
Submitted:
13 September 2024
Posted:
17 September 2024
You are already at the latest version
Abstract
Brain plasticity is the ability of the brain to change and adapt in response to experience or learning. Its hallmarks are developmental flexibility, complex interactions of genetic and environmental influences, and structure-functional changes comprising neurogenesis, axonal sprouting, and synaptic remodeling. Herein, we review brain plasticity association with non-coding RNAs (ncRNAs). Studies on brain plasticity have important practical implications. Numerous cerebral ncRNAs are linked with neurological disorders, which makes them putative targets for RNA-based diagnostics and therapeutics since molecular characteristics of brain plastic change may reveal disease course and rehabilitative potential of the patient. A new insight into the concept of brain plasticity will provide perspectives on functional recovery following brain damage. The knowledge of this phenomenon will enable physicians to exploit cerebral plastic potential and regulate eloquent networks with timely interventions. Future studies will provide pathophysiologic mechanisms of brain plasticity at macro- and microscopic levels to advance rehabilitation strategies and improve the quality of life in neurological disorders.
Keywords:
brain plasticity
; molecular biomarkers
; RNA diagnostics
; RNA therapeutics
; transcriptomics
; ncRNA
; lncRNA
; miRNA
; circRNA
; eRNA
; lincRNA
; piRNA
; yRNA
1. Introduction
Numerous studies showed that experience and lesions of the peripheral or central nervous system can modulate functional cortical organization [2]. Hence, the brain is a dynamic organ, which implies the ability of the network of neural connections for self-modification in response to experience [22]. Brain plasticity (BP) refers to its ability to optimize the functioning of brain networks through reorganization of neuronosynaptic maps [1,2]. BP is a continuous process and remodeling of the maps can be short-, middle- and long-term [23]. The capacity of the brain to change structurally and/or functionally allows the individual to learn, remember, forget and recover from injury [1,24]. Therefore, BP is a compensatory phenomenon [2]. BP changes across life: it is enhanced in children and reduced in adults [25]. Herein, we summarize views on pathophysiology of cerebral plasticity at sub-cellular, cellular, and synaptic map level.
1.1. Concept of Brain Plasticity
Neuroplasticity is the ability of the brain to change structurally and functionally [24]. Experience may produce multiple dissociable modifications to the neural system (see Figure 1). These refer to an increase in dendric length and glial activity, change in spine density, synapse formation, and altered metabolic activity. These variations change brain weight, cortical thickness, acetylcholine levels and dendric structures. The structural modulation impacts behavior. Age, hormonal profile, trophic factors, stress, and brain pathology also affect the functional outcomes.
The key principle of behavioral neuroscience is that experience can modify brain structure long after brain development is complete [24]. In response to behavioral demands, the mammalian brain can form new synapses, grow dendrites, and create new elements of supportive tissue: astrocytes, blood vessels [13,26]. Environmental enrichment studies showed large changes in various measures of cortical morphology. In these studies, a control group of animals is kept in laboratory cages and the experimental group is placed in large enclosures with visually stimulating objects and an opportunity to interact with the environment. The studies reported an increase in the dendritic fields of neurons by 20% relative to cage-reared animals. Dendric space correlates closely with synaptic numbers [27,28,29].
Besides, experience (environmental enrichment) modulates synapses by modifying the excitatory-inhibitory equilibrium. Specifically, the number of excitatory synapses per neuron increases and the number of inhibitory ones decreases. Changes in neuronal morphology require more active metabolism, blood supply and support from glial cells, especially from astrocytes [24].
Merely having exercise is not sufficient to induce neuronal changes. A more complex task increases neuronal processing, which results in a more active synapse formation [24]. Environmental enrichment increases both dendritic length and density of synaptic spines on the dendrites. Some authors found an association between the extent of dendritic arborization in a cortical language area and the amount of education [30]. In another experiment, children with developmental delay had spindly dendrites with a reduced spine density compared to average intelligence children [31].
1.2. Different Types of Plasticity
BP can be classified in different ways. For example, scientists mention ’activity-dependent plasticity’, and it talks to itself [1]. The brain’s ability to alter the structural and functional properties of neurons refers to structural and functional plasticity, respectively [32]. Researchers presume that synapse is the most likely place to identify neural changes associated with behavior [24]. Synaptic plasticity is the ability of a synapse to change over time through use or disuse. Meanwhile, dendritic plasticity occurs at postsynaptic sites, the dendrites, and spines of excitatory neurons [5].
In the glossary section, we listed a broader classification of BP into subtypes. Still, physicians mainly focus on post-lesional plasticity, which is the ability to adapt after damage to the peripheral or central nervous system, with functional reshaping underlying a partial or complete clinical recovery [2] Although BP is a dynamic construct, it should be stabilized through the mechanisms called homeostatic plasticity, otherwise, the system will not be functional [2].
2. Neuroanatomic and Neurophysiologic Bases of Brain Plasticity
2.1. Plasticity in the Periphery and at the Centrum of the Brain
In local anesthesia, amputation, and peripheral neuropathy, sensory deprivation is the major reason for cerebral reorganization. The adjacent regions of the cortex expand at the expense of the deprived cortex. The suggested mechanism of the expansion is as follows. Within certain minutes after the trauma, the acute reorganization happens due to the unveiling of latent intracortical connections. In the following months, additional remodeling occurs. In the primary motor cortex, a peripheral lesion also results in the expansion of the cortical areas in the vicinity of the representation of the body part that is injured [2].
After lesions of the primary somatosensory area, the damaged representations are redistributed both in remote regions and in the areas adjacent to the injury [2]. For the motor function, animal studies have also demonstrated a similar experience-dependent plasticity after the central lesions. The researchers showed a potential of rehabilitative training to shape remodelling in the adjacent undamaged cortex [2]. Hence, the recruitment of the intact motor cortex is a mechanism of motor recovery [33].
2.2. Natural Plasticity in Different Functional Areas
Plastic changes differ among functional areas. The primary motor cortex controls the kinetic and dynamic parameters of voluntary movements [34], and the cortical representations of muscles and movements have a mosaic structure [35]. Motor training reshapes the primary motor cortex, and the acquisition of new motor skills necessitates an extension of activation, which is reached by temporal or durable recruitment of adjacent sites [36].
The primary sensorimotor cortex integrates sensory and motor signals necessary for skilled movement, namely involved in cognitive functions: motor skill learning [37], calculation [38], and mental imagery [39]. Hence, the role of the sensorimotor cortex is more complex than the control of movements. Learning a skill modifies the activity of isolated neurons and brain regions. The synchronous activity of many neurons in the same cortical region may quickly change the time-course of the ensemble of neurons executing the movement [40,41]. The non-primary parts of the somatosensory network also undergo plasticity-related changes, and the effective connectivity within the whole functional network rises [42,43].
The functional areas of language and cognition are cortico-cortical and cortico-subcortical networks which function parallel. The areas have a hierarchy with both simultaneous and successive activation of the networks. Some of them are essential while others are compensative [44,45,46]. Plasticity implies the modification of spatio-temporal parameters of the functioning of the networks.
3. Pathophysiological Mechanisms Underlying Cerebral Plasticity
3.1. Plasticity Mechanisms at Microlevel
At the microscopic level, many ultrastructural and synaptic changes may take place. During neurodevelopment, these are cyto- and histogenesis, with proliferation and elaboration of dendritic and axonal branches; cell migration, formation of synapses, and cellular differentiation; precise organization of the circuitry, apoptosis, regression of axons, elimination of cells and synapses. At this stage, radial glia controls neuronal migration from the subventricular zone to the cortex, thus also contributing to developmental plasticity [47]. After the period of neurodevelopment, structural and functional reorganization of the brain may proceed with the major changes taking place at the synaptic level. The plasticity mechanisms include changes in the activity of isolated neurons, in synaptic efficacy, and in the temporal relations between ensembles of neurons in specific oscillation bands [48]. In a combination, these mechanisms can modulate the behavior [2,49].
The synapse is a dynamic rather than a static contract. Beyond its increase in size and number due to learning [50], one can see modulation of synaptic strength, which evidences the presence of plastic properties in these dynamic connections [51]. Once appear at the microscopic level, these modulations account for functional map reshaping observed at the macroscopic level [52]. The examples are the activity-dependent synaptic plasticity, auto-regulation of synapses which is called ‘metaplasticity‘.
The activity-dependent synaptic plasticity is a leading mechanism of memory formation. Repeated nerve impulses change synaptic transmission: frequent stimuli coming to the presynaptic membrane may increase or decrease the induced excitation of the postsynaptic neuron. The activity-dependent synaptic plasticity establishes a real-time control over the flow of information within neuronal networks [2]. This type of plasticity explains both long-term potentiation and long-term depression – the two opposite phenomena. The first one is a durable enlargement of the synaptic strength which is followed by a brief high-frequency stimulation. Otherwise, such stimulation might lead only to a short-term potentiation. The mechanism was demonstrated in the hippocampus and motor cortex, and it may underly functional plasticity in motor cortex [53]. The second phenomenon plays an important role in learning and memory [54].
Metaplasticity is an ability of synapses to auto-regulate themselves [55,56]. Different hypotheses were proposed to describe the mechanisms of memory formation through modulation of synapses. According to the synaptic plasticity and memory hypothesis, the induction of activity-dependent synaptic plasticity at appropriate synapses forms the memory [57]. However, little evidence supports the sufficiency of activity-dependent synaptic plasticity for storing memory [2]. According to Hebbian’s rule, the physiological bases for learning and memory are modifications of synaptic strength among neurons that are activated simultaneously when the task is repeated [11]. The rule is widely accepted in neuroscience [58]. Moreover, scientists have discovered a mechanism essential for balancing the processes of Hebbian learning. It is synaptic stabilization through a regulation of the AMPA receptors mediating fast synaptic transmission [59]. This self-regulation of neuronal excitability relative to network activity is called ’homeostatic’ plasticity - the term derives from two opposing concepts, and it means "staying the same through change".
Synchronization of episodic electrostimulation of cerebral ganglia is necessary for a massive reorganization of the cortex [60]. ’Effective’ connectivity refers to influences among brain regions. Biomathematical modeling is used to determine how a constrained set of brain regions influence each other in a specific task. Knowledge of these regions comes from neuroanatomy [12]. A study showed a synchronization of activity among different areas involved in a sensorimotor function due to training [61]. Hence, plasticity may appear as a modification in ’effective’ connectivity within the whole functional network [42].
Another major mechanism of short-term plasticity is a decrease in the inhibitory activity of the GABA interneurons that block horizontal connections in regular settings [62]. However, sensory deprivation or learning suppresses the GABA inhibition, which unmasks latent connections and transforms silent synapses to functional ones [11,63]. Thalamo-cortical networks facilitate the described process [64].
Glia can also affect synaptic transmission, coordinate activity across neuronal networks, and modulate neuronal activity in different ways. These include a release of neurotransmitters and other signaling molecules, neuro-vascular coupling which regulates energy metabolism [65]. Besides, glial cells can communicate with each other thus forming a glial network which is able to both listen and talk to neuronosynaptic circuits [66].
At the neuronal level, structural modifications include sprouting of the dendritic spine, growing of the axon and formation of new synapses (neosynaptogenesis). Experience or brain damage may initiate these modifications. The experience-dependent plasticity is based on the increased synapse turnover which denotes the accelerated formation and elimination of synapses. This mechanism underlies adaptive remodeling of neural circuits [67].
The post-injury plasticity is based on a rapid induction of changes in the number, size and shape of dendritic spines [68,69]. The suggested molecular mechanisms for this are protein synthesis [69], secretion of growth factors and neurotrophins [70]. AMPA receptors and integrins stabilize morphological changes through the mechanism of ’homeostatic’ plasticity [50,71]. Axons may also regenerate spontaneously and elongate [72]. Glia controls the number of synapses [73] and adjusts to meet modifications in the brain environment [74]. Both in physiologic conditions and after injury, the changes in the glia size and phenotype is quick (hours) [67,75], and it can be conveyed to other glial cells via connexin [76].
