
Submitted:
18 September 2024
Posted:
19 September 2024
Read the latest preprint version here
Abstract
This study evaluated the effects of replacing zinc bacitracin with a symbiotic diet on the biochemical profile, liver development, and Clostridium perfringens presence in laying hens at 90 weeks of age. A total of 198 Dekalb–White hens were assigned to six experimental diets: MBM (corn, soybean meal, and meat and bone meal), BAC (MBM + 0.05% zinc bacitracin), and SIMC (MBM + 0.1% symbiotic), provided from day 1 to 90 weeks. Additional treatments, SIMR and SIMP, received symbiotics from weeks 6 and 17, re-spectively. At 90 weeks, blood and tissue samples were analyzed. Results showed that symbiotic-treated hens had 20% heavier livers (P < 0.01) and lower concentrations of uric acid, total proteins, AST, ALT, and LDH (P < 0.01) compared to BAC-treated birds. Phosphorus, triglycerides, and HDL levels were better in symbiotic groups. C. perfringens counts and antibiotic resistance (ampicillin, erythromycin, aminoglyco-sides, lincomycin, tetracycline, bacitracin) were significantly higher in BAC groups (P < 0.01). Symbiotic supplementation improved intestinal health, reduced pathogenic bacterial colonization, and enhanced liver function compared to continuous antibiotic use, especially when administered from early life stages.
Keywords:
Prebiotic
; Probiotic
; Liver health
1. Introduction
Laying hens in the final cycle stage presents a significant challenge for the poultry industry, as they exhibit a decline in performance and egg quality [1]. During this phase, deterioration of intestinal health is a primary cause of immunological problems and issues related to nutrient digestion and absorption, which negatively affects laying performance [2]. Consequently, the poultry sector seeks solutions to enhance intestinal health, optimize laying yield, and extend the production period of mature laying hens to improve their productive performance [3].
With the prohibition of antibiotics as growth promoters, the poultry industry has turned to the use of symbiotics as a promising strategy to improve avian health and reduce bacterial resistance [4]. Symbiotics combine live microorganisms and substrates that promote the health of the host. The association between probiotics and prebiotics stimulates the growth of beneficial bacteria and consequently reduces bacterial resistance [5,6]. Probiotic supplementation in poultry feed is viewed as a performance enhancer because it increases nutrient digestibility [7,8], amplifies the cecal microbiota [9], improves the immune response [10], and reduces serum and yolk cholesterol levels in laying hens [11]. Adriani et al. [12] observed that the use of a probiotic compound in the diet of 90-week-old laying hens improved productive performance and reduced the presence of pathogenic bacteria in the intestine, thereby increasing immune cell activity.
Conversely, prebiotic ingestion modulates the colonic microbiota, increasing the proportion of probiotic bacteria such as lactobacilli and bifidobacteria [13], reducing pathogenic bacteria [14], and improving intestinal mineral absorption [15]. Considering that from 50 weeks of age, there is a predominance of potentially pathogenic bacterial genera (Bacteroidetes and Firmicutes) in the intestines of chickens [16], symbiotic products are a promising option for modulating the microbiota and reducing the presence of pathogenic bacteria such as C. perfringens [17,18,19].
The C. perfringens, a gram-positive anaerobic bacterium known for its toxins, can cause diseases in poultry, including necrotic enteritis [20]. The type G strain is responsible for significant economic losses in poultry farming [21], highlighting the importance of preventive strategies.
Symbiotics show promise in poultry farming, but a scientific consensus on the optimal supplementation timing remains elusive [22,23,24]. Current practices, largely based on empirical observations, highlight the need for rigorous research to determine effective administration strategies. The relationship between symbiotic supplementation timing and microbial colonization in laying hens is poorly understood, as is the long-term efficacy of continuous zinc bacitracin supplementation. Critical areas for investigation include the temporal dynamics of microbiome establishment, the impact of prolonged antimicrobial exposure on gut health and egg production, and potential interactions between symbiotics and host physiology across different life stages. Elucidating these factors is crucial for developing evidence-based protocols that optimize health and productivity in laying hens while mitigating concerns about antimicrobial resistance. Comprehensive studies that address these knowledge gaps are essential for advancing sustainable and effective poultry management practices.
Therefore, the present study aimed to evaluate the long-term effects of a symbiotic compound included in diets from different phases of poultry rearing, considering the serum biochemical profile, liver weight, isolation, and antibiotic resistance of C. perfringens.
2. Materials and Methods
Animal Ethics Statement
The research was approved by the Ethics Committee on Animal Use of the Federal Rural University of Pernambuco (CEUA, No. 060/2019), and all animal experiments complied with the guidelines.
Animals and Husbandry
A total of 198 one-day-old Dekalb–White lineage birds were housed from 1 to 16 weeks in a masonry shed equipped with metal cages measuring 100 x 80 x 50 cm. The cage dimensions were adjusted to 100 x 40 x 45 cm.
The average temperature and relative humidity data were recorded using a thermo-hygrometer, and specific values were indicated for each phase of the experiment. During weeks 1–5, the average temperature was 28.5°C and relative humidity was 70.23%. The temperature was achieved using heating lamps. During the rearing phase, the temperature was 27.7°C, and the relative humidity was 67%. In the production phase, the average temperature was 31°C, with a relative humidity of 72%.
The lighting program was adjusted according to each rearing phase of the birds. In the first week, the birds were exposed to 24 h of light. The light exposure was reduced by 1 h per day until 12 h of light was achieved after 6 weeks. The 12 h of light was maintained until the birds reached 16 weeks old. From week 17 forward, 12 h of natural light and 4 h of artificial light were provided, totaling 16 h of light.
Birds received diets formulated to meet their specific nutritional needs during each phase of the experiment, according to lineage recommendations. Feed supply was controlled to adequately meet the birds’ nutritional requirements. Water was provided ad libitum throughout the experimental period. The experiment had a total duration of 90 weeks.
Experimental Design
The birds were distributed in a completely randomized design across 5 experimental treatments with 6 replicates, consisting of 3 replicates of 6 birds and 3 replicates of 5 birds, totaling 165 animals.
The experimental diets are described in Table 1 and Table 2 consisted of a diets containing corn, soybean meal, and meat and bone meal. The treatments were:
-
Treatment MBM:
- ○
- Diet based on corn, soybean meal, and meat and bone meal administered from 1 to 90 weeks.
-
Treatment BAC:
- ○
- Diet based on corn, soybean meal, and meat and bone meal, with the addition of 0.05% zinc bacitracin administered from 1 to 90 weeks.
-
Treatment SIMC:
- ○
- Diet based on corn, soybean meal, and meat and bone meal, supplemented with 0.1% symbiotic, administered from 1 to 90 weeks.
-
Treatment SIMR:
- ○
- From 1 to 5 weeks: Diet based on corn, soybean meal, and meat and bone meal.
- ○
- From 6 to 90 weeks: Diet based on corn, soybean meal, and meat and bone meal, supplemented with 0.1% symbiotic.
-
Treatment SIMP:
- ○
- From 1 to 16 weeks: Diet based on corn, soybean meal, and meat and bone meal.
- ○
- From 17 to 90 weeks: Diet based on corn, soybean meal, and meat and bone meal, supplemented with 0.1% symbiotic.
Dietary Treatments
The composition of ingredients and concentrations of nutrients and energy are presented in Table 1 and Table 2. Diets were formulated based on the nutritional requirements recommended in the Dekalb–White lineage management guide, and the chemical composition and energy values of the feed ingredients were obtained from Rostagno et al. [25].
The symbiotic used had the following composition: prebiotics (mannans 52 g/kg and glucans 28 g/kg) and probiotics (Saccharomyces cerevisiae 2.00×10¹¹ CFU/kg, Bifidobacterium bifidum 2.00×10¹¹ CFU/kg, Bacillus subtilis 2.88×10¹¹ CFU/kg, Enterococcus faecium 2.08×10¹¹ CFU/kg, Lactobacillus acidophilus 1.04×10¹¹ CFU/kg).
Blood Collection and Biochemical Analysis
At 90 weeks, 2 birds per experimental plot were selected to collect 4 ml of blood from the ulnar vein. The blood samples were placed in tubes with a clot activator to obtain blood serum, which was then stored in 1.5 ml Eppendorf tubes. Serum samples were analyzed using commercial kits (Bioclin) and an automatic biochemical analyzer (BIOCLIN® 1000), which were previously calibrated according to the manufacturer’s methodology. The variables analyzed were uric acid (URI) activity, total proteins (TP), creatine kinase (CK), aspartate aminotransferase (AST), alanine aminotransferase (ALT), lactate dehydrogenase (LDH), phosphorus (PHOS), high-density lipoprotein (HDL), low-density lipoprotein (LDL), triglycerides (TRI), and total cholesterol (CHOL).
