
Submitted:
09 September 2024
Posted:
11 September 2024
You are already at the latest version
Abstract
Cell viability refers to the proportion of living, healthy cells within a given population. This information is crucial in any cell-based research, especially in toxicology. Over the past century, numerous methods for assessing cell viability have been developed. These methods can be categorized into five main types: non-invasive cell structure damage, invasive cell structure damage, cell growth, cellular metabolism, and cell membrane potential. Each method operates on different principles and has its own advantages and disadvantages, making it essential for researchers to choose the method that best fits their experimental design. Despite the variety of cell viability methods available, most rely on indirect measurements of cell viability, which may compromise their accuracy. In this sense, there is insufficient information regarding their accuracy, highlighting the need for researchers to investigate the accuracy of both existing and future methods.
Keywords:
Cell viability
; cell-based methods
; in vitro toxicology
1. Introduction
Cell viability, defined as the proportion of living, healthy cells within a given population[1], is crucial in various fields. In the pharmaceutical industry, it plays a pivotal role in screening potential therapeutic agents and determining their safe dosage ranges [2]. In toxicology, cell viability assays help assess the safety of chemicals and environmental pollutants, ensuring compliance with regulatory standards [2]. In cancer research, these assays are essential for testing the efficacy of anti-cancer drugs and developing effective treatment strategies [3]. Ultimately, cell viability tests advance our understanding of cellular processes and the biological effects of compounds[4].
At first, measuring cell viability may sound trivial but it is difficult to determine whether a cell is alive or dead [5]. A cell is considered viable if it can perform its essential functions, which are many [6]. On the contrary, the NCCD considers a cell to be dead when the plasma membrane’s barrier function is irreversible lost, the cell forms apoptotic bodies or the cell is engulfed by professional phagocytes [7]. Considering the many different aspects that define a cell to be death (or not alive, as both are exhaustive terms), there are many different methods to test cell viability [8]. Some methods measure metabolic activity, which is crucial for a cell’s essential functions, such as the MTT assay and ATP assay [9]. Other methods assess cell growth or proliferation, as a dividing cell is considered viable, using assays like the proliferation assay and BrdU [9]. Additionally, some methods focus on cell structure integrity, particularly cell membrane integrity, using indicators like propidium iodide and acridine orange [9]. To organize these methods, the OECD has compiled a list of cell viability testing methods, which we will follow in this manuscript [9]. Researchers should select a method based on their specific endpoint, available resources (instrumentation, cost, or necessary skills), and the characteristics of the method.
This paper aims to provide a comprehensive overview of cell viability assays, highlighting their principles, applications, and comparative advantages. By understanding the strengths and limitations of each method, researchers can choose the most appropriate assay for their experimental needs. Additionally, this overview may help clarify the current landscape of cell viability methods and highlight the potential need for new approaches.
2. Cell Viability Methods
We will use the OECD list to categorize the various cell viability methods, briefly describe them, and summarize their endpoints, advantages, and disadvantages [9].
2.1. Structural Cell Damage (Non-Invasive)
With an optical microscope we can see cells and can even see internal cellular structures such as mitochondria (on average about 2 μm by 0.5 μm) [10,11]. With this in mind, researchers may analyse the overall cell shape and size of the cells that may be changed due to the toxic effect of compounds and may be an indicator of the cell viability (Figure 1 a) [12,13,14,15]. Concretely, the overall shape and size of the cells may be changed due to the compound’s toxic effect in the cell’s cytoskeleton or cell membrane channels [16,17]. In this sense, there are programs that may aid to a quantitative determination of cell shape change (ImageJ/Fiji, Cell Profiler, Icy or IDEAS) [18,19]. However, there are various factors that induce a change in cell morphology not related to the toxic effects of the compounds; cell attachment, osmolarity, metabolism [20,21]. Thus, it is difficult to determine whether the cell changed its morphology due to the toxic effect of the test compound.
There are other tests that are also non-invasive and are used to determine the structural cell damage based on markers that leak out of the cytoplasm of dead cells into cell culture medium [8]. Within this category, researchers may measure the release of compounds into the supernatant due to cell membranes rupture (Figure 1 b) [8,22]. The most common compound used in this type of method is lactate dehydrogenase (LDH) [8], yet, there are other compounds which are also used under the trademark of different manufacturers: adenylate kinase (AK) (ToxiLight™, LONZA), dead-cell protease (not specify, CytoTox™, Promega), Glyceraldehyde-3 Phosphate Dehydrogenase (G3PDH) (aCella™ - TOX, Promega), glucose 6-phosphate dehydrogenase (G6PD) (Vybrant™ Cytotoxicity Assay Kit or CyQUANT™, Thermo Fisher Scientific). All these compounds are cytoplasmic enzymes that are present in all cells. They are rapidly released out of the cell when the plasma membrane is damaged [22]. Since membrane permanent disruption is the definitive act of cell death [7], there is a correlation between the amount of these compounds in cell supernatant and cell viability. [22]. But the cell membrane may also release these compounds despite being intact, thus, there is also evidence of their leakage under stress or metabolism change conditions [23,24]. Furthermore, high background levels of these compounds, specially LDH, were observed in untreated samples [25,26]. Interestingly, a study found that the LDH method underestimates cytotoxicity when assessing cell viability in coculture with bacteria[27]. Farhana and Lappin [28] reported issues with long-term assays involving medium changes. Azqueta et al. noted that the LDH method is not very sensitive for determining cytotoxicity [29]).
2.2. Structural Cell Damage (Invasive)
This cell viability category is also based on the principle of membrane integrity, but in this case, the molecules cross the membrane of non-viable cells in the opposite direction (inwards) [8].
One of the oldest methods known to determine cell viability has this characteristic, the trypan blue method [30]. Trypan blue only penetrates dead cells’ damaged plasma membrane while is impermeable to plasma membrane of viable cells (Figure 2 a). The mechanism of selective staining for dead cells may actually relate to the impermeability of trypan blue aggregates [31]. In sense, prolonged incubation with trypan blue can stain viable cells because of the dissociation of trypan blue aggregates [31]. With this in mind, the incubation periods with trypan blue are shortened, which may lead to underestimation of dead cells [32]. Although there are many different methods optimised to reduce the frequency of these events [32]. Moreover, the method is inexpensive and there are many different benchtop instruments to measure cell viability (hemocytometer, Bio Rad TC10 / TC20 Automated Cell Counter; Olympus Cell Counter model R1; ThermoFisher Scientific Countess II Automated Cell Counter (fluorescent); Roche Cedex HiRes Analyzer; Nexcelom Bioscience Cellometer Auto T4) [31].
Aside from trypan blue dye, there are various other dyes that are only transported inside the non-viable cells, and once inside the cell, bind to different target molecules. Upon binding to these intracellular molecules, their fluorescence emissions can be measured after excitation with a specific wavelength [33]. Each dye has a unique composition, and there are many different types within this category: propidium iodide, Hoechst 33342, DRAQ7, acridine orange [34]. And some proprietary dyes: CellTox (Promega), SYTOX, YO-YO, TO-PRO-3 Iodide from Thermo-Fisher Scientific [31]. There are many related papers corroborating the correlation between the presence of these dyes and the viability of the cells [13,31,33,34]. Nevertheless, there are also many references to false-positive events when assessing cell viability with these dyes [35,36,37,38,39]. This can occur because the dye may penetrate the cell during a change in osmolarity, change in metabolism, due to spontaneous invagination [40].
Within this category, there are also lipid-soluble dyes that may cross the cell membrane due to their hydrophobic nature and are transformed by cellular enzymes (esterases) into lipid-insoluble fluorescent compounds that cannot escape from cells with intact membranes (Figure 2 b) [41,42]. In this case, the presence of a modified dye (cleaved by the cellular enzymes) determines the viable cells. As the LDH may be leaked in viable cells, the transformed dyes may also leak through viable cells. More so, considering the higher cell membrane permeability to lipid soluble molecules [43]. Moreover, the enzymes that cleaved the dyes when the cell was in a cell death pathway but they were synthesised at the time the cell was viable [44].
There are many tests that combine the latter types of dyes (the dyes that crossed the membrane when it’s compromised and the lipid soluble dyes that are cleaved by esterases) to have a better correlation. An example of this kind of test is the Life/dead kit (Thermo-Fisher Scientific).
Within this category we also consider the antibodies that attach to molecules related with the activation of cell-death-associated pathways (Figure 2 c) [45,46]. In apoptotic pathways two main events are targeted: caspase activation (detectable through enzymatic analysis or cell staining) [46] and endonuclease activation (observable as DNA fragmentation at the population level) [47]. Additionally, chromatin condensation (visible via DNA staining) and the externalization of phosphatidylserine on the plasma membrane (detectable using annexin staining) strongly correlate with apoptotic cell death [48]. However, in a cell population there may be alternative subpopulations that has different cell death pathway [49] which may be undetected when using a single endpoint. Moreover, the pathways are interconnected and there is a crosstalk of mechanisms between different pathways which difficult the selection of specific markers [50].
2.3. Cell Growth
There are many ways of counting cells from a given samples (optical microscope, flow cytometer, haematocytometer) [51], which may also be used to determine cell growth and subsequently, determine cytotoxicity. When observing the difference on cell number of a sample before and after the treatment with a given test compound, we may determine the number of cells that lysed after the treatment (Figure 3 a). Subsequently, when comparing the difference in cell numbers between this sample and untreated samples, we may determine the toxicity of the test compound [52].
Figure 3.
Graphical scheme of the cell viability methods based on cell growth. a) The cell count of a sample is assessed before and after treatment using an inverted microscope, or alternatively, a flow cytometer. b) Brdu or EdU are integrated into the cell DNA, making them visible in daughter cells and allowing for the precise quantification of replicated cells. c) The absorbance of Sulforhodamine B, which binds to cellular proteins, can be measured with a spectrophotometer. This measurement allows for the quantification of cell mass.
Figure 3.
Graphical scheme of the cell viability methods based on cell growth. a) The cell count of a sample is assessed before and after treatment using an inverted microscope, or alternatively, a flow cytometer. b) Brdu or EdU are integrated into the cell DNA, making them visible in daughter cells and allowing for the precise quantification of replicated cells. c) The absorbance of Sulforhodamine B, which binds to cellular proteins, can be measured with a spectrophotometer. This measurement allows for the quantification of cell mass.
