al. 2021). Less germination, seedling injury, insufficient tillering, early booting, heading, flowering, and ripening stages were also found. Finally, unfilled, and small-sized grains were yielded (Sabagh et al. 2013; Alam et al. 2013a; Alam et al. 2013b).
2.3. RNS production under abiotic stress and their effects on wheat
RNSs are important signals in plants, serving as key regulators of a variety of processes including metabolism, growth, and development, response to abiotic and biotic stresses, solute transport, autophagy, and programmed cell death (Turkan 2018). RNS in plants were identified in the 1960s (Fewson and Nicholas 1960), but gained significant attention only in the late 1990s, particularly with the recognition of NO as a signaling molecule in plant pathogenesis. Unlike ROS, which were initially seen as damaging molecules, RNS were first acknowledged for their signaling role. The term “nitrosative stress” emerged in plant biology literature in the 2000s. RNS are now recognized as vital signals regulating various plant processes, even after over 50 years of research. Ongoing research continues to uncover new physiological and molecular functions for RNS, highlighting their central importance in plant cell functioning (Foyer and Noctor, 2015). NO in plants was initially discovered by Klepper (1979) in soybean plants exposed to photosynthetic inhibitors, darkness, or anoxic conditions. Subsequent research has revealed NO’s involvement in various physiological and metabolic plant processes, including adaptation to environmental stresses (Uchida et al., 2002). It reacts with O2•– to form peroxynitrite (ONOO2). ONOO2 can also be produced by nitrate reductase (NR) in the presence of O2 and NAD(P)H (Corpas et al. 2014). It is a powerful oxidant damaging DNA, lipids, and proteins, leading to cellular harm and cytotoxicity (Radi 2013). Protonation of ONOO2 produces peroxynitrous acid, a source of NO2 and OH− (Procházková et al. 2014). RNS, particularly S-nitrosylation, plays a crucial role in regulating cellular metabolism and signaling. It activates the production of GSNO, essential for abiotic stress response and plant immunity. Studies by Begara-Morales et al. (2018) address GSNO turnover mechanisms and methods for NO and SNO detection. Umbreen et al. (2018) delve into the specificity of NO signaling, emphasizing denitrosylation's significance in RNS and ROS interaction. Corpas et al. (2018) discuss RNS involvement during fruit ripening and its potential for improving pepper and tomato yield and quality through RNS interaction studies. The enzyme GSNO reductase (GSNOR) regulates cellular GSNO levels and NO content by reducing GSNO to oxidized glutathione (GSSG) and NH3 in an NADH-dependent manner. This process influences overall RSNO content. In plant cells, GSNOR serves a dual purpose: dampening GSNO-derived NO signaling and protecting against nitrosative stress by controlling excessive S-nitrosylation (Kovacs et al. 2016). NO facilitates post-translational modifications through nitration and S-nitrosylation. Nitration, a general chemical process, introduces a nitro group (NO2) into compounds, with tyrosine being a primary target. Tyrosine nitration, by adding a nitro group to one of its two orthocarbons, can alter protein function. It may enhance, inhibit, or leave the function unchanged. In plants, nitrotyrosine could indicate nitrosative stress during abiotic stress, similar to protein carbonylation or lipid peroxidation.
Heat and drought stresses reduce the relative water content of wheat plants, leading to increased generation of ROS and peroxynitrite (ONOO-), decreased NO production, and enhanced lipid peroxidation in stigmatic papilla cells. These physiological changes induce alterations in the morphology and anatomy of female reproductive organs, shorten the duration of gametogenesis and grain filling, and significantly reduce fertility and plant production (Fábián et al. 2019). In wheat roots, ABA synthesis increases in response to water deficiency, especially in the presence of NO donors and ROS. This indicates a synergistic action between ROS and NO. Moreover, exogenous application of the NO donor sodium nitroprusside (SNP) reduced water loss from detached wheat leaves, lowered transpiration rate, induced stomatal closure, and reduced ion leakage. These effects were abolished by a specific NO scavenger (Garcı´a-Mata and Lamattina 2001). Additionally, SNP improved photosynthetic performance and mitigated the negative impacts of drought on carbohydrate and nitrogen accumulation in marigold leaves (Tagetes erecta L.) (Liao et al. 2012). The accumulation of proline is a well-known adaptive response of plants to drought stress. In wheat, exogenous application of NO reduces osmotic stress by decreasing oxidative damage and stimulating proline accumulation (Tan et al. 2008). Additionally, NO increases the content of the low-molecular antioxidant ascorbate in Cd-treated rice leaves (Hsu and Kao 2004) and enhances glutathione peroxidase (GPX) activity in Cd-treated sunflower leaves (Laspina et al. 2005). However, NO treatment decreases GPX and catalase (CAT) activities in Cd-stressed wheat roots (Singh et al. 2008). Moreover, NO pretreatment improves wheat seed germination and alleviates oxidative stress under Cu-toxicity by increasing the activities of superoxide dismutase (SOD) and CAT, while decreasing lipoxygenase (LOX) activity and malondialdehyde synthesis (Hu et al. 2007). Singh et al. (2004) propose that the protective effect of NO may involve the suppression of O2•– production, subsequently reducing OH− production from O2•– and ONOO-. Excess aluminum creates toxicity in the root zone of plants, and it disrupts other parts. Al-stress in wheat affects redox balance and NO metabolism, leading to nitro-oxidative stress. The sensitive genotype, Yang-5, exhibits increased ROS, NO, and ONOO- levels, along with heightened activities of oxidizing enzymes. Conversely, the tolerant genotype, Jian-864, demonstrates lower oxidative damage and maintains redox balance through adjustments in its antioxidant defense system (Sun et al. 2017).
2.4. Effects of frost and cold temperature on wheat
Frost is very deleterious for both wheat’s growth and development. The impact of frost damage on wheat has been observed across all stages of growth, from seedlings to maturity (Hasan et al. 2021). However, Frederiks et al. (2012) found that the yield impact resulting from frost damage during the reproductive stage of growth is significantly greater for both spring and winter wheat compared to any other stage. Yue et al. (2016) found that spring frost affects a significant portion of the wheat-sown area worldwide every year, with approximately 85% of the total area impacted. This spring frost event typically occurs during March and April, coinciding with the early booting stage of wheat development. Cold conditions can indeed disrupt root water uptake, leading to water inadequacy in the stem and subsequent drought stress, as noted by Aroca et al. (2012). This drought condition, stemming from imbalanced water relations, can cause disturbances in nutrient uptake, decrease the rate of root ion absorption, and limit nutrient transport to other plant parts. Ultimately, this can result in stunted plant growth, as highlighted by Nezhadahmadi et al. (2013). Fuller et al. (2007) observed that frost during this period affects seedling survival. Frost can cause leaf damage, resulting in a scorched appearance of leaves. While frost damage during vegetative stages may impact seedling survival and cause leaf damage, its potential impact on yield is generally small because the growing point of wheat is typically located in the soil, protecting it from damage (Frederiks et al. 2015). Many wheat cultivars exhibit high levels of frost tolerance during the vegetative period. Some winter wheat types are tolerant to temperatures as low as -20°C (Frederiks et al. 2012). This tolerance is induced through a process of cold acclimation, which produces "hardened" wheat plants. Low-temperature acclimation in winter wheat is a genetically regulated, cumulative process initiated below 10°C (Hasan et al. 2021). A gradient in seedling survival for winter wheat, with non-acclimated plants starting to see declines in seedling survival at around -5°C, while acclimated winter wheat seedlings showed declines between -6°C and -8°C (Fuller et al. 2007). During the reproductive growth stages of wheat, a single frost event can have a significant impact on crop yields, ranging from moderate to severe (Barlow et al. 2015). Both spring and winter wheat are susceptible to frost damage during these stages, with sensitivity increasing as maturity progresses, particularly from spike emergence to anthesis (Frederiks et al. 2012). The transition from the vegetative to the reproductive stage is critical, as wheat's frost tolerance observed during the vegetative stages ceases with the onset of reproductive growth (Fuller et al. 2007). Around booting, the stem is most frequently damaged above the top node, potentially leading to the loss of the head (Frederiks et al. 2008). Stem damage lower in the plant can also occur, increasing the risk of lodging during grain filling. However, crops with frost-damaged stems during booting can still develop grain with minimal impact on yield, provided that other factors like water supply are not limiting and lodging does not occur (Rebbeck and Knell 2007). During the critical stages of wheat development, frost events can have various detrimental effects on the crop's yield, as highlighted by Frederiks et al. (2008), Cromey et al. (1998), Fuller et al. (2007), and Rebbeck and Knell (2007). Before emergence, frost can damage the head by causing sterility of florets, thereby reducing grain number. During ear emergence and early anthesis, frost can lead to the death of anthers and embryos, resulting in sterility of florets and entire spikelets. Frost occurring during grain filling may cause the death of partially filled grains and reduce grain weight, resulting in small, shriveled, shrunken, or blistered grains. Frost damage during ear emergence and anthesis typically occurs within a narrow temperature range. After reaching a threshold temperature, there is a sharp reduction in the grain set. Cromey et al. (1998) conducted experiments exposing wheat plants to freezing stress ranging from 0°C to -13°C in a controlled chamber for 2 hours. They observed that frost devastation commenced at -3°C, with complete burning of flag leaves and ears occurring at -7°C. Consequently, a substantial reduction in grain yield was observed. Similarly, Valluru et al. (2012) found that exposure to cold at the jointing stage of wheat growth leads to reduced leaf size, decreased leaf area, and lower shoot biomass, ultimately limiting the final output. Furthermore, applying freezing stress, particularly at the stem elongation stage, as investigated by Whaley et al. (2004), leads to several detrimental effects. This includes limiting internode extension, denaturing the spikelets, reducing assimilate transport, restricting dry matter accumulation, and ultimately causing a significant reduction in grain yield. Marcellos and Single (1984) identified the threshold canopy temperature to be between -4°C and -5°C for different cultivars, consistent with other studies noting good resistance down to around -5°C near anthesis. Once this threshold temperature is reached, even 1°C difference in nighttime minimum temperature can significantly increase crop damage, ranging from 10% to 90%. These findings underscore the vulnerability of wheat crops to frost damage during critical developmental stages and emphasize the importance of monitoring and mitigating the risks associated with frost events.