Organ Collection
At 90 weeks, one bird per experimental plot was selected based on the average weight (1658 ± 84g) of the plot and euthanized by cervical dislocation for organ collection. Livers were weighed using an analytical balance. Subsequently, the left cecum was separated with sterile scissors for later counting and isolation of C. perfringens, following the guidelines of the “Official Analytical Methods for Microbiological Analysis for Control of Animal-Origin Products and Water” (Brazil, 2003).
Isolation of C. perfringens and Antimicrobial Resistance
One gram of cecal content from each sample was collected and homogenized in 9 ml of 0.9% saline solution via vortexing for 30 s. From this dilution, 2 additional subsequent dilutions were performed in saline solution. The dilutions (10-1, 10-2, and 10-3) were placed on Petri dishes containing Sulfite Polymyxin Sulfadiazine agar using the spread plate technique. The plates containing the inoculated dilutions were incubated at 45°C for 24 h in anaerobic jars under microaerophilic conditions. Because the plates showed reduced counts, the counts were standardized at the first dilution (10-1). Dark colonies with diameters greater than 1 mm were counted and expressed as colony-forming units per gram (CFU/g).
Confirmatory tests for Gram staining, motility, carbohydrate fermentation (dextrose, lactose, and sucrose), indole, catalase, and beta-hemolysis were performed using the control strain of C. perfringens (ATCC 13124), which was provided by the Microbiology Laboratory of the Federal Laboratory of Agricultural Defense of Pernambuco (LFDA-PE). The disk diffusion method (Bauer et al., 1966) was used to determine the sensitivity of C. perfringens isolates to various antibiotics using Muller–Hinton agar according to the CLSI guidelines [26].
All colonies that grew on specific agar were transferred to blood agar and incubated at 37°C under microaerophilic conditions for 24 h. Subsequently, a pool of 10 colonies was created from each treatment group based on the cecal isolates. These colonies were suspended in 0.9% sodium chloride solution to achieve the MacFarland 0.5 standard and then seeded on Muller–Hinton agar plates. CECON discs containing ampicillin 2 µg, ciprofloxacin 5 µg, erythromycin 15 µg, gentamicin 10 µg, neomycin 30 µg, lincomycin 2 µg, tetracycline 30 µg, and bacitracin (10 IU) were used. The plates were incubated at 37°C for 24 h. The inhibition zones formed in response to each antibiotic were measured and expressed in millimeters. Based on the size of the inhibition zones, the isolates were classified as resistant (R), intermediate (I), or sensitive (S) to antibiotics according to the CLSI criteria.
Statistics
The normality and homoscedasticity assumptions were tested before the analysis of variance. Data analysis was performed using the PROC GLM procedure in the Statistical Analysis System (SAS) software, version 9.4 (SAS, 2008). To meet the homoscedasticity assumption, the data were log-transformed. In cases in which statistically significant differences were found, the means were compared using the Student-Newman-Keuls (SNK) test at a significance level of 5%. The statistical model used was as follows:
where:
Yij= μ+Ti+εij
Yij = observation
μ = overall mean constant common to all observations
Ti = Dietary effect
εij = random error term
3. Results
The treatments significantly affected the biochemical profiles of the birds (Table 3 and Table 4). Birds receiving symbiotics from the rearing phase (SIMR) had livers that were 20% heavier than those receiving BAC treatment (P < 0.01). Birds treated with MBM and BAC exhibited higher URI concentrations compared to those receiving symbiotics, regardless of the supplementation phase (P < 0.01). The BAC treatment resulted in an average 27% higher TP concentration compared with SIMP treatment (P < 0.01). The diets did not affect the serum CK levels of the birds (P = 0.95). Birds in the BAC treatment had higher serum AST concentrations than those in the MBM and SIMP treatments (P < 0.01). The BAC treatment also resulted in the highest serum ALT concentrations, whereas SIMR and SIMP treatments resulted in the lowest levels (P < 0.01). The serum LDH concentrations were highest in birds supplemented only with MBM (P < 0.01), differing from those in the other treatments. The BAC treatment resulted in the highest serum P concentrations (P = 0.02) compared to the other treatments.
Serum lipid concentration was affected by the treatments. The BAC and MBM treatments resulted in the lowest HDL concentrations (P < 0.01) compared to birds supplemented with symbiotics. Specifically, HDL concentrations were 61% and 48% higher in the SIMP group than in the BAC and MBM groups, respectively. Additionally, BAC treatment increased triglyceride concentrations (P = 0.03). Diets supplementation did not significantly influence CHO and LDL levels in birds (P = 0.60).
A significant difference was observed in the number of colony-forming units (CFU) of C. perfringens (Figure 1). Birds supplemented with MBM and BAC had higher numbers of CFUs than those receiving SIC treatment (P < 0.01). However, birds supplemented with symbiotics in the SIMR and SIMP treatments did not differ significantly from those in the BAC and MBM treatments.
Alongside these data (Figure 2), the antibiogram results indicated that C. perfringens isolates exhibited resistance to ampicillin in MBM and BAC groups, whereas isolates from the remaining treatments were sensitive to this antibiotic. Resistance to ciprofloxacin was not observed in any treatment. Resistance to erythromycin was detected in bacteria isolated in the BAC treatment. In the case of aminoglycosides, bacterial resistance was observed in the BAC treatment, with intermediate resistance to gentamicin and neomycin in the SIMR and SIMP treatments, respectively. Resistance to lincomycin was noted in the SIMR, MBM, and BAC treatments, whereas treatments containing symbiotics were either sensitive or showed intermediate resistance. For tetracycline and bacitracin, C. perfringens resistance was exclusively observed in birds in the BAC treatment.
4. Discussion
In laying hens, uric acid is the primary nitrogen metabolite produced by the kidneys and liver. This study found that birds fed symbiotic diets had lower uric acid concentrations than those receiving antibiotics. A similar reduction in serum uric acid levels was reported by Li et al. [27] in ducks fed a probiotic-based diet containing Bacillus subtilis, which was attributed to the probiotic’s ability to reduce urease-producing bacteria. Sugiharto et al. [28] reported similar results using a probiotic comprising various Bacillus strains. Probiotic microorganisms can utilize uric acid and other toxic compounds for growth [29]. Therefore, the significant decrease in uric acid levels in the symbiotic-treated groups indicates that the probiotics have beneficial effects on renal function [30].
Most plasma proteins, except immunoglobulins, are synthesized by the liver. These proteins primarily maintain blood volume through colloidal osmotic balance and help regulate blood pH through their buffering capacity. Additionally, they transport metabolites and hormones and play regulatory and catalytic roles in various biological processes. Some plasma proteins are essential for immunoinflammatory responses and tissue repair and restructuring [31]. Maintaining normal plasma protein levels is crucial for ensuring adequate colloidal osmotic pressure, which helps regulate blood pH and volume. In this study, birds consuming zinc bacitracin exhibited higher plasma protein activity, a finding that is in contrast with previous studies involving probiotics or prebiotics in poultry [28,32,33].
The AST is an enzyme involved in protein metabolism and plays a key role in cell membrane and amino acid synthesis [34]. The highest AST activity is found in the heart, liver, kidneys, muscles, and nervous tissues [35] because of the high metabolic activity in these tissues, which require continuous cellular function to maintain structural integrity. When cells in these organs begin to break down, AST is released into the bloodstream, leading to increased levels along with TP [36]. This study demonstrated that symbiotic supplementation reduced AST concentrations compared to antibiotics, along with lower TP levels, especially when supplemented in the production phase. Correlation between AST and TP levels indicates that prolonged antibiotic use can cause tissue damage.
Moreover, the presence and quantity of serum enzymes can indicate the degree of organ or tissue damage. Serum concentrations of liver enzymes, such as AST and ALT, are commonly used to evaluate avian liver function because these enzymes are synthesized in the liver [37]. In this study, laying hens fed with symbiotics exhibited lower AST and ALT activity, indicating good liver health. Tang et al. [38] reported similar results when symbiotics were included in the diets of laying hens aged 20–52 weeks. In a study by Attia et al. [39] on rabbits, continuous feeding with mannan oligosaccharide and zinc bacitracin resulted in increased liver enzyme levels (AST and ALT) with prolonged antibiotic use compared to the prebiotic. This increase was linked to histological evidence showing that continuous zinc bacitracin administration had harmful effects on the liver, kidney, and ileum, including multifocal dilation in liver tissue and atrophy of renal tubules.
