
Submitted:
12 September 2024
Posted:
13 September 2024
You are already at the latest version
Abstract
Background/Objectives: Chemerin, which is a multifunctional cytokine and adipokine, has been implicated in inflammatory and metabolic processes and might play a role in upper gastrointes-tinal (GI) malignancies, particularly gastric and esophageal cancer. The aim of this review is to explore the role of chemerin in the pathophysiology of upper GI cancers, as well as its potential as a biomarker for early detection and as a therapeutic target. Methods: A comprehensive review of recent studies about chemerin's biochemical properties and interaction with its receptors, as well as its effects on inflammatory responses, immune regulation and metabolic processes was con-ducted. Clinical implications of chemerin on gastric and esophageal cancer were analyzed, whereas the potential therapeutic strategies targeting chemerin were discussed. Results: Elevated chemerin levels are associated with poor prognosis in gastric cancer and promote invasiveness and metastasis in esophageal cancer. Chemerin receptor antagonists show promising results in inhibiting cancer cell migration, invasion and progression. Conclusions: Chemerin could repre-sent a valuable prognostic biomarker and therapeutic target for upper GI cancers. Future observa-tional studies should validate its clinical applications and investigate the efficacy of chemerin in-hibitors as potential therapeutic targets.
Keywords:
Chemerin
; Gastric cancer
; Esophageal cancer
; Obesity
; Biomarker
1. Introduction
1.1. Overview of Chemerin as a Multifunctional Cytokine and Adipokine
Chemerin, also known as retinoic acid receptor responder 2 (RARRES2), is a 163 amino acid protein, first identified in psoriatic skin lesions as a tazarotene-induced gene 2 protein (TIG2), and a few years later described as a natural ligand for chemokine-like receptor 1 (CMKLR1, previously known as ChemR23) [1]. Chemerin is widely found in endocrine, muscle, skin, lung, liver, and pancreas [2].
1.2. Rationale for Exploring Chemerin in the Context of Upper GI Cancers
Upper gastrointestinal (GI) cancers, specifically gastric and esophageal cancer, are significant global health burdens, particularly due to their high morbidity and mortality. These malignancies are closely linked with various modifiable risk factors, including dietary habits, smoking, and notably, obesity. Obesity, a major public health issue worldwide, is a known risk factor for several types of cancer, including those of the upper GI tract. The interplay between obesity and upper GI cancers is mediated by complex pathophysiologic mechanisms involving inflammation, insulin resistance and hormonal disruption. This connection emphasizes the need for advancing preventive strategies, early detection and effective therapeutic approaches. Research focused into biomarkers such as chemerin, which is implicated in both metabolic regulation and inflammation, could provide critical insights into the pathophysiology of upper GI cancers and offer new opportunities for targeted therapies, potentially improving outcomes for cancer patients.
The involvement of chemerin in upper GI cancers is of particular interest due to the association of these cancers with inflammatory and metabolic processes. The dual role of chemerin in modulating immune responses and metabolic pathways could potentially demonstrate an association between obesity, which is a known risk factor for upper GI cancers, and tumor development [3]. This makes chemerin a valuable biomarker for early detection, but also a promising novel target for therapeutic intervention. By targeting chemerin, the inflammatory and metabolic cascades, which contribute to tumor growth, might be disrupted. Therefore, investigating metabolic pathways of chemerin regarding upper GI tumor interaction, is critical to highlight its potential role in affecting disease progression and enhancing therapeutic outcomes for patients with such cancers.
2. Chemerin: Biochemistry and General Functions
2.1. Biochemical Properties and Synthesis of Chemerin
Chemerin is initially synthesized as a 163-amino acid precursor, preprochemerin, which undergoes proteolytic cleavage to remove a 20-amino acid N-terminal signal peptide, yielding a 143-amino acid protein, known as prochemerin. This precursor protein is further processed by several serine and cysteine proteases, resulting in different bioactive isoforms, including chemerinS157, chemerinF156 and others. These isoforms differ in their affinity for the chemerin receptors, particularly CMKLR1, with chemerinS157 and chemerinF156 exhibiting the highest binding affinity. The proteolytic processing of chemerin regulates its biological activity and also determines its function in different tissues. For example, chemerin in adipose tissue is crucial for adipogenesis and energy metabolism, whereas in the skin, it is involved in inflammatory responses and antimicrobial defense [4,5].
2.2. Main Receptors (CMKLR1, CCRL2, GPR1) and Signaling Pathways
Chemerin exerts its biological effects primarily by interacting with three G protein-coupled receptors (GPCRs): Chemokine-like receptor 1 (CMKLR1), C-C chemokine receptor-like 2 (CCRL2), and G protein-coupled receptor 1 (GPR1). Each of these receptors plays a unique role in mediating chemerin’s physiological processes and functions across different tissues.
First of all, chemokine-like receptor 1 (CMKLR1), is the most well-recognized receptor for chemerin. It is expressed in a variety of tissues, including adipose tissue, immune cells and endothelial cells [6,7]. Upon binding to chemerin, CMKLR1 activates several intracellular signaling pathways. These include the mobilization of Ca2+, inhibition of Cyclic Adenosine monophosphate (cAMP) accumulation and phosphorylation of Mitogen-Activated Protein Kinases (MAPKs), such as p42/p44 and p38 MAPK. These signaling events lead to diverse cellular responses, including chemotaxis, adipogenesis and modulation of inflammatory processes [7,8].
On the other hand, C-C chemokine receptor-like 2 (CCRL2) does not induce traditional GPCR signaling pathways like Ca2+ mobilization or cAMP inhibition upon chemerin binding. Instead, CCRL2 acts as a non-signaling “decoy” receptor that binds chemerin without initiating signal transduction [9]. This receptor is primarily involved in presenting chemerin to cells expressing CMKLR1, thereby enhancing local chemerin concentrations and facilitating CMKLR1-mediated responses [9,10]. CCRL2 is expressed in leukocytes, endothelial cells, and other immune cells; thus, its role is particularly significant in inflammatory conditions [10].
In addition, G protein-coupled receptor 1 (GPR1), the third known receptor for chemerin, shares structural similarities with CMKLR1, but exhibits distinct functional properties. Although GPR1 binds chemerin with high affinity, it induces only weak Ca2+ mobilization compared to CMKLR1. The primary signaling mechanism of GPR1 involves arrestin’s recruitment rather than traditional G protein-mediated pathways [11]. This arrestin-mediated signaling contrasts with the rapid, second messenger-driven responses typical of G protein activation. Instead, it leads to more sustained cellular responses, such as receptor internalization and activation of alternative pathways like MAPKs, which regulate long-term cellular processes [12]. GPR1 is predominantly expressed in the central nervous system and certain peripheral tissues, suggesting a role in modulating neuroendocrine and metabolic functions [13,14]. The specific physiological and pathological roles of GPR1 are still under investigation, but its distinct expression pattern indicates functions different from those mediated by CMKLR1 and CCRL2 [14].
Overall, the interaction of chemerin with its three receptors, each inducing distinct responses in different systems, underscores its multifaceted role in regulating immune responses, inflammation, adipogenesis and various metabolic processes.
2.3. Main Receptors (CMKLR1, CCRL2, GPR1) and Signaling Pathways
2.3.1. Inflammatory Response
Chemerin is intricately involved in the inflammatory response, acting through its receptor CMKLR1 to recruit neutrophils, macrophages and natural killer cells, thereby linking innate and adaptive immunity [15]. Elevated chemerin levels are observed in various inflammatory conditions, such as obesity [16], rheumatoid arthritis [17], psoriasis [18], and sepsis [19], correlating with disease severity. Under these circumstances, chemerin can modulate inflammation by inducing the production of pro-inflammatory cytokines, like Tumor Necrosis Factor Alpha (TNF-α) and Interleukin 6 (IL-6) through the Extracellular Signal-regulated Kinases 1/2 (ERK1/2), p38 MAPK, and Phosphoinositide 3-kinase-Akt (PI3K-Akt) signaling pathways [20]. For instance, in diabetic nephropathy, the chemerin/CMKLR1 axis promotes inflammation in glomerular endothelial cells, further exacerbating the disease by increasing TNF-α, IL-6, and Interleukin 8 (IL-8) levels [21]. Similarly, in intervertebral disc degeneration, chemerin facilitates the activation of the Nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) signaling pathway, leading to matrix degradation and cell aging [22]. Interestingly, chemerin exhibits dual roles, showing anti-inflammatory effects in some contexts. For example, it can inhibit neutrophil adhesion and promote the clearance of inflammatory cells from mucosal surfaces, as demonstrated in studies on epithelial cells with chemerin/resolvin E1 anti-adhesion phenotype [23]. Additionally, synthetic chemerin-derived peptides have been shown to suppress inflammation by inhibiting macrophage activation and reducing neutrophil and monocyte recruitment in in vitro models of peritonitis [24]. Thus, chemerin’s involvement in inflammation is multifaceted, acting both as a pro-inflammatory and anti-inflammatory mediator, depending on its interaction with the particular tissue environment and the specific inflammatory conditions.
