
Submitted:
18 September 2024
Posted:
20 September 2024
You are already at the latest version
Abstract
An untrained couch potato involuntarily forced to undergo a long strenuous exercise towards exhaustion will feel it at the end as a discomforting fatigue. A well-trained athlete will feel the same but after a longer time. Patients of various neurological diseases will do so much earlier. All may also finally experience muscle pain and soreness the next day.Muscle fatigue is an exercise-induced reduction in maximal voluntary muscle force. Muscle fatigability varies with muscle use, age and sex. It is commonly divided into two broad categories: peripheral and central fatigue. The first refers to the many processes in the fatiguing muscle(s), the second to subsequent processes in the central nervous system (CNS). Peripheral muscle fatigue can be caused by numerous different mechanisms, ranging from the accumulation of metabolites within muscle fibers to their damage. Central fatigue involves the inadequacy of the CNS to generate and maintain sufficiently strong motor command. There must of course be a mediator relating the two fatigue types, and this is the collection of small-diameter group III (Aδ) and IV (C) nerve fibers emanating from the muscle and distributing their activation effects throughout the CNS.Muscle fatigue develops during eventually exhausting exercise that goes along with activations of the cardio-vascular and respiratory system to supply oxygen and energy resources. While these functions are supported by a number of sensory afferents, fatigue-activated group III/IV muscle afferents have been proposed to contribute their share to cardio-vascular and ventilatory reflex responses that are mediated in the brainstem. Neural feedback from working skeletal muscle is a vital component in providing a high capacity for endurance exercise because muscle perfusion and O₂ delivery determine the fatigability of skeletal muscle (Amann 2012).The development of chronic muscle fatigue is related to muscle wasting mediated by aging, myopathies, muscle dystrophies, immobilization, insulin resistance, diseases associated with systemic inflammation (arthritis, sepsis, infections), trauma, cardio-vascular (heart failure) and respiratory disorders (chronic obstructive pulmonary disease (COPD)), chronic kidney failure, multiple sclerosis (MS), and, more recently, coronavirus disease 2019 (COVID-19).
Keywords:
Muscle Fatigue
; Peripheral and Central Fatigue
; Neurological Diseases
; Group III/IV Muscle Afferents
| CONTENTS | |
| Abstract | |
| 1 | Introduction |
| 2 | Peripheral Processes in Muscle Fatigue |
| 2.1 | Peripheral Muscle Fatigue in Health |
| 2.1.1 | Metabolic Factors in Muscle Fatigue |
| 2.1.2 | Disturbances in Excitation-contraction Coupling |
| 2.1.3 | Muscle Fatigue and Damage |
| 2.1.4 | Recovery from Muscle Fatigue |
| 2.2 | Peripheral Muscle Fatigue in Disease |
| 2.2.1 | Overview |
| 2.2.2 | Old Age |
| 2.2.3 | Myopathies |
| 2.2.4 | Disturbances in Excitation-contraction Coupling |
| 2.2.5 | Neurological Diseases |
| 3 | Central Processes in Muscle Fatigue |
| 3.1 | Central Fatigue |
| 3.1.1 | Muscle Wisdom |
| 3.1.2 | Potential Mechanisms |
| 3.2 | Spinal Muscle Fatigue in Health |
| 3.2.1 | Motor Unit (MU) Properties as a First Line of Defense |
| 3.2.2 | Neuronal Sensors of Muscle Fatigue |
| 3.2.2.1 | Fatigue-related Changes in Firing of Muscle Spindle Afferents |
| 3.2.2.2 | Fatigue-related Changes in Firing of GTO Afferents |
| 3.2.2.3 | Group III/IV Muscle Afferents |
| 3.2.3 | Spinal Fatigue-related Reflexes |
| 3.2.3.1 | Presynaptic Inhibition (PSI) |
| 3.2.3.2 | Monosynaptic Group Ia Afferent Excitation |
| 3.2.3.3 | Heteronymous Group Ib Afferent Inhibition |
| 3.2.3.4 | Recurrent Inhibition |