In healthy birds, plasma LDH activity typically remains below 1,000 IU/L, and increases are associated with hepatocellular disease or muscle injury. However, to confirm such diagnoses, it is essential to assess the concentrations of CK, AST, and ALT enzymes [36]. In this study, birds fed MBM exhibited abnormal LDH concentrations (1154 IU/L). Despite these changes, hepatic enzyme activity remained stable, indicating muscle and liver lesions. In contrast, birds receiving antibiotics had significantly higher LDH levels compared to those receiving symbiotics, along with alterations in hepatic enzyme activity, indicating potential liver damage. Michalska et al. [40] found that probiotics reduced LDH concentrations and challenge-induced lesions in C. perfringens-infected quails, attributing these effects to the anti-inflammatory and hepatoprotective properties of probiotics.
Plasma phosphorus levels are primarily regulated by renal excretion, stimulated by parathyroid hormone. In birds, hyperphosphatemia is defined as serum phosphorous concentrations exceeding 7.0 mg/dL [36]. This study found that phosphorus levels in the antibiotic-treated group were above the normal limit (11.45 mg/dL). Such elevations can occur in severe renal disease due to reduced glomerular filtration rate [36]. Long-term use of zinc bacitracin, as previously noted, can impair renal function and cause organ damage [39]. In this study, birds were evaluated at 90 weeks of age and those receiving BAC treatment from the first day of life. Elevated levels of enzymes such as AST, ALT, and uric acid were observed, indicating potential hepatotoxicity from continuous antibiotic administration. Conversely, birds that received symbiotics maintained healthier levels of these enzymes, indicating better liver function.
High-density lipoproteins (HDL) remove CHO from the arteries and transport it back to the liver, thereby preventing its accumulation [41]. The inclusion of symbiotics, regardless of age, resulted in increased HDL concentrations, corroborating the findings of Tang et al. [38], who reported that symbiotic inclusion (containing Lactobacillus acidophilus, Lactobacillus casei, Bifidobacterium bifidum, Streptococcus faecium, Aspergillus oryzae, and isomaltooligosaccharide) increased HDL levels in laying hens from 20 to 52 weeks. Although this study did not show a difference in serum cholesterol levels with symbiotic use, the increase in HDL likely contributes to reduced cholesterol levels [41]. These results indicate that probiotics have a hypocholesterolemic effect on the host, although the exact mechanism remains unclear. Possible explanations include the utilization of cholesterol by probiotics [42], inhibition of hydroxymethylglutaryl coenzyme A reductase [43], and conversion of cholesterol to coprostanol by cholesterol reductase produced by probiotics [44].
Treatment with zinc bacitracin resulted in higher cholesterol concentrations than treatment without additives, which only included meat and bone meals. This finding contradicts that of Ogboko [45] who reported that zinc bacitracin reduced serum cholesterol levels in broilers. However, since broilers have a short production cycle, this reduction may not reflect long-term effects. As previously reported, continuous zinc bacitracin administration can impair kidney and liver function, potentially disrupting cholesterol metabolism.
In this study, birds fed symbiotics from 6 or 17 weeks of age exhibited higher relative liver weights than those fed antibiotics. These findings agree with previous studies reporting similar increases in liver weight with probiotic use [46,47,48]. The correlation between symbiotic use and liver enlargement is not fully understood, but a study by Sharma, Bhardwaj, and Singh [49] found that administering probiotics (Lactobacillus casei and Bifidobacterium bifidum) to rats increased liver glycogen levels, which may explain the organ’s growth. In contrast, birds treated with antibiotics exhibited reduced relative liver weight compared to those treated with symbiotics. These findings are consistent with those of Attia et al. [39], who observed sinusoidal congestion and liver atrophy in rabbits treated continuously with zinc bacitracin.
By associating hepatic enzyme activity with liver size, this study found that birds treated with zinc bacitracin exhibited higher AST and ALT levels, implying possible liver damage. Conversely, birds supplemented with symbiotics showed improved enzyme levels, indicating larger and healthier livers. The administration of symbiotics from the first day of life resulted in a lower colony-forming unit (CFU/g) count in C. perfringens than in birds that received only zinc bacitracin. This effect was not observed when the symbiotics were introduced at 6 or 17 weeks of age. Abdelqader et al. [50] similarly demonstrated that a symbiotic comprising Bacillus subtilis and inulin reduced C. perfringens counts in the cecal content of laying hens treated from 64 to 75 weeks of age.
The beneficial effects of bacterial fermentation include modulating the gastrointestinal microbiota, promoting the growth of beneficial microorganisms, and producing advantageous postbiotics. Examples include bacteria of the genus Bifidobacterium spp. and lactic acid, which can inhibit the growth of pathogenic microorganisms such as Salmonella, Escherichia coli, and Clostridium perfringens [51,52]. However, the use of antibiotics, whether for therapeutic purposes or growth promotion, can increase microbial resistance in animals [53]. The development of resistant bacteria, along with an expanding range of antibiotic resistance genes, represents a significant concern, particularly in the poultry sector. Understanding the prevalence and dynamics of resistant microorganisms in this context is crucial [54].
Studies on the use of additives, especially symbiotics, in relation to antibiotic resistance in bacteria isolated from laying hens are limited. Roth et al. [54] evaluated the prevalence of antibiotic-resistant E. coli in broilers challenged with a multi-resistant E. coli strain and fed with ampicillin, organic acids (20% formic acid, 10% acetic acid and 5% propionic acid), or symbiotic. However, no studies have specifically addressed C. perfringens resistance in laying hens or broilers. Most research involving the isolation of C. perfringens from laying hens and the evaluation of its resistance profile has focused on field studies in poultry farms, where resistance is often attributed to the indiscriminate use of antibiotics in poultry nutrition [55,56,57].
This study demonstrated that symbiotics, regardless of the phase of inclusion, positively affected the susceptibility of C. perfringens to various antibiotics. Probiotics included in animal feed exert their effects through mechanisms such as competitive exclusion, bacteriocin production, intestinal structure reinforcement, immune function, and intestinal transit [57]. These mechanisms apply less selective pressure compared to antibiotics, potentially reducing the resistance of pathogenic bacteria.
Sáenz et al. [58] investigated the impact of broad-spectrum antibiotics on intestinal microbiota diversity, antimicrobial resistance genes, and mobile genetic elements in fish microbiota. They found that antibiotic treatment led to the dominance of pathogenic bacterial communities and resistance mechanisms, with antibiotic efflux positively correlated with the transfer of resistance via mobile genetic elements. However, these effects diminished after discontinuing antibiotic use. In the present study, since bacitracin was used continuously, the effects of gene transfer by mobile genetic elements may have been more pronounced, particularly due to the selective pressure exerted by the antibiotic.
C. perfringens is a commensal bacterium found in poultry, with a higher prevalence in the cecum [59]. This bacterium is frequently subjected to selective pressure from antibiotic treatments, which significantly contributes to the spread of resistance to these compounds [60]. Studies have shown that C. perfringens isolated from poultry exhibit resistance to aminoglycosides, polypeptides [55], erythromycin, tetracycline, lincomycin, bacitracin [61], and ampicillin [62].
Silva et al. [63] reported the antimicrobial susceptibility of C. perfringens strains isolated from poultry and identified resistance to bacitracin. They found that C. perfringens harbors resistance genes on plasmids, such as pCW3, which is associated with resistance to tetracycline and bacitracin and can be modified to influence resistance to other antibiotics [64,65]. This study found that introducing symbiotics only after the laying phase resulted in greater bacterial colonization, with an intermediate resistance profile to some antibiotics. Previous studies have not explored the relationship between the timing of symbiotic inclusion in animal feeds and bacterial resistance, making this the first to evaluate this aspect. Further research is needed to assess the influence of the timing of symbiotic inclusion on bacterial resistance pressure or to determine whether pre-established microbiota diminish the effectiveness of symbiotics in reducing C. perfringens susceptibility.
5. Conclusions
Symbiotic supplementation in the diets of laying hens significantly improves intestinal health, reduces pathogenic bacterial colonization, and enhances liver function compared to continuous antibiotic use. Specifically, symbiotics administered from early life stages reduce C. perfringens counts and antibiotic resistance. Symbiotics are a promising alternative to antibiotics for enhancing poultry health and productivity while minimizing the risks associated with prolonged antibiotic use.