2.5. Effects of abiotic stress on physiological processes
Plant pigments are broadly categorized by chlorophylls (Chl a, Chl b), anthocyanins, and carotenoids. They serve multiple functions, including light absorption for photosynthesis, protection against excess light, and attraction of pollinators. These pigment groups contribute to the diverse array of colors seen in the plant kingdom and serve important roles in various physiological processes, including photosynthesis, protection from environmental stresses, and attraction of pollinators. Abiotic stress reduces leaf Chl. Early leaf senescence caused by Chl loss under abiotic stress affected the Chl a:b ratio. Under abiotic stress, Chl molecules may disintegrate and produce ROS/RNS (Alam et al. 2021; Sabagh et al. 2021). Plant Chl content decreases with heat, dryness waterlogging, anaerobic, and salinity conditions of the plant. Abiotic stress-induced leaf senescence affected the Chl a: b ratio. ROS/RNS generation might be linked to Chl breakdown during abiotic stress (Pospíšil 2016; Alam et al. 2018b). Thylakoid membrane damage was exacerbated by high-temperature stress (Djanaguiraman et al. 2020). Because the characteristics of the thylakoid membranes, where photosystem (PS)II exists, altered with temperature, PSII is thought to be very temperature sensitive and its activity diminishes at high temperatures (Hameed et al. 2021). The oxygen-producing complex dissociates under high thermal stress, which produces an imbalance in the flow of electrons from the complex toward the acceptor side of the PSII reaction center (Khorobrykh et al. 2020). Under extreme temperature stress, the PSI system often performs better than the PS II (Rath et al. 2022). Either photosynthesis by closing stomata and decreasing CO2 flow into mesophyll tissue or directly impaired metabolic activities can be induced by high-temperature stress. Abiotic stress impairs photosynthetic electron transport, ribulose-1,5-bisphosphate (RuBP) regeneration capacity, and ribulose 1,5-bisphosphate carboxylase/oxygenase (Rubisco) protein content and activity (Scafaro et al. 2023). Decreased conductance through stomata is the main factor contributing to photosynthetic drop during the early phases of a stress period. Abiotic stress inhibits stomatal conductance and net photosynthesis due to decreased Rubisco activase enzyme (Scafaro et al. 2023). Low enzyme and CO2 affinity and increased oxygenase-type activity limit the net photosynthetic rate, while high temperature boosts the activities of the enzyme rubisco. While it's common knowledge that when temperature rises, so do respiratory rates, little is known about how respiration is controlled in response to extreme heat stress. Because photosynthesis is confined in time (only occurring throughout the day) and space (only occurring in green tissues), an appreciation of these reactions is crucial (Hill and Li 2022).
High light stress can lead to damage to the oxygen-evolving complex and PSII reaction center, causing photoinhibition by disrupting D1 and D2 proteins. Additionally, abiotic stresses trigger ROS production, upsetting chloroplast redox equilibrium, damaging reaction centers, and hindering synthesis and repair processes, ultimately leading to photoinhibition. Structural damage to chloroplasts, including lamellae and grana, and the breakdown of chlorophyll pigment and protein complexes can further inhibit photosynthesis. These stresses disrupt electron transfer, deactivate light reaction enzymes, and generate excess reduction power, all contributing to photosynthesis inhibition (Chauhan et al 2023).
Abiotic stress induces photoperiodic reactions in the plant. Plants perceive and respond to various aspects of light through photoreceptors and chloroplasts. Photoreceptors like phytochromes, cryptochromes, and UV RESISTANCE LOCUS8 (UVR8), sense different light wavelengths and are involved in circadian clock regulation. Chloroplasts also act as light sensors, altering their structure and function based on light conditions. They regulate starch accumulation and mobilization, adjusting to photoperiod changes to optimize energy usage. The molecular mechanisms behind these processes, including redox signaling and transcriptional control, are still being investigated. The circadian clock in plants regulates various physiological processes by measuring time through an internal mechanism, primarily influenced by light and temperature. In Arabidopsis thaliana, this clock involves interconnected transcription-translation feedback loops. Genes like CIRCADIAN CLOCK ASSOCIATED1 (CCA1) and LATE ELONGATED HYPOCOTYL (LHY) control morning expression and repress TOC1 during the day photoreceptor (Sanchez et al, 2020; Roeber et al, 2021). TIMING OF CAB EXPRESSION1 (TOC1), in turn, inhibits CCA1 and LHY, with its expression regulated by the evening complex comprising EARLY FLOWERING3 (ELF3), ELF4, and LUX ARRHYTHMO (LUX) ELF3, ELF4 and LUX. PRR9, PRR7, and PRR5 repress CCA1 and LHY throughout the day (Alabadi et al. 2001; Nakamichi et al. 2012). Other factors like PSEUDO-RESPONSE REGULATOR9 (PRR9), PRR7, and PRR5, LIGHT-REGULATED WD1 (LWD1) and LWD2 proteins, NIGHT LIGHT-INDUCIBLE and CLOCK-REGULATED GENE1 (LNK1) and LNK2 contribute to clock function. The clock influences plant responses to environmental stresses, with stress also affecting clock function (Rawat et al. 2011; Rugnone et al. 2013). A new web tool called ATTRACTOR aids in understanding transcriptional networks regulated by light and the circadian clock. One key clock-controlled gene, GIGANTEA (GI), plays a crucial role in photoperiod sensing by stabilizing the F-box protein ZEITLUPE, which targets clock components for degradation. This stabilization ensures the correct timing of clock output genes like CO2, essential for photoperiod-dependent flowering. The CO-FT-GI- CYCLING DOF FACTOR1 (CDF1) hub is conserved across flowering plants, indicating its importance in photoperiodic responses (Serrano-Bueno et al, 2021).
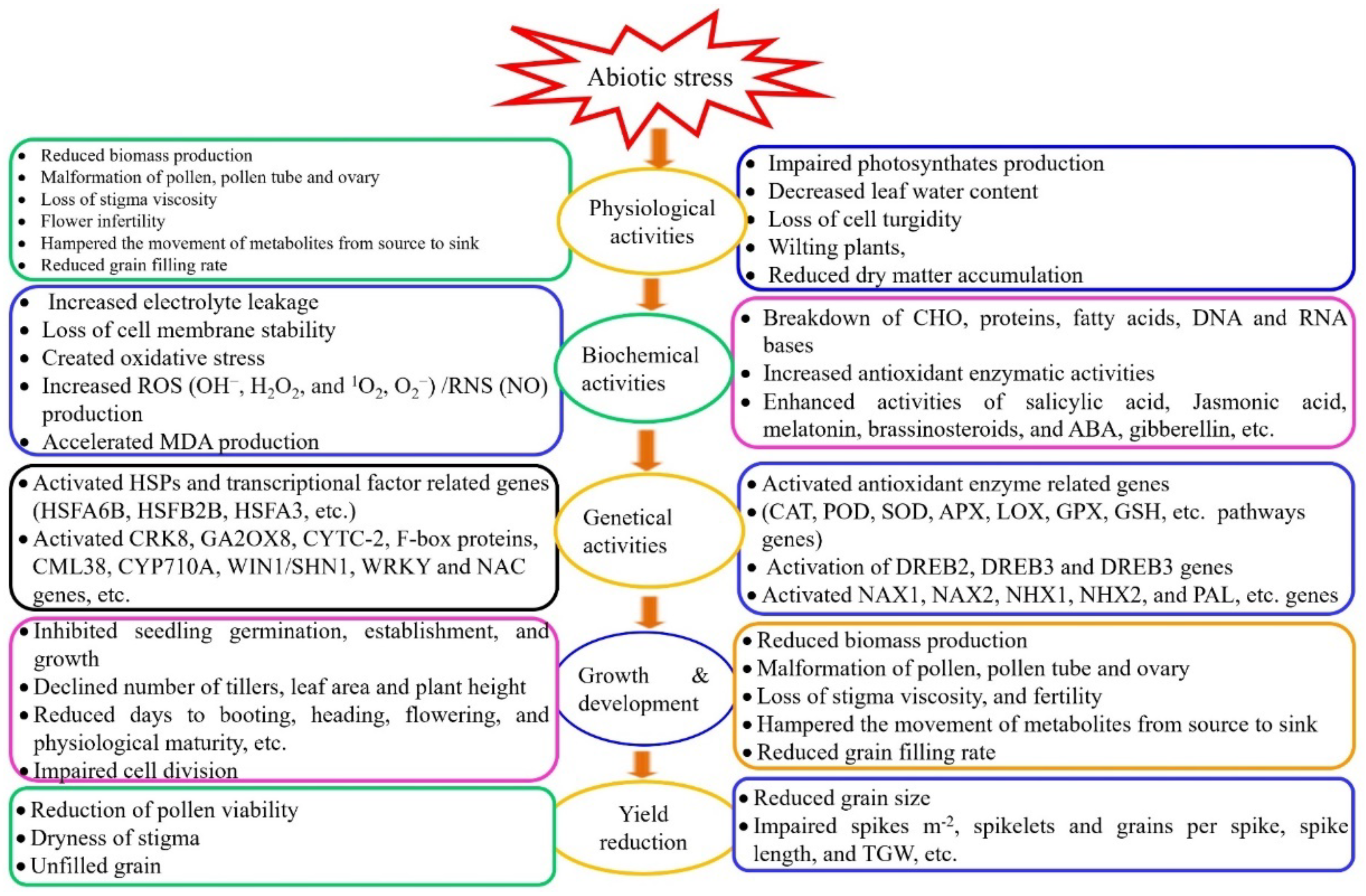
Figure 1.
Abiotic stress modulates physiological, biochemical, and genetical attributes of wheat which simultaneously affect its growth and development, resulting in yield reduction. CRK8 = Cysteine-rich RLK (RECEPTOR-like protein kinase) 8, GA2OX8 = Gibberellin 2-oxidase 8, CYTC-2 = Cytochrome C-2, F-box protein (F-box family protein), CML38 = Calmodulin38, CYP710A = Cytochrome P450, Family 710, Subfamily A, Polypeptide 1; WIN1 = Wax INDUCER1; SHN1 = SHINE1; NAC = NAM, ATAF and CUC; NAX1 and NAX2 are known as Na+ exclusive genes expressed as TmHKT1;4-A2 and TmHKT1;5-A, respectively found in durum wheat; NHX1 and NHX2 are depicted as Na+/H+ (K+) antiporter genes; PAL = Phenylalanine ammonia-lyase, DREB = Dehydration-responsive element binding protein, CAT = Catalase, POD = Peroxidase, SOD = Superoxide dismutase, APX = Ascorbate peroxidase, LOX = Lipoxygenase, GPX= Glutathione peroxidase, GSH = Glutathione (Figure was drawn by using MS Office 2021).