| 3.2.3.5 | Reciprocal Inhibition |
| 3.3 | Supraspinal Muscle Fatigue |
| 3.3.1 | Fatigue-induced Effects in Supraspinal Structures |
| 3.3.1.1 | Medulla Oblongata |
| 3.3.1.2 | Peri-aqueductal Gray (PAG) |
| 3.3.1.3 | Amygdala |
| 3.3.1.4 | Blood-Pressure and Respiratory Control in Medulla and Amygdala |
| 3.3.1.5 | Hypothalamus |
| 3.3.1.6 | Cerebellum |
| 3.3.1.7 | Cerebral Cortex |
| 3.3.1.8 | Cortico-cerebello-basal ganglia-thalamic System |
| 3.3.2 | A Long Way Down: From Cerebral Cortex to Spinal Cord |
| 3.3.2.1 | Insufficient Drive from Motor Cortex |
| 3.3.2.2 | Substantia Nigra Pars Compacta and Dopamine (DA) |
| 3.3.2.3 | Raphé Nuclei and Serotonin (5-HT) |
| 3.3.2.4 | Locus Coeruleus and Noradrenaline (NA) |
| 3.4 | Central Muscle Fatigue in Disease |
| 3.4.1 | Overview |
| 3.4.2 | Age |
| 3.4.3 | Motoneuron Diseases |
| 3.4.3.1 | Amyotrophic Lateral Sclerosis (ALS) |
| 3.4.3.2 | Spinal Muscle Atrophy (SMA) |
| 3.4.3.3 | Post-Polio Syndrome (PPS) |
| 3.4.4 | Spinal Cord Injury (SCI) |
| 3.4.5 | Stroke and spasticity |
| 3.4.6 | Parkinson’s Disease (PD) |
| 3.4.7 | Multiple Sclerosis (MS) |
| 3.4.8 | Guillain-Barré Syndrome (GBS) |
| 3.4.9 | Fibromyalgia and Chronic Fatigue Syndrome (CFS) |
| 3.4.10 | COVID-19 Neuromuscular Symptoms |
| 3.4.11 | Yawning-Fatigue Syndrome |
| 3.4.12 | Muscle Fatigue and Pain |
| 4 | Conclusions |
“... furthermore, incessant driving of the pen over paper causes intense fatigue of the hand and the whole arm, and because of this continuous and almost tonic stress of the muscles and tendons, which in the course of time results in failure of power of the right hand ...“ (Ramazzini 1713).
1. Introduction
“Exercise and its troubles” (Windhorst 2003).
From common experience, muscle fatigue appears to be a phenomenon that can easily defined as loss of muscle force, decreasing endurance, increasing shortness of breath, heartbeat rate and blood pressure. Looking below the cover for underlying mechanisms turns out to be mind-boggling.
What is fatigue? Fatigue is a term that is much used in various contexts. For scientific purposes, it must therefore be defined properly, although this effort has resulted in several definitions. A first division is between perceptual or mental fatigue and physical fatigue. Physical fatigue in turn can be classified as peripheral or central (spinal or supraspinal) fatigue (Tanaka and Watanabe 2012).
Muscle fatigue is a phenomenon of great practical importance in both health and disease. In normal individuals, it is of particular importance in physically demanding occupations and in athletic competitions. Muscle fatigue is influenced by age, gender, physical condition, type of food, latency to last meal, mental status, psychological conditions, personality type, life experience, and the health status of an individual (Finsterer and Mahjoub 2014). Sustained muscle contraction above a force level of ca 25% maximal force entails fatigue. During such a contraction, the muscle can maintain its force for some time, despite a decline in maximal muscle-fiber force, due to an increase in neural drive to the motor units (MUs) as reflected in increased electromyographic (EMG) activity.
Local or whole-body physical exercise has implications for the equilibrium of body’s internal environment. The contracting muscles generate force or power, metabolites and heat. This generated energy reduces the energy stocks within the body. Depending on the form of exercise, sooner or later sensations of fatigue and exhaustion will occur. Their role is the protection of the exercising subject from the deleterious effects of excessive exercise. The exercise intensity, exercise endurance time and type of exercise determine the different effects (Ament and Verkerke 2009).
