
Submitted:
10 September 2024
Posted:
10 September 2024
You are already at the latest version
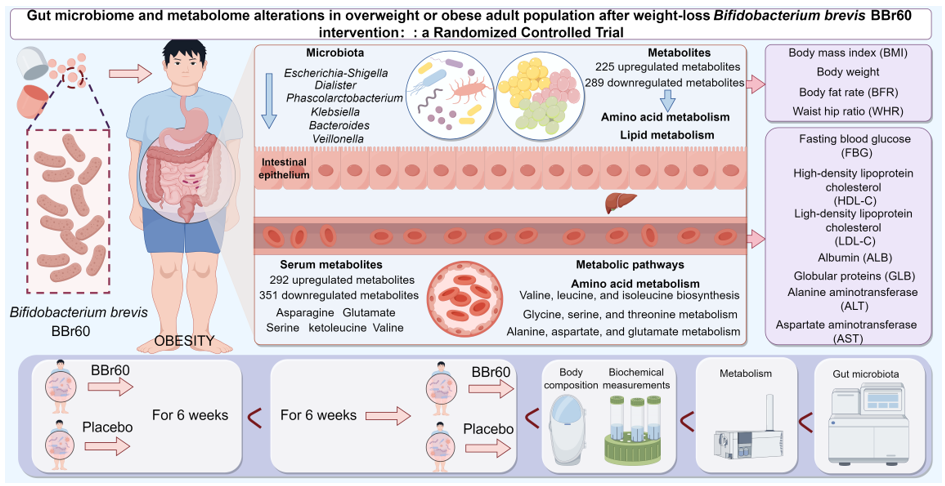
Abstract
Probiotics, known for regulating gut microbiota, may aid those with overweight or obesity, but their mechanisms require more research. This study involved 75 overweight or obese young adults, randomly assigned to either a Bifidobacterium brevis BBr60 group or a placebo group. Both groups received diet guidance and took either BBr60 (1×1010 CFU/day) or a placebo for 12 weeks. Re-searchers analyzed body composition, serum glucose, lipids, liver and kidney function, comprehensive metabolome, and intestinal homeostasis before and after the intervention. After 12 weeks, BBr60 significantly reduced weight and BMI compared to pre-treatment levels and outperformed the placebo. The BBr60 group also showed improved blood biochemistry, with notably lower fasting blood glucose (FBG) levels than the placebo group (p < 0.05). Additionally, BBr60 influenced vital serum and fecal metabolites related to three amino acid metabolic pathways and regulated the bacteria Dialister, Klebsiella, and Bacteroides, which correlated strongly with serum metabolites. These findings indicate that BBr60 can safely and effectively regulating BMI, body weight, serum glucose, lipids, and liver function markers, which may involve BBr60's impact on key gut bacteria, which influence metabolites related to the valine, leucine, and isoleucine biosynthesis; glycine, serine, and threonine metabolism; and alanine, aspartate, and glutamate metabolism.
Keywords:
Bifidobacterium brevis BBr60
; overweight or obese
; randomized controlled trial
; gut microbiota
; metabolic profile
1. Introduction
The global obesity epidemic has emerged as a significant public health concern due to the prevalence of poor dietary choices and sedentary lifestyles in conjunction with social advancements and improved living condition [1]. According to the World Health Organization (WHO), approximately 2.5 billion adults worldwide were classified as overweight (body mass index, BMI ≥ 25) in 2022, with 890 million individuals falling into the category of obesity (BMI ≥ 30). This equates to 43% of adults being overweight and 16% living with obesity[2]. In the past twenty years, there has been a significant increase in the prevalence of both general and abdominal obesity among adults of diverse age groups and genders in China[3]. The detrimental impacts of overweight or obesity extend to various bodily functions, leading to the development of numerous comorbidities such as hyperlipidemia, type 2 diabetes, hypertension, and other disorders[4]. Increasing evidence suggests that the initiation and progression of overweight/obesity are closely associated with the homeostatic regulation of overall metabolism. Especially, microbial metabolites, as the primary way that gut microbes interact with hosts, are important for maintaining the metabolic homeostasis of the organism[5]. According to the existing evidence, identified microbial metabolic signatures have been associated with obesity[6], such as short-chain fatty acids (SCFAs)[7],trimethylamine N-oxide (TMAO)[8], bile acids[9],linoleic acid[10] and branched chain fatty acids (BCFAs). The diverse metabolic products generated by the gut microbiota permeate into the intestinal mucosa, subtly influencing intestinal cellular activities. A subset of these metabolites possesses the capability to traverse the intestinal barrier, subsequently disseminating to distant organs through either circulatory routes or neural pathways[11].
Furthermore, overweight/obesity-associated microbial metabolites are regulated by intestinal microbiota to affecting the metabolic health of overweight/obese[12]. Multiple studies have demonstrated a notable disparity in the composition of intestinal bacteria between individuals of normal weight and those with overweight/obesity[13]. The alterations in microbiome associated with overweight/obesity, commonly referred to as ‘dysbiosis,’ are characterized by a deficiency in beneficial functions or the prevalence of detrimental microbial activity[14]. Lower proportion of Bifidobacteria, Faecalibacterium prausnitzii, Akkermansia muciniphila, F. prausnitzii, which are considered as the benefic strains, were found in overweight and obesity[15,16].
Probiotics are acknowledged as a valuable supplementary approach for regulating microbiota dysbiosis, consisting of viable microorganisms that can successfully modify imbalanced microbiota to enhance key obesity-related factors when given in precise doses of probiotic strains[17]. And, previous studies have identified Bifidobacterium, Lactobacillus, Enterococcus, Streptococcus, and Saccharomyces, as well as E. coli Nissle 1917 and the yeast Saccharomyces boulardii, as the primary probiotic genera [18,19]. Especially,Bifidobacterium species are frequently utilized as functional foods and dietary supplements, purportedly aiding in the prevention of dysmetabolic diseases through enhancement of metabolic function[20]. Furthermore, Bifidobacterium is one of the main producers of SCFAs in the intestinal flora to participate in regulating energy homeostasis[21]. As one kind of Bifidobacterium, the commercially available probiotic Bifidobacterium brevis BBr60 (BBr60) has demonstrated anti-inflammatory and antioxidant properties. Given its potential for addressing metabolic disorders, further research is needed to elucidate the mechanisms by which Bifidobacterium brevis BBr60 intervenes in metabolic disturbances associated with overweight/obesity, as well as to explore its potential application in clinical settings.
Hence, we carried out a double-blinded, randomized placebo-controlled trial to investigate the comparative efficacy and mechanism of Bifidobacterium brevis BBr60 in modulating disrupted metabolic profiles and gut microbiota in overweight/obese young individuals in China. Seventy overweight/obese adult subjects were randomly assigned to receive either Bifidobacterium brevis BBr60 or a placebo for a duration of 12 weeks, along with brief dietary counseling emphasizing a total daily energy intake of 1800 kcal. Serum and fecal samples were collected to assess glucose and lipid levels, gut metabolomics, and microflora following treatment in order to evaluate the efficacy and mechanism of regulating metabolic disturbances in overweight/obese individuals. Ultimately, this study provides a scientific rationale for the dietary and clinical utilization of Bifidobacterium brevis BBr60 in the prevention and management of overweight/obesity.
2. Materials and Methods
2.1. Ethics and Informed Consent
This study utilized a randomized, double-blind, placebo-controlled trial design conducted at the School of Food and Bioengineering, Henan University of Science and Technology from March 2023 to June 2024. The protocol adhered to the World Medical Association Declaration of Helsinki and was approved by the Ethics Committee of the First Affiliated Hospital of Henan University of Science and Technology (NCT06305650).
2.2. Study Design and Population
All participants in the study provided informed consent and met the designated inclusion criteria, which included being between the ages of 19 and 45, having a BMI of 28 kg/m2 or higher, and agreeing to participate after being informed about the study procedures and signing a written consent form. The exclusion criteria encompassed various factors that could potentially impact the validity of the results, such as short-term use of objects with similar functions to the test, recent administration of antibiotics, laxatives, or dietary supplements, history of alcohol or drug abuse, presence of serious medical illnesses (e.g., kidney or liver disease, neurological disorders), and pregnancy or lactation without the use of contraception. Studies were required to meet all of these criteria in order to be considered eligible for inclusion in the analysis.
2.3. Sample Size and Randomization
A total of 78 participants were screened, with 75 starting treatment and 65 completing the study. Eligible participants were randomly allocated to either the placebo group or the probiotics group using a random number table as shown in Figure 1. The trial adhered strictly to the initial protocol without modifications. Participants in the placebo group were administered daily maltodextrin (3 g), while those in the BBr60 group consumed daily BBr60 (1×1010 CFU, once a day, provided by Wecare Probiotics Co., Ltd.) for a duration of 12 weeks. All participants were instructed to reduce their daily energy intake by 1800 kcal and attended a nutrition information course covering topics such as the risks and causes of overweight and obesity, weight loss principles, dietary recommendations, and rest. Patients, study staff, clinical research associates, and statisticians were blinded to the randomization and study products. The residual products and medicines, and empty packing boxes were recovered.
2.4. Primary Outcome and Secondary Outcomes
The primary outcome of this study was the change in body mass index (BMI) from the baseline to 12 weeks after beginning the treatment. The key secondary outcomes were changes in waist hip ratio (WHR), and body fat rate (BFR) from the baseline to 12 weeks. Other secondary outcomes were the changes in serum biochemical indexes, fecal metabolism and gut microbiota from the baseline to 12 weeks.
2.5. Assessment of Body Composition
All subjects were weighed in light clothing without shoes. Body mass index (BMI), waist hip ratio (WHR), and body fat rate (BFR) were measured with body composition analyzer (InBody270, InBody, Tokyo, Japan).
2.6. Blood Sample Collection and Biochemical Measurements
Collection and biochemical measurements of blood were carried out at the hospital of Henan university of science and technology by clinical standard assays right after fasting blood sampling at baseline, and after 12 weeks intervention. Blood samples were collected following an overnight fast of at least 10 hours for clinical chemistry analyses. Serum samples were subsequently centrifuged and stored at -80°C until analysis. The following parameters were measured using automatic biochemical analyzer (KHB ZY-1280, Shanghai Kehua Bio-engineering Corporation, Shanghai, China): fasting blood glucose (FBG), alanine aminotransferase (ALT), aspartate aminotransferase (AST), alkaline phosphatase (ALP), total protein (TP), albumin (ALB), globular Proteins (ALP), total bilirubin (TB), blood urea nitrogen (BUN), and lipid profile including total triglycerides (TG), total cholesterol (TC), high-density lipoprotein cholesterol (HDL-C), and low-density lipoprotein cholesterol (LDL-C).
2.7. Serum and Fecal Metabolomic Analysis
Fasted serum samples were collected pre- and post-treatment, then centrifuged at 3000 g for 15 minutes. For metabolomics analysis, 100 μL of serum was mixed with 400 μL of extraction solution (MeOH: ACN, 1:1 (v/v)) containing deuterated internal standards. The mixture was vortexed for 30 s, sonicated for 10 min at 4℃, and incubated for 1 h at -40℃ to precipitate proteins. Finally, the samples were centrifuged at 12000 rpm (13800 g) for 15 min at 4℃. For analysis, the supernatant was transferred into a fresh vial of glass.
Fecal samples were mixed with beads and extraction solution, containing deuterated internal standards, and vortexed for 30 seconds.
Quality control (QC) samples were prepared by mixing an equal amount of supernatant from each sample. LC-MS/MS analysis was performed using an UHPLC system coupled to an Orbitrap MS. The mobile phase consisted of 25 mmol/L ammonium acetate and ammonia hydroxide in water. The auto-sampler was set at 4℃ with an injection volume of 2 μL. The Orbitrap Exploris 120 mass spectrometer was used in IDA mode with Xcalibur software, continuously evaluating the full scan MS spectrum. The acquisition software monitors the full scan MS spectrum continuously in this mode. ESI source conditions include sheath gas flow rate of 50 Arb, Aux gas flow rate of 15 Arb, capillary temperature of 320℃, full MS resolution of 60000, MS/MS resolution of 15000, collision energy settings of 20/30/40 SNCE, and spray voltage of 3.8 kV (positive) or -3.4 kV (negative). The raw data underwent conversion to the mzXML format through the utilization of ProteoWizard and were subsequently analyzed using a custom program developed in R and reliant on XCMS for peak detection, extraction, alignment, and integration. Metabolite identification was facilitated through the use of the R package and BiotreeDB (V3.0).