Researchers began to question the old dogma that the adult mammalian brain cannot develop new neurons. The olfactory bulb, the dentatus gyrus, and even the neocortex of adult primates are exceptions to this rule which turned out to be not absolute [26,77,78]. In vitro, multipotential progenitor cells of adult humans underwent neurogenesis. The cells were isolated from the temporal neocortex, hippocampus, and subcortical white matter [79,80,81]. Studies suggest that these newly created neurons may store memories and contribute to learning via changes in neuronosynaptic circuits, formation of new connections and networks [82]. The post-lesional plasticity can also be arranged in the way of the neurogenesis, as this was shown in adult rats. The animals generated endogenous neural precursor cells in situ and then to differentiate then into mature neurons replacing the damaged ones [83]. This fact favors the idea of neuronal replacement therapies.
3.2. Plasticity Mechanisms at Macrolevel
At the macroscopic level, functional reorganization is carried out through the mechanisms of diaschisis, functional reorganization of cortex within eloquent areas and networks, cross-modal plasticity, compensatory strategies, and macroscopic morphological changes. Diaschisis is a general term describing functional alteration outside of focal brain damage. These are electrophysiological, metabolical, and hemodynamic changes. Although diaschisis underlines initial functional impairment, the same mechanism accounts for spontaneous functional recovery after injury [84,85].
Another mechanism of functional reorganization after brain injury affects eloquent cortex. The eloquent areas have redundant representation of the same function within the same region, Within the eloquent areas, functions have multiple cortical representations within the same region. So, the eloquent site is discrete, and once partially destroyed, it is compensated by the adjacent redundant sites that are unmasked post injury [86,87]. However, in wide lesions, this mechanism does not provide sufficient compensation, therefore other cortical parts are recruited to restore the function [88]. These are regions of the same functional networks, remote ipsi-hemispheric structures and functional homologous structures in the contralateral hemisphere. The suppression of the regions is released step by step with the unmasking of each next region if the functional compensation is insufficient [89].
’Cross-modal plasticity’ refers to the compensation of functional alterations through the recruitment of the structures that did not belong to the eloquent circuit which was altered [8,9,10]. For example, deaf patients may activate auditory cortex during somatosensory tasks, and in this way, they get better tactile discrimination [90]. For the same reason, these individuals can benefit less from cochlear implants due to extensive cross-modal plasticity [91]. If the unimodal areas cannot be recruited after massive damage, heteromodal association areas are activated. Although the activation does not allow for a complete functional restoration, this mechanism can be considered as an elaboration of compensatory cognitive strategies [92].
Although happening mainly at the ultrastructural level, neurogenesis may result in the macrostructural changes that can be detected with the voxel-based morphometry [93]. With this technique, scientists showed that cortical regions, cerebellum hippocampus, and density of the white matter tracts in the predominant hemisphere can be enlarged to meet professional or educational demands [94,95,96,97,98,99]. In the grey matter, training can induce transient morphological changes [100].
4. Modulation of Experience-Dependent Change
4.1. Sex Hormones
Studies reported that the brain is more sensitive to experience in females than in males [101,102,103,104]. Same studies showed that hippocampus has the same gender disproportion: it is more sensitive to experience in females. However, these changes can be manipulated with hormonal replacement therapy [24]. A failure of dendric growth is a supposed pathophysiologic mechanism of developing dementia [105].
4.2. Neurodevelopment and Brain Plasticity in Childhood
A superior ability of children to learn a language and to recover from brain trauma demonstrate enhanced brain plasticity in comparison to adults [1]. During early years, several mechanisms account for enhanced brain plasticity. First, neurogenesis does not stop right after birth, although adult neurogenesis is missing in humans [106]. Second, programmed cell death (apoptosis) may eliminate neurons [107]. Third, the number of synapses may either increase or decrease and synaptic functioning can be refined by activity-dependent mechanisms [5,25].
In children, plasticity of the brain is maximal, and it can be classified into the following categories: adaptive, impaired, excessive plasticity, and plasticity that makes the brain vulnerable to injury [1]. The first category refers to adjustments in neuronal circuitry that allow the individual to compensate for injuries to the brain or develop a special skill with practice. The second one is linked with cognitive impairment when genetic or acquired disorders disrupt molecular plasticity pathways. In contrast to this, excessive plasticity leads to disability through the reorganization of maladaptive neuronal circuits in the developing brain. These new maladaptive brain circuits cause neurologic disorders such as partial seizures following mesial temporal sclerosis or focal dystonia. Finally, brain plasticity can become its ’Achilles’ heel’ and increase the vulnerability of the brain to injury. In energy failure or status epilepticus, the mechanisms regulating plasticity are over-stimulated, which leads to excitotoxic neuronal damage.
4.3. Brain Plasticity in Adulthood
The brain holds the potential for functional and structural rearrangement throughout life, which has been underestimated recently [108]. In adults, learning induces the elaboration of new circuits and the maintenance of neural networks. In elderly people, natural plasticity may resist negative outcomes of brain aging which typically results in neurocognitive slowing [22,109,110,111,112,113,114,115,116,117,118,119,120,121,122,123,124,125,126]. In normal aging, the number of synapses increases in the cortex, which allows middle-aged people to compensate for the loss of neurons with age and to maintain the number of synapses across life [24].
5. Non-Coding RNAs in Brain Plasticity
This section reveals the molecular alterations that may serve as biomarkers of plastic changes in the brain. These comprise non-coding RNAs, microRNAs, circular RNAs, long intergenic non-coding RNAs, Y RNAs. The recently discovered world of non-coding RNAs (ncRNAs) is continuously expanding to all areas of biomolecular interactions and variety of cellular processes including control of metabolism, gene regulation and protein turnover. Expectedly, multiple ncRNA players were found to be involved in brain plasticity (for several examples, see Table 1). Furthermore, interactions between different types of ncRNAs create multidimensional networks that respond to a range of endogenous and exogenous stimuli. Noncoding RNAs (ncRNAs) represent the major part of the transcriptome. Various classes of ncRNAs have emerged as critical regulators of transcription, epigenetic processes, gene silencing and play important roles in neural brain plasticity and cognitive processes [127]. ncRNAs regulate diverse intracellular and neuronal functions: modulate chromatin structure, act as chaperones, and contribute to synaptic remodeling and behavior [128].
Neurons are highly compartmentalized due to their morphological and functional complexity. This happens due to the transport of messenger RNA (mRNA) transcripts to specific subcellular areas, e.g. synaptic regions, for the local translation. Increasing evidence shows that highly expressed cerebral ncRNAs participate in the spatial and temporal control of the mRNA translation, therefore, in synaptic plasticity [129].
ncRNAs may contribute to the development of a variety of neuropsychiatric disorders, including schizophrenia, addiction, and fear-related anxiety disorders [127,128]. Moreover, the diversity of ncRNAs and their association with neurodegenerative diseases render them particularly interesting as putative targets of brain disease [130]. New RNA-based therapeutics can be developed due to the new knowledge on the ncRNA regulation and the downstream effects of their interactions in different pathologies.
Long non-coding RNAs (lncRNAs) act as scaffolds for biomolecule binding and mediate different RNA-protein interactions. LncRNAs are increasingly recognized for their involvement in neurodevelopmental processes including cell proliferation, neurite outgrowth, synaptogenesis, and neuroplasticity [159]. Neuronal lncRNAs are crucial for orchestrating neurogenesis, for tuning neuronal differentiation, and for the exact calibration of neuronal excitability [130]. Particularly, Malat1 is an lncRNA which is abundant in the nuclei of neurons. It promotes synapse formation by recruiting the serine/arginine splicing factors to the transcription sites of genes involved in synaptogenesis. Overexpression of Malat1 enhances the number of synapses in hippocampal neurons in vitro while its deficiency reduces number of synapses between dendrites and axons [160,161]. Gomafu is another lncRNA involved in ES cell, neuronal cell and retinal cell differentiation. The lack of Gomafu led to hyperactive phenotype and increased sensitivity to the psychostimulant MAP in Gomafu KO mice [131]. lncRNA Gas5 promotes neuronal differentiation of hippocampal NSCs and restores learning and memory in rats with cholinergic injury [132]. Furthermore, synapse-specific Gas5 KO led to impaired fear extinction memory [162].
MicroRNAs (miRNAs) are small endogenous RNAs about 20–25 nucleotides in length that regulate gene expression posttranscriptionally [163,164]. They are commonly expressed in specific brain regions and affect nervous system development, plasticity, and function [165]. For example, miR-9 has a critical role in hippocampal synaptic plasticity and memory [136], miR-34 regulates synaptogenesis [137], miR-132 participates in axon growth, neural migration, and plasticity [138]. In the temperament-character molecular integration network (TCMIN), only three miRNAs (hsa-miR-1-3p, hsa-miR-335-5p, hsa-miR-34a-5p) are sufficient to coordinate interactions between two gene networks in brain involved in self-regulation of emotional reactivity to extracellular stimuli (e.g., self-regulation of anxiety) and interpretations of meaning (e.g., production of concepts and language) [144].
Potential targeting or therapeutic use was demonstrated for several miRNAs [166]. Particularly, miR-17-92 cluster shown to enhace neuroplasticity [139] and regulate adult hippocampal neurogenesis, anxiety, and depression [140]. miR-144-5p is currently considered as a key target in major depressive disorder [141] and miRNA-145 was recently shown to enhance neural repair after spinal cord injury [142]. One of the highly conserved miRNAs in mice and humans, miRNA-153 stabilizes the neurogenesis of neural stem cells and enhances cognitive ability through the Notch signaling pathway [143].
Circular RNAs (circRNAs) are closed structural isoforms of linear mRNA. They are abundant in the brain and play a significant role in the development of the nervous system [167]. Cerebral circRNAs are linked with neurotransmitter function, synaptic activities, and neuronal maturation. Levels of ciRS-7, circRMST, and circFAT3 increased during the differentiation of human embryonic stem cells into rostral and caudal neural progenitor cells [145]. The level of a recently discovered circular RNAs - circIgfbp2 - is significantly increased in injured brain tissue. It is involved in neural plasticity and might be a future therapeutic target for anxiety and sleep disorders after traumatic brain injury [146]. At least four circRNAs (cirC_0000400, cirC_0000331, cirC_0000406, cirC_0000798) are involved in postoperative neurocognitive disorders [147]. In a rat model, a large number of circRNAs including 1167 cerebral circRNAs displayed a developmental-dependent expression pattern and may have important biological function in differentiation, development, and aging [148]. Enhancer RNAs (eRNAs) are long non-coding RNAs, bidirectionally transcribed by RNA polymerase II from enhancer regions of the genome. Generally, eRNAs are not spliced or polyadenylated [168,169,170]. Bdnf-Enhg1 and Bdnf-Enhg2 were characterised as novel enhancers that regulates Bdnf expression in developing neurons [149]. Conserved enhancer Evf2 was shown to functionally and spatially organizes megabase distant genes in the developing forebrain [150].
Long intergenic non-coding RNAs (lincRNAs) are biochemically identical to other lncRNAs but differ in their genomic organization as they reside in the space between genes [171]. Knockout of linc-Brn1b showed a reduced number of intermediate progenitor cells in the subventricular zone. This suggests that linc-Brn1b can be involved in the development of cortex [151]. Long non-coding RNA X-inactive specific transcript (XIST) was mentioned as a promising molecular target for SCI therapy [172] and may have a significant role in AD [152].
