
Submitted:
09 September 2024
Posted:
10 September 2024
You are already at the latest version
Abstract
Human Papillomavirus (HPV) related cancers account for a large percentage of cancers. The tumor microenvironment (TME) is a complex structure and contributes significantly to tumor development, progression, and invasion. The TME is particularly pronounced in HPV-related cancers and HPV is known to influence the TME. In cancer research, there is an urgent need for new models to deepen our understanding of the pathophysiology of the disease and to facilitate the development of innovative therapies. In this context, 3D cell culture has emerged as a promising method in cancer research, that offers significant potential compared to conventional approaches. 3D modelling can produce results that are more consistent with clinical outcomes without the ethical concerns and problems of animal models. In this review, our aim is to discuss the tumor microenvironment in HPV-related cancers and demonstrate the importance of advancing 3D cell culture models for this purpose.
Keywords:
3D-cell culture
; tumor microenvironment
; HPV-induced cancers
1. Introduction
Cancer is an urgent public health issue globally, and cancer types particularly associated with Human Papillomavirus (HPV) are gaining increasing importance. HPVs are small circular viruses with double-stranded DNA [1]. About 5% of all cancers are related to HPV infections [2]. HPV is classified into low and high-risk types based on oncogenic risk. Low-risk types (Types 6 and 11) are associated with skin and anogenital warts, whereas high-risk types (Types 16, 18, 31, and 45) are predisposed agents for anogenital cancers [3,4,5]. Understanding the interaction of HPV infection on carcinogenesis and cancer progression plays a vital role in the development of prevention, early detection, and treatment strategies. HPVs are the primary cause of nearly all cervical cancers (CVC) and a significant portion of other cancers in the mucosal lining of the anogenital tract, as well as in the head-neck, and even lung region [6]. HPV presents as a critical public health problem because of its ability to infect different anatomical regions and induce malignant transformations. Understanding the effects of HPV on the genetic material of the host cell is necessary to elucidate the mechanisms of cancer development in HPV-associated cancers [7,8,9,10].
In in vitro 3D cancer models, cancer cells are co-cultured with diverse stromal cell varieties, replicating tumor characteristics in a laboratory setting. This approach reveals increased invasiveness and resistance to anti-cancer therapies compared to traditional 2D cell culture, highlighting its promise for drug screening applications [11]. Modeling cancers represents a groundbreaking step in cancer research. These studies aim to explore further the effects of cancer in vitro environments, thereby working to develop more effective treatments and preventive measures for these types of cancers. The models provide a window to understand critical processes such as cancer development, metastasis, and progression, while also revealing the role of other cells and the extracellular matrix (ECM) in these processes, offering a more comprehensive view of cancer biology. Such research forms an important foundation for developing new strategies in cancer management. This review aims to explore the microenvironment in HPV-associated cancers and understand the current status of 3D models of HPV-associated cancers in the existing literature.
2. Overview of Human Papillomavirus (HPV) and Its Implications in Cancer Development
High risk HPV is a tumorigenic DNA virus that causes epithelial proliferation on skin and mucosal surfaces and is the most common sexually transmitted infection [12,13]. The most lethal type of cancer known in women is CVC, and its primary cause are HPV types 16, 18, 45 and 56 [14,15]. According to the National Cancer Institute (2019) data for 2023, there were 13,960 new CVC diagnoses with an estimated death count of 4,310. Although HPV is generally recognized as the predominant etiological factor in CVC, it has also been reported to play a role in the pathogenesis of other malignancies such as oral squamous cell carcinoma [16,17]. Squamous cell carcinomas of the head and neck, the seventh most diagnosed cancers worldwide, also account for 5% of all cancer cases [18,19]. Similarly, about 85% of anal cancers caused by HPV are reported to be squamous cell carcinomas [20].
In a study showing the strong association between HPV infection and colorectal cancer, HPV antigen was detected in 23% of normal colon samples, 60% of adenomas, and 97% of carcinomas [21]. Detection of HPV DNA in invasive penile cancer is relatively rare, and the incidence of penile cancer is lower compared to CVC [22,23,24]. The best-known skin cancers are melanoma and non-melanoma cancers, both of which are influenced by HPV in association with UV exposure [25,26].
HPV mediated carcinogenesis is mainly related with E6 and E7 proteins of HPV. They are intranuclear proteins and even though they have different properties, they act cooperatively in promotion of carcinogenesis [27]. E7 binds to retinoblastoma protein (Rb) and retinoblastoma pocket proteins which are p107 and p130. This binding causes inhibition or proteasomal degradation of Rb which results with the increased shift to S phase of cell cycle. This shift leads to an increase in cell proliferation and therefore, increase in viral gene transcription. E7 also contributes to G1-S phase entry by binding to p21 and p27. As a response to increased expression of proliferative proteins, p53 expressions normally increase to counter excessive proliferative activation [28]. Since p53 expression disrupts viral gene replication, HPV developed a counter mechanism against p53 using E6 protein. E6 can bind to p53 to prevent its tumor-suppressive effects [29]. Also, E6 causes degradation of p53 via the ubiquitin pathway [30]. Even though E6 mainly shows its activity by p53 regulation, there are p53-independent effects of E6 as well, such as mitogenic effects, inhibition of apoptosis and interfering with the organization of the tissue [31]. E5, another oncoprotein of HPV, plays an immunosuppressive role by negatively regulating anti-viral IFN response pathways, antigen processing and antigen presentation [32]. E5 hinders the transportation of HLA-1 molecule to cell surface for immune recognition and keep it inside the Golgi apparatus. This incident happens due to E5’s interaction with HLA-1’s heavy chain via E5’s first hydrophobic domain [33]. HPV 16 E5 is shown to diminish recognition by CD8+ T cells [34]. Figure 1 summarizes the effect of HPV on the cell through the E5, E6, and E7 proteins.
3. The role of HPV on the Tumor Microenvironment
The mechanisms by which HPV leads to cancer are quite complex and involve numerous molecular interactions. The infection itself, even with the virus’s integration into the host cell’s genome, is not sufficient for carcinogenesis [35,36]. However, it results in a series of events, such as genetic rearrangements, which lead to genomic instability and increase the risk of neoplastic transformation. HPV integration leads to the production of inflammatory mediators that promote cancer progression [37].
Active molecules in seminal fluid can induce inflammation in the cervical mucosa by attracting and activating leukocytes, thereby increasing inflammation [38,39]. Interestingly, it has been observed that IL-6 levels are regulated differently in HPV-positive squamous cell carcinoma of the head and neck than in HPV-negative cases. IL-6 has been found to increase in HPV-16 and/or HPV-18 positive cells, which is associated with a poor clinical prognosis of patients [40].
Regulation of the immunosuppressive cytokines IL-10 and TGF-β has been observed in HPV-positive keratinocytes [41,42]. The simultaneous expression of these cytokines is associated with the activation and polarization of immunosuppressive regulatory T cells [43].
Tumors are not only made up of cancer cells, but have a complex structure consisting of immune cells and various molecules that remodel the environment. This inner structure of tumors other than cancer cells, is called the tumor microenvironment (TME). The TME actively contributes to tumorigenesis, tumor progression, invasion and metastasis [44].
Lymphocytes are leukocytes that are identical in appearance but differ in their functions. There are three types of lymphocytes according to their functions: T lymphocytes, B lymphocytes and natural killer (NK) cells [45]. T lymphocytes develop in the thymus, hence the name. They are able to recognize antigens that are presented to them and react appropriately depending on their subgroup [46]. There are many subsets of T cells. The most important subsets that actively contribute to TME are CD4+ T cells (also called helper T cells, Th) and CD8+ T cells (also called cytotoxic T cells). CD4+ T cells are the mediators of antitumor immunity and show their effect by directly killing cancer cells, stimulating immune cells that play a role in innate immunity and preventing angiogenesis [47]. Regulatory T cells (Treg) are a remarkable subset of CD4+ T cells. They have a highly immunosuppressive profile and their abundance in TME is associated with unfavorable outcomes [48]. CD8+ T cells are the strongest effectors of the antitumor response and are considered the basis of immunotherapy. Despite the presence of CD8+ T cells, tumor growth is not prevented. This phenomenon is also known as the “Hellstrom paradox”, which states that tumor-reactive CD8+ T cells lose their function during tumor development [49]. NK cells are cytotoxic lymphocytes that, unlike CD8+ T cells, do not require prior antigen exposure to exert their antitumor effect [50]. Despite their different antitumor mechanisms, cancer cells have developed different strategies against the activity of NK cells and their infiltration into the TME [51,52].
Macrophages are one of the most important mediators of innate and adaptive inflammatory responses. There are two main types of macrophages. M1 macrophages, which have a proinflammatory function, exhibit antitumor effects through mechanisms such as efficient regulation of Th responses, promotion of T cell proliferation, IFNγ secretion, secretion of IL12, which has proinflammatory potent antitumor effects, and reduction in secretion of factors that contribute to cancer progression, such as VEGF, MMPs and CCL18 [53]. In contrast, M2 macrophages, which have anti-inflammatory effects, show tumor-supportive effects through various mechanisms such as immunosuppression, secretion of tumor-promoting growth factors such as PDGF, TGFβ1, HGF, promotion of angiogenesis, and facilitating the transformation of fibroblasts into cancer-associated fibroblasts [54]. Macrophages that contribute to TME are referred to as tumor-associated macrophages (TAMs). TAMs are in a constant state of flux between types M1 and M2 [55]. They play an important role in angiogenesis, migration, invasion, epithelial-to-mesenchymal transition, interaction with cancer stem cells, and immunosuppression. TAMs are associated with poor overall survival in CVC and other HPV-induced cancers [56]. TAMs remain poorly understood in HPV-positive tumors. However, the increasing three-dimensional (3D) models will enable more accurate research aimed at investigating the interaction of TAMs with TME [57].
Neutrophils are the most abundant type of leukocyte in the blood and act as the first line of defense under physiological conditions. There are three recognized mechanisms for neutrophils against pathogens: Phagocytosis, degranulation, and the formation of extracellular traps for neutrophils through the release of nuclear material. Beyond these functions, however, research describes many other mechanisms, including active involvement in cancer [58]. As in infectious and inflammatory diseases, an increase in peripheral neutrophil counts is also seen in cancer patients. Several studies have shown that an increased ratio of neutrophils to lymphocytes indicates a poor prognosis [59]. Neutrophils that engage in TME undergo phenotypic changes and are referred to as tumor-associated neutrophils (TANs). Similar to macrophages, there are two main phenotypes of TANs: antitumor (TAN1) and pro-tumor (TAN2) [60]. TAN1 cells target cancer cells directly by releasing their granule content, which contains elastase and myeloperoxidase, or indirectly by forming extracellular traps and secreting cytokines such as IFN-γ and IL-12. TAN2 cells promote tumorigenesis and progression, but their mechanisms are not fully understood [61].
Eosinophils are leukocytes that are mainly involved in parasitic infections. They also have important functions in various diseases such as asthma, eosinophilic gastrointestinal diseases, and systemic hypereosinophilic diseases [62]. In addition to their functions in physiological and pathological conditions, eosinophils regulate numerous immune cells such as lymphocytes, macrophages, and neutrophils [63]. Typically, eosinophils are not regarded as major contributors of TME. However, eosinophils have several roles in many stages of cancer. They may appear as anti-tumorigenic, pro-tumorigenic, or silent bystanders depending on cancer type [64]. Eosinophils are known to infiltrate the TME and this occurrence has been termed tumor-associated tissue eosinophilia (TATE). TATE predicts a better prognosis for patients with solid tumors [65]. Because of eosinophils’ involvement in the TME and their potential role in treatment, they are worthy of cancer research. The importance of TME in viral-induced carcinogenesis, especially in HPV-induced mucosal cancers, is paramount [66,67]. In addition, HIV-positive patients with HPV-related tumors have particular problems, such as earlier onset and resistance to standard treatments, highlighting the intricate interplay between HPV infection and the immune system within the TME [68].
Understanding these interactions is of central importance for the development of effective therapeutic strategies against HPV-induced carcinogenesis.
4. Characterization of the Tumor Microenvironment in HPV-Associated Cancers
The rise in global oral cancer cases has been confined to a 2.3% yearly increase specifically in diagnoses of tongue, tonsil, and oropharyngeal cancers linked to HPV [69]. In HPV-related oropharyngeal squamous cell carcinoma (OPSCC) patients, T cell infiltration is to a greater extent, and higher IFNγ, lower IL-4, and low transforming TGF-β expressions are seen which indicates a good prognosis [70]. A noteworthy increase in the percentage of effector T cells to naïve T cells is seen in HPV-positive OPSCC compared to HPV-negative OPSCC [71]. A notable increase in CD8+ T cells in circulation and TME has been found and this elevation significantly correlates with a better prognosis [72]. Literature has controversial results regarding the role of Tregs in OPSCCs and this may be related to inconsistent techniques that are used for detection of Tregs [73]. Elevated levels of M1/M2 and M2 macrophage infiltration are seen in tumor stroma of HPV-positive OPSCC patients compared to HPV-negative patients. Interestingly, a greater infiltration of CD163+ macrophages has been found in HPV-negative patients which indicates shorter metastasis-free survival and overall survival [74]. Similarly, another study proclaims that HPV positivity in OPSCC patients significantly indicates a better overall survival up to twenty years [75]. Al-Sahaf et al. have observed a significant increase in neutrophil levels in HPV-negative OPSCC patients compared to HPV-positive patients which suggests a better prognosis for HPV-positive patients [76]. Eosinophils were found to be a positive indicator of prognosis in HPV-positive patients and disparately a negative indicator in HPV-negative patients. Same study also revealed that total peripheral eosinophil count is higher in HPV-negative patients compared to HPV-positive patients [77].
