
Submitted:
17 September 2024
Posted:
17 September 2024
You are already at the latest version
Abstract
Limoncella of Mattinata, a rare and ancient Mediterranean citrus fruit, was investigated by sequence analysis of the ribosomal internal transcribed spacer regions, which assigns it as a variety of Citrus medica L. Morphological, chemical, and biomolecular approaches were used to characterize flavedo and albedo, parts usually rich of bioactive compounds. The morphological findings showed albedo and flavedo cellular structures as “reservoirs” of nutritional components. Both albedo and flavedo hydroalcoholic extracts were rich in polyphenols, but they were different in compounds and quantity. The flavedo is rich in p-coumaric acid and rutin, whereas the albedo contains high levels of hesperidin and quercitrin. Antioxidant, anti-inflammatory, and genoprotective effects for albedo and flavedo were found. The results confirmed the health properties of flavedo and highlighted that albedo is also a rich source of antioxidants. Moreover, this study valorizes Limoncella of Mattinata's nutritional properties, cueing its crops' repopulation.
Keywords:
albedo
; flavedo
; bioactive compound
; functional food
1. Introduction
Limoncella is the common name for a fruit tree of the Rutaceae family, considered a rare and ancient Mediterranean variety belonging to the Citrus genus. This family comprises 150 genera with approximately 2.000 species, 70 of which are Citrus. The taxonomy of Citrus still stirs up severe doubts in the scientific community because of the intergeneric sexual compatibility, the high frequency of bud mutations, and the long history of cultivation and spread [1]. Despite the difficulties in establishing a consensual classification of edible citrus, most authors now agree on the origin of cultivated forms. The use of molecular markers and genome sequencing have contributed to identifying four primary taxa: C. maxima (pummelos), C. medica (citrons), C. reticulata (mandarins), and C. micrantha (a wild Papeda species) [2].
Citrus species are economically and culturally significant, with their fruits widely consumed fresh, juiced, or used as flavorings in various culinary dishes. Italy is one of the world’s largest producers of citrus fruit and is in second place, after Spain, at the European level [3]. In 2022, Italy produced 3.09 tons of citrus fruits. The Italian citrus distribution is higher in southern regions, including Sicily and Calabria, with a value above 80%, as reported by ISTAT [4].
Citrus fruits also have a long history of use in pharmacology due to their rich content of bioactive compounds, particularly flavonoids, essential oils, and vitamin C. These components offer a range of therapeutic benefits and are utilized in various medicinal and health applications. Some species of Citrus have a broad spectrum of biological activities, including antibacterial, antiviral, antioxidant, antifungal, analgesic, and anti-inflammatory [5,6,7,8]. Citrus fruit peel (epicarp) is composed of flavedo (exocarp), the pigmented more external region, and albedo (mesocarp), the white layer before the pulp.
Many scientific studies refer to the citrus peel as a whole but must distinguish between the flavedo and albedo layers separately. The peel is often treated as a single entity in research, which can sometimes overlook the unique properties and functions of these individual components. Different studies conducted on various Citrus species have shown that peels have antioxidant properties, but less is known about the albedo fraction [9,10,11,12,13,14]. It is known that the albedo is characterized by high nutritional value due to the presence of functional compounds: phenolic acid, flavanones, and flavones [15]. In this study, we thoroughly characterized the flavedo and albedo extracts from sweet citrus popularly named Limoncella of Mattinata, a location on the Southern Italian coast of the Gargano in the Puglia Region. We investigated properties, including antioxidant and genoprotective activities, to valorize its flavedo and albedo as a natural resource and promote the cultivation of this traditional citrus tree on the verge of extinction.
2. Materials and Methods
2.1. Materials
Cyanidin-3-glucoside chloride, delphinidin-3,5-diglucoside chloride, delphinidin-3-galactoside chloride, petunidin-3-glucoside chloride, malvidin-3-galactoside chloride, quercetin-3-glucoside and kaempferol-3-glucoside were purchased from PhytoLab (Vestenbergsgreuth, Germany). The remaining 31 analytical standards of the 38 phenolic compounds were supplied by Sigma-Aldrich (Milan, Italy). Formic acid (99%) was obtained from Merck (Darmstadt, Germany). Analytical-grade hydrochloric acid (37%) was obtained from Carlo Erba Reagents (Milan, Italy). HPLC-grade methanol was supplied by Sigma-Aldrich (Milano, Italy). Deionized water (> 18 MΩ cm resistivity) was further purified using a Milli-Q SP Reagent Water System (Millipore, Bedford, MA, USA). All solvents and solutions were filtered through a 0.2 μm polyamide filter from Sartorius Stedim (Goettingen, Germany). Before HPLC analysis, all samples were dissolved in methanol and filtered with Phenex™ RC 4 mm 0.2 μm syringeless filter, Phenomenex (Castel Maggiore, BO, Italy). Reagents for sample preparation, antioxidant activity experiments, and genoprotective properties were purchased from Sigma Aldrich. Cell culture materials and reagents were from VWR International (Milan, Italy).
2.2. PCR Amplification of ITS Region
The molecular characterization was performed by amplification and sequencing the ITS (Internal Transcribed Spacer) region. According to the manufacturer's recommendation, the genomic DNA was obtained from 100 mg plant leaf using a QIAamp DNA mini kit (Qiagen, Milan, Italy). Universal primers ITS1F-(5'-TCCGTAGGTGAACCTGCGG-3’) and ITS4 R-(5'-TCCTCCGCTTATTGATATGC-3’), located at 3’ and 5’of 18S and 28 S of ribosomal genes, respectively were employed. PCR reactions were performed in 25 μl volume containing 50 ng of the genomic DNA, 0.4 μM of each of the above primers, 200 μM dNTP’s, 2.5 μl of 10x buffer, and 0.74 U of Taq DNA polymerase (Takara). Reaction conditions were: denaturation at 95 °C for 10 min followed by 35 cycles of 95 °C for 30 sec, 55 °C for 30 sec, and 72 °C for 30 sec, with final extension at 72 °C for 10 min. The amplification products were electrophoresed on 0.8 % agarose TBE gel and stained with ethidium bromide (0.3 μg/ml). PCR products were purified using the Gel Extraction Kit (Qiagen, Milano, Italy). Both strands of the products were sequenced using the pair of primers used in the PCR amplification. Sequences were run on an ABI 3700 automated sequencer, and the obtained sequences were aligned using BLAST as described in Supplementary Material.
2.3. Light and Transmission Electron Microscopy
For Light and Transmission Electron Microscopy (TEM), the samples were cut into small pieces, fixed in 2.5% glutaraldehyde, and post-fixed in 1% aqueous osmium tetroxide, both in phosphate buffer for one hour. The samples were rinsed in phosphate buffer, dehydrated in an increasing ethanol series (50, 70, 80, 90, 95, 100%, 15 min each), further dehydrated twice with propylene oxide for 15 min, and then incubated in epoxy resin at 60 °C for three days. Semi-thin sections were cut with an ultra-microtome (Ultratome LKB) into 1-mm semi-thin sections and stained with 1% Toluidine blue in distilled water, 0.2% Basic fuchsin in ethanol, Safranin O in 50% alcohol, 0.13 % toluidine blue + 0.02% Azur II. Basic fuchsin, Safranin O, and Azur II staining were performed on etched resin sections with sodium meta-periodate. Different dyes were used to highlight the various parenchyma morphological components. From the same sample block, sections were cut into 70-80 nm ultra-thin sections and placed on 400 mesh grids. Ultra-thin sections were stained for 45 min each with Uranyless followed by 30 min lead citrate and observed by Philips CM10 at 80 kV.
2.4. Scanning Electron Microscopy (SEM) and Environmental Scanning Electron Microscopy (ESEM)
Sample preparation for conventional SEM analysis involved fixation in 2.5% glutaraldehyde (Sigma-Aldrich) for 1 h at 4 °C and dehydration using 50%, 70%, 80%, 90%, and twice with 100% ethanol (Sigma-Aldrich) for 5 minutes at each concentration. The samples were treated with Hexamethyldisilazane (HMDS), transferred on aluminum stubs, and sputter-coated with an S-thin layer of gold, approximately 10 nm. Images were acquired using a Philips 515. ESEM technology represents an upgrade of the conventional scanning electron microscope (SEM), allowing the observation of samples at different vacuum levels, either prepared according to the conventional SEM method or natural, without dehydration or conductive coating. The latter opportunity is advantageous for biological samples, allowing morphological analysis without pre-treatment before observation. If equipped with a spectrometer for the dispersion of energy (EDS), ESEM allows the semiquantitative detection of chemical elements constituting the ultrastructural components of the sample through point or areal analysis. An FEI Quanta 200 FEG Environmental Scanning Electron Microscope (FEI, Hillsboro, OR, USA) was used with an energy-dispersive X-ray spectrometer (EDAX Inc., Mahwah, NJ, USA). The analyses used a focalized electron beam in a vacuum electron gun pressure of 5.0 e-6 bar. The ESEM was utilized in low vacuum mode, with a specimen chamber pressure set at about 0.80 mbar, an accelerating voltage of 25 kV, a working distance of 10 mm, a tilt angle of 0, a variable beam diameter, and a magnification between 100 and 40,000 x. The images were obtained utilizing the back-scattered electron detector to highlight the presence of particles or aggregates or the secondary electron detector to highlight the morphological features. The spectrometric unit has an ECON (Edax Carbon Oxygen Nitrogen) 6 UTW X-ray detector and Genesis Analysis software. Each sample was analyzed with a time count of 100 sec and an Amp Time of 51, while the probe current was 290 μA.
2.5. Preparation of Albedo and Flavedo Extracts
Albedo and flavedo of Limoncella lemons from Mattinata were separated and subject to drying at 60 °C overnight. After drying, albedo and peel were triturated in a mortar with liquid nitrogen. This procedure was repeated three times to reduce samples in a fine powder. Subsequently, 3 g of ground flavedo and albedo samples were extracted with 30 ml of 80% ethanol/water (80:20 v/v) at 4 °C under stirring. The albedo and flavedo extracts were then centrifuged at 400 g for 15 min, and the residues were extracted using two additional portions of the ethanol/water mixture. All supernatants were pooled, concentrated, and kept at -20 °C until use.
2.6. Quantification of Total Phenolics by Folin-Ciocalteu Method
Total polyphenol content was determined using the Folin-Ciocalteu method described by Singleton et al. [16]. The total volume of the reaction mixture was minimized to 1 ml. The extract solutions (10 μl) were mixed with Folin-Ciocalteu reagent (50 μl) and deionized water (90 μl). Three minutes later, 300 μl of 20% (w/v) sodium carbonate was added, and the mixture was brought up to 1 ml with distilled water. The tubes were vortex-mixed for 15 sec and allowed to stand for 30 min at room temperature for color development. Absorbance was then measured at 725 nm using a UV Beckman spectrophotometer. The amount of total phenolics was expressed as caffeic acid equivalents through the calibration curve of caffeic acid. The calibration curve ranged from 1 to 15 μg/ml (R2 = 0.9973).