Figure 1.
Abiotic stress modulates physiological, biochemical, and genetical attributes of wheat which simultaneously affect its growth and development, resulting in yield reduction. CRK8 = Cysteine-rich RLK (RECEPTOR-like protein kinase) 8, GA2OX8 = Gibberellin 2-oxidase 8, CYTC-2 = Cytochrome C-2, F-box protein (F-box family protein), CML38 = Calmodulin38, CYP710A = Cytochrome P450, Family 710, Subfamily A, Polypeptide 1; WIN1 = Wax INDUCER1; SHN1 = SHINE1; NAC = NAM, ATAF and CUC; NAX1 and NAX2 are known as Na+ exclusive genes expressed as TmHKT1;4-A2 and TmHKT1;5-A, respectively found in durum wheat; NHX1 and NHX2 are depicted as Na+/H+ (K+) antiporter genes; PAL = Phenylalanine ammonia-lyase, DREB = Dehydration-responsive element binding protein, CAT = Catalase, POD = Peroxidase, SOD = Superoxide dismutase, APX = Ascorbate peroxidase, LOX = Lipoxygenase, GPX= Glutathione peroxidase, GSH = Glutathione (Figure was drawn by using MS Office 2021).
3. Approaches of abiotic stress management
3.1. Biochemical adaptation under abiotic oxidative stress
Proline application Osmotic adjustment hydrates plants during drought stress. Osmotic correction requires proline, glycine betaine, sucrose, alcohols, soluble sugars, organic acids, chloride ions, calcium, and potassium. Proline is biosynthesized in the cytosol and plastid but degenerates in mitochondria (Dudziak et al. 2019). It protects plant cell membranes, proteins, cellular redox potential, free radicals, and oxidative stress from abiotic stressors. Glutamate and ornithine give plants proline (Yan et al. 2021). Glutamate controls proline production. The glutamate pathway, which produces proline, involves two important enzymes, pyrroline 5-carboxylate synthetase (P5CS) and pyrroline 5-carboxylate reductase (P5CR) (Dar et al. 2016; Dudziak et al. 2019). When water is stressed, plants produce proline to minimize cell damage. Even at high doses, enzyme activity is unaffected. Proline was more easily accumulated and mobilized during drought stress, indicating increased tolerance (Dudziak et al. 2019). Similar to this, proline was referred to as an anti-drought defense molecule. Proline content and tolerance to drought are positively connected in wheat genotypes (Hong-Bo et al. 2006). Proline treatment of seeds enhances wheat development and productivity. Exogenous proline treatment during drought stress boosted the amounts of total phenolics, Chl, and proline. Additionally, they observed that exogenous proline treatment increased grain weight and quantity (Kamran et al. 2019; Farooq et al. 2017).
Glycine betaine (GB) application
GB regulates cytoplasmic pH, cell membrane shape, and antioxidant enzyme activity under drought stress. Exogenous GB protects chloroplasts and mitochondria against oxidative stress (Huseynova et al. 2016). Few higher plant enzymes make GB from choline and glycine. Plants survive abiotic stress with GB. Drought-stressed wheat synthesizes Chl, proline, and sucrose with GB foliar spray (Gupta et al. 2014). Polyamines application. Higher plant polyamines, such as spermine, spermidine, putrescine, etc. molecules actively respond to salt, dryness, and temperatures to boost photosynthetic capacity and water balance through osmotic adjustment (Alcazar et al. 2011). Spermine and spermidine molecules existed more in drought-tolerant wheat cultivar leaves than those that are highly sensitive to drought stress. Exogenous polyamine treatment was tested on drought-stressed wheat genotypes (Liu et al. 2016).
Carbohydrates application
Numerous CHO work as osmoprotectants in stressful settings to maintain essential enzyme activity, remove excess ROS, safeguard cell membrane and DNA integrity, and regulate osmotic pressure (Keunen et al. 2013; Rosa et al. 2009; Bowne et al. 2012). To protect against drought stress, wheat plants produced much soluble CHO when they reached the grain formation stage. This synthesized CHO was observed much more at this stage of wheat than in other stages (Farshadfar et al. 2008). Phenylalanine, peroxidase, ascorbic acid oxidase, etc. synthesis was enhanced with exogenous trehalose application. Trehalose and maltose priming boosts proline, flavonoids, amino acids, and phenolics accumulation, which improves wheat's resistance to drought stress (Aldesuquy and Ghanem 2015; Ibrahim and Abdellatif 2016). Using mannose and mannitol to treat wheat seed increased the quantity of CHO in wheat leaves, to adjust osmotic pressure when plants got drought stress (Hameed and Iqbal 2014).

Table 1.
Several genes responsible for abiotic stress tolerance in plants.
| Gene Name | Gene Symbol | Function | Reference |
| ECERIFERUM1 | TaCER1 | It is the CER1 enzyme that is responsible for the production of the very long-chain alkanes that are found in large concentrations during the late stage of wheat development. | Li et al. 2019 |
| Dehydration-responsive element binding protein 2 Dehydration-responsive element binding protein 3 Dehydration-responsive element binding protein 6 |
TaDREB2 TaDREB3 TaDREB6 |
DREBs are transcription factors, Transcriptional regulates involved in plant responses to drought, cold, and salt tolerance | Sadau et al. 2024 |
| Heat shock protein | TaHSP | Molecular chaperones protecting the proteome against environmental stresses; thermometry | Hill and Li 2022 |
| Heat shock transcription factor | TaHSF | Portion of signal transduction chains mediating the activating of genes responsive to both heat stress and other stresses | Charng et al. 2007 |
| Sodium/hydrogen antiporter Sodium/hydrogen exchanger 1 | TaNHX1 | K+/H+ antiporter is highly expressed in gourd cells. It is found in plasma membranes, tonoplast membranes, mitochondrial membranes, chloroplast membranes, and intercellular membranes | Athar et al. 2022 |
| Sodium/hydrogen antiporter | TaNHX2 | K+ and Na+/H+ antiporters are highly expressed in gourd cells, resulting in salt-tolerant stomatal conductance and turgor pressure | Yu et al. 2023 |
| NAX1 and NAX2 | - | Both genes regulate the exclusion of Na+ from plant leaves in saline soil by removing Na+ from the xylem | James et al. 2011 |
| Calmodulin | CML38 | Senses calcium levels and relays signals to calcium-sensitive enzymes, ion channels, and other proteins | Yang et al. 1996 |
| Proton-inorganic pyrophosphatase(AVP1) | TaAVP1 | ROS scavenging gene | Menadue et al. 2021 |
| Phenylalanine ammonia-lyase (PAL) | TaPAL | Polyphenol compounds biosynthesis like flavonoids, phenylpropanoids, and lignin in plants | Mamrutha et al. 2017 |
| SHINE1 | TaSHN1 | It controls plant wax biosynthesis | Bi et al. 2016 |
| NAC transcription factor 47 | TaNAC47 | NAC proteins regulate stress-related transcriptional reprogramming | Wang et al. 2015 |
| WRKY transcription factor 44 | TaWRKY44 | Proteins known as WRKY play important roles in the growth, development, and metabolic processes of plants as well as their responses to abiotic stresses. | Yu et al. 2023 |
| Glutathione peroxidase | TaGPX | GPX reduces hydrogen peroxide to water and oxygen and peroxide radicals to alcohols and oxygen. | Ursini and Maiorino 2012 |
| Mn-superoxide dismutase | TaMn-SOD | ROS (O2•–) scavenging gene | Vijayaraghavaredy et al. 2022 |
| Catalase | TaCAT | Antioxidant pathway-related, ROS (H2O2) scavenging gene | Caverzan et al. 2016 |
| Ascorbate peroxidase | TaAPX | APX enzymes catalyze the conversion of H2O2 to H2O using ascorbate as a specific electron donor | Li 2023 |
| Cu/Zn-superoxide dismutase | TaCu/Zn-SOD | ROS (O2•–) scavenging gene | Tyagi et al. 2017 |
| Iron-superoxide dismutase | TaFe-SOD | ROS (O2•–) scavenging gene | Himi et al. 2016 |
3.2. Controlling abiotic stress through upkeeping of antioxidant enzymatic pathways
Plants have defenses to prevent ROS-related cell damage. ROS and antioxidant activity dictate plant cell damage. Enzymatic and non-enzymatic activities reduced ROS accumulation (Moller et al. 2007; Srivalli et al. 2003). APX, monodehydroascorbate reductase (MDHAR), Dehydroascorbate reductase (DHAR), Glutathione S-transferase (GST), glutathione reductase (GR), and glutathione peroxidase (GPX) scavenged ROS. On the other hand, photosynthetic cell membrane integrity during oxidative stress remained undamaged by the activities of tocopherol, GSH, carotenoids, ascorbate (AsA), alkaloids, non-protein amino acids, and phenolic acids, which are known as non-enzymes (Mittler et al. 2004; Hasanuzzaman et al. 2012). Tolerant wheat genotypes lowered ROS activity by the over-expressions of MnSOD, Cu/Zn SOD, FeSOD, CAT, APX, GPX, and GST genes (Table 1) (Dudziak et al. 2019). SOD lessened ROS accumulation. Under drought stress, superoxide ion (O2•–) production was enhanced in mitochondria, peroxisome, cytoplasm, and plastome. SOD converted O2•– into H2O2 and finally H2O2 turned into O2 in those organs. Therefore, the SOD enzyme had crucial for drought tolerance (Budak et al. 2013). GST isoforms detoxified via glutathione peroxidases, which aids oxidative stress responses. CAT detoxified mitochondrion and microbody H2O2 to reduce oxidative damage. N2O reduced heat-induced oxidative damage in wheat seedlings via increasing CAT (Hasanuzzaman et al. 2012). Jasmonic acid-treated wheat seedlings enhanced salt tolerance via CAT. GR or APX activity increased during water stress recovery to protect cells from ROS. APX scavenged water deficit induced H2O2 and O2•– in plant chloroplasts. Leaf growth and dryness affect wheat APX activity. Mild drought boosted leaf APX activity, whereas severe drought inhibited it because of higher malonic MDA (Hollander-Czytko et al. 2005; Gill and Tuteja 2010). To assist plants, cope with abiotic stresses, GR lowers GSH, AsA, and GSH/GSSG ratios (Wang and Frei 2011).