Muscle fatigue is commonly divided into two broad categories: peripheral and central fatigue. The first refers to the many processes in the fatiguing muscle(s), the second to subsequent processes in the CNS. Peripheral muscle fatigue can be caused by many different mechanisms, ranging from the accumulation of metabolites within muscle fibers to their damage. Central fatigue involves the inadequacy of the CNS to generate and maintain sufficiently strong motor command (Gandevia 2001; Taylor et al. 2016). There must of course be a mediator relating the two fatigue types, and this is the collection of small-diameter group III (Aδ) and IV (C) nerve fibers emanating from the muscle and distributing their activation effects throughout the CNS.
It might appear desirable that the CNS find ways to at least mitigate the effects of otherwise unavoidable loss of force and power – and the danger of muscle damage. Group III/IV muscle afferents facilitate `central fatigue’, this term referring to an insufficiency of the CNS to `drive’ motoneurons (MNs) so as to generate maximal force outputs (Gandevia 2001; Kent et al. 2016; Taylor et al. 2016).
The mechanisms that cause fatigue are specific to the task being performed (Enoka and Duchateau 2008). Moreover, the mechanisms underlying muscle fatigue do not occur in isolation in vivo. Rather, muscular work is supported by many complex physiological systems, any of which could fail during exercise and thus contribute to fatigue (Kent et al. 2016).
The development of chronic muscle fatigue is related to muscle wasting mediated by aging, immobilization, myopathies, insulin resistance (through high-fat dietary intake), diseases associated with systemic inflammation (arthritis, sepsis, infections), trauma, cardio-vascular (heart failure) and respiratory disorders (COPD), chronic kidney failure, MS, and, more recently, coronavirus disease 2019 (COVID-19) (Constantin-Teodosiu and Constantin 2021).
2. Peripheral Processes in Muscle Fatigue
During fatiguing exercise, alterations occur within the CNS and the muscle, but their relative importance depends on the severity of exercise. Specifically, neuromuscular responses to locomotor exercise differ in maximal, severe, heavy and moderate intensity, high-intensity intermittent exercise, and differences in neuromuscular responses between exercise modalities. Reductions in voluntary activation are greater during heavy and moderate intensity exercise. Although post-exercise reductions in voluntary activation following maximal and severe intensity exercise are generally modest, there may be some alterations at the cortical and/or spinal level. During maximal and severe intensity exercise, alterations in neuromuscular function reside primarily within the muscle. During prolonged heavy and moderate intensity exercise, impairments in contractile function are attenuated with respect to severe intensity exercise, but still occur widely (Brownstein et al. 2021).
2.1. Peripheral Muscle Fatigue in Health
Several definitions of muscle fatigue have been forwarded. For example, muscle fatigue is evidenced by a decrease in maximal isometric force, shortening velocity, twitch relaxation speed, or power output (Allen et al. 2008; Enoka and Duchateau 2008). The mechanisms contributing to muscle fatigue have not been completely elucidated because of interacting processes occurring at different levels (Kent-Braun et al. 2012). Some major hypotheses have been proposed to underlie skeletal muscle weakness: (i) glycogen depletion hypothesis, (ii) muscle damage hypothesis, (iii) inflammation hypothesis, and (iv) the oxidative stress hypothesis (Cheng et al. 2020).
Within and around muscle fibers, metabolic and physico-chemical changes are primarily involved, including changes in the neuromuscular transmission, sarcolemmal excitability, extracellular and intracellular ions, action potential (AP), excitation-contraction coupling (ECC), contractile mechanisms and metabolic energy supply (Allen et al. 2008; Bigland-Ritchie 1984; Gandevia 2001; Kent-Braun et al. 2012). The relative importance of factors and sites depends on muscle fiber types and composition, state of training and fitness, the intensity, type and duration of muscle activity, muscle temperature, age and sex (Allen et al. 2008; Enoka and Duchateau 2008; Fitts 1994; Green 1997). Among others, inorganic phosphate, protons, lactate and free Mg2+ accumulate within muscle fibers. They directly affect the mechanical machinery of the muscle cells. Furthermore, they negatively affect the different muscle-cell organelles that are involved in the transmission of neuronal signals. The contracting muscle fibers also release cytokines, which in turn create many effects in other organs, including the brain (Ament and Verkerke 2009).