2.7. HPLC-ESI-MS/MS Analysis
HPLC-MS/MS studies were performed using an Agilent 1290 Infinity series and a Triple Quadrupole 6420 from Agilent Technology (Santa Clara, CA) equipped with an electrospray ionization (ESI) source operating in negative and positive ionization modes by following a previously published method [17,18,19]. The separation of target compounds was achieved on a Synergy Polar–RP C18 analytical column (250 mm x 4.6 mm, 4 µm) from Phenomenex (Chesire, UK). The column was preceded by a Polar RP security guard cartridge (4 mm x 3 mm ID). The mobile phase was a mixture of water (A) and methanol (B), with formic acid 0.1% at a flow rate of 0.8 ml min−1 in gradient elution mode. The composition of the mobile phase varied as follows: 0–1 min, isocratic condition, 20% B; 1–25 min, 85% B; 25–26 min, isocratic condition, 85% B; 26–32 min, 20% B. The injection volume was 2 μl. The column temperature was 30 °C, and the drying gas temperature in the ionization source was 350 °C. The gas flow was 12 l/min, the nebulizer pressure was 55 psi, the capillary voltage was 4000 V, and a gas flow rate was maintained at 1,2000 ml/min. Detection was performed in the “dynamic-multiple reaction monitoring” (dynamic-MRM) mode and the dynamic-MRM peak areas were integrated for quantification. The most abundant product ion was used for quantitation, and the other for quantification. The selected ion transitions and the mass spectrometer parameters, including the specific time window for each compound (Δ retention time), are reported in Table S2 (Supplementary Materials).
2.8. Antioxidant Activity
2.8.1. DPPH (2,2-Diphenyl-1-picrylhydrazyl) Assay
The DPPH radical scavenging assay was conducted using the procedure previously described by Saltarelli et al. [20]. Fresh DPPH ethanol solution (850 µl, 100 μM) was mixed with the sample at different concentrations (150 µl, 0.2-3.5 mg/ml). The decreased absorbance at 517 nm was recorded after 10 min at room temperature. The scavenging activity was calculated as % = [(A0 – A)/A0] × 100, where A0 is the absorbance of the control reaction, and A is the absorbance in the presence of samples. The EC50 value was calculated from the plots as concentration extracts required to provide 50% free radical scavenging activity.
2.8.2. ABTS, 2,2′-Azino-bis(3-ethylbenzothiazoline-6-sulphonic acid) Assay
Antioxidant activity against ABTS radical was performed as described by Loizzo et al. with some modifications [21]. Briefly, the reaction mixture was prepared by mixing 7 mM ABTS solution and 2.45 mM potassium persulphate followed by 12–16 h incubation in the dark at room temperature to produce ABTS radical. Before use, the solution was diluted with ethanol to absorb 0.80 ± 0.05 at 734 nm. Aliquots of the sample at concentrations ranging from 0.01 to 0.4 mg/ml were added to 1 ml of ABTS ethanolic solution and incubated in the dark at room temperature for 6 min. The absorbance was then recorded at 734 nm using a UV Beckman spectrophotometer. The ABTS radical scavenging activity was calculated following the equation: ABTS scavenging activity (%) = [(A734 nm of blank – A734 nm of the sample)/A734 nm of blank] × 100. Results are reported as EC50 values (µg/mL). Trolox was a reference compound (0.5–5 μg/ml).
2.8.3. Lipoxygenase Inhibition Assay
Soybean 5-lipoxygenase was used for the assay, according to Saltarelli et al. [20]. Inhibition experiments were performed by measuring the loss of 5-lipoxygenase activity (0.18 µg/ml) with 100 µM linoleic acid as the substrate in 50 mM sodium phosphate, pH = 6.8. The reaction mixture was pre-equilibrated at 20 °C for 20 min without the enzyme. Inhibition studies in the presence of various extract concentrations (0.2-0.7 mg/ml) were recorded at 235 nm at 20 °C using a UV Beckman spectrophotometer. The lipoxygenase activity was calculated as % = 100-{[(Δ235 nm of blank - Δ235 nm of sample)/Δ235 nm of blank] x 100}. EC50 was determined by plotting the graph with the concentration of extracts versus the percentage of inhibition of linoleic acid peroxidation.
2.8.4. Chelating Capacity on Fe2+
Fe2+ chelating capacity was evaluated as described by Saltarelli et al. [20] with some modifications. Briefly, an aliquot of 20 μl of FeSO4 solution (2 mM) was added to 200 μl of sample (range 20-125 μg/ml) and incubated at room temperature for 5 min. After that, an aliquot of 40 μl of ferrozine solution (4 mM) was added to the reaction mixture, and the sample volume was diluted to 1 ml with deionized water, mixed, and incubated for 10 min in the dark at room temperature. The absorbance at 562 nm was then spectrophotometrically determined. The chelating activity was calculated as % = [(A562 nm of blank - A562 nm of the sample)/A562 nm of blank] × 100. EC50 is the concentration at which ferrous ions are chelated by 50% and was evaluated by plotting the sample concentration versus the chelating activity.
2.9. DNA Nicking Assay
The DNA nicking assay evaluated the protective effect of hydroalcoholic albedo and Limoncella flavedo extracts against oxidative DNA damage, which employs ferrous ions and dioxygen (Fe2+ + O2) to generate DNA strand breaks induced from free radicals [22]. The assay consists of a cell-free system composed of plasmid DNA (pEMBL8), which resembles the structure of mtDNA [23]. The hydroalcoholic solutions were sequentially diluted with water to obtain the final concentrations of 250 µg/ml, 125 µg/ml, 63 µg/ml and 31 µg/ml. Each extract was assayed in a final volume of 72 µl, consisting of PBS (Phosphate Buffer Solution), 7 µg/mL of pEMBL8, and 20 µl of the related dilution. The mixtures were added 8 μl of 3 mM FeSO4 freshly prepared and kept on ice. The tubes were incubated for 10 minutes at 37 °C, and the reaction stopped with 20 µl of loading buffer. The disappearance of the supercoiled form of the plasmid (CCC) was assessed on an ethidium bromide-stained agarose gel electrophoresis followed by quantification using Gel Doc 2000 and Quantity One software (Bio-Rad). In detail, 20 µl of each sample were loaded onto a 1.2% w/v agarose gel in TAE buffer (40 mM Tris-acetate and 1 mM EDTA, pH 8.0), run at 80 V for 30 min in a small electrophoresis chamber (Mini-Sub Cell GT Systems, Bio-Rad). The gel was stained with 0.3 μg/ml ethidium bromide (EtBr). The EC50 value was calculated by determining the concentration of the compound protecting half of the supercoiled plasmid.
2.10. Cell Cultures
The human keratinocyte cell line HaCaT was obtained from the Interlab Cell Line Collection (ICLC, Genoa, Italy). Cells were grown in DMEM medium supplemented with 10% fetal bovine serum, 2% glutamine, 1% sodium pyruvate, and 100 U/ml penicillin/streptomycin. Cells were maintained in an incubator at 37 °C and 5% CO2. RAW 264.7 murine macrophages were cultured in RPMI 1640 medium with 10%, 2 mM glutamine, and 1% 100 U/ml penicillin/streptomycin and maintained at 37 °C in a 5% CO2 atmosphere.
2.11. Cytotoxicity Assays
The cytotoxic effects of the Limoncella hydroalcoholic albedo and flavedo extract against HaCaT cells were analyzed by WST-8 and sulforhodamine B (SRB) assays, which evaluate cellular metabolic activity and cellular protein content, respectively [24]. In detail, cells (5 x 103/well) were seeded in 96-well plates and treated with water-diluted extracts from 110 to 7 µg/ml. After 24 hours of incubation, the test compounds were removed, and a fresh medium containing WST-8 (Sigma-Aldrich, Milan, Italy) was added to each well. Cells were further incubated at 37 °C for up to 4 hours, and color development was monitored at 450 nm in a microplate reader (Multiskan FC, Thermo Scientific) [25]. As previously published, the SRB assay was also performed in the same 96-well plate [26]. Briefly, cells were fixed with cold 50% trichloroacetic acid and stained with 0.4% SRB (Sigma-Aldrich, Milan, Italy) dissolved in 1% acetic acid. The protein-bound dye was subsequently solubilized with 10 mM Tris, and the absorbance was read at 570 nm in a microplate reader (Multiskan FC, Thermo Scientific). The concentration that caused 50% growth inhibition (IC50) was calculated and the data was expressed as a percentage (%) compared to that of untreated cells (controls).
2.12. Evaluation of Antioxidant Properties by DCFH-DA Assay
The antioxidant properties of albedo and flavedo extracts were analyzed in HaCaT cells using 2′,7′-dichlorofluorescein diacetate (DCFH-DA, Sigma-Aldrich, Milan, Italy), which transforms into 2′,7′-dichlorofluorescein (DCF) highly fluorescent after oxidation [25]. In detail, cells (1 x 104/well) were seeded into black 96-well plates and incubated for 2 h with a concentration of 25 µg/ml extracts in 100 µl DMEM. The medium was then removed and replaced with 50 µl DCFH-DA (5 µM in PBS), incubated for 30 minutes at 37 °C. After the removal of the excess probe, cells were treated with 100 µl of hydrogen peroxide (H2O2, 100 μM in PBS) for 30 min, and DCF fluorescence emission was measured at ex/em 485/520 nm in the FluoStar multiwell plate reader Optima (BMG Labtech, Germany). Data were expressed as relative oxidation compared to non-oxidized cells.
2.13. Determination of Nitric Oxide Production
The anti-inflammatory properties of both albedo and flavedo hydroalcoholic extracts were evaluated in RAW 264.7 cells (murine macrophages) stimulated by Lipopolysaccharide (LPS) (Sigma-Aldrich, Milan, Italy). Cells (3 x 104/well) were seeded into 96-well plates and treated with both Limoncella extracts (50 µg/mL) in the presence and absence of 1 µg/ml LPS for 24 h. The cells were also incubated alone in the presence and absence of LPS as a control. The drug dexamethasone at a 5 µg/ml concentration was used to validate the test. Subsequently, Nitric Oxide (NO) levels were determined in the supernatant medium using Griess reagent (Sigma-Aldrich, Milan, Italy) [27]. Absorbance was measured at 570 nm using a plate reader (BioRad Laboratories, Hercules, USA).
3. Results
3.1. PCR Amplification, DNA Sequencing, and Sequence Analysis of ITS Region
The ribosomal DNA region, including the ITS1, 5.8S, and ITS2, was amplified by polymerase chain reaction using universal primers located on the 3’ end of 18S and the 5’start of 28S. The electrophoretic patterns showed a ~700 bp amplicon PCR product, purified and sequenced. Sequence alignment analyses assign this fruit to Citrus medica species (Table S1, Supplementary Material).