Ascorbic acid (AA) and α-tocopherol
Ascorbic acid (AA), a water-soluble phytohormone, and tocopherol, a lipid-soluble phytohormone, protect plants from oxidative stress. Other four enzymes, such as DHAR, MDHAR, GR, and APX quench O2•– and H2O2 with ascorbate-glutathione activities oxidative stress (Hollander-Czytko et al. 2005; Wang and Frei 2011). AA and -tocopherol in wheat genotypes reduced leaf senescence by reducing H2O2 production during salt stress (Farouk 2011). Despite plants’ vulnerability to oxidative stress, carotenes are key antioxidant defense components. Beta-carotene is generated by chloroplast PSI and PSII. This chemical protects photosynthetic tissue by scavenging triplet Chl and prevents 1O2 formation. Carotenoids in drought-tolerant wheat genotypes reduce O2 oxidative damage (Wahid et al. 2007). Flavonoids protect plant cells against oxidative damage. Higher flavonoid concentrations in wheat leaves arise from drought-induced gene expression (Balouchi 2010).
3.3. Exogenous application of phytohormones in mitigating the effects of abiotic stress
Phytohormones are produced in small levels in many plant parts. The plant growth and development were boosted and adjusted to harsh conditions with phytohormones’ activities. Exogenous or endogenous Phytohormones can change plants’ physio-biological development to help them endure abiotic stress. IAA, GA, IBA, CK, ABA, ET, and SA are major phytohormones (Waadt et al. 2013; Sah et al. 2016).
Salicylic acid (SA)
By modulating the antioxidant defense system, SA can reduce oxidative stress. In abiotically stressed plants, SA influenced photosynthesis, nitrogen metabolism, proline metabolism, GB biosynthesis, and plant-water relations. Exogenous SA 10 mM alleviated wheat drought. SA (10 mM) eased stress by improving germination by 21%. Drought increased SA shoot length by 20%. SA increased water potential by 47%, proline 14%, and soluble sugar 25%. SA might enhance wheat growth in drought (Liu et al. 2022).
Abscisic acid (ABA)
Flowering plants contain growth-restricting phytohormone ABA. ABA, a plant signaling molecule, influences stress-induced growth (Xu et al. 2013). It affects flowering, stomatal opening, embryo morphogenesis, storage protein, lipid, and grain filling. ABA accumulates in chloroplasts. Light opens stomata; ABA and CO2 close them (Bano et al. 2012; Kim et al. 2010). ABA-activated wheat plants’ stress-response genes TIP1, (Temperature shock-inducible protein 1) (GLP-1) (Glucagon-like peptide-1), and MAPK4 (Mitogen-activated protein kinase 4-like). Plants face salinity, drought, and heat stresses; ABA synthesis is enhanced to ameliorate the stresses (Sah et al. 2016). Roots synthesize and transfer ABA to leaves to limit stomata opening and water loss during drought (Keskin et al. 2010; Schachtman and Goodger 2018). Exogenous ABA boosts antioxidant defense mechanisms and increases wheat's drought tolerance. Wheat's ABA synthesis improves root development and yield under drought (Xu et al. 2013; Bano et al. 2012).
Jasmonic acid (JA)
Jasmonic acid also known as jasmonate, is one of the most important cellular regulators. It is responsible for controlling a wide variety of physiological processes and for mediating responses to stress. Wheat plants mitigate the toxic effects of abiotic stress by increasing the synthesis of JA through overexpressing of JA biosynthetic gene (Qiu et al. 2014; Wang et al. 2020). JA (100µM) alleviated wheat drought. Drought reduced germination by 26%, although JA increased it by 27%. JA shoots grew 23% longer under drought. JA increased water potential by 60%, proline 14%, and soluble sugar 25%. JA can boost wheat growth in drought (Qiu et al. 2014; Wang et al.2020).
Gibberellic acid (GA)
Gibberellic acid promotes seed germination, leaf and stem growth, lateral root expansion, and blooming. It reduces free radical-caused lipid peroxidation. Plants generate GA fast when stressed (Gupta et al. 2013; Castro-Camba 2022). At 50ppm GA application for 24 hours, cross-Alborz and Sardari wheat cultivars demonstrated their maximum shoot and root length, as well as their maximum dry weight and germination rate. Osmo-priming for 12 hours boosted the length of the shoots and roots, as well as the dry weight and germination rate (Ghobadi et al. 2012).
Indole acetic acid (IAA)
IAA suppresses drought and salinity-induced lateral root development. IAA and Indole butyric acid (IBA) regulate plant cell division, elongation, differentiation, and apical domination Fu et al. 2015).
Ethylene (ET)
ET controls development under stress. ET in plant cells helps plants tolerate abiotic challenges such as temperature swings, salt, and drought (Khan and Khan 2014). Water stress increased wheat leaf ethylene production. Drought caused wheat leaf senescence (Narayana et al. 1991). ET is a plant growth regulator. It protects plants from stresses by regulating antioxidant enzyme activity (Khan and Khan 2014).
Melatonin
Melatonin is a ubiquitous, amphiphilic, and pleiotropic signaling chemical that protects plant and animal kingdoms from environmental challenges and has many cellular and physiological functions in wheat. Melatonin is a signal molecule during plant development and many abiotic stimuli. It protects plants from heat, cold, drought, salt, heavy metals, and UV radiation. As an antioxidant, it can scavenge free radicals (Turk et al. 2014; Alam et al. 2018b; Alam et al. 2021).
Brassinosteroids (BRs)
BRs regulate cell elongation, photomorphogenesis, xylem differentiation, seed germination, and adaptability to abiotic and biotic stressors (Alam et al. 2018a; Manghwar et al. 2022). 24-epibrassinolide (EBL) and 28-homobrassinolide are commercially available BRs. EBL enhanced K+, Ca2+, Mg2+, and NO3‾ uptake in the roots, which were transferred to the upper leaves. In younger leaves and roots, EBL improved K+/Na+, Ca2+/Na+, Mg2+/Na+, and NO3-/Cl homeostasis. BR-primed plants demonstrated higher antioxidative enzymes activities (SOD, CAT, POD, GR, APOX, MDHAR, and DHAR), Chl and protein content, and lowered ROS and RNS levels than controls (Fig 1) (Manghwar et al. 2022; Arora et al. 2012). At the cell surface, BR receptors can detect BR, which then activates Brassinazole-resistant 1 (BZR1) (Manghwar et al. 2022). BZR1 is responsible for regulating BR-responsive gene transcription in the nucleus.
3.4. Agronomic interventions
Salty soils can be recovered by mechanically scraping away salts, flushing, crusting, low-permeability soils with excellent water, and leaching saline soils with adequate structure and internal drainage. Alkali-sodic soils are recovered by draining excess sodium, deep plowing to mix in the calcareous subsoil, and adding acidifying minerals like pyrite and gypsum (Astolfi and Zuchi 2013). Plants with higher sulfur levels are salt-tolerant. Sulfate absorption plays a key function in salt-induced metabolic alterations. S-adenosyl methionine is a precursor of polyamines, which boost plant salt resistance (Chen et al. 2014).
3.5. Molecular strategies for mitigating abiotic stress effects on wheat crop
3.5.1. Molecular approaches to heat stress management
Most crops lack a single ‘thermotolerant’ gene. Different genes at different stages of crop growth may determine tolerance components. Quantitative trait loci (QTL) analysis can identify trait-causing genes. Once causal DNA is identified, it can be sequenced. This region’s DNA sequence can be compared to a database of known-function genes (Hill and Li 2022). Genetic diversity studies among cultivars, wild accessions, and ecotypes can help find novel QTL and alleles for high-temperature stress tolerance. Wild populations have high intraspecies variation in tolerance traits, which is absent in modern cultivars (Swamy et al. 2014). In contrast to a “constitutive” QTL, an “adaptive” QTL is one that is either only identified under certain environmental conditions or whose expression rises with the degree of environmental factors. Different wheat traits, including grain filling duration, canopy temperature depression, yield, and senescence, have QTL for high-temperature tolerance (Paliwal et al. 2012; Guan et al. 2018). Cellular membrane thermostability was correlated with seedling thermotolerance and yield of recombinant inbred line (RIL) grown in hot summer conditions (Fokar et al. 1998; Kumar et al. 2023). Mitochondrial respiration activity under high-temperature stress did not correlate with triphenyl-tetrazolium chloride reduction assay. Co-segregation analysis of winter wheat RILs suggests HSPs are involved in temperature tolerance. Crossing NW1014 and HUW468 produced RILs. Composite interval mapping found high-temperature tolerance in 2B, 7B, and 7D. Co-localized QTLs (in chromosomes 2B, 7B) elucidated 15% of phenotypic variation in 1000-grain weight, grain formation period, and lessened canopy temperature (Paliwal et al. 2012). A RIL population derived from Halberd and Cutter was evaluated 10 days after pollination. In the first year, the heat susceptibility index (HSI) was evaluated at 15% whereas 12% HSI was measured in the second year, resulting from two years of QTL analysis. Five QTL regions were discovered on chromosomes 1A-3B. These regions affected flag leaf length, width, and wax content. Under high-temperature stress, certain QTL areas revealed pleiotropic kernel number-weight trade-offs (Mason et al. 2010). Under high temperatures, the Babaxallele on linkage groups 1B, 3B, and 4A was associated with cooler canopy temperatures and higher grain yields. Seriallele on linkage group 2B caused cool canopy effects, but not yield. QTLs lowered canopy temperatures by 0.2 to 0.5°C (Pinto et al. 2008). QTL analysis was used to dissect maize RIL (T232 x CM37) high-temperature stress tolerance by analyzing cell-membrane stability (CMS) (Ottaviano et al. 1991). Six QTLs explained 53% of CMS genetic variability. In the same RIL population, researchers found five QTLs for high-temperature pollen germination and six for pollen tube growth (Frova and Sarigorla 1993). No significant correlation was found between HSPs and CMS. With additive, dominance, and over-dominance gene action, three maize QTLs linked to high seed drying temperature were found on chromosomes 6 and 8 and accounted for 39% of the trait's phenotypic variance (Salgado et al. 2008). Even when the growing conditions are similar, QTLs frequently do not connect well across genetic backgrounds, demonstrating lesser adaptation effects or disappearing entirely in different backgrounds. Candidate genes work better for QTL cloning when they regulate a related trait in another species (Ottaviano et al. 1991). An alternative to biological function is biochemical activity. The candidate gene approach works best when the biochemical activity underlying tolerance is known. Transcriptome profiling can be used to identify candidate genes (Alam et al. 2018a).