2.8. Gut Microbiota Analysis
DNA was extracted from fecal using CTAB following the manufacturer’s instructions. The reagent effectively extracted DNA from trace amounts of sample, particularly bacteria. Blank samples were prepared using nuclear-free water. The eluted DNA was stored at -80°C until PCR measurement. Primers were tagged with specific barcodes for each sample and sequencing universal primers. PCR was conducted in a 25 uL reaction mixture with 25 ng of template DNA, PCR Premix, primers, and water. The prokaryotic 16S fragments were amplified using specific PCR conditions, including denaturation, annealing, and extension steps. The PCR products were verified using agarose gel electrophoresis. During DNA extraction, ultrapure water was used as a negative control to prevent false-positive PCR results. PCR products were purified using AMPure XT beads and quantified with Qubit. Amplicon pools were prepared for sequencing and their size and quantity were assessed using Agilent 2100 Bioanalyzer and Library Quantification Kit for Illumina.
Samples were sequenced on an Illumina NovaSeq platform PE250 following manufacturer’s instructions. Paired-end reads were assigned to samples based on their unique barcode, merged using FLASH, and quality filtered using fqtrim (v0.94). Chimeric sequences were removed using Vsearch software (v2.3.4). Feature table and sequence were obtained after dereplication using DADA2. Alpha and beta diversity were calculated by randomly normalizing sequences. Feature abundance was normalized using relative abundance of each sample according to the SILVA classifier. Alpha diversity was analyzed using 5 indices in QIIME2, while beta diversity was calculated and visualized using R package. Blast was used for sequence alignment, and the feature sequences were annotated with SILVA database for each representative sequence. Other diagrams were implemented using the R package (v3.5.2). Blast was used for sequence alignment and SILVA database was used for annotation. Other diagrams were created using R package v3.5.2.
2.9. Safety Monitoring
Safety outcomes will be evaluated by monitoring vital signs and body weight at each visit. Participants will undergo blood routine tests, liver and kidney function tests, urine routine tests, and physical examinations at baseline and in the 12th week of treatment. Safety outcomes will be assessed through the evaluation of physical examinations, vital signs, hematological analyses, and reported adverse events or serious adverse events.
2.10. Statistical Analysis
Statistical analyses were performed using SPSS version 22.0 (SPSS, Chicago, IL, US) and Graphad Prism 8.0 (Graphpad Software, USA). Quantitative data following are reported as mean ± standard deviation (SD), with statistical significance assessed using a two-tailed T test for data conforming to a normal distribution. And, data not conforming to a normal distribution is analyzed using nonparametric tests such as the Mann-Whitney U test for between-group comparisons and the Wilcoxon signed-rank test for within-group comparisons. Categorical variables are presented as numbers (%) and compared using the chi-square test. Principal component analysis (PCA), orthogonal partial least-squared discriminant analysis (OPLS-DA) were conducted by the SIMCA software package (V18.0.1, Sartorius Stedim Data Analytics AB, Umea, Sweden). Pathway analysis was performed by databases including KEGG (http://www. genome. jp/kegg/) and MetaboAnalyst (http://www. metaboanalyst. ca/). Correlations between two variables were assessed through Spearman correlation analyses, with statistical significance defined as p < 0.05 and an alpha level (α) established at 0.05. The obtained p-values validate that the discrepancies observed between groups were not a result of random variation, thus bolstering the reliability of our results.
3. Results
3.1. Baseline Characteristics
Table 1 presents the baseline characteristics of the participants, including sex distribution, mean age, baseline BMI values, and waist-to-hip ratio (WHR). The distribution of sex did not show a significant difference between the BBr60 group, where 72.7% (n=24) were female, and the placebo group, where 59.4% (n=19) were female. The mean age was 27.88 ± 8.65 years in the BBr60 group and 30.38 ± 8.45 years in the placebo group. The baseline BMI values were 30.80 ± 3.21 kg/m2 and 31.96 ± 2.95 kg/m2, respectively. The WHR also did not exhibit a significant difference between groups, with values of 0.99 ± 0.05% in the BBr60 group and 1.01 ± 0.05% in the placebo group. There were no significant differences observed in lipid levels, renal and hepatic function indicators, anthropometric and body composition data, and biochemical parameters between the two groups.
3.2. Primary Outcome
Efficacy of BBr60 on Weight and BMI
The study measured the change in weight as a secondary outcome by comparing baseline weight to weight at 12 weeks. The BBr60 group experienced a statistically significant decrease in weight of 4.67 (p <0.0001) compared to a decrease of 2.82 in the placebo group (p = 0.0006), and the difference of body weight between 0 week-12 week in BBr60 group was significantly higher than that of placebo group (p = 0.047). Additionally, after 12 weeks, the BMI value in the BBr60 group was 86.19, which was 4.55 lower than the placebo group (p = 0.1137) as shown in Table 2 and Figure 2A.
The primary outcome of this study was the change in BMI, which was calculated as the difference in BMI from baseline to 12 weeks. The BMI significantly decreased with the BBr60 intervention (p <0.0001) and also decreased in the placebo group (p = 0.0019). At the end of the 12-week period, the BMI value was 29.32 in the BBr60 group, which was lower than the placebo group (p = 0.056) (Table 2, Figure 2B).
3.3. Secondary Outcomes
Efficacy of BBr60 on BFP and WHR
BFP and WHR are commonly used in the clinical evaluation of body fat distribution[22]. In the current study, BFP and WHR were significantly decreased in BBr60 and placebo groups after a 12-week intervention. Notably, BBr60 intervention presented more obvious effect than placebo, with lower p value (p < 0.0001). And the values of BFP and WHR in BBr60 group were both lower than those of placebo group in 12 weeks(Table 2, Figure 2C, D).
Efficacy of BBr60 on Blood Glucose and Lipid
In the BBr60 group, there was a significant reduction in fasting blood glucose (FBG) from a baseline value of 5.87 (SD 0.45) to 5.26 (SD 0.57) after the 12-week intervention period. Additionally, the FBG levels in the placebo group also showed a significant decrease; however, the mean FBG level in the BBr60 group was significantly lower than that of the placebo group after the 12-week intervention (p = 0.0381).
No clinically meaningful differences between BBr60 and placebo groups after a 12-week intervention were noted in plasma lipids, including TC, TG, HDL-C, HDL-C. Notably, HDL-C and LDL-C were could significantly regulated by BBr60 intervention with p < 0.0001. Furthermore, the level of HDL-C of BBr60 group was higher than that of placebo group and LDL-C of BBr60 group was lower than that of placebo group at week 12.
Efficacy of BBr60 on Liver Function and Renal Function
Liver function (TP, ALB, GLB, ALT, AST) and kidney function (BUN, CRE-E) were measured at week 12, And, the levels of ALB, GLB, ALT, AST were significantly reduced following BBr60 intervention, with levels in the BBr60 group being lower than those in the placebo group at week 12. No significant differences were observed for TP and BUN.
Efficacy of BBr60 on Metabolic Pathway
Alteration of Serum Metabolism after 12 Weeks of BBr60 Intervention
The aforementioned study demonstrated that, under the condition of effective regulation in the placebo group, which may be largely attributed to daily healthy diet recommendations, BBr60 more significantly improved clinic indexes of obesity, especially FBG and weight. Additionally, we explored the regulatory effects of BBr60 on serum metabolism in individuals with overweight or obesity. OPLS-DA was employed to identify metabolites that met the criterion of variable influence on projection (VIP) >1 between the BBr60-before and BBr60-after groups. The samples from BBr60-before and BBr60-after groups were distinctly separated in the OPLS-DA score plot (Figure 3A), with model parameters R2Y= 0.50 and Q2 = -0.89 as shown in Figure 3B. PCA was employed to evaluate the alterations in metabolic profiling between the BBr60-before and BBr60-after groups (Figure 3C). Although no distinct separation was observed, a substantial number of significantly regulated fecal metabolites (p < 0.05, VIP > 1) were identified in the volcano plot (Figure 3D). Specifically, 292 metabolites were significantly upregulated, while 351 serum metabolites were downregulated in the BBr60-before group compared to the BBr60-after group. Detailed information regarding these metabolites is provided in Supplementary Table 1 (Tables S1).
Furthermore, the analysis of 42 significant metabolic pathways (p < 0.05) associated with significant fecal metabolites (VIP > 1, p < 0.05 between BBr60-before and BBr60-after groups) was conducted (see Tables S1, S2, and Figure 3E-F). The top 15 KEGG pathways (p < 0.001) exhibited alterations in response to BBr60 treatment for 12-week intervention (Figure 3E). These pathways include central carbon metabolism in cancer, protein digestion and absorption, biosynthesis of amino acids, glyoxylate and dicarboxylate metabolism, pantothenate and CoA biosynthesis, citrate cycle (TCA cycle), carbon metabolism, valine, leucine and isoleucine biosynthesis, glycine, serine and threonine metabolism, aminoacyl-tRNA biosynthesis, 2-oxocarboxylic acid metabolism, alanine, aspartate and glutamate metabolism, mineral absorption, ABC transporters, beta-Alanine metabolism (Figure 3F). Biosynthesis of amino acids, carbon metabolism, and 2-oxocarboxylic acid metabolism categorized under global and overview maps, exhibited significant up-regulation in 15.69%, 11.76 and 12.75% of differential metabolites, respectively, when influenced by BBr60. Furthermore, the significantly regulated pathways associated with valine, leucine and isoleucine biosynthesis, glycine, serine and threonine metabolism as well as alanine, aspartate and glutamate metabolism were all categorized under amino acid metabolism with 5.88%, 7.84% and 5.88% of differential metabolites, respectively. The pathways of glyoxylate and dicarboxylate metabolism, as well as citrate cycle (TCA cycle), exhibited 9.8% and 5.88% differential metabolites among all metabolites in the pathway, which were significantly regulated by BBr60. These pathways are categorized under carbohydrate metabolism. In serum metabolism, 643 metabolites regulated by BBr60 were associated with 42 significant metabolic pathways. Notably, the pathways of glyoxylate and dicarboxylate metabolism, TCA cycle, valine, leucine and isoleucine biosynthesis, glycine, serine and threonine metabolism as well as alanine, aspartate and glutamate metabolism warrant further investigation.
Alteration of Fecal Metabolism after 12 Weeks of BBr60 Intervention
To analyze the underlying mechanisms of BBr60’s effective intervention on overweight or obesity, fecal metabolism was examined before and after BBr60 treatment. Orthogonal Partial Least Squares Discriminant Analysis (OPLS-DA) was utilized to identify metabolites with a variable influence on projection (VIP) score greater than 1, both prior to and following BBr60 intervention. The OPLS-DA score plot (Figure 4A) demonstrated a clear separation between the BBr60-before and BBr60-after groups, with model parameters R2Y = 0.75 and Q2 = -0.71, as depicted in Figure 4B. PCA was utilized to assess the changes in metabolic profiles between the BBr60-before and BBr60-after groups (Figure 4C). Despite the absence of a clear separation, 514 fecal metabolites exhibited significant regulation (p < 0.05, VIP > 1), as illustrated in the volcano plot (Figure 4D) and detailed in Table S3. Of these, 225 metabolites were significantly upregulated, whereas 289 metabolites were downregulated following the 12-week BBr60 intervention.
