
Submitted:
11 September 2024
Posted:
12 September 2024
You are already at the latest version
Abstract
Pluripotent stem cells (PSCs) hold many potential research and clinical benefits due to their ability to differentiate into nearly every cell type in the body. They are often used as model systems for early stages of ontogenesis to better understand key developmental pathways, and for drug screening. However, in order to fully realise the potential of PSCs and their translational applications, a deeper understanding of developmental pathways, especially in humans, is required. Several signalling molecules play important roles during development and are required for proper differentiation of PSCs. The concentration and timing of signal activation is important, with perturbations resulting in improper development and/or pathology. Bone morphogenetic protein (BMP) is one such key signalling molecule involved in the specification and differentiation of various cell types and tissues in the human body including tooth and otic development. In this review, we describe the role of BMP signalling and its regulation, the consequences of BMP dysregulation in disease and differentiation, and how PSCs can be used to investigate the effects of BMP modulation during development, mainly focusing on otic development. Finally, we emphasise the unique role of BMP4 in otic specification and how refined knowledge in controlling its regulation could lead to the generation of more robust and reproducible human PSC-derived otic organoids for research and translational applications.
Keywords:
bone morphogenetic proteins
; human pluripotent stem cells
; human cell models
; organoids
; preplacodal ectoderm
; otic lineage
1. Introduction
Many features of early human development can be recapitulated in vitro using pluripotent stem cells (PSCs). Embryonic specification of various domains arising from the different germ layers is achieved by the activation of complex and pleiotropic signalling pathways and inhibitors which interact with one another at critical times during development. Similarly, addition of numerous small molecules that can either activate or inhibit signalling pathways in cultures of PSCs can lead to their specification and subsequent differentiation into any cell types of the three germ layers: endoderm, mesoderm or ectoderm.
Several in vitro protocols have been developed and refined in recent years for the generation of diverse cell types from human PSCs into complex tissue-like structures and organoids, including brain [1,2], cardiac [3,4,5,6], blood vessel [7], retina [8,9,10,11,12], lens [13,14,15,16], inner ear [17,18,19,20,21,22,23,24], etc. However, variations either across labs or between cell lines exist, suggesting the need for further optimisation of differentiation protocols in order to better understand the complex dynamics of signalling molecules involved in specific developmental pathways.
In the case of bone morphogenetic protein (BMP) signalling in particular, endogenous levels of BMP expression and activity within cell lines have previously been shown to affect the concentration of BMP that is required to be added to cultures of PSCs in order to directly differentiation into specific lineages, such as otic lineages [17,18,20,24]. Different levels of BMP and Activin/Nodal signalling are also required for cardiac differentiation of many mouse and human PSC lines [25,26]. A similar observation has been made in human retinal organoids derived from induced PSCs, whereby activation of BMP4 had different effects on different PSC lines, resulting in the generation of different retinal cell types in organoids from each cell line [10]. Differing levels of endogenous BMP4 and BMP4 signalling activity have also been shown to affect differentiation of PSC lines to corneal epithelial-like cells [12]. This suggests that the effects of BMP4 are dependent on the PSC line used and in vitro differentiation protocols that are both robust and efficient require optimisation for each cell line.
In addition, the interplay between BMP signalling and other signalling pathways is complex and likely important for determining cell fate. Indeed, induction of ectodermal placodal fate by exogenous BMP4 in human ES cell lines can be abolished by addition of WNT3a, and rescued when the concentration of BMP4 is increased relative to that of WNT3a [27]. Similarly, Camacho-Aguilar et al. [28] demonstrated the requirement of upregulation of WNT signalling in addition to BMP for conversion of human PSCs from pluripotent state to mesodermal and extraembryonic fates. Importantly, they observed that the timing of BMP exposure was critical for specification of different fates, with long and medium culture period duration of exposure to BMP4 driving extraembryonic and mesodermal fates, respectively, due to activation of endogenous WNT, while short pulses of BMP4 caused cells to remain in the pluripotent state. Indeed, recent studies have indicated that it is not necessarily the concentration of BMP that is important for determining cell fate per se, but rather the integrated signalling level (i.e. concentration and duration) that determines cell fate. Recent, elegant experiments performed by Teague et al. [29] demonstrated that lower levels of BMP signalling for long durations resulted in similar differentiation of hPSCs exposed to higher signalling levels for shorter durations, highlighting that the timing of BMP signalling also needs to be taken into account when designing hPSC in vitro differentiation protocols.
Most BMPs are members of the transforming growth factor β (TGFβ) superfamily of ligands that play critical roles in a multitude of processes during specification and development of almost every tissue and cell type. Originally named for their ability to induce bone and cartilage formation [30], they have since been found to be involved in many aspects of development, such as extraembryonic and mesodermal specification [27,28], dorso-ventral axis formation (reviewed in [31]), ectodermal patterning and subsequent specification of ectodermal fates including neuronal, epidermal, and pre-placodal lineages [27,32,33,34,35]. Several types of BMP (BMP1, BMP2, BMP4, BMP6, BMP7) ligands have additional roles during early development and interact with multiple receptors (BMPR1A, BMPR1B, BMPR2) and mediators (SMAD proteins) for further specification of various cell types.
2. Role and Function of BMP Signalling during Development
2.1. Dorsoventral Patterning and Ectodermal Derivatives
A gradient of BMP signalling is required for the mechanisms of dorsoventral axis determination during gastrulation (reviewed in [31]). In zebrafish, overexpression of BMP can rescue dorsalised mutants [36], while inhibiting BMP by overexpression of either human TAPT1 or zebrafish tapt1a/tapt1b results in dorsalised embryos [37]. Similarly in Xenopus, inhibition of BMP signalling by injection of the BMP antagonist USAG1 into embryos causes them to become more dorsalised [38], supporting a conserved role of BMPs in dorsal-ventral axis patterning during embryonic development. R-spondin 2 (RSPO2) has also been shown to regulate dorsoventral axis formation in Xenopus by antagonising BMP signalling [39,40]; whether R-spondins are similarly involved in mammalian dorsoventral patterning is not known. Later in development, this BMP gradient appears to be inverted in the ectodermal layer, where specification of epidermis occurs in the most dorsal region where BMP concentration is highest, non-neural ectoderm (NNE) (giving rise to pre-placodal ectoderm (PPE) and neural crest in the ventral underlying regions where BMP concentration is lower, and neuronal cell fate where BMP concentration is the lowest [41] (Figure 1).
BMP is required for the expression of NNE genes and PPE competence factors, and previous work with human stem cell lines has demonstrated that this expression requires transient BMP signalling at an optimal concentration for the generation of the desired cell types [27,33,42]. Blocking BMP signalling by addition of Noggin abolished expression of PPE competence genes and induced expression of the neural precursor marker HES5 [33], in agreement with the notion that a reduction of BMP induces neuronal fate. Conversely, removal of Noggin in human PSCs undergoing neural induction using dual-SMAD inhibition resulted in induction of placodal fate at the expense of neuronal fate [14]. Neuroectodermal cells express several transcription factors that regulate their competency to respond to neural inducing signals and inhibit the effects of BMP and WNT signalling (reviewed in [43,44]). In contrast, high BMP levels have been found to induce epithelial differentiation of human ES cells, and addition of Noggin to these cells can increase the population of Nestin-positive neuroectodermal cells in culture at the expensive of keratinocyte differentiation [45,46]. BMP appears to block neural differentiation, possibly through induction of DeltaNp63, a transcriptional target of BMP signalling that can block neuronal development in zebrafish upon its forced expression in this model organism [47].
The PPE in turn gives rise to the sensory placodes of the head region, including the lens, inner ear, olfactory epithelium, etc. (reviewed in [48]. The development of the placodal structures and their subsequent tissues involves BMP signalling and will be discussed in detail below. BMP signalling, in conjunction with WNT, is also important for development of the neural crest [42], and WNT signalling appears to be key for determining whether ectodermal cells become NNE/PPE or neural crest. Neural crest cells localised at dorsolateral position of the neural tube give rise to the neurons and glia of the peripheral nervous system, the enteric nervous system as well as non-neural derivatives. Low concentration of BMP4 in combination with WNT activation is able to generate SOX10-expressing neural crest cells from human PSC cultures [49]. Similarly, treatment of neural crest stem cell-like cells isolated from human skin with BMP2 and an activator of WNT signalling improves their multipotency and differentiation potential to neural crest lineage cells [27]. Conversely, in cultures of human ES cells, BMP signalling in combination with inhibition of WNT signalling resulted in increased expression of SIX1-positive PPE cells and reduced number of cells expressing PAX3 and SOX9 neural crest markers [47].
2.2. Placodal Lineages
Sensory placodes derived from the PPE reuse many of the same signalling molecules and pathways as during earlier development to achieve a diverse range of cell and tissue types, including anterior pituitary gland, lens, olfactory epithelium, trigeminal ganglia, otic epithelium, and epibranchial neurons (reviewed in [32,50]. Studies in Xenopus and zebrafish have shown that once the PPE has been specified, BMP must then be inhibited by dorsally-expressed BMP antagonists in order for placodal development to occur [14,33]. PPE cells generated from human iPSCs and ES cells can be further differentiated to give rise to various placodally-derived cells, including trigeminal ganglia, lens fibres, and anterior pituitary hormone-producing cells [33].
2.2.1. Lens Development
Specification of the PPE into anterior placode is required for development of the eye. Experiments with human ES cell lines have shown that BMP inhibition is required for induction of the anterior placode from PPE, and that addition of BMP4 is subsequently required for induction of lens placode from these cells [14]. Addition of recombinant BMP4 or inhibition of FGF signalling were both also able to induce expression of the lens precursor marker PITX3 in human PSC-derived pre-placodal cells, which could be further differentiated into crystalline-positive cells containing mature lens fibres [16]. Lentoid bodies can also be generated from hiPSCs and ES cells using continuous stimulation with BMP4 and BMP7 followed by WNT activation [13,15,16,51,52,53] and have recently been employed in drug screening for cataract treatments [52].
2.2.2. Olfactory EPITHELIUM DEVELOPMENT
The olfactory epithelium is also derived from the anterior placode. BMPs play important roles in the development of the embryonic olfactory epithelium and bulb, where they are expressed along with their receptors [54]. Studies in mouse and chick embryos demonstrate that expression of SOX2 is required to downregulate BMP4 in the developing olfactory epithelium for subsequent formation of the olfactory pit [53]. Downregulation of BMP also appears to be required for the development of odorant-responsive olfactory sensory neurons derived from hiPSCs [17,18,20,21,22,23,24]. Nevertheless, BMPs continue to be expressed in the olfactory epithelium throughout adulthood, where they are thought to be important for adult neurogenesis in the olfactory system [55].
2.2.3. Inner Ear Development
The posterior placodal region gives rise to the otic-epibranchial progenitor domain (OEPD), from which both the otic and epibranchial placodes are generated. The otic placode invaginates into the underlying mesenchyme to form the otic vesicle. This involves inhibition of BMP signalling, which is recapitulated in human pluripotent stem cell-derived otic organoids using the BMP inhibitor LDN193189 [56]. Such inhibition of BMP signalling could be mediated by LMO4, which was recently found to negatively regulate BMP2 and BMP4 signalling in the zebrafish inner ear [57].
BMP signalling is also important at later stages of otic development. In chick otic vesicles dissected from E3.5-4 embryos, addition of recombinant BMP4 reduced the number of hair cells due to decreased proliferation of otic progenitor cells and increased cell death, while addition of the BMP inhibitor Noggin increased the number of sensory hair cells [58]. Similarly, treatment of chick organotypic cultures with BMP4 during hair cell destruction prevented regeneration of hair cells from supporting cells, while Noggin was able to increase the number of regenerated hair cells [59]. In contrast, another study using chick otocyst cultures reported that blocking BMP signalling reduced generation of hair cells and supporting cells, and that exogenous BMP4 treatment increased the number of hair cells by down-regulation of PAX2 in proliferating sensory epithelial progenitor cells [18,22]. It has been proposed that differences in the concentrations of BMP4 might be responsible for these discrepancies between studies, as the concentration of BMP4 is also found to affect patterning of sensory and nonsensory tissue in the mouse cochlea, with intermediate levels of BMP signalling required to increase the number of sensory hair cells [22]. Similar experiments have not yet been performed in stem cell-derived otic organoids to investigate whether modulation of BMP signalling could alter the number of hair cells (or indeed other otic cell types) produced within these 3D-cell structures.
2.2.4. Epibranchial Placodes
The epibranchial placodes derived from the posterior placode give rise to sensory neurons for ganglia associated with facial, glossopharyngeal and vagal nerves. While OEPD is routinely generated during production of otic organoids and generation of epibranchial-like neurons has been reported in these cultures [18], there are currently no known established models for specific and directed differentiation of epibranchial neurons from human pluripotent stem cells. Interestingly, development of epibranchial-like neurons (and other off-target neurons including neural crest) appears to occur earlier than otic neurons in these cell culture systems [60]. Treatment of stem cell aggregates with FGF, the TGFβ inhibitor SB431542, and the pan-BMP inhibitor LDN193189 was found to be sufficient for the generation of cells expressing posterior placodal markers including PAX8, SOX2, TFAP2A, ECAD, and NCAD, but not the otic marker PAX2 [14], suggesting it may be possible to generate epibranchial neurons separately from otic cells. Moreover, these cells could mature into BRN3A/POU4F1 and TUJ1-positive sensory-like neurons with morphology more similar to that of epibranchial neurons than inner ear ganglia neurons. More directed differentiation and maturation of these neurons has not been investigated, although BMP signalling could be involved. Recent experiments in mice have found that blocking BMP signalling using LDN193189 strongly reduced the numbers of neuroblasts in epibranchial placode 1 and moderately in epibranchial placode 3 [14], suggesting a differential requirement of BMP signalling in neurogenesis in the epibranchial placodes.
2.2.5. Trigeminal Neurons
The trigeminal ganglia are derived from the intermediate placode and contains neurons responsible for transmitting sensory information such as pain and temperature from the face. Trigeminal sensory neurons have been generated from hiPSCs by initial activation of BMP signalling. In one protocol, trigeminal fate was subsequently induced by maintaining cells in N2 medium supplemented with ascorbic acid and BDNF [61], while another protocol used CHIR to activate WNT signalling followed by maturation in neurobasal medium supplemented with NGF, BDNF, and GDNF [62,63]. Engraftment of hiPSC-derived trigeminal ganglia into chick and mouse have shown their survival and ability to establish axonal projections to their target regions [64].