Author Contributions
W.A.S., and D.P.d.O.: investigation; C.B-V.R., J.C.d.S.N., and L.F.A.S.,P: supervision; W.A.d.S., L.F.A.d.S and M.J.B.d.S.: writing-original draft preparation, A.G.R., D.A.d.S., and S.M.d.S.C data curation; L.F.A.d.S., and M.J.B.d.S.: analysis; L.F.A.d.S., and C.B-V.R: funding acquisition; M.R.B., M.N.T., A.G.R., D.A.d.S., and S.M.d.S.C: writing-review and editing.
Funding
This research was funded by Coordination for the Improvement of Higher Education Personnel (CAPES) for the fellowship grant and financial support for this study.
Institutional Review Board Statement
The study was certified by the Ethics Committee on the Use of Animals (CEUA) of the Universidade Federal Rural de Pernambuco (UFRPE), number 23082.011622/2019-14 with license 060/2019.
Data Availability Statement
The data that support the findings of this study are available on request from the corresponding author.
Acknowledgments
The authors would like to thank the Federal Rural University of Pernambuco (UFRPE) for the support of the technical team and the use of laboratories to carry out the other analyses in this research. Coordination for the Improvement of Higher Education Personnel (CAPES) for financial support.
Conflicts of Interest
The authors declare no conflicts of interest
References
- Hester, P.Y.; Enneking, S.A.; Jefferson-Moore, K.; Einstein, M.; Cheng, H.W.; Rubin, D.A. The effect of perches in cages during pullet rearing and egg laying on hen performance, foot health, and plumage. Poultry Science 2013, 92, 310–320. [CrossRef]
- Jing, M.; Munyaka, P.M.; Tactacan, G.B.; Rodriguez-Lecompte, J.C.; O, K.; House, J.D. Performance, serum biochemical responses, and gene expression of intestinal folate transporters of young and older laying hens in response to dietary folic acid supplementation and challenge with Escherichia coli lipopolysaccharide. Poultry Science 2014, 93, 122–131. [CrossRef] [PubMed]
- Adetunji, C.O.; Adejumo, I.O. Potency of agricultural wastes in mushroom (Pleurotus sajor-caju) biotechnology for feeding broiler chicks (Arbor acre). International Journal of Recycling of Organic Waste in Agriculture 2018, 8, 37–45. [CrossRef]
- FAO. Outputs and activities of FAO Project FMM/RAS/298/MUL and summary of FAO’s recent work on antimicrobial resistance in aquaculture. FAO 2020.
- Hamasalim, H.J. Synbiotic as feed additives relating to animal health and performance. Advances in Microbiology 2016, 6, 288–302. [CrossRef]
- Wanson, K.S.; Gibson, G.R.; Hutkins, R.; Reimer, R.A.; Reid, G.; Verbeke, K.; Scott, K.P.; Holscher, H.D.; Azad, M.B.; Delzenne, N.M. The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of synbiotics. Nature Reviews Gastroenterology & Hepatology 2020, 17, 687–701.
- Mountzouris, K.C.; Tsitrsikos, P.; Palamidi, I.; Arvaniti, A.; Mohnl, M.; Schatzmayr, G.; Fegeros, K. Effects of probiotic inclusion levels in broiler nutrition on growth performance, nutrient digestibility, plasma immunoglobulins, and cecal microflora composition. Poultry Science 2010, 89, 58–67. [CrossRef]
- Sjofjan, O.; Natsir, M.H.; Adli, D.N.; Adelina, D.D.; Triana, L.M. Effect of symbiotic flour (Lactobacillus Sp. and FOS) to the egg quality and performance of laying hens. IOP Conference Series: Earth and Environmental Science 2020, 465, 012033.
- Lv, J.; Guo, L.; Chen, B.; Hao, K.; Ma, H.; Liu, Y.; Min, Y. Effects of different probiotic fermented feeds on production performance and intestinal health of laying hens. Poultry Science 2022, 101, 101570. [CrossRef]
- Khattab, A.A.; Basuini, M.F.M.E.; El-Ratel, I.T.; Fouda, S.F. Dietary probiotics as a strategy for improving growth performance, intestinal efficacy, immunity, and antioxidant capacity of white Pekin ducks fed with different levels of CP. Poultry Science 2021, 100, 100898. [CrossRef]
- Deng, Q.; Shi, H.; Luo, Y.; Liu, N.; Deng, X. Dietary lactic acid bacteria modulate yolk components and cholesterol metabolism by Hmgr pathway in laying hens. Brazilian Journal of Poultry Science 2020, 22, 1–8. [CrossRef]
- Adriani, L.; Latipudin, D.; Joni, I.M.; Panatarani, C.; Sania, G. Hematological status and egg production of laying hen with probiotic powder as feed supplements. IOP Conference Series: Earth and Environmental Science 2021, 902, 012032.
- Mookiah, S.; Sieo, C.C.; Ramasamy, K.; Abdullah, N.; Ho, Y.W. Effects of dietary prebiotics, probiotic, and synbiotics on performance, caecal bacterial populations, and caecal fermentation concentrations of broiler chickens. Journal of the Science of Food and Agriculture 2013, 94, 341–348. [CrossRef] [PubMed]
- Iji, P.A.; Tivey, D.R. Natural and synthetic oligosaccharides in broiler chicken diets. World’s Poultry Science Journal 1998, 54, 129–143.
- Yan, F.F.; Mohammed, A.A.; Murugesan, G.R.; Cheng, H.W. Effects of a dietary synbiotic inclusion on bone health in broilers subjected to cyclic heat stress episodes. Poultry Science 2019, 98, 1083–1089. [CrossRef]
- Dai, D.; Qi, G.; Wang, J.; Zhang, H.; Qiu, K.; Wu, S. Intestinal microbiota of layer hens and its association with egg quality and safety. Poultry Science 2022, 101, 102008. [CrossRef]
- Biggs, P.; Parsons, C.M.; Fahey, G.C. The effects of several oligosaccharides on growth performance, nutrient digestibilities, and cecal microbial populations in young chicks. Poultry Science 2007, 86, 2327–2336. [CrossRef] [PubMed]
- Śliżewska, K.; Markowiak-Kopeć, P.; Śbikowski, A.; Szeleszczuk, P. The effect of synbiotic preparations on the intestinal microbiota and her metabolism in broiler chickens. Scientific Reports 2020, 10, 1–13. [CrossRef] [PubMed]
- Lamora, Z.V.D.; Nuño, K.; Vázquez-Paulino, O.; Avalos, H.; Castro-Rosas, J.; Gómez-Aldapa, C.; Angulo, C.; Ascencio, F.; Villarruel-López, A. Effect of a synbiotic mix on intestinal structural changes and Salmonella Typhimurium and Clostridium Perfringens colonization in broiler chickens. Animals 2019, 9, 777. [CrossRef] [PubMed]
- Kiu, R.; Hall, L.J. An update on the human and animal enteric pathogen Clostridium Perfringens. Emerging Microbes & Infections 2018, 7, 1–15.
- Latorre, J.D.; Adhikari, B.; Park, S.H.; Teague, K.D.; Graham, L.E.; Mahaffey, B.D.; Baxter, M.F.A.; Hernandez-Velasco, X.; Kwon, Y.M.; Ricke, S.C. Evaluation of the epithelial barrier function and ileal microbiome in an established necrotic enteritis challenge model in broiler chickens. Frontiers in Veterinary Science 2018, 5, 1–11. [CrossRef] [PubMed]
- Bouassi, T.; Libanio, D.; Mesa, M.D.; Oke, O.E.; Gil, A.H.; Tona, K.; Ameyapoh, Y. Supplementation with liquid whey and ACIDAL® ML in drinking water affect gut pH and microflora and productive performance in laying hens. British Poultry Science 2020, 62, 138–146. [CrossRef] [PubMed]
- Kimminau, E.A.; Karnezos, T.P.; Berghaus, R.D.; Jones, M.K.; Baxter, J.A.; Hofacre, C.L. Combination of probiotic and prebiotic impacts Salmonella Enteritidis infection in layer hens. Journal of Applied Poultry Research 2021, 30, 100200. [CrossRef]
- Yaqoob, M.U.; El-Hack, M.E.; Hassan, F.; El-Saadony, M.T.; Khafaga, A.F.; Batiha, G.E.; Yehia, N.; Elnesr, S.S.; Alagawany, M.; El-Tarabilly, K.A. The potential mechanistic insights and future implications for the effect of prebiotics on poultry performance, gut microbiome, and intestinal morphology. Poultry Science 2021, 100, 101143. [CrossRef]
- Rostagno, H.S.; Albino, L.F.T.; Hannas, M.I.; et al. Tabelas Brasileiras para Aves e Suínos, 4th ed.; UFV/Departamento de Zootecnia: Viçosa, Brazil, 2017.