Additionally, a comprehensive analysis of the metabolic pathways associated with these significantly altered fecal metabolites (VIP > 1, p < 0.05 between BBr60-before and BBr60-after groups) was performed (Tables S3, S4, and Figure 5). A total of 52 metabolic pathways exhibited alterations in response to BBr60 treatment compared to the pre-treatment condition (Table S4). Among these pathways, 25 demonstrated statistical significance (p < 0.05). Notably, 15 pathways, which were down-regulated by BBr60 over a 12-week period, met a more stringent significance threshold (p < 0.01) and are illustrated in Figure 5A. These pathways encompass ABC transporters, mineral absorption, central carbon metabolism in cancer, biosynthesis of amino acids, valine, leucine and isoleucine biosynthesis, nucleotide metabolism, protein digestion and absorption, glycine, serine and threonine metabolism, aminoacyl-tRNA biosynthesis, 2-oxocarboxylic acid metabolism, linoleic acid metabolism, pyrimidine metabolism, cysteine and methionine metabolism, valine, leucine and isoleucine degradation, glycerophospholipid metabolism (Figure 5B). Biosynthesis of amino acids, nucleotide metabolism, and 2-oxocarboxylic acid metabolism, which are categorized under global and overview maps, exhibited significant down-regulation in 15.38%, 10.77%, and 13.85% of differential metabolites, respectively, when influenced by BBr60. Additionally, four metabolic pathways were classified under amino acid metabolism, including valine, leucine, and isoleucine biosynthesis; glycine, serine, and threonine metabolism; cysteine and methionine metabolism; and valine, leucine, and isoleucine degradation. Furthermore, two metabolic pathways, including linoleic acid metabolism and glycerophospholipid metabolism, were categorized as lipid metabolism.
In fecal metabolism, 514 fecal metabolites exhibited significant regulation (p < 0.05, VIP > 1) and related to 25 significant metabolic pathways including amino acid metabolism and lipid metabolism on BBr60-before vs. BBr60-after groups.

Table 4.
Changes in liver and renal function in the BBr60 and Placebo Groups after 12 Weeks.
| Variables | BBr60 (n=33) | Placebo (n=32) | p-Value | ||||
|---|---|---|---|---|---|---|---|
| Before (0 week) |
After (12 week) |
p-Value | Before (0 week) |
After (12 week) |
p-Value | ||
| TP, g/L | 74.45±9.70 | 72.85±3.70 | 0.601 | 72.47±3.46 | 73.19±3.39 | 0.404 | 0.701 |
| ALB, g/L | 47.76±3.93 | 50.09±3.18 | 0.0003 | 47.41±3.07 | 49.31±2.01 | 0.0058 | 0.2037 |
| GLB, g/L | 26.70±7.28 | 22.76±3.85 | 0.0089 | 25.06±3.05 | 23.94±2.96 | 0.0499 | 0.170 |
| ALT, IU/L | 39.27±26.55 | 26.03±17.13 | <0.0001 | 41.53±21.84 | 27.22±15.69 | <0.0001 | 0.631 |
| AST, IU/L | 61.18±58.70 | 38.27±17.75 | 0.0002 | 49.56±17.88 | 39.59±17.49 | 0.0089 | 0.636 |
| BUN, mg/dL | 4.48±1.24 | 4.19±0.83 | 0.219 | 4.65±1.26 | 4.73±1.24 | 0.418 | 0.093 |
Abbreviations: ALT, Alanine aminotransferase; AST, Aspartate aminotransferase; TP, Total Protein; ALB, Albumin; GLB, Globular Proteins; BUN, Blood urea nitrogen.
Efficacy of BBr60 on Gut Microbiota
The richness, diversity, and evenness of the gut microbiota in the BBr60 placebo groups at the 12th week were assessed using α and β diversity analyses (Figure 6A-J). The α diversity was significantly increased following a 12-week BBr60 intervention, as indicated by metrics such as goods_coverage, pielou_e, and the Shannon index. Additionally, a clear separation of samples from the BBr60-before and BBr60-after groups was observed based on jaccard_distance and unweighted_unifrac_distance. At the phylum level, Firmicutes and Actinobacteria were the predominant species in both the placebo-after and BBr60-after groups, followed by Proteobacteria and Bacteroidota (Figure 7A and Table S5). As shown in the Figure 7B and Table S6, the dominant bacterial groups in each group are mainly Bifidobacterium, Streptococcus, Agathobacter, Escherichia-Shigella, Faecalibacterium, Megamonas, Erysipelotrichaceae_UCG-003, Dialister at the genus level. The relative abundance of Escherichia-Shigella, Dialister, Phascolarctobacterium, Klebsiella, Bacteroides, Veillonella in the BBr60-after group was higher than that of BBr60-before the group (Figure 7C). The 12-week intervention with BBr60 resulted in a statistically significant decrease in the relative abundance of the class Bacilli (p<0.05). These results indicate that BBr60 can effectively increase the abundance of Escherichia-Shigella, Dialister, Phascolarctobacterium, Klebsiella, Bacteroides, and Veillonella, while concomitantly reducing the abundance of the class Bacilli. These microbial species may serve as key indicators in the regulation of obesity and could potentially aid in differentiating between the gut microbiota of obese and normal-weight individuals.
3.4. Correlation Analysis
The potential relationships of gut flora, metabolic profiling and clinic index were conducted by Spearman correlation analysis (Figure 8A-C). Escherichia-Shigella, Dialister, Phascolarctobacterium, Klebsiella, Bacteroides, and Veillonella, the vital genera significantly regulated by BBr60, showed the significantly certain metabolites (Figure 8A-B). Specially, Dialister and Bacteroides exhibited obvious correlation with most of serum metabolites. They both showed the negative relationship with the serum metabolites, including butanoic acid, isovaleric acid, oxalic acid, 3-hydroxypyruvic acid, malonic acid, o-acetylserine, valine, beta-alanine, ketoleucine, as well as, exhibited positive relationship with glutamate, serine, asparagine (Figure 8A). And those metabolites were involved in 14 pathways of 2-oxocarboxylic acid metabolism, ABC transporters, alanine, aspartate and glutamate metabolism, aminoacyl-tRNA biosynthesis, beta-alanine metabolism, biosynthesis of amino acids, carbon metabolism, central carbon metabolism in cancer, glyoxylate and dicarboxylate metabolism, mineral absorption, pantothenate and CoA biosynthesis, protein digestion and absorption, valine, leucine and isoleucine biosynthesis, glycine, serine and threonine metabolism. 3 out of 14 were categorized under global and overview maps including biosynthesis of amino acids, 2-oxocarboxylic acid metabolism, and carbon metabolism. Furthermore, 3 out of 11 were categorized under amino acid metabolism including valine, leucine and isoleucine biosynthesis, glycine, serine and threonine metabolism as well as alanine, aspartate and glutamate metabolism. And three pathways of amino acid metabolism were also significantly regulated by BBr60 on fecal metabolism. Besides, among metabolites significantly correlated with Dialister and Bacteroides, 5 amino acids (asparagine, serine, glutamate, ketoleucine, valine), which were associated with three pathways of amino acid metabolism, exhibited significant relationships with the certain clinic indexes of obesity, such as FBG, ALB, LDL-C, HDL-C (Figure 8C).
3.5. Safety and Tolerability
There was no serious adverse event in any of the groups during the study period, suggesting a favorable safety profile for BBr60.
4. Discussion
Obesity is a multifaceted condition with a complex pathogenesis involving socioeconomic, hormonal, and neuronal mechanisms, as well as unhealthy lifestyle choices and genetic and epigenetic factors[23]. The World Health Organization predicts that by 2035, 39% of the global adult population will be affected by obesity[2]. Research suggests that obesity and related metabolic disorders are linked to changes in gut microbiota function and composition, which play a significant role in regulating the body’s energy metabolism[24]. Furthermore, alterations in the gut microbiota composition have been linked to the onset of obesity and its related metabolic conditions[25]. Utilizing probiotics to manipulate the gut microbiome may serve as a potential approach for managing metabolic syndrome and obesity-associated complications, such as dyslipidemia and insulin resistance[26]. Nevertheless, the efficacy of probiotics is contingent upon the specific species and dosages employed, as well as the underlying medical condition[27]. Therefore, the current study investigated the effectiveness of BBr60 (1×1010 CFU, once a day) for 12 weeks on body composition, serum glucose, lipid, liver and kidney functions in overweight or obese adult population with BMI ≥ 28 kg/m2. After a 12-week intervention, BMI was significantly decreased by BBr60 intervention, with levels in the BBr60 group being lower than that in the placebo group at week 12. Similar trends were seen for body weight, BFP, WHR, FBG, HDL-C, LDL-C and liver function indexes (ALB, GLB, ALT, AST) in the BBr60 group.
Bifidobacterium is a typical probiotic with ability of reducing intestinal lipopolysaccharide and fortifying intestinal barrier function, and have been widely used as probiotic preparations for the treatment of intestinal microecological disorders[28,29]. Bifidobacterium brevis BBr60 is one kind of Bifidobacterium, have been commercialized for anti-inflammatory and antioxidant properties. In this study, BBr60 presented the effectiveness on BMI, body weight, BFP, WHR after a 12-week intervention, and scientific evidence also suggests that alterations in the gut microbiota through the use of probiotics may play a role in changes in body weight and composition[30]. Specifically, administering Bifidobacterium to individuals with over-weight or obesity (BMI >24.9 kg/m2) resulted in significant reductions in body fat mass (p = 0.006), body fat percentage (p = 0.02), waist circumference (p < 0.00001), and visceral fat area (p = 0.003)[31]. A meta-analysis of 15 studies on probiotics demonstrated significant changes in body weight and body fat among obese individuals with a BMI exceeding 25 kg/m2, with an average weight loss of 0.6 kg and a BMI reduction of 0.27 kg/m2[32]. The admonition of Bifidobacterium breve B-3 (20 billion CFU/day) for 12 weeks significantly reduced the body fat in pre-obese adults without any adverse effects[33]. In addition, the reducing weight, BMI, BPF and WHR in placebo group, the reason may be associated with the increased awareness of weight loss by dietary recommendations once a week for all the participants during the trial, and similar results were found in many clinical study[34,35].
Several studies have demonstrated that probiotic intervention may be beneficial in the management of obesity, as well as various metabolic abnormalities such as dysglycemia, insulin resistance, and dyslipidemia[36]. Additionally, probiotic supplementation has been shown to improve fasting blood glucose levels, insulin sensitivity, and hyperlipidemia[37]. In the present study, levels of fasting blood glucose and LDL-C were significantly reduced and HDL-C was effectively increased following a 12-week intervention with BBr60. Accordingly, previous studies also presented that Bifidobacteria supplementation (50 × 109 CFU/day) for 12 weeks effectively ameliorating hyperglycemia, dyslipidemia by decreasing serum LDL, TG and glycosylated hemoglobin concentration in type 2 diabetic patients[38]. Administration Bifidobacterium animalis IPLA R1 decreased serum insulin level with no significant variation in FGB and HOMA index in mice of a short-term diet-induced obesity[39].
The liver plays a crucial role in regulating whole-body cholesterol levels, while the kidney is essential for maintaining overall homeostasis[40,41]. Impaired kidney function and liver dysfunction are commonly observed in individuals with severe obesity[42]. Assessment of liver and kidney function serves as a valid measure for evaluating psychological states in obesity. In a study involving overweight and obese adults undergoing a 12-week BBr60 intervention, liver function indicators (TP, ALP, GLB, ALT, AST) and renal function marker (BUN) were examined to assess the impact of BBr60 supplementation. Furthermore, supplementation with BBr60 led to notable decreases in ALB, GLB, ALT, and AST levels. Previous research has shown that supplementation with Bifidobacterium breve resulted in significant reductions in BUN and creatinine levels compared to a placebo group[38]. Additionally, mice that were gavaged with Bifidobacterium pseudolongum exhibited a lower liver-to-body weight ratio and reduced serum levels of ALT, AST, and hepatic triglycerides in cases of non-alcoholic fatty liver disease-associated hepatocellular carcinoma[43]. These findings suggest the efficacy of BBr60 in improving certain clinical indicators in overweight/obese individuals.