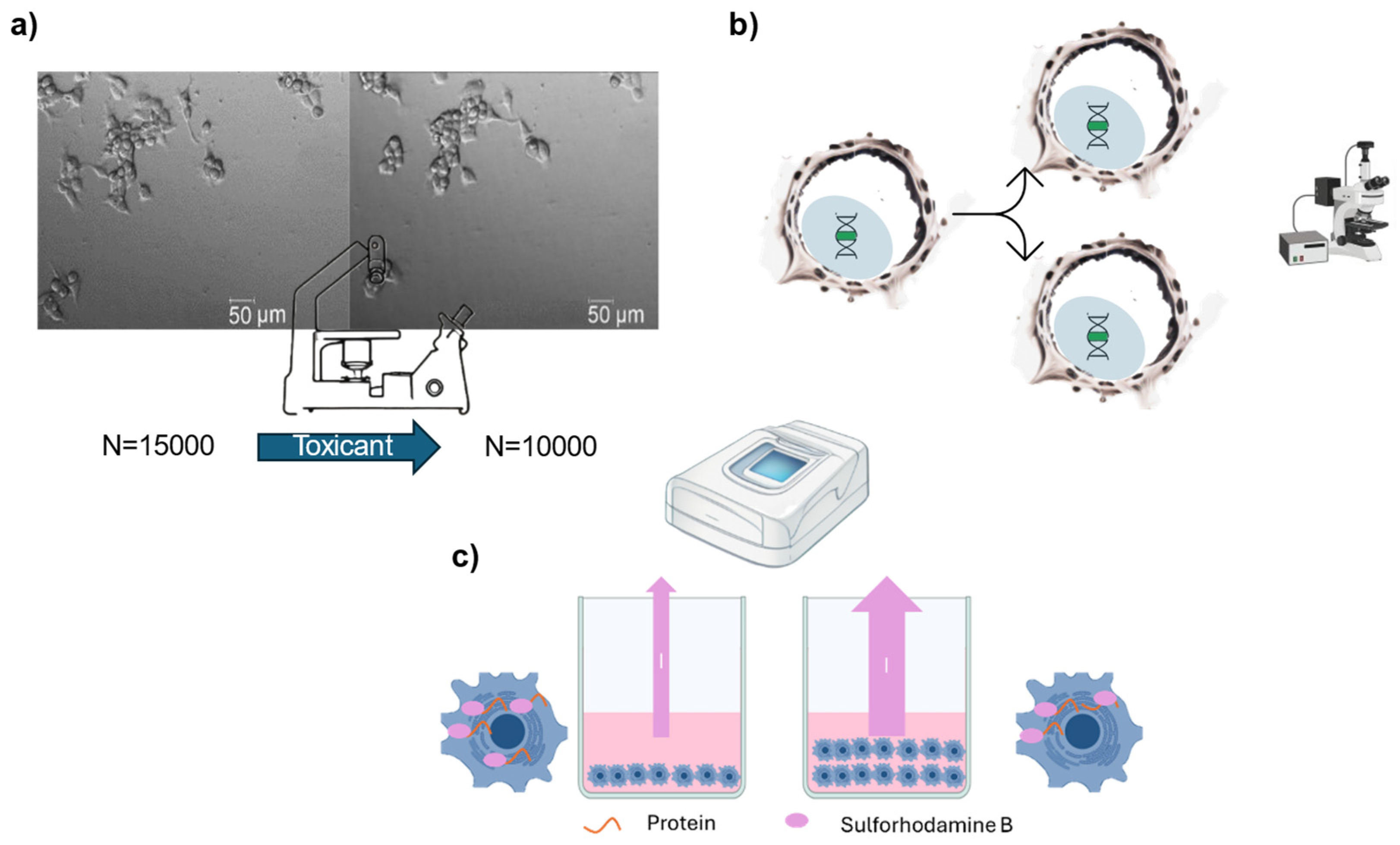
Measures new DNA synthesis based on incorporation of the easily detectable nucleoside analogues like Bromodeoxyuridine (BrdU) or EdU into DNA [53]. BrdU is an analogue of thymidine, a nucleoside involved in DNA synthesis. During the S-phase of the cell cycle, BrdU is incorporated into replicating DNA (Figure 3 b). BrdU or EdU can be detected e.g., by fluorescent-labelled antibodies in permeabilised cells [53]. BrdU/EdU cell division gives a very exact result about the number of cells that divided because the compound intercalates in the DNA at a known rate [53]. However, BrdU and Edu are cytotoxic and temper with the proliferation ability of the cells, thus giving unrealistic results [54,55]. The BrdU protocol is more time consuming than the EdU protocol, which doesn’t need no DNA denaturation step [53].
Cell growth may also be measured with fluorescence dyes that bind to cellular proteins or nucleic acids within cells, allowing for the quantification of cell density and protein content. Among the most common cytotoxicity tests within this category are Sulforhodamine B (SRB) and crystal violet (CV) [56,57]. Protein staining serves as a proxy for actual cell count, so this method measures the population mass rather than individual cells (Figure 3 c) [56,57], which may result in lower accuracy. Moreover, the use of crystal violet should be limited because its classification as a toxic and CMR (carcinogenic, mutagenic, or toxic for reproduction) chemical [58].
2.4. Cellular Metabolism
The used of dyes that are metabolized by the cell during its biochemical activities is widely used for the determination of cell viability. Most commonly the mitochondrial metabolism is targeted, concretely, the production of reducing equivalents such as NAD(P)H led to the reduction of the tetrazolium dye in viable cells. The resulting formazan is extracted and measured spectrophotometrically. The rate of formazan formation reflects the activity of essential cellular processes such as respiration [25,59]. These methods are high throughput, easy, low cost and have a high sensitivity. Moreover, there are very versatile, and can be even used for tissue constructs. There are many references to studies made with these methods and are used in several ISO standards and OECD test guidelines [59,60,61]. Still, this method measures the number of viable cells by indirect measurement, which has commonly known drawbacks [62]. These are especially related to the cells with reduced mitochondrial function, which may appear non-viable [62]. Inhibition of cell metabolism by the test item causes low values in the assay which is not necessarily related to cell viability [40]. Moreover, some test items interfere with the assay e.g., by reducing the dye because interference testing is recommended [62]. As we discuss with the SRB and CV methods, the measurements are not made in single cells and they rather show population wise metabolism (in this case) [63]. Some cell cultures need long time to reduce sufficient amount of dye (no sharp time point for viability definition) [64]. Assessment of kinetic of the reduction may be necessary to ensure proper selection of incubation time with a tetrazolium dye (to avoid reaching plateau of OD)[65].
Figure 4.
Graphical scheme of the cell viability methods based on cellular metabolism. a) Molecules are metabolized in the mitochondria at varying rates depending on the cell’s metabolism (viability). The viable cell on the left has a higher number of reduced molecules compared to the non-viable cell on the right, resulting in lower absorbance or fluorescence intensity in the latter. b) The mitochondrial membrane potential is measured using a dye that emits different intensities and bandwidths based on the mitochondrial membrane potential. c) Neutral red is transported within the cell by lysosomes. In viable cells, this transport is unaltered, while in non-viable cells, it is disrupted. The amount of neutral red within the cell is then measured, with higher levels found in viable cells. d) Viable cells produce more ATP than non-viable cells. After cell lysis, ATP content is measured using luminometric assays.
Figure 4.
Graphical scheme of the cell viability methods based on cellular metabolism. a) Molecules are metabolized in the mitochondria at varying rates depending on the cell’s metabolism (viability). The viable cell on the left has a higher number of reduced molecules compared to the non-viable cell on the right, resulting in lower absorbance or fluorescence intensity in the latter. b) The mitochondrial membrane potential is measured using a dye that emits different intensities and bandwidths based on the mitochondrial membrane potential. c) Neutral red is transported within the cell by lysosomes. In viable cells, this transport is unaltered, while in non-viable cells, it is disrupted. The amount of neutral red within the cell is then measured, with higher levels found in viable cells. d) Viable cells produce more ATP than non-viable cells. After cell lysis, ATP content is measured using luminometric assays.
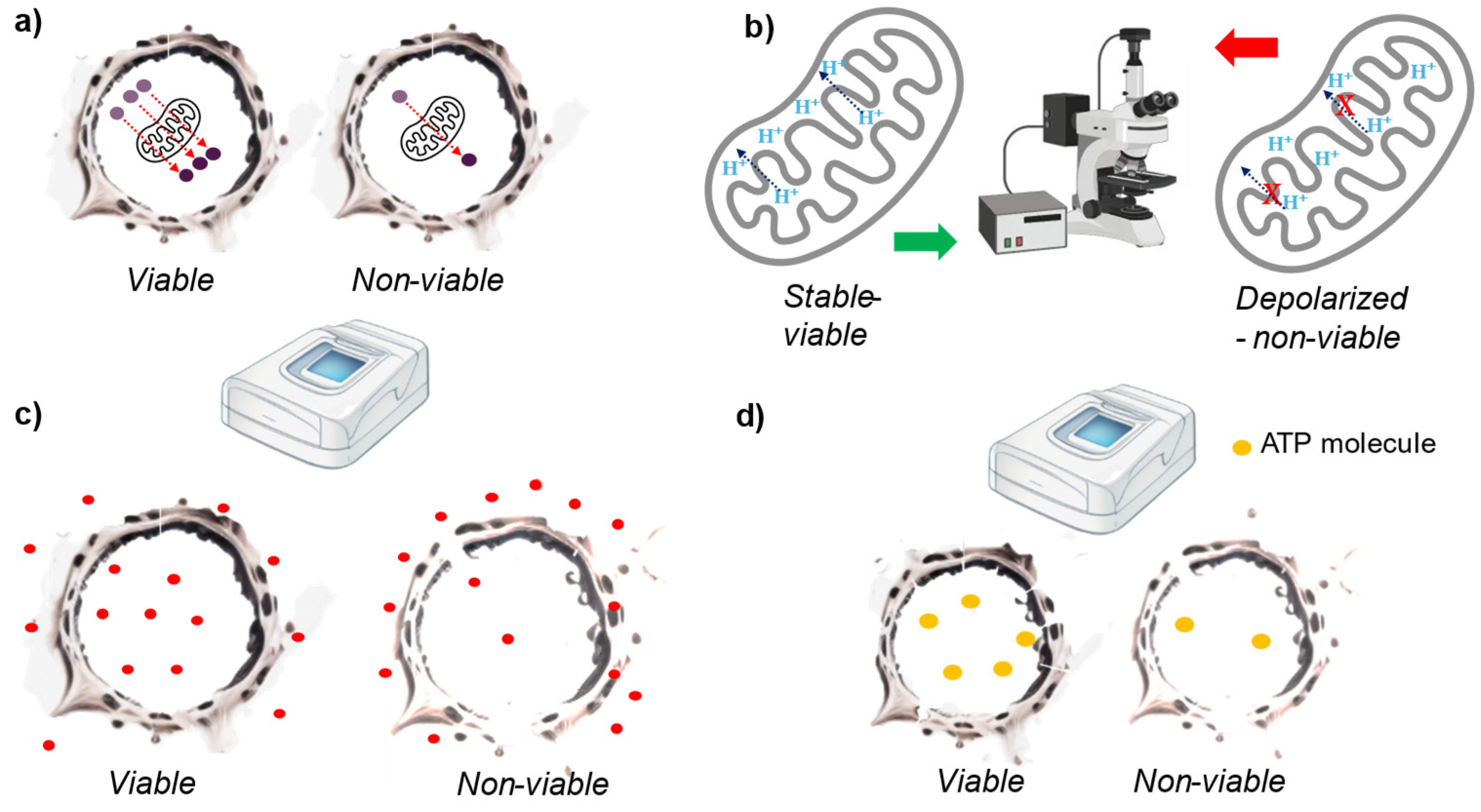
Resazurin reduction assay (sometimes called Alamar blue) is similar to tetrazolium reduction assays because it’s based on the metabolic activity of the mitochondria. In this case, resazurin is reduced to resorufin, which has an excitation peak at 571 nm and an emission peak at 584 nm [61]. Considering the similarity with the tetrazolium reduction assays this method have similar advantages and drawbacks as the tetrazolium reduction assay. The tests have a high sensitivity, it can be performed rapidly in multi-well dishes and is relatively low cost [61]. Although, cells with reduced mitochondrial function may appear non-viable and there are certain endogenous molecules that interfere with the assay (e.g., superoxide also reduces the dye) [66]. Furthermore, some cell cultures need a long time to reduce sufficient resazurin (no sharp time point for viability definition).