2.3.2. Immune Regulation
Chemerin plays a critical role in immune regulation by modulating the activities of various immune cells and signaling pathways. Through its receptor CMKLR1, chemerin not only recruits immune cells, like dendritic cells and macrophages to inflammation sites, but also influences their activation and maturation [15]. This recruitment and activation are essential for the initiation and resolution of immune responses. Chemerin also impacts the differentiation and function of T cells, thereby linking innate and adaptive immunity [25]. For instance, chemerin can promote the development of regulatory T cells, which are vital for maintaining immune tolerance and preventing autoimmune responses [26]. Furthermore, chemerin influences the balance between pro-inflammatory and anti-inflammatory cytokines, modulating the immune response’s intensity and duration [3]. By regulating the immune cell infiltration and cytokine environment, chemerin ensures a balanced immune response, which is crucial for effective pathogen defense and tissue homeostasis.
2.3.3. Metabolic Processes
Chemerin is a multifunctional adipokine that significantly impacts metabolic processes, including glucose and lipid metabolism as well as energy homeostasis. It is highly expressed in adipose tissue, where it plays a crucial role in adipogenesis—the differentiation of preadipocytes into mature adipocytes. Elevated levels of chemerin are associated with obesity and metabolic syndrome, contributing to insulin resistance and glucose uptake regulation in the liver and skeletal muscle [27]. Chemerin also influences lipid metabolism by promoting adipocyte differentiation and enhancing lipid storage capacity [28]. Additionally, it acts as a chemotactic factor, recruiting immune cells to adipose tissue and contributing to the chronic low-grade inflammation observed in obesity, which exacerbates metabolic dysfunctions, such as insulin resistance [29]. Chemerin levels correlate with several metabolic syndrome components, including body mass index (BMI), triglycerides, and blood pressure [30]. Furthermore, chemerin influences hepatic lipid metabolism, leading to increased lipid accumulation in liver cells via the receptor GPR1, which can contribute to non-alcoholic fatty liver disease (NAFLD) [31]. Thus, the role of chemerin in metabolic processes highlights its importance as a key regulator of energy balance and metabolic homeostasis.
3. Chemerin in Esophageal Cancer
Esophageal cancer, one of the most lethal cancers globally, is known for its aggressive nature and poor prognosis [32]. It ranks as the sixth leading cause of cancer-related deaths worldwide and its prevalence is rising, particularly in developed countries, where adenocarcinoma has become more common than squamous cell carcinoma [33]. There are two main types of esophageal cancer: squamous cell carcinoma, typically linked to smoking, alcohol consumption, and low intake of fruits and vegetables [34]; and adenocarcinoma, often associated with gastroesophageal reflux disease, obesity and Barrett’s esophagus [33]. Other risk factors include hot beverage consumption, poor oral health and low socioeconomic status [32]. The role of chemerin in esophageal cancer is still under investigation, with limited data pointing to an oncogenic role of chemerin for this malignancy as shown in Figure 1.
The study by Somja et al. explored the role of dendritic cells (DCs) in the progression of Barrett’s esophagus (BE) to esophageal adenocarcinoma (EAC), emphasizing the role of chemerin [35]. The researchers found that both myeloid dendritic cells (mDCs) and plasmacytoid dendritic cells (pDCs) are recruited during the metaplasia-dysplasia-carcinoma (MDC) sequence of BE and chemo-attractants, such as chemerin and macrophage inflammatory protein 3a (MIP3a) are responsible for this recruitment. Chemerin is increasingly being expressed throughout the disease progression. In BE, chemerin expression was weak, but it was increased significantly in low-grade dysplasia (LGBE), high-grade dysplasia (HGBE) and EAC. Specifically, chemerin expression scores increased from 1 in BE to 3 in EAC (p<0.001) and this increased expression was correlated with the recruitment of mDCs and pDCs. Functionally, mDCs which were co-cultured with BE and EAC cell lines displayed reduced expression of antigen-presenting molecules such as CD80, CD83, and CD86. Additionally, these mDCs showed increased secretion of the anti-inflammatory cytokine IL-10 and decreased secretion of the pro-inflammatory cytokine IL-12, further promoting an immunosuppressive tumor environment.
A study by Kumar et al. investigated the role of chemerin in recruiting Mesenchymal Stromal Cells (MSCs) to the tumor microenvironment of esophageal squamous cell carcinoma (ESCC) [36]. This study demosntrated chemerin as significantly overexpressed in Cancer-Associated Myofibroblasts (CAMs) compared to adjacent tissue myofibroblasts (ATMs). Conditioned media (CM) from CAMs increased MSC migration by 2.1-fold compared to ATM-CM. This effect was significantly reduced by chemerin-neutralizing antibody, which decreased migration by 60%, as well as chemerin siRNA, ChemR23 siRNA, and the ChemR23 receptor antagonist CCX832, which inhibited MSC migration by 50%. The authors demonstrated that chemerin stimulated MSC migration by activating several signaling pathways, including the phosphorylation of p42/44, p38 and c-Jun NH2-Terminal Kinase 2 (JNK2) kinases. Inhibitors of these kinases and protein kinase C (PKC) reversed chemerin-stimulated MSC migration. In a xenograft model with OE21 esophageal cancer cells and CAMs, MSC migration was increased by 75% when CAMs were present, and this effect was inhibited by CCX832 by 70%, showing both in vitro and in vivo in a xenograft model that CCX832 inhibits MSC migration in response to CAMs.
A follow up study conducted by Kumar et al. explored the role of chemerin and its receptor ChemR23 in promoting the invasion of squamous esophageal cancer (OSC) cells [37]. The study confirmed the expression of ChemR23 in OSC tissues and the OSC cell line OE21 through immunohistochemistry. Functional assays revealed that chemerin significantly enhances the migration, invasion and proliferation of OE21 cells in vitro. Additionally, conditioned media (CM) from CAMs, which is rich in chemerin, markedly increased OE21 cell invasion, an effect that was significantly reduced by chemerin immuno-neutralisation, the ChemR23 antagonist CCX832 and chemerin-specific Small interfering RNA (siRNA). The study further investigated the mechanisms behind chemerin’s role in OSC cell invasion. It was found that chemerin increased the expression and activity of several Matrix Metalloproteinases (MMPs), particularly MMP-1, MMP-2, and MMP-3, in OE21 cells, facilitating cancer cell invasion. The induction of MMPs by chemerin was mediated through Protein Kinase C (PKC) and Mitogen-Activated Protein (MAP) kinase pathways, as inhibitors of these pathways significantly reduced MMP expression. The data suggested that chemerin released by CAMs acts on ChemR23 expressed on OSC cells, promoting an aggressive invasive phenotype via the upregulation of MMPs.