The deep mechanism was further analyzed after the effective regulation of BBr60 on clinic indictors of obesity. The metabolic status of the organism was investigated by serum and fecal metabolism as well as gut microbiota, and the potential relationship between metabolic profiles and clinic indexes of obesity. 42 significant metabolic pathways associated with 643 serum metabolites and 25 vital metabolic pathways related to 514 fecal metabolites were obviously regulated by BBr60, and the amino acid metabolism pathways of valine, leucine and isoleucine biosynthesis, glycine, serine and threonine metabolism as well as alanine, aspartate and glutamate metabolism were regulated in serum metabolism and fecal metabolism. And significant associations between metabolites involved in three pathways and predominant genera or clinic indexes (such as LDI-C and HDL-C) of BBr60 intervention were found. Extensive research demonstrates that distinct metabolism of amino acids has long been recognized as a feature of obesity[44,45] and probiotics can improve intestinal flora imbalance and regulate intestinal microbial metabolites[46,47] to affect amino acids metabolism and other metabolic pathways[48] to further alleviating overweight or obesity[49]. The potential mechanism of probiotics on overweight or obesity have been discussed in previous studies[47], more researches have focused on gut inflammation and lipid metabolism[50]. The enhancement of glucolipid metabolism by probiotics may be linked to a reduction in bacterial lipopolysaccharides (LPSs), which are known to induce inflammation and obesity[51,52]. While the role of lipopolysaccharides in the regulation of obesity through amino acid metabolism remains unclear, numerous studies have identified significant associations between specific probiotic strains and particular amino acids[53]. Host glutamate levels could be influenced by B. thetaiotaomicron colonization increases the levels of mRNAs encoding glutamate decarboxylase and glutamate transporter in epithelial cells and the concentration of plasma glutamate reduces by gavage with B. thetaiotaomicron in mice[12,54]. Also, the abundance of B. thetaiotaomicron showed a negative correlation with the circulation of glutamate in previous study[12]. Consistently, our study revealed that the plasma glutamate levels, which are involved in the alanine, aspartate, and glutamate metabolism pathway, exhibited significant associations with dominant bacterial strains (Klebsiella, Bacteroides, and Dialister) following the BBr60 intervention, as well as with clinical indices such as fasting blood glucose (FBG) and albumin (ALB). Additionally, other amino acids, including asparagine, serine, ketoleucine, and valine, which are implicated in pathways of valine, leucine and isoleucine biosynthesis, glycine, serine and threonine metabolism, as well as alanine, aspartate and glutamate metabolism, demonstrated significant correlations with the bacterial strains Dialister and Bacteroides, and with specific clinical indices such as low-density lipoprotein cholesterol (LDL-C). Generally, the effective regulating of FBG, LDL-C, HDL-C and ALB may be attributed to the regulation of BBr60 on vital genus (Klebsiella, Bacteroides, and Dialişter) to affect the vital metabolites associated with the pathways of biosynthesis pathways of valine, leucine, and isoleucine, as well as in glycine, serine, and threonine metabolism and alanine, aspartate, and glutamate metabolism.
5. Conclusions
Collectively, these findings suggest that BBr60 can safely and effectively reduce body weight, BMI, BFP, WHR, FBG, LDL-C, HDL-C, and improve liver function (ALB, GLB, ALT, AST). Furthermore, the potential mechanism underlying the effective regulation of clinical indices may be attributed to BBr60’s influence on key genera (Klebsiella, Bacteroides, and Dialister), which in turn affect critical metabolites associated with the pathways of valine, leucine and isoleucine biosynthesis, glycine, serine and threonine metabolism, as well as alanine, aspartate and glutamate metabolism. Future research should focus on elucidating the deeper mechanisms by which BBr60 affects these key genera and specific metabolites to regulate glucolipid metabolism in murine models.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Funding
Figures of Graphical abstract in the manuscript were drawn in Figdraw. The authors gratefully acknowledge the financial support from Major Science and Technology Special Projects in Henan Province (231100310200);Key R&D Projects in Henan Province (241111314200); Key R&D Projects in Henan Province (221111111400).
Authors’ contributions
ZYB: Investigation, Methodology, Writing - original draft, Software, Formal analysis, Writing - review & editing; YW: Writing - review & editing, Resources, Supervision; DEG: Investigation, Methodology, Writing - original draft, Conceptualization, Data curation; YD: Resources, Supervision; YJP: Investigation, Methodology; SBG: Project administration, Resources, Supervision, Validation, Visualization.
Ethics approval and consent to participate
This study was performed in line with the principles of the Declaration of Helsinki. The study protocol was approved by the First Affiliated Hospital of Henan University of Science and Technology (NCT06305650). All participants provided written informed consent prior to study enrolment.
Competing interests
The authors declare no competing interests.
Data Share Statement
Data described in the manuscript, code book, and analytic code will be made available upon request pending application and approval.
Abbreviations
ALB: Albumin; A/G, Albumin/ Globulin; ALT, Alanine aminotransferase; AST, Aspartate aminotransferase; BBr60, Bifidobacterium brevis BBr60; BFP, Body fat percentage; BMI, Body mass index; BUN, Blood urea nitrogen; CRE, Creatinine; FBG, Fasting blood glucose; GLB, Globular Proteins; HDL-C, high-density lipoprotein cholesterol; LDL-C, low-density lipoprotein cholesterol; OPLS-DA, Orthogonal Partial Least Squares Discriminant Analysis; PCA, Principal component analysis; TB, Total bilirubin; TC, Total cholesterol; TCA cycle, citrate cycle; TG, Triglyceride; TP, Total Protein; UA, Uric acid; WHR, Waist-to-Hip Ratio.
References
- Zhao, Z.; Zhen, S.; Yan, Y.; Liu, N.; Ding, D.; Kong, J. Association of dietary patterns with general and central obesity among Chinese adults: a longitudinal population-based study. BMC Public Heal. 2023, 23, 1–10. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Obesity and Overweight. Available online: http://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight. (accessed on 16 July 2021).
- Chen, Y.; Peng, Q.; Yang, Y.; Zheng, S.; Wang, Y.; Lu, W. The prevalence and increasing trends of overweight, general obesity, and abdominal obesity among Chinese adults: a repeated cross-sectional study. BMC Public Heal. 2019, 19, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Château-Degat, M.-L.; Dewailly, E.; Charbonneau, G.; Laouan-Sidi, E.A.; Tremblay, A.; Egeland, G.M. Obesity risks: towards an emerging Inuit pattern. Int. J. Circumpolar Heal. 2011, 70, 166–177. [Google Scholar] [CrossRef] [PubMed]
- Rezgui, R.; Walia, R.; Sharma, J.; Sidhu, D.; Alshagadali, K.; Chaudhuri, S.R.; Saeed, A.; Dey, P. Chemically Defined Lactobacillus plantarum Cell-Free Metabolites Demonstrate Cytoprotection in HepG2 Cells through Nrf2-Dependent Mechanism. Antioxidants 2023, 12, 930. [Google Scholar] [CrossRef]
- Sowah, S.A.; Milanese, A.; Schübel, R.; Wirbel, J.; Kartal, E.; Johnson, T.S.; Hirche, F.; Grafetstätter, M.; Nonnenmacher, T.; Kirsten, R.; et al. Calorie restriction improves metabolic state independently of gut microbiome composition: a randomized dietary intervention trial. Genome Med. 2022, 14, 1–21. [Google Scholar] [CrossRef]
- Chambers, E.S.; Preston, T.; Frost, G.; Morrison, D.J. Role of Gut Microbiota-Generated Short-Chain Fatty Acids in Metabolic and Cardiovascular Health. Curr. Nutr. Rep. 2018, 7, 198–206. [Google Scholar] [CrossRef]
- Schugar, R.C.; Shih, D.M.; Warrier, M.; Helsley, R.N.; Burrows, A.; Ferguson, D.; Brown, A.L.; Gromovsky, A.D.; Heine, M.; Chatterjee, A.; et al. The TMAO-Producing Enzyme Flavin-Containing Monooxygenase 3 Regulates Obesity and the Beiging of White Adipose Tissue. Cell Rep. 2017, 19, 2451–2461. [Google Scholar] [CrossRef]
- Pang, Y.; Zheng, Y.; Yang, N.; Zan, M.; Zhang, L.; Ding, W. Potential novel biomarkers in small intestine for obesity/obesity resistance revealed by multi-omics analysis. Lipids Heal. Dis. 2022, 21, 1–15. [Google Scholar] [CrossRef]
- Yamazaki, T.; Yamada, S.; Ohshio, K.; Sugamata, M.; Morita, Y. Lactobacillus paracasei KW3110 Prevents Inflammatory-Stress-Induced Mitochondrial Dysfunction in Mouse Macrophages. Int. J. Mol. Sci. 2022, 23, 1443. [Google Scholar] [CrossRef]
- Ezzine, C.; Loison, L.; Montbrion, N.; Bôle-Feysot, C.; Déchelotte, P.; Coëffier, M.; Ribet, D. Fatty acids produced by the gut microbiota dampen host inflammatory responses by modulating intestinal SUMOylation. Gut Microbes 2022, 14, 2108280. [Google Scholar] [CrossRef]
- Liu, R.; Hong, J.; Xu, X.; Feng, Q.; Zhang, D.; Gu, Y.; Shi, J.; Zhao, S.; Liu, W.; Wang, X.; et al. Gut microbiome and serum metabolome alterations in obesity and after weight-loss intervention. Nat. Med. 2017, 23, 859–868. [Google Scholar] [CrossRef] [PubMed]
- Ley, R.E., et al., Microbial ecology: human gut microbes associated with obesity. Nature 2006. 444(7122): p. 1022-3.
- Sze, M.A.; Schloss, P.D. Looking for a Signal in the Noise: Revisiting Obesity and the Microbiome. mBio 2016, 7, e01018-16. [Google Scholar] [CrossRef] [PubMed]
- Santacruz, A.; Collado, M.C.; García-Valdés, L.; Segura, M.T.; Martín-Lagos, J.A.; Anjos, T.; Martí-Romero, M.; Lopez, R.M.; Florido, J.; Campoy, C.; et al. Gut microbiota composition is associated with body weight, weight gain and biochemical parameters in pregnant women. Br. J. Nutr. 2010, 104, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Kaźmierczak-Siedlecka, K., et al., Next-generation probiotics - do they open new therapeutic strategies for cancer patients? Gut Microbes 2022. 14(1): p. 2035659.
- Gomaa, E.Z. Human gut microbiota/microbiome in health and diseases: a review. Antonie van Leeuwenhoek 2020, 113, 2019–2040. [Google Scholar] [CrossRef] [PubMed]
- Suez, J.; Zmora, N.; Segal, E.; Elinav, E. The pros, cons, and many unknowns of probiotics. Nat. Med. 2019, 25, 716–729. [Google Scholar] [CrossRef]
- McFarland, L.V.; Evans, C.T.; Goldstein, E.J.C. Strain-Specificity and Disease-Specificity of Probiotic Efficacy: A Systematic Review and Meta-Analysis. Front. Med. (Lausanne) 2018, 5, 124. [Google Scholar] [CrossRef]
- Ben Othman, M.; Sakamoto, K. Effect of inactivated Bifidobacterium longum intake on obese diabetes model mice (TSOD). Food Res. Int. 2020, 129, 108792. [Google Scholar] [CrossRef]
- Liu, N.; Yang, C.; Liang, X.; Cao, K.; Xie, J.; Luo, Q.; Luo, H. Mesoporous silica nanoparticle-encapsulated Bifidobacterium attenuates brain Aβ burden and improves olfactory dysfunction of APP/PS1 mice by nasal delivery. J. Nanobiotechnology 2022, 20, 1–17. [Google Scholar] [CrossRef]
- Zhou, Y.-Y.; Qiu, H.-M.; Yang, Y.; Han, Y.-Y. Analysis of risk factors for carotid intima-media thickness in patients with type 2 diabetes mellitus in Western China assessed by logistic regression combined with a decision tree model. Diabetol. Metab. Syndr. 2020, 12, 1–13. [Google Scholar] [CrossRef]
- Hruby, A.; Hu, F.B. The Epidemiology of Obesity: A Big Picture. PharmacoEconomics 2014, 33, 673–689. [Google Scholar] [CrossRef]
- Chen, B.; Bai, Y.; Tong, F.; Yan, J.; Zhang, R.; Zhong, Y.; Tan, H.; Ma, X. Glycoursodeoxycholic acid regulates bile acids level and alters gut microbiota and glycolipid metabolism to attenuate diabetes. Gut Microbes 2023, 15, 2192155. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.-J.; Lin, C.-S.; Lu, C.-C.; Martel, J.; Ko, Y.-F.; Ojcius, D.M.; Tseng, S.-F.; Wu, T.-R.; Chen, Y.-Y.M.; Young, J.D.; et al. Ganoderma lucidum reduces obesity in mice by modulating the composition of the gut microbiota. Nat. Commun. 2015, 6, 7489. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.Y.; Shin, M.J.; Youn, G.S.; Yoon, S.J.; Choi, Y.R.; Kim, H.S.; Gupta, H.; Han, S.H.; Kim, B.K.; Lee, D.Y.; et al. Lactobacillus attenuates progression of nonalcoholic fatty liver disease by lowering cholesterol and steatosis. Clin. Mol. Hepatol. 2021, 27, 110–124. [Google Scholar] [CrossRef] [PubMed]
- Sharifi-Rad, J.; Rodrigues, C.F.; Stojanović-Radić, Z.; Dimitrijević, M.; Aleksić, A.; Neffe-Skocińska, K.; Zielińska, D.; Kołożyn-Krajewska, D.; Salehi, B.; Prabu, S.M.; et al. Probiotics: Versatile Bioactive Components in Promoting Human Health. Medicina 2020, 56, 433. [Google Scholar] [CrossRef]
- Gong, H.; Gao, H.; Ren, Q.; He, J. The abundance of bifidobacterium in relation to visceral obesity and serum uric acid. Sci. Rep. 2022, 12, 1–7. [Google Scholar] [CrossRef]
- Engevik, M.A.; Luk, B.; Chang-Graham, A.L.; Hall, A.; Herrmann, B.; Ruan, W.; Endres, B.T.; Shi, Z.; Garey, K.W.; Hyser, J.M.; et al. Bifidobacterium dentium Fortifies the Intestinal Mucus Layer via Autophagy and Calcium Signaling Pathways. mBio 2019, 10. [Google Scholar] [CrossRef]
- Rustanti, N.; Murdiati, A.; Juffrie, M.; Rahayu, E.S. Effect of Probiotic Lactobacillus plantarum Dad-13 on Metabolic Profiles and Gut Microbiota in Type 2 Diabetic Women: A Randomized Double-Blind Controlled Trial. Microorganisms 2022, 10, 1806. [Google Scholar] [CrossRef]
- Riveros, N.F.H.; García-Corredor, L.; Martínez-Solare, M.; González-Clavijo, A. Effect of Bifidobacterium Intake on Body Weight and Body Fat in Overweight and Obese Adult Subjects: A Systematic Review and Meta-Analysis. J. Am. Nutr. Assoc. 2024, 1–13. [Google Scholar] [CrossRef]
- Borgeraas, H., et al., Effects of probiotics on body weight, body mass index, fat mass and fat percentage in subjects with overweight or obesity: a systematic review and meta-analysis of randomized controlled trials. Obes Rev 2018. 19(2): p. 219-232.