2.3. Tooth Development
Teeth are another ectodermally-derived tissue, and their development requires reciprocal interactions between the epithelium and mesenchyme [65,66]. BMPs in particular are involved and have been shown to interact with other signalling pathways such as SHH [65,67] and WNT [66,67] for tooth development. Experiments in mice at embryonic days E14 and E15 have confirmed the expression of BMP2 in the oral epithelium, and of BMP4, BMP6, and BMP7 in both the epithelium and mesenchyme [65]. Uterine sensitization associated gene-1 (USAG1) is an antagonist of BMP signalling which is also expressed in the epithelium and mesenchyme during tooth formation [68]. Mice lacking USAG1 have an increased number of teeth (supernumerary teeth) which is due to enhanced BMP signalling [69], suggesting a role of BMP in regulating tooth number. Indeed, topical administration of BMP7 can result in partial supernumerary incisor formation in mouse dental explant cultures [70]. Modulation of BMP signalling has also been used to recover tooth development in mice [71]. By using antibodies to block USAG1 in a mouse model of tooth agenesis, Murashima-Suginami and colleagues were able to induce tooth formation in these mice.
Human ES cells have been used to generate oral ectoderm and dental epithelium following a differentiation protocol with increasing concentration of BMP4 [72]. These cells could be mixed with cultures of mouse dental mesenchyme, and when transplanted into murine hosts were capable of forming tooth-like structures in vivo. Recently developed in vitro protocols have enabled the rapid generation of dental epithelial cells in just over one week from hiPSCs by simultaneously inhibiting BMP signalling and activating SHH signalling to generate oral ectoderm from NNE, followed by activation of BMP and SHH pathways and inhibition of WNT signalling [3]. It is not clear why induction of Pitx1-expressing oral epithelium required low concentration of BMP in one protocol and inhibition of BMP in the other, although differences in endogenous BMP signalling and activity between the cell lines used in these studies might be involved.
2.4. Cardiac Development
BMPs act with other signalling pathways, including WNT, Nodal, and FGF, to induce early mesoderm (reviewed in [5]). Specification of later mesodermal fates, such as cardiac, requires additional BMP signalling. BMP2 and BMP4 are involved in cardiomyogenesis, with exogeneous application of either BMP2 or BMP4 proving sufficient to induce ectopic cardiomyocyte differentiation in chick embryos [73,74]. Experiments performed in precardiac spheroids generated from PSCs found that the specification of two separate populations of cardiac progenitor cells (termed first and second heart fields) requires BMP signalling, but that cells of the first heart field are specified via the BMP/SMAD pathway, while cells of the second heart field are specified through a SMAD-independent BMP/WNT pathway [75,76,77,78]. Moreover, blocking BMP signalling abolished the specification of both populations of cardiac progenitor cells, highlighting the importance of BMP in early cardiac development. Nevertheless, modulation of WNT signalling is sufficient to generate heart organoids from PSCs, although addition of BMP4 and Activin A were found to improve the size and vascularisation of organoids [75,77].
Certain cardiac structures, such as the cardiac outflow tract and aortic arch, are derived from neural crest cells (reviewed in [79]. Cardiac neural crest cells have also been proposed to contribute to regeneration of the myocardium following injury in zebrafish and mouse [80,81,82]. In mice, cKit-positive cardiac neural crest cells possess full cardiomyogenic capacity and give rise to several cardiac cell types, which is dependent on BMP antagonism [83]. Suppression of BMP activity is also involved in fate specficiation of cardiac neural crest cells via Adam19-mediated cleavage of ACVR1 and suppression of the BMP-SOX9 cascade [84]. In contrast, BMP activity is required for delamination of neural crest cells from the dorsal neural tube [85,86], via cleavage of N-cadherin allowing these cells to migrate [87]. Stem cell therapies based on cardiac neural crest cells derived from hiPSCs could offer a promising therapy for heart repair following disease or injury, but requires further investigation to better understand the processes involved in specification of cardiac neural crest cells as opposed to other types of neural crest cells, and how to differentiate these cells into the different cardiac cell types.
2.5. Bone
The role of BMPs in bone development, homeostasis, and remodelling has been extensively reviewed elsewhere [88]. Exposure of mesenchymal stem cells (MSCs) to BMP2 is able to induce osteogenic differentiation of these cells both in vitro and in vivo and promote bone formation [89]. Hydrogels containing BMP2 mimetics were found to induce bone formation when injected into rats, which was enhanced when these hydrogels are injected in combination with MSCs [90]. As well as BMP2, BMP9 may also be important for bone formation and regeneration. Overexpression of BMP9 in MSCs increased their osteogenic potential and in resulted in increased bone formation and bone mineral density when injected into rats with calvarial bone defects [91]. This BMP9-induced differentiation of MSCs towards osteogenic fate seems to require Notch signalling, as inhibiting Notch prevents BMP9-induced osteogenic differentiation [92]. A recent study revealed that conditioned media from MSCs overexpressing BMP9 also enhanced bone repair of mouse calvarial defects, compared with media from MSCs that did not overexpress BMP9 [93], suggesting the presence of additional trophic factors released from these cells. BMP9 was additionally able to induce osteogenic differentiation in spheroids derived from human gingival stem cells [94,95], indicating that osteogenesis can be induced in several types of stem cells.
More recently, attempts have been made to induce bone formation from iPSCs, due to their greater proliferative and differentiation capabilities over MSCs. Bone formation has successfully been induced in hiPSCs using retinoic acid, which results in activation of BMP and WNT signalling pathways and differentiation of hiPSCs into osteoblast-like and osteocyte-like cells [58,96]. These cells were able to form bone tissue when injected into mice with calvarial defects, and also recapitulated the phenotype of osteogenesis-imperfecta when cultured from patient-derived iPSCs. Undifferentiated muscle-derived hiPSCs loaded onto an osteoconductive scaffold and implanted into mice can induce ectopic bone formation [38,66,67,68]. Analysis of the scaffolds at 15 and 30 days post-implantation revealed the absence of mRNA of human origin, suggesting that the implanted cells were able to induce bone formation via a paracrine communication. Indeed, conditioned media from these cells was able to induce expression of osteogenesis related genes, upregulation of BMP2, BMP4 and BMP6, increased phosphorylation of SMAD 1/5/8, and the appearance of calcium-containing deposits in the extracellular matrix of cultured human MSCs. Further analysis of these undifferentiated hiPSCs in culture revealed higher expression of BMPs relative to expression in fibroblasts, with BMP2 levels being particularly high. It is not clear whether this high expression of BMPs is because these cells were muscle-derived, and whether hiPSCs derived from other cell types would have similarly high expression of BMPs. It also cannot be ruled out that the high BMP levels are a feature of the cell line that was used in the study. Further experiments in additional hiPSC lines derived from cells of different origins would help to clarify this issue.
Unlike bone formation in the rest of the body, the bone and cartilage of craniofacial structures are derived from cranial neural crest cells, a process which relies heavily on BMP signalling (reviewed in [97]). Treatment of human PSCs with BMP4 from day 8 after specification of neural crest induces expression of cranial neural crest markers such as TFAP2A, MSX1, and DLX1 [98]. Increased BMP signalling in cranial neural crest cells causes premature fusion of cranial sutures and deformities in the skull base in mice [99,100,101]. As a result of this difference in embryonic origin, the MSCs found in cranial structures have different characteristics to those of the long bones. For instance, orofacial MSCs and iliac crest MSCs from the same donor have been found to behave differently when cultured in vitro. Orofacial MSCs proliferated more rapidly and had delayed senescence compared with iliac crest MSCs. Moreover, iliac crest MSCs were more responsive to osteogenic and adipogenic inductions than orofacial MSCs [102]. Recently, ectodermal MSCs, derived from human ES cells via a neural crest intermediate, have been compared with adult bone marrow-derived MSCs. They were found to have comparable osteogenic and chondrogenic ability in culture, although ectodermal MSCs had greater proliferation and formed more dense osseous constructs in a rat calvarial defect model [103].
3. Regulation of BMP Signalling during Development
3.1. BMP Signalling Pathways and Downstream Effects on Gene Expression
BMPs act on their receptors which are typically heterotetrameric complexes composed of type I and type II serine/threonine kinase receptors. Upon ligand binding, type II receptors phosphorylate type I receptors, which then activate SMAD1, SMAD5, and SMAD8 (Figure 2). These receptor-regulated SMADs pair with SMAD4 and translocate to the nucleus to influence the transcription of target genes. This signalling affects gene expression linked to cell growth, differentiation, and apoptosis, which is crucial during embryonic development [37]. Certain subclasses of BMPs, such as BMP4, have specific effects on developmental pathways, including that of inner ear hair cells and spiral ganglion neurons, highlighting its importance in neurosensory differentiation [39,40].
3.2. Endogenous Activators and Inhibitors of BMP Signalling
BMP signalling is finely tuned by endogenous molecules and its role in the differentiation of many cell types, including neural differentiation, is complex. BMPs are generally antagonistic to neural differentiation at early stages of development but promote the formation of autonomic and sensory neurons from neural crest progenitors at later stages. Extracellular antagonists like Noggin, Chordin, Gremlin, and Follistatin bind BMP proteins (Figure 2), inhibiting receptor interaction and modulating processes such as neural and limb development. Conversely, modulators like Twisted Gastrulation (TWSG1) can either enhance or inhibit BMP signalling according to the developmental context. For instance, TWSG1 can enhance BMP signalling in the context of early neural development, promoting neural crest cell formation, while it can inhibit BMP signalling during limb formation to prevent excessive growth. USAG1 directly binds to BMPs to antagonise BMP signalling and has been shown to be important for tooth and kidney development [104,105]. Transmembrane anterior posterior transformation 1 (TAPT1), involved in axial skeletal patterning, causes proteasomal degradation of SMAD1/5, thereby inhibiting BMP signalling [106,107,108]. R-spondin 2 and 3 (RSPO2 and RSPO3) act as BMP antagnoists by binding to the BMP receptor BMPR1A resulting in their internalisation and degradation [109,110]. Intracellular inhibitors, such as SMAD6 and SMAD7, prevent R-SMAD phosphorylation or promote receptor degradation, ensuring balanced BMP4 activity for normal development [111,112]. BMP4 specifically promotes glial differentiation while inhibiting oligodendrocyte formation and can be overridden by Notch signalling, which favours Schwann cell differentiation [113].
4. Consequences of BMP Dysregulation
Because of their diverse roles in development and differentiation of many cell types, dysregulation of BMPs, their receptors, and their endogenous modulators can have a spectrum of effects on nearly every tissue type involving all three germ layers. Indeed, BMPs are essential for development, with embryonic lethality reported in mice lacking expression of either BMP2 or BMP4 [114,115,116,117,118], and mice deficient in BMP7 have eye, kidney, and skeletal patterning defects and die shortly after birth [113]. Expression of BMP4 is found to be strong in mouse caudal tissues, and loss of BMP4 in this region resulted in hindlimb fusion and lethality [119].
The effects of dysregulation of BMP signalling and its link to various diseases have been extensively reviewed elsewhere [120] and highlight the need for improved understanding of the roles of BMPs, their receptors, and their modulators in development and disease. Considering the importance of BMP signalling in development and differentiation of tissues, studying the effects of BMPs in whole model organisms is challenging due to the lack of viability and early arrest of growth and development of embryos following perturbation of BMP signalling. Moreover, many different tissues and organs may be affected, which further complicates interpretation of the effects of loss, mutation, or forced expression of BMP and/or its receptors and modulators in the whole organism. Some of these effects are likely to be secondary as a result of gross defects arising from dysregulation of BMP signalling, rather than as a direct consequence of BMP signalling. Conditional knockouts (and other similar targeting of specific tissues) might overcome some of these limitations. For instance, while loss of BMP4 results in embryonic lethality, Suzuki et al. [121,122,123] were able to use a conditional knockout Isl1-Cre mouse line in which BMP4 expression was reduced in the caudal body region only, allowing their mice to survive to a developmental stage late enough to investigate the caudalising effects of BMP4. Likewise, Chang et al. [124] conditionally deleted BMP4 expression in the mouse inner ear and were able to demonstrate the importance of BMP4 in the formation of the vestibular cristae and canals. They also succeeded in electroporating expression vectors to inhibit BMP signalling directly into the otocyst of the developing chick and observed that down-regulation of BMP resulted in patterning defects in the crista, although they cautioned that some of the effects could also be due to electroporation rather than reduced BMP signalling.
Using pluripotent stem cells to investigate BMP signalling could also be used to overcome some of the limitations mentioned above, although care must be taken to ensure that perturbations of BMP signalling do not affect their overall survival, maintenance, and differentiation potential. Indeed, the ability of cells to differentiate towards the desired lineage is likely to be affected in conditions of abnormal BMP signalling or if cells are unable to respond to exogenous BMP. Maintenance of murine ES cell pluripotency has been shown to require BMP4, which induces expression of Klf2 [125], although direct BMP signalling is unlikely to be involved in maintenance of human ES and iPS cell pluripotency, as these cells are primed unlike mouse ES and require Activin A for their maintenance [126].
Generation of pluripotent stem cells may also be affected by perturbation of BMP activity. For instance, fibroblasts from fibrodysplasia ossificans progressiva patients carrying a mutation in the ACVR1 gene which resulted in hyperactivation of BMP-SMAD signalling were found to have increased iPSC reprogramming efficiency [127]. Addition of exogenous BMP4 to cultures during early stages of reprogramming was found to have a similar effect. Recently, modulation of the stiffness of the hydrogels on which fibroblasts undergoing reprogramming to iPSCs were cultured was found to upregulate BMP2 and several genes involved in BMP signalling, as well as improve reprogramming of the cells. Increased hydrogel stiffness upregulated Phactr3, which then resulted in increased BMP2 and improved reprogramming efficiency [128]. How Phactr3 causes an increase in BMP2 is not known, although Phactr3 is known to associate with nuclear nonchromatin structure [129,130], where it might influence expression of genes involved in reprogramming. Alternatively, Phactr3 may exert its effects by inhibiting polymerisation of actin in the cell, resulting in increased cell spreading [131,132], as changes in cell shape have recently been shown to affect distribution of BMP receptors on the cell membrane [133,134].
5. Uses of Pluripotent Stem Cells to Investigate the Role of BMP in Development
5.1. Advantages and Limitations of Human PSCs
Stem cells, and more specifically human-derived stem cells, are a valuable tool to study development and disease mechanisms without the need for fetal samples that are difficult to acquire. They offer the possibility to study aspects of development that are specific to humans. Indeed, previous studies have demonstrated differences in development and disease mechanisms between humans and animal models [135]. For instance, in the case of the inner ear, development and maturation is almost completed by around 36 gestation weeks in humans [136,137], while in mice the cells of the cochlea of the inner ear continue to develop and mature after birth until about the third post-natal week [138]. This highlights the need for human-specific models to study development. Moreover, patient-derived stem cells can be used to study development in a patient-specific or disease context, without the need for generating mutant cell lines that might not behave in the same manner or fail to recapitulate some aspects of the disease. Gene correction of such patient-derived stem cells can also be used to correct mutations to investigate whether proper functioning of the gene is regained, opening the way for gene therapy treatments. For instance, patient-derived hiPSCs have recently been used to model mutations in TMC1, which are associated with a type of progressive hearing loss termed DFNA36 [139]. While differentiation of pluripotent stem cells to sensory hair cells was not affected by the TMC1 mutation, the morphology and electrophysiological properties of the derived hair cells were altered. Moreover, using CRISPR/Cas9 genome editing technique to generate an isogenic cell line, in which the mutated gene was corrected, resulted in recovery of hair cell morphology and electrophysiology. Similar works have been done using patient-derived lines carrying mutations for several genes associated with hearing loss including USH2A [140], TRMU [141], ELMOD3 [142,143,144], MYO7A [145], and AIFM1 [146,147], highlighting the potential strength of this approach for therapeutic genome editing.