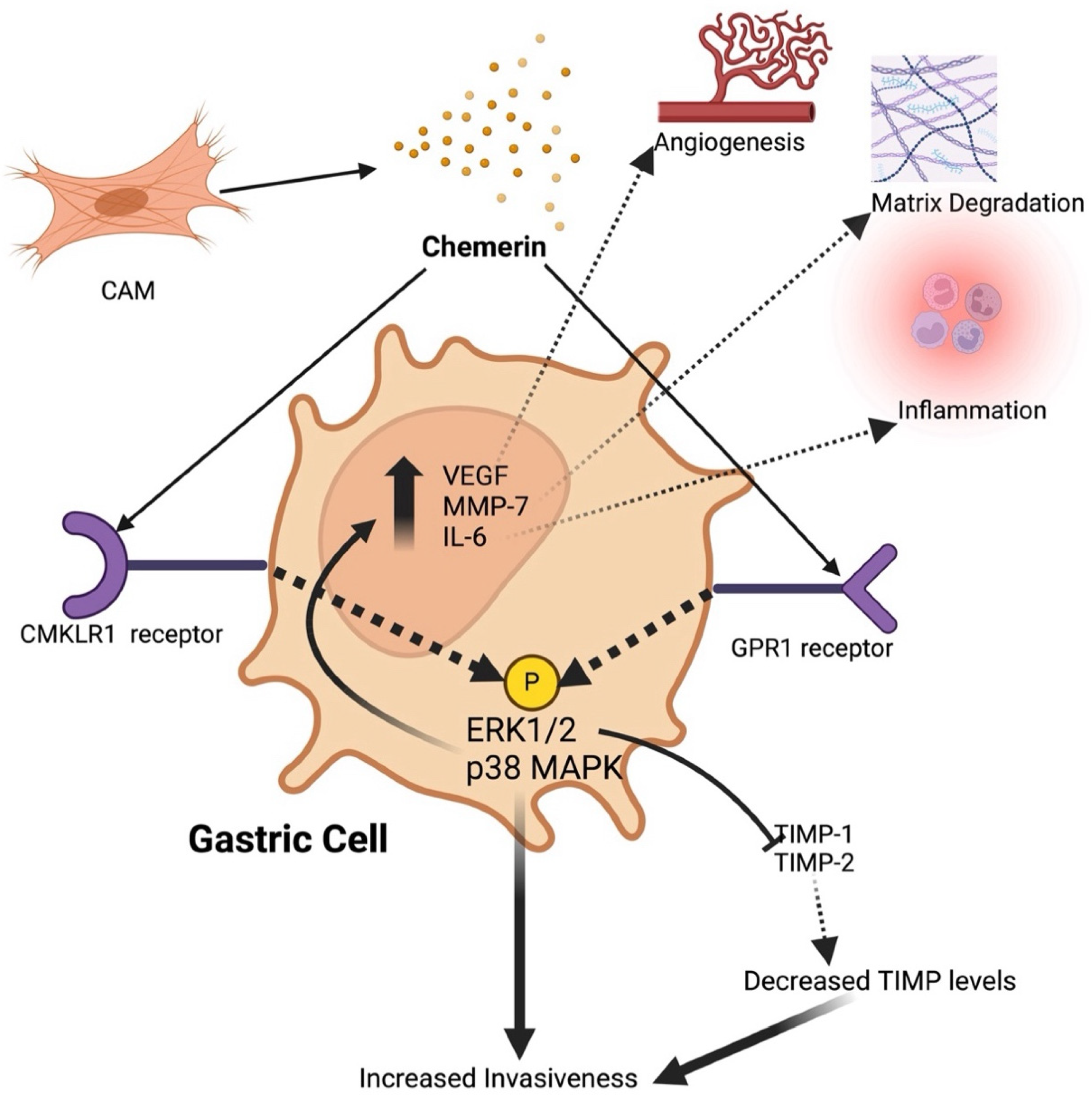
4. Chemerin in Gastric Cancer
Gastric cancer, one of the most common and deadly cancers worldwide, is characterized by its high incidence and poor prognosis. It ranks as the fifth most common cancer and the third leading cause of cancer-related deaths globally, with over 1 million new cases diagnosed annually [38,39]. The prevalence of gastric cancer is particularly high in Eastern Asia, Eastern Europe, and South America, with significant variations in histological subtypes and risk factors between different geographical regions [40]. Gastric cancer is primarily classified into two main types: intestinal and diffuse. Intestinal-type gastric cancer is often associated with chronic infection by Helicobacter pylori, dietary factors such as high salt intake, and smoking [41]. On the other hand, diffuse-type gastric cancer, is linked to genetic predispositions and is less influenced by environmental factors [42]. Other risk factors for gastric cancer include obesity, Epstein-Barr virus infection and pernicious anemia [43]. The prognosis for gastric cancer is poor, with 1-year and 5-year survival rates varying significantly based on the stage at diagnosis. The 5-year survival rate is less than 10% when diagnosed at an advanced stage but can be as high as 85% if detected early [44]. Unlike esophageal cancer, where the role of chemerin is still under investigation, there is more data available on the role of chemerin in gastric cancer. Research indicates that chemerin might play a significant role in the progression and metastasis of gastric cancer, potentially serving as a biomarker for diagnosis and a target for therapeutic interventions as shown in Figure 2.
In their study, Zhang et al. evaluated the prognostic significance of preoperative plasma chemerin levels in gastric cancer patients [45]. Researchers recruited 196 gastric cancer patients and 196 age- and gender-matched healthy controls, measuring preoperative plasma chemerin levels. They found that gastric cancer patients had significantly higher plasma chemerin levels compared to healthy controls. Elevated chemerin levels were identified as an independent predictor of 5-year mortality (OR, 2.718) and adverse events (OR, 2.982), with high area under the ROC curve (AUC) values for both outcomes, indicating strong predictive capability. Furthermore, high chemerin levels were associated with shorter overall survival (HR, 1.788) and disease-free survival (HR, 2.016). The study concluded that high plasma chemerin levels correlated with poor prognosis in gastric cancer, making it as a valuable prognostic biomarker. Patients with higher chemerin levels had increased risk of mortality and adverse events, as well as significantly shorter overall and disease-free survival intervals.
Wang et al. investigated the serum levels of chemerin in gastric cancer patients and its biological effects on gastric cancer cells [46]. The study included 36 gastric cancer patients and 40 healthy subjects, measuring serum chemerin levels before surgical resection of gastric cancer. Results showed significantly higher serum chemerin levels in gastric cancer patients compared to healthy subjects, while increased levels were associated with advanced clinical stages and non-intestinal type of gastric cancer. The study further revealed that serum chemerin levels were significantly higher in patients with stage II and stage III + IV gastric cancer compared to stage I and healthy subjects. Additionally, in vitro experiments demonstrated that chemerin increased the invasiveness of gastric cancer cells, induced phosphorylation of p38 and ERK1/2 MAPKs, while it upregulated pro-invasive genes, such as VEGF, MMP-7 and IL-6. Notably, inhibition of ERK1/2 phosphorylation abolished the pro-invasive effects of chemerin, highlighting a novel action of chemerin in promoting gastric cancer cell invasiveness. Chemerin exposure increased gastric cancer cell invasiveness without affecting cell proliferation and this effect was mediated through the activation of the ERK1/2 MAPK pathway. Pharmacological inhibition of this pathway prevented chemerin-induced invasiveness and upregulation of pro-invasive genes. These findings suggest that increased serum chemerin levels are associated with advanced stages and aggressive types of gastric cancer, and that chemerin may serve as a potential target for therapeutic intervention in gastric cancer.
In another study, Kumar et al. investigated the role of chemerin and its receptors, CMKLR1 and GPR1, in promoting migration and invasion of gastric cancer cells [47]. Immunohistochemical analysis of gastric cancer cells from 15 patients revealed high expression of CMKLR1 and GPR1 levels in nearly all cancer cells, with no evident difference between intestinal, diffuse or mixed gastric cancers or TNM stage. Chemerin was found to be secreted by gastric cancer-associated myofibroblasts (CAMs) but not by gastric cancer cells. The study demonstrated that chemerin significantly increased the in vitro invasiveness of gastric cancer cells, in Boyden chamber assays, with a 5-fold increase in migration at a chemerin concentration of 0.1 ng/ml, without affecting their proliferation. This effect was found to be mediated through the activation of the ERK1/2 MAPK pathway. The investigators demonstrate that both CMKLR1 and GPR1 are involved in these processes, by selectively inhibiting these receptors and observing a significant but not complete suppression of cancer cells migration. Furthermore, the authors suggest that chemerin promotes gastric cancer cell invasion partly by suppressing Tissue Inhibitors of Metalloproteinases TIMP-1 and TIMP-2. This was established by correlating reduction in TIMP levels with increased cancer cell invasiveness, while supplementation with TIMP-1 (2.1 nM) and TIMP-2 (2.5 nM) significantly inhibited the chemerin-induced migration and invasion responses. These findings suggested that chemerin promotes gastric cancer cell invasion partly by suppressing TIMP-1 and TIMP-2, highlighting the potential role of chemerin receptor antagonists as therapeutic agents to inhibit gastric cancer progression.