Muscle fatigue has been categorized into different types, which depend on the type of muscle affected, the stimulus pattern, type of contractile activity, state of fitness and various neural factors (Allen et al. 2008; Amann et al. 2015; Kent-Braun et al. 2012).
High-frequency Fatigue. Continuous high-frequency muscle activation, which generates near-maximal force, typically leads to rapid force decline followed by rapid recovery after stimulation offset. Above about 30-50% of maximal force development, part of the fatigue is due to occlusion of blood circulation. Fatigue develops more dramatically during dynamic exercise involving shortening contractions than during isometric contractions (Jones 1993).
Repeated Short Tetani entail slower fatigue development that depends on stimulation rate and duty cycle (fraction of time covered by contraction). This stimulation pattern simulates natural rhythmic activities like walking, running, cycling, breathing etc. Initially after activity starts, fast-twitch muscle fibers show a rapid decrease in force (phase 1), followed by a relatively constant force (phase 2), which ends with a rapid decline in force (phase 3) (Allen et al. 2008).
Low-frequency Fatigue develops after long-lasting low-frequency tetanic stimulation, after which force remains depressed over long periods (Kent-Braun et al. 2012).
Strenuous exercise induces beneficial skeletal muscle adaptations, ranging from increased endurance due to mitochondrial biogenesis and angiogenesis, to increased strength from hypertrophy. While exercise is necessary to trigger and stimulate muscle adaptations, the post-exercise recovery period is equally critical in providing sufficient time for metabolic and structural adaptations to occur within skeletal muscle (Cheng et al. 2020).
2.1.1. Metabolic Factors in Muscle Fatigue
When a slow mammalian muscle is maximally activated from rest, its metabolic rate increases strongly by a factor of approximately 20, the increase being specific to animal species, muscle-fiber type and form of exercise (Ball 2015; Westerblad et al. 1991). The metabolic changes alter intra- and extracellular ion and metabolite concentrations that can influence muscle fiber function (Debold et al. 2016). Factors influencing these changes include the type of experimental preparation, pattern of fatiguing stimulation, type of muscle fiber, aerobic versus anaerobic conditions, and isometric vs. shortening contractions (Allen et al. 2008; Jones 2010; Lännergren et al. 1993). It should be noted that fatigue may spread from one part to adjacent inactive parts within the muscle but not between whole muscles (Kostyukov et al. 2002).
Glycogenolysis. Glycogen is rapidly depleted during intense exercise by anaerobic glycolysis and more slowly in aerobic exercise. Glycogen depletion may decrease Ca2+ release from the sarcoplasmic reticulum (SR) by putative mechanisms mentioned below (Allen et al. 2008; Ørtenblad et al. 2013).
Anaerobic Glycolysis is turned on rapidly and leads to breakdown of glucose into lactate and protons (H+), i.e., early acidosis. A significant decrease in intracellular pH can reduce the Ca2+ sensitivity of the myofibrils, the number of high-force cross-bridges in fast muscle fibers and the force per cross-bridge in fast and slow fibers (Fitts 2008).
Intracellular ATP Concentration is low and kept fairly constant by resynthesis from creatine phosphate (CrP), glycogen and fatty acids (Ball 2015). Adenosin-triphosphate (ATP) concentration falls late in fatiguing muscle because initially it is reconstituted by CrP. Fast muscles consume ATP, producing adenosin-diphosphate (ADP) and inorganic phosphate (Pi), much faster than regenerating it. The late decline in ATP (and concomitantly in [CrP]) can reduce Ca2+ pumping back into the SR, resulting in augmented resting [Ca2+]i. Transient increases in [ADP]i appear to be involved in reducing the maximal shortening velocity and in slowing relaxation (Allen et al. 2008).
Creatine and Inorganic Phosphate Pi Concentrations [Cr] and [Pi] increase early as a consequence of increased CrP breakdown. Whereas [Cr] does not appear to play a significant role in fatigue, the increase in [Pi] is a major contributor. [Pi] concentration can significantly and rapidly increase during intense fatigue, decreasing Ca2+ release from the SR, Ca2+ sensitivity of the myofibrils, and myofibrillar force production by a direct action on cross-bridge function, and may thus contribute to slow down the rate of muscle relaxation (Allen and Trajanovska 2012; Fitts 2008).