In spite of recent developments and advances in stem cell technologies, numerous barriers must still be overcome before stem cells can reach their optimal potential in research and in the clinic. Many stem cell differentiation protocols result in batch-to-batch variability and also variation between labs and require further refinements to produce more uniform and homogenous populations of the desired cell and tissue types being investigated. Furthermore, unlike studying development in animal models, such in vitro differentiation often occurs in isolation from other cell types, which might provide trophic and supportive factors beneficial for generating the cells under investigation. For instance, differentiation and development of inner ear hair cells requires the support of the surrounding connective tissue and mesenchymal cells [148]. Indeed, otic mesenchyme cells comprise a diverse cell type that make up several important cell types in the inner ear, including spiral limbus fibrocytes and modiolar osteoblasts [149]. Because hair cells require neurons to transmit auditory signals to the brain, co-culture of stem cell-derived hair cells with spiral ganglion neurons should be considered to make functional circuits. The generation of vascularised organoids would additionally be beneficial to enable the growth of larger and healthier organoids. Incorporation of such tissues in the form of co-cultures and assembloids can lead to the development of more robust and mature models which are already showing promising results [150], but would also complicate the system, especially if looking for populations of pure and mature cells with the intention of being able to transplant the generated cells into patients. Finally, different biomaterials should also be tested to investigate their roles and potential benefits in constructing more physiologically relevant 3D culture systems that better recapitulate the tissue microenvironment (reviewed in [17,18,20,22,24,151]), as remodelling of the extracellular matrix plays an important role during maturation of the cochlea (reviewed in [20]).
5.2. Otic Neurosensory Specification as a Model to Study BMP4 Signalling
Because BMPs are involved in many steps of inner ear development and are required at specific concentrations for precise durations [152], otic lineages provide an interesting model system to investigate the effects of BMP signalling during development. Human PSCs can be differentiated under either 2D or 3D culture systems to give rise to otic progenitors that express several of the markers and components of activation pathways found during early otic development, and eventually hair cells, supporting cells, and neurons in inner ear otic organoids that have been allowed to mature in long-term culture [153]. Recent advances in 3D-otic organoids have additionally been able to generate both cochlear and vestibular type hair cells [154], demonstrating the ability to finely control generation of inner ear hair cells in such 3D-cell culture systems.
Mutations in some genes involved in BMP signalling are associated with hearing loss (Table 1). Nager syndrome is associated with hearing loss as a result of mutations in the SF3B4 gene, which codes for a spliceosome that affects expression of Noggin and BMP and may be directly involved in neural crest and otic development [155]. Mutations in chondroitin synthase 1 (CHSY1), involved in the synthesis of chondroitin sulfate, are characterised by limb malformations, short stature, and hearing loss [156,157,158,159,160,161,162,163,164,165], and studies in the inner ears of zebrafish larvae have found that Chsy1 expression is similar to that of the BMP inhibitor dan and complementary to Bmp2b expression, suggesting a role for this gene in BMP signalling and otic development [152]. In cultures of mouse chondrocytes, knockdown of Chsy1 resulted in increased BMP signalling, while overexpression of Chsy1 reduced BMP signalling [166,167,168,169,170,171,172]. Whether similar effects of Sf3b4, Chsy1, and other genes potentially involved in BMP signalling (Table 1) can be observed in cultures of PSC-derived otic progenitors remains to be investigated.
One of the consequences of suboptimal BMP4 signalling during early specification of otic progenitors under these pluripotent cell culture systems is the generation of off-target cell types such as neurons and surface epidermis [96]. Current methods for detecting subtle differences in off-target differentiation are mostly restricted to immunolabelling and qPCR analyses for off-target genes, most of which are transcription factors. It has also been reported that the epithelial thickness of organoids after just 3 days in vitro can be used as a proxy to optimise BMP4 concentration in such cultures [109,110]. However, the link between BMP4 concentration and epidermal thickness is not clear, and this method requires the production and screening of many otic organoids. New methods that can allow for the detection of off-target differentiation more quickly and using fewer samples would enable researchers to detect such off-target effects more quickly and to better understand the variations between different lineages beyond the expression of transcription factors, for example, in the biochemical and metabolic properties of such in vitro differentiated cells.
Pluripotent stem cells also offer the opportunity to study the effects of BMP signalling at later stages of development. Several studies have reported conductive hearing loss in patients with mutations in the NOG gene, which encodes for the BMP antagonist Noggin, as a result of auditory-ossicle fusion [111,112]. As these patients display additional symptoms, including bone and joint disorders and digital anomalies, patient-derived stem cells may allow for the study of these mutations specifically in inner ear development, and to determine at which stages in development these symptoms begin to appear, as well as follow disease progression and test the effects of potential therapeutics. PSC-derived otic organoids could also be used to investigate the role of different BMPs in cochlear and vestibular development in humans, as these organs have been shown to require differential BMP signalling in chick [113].
6. Conclusions and Future Perspectives
Advances in stem cell research have greatly expanded our knowledge and understanding of development and the signalling pathways involved in developmental processes and have prompted new questions and lines of investigation. Nevertheless, as the situation with BMP signalling in development of the inner ear and other tissues has demonstrated, further work is needed to better understand general human-specific developmental and disease pathways and mechanisms, rather than potentially batch or cell line-specific features. As signalling pathways other than BMP are likely to differ between cell types and also perhaps culture conditions, the starting state of stem cell cultures should be determined before initiating any cell differentiation protocol, in order to ensure that the optimal conditions for differentiation of the desired tissues are being met. New technologies could help to simplify with determining the endogenous levels of signalling molecules and signalling activity in cell lines, allowing for more robust and homogeneous cultures that better recapitulate the in vivo situation.
Author Contributions
Conceptualization, A.Z. and K.C.; writing—original draft preparation, K.C. and M.M.; writing—review and editing, K.C., A.Z., M.M., L.R.; visualization, K.C., A.Z., M.M., L.R.; supervision, A.Z.; project administration, A.Z.; funding acquisition, A.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research has been financially supported by la Fondation pour l’Audition (Paris) to A.Z, grant number FPA RD-2022-8.
Acknowledgments
We sincerely apologize for the articles that could not be referenced due to space limitations. We thank several collaborators, in particular Frederic Cuisinier (LBN, University of Montpellier), Hanae Lahlou, and Albert Edge (Harvard Medical School).
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
| ACVR1 | Activin A receptor type 1 |
| BMP | Bone morphogenetic protein |
| CHSY1 | Chondroitin synthase 1 |
| ES | Embryonic stem cell |
| FGF | Fibroblast growth factor |
| hiPSC | Human induced pluripotent stem cell |
| OEPD | Otic-epibranchial progenitor domain |
| MSC | Mesenchymal stem cell |
| NNE | Non-neural ectoderm |
| PPE | Pre-placodal ectoderm |
| PSC | Pluripotent stem cell |
| SHH | Sonic hedgehog |
| TAPT1 | Transmembrane anterior posterior transformation 1 |
| TGFβ | Transforming growth factor beta |
| TMEM53 | Transmembrane protein 53 |
| TWSG1 | Twisted Gastrulation 1 |
| USAG1 | Uterine sensitization associated gene-1 |
References
- Chiaradia, I.; Lancaster, M.A. Brain Organoids for the Study of Human Neurobiology at the Interface of in Vitro and in Vivo. Nat. Neurosci. 2020, 23, 1496–1508. [Google Scholar] [CrossRef] [PubMed]
- Lancaster, M.A.; Renner, M.; Martin, C.-A.; Wenzel, D.; Bicknell, L.S.; Hurles, M.E.; Homfray, T.; Penninger, J.M.; Jackson, A.P.; Knoblich, J.A. Cerebral Organoids Model Human Brain Development and Microcephaly. Nature 2013, 501, 373–379. [Google Scholar] [CrossRef] [PubMed]
- Andersen, P.; Tampakakis, E.; Jimenez, D. V.; Kannan, S.; Miyamoto, M.; Shin, H.K.; Saberi, A.; Murphy, S.; Sulistio, E.; Chelko, S.P.; et al. Precardiac Organoids Form Two Heart Fields via Bmp/Wnt Signaling. Nat. Commun. 2018, 9, 3140. [Google Scholar] [CrossRef] [PubMed]
- Kostina, A.; Lewis-Israeli, Y.R.; Abdelhamid, M.; Gabalski, M.A.; Kiselev, A.; Volmert, B.D.; Lankerd, H.; Huang, A.R.; Wasserman, A.H.; Lydic, T.; et al. ER Stress and Lipid Imbalance Drive Diabetic Embryonic Cardiomyopathy in an Organoid Model of Human Heart Development. Stem Cell Reports 2024, 19, 317–330. [Google Scholar] [CrossRef] [PubMed]
- Lewis-Israeli, Y.R.; Wasserman, A.H.; Gabalski, M.A.; Volmert, B.D.; Ming, Y.; Ball, K.A.; Yang, W.; Zou, J.; Ni, G.; Pajares, N.; et al. Self-Assembling Human Heart Organoids for the Modeling of Cardiac Development and Congenital Heart Disease. Nat. Commun. 2021, 12, 5142. [Google Scholar] [CrossRef]
- Lewis-Israeli, Y.R.; Abdelhamid, M.; Olomu, I.; Aguirre, A. Modeling the Effects of Maternal Diabetes on the Developing Human Heart Using Pluripotent Stem Cell–Derived Heart Organoids. Curr. Protoc. 2022, 2. [Google Scholar] [CrossRef]
- Wimmer, R.A.; Leopoldi, A.; Aichinger, M.; Wick, N.; Hantusch, B.; Novatchkova, M.; Taubenschmid, J.; Hämmerle, M.; Esk, C.; Bagley, J.A.; et al. Human Blood Vessel Organoids as a Model of Diabetic Vasculopathy. Nature 2019, 565, 505–510. [Google Scholar] [CrossRef]
- Agarwal, D.; Dash, N.; Mazo, K.W.; Chopra, M.; Avila, M.P.; Patel, A.; Wong, R.M.; Jia, C.; Do, H.; Cheng, J.; et al. Human Retinal Ganglion Cell Neurons Generated by Synchronous BMP Inhibition and Transcription Factor Mediated Reprogramming. npj Regen. Med. 2023, 8, 55. [Google Scholar] [CrossRef]
- Capowski, E.E.; Samimi, K.; Mayerl, S.J.; Phillips, M.J.; Pinilla, I.; Howden, S.E.; Saha, J.; Jansen, A.D.; Edwards, K.L.; Jager, L.D.; et al. Reproducibility and Staging of 3D Human Retinal Organoids across Multiple Pluripotent Stem Cell Lines. Development 2018. [Google Scholar] [CrossRef]
- Chichagova, V.; Hilgen, G.; Ghareeb, A.; Georgiou, M.; Carter, M.; Sernagor, E.; Lako, M.; Armstrong, L. Human IPSC Differentiation to Retinal Organoids in Response to IGF1 and BMP4 Activation Is Line- and Method-Dependent. Stem Cells 2020, 38, 195–201. [Google Scholar] [CrossRef]
- Hallam, D.; Hilgen, G.; Dorgau, B.; Zhu, L.; Yu, M.; Bojic, S.; Hewitt, P.; Schmitt, M.; Uteng, M.; Kustermann, S.; et al. Human-Induced Pluripotent Stem Cells Generate Light Responsive Retinal Organoids with Variable and Nutrient-Dependent Efficiency. Stem Cells 2018, 36, 1535–1551. [Google Scholar] [CrossRef] [PubMed]