Untreated HPV infections are the cause of 95% of all CVCs [78]. There is an unanimity that CVC is an immunogenic tumor since it’s caused by persistent infection with HPV [79]. CVC patients either lack or have severely diminished HPV-16 specific CD4+ T-cell response [80]. Inhibition of CD4+ and CD8+ T cell responses to HPV are associated with the CVC development [81]. Also, CD4+/CD8+ ratio is found to be significantly lower in CVC tumor than in peripheral blood [82]. Decreased CD4+ T cell ratio is strongly correlated with rapid tumor growth and lymph node metastasis in CVC [83]. Treg infiltration correlates with CVC progression and decreased Treg levels show improved outcomes [84]. NK cell ratio is greater in stage II CVC in comparison to stage I CVC patients. Also, patients with NK cell population greater than 15% have significantly higher clinical stage and invasion depth compared to those with lesser than 15% [85]. Elevated levels of CD56high NK cells in tumor tissue compared to peripheral blood in CVC patients have been demonstrated. Conversely, CD56low NK cells are present in peripheral blood to a greater extent [86]. Macrophages in the TME of CVC are mainly polarized towards the M2 phenotype and this polarization also correlates with poor response to chemoradiation [87]. Increased TAM density is markedly associated with poor survival in CVC [88]. Also, there is a modulation of TAMs by CVC cells via lactate secretion [89]. In terms of distinguishing precancerous lesions from CVC, increased neutrophil-to-lymphocyte ratio is predictive for CVC [90]. A recent study has established that high peripheral neutrophil to lymphocyte ratio is correlated with poor prognosis in early CVC [91]. Increased eosinophil levels and eosinophil-to-lymphocyte ratio indicate a favorable overall survival in CVC patients [92]. Microenvironment of CVC can be summarized as shown in Figure 2.
Penile cancer is a rare condition in industrialized nations; however, they have high incidences in developing regions [93]. Limited availability and access to medical healthcare in these regions may benefit greatly from advances in treatment. Detection of HPV in penile cancers is highly inconsistent which may explain the heterogeneity of literature considering the presence of HPV in penile cancers [94]. There is an increase of cytotoxic T cells in HPV-positive patients compared to HPV-negative patients. This increase is associated with poor overall survival [95]. Increased CD8+ T cell infiltration rate is related with decreased lymph node metastasis in penile cancer patients [96]. Literature investigating Tregs in penile cancers is scant [97]. NK cell activity has shown a noteworthy decrease in penile carcinoma [98]. CD68+ macrophages are found to be in high densities, and they are associated with a more favorable overall survival and decreased risk of regional recurrence [95]. Stromal CD68+ macrophages are inversely associated with lymph node metastasis [99]. Increased intratumoral CD163+ M2 macrophages are significantly associated with higher lymph node metastasis [96]. Higher neutrophil-to-lymphocyte ratio is associated with worse prognosis and lymph node metastasis in penile cancer patients [100]. Eosinophil infiltration has been demonstrated in penile cancers and seems to improve the 5-year survival, however researchers point out the small sample group and suggest broader research in this subject [101].
Colorectal cancer (CRC) is the second leading cause of cancer-related deaths in the United States. Although HPV is not considered a major risk factor for CRC, the presence of HPV infection is associated with a higher risk of CRC [102]. In addition, the overall prevalence of HPV in CRC patients is 31.9%, further supporting the association between HPV and CRC [103]. In HPV-positive CRC patients, there is an increase in T cells compared to HPV-negative patients [104]. An increase in Tregs was also observed [105]. M1 macrophage infiltration is increased in HPV-positive CRC. The same study also showed that the expression of MHC-II by M1 macrophages is increased and that these findings together directly increase the antitumor response [104]. The literature investigating the HPV-associated CRC microenvironment is insufficient and new studies may shed light on our understanding and thus improve treatment options.
Anal cancer is one of the rare cancers however its incidence increases every year by 2.7%. Anal squamous cell cancer (ASCC) is the most common form of anal cancer [106]. Approximately 90% of anal cancers are associated with HPV infection [107]. High amounts of tumor-infiltrating lymphocytes are related to better prognosis in ASCC patients [108]. As a result, IFNγ secretion and increased PD-L1 expression are seen within TME [109]. Studies regarding Tregs in ASCC are limited and have controversial results. The roles of TAMs in ASCC remain mostly unresolved [110]. Neutrophilia strongly predicts overall survival, progression, and failure-free survival in APSCC patients [111]. Increased neutrophil-to-lymphocyte ratio is associated with poor overall survival and cancer-specific survival [112]. In HPV-positive ASCC patients, low eosinophil levels correlate with better disease-free survival [77].
Normally, the supporting structure, the so-called stroma, has been regarded merely as a scaffold on which the epithelial cells “sit”. However, it is known that the communication between stromal cells and HPV-infected epithelial cells plays a crucial role in carcinogenesis. Stromal fibroblasts, which are the most important cellular component of connective tissue, provide signals that significantly influence the development and progression of cancer. Cancer-associated fibroblasts (CAFs) are fibroblasts that are activated by paracrine mediators produced by cancer cells. They can promote HPV-mediated carcinogenesis through various mechanisms via stromal-epithelial interactions [113]. The ability of CAFs to promote immunosuppression in HPV-associated cancer, as well as their ability to enhance cell growth and metastasis, has been reported several times [114,115,116].
It has been indicated that bidirectional communication between epithelial cells and the TME affects a range of processes from tumor initiation to neoplastic progression, and even to metastasis and drug resistance. Researchers believe that in response to this communication, the TME alters various stromal-epithelial signaling activities crucial for HPV-positive epithelial cell growth, disease initiation, and progression. Therefore, there is a clear need for 3D in vitro model systems to better understand the interactions between HPV-infected tissue and the surrounding TME. These systems are crucial for comprehending the fundamental mechanisms of cancer and developing new treatment strategies.
5. Utilization of 3D In Vitro Models in HPV-Related Cancer Research
For about a century, cancer research has progressed by focusing on the cellular structures of tumors [117]. Cell lines derived from various cancer types have been developed. These cell lines can be easily multiplied in culture and reflect the characteristics of specific cancer types. They have become a significant tool in cancer research [118]. However, commercially available tumor-derived cell lines often have genetic characteristics that differ from the original tumor cells [117]. Furthermore, cell lines undergoing extended cultures may exhibit heightened susceptibility to genetic alterations, varied morphological characteristics, and increased vulnerability to contamination [119,120]. Although extensive evidence supports the superiority of 3D cell models, engineered with materials optimized for cellular growth in the laboratory over traditional two-dimensional (2D) cultures [121], preclinical research predominantly utilizes 2D cultures due to their simpler cultivation process [122].
Culturing commercial cell lines which are modified for easier replication and using materials that don’t harm these cells in cell culture is practical in terms of in vitro cancer studies. However, these factors often remain inadequate and unreliable to mimic an organism. The significant gap between 2D cell cultures studied in the laboratory and clinical studies is attempted to be filled with animal model experiments; however, both the inconsistency between human and animal responses and substantial time and cost expenditures are incurred as a result [123]. In 2D cell cultures, the cells grow on polystyrene plates and form individual layers. Therefore, the culture lacks the complex interactions between cells and the surrounding ECM as well as the interactions between neighboring cells that are present in 3D cell culture models [124]. Anticancer drugs used in 2D cell cultures usually reach the cells without any physical barriers. However, the drugs have to overcome many factors in the organism that prevent their spread throughout the tumor [125]. 3D organization of the tumor mass and associated stroma fundamentally alters the tumor spread profile via cell-cell contacts and cell-matrix interactions [126].
In the literature, mice have been considered a good model for mimicking human cancers, particularly for animal model approaches. However, the significant differences among tumors and ethical concerns have emphasized the need for a new model [127,128]. Therefore, in the literature, studies are attempting to mimic the ECM by using different biomaterials on the lung [129], breast [130], prostate [131], and kidney [132] to investigate such cancer development.
While different forms of biomaterials affect cell culture efficiency, 3D culture methods contribute to the advancement of many applications, particularly in the fields of cancer and stem cell research, as well as drug and toxicity screening [133]. Cells in 2D are exposed to a homogeneous environment with sufficient oxygen and nutrients, whereas cells in solid tumors are exposed to gradients of critical chemical and biological signals, which can have both stimulating and inhibitory effects on tumor progression [134]. When comparing 3D and 2D gene expressions, the crucial difference emerges in culture systems rather than cell lines [135]. Schematic illustration of the comparison between 2D and 3D culture models Figure 3.
In this field of research, there is considerable amount of interest in identifying 3D models that best reflect the specific characteristics of the TME. With the increase in studies in this area over the past decade, interest in 3D modeling has grown exponentially. Approximately 18,000 publications that have been found in the search for “3D cancer models” in 2024 can be seen as a precursor to the emergence of new and valuable techniques for studying tumor and TME interactions.
6. 3D Modelling of HPV-Induced Cancers
According to the National Cancer Institute (NCI), major HPV-related cancers are anal cancer, cervical cancer, oropharyngeal cancer, penile cancer, vaginal cancer, and vulvar cancer. Also, other cancers such as colorectal cancer, skin cancer, and lung cancer may be related to HPV infection. 3D models of HPV-related cancer models are summarized in Table 1.
Cervical Cancers
When examining 3D printing articles mainly related to CVC, studies are aimed at analyzing cell behavior and simulations designed to visualize CVC. The second type of 3D CVC cancer model provides visual interaction between doctors and patients, facilitating surgical simulation for medical personnel [136,137]. However, in this review, we will focus on the studies of the first type. Zhao et al.‘s (2014) study aimed to create cervical tumor models using HeLa cells and gelatin/alginate/fibrinogen hydrogels with 3D printing technique and reported the comparison of these models with traditional 2D cultures. The results indicated that printed 3D models offered successfully simulated tumor properties such as higher proliferation rate, cellular spheroid formation, elevated MMP protein expression, and chemoresistance, suggesting a broad application potential of the novel 3D cell printing technology in tumor biology and tumor heterogeneity studies [138]. Muniandy et al. (2021) successfully established 3D spheroids of CVC cell lines using the liquid overlay method, revealing distinct behaviors in different cell types: HeLa and CaSki formed compact spheroids while SiHa formed flat ones. HeLa and CaSki spheroids showed aggressive invasion, while SiHa spheroids displayed incremental growth and invasion [139]. Zuk and colleagues created 3D artificial tissues in de-epidermized dermal scaffolds using normal and cancerous cervical cell lines in their study in 2017. While the normal tissue model exhibited distinct epithelial layers and specific cytokeratin expressions, the cancer model demonstrated dysplastic changes and different cytokeratin expressions [140]. Additionally, De Gregorio et al. developed a novel organotypic model for cervical tumors and identified key molecules involved in the epithelial-mesenchymal transition process. The systematic characterization of stromal factors highlighted the significance of potential therapeutic targets and emphasized the importance of treatments targeting not only cancer cells but also the stroma [141]. Thippabhotla et al. (2019) compared extracellular vehicles derived from 2D and 3D cell cultures and found that 3D culture-derived extracellular vesicles (EVs) more closely resembled in vivo circulating EVs from CVC patients, suggesting a specific sorting process [64]. In the study conducted by Lin and colleagues (2024), it is demonstrated that C3 PLP2+ Tumor Epithelial Progenitor Cells play a significant role in cervical carcinoma in the generated 3D model, and the PLP2+ Tumor EPC score is shown to be an independent prognostic factor [142]. Another study was conducted by Jackson et al. (2020), their method involved the successful creation of a 3D culture by growing primary human epithelial cells on a dermal equivalent using collagen fibers, derived from both HPV-negative and HPV-positive oral and cervical tissues [143]. The studies conducted by Mao et al., on human CVC, including research on cell proliferation, epithelial-mesenchymal transition, and drug resistance, demonstrated that 3D culture models yield much more effective results compared to 2D culture systems [144].