Piwi-interacting RNAs (piRNAs) are a class of Piwi-associated, 26–32 nucleotide small non-coding RNAs that, unlike other small RNAs, are generated from long genomic clusters [173,174,175]. piRNAs are part of a gene regulatory mechanism responsible for establishing stable long-term changes in neurons and the persistence of memory in brain synaptic plasticity [153]. The main molecular function of piRNAs is to regulate transposons. The co-existence of piRNA and retrotransposons might play important roles in the brain development and the adult brains [154]. A number of piRNAs across brain transcriptome are associated with Alzheimer’s disease [155].
Y RNAs (yRNAs) are a class of non-coding RNA often found abundantly expressed in brain and neuronal tissues. Y RNAs are linked to neuronal stress and very often associated with neuronal ELAV-like proteins in Alzheimer’s disease [156] and could serve as biomarkers in glioma [157]. Recent study suggested that the strong tendency of Y RNAs to bind nELAVL proteins to in response to stress conditions might prevent these proteins from associating with their normal messenger RNA targets [156].
6. Implications for Medical Practice
6.1. Pharmacology
In a study, the administration of piracetam potentiated post-lesional plasticity, thus playing a neuroprotective role [176]. Transgenic mice models enabled modulation of plasticity. In humans, different drugs can improve brain reshaping after stroke or brain trauma [177]. The beneficial effect was observed in different functional areas. Norepinephrine, fluoxetine, paroxetine, scopolamine, and lorazepam improve cortical motor plasticity [178,179,180,181,182]. Meanwhile, amphetamine, bromocriptine, and piracetam reactivate brain parts in the left hemisphere and facilitate recovery after aphasia by modulating activity in language centers [183,184,185].
6.2. Transcranial Magnetic Stimulation
In post-stroke rehabilitation, transcranial magnetic stimulation (TMS) can potentiate motor learning [186]. It can rapidly elevate the excitability of primary motor cortex with a long-lasting effect [187]. The same technique modulates sensory maps: it can eliminate the deficit of spatial awareness for the contralesional space [188,189]. TMS can facilitate cognitive rehabilitation by improving memory and performance in picture-naming, analogic reasoning, and decision-making tasks [186,190,191,192,193]. Its mechanism of action is based on a modulation of the effective connectivity [194]. A combination of TMS, pharmacological intervention and rehabilitation is suggested.
6.3. Surgery
Cortical stimulation is an intervention performed for different indications. High-frequency chronic cortical stimulation efficiently modulates functional networks in movement disorders and chronic pain [195,196,197] This technique improves the functioning of the subcortico-cortical loops, which relieves motor, cognitive, and behavioral symptoms in Parkinson’s disease [198,199].
Surgical resection redistributes functional activity throughout latent networks. Hence, an incomplete removal of a tumor in eloquent areas reshapes the eloquent maps and extends functional sites. In a few years, the extended resection during the second surgery will not induce sequelae due to recruiting latent networks, unmasked after the first resection [200,201,202,203,204,205,206]. This approach allows to extend indications for surgeries in ’non-operable’ eloquent regions (sensory motor and language). Still, cortical plasticity may manifest only if subcotrical connectivity is not altered. Therefore, a stroke can cause permanent deficit due to the damage to the white matter [207]. For the same reason, resection of subcortical pathways may display sequelae despite a plastic potential of the cortex [208,209,210].
6.4. Transplantation
Observation of neural grafts explains how environment and experience can modulate brain function [211]. For example, the transplantation of neuroblasts from the fetal striatum to the same brain region may treat Huntington’s disease. The graft enhances cognitive performance and motor functioning by strengthening connections of the striato-cortical loop [212]. In Parkinson’s disease, the transfer of dopaminergic neural cells to putamen showed promising results [213]. After the basal ganglia infarction, the graft comprising cultured human neuronal cells can reduce motor deficit [214].
Conclusion
- The brain is a dynamic construct that changes structurally and/or functionally and constitutes interactive distributed glial-neuro-synaptic networks. Behavioral consequences of the changes may vary as a function of their effective connectivity, but the overall system remains stable due to homeostatic plasticity.
- A new insight into the concept of brain plasticity will provide perspectives on functional recovery following brain damage. The knowledge of this phenomenon will enable physicians to exploit cerebral plastic potential and regulate eloquent networks with timely interventions. Future studies will reveal pathophysiologic mechanisms of brain plasticity at microscopic and macroscopic levels, which will advance rehabilitation strategies and improve the quality of life in neurological diseases.
- Non-coding RNAs are optimal candidates for elucidating the molecular pathways underlying the phenomenon of brain plasticity. The candidates may signal the development of various neuropsychiatric disorders comprising schizophrenia, addiction, and fear-related anxiety disorders. The diversity of ncRNAs and their association with neurodegenerative diseases render them particularly interesting targets for new therapeutic approaches. New RNA-based therapeutics can arise from new data on the ncRNA regulation and the downstream effects of their interactions.
Author Contributions
conceptualization — M.L.; writing (original draft preparation) — Y.S. and N.V.K.; writing (review and editing)— N.V.K., Y.S.; subject investigation — Y.S. and N.V.K.; figure preparation – N.K.; supervision — M.L.; project administration — Y.S. and M.L.; funding acquisition — M.L. All authors contributed to the article and approved the submitted version.
Funding
The study was supported by ASPIRE, the technology program management pillar of Abu Dhabi’s Advanced Technology Research Council (ATRC), via the ASPIRE Precision Medicine Research Institute Abu Dhabi (ASPIREPMRIAD) award grant number VRI-20-10.
Conflicts of Interest
The research will be carried out without any financial or commercial ties that might be viewed as having a possible conflict of interest, according to the authors.
Patients and Public Involvement
Patients or the general public are not participants in the study.
Ethics and Dissemination
An ethics approval is not required for the review. The results of the study will be presented at scientific conferences as a poster or presentation in addition to being published in a peer-reviewed journal.
Abbreviations
The following abbreviations are used in this manuscript:
| AD | Alzheimer’s disease |
| AMPA | -amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid |
| BP | brain plasticity |
| BDNF | brain-derived neurotrophic factor |
| circRNA | circular RNA |
| ELAVL | embryonic lethal, abnormal vision, Drosophila-like |
| eRNA | enhancer RNA |
| FAT3 | FAT atypical cadherin 3 |
| GABA | gamma-aminobutyric acid |
| GAS5 | growth arrest specific 5 |
| HOTAIR | HOX transcript antisense RNA |
| Igfbp2 | insulin like growth factor binding protein 2 |
| lincRNA | long intergenic non-coding RNA |
| lncRNA | long non-coding RNA |
| MALAT1 | metastasis associated lung adenocarcinoma transcript 1 |
| mRNA | messengerRNA |
| miRNA | microRNA |
| ncRNA | non-coding RNA |
| piRNA | Piwi-interacting RNA |
| RMST | rhabdomyosarcoma 2 associated transcript |
| RNA | ribonucleic acid |
| TMS | transcranial magnetic stimulation |
| XIST | X inactive specific transcript |
| yRNA | Y RNA |
Glossary
References
- Johnston, M.V. Clinical disorders of brain plasticity. Brain and Development 2004, 26, 73–80. [Google Scholar] [CrossRef]
- Duffau, H. Brain plasticity: from pathophysiological mechanisms to therapeutic applications. Journal of clinical neuroscience 2006, 13, 885–897. [Google Scholar] [CrossRef]
- Kolb, B.; Gibb, R. Brain plasticity and behaviour in the developing brain. Journal of the Canadian Academy of Child and Adolescent Psychiatry 2011, 20, 265. [Google Scholar]
- Fischer, T.M.; Blazis, D.E.; Priver, N.A.; Carew, T.J. Metaplasticity at identified inhibitory synapses in Aplysia. Nature 1997, 389, 860–865. [Google Scholar] [CrossRef]
- Forrest, M.P.; Parnell, E.; Penzes, P. Dendritic structural plasticity and neuropsychiatric disease. Nature Reviews Neuroscience 2018, 19, 215–234. [Google Scholar] [CrossRef]
- Nature Porfolio. Spine Plasticity — Nature, Subjects, 2024. [Online; accessed 22-June-2024].
- Wikipedia contributors. Homeostatic plasticity — Wikipedia, The Free Encyclopedia, 2024. [Online; accessed 22-June-2024].
- Kujala, T.; Alho, K.; Näätänen, R. Cross-modal reorganization of human cortical functions. Trends in neurosciences 2000, 23, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Shimojo, S.; Shams, L. Sensory modalities are not separate modalities: plasticity and interactions. Current opinion in neurobiology 2001, 11, 505–509. [Google Scholar] [CrossRef]
- Bavelier, D.; Neville, H.J. Cross-modal plasticity: where and how? Nature Reviews Neuroscience 2002, 3, 443–452. [Google Scholar] [CrossRef]
- Rioult-Pedotti, M.S.; Friedman, D.; Donoghue, J.P. Learning-induced LTP in neocortex. science 2000, 290, 533–536. [Google Scholar] [CrossRef]
- Stephan, K.E.; Friston, K.J. Analyzing effective connectivity with functional magnetic resonance imaging. Wiley Interdisciplinary Reviews: Cognitive Science 2010, 1, 446–459. [Google Scholar] [CrossRef] [PubMed]
- La Rosa, C.; Parolisi, R.; Bonfanti, L. Brain structural plasticity: from adult neurogenesis to immature neurons. Frontiers in neuroscience 2020, 14, 512123. [Google Scholar] [CrossRef] [PubMed]
- Sale, A.; Berardi, N.; Maffei, L. Environment and brain plasticity: towards an endogenous pharmacotherapy. Physiological reviews 2014, 94, 189–234. [Google Scholar] [CrossRef] [PubMed]
- Mandolesi, L.; Gelfo, F.; Serra, L.; Montuori, S.; Polverino, A.; Curcio, G.; Sorrentino, G. Environmental factors promoting neural plasticity: insights from animal and human studies. Neural plasticity 2017, 2017, 7219461. [Google Scholar] [CrossRef] [PubMed]
- Buttelmann, F.; Karbach, J. Development and plasticity of cognitive flexibility in early and middle childhood. Frontiers in psychology 2017, 8, 258078. [Google Scholar] [CrossRef] [PubMed]
- Stampanoni Bassi, M.; Iezzi, E.; Gilio, L.; Centonze, D.; Buttari, F. Synaptic plasticity shapes brain connectivity: implications for network topology. International journal of molecular sciences 2019, 20, 6193. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Zheng, B. Axon plasticity in the mammalian central nervous system after injury. Trends in neurosciences 2014, 37, 583–593. [Google Scholar] [CrossRef]
- Marshall, K.L.; Farah, M.H. Axonal regeneration and sprouting as a potential therapeutic target for nervous system disorders. Neural Regeneration Research 2021, 16, 1901–1910. [Google Scholar]
- El Waly, B.; Macchi, M.; Cayre, M.; Durbec, P. Oligodendrogenesis in the normal and pathological central nervous system. Frontiers in neuroscience 2014, 8, 87323. [Google Scholar] [CrossRef]
- Fletcher, J.L.; Makowiecki, K.; Cullen, C.L.; Young, K.M. Oligodendrogenesis and myelination regulate cortical development, plasticity and circuit function. Seminars in cell & developmental biology. Elsevier, 2021, Vol. 118, pp. 14–23.