- Kamarudin, T.A.; Bojic, S.; Collin, J.; Yu, M.; Alharthi, S.; Buck, H.; Shortt, A.; Armstrong, L.; Figueiredo, F.C.; Lako, M. Differences in the Activity of Endogenous Bone Morphogenetic Protein Signaling Impact on the Ability of Induced Pluripotent Stem Cells to Differentiate to Corneal Epithelial-Like Cells. Stem Cells 2018, 36, 337–348. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.; Kabir, F.; Thomson, J.J.; Ma, Y.; Qiu, C.; Delannoy, M.; Khan, S.Y.; Riazuddin, S.A. Comparative Transcriptome Analysis of HESC- and IPSC-Derived Lentoid Bodies. Sci. Rep. 2019, 9, 18552. [Google Scholar] [CrossRef] [PubMed]
- Dincer, Z.; Piao, J.; Niu, L.; Ganat, Y.; Kriks, S.; Zimmer, B.; Shi, S.-H.; Tabar, V.; Studer, L. Specification of Functional Cranial Placode Derivatives from Human Pluripotent Stem Cells. Cell Rep. 2013, 5, 1387–1402. [Google Scholar] [CrossRef]
- Fu, Q.; Qin, Z.; Jin, X.; Zhang, L.; Chen, Z.; He, J.; Ji, J.; Yao, K. Generation of Functional Lentoid Bodies From Human Induced Pluripotent Stem Cells Derived From Urinary Cells. Investig. Opthalmology Vis. Sci. 2017, 58, 517. [Google Scholar] [CrossRef]
- Zhang, L.; Qin, Z.; Lyu, D.; Lu, B.; Chen, Z.; Fu, Q.; Yao, K. Postponement of the Opacification of Lentoid Bodies Derived from Human Induced Pluripotent Stem Cells after Lanosterol Treatment—the First Use of the Lens Aging Model in Vitro in Cataract Drug Screening. Front. Pharmacol. 2022, 13. [Google Scholar] [CrossRef]
- Doda, D.; Alonso Jimenez, S.; Rehrauer, H.; Carreño, J.F.; Valsamides, V.; Di Santo, S.; Widmer, H.R.; Edge, A.; Locher, H.; van der Valk, W.H.; et al. Human Pluripotent Stem Cell-Derived Inner Ear Organoids Recapitulate Otic Development in Vitro. Development 2023, 150. [Google Scholar] [CrossRef]
- Koehler, K.R.; Nie, J.; Longworth-Mills, E.; Liu, X.-P.; Lee, J.; Holt, J.R.; Hashino, E. Generation of Inner Ear Organoids Containing Functional Hair Cells from Human Pluripotent Stem Cells. Nat. Biotechnol. 2017, 35, 583–589. [Google Scholar] [CrossRef]
- Kurihara, S.; Fujioka, M.; Hirabayashi, M.; Yoshida, T.; Hosoya, M.; Nagase, M.; Kato, F.; Ogawa, K.; Okano, H.; Kojima, H.; et al. Otic Organoids Containing Spiral Ganglion Neuron-like Cells Derived from Human-Induced Pluripotent Stem Cells as a Model of Drug-Induced Neuropathy. Stem Cells Transl. Med. 2022, 11, 282–296. [Google Scholar] [CrossRef]
- Moore, S.T.; Nakamura, T.; Nie, J.; Solivais, A.J.; Aristizábal-Ramírez, I.; Ueda, Y.; Manikandan, M.; Reddy, V.S.; Romano, D.R.; Hoffman, J.R.; et al. Generating High-Fidelity Cochlear Organoids from Human Pluripotent Stem Cells. Cell Stem Cell 2023, 30, 950–961. [Google Scholar] [CrossRef]
- Nie, J.; Ueda, Y.; Solivais, A.J.; Hashino, E. CHD7 Regulates Otic Lineage Specification and Hair Cell Differentiation in Human Inner Ear Organoids. Nat. Commun. 2022, 13, 7053. [Google Scholar] [CrossRef] [PubMed]
- Steinhart, M.R.; van der Valk, W.H.; Osorio, D.; Serdy, S.A.; Zhang, J.; Nist-Lund, C.; Kim, J.; Moncada-Reid, C.; Sun, L.; Lee, J.; et al. Mapping Oto-Pharyngeal Development in a Human Inner Ear Organoid Model. Development 2023, 150. [Google Scholar] [CrossRef] [PubMed]
- Ueda, Y.; Nakamura, T.; Nie, J.; Solivais, A.J.; Hoffman, J.R.; Daye, B.J.; Hashino, E. Defining Developmental Trajectories of Prosensory Cells in Human Inner Ear Organoids at Single-Cell Resolution. Development 2023, 150. [Google Scholar] [CrossRef] [PubMed]
- van der Valk, W.H.; van Beelen, E.S.A.; Steinhart, M.R.; Nist-Lund, C.; Osorio, D.; de Groot, J.C.M.J.; Sun, L.; van Benthem, P.P.G.; Koehler, K.R.; Locher, H. A Single-Cell Level Comparison of Human Inner Ear Organoids with the Human Cochlea and Vestibular Organs. Cell Rep. 2023, 42, 112623. [Google Scholar] [CrossRef] [PubMed]
- Kattman, S.J.; Witty, A.D.; Gagliardi, M.; Dubois, N.C.; Niapour, M.; Hotta, A.; Ellis, J.; Keller, G. Stage-Specific Optimization of Activin/Nodal and BMP Signaling Promotes Cardiac Differentiation of Mouse and Human Pluripotent Stem Cell Lines. Cell Stem Cell 2011, 8, 228–240. [Google Scholar] [CrossRef]
- Sa, S.; McCloskey, K.E. Activin A and BMP4 Signaling for Efficient Cardiac Differentiation of H7 and H9 Human Embryonic Stem Cells. J. Stem Cells Regen. Med. 2012, 8, 198–202. [Google Scholar] [CrossRef]
- Britton, G.; Heemskerk, I.; Hodge, R.; Qutub, A.A.; Warmflash, A. A Novel Self-Organizing Embryonic Stem Cell System Reveals Signaling Logic Underlying the Patterning of Human Ectoderm. Development 2019. [Google Scholar] [CrossRef]
- Camacho-Aguilar, E.; Yoon, S.T.; Ortiz-Salazar, M.A.; Du, S.; Guerra, M.C.; Warmflash, A. Combinatorial Interpretation of BMP and WNT Controls the Decision between Primitive Streak and Extraembryonic Fates. Cell Syst. 2024, 15, 445–461. [Google Scholar] [CrossRef]
- Teague, S.; Primavera, G.; Chen, B.; Liu, Z.-Y.; Yao, L.; Freeburne, E.; Khan, H.; Jo, K.; Johnson, C.; Heemskerk, I. Time-Integrated BMP Signaling Determines Fate in a Stem Cell Model for Early Human Development. Nat. Commun. 2024, 15, 1471. [Google Scholar] [CrossRef]
- Urist, M.R.; Strates, B.S. Bone Morphogenetic Protein. J. Dent. Res. 1971, 50, 1392–1406. [Google Scholar] [CrossRef]
- Yan, Y.; Wang, Q. BMP Signaling: Lighting up the Way for Embryonic Dorsoventral Patterning. Front. Cell Dev. Biol. 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.-J.; Bhat, N.; Sweet, E.M.; Cornell, R.A.; Riley, B.B. Identification of Early Requirements for Preplacodal Ectoderm and Sensory Organ Development. PLoS Genet. 2010, 6, e1001133. [Google Scholar] [CrossRef] [PubMed]
- Leung, A.W.; Kent Morest, D.; Li, J.Y.H. Differential BMP Signaling Controls Formation and Differentiation of Multipotent Preplacodal Ectoderm Progenitors from Human Embryonic Stem Cells. Dev. Biol. 2013, 379, 208–220. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Liu, C.; Biechele, S.; Zhu, Q.; Song, L.; Lanner, F.; Jing, N.; Rossant, J. Location of Transient Ectodermal Progenitor Potential in Mouse Development. Development 2013, 140, 4533–4543. [Google Scholar] [CrossRef] [PubMed]
- Reichert, S.; Randall, R.A.; Hill, C.S. A BMP Regulatory Network Controls Ectodermal Cell Fate Decisions at the Neural Plate Border. Development 2013, 140, 4435–4444. [Google Scholar] [CrossRef]
- Kishimoto, Y.; Lee, K.-H.; Zon, L.; Hammerschmidt, M.; Schulte-Merker, S. The Molecular Nature of Zebrafish Swirl : BMP2 Function Is Essential during Early Dorsoventral Patterning. Development 1997, 124, 4457–4466. [Google Scholar] [CrossRef]
- Wang, B.; Zhao, Q.; Gong, X.; Wang, C.; Bai, Y.; Wang, H.; Zhou, J.; Rong, X. Transmembrane Anterior Posterior Transformation 1 Regulates BMP Signaling and Modulates the Protein Stability of SMAD1/5. J. Biol. Chem. 2022, 298, 102684. [Google Scholar] [CrossRef]
- Yanagita, M.; Oka, M.; Watabe, T.; Iguchi, H.; Niida, A.; Takahashi, S.; Akiyama, T.; Miyazono, K.; Yanagisawa, M.; Sakurai, T. USAG-1: A Bone Morphogenetic Protein Antagonist Abundantly Expressed in the Kidney. Biochem. Biophys. Res. Commun. 2004, 316, 490–500. [Google Scholar] [CrossRef]
- Lee, H.; Seidl, C.; Sun, R.; Glinka, A.; Niehrs, C. R-Spondins Are BMP Receptor Antagonists in Xenopus Early Embryonic Development. Nat. Commun. 2020, 11, 5570. [Google Scholar] [CrossRef]
- Lee, H.; Sun, R.; Niehrs, C. Uncoupling the BMP Receptor Antagonist Function from the WNT Agonist Function of R-Spondin 2 Using the Inhibitory Peptide Dendrimer RWd. J. Biol. Chem. 2022, 298, 101586. [Google Scholar] [CrossRef]
- Wilson, P.A.; Lagna, G.; Suzuki, A.; Hemmati-Brivanlou, A. Concentration-Dependent Patterning of the Xenopus Ectoderm by BMP4 and Its Signal Transducer Smad1. Development 1997, 124, 3177–3184. [Google Scholar] [CrossRef] [PubMed]
- Tchieu, J.; Zimmer, B.; Fattahi, F.; Amin, S.; Zeltner, N.; Chen, S.; Studer, L. A Modular Platform for Differentiation of Human PSCs into All Major Ectodermal Lineages. Cell Stem Cell 2017, 21, 399–410. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-K.; Lee, H.-S.; Moody, S.A. Neural Transcription Factors: From Embryos to Neural Stem Cells. Mol. Cells 2014, 37, 705–712. [Google Scholar] [CrossRef] [PubMed]
- Zine, A.; Fritzsch, B. Early Steps towards Hearing: Placodes and Sensory Development. Int. J. Mol. Sci. 2023, 24, 6994. [Google Scholar] [CrossRef] [PubMed]
- Bakkers, J.; Hild, M.; Kramer, C.; Furutani-Seiki, M.; Hammerschmidt, M. Zebrafish ΔNp63 Is a Direct Target of Bmp Signaling and Encodes a Transcriptional Repressor Blocking Neural Specification in the Ventral Ectoderm. Dev. Cell 2002, 2, 617–627. [Google Scholar] [CrossRef]
- Metallo, C.M.; Ji, L.; de Pablo, J.J.; Palecek, S.P. Retinoic Acid and Bone Morphogenetic Protein Signaling Synergize to Efficiently Direct Epithelial Differentiation of Human Embryonic Stem Cells. Stem Cells 2008, 26, 372–380. [Google Scholar] [CrossRef]
- Conti, E.; Harschnitz, O. Human Stem Cell Models to Study Placode Development, Function and Pathology. Development 2022, 149. [Google Scholar] [CrossRef]
- Raible, D.W.; Ragland, J.W. Reiterated Wnt and BMP Signals in Neural Crest Development. Semin. Cell Dev. Biol. 2005, 16, 673–682. [Google Scholar] [CrossRef]
- Mehrotra, P.; Ikhapoh, I.; Lei, P.; Tseropoulos, G.; Zhang, Y.; Wang, J.; Liu, S.; Bronner, M.E.; Andreadis, S.T. Wnt/BMP Mediated Metabolic Reprogramming Preserves Multipotency of Neural Crest-Like Stem Cells. Stem Cells 2023, 41, 287–305. [Google Scholar] [CrossRef]
- Ahrens, K.; Schlosser, G. Tissues and Signals Involved in the Induction of Placodal Six1 Expression in Xenopus Laevis. Dev. Biol. 2005, 288, 40–59. [Google Scholar] [CrossRef]
- Ito, A.; Miller, C.; Imamura, F. Suppression of BMP Signaling Restores Mitral Cell Development Impaired by FGF Signaling Deficits in Mouse Olfactory Bulb. Mol. Cell. Neurosci. 2024, 128, 103913. [Google Scholar] [CrossRef] [PubMed]
- Panaliappan, T.K.; Wittmann, W.; Jidigam, V.K.; Mercurio, S.; Bertolini, J.A.; Sghari, S.; Bose, R.; Patthey, C.; Nicolis, S.K.; Gunhaga, L. Sox2 Is Required for Olfactory Pit Formation and Olfactory Neurogenesis through BMP Restriction and Hes5 Upregulation. Development 2018, 145. [Google Scholar] [CrossRef] [PubMed]
- Peretto, P.; Cummings, D.; Modena, C.; Behrens, M.; Venkatraman, G.; Fasolo, A.; Margolis, F.L. BMP MRNA and Protein Expression in the Developing Mouse Olfactory System. J. Comp. Neurol. 2002, 451, 267–278. [Google Scholar] [CrossRef] [PubMed]
- Kikuta, H.; Tanaka, H.; Ozaki, T.; Ito, J.; Ma, J.; Moribe, S.; Hirano, M. Spontaneous Differentiation of Human Induced Pluripotent Stem Cells to Odorant-Responsive Olfactory Sensory Neurons. Biochem. Biophys. Res. Commun. 2024, 719, 150062. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Ping, L.; Gao, R.; Zhang, B.; Chen, X. Lmo4a Contributes to Zebrafish Inner Ear and Vestibular Development via Regulation of the Bmp Pathway. Genes (Basel). 2023, 14, 1371. [Google Scholar] [CrossRef]
- Pujades, C.; Kamaid, A.; Alsina, B.; Giraldez, F. BMP-Signaling Regulates the Generation of Hair-Cells. Dev. Biol. 2006, 292, 55–67. [Google Scholar] [CrossRef]
- Lewis, R.M.; Keller, J.J.; Wan, L.; Stone, J.S. Bone Morphogenetic Protein 4 Antagonizes Hair Cell Regeneration in the Avian Auditory Epithelium. Hear. Res. 2018, 364, 1–11. [Google Scholar] [CrossRef]
- Li, H.; Corrales, C.E.; Wang, Z.; Zhao, Y.; Wang, Y.; Liu, H.; Heller, S. BMP4 Signaling Is Involved in the Generation of Inner Ear Sensory Epithelia. BMC Dev. Biol. 2005, 5, 16. [Google Scholar] [CrossRef]
- Ohyama, T.; Basch, M.L.; Mishina, Y.; Lyons, K.M.; Segil, N.; Groves, A.K. BMP Signaling Is Necessary for Patterning the Sensory and Nonsensory Regions of the Developing Mammalian Cochlea. J. Neurosci. 2010, 30, 15044–15051. [Google Scholar] [CrossRef]