5. Clinical Implications of Chemerin
5.1. Prognostic Value of Chemerin Levels
In gastric cancer, elevated serum chemerin levels are significantly higher compared to healthy controls, correlating with advanced clinical stages, non-intestinal types of gastric cancer and increased cellular invasiveness. Higher chemerin levels are associated with increased expression of VEGF, MMP-7 and IL-6, which promote tumor metastasis [46]. Moreover, chemerin levels serve as an independent predictor of 5-year mortality, overall survival (OS) and disease-free survival (DFS), underscoring its value as a prognostic biomarker in gastric cancer [45]. Similarly to gastric cancer, chemerin has demonstrated significant prognostic value in gastrointestinal cancers, particularly concerning metastasis. In colorectal cancer (CRC), chemerin levels are usually increased with TNM tumor stage progression and have been correlated with elevated CRP, CEA, and CA 19-9 levels, indicating a pathogenetic and prognostic role [48]. Additionally, higher circulating chemerin levels have been linked to an increased risk of developing colorectal adenoma, a precursor to colorectal cancer, highlighting its importance in early detection and prevention [49]. Furthermore, higher circulating chemerin levels have been associated with increased inflammatory markers such as C-Reactive Protein (CRP), Erythrocyte Sedimentation Rate (ESR), and fibrinogen in CRC patients, linking inflammation to cancer progression and metastasis [50]. Overall, elevated chemerin levels are associated with advanced disease stages, metastasis and poorer survival outcomes in GI cancers, making it a valuable prognostic biomarker for these malignancies.
5.2. Therapeutic Targeting of Chemerin and its Receptors
Considering its role in promoting cancer cell invasiveness and progression, chemerin and its receptors represent potential therapeutic targets. Studies on the chemerin receptor antagonist, CCX832, have shown promising results in inhibiting cancer cell invasion and progression. In squamous esophageal cancer (OSC), the chemerin receptor ChemR23 is expressed in OSC cells, where chemerin stimulates invasion. CCX832 inhibits this invasion by reducing the activity of matrix metalloproteinases (MMPs), which are essential for cancer cell invasion [37]. In gastric cancer, chemerin promotes migration and invasion of cancer cells through its receptors CMKLR1 and GPR1. CCX832 inhibits these effects by downregulating tissue inhibitors of metalloproteinases (TIMP-1 and TIMP-2), highlighting chemerin’s role in enhancing cancer cell invasiveness [47]. Furthermore, in esophageal squamous cancer, chemerin is overexpressed in cancer-associated myofibroblasts (CAMs). CCX832 inhibits chemerin-induced recruitment of mesenchymal stromal cells (MSCs) to the tumor site, potentially delaying tumor progression by preventing the recruitment of supportive stromal cells [36]. These studies collectively highlight the potential role of CCX832 in reducing cancer cell invasion and progression by inhibiting the chemerin/ChemR23 signaling pathway, offering a promising therapeutic strategy for managing gastrointestinal cancers.
5.3. Integration into Current Clinical Practice
Chemerin-targeted therapies could enhance cancer treatment by complementing existing modalities such as chemotherapy, radiotherapy, and immunotherapy [51]. These therapies can improve treatment efficacy and reduce side effects by upregulating Phosphatase and Tensin Homolog (PTEN) and suppressing Programmed death-ligand 1 (PD-L1) in tumor cells, thereby boosting immune responses and recruiting NK and T cells to the tumor microenvironment [51,52]. Synthetic derivatives of chemerin, which are metabolically stable, can serve as precision carriers for chemotherapeutic agents against tumor cells [53]. Furthermore, combining chemerin with existing therapies can prevent chemotherapy-induced cachexia and enhance immune clearance of tumor cells [54]. Overall, chemerin-targeted therapies represent a significant advancement in personalized cancer treatment, offering a comprehensive and potent approach to tumor suppression [55]. Combining chemerin receptor antagonists with standard chemotherapy could enhance the overall response rate by simultaneously targeting tumor cells and the tumor microenvironment, presenting an extremely promising and emerging therapeutic approach [56].
6. Future Directions and Research Needs
6.1. What Is Missing About Chemerin’s Role in Upper GI Cancers?
While substantial progress has been made in understanding the role of chemerin in upper gastrointestinal cancers, several critical gaps still remain. One significant gap is the lack of comprehensive data on chemerin levels and their association with surgical interventions or chemotherapy, which are the gold standard for esophageal and gastric cancer. Understanding how chemerin levels fluctuate in response to these treatments could provide valuable insights into its potential role as a biomarker for treatment efficacy and disease progression [45] Specifically, evaluating changes in chemerin levels as a response to chemotherapy could be particularly valuable in assessing treatment effectiveness and guiding therapeutic adjustments [46].
Another critical gap is the need for detailed studies on chemerin levels in different tissues, including adipose tissue and cancer tissue. Both omental and subcutaneous fat can be used for these assessments, as these adipose tissue depots might provide valuable information on the local production and action of chemerin within the tumor microenvironment. Notably, studies have shown that chemerin mRNA expression is significantly higher in subcutaneous adipose tissue compared to visceral (omental) adipose tissue, indicating depot-specific differences in chemerin expression [57]. Additionally, it is unclear whether blood levels of chemerin are as reliable as tissue levels in reflecting disease status, which also highlights the need for comparative studies to determine the most accurate biomarkers for monitoring treatment efficacy and disease progression.
Furthermore, the correlation between cancer and the potential rise in chemerin levels due to obesity remains unclear, necessitating further research to elucidate this relationship and its implications for cancer progression and treatment [58,59]. Conversely, investigating the relationship between chemerin levels and cachexia could uncover mechanisms by which chemerin influences muscle wasting and weight loss, which represent common complications in advanced cancer stages [54]. This information could lead to more personalized and precise treatment strategies, improving patient outcomes and optimizing therapeutic regimens.
Lastly, the association of chemerin levels with cancer stage, morbidity and mortality as well as patient cachexia and nutritional status in upper GI cancer patients requires further investigation. Understanding these associations could provide crucial insights into the prognostic value of chemerin as a biomarker. Specifically, correlating chemerin levels with TNM staging could potentially identify the extent of cancer progression and metastasis (38,39). Exploring the link between chemerin levels and morbidity and mortality rates may reveal its role in predicting patient outcomes and survival rates [45]. Such detailed studies could help in stratifying patients based on their risk profiles and tailoring personalized treatment approaches, ultimately improving clinical management and outcomes for upper GI cancer patients.
6.2. State-of-the-Art Assessment of Chemerin
Advancements in technology and methodology can significantly enhance the assessment of chemerin in both clinical and research settings. Standardized measurement techniques for novel adipokines, like chemerin, are essential for reducing costs and promoting consistency across studies. The development of advanced ELISA kits and mass spectrometry-based assays can improve the accuracy and reliability of chemerin measurements in blood and tissue samples. Furthermore, the potential of saliva tests to become widely available in the future could offer a non-invasive and convenient method for chemerin assessment, broadening its application in routine clinical practice and large-scale epidemiological studies [60].
Additionally, adipose tissue biopsy could be explored as a viable method for assessing chemerin levels, particularly if adipose tissue proves to be a more reliable source than blood samples. Both omental and subcutaneous fat biopsies could be considered to determine the most effective site for chemerin assessment. This approach could provide a more direct measurement of chemerin’s role in metabolic and inflammatory processes within the tumor microenvironment [57,61].