- Minami, J.; Iwabuchi, N.; Tanaka, M.; Yamauchi, K.; Xiao, J.-Z.; Abe, F.; Sakane, N. Effects of Bifidobacterium breve B-3 on body fat reductions in pre-obese adults: a randomized, double-blind, placebo-controlled trial. Biosci. Microbiota, Food Health 2018, 37, 67–75. [Google Scholar] [CrossRef]
- Dixon, A.E.; Subramanian, M.; DeSarno, M.; Black, K.; Lane, L.; Holguin, F. A pilot randomized controlled trial of pioglitazone for the treatment of poorly controlled asthma in obesity. Respir. Res. 2015, 16, 143. [Google Scholar] [CrossRef]
- Sung, H.K., et al., Body Fat Reduction Effect of Bifidobacterium breve B-3: A Randomized, Double-Blind, Placebo Comparative Clinical Trial. Nutrients 2023. 15(1): p. 28.
- Agius, R.; Pace, N.P.; Fava, S. Reduced leukocyte mitochondrial copy number in metabolic syndrome and metabolically healthy obesity. Front. Endocrinol. 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Tiderencel, K.A.; Hutcheon, D.A.; Ziegler, J. Probiotics for the treatment of type 2 diabetes: A review of randomized controlled trials. Diabetes/Metabolism Res. Rev. 2019, 36, e3213. [Google Scholar] [CrossRef] [PubMed]
- Chaiyasut, C.; Sivamaruthi, B.S.; Lailerd, N.; Sirilun, S.; Thangaleela, S.; Khongtan, S.; Bharathi, M.; Kesika, P.; Saelee, M.; Choeisoongnern, T.; et al. Influence of Bifidobacterium breve on the Glycaemic Control, Lipid Profile and Microbiome of Type 2 Diabetic Subjects: A Preliminary Randomized Clinical Trial. Pharmaceuticals 2023, 16, 695. [Google Scholar] [CrossRef] [PubMed]
- Salazar, N.; Neyrinck, A.M.; Bindels, L.B.; Druart, C.; Ruas-Madiedo, P.; Cani, P.D.; Reyes-Gavilán, C.G.d.L.; Delzenne, N.M. Functional Effects of EPS-Producing Bifidobacterium Administration on Energy Metabolic Alterations of Diet-Induced Obese Mice. Front. Microbiol. 2019, 10, 1809. [Google Scholar] [CrossRef]
- Sun, H.; Liu, Q.; Hu, H.; Jiang, Y.; Shao, W.; Wang, Q.; Jiang, Z.; Gu, A. Berberine ameliorates blockade of autophagic flux in the liver by regulating cholesterol metabolism and inhibiting COX2-prostaglandin synthesis. Cell Death Dis. 2018, 9, 824. [Google Scholar] [CrossRef]
- Lyu, M.; Su, C.-C.; Miyagi, M.; Yu, E.W. Simultaneous solving high-resolution structures of various enzymes from human kidney microsomes. Life Sci. Alliance 2022, 6, e202201580. [Google Scholar] [CrossRef]
- Pi-Sunyer, F.X., Health implications of obesity. The American Journal of Clinical Nutrition 1991. 53(6): p. 1595S-1603S.
- Song, Q.; Zhang, X.; Liu, W.; Wei, H.; Liang, W.; Zhou, Y.; Ding, Y.; Ji, F.; Cheung, A.H.-K.; Wong, N.; et al. Bifidobacterium pseudolongum-generated acetate suppresses non-alcoholic fatty liver disease-associated hepatocellular carcinoma. J. Hepatol. 2023, 79, 1352–1365. [Google Scholar] [CrossRef]
- White, P.J.; Lapworth, A.L.; An, J.; Wang, L.; McGarrah, R.W.; Stevens, R.D.; Ilkayeva, O.; George, T.; Muehlbauer, M.J.; Bain, J.R.; et al. Branched-chain amino acid restriction in Zucker-fatty rats improves muscle insulin sensitivity by enhancing efficiency of fatty acid oxidation and acyl-glycine export. Mol. Metab. 2016, 5, 538–551. [Google Scholar] [CrossRef]
- Colom-Pellicer, M.; Rodríguez, R.M.; Soliz-Rueda, J.R.; de Assis, L.V.M.; Navarro-Masip, .; Quesada-Vázquez, S.; Escoté, X.; Oster, H.; Mulero, M.; Aragonès, G. Proanthocyanidins Restore the Metabolic Diurnal Rhythm of Subcutaneous White Adipose Tissue According to Time-Of-Day Consumption. Nutrients 2022, 14, 2246. [CrossRef]
- Guo, M., et al., Lactobacillus rhamnosus GG ameliorates osteoporosis in ovariectomized rats by regulating the Th17/Treg balance and gut microbiota structure. Gut Microbes 2023. 15(1): p. 2190304.
- He, H. , et al., Arabinogalactan, Bifidobacterium longum, and Faecalibacterium prausnitzii improve insulin resistance in high-fat diet-induced C57BL/6J mice. eFood 2022. 3(1-2): p. e1.
- Desbonnet, L.; Garrett, L.; Clarke, G.; Bienenstock, J.; Dinan, T.G. The probiotic Bifidobacteria infantis: An assessment of potential antidepressant properties in the rat. J. Psychiatr. Res. 2008, 43, 164–174. [Google Scholar] [CrossRef]
- Vallianou, N.; Stratigou, T.; Christodoulatos, G.S.; Tsigalou, C.; Dalamaga, M. Probiotics, Prebiotics, Synbiotics, Postbiotics, and Obesity: Current Evidence, Controversies, and Perspectives. Curr. Obes. Rep. 2020, 9, 179–192. [Google Scholar] [CrossRef] [PubMed]
- Salazar, N.; Neyrinck, A.M.; Bindels, L.B.; Druart, C.; Ruas-Madiedo, P.; Cani, P.D.; Reyes-Gavilán, C.G.d.L.; Delzenne, N.M. Functional Effects of EPS-Producing Bifidobacterium Administration on Energy Metabolic Alterations of Diet-Induced Obese Mice. Front. Microbiol. 2019, 10, 1809. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z. and V.B. Kraus, Does lipopolysaccharide-mediated inflammation have a role in OA? Nat Rev Rheumatol 2016. 12(2): p. 123-9.
- Jeon, M.S.; Choi, Y.Y.; Mo, S.J.; Ha, J.H.; Lee, Y.S.; Lee, H.U.; Park, S.D.; Shim, J.-J.; Lee, J.-L.; Chung, B.G. Contributions of the microbiome to intestinal inflammation in a gut-on-a-chip. Nano Converg. 2022, 9, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Han, X.; Fang, J.; Jiang, H. Role of dietary amino acids and microbial metabolites in the regulation of pig intestinal health. Anim. Nutr. 2021, 9, 1–6. [Google Scholar] [CrossRef]
- Hooper, L.V.; Wong, M.H.; Thelin, A.; Hansson, L.; Falk, P.G.; Gordon, J.I. Molecular Analysis of Commensal Host-Microbial Relationships in the Intestine. Science 2001, 291, 881–884. [Google Scholar] [CrossRef]
Figure 1.
Flowchart of the study selection.
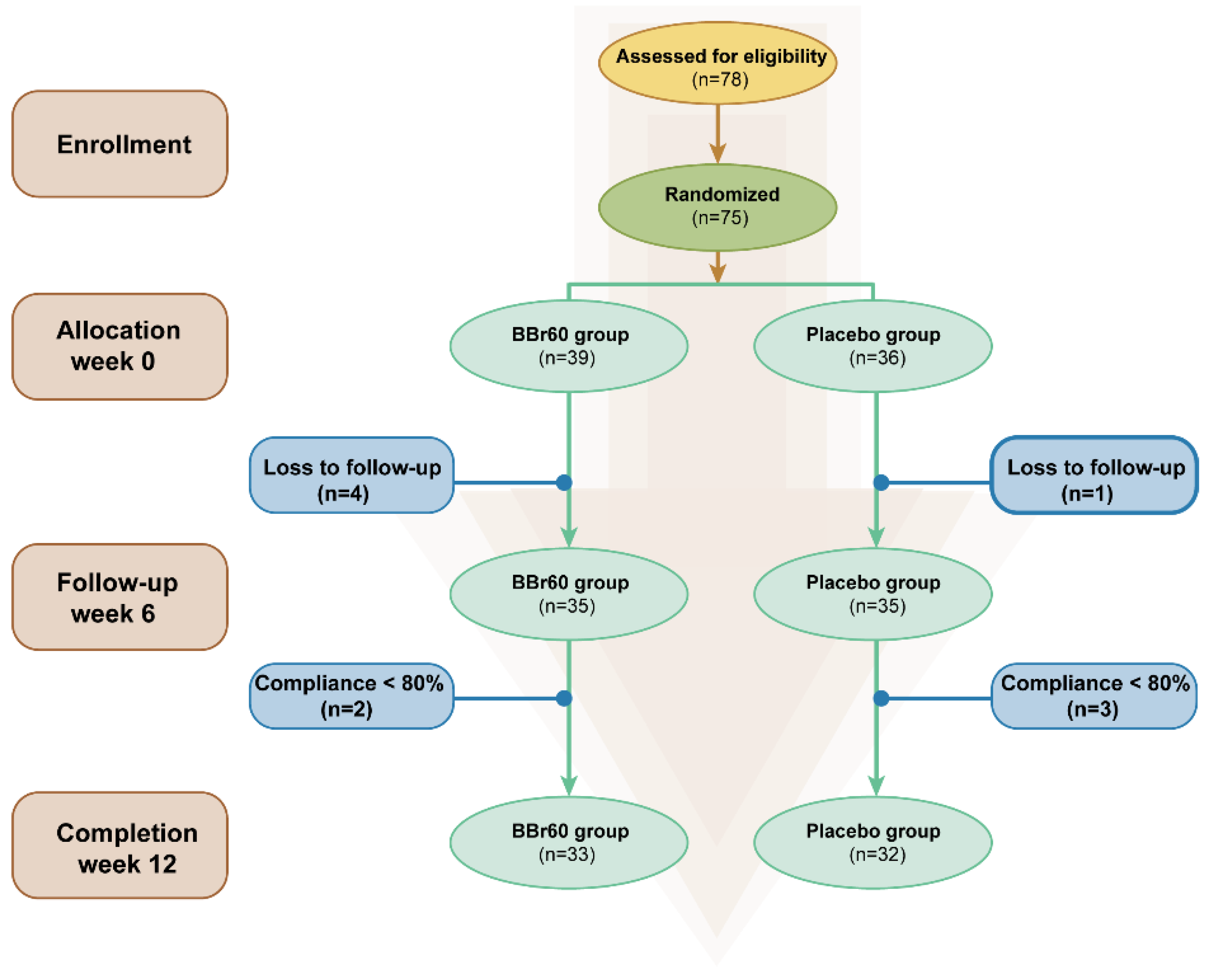
Figure 2.
the effect of BBr60 on body composition in overweight or obese adult population.