- Washausen, S.; Knabe, W. Responses of Epibranchial Placodes to Disruptions of the FGF and BMP Signaling Pathways in Embryonic Mice. Front. Cell Dev. Biol. 2021, 9. [Google Scholar] [CrossRef]
- Thesleff, I. Epithelial-Mesenchymal Signalling Regulating Tooth Morphogenesis. J. Cell Sci. 2003, 116, 1647–1648. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Feng, J.; Liu, Y.; Ho, T.-V.; Grimes, W.; Ho, H.A.; Park, S.; Wang, S.; Chai, Y. BMP-SHH Signaling Network Controls Epithelial Stem Cell Fate via Regulation of Its Niche in the Developing Tooth. Dev. Cell 2015, 33, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Zimmer, B.; Ewaleifoh, O.; Harschnitz, O.; Lee, Y.-S.; Peneau, C.; McAlpine, J.L.; Liu, B.; Tchieu, J.; Steinbeck, J.A.; Lafaille, F.; et al. Human IPSC-Derived Trigeminal Neurons Lack Constitutive TLR3-Dependent Immunity That Protects Cortical Neurons from HSV-1 Infection. Proc. Natl. Acad. Sci. 2018, 115. [Google Scholar] [CrossRef] [PubMed]
- Yuan, G.; Yang, G.; Zheng, Y.; Zhu, X.; Chen, Z.; Zhang, Z.; Chen, Y. The Non-Canonical BMP and Wnt/β-Catenin Signaling Pathways Orchestrate Early Tooth Development. Development 2015, 142, 128–139. [Google Scholar] [CrossRef]
- Kiso, H.; Takahashi, K.; Saito, K.; Togo, Y.; Tsukamoto, H.; Huang, B.; Sugai, M.; Shimizu, A.; Tabata, Y.; Economides, A.N.; et al. Interactions between BMP-7 and USAG-1 (Uterine Sensitization-Associated Gene-1) Regulate Supernumerary Organ Formations. PLoS One 2014, 9, e96938. [Google Scholar] [CrossRef]
- Murashima-Suginami, A.; Takahashi, K.; Sakata, T.; Tsukamoto, H.; Sugai, M.; Yanagita, M.; Shimizu, A.; Sakurai, T.; Slavkin, H.C.; Bessho, K. Enhanced BMP Signaling Results in Supernumerary Tooth Formation in USAG-1 Deficient Mouse. Biochem. Biophys. Res. Commun. 2008, 369, 1012–1016. [Google Scholar] [CrossRef]
- Murashima-Suginami, A.; Takahashi, K.; Kawabata, T.; Sakata, T.; Tsukamoto, H.; Sugai, M.; Yanagita, M.; Shimizu, A.; Sakurai, T.; Slavkin, H.C.; et al. Rudiment Incisors Survive and Erupt as Supernumerary Teeth as a Result of USAG-1 Abrogation. Biochem. Biophys. Res. Commun. 2007, 359, 549–555. [Google Scholar] [CrossRef]
- Murashima-Suginami, A.; Kiso, H.; Tokita, Y.; Mihara, E.; Nambu, Y.; Uozumi, R.; Tabata, Y.; Bessho, K.; Takagi, J.; Sugai, M.; et al. Anti-USAG-1 Therapy for Tooth Regeneration through Enhanced BMP Signaling. Sci. Adv. 2021, 7. [Google Scholar] [CrossRef]
- Li, Q.; Zhang, S.; Sui, Y.; Fu, X.; Li, Y.; Wei, S. Sequential Stimulation with Different Concentrations of BMP4 Promotes the Differentiation of Human Embryonic Stem Cells into Dental Epithelium with Potential for Tooth Formation. Stem Cell Res. Ther. 2019, 10, 276. [Google Scholar] [CrossRef]
- Zhu, X.; Li, Y.; Dong, Q.; Tian, C.; Gong, J.; Bai, X.; Ruan, J.; Gao, J. Small Molecules Promote the Rapid Generation of Dental Epithelial Cells from Human-Induced Pluripotent Stem Cells. Int. J. Mol. Sci. 2024, 25, 4138. [Google Scholar] [CrossRef]
- Galdos, F.X.; Guo, Y.; Paige, S.L.; VanDusen, N.J.; Wu, S.M.; Pu, W.T. Cardiac Regeneration. Circ. Res. 2017, 120, 941–959. [Google Scholar] [CrossRef] [PubMed]
- Schultheiss, T.M.; Burch, J.B.; Lassar, A.B. A Role for Bone Morphogenetic Proteins in the Induction of Cardiac Myogenesis. Genes Dev. 1997, 11, 451–462. [Google Scholar] [CrossRef]
- Lowery, J.W.; Rosen, V. The BMP Pathway and Its Inhibitors in the Skeleton. Physiol. Rev. 2018, 98, 2431–2452. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Wu, S.; Chen, W.; Li, Y.-P. The Roles and Regulatory Mechanisms of TGF-β and BMP Signaling in Bone and Cartilage Development, Homeostasis and Disease. Cell Res. 2024, 34, 101–123. [Google Scholar] [CrossRef] [PubMed]
- Gultian, K.A.; Gandhi, R.; DeCesari, K.; Romiyo, V.; Kleinbart, E.P.; Martin, K.; Gentile, P.M.; Kim, T.W.B.; Vega, S.L. Injectable Hydrogel with Immobilized BMP-2 Mimetic Peptide for Local Bone Regeneration. Front. Biomater. Sci. 2022, 1. [Google Scholar] [CrossRef] [PubMed]
- Hanada, K.; Dennis, J.E.; Caplan, A.I. Stimulatory Effects of Basic Fibroblast Growth Factor and Bone Morphogenetic Protein-2 on Osteogenic Differentiation of Rat Bone Marrow-Derived Mesenchymal Stem Cells. J. Bone Miner. Res. 1997, 12, 1606–1614. [Google Scholar] [CrossRef]
- Love, S.A.; Gultian, K.A.; Jalloh, U.S.; Stevens, A.; Kim, T.W.B.; Vega, S.L. Mesenchymal Stem Cells Enhance Targeted Bone Growth from Injectable Hydrogels with BMP-2 Peptides. J. Orthop. Res. 2024, 42, 1599–1607. [Google Scholar] [CrossRef]
- Moutsatsos, I.K.; Turgeman, G.; Zhou, S.; Kurkalli, B.G.; Pelled, G.; Tzur, L.; Kelley, P.; Stumm, N.; Mi, S.; Müller, R.; et al. Exogenously Regulated Stem Cell-Mediated Gene Therapy for Bone Regeneration. Mol. Ther. 2001, 3, 449–461. [Google Scholar] [CrossRef]
- Yamagishi, H. Cardiac Neural Crest. Cold Spring Harb. Perspect. Biol. 2021, 13, a036715. [Google Scholar] [CrossRef]
- Sande-Melón, M.; Marques, I.J.; Galardi-Castilla, M.; Langa, X.; Pérez-López, M.; Botos, M.-A.; Sánchez-Iranzo, H.; Guzmán-Martínez, G.; Ferreira Francisco, D.M.; Pavlinic, D.; et al. Adult Sox10+ Cardiomyocytes Contribute to Myocardial Regeneration in the Zebrafish. Cell Rep. 2019, 29, 1041–1054. [Google Scholar] [CrossRef]
- Tamura, Y.; Matsumura, K.; Sano, M.; Tabata, H.; Kimura, K.; Ieda, M.; Arai, T.; Ohno, Y.; Kanazawa, H.; Yuasa, S.; et al. Neural Crest–Derived Stem Cells Migrate and Differentiate Into Cardiomyocytes After Myocardial Infarction. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 582–589. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; Martik, M.L.; Li, Y.; Bronner, M.E. Cardiac Neural Crest Contributes to Cardiomyocytes in Amniotes and Heart Regeneration in Zebrafish. Elife 2019, 8. [Google Scholar] [CrossRef]
- Hatzistergos, K.E.; Takeuchi, L.M.; Saur, D.; Seidler, B.; Dymecki, S.M.; Mai, J.J.; White, I.A.; Balkan, W.; Kanashiro-Takeuchi, R.M.; Schally, A. V.; et al. CKit + Cardiac Progenitors of Neural Crest Origin. Proc. Natl. Acad. Sci. 2015, 112, 13051–13056. [Google Scholar] [CrossRef] [PubMed]
- Arai, H.N.; Sato, F.; Yamamoto, T.; Woltjen, K.; Kiyonari, H.; Yoshimoto, Y.; Shukunami, C.; Akiyama, H.; Kist, R.; Sehara-Fujisawa, A. Metalloprotease-Dependent Attenuation of BMP Signaling Restricts Cardiac Neural Crest Cell Fate. Cell Rep. 2019, 29, 603–616. [Google Scholar] [CrossRef] [PubMed]
- Sela-Donenfeld, D.; Kalcheim, C. Regulation of the Onset of Neural Crest Migration by Coordinated Activity of BMP4 and Noggin in the Dorsal Neural Tube. Development 1999, 126, 4749–4762. [Google Scholar] [CrossRef] [PubMed]
- Burstyn-Cohen, T.; Stanleigh, J.; Sela-Donenfeld, D.; Kalcheim, C. Canonical Wnt Activity Regulates Trunk Neural Crest Delamination Linking BMP/Noggin Signaling with G1/S Transition. Development 2004, 131, 5327–5339. [Google Scholar] [CrossRef]
- Shoval, I.; Ludwig, A.; Kalcheim, C. Antagonistic Roles of Full-Length N-Cadherin and Its Soluble BMP Cleavage Product in Neural Crest Delamination. Development 2007, 134, 491–501. [Google Scholar] [CrossRef]
- Freitas, G.P.; Lopes, H.B.; Souza, A.T.P.; Gomes, M.P.O.; Quiles, G.K.; Gordon, J.; Tye, C.; Stein, J.L.; Stein, G.S.; Lian, J.B.; et al. Mesenchymal Stem Cells Overexpressing BMP-9 by CRISPR-Cas9 Present High in Vitro Osteogenic Potential and Enhance in Vivo Bone Formation. Gene Ther. 2021, 28, 748–759. [Google Scholar] [CrossRef]
- Cui, J.; Zhang, W.; Huang, E.; Wang, J.; Liao, J.; Li, R.; Yu, X.; Zhao, C.; Zeng, Z.; Shu, Y.; et al. BMP9-Induced Osteoblastic Differentiation Requires Functional Notch Signaling in Mesenchymal Stem Cells. Lab. Investig. 2019, 99, 58–71. [Google Scholar] [CrossRef]
- Calixto, R.D.; Freitas, G.P.; Souza, P.G.; Ramos, J.I.R.; Santos, I.C.; de Oliveira, F.S.; Almeida, A.L.G.; Rosa, A.L.; Beloti, M.M. Effect of the Secretome of Mesenchymal Stem Cells Overexpressing BMP-9 on Osteoblast Differentiation and Bone Repair. J. Cell. Physiol. 2023, 238, 2625–2637. [Google Scholar] [CrossRef]
- Lee, S.-B.; Lee, H.-J.; Park, J.-B. Bone Morphogenetic Protein-9 Promotes Osteogenic Differentiation and Mineralization in Human Stem-Cell-Derived Spheroids. Medicina (B. Aires). 2023, 59, 1315. [Google Scholar] [CrossRef] [PubMed]
- Kawai, S.; Yoshitomi, H.; Sunaga, J.; Alev, C.; Nagata, S.; Nishio, M.; Hada, M.; Koyama, Y.; Uemura, M.; Sekiguchi, K.; et al. In Vitro Bone-like Nodules Generated from Patient-Derived IPSCs Recapitulate Pathological Bone Phenotypes. Nat. Biomed. Eng. 2019, 3, 558–570. [Google Scholar] [CrossRef] [PubMed]
- Oudina, K.; Paquet, J.; Moya, A.; Massourides, E.; Bensidhoum, M.; Larochette, N.; Deschepper, M.; Pinset, C.; Petite, H. The Paracrine Effects of Human Induced Pluripotent Stem Cells Promote Bone-like Structures via the Upregulation of BMP Expression in a Mouse Ectopic Model. Sci. Rep. 2018, 8, 17106. [Google Scholar] [CrossRef] [PubMed]
- Coucouvanis, E.; Martin, G.R. BMP Signaling Plays a Role in Visceral Endoderm Differentiation and Cavitation in the Early Mouse Embryo. Development 1999, 126, 535–546. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.N.; Green, J.; Wang, Z.; Deng, Y.; Qiao, M.; Peabody, M.; Zhang, Q.; Ye, J.; Yan, Z.; Denduluri, S.; et al. Bone Morphogenetic Protein (BMP) Signaling in Development and Human Diseases. Genes Dis. 2014, 1, 87–105. [Google Scholar] [CrossRef]
- Oh, S.-H.; Johnson, R.; Wu, D.K. Differential Expression of Bone Morphogenetic Proteins in the Developing Vestibular and Auditory Sensory Organs. J. Neurosci. 1996, 16, 6463–6475. [Google Scholar] [CrossRef]
- Graf, D.; Malik, Z.; Hayano, S.; Mishina, Y. Common Mechanisms in Development and Disease: BMP Signaling in Craniofacial Development. Cytokine Growth Factor Rev. 2016, 27, 129–139. [Google Scholar] [CrossRef]
- Mimura, S.; Suga, M.; Okada, K.; Kinehara, M.; Nikawa, H.; Furue, M.K. Bone Morphogenetic Protein 4 Promotes Craniofacial Neural Crest Induction from Human Pluripotent Stem Cells. Int. J. Dev. Biol. 2016, 60, 21–28. [Google Scholar] [CrossRef]
- Komatsu, Y.; Yu, P.B.; Kamiya, N.; Pan, H.; Fukuda, T.; Scott, G.J.; Ray, M.K.; Yamamura, K.; Mishina, Y. Augmentation of Smad-Dependent BMP Signaling in Neural Crest Cells Causes Craniosynostosis in Mice. J. Bone Miner. Res. 2013, 28, 1422–1433. [Google Scholar] [CrossRef]
- Ueharu, H.; Pan, H.; Liu, X.; Ishii, M.; Pongetti, J.; Kulkarni, A.K.; Adegbenro, F.E.; Wurn, J.; Maxson, R.E.; Sun, H.; et al. Augmentation of <scp>BMP</Scp> Signaling in Cranial Neural Crest Cells Leads to Premature Cranial Sutures Fusion through Endochondral Ossification in Mice. JBMR Plus 2023, 7. [Google Scholar] [CrossRef]
- Ueharu, H.; Pan, H.; Hayano, S.; Zapien-Guerra, K.; Yang, J.; Mishina, Y. Augmentation of Bone Morphogenetic Protein Signaling in Cranial Neural Crest Cells in Mice Deforms Skull Base Due to Premature Fusion of Intersphenoidal Synchondrosis. genesis 2023, 61. [Google Scholar] [CrossRef] [PubMed]
- Akintoye, S.O.; Lam, T.; Shi, S.; Brahim, J.; Collins, M.T.; Robey, P.G. Skeletal Site-Specific Characterization of Orofacial and Iliac Crest Human Bone Marrow Stromal Cells in Same Individuals. Bone 2006, 38, 758–768. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, A.; Teo, N.; Poon, K.J.; Tiwari, P.; Ravichandran, A.; Wen, F.; Teoh, S.H.; Lim, T.C.; Toh, Y.-C. Comparative Craniofacial Bone Regeneration Capacities of Mesenchymal Stem Cells Derived from Human Neural Crest Stem Cells and Bone Marrow. ACS Biomater. Sci. Eng. 2021, 7, 207–221. [Google Scholar] [CrossRef] [PubMed]
- Solloway, M.J.; Robertson, E.J. Early Embryonic Lethality in Bmp5;Bmp7 Double Mutant Mice Suggests Functional Redundancy within the 60A Subgroup. Development 1999, 126, 1753–1768. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.Y.; Hill, C.S. TGF-β Superfamily Signaling in Embryonic Development and Homeostasis. Dev. Cell 2009, 16, 329–343. [Google Scholar] [CrossRef]