- Scott, H. Brain Plasticity Influencing Phantom Limb and Prosthetics. Outstanding Honors Theses 2011. [Google Scholar]
- Duffau, H. New insights into functional mapping in cerebral tumor surgery: study of the dynamic interactions between the lesion and the brain. Focus on Brain Mapping Research 2006, 1. [Google Scholar]
- Kolb, B.; Whishaw, I.Q. Brain plasticity and behavior. Annual review of psychology 1998, 49, 43–64. [Google Scholar] [CrossRef] [PubMed]
- Johnston, M.V.; Nishimura, A.; Harum, K.; Pekar, J.; Blue, M.E. Sculpting the developing brain. Advances in pediatrics 2001, 48, 1–40. [Google Scholar] [CrossRef] [PubMed]
- Gould, E.; Reeves, A.J.; Graziano, M.S.; Gross, C.G. Neurogenesis in the neocortex of adult primates. Science 1999, 286, 548–552. [Google Scholar] [CrossRef]
- Greenough, W.T. Structural correlates of information storage in the mammalian brain: a review and hypothesis. Trends in Neurosciences 1984, 7, 229–233. [Google Scholar] [CrossRef]
- Greenough, W.T.; Hwang, H.; Gorman, C. Evidence for active synapse formation or altered postsynaptic metabolism in visual cortex of rats reared in complex environments. Proceedings of the National Academy of Sciences 1985, 82, 4549–4552. [Google Scholar] [CrossRef]
- Turner, A.M.; Greenough, W.T. Differential rearing effects on rat visual cortex synapses. I. Synaptic and neuronal density and synapses per neuron. Brain research 1985, 329, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, B.; Schall, M.; Scheibel, A.B. A quantitative dendritic analysis of Wernicke’s area in humans. II. Gender, hemispheric, and environmental factors. Journal of Comparative Neurology 1993, 327, 97–111. [Google Scholar] [CrossRef]
- Purpura, D.P. Dendritic spine" dysgenesis" and mental retardation. Science 1974, 186, 1126–1128. [Google Scholar] [CrossRef]
- Wikipedia contributors. Neuroplasticity — Wikipedia, The Free Encyclopedia, 2024. [Online; accessed 24-June-2024].
- Nudo, R.J.; Wise, B.M.; SiFuentes, F.; Milliken, G.W. Neural substrates for the effects of rehabilitative training on motor recovery after ischemic infarct. Science 1996, 272, 1791–1794. [Google Scholar] [CrossRef]
- Georgopoulos, A.P. News in motor cortical physiology. News Physiol Sci 1999, 14, 64–8. [Google Scholar] [CrossRef]
- Sanes, J.N.; Schieber, M.H. Orderly somatotopy in primary motor cortex: does it exist? NeuroImage 2001, 13, 968–74. [Google Scholar] [CrossRef] [PubMed]
- Karni, A.; Meyer, G.; Rey-Hipolito, C.; Jezzard, P.; Adams, M.M.; Turner, R.; Ungerleider, L.G. The acquisition of skilled motor performance: fast and slow experience-driven changes in primary motor cortex. Proceedings of the National Academy of Sciences of the United States of America 1998, 95, 861–8. [Google Scholar] [CrossRef] [PubMed]
- Hlustik, P.; Solodkin, A.; Gullapalli, R.; Noll, D.C.; Small, S.L. Hand motor skill learning generalizes anatomically and behaviorally. NeuroImage 2000, 11, S866. [Google Scholar] [CrossRef]
- Pesenti, M.; Thioux, M.; Seron, X.; De Volder, A. Neuroanatomical substrates of arabic number processing, numerical comparison, and simple addition: a PET study. Journal of cognitive neuroscience 2000, 12, 461–79. [Google Scholar] [CrossRef] [PubMed]
- Ganis, G.; Keenan, J.P.; Kosslyn, S.M.; Pascual-Leone, A. Transcranial magnetic stimulation of primary motor cortex affects mental rotation. Cerebral cortex (New York, N.Y. : 1991) 2000, 10, 175–80. [Google Scholar] [CrossRef]
- Laubach, M.; Wessberg, J.; Nicolelis, M.A. Cortical ensemble activity increasingly predicts behaviour outcomes during learning of a motor task. Nature 2000, 405, 567–71. [Google Scholar] [CrossRef]
- Salenius, S.; Hari, R. Synchronous cortical oscillatory activity during motor action. Current opinion in neurobiology 2003, 13, 678–84. [Google Scholar] [CrossRef] [PubMed]
- Buchel, C.; Coull, J.; Friston, K.J. The predictive value of changes in effective connectivity for human learning. Science 1999, 283, 1538–1541. [Google Scholar] [CrossRef]
- Andres, F.G.; Gerloff, C. Coherence of sequential movements and motor learning. Journal of clinical neurophysiology : official publication of the American Electroencephalographic Society 1999, 16, 520–7. [Google Scholar] [CrossRef] [PubMed]
- Duffau, H.; Capelle, L.; Sichez, N.; Denvil, D.; Lopes, M.; Sichez, J.P.; Bitar, A.; Fohanno, D. Intraoperative mapping of the subcortical language pathways using direct stimulations. An anatomo-functional study. Brain 2002, 125, 199–214. [Google Scholar] [CrossRef]
- Duffau, H.; Gatignol, P.; Mandonnet, E.; Peruzzi, P.; Tzourio-Mazoyer, N.; Capelle, L. New insights into the anatomo-functional connectivity of the semantic system: A study using cortico-subcortical electrostimulations. Brain 2005, 128, 797–810. [Google Scholar] [CrossRef]
- McClelland, J.L.; Rogers, T.T. The parallel distributed processing approach to semantic cognition. Nature reviews neuroscience 2003, 4, 310–322. [Google Scholar] [CrossRef]
- Hatten, M.E. New directions in neuronal migration. Science 2002, 297, 1660–1663. [Google Scholar] [CrossRef] [PubMed]
- Gandolfo, F.; Li, C.S.; Benda, B.; Schioppa, C.P.; Bizzi, E. Cortical correlates of learning in monkeys adapting to a new dynamical environment. Proceedings of the National Academy of Sciences 2000, 97, 2259–2263. [Google Scholar] [CrossRef] [PubMed]
- Laubach, M.; Wessberg, J.; Nicolelis, M.A. Cortical ensemble activity increasingly predicts behaviour outcomes during learning of a motor task. Nature 2000, 405, 567–571. [Google Scholar] [CrossRef] [PubMed]
- Lamprecht, R.; LeDoux, J. Structural plasticity and memory. Nature Reviews Neuroscience 2004, 5, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Byrne, J.H. Synapses Plastic plasticity. Nature 1997, 389, 791–792. [Google Scholar] [CrossRef]
- Buonomano, D.V.; Merzenich, M.M. Cortical plasticity: from synapses to maps. Annual review of neuroscience 1998, 21, 149–186. [Google Scholar] [CrossRef] [PubMed]
- Aroniadou, V.A.; Keller, A. Mechanisms of LTP induction in rat motor cortex in vitro. Cerebral cortex 1995, 5, 353–362. [Google Scholar] [CrossRef]
- Braunewell, K.H.; Manahan-Vaughan, D. Long-term depression: a cellular basis for learning? Reviews in the Neurosciences 2001, 12, 121–140. [Google Scholar] [CrossRef] [PubMed]
- Fischer, T.M.; Blazis, D.E.; Priver, N.A.; Carew, T.J. Metaplasticity at identified inhibitory synapses in Aplysia. Nature 1997, 389, 860–5. [Google Scholar] [CrossRef]
- Martin, S.J.; Grimwood, P.D.; Morris, R.G. Synaptic plasticity and memory: an evaluation of the hypothesis. Annual review of neuroscience 2000, 23, 649–711. [Google Scholar] [CrossRef] [PubMed]
- Martin, S.J.; Grimwood, P.D.; Morris, R.G. Synaptic plasticity and memory: an evaluation of the hypothesis. Annual review of neuroscience 2000, 23, 649–711. [Google Scholar] [CrossRef]
- Widrow, B.; Kim, Y.; Park, D.; Perin, J.K. Nature’s learning rule: The Hebbian-LMS algorithm. In Artificial Intelligence in the Age of Neural Networks and Brain Computing; Elsevier, 2024; pp. 11–40.
- Cruikshank, S.J.; Weinberger, N.M. Evidence for the Hebbian hypothesis in experience-dependent physiological plasticity of neocortex: a critical review. Brain Research Reviews 1996, 22, 191–228. [Google Scholar] [CrossRef] [PubMed]
- Kilgard, M.P.; Merzenich, M.M. Cortical map reorganization enabled by nucleus basalis activity. Science 1998, 279, 1714–1718. [Google Scholar] [CrossRef]
- Andres, F.G.; Gerloff, C. Coherence of sequential movements and motor learning. Journal of clinical neurophysiology 1999, 16, 520. [Google Scholar] [CrossRef]
- Blitz, D.M.; Foster, K.A.; Regehr, W.G. Short-term synaptic plasticity: a comparison of two synapses. Nature Reviews Neuroscience 2004, 5, 630–640. [Google Scholar] [CrossRef] [PubMed]
- Malenka, R.C.; Nicoll, R.A. Silent synapses speak up. Neuron 1997, 19, 473–476. [Google Scholar] [CrossRef] [PubMed]
- Ridding, M.; Brouwer, B.; Miles, T.; Pitcher, J.; Thompson, P. Changes in muscle responses to stimulation of the motor cortex induced by peripheral nerve stimulation in human subjects. Experimental brain research 2000, 131, 135–143. [Google Scholar] [CrossRef]
- Fields, R.D.; Stevens-Graham, B. New insights into neuron-glia communication. Science 2002, 298, 556–562. [Google Scholar] [CrossRef]
- Haydon, P.G. GLIA: listening and talking to the synapse. Nature Reviews Neuroscience 2001, 2, 185–193. [Google Scholar] [CrossRef]
- Trachtenberg, J.T.; Chen, B.E.; Knott, G.W.; Feng, G.; Sanes, J.R.; Welker, E.; Svoboda, K. Long-term in vivo imaging of experience-dependent synaptic plasticity in adult cortex. Nature 2002, 420, 788–794. [Google Scholar] [CrossRef] [PubMed]
- Ivanco, T.L.; Greenough, W.T. Physiological consequences of morphologically detectable synaptic plasticity: potential uses for examining recovery following damage. Neuropharmacology 2000, 39, 765–776. [Google Scholar] [CrossRef]
- by Synaptic, H.D.I. Rapid Dendritic Morphogenesis in CA1. Cold Spring Harb. Symp. Quant. Biol, 1987, Vol. 52, p. 825.