Other type of dyes directly measure mitochondrial activity rather than through molecule reduction. Specifically, dyes such as JC-1, TMRE, and MitoTracker assess mitochondrial membrane potential [67]. Changes in this potential are indicative of cell viability loss. Quantification is performed using HCI (High Content Imaging) or FACS (Fluorescence-Activated Cell Sorting) [67]. These methods provide single cell information, and they are also fast, inexpensive and high throughput [67]. Although, they are not as sensitive as the methods describe above. There are many artefacts that affect plasma membrane potential due to changes in shape and clustering of mitochondria [68]. Moreover, these dyes are prone to bleaching, quenching and unquenching. And specially JC-1 is toxic to the mitochondria [69].
Neutral red is used in a cell organelle function assay to assess lysosomal function. Active cells accumulate this red dye in their lysosomes while dying cells don’t. The dye incorporation is measured through spectrophotometric analysis [70]. Is low cost and is utilized in various ISO standards and OECD test guidelines, with a substantial historical database [71]. Typically provides information at the population level and is not suitable for tissue constructs or certain cell lines, especially for testing items that affect lysosome function [72,73]. Quantitative measurements require normalization, such as with protein content [71].
Dying cells fail to produce ATP, consume more ATP, and may lose ATP through plasma membrane perforations. By comparing the ATP levels of known viable cells, we can assess the viability of test cells. This involves preparing cell lysates and measuring the total ATP content using a luminometric assay [74]. Rapid and high-throughput test. The test is not invasive, and the cells are kept intact, which enables monitoring the cells for longer periods. Similar to most of the tests within this category, it measures viable cell mass rather than directly assessing cytotoxicity [8].
2.5. Membrane Potential
Despite the constant upgrade of existing cell viability methods and the designing of novel cell viability methods, these methods fall under the categories determine by the OECD. But this category is based on a new approach and thus, it is out of the scope of the OECD list. This approach aims to avoid the major drawbacks of the existing cell viability methods, the use of indirect measurements to determine cell viability. The methods of this category are based on direct measurement of cell viability. Concretely, both methods consider the cell death definition establish at the NCCD in 2015 that considers a cell as dead when the cell membrane losses its integrity [7]. In this sense, both methods measure the cell membrane potential to determine the cell membrane integrity (Figure 5).
2.5.1. The Membrane Potential Cell Viability Assay (MPCVA)
The Membrane Potential Cell Viability Assay (MPCVA) determines that a cell is dead when its cell membrane depolarizes for a long period of time (permanently). For that purpose, the method encompasses a fluorescence dye that anchors in the cell membrane and changes its intensity according to the membrane potential. Simultaneously, a cell membrane permeable DNA stain is added to measure the DNA content. This last step it’s important to improve the cell ability assessment accuracy, since doubling cells also depolarized for a relatively long periods of time [40]. The method is based on a direct approach to determine cell viability, thus less artifacts influence its cell viability determination in comparison to the methods based on an indirect approach [40]. The method is also compatible with live imaging because the fluorescence dye that anchors to the cell membrane is not toxic and don’t influence the courses of the cell [40]. In this case, there is no DNA staining because doubling cells are determined by simple observation (two daughter cells develop from a parent cell). Anyhow, the method is new and it hasn’t been tested in many alternative situations, where unpredicted artifacts may occur [40].
2.5.2. The DD Cell-Tox Method
The DD Cell-Tox method it’s also based on the cell membrane potential but in this case a more dynamic approach is considered [75]. Moreover, this method has a more holistic approach because it considers the various outcomes the cell has after exposing them to a toxic agent [75]. The cell number of a cell population is measured before and after the exposure of the toxic agents to them. After the exposure, the number of dead cells and doubling cells is determined with a membrane potential sensing dye and a DNA stain dye [75]. By combining these three elements (cell number before and after the exposure and the doubling cells) a prediction for the cell population dynamics is made [75]. The method assess the toxic effect of the compounds more accurately because it considers the various outcomes triggered by the toxic compounds [75]. The method also considers the cell population dynamics, and thus, provides valuable information about the effect of the toxic agent to the cell population [75]. The method it’s not compatible with live imaging unless there is a DNA dye that doesn’t influence the cell processes. The method has not been yet tested in different alternative situations, thus, there are many unknowns yet to be cover [75].
3. Discussion
This manuscript presented many different cell viability methods, their advantages and disadvantages. Among all the different methods, it’s up to the researcher to determine which method suits better the experimental design he determined.
If the experimental design requires monitoring the cells, a non-invasive method should be chosen because any alteration of the cell itself or its processes will influence the outcome of the study [101]. However, these categories of methods are not very sensitive [25,26] and if high sensitivity is required alternate methods should be chosen (Figure 3,a).
Figure 3.
Suggested decision tree for the desired method.
A metabolism-based method may be used if the experimental design does not require monitoring and the pathway triggered by the toxic compound is unknown. These methods are not specific to any cell death pathway and they are reliable [61] (Figure 3,b).
While, if the experimental design foresees metabolism changes another approach should be chosen. There are methods that rely on the cell membrane integrity, dyes that crossed the cell membrane when it’s allegedly disrupted are most commonly used (propidium iodide) [9] (Figure 3,c). However, there have been many studies that observed these dyes to cross intact cell membranes [35,36,37,38,39]. In this sense, the combination of these dyes with dyes that are esterified within the cell are widely used to avoid such issues, despite they too have drawbacks [102]. It’s worth noting that the esterases responsible for cleaving these dyes may have been synthesized when the cells were viable, even if the cells are currently undergoing a cell death pathway (but still have active esterases)[103].
Under similar conditions (unknown cell death pathway) the BrdU/EdU methods provide exact result about the number of cells that divided as these dyes are intercalated in the DNA [53]. But they are toxic and they may influence the cell viability [54,55]. Within the same principle (cell growth), there are fluorescence dyes that bind to cellular proteins, but there less accurate and have similar drawbacks [58].
If the pathway is known, a more specific method (using antibodies) may be selected to have a more comprehensive understanding of the expected cell death pathway [45,46]. Although, this method does not encompassed the alternative cell death pathway that may occur within the same cell population [49]. Thus, combining it with a general cytotoxicity test is necessary since techniques with antibodies may unintentionally select specific subpopulations and overestimate cell viability [45].
In this sense, a more universal method may aid the researchers evaluating the viability of all the subpopulations within a cell population, viability methods based on the cell membrane potential [5,40,75]. These methods should also have less artifacts since they assess cell death directly [40,75]. The MPCVA has the advantage of being compatible with live-imaging while the DD cell tox method is of special interest because encompass all the possible outcomes triggered by the tested molecule [40,75]. This is of special interest in the field of cancer research for testing anti-cancerogenic drug biological effect in the cell [75]. However, both methods are new and their accuracy and reliability needs to be tested in alternative situations.
It is also worth mentioning that new technologies aid cell viability assessment methods. In this sense, fluorescence emission may also be measured in different instruments with these capabilities (VARIOSKAN) [104]. Likewise new spectrophotometers are also capable of measuring luminescence, so the luminescence measurement is more available (VARIOSKAN). Nowadays, instruments capable of live-imaging are also more easily available, which are a powerful tool for cell viability studies [105]. Of special interest is also the multispectral imaging flow cytometer, known as Image stream MK2, which combines the flow cytometer and fluorescent microscope techniques [19].
Therefore, there are many methods and instrumentation to determine cell viability, and also many studies where the different cell viability methods are compared to each other [1,106,107]. Yet, there are not many studies where the accuracy of the methods is contested against a standard method [106]. The method should have single cell approach and consider that the cell death pathways may last up to 48 hours, with little to no false positives and false negatives [40]. Counting the cells 48 hours after their exposure to a toxic agent fulfils all these requirements and was successfully tested when testing the MPCVA method accuracy [40]. The method it’s time consuming but it is a great candidate to use as a standard method to test the accuracy of the cell viability methods, which are less time consuming. Then, if the tested methods have a comparable viability assessment, they may be used further for the cell viability determination under the same conditions.
4. Conclusions
Many cell viability methods have been developed during the last 100 years and some of them have been upgraded in the past years. New methods also have been developed taking advantage of the new knowledge and developed technology. Thus, there are many cell viability methods with different principles that may be suitable for various experimental designs. Each researcher may choose the one which is the most suitable for its experimental design depending on their needs. Yet, most of them rely on indirect measurement rather than measuring it directly. This raises a concern because the probability of having artifacts unrelated to cell viability that tempers with the assays, and ultimately, their accuracy. Moreover, there is no sufficient information regarding the accuracy of the cell viability methods and their accuracy assessment is done comparing the viability assessment between each method. A known reliable method should be selected to compare the rest of the methods, which may be the cell count method because it has the highest little to no false positives and false negatives [40].
5. Future Directions
Researchers should develop new cell viability methods based on new approaches that measures the cell viability directly. And they should challenge these new methods against a standard cell viability method that has high accuracy.