3.2. Morphological and Semi-Quantitative Data
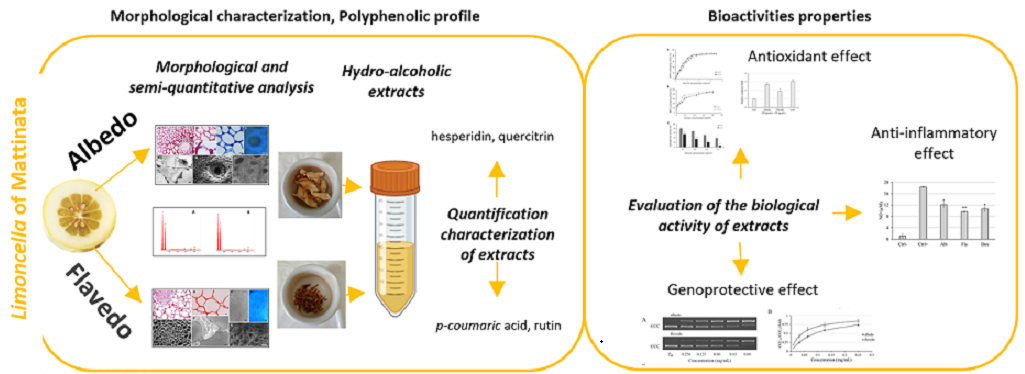
All morphological approaches that allowed to observe flavedo (Figure 1 A) consist of undifferentiated tightly packed parenchyma tissue with slightly turgid cells, which present numerous large oil glands (A, E, H, I), formed inside by cells with very thin walls, as demonstrated in A. Some plastids (F, G) and plasmodesmata structures (B, C) can be observed. The albedo parenchymal cells (Figure 1 B) show a thin wall and a considerable central vacuole. Plasmodesmata or intercellular bridges that connect plant cells are cylindrical channels approximately 40 nm in diameter, which are characteristic structures that enhance cell communication through the secondary wall (B, F). Semi-quantitative chemical analyses showed that both albedo (A) and flavedo (B) contained carbon (A 73.79%; B 63.75%), oxygen (A 25.47%; B 35.96%), chloride (A 0.16%), potassium (A 0.27%; B 0.34%) and calcium (A 0.31%; B 0.50%) (Figure 1 C).
3.3. Characterization and Quantification of Phenolic Compounds
The folin-Ciocalteu method (FC) and HPLC-ESI-MS/MS were used to evaluate the polyphenol content. Both methods showed that the total phenol content was comparable in both extracts (Table 1). In particular, total phenol content measured by FC was 18700 ± 323 and 22040 ± 853 mg/kg dw in albedo and flavedo extracts, respectively.
In contrast, the values obtained by HPLC were slightly lower than those observed by FC (14863.69 and 11392.24 mg/kg dw for albedo and flavedo extracts, respectively). The phenolic compounds were characterized by HPLC-ESI-MS/MS testing the presence of 38 compounds, as reported in Table 1. It is worth underlining the higher concentration of hesperidin (15-fold) and quercitrin (7-fold) in the albedo compared to flavedo.
3.4. Antioxidant Activities
The antioxidant capacity of albedo and flavedo extracts was evaluated using different chemical tests: the DPPH assay, the ABTS assay, the chelating ability of Fe2+, and the lipoxygenase inhibition assay. Figure 2 shows the antioxidant activity of the albedo and flavedo extracts from Limoncella. The free-radical scavenging effect of Limoncella extracts was evaluated using DPPH and ABTS assays. As reported in Figure 2 A and B, albedo and flavedo extracts showed scavenging activity in a dose-dependent manner with an inhibition range of 80-85% at a concentration > 0.2 and 2.0 mg/ml in ABTS and DPPH tests, respectively. Both parts of the fruit prevented oxidative damage mediated by free radicals since there was no statistically significant difference between albedo and flavedo extracts. The albedo extract did not show the chelating effect on Fe2+, and flavedo displayed only a small chelating activity: about 20% at 15 mg/ml (data not shown). Furthermore, the Limoncella extracts inhibited the oxidation of linoleic acid catalyzed by lipoxygenase in a dose-dependent manner (Figure 2 C). However, the flavedo extract was significantly more effective than albedo since at 0.7 mg/ml of extract concentration, lipoxygenase activity was 5.5 ± 2.5 and 37.7 ± 6.2, respectively.
EC50 values (Table 2) for ABTS and DPPH scavenging activity were comparable in albedo and flavedo extracts. However, ABTS EC50 measurements were lower than the ones obtained for DPPH EC50 (about 0.063 mg/ml and 0.971-0.996 mg/ml, respectively). The antioxidant capacities of albedo and flavedo extracts were 35.42 ± 3.31 and 40.41 ± 10.15 µg Trolox Equivalent/mg, respectively. A different EC50 level between albedo and flavedo extracts was detected for lipoxygenase inhibition; in particular, the EC50 of flavedo was significantly lower than that reported for albedo (Table 2).
3.5. Genoprotective Activity
DNA nicking assay is employed to check the ability of albedo and flavedo extracts against the plasmid DNA damage caused by free radicals generated in vitro by Fe2+ + O2. Figure 3(A) depicts the ability of the extracts to protect supercoiled DNA form (CCC) during the incubation with 3 mM Fe2+, and Figure 3(B) reports the quantification analyses expressed as the ratio between the CCC after treatment and CCC at time 0. From Figure 3(A), the protection of the supercoiled form of DNA is observed as the concentrations of the two extracts increase in the presence of the oxidant. This protection is higher with flavedo extracts compared to albedo. The bands of the open circular form (OC), formed in damaged DNA, are more evident with the albedo extract. This indicates that flavedo offers more excellent protection against DNA oxidation. The bands were quantified, and the densitometric analysis showed a dose-dependent response with a higher percentage of protection in flavedo (Figure 3, B). The EC50 values obtained in the DNA nicking assays were 0.090 mg/ml and 0.042 mg/ml for albedo and flavedo, respectively.
3.6. Evaluation of the Cytotoxic Effects
The evaluation of the cytotoxicity of the hydroalcoholic extracts of Limoncella using the sulforhodamine B (SRB) test revealed a significant reduction in the viability of cultured human keratinocytes cells at concentrations higher than 200 µg/ml (Figure 4). The calculated IC50 values were 250 µg/mL and 161 µg/mL for albedo and flavedo, respectively.
3.7. Radical Scavenging Properties of Limoncella Extracts by Cell-Based Assay
Radical scavenging properties of Limoncella extracts were also tested in a cell system. As reported in Figure 5, the administration of H2O2 to HaCaT cells (Ctrl+) led to a significant increase in DCF fluorescence emission compared to untreated cells (Ctrl-). When cells were pretreated for 2 hours with 50 µg/ml of both Limoncella preparations, a highly significant reduction in H2O2-induced free radicals in flavedo extract was observed (-37-fold vs. Ctrl+), while a reduction less significant was observed in albedo extract (-12.5-fold vs Ctrl+), further supporting their action as a protective antioxidant against oxidative stress.
3.8. Anti-Inflammatory Activity
As reported in Figure 6, the stimulation of RAW 264.7 cells (murine macrophages) by one µg/ml lipopolysaccharide (Ctrl+) led to a robust extracellular release of the inflammatory response mediator NO as compared to unstimulated control cells (Ctrl-). When LPS-exposed cells were co-incubated with 50 µg/ml of both extracts, a decrement of NO production was observed; albedo extract reduced the production of NO at 12.1 µM and flavedo at 9.6 µM respect to positive control at 18.3 µM. The anti-inflammatory activity of both flavedo and albedo extract was comparable to that of the reference drug dexamethasone 5 µg/mL (10.6 µM vs Ctrl+).
4. Discussion
The various species of Citrus are mainly used in the food and cosmetic fields. However, due to their antioxidant, antimicrobial, and anticancer properties, they have also found applications in the therapeutic field [28,29,30,31]. Most studies have been performed on the flavedo or the entire peel, while little is known about the beneficial properties of the albedo [14,32,33,34]. The present work represents an attempt to appreciate this part of the fruit. For the present research, we selected a rare, ancient Mediterranean Citrus variety from Southern Italy to establish its phylogenetic position and appreciate its health properties, thus promoting its cultivation and avoiding extinction. The sequence analysis of the ribosomal internal transcribed spacer regions assigns Limoncella as a variety of Citrus medica L. The different cultivars of C. medica L. are generally divided into “acidic” and “sweet” cedars, and Limoncella fruits have a sweeter pulp. Morphological approaches allowed the structure and ultrastructure of Limoncella to be studied by analyzing different cellular components, supporting the correlation between the nutritional characteristics and the structural organization of the Limoncella [35] In particular, the morphological evidence obtained utilizing different and complementary approaches allowed us to understand better the albedo’s and flavedo’s components that constitute the “reservoir” of nutritional substances and the parts of them that do not interfere with the extractive procedure. Therefore, all the cellular structures and remnants after the extraction can be considered a resource (in terms of wide-scale industrial extraction) as fertilizer in a fully circular economy of fruit treatment. Moreover, the well-defined morphological Limoncella characterization represents a comparable model for studying properties similar to those of other citrus fruit types. Concerning the chemical characterization of flavedo and albedo alcoholic extracts, herein, we report that they are rich in polyphenols, quite different in compounds and quantity (Table 1), although their total polyphenol content fall within the range of those previously reported for 35 cultivars of Citrus reticulata Blanco [36] and Citrus medica L. [37]. In particular, the Folin-Ciocalteu method (FC) and HPLC-ESI-MS/MS were used to evaluate the polyphenol content. Both methods showed that the total phenol content was comparable in both extracts, indicating that albedo is also a good source of these compounds. However, higher concentrations were systematically found with FC compared to those obtained by HPLC analysis [38]. The values obtained by HPLC were 14863.69 and 11392.24 mg/kg dw for albedo and flavedo extracts, respectively, and they were slightly lower than those observed by FC (18700 ± 323 and 22040 ± 853 mg/kg dw in albedo and flavedo extracts, respectively). This outcome could be explained considering that the FC method estimates the content of reducing compounds, phenolic and non-phenolic. In contrast, the HPLC analysis only measures the content of the major phenolics in both extracts. Furthermore, in flavedo, p-coumaric acid and rutin prevail, while in albedo, hesperidin and quercitrin. Hesperidin and quercitrin exceed the corresponding content of flavedo by 15 and 7 times, respectively. The qualitative and quantitative differences of the polyphenolic compounds also explain the different activities or functions of the phytocomplexes of these two parts of the fruit [39]. Hesperidin is a flavanone glycoside with various biological effects. This compound has strong antioxidant and anti-inflammatory potential against many lifestyle-linked metabolic syndromes [40]. It has been found to benefit cardiovascular and cutaneous functions, prevent bone resorption and type II diabetes, as well as anticancer and anti-inflammatory effects. The antioxidant activity of hesperidin was limited to its radical scavenging activity, and the antioxidant cellular defenses were augmented via the ERK/Nrf2 signaling pathway [41]. Increasing evidence shows the benefits of hesperidin in central nervous system disorders based on its neuroprotective effect [42]. Several cellular and animal models have been developed to evaluate the underlying neuropharmacological mechanisms of hesperidin. Additionally, clinical evidence has confirmed its neuroprotective function. Hesperidin exerts its neuroprotective properties by decreasing neuro-inflammatory and apoptotic pathways. Hesperidin can effectively alleviate depression and improve cognition and memory. Quercitrin is a glycoside derived from the flavonoid quercetin and the deoxy sugar rhamnose, commonly used as a dietary ingredient and supplement [43] compound, compared with quercetin, possesses different physical and chemical properties. Indeed, quercetin glycoside form, being more soluble in polar solvents, is better absorbed than quercetin and another form of quercetin at the intestinal mucosa level through glucose transporters [44]. Quercitrin has pharmacological properties, including antioxidants, antiinflammation, anti-microorganisms, immunomodulation, analgesia, wound healing, and vasodilation. These functions identify quercitrin as a potential therapeutic agent for several diseases, such as bone metabolic, gastrointestinal, cardiovascular, and cerebrovascular [45].