Oropharyngeal Cancers
Miserocchi et al. (2021), who created a 3D model with a collagen-based scaffold using HPV-positive and HPV-negative oropharyngeal squamous cell carcinoma cells (OSCC), emphasized the in vitro model’s ability to represent the different pathophysiological nature and drug sensitivity of the two tumor types [145]. Colley et al. (2011) investigated where three different in vitro models demonstrating the transition from dysplasia to in situ carcinoma histopathologically were utilized for the tissue engineering of oral mucosa. These models were further examined through in vivo comparisons. In this context, they demonstrated that when natural dermis was used, it was possible to culture dysplasia and tumor cell invasion in vitro, mimicking the morphological appearance and histological features observed in vivo [146]. In another study, Almela et al. (2018) printed a scaffold with suitable pore sizes in 3D using an injectable β-tricalcium phosphate paste. Human primary oral squamous cell carcinoma cells were seeded onto this scaffold, successfully establishing a short-term in vitro model, although not long-lasting [147]. In another study, Noi and colleagues (2018) used a commercially purchased 3D culture system. They suggest that signaling pathways are significantly altered only through this method, contributing to invadopodium formation and the development of oral cancer in experiments conducted within the 3D culture system [148]. In another study, Vincent-Chong and Seshadri (2020) emphasized the importance of strategies aiming to successfully identify the response of the subtype of oral cancer and sensitize tumors to radiation, stating that the 3D organoid platform is a valuable tool for screening radiosensitizers and investigating the efficacy of novel immunotherapeutic strategies [149]. Lei et al. developed a perfusion 3D cell culture microfluidic chip for real-time and non-invasive monitoring of cell proliferation and chemosensitivity. They demonstrated that oral cancer cells proliferation and chemosensitivity in 3D cell culture format can be monitored by impedance measurement [150]. In another investigation anticipating the prospective importance of personalized medicine, Igawa et al. (2023) have established a 3D oral cancer model incorporating oral cancer cells and normal oral fibroblasts [151].
In their study, Tanaka et al. (2018) established three well-characterized primary head and neck squamous cell carcinoma HPV-positive organoid lines, from which 2D cell lines can also be easily obtained. They demonstrated that by conducting drug resistance studies on cells derived from organoids, they could obtain the best possible response similar to the original tumor [152]. In another study, Sawant et al. (2016) created a 3D culture by embedding primary keratinocytes in a collagen matrix with fibroblasts and, for comparison, seeding them in a flat collagen matrix without growth factors, for coculture with fibroblasts. While the 3D coculture containing fibroblasts yielded results closer to in vivo conditions, they noted some limitations such as limited sample quantity due to being obtained from humans, contamination based on poor hygiene, and inter-donor variability [153].

Table 1.
3D models to mimic tumor and TME in HPV related cancers.
| HPV-related cancer types | Culturing Model | Used Cells | ECM Matrix | References |
|---|---|---|---|---|
| CVC | Hydrogel | Hela cells | Gelatin/alginate/fibrinogen | [138] |
| CVC | Spheroid System | HeLa (HPV18), CaSki (HPV16), SiHa (HPV16), C33A (non-HPV), HT3 (non-HPV) |
Bovine collagen I matrix | [139] |
| CVC | Scaffold | human keratinocyte cell line (NTERT), cervical cancer cell line (C33A) |
Human skin (Euro Skin Bank, Netherlands) | [140] |
| CVC | Scaffold | SiHa (ATCC® HTB-35), Cervical CAF isolated from squamous intraepithelial lesion, and invasive cervical carcinoma biopsies |
Fibronectin solution human fibroblasts | [141] |
| CVC | Hydrogel | Hela cells (ATCC® CCL-2™) | Peptide-based | [64] |
| CVC | Hydrogel | SiHa and Hela cells | Matrigel matrix (BD Biosciences, USA) | [142] |
| CVC | Scaffold | primary human cervical carcinoma | Collagen, glycosaminoglycan, and elastin | [144] |
| CVC and OSCC | Organotypic Raft | primary epithelial cells (human tonsillar keratinocytes or human cervical keratinocytes), J2 fibroblasts, Human fibroblasts hTERT |
Collagen-fibroblast mixture | [143] |
| OSCC | Scaffold | UPCI:SCC090 (CRL-3239), UM-SCC-6 Squamous Carcinoma Cell Line |
Type I collagen | [145] |
| OSCC | Scaffold | Normal human oral keratinocytes and fibroblasts were isolated from oral mucosa biopsies, Primary alveolar human osteoblasts were isolated from aspirated waste bone chips |
Collagen mixture and fibrin adhesive sealant | [147] |
| OSCC | Spheroid System | D20, FaDu, DOK, Cal27 (CRL-2095) and, Normal oral keratinocytes and fibroblasts were isolated from biopsies |
Human cadaver skin(Euroskin, Beverwijk, Holland) | [146] |
| HNSCC | Organoid | Individual patient’s tumors | - | [152] |
| Human tongue squamous cell carcinoma | Organotypic Culture | Individual patient’s tumors (keratinocytes and fibroblasts) | Collagen Matrix | [153] |
| Human tongue cancer tissue | Silica fibre sheet | HSC-3 and HSC-4 | - | [148] |
| Human oral cancer | Microfluidic chip | OEC-M1 | - | [150] |
| Human oral cancer | Hydrogel | Human primary normal oral fibroblasts human oral squamous carcinoma cell lines |
Collagen Matrix | [151] |
In addition to the major cancers affected by HPV, the 3D models of the cancer types that are minorly affected by HPV are as follows:
Colorectal Cancer
The organoid model of the mouse intestine developed by Sato et al. in 2009 has provided important progress in CRC organoid models. Sato et al. also improved their model in 2011 to adapt it for human small intestine and colon [154]. McGuckin et al. have achieved to produce the first long-term 3D CRC model. Abbas et al. (2023) found significant differences in cell proliferation, death phase profiles, tumor-related gene expression, and drug sensitivity in 3D colorectal cancer cultures compared to 2D cultures [155]. Another study on drug resistance was conducted by Koch et al. in 2021, which found that 3D colorectal cancer cells were more resistant to radiation and certain chemotherapy drugs compared to 2D cell cultures [156]. Magdeldin et al. (2014) developed and established a physiologically relevant model for colorectal cancer using HT29 and HCT116 cell lines, focusing on 3D in vitro model, gene expression profiles, oxygen and nutrient gradients, and pharmacokinetics [157].
Skin Cancer
Skin cancer can be categorized into two main types, nonmelanoma and melanoma, based on the histological characteristics of the malignant cells [158]. Berning et al. (2015) constructed a complex dermal equivalent using normal human dermal fibroblasts, enabling long-term epidermal regeneration. Furthermore, by incorporating cells from various stages of malignancy using inserts, they developed a human skin cancer model reflecting tumor-specific growth and invasion profiles [159]. In another study highlighting significant differences in treatment outcomes between 2D and 3D cultures, Vörsmann et al. (2013) developed an organotypic human skin-melanoma model [160]. In another study, they proposed a collagen-based membrane for early invasion investigations of human melanoma and various drug trials [161].
7. Limitations and Future Directions in 3D Model-Based Cancer Research
3D culture systems offer a significant opportunity for investigating cancer immunology; however, they also come with some limitations. The increased complexity of these systems can create challenges in replicating and interpreting results across experiments and within them. Additionally, compared to 2D culture systems, 3D culture systems can be more costly and less accessible. Researchers may struggle to capture images of 3D cultures by some microscopy techniques due to their depth and limited transparency. When it comes to studying immunological environment of cancer using 3D culture systems, it can be challenging to culture primary immune cells long-term in both 2D and 3D environments. Additionally, existing 3D models of the tumor microenvironment might not fully capture the complexity of human tumors, which often involve various immune and stromal cell populations. Accurately replicating this complexity and ensuring that immunotherapeutic agents can effectively penetrate the ECM are critical considerations [162]. The movement of immune cells is vital to their function and activity in the tumor microenvironment, so it must be a primary consideration in 3D cell culture assays [163]. Moreover, along with the multi-step interactions between various cell types, the extraordinary complexity of the immune system may limit the applicability of simple heterotypic or multicellular culture methods. Before starting 3D culture, a tumor model must be sufficiently characterized compared to the original tumor tissue to ensure accurate interpretation of results. Each experiment should include 3D models representing different regions of the tumor to mimic tumor heterogeneity. The availability of sufficient tissue material and ethical issues related to tumors taken from patients are among the difficulties of 3D models. A significant disadvantage is the inability to replicate ex vivo results from individual tumors in an in vivo environment. Therefore, large patient samples, as in clinical trials, are required to obtain reliable results.
The in vitro utilization of 3D models highlights the ability to provide a realistic simulation of the tumor microenvironment by replicating cellular heterogeneity, gene expression patterns, cell differentiation, hypoxia induction, activation of cell signaling pathways, and cell-cell/cell-ECM interactions. These models enable the in vitro assessment of various aspects of tumor biology such as chemoresistance, migration, and invasion, making them a valuable tool in cancer research [164]. Interestingly, tumor cells in some cancer patients exhibit intrinsic resistance to a wide spectrum of chemotherapeutic drugs without prior exposure to these cytotoxic agents [165]. This intrinsic drug resistance has been assigned to the increased expression of multidrug resistance (MDR) proteins by cancer cells [166,167]. Hypoxia [168], low nutrient availability, and low pH which are characteristics of tumor tissue [169] have been suggested to increase the expression of MDR proteins through specific cellular signaling pathways. While an MDR-permissive environment can be partially recreated in 2D cultures, the absence of 3D architectural context hinders the recapitulation and conservation of MDR behaviors [170].
Upon reviewing the literature, it is notable that particularly HPV-infected 3D cell cultures have not been extensively developed. Furthermore, studies on the microenvironment in many HPV-induced cancer types remain scarce. Looking ahead, we envision that the penetration of various drugs through 3D models, drug absorption according to ECM and TME, and assessment of potential drug candidates can be more accurately evaluated in HPV-induced tumors.
8. Conclusions
In recent years, cancer researchers have been emphasizing the importance of the 3D TME that influences tumor cell behavior. However, much of cancer research, especially in the field of drug screening, remains confined to the 2D cell culture paradigm. This review, as evidenced by various publications, has robustly demonstrated the superiority of 3D growth in modeling tumor cell behavior and drug response. Additionally, many studies indicate the need for more accurate modeling of tumor stroma, as the ECM serves as a regulator of tumor cell behavior and is one of the new cancer treatment targets. 3D in vitro culture systems successfully mimicking cancer cell behavior in vivo have become important tools for drug screening, angiogenesis, immunotherapy, and hypoxic conditions. 3D environments developed with high precision and accuracy, utilizing hydrogel materials containing various growth stimulants or inhibitors, may play a significant role in future cancer treatments.
On the other hand, despite gaining deeper insights into HPV-mediated cancer formation, many questions regarding virus-host interactions remain unanswered. Particularly, uncertainties persist regarding how HPV affects antitumoral immunity in the context of radiotherapy and immune checkpoint blockade. In this review, we have examined significant progress made in creating in vitro models that better reflect HPV-associated cancer types. We have focused on 3D models expected to more closely resemble the in vitro environment than the standard 2D settings used in many new models. We believe that in future cancers or non-cancerous diseases of all kinds, the penetration of various drugs through 3D models, the absorption of drugs according to ECM and TME, and potential drug candidates can be more accurately evaluated.
Author Contributions
Conceptualization, S.T.A., B.K., G.H.A., M.O., O.Ç. and B.K.Y.; methodology, S.T.A., B.K. and B.K.Y.; investigation, S.T.A. and B.K.; data curation, S.T.A. and B.K.; writing—original draft preparation, S.T.A. and B.K.; writing—review and editing, S.T.A., B.K., B.K.Y. and O.Ç. G.H.A., M.O.; supervision, B.K.Y., O.Ç.; project administration, B.K.Y. All authors have read and agreed to the published version of the manuscript.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable because this review article examines the current literature therefore no new data to literature is available.
Acknowledgments
-.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Riou, G.; Bourhis, J.; Favre, M.; Orth, G.; Jeannel, D.; Le Doussal, V. Association between Poor Prognosis in Early-Stage Invasive Cervical Carcinomas and Non-Detection of HPV DNA. The Lancet 1990, 335, 1171–1174. [Google Scholar] [CrossRef] [PubMed]
- Canfell, K.; Kim, J.J.; Brisson, M.; Keane, A.; Simms, K.T.; Caruana, M.; Burger, E.A.; Martin, D.; Nguyen, D.T.N.; Bénard, É.; et al. Mortality Impact of Achieving WHO Cervical Cancer Elimination Targets: A Comparative Modelling Analysis in 78 Low-Income and Lower-Middle-Income Countries. The Lancet 2020, 395, 591–603. [Google Scholar] [CrossRef] [PubMed]
- Pina Brianti 1, E.D.F. 1 Review of HPV-Related Diseases and Cancers - PubMed Available online:. Available online: https://pubmed.ncbi.nlm.nih.gov/28368072/ (accessed on 19 April 2024).