3.5.2. Molecular approaches to salt stress management
Plant Na+, K+/H+ antiporter genes were difficult to find. Genome sequencing shows that plants have many potential cation/proton antiporters, but their function is unknown (Rodríguez-Rosales et al. 2009). NHX family was first explored in plants. The original component, NHX1, catalyzes vacuole Na+ accumulation and salt tolerance. NHX1 and NHX2 regulate pH, K+, vesicle trafficking, and cell proliferation for plant development (Table 1) (Athar et al. 2022; Yu et al. 2023; Rodríguez-Rosales et al. 2009). Salty soil impacts plant growth from germination to harvest. Salinity affects germination, growth, water and nutrient intake, photosynthesis, yield, and enzyme activity. Higher salinity reduces germination, photosynthesis, transpiration, Na+ and Cl− ion buildup, and wheat's metabolic activities (Fig 1) (Yu et al. 2023). It may speed up ion entry, creating osmotic stress, limiting cell proliferation in early leaves and root tips, and closing stomata (Munns et al. 2008). Root-zone soil osmotic capability reduces salinity-induced crop growth. Na+ exclusion failure kills older leaves days or weeks after it emerges (James et al. 2011). Salt stress impacts all wheat stages. wheat's salt stress response at each stage helps find salt-tolerant genotypes. Leaves, roots, and vegetative plant elements (plant height, diameter, fresh and dry biomass) are affected. Ramified roots boost above-ground biomass in salinity. Early plant development is aided by a developed root system. Salt stress lowers leaf area, slowing photosynthesis and biomass production (Kingsbury et al. 19984). Ionic and osmotic pressure from salinity reduces wheat's physiological activity. Plant development, mineral dispersion, calcium-by-sodium separation, and membrane permeability (Mehta et al. 1983). Proline, glycine betaine, abscisic acid, and polyol protect vacuole osmotic potential from ion toxicity in plants. Solutes or metabolites protect plants from salt stress (Sharma and Garg 1983). Overproducing enzymes and solutes help select salt-resistant genotypes. Low Na+ concentrations and high proline and chlorophyll contents at the seedling stage affect salt stress yield and can be used as a breeding criterion. Salt-related traits improve plant genetics under saline conditions. They examined coastal salt-tolerant rice grain yield and related traits (Manohara et al. 2020). Transgenic approaches for the mitigation of abiotic stress. Stress activates enzymatic, structural, and regulatory genes. Transgenic investigations started with single-action genes. The first targets were water channel proteins, osmolyte biosynthesis (proline, betaine, sugars like trehalose, and polyamines), detoxifying enzymes, and transport proteins. However, stress tolerance is regulated by multiple genes simultaneously, thus single-gene tolerance is implausible. Plants have stress-induced regulatory genes. One gene can regulate many stress-related proteins. In all species, temperature boosts HSP transcription (Schramm et al. 2007; Charng et al. 2007). Stress converts a non-DNA-binding monomeric form of HSF into a DNA-binding trimeric form (Table 1). High temperatures create HSPs (ul Haq et al. 2019). Transgenic plants with higher plastid EF-Tu reduced thylakoid membrane damage and boosted photosynthetic rate and grain yield during high-temperature stress. EF-Tu may protect photosynthetic membranes and photosynthesis-related enzymes from high-temperature stress, boosting CO2 fixation. Detox gene overexpression of CBF1/DREB1B genes increased rice, wheat, and canola’s temperature tolerance (Table 1) (Sadau et al. 2024). Co-expression of Escherichia coli P5C biosynthetic enzymes GK74 and GPR in Arabidopsis and tobacco increased proline biosynthesis. Transgenic plants have increased high-temperature stress tolerance due to cell wall proline-rich proteins. Simultaneous co-expression of DHAR, GR or GST, and GR in tobacco plants boosted temperature stress tolerance (Table 1) (Reguera et al. 2012). ROS are poisonous molecules that can cause oxidative damage to lipids, proteins, and DNA. Overexpression of Mn-superoxide dismutase in wheat increased temperature tolerance and field yields. APX and CAT detoxify H2O2. The cAPX gene improves tomato’s heat tolerance (Gusta et al. 2009). Combining antioxidant enzyme expression may improve high-temperature stress resistance. Constitutive expression of MBF1c in Arabidopsis thaliana increases transgenic plants’ tolerance to bacterial infection, high temperature, and osmotic stress. Enhanced transgenic plant resistance to osmotic and high-temperature stress was maintained when combined (Tian et al. 2013). Transcriptomic profiling and inhibitor experiments reveal that MBF1c expression increases transgenic plant tolerance to high temperature and osmotic stress by partially activating the ethylene-response signal transduction system (Suzuki and Mittler 2006). Katiyar-Agarwal et al. (2003) improved a high-temperature-tolerant transgenic rice line (Pusa basmati). Almost all transgenic plants recovered after 45-50°C heat stress and grew vigorously at 28°C, while untransformed plants did not. Overexpressing sHSP17.7 produces high-temperature-tolerant rice plants. mtHSP70 overexpression reduces programmed cell death in rice protoplasts via preserving mitochondrial membrane potential and limiting ROS signal amplification (Murakami et al. 2004).
Conclusions and Recommendations
Wheat plants are oxidized by abiotic stress. ROS/RNS damage cell membranes and photosynthetic equipment, decreasing wheat growth and production. Wheat plants have antioxidants both enzymatic and non-enzymatic that increase their resistance to stress. Abiotic stress reduces grain yield affecting physiological, growth, and yield processes (Fig 1). Wheat’s ability to withstand the effects of abiotic stress can be improved, leading to increased productivity and improved food security. Abiotic stress tolerance breeding is crucial not just in current climate change, but also in future climatic alteration with harsh environmental stress episodes. Wild type and landraces may contribute to abiotic stress-tolerant wheat; however, many germplasm collections lack the most tolerant wild accessions and landraces. Innovations in molecular technology are helping to identify wild-type wheat and generate new and improved cultivars in the future. Many of these resources are unexplored but valuable. Wild genetic resources can give chances for introgression, cloning, and transformation of essential genes and QTLs from wild type to cultivated crops using classical and current techniques for crop development. The lack of wild relatives in our collections is a major obstruction. The need to find genetic variability and new sources of tolerance, as well as to use all existing genetic resources, identify new resources, and protect them for the future, is heightened by the fact that environmental conditions are constantly shifting and climate variability is increasing in intensity. Species fitness relates to the germination and vitality of seeds in varied environments. For offspring (seeds) to pass on their quality to the next generation, they must live. Few data exist on the consequences of climate change, especially on appropriateness. Further research is needed to determine the role of wheat stress-induced.
Author Contributions
MNA and MZI were designed to write the manuscript. MNA, MTA, and MMH wrote it. MAF, ZC, MZI, and MMA revised, suggested, and improved it. All authors read and approved it for publication. All authors have read and agreed to the published version of the manuscript.
Funding
The Ministry of Agriculture, The Government of the People’s Republic of Bangladesh [Grant number 135015200; Fiscal year: 2023-24] funded to the Bangladesh Wheat and Maize Research Institute, Nashipur, Dinajpur, Bangladesh for research.
Acknowledgments
Sincere gratitude to Dr. Golam Faruq, Director General, Bangladesh Wheat and Maize Research Institute, Dinajpur, Bangladesh for valuable guidelines, suggestions, and comments for writing the paper.
Conflicts of Interests
The authors declare that they have no conflicts of interest.
References
- Abhinandan K, Skori L, Stanic M, Hickerson NMN, Jamshed M, Samuel MA (2018) Abiotic stress signaling in wheat-An inclusive overview of hormonal interactions during abiotic stress responses in wheat. Front Plant Sci 9:1-25. [CrossRef]
- Alabadi D, Oyama T, Yanovsky MJ, et al. (2001) Reciprocal regulation between TOC1 and LHY/CCA1 within the Arabidopsis circadian clock. Science 293:880–883. [CrossRef]
- Alam MN, Bodruzzaman M, Hossain MM, Sadekuzzaman M (2014) Growth performance of spring wheat under heat stress conditions. Int J Agron Agric Res 4(6):91-103.
- Alam MN, Akhter MM, Hossain MM, Rokonuzzaman (2013c). Performance of different genotypes of wheat (Triticum aestivum L.) in heat stress conditions. Intl J Biosci 3(8):295-306. =: =. [CrossRef]
- Alam MN, Akhter MM, Hossain MM, Mahbubul SM (2013b) Phenological changes of different wheat genotypes (Triticum aestivum L.) in high temperature imposed by late seeding. J Biodivers Environ Sci 3(8):83-93.
- Alam MN, Mannaf MA, Sarker MAZ, Akhter MM (2013a) Effect of terminal high temperature imposed by late sowing on phenological traits of wheat (Triticum aestivum L.). Intl J Agron Agric Res 3(3):6-10.
- Alam MN, Wang Y, Chan Z (2018b) Physiological and biochemical analyses reveal drought tolerance in cool-season tall fescue (Festuca arundinacea) turf grass with the application of melatonin. Crop Past Sci 69(10):1041-1049. [CrossRef]
- Alam MN, Yang L, Yi X, Wang Q-F, Robin AHK (2021) Role of melatonin in inducing the physiological and biochemical processes associated with heat stress tolerance in tall fescue (Festuca arundinaceous). J Plant Growth Regul. [CrossRef]
- Alam MN, Zhang L, Yang L, Islam MR, Liu Y, Luo H, et al. (2018a) Transcriptomic profiling of tall fescue in response to heat stress and improved thermotolerance by melatonin and 24- epibrassinolide. BMC Genomics 19:224. [CrossRef]
- Alcazar R, Cuevas JC, Planas J, Zarza X, Bortolotti C, Carrasco P, et al. (2011) Integration of polyamines in the cold acclimation response. Plant Sci 180:31-38. [CrossRef]
- Aldesuquy HS, Ghanem HE (2015) Exogenous salicylic acid and trehalose ameliorate short-term drought stress in wheat cultivars by up-regulating membrane characteristics and antioxidant defense system. Plant Omics 2(2):1-10. [CrossRef]
- Aroca R, Porcel R, Ruiz-Lozano JM (2012) Regulation of root water uptake under abiotic stress conditions. J Exp Bot 63:43–57. [CrossRef]
- Arora P, Bhardwaj R, Kanwar MK (2012) Effect of 24-epibrassinolide on growth, protein content and antioxidative defense system of Brassica juncea L. subjected to cobalt ion toxicity. Acta Physiol Plant 34(5):2007-2017. [CrossRef]
- Astolfi S, Zuchi S (2013) Adequate S supply protects barley plants from adverse effects of salinity stress by increasing thiol contents. Acta Physiol Plant 35(1):175-181. [CrossRef]
- Atanga RA, Tankpa V (2021) Climate change, flood disaster risk and food security nexus in northern Ghana. Front Sustain Food Sys. 5. [CrossRef]
- Athar R, Zulfiqar, F, Moosa A, et al. (2022) Salt stress proteins in plants: An overview. Front Plant Sci 13. [CrossRef]
- Balouchi HR (2010) Screening wheat parents of mapping population for heat and drought tolerance, detection of wheat genetic variation. Int J Bio Life Sci 4:63–73.