- Poo, M.m. Neurotrophins as synaptic modulators. Nature reviews neuroscience 2001, 2, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Turrigiano, G.G.; Nelson, S.B. Homeostatic plasticity in the developing nervous system. Nature reviews neuroscience 2004, 5, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Selzer, M.E. Promotion of axonal regeneration in the injured CNS. The Lancet Neurology 2003, 2, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Ullian, E.M.; Sapperstein, S.K.; Christopherson, K.S.; Barres, B.A. Control of synapse number by glia. science 2001, 291, 657–661. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.; McCarthy, K.D. Plasticity of astrocytes. Glia 1994, 11, 147–155. [Google Scholar] [CrossRef]
- Langle, S.L.; Poulain, D.A.; Theodosis, D.T. Neuronal–glial remodeling: a structural basis for neuronal–glial interactions in the adult hypothalamus. Journal of physiology-Paris 2002, 96, 169–175. [Google Scholar] [CrossRef]
- Zhang, W.; Couldwell, W.T.; Simard, M.F.; Song, H.; Lin, J.H.; Nedergaard, M. Direct gap junction communication between malignant glioma cells and astrocytes. Cancer research 1999, 59, 1994–2003. [Google Scholar] [PubMed]
- Steindler, D.A.; Pincus, D.W. Stem cells and neuropoiesis in the adult human brain. The Lancet 2002, 359, 1047–1054. [Google Scholar] [CrossRef]
- Sanai, N.; Tramontin, A.D.; Quinones-Hinojosa, A.; Barbaro, N.M.; Gupta, N.; Kunwar, S.; Lawton, M.T.; McDermott, M.W.; Parsa, A.T.; Manuel-García Verdugo, J.; others. Unique astrocyte ribbon in adult human brain contains neural stem cells but lacks chain migration. Nature 2004, 427, 740–744. [Google Scholar] [CrossRef] [PubMed]
- Pincus, D.; Harrison-Restelli, C.; Barry, J.; Goodman, R.; Fraser, R.; Nedergaard, M.; Goldman, S. In vitro neurogenesis by adult human epileptic temporal neocortex. Clinical neurosurgery 1997, 44, 17–25. [Google Scholar] [PubMed]
- Roy, N.S.; Wang, S.; Jiang, L.; Kang, J.; Benraiss, A.; Harrison-Restelli, C.; Fraser, R.A.; Couldwell, W.T.; Kawaguchi, A.; Okano, H.; others. In vitro neurogenesis by progenitor cells isolated from the adult human hippocampus. Nature medicine 2000, 6, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Nunes, M.C.; Roy, N.S.; Keyoung, H.M.; Goodman, R.R.; McKhann, G.; Jiang, L.; Kang, J.; Nedergaard, M.; Goldman, S.A. Identification and isolation of multipotential neural progenitor cells from the subcortical white matter of the adult human brain. Nature medicine 2003, 9, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Gross, C.G. Neurogenesis in the adult brain: death of a dogma. Nature Reviews Neuroscience 2000, 1, 67–73. [Google Scholar] [CrossRef]
- Magavi, S.S.; Macklis, J.D. Induction of neuronal type-specific neurogenesis in the cerebral cortex of adult mice: manipulation of neural precursors in situ. Developmental brain research 2002, 134, 57–76. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.K.; Botez, M. Diaschisis and neurobehavior. Canadian journal of neurological sciences 1998, 25, 5–12. [Google Scholar] [CrossRef]
- Seitz, R.J.; Azari, N.P.; Knorr, U.; Binkofski, F.; Herzog, H.; Freund, H.J. The role of diaschisis in stroke recovery. Stroke 1999, 30, 1844–1850. [Google Scholar] [CrossRef]
- Duffau, H.; Sichez, J.P.; Lehéricy, S. Intraoperative unmasking of brain redundant motor sites during resection of a precentral angioma: evidence using direct cortical stimulation. Annals of neurology 2000, 47, 132–135. [Google Scholar] [CrossRef]
- Duffau, H. Acute functional reorganisation of the human motor cortex during resection of central lesions: a study using intraoperative brain mapping. Journal of Neurology, Neurosurgery & Psychiatry 2001, 70, 506–513. [Google Scholar]
- Rijntjes, M.; Weiller, C. Recovery of motor and language abilities after stroke: the contribution of functional imaging. Progress in neurobiology 2002, 66, 109–122. [Google Scholar] [CrossRef] [PubMed]
- Krainik, A.; Duffau, H.; Capelle, L.; Cornu, P.; Boch, A.L.; Mangin, J.F.; Le Bihan, D.; Marsault, C.; Chiras, J.; Lehéricy, S. Role of the healthy hemisphere in recovery after resection of the supplementary motor area. Neurology 2004, 62, 1323–1332. [Google Scholar] [CrossRef] [PubMed]
- Levänen, S.; Jousmäki, V.; Hari, R. Vibration-induced auditory-cortex activation in a congenitally deaf adult. Current Biology 1998, 8, 869–872. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.S.; Lee, J.S.; Oh, S.H.; Kim, S.K.; Kim, J.W.; Chung, J.K.; Lee, M.C.; Kim, C.S. Cross-modal plasticity and cochlear implants. Nature 2001, 409, 149–50. [Google Scholar] [CrossRef] [PubMed]
- Rossini, P.M.; Dal Forno, G. Integrated technology for evaluation of brain function and neural plasticity. Physical medicine and rehabilitation clinics of North America 2004, 15, 263–306. [Google Scholar] [CrossRef] [PubMed]
- Luders, E.; Gaser, C.; Jancke, L.; Schlaug, G. A voxel-based approach to gray matter asymmetries. Neuroimage 2004, 22, 656–664. [Google Scholar] [CrossRef]
- Josse, G.; Mazoyer, B.; Crivello, F.; Tzourio-Mazoyer, N. Left planum temporale: an anatomical marker of left hemispheric specialization for language comprehension. Cognitive Brain Research 2003, 18, 1–14. [Google Scholar] [CrossRef]
- Sluming, V.; Barrick, T.; Howard, M.; Cezayirli, E.; Mayes, A.; Roberts, N. Voxel-based morphometry reveals increased gray matter density in Broca’s area in male symphony orchestra musicians. Neuroimage 2002, 17, 1613–1622. [Google Scholar] [CrossRef]
- Hutchinson, S.; Lee, L.H.L.; Gaab, N.; Schlaug, G. Cerebellar volume of musicians. Cerebral cortex 2003, 13, 943–949. [Google Scholar] [CrossRef]
- Mackay, C.E.; Roberts, N.; Mayes, A.R.; Downes, J.J.; Foster, J.K.; Mann, D. An exploratory study of the relationship between face recognition memory and the volume of medial temporal lobe structures in healthy young males. Behavioural Neurology 1998, 11, 3–20. [Google Scholar] [CrossRef]
- Maguire, E.A.; Gadian, D.G.; Johnsrude, I.S.; Good, C.D.; Ashburner, J.; Frackowiak, R.S.; Frith, C.D. Navigation-related structural change in the hippocampi of taxi drivers. Proceedings of the National Academy of Sciences 2000, 97, 4398–4403. [Google Scholar] [CrossRef]
- Paus, T.; Zijdenbos, A.; Worsley, K.; Collins, D.L.; Blumenthal, J.; Giedd, J.N.; Rapoport, J.L.; Evans, A.C. Structural maturation of neural pathways in children and adolescents: in vivo study. Science 1999, 283, 1908–1911. [Google Scholar] [CrossRef]
- Draganski, B.; Gaser, C.; Busch, V.; Schuierer, G.; Bogdahn, U.; May, A. Changes in grey matter induced by training. Nature 2004, 427, 311–312. [Google Scholar] [CrossRef]
- Juraska, J.M. Sex differences in dendritic response to differential experience in the rat visual cortex. Brain Research 1984, 295, 27–34. [Google Scholar] [CrossRef]
- JURASKA, J.M. Sex differences in developmental plasticity of behavior and the brain. Developmental neuropsychobiology 1986, 409–422. [Google Scholar]
- Juraska, J.M.; Fitch, J.M.; Henderson, C.; Rivers, N. Sex differences in the dendritic branching of dentate granule cells following differential experience. Brain research 1985, 333, 73–80. [Google Scholar] [CrossRef]
- Juraska, J.M. The structure of the rat cerebral cortex: Effects of gender and the environment. 1990.
- Coleman, P.; Buell, S. Regulation of dendritic extent in developing and aging brain. Synaptic plasticity 1985, 311–333. [Google Scholar]
- Duque, A.; Arellano, J.I.; Rakic, P. An assessment of the existence of adult neurogenesis in humans and value of its rodent models for neuropsychiatric diseases. Molecular psychiatry 2022, 27, 377–382. [Google Scholar] [CrossRef]
- Raff, M.C.; Barres, B.A.; Burne, J.F.; Coles, H.S.; Ishizaki, Y.; Jacobson, M.D. Programmed cell death and the control of cell survival: lessons from the nervous system. Science 1993, 262, 695–700. [Google Scholar] [CrossRef] [PubMed]
- Johansson, B.B. Brain plasticity in health and disease. The Keio journal of medicine 2004, 53, 231–246. [Google Scholar] [CrossRef] [PubMed]
- Statsenko, Y.; Habuza, T.; Charykova, I.; Gorkom, K.N.V.; Zaki, N.; Almansoori, T.M.; Baylis, G.; Ljubisavljevic, M.; Belghali, M. Predicting age from behavioral test performance for screening early onset of cognitive decline. Frontiers in Aging Neuroscience 2021, 13, 661514. [Google Scholar] [CrossRef]
- Statsenko, Y.; Habuza, T.; Gorkom, K.N.V.; Zaki, N.; Almansoori, T.M.; Al Zahmi, F.; Ljubisavljevic, M.R.; Belghali, M. Proportional changes in cognitive subdomains during normal brain aging. Frontiers in Aging Neuroscience 2021, 13, 673469. [Google Scholar] [CrossRef] [PubMed]
- Statsenko, Y.; Habuza, T.; Smetanina, D.; Simiyu, G.L.; Uzianbaeva, L.; Neidl-Van Gorkom, K.; Zaki, N.; Charykova, I.; Al Koteesh, J.; Almansoori, T.M.; others. Brain morphometry and cognitive performance in normal brain aging: age-and sex-related structural and functional changes. Frontiers in aging neuroscience 2022, 13, 713680. [Google Scholar] [CrossRef]
- Belghali, M.; Statsenko, Y.; Laver, V. Stroop switching card test: brief screening of executive functions across the lifespan. Aging, Neuropsychology, and Cognition 2022, 29, 14–33. [Google Scholar] [CrossRef]
- Statsenko, Y.; Habuza, T.; Uzianbaeva, L.; Gorkom, K.; Belghali, M.; Charykova, I.; others. Correlation between lifelong dynamics of psychophysiological performance and brain morphology. ESNR 2021. Neuroradiology 2021, 63, 41–42. [Google Scholar]
- Habuza, T.; Statsenko, Y.; Uzianbaeva, L.; Gorkom, K.; Zaki, N.; Belghali, M.; others. Models of brain cognitive and morphological changes across the life: machine learning-based approach. ESNR 2021. Neuroradiology 2021, 63, 10–26226. [Google Scholar]
- Uzianbaeva, L.; Statsenko, Y.; Habuza, T.; Gorkom, K.; Belghali, M.; Charykova, I.; others. Effects of sex age-related changes in brain morphology. ESNR 2021. Neuroradiology 2021, 63, 42–43. [Google Scholar]
- Gorkom, K.; Statsenko, Y.; Habuza, T.; Uzianbaeva, L.; Belghali, M.; Charykova, I.; others. Comparison of brain volumetric changes with functional outcomes in physiologic brain aging. ESNR 2021. Neuroradiology 2021, 63, 43–44. [Google Scholar]
- Statsenko, Y.; Habuza, T.; Charykova, I.; Gorkom, K.; Zaki, N.; Almansoori, T.; Ljubisavljevic, M.; Szolics, M.; Al Koteesh, J.; Ponomareva, A.; others. AI models of age-associated changes in CNS composition identified by MRI. Journal of the Neurological Sciences 2021, 429. [Google Scholar] [CrossRef]
- Habuza, T.; Zaki, N.; Statsenko, Y.; Elyassami, S. MRI and cognitive tests-based screening tool for dementia. Journal of the Neurological Sciences 2021, 429. [Google Scholar] [CrossRef]
- Statsenko, Y.; Habuza, T.; Gorkom, K.N.V.; Zaki, N.; Almansoori, T.M. Applying the inverse efficiency score to visual–motor task for studying speed-accuracy performance while aging. Frontiers in Aging Neuroscience 2020, 12, 574401. [Google Scholar] [CrossRef]
- Habuza, T.; Zaki, N.; Statsenko, Y.; Alnajjar, F.; Elyassami, S. Predicting the diagnosis of dementia from MRI data: added value to cognitive tests. The 7th Annual International Conference on Arab Women in Computing in Conjunction with the 2nd Forum of Women in Research, 2021, pp. 1–7.