- Muñoz, N.; Castellsagué, X.; de González, A.B.; Gissmann, L. Chapter 1: HPV in the Etiology of Human Cancer. Vaccine 2006, 24 (Suppl 3). [Google Scholar] [CrossRef]
- Stanley, M.A.; Pett, M.R.; Coleman, N. HPV: From Infection to Cancer. Biochem Soc Trans 2007, 35, 1456–1460. [Google Scholar] [CrossRef]
- de Sanjose, S.; Serrano, B.; Tous, S.; Alejo, M.; Loveras, B.L.; Quiros, B.; Clavero, O.; Vidal, A.; Ferrandiz-Pulido, C.; Pavon, M.A.; et al. Burden of Human Papillomavirus (HPV)-Related Cancers Attributable to HPVs 6/11/16/18/31/33/45/52 and 58. JNCI Cancer Spectr 2018, 2. [Google Scholar] [CrossRef]
- Potale, Y.; Kasat, Y.K.; Kumar, A.; Ahmad, F. Unravelling the Impact of Human Papillomavirus (HPV): A Comprehensive Exploration of Its Role in Cancer Progression and Global Health Challenges. [CrossRef]
- Fitzmaurice, C.; Allen, C.; Barber, R.M.; Barregard, L.; Bhutta, Z.A.; Brenner, H.; Dicker, D.J.; Chimed-Orchir, O.; Dandona, R.; Dandona, L.; et al. Global, Regional, and National Cancer Incidence, Mortality, Years of Life Lost, Years Lived With Disability, and Disability-Adjusted Life-Years for 32 Cancer Groups, 1990 to 2015: A Systematic Analysis for the Global Burden of Disease Study. JAMA Oncol 2017, 3, 524–548. [Google Scholar] [CrossRef]
- Human Papillomavirus (HPV) Available online:. Available online: https://www.who.int/teams/health-product-policy-and-standards/standards-and-specifications/vaccine-standardization/human-papillomavirus (accessed on 29 February 2024).
- Krasniqi, E.; Barba, M.; Venuti, A.; Pizzuti, L.; Cappuzzo, F.; Landi, L.; Carpano, S.; Marchetti, P.; Villa, A.; Vizza, E.; et al. Circulating HPV DNA in the Management of Oropharyngeal and Cervical Cancers: Current Knowledge and Future Perspectives. J Clin Med 2021, 10, 1525. [Google Scholar] [CrossRef]
- Costa, E.C.; Moreira, A.F.; de Melo-Diogo, D.; Gaspar, V.M.; Carvalho, M.P.; Correia, I.J. 3D Tumor Spheroids: An Overview on the Tools and Techniques Used for Their Analysis. Biotechnol Adv 2016, 34, 1427–1441. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, N. Human Papillomavirus and Cancer: The Epidemiological Evidence. J Clin Virol 2000, 19, 1–5. [Google Scholar] [CrossRef]
- Muñoz, N.; Bosch, F.X.; de Sanjosé, S.; Herrero, R.; Castellsagué, X.; Shah, K. V.; Snijders, P.J.F.; Meijer, C.J.L.M. Epidemiologic Classification of Human Papillomavirus Types Associated with Cervical Cancer. N Engl J Med 2003, 348, 518–527. [Google Scholar] [CrossRef]
- Zibako, P.; Tsikai, N.; Manyame, S.; Ginindza, T.G. Cervical Cancer Management in Zimbabwe (2019–2020). PLoS One 2022, 17, e0274884. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization Global Strategy to Accelerate the Elimination of Cervical Cancer as a Public Health Problem. World Health Organization 2020, 1–56.
- Hardefeldt, H.A.; Cox, M.R.; Eslick, G.D. Association between Human Papillomavirus (HPV) and Oesophageal Squamous Cell Carcinoma: A Meta-Analysis. Epidemiol Infect 2014, 142, 1119–1137. [Google Scholar] [CrossRef] [PubMed]
- Melo, B.A. de C.; Vilar, L.G.; Oliveira, N.R. de; Lima, P.O. de; Pinheiro, M. de B.; Domingueti, C.P.; Pereira, M.C. Human Papillomavirus Infection and Oral Squamous Cell Carcinoma - a Systematic Review. Braz J Otorhinolaryngol 2021, 87, 346–352. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J Clin 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Tornesello, M.L.; Perri, F.; Buonaguro, L.; Ionna, F.; Buonaguro, F.M.; Caponigro, F. HPV-Related Oropharyngeal Cancers: From Pathogenesis to New Therapeutic Approaches. Cancer Lett 2014, 351, 198–205. [Google Scholar] [CrossRef]
- Daling, J.R.; Madeleine, M.M.; Johnson, L.G.; Schwartz, S.M.; Shera, K.A.; Wurscher, M.A.; Carter, J.J.; Porter, P.L.; Galloway, D.A.; McDougall, J.K. Human Papillomavirus, Smoking, and Sexual Practices in the Etiology of Anal Cancer. Cancer 2004, 101, 270–280. [Google Scholar] [CrossRef]
- Kirgan, D.; Manalo, P.; Hall, M.; Mcgregor, B. Association of Human Papillomavirus and Colon Neoplasms. Archives of Surgery 1990, 125, 862–865. [Google Scholar] [CrossRef]
- Burgers, J.K.; Badalament, R.A.; Drago, J.R. PENILE CANCER: Clinical Presentation, Diagnosis, and Staging. Urologic Clinics of North America 1992, 19, 247–256. [Google Scholar] [CrossRef]
- Gregoire, L.; Cubilla, A.L.; Reuter, V.E.; Haas, G.P.; Lancaster, W.D. Preferential Association of Human Papillomavirus With High-Grade Histologic Variants of Penile-Invasive Squamous Cell Carcinoma. JNCI: Journal of the National Cancer Institute 1995, 87, 1705–1709. [Google Scholar] [CrossRef]
- Rubin, M.A.; Kleter, B.; Zhou, M.; Ayala, G.; Cubilla, A.L.; Quint, W.G.V.; Pirog, E.C. Detection and Typing of Human Papillomavirus DNA in Penile Carcinoma: Evidence for Multiple Independent Pathways of Penile Carcinogenesis. Am J Pathol 2001, 159, 1211–1218. [Google Scholar] [CrossRef] [PubMed]
- Harwood, C.A.; McGregor, J.M.; Proby, C.M.; Breuer, J. Human Papillomavirus and the Development of Non-Melanoma Skin Cancer. J Clin Pathol 1999, 52, 249–253. [Google Scholar] [CrossRef] [PubMed]
- Apalla, Z.; Nashan, D.; Weller, R.B.; Castellsagué, X. Skin Cancer: Epidemiology, Disease Burden, Pathophysiology, Diagnosis, and Therapeutic Approaches. Dermatol Ther (Heidelb) 2017, 7, 5–19. [Google Scholar] [CrossRef]
- Tomaić, V. Functional Roles of E6 and E7 Oncoproteins in HPV-Induced Malignancies at Diverse Anatomical Sites. Cancers (Basel) 2016, 8. [Google Scholar] [CrossRef] [PubMed]
- Cosper, P.F.; Bradley, S.; Luo, L.; Kimple, R.J. Biology of HPV Mediated Carcinogenesis and Tumor Progression. Semin Radiat Oncol 2021, 31, 265. [Google Scholar] [CrossRef]
- Lechner, M.S.; Laimins, L.A. Inhibition of P53 DNA Binding by Human Papillomavirus E6 Proteins. J Virol 1994, 68, 4262–4273. [Google Scholar] [CrossRef]
- Thomas, M.; David, P.; Banks, L. The Role of the E6-P53 Interaction in the Molecular Pathogenesis of HPV. Oncogene 1999, 18, 7690–7700. [Google Scholar] [CrossRef]
- Mantovani, F.; Banks, L. The Human Papillomavirus E6 Protein and Its Contribution to Malignant Progression. Oncogene 2001, 20, 7874–7887. [Google Scholar] [CrossRef]
- Miyauchi, S.; Kim, S.S.; Jones, R.N.; Zhang, L.; Guram, K.; Sharma, S.; Schoenberger, S.P.; Cohen, E.E.W.; Califano, J.A.; Sharabi, A.B. Human Papillomavirus E5 Suppresses Immunity via Inhibition of the Immunoproteasome and STING Pathway. Cell Rep 2023, 42. [Google Scholar] [CrossRef]
- Ashrafi, G.H.; Haghshenas, M.; Marchetti, B.; Campo, M.S. E5 Protein of Human Papillomavirus 16 Downregulates HLA Class I and Interacts with the Heavy Chain via Its First Hydrophobic Domain. Int J Cancer 2006, 119, 2105–2112. [Google Scholar] [CrossRef]
- Campo, M.S.; Graham, S. V.; Cortese, M.S.; Ashrafi, G.H.; Araibi, E.H.; Dornan, E.S.; Miners, K.; Nunes, C.; Man, S. HPV-16 E5 down-Regulates Expression of Surface HLA Class I and Reduces Recognition by CD8 T Cells. Virology 2010, 407, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Sharkey, D.J.; Tremellen, K.P.; Jasper, M.J.; Gemzell-Danielsson, K.; Robertson, S.A. Seminal Fluid Induces Leukocyte Recruitment and Cytokine and Chemokine MRNA Expression in the Human Cervix after Coitus. The Journal of Immunology 2012, 188, 2445–2454. [Google Scholar] [CrossRef] [PubMed]
- Kovacic, M.B.; Katki, H.A.; Kreimer, A.R.; Sherman, M.E. Epidemiologic Analysis of Histologic Cervical Inflammation: Relationship to Human Papillomavirus Infections. Hum Pathol 2008, 39, 1088–1095. [Google Scholar] [CrossRef] [PubMed]
- Potale, Y.; Kasat, Y.K.; Kumar, A.; Ahmad, F. Unravelling the Impact of Human Papillomavirus (HPV): A Comprehensive Exploration of Its Role in Cancer Progression and Global Health Challenges. [CrossRef]
- Faja, F.; Pallotti, F.; Bianchini, S.; Buonacquisto, A.; Cicolani, G.; Conflitti, A.C.; Fracella, M.; Cavallari, E.N.; Sciarra, F.; Pierangeli, A.; et al. Molecular Study of the Presence and Transcriptional Activity of HPV in Semen. J Endocrinol Invest 2024, 47, 557–570. [Google Scholar] [CrossRef]
- Capra, G.; Schillaci, R.; Bosco, L.; Roccheri, M.C.; Perino, A.; Ragusa, M.A. HPV Infection in Semen: Results from a New Molecular Approach. Epidemiol Infect 2019, 147, e177. [Google Scholar] [CrossRef]
- Pérez-Soto, E.; Fernández-Martínez, E.; Oros-Pantoja, R.; Medel-Flores, O.; Miranda-Covarrubias, J.C.; Sánchez-Monroy, V. Proinflammatory and Oxidative Stress States Induced by Human Papillomavirus and Chlamydia Trachomatis Coinfection Affect Sperm Quality in Asymptomatic Infertile Men. Medicina 2021, Vol. 57, Page 862 2021, 57, 862. [Google Scholar] [CrossRef]
- Giannini, S.L.; Al-Saleh, W.; Piron, H.; Jacobs, N.; Doyen, J.; Boniver, J.; Delvenne, P. Cytokine Expression in Squamous Intraepithelial Lesions of the Uterine Cervix: Implications for the Generation of Local Immunosuppression. Clin Exp Immunol 1998, 113, 183–189. [Google Scholar] [CrossRef]
- Torres-Poveda, K.; Bahena-Román, M.; Madrid-González, C.; Burguete-García, A.I.; Bermúdez-Morales, V.H.; Peralta-Zaragoza, O.; Madrid-Marina, V. Role of IL-10 and TGF-Β1 in Local Immunosuppression in HPV-Associated Cervical Neoplasia. World J Clin Oncol 2014, 5, 753. [Google Scholar] [CrossRef]
- Pahne-Zeppenfeld, J.; Schröer, N.; Walch-Rückheim, B.; Oldak, M.; Gorter, A.; Hegde, S.; Smola, S. Cervical Cancer Cell-Derived Interleukin-6 Impairs CCR7-Dependent Migration of MMP-9-Expressing Dendritic Cells. Int J Cancer 2014, 134, 2061–2073. [Google Scholar] [CrossRef]
- Anderson, N.M.; Simon, M.C. The Tumor Microenvironment. Current Biology 2020, 30, R921–R925. [Google Scholar] [CrossRef]
- LaRosa, D.F.; Orange, J.S. 1. Lymphocytes. J Allergy Clin Immunol 2008, 121. [Google Scholar] [CrossRef] [PubMed]
- Ruffell, B.; DeNardo, D.G.; Affara, N.I.; Coussens, L.M. Lymphocytes in Cancer Development: Polarization towards pro-Tumor Immunity. Cytokine Growth Factor Rev 2010, 21, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Kravtsov, D.S.; Erbe, A.K.; Sondel, P.M.; Rakhmilevich, A.L. Roles of CD4+ T Cells as Mediators of Antitumor Immunity. Front Immunol 2022, 13, 972021. [Google Scholar] [CrossRef] [PubMed]
- Togashi, Y.; Shitara, K.; Nishikawa, H. Regulatory T Cells in Cancer Immunosuppression - Implications for Anticancer Therapy. Nat Rev Clin Oncol 2019, 16, 356–371. [Google Scholar] [CrossRef]
- Philip, M.; Schietinger, A. CD8+ T Cell Differentiation and Dysfunction in Cancer. Nat Rev Immunol 2022, 22, 209–223. [Google Scholar] [CrossRef]
- Abel, A.M.; Yang, C.; Thakar, M.S.; Malarkannan, S. Natural Killer Cells: Development, Maturation, and Clinical Utilization. Front Immunol 2018, 9. [Google Scholar] [CrossRef]
- Fantini, M.; Arlen, P.M.; Tsang, K.Y. Potentiation of Natural Killer Cells to Overcome Cancer Resistance to NK Cell-Based Therapy and to Enhance Antibody-Based Immunotherapy. Front Immunol 2023, 14. [Google Scholar] [CrossRef]