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing; Twenty-first Informational Supplement; CLSI: Wayne, PA, USA, 2014.
- Li, X.; Liu, L.; Li, K.; Hao, K.; Xu, C. Effect of fructooligosaccharides and antibiotics on laying performance of chickens and cholesterol content of egg yolk. British Poultry Science 2007, 48, 185–189. [CrossRef]
- Sugiharto, S.; Isroli, I.; Yudiarti, T.; Widiastuti, E. The effect of supplementation of multistrain probiotic preparation in combination with vitamins and minerals to the basal diet on the growth performance, carcass traits, and physiological response of broilers. Veterinary World 2018, 11, 240–247. [CrossRef] [PubMed]
- Chauhan, S.S.; Sharma, R.K.; Singh, D.V.; Shukla, S.K.; Palod, J.; Singh, M.K. Studies on serum mineral profile and kidney function of broiler chickens fed diets containing different supplements. Indian Journal of Animal Research 2020, 189–192. [CrossRef]
- Hatab, M.H.; Elsayed, M.A.; Ibrahim, N.S. Effect of some biological supplementation on productive performance, physiological, and immunological response of layer chicks. Journal of Radiation Research and Applied Sciences 2016, 9, 185–192. [CrossRef]
- Melillo, A. Applications of serum protein electrophoresis in exotic pet medicine. Veterinary Clinics of North America: Exotic Animal Practice 2013, 16, 211–225.
- Rashidi, N.; Khatibjoo, A.; Taherpour, K.; Akbari-Gharaei, M.; Shirzadi, H. Effects of licorice extract, probiotic, toxin binder, and poultry litter biochar on performance, immune function, blood indices, and liver histopathology of broilers exposed to aflatoxin-B1. Poultry Science 2020, 99, 5896–5906. [CrossRef]
- Xu, Q.; Azzam, M.M.M.; Zou, X.; Dong, X. Effects of chitooligosaccharide supplementation on laying performance, egg quality, blood biochemistry, antioxidant capacity, and immunity of laying hens during the late laying period. Italian Journal of Animal Science 2020, 19, 1180–1187. [CrossRef]
- Khabirov, A.; Khaziakhmetov, F.; Kuznetsov, V.; Tagirov, H.; Rebezov, M.; Andreyeva, A.; Basharov, A.; Yessimbekov, Z.; Ayaz, M. Effect of Normosil probiotic supplementation on the growth performance and blood parameters of broiler chickens. Indian Journal of Pharmaceutical Education and Research 2020, 54, 1046–1055. [CrossRef]
- Ndrepepa, G. Aspartate aminotransferase and cardiovascular disease—a narrative review. Journal of Laboratory and Precision Medicine 2021, 6, 6. [CrossRef]
- Thrall, A.M. Hematologia e Bioquímica Clínica Veterinária, 2nd ed.; Guanabara Koogan: Rio de Janeiro, Brazil, 2015.
- González, F.; Silva, S.C. Perfil Bioquímico Sanguíneo: Introdução à Bioquímica Clínica Veterinária; [n.p.]: Rio Grande do Sul, Brazil, 2006.
- Tang, S.G.H.; Sieo, C.C.; Ramasamy, K.; Saad, W.Z.; Wong, H.K.; Ho, Y.W. Performance, biochemical, and haematological responses, and relative organ weights of laying hens fed diets supplemented with prebiotic, probiotic, and synbiotic. BMC Veterinary Research 2017, 13, 1–12. [CrossRef]
- Attia, Y.; Hamed, R.; El-Hamid, A.; Al-Harthi, M.; Shahba, H.; Bovera, F. Performance, blood profile, carcass and meat traits, and tissue morphology in growing rabbits fed mannanoligosaccharides and zinc-bacitracin continuously or intermittently. Animal Science Papers and Reports 2015, 33, 85–101.
- Michalska, K.; Gesek, M.; Sokoł, R.; Murawska, D.; Mikiewicz, M.; Chłodowska, A. Effective microorganisms (EM) improve internal organ morphology, intestinal morphometry, and serum biochemical activity in Japanese quails under Clostridium Perfringens challenge. Molecules 2021, 26, 2786. [CrossRef]
- Alvarenga, R.R.; Zangeronimo, M.G.; Pereira, L.J.; Rodrigues, P.B.; Gomide, E.M. Lipoprotein metabolism in poultry. World’s Poultry Science Journal 2011, 67, 431–440.
- Gilliland, S.E.; Nelson, C.R.; Maxwell, C. Assimilation of cholesterol by Lactobacillus acidophilus. Applied and Environmental Microbiology 1985, 49, 377–381. [CrossRef]
- Fukushima, M.; Nakano, M. The effect of a probiotic on faecal and liver lipid classes in rats. British Journal of Nutrition 1995, 73, 701–710. [CrossRef] [PubMed]
- Ooi, L.G.; Liong, M.T. Cholesterol-lowering effects of probiotics and prebiotics: A review of in vivo and in vitro findings. International Journal of Molecular Sciences 2010, 11, 2499–2522. [CrossRef]
- Ogboko, B. Lipid profile of broilers fed zinc bacitracin on plant and animal protein diets. International Journal of Poultry Science 2011, 10, 567–573. [CrossRef]
- Reuben, R.C.; Sarkar, S.L.; Ibnat, H.; Setu, M.A.; Roy, P.C.; Jahid, I.K. Novel multi-strain probiotics reduces Pasteurella multocida induced fowl cholera mortality in broilers. Scientific Reports 2021, 11, 1–16. [CrossRef]
- Wang, W.; Yang, H.; Wang, Z.; Han, J.; Zhang, D.; Sun, H.; Zhang, F. Effects of prebiotic supplementation on growth performance, slaughter performance, growth of internal organs and small intestine, and serum biochemical parameters of broilers. Journal of Applied Animal Research 2014, 43, 33–38. [CrossRef]
- Yu, W.; Hao, X.; Zhiyue, W.; Haiming, Y.; Lei, X. Evaluation of the effect of Bacillus subtilis and Pediococcus acidilactici mix on serum biochemistry growth, promotion of body, and visceral organs in Lohmann Brown chicks. Brazilian Journal of Poultry Science 2020, 22, 1–8. [CrossRef]
- Sharma, P.; Bhardwaj, P.; Singh, R. Administration of Lactobacillus casei and Bifidobacterium bifidum ameliorated hyperglycemia, dyslipidemia, and oxidative stress in diabetic rats. International Journal of Preventive Medicine 2016, 7, 102.
- Abdelqader, A.; Al-Fataftah, A.R.; Daş, G. Effects of dietary Bacillus subtilis and inulin supplementation on performance, eggshell quality, intestinal morphology, and microflora composition of laying hens in the late phase of production. Animal Feed Science and Technology 2013, 179, 103–111. [CrossRef]
- Cummings, J.H.; Macfarlane, G.T. Gastrointestinal effects of prebiotics. British Journal of Nutrition 2002, 87, S145–S151. [CrossRef]
- Lone, A.; Mottawea, W.; Chait, Y.A.; Hammami, R. Dual inhibition of Salmonella enterica and Clostridium perfringens by new probiotic candidates isolated from chicken intestinal mucosa. Microorganisms 2021, 9, 166. [CrossRef]
- Pal, C.; Bengtsson-Palme, J.; Kristiansson, E.; Larsson, D.G.J. The structure and diversity of human, animal, and environmental resistomes. Microbiome 2016, 4, 1–15. [CrossRef] [PubMed]
- Roth, N.; Hofacre, C.; Zitz, U.; Mathis, G.F.; Moder, K.; Doupovec, B.; Berghouse, R.; Domig, K.J. Prevalence of antibiotic-resistant E. coli in broilers challenged with a multi-resistant E. coli strain and received ampicillin, an organic acid-based feed additive, or a synbiotic preparation. Poultry Science 2019, 98, 2598–2607.