Clinical studies of quercitrin are few, although molecular mechanisms in treating diseases and the dose-effect relationships are partially known [45]. Also, novel preparations that are useful for clinical research and as functional food are available [46] The qualitative/quantitative characterization of the two ethanolic extracts led us to test their antioxidant activity, which cell-free and cell-based models performed. DPPH and ABTS assay are cell-free methods showing that either flavedo or albedo exhibit similar radical scavenging properties. Furthermore, these values are comparable to those previously reported for Citrus lumia Risso [14]. The flavedo extract is more performant from the assays investigating the effect of Fe2+ chelating, lipoxygenase activity, and the level of ROS.
We evaluated the antioxidant properties of two extracts using a cell-based method with the DCFA-DA probe in H2O2-treated HACAT cells. This test requires previous viability assays to select the range of possible non-cytotoxic concentrations and the toxicity threshold. Cell viability was assessed using the WST-8 and SRB assays. While the SRB assay gave significant results for both samples, the WST-8 assay gave nonrealistic results; in fact, plant extract contains compounds that may interfere with reducing WST-8 or react with the formazan dye, leading to false results [47]. The evaluation of the calculated IC50 of the hydroalcoholic extracts of Limoncella using the sulforhodamine B (SRB) test revealed cell viability and classified both extracts as weakly cytotoxic [48]. Both Limoncella preparations gave a highly significant reduction in H2O2-induced free radicals in flavedo extract and a less significant reduction in albedo extract, further supporting their action as a protective antioxidant against oxidative stress. The lipoxygenase enzymes contribute to the emergence of inflammation and allergic reactions by producing leukotrienes. Soybean lipoxygenase has substrate selectivity and inhibitory properties similar to humans, exhibits good stability, and was used in inhibition assay [49]. Albedo and flavedo extracts significantly inhibited lipoxygenase action with an EC50 of 0.54 ± 0.10 0.14 ± 0.06 for albedo and flavedo, respectively. In literature, it was reported that coumarin and hesperidin show considerable lipoxygenase inhibitory activity [49,50], and as reported above, these phenolic compounds are present in large amounts in Limoncella extracts. The results regarding the oxidation of linoleic acid catalyzed by lipoxygenase suggested a possible anti-inflammatory activity by these extracts, which was investigated by choosing the Griess test, a cell-based model that uses the RAW264.7 cells, an appropriate macrophage model to study the inflammatory cell responses rapidly [51]. Also, this assay showed that the flavedo exhibits more anti-inflammatory activity than the albedo, -0.54-fold and -0.67-fold versus Ctrl+, respectively. Both extracts show properties similar to the drug dexamethasone (-0.60-fold). On the other hand, anti-inflammatory activity is not surprising, as polyphenols can also have anti-inflammatory activity. The anti-inflammatory properties of lemon peel extract could be explained by the presence of p-coumaric acid, a phenolic acid, ten times more represented in flavedo vs albedo extract, showing immunosuppressive effects by reducing cell-mediated immune responses and macrophage phagocytic activity in rats [52]. It also shows that anti-inflammatory action decreased the expression of the inflammatory mediator tumor necrosis factor-alpha (TNF-α) and circulating immune complexes in adjuvant-induced arthritic rats. The rutin, a bioflavonoid, 40 times more present in flavedo, shows anti-inflammatory properties by inhibiting inflammatory mediators and enzymes, which help reduce inflammation in the body and can alleviate symptoms of inflammatory conditions like arthritis and may contribute to preventing chronic diseases [53]. In addition, other compounds with anti-inflammatory properties that still need to be identified may be present in the two extracts. Fruits and vegetables are the main anticancer foods, rich in antioxidants, Vitamins C and E, beta-carrots, and lipothin. Citrus fruits, in particular, play an essential role as genoprotective agents. We evaluate the genoprotective effects against oxidative damage by DNA nicking assay. The results proved that albedo and flavedo provided significant antioxidant protection against oxidative damage to biomolecules, acting on pathogenic hydroxyl radicals generated in vitro by Fe2+ + O2 [54,55,56]. However, their cytotoxicity in HaCaT (IC50 albedo, 250 g/ml; IC50 flavedo, 161 µg/ml) is much higher than the EC50 obtained in the nicking assay (albedo, 90 µg/mL; flavedo, 42 µg/mL), and both extracts are indeed safe when expressing protective capacity against hydroxyl radicals. Hydroxyl radicals are mainly produced in mitochondria during oxidative phosphorylation, causing oxidative damage to mtDNA, which is implicated in physiological senescence and age-related disorders [57]. Our experimental model reveals that both phytoextracts protect plasmid DNA (pEMBL8), which resembles the structure of mtDNA, from oxidation by hydroxyl radicals. Thus, although flavedo was found superior, albedo protects against oxidative DNA damage and can counteract mutagenic and carcinogenic agents. Many studies have been performed on the genoprotective properties of lemon juice and peel. A study conducted on Citrus medica L. through the Ames test demonstrated the antimutagenic and anticarcinogenic effects of the juice [58]. Koolaji et al. found that citrus peel contains numerous bioactive compounds, including carotenes, essential oils, pectins, and phenolic compounds, which may reduce cancer risk. The peel extract of the genus Citrus has a higher antitumor activity than the single isolated compound.
In particular, it has been shown that the compounds contained in the peel extract of these citrus fruits have an antitumor activity, which is manifested through suppression of cell proliferation, the inhibition of the cell cycle, the induction of apoptosis, the reduction of the formation of metastases and the inhibition of angiogenesis. The polymethoxyflavonones (PMFs) and other compounds found in the citrus peel can increase apoptotic activity through the intrinsic pathway, eliminating cancer cells [59].
Furthermore, the flavonoids are also antiangiogenic, inhibiting the expression of vascular endothelial growth factor (VEGF) and suppressing endothelial cell migration. Koolaji et al. also showed that the peel bioactive compounds of these citrus fruits reduce the formation of metastases, decreasing the expression and activity of MMP-2 and MMP-9, the main proteins involved in forming metastases [60]. Therefore, the flavonoids present in citrus peels act as free radical scavengers and modulators of several fundamental molecular events implicated in cancer development [36,59,60,61]. The present study, for the first time, highlights the genoprotective property of albedo and suggests that every part of the fruit can be used as a source of compounds beneficial to human health.
5. Conclusions
Limoncella is particularly interesting since it is an ancient variety of Citrus medica L. that has yet to be investigated, and this study aims to valorize this variety and its bioactive compounds.
The present study demonstrated that the two tissues constituting the citrus peel, flavedo, and albedo, possess advantageous functional properties. In particular, we found that the extract derived from albedo is a rich source of antioxidants, which could be utilized in developing various functional food products with significant health benefits, including antioxidative, anti-inflammatory, and anti-mutagenic properties. These results reinforce the importance of further research on Limoncella from Mattinata and its integration into personalized nutrition and disease prevention strategies. Moreover, such research could promote the repopulation of Limoncella crops and consumption of the fruit, including albedo, and could maintain the biodiversity and the properties of this ancient fruit.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: Scores and information from a BLAST search using as query the ITS sequence of Citrus medica var Limoncella; Table S2: HPLC–MS/MS acquisition parameters (dynamic-MRM mode) used for the analysis of the 38 marker compounds.
Author Contributions
Conceptualization, L.P., L.D.P. and E.B.; methodology, R.S., F.P., L.D.P., S.B., and L.V.; validation, R.S., L.P., F.P., L.D.P., P.G., G.C., and A.S.; formal analysis, R.S., F.P., S.B., L.V.; investigation, R.S., L.P., F.P., L.D.P., G.A., S.B., L.V., G.L., and A.S.; resources, R.S., L.P., and E.B.; data curation, L.P., F.P., and P.G.; writing—original draft preparation, R.S., L.P., F.P., S.P., L.V., G.C., and A.S.; writing—review and editing, L.P., F.P., S.V., and E.B.; visualization, R.S., L.P., S.B., and G.C.; supervision, L.P., G.A., and S.V.; project administration, E.B.; funding acquisition, E.B. All authors have read and agreed to the published version of the manuscript
Funding
This research received no specific grant from public, commercial, or not-for-profit funding agencies. Funds from the University of Urbino Carlo Bo funded it. The European Union partially supports it – NextGenerationEU - National Recovery and Resilience Plan, Mission 4 Education and Research - Component 2 From research to business - Investment 1.5, ECS\_00000041-VITALITY - Innovation, digitalization, and sustainability for the diffused economy in Central Italy, CUP J13C22000430001.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable
Data Availability Statement
The original publication, which contains comprehensive research findings and detailed analyses, will be made available on the University of Urbino’s official website. Those who are keen on this research can access it at the following address: https://ora.uniurb.it/. This platform ensures that the publication is easily accessible to students, researchers, and any other reader, fostering a broader dissemination of knowledge and facilitating academic engagement.
Acknowledgments
The authors thank Dr. Serena Benedetti for her assistance with the cell models; her expertise supported our research. They also acknowledge Dr. Roberto Politi for aid in the use of instrumentation, which is crucial for the experiments reported. Special thanks to the Biochemical and Biotechnology Laboratory for providing the necessary resources and facilities. Finally, the authors appreciate the support from the Department of Biomolecular Science for their administrative support.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Nicolosi, E.; Deng, Z.N.; Gentile, A.; La Malfa, S.; Continella, G.; Tribulato, E. Citrus Phylogeny and Genetic Origin of Important Species as Investigated by Molecular Markers. Theoretical and Applied Genetics 2000, 100, 1155–1166. [Google Scholar] [CrossRef]
- Zech-Matterne, V.; Fiorentino, G. AGRUMED: Archaeology and History of Citrus Fruit in the Mediterranean. AGRUMED: Archaeology and history of citrus fruit in the Mediterranean 2017. [Google Scholar] [CrossRef]
- Falcone, G.; Stillitano, T.; De Luca, A.I.; Di Vita, G.; Iofrida, N.; Strano, A.; Gulisano, G.; Pecorino, B.; D’Amico, M. Energetic and Economic Analyses for Agricultural Management Models: The Calabria PGI Clementine Case Study. Energies 2020, 13, 1289. [Google Scholar] [CrossRef]
- Coltivazioni: Coltivazioni Legnose Fruttifere. Available online: http://dati.istat.it/Index.aspx?QueryId=33705 (accessed on 12 August 2024).