Metabolisation of Pyruvate to Lactic Acid. Insufficient muscle oxygenation, for example during strong muscle contractions that occlude blood flow, promote anaerobic glycolysis. Consequently, pyruvate is metabolized to lactic acid, leading to intra- and extracellular acidosis (Allen et al. 2008). Traditionally, lactic acid has been called a waste product of anaerobic metabolism and has been believed to be responsible for the uncomfortable `burn’ of intense exercise and directly responsible for the metabolic acidosis of exercise, leading to decreased muscle contractility. However, contemporary lactate theories argue for positive effects, including lactate’s role in energy production, its contributions to metabolic acidosis, and its function as an energy substrate for a variety of tissues (Hall et al. 2016). In fact, there is evidence to show that lactate/H+, rather than having detrimental effects on muscle performance during high-intensity exercise, may even improve it. However, induced acidosis can exacerbate fatigue during whole-body dynamic exercise and alkalosis can improve exercise performance in events lasting 1-10 minutes. It has therefore been hypothesized that a severe plasma acidosis in humans might impair exercise performance by causing a reduced CNS drive to muscle (Cairns 2006; see also Hall et al. 2016).
Production of Reactive Oxygen Species and Reactive Nitrogen Species (ROS/RNS). In rested muscle fibers, mild and acute exposure to exogenous ROS/RNS generally increases myofibrillar sub-maximal force, while stronger or prolonged exposure has the opposite effect. Endogenous production of ROS/RNS seems to preferentially decrease sub-maximal force, and positive effects of anti-oxidants are mainly observed during fatigue induced by submaximal contractions. Fatigued muscle fibers frequently undergo a prolonged state of reduced sub-maximal force, which impairs SR Ca2+ release, depresses myofibrillar Ca2+ sensitivity, and depresses Na+-K+ pump activity (Allen et al. 2008; Cheng et al. 2016; Debold 2015; Fitts 2008; Powers and Jackson 2008; McKenna et al. 2008). Low physiological concentrations of ROS are required for normal force production, whereas high concentrations promote contractile dysfunction (Powers and Jackson 2008). This contraction-induced ROS generation is associated with (i) oxidant damage in several tissues, (ii) accelerated muscle fatigue, and (iii) activation of biochemical signaling pathways that contribute to exercise-induced adaptation in the contracting muscle fibers. It still remains open whether exercise-induced increases in ROS production are beneficial or harmful to health (Powers et al. 2020). In conclusion, increased ROS/RNS production have both beneficial and detrimental effects on skeletal muscle function and the outcome depends on a combination of factors: the type of ROS/RNS; the magnitude, duration and location of ROS/RNS production; and the defense systems, including both endogenous and exogenous antioxidants (Cheng et al. 2016).
Alterations in Intracellular Ca2+ Exchange. Dysregulation of Ca2+ release from the SR has been associated with impaired muscle function induced by a wide range of stressors, from dystrophy to heart failure to muscle fatigue.. Early stages of muscle fatigue involve impaired myofibrillar function, whereas decreased Ca2+ release from the SR becomes more important later. SR Ca2+ release can also decline with more prolonged, lower intensity exercise, and it is then related to glycogen depletion. Increased ROS/RNS species can cause long-lasting impairments in SR Ca2+ release resulting in a prolonged force depression after exercise (Cheng et al. 2018). The rates of Ca2+ uptake and release by the SR is reduced, due to the fact that fatiguing activity causes intrinsic alterations in the functional properties of the SR. It has been proposed that fatigue-induced alterations in Ca2+ exchange may be beneficial by reducing the rate of energy utilization by the muscle fiber and preventing irreversible damage to the cell (Williams and Klug 1995). Altered regulation of SR Ca2+ release during chronic stress may be associated with the SR Ca2+ release channel known as the type 1 ryanodine receptor (Bellinger et al. 2008).