- Achakzai, R.; Taj, M.K.; Achakzai, K.B. Microbiological studies on Clostridium perfringens isolated from commercial poultry of Balochistan. Asian Journal of Biological and Life Sciences 2020, 9, 204–208. [CrossRef]
- Li, J.; Zhou, Y.; Yang, D.; Zhang, S.; Sun, Z.; Wang, Y.; Wang, S.; Wu, C. Prevalence and antimicrobial susceptibility of Clostridium perfringens in chickens and pigs from Beijing and Shanxi, China. Veterinary Microbiology 2021, 252, 108932. [CrossRef]
- Xu, W.; Wang, H.; Liu, L.; Miao, Z.; Huo, Y.; Zhong, Z. Prevalence and characterization of Clostridium perfringens isolated from different chicken farms in China. Anaerobe 2021, 72, 102467. [CrossRef]
- Sáenz, J.S.; Marques, T.V.; Barone, R.S.C.; Cyrino, J.E.P.; Kublik, S.; Nesme, J.; Schloter, M.; Rath, S.; Vestergaard, G. Oral administration of antibiotics increased the potential mobility of bacterial resistance genes in the gut of the fish Piaractus mesopotamicus. Microbiome 2019, 7, 1–14. [CrossRef] [PubMed]
- Van Immerseel, F.; De Buck, J.; Pasmans, F.; Huyghebaert, G.; Haesebrouck, F.; Ducatelle, R. Clostridium perfringens in poultry: An emerging threat for animal and public health. Avian Pathology 2004, 33, 537–549. [CrossRef]
- Khademi, F.; Sahebkar, A. The prevalence of antibiotic-resistant Clostridium species in Iran: A meta-analysis. Pathogens and Global Health 2019, 113, 58–66. [CrossRef]
- Mwangi, S.; Timmons, J.; Fitz-Coy, S.; Parveen, S. Characterization of Clostridium perfringens recovered from broiler chicken affected by necrotic enteritis. Poultry Science 2019, 98, 128–135. [CrossRef]
- Jang, Y.-S.; Kim, D.-H.; Bae, D.; Kim, S.-H.; Kim, H.; Moon, J.-S.; Song, K.-Y.; Chon, J.-W.; Seo, K.-H. Prevalence, toxin-typing, and antimicrobial susceptibility of Clostridium perfringens from retail meats in Seoul, Korea. Anaerobe 2020, 64, 102235. [CrossRef] [PubMed]
- Silva, R.O.S.; Salvarani, F.M.; Assis, R.A.; Martins, N.R.S.; Pires, P.S.; Lobato, F.C.F. Antimicrobial susceptibility of Clostridium perfringens strains isolated from broiler chickens. Brazilian Journal of Microbiology 2009, 40, 262–264. [CrossRef] [PubMed]
- Adams, V.; Han, X.; Lyras, D.; Rood, J.I. Antibiotic resistance plasmids and mobile genetic elements of Clostridium perfringens. Plasmid 2018, 99, 32–39. [CrossRef] [PubMed]
- Han, X.; Du, X.-D.; Southey, L.; Bulach, D.M.; Seemann, T.; Yan, X.-X.; Bannam, T.L.; Rood, J.I. Functional analysis of a bacitracin resistance determinant located on ICE Cp1, a novel Tn916-like element from a conjugative plasmid in Clostridium perfringens. Antimicrobial Agents and Chemotherapy 2015, 59, 6855–6865. [CrossRef]
Figure 1.
Average counts of C. perfringens in the cecal contents of laying hens subjected to long-term supplementation with zinc bacitracin or a symbiotic compound included in the diet at different rearing phases. MBM: corn, soybean meal, and meat and bone meal diet from 1 day to 90 weeks; BAC: corn, soybean meal, and meat and bone meal diet with 0.05% zinc bacitracin from 1 day to 90 weeks; SIMC: corn, soybean meal, and meat and bone meal diet with 0.1% symbiotic from 1 day to 90 weeks; SIMR: corn, soybean meal, and meat and bone meal diet with 0.1% symbiotic from 6 to 90 weeks; SIMP: corn, soybean meal, and meat and bone meal diet with 0.1% symbiotic from 17 to 90 weeks. a,b Within a column, values with different letters differ significantly according to SNK’s test (P < 0.05).
Figure 1.
Average counts of C. perfringens in the cecal contents of laying hens subjected to long-term supplementation with zinc bacitracin or a symbiotic compound included in the diet at different rearing phases. MBM: corn, soybean meal, and meat and bone meal diet from 1 day to 90 weeks; BAC: corn, soybean meal, and meat and bone meal diet with 0.05% zinc bacitracin from 1 day to 90 weeks; SIMC: corn, soybean meal, and meat and bone meal diet with 0.1% symbiotic from 1 day to 90 weeks; SIMR: corn, soybean meal, and meat and bone meal diet with 0.1% symbiotic from 6 to 90 weeks; SIMP: corn, soybean meal, and meat and bone meal diet with 0.1% symbiotic from 17 to 90 weeks. a,b Within a column, values with different letters differ significantly according to SNK’s test (P < 0.05).

Figure 2.
Cecal C. perfringens resistance to ampicillin (AMP), ciprofloxacin (CIP), erythromycin (ERI), gentamicin (GEN), neomycin (NEO), lincomycin (LIN), tetracycline (TET), and bacitracin (BAC) in laying hens subjected to long-term supplementation with zinc bacitracin or a symbiotic compound included in the diet at different rearing phases. Resistant (R), susceptible (S), and intermediate (I). MBM: corn, soybean meal, and meat and bone meal diet from 1 day to 90 weeks; BAC: corn, soybean meal, and meat and bone meal diet with 0.05% zinc bacitracin from 1 day to 90 weeks; SIMC: corn, soybean meal, and meat and bone meal diet with 0.1% symbiotic from 1 day to 90 weeks; SIMR: corn, soybean meal, and meat and bone meal diet with 0.1% symbiotic from 6 to 90 weeks; SIMP: corn, soybean meal, and meat and bone meal diet with 0.1% symbiotic from 17 to 90 weeks.
Figure 2.
Cecal C. perfringens resistance to ampicillin (AMP), ciprofloxacin (CIP), erythromycin (ERI), gentamicin (GEN), neomycin (NEO), lincomycin (LIN), tetracycline (TET), and bacitracin (BAC) in laying hens subjected to long-term supplementation with zinc bacitracin or a symbiotic compound included in the diet at different rearing phases. Resistant (R), susceptible (S), and intermediate (I). MBM: corn, soybean meal, and meat and bone meal diet from 1 day to 90 weeks; BAC: corn, soybean meal, and meat and bone meal diet with 0.05% zinc bacitracin from 1 day to 90 weeks; SIMC: corn, soybean meal, and meat and bone meal diet with 0.1% symbiotic from 1 day to 90 weeks; SIMR: corn, soybean meal, and meat and bone meal diet with 0.1% symbiotic from 6 to 90 weeks; SIMP: corn, soybean meal, and meat and bone meal diet with 0.1% symbiotic from 17 to 90 weeks.
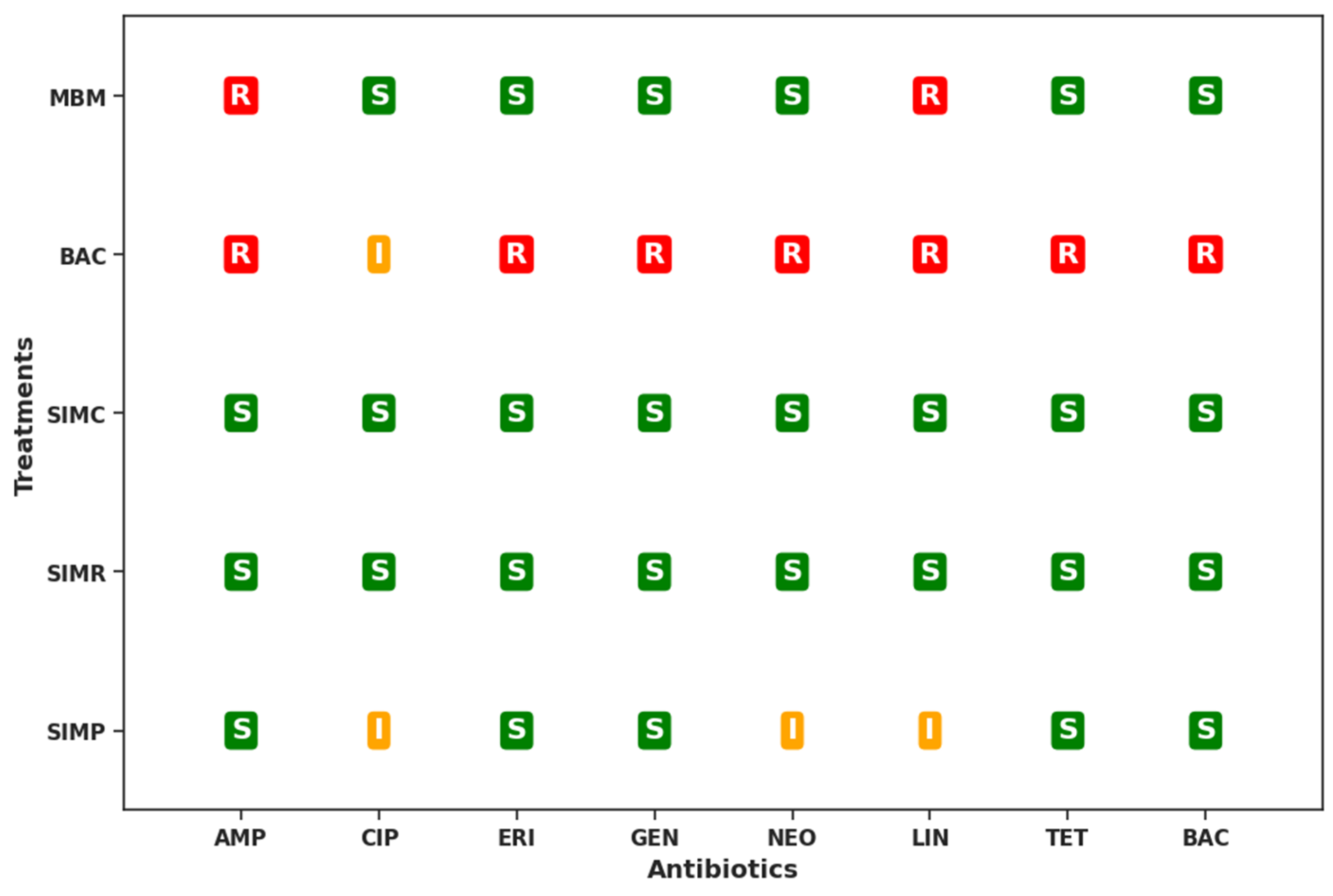

Table 1.