- Portale, F.; Di Mitri, D. NK Cells in Cancer: Mechanisms of Dysfunction and Therapeutic Potential. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef]
- Cendrowicz, E.; Sas, Z.; Bremer, E.; Rygiel, T.P. The Role of Macrophages in Cancer Development and Therapy. Cancers 2021, Vol. 13, Page 1946 2021, 13, 1946. [Google Scholar] [CrossRef]
- Liu, J.; Geng, X.; Hou, J.; Wu, G. New Insights into M1/M2 Macrophages: Key Modulators in Cancer Progression. Cancer Cell Int 2021, 21, 1–7. [Google Scholar] [CrossRef]
- Pan, Y.; Yu, Y.; Wang, X.; Zhang, T. Tumor-Associated Macrophages in Tumor Immunity. Front Immunol 2020, 11, 583084. [Google Scholar] [CrossRef] [PubMed]
- Lechien, J.R.; Descamps, G.; Seminerio, I.; Furgiuele, S.; Dequanter, D.; Mouawad, F.; Badoual, C.; Journe, F.; Saussez, S. HPV Involvement in the Tumor Microenvironment and Immune Treatment in Head and Neck Squamous Cell Carcinomas. Cancers 2020, Vol. 12, Page 1060 2020, 12, 1060. [Google Scholar] [CrossRef]
- Gorvel, L.; Olive, D. Tumor Associated Macrophage in HPV+ Tumors: Between Immunosuppression and Inflammation. Semin Immunol 2023, 65, 101671. [Google Scholar] [CrossRef]
- Rosales, C. Neutrophil: A Cell with Many Roles in Inflammation or Several Cell Types? Front Physiol 2018, 9, 324475. [Google Scholar] [CrossRef]
- Wu, L.; Saxena, S.; Singh, R.K. Neutrophils in the Tumor Microenvironment. Adv Exp Med Biol 2020, 1224, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Que, H.; Fu, Q.; Lan, T.; Tian, X.; Wei, X. Tumor-Associated Neutrophils and Neutrophil-Targeted Cancer Therapies. Biochimica et Biophysica Acta (BBA) - Reviews on Cancer 2022, 1877, 188762. [Google Scholar] [CrossRef]
- Dutta, A.; Bhagat, S.; Paul, S.; Katz, J.P.; Sengupta, D.; Bhargava, D. Neutrophils in Cancer and Potential Therapeutic Strategies Using Neutrophil-Derived Exosomes. Vaccines 2023, Vol. 11, Page 1028 2023, 11, 1028. [Google Scholar] [CrossRef]
- Wechsler, M.E.; Munitz, A.; Ackerman, S.J.; Drake, M.G.; Jackson, D.J.; Wardlaw, A.J.; Dougan, S.K.; Berdnikovs, S.; Schleich, F.; Matucci, A.; et al. Eosinophils in Health and Disease: A State-of-the-Art Review. Mayo Clin Proc 2021, 96, 2694–2707. [Google Scholar] [CrossRef] [PubMed]
- Wen, T.; Rothenberg, M.E. The Regulatory Function of Eosinophils. Microbiol Spectr 2016, 4. [Google Scholar] [CrossRef]
- Thippabhotla, S.; Zhong, C.; He, M. 3D Cell Culture Stimulates the Secretion of in Vivo like Extracellular Vesicles. Sci Rep 2019, 9. [Google Scholar] [CrossRef]
- Ghaffari, S.; Rezaei, N. Eosinophils in the Tumor Microenvironment: Implications for Cancer Immunotherapy. J Transl Med 2023, 21. [Google Scholar] [CrossRef] [PubMed]
- Schiffman, M.; Castle, P.E. Human Papillomavirus: Epidemiology and Public Health. Arch Pathol Lab Med 2003, 127, 930–934. [Google Scholar] [CrossRef] [PubMed]
- Brianti, P.; De Flammineis, E.; Mercuri, S.R. Review of HPV-Related Diseases and Cancers.
- Reusser, N.M.; Downing, C.; Guidry, J.; Tyring, S.K. HPV Carcinomas in Immunocompromised Patients. J Clin Med 2015, 4, 260–281. [Google Scholar] [CrossRef] [PubMed]
- Siegel Mph, R.L.; Giaquinto, A.N.; Ahmedin, |; Dvm, J.; Siegel, R.L. Cancer Statistics, 2024. CA Cancer J Clin 2024, 74, 12–49. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.M.; Lin, L.W.; Chen, Y.W.; Ye, Y.L. The Expression and Prognostic Impact of Proinflammatory Cytokines and Their Associations with Carcinogens in Oropharyngeal Squamous Cell Carcinoma. Cancer Immunology, Immunotherapy 2020, 69, 549–558. [Google Scholar] [CrossRef]
- Turksma, A.; Bontkes, H.; van den Heuvel, H.; De Gruijl, T.; Von Blomberg, B.; Braakhuis, B.; Leemans, C.; Bloemena, E.; Meijer, C.; Hooijberg, E. Effector Memory T-Cell Frequencies in Relation to Tumour Stage, Location and HPV Status in HNSCC Patients. Oral Dis 2013, 19, 577–584. [Google Scholar] [CrossRef] [PubMed]
- Matlung, S.E.; Wilhelmina Van Kempen, P.M.; Bovenschen, N.; Baarle, D. Van; Willems, S.M. Differences in T-Cell Infiltrates and Survival between HPV+ and HPV- Oropharyngeal Squamous Cell Carcinoma. Future Sci OA 2016, 2. [Google Scholar] [CrossRef]
- O’Higgins, C.; Ward, F.J.; Eid, R.A. Deciphering the Role of Regulatory CD4 T Cells in Oral and Oropharyngeal Cancer: A Systematic Review. Front Oncol 2018, 8, 402601. [Google Scholar] [CrossRef]
- Snietura, M.; Brewczynski, A.; Kopec, A.; Rutkowski, T. Infiltrates of M2-Like Tumour-Associated Macrophages Are Adverse Prognostic Factor in Patients with Human Papillomavirus-Negative but Not in Human Papillomavirus-Positive Oropharyngeal Squamous Cell Carcinoma. Pathobiology 2020, 87, 75–86. [Google Scholar] [CrossRef]
- Ljokjel, B.; Lybak, H.; Moe, S.E.; Berge, J.E.; Vintermyr, O.K.; Helgeland, L.; Haave, H.; Ljokjel, B.; Lybak, H.; Moe, S.E.; et al. Tumor HPV Status, Level of Regulatory T Cells and Macrophage Infiltration Predict up to 20-Year Non-Disease-Specific Survival in Oropharynx Squamous Cell Carcinoma Patients. Biomedicines 2022, Vol. 10, Page 2484 2022, 10, 2484. [Google Scholar] [CrossRef]
- Al-Sahaf, S.; Hendawi, N.B.; Ollington, B.; Bolt, R.; Ottewell, P.D.; Hunter, K.D.; Murdoch, C. Increased Abundance of Tumour-Associated Neutrophils in HPV-Negative Compared to HPV-Positive Oropharyngeal Squamous Cell Carcinoma Is Mediated by IL-1R Signalling. Frontiers in Oral Health 2021, 2, 604565. [Google Scholar] [CrossRef] [PubMed]
- Rimini, M.; Franco, P.; Bertolini, F.; Berardino, D.B.; giulia, Z.M.; Stefano, V.; Andrikou, K.; Arcadipane, F.; Napolitano, M.; Buno, L.V.; et al. The Prognostic Role of Baseline Eosinophils in HPV-Related Cancers: A Multi-Institutional Analysis of Anal SCC and OPC Patients Treated with Radical CT-RT. J Gastrointest Cancer 2023, 54, 662–671. [Google Scholar] [CrossRef] [PubMed]
- Cervical Cancer Available online:. Available online: https://www.who.int/news-room/fact-sheets/detail/cervical-cancer#:~:text=Key%20facts,%2D%20and%20middle%2Dincome%20countries. (accessed on 28 April 2024).
- van Luijk, I.F.; Smith, S.M.; Marte Ojeda, M.C.; Oei, A.L.; Kenter, G.G.; Jordanova, E.S. A Review of the Effects of Cervical Cancer Standard Treatment on Immune Parameters in Peripheral Blood, Tumor Draining Lymph Nodes, and Local Tumor Microenvironment. J Clin Med 2022, 11. [Google Scholar] [CrossRef]
- De Jong, A.; Van Poelgeest, M.I.E.; Van Der Hulst, J.M.; Drijfhout, J.W.; Fleuren, G.J.; Melief, C.J.M.; Renter, G.; Offringa, R.; Van Der Burg, S.H. Human Papillomavirus Type 16-Positive Cervical Cancer Is Associated with Impaired CD4+ T-Cell Immunity against Early Antigens E2 and E6. Cancer Res 2004, 64, 5449–5455. [Google Scholar] [CrossRef] [PubMed]
- I, A.; RO, A.; A, W.A.; S, A.A. Role of T Cells in Cervical Cancer. Bioinformation 2023, 19, 556–561. [Google Scholar] [CrossRef]
- Das, D.; Sarkar, B.; Mukhopadhyay, S.; Banerjee, C.; Mondal, S.B. An Altered Ratio of CD4+ And CD8+ T Lymphocytes in Cervical Cancer Tissues and Peripheral Blood – A Prognostic Clue? Asian Pac J Cancer Prev 2018, 19, 471. [Google Scholar] [CrossRef]
- B C Sheu 1, S.M.H. Reversed CD4/CD8 Ratios of Tumor-Infiltrating Lymphocytes Are Correlated with the Progression of Human Cervical Carcinoma - PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/10526283/ (accessed on 28 April 2024).
- Litwin, T.R.; Irvin, S.R.; Chornock, R.L.; Sahasrabuddhe, V. V.; Stanley, M.; Wentzensen, N. Infiltrating T-Cell Markers in Cervical Carcinogenesis: A Systematic Review and Meta-Analysis. Br J Cancer 2021, 124, 831–841. [Google Scholar] [CrossRef]
- Jang, S.; Jang, S.; Nam, E.; Lee, D.; Hong, J.; Kim, S.; Kim, S.; Kim, J.; Kim, Y.; Kim, J. Relationship between the Proportion of Natural Killer Cells in Peripheral Blood Lymphocytes and Risk Factors in Cervical Cancer. 2008, 26, 16546–16546. [Google Scholar] [CrossRef]
- Wu, Y.; Ye, S.; Goswami, S.; Pei, X.; Xiang, L.; Zhang, X.; Yang, H. Clinical Significance of Peripheral Blood and Tumor Tissue Lymphocyte Subsets in Cervical Cancer Patients. BMC Cancer 2020, 20, 1–12. [Google Scholar] [CrossRef]
- Petrillo, M.; Zannoni, G.F.; Martinelli, E.; Anchora, L.P.; Ferrandina, G.; Tropeano, G.; Fagotti, A.; Scambia, G. Polarisation of Tumor-Associated Macrophages toward M2 Phenotype Correlates with Poor Response to Chemoradiation and Reduced Survival in Patients with Locally Advanced Cervical Cancer. PLoS One 2015, 10. [Google Scholar] [CrossRef]
- Lee, H.W.; Choi, H.J.; Ha, S.J.; Lee, K.T.; Kwon, Y.G. Recruitment of Monocytes/Macrophages in Different Tumor Microenvironments. Biochim Biophys Acta 2013, 1835, 170–179. [Google Scholar] [CrossRef] [PubMed]
- Stone, S.C.; Rossetti, R.A.M.; Alvarez, K.L.F.; Carvalho, J.P.; Margarido, P.F.R.; Baracat, E.C.; Tacla, M.; Boccardo, E.; Yokochi, K.; Lorenzi, N.P.; et al. Lactate Secreted by Cervical Cancer Cells Modulates Macrophage Phenotype. J Leukoc Biol 2019, 105, 1041–1054. [Google Scholar] [CrossRef] [PubMed]
- Tas, M.; Yavuz, A.; Ak, M.; Ozcelik, B. Neutrophil-to-Lymphocyte Ratio and Platelet-to-Lymphocyte Ratio in Discriminating Precancerous Pathologies from Cervical Cancer. J Oncol 2019, 2019. [Google Scholar] [CrossRef] [PubMed]
- Du, J.Q.; Zhang, F.; Wang, C.Q.; Zhu, J.F.; Xu, L.X.; Yang, Y.H.; Han, M.F.; Hu, Y. Effects of Peripheral Blood Neutrophil/Lymphocyte Ratio Levels and Their Changes on the Prognosis of Patients with Early Cervical Cancer. Front Oncol 2023, 13, 1139809. [Google Scholar] [CrossRef]
- Holub, K.; Biete, A. Impact of Systemic Inflammation Biomarkers on the Survival Outcomes of Cervical Cancer Patients. Clin Transl Oncol 2019, 21, 836–844. [Google Scholar] [CrossRef]
- Chadha, J.; Chahoud, J.; Spiess, P.E. An Update on Treatment of Penile Cancer. Ther Adv Med Oncol 2022, 14. [Google Scholar] [CrossRef]
- L, I.; R, D.M.; C, C.D.; A, M.A.S.; A, P.S.; D, L.D.M.; M, S.; S, B.; T, C.; D, B.; et al. Penile Carcinoma and HPV Infection (Review). Exp Ther Med 2020, 20. [Google Scholar] [CrossRef]
- Chahoud, J.; Netto, F.; Lazcano Segura, R.; Parra Cuentas, E.R.; Lu, X.; Rao, P.; Wistuba, I.I.; Pickering, C.R.; Pettaway, C.A. Tumor Immune Microenvironment Alterations in Penile Squamous Cell Carcinoma Using Multiplex Immunofluorescence and Image Analysis Approaches. Journal of Clinical Oncology 2020, 38, 4–4. [Google Scholar] [CrossRef]
- Ottenhof, S.R.; Djajadiningrat, R.S.; Thygesen, H.H.; Jakobs, P.J.; Józwiak, K.; Heeren, A.M.; de Jong, J.; Sanders, J.; Horenblas, S.; Jordanova, E.S. The Prognostic Value of Immune Factors in the Tumor Microenvironment of Penile Squamous Cell Carcinoma. Front Immunol 2018, 9. [Google Scholar] [CrossRef]
- Matthias Walter Tumor Microenvironment in Penile Cancer. 2020.