Alterations in K+. Perturbations in potassium (K+) are a key factor in skeletal muscle fatigue, but the issue is complicated (Renaud et al. 2023). One proposed concept suggests that K+ has two physiological roles: (i) K+-induced potentiation and (ii) K+-induced force depression. During low-moderate intensity muscle contractions, the K+-induced potentiation of sub-maximal tetanic contractions dominates, thereby optimizing muscle performance. When ATP demand exceeds supply, creating metabolic stress, KATP channels contribute to force reductions by lowering sarcolemmal generation of APs. The ultimate function of these changes is to preserve the remaining ATP to prevent damaging ATP depletion (Renaud et al. 2023). Alterations in K+ transport proteins contribute to, or modulate, K+ disturbances during exercise. Muscle and plasma K+ balance are markedly altered during and after high-intensity dynamic exercise, static contractions and ischemia. Moderate elevations of plasma and interstitial [K+] during exercise have beneficial effects on multiple physiological systems. Severe reductions of the trans-sarcolemmal [K+] gradient likely contributes to muscle and whole-body fatigue (Lindinger and Cairns 2021).
2.1.2. Disturbances in Excitation-Contraction Coupling
Muscle contractions involve critical interactions between myofibril [Ca2+]i and the muscle proteins actin and myosin, control of myofibril Na+ and K+ concentrations, proper functioning of the T-tubular voltage sensor and regulation of Ca2+ release and re-uptake by the SR. Malfunctioning of one or more of these processes will result in disturbances of ECC (Allen et al. 2008; Kent-Braun et al. 2012; McKenna et al. 2008).
Altered Excitability of the Sarcolemma and/or T-tubular System. According to the membrane hypothesis of muscle fatigue, AP propagation along the sarcolemma and into the T-tubules is compromised. AP generation and propagation are influenced by many factors, such as internal and external resistances of the muscle fiber, membrane resistance and capacity, and resting membrane potential, which in turn is influenced by extra- and intracellular ion concentrations. Changes in the latter are most marked with continuous high-frequency stimulation (Allen et al. 2008; Kent-Braun et al. 2012; McKenna et al. 2008; Sejersted and Sjøgaard 2000).
Changes in K+ and Na+ Concentrations. Intense, repeated stimulus-evoked muscle contractions can increase intracellular Na+ concentration, reduce intracellular K+ concentration and elevate extracellular K+ concentration, leading to cell membrane depolarization, Na+-channel inactivation, impaired AP generation and propagation, which in turn depress the Ca2+ release from the SR (Allen et al. 2008; Kent-Baum et al. 2012; McKenna et al. 2008).
Impaired T-tubular Voltage-sensor Activation. Voltage-sensor activation may be reduced by a decreased AP and by voltage-sensor inactivation (Allen et al. 2008; Kent-Baum et al. 2012).
Changes in Ca2+ Dynamics. The Ca2+ dynamics and effects may be impaired by a reduced Ca2+ release from the sarcoplasmic reticulum and slowed Ca2+ re-uptake into the SR, as well as a decrease in Ca2+ sensitivity of myofibrillar proteins, which in turn may result from electrical and chemical changes listed above (Allen et al. 2008; Kent-Baum et al. 2012).
Inhibition of Actin-Myosin Interactions. Increased Pi and H+ concentrations may inhibit the transition from the low- to high-force cross-bridge state and the release of ADP from the cross-bridge (Kent-Baum et al. 2012).
2.1.3. Muscle Fatigue and Damage
Prolonged, intense, fatiguing exercise may cause muscle damage, in particular after frequent unaccustomed eccentric exercise, where the contracting muscle is forcibly lengthened. Greater and longer-lasting fatigue occurs after eccentric compared with concentric and isometric exercise. One day after intense exercise involving stretched contractions, muscles are weak, sore and tender, and this damage can take a week to recover. In this condition, sarcomeres are disorganized and there are increases in resting intracellular Ca2+ and Na+ concentrations (Allen 2004). The initial damage has been ascribed to mechanical disruption of the fiber (sarcomeres, myofibrillar disorientation, and damage to the cytoskeletal framework), preferential fiber-type damage, and impaired muscle metabolism, and subsequent damage is linked to inflammatory processes and to changes in ECC within the muscle. Reductions occur in the ability to generate power output during single-joint movements as well as during cycling and vertical jump movements. Higher forces are achieved during eccentric contractions with less ATP usage and greater increases in temperature (Byrne et al. 2004; Clarkson and Hubal 2002; Clarkson and Newham 1995; Green 1997; Proske 2019). In view of the intense nature of the activity and the need for extensive, high-frequency recruitment of muscle fibers and MUs in a range of synergistic muscles, there is limited opportunity for compensatory strategies to enable performance to be sustained (Green 1997). Indeed, at the single MU level, eccentric exercise causes substantial changes in recruitment thresholds, discharge rates, MU conduction velocities and synchronization, which can last for up to 1 week after eccentric exercise (Semmler 2014).