- De Freitas, R.M.; Campêlo, L.M.L.; De Almeida, A.A.C.; De Freitas, R.L.M.; Cerqueira, G.S.; De Sousa, G.F.; Saldanha, G.B.; Feitosa, C.M. Antioxidant and Antinociceptive Effects of Citrus Limon Essential Oil in Mice. J Biomed Biotechnol 2011, 2011. [Google Scholar] [CrossRef]
- Ladaniya, M.S. NUTRITIVE AND MEDICINAL VALUE OF CITRUS FRUITS. Citrus Fruit 2008, 501–514. [Google Scholar] [CrossRef]
- Ballistreri, G.; Fabroni, S.; Romeo, F.V.; Timpanaro, N.; Amenta, M.; Rapisarda, P. Anthocyanins and Other Polyphenols in Citrus Genus: Biosynthesis, Chemical Profile, and Biological Activity. Polyphenols in Plants 2019, 191–215. [Google Scholar] [CrossRef]
- Zou, Z.; Xi, W.; Hu, Y.; Nie, C.; Zhou, Z. Antioxidant Activity of Citrus Fruits. Food Chem 2016, 196, 885–896. [Google Scholar] [CrossRef]
- Castro-Vazquez, L.; Alañón, M.E.; Rodríguez-Robledo, V.; Pérez-Coello, M.S.; Hermosín-Gutierrez, I.; Díaz-Maroto, M.C.; Jordán, J.; Galindo, M.F.; Arroyo-Jiménez, M.D.M. Bioactive Flavonoids, Antioxidant Behaviour, and Cytoprotective Effects of Dried Grapefruit Peels (Citrus Paradisi Macf.). Oxid Med Cell Longev 2016, 2016. [Google Scholar] [CrossRef]
- Esparza-Martínez, F.J.; Miranda-López, R.; Mata-Sánchez, S.M.; Guzmán-Maldonado, S.H. Extractable and Non-Extractable Phenolics and Antioxidant Capacity of Mandarin Waste Dried at Different Temperatures. Plant Foods Hum Nutr 2016, 71, 294–300. [Google Scholar] [CrossRef]
- Pepe, G.; Pagano, F.; Adesso, S.; Sommella, E.; Ostacolo, C.; Manfra, M.; Chieppa, M.; Sala, M.; Russo, M.; Marzocco, S.; et al. Bioavailable Citrus Sinensis Extract: Polyphenolic Composition and Biological Activity. Molecules 2017, 22, 623. [Google Scholar] [CrossRef]
- Sharma, K.; Mahato, N.; Cho, M.H.; Lee, Y.R. Converting Citrus Wastes into Value-Added Products: Economic and Environmently Friendly Approaches. Nutrition 2017, 34, 29–46. [Google Scholar] [CrossRef] [PubMed]
- Diab, K.A.E. In Vitro Studies on Phytochemical Content, Antioxidant, Anticancer, Immunomodulatory, and Antigenotoxic Activities of Lemon, Grapefruit, and Mandarin Citrus Peels. Asian Pacific Journal of Cancer Prevention 2016, 17, 3559–3567. [Google Scholar]
- Smeriglio, A.; Cornara, L.; Denaro, M.; Barreca, D.; Burlando, B.; Xiao, J.; Trombetta, D. Antioxidant and Cytoprotective Activities of an Ancient Mediterranean Citrus (Citrus Lumia Risso) Albedo Extract: Microscopic Observations and Polyphenol Characterization. Food Chem 2019, 279, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Wang, H.; Xu, W.; Liu, X.; Zhao, C.; Chen, J.; Wang, D.; Xu, S.; Cao, J.; Sun, C.; et al. Metabolome and Transcriptome Analysis Revealed the Basis of the Difference in Antioxidant Capacity in Different Tissues of Citrus Reticulata ‘Ponkan’. Antioxidants 2024, 13, 243. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Methods in Enzymology—Oxidants and Antioxidants Part A. Methods Enzymol 1999, 299, 152–178. [Google Scholar] [CrossRef]
- Mustafa, A.M.; Angeloni, S.; Abouelenein, D.; Acquaticci, L.; Xiao, J.; Sagratini, G.; Maggi, F.; Vittori, S.; Caprioli, G. A New HPLC-MS/MS Method for the Simultaneous Determination of 36 Polyphenols in Blueberry, Strawberry and Their Commercial Products and Determination of Antioxidant Activity. Food Chem 2022, 367. [Google Scholar] [CrossRef] [PubMed]
- Santanatoglia, A.; Cespi, M.; Perinelli, D.R.; Fioretti, L.; Sagratini, G.; Vittori, S.; Caprioli, G. Impact of the Human Factor on the Reproducibility of Different Coffee Brewing Methods. Journal of Food Composition and Analysis 2023, 124, 105698. [Google Scholar] [CrossRef]
- Santanatoglia, A.; Caprioli, G.; Cespi, M.; Ciarlantini, D.; Cognigni, L.; Fioretti, L.; Maggi, F.; Mustafa, A.M.; Nzekoue, F.; Vittori, S. A Comprehensive Comparative Study among the Newly Developed Pure Brew Method and Classical Ones for Filter Coffee Production. LWT 2023, 175, 114471. [Google Scholar] [CrossRef]
- Saltarelli, R.; Palma, F.; Gioacchini, A.M.; Calcabrini, C.; Mancini, U.; De Bellis, R.; Stocchi, V.; Potenza, L. Phytochemical Composition, Antioxidant and Antiproliferative Activities and Effects on Nuclear DNA of Ethanolic Extract from an Italian Mycelial Isolate of Ganoderma Lucidum. J Ethnopharmacol 2019, 231, 464–473. [Google Scholar] [CrossRef] [PubMed]
- Loizzo, M.R.; Sicari, V.; Tundis, R.; Leporini, M.; Falco, T.; Calabrò, V. The Influence of Ultrafiltration of Citrus Limon L. Burm. Cv Femminello Comune Juice on Its Chemical Composition and Antioxidant and Hypoglycemic Properties. Antioxidants 2019, 8, 23. [Google Scholar] [CrossRef] [PubMed]
- Flemmig, J.; Arnhold, J. Ferrous Ion-Induced Strand Breaks in the DNA Plasmid PBR322 Are Not Mediated by Hydrogen Peroxide. European Biophysics Journal 2007, 36, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Benedetti, S.; Catalani, S.; Palma, F.; Canestrari, F. The Antioxidant Protection of CELLFOOD® against Oxidative Damage in Vitro. Food and Chemical Toxicology 2011, 49, 2292–2298. [Google Scholar] [CrossRef]
- Adan, A.; Kiraz, Y.; Baran, Y. Cell Proliferation and Cytotoxicity Assays. Curr Pharm Biotechnol 2016, 17, 1213–1221. [Google Scholar] [CrossRef] [PubMed]
- Catalani, S.; Palma, F.; Battistelli, S.; Benedetti, S. Oxidative Stress and Apoptosis Induction in Human Thyroid Carcinoma Cells Exposed to the Essential Oil from Pistacia Lentiscus Aerial Parts. PLoS One 2017, 12, e0172138. [Google Scholar] [CrossRef]
- Benedetti, S.; Catalani, S.; Canonico, B.; Nasoni, M.G.; Luchetti, F.; Papa, S.; Potenza, L.; Palma, F. The Effects of Acyclovir Administration to NCI-H1975 Non-Small Cell Lung Cancer Cells. Toxicology in Vitro 2022, 79, 105301. [Google Scholar] [CrossRef]
- Bryan, N.S.; Grisham, M.B. Methods to Detect Nitric Oxide and Its Metabolites in Biological Samples. Free Radic Biol Med 2007, 43, 645–657. [Google Scholar] [CrossRef] [PubMed]
- Saini, R.K.; Ranjit, A.; Sharma, K.; Prasad, P.; Shang, X.; Gowda, K.G.M.; Keum, Y.S. Bioactive Compounds of Citrus Fruits: A Review of Composition and Health Benefits of Carotenoids, Flavonoids, Limonoids, and Terpenes. Antioxidants 2022, 11, 239. [Google Scholar] [CrossRef]
- Borghi, S.M.; Pavanelli, W.R. Antioxidant Compounds and Health Benefits of Citrus Fruits. Antioxidants 2023, 12, 1526. [Google Scholar] [CrossRef] [PubMed]
- Richa, R.; Kohli, D.; Vishwakarma, D.; Mishra, A.; Kabdal, B.; Kothakota, A.; Richa, S.; Sirohi, R.; Kumar, R.; Naik, B. Citrus Fruit: Classification, Value Addition, Nutritional and Medicinal Values, and Relation with Pandemic and Hidden Hunger. J Agric Food Res 2023, 14, 100718. [Google Scholar] [CrossRef]
- González-Molina, E.; Domínguez-Perles, R.; Moreno, D.A.; García-Viguera, C. Natural Bioactive Compounds of Citrus Limon for Food and Health. J Pharm Biomed Anal 2010, 51, 327–345. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.; Singh, J.P.; Kaur, A.; Singh, N. Phenolic Composition, Antioxidant Potential and Health Benefits of Citrus Peel. Food Research International 2020, 132, 109114. [Google Scholar] [CrossRef] [PubMed]
- De Moraes Barros, H.R.; De Castro Ferreira, T.A.P.; Genovese, M.I. Antioxidant Capacity and Mineral Content of Pulp and Peel from Commercial Cultivars of Citrus from Brazil. Food Chem 2012, 134, 1892–1898. [Google Scholar] [CrossRef] [PubMed]
- Multari, S.; Licciardello, C.; Caruso, M.; Anesi, A.; Martens, S. Flavedo and Albedo of Five Citrus Fruits from Southern Italy: Physicochemical Characteristics and Enzyme-Assisted Extraction of Phenolic Compounds. Journal of Food Measurement and Characterization 2021, 15, 1754–1762. [Google Scholar] [CrossRef]
- Ninfali, P.; Panato, A.; Bortolotti, F.; Valentini, L.; Gobbi, P. Morphological Analysis of the Seeds of Three Pseudocereals by Using Light Microscopy and ESEM-EDS. European Journal of Histochemistry 2020, 64, 3075. [Google Scholar] [CrossRef]