Conflicts of Interest
The authors declare no conflicts of interest
References
- Adan, A.; Kiraz, Y.; Baran, Y. , Cell Proliferation and Cytotoxicity Assays. Curr Pharm Biotechnol 2016, 17, 1213–1221. [Google Scholar] [CrossRef] [PubMed]
- Pognan, F.; Beilmann, M.; Boonen, H. C. M.; Czich, A.; Dear, G.; Hewitt, P.; Mow, T.; Oinonen, T.; Roth, A.; Steger-Hartmann, T.; Valentin, J. P.; Van Goethem, F.; Weaver, R. J.; Newham, P. , The evolving role of investigative toxicology in the pharmaceutical industry. Nat Rev Drug Discov 2023, 22, 317–335. [Google Scholar] [CrossRef] [PubMed]
- Mirzayans, R.; Andrais, B.; Murray, D. , Viability Assessment Following Anticancer Treatment Requires Single-Cell Visualization. Cancers (Basel) 2018, 10. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, Stuart L. ; Kotz, Joanne D.; Li, M.; Aubé, J.; Austin, Christopher P.; Reed, John C.; Rosen, H.; White, E. L.; Sklar, Larry A.; Lindsley, Craig W.; Alexander, Benjamin R.; Bittker, Joshua A.; Clemons, Paul A.; de Souza, A.; Foley, Michael A.; Palmer, M.; Shamji, Alykhan F.; Wawer, Mathias J.; McManus, O.; Wu, M.; Zou, B.; Yu, H.; Golden, Jennifer E.; Schoenen, Frank J.; Simeonov, A.; Jadhav, A.; Jackson, Michael R.; Pinkerton, Anthony B.; Chung, Thomas D. Y.; Griffin, Patrick R.; Cravatt, Benjamin F.; Hodder, Peter S.; Roush, William R.; Roberts, E.; Chung, D.-H.; Jonsson, Colleen B.; Noah, James W.; Severson, William E.; Ananthan, S.; Edwards, B.; Oprea, Tudor I.; Conn, P. J.; Hopkins, Corey R.; Wood, Michael R.; Stauffer, Shaun R.; Emmitte, Kyle A.; Brady, Linda S.; Driscoll, J.; Li, Ingrid Y.; Loomis, Carson R.; Margolis, Ronald N.; Michelotti, E.; Perry, Mary E.; Pillai, A.; Yao, Y., Advancing Biological Understanding and Therapeutics Discovery with Small-Molecule Probes. Cell 2015, 161, 1252–1265. [Google Scholar] [CrossRef]
- Madorran, E.; Stozer, A.; Bevc, S.; Maver, U. , In vitro toxicity model: Upgrades to bridge the gap between preclinical and clinical research. Bosn J Basic Med Sci 2019. [Google Scholar] [CrossRef]
- Oltvai, Z. N.; Barabási, A.-L.; Jeong, H.; Tombor, B.; Albert, R. , The large-scale organization of metabolic networks. Nature 2000, 407, 651–654. [Google Scholar] [CrossRef]
- Galluzzi, L.; Pedro, J. M. B.-s.; Vitale, I.; Aaronson, S. A.; Abrams, J. M.; Adam, D.; Alnemri, E. S.; Altucci, L.; Andrews, D. , Essential versus accessory aspects of cell death : recommendations of the NCCD 2015. 2015, 58-73. [CrossRef]
- Riss, T.; Niles, A.; Moravec, R.; Karassina, N.; Vidugiriene, J. , Markossian, S.; Grossman, A.; Arkin, M.; Auld, D.; Austin, C.; Baell, J.; Brimacombe, K.; Chung, T. D. Y.; Coussens, N. P.; Dahlin, J. L.; Devanarayan, V.; Foley, T. L.; Glicksman, M.; Gorshkov, K.; Haas, J. V.; Hall, M. D.; Hoare, S.; Inglese, J.; Iversen, P. W.; Lal-Nag, M.; Li, Z.; Manro, J. R.; McGee, J.; McManus, O.; Pearson, M.; Riss, T.; Saradjian, P.; Sittampalam, G. S.; Tarselli, M.; Trask, O. J., Jr.; Weidner, J. R.; Wildey, M. J.; Wilson, K.; Xia, M.; Xu, X., Eds. Eli Lilly & Company and the National Center for Advancing Translational Sciences: Bethesda (MD), 2004.Cells. In Assay Guidance Manual, Markossian, S.; Grossman, A., Arkin, M., Auld, D., Austin, C., Baell, J., Brimacombe, K., Chung, T. D. Y., Coussens, N. P., Dahlin, J. L., Devanarayan, V., Foley, T. L., Glicksman, M., Gorshkov, K., Haas, J. V., Hall, M. D., Hoare, S., Inglese, J., Iversen, P. W., Lal-Nag, M., Li, Z., Manro, J. R., McGee, J., McManus, O., Pearson, M., Riss, T., Saradjian, P., Sittampalam, G. S., Tarselli, M., Trask, O. J., Jr., *!!! REPLACE !!!*, Weidner, J. R., Wildey, M. J., Wilson, K., Xia, M., Eds.; Xu, X., Eds. Eli Lilly & Company and the National Center for Advancing Translational Sciences: Bethesda (MD), 2004; Xu, X., Eds. Eli Lilly &. [Google Scholar]
- OECD, In vitro toxicological tests. 2016.
- Uzman, A. , Molecular biology of the cell (4th ed.): Alberts, B., Johnson, A., Lewis, J., Raff, M., Roberts, K., and Walter, P. Biochemistry and Molecular Biology Education 2003, 31, 212–214. [Google Scholar] [CrossRef]
- Chalmers, S.; Saunter, C.; Wilson, C.; Coats, P.; Girkin, J. M.; McCarron, J. G. , Mitochondrial motility and vascular smooth muscle proliferation. Arterioscler Thromb Vasc Biol 2012, 32, 3000–3011. [Google Scholar] [CrossRef]
- Seal, S.; Yang, H.; Vollmers, L.; Bender, A. , Comparison of Cellular Morphological Descriptors and Molecular Fingerprints for the Prediction of Cytotoxicity- and Proliferation-Related Assays. Chemical Research in Toxicology 2021, 34, 422–437. [Google Scholar] [CrossRef]
- Munoz, L. E.; Maueröder, C.; Chaurio, R.; Berens, C.; Herrmann, M.; Janko, C. , Colourful death: six-parameter classification of cell death by flow cytometry--dead cells tell tales. Autoimmunity 2013, 46, 336–341. [Google Scholar] [CrossRef]
- Kühn, J.; Shaffer, E.; Mena, J.; Breton, B.; Parent, J.; Rappaz, B.; Chambon, M.; Emery, Y.; Magistretti, P.; Depeursinge, C.; Marquet, P.; Turcatti, G. , Label-free cytotoxicity screening assay by digital holographic microscopy. Assay Drug Dev Technol 2013, 11, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Kasprowicz, R.; Suman, R.; O’Toole, P. , Characterising live cell behaviour: Traditional label-free and quantitative phase imaging approaches. The International Journal of Biochemistry & Cell Biology 2017, 84, 89–95. [Google Scholar] [CrossRef]
- Louzao, M. C.; Ares, I. R.; Vieytes, M. R.; Valverde, I.; Vieites, J. M.; Yasumoto, T.; Botana, L. M. , The cytoskeleton, a structure that is susceptible to the toxic mechanism activated by palytoxins in human excitable cells. Febs j 2007, 274, 1991–2004. [Google Scholar] [CrossRef] [PubMed]
- Restrepo-Angulo, I.; De Vizcaya-Ruiz, A.; Camacho, J. , Ion channels in toxicology. Journal of Applied Toxicology 2010, 30, 497–512. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, A. B.; Dobson, E. T. A.; Rueden, C. T.; Tomancak, P.; Jug, F.; Eliceiri, K. W. , The ImageJ ecosystem: Open-source software for image visualization, processing, and analysis. Protein Sci 2021, 30, 234–249. [Google Scholar] [CrossRef]
- Zuba-Surma, E. K.; Kucia, M.; Abdel-Latif, A.; Lillard, J. W., Jr.; Ratajczak, M. Z. , The ImageStream System: a key step to a new era in imaging. Folia Histochem Cytobiol 2007, 45, 279–290. [Google Scholar]
- Urbanska, M.; Muñoz, H. E.; Shaw Bagnall, J.; Otto, O.; Manalis, S. R.; Di Carlo, D.; Guck, J. , A comparison of microfluidic methods for high-throughput cell deformability measurements. Nature Methods 2020, 17, 587–593. [Google Scholar] [CrossRef]
- Devendran, C.; Carthew, J.; Frith, J. E.; Neild, A. , Cell Adhesion, Morphology, and Metabolism Variation via Acoustic Exposure within Microfluidic Cell Handling Systems. Adv Sci (Weinh) 2019, 6, 1902326. [Google Scholar] [CrossRef]
- Kumar, P.; Nagarajan, A.; Uchil, P. D. , Analysis of Cell Viability by the Lactate Dehydrogenase Assay. Cold Spring Harb Protoc.
- Legrand, C.; Bour, J. M.; Jacob, C.; Capiaumont, J.; Martial, A.; Marc, A.; Wudtke, M.; Kretzmer, G.; Demangel, C.; Duval, D.; et al. , Lactate dehydrogenase (LDH) activity of the cultured eukaryotic cells as marker of the number of dead cells in the medium [corrected]. J Biotechnol 1992, 25, 231–243. [Google Scholar] [CrossRef]
- Mishra, D.; Banerjee, D. , Lactate Dehydrogenases as Metabolic Links between Tumor and Stroma in the Tumor Microenvironment. Cancers (Basel). [CrossRef]
- Fotakis, G.; Timbrell, J. A. , In vitro cytotoxicity assays: comparison of LDH, neutral red, MTT and protein assay in hepatoma cell lines following exposure to cadmium chloride. Toxicol Lett 2006, 160, 171–177. [Google Scholar] [CrossRef]
- Galluzzi, L.; Aaronson, S. A.; Abrams, J.; Alnemri, E. S.; Andrews, D. W.; Baehrecke, E. H.; Bazan, N. G.; Blagosklonny, M. V.; Blomgren, K.; Borner, C.; Bredesen, D. E.; Brenner, C.; Castedo, M.; Cidlowski, J. A.; Ciechanover, A.; Cohen, G. M.; De Laurenzi, V.; De Maria, R.; Deshmukh, M.; Dynlacht, B. D.; El-Deiry, W. S.; Flavell, R. A.; Fulda, S.; Garrido, C.; Golstein, P.; Gougeon, M. L.; Green, D. R.; Gronemeyer, H.; Hajnoczky, G.; Hardwick, J. M.; Hengartner, M. O.; Ichijo, H.; Jaattela, M.; Kepp, O.; Kimchi, A.; Klionsky, D. J.; Knight, R. A.; Kornbluth, S.; Kumar, S.; Levine, B.; Lipton, S. A.; Lugli, E.; Madeo, F.; Malomi, W.; Marine, J. C.; Martin, S. J.; Medema, J. P.; Mehlen, P.; Melino, G.; Moll, U. M.; Morselli, E.; Nagata, S.; Nicholson, D. W.; Nicotera, P.; Nunez, G.; Oren, M.; Penninger, J.; Pervaiz, S.; Peter, M. E.; Piacentini, M.; Prehn, J. H.; Puthalakath, H.; Rabinovich, G. A.; Rizzuto, R.; Rodrigues, C. M.; Rubinsztein, D. C.; Rudel, T.; Scorrano, L.; Simon, H. U.; Steller, H.; Tschopp, J.; Tsujimoto, Y.; Vandenabeele, P.; Vitale, I.; Vousden, K. H.; Youle, R. J.; Yuan, J.; Zhivotovsky, B.; Kroemer, G. , Guidelines for the use and interpretation of assays for monitoring cell death in higher eukaryotes. Cell death and differentiation 2009, 16, 1093–1107. [Google Scholar] [CrossRef] [PubMed]
- Van den Bossche, S.; Vandeplassche, E.; Ostyn, L.; Coenye, T.; Crabbé, A. , Bacterial Interference With Lactate Dehydrogenase Assay Leads to an Underestimation of Cytotoxicity. Front Cell Infect Microbiol 2020, 10, 494. [Google Scholar] [CrossRef] [PubMed]
- Farhana, A.; Lappin, S. L. , Biochemistry, Lactate Dehydrogenase. In StatPearls, StatPearls Publishing Copyright © 2024, StatPearls Publishing LLC.: Treasure Island (FL) ineligible companies. Disclosure: Sarah Lappin declares no relevant financial relationships with ineligible companies., 2024.