Emerging imaging technologies, such as PET scans using radiolabeled chemerin analogs, could offer non-invasive methods to assess chemerin distribution and activity in vivo, as proposed for breast cancer [62]. These technologies could provide real-time insights into chemerin dynamics during disease progression and treatment, potentially guiding more effective therapeutic interventions.
6.3. Proposals for Future Research Focusing on Translational and Clinical Studies
Despite significant progress, our understanding of the role of chemerin in upper GI cancer remains limited, especially when compared to more well-studied adipokines. Translational studies should explore the mechanisms by which chemerin influences tumor biology and the tumor microenvironment. This could uncover new pathways and molecular targets for intervention, broadening the scope of therapeutic strategies available for upper gastrointestinal (GI) cancers.
Future research should aim to translate the current understanding of chemerin into clinical applications that can improve patient outcomes. Firstly, large-scale clinical trials are necessary to validate chemerin both as a prognostic biomarker and a therapeutic target. These trials should involve diverse patient populations to assess how chemerin levels influence treatment outcomes, particularly with chemotherapy and surgical interventions.
Additionally, initial studies should aim to establish the safety and efficacy of chemerin inhibitors using preclinical models. This includes using animal models to investigate the effects of chemerin inhibitors on tumor growth, metastasis and responses to standard therapies in gastric and esophageal cancers. Furthermore, research should investigate the potential synergistic effects of combining chemerin inhibitors with other targeted therapies or immunotherapies, as well as conventional chemotherapy.
Collaborative efforts between academic institutions, clinical researchers, and the pharmaceutical industry are essential to accelerate the development and clinical application of chemerin-targeted therapies. Such collaborations can facilitate the sharing of resources, expertise, and data, ultimately advancing the field of cancer research and improving patient care.
In summary, addressing these research needs and leveraging emerging technologies can significantly enhance our understanding of chemerin’s role in upper GI cancers, paving the way for innovative and effective clinical applications.
7. Conclusions
Chemerin, a multifunctional cytokine and adipokine, plays a significant role in inflammatory and metabolic processes, making it a promising prognostic biomarker and therapeutic target for upper GI cancers. To fully realize its potential, comprehensive translational studies and large-scale clinical trials are needed to validate its clinical applications. Research should focus on understanding chemerin’s influence on tumor biology and exploring the efficacy of chemerin inhibitors, especially in combination with existing therapies. Collaborative efforts among researchers and the pharmaceutical industry are crucial to advancing chemerin-targeted therapies, ultimately improving outcomes for patients with upper GI cancer.
Funding
This research received no external funding.
Institutional Review Board Statement
Not Applicable.
Informed Consent Statement
Not Applicable.
Data Availability Statement
Not Applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Jacenik, D.; Fichna, J. Chemerin in Immune Response and Gastrointestinal Pathophysiology. Clin Chim Acta 2020, 504, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Towards an Integrative Approach to Understanding the Role of Chemerin in Human Health and Disease—Rourke—2013—Obesity Reviews—Wiley Online Library Available online:. Available online: https://onlinelibrary.wiley.com/doi/10.1111/obr.12009 (accessed on 6 May 2024).
- Ernst, M.C.; Sinal, C.J. Chemerin: At the Crossroads of Inflammation and Obesity. Trends Endocrinol Metab 2010, 21, 660–667. [Google Scholar] [CrossRef] [PubMed]
- Zabel, B.A.; Allen, S.J.; Kulig, P.; Allen, J.A.; Cichy, J.; Handel, T.M.; Butcher, E.C. Chemerin Activation by Serine Proteases of the Coagulation, Fibrinolytic, and Inflammatory Cascades. J Biol Chem 2005, 280, 34661–34666. [Google Scholar] [CrossRef]
- Du, X.-Y.; Zabel, B.A.; Myles, T.; Allen, S.J.; Handel, T.M.; Lee, P.P.; Butcher, E.C.; Leung, L.L. Regulation of Chemerin Bioactivity by Plasma Carboxypeptidase N, Carboxypeptidase B (Activated Thrombin-Activable Fibrinolysis Inhibitor), and Platelets. J Biol Chem 2009, 284, 751–758. [Google Scholar] [CrossRef]
- Wittamer, V.; Franssen, J.-D.; Vulcano, M.; Mirjolet, J.-F.; Le Poul, E.; Migeotte, I.; Brézillon, S.; Tyldesley, R.; Blanpain, C.; Detheux, M.; et al. Specific Recruitment of Antigen-Presenting Cells by Chemerin, a Novel Processed Ligand from Human Inflammatory Fluids. Journal of Experimental Medicine 2003, 198, 977–985. [Google Scholar] [CrossRef]
- Goralski, K.B.; McCarthy, T.C.; Hanniman, E.A.; Zabel, B.A.; Butcher, E.C.; Parlee, S.D.; Muruganandan, S.; Sinal, C.J. Chemerin, a Novel Adipokine That Regulates Adipogenesis and Adipocyte Metabolism. J Biol Chem 2007, 282, 28175–28188. [Google Scholar] [CrossRef]
- Parolini, S.; Santoro, A.; Marcenaro, E.; Luini, W.; Massardi, L.; Facchetti, F.; Communi, D.; Parmentier, M.; Majorana, A.; Sironi, M.; et al. The Role of Chemerin in the Colocalization of NK and Dendritic Cell Subsets into Inflamed Tissues. Blood 2007, 109, 3625–3632. [Google Scholar] [CrossRef] [PubMed]
- Zabel, B.A.; Nakae, S.; Zúñiga, L.; Kim, J.-Y.; Ohyama, T.; Alt, C.; Pan, J.; Suto, H.; Soler, D.; Allen, S.J.; et al. Mast Cell-Expressed Orphan Receptor CCRL2 Binds Chemerin and Is Required for Optimal Induction of IgE-Mediated Passive Cutaneous Anaphylaxis. J Exp Med 2008, 205, 2207–2220. [Google Scholar] [CrossRef]
- Monnier, J.; Lewén, S.; O’Hara, E.; Huang, K.; Tu, H.; Butcher, E.C.; Zabel, B.A. Expression, Regulation, and Function of Atypical Chemerin Receptor CCRL2 on Endothelial Cells. J Immunol 2012, 189, 956–967. [Google Scholar] [CrossRef]
- Barnea, G.; Strapps, W.; Herrada, G.; Berman, Y.; Ong, J.; Kloss, B.; Axel, R.; Lee, K.J. The Genetic Design of Signaling Cascades to Record Receptor Activation. Proceedings of the National Academy of Sciences 2008, 105, 64–69. [Google Scholar] [CrossRef]
- Gurevich, V.V.; Gurevich, E.V. GPCR Signaling Regulation: The Role of GRKs and Arrestins. Front. Pharmacol. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Marchese, A.; Docherty, J.M.; Nguyen, T.; Heiber, M.; Cheng, R.; Heng, H.H.; Tsui, L.C.; Shi, X.; George, S.R.; O’Dowd, B.F. Cloning of Human Genes Encoding Novel G Protein-Coupled Receptors. Genomics 1994, 23, 609–618. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, N.; Soda, Y.; Kanbe, K.; Liu, H.Y.; Jinno, A.; Kitamura, T.; Hoshino, H. An Orphan G Protein-Coupled Receptor, GPR1, Acts as a Coreceptor to Allow Replication of Human Immunodeficiency Virus Types 1 and 2 in Brain-Derived Cells. J Virol 1999, 73, 5231–5239. [Google Scholar] [CrossRef] [PubMed]