- Wang, Y.; Qian, J.; Cao, J.; Wang, D.; Liu, C.; Yang, R.; Li, X.; Sun, C. Antioxidant Capacity, Anticancer Ability and Flavonoids Composition of 35 Citrus (Citrus Reticulata Blanco) Varieties. Molecules 2017, 22, 1114. [Google Scholar] [CrossRef]
- Mateus, A.R.S.; Teixeira, J.D.; Barros, S.C.; Almeida, C.; Silva, S.; Sanches-Silva, A. Unlocking the Potential of Citrus Medica L.: Antioxidant Capacity and Phenolic Profile across Peel, Pulp, and Seeds. Molecules 2024, 29, 3533. [Google Scholar] [CrossRef]
- Vidal-Casanella, O.; Moreno-Merchan, J.; Granados, M.; Nuñez, O.; Saurina, J.; Sentellas, S. Total Polyphenol Content in Food Samples and Nutraceuticals: Antioxidant Indices versus High Performance Liquid Chromatography. Antioxidants 2022, 11, 324. [Google Scholar] [CrossRef]
- Chhikara, N.; Kour, R.; Jaglan, S.; Gupta, P.; Gat, Y.; Panghal, A. Citrus Medica: Nutritional, Phytochemical Composition and Health Benefits—A Review. Food Funct 2018, 9, 1978–1992. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, A.; Afzaal, M.; Saeed, F.; Ali, S.W.; Imran, A.; Zaidi, S.Y.R.; Saleem, M.A.; Hussain, M.; Al Jbawi, E. A Comprehensive Review of the Therapeutic Potential of Citrus Bioflavonoid Hesperidin against Lifestyle-Related Disorders. Cogent Food Agric 2023, 9. [Google Scholar] [CrossRef]
- Parhiz, H.; Roohbakhsh, A.; Soltani, F.; Rezaee, R.; Iranshahi, M. Antioxidant and Anti-Inflammatory Properties of the Citrus Flavonoids Hesperidin and Hesperetin: An Updated Review of Their Molecular Mechanisms and Experimental Models. Phytotherapy Research 2015, 29, 323–331. [Google Scholar] [CrossRef]
- Li, X.; Huang, W.; Tan, R.; Xu, C.; Chen, X.; Li, S.; Liu, Y.; Qiu, H.; Cao, H.; Cheng, Q. The Benefits of Hesperidin in Central Nervous System Disorders, Based on the Neuroprotective Effect. Biomedicine & Pharmacotherapy 2023, 159, 114222. [Google Scholar] [CrossRef]
- Boots, A.W.; Haenen, G.R.M.M.; Bast, A. Health Effects of Quercetin: From Antioxidant to Nutraceutical. Eur J Pharmacol 2008, 585, 325–337. [Google Scholar] [CrossRef]
- Morand, C.; Manach, C.; Crespy, V.; Remesy, C. Quercetin 3-O-Beta-Glucoside Is Better Absorbed than Other Quercetin Forms and Is Not Present in Rat Plasma. Free Radic Res 2000, 33, 667–676. [Google Scholar] [CrossRef]
- Chen, J.; Li, G.; Sun, C.; Peng, F.; Yu, L.; Chen, Y.; Tan, Y.; Cao, X.; Tang, Y.; Xie, X.; et al. Chemistry, Pharmacokinetics, Pharmacological Activities, and Toxicity of Quercitrin. Phytother Res 2022, 36, 1545–1575. [Google Scholar] [CrossRef]
- Osonga, F.J.; Akgul, A.; Miller, R.M.; Eshun, G.B.; Yazgan, I.; Akgul, A.; Sadik, O.A. Recent Advances in Potential Health Benefits of Quercetin. Pharmaceuticals 2023, 16, 1020. [Google Scholar] [CrossRef]
- Batool, S.; Javaid, S.; Javed, H.; Asim, L.; Shahid, I.; Khan, M.; Muhammad, A. Addressing Artifacts of Colorimetric Anticancer Assays for Plant-Based Drug Development. Medical Oncology 2022, 39, 198. [Google Scholar] [CrossRef] [PubMed]
- Geran, R.I.; Greenberg, N.H.; Donald, M.M. Mac PROTOCOLS FOR SCREENING CHEMICAL AGENTS AND NATURAL PRODUCTS AGAINST ANIMAL TUMORS AND OTHER BIOLOGICAL SYSTEMS. Cancer Chemother Rep 1972, 3, 1–103. [Google Scholar]
- Gözcü, S.; Polat, H.K.; Gültekin, Y.; Ünal, S.; Karakuyu, N.F.; Şafak, E.K.; Doğan, O.; Pezik, E.; Haydar, M.K.; Aytekin, E.; et al. Formulation of Hesperidin-Loaded in Situ Gel for Ocular Drug Delivery: A Comprehensive Study. J Sci Food Agric 2024. [Google Scholar] [CrossRef] [PubMed]
- Lončarić, M.; Strelec, I.; Moslavac, T.; Šubarić, D.; Pavić, V.; Molnar, M. Lipoxygenase Inhibition by Plant Extracts. Biomolecules 2021, 11, 152. [Google Scholar] [CrossRef]
- Taciak, B.; Białasek, M.; Braniewska, A.; Sas, Z.; Sawicka, P.; Kiraga, Ł.; Rygiel, T.; Król, M. Evaluation of Phenotypic and Functional Stability of RAW 264.7 Cell Line through Serial Passages. PLoS One 2018, 13, e0198943. [Google Scholar] [CrossRef]
- Pragasam, S.J.; Venkatesan, V.; Rasool, M. Immunomodulatory and Anti-Inflammatory Effect of p-Coumaric Acid, a Common Dietary Polyphenol on Experimental Inflammation in Rats. Inflammation 2013, 36, 169–176. [Google Scholar] [CrossRef]
- Iriti, M.; Varoni, E.M.; Vitalini, S. Bioactive Compounds in Health and Disease—Focus on Rutin. Bioactive Compounds in Health and Disease—Online ISSN: 2574-0334; Print ISSN: 2769-2426 2023, 6, 235–242. [Google Scholar] [CrossRef]
- Mari, G.; Catalani, S.; Antonini, E.; De Crescentini, L.; Mantellini, F.; Santeusanio, S.; Lombardi, P.; Amicucci, A.; Battistelli, S.; Benedetti, S.; et al. Synthesis and Biological Evaluation of Novel Heteroring-Annulated Pyrrolino-Tetrahydroberberine Analogues as Antioxidant Agents. Bioorg Med Chem 2018, 26, 5037–5044. [Google Scholar] [CrossRef]
- Zhao, Z. Hydroxyl Radical Generations Form the Physiologically Relevant Fenton-like Reactions. Free Radic Biol Med 2023, 208, 510–515. [Google Scholar] [CrossRef] [PubMed]
- Benedetti, S.; Nasoni, M.G.; Luchetti, F.; Palma, F. New Insights into the Cytotoxic Effects of Thymus Vulgaris Essential Oil on the Human Triple-Negative Breast Cancer Cell Line MDA-MB-231. Toxicol In Vitro 2023, 93. [Google Scholar] [CrossRef]
- Balaban, R.S.; Nemoto, S.; Finkel, T. Mitochondria, Oxidants, and Aging. Cell 2005, 120, 483–495. [Google Scholar] [CrossRef] [PubMed]
- Entezari, M.; Majd, A.; Falahian, F.; Mehrabian, S.; Hashemi, M.; Lajimi, A.A. Antimutagenicity and Anticancer Effects of Citrus Medica Fruit Juice. Acta Med Iran 2009, 47, 373–377. [Google Scholar]
- Mazaki, M.; Ishii, T.; Uyeta, M. Mutagenicity of Hydrolysates of Citrus Fruit Juices. Mutation Research/Genetic Toxicology 1982, 101, 283–291. [Google Scholar] [CrossRef] [PubMed]
- Koolaji, N.; Shammugasamy, B.; Schindeler, A.; Dong, Q.; Dehghani, F.; Valtchev, P. Citrus Peel Flavonoids as Potential Cancer Prevention Agents. Curr Dev Nutr 2020, 4. [Google Scholar] [CrossRef]
- Jacob, R.; Hasegawa, S.; Manners, G.D. The Potential of Citrus Limonoids as Anticancer Agents. 2000. [Google Scholar]
Figure 1.
Morphological analysis and semi-quantitative data of flavedo and albedo from Limoncella. A: Flavedo parenchymal cells were observed by LM (A-D), fluorescent (E), TEM (F, G), SEM (H) and ESEM (I) microscopy. Basic fuchsin (A-C) staining highlighted cellular wall and plasmodesmata Toluidine Blue (B) evidenced cytoplasmic organelles and central vacuoles. In F and G, some plastids were visible. A, H, I oil glands morphology. (A, E) Bar = 5 μm; (B, F) Bar = 2.5 μm; (D) Bar = 1 μm; (G) Bar = 0.5 mm; (H) Bar = 50 μm; (I) Bar = 100 μm.pp. B: Albedo parenchymal cells were observed by LM (A, B), differential interference contrast (C), fluorescent (D), SEM (E), TEM (F), and ESEM (G) microscopy. Basic fuchsin (A) and safranin (B) staining are used to identify cellular walls. In F, plasmodesmata structures were visible. (A, B, E, G) Bar = 20 μm; (C, D) Bar = 0.2 μm; (F) Bar = 2 μm. C: Semi-quantitative analysis performed on the albedo (A) and flavedo (B) surfaces.
Figure 1.
Morphological analysis and semi-quantitative data of flavedo and albedo from Limoncella. A: Flavedo parenchymal cells were observed by LM (A-D), fluorescent (E), TEM (F, G), SEM (H) and ESEM (I) microscopy. Basic fuchsin (A-C) staining highlighted cellular wall and plasmodesmata Toluidine Blue (B) evidenced cytoplasmic organelles and central vacuoles. In F and G, some plastids were visible. A, H, I oil glands morphology. (A, E) Bar = 5 μm; (B, F) Bar = 2.5 μm; (D) Bar = 1 μm; (G) Bar = 0.5 mm; (H) Bar = 50 μm; (I) Bar = 100 μm.pp. B: Albedo parenchymal cells were observed by LM (A, B), differential interference contrast (C), fluorescent (D), SEM (E), TEM (F), and ESEM (G) microscopy. Basic fuchsin (A) and safranin (B) staining are used to identify cellular walls. In F, plasmodesmata structures were visible. (A, B, E, G) Bar = 20 μm; (C, D) Bar = 0.2 μm; (F) Bar = 2 μm. C: Semi-quantitative analysis performed on the albedo (A) and flavedo (B) surfaces.