- Bano A, Ullah F, Nosheen A (2012) Role of abscisic acid and drought stress on the activities of antioxidant enzymes in wheat. Plant, Soil Environ 58:181–185. [CrossRef]
- Barlow KM, Christy BP, O’Leary GJ, Riffkin PA, Nuttall JG (2015) Simulating the impact of extreme heat and frost events on wheat crop production: A review. Field Crops Res 171:109-119. [CrossRef]
- Begara-Morales JC, Chaki M, et al. (2018) Nitric oxide buffering and conditional nitric oxide release in stress response. J Exp Bot 69:3425–3438. [CrossRef]
- Kumar H, Chugh, V, Kumar M, et al. (2023) Investigating the impact of terminal heat stress on contrasting wheat cultivars: A comprehensive analysis of phenological, physiological, and biochemical traits. Front Plant Sci 14. [CrossRef]
- Bowne JB, Erwin TA, Juttner J, et al. (2012) Drought responses of leaf tissues from wheat cultivars of differing drought tolerance at the metabolite level. Mol Plant 5:418–429.
- Budak H, Kantar M, Kurtoglu KY (2013) Drought tolerance in modern and wild wheat. Sci World J 548246. [CrossRef]
- Castro-Camba R, Sánchez C, Vidal N, Vielba JM (2022) Plant Development and Crop Yield: The Role of Gibberellins. Plants 11(19):2650. [CrossRef]
- Chauhan J, Prathibha M, Singh P, et al. (2023) Plant photosynthesis under abiotic stresses: Damages, adaptive, and signaling mechanisms. Plant Stress 10:100296. [CrossRef]
- Charng Y, Liu H, Liu N, Chi W, Wang C, Chang S, Wang T (2007) A heat-inducible transcription factor, hsfa2, is required for extension of acquired thermotolerance in Arabidopsis. Plant Physiol 143:251–262. [CrossRef]
- Caverzan A, Casassola A, Brammer SP (2016) Antioxidant responses of wheat plants under stress. Genet Mol Biol 39(1):1-6. [CrossRef]
- Chele KH, Tinte MM, Piater LA, Dubery IA, Tugizimana F (2021) Soil salinity, a serious environmental issue and plant responses: A metabolomics perspective. metabolites. 11(11):724. [CrossRef]
- Chen M, Chen J, Fang J, Guo Z, Lu S (2014) Down-regulation of S-adenosylmethionine decarboxylase genes results in reduced plant length, pollen viability, and abiotic stress tolerance. Plant Cell Tissue Organ Cult 116(3):311-322. [CrossRef]
- Chen Y, Guo Y, Guan P, et al. (2023) A wheat integrative regulatory network from large-scale complementary functional datasets enables trait-associated gene discovery for crop improvement. Mol Plant 16:393–414.
- Corpas FJ, Barroso JB (2014) Peroxynitrite (ONOO-) is endogenously produced in Arabidopsis peroxisomes and is overproduced under cadmium stress. Ann Bot 113(1):87-96. [CrossRef]
- Corpas FJ, Freschi L, Rodríguez-Ruiz M, et al (2018) Nitro-oxidative metabolism during fruit ripening. J Exp Bot 69:3449–3463. [CrossRef]
- Cromey MG, Wright DSC, Boddington HJ (1998) Effects of frost during grain filling on wheat yield and grain structure. New Zeal. J Crop Hortic Sci 26:279–290. [CrossRef]
- Dar MI, Naikoo MI, Rehman F, Naushin F, Khan FA (2016) Proline accumulation in plants: Roles in stress Emerging Omics Technologies; Springer: Berlin/Heidelberg, Germany, pp.155–166.
- Djanaguiraman M, Narayanan S, Erdayani E, Prasad PVV (2020) Effects of high-temperature stress during anthesis and grain filling periods on photosynthesis, lipids and grain yield in wheat. BMC Plant Biol 20(1):268. [CrossRef]
- Dudziak K, Zapalska M, Börner A, et al. (2019) Analysis of wheat gene expression related to the oxidative stress response and signal transduction under short-term osmotic stress. Sci Rep 9:2743. [CrossRef]
- Fábián A, Sáfrán E, Szabó-Eitel G, Barnabás B, Jäger K (2019) Stigma Functionality and Fertility Are Reduced by Heat and Drought Co-stress in Wheat. Front. Plant Sci 10:2019. [CrossRef]
- Farooq M, Nawaz A, Chaudhry MAM, Indrasti R, Rehman A (2017) Improving resistance against terminal drought in bread wheat by exogenous application of proline and gamma-aminobutyric acid. J Agron Crop Sci 203:464–472. [CrossRef]
- Farouk S (2011) Ascorbic acid and α-tocopherol minimize salt-induced wheat leaf senescence. J Stress Physiol Biochem 7:58-79. https://doaj.org/article/769740fc01124310b03f58b085b089a8.
- Farshadfar E, Ghasempour H, Vaezi H (2008) Molecular aspects of drought tolerance in bread wheat (T. aestivum). Pakistan Journal of Biol Sci 11:118-122. [CrossRef]
- Fewson CA, Nicholas DJ (1960) Utilization of nitric oxide by microorganisms and higher plants. Nature 188:794-796. [CrossRef]
- Fokar M, Nguyen HT, Blum A (1998) Heat tolerance in spring wheat. I. Estimating cellular thermotolerance and its heritability. Euphytica 104(1):1-8. [CrossRef]
- Foyer CH, Noctor G (2003) Redox sensing and signaling associated with reactive oxygen in chloroplasts, peroxisomes, and mitochondria. Physiol Plant 119:355–364. [CrossRef]
- Frederiks TM, Christopher JT, Borrell AK (2008) Low temperature adaption of wheat post head-emergence in northern Australia. In: (eds) Appels R, Eastwood R, Lagudah E, Langridge P, Mackay-Lynne M, The 11th International Wheat Genetics Symposium Proceedings. Sydney University Press.
- Frederiks TM, Christopher JT, Harvey GL, Sutherland MW, Borrell AK (2012) Current and emerging screening methods to identify post-head-emergence frost adaptation in wheat and barley. J Exp Bot 63:5405–5416. [CrossRef]
- Frederiks TM, Christopher JT, Sutherland MW, Borrell AK (2015) Post-head-emergence frost in wheat and barley: defining the problem, assessing the damage, and identifying resistance. J Exp Bot 66:3487–3498. [CrossRef]
- Frova C, Sarigorla M (1993) Quantitative expression of maize HSPs: genetic dissection and association with thermotolerance. Theor Appl Genet 86:213-220. [CrossRef]
- Fu SF, Wei JY, Chen HW, Liu YY, Lu HY, Chou JY (2015) Indole-3-acetic acid: A widespread physiological code in interactions of fungi with other organisms. Plant Signal Behavior 10(8). [CrossRef]
- Fuller MP, Fuller AM, Kaniouras S, Christophers J, and Fredericks T (2007) The freezing characteristics of wheat at ear emergence. Eur J Agron 26: 435–441. [CrossRef]
- Ghobadi M, Shafiei Abnavi M, et al. (2012) Effect of Hormonal Priming (GA3) and Osmo-priming on Behavior of Seed Germination in Wheat (Triticum aestivum L.). J Agric Sci 4(9). [CrossRef]
- Gill SS, Tuteja N (2010) Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol Biochem 48:909–930. [CrossRef]
- Guan P, Lu L, Jia L, et al. (2018) Global QTL Analysis identifies genomic regions on chromosomes 4A and 4B harboring stable loci for yield-related traits across different environments in wheat (Triticum aestivum L.). Front Plant Sci 9:529. [CrossRef]
- Gull A, Lone AA, Wani NUI (2019) Biotic and abiotic stresses in plants. In abiotic and biotic stress in plants; De Oliveira AB, Ed.; IntechOpen: London, UK.
- Gupta N, Sanjeev KT, Navtej SB (2014) Glycine betaine application modifies biochemical attributes of osmotic adjustment in drought-stressed wheat. Plant Growth Regul 72:221-228. [CrossRef]
- Gupta R, Chakrabarty SK (2013) Gibberellic acid in plant: still a mystery unresolved. Plant Signal Behav 8(9):e25504. [CrossRef]
- Gusta L, Benning N, Wu G, et al. (2009) Superoxide dismutase: an all-purpose gene for agri-biotechnology. Mol Breed 24:103–115. [CrossRef]
- Hameed A, Ahmed MZ, Hussain T, et al. (2021) Effects of salinity stress on chloroplast structure and function. Cells 10(8). [CrossRef]
- Hameed A, Iqbal N (2014) Chemo-priming with mannose, mannitol, and H2O2 mitigate drought stress in wheat. Cereal Res Commun 42:450. [CrossRef]
- Hasanuzzaman M, Nahar K, Alam MM, Fujita M (2012) Exogenous nitric oxide alleviates high temperature induced oxidative stress in wheat (Triticum aestivum L.) seedlings by modulating the antioxidant defense and glyoxalase system. Aust J Crop Sci 6:1314–1323.
- Hill CB, Li C (2022) Genetic improvement of heat stress tolerance in cereal crops. Agronomy 12(5):1205. [CrossRef]
- Hollander-Czytko H, Grabowski J, et al. (2005) Tocopherol content and activities of tyrosine aminotransferase and cystine lyase in Arabidopsis under stress conditions. J Plant Physiol 162:767–770. [CrossRef]
- Hong-Bo S, Xiao-Yan C, Li-Ye C, et al. (2006) Investigation on the relationship of proline with wheat anti-drought under soil water deficits. Colloids Surf B Biointerfaces. 53(1):113-9. [CrossRef]
- Hossain A, Islam MT, Islam MT (2020) Wheat (Triticum aestivum L.) in the rice-wheat systems of South Asia is influenced by terminal heat stress at late sown condition: A case in Bangladesh. In Plant Stress Physiology; IntechOpen: London, UK.