Muscle Adaptation. Repeated strenuous exercise triggers beneficial skeletal muscle adaptations, ranging from increased endurance due to mitochondrial biogenesis and angiogenesis, to increased strength from hypertrophy. The post-exercise recovery period is critical in providing sufficient time for metabolic and structural adaptations. These alternations between exhausting exercise and recovery underlie any effective exercise training to improve muscle endurance and strength (Cheng et al. 2020).
Proprioception and Kinesthesia. After intense exercise, people are clumsy in the execution of skilled movements. Eccentric exercise produces muscle damage (Clarkson and Hubal 2002), and the damage might extend to muscle spindles, producing a disturbance of limb position sense. However, provided the exercise was sufficiently severe (20-30% fall in muscle force), comparing eccentric exercise with concentric exercise, where no damage ensues, there was no difference in the effects on position sense. After exercise of elbow muscles, the forearm was always perceived as more extended than its actual position. This prompted a question: After exercise, did the extra effort required to lift the fatigued arm provide a position signal? Experimental findings led to the conclusion that post-exercise disturbances to position sense, did not involve peripheral receptors, and that the effect arose within the brain (Proske 2019).
2.1.4. Recovery from Muscle Fatigue
Force recovery after fatiguing stimulation shows different time courses depending on the pattern of stimulation. After brief, high-intensity exercise in humans, voluntary force is typically restored within a few minutes due to recovery of central fatigue and peripheral fatigue associated with ECC and reperfusion of muscles. But recovery may remain incomplete for some hours. After long-lasting, low-intensity exercise, voluntary force typically shows rapid but partial recovery within the first few minutes, mainly due to recovery from central fatigue (Carroll et al. 2017). There is suggestive evidence that different time courses of recovery are associated with [K+] and [Ca2+] concentration gradients. It has been proposed that the fast component of recovery represents diffusion of elevated [K+] from the T-tubules. In addition, the time course of recovery of central Ca2+ release and force are both consistent with the time for diffusion of small ions out of the T-tubule (Allen et al. 2008).
2.2. Peripheral Muscle Fatigue in Disease
2.2.1. Overview
Numerous health problems, including acute critical illness, cancer, diseases associated with chronic inflammation and neurological disorders, often result in skeletal muscle wasting, weakness and fatigue. More specifically, such conditions include aging, immobilization, myopathies [muscle dystrophies (e.g., Duchenne and Becker's muscular dystrophy and facio- scapulo-humeral dystrophy), myotonic syndromes (myotonia congenita, paramyotonia congenita, hypokalemic and hyperkalemic periodic paralysis), myasthenia gravis among others], hereditary motor and sensory neuropathy, post-polio syndrome (PPS), Guillain-Barré syndrome (GBS), insulin resistance, diseases associated with systemic inflammation (arthritis among other rheumatic diseases, sepsis, infections), trauma, cardio-vascular (heart failure, stroke), and respiratory disorders [COPD], chronic kidney failure, MS, and, more recently, coronavirus disease 2019 (COVID-19). Skeletal muscle atrophy is also associated with increased morbidity and mortality of patients (Constantin-Teodosiu and Constantin 2021; Pearson and Young 1993; Powers et al. 2016; Zwarts et al. 2008).