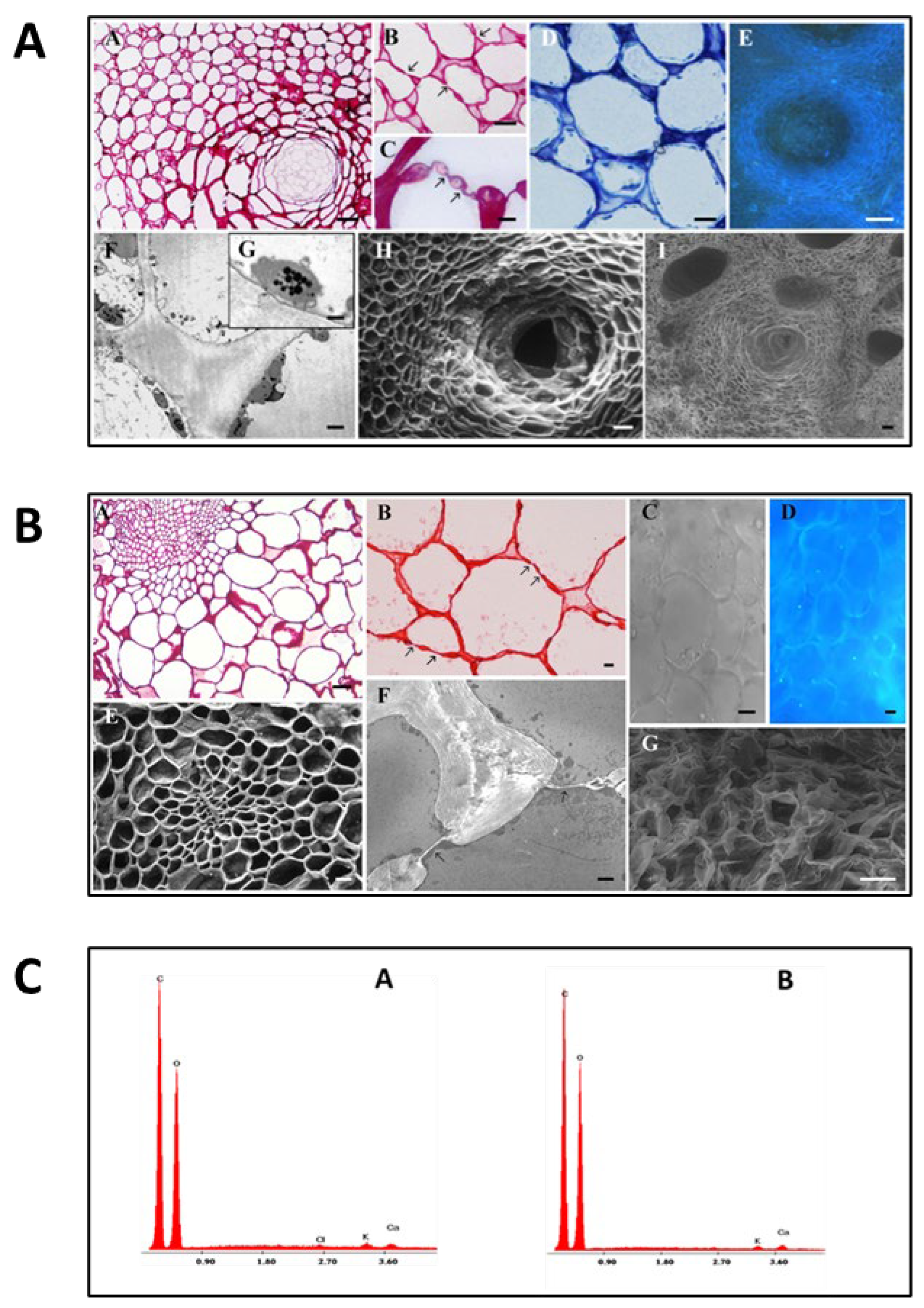
Figure 2.
Antioxidant capacity of albedo and flavedo Limoncella extracts. Scavenging effect on DPPH (A) and ABTS (B) tests. Effect on lipoxygenase activity in vitro (C). The data represent the inhibition percentage induced by increasing albedo and flavedo extracts. Data are expressed as mean ± SD (n = 3). * p < 0.05.
Figure 2.
Antioxidant capacity of albedo and flavedo Limoncella extracts. Scavenging effect on DPPH (A) and ABTS (B) tests. Effect on lipoxygenase activity in vitro (C). The data represent the inhibition percentage induced by increasing albedo and flavedo extracts. Data are expressed as mean ± SD (n = 3). * p < 0.05.
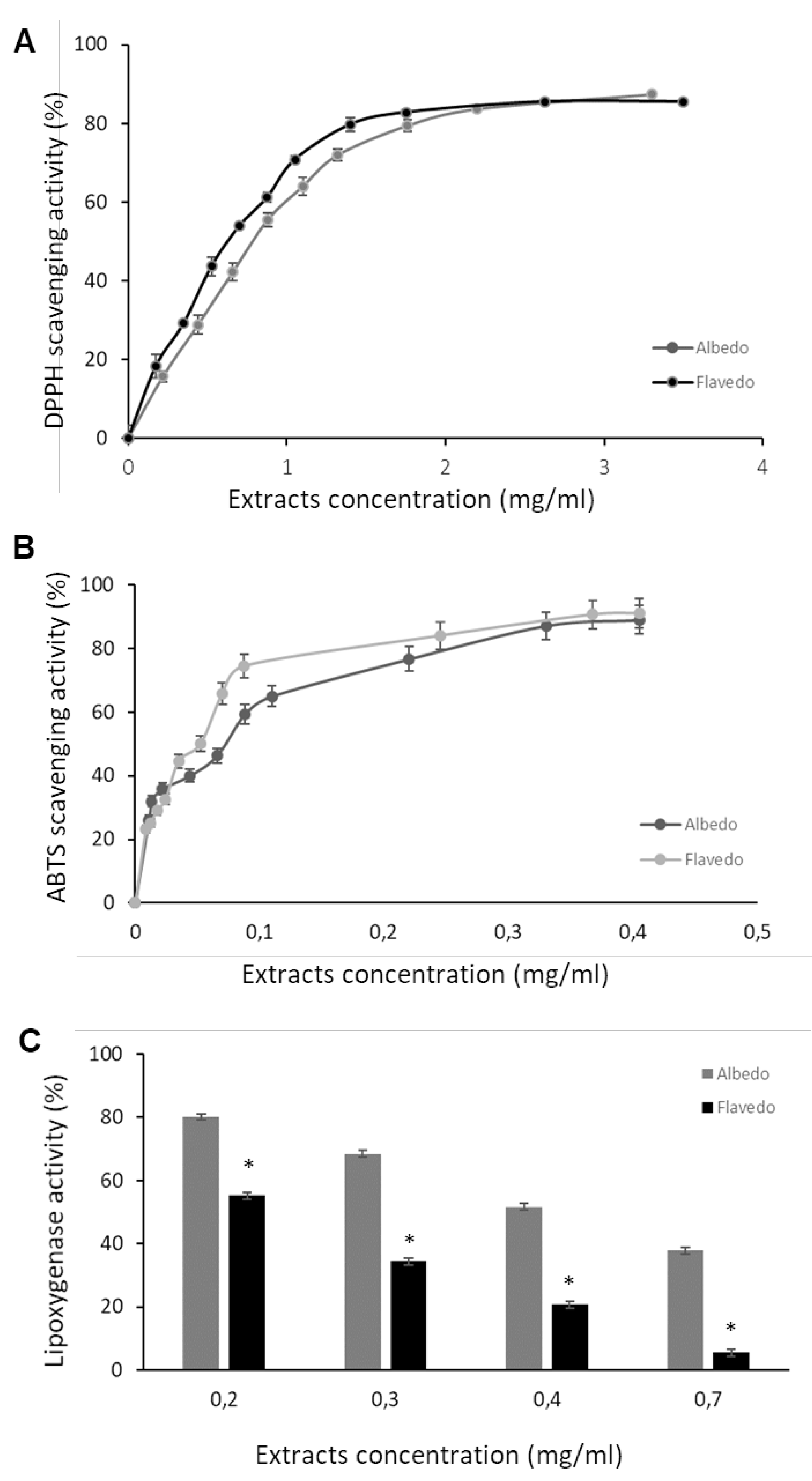
Figure 3.
Protective activity of albedo and flavedo extract versus oxidative DNA damage evaluated by DNA nicking assay. Square A reports a representative agarose gel electrophoresis of the pEMBL8 samples before (T0) and after treatment with different concentrations of hydroalcoholic extracts. CCC is a supercoiled form of a plasmid. Square B reports the quantification analyses expressed as the ratio between the CCC after treatment and CCC at time 0.
Figure 3.
Protective activity of albedo and flavedo extract versus oxidative DNA damage evaluated by DNA nicking assay. Square A reports a representative agarose gel electrophoresis of the pEMBL8 samples before (T0) and after treatment with different concentrations of hydroalcoholic extracts. CCC is a supercoiled form of a plasmid. Square B reports the quantification analyses expressed as the ratio between the CCC after treatment and CCC at time 0.
Figure 4.
Cell viability evaluation by SRB colorimetric assay upon Limoncella extracts administration (from 14 to 224 µg/ml) to HaCaT cell lines for 24 h. The cell viability was expressed as the ratio between absorbance at 570 nm of treated and untreated samples.
Figure 4.
Cell viability evaluation by SRB colorimetric assay upon Limoncella extracts administration (from 14 to 224 µg/ml) to HaCaT cell lines for 24 h. The cell viability was expressed as the ratio between absorbance at 570 nm of treated and untreated samples.
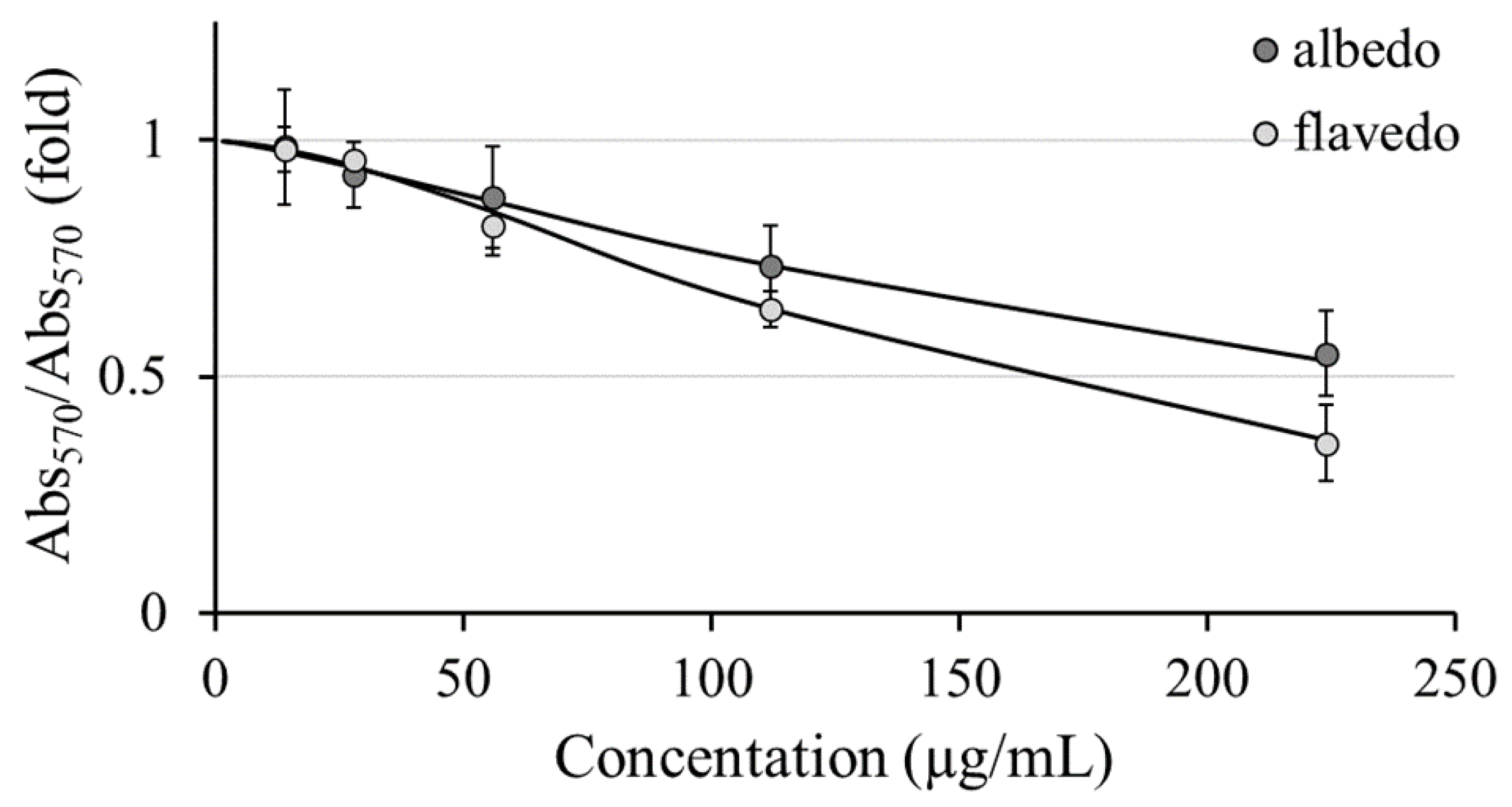
Figure 5.