- Wittamer, V.; Bondue, B.; Guillabert, A.; Vassart, G.; Parmentier, M.; Communi, D. Neutrophil-Mediated Maturation of Chemerin: A Link between Innate and Adaptive Immunity. J Immunol 2005, 175, 487–493. [Google Scholar] [CrossRef]
- Rowicka, G.; Dyląg, H.; Chełchowska, M.; Weker, H.; Ambroszkiewicz, J. Serum Calprotectin and Chemerin Concentrations as Markers of Low-Grade Inflammation in Prepubertal Children with Obesity. Int J Environ Res Public Health 2020, 17, 7575. [Google Scholar] [CrossRef]
- Gonzalez-Ponce, F.; Gamez-Nava, J.I.; Perez-Guerrero, E.E.; Saldaña-Cruz, A.M.; Vazquez-Villegas, M.L.; Ponce-Guarneros, J.M.; Huerta, M.; Trujillo, X.; Contreras-Haro, B.; Rocha-Muñoz, A.D.; et al. Serum Chemerin Levels: A Potential Biomarker of Joint Inflammation in Women with Rheumatoid Arthritis. PLoS One 2021, 16, e0255854. [Google Scholar] [CrossRef]
- Gisondi, P.; Lora, V.; Bonauguri, C.; Russo, A.; Lippi, G.; Girolomoni, G. Serum Chemerin Is Increased in Patients with Chronic Plaque Psoriasis and Normalizes Following Treatment with Infliximab. Br J Dermatol 2013, 168, 749–755. [Google Scholar] [CrossRef]
- Horn, P.; Metzing, U.B.; Steidl, R.; Romeike, B.; Rauchfuß, F.; Sponholz, C.; Thomas-Rüddel, D.; Ludewig, K.; Birkenfeld, A.L.; Settmacher, U.; et al. Chemerin in Peritoneal Sepsis and Its Associations with Glucose Metabolism and Prognosis: A Translational Cross-Sectional Study. Crit Care 2016, 20, 39. [Google Scholar] [CrossRef]
- Ye, Y.; Fang, L.; Li, J.; Wu, H.; Tan, X.; Luo, H.; Li, X.; Huang, L. Chemerin/ChemR23 Regulates Cementoblast Function and Tooth Resorption in Mice via Inflammatory Factors. J Periodontol 2021, 92, 1470–1482. [Google Scholar] [CrossRef]
- Shang, J.; Wang, L.; Zhang, Y.; Zhang, S.; Ning, L.; Zhao, J.; Cheng, G.; Liu, D.; Xiao, J.; Zhao, Z. Chemerin/ChemR23 Axis Promotes Inflammation of Glomerular Endothelial Cells in Diabetic Nephropathy. J Cell Mol Med 2019, 23, 3417–3428. [Google Scholar] [CrossRef]
- Hu, S.; Shao, Z.; Zhang, C.; Chen, L.; Mamun, A.A.; Zhao, N.; Cai, J.; Lou, Z.; Wang, X.; Chen, J. Chemerin Facilitates Intervertebral Disc Degeneration via TLR4 and CMKLR1 and Activation of NF-kB Signaling Pathway. Aging (Albany NY) 2020, 12, 11732–11753. [Google Scholar] [CrossRef] [PubMed]
- Campbell, E.L.; Louis, N.A.; Tomassetti, S.E.; Canny, G.O.; Arita, M.; Serhan, C.N.; Colgan, S.P. Resolvin E1 Promotes Mucosal Surface Clearance of Neutrophils: A New Paradigm for Inflammatory Resolution. FASEB J 2007, 21, 3162–3170. [Google Scholar] [CrossRef] [PubMed]
- Cash, J.L.; Hart, R.; Russ, A.; Dixon, J.P.C.; Colledge, W.H.; Doran, J.; Hendrick, A.G.; Carlton, M.B.L.; Greaves, D.R. Synthetic Chemerin-Derived Peptides Suppress Inflammation through ChemR23. J Exp Med 2008, 205, 767–775. [Google Scholar] [CrossRef] [PubMed]
- Bondue, B.; Wittamer, V.; Parmentier, M. Chemerin and Its Receptors in Leukocyte Trafficking, Inflammation and Metabolism. Cytokine & Growth Factor Reviews 2011, 22, 331–338. [Google Scholar] [CrossRef]
- Su, X.; Cheng, Y.; Zhang, G.; Wang, B. Chemerin in Inflammatory Diseases. Clinica Chimica Acta 2021, 517, 41–47. [Google Scholar] [CrossRef]
- Bozaoglu, K.; Bolton, K.; McMillan, J.; Zimmet, P.; Jowett, J.; Collier, G.; Walder, K.; Segal, D. Chemerin Is a Novel Adipokine Associated with Obesity and Metabolic Syndrome. Endocrinology 2007, 148, 4687–4694. [Google Scholar] [CrossRef] [PubMed]
- Helfer, G.; Wu, Q.-F. Chemerin: A Multifaceted Adipokine Involved in Metabolic Disorders. The Journal of Endocrinology 2018, 238, 79–94. [Google Scholar] [CrossRef]
- Lehrke, M.; Becker, A.; Greif, M.; Stark, R.; Laubender, R.P.; von Ziegler, F.; Lebherz, C.; Tittus, J.; Reiser, M.; Becker, C.; et al. Chemerin Is Associated with Markers of Inflammation and Components of the Metabolic Syndrome but Does Not Predict Coronary Atherosclerosis. European Journal of Endocrinology 2009, 161, 339–344. [Google Scholar] [CrossRef]
- Stejskal, D.; Karpisek, M.; Hanulova, Z.; Svestak, M. CHEMERIN IS AN INDEPENDENT MARKER OF THE METABOLIC SYNDROME IN A CAUCASIAN POPULATION—A PILOT STUDY. Biomedical Papers 2008, 152, 217–221. [Google Scholar] [CrossRef]
- Zhu, L.; Huang, J.; Wang, Y.; Yang, Z.; Chen, X. Chemerin Causes Lipid Metabolic Imbalance and Induces Passive Lipid Accumulation in Human Hepatoma Cell Line via the Receptor GPR1. Life Sciences 2021, 278, 119530. [Google Scholar] [CrossRef]
- Zhang, Y. Epidemiology of Esophageal Cancer. World Journal of Gastroenterology 2013, 19, 5598–5606. [Google Scholar] [CrossRef] [PubMed]
- Uhlenhopp, D.J.; Then, E.O.; Sunkara, T.; Gaduputi, V. Epidemiology of Esophageal Cancer: Update in Global Trends, Etiology and Risk Factors. Clin J Gastroenterol 2020, 13, 1010–1021. [Google Scholar] [CrossRef] [PubMed]
- Arnal, M.J.D.; Arenas, Á.F.; Arbeloa, Á.L. Esophageal Cancer: Risk Factors, Screening and Endoscopic Treatment in Western and Eastern Countries. World Journal of Gastroenterology 2015, 21, 7933–7943. [Google Scholar] [CrossRef]
- Somja, J.; Demoulin, S.; Roncarati, P.; Herfs, M.; Bletard, N.; Delvenne, P.; Hubert, P. Dendritic Cells in Barrett’s Esophagus Carcinogenesis: An Inadequate Microenvironment for Antitumor Immunity? Am J Pathol 2013, 182, 2168–2179. [Google Scholar] [CrossRef]
- Kumar, J.D.; Holmberg, C.; Kandola, S.; Steele, I.; Hegyi, P.; Tiszlavicz, L.; Jenkins, R.; Beynon, R.J.; Peeney, D.; Giger, O.T.; et al. Increased Expression of Chemerin in Squamous Esophageal Cancer Myofibroblasts and Role in Recruitment of Mesenchymal Stromal Cells. PLoS One 2014, 9, e104877. [Google Scholar] [CrossRef] [PubMed]
- Kumar, J.D.; Kandola, S.; Tiszlavicz, L.; Reisz, Z.; Dockray, G.J.; Varro, A. The Role of Chemerin and ChemR23 in Stimulating the Invasion of Squamous Oesophageal Cancer Cells. Br J Cancer 2016, 114, 1152–1159. [Google Scholar] [CrossRef]
- Thrift, A.P.; El-Serag, H.B. Burden of Gastric Cancer. Clinical Gastroenterology and Hepatology 2020, 18, 534–542. [Google Scholar] [CrossRef]
- Rawla, P.; Barsouk, A. Epidemiology of Gastric Cancer: Global Trends, Risk Factors and Prevention. Gastroenterology Rev 2018, 14, 26–38. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Ying, X.; Liu, S.; Lyu, G.; Xu, Z.; Zhang, X.; Li, H.; Li, Q.; Wang, N.; Ji, J. Gastric Cancer: Epidemiology, Risk Factors and Prevention Strategies. CJCR 2020, 32, 695–704. [Google Scholar] [CrossRef]
- Venerito, M.; Vasapolli, R.; Rokkas, T.; Malfertheiner, P. Gastric Cancer: Epidemiology, Prevention, and Therapy. Helicobacter 2018, 23, e12518. [Google Scholar] [CrossRef]