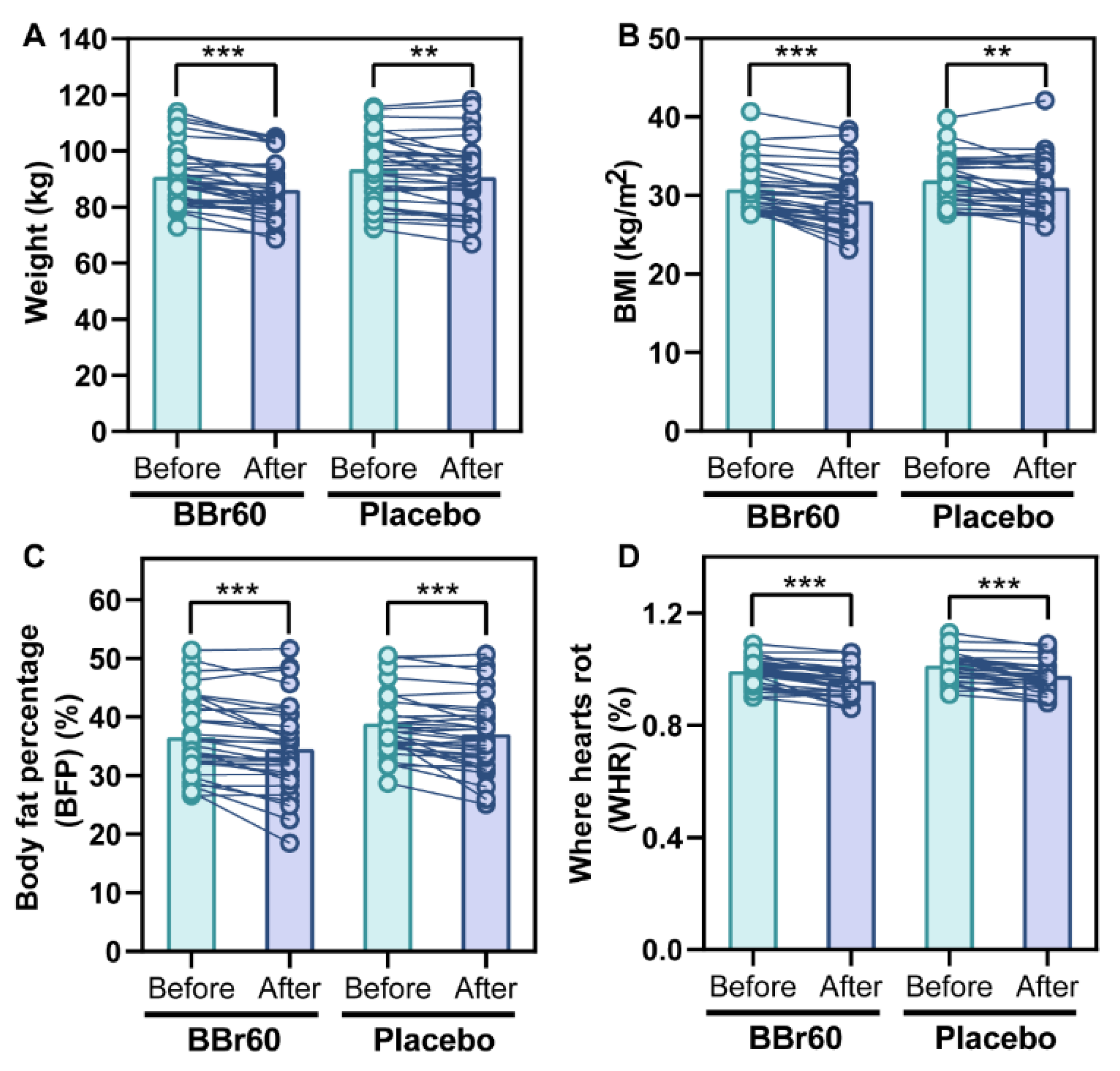
Figure 3.
serum metabolic profile between BBr60-after and BBr60-before groups. (A) The OPLS-DA scores of BBr60 vs. placebo group. (B) The OPLS-DA permutation test in BBr60 vs. placebo group. (C) The PCA scores of BBr60 vs. placebo group. (D) The volcano plot of placebo vs. BBr60 group. Significantly up-regulated metabolites are represented by red points, and significantly down-regulated metabolites and nonsignificant different ones are represented by blue or gray points, respectively. (E) KEGG classification plot for BBr60 vs. placebo group. (F) Differential abundance score plot for BBr60 vs. placebo group.
Figure 3.
serum metabolic profile between BBr60-after and BBr60-before groups. (A) The OPLS-DA scores of BBr60 vs. placebo group. (B) The OPLS-DA permutation test in BBr60 vs. placebo group. (C) The PCA scores of BBr60 vs. placebo group. (D) The volcano plot of placebo vs. BBr60 group. Significantly up-regulated metabolites are represented by red points, and significantly down-regulated metabolites and nonsignificant different ones are represented by blue or gray points, respectively. (E) KEGG classification plot for BBr60 vs. placebo group. (F) Differential abundance score plot for BBr60 vs. placebo group.
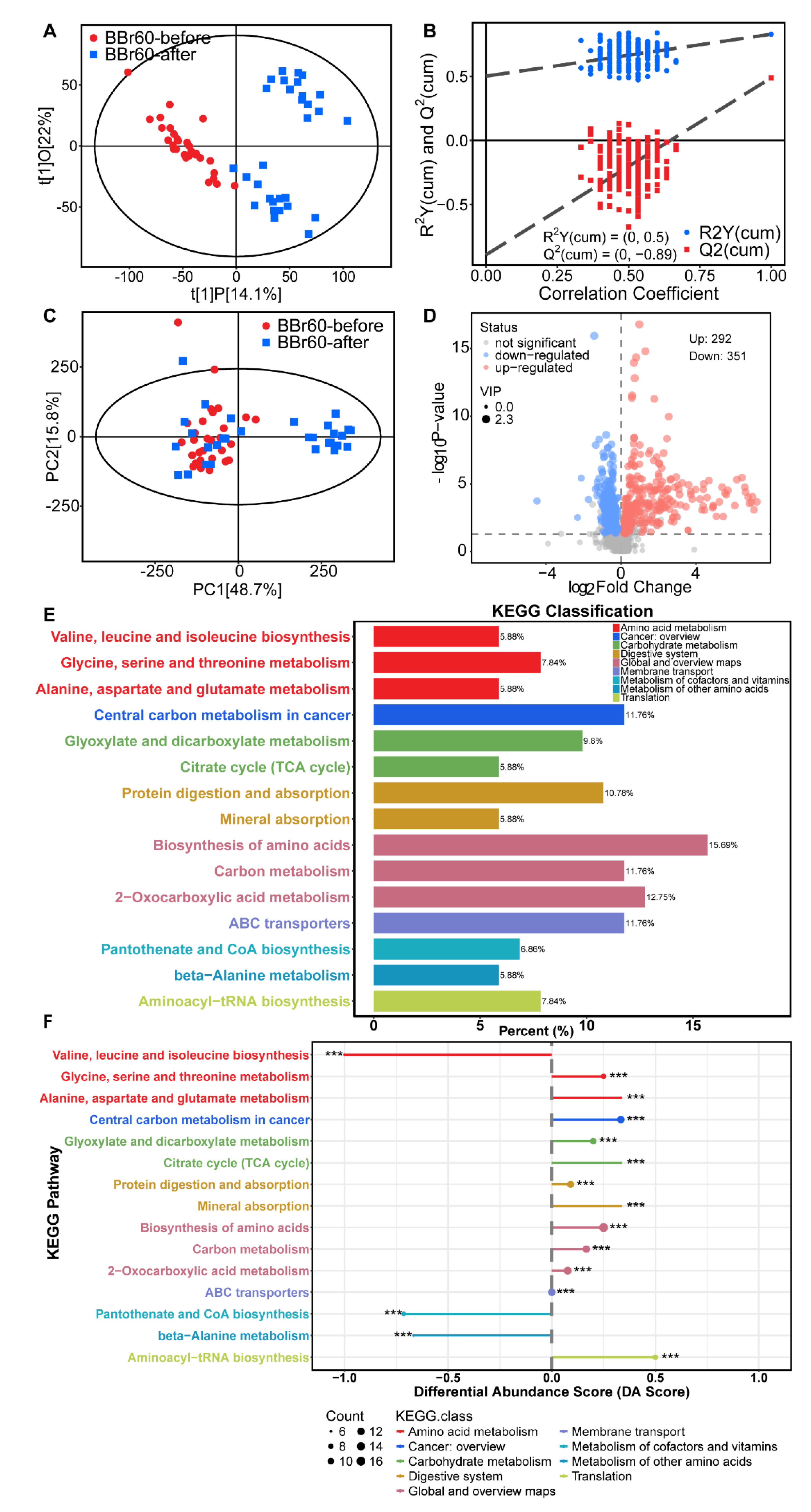
Figure 4.
Fecal metabolic profile between BBr60-after and BBr60-before groups. (A) The OPLS-DA scores of BBr60 vs. placebo group. (B) The OPLS-DA permutation test in BBr60 vs. placebo group. (C) The PCA scores of BBr60 vs. placebo group. (D) The volcano plot of placebo vs. BBr60 group. Significantly up-regulated metabolites are represented by red points, and significantly down-regulated metabolites and nonsignificant different ones are represented by blue or gray points, respectively.
Figure 4.
Fecal metabolic profile between BBr60-after and BBr60-before groups. (A) The OPLS-DA scores of BBr60 vs. placebo group. (B) The OPLS-DA permutation test in BBr60 vs. placebo group. (C) The PCA scores of BBr60 vs. placebo group. (D) The volcano plot of placebo vs. BBr60 group. Significantly up-regulated metabolites are represented by red points, and significantly down-regulated metabolites and nonsignificant different ones are represented by blue or gray points, respectively.