- Campos, M.M.; Ornellasde Souza, M.H.; Pires, V.; Scheiner, M.A.M.; Esteves, E.B.; Ornellas, A.A. Clinical Implications of Natural Killer Cytotoxicity in Patients with Squamous Cell Carcinoma of the Penis. Nat Immun 1998, 16, 256–262. [Google Scholar] [CrossRef]
- Chu, C.; Yao, K.; Lu, J.; Zhang, Y.; Chen, K.; Lu, J.; Zhang, C.Z.; Cao, Y. Immunophenotypes Based on the Tumor Immune Microenvironment Allow for Unsupervised Penile Cancer Patient Stratification. Cancers (Basel) 2020, 12, 1–17. [Google Scholar] [CrossRef] [PubMed]
- de Vries, H.M.; Ottenhof, S.R.; Horenblas, S.; van der Heijden, M.S.; Jordanova, E.S. Defining the Tumor Microenvironment of Penile Cancer by Means of the Cancer Immunogram. Eur Urol Focus 2019, 5, 718–721. [Google Scholar] [CrossRef] [PubMed]
- Ono, Y.; Ozawa, M.; Tamura, Y.; Suzuki, T.; Suzuki, K.; Kurokawa, K.; Fukabori, Y.; Yamanaka, H. Tumor-Associated Tissue Eosinophilia of Penile Cancer. International Journal of Urology 2002, 9, 82–87. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.H.; Lin, Y.J.; Chen, Y.C.; Liu, I.L.; You, S.L.; Hu, J.M.; Lin, T.C.; Chang, P.K.; Chen, C.Y.; Chou, Y.C.; et al. Human Papillomavirus and Risk of Colorectal Cancer: An Analysis of Nationwide Claims Data. Medicina (Kaunas) 2022, 58. [Google Scholar] [CrossRef]
- Damin, D.C.; Ziegelmann, P.K.; Damin, A.P. Human Papillomavirus Infection and Colorectal Cancer Risk: A Meta-Analysis. Colorectal Dis 2013, 15. [Google Scholar] [CrossRef]
- Zhou, J.; Liu, Y.; Zhang, Y.; Ling, F.; Zheng, J.; Yao, X.; Lyu, Z.; Feng, H.; Li, Y. Comprehensive Analysis of a Novel Subtype of Immune Microenvironment-Derived HPV-Infected Colorectal Cancer. Microbes Infect 2024, 105315. [Google Scholar] [CrossRef]
- Ambrosio, M.R.; Niccolai, E.; Petrelli, F.; Di Gloria, L.; Bertacca, G.; Giusti, A.; Baldi, S.; Cavazzana, A.; Palmeri, M.; Perotti, B.; et al. Immune Landscape and Oncobiota in HPV-Associated Colorectal Cancer: An Explorative Study. Clin Exp Med 2023, 23, 5101–5112. [Google Scholar] [CrossRef]
- Gondal, T.A.; Chaudhary, N.; Bajwa, H.; Rauf, A.; Le, D.; Ahmed, S. Anal Cancer: The Past, Present and Future. Curr Oncol 2023, 30, 3232–3250. [Google Scholar] [CrossRef] [PubMed]
- Elnaggar, J.H.; Huynh, V.O.; Lin, D.; Hillman, R.T.; Abana, C.O.; El Alam, M.B.; Tomasic, K.C.; Karpinets, T. V.; Kouzy, R.; Phan, J.L.; et al. HPV-Related Anal Cancer Is Associated with Changes in the Anorectal Microbiome during Cancer Development. Front Immunol 2023, 14, 1051431. [Google Scholar] [CrossRef]
- Balermpas, P.; Martin, D.; Wieland, U.; Rave-Fränk, M.; Strebhardt, K.; Rödel, C.; Fokas, E.; Rödel, F. Human Papilloma Virus Load and PD-1/PD-L1, CD8+ and FOXP3 in Anal Cancer Patients Treated with Chemoradiotherapy: Rationale for Immunotherapy. Oncoimmunology 2017, 6. [Google Scholar] [CrossRef]
- Dhawan, N.; Afzal, M.Z.; Amin, M. Immunotherapy in Anal Cancer. Curr Oncol 2023, 30, 4538–4550. [Google Scholar] [CrossRef] [PubMed]
- Martin, D.; Rödel, F.; Balermpas, P.; Rödel, C.; Fokas, E. The Immune Microenvironment and HPV in Anal Cancer: Rationale to Complement Chemoradiation with Immunotherapy. Biochim Biophys Acta Rev Cancer 2017, 1868, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Schernberg, A.; Escande, A.; Rivin Del Campo, E.; Ducreux, M.; Nguyen, F.; Goere, D.; Chargari, C.; Deutsch, E. Leukocytosis and Neutrophilia Predicts Outcome in Anal Cancer. Radiother Oncol 2017, 122, 137–145. [Google Scholar] [CrossRef]
- Toh, E.; Wilson, J.; Sebag-Montefiore, D.; Botterill, I. Neutrophil:Lymphocyte Ratio as a Simple and Novel Biomarker for Prediction of Locoregional Recurrence after Chemoradiotherapy for Squamous Cell Carcinoma of the Anus. Colorectal Dis 2014, 16. [Google Scholar] [CrossRef] [PubMed]
- Erez, N.; Truitt, M.; Olson, P.; Hanahan, D. Article Cancer-Associated Fibroblasts Are Activated in Incipient Neoplasia to Orchestrate Tumor-Promoting Inflammation in an NF-KB-Dependent Manner. Cancer Cell 17 135–147. [CrossRef]
- Barros, M.R.; De Melo, C.M.L.; Barros, M.L.C.M.G.R.; De Cássia Pereira De Lima, R.; De Freitas, A.C.; Venuti, A. Activities of Stromal and Immune Cells in HPV-Related Cancers. Journal of Experimental & Clinical Cancer Research 2018 37:1 2018, 37, 1–18. [Google Scholar] [CrossRef]
- Cirri, P.; Chiarugi, P. Cancer-Associated-Fibroblasts and Tumour Cells: A Diabolic Liaison Driving Cancer Progression. Cancer and Metastasis Reviews 2011 31:1 2011, 31, 195–208. [Google Scholar] [CrossRef]
- Id, K.S.; Mcbride, A.A.; Munger, K. Changing Stem Cell Dynamics during Papillomavirus Infection: Potential Roles for Cellular Plasticity in the Viral Lifecycle and Disease. Viruses 2017, Vol. 9, Page 221 2017, 9, 221. [Google Scholar] [CrossRef]
- Greshock, J.; Nathanson, K.; Martin, A.M.; Zhang, L.; Coukos, G.; Weber, B.L.; Zaks, T.Z. Cancer Cell Lines as Genetic Models of Their Parent Histology: Analyses Based on Array Comparative Genomic Hybridization. Cancer Res 2007, 67, 3594–3600. [Google Scholar] [CrossRef]
- Perkel, J.M. Curing Cell Lines. Biotechniques 2011, 51, 85–90. [Google Scholar] [CrossRef]
- Kasai, F.; Hirayama, N.; Ozawa, M.; Iemura, M.; Kohara, A. Changes of Heterogeneous Cell Populations in the Ishikawa Cell Line during Long-Term Culture: Proposal for an in Vitro Clonal Evolution Model of Tumor Cells. Genomics 2016, 107, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Cooper, J.R.; Abdullatif, M.B.; Burnett, E.C.; Kempsell, K.E.; Conforti, F.; Tolley, H.; Collins, J.E.; Davies, D.E. Long Term Culture of the A549 Cancer Cell Line Promotes Multilamellar Body Formation and Differentiation towards an Alveolar Type II Pneumocyte Phenotype. PLoS One 2016, 11, e0164438. [Google Scholar] [CrossRef] [PubMed]
- Barrett, T.A.; Wu, A.; Zhang, H.; Levy, M.S.; Lye, G.J. Microwell Engineering Characterization for Mammalian Cell Culture Process Development. Biotechnol Bioeng 2010, 105, 260–275. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Liu, A.; Yang, X.; Gong, J.; Yu, M.; Yao, X.; Wang, H.; He, Y. 3D Cell Culture—Can It Be As Popular as 2D Cell Culture? Adv Nanobiomed Res 2021, 1, 2000066. [Google Scholar] [CrossRef]
- Huh, D.; Hamilton, G.A.; Ingber, D.E. From 3D Cell Culture to Organs-on-Chips. Trends Cell Biol 2011, 21, 745–754. [Google Scholar] [CrossRef]
- Bielecka, Z.F.; Maliszewska-Olejniczak, K.; Safir, I.J.; Szczylik, C.; Czarnecka, A.M. Three-Dimensional Cell Culture Model Utilization in Cancer Stem Cell Research. Biological Reviews 2017, 92, 1505–1520. [Google Scholar] [CrossRef] [PubMed]
- Goodman, T.T.; Chee, P.N.; Suzie, H.P. 3-D Tissue Culture Systems for the Evaluation and Optimization of Nanoparticle-Based Drug Carriers. Bioconjug Chem 2008, 19, 1951–1959. [Google Scholar] [CrossRef]
- Chauhan, V.P.; Stylianopoulos, T.; Boucher, Y.; Jain, R.K. Delivery of Molecular and Nanoscale Medicine to Tumors: Transport Barriers and Strategies. Annu Rev Chem Biomol Eng 2011, 2, 281–298. [Google Scholar] [CrossRef]
- Larue, L.; Beermann, F. Cutaneous Melanoma in Genetically Modified Animals. Pigment Cell Res 2007, 20, 485–497. [Google Scholar] [CrossRef]
- Cheon, D.J.; Orsulic, S. Mouse Models of Cancer. Annual Review of Pathology: Mechanisms of Disease 2011, 6, 95–119. [Google Scholar] [CrossRef]
- Herreros-Pomares, A.; Zhou, X.; Calabuig-Fariñas, S.; Lee, S.J.; Torres, S.; Esworthy, T.; Hann, S.Y.; Jantus-Lewintre, E.; Camps, C.; Zhang, L.G. 3D Printing Novel in Vitro Cancer Cell Culture Model Systems for Lung Cancer Stem Cell Study. Materials Science and Engineering: C 2021, 122, 111914. [Google Scholar] [CrossRef]
- Geiger, P.; Mayer, B.; Wiest, I.; Schulze, S.; Jeschke, U.; Weissenbacher, T. Binding of Galectin-1 to Breast Cancer Cells MCF7 Induces Apoptosis and Inhibition of Proliferation in Vitro in a 2D- and 3D- Cell Culture Model. BMC Cancer 2016, 16, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Mosaad, E.; Chambers, K.; Futrega, K.; Clements, J.; Doran, M.R. Using High Throughput Microtissue Culture to Study the Difference in Prostate Cancer Cell Behavior and Drug Response in 2D and 3D Co-Cultures. BMC Cancer 2018, 18, 1–11. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, D.; Wu, G.; Wu, J.; Lu, S.; Lo, J.; He, Y.; Zhao, C.; Zhao, X.; Zhang, H.; et al. Metastasis-on-a-Chip Mimicking the Progression of Kidney Cancer in the Liver for Predicting Treatment Efficacy. Theranostics 2020, 10, 300. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yu, Y.; Guo, J.; Lu, W.; Wei, Q.; Zhao, Y. The Construction and Application of Three-Dimensional Biomaterials. Adv Biosyst 2020, 4, 1900238. [Google Scholar] [CrossRef]
- Mehta, G.; Hsiao, A.Y.; Ingram, M.; Luker, G.D.; Takayama, S. Opportunities and Challenges for Use of Tumor Spheroids as Models to Test Drug Delivery and Efficacy. Journal of Controlled Release 2012, 164, 192–204. [Google Scholar] [CrossRef]
- Jang, S.; Jang, S.; Nam, E.; Lee, D.; Hong, J.; Kim, S.; Kim, S.; Kim, J.; Kim, Y.; Kim, J. Relationship between the Proportion of Natural Killer Cells in Peripheral Blood Lymphocytes and Risk Factors in Cervical Cancer. 2008, 26, 16546–16546. [Google Scholar] [CrossRef]