- Morrison, S.J.; Perez, S.E.; Qiao, Z.; Verdi, J.M.; Hicks, C.; Weinmaster, G.; Anderson, D.J. Transient Notch Activation Initiates an Irreversible Switch from Neurogenesis to Gliogenesis by Neural Crest Stem Cells. Cell 2000, 101, 499–510. [Google Scholar] [CrossRef]
- Lim, D.A.; Tramontin, A.D.; Trevejo, J.M.; Herrera, D.G.; García-Verdugo, J.M.; Alvarez-Buylla, A. Noggin Antagonizes BMP Signaling to Create a Niche for Adult Neurogenesis. Neuron 2000, 28, 713–726. [Google Scholar] [CrossRef]
- Gomes, W.A.; Mehler, M.F.; Kessler, J.A. Transgenic Overexpression of BMP4 Increases Astroglial and Decreases Oligodendroglial Lineage Commitment. Dev. Biol. 2003, 255, 164–177. [Google Scholar] [CrossRef]
- Winnier, G.; Blessing, M.; Labosky, P.A.; Hogan, B.L. Bone Morphogenetic Protein-4 Is Required for Mesoderm Formation and Patterning in the Mouse. Genes Dev. 1995, 9, 2105–2116. [Google Scholar] [CrossRef]
- Zhang, H.; Bradley, A. Mice Deficient for BMP2 Are Nonviable and Have Defects in Amnion/Chorion and Cardiac Development. Development 1996, 122, 2977–2986. [Google Scholar] [CrossRef]
- Dudley, A.T.; Lyons, K.M.; Robertson, E.J. A Requirement for Bone Morphogenetic Protein-7 during Development of the Mammalian Kidney and Eye. Genes Dev. 1995, 9, 2795–2807. [Google Scholar] [CrossRef] [PubMed]
- Luo, G.; Hofmann, C.; Bronckers, A.L.; Sohocki, M.; Bradley, A.; Karsenty, G. BMP-7 Is an Inducer of Nephrogenesis, and Is Also Required for Eye Development and Skeletal Patterning. Genes Dev. 1995, 9, 2808–2820. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Adachi, Y.; Numata, T.; Nakada, S.; Yanagita, M.; Nakagata, N.; Evans, S.M.; Graf, D.; Economides, A.; Haraguchi, R.; et al. Reduced BMP Signaling Results in Hindlimb Fusion with Lethal Pelvic/Urogenital Organ Aplasia: A New Mouse Model of Sirenomelia. PLoS One 2012, 7, e43453. [Google Scholar] [CrossRef] [PubMed]
- Correns, A.; Zimmermann, L.M.A.; Baldock, C.; Sengle, G. BMP Antagonists in Tissue Development and Disease. Matrix Biol. Plus 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Puerto, M.C.; Iyengar, P.V.; García de Vinuesa, A.; ten Dijke, P.; Sanchez-Duffhues, G. Bone Morphogenetic Protein Receptor Signal Transduction in Human Disease. J. Pathol. 2019, 247, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Duffhues, G.; Hiepen, C. Human IPSCs as Model Systems for BMP-Related Rare Diseases. Cells 2023, 12, 2200. [Google Scholar] [CrossRef]
- Tang, J.; Tan, M.; Liao, S.; Pang, M.; Li, J. Recent Progress in the Biology and Physiology of BMP-8a. Connect. Tissue Res. 2023, 64, 219–228. [Google Scholar] [CrossRef]
- Zhang, Y.; Que, J. BMP Signaling in Development, Stem Cells, and Diseases of the Gastrointestinal Tract. Annu. Rev. Physiol. 2020, 82, 251–273. [Google Scholar] [CrossRef]
- Chang, W.; Lin, Z.; Kulessa, H.; Hebert, J.; Hogan, B.L.M.; Wu, D.K. Bmp4 Is Essential for the Formation of the Vestibular Apparatus That Detects Angular Head Movements. PLoS Genet. 2008, 4, e1000050. [Google Scholar] [CrossRef]
- Morikawa, M.; Koinuma, D.; Mizutani, A.; Kawasaki, N.; Holmborn, K.; Sundqvist, A.; Tsutsumi, S.; Watabe, T.; Aburatani, H.; Heldin, C.-H.; et al. BMP Sustains Embryonic Stem Cell Self-Renewal through Distinct Functions of Different Krüppel-like Factors. Stem Cell Reports 2016, 6, 64–73. [Google Scholar] [CrossRef]
- Hirai, H.; Karian, P.; Kikyo, N. Regulation of Embryonic Stem Cell Self-Renewal and Pluripotency by Leukaemia Inhibitory Factor. Biochem. J. 2011, 438, 11–23. [Google Scholar] [CrossRef] [PubMed]
- TOMIZAWA, M.; SHINOZAKI, F.; SUGIYAMA, T.; YAMAMOTO, S.; SUEISHI, M.; YOSHIDA, T. Activin A Maintains Pluripotency Markers and Proliferative Potential of Human Induced Pluripotent Stem Cells. Exp. Ther. Med. 2011, 2, 405–408. [Google Scholar] [CrossRef] [PubMed]
- Tomizawa, M.; Shinozaki, F.; Sugiyama, T.; Yamamoto, S.; Sueishi, M.; Yoshida, T. Activin A Is Essential for Feeder-free Culture of Human Induced Pluripotent Stem Cells. J. Cell. Biochem. 2013, 114, 584–588. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, Y.; Hsiao, E.C.; Sami, S.; Lancero, M.; Schlieve, C.R.; Nguyen, T.; Yano, K.; Nagahashi, A.; Ikeya, M.; Matsumoto, Y.; et al. BMP-SMAD-ID Promotes Reprogramming to Pluripotency by Inhibiting P16/INK4A-Dependent Senescence. Proc. Natl. Acad. Sci. 2016, 113, 13057–13062. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, M.M.; Zimmerman, S.; Leeson, H.; Nefzger, C.M.; Mar, J.C.; Laslett, A.; Polo, J.M.; Wolvetang, E.; Cooper-White, J.J. Superior Induced Pluripotent Stem Cell Generation through Phactr3-Driven Mechanomodulation of Both Early and Late Phases of Cell Reprogramming. Biomater. Res. 2024, 28. [Google Scholar] [CrossRef]
- Sagara, J.; Higuchi, T.; Hattori, Y.; Moriya, M.; Sarvotham, H.; Shima, H.; Shirato, H.; Kikuchi, K.; Taniguchi, S. Scapinin, a Putative Protein Phosphatase-1 Regulatory Subunit Associated with the Nuclear Nonchromatin Structure. J. Biol. Chem. 2003, 278, 45611–45619. [Google Scholar] [CrossRef]
- Sagara, J.; Arata, T.; Taniguchi, S. Scapinin, the Protein Phosphatase 1 Binding Protein, Enhances Cell Spreading and Motility by Interacting with the Actin Cytoskeleton. PLoS One 2009, 4, e4247. [Google Scholar] [CrossRef]
- Boog, H.; Medda, R.; Cavalcanti-Adam, E.A. Single Cell Center of Mass for the Analysis of BMP Receptor Heterodimers Distributions. J. Imaging 2021, 7, 219. [Google Scholar] [CrossRef]
- Gabdoulline, R.; Kaisers, W.; Gaspar, A.; Meganathan, K.; Doss, M.X.; Jagtap, S.; Hescheler, J.; Sachinidis, A.; Schwender, H. Differences in the Early Development of Human and Mouse Embryonic Stem Cells. PLoS One 2015, 10, e0140803. [Google Scholar] [CrossRef]
- Rossant, J. Mouse and Human Blastocyst-Derived Stem Cells: Vive Les Differences. Development 2015, 142, 9–12. [Google Scholar] [CrossRef]
- Johnson Chacko, L.; Wertjanz, D.; Sergi, C.; Dudas, J.; Fischer, N.; Eberharter, T.; Hoermann, R.; Glueckert, R.; Fritsch, H.; Rask-Andersen, H.; et al. Growth and Cellular Patterning during Fetal Human Inner Ear Development Studied by a Correlative Imaging Approach. BMC Dev. Biol. 2019, 19, 11. [Google Scholar] [CrossRef] [PubMed]
- Richard, C.; Courbon, G.; Laroche, N.; Prades, J.M.; Vico, L.; Malaval, L. Inner Ear Ossification and Mineralization Kinetics in Human Embryonic Development - Microtomographic and Histomorphological Study. Sci. Rep. 2017, 7, 4825. [Google Scholar] [CrossRef] [PubMed]
- Kopecky, B.; Johnson, S.; Schmitz, H.; Santi, P.; Fritzsch, B. Scanning Thin-sheet Laser Imaging Microscopy Elucidates Details on Mouse Ear Development. Dev. Dyn. 2012, 241, 465–480. [Google Scholar] [CrossRef] [PubMed]
- Shnerson, A.; Pujol, R. Age-Related Changes in the C57BL/6J Mouse Cochlea. I. Physiological Findings. Dev. Brain Res. 1981, 2, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Wu, K.; Zhang, X.; Wang, H.; Wang, Q. Genetic Correction of Induced Pluripotent Stem Cells from a DFNA36 Patient Results in Morphologic and Functional Recovery of Derived Hair Cell-like Cells. Stem Cell Res. Ther. 2024, 15, 4. [Google Scholar] [CrossRef]
- Liu, X.; Lillywhite, J.; Zhu, W.; Huang, Z.; Clark, A.M.; Gosstola, N.; Maguire, C.T.; Dykxhoorn, D.; Chen, Z.-Y.; Yang, J. Generation and Genetic Correction of USH2A c.2299delG Mutation in Patient-Derived Induced Pluripotent Stem Cells. Genes (Basel). 2021, 12, 805. [Google Scholar] [CrossRef]
- Sanjurjo-Soriano, C.; Erkilic, N.; Baux, D.; Mamaeva, D.; Hamel, C.P.; Meunier, I.; Roux, A.-F.; Kalatzis, V. Genome Editing in Patient IPSCs Corrects the Most Prevalent USH2A Mutations and Reveals Intriguing Mutant MRNA Expression Profiles. Mol. Ther. - Methods Clin. Dev. 2020, 17, 156–173. [Google Scholar] [CrossRef]
- Chen, C.; Guan, M.-X. Genetic Correction of TRMU Allele Restored the Mitochondrial Dysfunction-Induced Deficiencies in IPSCs-Derived Hair Cells of Hearing-Impaired Patients. Hum. Mol. Genet. 2022, 31, 3068–3082. [Google Scholar] [CrossRef]
- Liu, X.; Wen, J.; Liu, X.; Chen, A.; Li, S.; Liu, J.; Sun, J.; Gong, W.; Kang, X.; Feng, Z.; et al. Gene Regulation Analysis of Patient-Derived IPSCs and Its CRISPR-Corrected Control Provides a New Tool for Studying Perturbations of ELMOD3 c.512A>G Mutation during the Development of Inherited Hearing Loss. PLoS One 2023, 18, e0288640. [Google Scholar] [CrossRef]
- Tang, Z.-H.; Chen, J.-R.; Zheng, J.; Shi, H.-S.; Ding, J.; Qian, X.-D.; Zhang, C.; Chen, J.-L.; Wang, C.-C.; Li, L.; et al. Genetic Correction of Induced Pluripotent Stem Cells From a Deaf Patient With MYO7A Mutation Results in Morphologic and Functional Recovery of the Derived Hair Cell-Like Cells. Stem Cells Transl. Med. 2016, 5, 561–571. [Google Scholar] [CrossRef]
- Qiu, Y.; Wang, H.; Fan, M.; Pan, H.; Guan, J.; Jiang, Y.; Jia, Z.; Wu, K.; Zhou, H.; Zhuang, Q.; et al. Impaired AIF-CHCHD4 Interaction and Mitochondrial Calcium Overload Contribute to Auditory Neuropathy Spectrum Disorder in Patient-IPSC-Derived Neurons with AIFM1 Variant. Cell Death Dis. 2023, 14, 375. [Google Scholar] [CrossRef] [PubMed]
- Doetzlhofer, A.; White, P.M.; Johnson, J.E.; Segil, N.; Groves, A.K. In Vitro Growth and Differentiation of Mammalian Sensory Hair Cell Progenitors: A Requirement for EGF and Periotic Mesenchyme. Dev. Biol. 2004, 272, 432–447. [Google Scholar] [CrossRef] [PubMed]
- Huh, S.-H.; Warchol, M.E.; Ornitz, D.M. Cochlear Progenitor Number Is Controlled through Mesenchymal FGF Receptor Signaling. Elife 2015, 4. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Zuo, N.; Wu, C.; Chen, P.; Ma, J.; Wang, C.; Li, W.; Liu, S. Role of the Periotic Mesenchyme in the Development of Sensory Cells in Early Mouse Cochlea. J. Otol. 2020, 15, 138–143. [Google Scholar] [CrossRef] [PubMed]
- Rose, K.P.; Manilla, G.; Milon, B.; Zalzman, O.; Song, Y.; Coate, T.M.; Hertzano, R. Spatially Distinct Otic Mesenchyme Cells Show Molecular and Functional Heterogeneity Patterns before Hearing Onset. iScience 2023, 26, 107769. [Google Scholar] [CrossRef]
- Moeinvaziri, F.; Shojaei, A.; Haghparast, N.; Yakhkeshi, S.; Nemati, S.; Hassani, S.-N.; Baharvand, H. Towards Maturation of Human Otic Hair Cell–like Cells in Pluripotent Stem Cell–Derived Organoid Transplants. Cell Tissue Res. 2021, 386, 321–333. [Google Scholar] [CrossRef]
- Xia, M.; Ma, J.; Wu, M.; Guo, L.; Chen, Y.; Li, G.; Sun, S.; Chai, R.; Li, H.; Li, W. Generation of Innervated Cochlear Organoid Recapitulates Early Development of Auditory Unit. Stem Cell Reports 2023, 18, 319–336. [Google Scholar] [CrossRef]
- Lu, J.; Wang, M.; Meng, Y.; An, W.; Wang, X.; Sun, G.; Wang, H.; Liu, W. Current Advances in Biomaterials for Inner Ear Cell Regeneration. Front. Neurosci. 2024, 17. [Google Scholar] [CrossRef]
- Pressé, M.T.; Malgrange, B.; Delacroix, L. The Cochlear Matrisome: Importance in Hearing and Deafness. Matrix Biol. 2024, 125, 40–58. [Google Scholar] [CrossRef]
- Ma, J.; You, D.; Li, W.; Lu, X.; Sun, S.; Li, H. Bone Morphogenetic Proteins and Inner Ear Development. J. Zhejiang Univ. B 2019, 20, 131–145. [Google Scholar] [CrossRef]
- Saeki, T.; Yoshimatsu, S.; Ishikawa, M.; Hon, C.-C.; Koya, I.; Shibata, S.; Hosoya, M.; Saegusa, C.; Ogawa, K.; Shin, J.W.; et al. Critical Roles of FGF, RA, and WNT Signalling in the Development of the Human Otic Placode and Subsequent Lineages in a Dish. Regen. Ther. 2022, 20, 165–186. [Google Scholar] [CrossRef] [PubMed]
- Maharana, S.K.; Saint-Jeannet, J.-P. Molecular Mechanisms of Hearing Loss in Nager Syndrome. Dev. Biol. 2021, 476, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Sher, G.; Naeem, M. A Novel CHSY1 Gene Mutation Underlies Temtamy Preaxial Brachydactyly Syndrome in a Pakistani Family. Eur. J. Med. Genet. 2014, 57, 21–24. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Laue, K.; Temtamy, S.; Aglan, M.; Kotan, L.D.; Yigit, G.; Canan, H.; Pawlik, B.; Nürnberg, G.; Wakeling, E.L.; et al. Temtamy Preaxial Brachydactyly Syndrome Is Caused by Loss-of-Function Mutations in Chondroitin Synthase 1, a Potential Target of BMP Signaling. Am. J. Hum. Genet. 2010, 87, 757–767. [Google Scholar] [CrossRef] [PubMed]