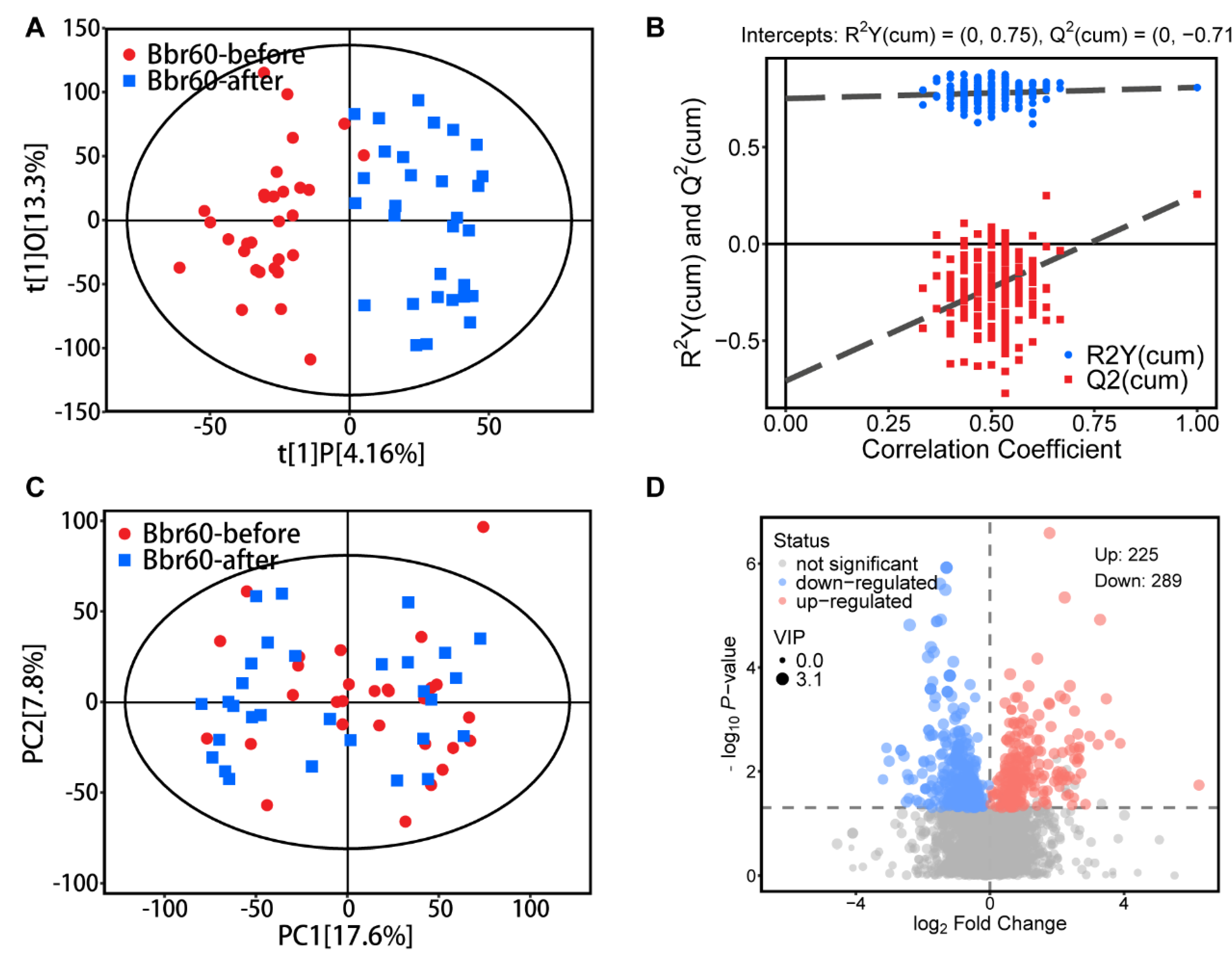
Figure 5.
Fecal metabolic pathways between BBr60-after and BBr60-before groups. (A) KEGG classification plot for BBr60-after vs. BBr60-before groups. (B) Differential abundance score plot for BBr60-after vs. BBr60-before groups.
Figure 5.
Fecal metabolic pathways between BBr60-after and BBr60-before groups. (A) KEGG classification plot for BBr60-after vs. BBr60-before groups. (B) Differential abundance score plot for BBr60-after vs. BBr60-before groups.
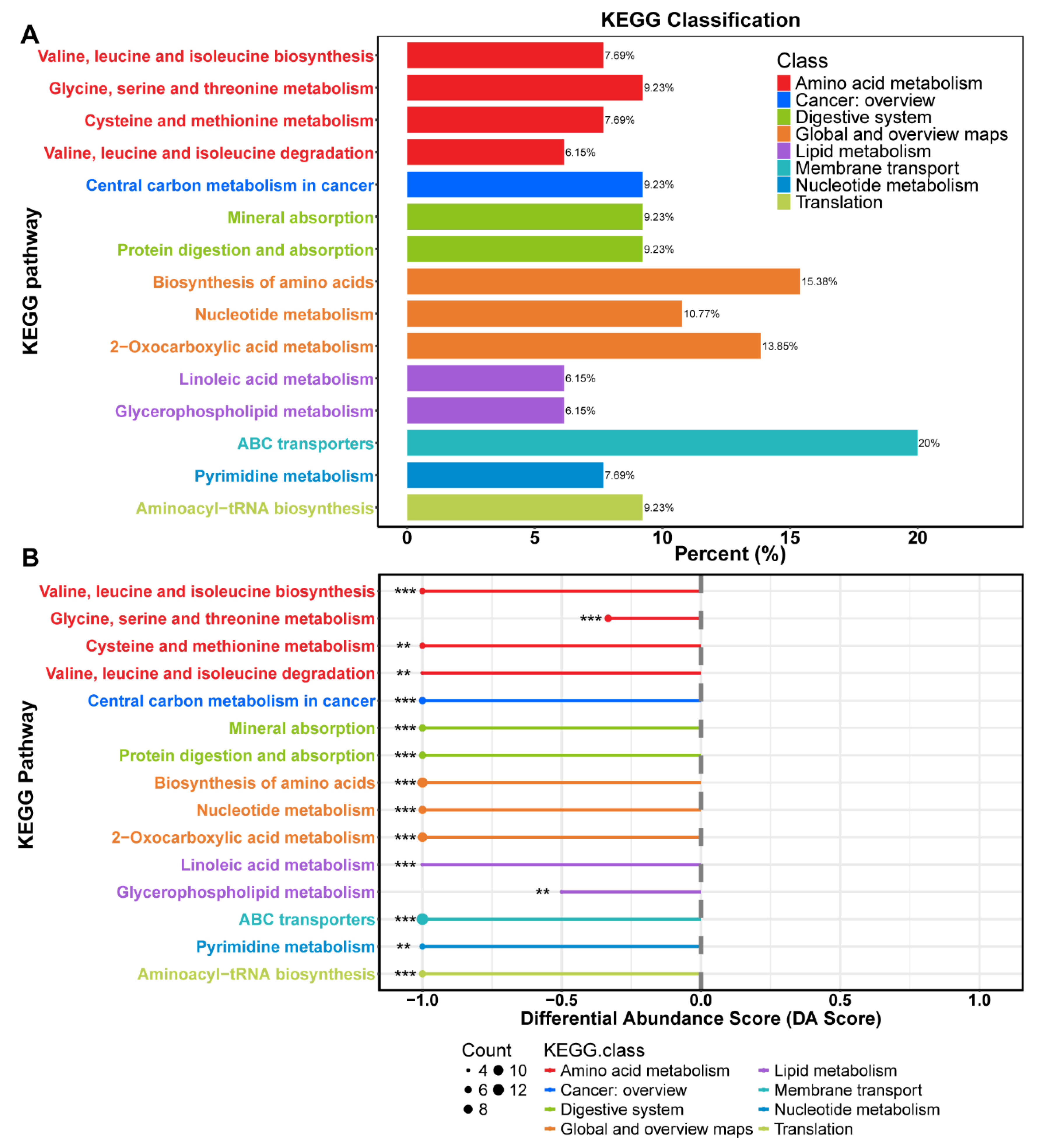
Figure 6.
α and β diversity analysis of BBr60-after and BBr60-before group based on ace (A), chao1 (B), goods_coverage (C), observed_otus (D), pielou-e (E), Shannon (F), bray_curtis_distance (G), jaccard_distance (H), unweighted_unifrac_distance (I) weighted_unifrac (J) between BBr60 and placebo groups in the 12th week.
Figure 6.
α and β diversity analysis of BBr60-after and BBr60-before group based on ace (A), chao1 (B), goods_coverage (C), observed_otus (D), pielou-e (E), Shannon (F), bray_curtis_distance (G), jaccard_distance (H), unweighted_unifrac_distance (I) weighted_unifrac (J) between BBr60 and placebo groups in the 12th week.
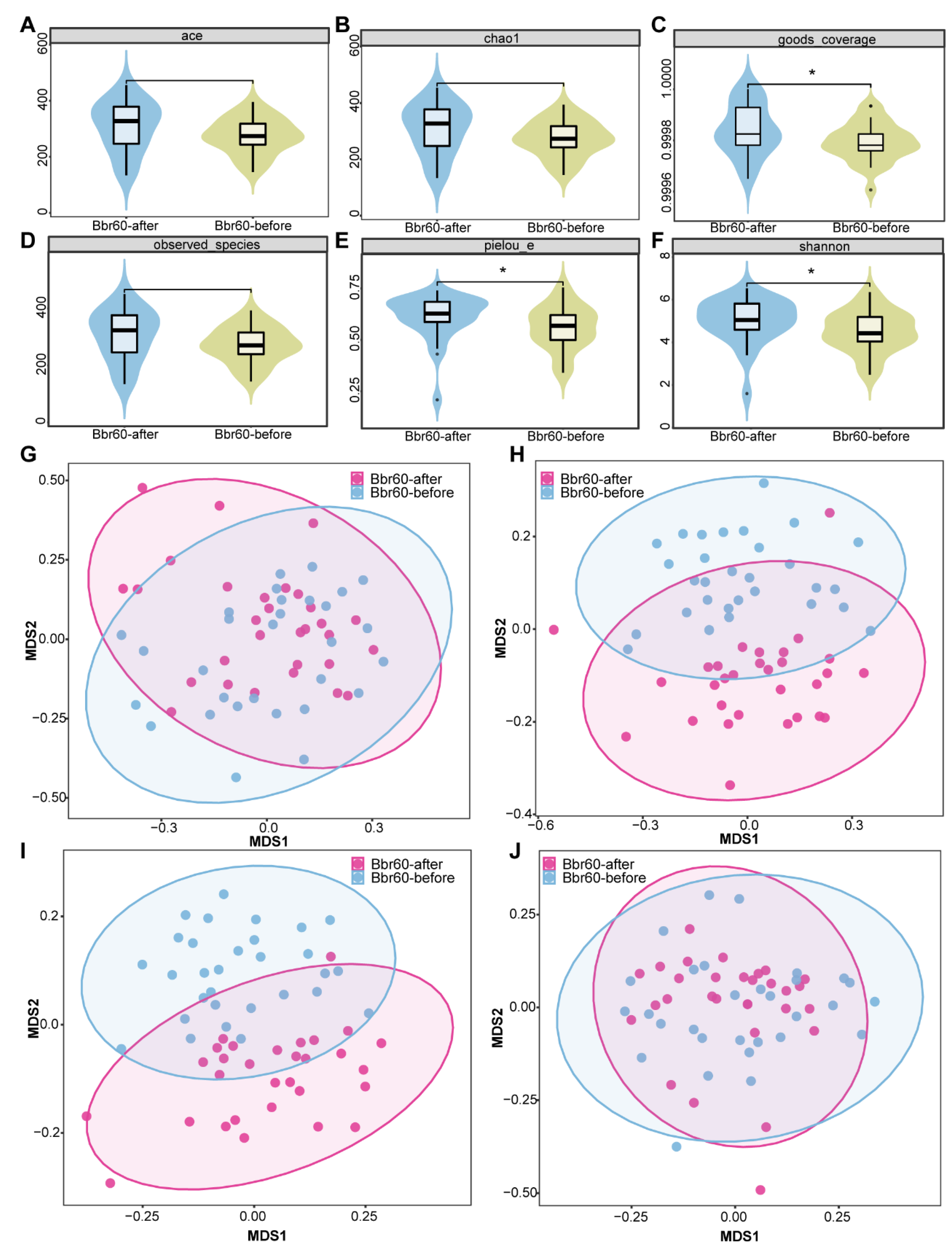
Figure 7.
Bacterial compositions of BBr60-after and BBr60-before groups. (A) Composition of intestinal microbiota in the three groups at phylum level. (B) Composition of intestinal microbiota in the three groups at genus level. (C) Histogram of linear discriminant analysis (LDA) value distribution of intestinal microflora.
Figure 7.
Bacterial compositions of BBr60-after and BBr60-before groups. (A) Composition of intestinal microbiota in the three groups at phylum level. (B) Composition of intestinal microbiota in the three groups at genus level. (C) Histogram of linear discriminant analysis (LDA) value distribution of intestinal microflora.