- Baek, M.H.; Kim, D.Y.; Kim, N.; Rhim, C.C.; Kim, J.H.; Nam, J.H. Incorporating a 3-Dimensional Printer into the Management of Early-Stage Cervical Cancer. J Surg Oncol 2016, 114, 150–152. [Google Scholar] [CrossRef] [PubMed]
- Huo, W.; Ding, Y.; Sheng, C.; Pi, Y.; Guo, Y.; Wu, A.; Zhang, Z. Application of 3D Printing in Cervical Cancer Brachytherapy. J Radiat Res Appl Sci 2022, 15, 18–24. [Google Scholar] [CrossRef]
- Zhao, Y.; Yao, R.; Ouyang, L.; Ding, H.; Zhang, T.; Zhang, K.; Cheng, S.; Sun, W. Three-Dimensional Printing of Hela Cells for Cervical Tumor Model in Vitro. Biofabrication 2014, 6, 035001. [Google Scholar] [CrossRef]
- Kalaivani, Muniandy; Annabel Dass, S.; Shamsuddin, S.; Mohana-Kumaran, N.; Balakrishnan, V.; Muniandy, K.; Asra Ahmad, Z.; Mohana Kumaran, N.; Author, C. Growth and Invasion of 3D Spheroid Tumor of HeLa and CasKi Cervical Cancer Cells. 2021. [Google Scholar] [CrossRef]
- Zuk, A.K.; Wen, X.; Dilworth, S.; Li, D.; Ghali, L. Modeling and Validating Three Dimensional Human Normal Cervix and Cervical Cancer Tissues in Vitro. J Biomed Res 2017, 31, 240. [Google Scholar] [CrossRef]
- De Gregorio, V.; La Rocca, A.; Urciuolo, F.; Annunziata, C.; Tornesello, M.L.; Buonaguro, F.M.; Netti, P.A.; Imparato, G. Modeling the Epithelial-Mesenchymal Transition Process in a 3D Organotypic Cervical Neoplasia. Acta Biomater 2020, 116, 209–222. [Google Scholar] [CrossRef]
- Lin, Z.; Li, X.; Shi, H.; Cao, R.; Zhu, L.; Dang, C.; Sheng, Y.; Fan, W.; Yang, Z.; Wu, S. Decoding the Tumor Microenvironment and Molecular Mechanism: Unraveling Cervical Cancer Subpopulations and Prognostic Signatures through ScRNA-Seq and Bulk RNA-Seq Analyses. Front Immunol 2024, 15, 1351287. [Google Scholar] [CrossRef]
- Jackson, R.; Maarsingh, J.D.; Herbst-Kralovetz, M.M.; Van Doorslaer, K. 3D Oral and Cervical Tissue Models for Studying Papillomavirus Host-Pathogen Interactions. Curr Protoc Microbiol 2020, 59, e129. [Google Scholar] [CrossRef] [PubMed]
- Mao, J.; Yang, W.; Guo, H.; Dong, R.; Ren, L.; Li, S. A Cervical Cancer Tissue-Derived Decellularized Extracellular Matrix Scaffold for Cervical Cancer Tissue Reconstruction in Vitro. Journal of Southern Medical University 2023, 43, 157. [Google Scholar] [CrossRef]
- Miserocchi, G.; Cocchi, C.; De Vita, A.; Liverani, C.; Spadazzi, C.; Calpona, S.; Di Menna, G.; Bassi, M.; Meccariello, G.; De Luca, G.; et al. Three-Dimensional Collagen-Based Scaffold Model to Study the Microenvironment and Drug-Resistance Mechanisms of Oropharyngeal Squamous Cell Carcinomas. Cancer Biol Med 2021, 18, 502. [Google Scholar] [CrossRef] [PubMed]
- Colley, H.E.; Hearnden, V.; Jones, A. V.; Weinreb, P.H.; Violette, S.M.; MacNeil, S.; Thornhill, M.H.; Murdoch, C. Development of Tissue-Engineered Models of Oral Dysplasia and Early Invasive Oral Squamous Cell Carcinoma. British Journal of Cancer 2011 105:10 2011, 105, 1582–1592. [Google Scholar] [CrossRef]
- Almela, T.; Al-Sahaf, S.; Brook, I.M.; Khoshroo, K.; Rasoulianboroujeni, M.; Fahimipour, F.; Tahriri, M.; Dashtimoghadam, E.; Bolt, R.; Tayebi, L.; et al. 3D Printed Tissue Engineered Model for Bone Invasion of Oral Cancer. Tissue Cell 2018, 52, 71–77. [Google Scholar] [CrossRef]
- Noi, M.; Mukaisho, K.I.; Yoshida, S.; Murakami, S.; Koshinuma, S.; Adachi, T.; Machida, Y.; Yamori, M.; Nakayama, T.; Yamamoto, G.; et al. ERK Phosphorylation Functions in Invadopodia Formation in Tongue Cancer Cells in a Novel Silicate Fibre-Based 3D Cell Culture System. International Journal of Oral Science 2018 10:4 2018, 10, 1–10. [Google Scholar] [CrossRef]
- Vincent-Chong, V.K.; Seshadri, M. Development and Radiation Response Assessment in A Novel Syngeneic Mouse Model of Tongue Cancer: 2D Culture, 3D Organoids and Orthotopic Allografts. Cancers 2020, Vol. 12, Page 579 2020, 12, 579. [Google Scholar] [CrossRef] [PubMed]
- Lei, K.F.; Wu, M.H.; Hsu, C.W.; Chen, Y.D. Real-Time and Non-Invasive Impedimetric Monitoring of Cell Proliferation and Chemosensitivity in a Perfusion 3D Cell Culture Microfluidic Chip. Biosens Bioelectron 2014, 51, 16–21. [Google Scholar] [CrossRef]
- Igawa, K.; Izumi, K.; Sakurai, Y. Development of the Follow-Up Human 3D Oral Cancer Model in Cancer Treatment. BioTech 2023, Vol. 12, Page 35 2023, 12, 35. [Google Scholar] [CrossRef]
- Tanaka, N.; Osman, A.A.; Takahashi, Y.; Lindemann, A.; Patel, A.A.; Zhao, M.; Takahashi, H.; Myers, J.N. Head and Neck Cancer Organoids Established by Modification of the CTOS Method Can Be Used to Predict in Vivo Drug Sensitivity. Oral Oncol 2018, 87, 49–57. [Google Scholar] [CrossRef]
- Sawant, S.; Dongre, H.; Singh, A.K.; Joshi, S.; Costea, D.E.; Mahadik, S.; Ahire, C.; Makani, V.; Dange, P.; Sharma, S.; et al. Establishment of 3D Co-Culture Models from Different Stages of Human Tongue Tumorigenesis: Utility in Understanding Neoplastic Progression. PLoS One 2016, 11, e0160615. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Stange, D.E.; Ferrante, M.; Vries, R.G.J.; Van Es, J.H.; Van Den Brink, S.; Van Houdt, W.J.; Pronk, A.; Van Gorp, J.; Siersema, P.D.; et al. Long-Term Expansion of Epithelial Organoids from Human Colon, Adenoma, Adenocarcinoma, and Barrett’s Epithelium. Gastroenterology 2011, 141, 1762–1772. [Google Scholar] [CrossRef] [PubMed]
- Abbas, Z.N.; Al-Saffar, A.Z.; Jasim, S.M.; Sulaiman, G.M. Comparative Analysis between 2D and 3D Colorectal Cancer Culture Models for Insights into Cellular Morphological and Transcriptomic Variations. Scientific Reports 2023 13:1 2023, 13, 1–16. [Google Scholar] [CrossRef]
- Koch, J.; Mönch, D.; Maaß, A.; Gromoll, C.; Hehr, T.; Leibold, T.; Schlitt, H.J.; Dahlke, M.H.; Renner, P. Three Dimensional Cultivation Increases Chemo- and Radioresistance of Colorectal Cancer Cell Lines. PLoS One 2021, 16, e0244513. [Google Scholar] [CrossRef] [PubMed]
- Magdeldin, T.; López-Dávila, V.; Villemant, C.; Cameron, G.; Drake, R.; Cheema, U.; Loizidou, M. The Efficacy of Cetuximab in a Tissue-Engineered Three-Dimensional in Vitro Model of Colorectal Cancer. J Tissue Eng 2014, 5. [Google Scholar] [CrossRef]
- Cives, M.; Mannavola, F.; Lospalluti, L.; Sergi, M.C.; Cazzato, G.; Filoni, E.; Cavallo, F.; Giudice, G.; Stucci, L.S.; Porta, C.; et al. Non-Melanoma Skin Cancers: Biological and Clinical Features. Int J Mol Sci 2020, 21, 1–24. [Google Scholar] [CrossRef]
- Berning, M.; Prätzel-Wunder, S.; Bickenbach, J.R.; Boukamp, P. Three-Dimensional In Vitro Skin and Skin Cancer Models Based on Human Fibroblast-Derived Matrix. https://home.liebertpub.com/tec 2015, 21, 958–970. [Google Scholar] [CrossRef] [PubMed]
- Vörsmann, H.; Groeber, F.; Walles, H.; Busch, S.; Beissert, S.; Walczak, H.; Kulms, D. Development of a Human Three-Dimensional Organotypic Skin-Melanoma Spheroid Model for in Vitro Drug Testing. Cell Death & Disease 2013 4:7 2013, 4, e719–e719. [Google Scholar] [CrossRef]
- Hill, D.S.; Robinson, N.D.P.; Caley, M.P.; Chen, M.; O’Toole, E.A.; Armstrong, J.L.; Przyborski, S.; Lovat, P.E. A Novel Fully Humanized 3D Skin Equivalent to Model Early Melanoma Invasion. Mol Cancer Ther 2015, 14, 2665–2673. [Google Scholar] [CrossRef]
- Visalakshan, R.M.; Lowrey, M.K.; Sousa, M.G.C.; Helms, H.R.; Samiea, A.; Schutt, C.E.; Moreau, J.M.; Bertassoni, L.E. Opportunities and Challenges to Engineer 3D Models of Tumor-Adaptive Immune Interactions. Front Immunol 2023, 14, 1162905. [Google Scholar] [CrossRef]
- Lee, J.Y.; Chaudhuri, O. Modeling the Tumor Immune Microenvironment for Drug Discovery Using 3D Culture. APL Bioeng 2021, 5, 10903. [Google Scholar] [CrossRef] [PubMed]
- Pinto, B.; Henriques, A.C.; Silva, P.M.A.; Bousbaa, H. Three-Dimensional Spheroids as In Vitro Preclinical Models for Cancer Research. Pharmaceutics 2020, Vol. 12, Page 1186 2020, 12, 1186. [Google Scholar] [CrossRef]
- Zhu, H.; Luo, S.F.; Wang, J.; Li, X.; Wang, H.; Pu, W.Y.; Zhang, H.; Zhuang, Z.X. Effect of Environmental Factors on Chemoresistance of HepG2 Cells by Regulating Hypoxia-Inducible Factor-1α. Chin Med J (Engl) 2012, 125, 1095–1103. [Google Scholar] [CrossRef]
- Zhu, H.; Chen, X.P.; Luo, S.F.; Guan, J.; Zhang, W.G.; Zhang, B.X. Involvement of Hypoxia-Inducible Factor-1-Alpha in Multidrug Resistance Induced by Hypoxia in HepG2 Cells. J Exp Clin Cancer Res 2005, 24, 565–574. [Google Scholar]
- Wartenberg, M.; Frey, C.; Diedershagen, H.; Ritgen, J.; Hescheler, J.; Sauer, H. DEVELOPMENT OF AN INTRINSIC P-GLYCOPROTEIN-MEDIATED DOXORUBICIN RESISTANCE IN QUIESCENT CELL LAYERS OF LARGE, MULTICELLULAR PROSTATE TUMOR SPHEROIDS. J. Cancer 1998, 75, 855–863. [Google Scholar] [CrossRef]
- Milane, L.; Duan, Z.; Amiji, M. Role of Hypoxia and Glycolysis in the Development of Multi-Drug Resistance in Human Tumor Cells and the Establishment of an Orthotopic Multi-Drug Resistant Tumor Model in Nude Mice Using Hypoxic Pre-Conditioning. Cancer Cell Int 2011, 11, 1–16. [Google Scholar] [CrossRef]
- Webb, B.A.; Chimenti, M.; Jacobson, M.P.; Barber, D.L. Dysregulated PH: A Perfect Storm for Cancer Progression. Nature Reviews Cancer 2011 11:9 2011, 11, 671–677. [Google Scholar] [CrossRef] [PubMed]
- Correia, A.L.; Bissell, M.J. The Tumor Microenvironment Is a Dominant Force in Multidrug Resistance. Drug Resistance Updates 2012, 15, 39–49. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Illustration of HPV E5/E6/E7-mediated cellular transformation.