- Hsu YT and Kao CH (2004) Cadmium toxicity is reduced by nitric oxide in rice leaves. Plant Growth Regul 42:227-238. [CrossRef]
- Hu KD, Hu LY, Li YH, Zhang FQ, Zhang H, 2007. Protective roles of nitric oxide on germination and antioxidant metabolism in wheat seeds under copper stress. Plant Growth Regul 53:173-183. [CrossRef]
- Huseynova IM, Rustamova SM, Suleymanov SY, et al. (2016) Drought-induced changes in pho-to synthetic apparatus and antioxidant components of wheat (Triticum durum Desf.) varieties. Photosynth Res 130:215–223. [CrossRef]
- Ibrahim HA, Abdellatif YMR (2016) Effect of maltose and trehalose on growth, yield, and some biochemical components of wheat plant under water stress. Ann Agric Sci 61:267–274. [CrossRef]
- IPCC (Intergovernmental Panel on Climate Change) (2014) The physical science basis: working group I contribution to the fifth assessment reports of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge, United Kingdom and New York, NY, USA.
- Islam T, Croll D, Gladieux P, et al. (2016) Emergence of wheat blast in Bangladesh was caused by a South American lineage of Magnaporthe oryzae. BMC Biol 14:84. [CrossRef]
- James RA, Blake C, Byrt CS, Munns R (2011) Major genes for Na+ exclusion, Nax1 and Nax2 (wheat HKT1;4 and HKT1;5), decrease Na+ accumulation in bread wheat leaves under saline and waterlogged conditions. J Exp Bot 62(8):2939-47. [CrossRef]
- Kamran M, Shahbaz M, Ashraf M, Akram NA (2019) Alleviation of drought-induced adverse effects in spring wheat (Triticum aestivum L.) using proline as a pre-sowing seed treatment. Pak J Bot 41:621–632.
- Katiyar-Agarwal S, Agarwal M, Grover A (2003) Heat tolerant basmati rice engineered by over-expression of HSP101. Plant Mol Bio 51:677–686. [CrossRef]
- Keskin BC, Sarikaya AT, Yuksel B, Memon AR (2010) Abscisic acid regulated gene expression in bread wheat. Aust J Crop Sci 4:617–625.
- Keunen ELS, Darin P, Jaco V, Van Den Ende WIM, Cuypers ANN (2013) Plant sugars are crucial players in the oxidative challenge during abiotic stress: Extending the traditional concept. Plant Cell Environ 36:1242–1255. [CrossRef]
- Khan NA, Khan MIR (2014) The Ethylene: From senescence hormone to key player in plant metabolism. J Plant Biochem Physiol 2:e124. [CrossRef]
- Khorobrykh S, Havurinne V, Mattila H, Tyystjärvi E. Oxygen and ROS in photosynthesis. Plants (2020) 9(1):91. [CrossRef]
- Kim TH, Bohmer M, Hu H, Nishimura N, Schroeder J (2010) Guard cell signal transduction network: Advances in understanding abscisic acid, CO2, and Ca2+ signaling. Ann Rev Plant Bio 61:561–591.
- Kingsbury R, Epstein E (1984) Selection for salt-resistant spring wheat1. Crop Sci 24(2):310-315. [CrossRef]
- Klepper LA (1975) Evolution of nitrogen oxide gases from herbicide treated plant tissues. WSSA Abstracts 184:70.
- Laspina NV, Groppa MD, Tomaro ML, Benavides MP (2005) Nitric oxide protects sunflower leaves against Cd-induced oxidative stress. Plant Sci 169:323-330. [CrossRef]
- Li T, Sun Y, Liu T, Wu H, An P, Shui Z, et al. (2019) TaCER1- 1A is involved in cuticular wax alkane biosynthesis in hexaploid wheat and responds to plant abiotic stresses. Plant, Cell Environ. [CrossRef]
- Li S (2023) Novel insight into functions of ascorbate peroxidase in higher plants: More than a simple antioxidant enzyme. Redox Biol 64:102789. [CrossRef]
- Liao WB, Huang GB, Yu JH, Zhang ML (2012) Nitric oxide and hydrogen peroxide alleviate drought stress in marigold explants and promote its adventitious root development. Plant Physiol. Biochem 58:6-15. [CrossRef]
- Liu J, Qiu G, Liu C, Li H, Chen X, Fu Q, Lin Y, Guo B (2022) Salicylic acid, a multifaceted hormone, combats abiotic stresses in plants. Life (Basel) 12(6):886. [CrossRef]
- Liu Q, Nishibori N, Imai I, Hollibaugh JT (2016) Response of polyamine pools in marine phytoplankton to nutrient limitation and variation in temperature and salinity. Mar Ecol Prog Ser 544:93-105. [CrossRef]
- Ma Z, Hu L Jiang W (2024) Understanding AP2/ERF transcription factor responses and tolerance to various abiotic stresses in plants: a comprehensive review. Int J Mol Sci 25(2):893. [CrossRef]
- Menadue DJ, Riboni M, Baumann U, et al. (2021) Proton-pumping pyrophosphatase homeolog expression is a dynamic trait in bread wheat (Triticum aestivum). Plant Direct 5(10):e354. [CrossRef]
- Mamrutha HM, Kumar R, Kaur A, Kumar M (2017) Expressed sequence tags of PAL gene from Triticum aestivum L. cv. DPW. 621-650. https://www.ncbi.nlm.nih.gov/nuccore/JZ970255.
- Manghwar H, Hussain A, Ali Q, Liu F (2022) Brassinosteroids (BRs) Role in plant development and coping with different stresses. Int J Mol Sci 23(3). [CrossRef]
- Manohara K, Morajkar S, Shanbagh Y (2020) Genetic analysis of grain yield and its associated traits in diverse salt-tolerant rice genotypes under coastal salinity condition. J Cereal Res 12(3):290-296. [CrossRef]
- Marcellos H, Single WV (1984) Frost injury in wheat ears after ear emergence. Aust J Plant Physio l11:7-15. [CrossRef]
- Mason RE, Mondal S, Beecher FW, Pacheco A, Jampala B., et al. (2010) QTL associated with heat susceptibility index in wheat (Triticum aestivum L.) under short-term reproductive stage heat stress. Euphytica 174:423-436. [CrossRef]
- Mathur S, Allakhverdiev SI, Jajoo A (2011) Analysis of high temperature stress on the dynamics of antenna size and reducing side heterogeneity of Photosystem II in wheat leaves (Triticum aestivum). Biochim Biophys Acta Bioenerg 1807(1):22. [CrossRef]
- Mehta P, Jajoo A, Mathur S, Bharti S (2010) Chlorophyll a fluorescence study revealing effects of high salt stress on Photosystem II in wheat leaves. Plant Physiol Biochem 48(1):16-20. [CrossRef]
- Mittler R, Vanderauwera S, Gollery M, Van Breusegem F (2004) Reactive oxygen gene network of plants. Trends Plant Sci 9:490–498. [CrossRef]
- Miura K, Furumoto T (2013) Cold signaling and cold response in plants. Int J Mol Sci 14(3):5312-5337. [CrossRef]
- Moller IM, Jensen PE, Hansson A (2007) Oxidative modifications to cellular components in plants. Annu Rev Plant Bio 58:459–481. [CrossRef]
- Mukherjee A, Wang S-YS, Promchote P (2019) Examination of the climate factors that reduced wheat yield in Northwest India during the 2000s. Water 11:343. [CrossRef]
- Munns R, Tester M (2008) Mechanism of salinity tolerance. Ann Rev Plant Biol 59:651-681. [CrossRef]
- Murakami T, Matsuba S, Funatsuki H, et al. (2004) Overexpression of a small heat shock protein, sHSP17.7 confers both heat tolerance and UV-B resistance to rice plants. Mol Breed 13:165–175. [CrossRef]
- Mwadzingeni L, Hussein S, Tsilo TJ (2017) Variance components and heritability of yield and yield components of wheat under under drought-stressed and non-stressed conditions. Aust J Crop Sci 11(11): 1425-1430. [CrossRef]
- Narayana Y, Lalonde S, Saini HS (1991) Water-stress induced ethylene production in wheat. A fact or artifact? Plant Physiol 96:406–410. [CrossRef]
- Nezhadahmadi A, Prodhan ZH, Faruq G (2013) Drought tolerance in wheat. Sci World J 2–7. [CrossRef]
- Nakamichi N, Kiba T, Kamioka, M, et al. (2012) Transcriptional repressor PRR5 directly regulates clock-output pathways. Proc Natl Acad Sci U.S.A. 109:17123–17128. [CrossRef]
- Nover L, Bharti K, Döring P, et al. (2001) Arabidopsis and the heat stress transcription factor world: how many heat stress transcription factors do we need? Cell Stress. 6(3):177-89. [CrossRef]
- Ottaviano E, Sari-Gorla M, Pe E, Frova C (1991) Molecular markers (RFLPs and HSPs) for the genetic dissection of thermotolerance in maize. Theor Appl Genet 81:713–719. [CrossRef]
- Paliwal R, Röder MS, Kumar U, Srivastava JP, Joshi AK (2012) QTL Mapping of terminal heat tolerance in hexaploid wheat (T. aestivum L.). Theor Appl Genet 125:561-575. [CrossRef]
- Pinto S, Chapman SC, McIntyre CL, Shorter R. Reynolds M (2008) QTL for canopy temperature response related to yield in both heat and drought environments. In: (eds) Appels R, Eastwood R, Lagudah E, Langridge P, Mackay M. et al. (eds) Proceedings of the 11th International Wheat Genetics Symposium, 24–28 August. Brisbane, Australia. Sydney University Press, Sydney. Available at: http://ses. library.usyd.edu.au/bitstream/2123/3351/1/ P172.pdf.
- Procházková D, Sumaira J, Wilhelmová N, et al. (2014) Reactive nitrogen species and the role of no in abiotic stress. In: (eds) Ahmad P, Rasool S, Emerging technologies and management of crop stress tolerance, Academic Press, pp.249-266.