- Lyu, Z.; Da, Y.; Liu, H.; Wang, Z.; Zhu, Y.; Tian, J. Chsy1 Deficiency Reduces Extracellular Matrix Productions and Aggravates Cartilage Injury in Osteoarthritis. Gene 2022, 827, 146466. [Google Scholar] [CrossRef] [PubMed]
- Carlson, R.J.; Quesnel, A.; Wells, D.; Brownstein, Z.; Gilony, D.; Gulsuner, S.; Leppig, K.A.; Avraham, K.B.; King, M.-C.; Walsh, T.; et al. Genetic Heterogeneity and Core Clinical Features of NOG-Related-Symphalangism Spectrum Disorder. Otol. Neurotol. 2021, 42, e1143–e1151. [Google Scholar] [CrossRef] [PubMed]
- Hirshoren, N.; Gross, M.; Banin, E.; Sosna, J.; Bargal, R.; Raas-Rothschild, A. P35S Mutation in the NOG Gene Associated with Teunissen–Cremers Syndrome and Features of Multiple NOG Joint-Fusion Syndromes. Eur. J. Med. Genet. 2008, 51, 351–357. [Google Scholar] [CrossRef]
- Ishino, T.; Takeno, S.; Hirakawa, K. Novel NOG Mutation in Japanese Patients with Stapes Ankylosis with Broad Thumbs and Toes. Eur. J. Med. Genet. 2015, 58, 427–432. [Google Scholar] [CrossRef]
- Masuda, S.; Namba, K.; Mutai, H.; Usui, S.; Miyanaga, Y.; Kaneko, H.; Matsunaga, T. A Mutation in the Heparin-Binding Site of Noggin as a Novel Mechanism of Proximal Symphalangism and Conductive Hearing Loss. Biochem. Biophys. Res. Commun. 2014, 447, 496–502. [Google Scholar] [CrossRef]
- Nakashima, T.; Ganaha, A.; Tsumagari, S.; Nakamura, T.; Yamada, Y.; Nakamura, E.; Usami, S.; Tono, T. Is the Conductive Hearing Loss in NOG-Related Symphalangism Spectrum Disorder Congenital? ORL 2021, 83, 196–202. [Google Scholar] [CrossRef]
- Pang, X.; Wang, Z.; Chai, Y.; Chen, H.; Li, L.; Sun, L.; Jia, H.; Wu, H.; Yang, T. A Novel Missense Mutation of NOG Interferes With the Dimerization of NOG and Causes Proximal Symphalangism Syndrome in a Chinese Family. Ann. Otol. Rhinol. Laryngol. 2015, 124, 745–751. [Google Scholar] [CrossRef] [PubMed]
- Sonoyama, T.; Ishino, T.; Ogawa, Y.; Oda, T.; Takeno, S. Identification of a Novel Nonsense NOG Mutation in a Patient with Stapes Ankylosis and Symphalangism Spectrum Disorder. Hum. Genome Var. 2023, 10, 12. [Google Scholar] [CrossRef] [PubMed]
- Takano, K.; Ogasawara, N.; Matsunaga, T.; Mutai, H.; Sakurai, A.; Ishikawa, A.; Himi, T. A Novel Nonsense Mutation in the NOG Gene Causes Familial NOG-Related Symphalangism Spectrum Disorder. Hum. Genome Var. 2016, 3, 16023. [Google Scholar] [CrossRef] [PubMed]
- Thomeer, H.G.X.M.; Admiraal, R.J.C.; Hoefsloot, L.; Kunst, H.P.M.; Cremers, C.W.R.J. Proximal Symphalangism, Hyperopia, Conductive Hearing Impairment, and the NOG Gene. Otol. Neurotol. 2011, 32, 632–638. [Google Scholar] [CrossRef] [PubMed]
- Yu, R.; Jiang, H.; Liao, H.; Luo, W. Genetic and Clinical Phenotypic Analysis of Familial Stapes Sclerosis Caused by an NOG Mutation. BMC Med. Genomics 2020, 13, 187. [Google Scholar] [CrossRef]
- Caputo, V.; Bocchinfuso, G.; Castori, M.; Traversa, A.; Pizzuti, A.; Stella, L.; Grammatico, P.; Tartaglia, M. Novel SMAD4 Mutation Causing Myhre Syndrome. Am. J. Med. Genet. Part A 2014, 164, 1835–1840. [Google Scholar] [CrossRef]
- Cătană, A.; Simonescu-Colan, R.; Cuzmici-Barabaș, Z.; Militaru, D.; Iordănescu, I.; Militaru, M. First Documented Case of Myhre Syndrome in Romania: A Case Report. Exp. Ther. Med. 2022, 23, 323. [Google Scholar] [CrossRef]
- Le Goff, C.; Mahaut, C.; Abhyankar, A.; Le Goff, W.; Serre, V.; Afenjar, A.; Destrée, A.; di Rocco, M.; Héron, D.; Jacquemont, S.; et al. Mutations at a Single Codon in Mad Homology 2 Domain of SMAD4 Cause Myhre Syndrome. Nat. Genet. 2012, 44, 85–88. [Google Scholar] [CrossRef]
- Lin, A.E.; Alali, A.; Starr, L.J.; Shah, N.; Beavis, A.; Pereira, E.M.; Lindsay, M.E.; Klugman, S. Gain-of-function Pathogenic Variants in SMAD4 Are Associated with Neoplasia in Myhre Syndrome. Am. J. Med. Genet. Part A 2020, 182, 328–337. [Google Scholar] [CrossRef]
- Michot, C.; Le Goff, C.; Mahaut, C.; Afenjar, A.; Brooks, A.S.; Campeau, P.M.; Destree, A.; Di Rocco, M.; Donnai, D.; Hennekam, R.; et al. Myhre and LAPS Syndromes: Clinical and Molecular Review of 32 Patients. Eur. J. Hum. Genet. 2014, 22, 1272–1277. [Google Scholar] [CrossRef]
- Yang, K.; Wang, X.; Wang, W.; Han, M.; Hu, L.; Kang, D.; Yang, J.; Liu, M.; Gao, X.; Yuan, Y.; et al. A Newborn Male with Myhre Syndrome, Hearing Loss, and Complete Syndactyly of Fingers 3–4. Mol. Genet. Genomic Med. 2023, 11. [Google Scholar] [CrossRef] [PubMed]
- Caputo, V.; Cianetti, L.; Niceta, M.; Carta, C.; Ciolfi, A.; Bocchinfuso, G.; Carrani, E.; Dentici, M.L.; Biamino, E.; Belligni, E.; et al. A Restricted Spectrum of Mutations in the SMAD4 Tumor-Suppressor Gene Underlies Myhre Syndrome. Am. J. Hum. Genet. 2012, 90, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Iida, A.; Bhavani, G.S.; Gowrishankar, K.; Wang, Z.; Xue, J.; Wang, J.; Miyake, N.; Matsumoto, N.; Hasegawa, T.; et al. Deficiency of TMEM53 Causes a Previously Unknown Sclerosing Bone Disorder by Dysregulation of BMP-SMAD Signaling. Nat. Commun. 2021, 12, 2046. [Google Scholar] [CrossRef] [PubMed]
- Whyte, M.P.; Weinstein, R.S.; Phillips, P.H.; McAlister, W.H.; Ramakrishnaiah, R.H.; Schaefer, G.B.; Cai, R.; Hutchison, M.R.; Duan, S.; Gottesman, G.S.; et al. Transmembrane Protein 53 Craniotubular Dysplasia (OMIM # 619727): The Skeletal Disease and Consequent Blindness of This New Disorder. Bone 2024, 188, 117218. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, F.S.; Xu, M.; Seemann, P.; Connor, J.M.; Glaser, D.L.; Carroll, L.; Delai, P.; Fastnacht-Urban, E.; Forman, S.J.; Gillessen-Kaesbach, G.; et al. Classic and Atypical Fibrodysplasia Ossificans Progressiva (FOP) Phenotypes Are Caused by Mutations in the Bone Morphogenetic Protein (BMP) Type I Receptor ACVR1. Hum. Mutat. 2009, 30, 379–390. [Google Scholar] [CrossRef]
- Kaplan, F.S.; Kobori, J.A.; Orellana, C.; Calvo, I.; Rosello, M.; Martinez, F.; Lopez, B.; Xu, M.; Pignolo, R.J.; Shore, E.M.; et al. Multi-system Involvement in a Severe Variant of Fibrodysplasia Ossificans Progressiva ( ACVR1 c.772G>A; R258G): A Report of Two Patients. Am. J. Med. Genet. Part A 2015, 167, 2265–2271. [Google Scholar] [CrossRef]
- Levy, C.E.; Lash, A.T.; Janoff, H.B.; Kaplan, F.S. Conductive Hearing Loss in Individuals with Fibrodysplasia Ossificans Progressiva. Am. J. Audiol. 1999, 8, 29–33. [Google Scholar] [CrossRef]
- Hasegawa, K.; Tanaka, H.; Futagawa, N.; Miyahara, H.; Tsukahara, H. Rapid Progression of Heterotopic Ossification in Severe Variant of Fibrodysplasia Ossificans Progressiva with p.Arg258Gly in ACVR1: A Case Report and Review of Clinical Phenotypes. Case Rep. Genet. 2022, 2022, 1–6. [Google Scholar] [CrossRef]
- Ealy, M.; Meyer, N.C.; Corchado, J.C.; Schrauwen, I.; Bress, A.; Pfister, M.; Van Camp, G.; Smith, R.J.H. Rare Variants in BMP2 and BMP4 Found in Otosclerosis Patients Reduce Smad Signaling. Otol. Neurotol. 2014, 35, 395–400. [Google Scholar] [CrossRef]
- Hansdah, K.; Singh, N.; Bouzid, A.; Priyadarshi, S.; Ray, C.S.; Desai, A.; Panda, K.C.; Choudhury, J.C.; Biswal, N.C.; Tekari, A.; et al. Evaluation of the Genetic Association and MRNA Expression of the COL1A1, BMP2, and BMP4 Genes in the Development of Otosclerosis. Genet. Test. Mol. Biomarkers 2020, 24, 343–351. [Google Scholar] [CrossRef]
- Priestley, J.R.C.; Deshwar, A.R.; Murthy, H.; D’Agostino, M.D.; Dupuis, L.; Gangaram, B.; Gray, C.; Jobling, R.; Pannia, E.; Platzer, K.; et al. Monoallelic Loss-of-Function BMP2 Variants Result in BMP2-Related Skeletal Dysplasia Spectrum. Genet. Med. 2023, 25, 100863. [Google Scholar] [CrossRef] [PubMed]
- Schrauwen, I.; Thys, M.; Vanderstraeten, K.; Fransen, E.; Dieltjens, N.; Huyghe, J.R.; Ealy, M.; Claustres, M.; Cremers, C.R.; Dhooge, I.; et al. Association of Bone Morphogenetic Proteins With Otosclerosis. J. Bone Miner. Res. 2008, 23, 507–516. [Google Scholar] [CrossRef] [PubMed]
- Tan, T.Y.; Gonzaga-Jauregui, C.; Bhoj, E.J.; Strauss, K.A.; Brigatti, K.; Puffenberger, E.; Li, D.; Xie, L.; Das, N.; Skubas, I.; et al. Monoallelic BMP2 Variants Predicted to Result in Haploinsufficiency Cause Craniofacial, Skeletal, and Cardiac Features Overlapping Those of 20p12 Deletions. Am. J. Hum. Genet. 2017, 101, 985–994. [Google Scholar] [CrossRef]
- Nixon, T.R.W.; Richards, A.; Towns, L.K.; Fuller, G.; Abbs, S.; Alexander, P.; McNinch, A.; Sandford, R.N.; Snead, M.P. Bone Morphogenetic Protein 4 (BMP4) Loss-of-Function Variant Associated with Autosomal Dominant Stickler Syndrome and Renal Dysplasia. Eur. J. Hum. Genet. 2019, 27, 369–377. [Google Scholar] [CrossRef] [PubMed]
- Wyatt, A.W.; Osborne, R.J.; Stewart, H.; Ragge, N.K. Bone Morphogenetic Protein 7 (BMP7) Mutations Are Associated with Variable Ocular, Brain, Ear, Palate, and Skeletal Anomalies. Hum. Mutat. 2010, 31, 781–787. [Google Scholar] [CrossRef] [PubMed]
- Adeyemo, A.; Faridi, R.; Chattaraj, P.; Yousaf, R.; Tona, R.; Okorie, S.; Bharadwaj, T.; Nouel-Saied, L.M.; Acharya, A.; Schrauwen, I.; et al. Genomic Analysis of Childhood Hearing Loss in the Yoruba Population of Nigeria. Eur. J. Hum. Genet. 2022, 30, 42–52. [Google Scholar] [CrossRef]
- Driesen, J.; Van Hoecke, H.; Maes, L.; Janssens, S.; Acke, F.; De Leenheer, E. CHD7 Disorder—Not CHARGE Syndrome—Presenting as Isolated Cochleovestibular Dysfunction. Genes (Basel). 2024, 15, 643. [Google Scholar] [CrossRef]
- Roux, I.; Fenollar-Ferrer, C.; Lee, H.J.; Chattaraj, P.; Lopez, I.A.; Han, K.; Honda, K.; Brewer, C.C.; Butman, J.A.; Morell, R.J.; et al. CHD7 Variants Associated with Hearing Loss and Enlargement of the Vestibular Aqueduct. Hum. Genet. 2023, 142, 1499–1517. [Google Scholar] [CrossRef]
- Akahira-Azuma, M.; Enomoto, Y.; Nakamura, N.; Yokoi, T.; Minatogawa, M.; Harada, N.; Tsurusaki, Y.; Kurosawa, K. Novel COL2A1 Variants in Japanese Patients with Spondyloepiphyseal Dysplasia Congenita. Hum. Genome Var. 2022, 9, 16. [Google Scholar] [CrossRef]
- Terhal, P.A.; Nievelstein, R.J.A.J.; Verver, E.J.J.; Topsakal, V.; van Dommelen, P.; Hoornaert, K.; Le Merrer, M.; Zankl, A.; Simon, M.E.H.; Smithson, S.F.; et al. A Study of the Clinical and Radiological Features in a Cohort of 93 Patients with a COL2A1 Mutation Causing Spondyloepiphyseal Dysplasia Congenita or a Related Phenotype. Am. J. Med. Genet. Part A 2015, 167, 461–475. [Google Scholar] [CrossRef]
- Wu, K.; Li, Z.; Zhu, Y.; Wang, X.; Chen, G.; Hou, Z.; Zhang, Q. Discovery of Sensorineural Hearing Loss and Ossicle Deformity in a Chinese Li Nationality Family with Spondyloepiphyseal Dysplasia Congenita Caused by p.G504S Mutation of COL2A1. BMC Med. Genomics 2021, 14, 170. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Tabouni, M.; Kizhakkedath, P.; Baydoun, I.; Allam, M.; John, A.; Busafared, F.; Alnuaimi, A.; Al-Jasmi, F.; Alblooshi, H. Spectrum of Genetic Variants in Bilateral Sensorineural Hearing Loss. Front. Genet. 2024, 15. [Google Scholar] [CrossRef] [PubMed]
- Clarke, R.A.; Fang, Z.; Murrell, D.; Sheriff, T.; Eapen, V. GDF6 Knockdown in a Family with Multiple Synostosis Syndrome and Speech Impairment. Genes (Basel). 2021, 12, 1354. [Google Scholar] [CrossRef] [PubMed]
- Terhal, P.A.; Verbeek, N.E.; Knoers, N.; Nievelstein, R.J.A.J.; van den Ouweland, A.; Sakkers, R.J.; Speleman, L.; van Haaften, G. Further Delineation of the GDF6 Related Multiple Synostoses Syndrome. Am. J. Med. Genet. Part A 2018, 176, 225–229. [Google Scholar] [CrossRef]
- Wang, J.; Yu, T.; Wang, Z.; Ohte, S.; Yao, R.; Zheng, Z.; Geng, J.; Cai, H.; Ge, Y.; Li, Y.; et al. A New Sub type of Multiple Synostoses Syndrome Is Caused by a Mutation in GDF6 That Decreases Its Sensitivity to Noggin and Enhances Its Potency as a BMP Signal. J. Bone Miner. Res. 2016, 31, 882–889. [Google Scholar] [CrossRef]
Figure 1.