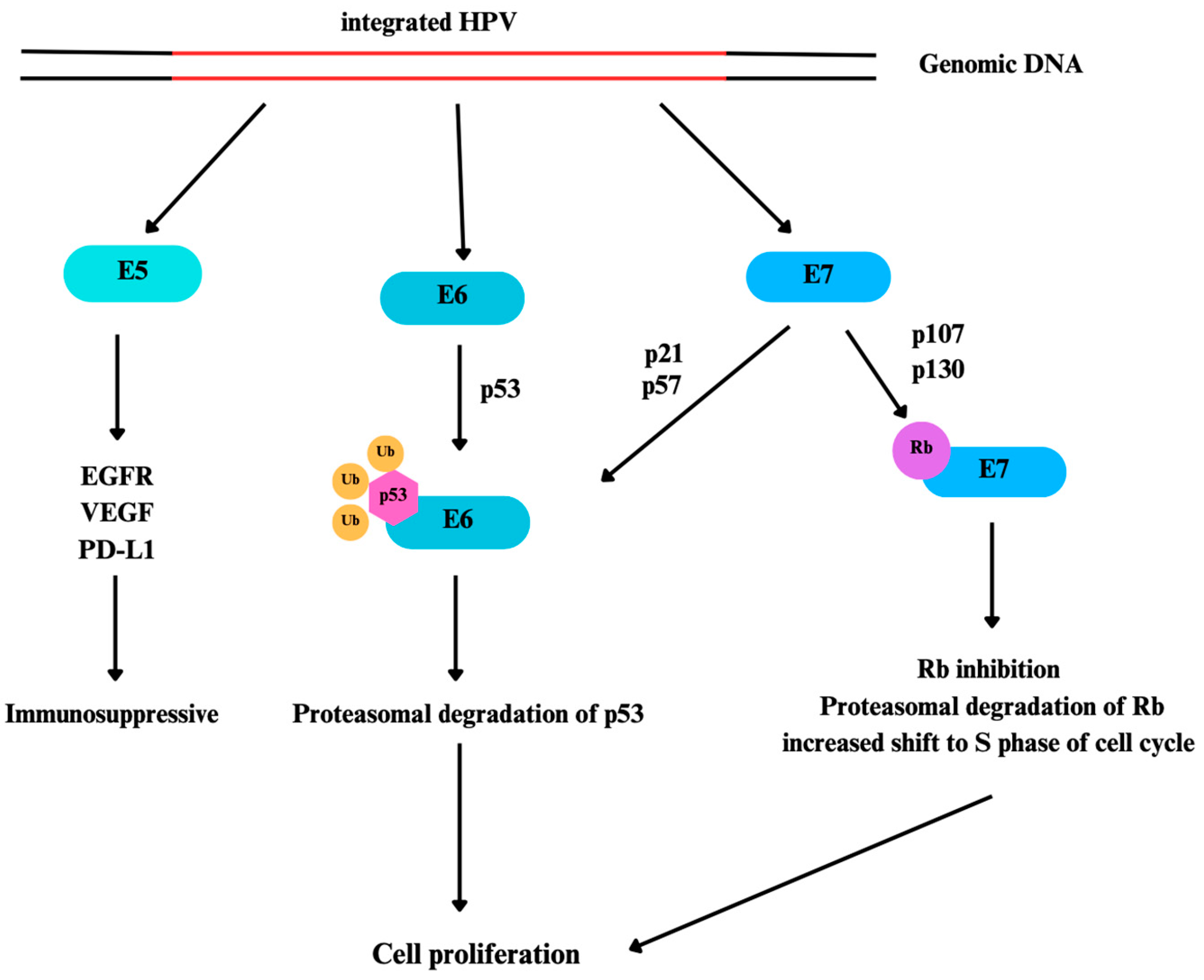
Figure 2.
Schematic illustration of TME of CVC. Many immune cells and CAFs contribute to TME. Also, extracellular matrix plays an important role in TME. Taken together, TME has vital roles in carcinogenesis, cancer progression, invasion, and metastasis in CVC.
Figure 2.
Schematic illustration of TME of CVC. Many immune cells and CAFs contribute to TME. Also, extracellular matrix plays an important role in TME. Taken together, TME has vital roles in carcinogenesis, cancer progression, invasion, and metastasis in CVC.
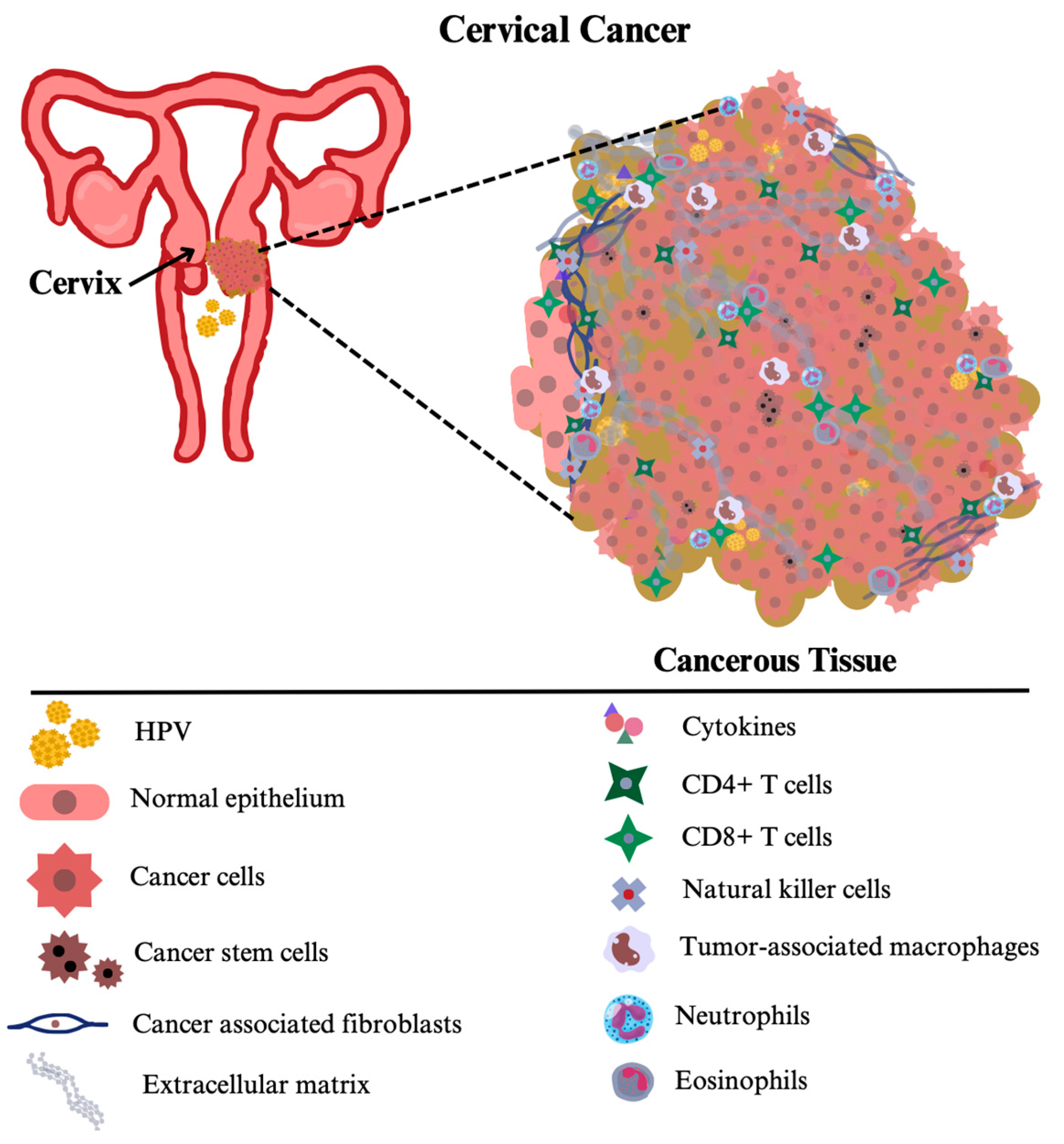
Figure 3.
2D cell culture systems grow cells in a flat, single-layer environment, which restricts them to only cell-cell interactions. In contrast, 3D cell culture models better mimic the tumor microenvironment by allowing cells to interact with each other and the surrounding matrix in a three-dimensional space. This 3D approach can support multiple cell types within one environment, reflecting the complexity and diversity of actual tumors.
Figure 3.
2D cell culture systems grow cells in a flat, single-layer environment, which restricts them to only cell-cell interactions. In contrast, 3D cell culture models better mimic the tumor microenvironment by allowing cells to interact with each other and the surrounding matrix in a three-dimensional space. This 3D approach can support multiple cell types within one environment, reflecting the complexity and diversity of actual tumors.
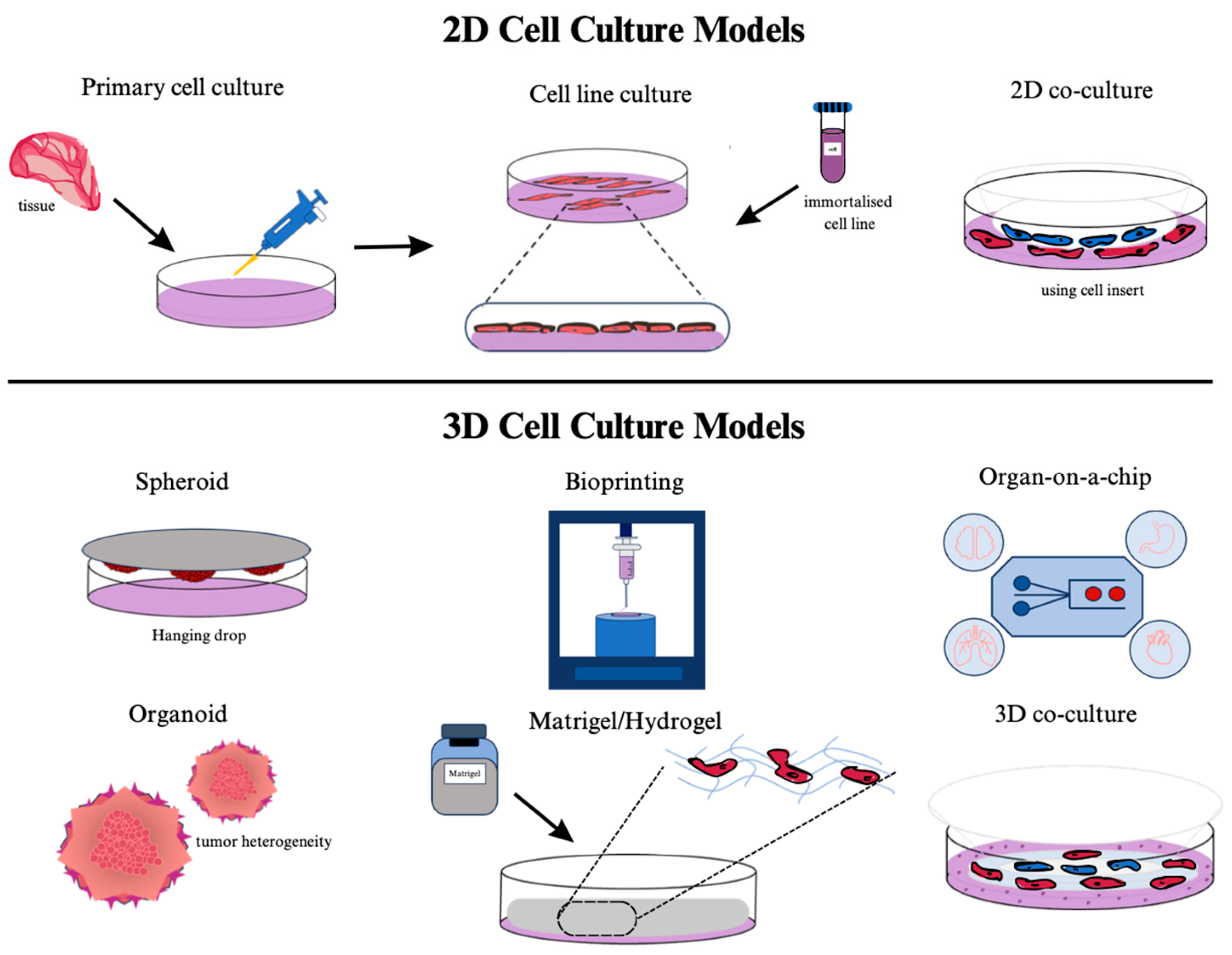
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