- Qiu Z, Guo J, Zhu A, Zhang L, Zhang M (2014) Exogenous jasmonic acid can enhance tolerance of wheat seedlings to salt stress Ecotox Environ Safety 104:202–208. [CrossRef]
- Radi R (2013) Peroxynitrite, a stealthy biological oxidant. J Biol Chem 288(37):26464-26472. [CrossRef]
- Rath JR, Pandey J, Yadav RM, et al. (2022) Temperature-induced reversible changes in photosynthesis efficiency and organization of thylakoid membranes from pea (Pisum sativum). Plant Physiol Biochem 185:144 154. [CrossRef]
- Raza A, Razzaq A, Mehmood SS, et al. (2019) Impact of climate change on crops adaptation and strategies to tackle its outcome: A review. Plants 8:34. [CrossRef]
- Reguera M, Peleg Z, Blumwald E (2012) Targeting metabolic pathways for genetic engineering abiotic stress-tolerance in crops. Biochim Biophy Acta 1819:186–194. [CrossRef]
- Rawat R, Takahashi N, Hsu PY, et al. (2011) REVEILLE8 and PSEUDO-REPONSE REGULATOR5 form a negative feedback loop within the Arabidopsis circadian clock. PLoS Genet. 7:e1001350. [CrossRef]
- Röder M, Thornley P, Campbell G, Bows-Larkin A (2014) Emissions associated with meeting the future global wheat demand: a case study of UK production under climate change constraints. Environ Sci Policy 39: 13–24. [CrossRef]
- Roeber VM, Bajaj I, Rohde M, Schelling T, Cortleven A (2021) Light acts as a stressor and influences abiotic and biotic stress responses in plants. Plant Cell Environ. 44:645–664. [CrossRef]
- Rodríguez-Rosales MP, Gálvez FJ, Huertas R, et al. (2009) Plant NHX cation/proton antiporters. Plant Signal Behav 4(4):265-276. [CrossRef]
- Rosa M, Hilal M, Juan AG, Fernando EP (2009) Low-temperature effect on enzyme activities involved in sucrose–starch partitioning in salt-stressed and salt-acclimated cotyledons of quinoa (Chenopodium quinoa Willd.) seedlings. Plant Physiol Biochem 47:300–307. [CrossRef]
- Rugnone ML, Faigon Soverna A, Sanchez SE, et al. (2013) LNK genes integrate light and clock signaling networks at the core of the Arabidopsis oscillator. Proc Natl Acad Sci USA. 110:12120–12125. [CrossRef]
- Sabagh EL, Islam A, Skalicky MS, et al. (2021) Salinity stress in wheat (Triticum aestivum L.) in the changing climate: Adaptation and Management Strategies. Front Agron 3. [CrossRef]
- Sadau SB, Liu Z, Ninkuu V, Guan L, Sun X (2024) DREB transcription factors are crucial regulators of abiotic stress responses in Gossypium spp. Plant Stress 11:100350. [CrossRef]
- Sah SK, Reddy KR, Li J (2016) Abscisic acid and abiotic stress tolerance in crop plants. Front Plant Sci 7:571. [CrossRef]
- Salgado KCPC, Von Pinho EVR, Guimaraes CT, et al. (2008) Mapping of quantitative trait locus associated with maize tolerance to high seed drying temperature. Genet Mol Res 7:1319–1326. [CrossRef]
- Sanchez A, Shin J, Davis SJ (2011) Abiotic stress and the plant circadian clock. Plant Signal. Beha. 6:223–231. [CrossRef]
- Sharma S, Garg O (1983) Comparative study of osmotic and salt stress effects on nitrate assimilation in wheat. Curr Agric 7:36-40.
- Shrivastava P, Kumar R (2015) Soil salinity: A serious environmental issue and plant growth promoting bacteria as one of the tools for its alleviation. Saudi J Biol Sci 22(2):123-131. [CrossRef]
- Singh C (1988) Modern techniques of raising field crops, 4th ed.; Oxford and IBH publishing Co. Pvt. Ltd.: New Delhi, India, p.46.
- Singh AK, Sharma L, Mallick N (2004) Antioxidative role of nitric oxide on copper toxicity to a chlorophycean alga, Chlorella. Ecotoxicol Environ Saf 59(2):223-7. [CrossRef]
- Serrano-Bueno G, Sanchez de Medina Hernandez V, and Valverde F (2021) Photoperiodic signaling and senescence, an ancient solution to a modern problem? Front. Plant Sci 12:634393. [CrossRef]
- Singh HP, Batish DR, Kaur G, Arora K, Kohli RK (2008) Nitric oxide (as sodium nitroprusside) supplementation ameliorates Cd toxicity in hydroponically grown wheat roots. Environ Exp Bot 63(1-3):158-167. [CrossRef]
- Srivalli B, Chinnusami V, Renu KC (2003) Antioxidant defense in response to abiotic stresses in plants. J Plant Bio 30:121–139.
- Sun C, Liu L, Zhou W, Lingli Lu, Jin C, Lin X (2017) Aluminum induces distinct changes in the metabolism of reactive oxygen and nitrogen species in the roots of two wheat genotypes with different aluminum resistance. J Agric Food Chem, 65 (43): 9419–9427. [CrossRef]
- Suzuki N, Mittler R (2006) Reactive oxygen species and temperature stresses: A delicate balance between signaling and destruction. Physiol Plant 126:45–51. [CrossRef]
- Swamy BP, Kaladhar K, Ashok RG, Viraktamath BC, Sarla N (2014) Mapping and introgression QTLs for yield and related traits in two backcross populations derived from O. sativa cv Swarna and two accessions of O. nivara. J Genet 93:643–654. [CrossRef]
- Tan J, Zhao H, Hong J, Han, Y, Li H, Zhao W (2008) Effects of exogenous nitric oxide on photosynthesis, antioxidant capacity and proline accumulation in wheat seedlings subjected to osmotic stress. World J Agric Sci 4:307-313.
- Tian F, Gong J, Zhang J, Zhang M, Wang G, Li A, Wang W (2013) Enhanced stability of thylakoid membrane proteins and antioxidant competence contribute to drought stress resistance in the tasg1 wheat stay-green mutant. J Exp Bot 64(6):1509-20. [CrossRef]
- Turk H, Erdal S, Genisel M, Atici O, Yavuz Demir Y, Yanmis D (2014) The regulatory effect of melatonin on physiological, biochemical, and molecular parameters in cold-stressed wheat seedlings. Plant Growth Regul 74:139-152. [CrossRef]
- Tyagi S, Sharma S, Taneja M, Kumar R, Sembi JK, Upadhyay SK (2017) Superoxide dismutases in bread wheat (Triticum aestivum L.): Comprehensive characterization and expression analysis during development and, biotic and abiotic stresses. Agri Gene 6:1-13. [CrossRef]
- ul Haq S, Khan A, Ali M, et al. (2019) Heat Shock Proteins: Dynamic biomolecules to counter plant biotic and abiotic stresses. Int J Mol Sci 20(21):5321. [CrossRef]
- Umbreen S, Lubega J, Cui B, Pan Q, Jiang J, Loake G (2018) Specificity in nitric oxide signaling. J Exp Bot 69:3439–3448. [CrossRef]
- Ursini F, Maiorino M (2012) Glutathione Peroxidases. In: Encyclopedia of Biological Chemistry,399-404. [CrossRef]
- Valluru R, Link J, Claupein W (2012) Consequences of early chilling stress in two Triticum species: plastic responses and adaptive significance. Plant Biol 14:641–651. [CrossRef]
- Vijayaraghavaredy P, Lekshmy SV, Struik PC, et al. (2022) Production and scavenging of reactive oxygen species confer to differential sensitivity of rice and wheat to drought stress. Crop Environ, 1(1):15-23. [CrossRef]
- Waadt R, Hsu PK, Schroeder JI (2015) Abscisic acid and other plant hormones: Methods to visualize distribution and signaling. Bioessays 37(12):1338-49. [CrossRef]
- Wahid A, Gelani S, Ashraf M, Foolad MR (2007) Heat tolerance in plants: An overview. Environ Exp Bot 61:199–223. [CrossRef]
- Wang X, Li Q, Xie J, Huang M, Cai J, Zhou Q, et al. (2020) Abscisic acid and jasmonic acid are involved in drought priming-induced tolerance to drought in wheat. [CrossRef]
- Wang X, Zeng J, Li Y, Rong X, Sun J, Sun T, et al. (2015) Expression of TaWRKY44, a wheat WRKY gene, in transgenic tobacco confers multiple abiotic stress tolerances. Front Plant Sci 6:615. [CrossRef]
- Wang, Y. and Frei, M. (2011) Stressed Food—The Impact of Abiotic Environmental Stresses on Crop Quality. Agriculture, Ecosystems & Environment, 141, 271-286. [CrossRef]
- Wardlaw IF, Dawson IA, Munibi P, Fewster R (1989) The tolerance of wheat to high temperatures during reproductive growth. I. Survey procedures and general response patterns. Aust J Agric Res 40:1–13. [CrossRef]
- Whaley JM, Kirby EJM, et al. (2004) Frost damage to winter wheat in the UK: the effect of plant population density. Eur J Agron 21:105–115. [CrossRef]
- Xu W, Jia L, Shi W, Liang J, Zhou F, Li Q, Zhang J (2013) Abscisic acid accumulation modulates auxin transport in the root tip to enhance proton secretion for maintaining root growth under moderate water stress. New Phytol 197:139–150. [CrossRef]
- Yadav S, Modi P, Dave A, Vijapura A, Patel D, Patel M (2020) Effect of abiotic stress on crops. In: (eds) Hasanuzzaman M, Minhoto MC, Filho T, Fujita M, Nogueira TAR Sustainable Crop Production, IntechOpen. [CrossRef]
- Yan L, Li S, Riaz M, Jiang C (2021) Proline metabolism and biosynthesis behave differently in response to boron deficiency and toxicity in Brassica napus. Plant Physiol Biochem 167:529-540. [CrossRef]
- Yang T, Segal G, Abbo S. Feldman M, Fromm H (1996) Characterization of the calmodulin gene family in wheat: structure, chromosomal location, and evolutionary aspects. Mol Gen Genet 252:684–694. [CrossRef]
- Yu Y, Song T, Wang Y, et al. (2023) The wheat WRKY transcription factor TaWRKY1-2D confers drought resistance in transgenic Arabidopsis and wheat (Triticum aestivum L.). Int J Biol Macromol 226:1203–1217. [CrossRef]
- Yue R, Zhou BO, Shimada IS, Zhao Z, Morrison SJ (2016) Leptin receptor promotes adipogenesis and reduces osteogenesis by regulating mesenchymal stromal cells in the adult bone marrow. Cell Stem Cell 18(6):782-796. [CrossRef]
- Zhao C, Liu B, Piao S, Wang X, Lobell DB, Huang Y, et al. (2017) Temperature increase reduces global yields of major crops in four independent estimates. Proc Natl Acad Sci USA, 114:9326-9331. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