Composition of the experimental diet on starter e growth phase.
| Ingredients | Starter | Grower I | Grower II | ||||||
|---|---|---|---|---|---|---|---|---|---|
| MBM | BAC | SIMC | MBM | BAC | SIMR | MBM | BAC | SIMR | |
| Corn | 59.4 | 59.4 | 59.4 | 63.83 | 63.83 | 63.83 | 65.06 | 65.06 | 65.06 |
| Soybean Meal, 46% | 33.8 | 33.8 | 33.8 | 27.04 | 27.04 | 27.04 | 23.85 | 23.85 | 23.85 |
| Meat and Bone Meal | 2.51 | 2.51 | 2.51 | 2.51 | 2.51 | 2.51 | 2.01 | 2.01 | 2.01 |
| Soybean Oil | 0.54 | 0.54 | 0.54 | ---- | ---- | ---- | ---- | ---- | ---- |
| Limestone | 0.80 | 0.80 | 0.80 | 0.81 | 0.81 | 0.81 | 1.15 | 1.15 | 1.15 |
| Salt | 0.17 | 0.17 | 0.17 | 0.15 | 0.15 | 0.15 | 0.13 | 0.13 | 0.13 |
| Sodium bicarbonate | 0.15 | 0.15 | 0.15 | 0.15 | 0.15 | 0.15 | 0.15 | 0.15 | 0.15 |
| Vitamin Premix1 | 0.15 | 0.15 | 0.15 | 0.15 | 0.15 | 0.15 | 0.15 | 0.15 | 0.15 |
| Mineral Premix2 | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 |
| Zinc Bacitracin | ---- | 0.05 | ---- | ---- | 0.05 | ---- | ---- | 0.05 | ---- |
| Symbiotic | ---- | ---- | 1,0 | ---- | ---- | 1,0 | ---- | ---- | 1.0 |
| DL-Methionine 99% | 0.25 | 0.25 | 0.25 | 0.25 | 0.25 | 0.25 | 0.089 | 0.089 | 0.089 |
| L-lysine HCl, 78.8% | 0.19 | 0.19 | 0.19 | 0.19 | 0.19 | 0.19 | 0.02 | 0.02 | 0.02 |
| L-Threonine 98.5% | 0.02 | 0.02 | 0.02 | 0.01 | 0.01 | 0.01 | ---- | ---- | ---- |
| Phytase | 0.006 | 0.006 | 0.006 | 0.006 | 0.006 | 0.006 | 0.006 | 0.006 | 0.006 |
| Inert | 2 | 2 | 2 | 4.85 | 4.85 | 4.85 | 9.24 | 9.24 | 9.24 |
| Total | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| Calculated Nutritional Composition (%) | |||||||||
| ME (kcal/kg) | 2950 | 2950 | 2950 | 2900 | 2900 | 2900 | 2800 | 2800 | 2800 |
| Crude Protein | 21.4 | 21.4 | 21.4 | 18.7 | 18.7 | 18.7 | 16.0 | 16.0 | 16.0 |
| Calcium | 0.97 | 0.97 | 0.97 | 0.95 | 0.95 | 0.95 | 1.0 | 1.0 | 1.0 |
| Available Phosphorus | 0.45 | 0.45 | 0.45 | 0.44 | 0.44 | 0.44 | 0.40 | 0.40 | 0.40 |
| Sodium | 0.18 | 0.18 | 0.18 | 0.17 | 0.17 | 0.17 | 0.16 | 0.16 | 0.16 |
| Chlorine | 0.19 | 0.19 | 0.19 | 0.17 | 0.17 | 0.17 | 0.16 | 0.16 | 0.16 |
| Potassium | 0.82 | 0.82 | 0.82 | 0.71 | 0.71 | 0.71 | 0.62 | 0.62 | 0.62 |
| Digestible amino acids (%) | |||||||||
| Methionine + Cystine | 0.86 | 0.86 | 0.86 | 0.80 | 0.80 | 0.80 | 0.58 | 0.58 | 0.58 |
| Methionine | 0.54 | 0.54 | 0.54 | 0.50 | 0.50 | 0.50 | 0.32 | 0.32 | 0.32 |
| Lysine | 1.16 | 1.16 | 1.16 | 1.00 | 1.00 | 1.00 | 0.73 | 0.73 | 0.73 |
| Threonine | 0.78 | 0.78 | 0.78 | 0.68 | 0.68 | 0.68 | 0,58 | 0,58 | 0,58 |
| Tryptophan | 0.26 | 0.26 | 0.26 | 0.23 | 0.23 | 0.23 | 0.19 | 0.19 | 0.19 |
1Vitamin Premix (supplies per kilogram of product): vit. D3, 2,500,000.00 IU; vit. A, 9,000.00 IU; vit.; vit. And, 20,000.00 IU; vit. K3 (Menadione) 2500.00 mg; vit. B1 (Thiamine) 2000.00 mg; B2 (Riboflavin) 6,000.00 mg; B6 (Pyridoxine) 3000.38 mg; B12 Cobalamin) 15,000.00 mg; Niacin (Ac. Nicotinico) 35,000.00 mg; Pantothenic Acid, 12,000,000 mg; Folic Acid, 1,500.00 mg; Selenium, 250.00 mg; Biotin, 100,000 mg. 2Premix Mineral (provides per kilogram of product): Copper, 20,000,000 mg; Iron, 100,000,000 mg; Manganese, 130,000,000 mg; Iodine, 2000.00 mg; Zinc, 130,000,000 mg. 3Phytase: 10,000 FTU/g.
Table 2.
Composition of the experimental diet on pre-laying phase and production phase.