- Azqueta, A.; Stopper, H.; Zegura, B.; Dusinska, M.; Møller, P. , Do cytotoxicity and cell death cause false positive results in the in vitro comet assay? Mutat Res Genet Toxicol Environ Mutagen 2022, 881, 503520. [Google Scholar] [CrossRef] [PubMed]
- Pappenheimer, A. M. , EXPERIMENTAL STUDIES UPON LYMPHOCYTES : I. THE REACTIONS OF LYMPHOCYTES UNDER VARIOUS EXPERIMENTAL CONDITIONS. J Exp Med 1917, 25, 633–650. [Google Scholar] [CrossRef]
- Owen, S. C.; Doak, A. K.; Ganesh, A. N.; Nedyalkova, L.; McLaughlin, C. K.; Shoichet, B. K.; Shoichet, M. S. , Colloidal drug formulations can explain “bell-shaped” concentration-response curves. ACS Chem Biol 2014, 9, 777–784. [Google Scholar] [CrossRef]
- Kuijpers, L.; van Veen, E.; van der Pol, L. A.; Dekker, N. H. , Automated cell counting for Trypan blue-stained cell cultures using machine learning. PLoS One 2023, 18, e0291625. [Google Scholar] [CrossRef]
- Chiaraviglio, L.; Kirby, J. E. , Evaluation of impermeant, DNA-binding dye fluorescence as a real-time readout of eukaryotic cell toxicity in a high throughput screening format. Assay Drug Dev Technol 2014, 12, 219–228. [Google Scholar] [CrossRef]
- Foglieni, C.; Meoni, C.; Davalli, A. M. , Fluorescent dyes for cell viability: an application on prefixed conditions. Histochem Cell Biol 2001, 115, 223–229. [Google Scholar] [CrossRef]
- Boyd, V.; Cholewa, O. M.; Papas, K. K. , Limitations in the Use of Fluorescein Diacetate/Propidium Iodide (FDA/PI) and Cell Permeable Nucleic Acid Stains for Viability Measurements of Isolated Islets of Langerhans. Curr Trends Biotechnol Pharm 2008, 2, 66–84. [Google Scholar]
- Inde, Z.; Rodencal, J.; Dixon, S. J. , Quantification of drug-induced fractional killing using high-throughput microscopy. STAR Protoc 2021, 2, 100300. [Google Scholar] [CrossRef]
- Darzynkiewicz, Z.; Bedner, E.; Traganos, F. , Difficulties and pitfalls in analysis of apoptosis. Methods Cell Biol 2001, 63, 527–546. [Google Scholar] [CrossRef] [PubMed]
- Vitale, M.; Zamai, L.; Mazzotti, G.; Cataldi, A.; Falcieri, E. , Differential kinetics of propidium iodide uptake in apoptotic and necrotic thymocytes. Histochemistry 1993, 100, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Zamai, L.; Falcieri, E.; Marhefka, G.; Vitale, M. , Supravital exposure to propidium iodide identifies apoptotic cells in the absence of nucleosomal DNA fragmentation. Cytometry 1996, 23, 303–311. [Google Scholar] [CrossRef]
- Madorran, E.; Stožer, A.; Arsov, Z.; Maver, U.; Rožanc, J. , A Promising Method for the Determination of Cell Viability: The Membrane Potential Cell Viability Assay. 2022, 11, 2314. [CrossRef]
- Castro-Concha, L. A.; Escobedo, R. M.; de Miranda-Ham, M. L. , Loyola-Vargas, V. M.; Vázquez-Flota, F., Eds. Humana Press: Totowa, NJ, 2006; pp 71-76.Cultures. In Plant Cell Culture Protocols, Loyola-Vargas, V. M.; Vázquez-Flota, F., Eds. Humana Press: Totowa, NJ, 2006; Vázquez-Flota, F., Eds. Humana Press: Totowa, NJ, 2006. [Google Scholar]
- Kumar, P.; Srivastava, N.; Pande, M.; Prasad, J. K.; Sirohi, A. S. , Srivastava, N.; Pande, M., Eds. Springer Singapore: Singapore, 2017; pp 57-71.Integrity. In Protocols in Semen Biology (Comparing Assays), Srivastava, N.; Pande, M., Eds. Springer Singapore: Singapore, 2017; Pande, M., Eds. Springer Singapore: Singapore, 2017. [Google Scholar]
- Michel, C. C. , Chapter 9 - Microvascular Permeability and the Exchange of Water and Solutes Across Microvascular Walls. In Seldin and Giebisch’s The Kidney (Fifth Edition), Alpern, R. J.; Moe, O. W.; Caplan, M., Eds. Academic Press: 2013; pp 263-290.
- Heymann, E.; Mentlein, R.; Schmalz, R.; Schwabe, C.; Wagenmann, F. , A method for the estimation of esterase synthesis and degradation and its application to evaluate the influence of insulin and glucagon. Eur J Biochem 1979, 102, 509–519. [Google Scholar] [CrossRef] [PubMed]
- Kari, S.; Subramanian, K.; Altomonte, I. A.; Murugesan, A.; Yli-Harja, O.; Kandhavelu, M. , Programmed cell death detection methods: a systematic review and a categorical comparison. Apoptosis 2022, 27, 482–508. [Google Scholar] [CrossRef]
- Ai, X.; Butts, B.; Vora, K.; Li, W.; Tache-Talmadge, C.; Fridman, A.; Mehmet, H. , Generation and characterization of antibodies specific for caspase-cleaved neo-epitopes: a novel approach. Cell death & disease 2011, 2, e205. [Google Scholar] [CrossRef]
- Bajt, M. L.; Cover, C.; Lemasters, J. J.; Jaeschke, H. , Nuclear Translocation of Endonuclease G and Apoptosis-Inducing Factor during Acetaminophen-Induced Liver Cell Injury. Toxicological Sciences 2006, 94, 217–225. [Google Scholar] [CrossRef]
- Rieger, A. M.; Nelson, K. L.; Konowalchuk, J. D.; Barreda, D. R. , Modified annexin V/propidium iodide apoptosis assay for accurate assessment of cell death. Journal of visualized experiments : JoVE.
- Gao, J.; Xiong, A.; Liu, J.; Li, X.; Wang, J.; Zhang, L.; Liu, Y.; Xiong, Y.; Li, G.; He, X. , PANoptosis: bridging apoptosis, pyroptosis, and necroptosis in cancer progression and treatment. Cancer Gene Therapy 2024, 31, 970–983. [Google Scholar] [CrossRef]
- Kist, M.; Vucic, D. , Cell death pathways: intricate connections and disease implications. The EMBO journal 2021, 40, e106700. [Google Scholar] [CrossRef]
- Vembadi, A.; Menachery, A.; Qasaimeh, M. A. , Cell Cytometry: Review and Perspective on Biotechnological Advances. Front Bioeng Biotechnol 2019, 7, 147. [Google Scholar] [CrossRef]
- Luk, H.-Y.; McFarlin, B. K.; Vingren, J. L. , Using image-based flow cytometry to monitor satellite cells proliferation and differentiation in vitro. Methods (San Diego, Calif.) 2017, 112, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Flomerfelt, F. A.; Gress, R. E. , Analysis of Cell Proliferation and Homeostasis Using EdU Labeling. Methods in molecular biology (Clifton, N.J.) 2016, 1323, 211–220. [Google Scholar] [PubMed]
- Levkoff, L. H.; Marshall, G. P., 2nd; Ross, H. H.; Caldeira, M.; Reynolds, B. A.; Cakiroglu, M.; Mariani, C. L.; Streit, W. J.; Laywell, E. D. , Bromodeoxyuridine inhibits cancer cell proliferation in vitro and in vivo. Neoplasia 2008, 10, 804–816. [Google Scholar] [CrossRef] [PubMed]
- Haskins, J. S.; Su, C.; Maeda, J.; Walsh, K. D.; Haskins, A. H.; Allum, A. J.; Froning, C. E.; Kato, T. A. , Evaluating the Genotoxic and Cytotoxic Effects of Thymidine Analogs, 5-Ethynyl-2’-Deoxyuridine and 5-Bromo-2’-Deoxyurdine to Mammalian Cells. International journal of molecular sciences.