- Sitarz, R.; Skierucha, M.; Mielko, J.; Offerhaus, G.J.A.; Maciejewski, R.; Polkowski, W.P. Gastric Cancer: Epidemiology, Prevention, Classification, and Treatment. CMAR 2018, 10, 239–248. [Google Scholar] [CrossRef] [PubMed]
- Machlowska, J.; Baj, J.; Sitarz, M.; Maciejewski, R.; Sitarz, R. Gastric Cancer: Epidemiology, Risk Factors, Classification, Genomic Characteristics and Treatment Strategies. International Journal of Molecular Sciences 2020, 21, 4012. [Google Scholar] [CrossRef] [PubMed]
- Smyth, E.C.; Nilsson, M.; Grabsch, H.I.; Grieken, N.C. van; Lordick, F. Gastric Cancer. The Lancet 2020, 396, 635–648. [Google Scholar] [CrossRef]
- Zhang, J.; Jin, H.-C.; Zhu, A.-K.; Ying, R.-C.; Wei, W.; Zhang, F.-J. Prognostic Significance of Plasma Chemerin Levels in Patients with Gastric Cancer. Peptides 2014, 61, 7–11. [Google Scholar] [CrossRef]
- Wang, C.; Wu, W.K.K.; Liu, X.; To, K.-F.; Chen, G.G.; Yu, J.; Ng, E.K.W. Increased Serum Chemerin Level Promotes Cellular Invasiveness in Gastric Cancer: A Clinical and Experimental Study. Peptides 2014, 51, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Kumar, J.D.; Aolymat, I.; Tiszlavicz, L.; Reisz, Z.; Garalla, H.M.; Beynon, R.; Simpson, D.; Dockray, G.J.; Varro, A. Chemerin Acts via CMKLR1 and GPR1 to Stimulate Migration and Invasion of Gastric Cancer Cells: Putative Role of Decreased TIMP-1 and TIMP-2. Oncotarget 2019, 10, 98–112. [Google Scholar] [CrossRef]
- Alkady, M.M.; Abdel-Messeih, P.L.; Nosseir, N.M. Assessment of Serum Levels of the Adipocytokine Chemerin in Colorectal Cancer Patients. J Med Biochem 2018, 37, 313–319. [Google Scholar] [CrossRef]
- Yagi, M.; Sasaki, Y.; Abe, Y.; Yaoita, T.; Sakuta, K.; Mizumoto, N.; Shoji, M.; Onozato, Y.; Kon, T.; Nishise, S.; et al. Association between High Levels of Circulating Chemerin and Colorectal Adenoma in Men. Digestion 2019, 101, 571–578. [Google Scholar] [CrossRef]
- Erdogan, S.; Yilmaz, F.M.; Yazici, O.; Yozgat, A.; Sezer, S.; Ozdemir, N.; Uysal, S.; Purnak, T.; Sendur, M.A.; Ozaslan, E. Inflammation and Chemerin in Colorectal Cancer. Tumor Biol. 2016, 37, 6337–6342. [Google Scholar] [CrossRef]
- Rennier, K.; Shin, W.J.; Krug, E.; Virdi, G.; Pachynski, R.K. Chemerin Reactivates PTEN and Suppresses PD-L1 in Tumor Cells via Modulation of a Novel CMKLR1-Mediated Signaling Cascade. Clin Cancer Res 2020, 26, 5019–5035. [Google Scholar] [CrossRef]
- Pachynski, R.K.; Wang, P.; Salazar, N.; Zheng, Y.; Nease, L.; Rosalez, J.; Leong, W.-I.; Virdi, G.; Rennier, K.; Shin, W.J.; et al. Chemerin Suppresses Breast Cancer Growth by Recruiting Immune Effector Cells Into the Tumor Microenvironment. Front. Immunol. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Fischer, T.F.; Czerniak, A.S.; Weiß, T.; Zellmann, T.; Zielke, L.; Els-Heindl, S.; Beck-Sickinger, A.G. Cyclic Derivatives of the Chemerin C-Terminus as Metabolically Stable Agonists at the Chemokine-like Receptor 1 for Cancer Treatment. Cancers 2021, 13, 3788. [Google Scholar] [CrossRef] [PubMed]
- Klose, R.; Krzywinska, E.; Castells, M.; Gotthardt, D.; Putz, E.M.; Kantari-Mimoun, C.; Chikdene, N.; Meinecke, A.-K.; Schrödter, K.; Helfrich, I.; et al. Targeting VEGF-A in Myeloid Cells Enhances Natural Killer Cell Responses to Chemotherapy and Ameliorates Cachexia. Nat Commun 2016, 7, 12528. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Lee, J.-H.; Lee, S.K.; Song, N.-Y.; Son, S.H.; Kim, K.R.; Chung, W.-Y. Chemerin Treatment Inhibits the Growth and Bone Invasion of Breast Cancer Cells. International Journal of Molecular Sciences 2020, 21, 2871. [Google Scholar] [CrossRef]
- Xiao, Y.; Yu, D. Tumor Microenvironment as a Therapeutic Target in Cancer. Pharmacol Ther 2021, 221, 107753. [Google Scholar] [CrossRef]
- Alfadda, A.A.; Sallam, R.M.; Chishti, M.A.; Moustafa, A.S.; Fatma, S.; Alomaim, W.S.; Al-Naami, M.Y.; Bassas, A.F.; Chrousos, G.P.; Jo, H. Differential Patterns of Serum Concentration and Adipose Tissue Expression of Chemerin in Obesity: Adipose Depot Specificity and Gender Dimorphism. Molecules and Cells 2012, 33, 591–596. [Google Scholar] [CrossRef]
- Goralski, K.B.; Jackson, A.E.; McKeown, B.T.; Sinal, C.J. More Than an Adipokine: The Complex Roles of Chemerin Signaling in Cancer. Int J Mol Sci 2019, 20, 4778. [Google Scholar] [CrossRef]
- Sell, H.; Divoux, A.; Poitou, C.; Basdevant, A.; Bouillot, J.-L.; Bedossa, P.; Tordjman, J.; Eckel, J.; Clément, K. Chemerin Correlates with Markers for Fatty Liver in Morbidly Obese Patients and Strongly Decreases after Weight Loss Induced by Bariatric Surgery. The Journal of Clinical Endocrinology & Metabolism 2010, 95, 2892–2896. [Google Scholar] [CrossRef]
- Waniczek, D.; Swiętochowska, E.; Lorenc, Z. Serum and Salivary Chemerin Concentrations in Patients with Colorectal Cancer and Obesity. Ann. Acad. Med. Siles. 2021, 75, 11–17. [Google Scholar] [CrossRef]
- Buechler, C.; Feder, S.; Haberl, E.M.; Aslanidis, C. Chemerin Isoforms and Activity in Obesity. Int J Mol Sci 2019, 20, 1128. [Google Scholar] [CrossRef]
- Erdmann, S.; Niederstadt, L.; Koziolek, E.J.; Gómez, J.D.C.; Prasad, S.; Wagener, A.; von Hacht, J.L.; Reinicke, S.; Exner, S.; Bandholtz, S.; et al. CMKLR1-Targeting Peptide Tracers for PET/MR Imaging of Breast Cancer. Theranostics 2019, 9, 6719–6733. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The mechanisms of tumor-promoting effects of Chemerin in esophageal cancer: Chemerin, released from cancer-associated myofibroblasts (CAMs), acts on CMKLR1 receptors present on esophageal cancer cells and mesenchymal stromal cells (MSCs), activating several intracellular signaling pathways, including ERK1/2, p38 MAPK, JNK-II, and PKC. This signaling leads to increased expression and activity of matrix metalloproteinases (MMP-1, MMP-2, MMP-3), resulting in matrix degradation and enhanced invasiveness of esophageal cancer cells. Chemerin also recruits MSCs to the tumor site, promoting further tumor progression. Additionally, chemerin influences the tumor microenvironment by recruiting myeloid dendritic cells (mDCs) and plasmacytoid dendritic cells (pDCs), which display reduced expression of antigen-presenting molecules (CD80, CD83, CD86) and altered cytokine secretion (increased IL-10, decreased IL-12), creating an immunosuppressive environment. These effects collectively contribute to the progression of esophageal cancer through increased tumor cell migration, invasion, and immune evasion. Key components include ECM (extracellular matrix), ERK1/2 (extracellular signal-regulated kinase 1/2), IL-10 (interleukin 10), IL-12 (interleukin 12), MAPK (mitogen-activated protein kinase), PKC (protein kinase C), and MMPs (matrix metalloproteinases). Created with BioRender.com.