Effect of BMP concentration on fate of ectodermal cells to induce epidermal, placodal, neural crest, and neural derivatives. Exposure of pluripotent stem cells to different concentrations of BMP4 results in differentiation towards different cell fates via activation of various downstream genes. High concentration of BMP4 results in activation of genes such as K18, which causes cells to differentiate towards epidermal fate. Medium concentration of BMP4, which can be due to the presence of some inhibitors such as Noggin, causes activation of SIX1 for differentiation towards pre-placodal ectoderm and subsequent placodal lineages including lens, olfactory, and otic placodes. However, in the presence of WNT, neural crest fate is induced. Activation of genes such as Hes1 can have an inhibitory effect this pathway. Low concentration of BMP4, which can be the result of high levels of Noggin due to Shh signalling or the presence of Chordin, results in activation of PAX6, NCAD, and other genes that results in neural fate. (Generated using Biorender accessed on Setpember 2024.).
Figure 1.
Effect of BMP concentration on fate of ectodermal cells to induce epidermal, placodal, neural crest, and neural derivatives. Exposure of pluripotent stem cells to different concentrations of BMP4 results in differentiation towards different cell fates via activation of various downstream genes. High concentration of BMP4 results in activation of genes such as K18, which causes cells to differentiate towards epidermal fate. Medium concentration of BMP4, which can be due to the presence of some inhibitors such as Noggin, causes activation of SIX1 for differentiation towards pre-placodal ectoderm and subsequent placodal lineages including lens, olfactory, and otic placodes. However, in the presence of WNT, neural crest fate is induced. Activation of genes such as Hes1 can have an inhibitory effect this pathway. Low concentration of BMP4, which can be the result of high levels of Noggin due to Shh signalling or the presence of Chordin, results in activation of PAX6, NCAD, and other genes that results in neural fate. (Generated using Biorender accessed on Setpember 2024.).
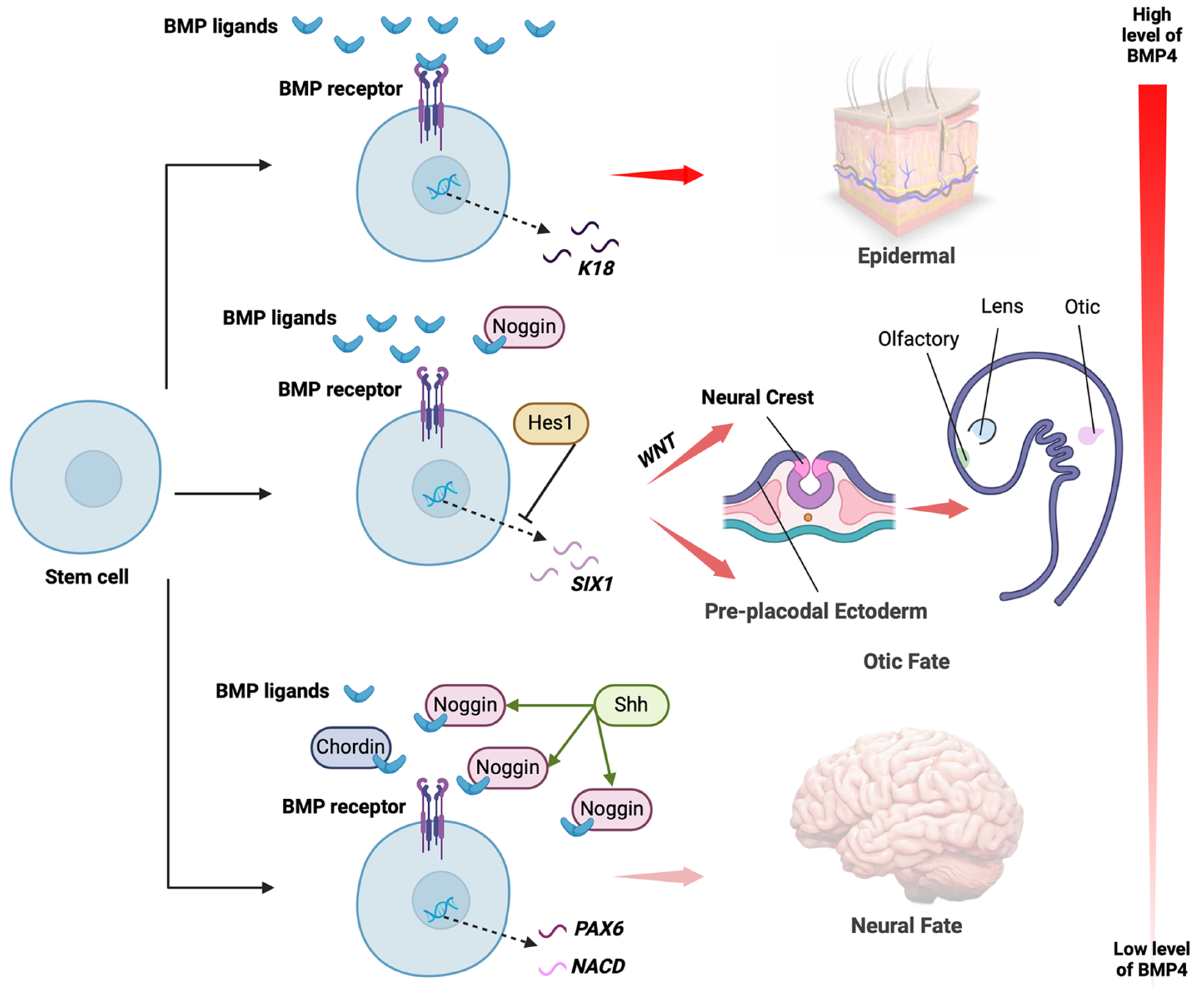
Figure 2.
Overview of BMP signalling pathway and modulators during development of pre-placodal ectoderm. BMPs such as BMP4 bind to their receptors BMPR1A, BMPR1B, and BMPR2 on the cell surface, resulting in activation of SMADs which translocate to the nucleus to influence transcription of genes controlling cell fate towards non-neural ectoderm/pre-placodal fate, and inhibiting differentiation towards neural fate. The presence of antagonists and modulators such as Noggin, Follistatin, Chordin, Gremlin, and TWSG1 alters the level of BMP activity on the cell and hence can also influence cell fate. (Generated using Biorender accessed on September 2024.).
Figure 2.
Overview of BMP signalling pathway and modulators during development of pre-placodal ectoderm. BMPs such as BMP4 bind to their receptors BMPR1A, BMPR1B, and BMPR2 on the cell surface, resulting in activation of SMADs which translocate to the nucleus to influence transcription of genes controlling cell fate towards non-neural ectoderm/pre-placodal fate, and inhibiting differentiation towards neural fate. The presence of antagonists and modulators such as Noggin, Follistatin, Chordin, Gremlin, and TWSG1 alters the level of BMP activity on the cell and hence can also influence cell fate. (Generated using Biorender accessed on September 2024.).
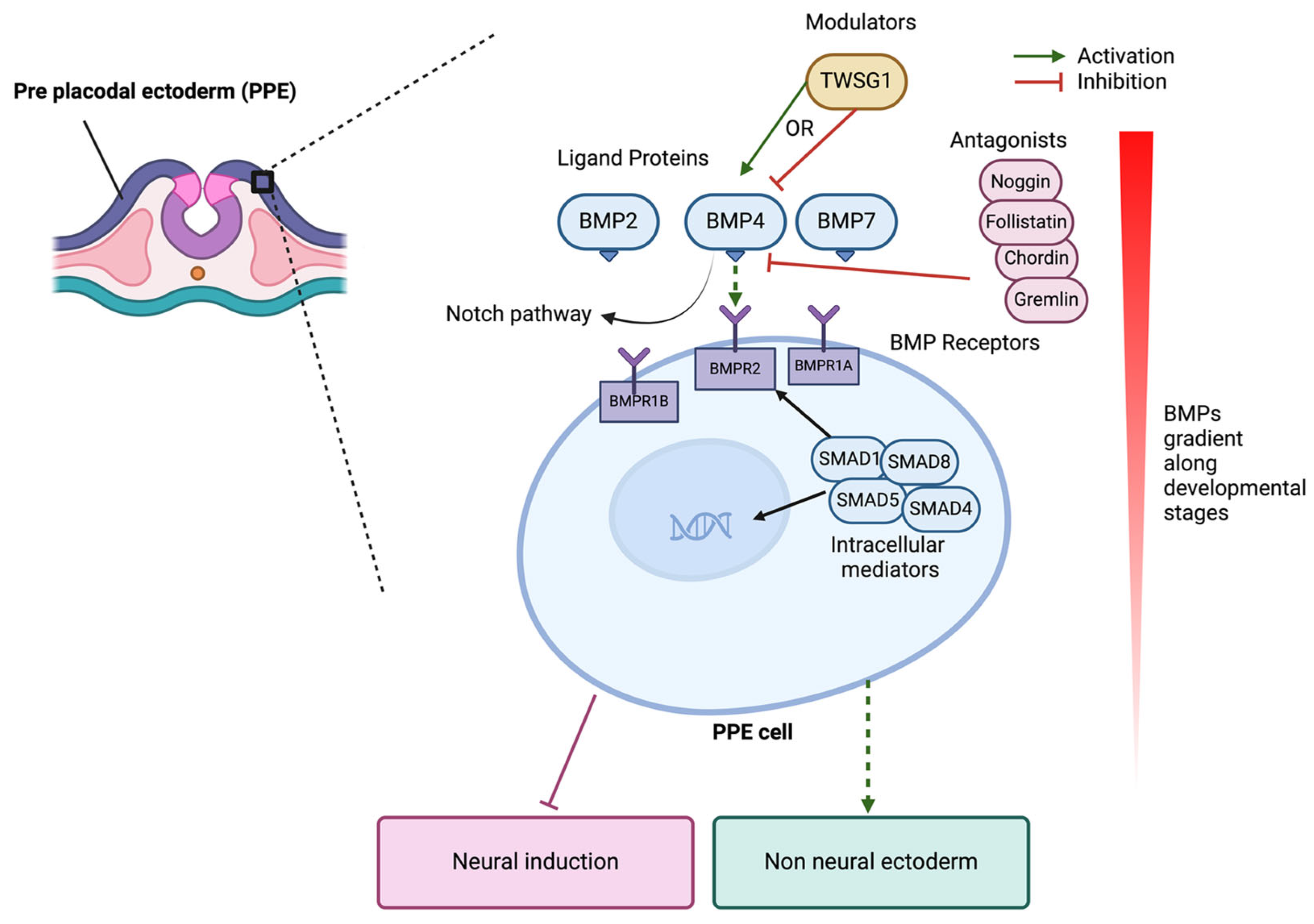

Table 1.
Genes involved in BMP signalling that are associated with hearing loss in humans.
| Gene | Role in BMP signalling | Inner ear deficits | Additional symptoms | References |
|---|---|---|---|---|
| ACVR1 (Activin A receptor type 1) | Type 1 BMP receptor | Sensorineural hearing loss; Conductive hearing loss | Bone and skeletal disorders | [185] |
| BMP2 | BMP ligand | Conductive hearing loss (otosclerosis) | Craniofacial, cardiac, and skeletal anomalies | [186,187,188] |
| BMP4 | BMP ligand | Sensorineural hearing loss; Conductive hearing loss (otosclerosis) | Eye, joint, and craniofacial disorders, renal dysplasia | [153] |
| BMP7 | BMP ligand | Sensorineural hearing loss | Eye anomalies, developmental delay, scoliosis, cleft palate | [189,190,191] |
| CHD7 | Promotes Col2a1 expression; Regulation of BMPR1B expression | Sensorineural hearing loss; some conductive hearing loss due to enlargement of vestibular aqueduct | Vestibular dysfunctions, hypogonadotropic hypogonadism | [192,193,194,195] |
| CHSY1 (Chondroitin synthase 1) | BMP inhibition | Sensorineural hearing loss | Facial dysmorphism, dental anomalies, digital anomalies, delayed motor development, delayed mental development, growth retardation | [17,18,20,24] |
| COL2A1 | Binds BMP | Sensorineural hearing loss | Short stature, bone and joint dysplasias, ocular problems | [24] |
| GDF6 (Growth and differentiation factor 6) | Forms heterodimers with BMPs | Conductive hearing loss (otosclerosis); Cochlear aplasia | Wrist and ankle deformities, tarsal-carpal fusion, vertebral fusion, speech impairment | [156,157,158,159,160,161,162,163,164,165] |
| NOG (Noggin) | BMP antagonist | Conductive hearing loss (stapes ankylosis and incus short process fixation) | Bone and joint disorders, digital and eye anomalies | [173,174] |
| SF3B4 (Splicing factor 3B subunit 4) | Spliceosome that affects Noggin and BMP expression | Conductive, sensorineural, and mixed hearing loss | Craniofacial defects, limb defects | [175,176,177,178] |
| SMAD4 | Downstream effector of BMP signalling | Conductive, sensorineural, and mixed hearing loss | Short stature, facial dysmorphism, muscular hypertrophy, cognitive delay | [179,180,181,182,183] |
| TMEM53 (Transmembrane protein 53) | Inhibits BMP/SMAD signalling | Sensorineural hearing loss | Bone and eye disorders | [179,180,182,184] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