- Vichai, V.; Kirtikara, K. , Sulforhodamine B colorimetric assay for cytotoxicity screening. Nat Protoc 2006, 1, 1112–1116. [Google Scholar] [CrossRef] [PubMed]
- Feoktistova, M.; Geserick, P.; Leverkus, M. , Crystal Violet Assay for Determining Viability of Cultured Cells. Cold Spring Harbor Protocols, 0873. [Google Scholar] [CrossRef]
- Mani, S.; Bharagava, R. N. , Exposure to Crystal Violet, Its Toxic, Genotoxic and Carcinogenic Effects on Environment and Its Degradation and Detoxification for Environmental Safety. Rev Environ Contam Toxicol 2016, 237, 71–104. [Google Scholar] [CrossRef]
- van Meerloo, J.; Kaspers, G. J.; Cloos, J. , Cell sensitivity assays: the MTT assay. Methods in molecular biology (Clifton, N.J.) 2011, 731, 237–245. [Google Scholar] [CrossRef]
- Alépée, N.; Grandidier, M. H.; Cotovio, J. , Sub-categorisation of skin corrosive chemicals by the EpiSkin™ reconstructed human epidermis skin corrosion test method according to UN GHS: revision of OECD Test Guideline 431. Toxicol In Vitro 2014, 28, 131–145. [Google Scholar] [CrossRef]
- Hamid, R.; Rotshteyn, Y.; Rabadi, L.; Parikh, R.; Bullock, P. , Comparison of alamar blue and MTT assays for high through-put screening. 2004, 18, 703–710. [CrossRef]
- Ghasemi, M.; Turnbull, T.; Sebastian, S.; Kempson, I. , The MTT Assay: Utility, Limitations, Pitfalls, and Interpretation in Bulk and Single-Cell Analysis. International journal of molecular sciences. [CrossRef]
- Karakaş, D.; Ari, F.; Ulukaya, E. , The MTT viability assay yields strikingly false-positive viabilities although the cells are killed by some plant extracts. Turk J Biol 2017, 41, 919–925. [Google Scholar] [CrossRef]
- Wang, P.; Henning, S. M.; Heber, D. , Limitations of MTT and MTS-based assays for measurement of antiproliferative activity of green tea polyphenols. PLoS One 2010, 5, e10202. [Google Scholar] [CrossRef]
- Rai, Y.; Pathak, R.; Kumari, N.; Sah, D. K.; Pandey, S.; Kalra, N.; Soni, R.; Dwarakanath, B. S.; Bhatt, A. N. , Mitochondrial biogenesis and metabolic hyperactivation limits the application of MTT assay in the estimation of radiation induced growth inhibition. Scientific reports 2018, 8, 1531. [Google Scholar] [CrossRef]
- Damiani, E.; Solorio, J. A.; Doyle, A. P.; Wallace, H. M. , How reliable are in vitro IC(50) values? Values vary with cytotoxicity assays in human glioblastoma cells. Toxicol Lett 2019, 302, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Perelman, A.; Wachtel, C.; Cohen, M.; Haupt, S.; Shapiro, H.; Tzur, A. , JC-1 : alternative excitation wavelengths facilitate mitochondrial membrane potential cytometry. 2012, 3, e430–e437. [CrossRef]
- Keil, V. C.; Funke, F.; Zeug, A.; Schild, D.; Müller, M. , Ratiometric high-resolution imaging of JC-1 fluorescence reveals the subcellular heterogeneity of astrocytic mitochondria. Pflugers Arch 2011, 462, 693–708. [Google Scholar] [CrossRef] [PubMed]
- Perry, S. W.; Norman, J. P.; Barbieri, J.; Brown, E. B.; Gelbard, H. A. , Mitochondrial membrane potential probes and the proton gradient: a practical usage guide. Biotechniques 2011, 50, 98–115. [Google Scholar] [CrossRef] [PubMed]
- Zuang, V. , The neutral red release assay: a review. Altern Lab Anim 2001, 29, 575–599. [Google Scholar] [CrossRef]
- Ates, G.; Vanhaecke, T.; Rogiers, V.; Rodrigues, R. M. , Assaying Cellular Viability Using the Neutral Red Uptake Assay. Methods in molecular biology (Clifton, N.J.) 2017, 1601, 19–26. [Google Scholar] [CrossRef]
- Seglen, P. O. , Inhibitors of lysosomal function. Methods in enzymology 1983, 96, 737–764. [Google Scholar] [CrossRef]
- Hu, W.; Culloty, S.; Darmody, G.; Lynch, S.; Davenport, J.; Ramirez-Garcia, S.; Dawson, K.; Lynch, I.; Doyle, H.; Sheehan, D. , Neutral red retention time assay in determination of toxicity of nanoparticles. Marine environmental research 2015, 111, 158–161. [Google Scholar] [CrossRef]
- Fan, F.; Wood, K. V. , Bioluminescent Assays for High-Throughput Screening. ASSAY and Drug Development Technologies 2007, 5, 127–136. [Google Scholar] [CrossRef]
- Madorran, E.; Kocbek Šaherl, L.; Rakuša, M.; Takač, I.; Munda, M. , Finding a Direct Method for a Dynamic Process: The DD (Direct and Dynamic) Cell-Tox Method. International journal of molecular sciences 2024, 25, 5133. [Google Scholar] [CrossRef]
- Cho, M. H.; Niles, A.; Huang, R.; Inglese, J.; Austin, C. P.; Riss, T.; Xia, M. , A bioluminescent cytotoxicity assay for assessment of membrane integrity using a proteolytic biomarker. Toxicol In Vitro 2008, 22, 1099–1106. [Google Scholar] [CrossRef]
- Zhu, S.; Barbe, M. F.; Liu, C.; Hadjiargyrou, M.; Popoff, S. N.; Rani, S.; Safadi, F. F.; Litvin, J. , Periostin-like-factor in osteogenesis. Journal of cellular physiology 2009, 218, 584–592. [Google Scholar] [CrossRef] [PubMed]
- Strober, W. , Trypan Blue Exclusion Test of Cell Viability. Curr Protoc Immunol 2015, 111, A3. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Yang, J.; Zhang, Y.; Xi, Y.; Wen, X.; Yuan, D.; Wang, Y.; Wei, C.; Wang, R.; Wu, L.; Li, H.; Xu, C. , Exogenous H(2)S restores ischemic post-conditioning-induced cardioprotection through inhibiting endoplasmic reticulum stress in the aged cardiomyocytes. Cell & bioscience 2017, 7, 67. [Google Scholar] [CrossRef]
- Chazotte, B. , Labeling nuclear DNA with hoechst 33342. Cold Spring Harb Protoc, 5557. [Google Scholar] [CrossRef]
- Wlodkowic, D.; Akagi, J.; Dobrucki, J.; Errington, R.; Smith, P. J.; Takeda, K.; Darzynkiewicz, Z. , Kinetic viability assays using DRAQ7 probe. Current protocols in cytometry, 9. [CrossRef]
- Da Costa, R.; Redmann, K.; Schlatt, S. , Simultaneous detection of sperm membrane integrity and DNA fragmentation by flow cytometry: A novel and rapid tool for sperm analysis. Andrology 2021, 9, 1254–1263. [Google Scholar] [CrossRef]
- Jannoo, R.; Xia, Z.; Row, P. E.; Kanamarlapudi, V. , Targeting of the Interleukin-13 Receptor (IL-13R)α2 Expressing Prostate Cancer by a Novel Hybrid Lytic Peptide. Biomolecules. [CrossRef]
- Wlodkowic, D.; Faley, S.; Darzynkiewicz, Z.; Cooper, J. M. , Real-time cytotoxicity assays. Methods in molecular biology (Clifton, N.J.) 2011, 731, 285–291. [Google Scholar]
- Gustafson, D. L.; Viola, L. O.; Towers, C. G.; Das, S.; Duval, D. L.; Van Eaton, K. M. , Sensitivity of osteosarcoma cell lines to autophagy inhibition as determined by pharmacologic and genetic manipulation. Veterinary and comparative oncology 2023, 21, 726–738. [Google Scholar] [CrossRef]
- Lee-MacAry, A. E.; Ross, E. L.; Davies, D.; Laylor, R.; Honeychurch, J.; Glennie, M. J.; Snary, D.; Wilkinson, R. W. , Development of a novel flow cytometric cell-mediated cytotoxicity assay using the fluorophores PKH-26 and TO-PRO-3 iodide. J Immunol Methods. [CrossRef]
- Neri, S.; Mariani, E.; Meneghetti, A.; Cattini, L.; Facchini, A. , Calcein-acetyoxymethyl cytotoxicity assay: standardization of a method allowing additional analyses on recovered effector cells and supernatants. Clinical and diagnostic laboratory immunology 2001, 8, 1131–1135. [Google Scholar] [CrossRef]
- Yang, W.; Mu, B.; You, J.; Tian, C.; Bin, H.; Xu, Z.; Zhang, L.; Ma, R.; Wu, M.; Zhang, G.; Huang, C.; Li, L.; Shao, Z.; Dai, L.; Désaubry, L.; Yang, S. , Non-classical ferroptosis inhibition by a small molecule targeting PHB2. Nature Communications 2022, 13, 7473. [Google Scholar] [CrossRef]
- Virág, L.; Kerékgyártó, C.; Fachet, J. , A simple, rapid and sensitive fluorimetric assay for the measurement of cell-mediated cytotoxicity. Journal of Immunological Methods 1995, 185, 199–208. [Google Scholar] [CrossRef]
- Masson-Meyers, D. S.; Bumah, V. V.; Enwemeka, C. S. , A comparison of four methods for determining viability in human dermal fibroblasts irradiated with blue light. J Pharmacol Toxicol Methods 2016, 79, 15–22. [Google Scholar] [CrossRef]
- Im, K.; Mareninov, S.; Diaz, M. F. P.; Yong, W. H. , An Introduction to Performing Immunofluorescence Staining. Methods in molecular biology (Clifton, N.J.) 2019, 1897, 299–311. [Google Scholar] [CrossRef] [PubMed]
- Mfouo-Tynga, I.; Houreld, N. N.; Abrahamse, H. , Evaluation of cell damage induced by irradiated Zinc-Phthalocyanine-gold dendrimeric nanoparticles in a breast cancer cell line. Biomedical journal 2018, 41, 254–264. [Google Scholar] [CrossRef] [PubMed]
- Lakshmanan, I.; Batra, S. K. , Protocol for Apoptosis Assay by Flow Cytometry Using Annexin V Staining Method. Bio-protocol. [CrossRef]
- Huang, Q.; Wang, L.; Ran, Q.; Wang, J.; Wang, C.; He, H.; Li, L.; Qi, H. , Notopterol-induced apoptosis and differentiation in human acute myeloid leukemia HL-60 cells. Drug design, development and therapy 2019, 13, 1927–1940. [Google Scholar] [CrossRef] [PubMed]
- Peters, T.; Lindenmaier, H.; Haefeli, W. E.; Weiss, J. , Interaction of the mitotic kinesin Eg5 inhibitor monastrol with P-glycoprotein. Naunyn-Schmiedeberg’s Archives of Pharmacology 2006, 372, 291–299. [Google Scholar] [CrossRef]
- Ngamwongsatit, P.; Banada, P. P.; Panbangred, W.; Bhunia, A. K. , WST-1-based cell cytotoxicity assay as a substitute for MTT-based assay for rapid detection of toxigenic Bacillus species using CHO cell line. Journal of Microbiological Methods 2008, 73, 211–215. [Google Scholar] [CrossRef]
- Huyck, L.; Ampe, C.; Van Troys, M. , The XTT cell proliferation assay applied to cell layers embedded in three-dimensional matrix. Assay Drug Dev Technol 2012, 10, 382–392. [Google Scholar] [CrossRef]
- Bonnier, F.; Keating, M. E.; Wróbel, T. P.; Majzner, K.; Baranska, M.; Garcia-Munoz, A.; Blanco, A.; Byrne, H. J. , Cell viability assessment using the Alamar blue assay: A comparison of 2D and 3D cell culture models. Toxicology in Vitro 2015, 29, 124–131. [Google Scholar] [CrossRef]
- Desai, S.; Grefte, S.; van de Westerlo, E.; Lauwen, S.; Paters, A.; Prehn, J. H. M.; Gan, Z.; Keijer, J.; Adjobo-Hermans, M. J. W.; Koopman, W. J. H. , Performance of TMRM and Mitotrackers in mitochondrial morphofunctional analysis of primary human skin fibroblasts. Biochimica et Biophysica Acta (BBA) - Bioenergetics 2024, 1865, 149027. [Google Scholar] [CrossRef]
- Repetto, G.; del Peso, A.; Zurita, J. L. , Neutral red uptake assay for the estimation of cell viability/cytotoxicity. Nat Protoc 2008, 3, 1125–1131. [Google Scholar] [CrossRef]
- Caldwell, J. C. , cell death, or nutrient supply. In Tumour Site Concordance and Mechanisms of Carcinogenesis, Baan, R. A.; Stewart, B. W., Ed.; Straif, K., Eds. International Agency for Research on Cancer © International Agency for Research on Cancer, 2019. For more information contact publications@iarc.fr.: Lyon (FR), 2019. [Google Scholar]
- Zhao, J.; Yu, H. Q.; Ge, F. Q.; Zhang, M. R.; Song, Y. C.; Guo, D. D.; Li, Q. H.; Zhu, H.; Hang, P. Z. , 7,8,3’-Trihydroxyflavone prevents doxorubicin-induced cardiotoxicity and mitochondrial dysfunction via activating Akt signaling pathway in H9c2 cells. Cell Signal 2023, 112, 110924. [Google Scholar] [CrossRef]
- Elmore, S. , Apoptosis: a review of programmed cell death. Toxicol Pathol 2007, 35, 495–516. [Google Scholar] [CrossRef] [PubMed]
- Rainieri, S.; Conlledo, N.; Langerholc, T.; Madorran, E.; Sala, M.; Barranco, A. , Toxic effects of perfluorinated compounds at human cellular level and on a model vertebrate. Food and Chemical Toxicology 2017. [Google Scholar] [CrossRef] [PubMed]
- Ettinger, A.; Wittmann, T. , Fluorescence live cell imaging. Methods Cell Biol 2014, 123, 77–94. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Prochazkova, M.; Kim, Y. S.; Jiang, C.; Ma, J.; Moses, L.; Martin, K.; Pham, V.; Zhang, N.; Highfill, S. L.; Somerville, R. P.; Stroncek, D. F.; Jin, P. , Assessment and comparison of viability assays for cellular products. Cytotherapy 2024, 26, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Afjei, R.; Massoud, T. F.; Paulmurugan, R. , Comparison of cell-based assays to quantify treatment effects of anticancer drugs identifies a new application for Bodipy-L-cystine to measure apoptosis. Scientific reports 2018, 8, 16363. [Google Scholar] [CrossRef]
Figure 1.