- Habuza, T.; Zaki, N.; Mohamed, E.A.; Statsenko, Y. Deviation from model of normal aging in alzheimer’s disease: Application of deep learning to structural MRI data and cognitive tests. IEEE Access 2022, 10, 53234–53249. [Google Scholar] [CrossRef]
- Statsenko, Y.; Meribout, S.; Habuza, T.; Almansoori, T.M.; Gorkom, K.N.V.; Gelovani, J.G.; Ljubisavljevic, M. Patterns of structure-function association in normal aging and in Alzheimer’s disease: Screening for mild cognitive impairment and dementia with ML regression and classification models. Frontiers in Aging Neuroscience 2023, 14, 943566. [Google Scholar] [CrossRef]
- Smetanina, D.; Meribout, S.; Habuza, T.; Simiyu, G.; Ismail, F.; Zareba, K.; Gorkom, K.; Almansoori, T.; Gelovani, J.; Ljubisavljevic, M.; others. Reference curves of age-related volumetric changes in hippocampus and brain ventricles in healthy population. Journal of the Neurological Sciences 2023, 455. [Google Scholar] [CrossRef]
- Meribout, S.; Habuza, T.; Smetanina, D.; Simiyu, G.; Ismail, F.; Gorkom, K.; Almansoori, T.; Gelovani, J.; Statsenko, Y.; Ljubisavljevic, M. Rate and onset of cognitive decline and cortical atrophy in normal and pathological ageing. Journal of the Neurological Sciences 2023, 455. [Google Scholar] [CrossRef]
- Meribout, S.; Habuza, T.; Smetanina, D.; Simiyu, G.; Ismail, F.; Gorkom, K.; Almansoori, T.; Gelovani, J.; Statsenko, Y.; Ljubisavljevic, M. Functional changes in age-related neurocognitive slowing and disease-related cognitive decline: Evidence from global cognitive tests and psychophysiological tests. Journal of the Neurological Sciences 2023, 455. [Google Scholar] [CrossRef]
- Statsenko, Y.; Meribout, S.; Habuza, T.; Smetanina, D.; Simiyu, G.; Ismail, F.; Gorkom, K.; Almansoori, T.; Gelovani, J.; Ljubisavljevic, M. Structure-function association patterns of the brain in individuals with different level of cognitive impairment. Journal of the Neurological Sciences 2023, 455. [Google Scholar] [CrossRef]
- Spadaro, P.A.; Bredy, T.W. Emerging role of non-coding RNA in neural plasticity, cognitive function, and neuropsychiatric disorders. Frontiers in genetics 2012, 3, 132. [Google Scholar] [CrossRef]
- Kyzar, E.J.; Bohnsack, J.P.; Pandey, S.C. Current and future perspectives of noncoding RNAs in brain function and neuropsychiatric disease. Biological psychiatry 2022, 91, 183–193. [Google Scholar] [CrossRef]
- Earls, L.R.; Westmoreland, J.J.; Zakharenko, S.S. Non-coding RNA regulation of synaptic plasticity and memory: implications for aging. Ageing research reviews 2014, 17, 34–42. [Google Scholar] [CrossRef]
- Keihani, S.; Kluever, V.; Fornasiero, E.F. Brain long noncoding RNAs: multitask regulators of neuronal differentiation and function. Molecules 2021, 26, 3951. [Google Scholar] [CrossRef]
- Ip, J.Y.; Sone, M.; Nashiki, C.; Pan, Q.; Kitaichi, K.; Yanaka, K.; Abe, T.; Takao, K.; Miyakawa, T.; Blencowe, B.J.; others. Gomafu lncRNA knockout mice exhibit mild hyperactivity with enhanced responsiveness to the psychostimulant methamphetamine. Scientific reports 2016, 6, 27204. [Google Scholar] [CrossRef]
- Zhao, H.; Jin, T.; Cheng, X.; Qin, J.; Zhang, L.; He, H.; Xue, J.; Jin, G. GAS5 which is regulated by Lhx8 promotes the recovery of learning and memory in rats with cholinergic nerve injury. Life sciences 2020, 260, 118388. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zhuang, Y.; Zhao, X.; Li, X. Long non-coding RNA in neuronal development and neurological disorders. Frontiers in genetics 2019, 9, 436598. [Google Scholar] [CrossRef]
- Bhan, A.; Soleimani, M.; Mandal, S.S. Long noncoding RNA (lncRNA): Functions in health and disease. Gene Regulation, Epigenetics and Hormone Signaling 2017, 169–208. [Google Scholar]
- Khalifa, F.N.; Hussein, R.F.; Mekawy, D.M.; Elwi, H.M.; Alsaeed, S.A.; Elnawawy, Y.; Shaheen, S.H. Potential role of the lncRNA. Molecular Biology Reports 2024, 51, 1–11. [Google Scholar]
- Sim, S.E.; Lim, C.S.; Kim, J.I.; Seo, D.; Chun, H.; Yu, N.K.; Lee, J.; Kang, S.J.; Ko, H.G.; Choi, J.H.; others. The brain-enriched microRNA miR-9-3p regulates synaptic plasticity and memory. Journal of Neuroscience 2016, 36, 8641–8652. [Google Scholar] [CrossRef]
- McNeill, E.M.; Warinner, C.; Alkins, S.; Taylor, A.; Heggeness, H.; DeLuca, T.F.; Fulga, T.A.; Wall, D.P.; Griffith, L.C.; Van Vactor, D. The conserved microRNA miR-34 regulates synaptogenesis via coordination of distinct mechanisms in presynaptic and postsynaptic cells. Nature communications 2020, 11, 1092. [Google Scholar] [CrossRef]
- Qian, Y.; Song, J.; Ouyang, Y.; Han, Q.; Chen, W.; Zhao, X.; Xie, Y.; Chen, Y.; Yuan, W.; Fan, C. Advances in roles of miR-132 in the nervous system. Frontiers in Pharmacology 2017, 8, 770. [Google Scholar] [CrossRef]
- Xin, H.; Katakowski, M.; Wang, F.; Qian, J.Y.; Liu, X.S.; Ali, M.M.; Buller, B.; Zhang, Z.G.; Chopp, M. MicroRNA-17–92 cluster in exosomes enhance neuroplasticity and functional recovery after stroke in rats. Stroke 2017, 48, 747–753. [Google Scholar] [CrossRef]
- Jin, J.; Kim, S.N.; Liu, X.; Zhang, H.; Zhang, C.; Seo, J.S.; Kim, Y.; Sun, T. miR-17-92 cluster regulates adult hippocampal neurogenesis, anxiety, and depression. Cell reports 2016, 16, 1653–1663. [Google Scholar] [CrossRef]
- Wu, X.; Zhang, Y.; Wang, P.; Li, X.; Song, Z.; Wei, C.; Zhang, Q.; Luo, B.; Liu, Z.; Yang, Y.; others. Clinical and preclinical evaluation of miR-144-5p as a key target for major depressive disorder. CNS Neuroscience & Therapeutics 2023, 29, 3598–3611. [Google Scholar]
- Gao, C.; Yin, F.; Li, R.; Ruan, Q.; Meng, C.; Zhao, K.; Zhu, Q. MicroRNA-145-Mediated KDM6A Downregulation Enhances Neural Repair after Spinal Cord Injury via the NOTCH2/Abcb1a Axis. Oxidative Medicine and Cellular Longevity 2021, 2021, 2580619. [Google Scholar] [CrossRef]
- Qiao, J.; Zhao, J.; Chang, S.; Sun, Q.; Liu, N.; Dong, J.; Chen, Y.; Yang, D.; Ye, D.; Liu, X.; others. MicroRNA-153 improves the neurogenesis of neural stem cells and enhances the cognitive ability of aged mice through the notch signaling pathway. Cell Death & Differentiation 2020, 27, 808–825. [Google Scholar]
- Del Val, C.; Díaz de la Guardia-Bolívar, E.; Zwir, I.; Mishra, P.P.; Mesa, A.; Salas, R.; Poblete, G.F.; de Erausquin, G.; Raitoharju, E.; Kähönen, M. ; others. Gene expression networks regulated by human personality. Molecular Psychiatry 2024, 1–20. [Google Scholar]
- Seeler, S.; Andersen, M.S.; Sztanka-Toth, T.; Rybiczka-Tešulov, M.; van den Munkhof, M.H.; Chang, C.C.; Maimaitili, M.; Venø, M.T.; Hansen, T.B.; Pasterkamp, R.J.; others. A circular RNA expressed from the FAT3 locus regulates neural development. Molecular Neurobiology 2023, 60, 3239–3260. [Google Scholar] [CrossRef]
- Du, M.; Wu, C.; Yu, R.; Cheng, Y.; Tang, Z.; Wu, B.; Fu, J.; Tan, W.; Zhou, Q.; Zhu, Z.; others. A novel circular RNA, circIgfbp2, links neural plasticity and anxiety through targeting mitochondrial dysfunction and oxidative stress-induced synapse dysfunction after traumatic brain injury. Molecular Psychiatry 2022, 27, 4575–4589. [Google Scholar] [CrossRef]
- Bao, N.; Liu, J.; Peng, Z.; Zhang, R.; Ni, R.; Li, R.; Wu, J.; Liu, Z.; Pan, B. Identification of circRNA-miRNA-mRNA networks to explore the molecular mechanism and immune regulation of postoperative neurocognitive disorder. Aging (Albany NY) 2022, 14, 8374. [Google Scholar] [CrossRef]
- Mahmoudi, E.; Fitzsimmons, C.; Geaghan, M.P.; Shannon Weickert, C.; Atkins, J.R.; Wang, X.; Cairns, M.J. Circular RNA biogenesis is decreased in postmortem cortical gray matter in schizophrenia and may alter the bioavailability of associated miRNA. Neuropsychopharmacology 2019, 44, 1043–1054. [Google Scholar] [CrossRef]
- Brookes, E.; Alan Au, H.Y.; Varsally, W.; Barrington, C.; Hadjur, S.; Riccio, A. A novel enhancer that regulates Bdnf expression in developing neurons. bioRxiv 2021. pp. 2021–11. [Google Scholar]
- Cajigas, I.; Chakraborty, A.; Swyter, K.R.; Luo, H.; Bastidas, M.; Nigro, M.; Morris, E.R.; Chen, S.; VanGompel, M.J.; Leib, D.; others. The Evf2 ultraconserved enhancer lncRNA functionally and spatially organizes megabase distant genes in the developing forebrain. Molecular cell 2018, 71, 956–972. [Google Scholar] [CrossRef]
- Sauvageau, M.; Goff, L.A.; Lodato, S.; Bonev, B.; Groff, A.F.; Gerhardinger, C.; Sanchez-Gomez, D.B.; Hacisuleyman, E.; Li, E.; Spence, M.; others. Multiple knockout mouse models reveal lincRNAs are required for life and brain development. elife 2013, 2, e01749. [Google Scholar] [CrossRef] [PubMed]