Evaluation of the antioxidant properties of albedo and flavedo extract in HaCaT cells. The relative intracellular oxidation levels were obtained by incubating the cells with 50 µg/ml of each extract. The untreated and non-oxidized cells were reported as Ctrl- while the untreated and oxidized (H2O2) cells were Ctrl+. Data are expressed as mean ± SD (n = 3). * p < 0.05 vs. Ctrl-. ** p < 0.01 vs. Ctrl-.
Figure 5.
Evaluation of the antioxidant properties of albedo and flavedo extract in HaCaT cells. The relative intracellular oxidation levels were obtained by incubating the cells with 50 µg/ml of each extract. The untreated and non-oxidized cells were reported as Ctrl- while the untreated and oxidized (H2O2) cells were Ctrl+. Data are expressed as mean ± SD (n = 3). * p < 0.05 vs. Ctrl-. ** p < 0.01 vs. Ctrl-.
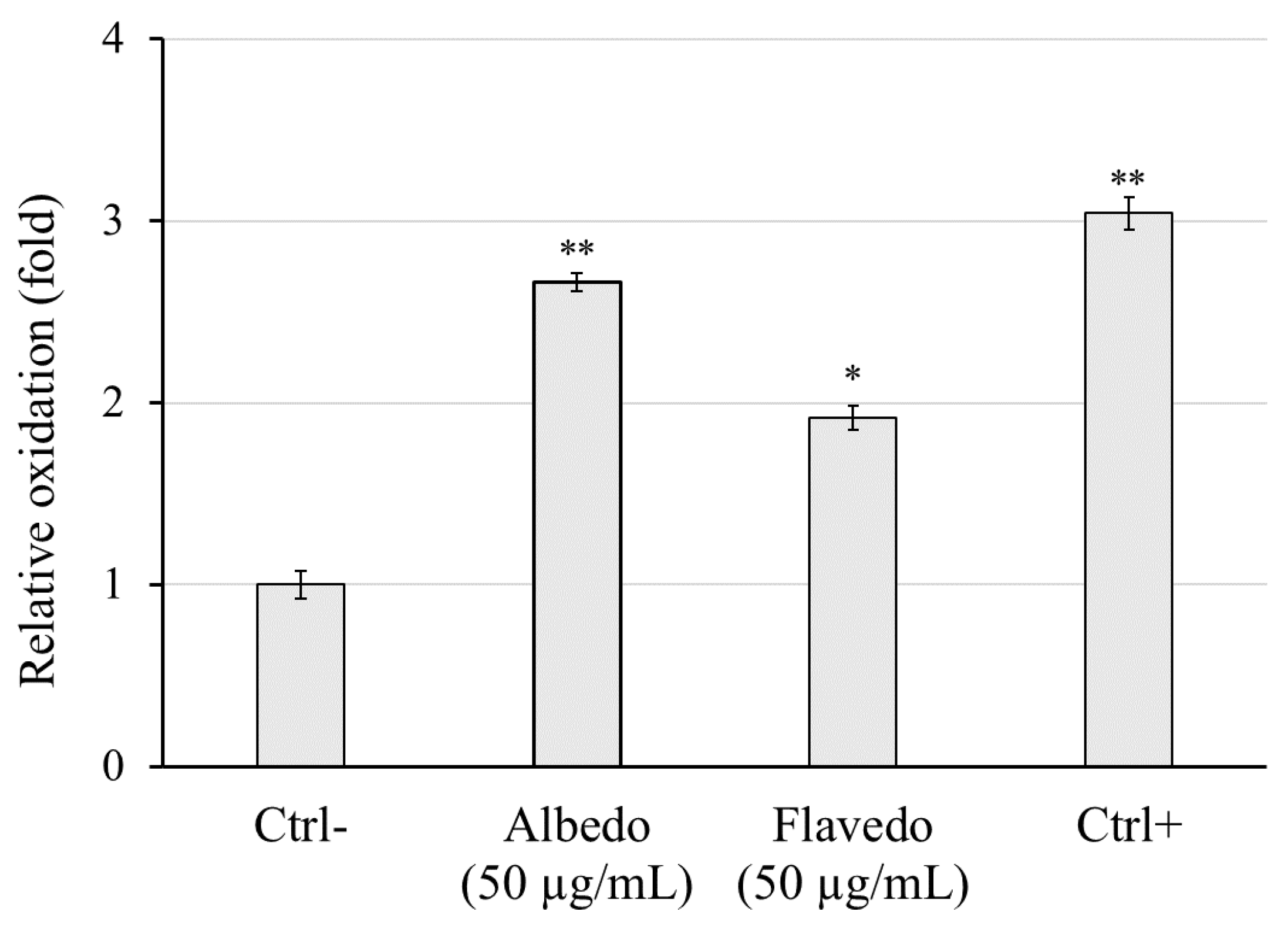
Figure 6.
Extracellular NO release after RAW 264.7 stimulation by LPS for 24 h in the presence of albedo and flavedo extracts (50 µg/mL). Ctrl-: cell without stimulus or extract; Ctrl+: cell stimulated by 1 µg/mL LPS; Alb: 50 µg/mL albedo + 1 µg/mL LPS; Fla: 50 µg/mL flavedo + 1 µg/mL LPS; Dex: 5 µg/mL dexamethasone + 1 µg/mL LPS. Data are expressed as the mean ± SD (n = 3). * p < 0.05, ** p < 0.01 vs. Ctrl+ (Tukey’s post hoc test).
Figure 6.
Extracellular NO release after RAW 264.7 stimulation by LPS for 24 h in the presence of albedo and flavedo extracts (50 µg/mL). Ctrl-: cell without stimulus or extract; Ctrl+: cell stimulated by 1 µg/mL LPS; Alb: 50 µg/mL albedo + 1 µg/mL LPS; Fla: 50 µg/mL flavedo + 1 µg/mL LPS; Dex: 5 µg/mL dexamethasone + 1 µg/mL LPS. Data are expressed as the mean ± SD (n = 3). * p < 0.05, ** p < 0.01 vs. Ctrl+ (Tukey’s post hoc test).

Table 1.
Concentration (mg kg-1) of bioactive compounds in Albedo and Flavedo extract (n = 3).
| No | Compounds | Albedo Concentration mg kg-1 | Flavedo Concentration mg kg-1 |
|---|---|---|---|
| 1 | Gallic acid | 5.57 | 41.20 |
| 2 | Neochlorogenic acid | n.d. a | n.d. |
| 3 | Delphinidin-3-galactoside | n.d. | 7.35 |
| 4 | (+)-Catechin | n.d. | n.d. |
| 5 | Procyanidin B2 | n.d. | n.d. |
| 6 | Chlorogenic acid | 0.80 | 1.54 |
| 7 | p-Hydroxybenzoic acid | 411.34 | 895.60 |
| 8 | (-)-Epicatechin | n.d. | 0.69 |
| 9 | Cyanidin-3-glucoside | n.d. | n.d. |
| 10 | Petunidin-3-glucoside | 0.45 | 1.82 |
| 11 | 3-Hydroxybenzoic acid | n.d. | n.d. |
| 12 | Caffeic acid | 6.92 | n.d. |
| 13 | Vanillic acid | 134.39 | n.d. |
| 14 | Resveratrol | n.d. | n.d. |
| 15 | Pelargonidin-3-glucoside | n.d. | 0.50 |
| 16 | Pelagonidin-3-rutinoside | n.d. | n.d. |
| 17 | Malvidin-3-galactoside | n.d. | n.d. |
| 18 | Syringic acid | n.d. | 36.43 |
| 19 | Procyanidin A2 | n.d. | n.d. |
| 20 | p-Coumaric acid | 143.89 | 1566.65 |
| 21 | Ferulic acid | 147.86 | 609.96 |
| 22 | 3,5-Dicaffeoylquinic acid | n.d. | n.d. |
| 23 | Rutin | 137.04 | 5996.78 |
| 24 | Hyperoside | n.d. | 245.49 |
| 25 | Isoquercitrin | 9.73 | 216.05 |
| 26 | Delphindin-3,5-diglucoside | 68.98 | 212.24 |
| 27 | Phloridzin | 0.82 | n.d. |
| 28 | Quercitrin | 1961.17 | 267.76 |
| 29 | Myricetin | 0.41 | n.d. |
| 30 | Naringin | 3.86 | 1.65 |
| 31 | Kaempferol-3-glucoside | 47.45 | 4.73 |
| 32 | Hesperidin | 11625.45 | 778.88 |
| 33 | Ellagic acid | n.d. | 24.49 |
| 34 | trans-cinnamic acid | 13.23 | 4.09 |
| 35 | Quercetin | 12.92 | 73.08 |
| 36 | Phloretin | n.d. | n.d. |
| 37 | Kaempferol | 128.20 | 246.73 |
| 38 | Isorhamnetin | 3.22 | 158.56 |
| Total content (mg kg-1) | 14863.69 | 11392.24 | |
| Total content (%) | 1.48 | 1.14 | |
| Total content (mg kg-1) Folin-Ciocalteu method | 18700 | 22040 | |
a n.d., not detectable. Relative standard deviation (RSD) for all compounds ranged from 2.54 to 10.31%.
Table 2.
Antioxidant activities of albedo and flavedo extracts.
| Part of fruit | DPPH EC50 (mg/ml) | ABTS EC50 (mg/ml) | Lipoxygenase EC50 (mg/ml) | Total Antioxidant Capacity (µg TE/mg) |
|---|---|---|---|---|
| Albedo | 0.996 ± 0.17 | 0.063 ± 0.029 | 0.54 ± 0.10 | 35.42 ± 3.31 |
| Flavedo | 0.971 ± 0.34 | 0.063 ± 0.012 | 0.14 ± 0.06 * | 40.41 ± 10.15 |
Values are expressed as mean ± standard error of the mean (n = 3). * P < 0.01 DPPH: 2,2-Diphenyl-1-picrylhydrazyl; ABTS: 2,2′-Azino-bis(3-ethylbenzothiazoline-6-sulfonic acid)
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