Graphical scheme of the cell viability methods based on structural cell damage (non-invasive). a) Cell morphology changes between two samples with an inverted microscope. The left image shows viable cells, while the right image displays cells treated with 12.5 mg/L of Arsenic V. b) Measurement of leaking molecules from viable and non-viable cells. In the caption, an example with the LDH molecule is made. The example provided uses the LDH molecule, which leaks in greater amounts from non-viable cells due to membrane disruption. The supernatant (extracellular fluid) is collected and measured with a spectrophotometer, with higher values indicating non-viable cells.
Figure 1.
Graphical scheme of the cell viability methods based on structural cell damage (non-invasive). a) Cell morphology changes between two samples with an inverted microscope. The left image shows viable cells, while the right image displays cells treated with 12.5 mg/L of Arsenic V. b) Measurement of leaking molecules from viable and non-viable cells. In the caption, an example with the LDH molecule is made. The example provided uses the LDH molecule, which leaks in greater amounts from non-viable cells due to membrane disruption. The supernatant (extracellular fluid) is collected and measured with a spectrophotometer, with higher values indicating non-viable cells.
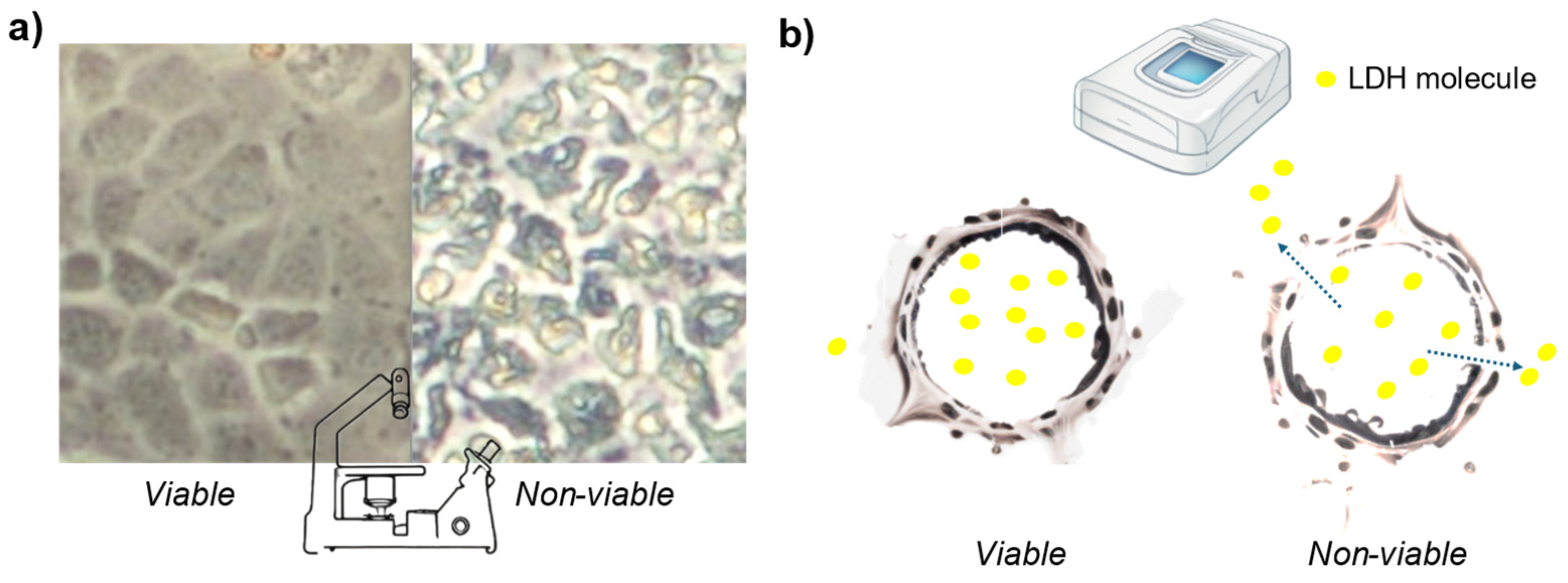
Figure 2.
Graphical scheme of the cell viability methods based on structural cell damage (invasive). a) In non-viable cells, a molecule like PI is transported within the cell. While it can also be transported in viable cells, it occurs to a much lesser extent. Once PI binds to RNA or DNA, it emits fluorescence upon excitation. b). Lipophilic dyes pass through the cell membrane and are cleaved by esterases within viable cells, occurring more frequently in viable cells. After cleavage, they emit fluorescence upon excitation. c) Antibodies pass through permeabilized cells and attach to specific molecules related to cell death pathways, such as caspase. If the target molecule is present, the antibodies bind to them, allowing visualization or quantification using fluorescence measurement instruments.
Figure 2.
Graphical scheme of the cell viability methods based on structural cell damage (invasive). a) In non-viable cells, a molecule like PI is transported within the cell. While it can also be transported in viable cells, it occurs to a much lesser extent. Once PI binds to RNA or DNA, it emits fluorescence upon excitation. b). Lipophilic dyes pass through the cell membrane and are cleaved by esterases within viable cells, occurring more frequently in viable cells. After cleavage, they emit fluorescence upon excitation. c) Antibodies pass through permeabilized cells and attach to specific molecules related to cell death pathways, such as caspase. If the target molecule is present, the antibodies bind to them, allowing visualization or quantification using fluorescence measurement instruments.
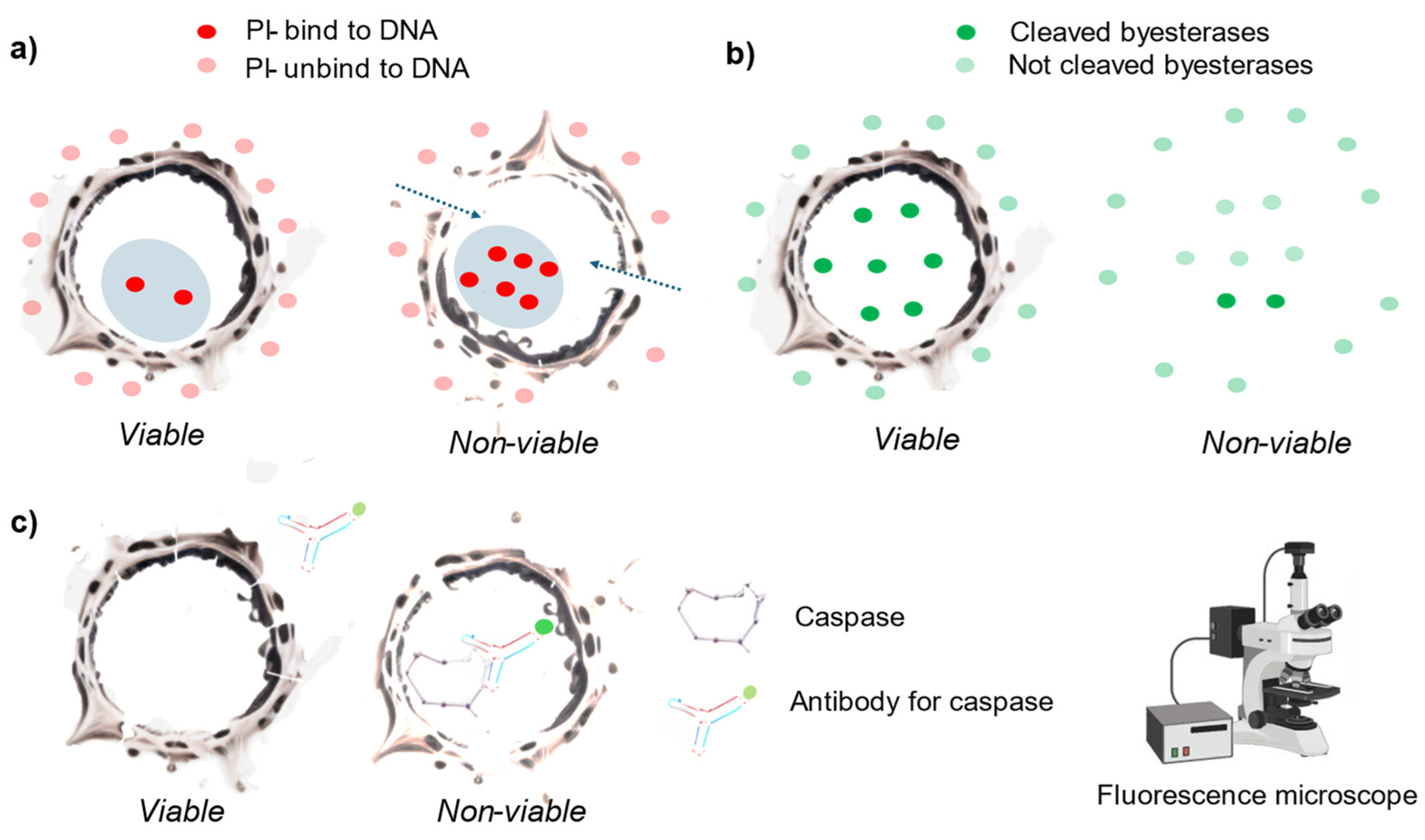
Figure 5.