| Ingredients | Pre-laying phase | Peak phase | Post-peak phase | ||||||
|---|---|---|---|---|---|---|---|---|---|
| MBM | BAC | SIMP | MBM | BAC | SIMP | MBM | BAC | SIMP | |
| Corn | 65.17 | 65.17 | 65.17 | 60.2 | 60.2 | 60.2 | 60.1 | 60.1 | 60.1 |
| Soybean Meal, 46% | 21.99 | 21.99 | 21.99 | 24.5 | 24.5 | 24.5 | 22.9 | 22.9 | 22.9 |
| Meat and Bone Meal | 2.54 | 2.54 | 2.54 | 1.41 | 1.41 | 1.41 | 1.49 | 1.49 | 1.49 |
| Soybean Oil | 2.34 | 2.34 | 2.34 | 1.0 | 1.0 | 1.0 | 1.05 | 1.05 | 1.05 |
| Limestone | 4.14 | 4.14 | 4.14 | 10.4 | 10.4 | 10.4 | 10.6 | 10.6 | 10.6 |
| Salt | 0.15 | 0.15 | 0.15 | 0.26 | 0.26 | 0.26 | 0.25 | 0.25 | 0.25 |
| Sodium Bicarbonate | 0.15 | 0.15 | 0.15 | 0.15 | 0.15 | 0.15 | 0.15 | 0.15 | 0.15 |
| Vitamin Premix | 0.15 | 0.15 | 0.15 | 0.15 | 0.15 | 0.15 | 0.15 | 0.15 | 0.15 |
| Mineral Premix | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 |
| Zinc Bacitracin | ---- | 0.05 | 0.05 | ---- | 0.05 | ---- | ---- | 0.05 | ---- |
| Symbiotic | ---- | ---- | 1.0 | ---- | ---- | 1.0 | ---- | ---- | 1.0 |
| DL-Methionine 99% | 0.19 | 0.19 | 0.19 | 0.27 | 0.27 | 0.27 | 0.26 | 0.26 | 0.26 |
| L-lysine HCl, 78.8% | 0.15 | 0.15 | 0.15 | 0.04 | 0.04 | 0.04 | 0.05 | 0.05 | 0.05 |
| L-Threonine 98.5% | 0.014 | 0.014 | 0.014 | ---- | ---- | ---- | ---- | ---- | ---- |
| Phytase | 0.006 | 0.006 | 0.006 | 0.006 | 0.006 | 0.006 | 0.006 | 0.006 | 0.006 |
| Inert | 2.92 | 2.92 | 2.92 | 1.62 | 1.62 | 1.62 | 2.73 | 2.73 | 2.73 |
| Total | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| Calculated Nutritional Composition (%) | |||||||||
| ME (kcal/kg) | 2799 | 2799 | 2799 | 2780 | 2780 | 2780 | 2750 | 2750 | 2750 |
| Crude Protein | 16.50 | 16.50 | 16.50 | 16.7 | 16.7 | 16.7 | 15.89 | 15.89 | 15.89 |
| Calcium | 2.2 | 2.2 | 2.2 | 4.4 | 4.4 | 4.4 | 4.5 | 4.5 | 4.5 |
| Available Phosphorus | 0.44 | 0.44 | 0.44 | 0.37 | 0.37 | 0.37 | 0.37 | 0.37 | 0.37 |
| Sodium | 0.18 | 0.18 | 0.18 | 0.21 | 0.21 | 0.21 | 0.20 | 0.20 | 0.20 |
| Chlorine | 0.15 | 0.15 | 0.15 | 0.23 | 0.23 | 0.23 | 0.22 | 0.22 | 0.22 |
| Potassium | 1.04 | 1.04 | 1.04 | 0.65 | 0.65 | 0.65 | 0.62 | 0.62 | 0.62 |
| Digestible amino acids (%) | |||||||||
| Methionine + Cystine | 0.68 | 0.68 | 0.68 | 0.77 | 0.77 | 0.77 | 0.74 | 0.74 | 0.74 |
| Methionine | 0.41 | 0.41 | 0.41 | 0.50 | 0.50 | 0.50 | 0.48 | 0.48 | 0.48 |
| Lysine | 0.83 | 0.83 | 0.83 | 0.79 | 0.79 | 0.79 | 0.76 | 0.76 | 0.76 |
| Threonine | 0.57 | 0.57 | 0.57 | 0.61 | 0.61 | 0.61 | 0.59 | 0.59 | 0.59 |
| Tryptophan | 0.18 | 0.18 | 0.18 | 0.20 | 0.20 | 0.20 | 0.20 | 0.20 | 0.20 |
1Vitamin Premix (supplies per kilogram of product): vit. D3, 2,500,000.00 IU; vit. A, 9,000.00 IU; vit.; vit. And, 20,000.00 IU; vit. K3 (Menadione) 2500.00 mg; vit. B1 (Thiamine) 2000.00 mg; B2 (Riboflavin) 6,000.00 mg; B6 (Pyridoxine) 3000.38 mg; B12 Cobalamin) 15,000.00 mg; Niacin (Ac. Nicotinico) 35,000.00 mg; Pantothenic Acid, 12,000,000 mg; Folic Acid, 1,500.00 mg; Selenium, 250.00 mg; Biotin, 100,000 mg. 2Premix Mineral (provides per kilogram of product): Copper, 20,000,000 mg; Iron, 100,000,000 mg; Manganese, 130,000,000 mg; Iodine, 2000.00 mg; Zinc, 130,000,000 mg. 3Phytase: 10,000 FTU/g.
Table 3.
Average values of relative liver weight (LIV), uric Acid (URI), total proteins (TP), creatine Kinase (CK), aspartate aminotransferase (AST), alanine aminotransferase (ALT), Lactate Dehydrogenase (LDH), and Phosphorus (PHOS) in the serum of laying hens subjected to long-term supplementation with zinc bacitracin or a symbiotic compound included in the diet at different rearing phases.
Table 3.
Average values of relative liver weight (LIV), uric Acid (URI), total proteins (TP), creatine Kinase (CK), aspartate aminotransferase (AST), alanine aminotransferase (ALT), Lactate Dehydrogenase (LDH), and Phosphorus (PHOS) in the serum of laying hens subjected to long-term supplementation with zinc bacitracin or a symbiotic compound included in the diet at different rearing phases.
| Treatments | Liv (%) | URI | TP | CK | AST | ALT | LDH | Phos |
| (mg/dL) | (mg/dL) | (UI/L) | (UI/L) | (UI/L) | (UI/L) | (mg/dL) | ||
| MBM | 2.64ab | 5.45a | 6.21ab | 1195 | 197b | 18b | 1154a | 6.95b |
| BAC | 2.30b | 6.54a | 6.77a | 1154 | 253a | 37a | 873b | 11.0a |
| SIMC | 2.51ab | 3.68b | 5.40bc | 1259 | 171ab | 14c | 764b | 5.52b |
| SIMR | 2.88a | 3.59b | 5.32bc | 1194 | 173ab | 11d | 749b | 5.18b |
| SIMP | 2.77ab | 2.77b | 4.93c | 1106 | 147b | 11d | 643b | 6.22b |
| Mean | 2.62 | 4.41 | 5.72 | 1181 | 188 | 18 | 836 | 7.06b |
| P-value | 0.049 | 0.001 | 0.001 | 0.953 | 0.001 | 0.001 | 0.009 | 0.017 |
| SEM | 0.10 | 0.69 | 0.33 | 25 | 18 | 4.85 | 87 | 1.14 |
SEM = Standard Error of the Mean. MBM: corn, soybean meal, and meat and bone meal diet from 1 day to 90 weeks; BAC: corn, soybean meal, and meat and bone meal diet with 0.05% zinc bacitracin from 1 day to 90 weeks; SIMC: corn, soybean meal, and meat and bone meal diet with 0.1% symbiotic from 1 day to 90 weeks; SIMR: corn, soybean meal, and meat and bone meal diet with 0.1% symbiotic from 6 to 90 weeks; SIMP: corn, soybean meal, and meat and bone meal diet with 0.1% symbiotic from 17 to 90 weeks. a,b Within a column, values with different letters differ significantly according to SNK’s test (P < 0.05).
Table 4.
Average values of High-Density Lipoprotein (HDL), Low-Density Lipoprotein (LDL), Triglycerides (TRI), and Total Cholesterol (COL) in the serum of laying hens subjected to long-term supplementation with zinc bacitracin or a symbiotic compound included in the diet at different rearing phases.
Table 4.
Average values of High-Density Lipoprotein (HDL), Low-Density Lipoprotein (LDL), Triglycerides (TRI), and Total Cholesterol (COL) in the serum of laying hens subjected to long-term supplementation with zinc bacitracin or a symbiotic compound included in the diet at different rearing phases.
| Treatments | HDL | LDL | TRI | COL |
|---|---|---|---|---|
| (mg/dL) | (mg/dL) | (mg/dL) | (mg/dL) | |
| MBM | 32.3c | 18.5 | 1004a | 119 |
| BAC | 24.2d | 35.8 | 989ab | 171 |
| SIMC | 46.9b | 7.96 | 977b | 144 |
| SIMR | 45.7b | 11.9 | 989ab | 137 |
| SIMP | 61.6a | 11.1 | 984b | 173 |
| Média | 42 | 17 | 989 | 149 |
| P-value | <0.001 | 0.649 | 0.031 | 0.121 |
| SEM | 6.46 | 4.99 | 4.62 | 10 |
MBM: corn, soybean meal, and meat and bone meal diet from 1 day to 90 weeks; BAC: corn, soybean meal, and meat and bone meal diet with 0.05% zinc bacitracin from 1 day to 90 weeks; SIMC: corn, soybean meal, and meat and bone meal diet with 0.1% symbiotic from 1 day to 90 weeks; SIMR: corn, soybean meal, and meat and bone meal diet with 0.1% symbiotic from 6 to 90 weeks; SIMP: corn, soybean meal, and meat and bone meal diet with 0.1% symbiotic from 17 to 90 weeks. a,b Within a column, values with different letters differ significantly according to SNK’s test (P < 0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