Figure 8.
correlation analysis of vital metabolites, intestinal bacteria and clinic indexes before and after BBr60 intervention in overweight or obesity. (A) correlation analysis between serum metabolites associated with the top 15 changed metabolic pathways and with the top 30 intestinal bacteria before and after BBr60 intervention in overweight or obesity. The R values are represented by gradient colors, where blue and green cells indicate positive and negative correlations, respectively; *p < 0.05 and **p < 0.01. (B) correlation analysis between serum metabolites associated with the top 15 changed metabolic pathways and with the top 30 intestinal bacteria before and after BBr60 intervention in overweight or obesity. The R values are represented by gradient colors, where blue and red cells indicate positive and negative correlations, respectively; *p < 0.05 and **p < 0.01. (C) correlation analysis between serum metabolites associated with the top 15 changed metabolic pathways and clinic indexes in overweight or obesity. The R values are represented by gradient colors, where blue and red cells indicate positive and negative correlations, respectively; *p < 0.05 and **p < 0.01. Abbreviations: BMI, body mass index; BFP, body fat percentage; WHR, Waist-to-Hip Ratio; FBG, Fasting blood glucose; TC, Total cholesterol; TG, Triglyceride; HDL-C, high-density lipoprotein cholesterol; LDL-C, low-density lipoprotein cholesterol; TP, Total Protein; ALB, Albumin; GLB, Globular Proteins; ALT, Alanine aminotransferase; AST, aspartate aminotransferase; BUN, Blood urea nitrogen.
Figure 8.
correlation analysis of vital metabolites, intestinal bacteria and clinic indexes before and after BBr60 intervention in overweight or obesity. (A) correlation analysis between serum metabolites associated with the top 15 changed metabolic pathways and with the top 30 intestinal bacteria before and after BBr60 intervention in overweight or obesity. The R values are represented by gradient colors, where blue and green cells indicate positive and negative correlations, respectively; *p < 0.05 and **p < 0.01. (B) correlation analysis between serum metabolites associated with the top 15 changed metabolic pathways and with the top 30 intestinal bacteria before and after BBr60 intervention in overweight or obesity. The R values are represented by gradient colors, where blue and red cells indicate positive and negative correlations, respectively; *p < 0.05 and **p < 0.01. (C) correlation analysis between serum metabolites associated with the top 15 changed metabolic pathways and clinic indexes in overweight or obesity. The R values are represented by gradient colors, where blue and red cells indicate positive and negative correlations, respectively; *p < 0.05 and **p < 0.01. Abbreviations: BMI, body mass index; BFP, body fat percentage; WHR, Waist-to-Hip Ratio; FBG, Fasting blood glucose; TC, Total cholesterol; TG, Triglyceride; HDL-C, high-density lipoprotein cholesterol; LDL-C, low-density lipoprotein cholesterol; TP, Total Protein; ALB, Albumin; GLB, Globular Proteins; ALT, Alanine aminotransferase; AST, aspartate aminotransferase; BUN, Blood urea nitrogen.
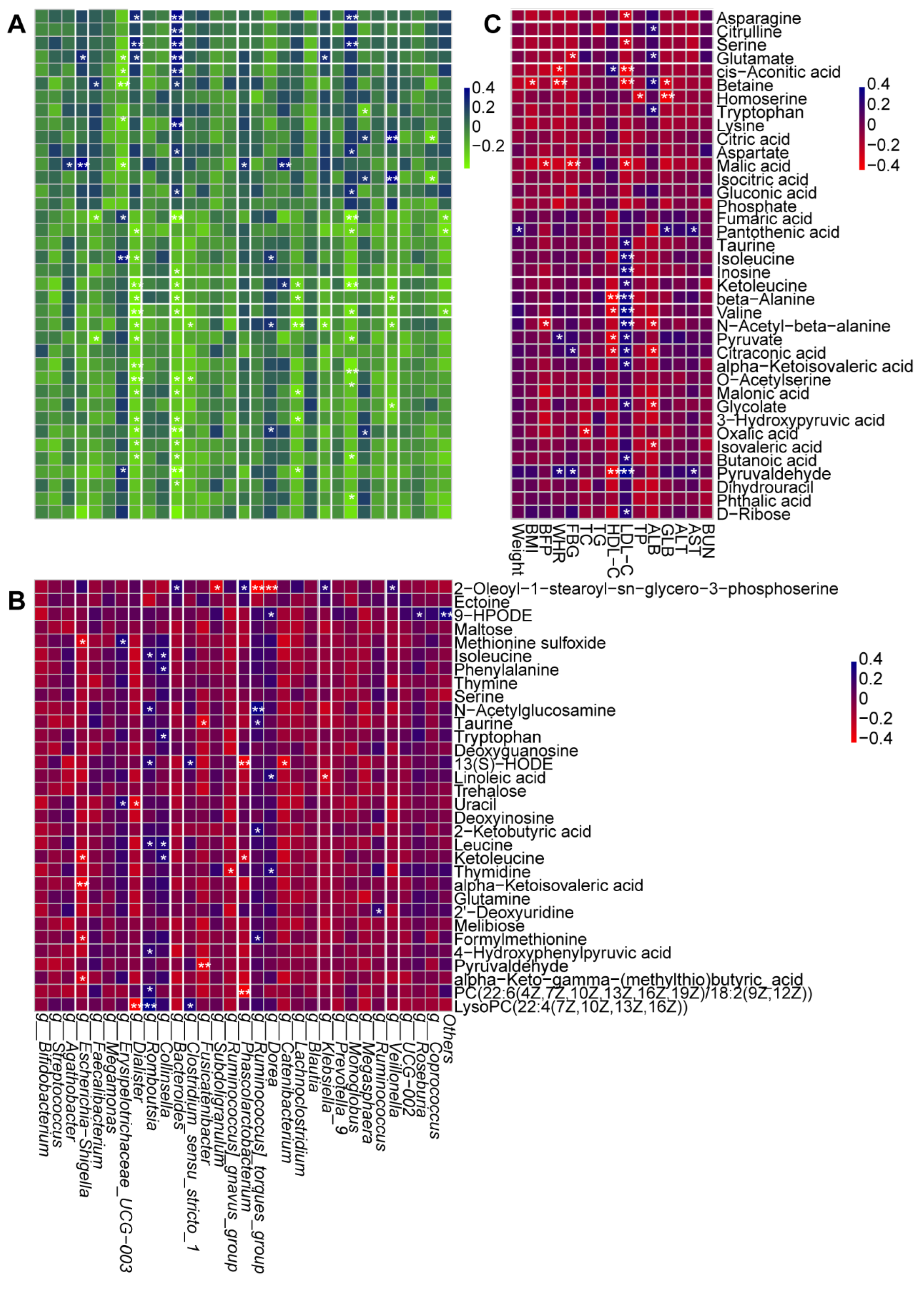
Table 1.
Baseline characteristics in the probiotics and placebo group.
| Project | Unit | BBr60 (33) | Placebo (32) | P value |
|---|---|---|---|---|
| Woman | - | 24 (72.7%) | 19 (59.4%) | 0.255 |
| Man | - | 9 (27.3%) | 13 (40.6%) | |
| Age | year | 27.88±8.65 | 30.38±8.45 | 0.150 |
| Weight | kg | 90.86 ± 10.45 | 93.56 ± 12.04 | 0.251 |
| BMI | kg/m2 | 30.80 ± 3.21 | 31.96 ± 2.95 | 0.068 |
| FP | % | 36.57 ± 6.77 | 38.91 ± 5.69 | 0.203 |
| WHR | % | 0.99 ± 0.05 | 1.01 ± 0.05 | 0.500 |
| FBG | mg/dL | 6.27±0.87 | 5.87±0.45 | 0.120 |
| TC | mg/dL | 4.31±0.90 | 4.82±0.94 | 0.058 |
| TG | mg/dL | 2.13±1.78 | 1.62±0.71 | 0.345 |
| HDL-C | mg/dL | 1.15±0.27 | 1.29±0.31 | 0.484 |
| LDL-C | mg/dL | 2.26±0.63 | 2.62±0.62 | 0.970 |
| ALT | IU/L | 39.27±26.55 | 41.53±21.84 | 0.618 |
| AST | IU/L | 61.18±58.70 | 49.56±17.88 | 0.969 |
| TP | g/L | 74.45±9.70 | 72.47±3.46 | 0.787 |
| ALB | g/L | 47.76±3.93 | 47.41±3.07 | 0.905 |
| GLB | g/L | 26.70±7.28 | 25.06±3.05 | 0.697 |
| A/G | - | 1.90±0.41 | 1.92±0.29 | 0.273 |
| TB | mg/dL | 13.58±6.58 | 17.38±14.83 | 0.453 |
| BUN | mg/dL | 4.48±1.24 | 4.65±1.26 | 0.846 |
| UA | mg/dL | 427.09±76.69 | 433.59±103.49 | 0.069 |
| CRE | mg/dL | 76.48±14.80 | 71.16±15.77 | 0.543 |
Abbreviations: BMI, Body mass index; BFP, Body fat percentage; WHR, Waist-to-Hip Ratio; FBG, Fasting blood glucose; TC, Total cholesterol; TG, Triglyceride; HDL-C, high-density lipoprotein cholesterol; LDL-C, low-density lipoprotein cholesterol; ALT, Alanine aminotransferase; AST, Aspartate aminotransferase; TP, Total Protein; ALB, Albumin; GLB, Globular Proteins; A/G, Albumin/ Globulin; TB, Total bilirubin; BUN, Blood urea nitrogen; UA, Uric acid; CRE, Creatinine.
Table 2.
Changes in Body Composition in the BBr60 and Placebo Groups after 12 Week.
| Variables | BBr60 (n=33) | Placebo (n=32) | p-Value | ||||
|---|---|---|---|---|---|---|---|
| Before (0 week) |
After (12 week) |
p-Value | Before (0 week) |
After (12 week) |
p-Value | ||
| Weight (kg) | 90.86±10.45 | 86.19±9.82 | <0.0001 | 93.56±12.04 | 90.74±12.77 | 0.0006 | 0.114 |
| Weight (12-0 week) | -4.67±4.40 | -2.82±4.17 | 0.047 | ||||
| BMI (kg/m²) | 30.80±3.21 | 29.32±3.63 | <0.0001 | 31.96±2.95 | 31.03±3.49 | 0.0019 | 0.057 |
| BMI (12-0 week) | -1.49±1.37 | -0.93±1.55 | 0.135 | ||||
| BFP (%) | 36.57±6.77 | 34.54±7.50 | <0.0001 | 38.91±5.69 | 37.11±6.70 | 0.0003 | 0.150 |
| BFP (12-0 week) | -2.03±2.54 | -1.80±2.47 | 0.684 | ||||
| WHR (%) | 0.99±0.05 | 0.96±0.04 | <0.0001 | 1.01±0.05 | 0.98±0.05 | <0.0001 | 0.149 |
| WHR (12-0 week) | -0.036±0.03 | -0.038±0.03 | 0.821 | ||||
Abbreviations: BMI, body mass index; BFP, Body fat percentage; WHR, Where hearts rot.
Table 3.
Changes in serum glucose and lipids in the BBr60 and Placebo Groups after 12 Weeks.
| Variables | BBr60 (n=33) | Placebo (n=32) | p-Value | ||||
|---|---|---|---|---|---|---|---|
| Before (0 week) |
After (12 week) |
p-Value | Before (0 week) |
After (12 week) |
p-Value | ||
| FBG, mg/dL | 5.87±0.45 | 5.26±0.57 | <0.0001 | 6.27±0.87 | 5.69±0.86 | <0.0001 | 0.0381 |
| TC, mg/dL | 4.31±0.90 | 4.38±0.75 | 0.618 | 4.82±0.94 | 4.55±0.90 | 0.0058 | 0.4181 |
| TG, mg/dL | 2.13±1.78 | 1.98±1.12 | 0.8566 | 1.62±0.71 | 2.09±1.34 | 0.0984 | 0.9870 |
| HDL-C, mg/dL | 1.15±0.27 | 1.45±0.28 | <0.0001 | 1.29±0.31 | 1.47±0.26 | 0.0071 | 0.7589 |
| LDL-C, mg/dL | 2.26±0.63 | 1.44±0.52 | <0.0001 | 2.62±0.62 | 1.57±0.53 | <0.0001 | 0.3483 |
Abbreviations: FBG, Fasting blood glucose; TC, Total cholesterol; TG, Triglyceride; HDL-C, high-density lipoprotein cholesterol; LDL-C, low-density lipoprotein cholesterol.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