- Chanda, K.; Mukhopadhyay, D. LncRNA Xist, X-chromosome instability and Alzheimer’s disease. Current Alzheimer Research 2020, 17, 499–507. [Google Scholar] [CrossRef]
- Rajasethupathy, P.; Antonov, I.; Sheridan, R.; Frey, S.; Sander, C.; Tuschl, T.; Kandel, E.R. A role for neuronal piRNAs in the epigenetic control of memory-related synaptic plasticity. Cell 2012, 149, 693–707. [Google Scholar] [CrossRef] [PubMed]
- Zuo, L.; Wang, Z.; Tan, Y.; Chen, X.; Luo, X. piRNAs and their functions in the brain. International journal of human genetics 2016, 16, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Qiu, W.; Guo, X.; Lin, X.; Yang, Q.; Zhang, W.; Zhang, Y.; Zuo, L.; Zhu, Y.; Li, C.S.R.; Ma, C.; others. Transcriptome-wide piRNA profiling in human brains of Alzheimer’s disease. Neurobiology of aging 2017, 57, 170–177. [Google Scholar] [CrossRef]
- Scheckel, C.; Drapeau, E.; Frias, M.A.; Park, C.Y.; Fak, J.; Zucker-Scharff, I.; Kou, Y.; Haroutunian, V.; Ma’ayan, A.; Buxbaum, J.D.; others. Regulatory consequences of neuronal ELAV-like protein binding to coding and non-coding RNAs in human brain. Elife 2016, 5, e10421. [Google Scholar] [CrossRef]
- Wei, Z.; Batagov, A.O.; Schinelli, S.; Wang, J.; Wang, Y.; El Fatimy, R.; Rabinovsky, R.; Balaj, L.; Chen, C.C.; Hochberg, F.; others. Coding and noncoding landscape of extracellular RNA released by human glioma stem cells. Nature communications 2017, 8, 1145. [Google Scholar] [CrossRef]
- Haack, F.; Trakooljul, N.; Gley, K.; Murani, E.; Hadlich, F.; Wimmers, K.; Ponsuksili, S. Deep sequencing of small non-coding RNA highlights brain-specific expression patterns and RNA cleavage. RNA biology 2019, 16, 1764–1774. [Google Scholar] [CrossRef]
- Zimmer-Bensch, G. Emerging roles of long non-coding RNAs as drivers of brain evolution. Cells 2019, 8, 1399. [Google Scholar] [CrossRef]
- Bernard, D.; Prasanth, K.V.; Tripathi, V.; Colasse, S.; Nakamura, T.; Xuan, Z.; Zhang, M.Q.; Sedel, F.; Jourdren, L.; Coulpier, F.; others. A long nuclear-retained non-coding RNA regulates synaptogenesis by modulating gene expression. The EMBO journal 2010, 29, 3082–3093. [Google Scholar] [CrossRef]
- Tripathi, V.; Ellis, J.D.; Shen, Z.; Song, D.Y.; Pan, Q.; Watt, A.T.; Freier, S.M.; Bennett, C.F.; Sharma, A.; Bubulya, P.A.; others. The nuclear-retained noncoding RNA MALAT1 regulates alternative splicing by modulating SR splicing factor phosphorylation. Molecular cell 2010, 39, 925–938. [Google Scholar] [CrossRef]
- Liau, W.S.; Zhao, Q.; Bademosi, A.; Gormal, R.S.; Gong, H.; Marshall, P.R.; Periyakaruppiah, A.; Madugalle, S.U.; Zajaczkowski, E.L.; Leighton, L.J.; others. Fear extinction is regulated by the activity of long noncoding RNAs at the synapse. Nature Communications 2023, 14, 7616. [Google Scholar] [CrossRef]
- Lu, T.X.; Rothenberg, M.E. MicroRNA. Journal of allergy and clinical immunology 2018, 141, 1202–1207. [Google Scholar] [CrossRef]
- Starega-Roslan, J.; Krol, J.; Koscianska, E.; Kozlowski, P.; Szlachcic, W.J.; Sobczak, K.; Krzyzosiak, W.J. Structural basis of microRNA length variety. Nucleic acids research 2011, 39, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Fiore, R.; Siegel, G.; Schratt, G. MicroRNA function in neuronal development, plasticity and disease. Biochimica et Biophysica Acta (BBA)-Gene Regulatory Mechanisms 2008, 1779, 471–478. [Google Scholar] [CrossRef] [PubMed]
- Martins, H.C.; Schratt, G. MicroRNA-dependent control of neuroplasticity in affective disorders. Translational Psychiatry 2021, 11, 263. [Google Scholar] [CrossRef] [PubMed]
- Constantin, L. Circular RNAs and neuronal development. Circular RNAs: Biogenesis and Functions 2018, 205–213. [Google Scholar]
- Kim, T.K.; Hemberg, M.; Gray, J.M.; Costa, A.M.; Bear, D.M.; Wu, J.; Harmin, D.A.; Laptewicz, M.; Barbara-Haley, K.; Kuersten, S.; others. Widespread transcription at neuronal activity-regulated enhancers. Nature 2010, 465, 182–187. [Google Scholar] [CrossRef]
- Arner, E.; Daub, C.O.; Vitting-Seerup, K.; Andersson, R.; Lilje, B.; Drabløs, F.; Lennartsson, A.; Rönnerblad, M.; Hrydziuszko, O.; Vitezic, M.; others. Transcribed enhancers lead waves of coordinated transcription in transitioning mammalian cells. Science 2015, 347, 1010–1014. [Google Scholar] [CrossRef] [PubMed]
- Gray, J.M.; Kim, T.K.; West, A.E.; Nord, A.S.; Markenscoff-Papadimitriou, E.; Lomvardas, S. Genomic views of transcriptional enhancers: essential determinants of cellular identity and activity-dependent responses in the CNS. Journal of Neuroscience 2015, 35, 13819–13826. [Google Scholar] [CrossRef]
- Roberts, T.C.; Morris, K.V.; Wood, M.J. The role of long non-coding RNAs in neurodevelopment, brain function and neurological disease. Philosophical Transactions of the Royal Society B: Biological Sciences 2014, 369, 20130507. [Google Scholar] [CrossRef] [PubMed]
- Gu, S.; Xie, R.; Liu, X.; Shou, J.; Gu, W.; Che, X. Long coding RNA XIST contributes to neuronal apoptosis through the downregulation of AKT phosphorylation and is negatively regulated by miR-494 in rat spinal cord injury. International journal of molecular sciences 2017, 18, 732. [Google Scholar] [CrossRef]
- Aravin, A.; Gaidatzis, D.; Pfeffer, S.; Lagos-Quintana, M.; Landgraf, P.; Iovino, N.; Morris, P.; Brownstein, M.J.; Kuramochi-Miyagawa, S.; Nakano, T.; others. A novel class of small RNAs bind to MILI protein in mouse testes. Nature 2006, 442, 203–207. [Google Scholar] [CrossRef]
- Girard, A.; Sachidanandam, R.; Hannon, G.J.; Carmell, M.A. A germline-specific class of small RNAs binds mammalian Piwi proteins. Nature 2006, 442, 199–202. [Google Scholar] [CrossRef]
- Betel, D.; Sheridan, R.; Marks, D.S.; Sander, C. Computational analysis of mouse piRNA sequence and biogenesis. PLoS computational biology 2007, 3, e222. [Google Scholar] [CrossRef]
- Xerri, C.; Zennou-Azogui, Y. Influence of the postlesion environment and chronic piracetam treatment on the organization of the somatotopic map in the rat primary somatosensory cortex after focal cortical injury. Neuroscience 2003, 118, 161–177. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, L.B. Neuropharmacology of TBI-induced plasticity. Brain injury 2003, 17, 685–94. [Google Scholar] [CrossRef] [PubMed]
- Plewnia, C.; Hoppe, J.; Cohen, L.G.; Gerloff, C. Improved motor skill acquisition after selective stimulation of central norepinephrine. Neurology 2004, 62, 2124–6. [Google Scholar] [CrossRef] [PubMed]
- Loubinoux, I.; Pariente, J.; Boulanouar, K.; Carel, C.; Manelfe, C.; Rascol, O.; Celsis, P.; Chollet, F. A single dose of the serotonin neurotransmission agonist paroxetine enhances motor output: double-blind, placebo-controlled, fMRI study in healthy subjects. NeuroImage 2002, 15, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Dam, M.; Tonin, P.; De Boni, A.; Pizzolato, G.; Casson, S.; Ermani, M.; Freo, U.; Piron, L.; Battistin, L. Effects of fluoxetine and maprotiline on functional recovery in poststroke hemiplegic patients undergoing rehabilitation therapy. Stroke; a journal of cerebral circulation 1996, 27, 1211–4. [Google Scholar] [CrossRef] [PubMed]
- Sawaki, L.; Boroojerdi, B.; Kaelin-Lang, A.; Burstein, A.H.; Bütefisch, C.M.; Kopylev, L.; Davis, B.; Cohen, L.G. Cholinergic influences on use-dependent plasticity. Journal of neurophysiology 2002, 87, 166–71. [Google Scholar] [CrossRef] [PubMed]
- Ziemann, U.; Lönnecker, S.; Steinhoff, B.J.; Paulus, W. The effect of lorazepam on the motor cortical excitability in man. Experimental brain research 1996, 109, 127–35. [Google Scholar] [CrossRef]
- Walker-Batson, D.; Curtis, S.; Natarajan, R.; Ford, J.; Dronkers, N.; Salmeron, E.; Lai, J.; Unwin, D.H. A double-blind, placebo-controlled study of the use of amphetamine in the treatment of aphasia. Stroke; a journal of cerebral circulation 2001, 32, 2093–8. [Google Scholar] [CrossRef] [PubMed]
- Bragoni, M.; Altieri, M.; Di Piero, V.; Padovani, A.; Mostardini, C.; Lenzi, G.L. Bromocriptine and speech therapy in non-fluent chronic aphasia after stroke. Neurological sciences : official journal of the Italian Neurological Society and of the Italian Society of Clinical Neurophysiology 2000, 21, 19–22. [Google Scholar] [CrossRef]
- Kessler, J.; Thiel, A.; Karbe, H.; Heiss, W.D. Piracetam improves activated blood flow and facilitates rehabilitation of poststroke aphasic patients. Stroke; a journal of cerebral circulation 2000, 31, 2112–6. [Google Scholar] [CrossRef]