Graphical scheme of the cell viability methods based on cell membrane potential. a) The intensity of dye fluorescence is proportional to the membrane potential. Previous studies utilized Fluovolt™ (FV) for this purpose. b) Cell viability is assessed by measuring both the cell membrane potential and DNA content, using the dye Vybrant™. Viable cells and cells in cell cycle arrest exhibit stable FV intensity, while apoptotic and dividing cells show higher intensity. Necrotic cells have the lowest fluorescence emission due to dye leakage. Dividing cells and cells in cycle arrest emit higher fluorescence intensity corresponding to 2n DNA.
Figure 5.
Graphical scheme of the cell viability methods based on cell membrane potential. a) The intensity of dye fluorescence is proportional to the membrane potential. Previous studies utilized Fluovolt™ (FV) for this purpose. b) Cell viability is assessed by measuring both the cell membrane potential and DNA content, using the dye Vybrant™. Viable cells and cells in cell cycle arrest exhibit stable FV intensity, while apoptotic and dividing cells show higher intensity. Necrotic cells have the lowest fluorescence emission due to dye leakage. Dividing cells and cells in cycle arrest emit higher fluorescence intensity corresponding to 2n DNA.
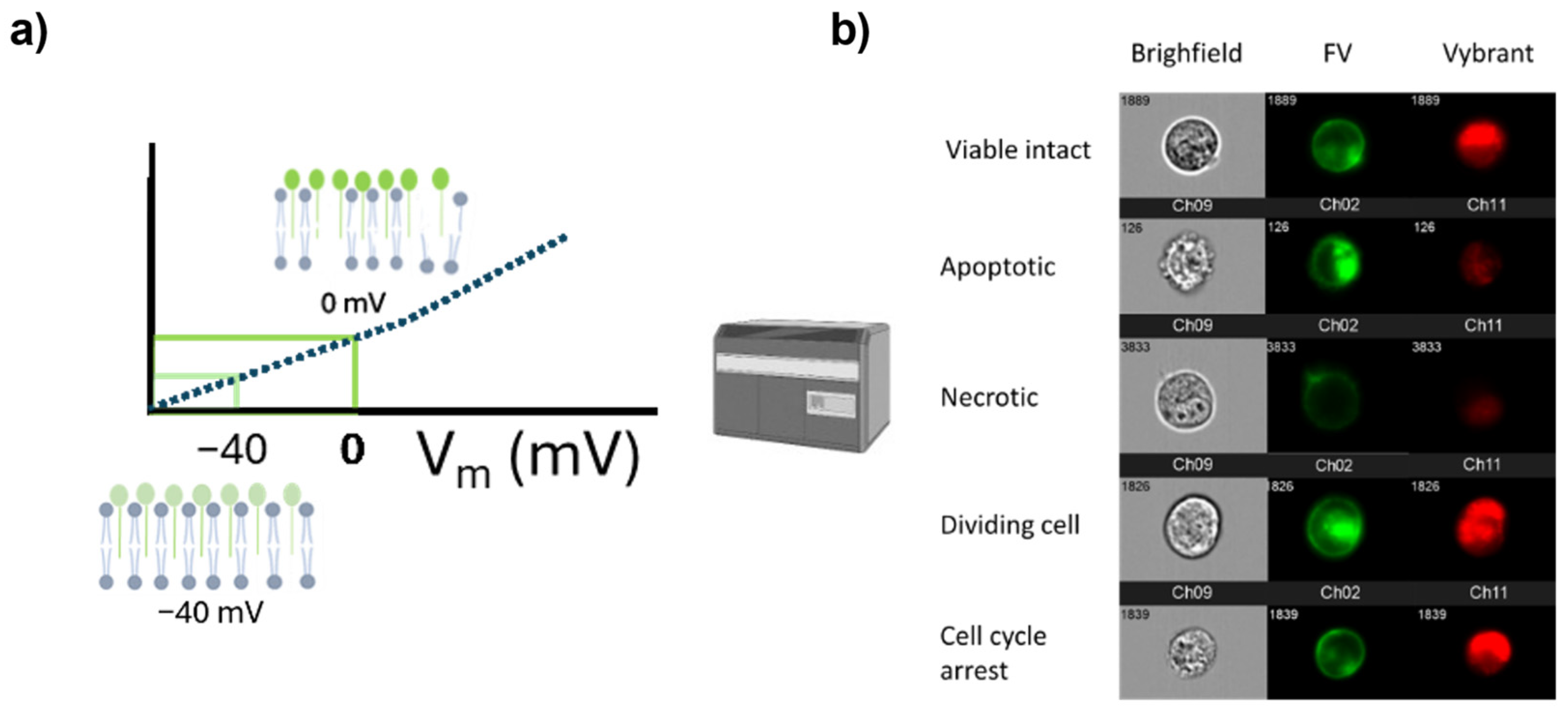

Table 1.
This is a table. Tables should be placed in the main text near to the first time they are cited.
Table 1.
This is a table. Tables should be placed in the main text near to the first time they are cited.
| Method (Group) | Principle | Endpoint (Instrument) |
Advantages | Disadvantages |
|---|---|---|---|---|
| Structural cell damage (non-invasive) | ||||
| Optical microscope (1) | Changes in the morphology of the cell. | Observe changes in the morphology of the cell (optical microscope). | Quick, cheap and it could be automatized. | Many artifacts affect the accuracy of the assay. |
| Release of intracellular compounds (2) | Measure the release of compounds into the cell culture medium due to damaged cell membrane. | Absorbance or luminescence of the compound (spectrophotometer or luminometer). | Quick, non-invasive. | High background level and potential false positive events. |
| Structural cell damage | ||||
| Trypan blue (3) | Viable cells exclude the trypan blue dye. | Observation of stained cells (optical microscope). | Cheap, it could be automatized. | High rates of false positives and false negatives. |
| Lipid-soluble dyes (4) | Hydrolysis of the dye by intracellular esterase. | Fluorescence intensity of reduced products (Fluorescence microscope, Flow cytometer). | Quick, versatile (analysis of single cell or cell population in different instruments) | High rate of false-positives. |
| Propidium Iodide (5) | The dyes are impermeant only to live cell. | Fluorescence intensity of the internalized dye (Fluorescence microscope, Flow cytometer). | Quick, many references. | High rate of false-positives. |
| Live/Dead assay (6) | It has two dyes: one impermeant only to live cells and the second is cleaved by esterases within the cell. | Fluorescence intensity of internalized dye and reduced dye (Fluorescence microscope, Flow cytometer). | Quick, versatile, better accuracy than single dyes alone. | High rate of false-positives |
| Antibodies to cell-death-associated pathways (7) | Determine the presence of molecules associated to cell death pathways. | Fluorescence intensity of the dye attached to the antibody (Fluorescence microscope, Flow cytometer). | Specific, determines the type of the program cell death. | Expensive, unable to determine the viability of the cells with a different cell death pathway. |
| Cell growth | ||||
| Cell division (8) | Difference in cell number before and after the exposure | Counting cell number in a population (microscope, flow cytometer, cell counter) | Cheap, accurate and straightforward. | Time-consuming. |
| BrdU/EdU (9) | Daughter cells contained BrdU intercalated in their DNA. | Fluorescence intensity of BrdU (Fluorescence microscope, Flow cytometer). | High sensitivity no. | Toxic, impairs cell division. |
| Sulforhodamine B (SRB) and crystal violet (10) | The amount of dye is proportional to the cells (cell proliferation). | Absorbance of the dye (spectrophotometer) | Quick, cheap. | Toxic. Measures cell mass, not cell viability. |
| Cellular metabolism | ||||
| MTT (11) | Reduction of the tetrazolium dye to formazan. | Absorbance of the formazan product (spectrophotometer). | Quick, cheap, high throughput and many references. | Many artifacts affect the accuracy of the assay. |
| Alamar blue (12) | Resazurin reduction to resorufin. | Luminescence measurement of resorufin (Fluorescence microscope and Flow cytometer). | Quick, cheap and high throughput | Naturally occurring molecules disrupt the assay. |
| JC-1, TMRE, MitoTracker (13) | Mitochondrial membrane potential (lost in non-viable cells). | Fluorescence emission of the dye proportional to the mitochondrial membrane potential (Fluorescence microscope and Flow cytometer). | Fast, cheap and high throughput. | Prone to bleaching, quenching and unquenching. Accuracy issues in certain situations. |
| Neutral Red (14) | Viable cells incorporate and bind the neutral red dye. | Absorbance of the incorporated dye (spectrophotometer). | Quick, cheap and standardize. | Not a good correlation. Lysosomal activity affects its accuracy. |
| ATP production (15) | ATP production is correlated to cell viability. | Luminescence measurement of released ATP (Luminometer). | Live imaging, non-invasive and high throughput | Many artifacts affect the accuracy of the assay. |
| Cell membrane potential | ||||
| MPCVA (16) | The cell membrane potential determines cell membrane integrity. | Fluorescence intensity of the dyes (Fluorescence microscope, Flow cytometer). | Direct determination. Live imaging | Not tested in various alternative situations. |
| DD Cell-Tox (17) | The cell membrane potential determines cell membrane integrity and DNA content the doubling cells. | Fluorescence intensity of the dyes (Fluorescence microscope, Flow cytometer). | Direct determination. Considers the cell population dynamics, and the various outcomes triggered by toxic compounds. | Not tested in various alternative situations. Not suitable for long periods of cultivation. |
Table 2.
Cell viability method list.
| Group | Method | Reference |
|---|---|---|
| 1 | Label-free imaging | [15] |
| 2 | LDH | [23] |
| 2 | CytoToxTM | [76] |
| 2 | Toxi-Light®® | [76] |
| 2 | aCella™ - TOX | [76] |
| 2 | CyQUANT™ | [77] |
| 3 | Trypan blue | [78] |
| 4 | Propidium iodide | [48] |
| 4 | Hoechst 33342 | [79,80] |
| 4 | DRAQ7 | [81] |
| 4 | Acridine orange | [82] |
| 4 | CellTox | [83] |
| 4 | SYTOX | [84] |
| 4 | YO-YO | [85] |
| 4 | TO-PRO-3 Iodide | [86] |
| 5 | Calcein AM | [87] |
| 5 | CytoCalcein™ | [88] |
| 5 | 4-methylumbelliferyl heptanoate (MUH) assay | [89] |
| 6 | Live/Dead assay | [90] |
| 7 | Antibodies | [91] |
| 7 | ApoTox-Glo | [92] |
| 7 | Annexin V | [93] |
| 8 | Cell division counting | [52] |
| 9 | BrdU assay | [53] |
| 9 | Edu assay | [53] |
| 10 | Sulforhodamine B | [94] |
| 10 | Crystal Violet | [95] |
| 11 | MTT assay | [59] |
| 11 | WST-1 assay | [96] |
| 11 | MTS assay | [8] |
| 11 | XTT assay | [97] |
| 12 | Alamar Blue | [98] |
| 13 | JC-1 | [67] |
| 13 | TMRE | [99] |
| 13 | Mitotracker | [99] |
| 14 | Neutral red | [100] |
| 15 | ATP assay | [8] |
| 16 | MPCVA | [40] |
| 17 | DD Cell-Tox | [75] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