Figure 1.
The mechanisms of tumor-promoting effects of Chemerin in esophageal cancer: Chemerin, released from cancer-associated myofibroblasts (CAMs), acts on CMKLR1 receptors present on esophageal cancer cells and mesenchymal stromal cells (MSCs), activating several intracellular signaling pathways, including ERK1/2, p38 MAPK, JNK-II, and PKC. This signaling leads to increased expression and activity of matrix metalloproteinases (MMP-1, MMP-2, MMP-3), resulting in matrix degradation and enhanced invasiveness of esophageal cancer cells. Chemerin also recruits MSCs to the tumor site, promoting further tumor progression. Additionally, chemerin influences the tumor microenvironment by recruiting myeloid dendritic cells (mDCs) and plasmacytoid dendritic cells (pDCs), which display reduced expression of antigen-presenting molecules (CD80, CD83, CD86) and altered cytokine secretion (increased IL-10, decreased IL-12), creating an immunosuppressive environment. These effects collectively contribute to the progression of esophageal cancer through increased tumor cell migration, invasion, and immune evasion. Key components include ECM (extracellular matrix), ERK1/2 (extracellular signal-regulated kinase 1/2), IL-10 (interleukin 10), IL-12 (interleukin 12), MAPK (mitogen-activated protein kinase), PKC (protein kinase C), and MMPs (matrix metalloproteinases). Created with BioRender.com.
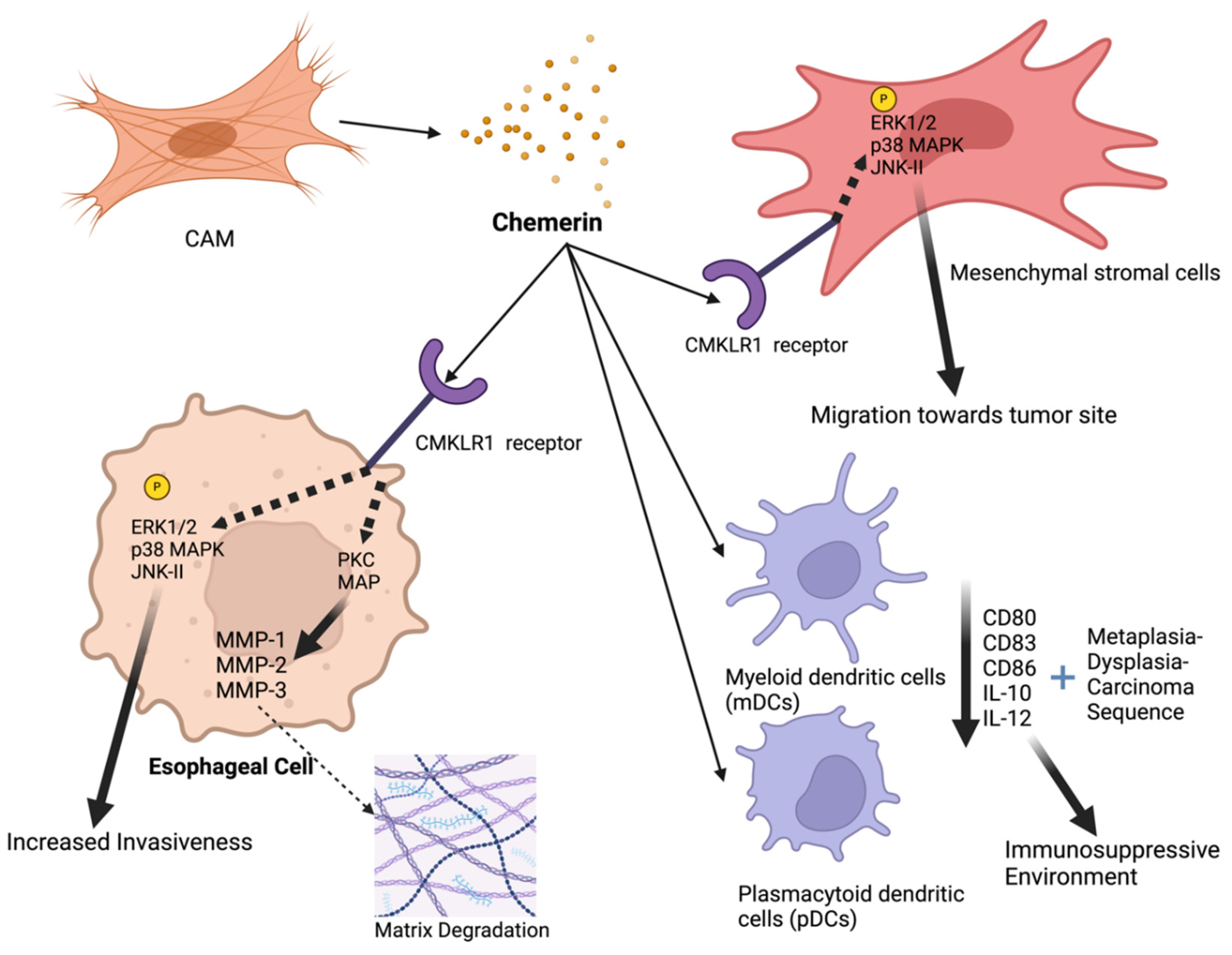
Figure 2.
The mechanisms of tumor-promoting effects of chemerin in gastric cancer: Chemerin is released from cancer-associated myofibroblasts (CAMs) and acts on CMKLR1 and GPR1 receptors present on gastric carcinoma cells to activate several intracellular signaling pathways. Functionally, this signaling leads to increased expression of pro-invasive genes such as VEGF, MMP-7, and IL-6, reduced secretion of tissue inhibitors of metalloproteinases (TIMP-1 and TIMP-2), and enhanced production of matrix metalloproteinases (MMPs). These effects collectively result in migration and invasion of tumor cells and tumor cell transformation resembling an epithelial-to-mesenchymal transformation (EMT). Additionally, chemerin signaling promotes angiogenesis, matrix degradation, and inflammation, contributing to the tumor-promoting environment. Key components include ERK1/2 (extracellular signal-regulated kinase 1/2), IL-6 (interleukin 6), MAPK (mitogen-activated protein kinase), PKC (protein kinase C), TIMP (tissue inhibitors of metalloproteinases) and VEGF (vascular endothelial growth factor). Created with BioRender.com.
Figure 2.
The mechanisms of tumor-promoting effects of chemerin in gastric cancer: Chemerin is released from cancer-associated myofibroblasts (CAMs) and acts on CMKLR1 and GPR1 receptors present on gastric carcinoma cells to activate several intracellular signaling pathways. Functionally, this signaling leads to increased expression of pro-invasive genes such as VEGF, MMP-7, and IL-6, reduced secretion of tissue inhibitors of metalloproteinases (TIMP-1 and TIMP-2), and enhanced production of matrix metalloproteinases (MMPs). These effects collectively result in migration and invasion of tumor cells and tumor cell transformation resembling an epithelial-to-mesenchymal transformation (EMT). Additionally, chemerin signaling promotes angiogenesis, matrix degradation, and inflammation, contributing to the tumor-promoting environment. Key components include ERK1/2 (extracellular signal-regulated kinase 1/2), IL-6 (interleukin 6), MAPK (mitogen-activated protein kinase), PKC (protein kinase C), TIMP (tissue inhibitors of metalloproteinases) and VEGF (vascular endothelial growth factor). Created with BioRender.com.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