- Rossi, S.; Rossini, P.M. TMS in cognitive plasticity and the potential for rehabilitation. Trends in cognitive sciences 2004, 8, 273–9. [Google Scholar] [CrossRef] [PubMed]
- Stefan, K.; Kunesch, E.; Cohen, L.G.; Benecke, R.; Classen, J. Induction of plasticity in the human motor cortex by paired associative stimulation. Brain : a journal of neurology 2000, 123 Pt 3, 572–84. [Google Scholar] [CrossRef] [PubMed]
- Oliveri, M.; Bisiach, E.; Brighina, F.; Piazza, A.; La Bua, V.; Buffa, D.; Fierro, B. rTMS of the unaffected hemisphere transiently reduces contralesional visuospatial hemineglect. Neurology 2001, 57, 1338–40. [Google Scholar] [CrossRef] [PubMed]
- Brighina, F.; Bisiach, E.; Oliveri, M.; Piazza, A.; La Bua, V.; Daniele, O.; Fierro, B. 1 Hz repetitive transcranial magnetic stimulation of the unaffected hemisphere ameliorates contralesional visuospatial neglect in humans. Neuroscience Letter 2003, 336, 131–3. [Google Scholar] [CrossRef] [PubMed]
- Mottaghy, F.M.; Hungs, M.; Brügmann, M.; Sparing, R.; Boroojerdi, B.; Foltys, H.; Huber, W.; Töpper, R. Facilitation of picture naming after repetitive transcranial magnetic stimulation. Neurology 1999, 53, 1806–12. [Google Scholar] [CrossRef] [PubMed]
- Grafman, J.; Wassermann, E. Transcranial magnetic stimulation can measure and modulate learning and memory. Neuropsychologia 1999, 37, 159–67. [Google Scholar] [CrossRef] [PubMed]
- Boylan, L.S. Enhancing analogic reasoning with rTMS over the left prefrontal cortex. Neurology 2001, 57, 1349. [Google Scholar] [CrossRef]
- Evers, S.; Böckermann, I.; Nyhuis, P.W. The impact of transcranial magnetic stimulation on cognitive processing: an event-related potential study. Neuroreport 2001, 12, 2915–8. [Google Scholar] [CrossRef]
- Lee, L.; Siebner, H.R.; Rowe, J.B.; Rizzo, V.; Rothwell, J.C.; Frackowiak, R.S.J.; Friston, K.J. Acute remapping within the motor system induced by low-frequency repetitive transcranial magnetic stimulation. The Journal of neuroscience : the official journal of the Society for Neuroscience 2003, 23, 5308–18. [Google Scholar] [CrossRef]
- Canavero, S.; Bonicalzi, V.; Paolotti, R.; Castellano, G.; Greco-Crasto, S.; Rizzo, L.; Davini, O.; Maina, R. Therapeutic extradural cortical stimulation for movement disorders: a review. Neurological research 2003, 25, 118–22. [Google Scholar] [CrossRef]
- Tsubokawa, T.; Katayama, Y.; Yamamoto, T.; Hirayama, T.; Koyama, S. Chronic motor cortex stimulation in patients with thalamic pain. Journal of neurosurgery 1993, 78, 393–401. [Google Scholar] [CrossRef] [PubMed]
- Canavero, S.; Bonicalzi, V. Extradural cortical stimulation for central pain. Acta neurochirurgica. Supplement 2007, 97, 27–36. [Google Scholar] [PubMed]
- Benabid, A. Deep brain stimulation for Parkinson’s disease. Current opinion in neurobiology 2003, 13, 696–706. [Google Scholar] [CrossRef] [PubMed]
- Lozano, A.M.; Dostrovsky, J.; Chen, R.; Ashby, P. Deep brain stimulation for Parkinson’s disease: disrupting the disruption. The Lancet. Neurology 2002, 1, 225–31. [Google Scholar] [CrossRef]
- Duffau, H.; Capelle, L.; Lopes, M.; Faillot, T.; Sichez, J.P.; Fohanno, D. The insular lobe: physiopathological and surgical considerations. Neurosurgery 2000, 47, 801–10, discussion 810–1. [Google Scholar] [CrossRef]
- Duffau, H.; Bauchet, L.; Lehéricy, S.; Capelle, L. Functional compensation of the left dominant insula for language. Neuroreport 2001, 12, 2159–2163. [Google Scholar] [CrossRef]
- Duffau, H.; Denvil, D.; Capelle, L. Absence of movement disorders after surgical resection of glioma invading the right striatum. Journal of neurosurgery 2002, 97, 363–9. [Google Scholar] [CrossRef] [PubMed]
- Duffau, H.; Capelle, L.; Denvil, D.; Sichez, N.; Gatignol, P.; Lopes, M.; Mitchell, M.C.; Sichez, J.P.; Van Effenterre, R. Functional recovery after surgical resection of low grade gliomas in eloquent brain: hypothesis of brain compensation. Journal of neurology, neurosurgery, and psychiatry 2003, 74, 901–7. [Google Scholar] [CrossRef]
- Duffau, H.; Capelle, L.; Denvil, D.; Gatignol, P.; Sichez, N.; Lopes, M.; Sichez, J.P.; Van Effenterre, R. The role of dominant premotor cortex in language: a study using intraoperative functional mapping in awake patients. NeuroImage 2003, 20, 1903–14. [Google Scholar] [CrossRef]
- Duffau, H.; Khalil, I.; Gatignol, P.; Denvil, D.; Capelle, L. Surgical removal of corpus callosum infiltrated by low-grade glioma: functional outcome and oncological considerations. Journal of neurosurgery 2004, 100, 431–7. [Google Scholar] [CrossRef]
- Duffau, H.; Denvil, D.; Capelle, L. Long term reshaping of language, sensory, and motor maps after glioma resection: a new parameter to integrate in the surgical strategy. Journal of neurology, neurosurgery, and psychiatry 2002, 72, 511–6. [Google Scholar] [PubMed]
- Naeser, M.A.; Palumbo, C.L.; Helm-Estabrooks, N.; Stiassny-Eder, D.; Albert, M.L. Severe nonfluency in aphasia. Role of the medial subcallosal fasciculus and other white matter pathways in recovery of spontaneous speech. Brain : a journal of neurology 1989, 112 Pt 1, 1–38. [Google Scholar]
- Holodny, A.I.; Schwartz, T.H.; Ollenschleger, M.; Liu, W.C.; Schulder, M. Tumor involvement of the corticospinal tract: diffusion magnetic resonance tractography with intraoperative correlation. Journal of neurosurgery 2001, 95, 1082. [Google Scholar] [CrossRef] [PubMed]
- Peraud, A.; Meschede, M.; Eisner, W.; Ilmberger, J.; Reulen, H.J. Surgical resection of grade II astrocytomas in the superior frontal gyrus. Neurosurgery 2002, 50, 966–75, discussion 975–7. [Google Scholar] [PubMed]
- Duffau, H. Intraoperative cortico–subcortical stimulations in surgery of low-grade gliomas. Expert review of neurotherapeutics 2005, 5, 473–485. [Google Scholar] [CrossRef] [PubMed]
- Döbrössy, M.D.; Dunnett, S.B. The influence of environment and experience on neural grafts. Nature reviews. Neuroscience 2001, 2, 871–9. [Google Scholar] [CrossRef]
- Bachoud-Lévi, A.C.; Rémy, P.; Nguyen, J.P.; Brugières, P.; Lefaucheur, J.P.; Bourdet, C.; Baudic, S.; Gaura, V.; Maison, P.; Haddad, B.; Boissé, M.F.; Grandmougin, T.; Jény, R.; Bartolomeo, P.; Dalla Barba, G.; Degos, J.D.; Lisovoski, F.; Ergis, A.M.; Pailhous, E.; Cesaro, P.; Hantraye, P.; Peschanski, M. Motor and cognitive improvements in patients with Huntington’s disease after neural transplantation. Lancet 2000, 356, 1975–9. [Google Scholar] [CrossRef] [PubMed]
- Lindvall, O. Stem cells for cell therapy in Parkinson’s disease. Pharmacological research : the official journal of the Italian Pharmacological Society 2003, 47, 279–287. [Google Scholar] [CrossRef]
- Buchan, A.M.; Warren, D.; Burnstein, R. Transplantation of cultured human neuronal cells for patients with stroke. Neurology 2001, 56, 821–2. [Google Scholar] [CrossRef]
Figure 1.
Hallmarks of brain plasticity: developmental flexibility, complex interactions of genetic and environmental influences, and structure-function changes comprising neurogenesis, axonal sprouting, and synaptic remodeling.
Figure 1.
Hallmarks of brain plasticity: developmental flexibility, complex interactions of genetic and environmental influences, and structure-function changes comprising neurogenesis, axonal sprouting, and synaptic remodeling.
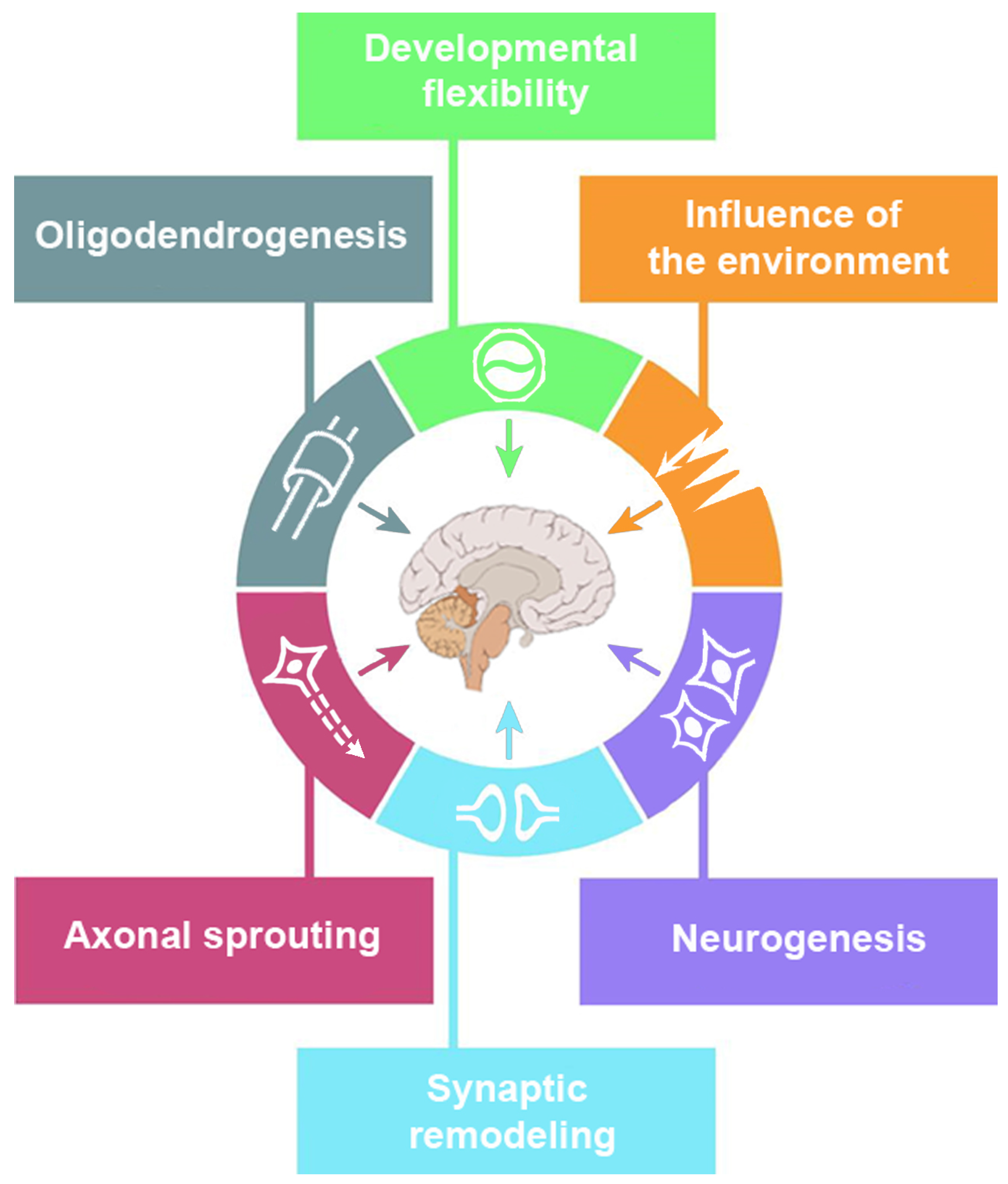

Table 1.
Examples of non-coding RNAs involved in brain plasticity
| No | Name (acronym) | Molecular species | References |
|---|---|---|---|
| 1 | long non-coding RNA (lncRNA) |
Gomafu, GAS5, MALAT1, HOTAIR | [131,132,133,134,135] |
| 2 | microRNA (miRNA) | miR-9, miR-34, miR-132 | [136,137,138] |
| miR-17-92 cluster | [139,140] | ||
| miR-144-5p, miR-145, miR-153 | [141,142,143] | ||
| hsa-miR-1-3p, hsa-miR-335-5p, hsa-miR-34a-5p |
[144] | ||
| 3 | circular RNA (circRNA) | ciRS-7, circRMST, circFAT3 | [145] |
| circIgfbp2 | [146] | ||
| nearly 1167 cerebral circRNAs | [147] | ||
| cirC_0000400, cirC_0000331, cirC_0000406, cirC_0000798 |
[148] | ||
| 4 | enhancer RNA (eRNA) | Bdnf-Enhg1, Bdnf-Enhg2 | [149] |
| Evf2 | [150] | ||
| 5 | long intergenic non-coding RNA (lincRNA) |
linc-Brn1b | [151] |
| Xist | [152] | ||
| 6 | Piwi-interacting RNA (piRNA) |
list of 1251 brain-specific piRNAs piR-hsa-1281, piR-hsa-1280, piR-hsa-1282, piR-hsa-27492 |
[153,154,155] |
| 7 | Y RNA (yRNA) | nELAVL/Y RNA complex hY1, hY4, hY5 |
[156,157,158] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