2.2.2. Old Age
Motor performance declines with old age, particularly after 80, including (i) reduced maximal strength and power, slower contraction velocity and increased fatigability, and (ii) increased variability during and between motor tasks, including decreased force steadiness and increased variability of contraction velocity and torque over repeat contractions. The increase in variability seems to involve reduced and more variable synaptic inputs that drive MN activation, fewer and larger MUs, smaller and slower skeletal muscle fibers, less stable neuromuscular junctions, lower and more variable MU AP discharge rates. In old subjects, physical activity may modify MU properties and function (Hunter et al. 2016).
Sarcopenia is a loss of muscle mass and function. In aging, it is due to mild chronic inflammation but also to fiber-intrinsic defects, such as mitochondrial dysfunction. Age-related sarcopenia is associated with physical disability. In addition to skeletal muscle, the nervous tissue is also affected in elderly people. With aging, type 2 fast fibers preferentially undergo denervation and are reinnervated by slow-twitch MNs. They spread forming new neuromuscular junctions with the denervated fibers, resulting in an increased proportion of slow fibers (Coletti et al. 2022).
2.2.3. Myopathies
Myopathies are muscle diseases that have no neuronal causes. The affected muscle groups usually show weakness due to structural alterations and/or functional impairments. Primary myopathies such as muscle dystrophies, myotonic syndromes mitochondrial myopathies and congenital myopathies are differentiated from myopathies caused by systemic inflammatory, endocrine, or toxic disorders.
Inherited Muscle Dysfunction. As an example, mutations in the gene encoding RyR1 have been linked to skeletal myopathies, the basis for which is presumably altered Ca2+ homeostasis. RyR1 mutations cause, among others, malignant hyperthermia (MH), which features susceptibility to a severe, life-threatening rise in body temperature and muscle rigidity upon exposure to, e.g., halothane. This potentially fatal disorder results from the triggering of a rapid and sustained rise in intracellular Ca2+ in muscle due to hyperactivation of RyR1 channels. In contrast to MH, which presents as an acute crisis in an otherwise normal individual, another group of RyR1 mutations cause central-core disease, a congenital myopathy that presents in infancy and is characterized by poor muscle tone, muscle weakness, and musculo-skeletal abnormalities (Bellinger et al. 2008).
Metabolic Myopathies include several genetic disorders that result from abnormalities in metabolic pathways. These conditions result in impaired energy production and utilization within muscle cells, leading to limitations in muscle function with concomitant occurrence of related signs and symptoms, among which fatigue is one of the most frequently reported. The key biomolecules involved in muscle fatigue in metabolic myopathies include energy substrates, enzymes, ion channels, and signaling molecules (Schirinzi et al. 2023).
Mitochondrial Myopathies. In the rare cases of mitochondrial myopathies, which can lead to lactic acidosis (typically in MELAS syndrome), symptoms of severe exhaustion, weakness and muscle fatigue during exercise are predominant features (Allen et al. 2008).
Duchenne Muscular Dystrophy (DMD) and Becker Muscular Dystrophy (BMD). DMD is a lethal degenerative disease of muscles in which the protein dystrophin is absent. Dystrophic muscles in DMD are more susceptible to stretch-induced muscle damage and the stretch-activated channel seems to be one pathway for the increases in intracellular Ca2+ and Na+ that are a feature of this disease (Allen 2004). Altered Ca2+ homeostasis is probably key to the pathogenesis of the most common forms of muscular dystrophy caused by either a lack or only partial expression of dystrophin, DMD and Becker muscular dystrophy (BMD), respectively. Increased flux of Ca2+ across the sarcolemma and activation of cell-death pathways has been proposed as a mechanism that contributes to the pathogenesis of DMD. Increased mitochondrial Ca2+ uptake resulting in cell death has been proposed as a contributing factor to the muscle weakness associated with DMD (Bellinger et al. 2008).
Myotonic Dystrophy. In addition to symptoms such as myotonia and muscle weakness, patients with myotonic dystrophy experience muscle fatigue. In particular, peripheral fatigue in patients with myotonic dystrophy type 1 has more evident effects on muscle relaxation than on contraction characteristics (Esposito et al. 2017).
Myasthenia Gravis. Patients suffering from myasthenia gravis exhibit peripheral fatigue, but in contrast to MS, the mechanisms of weakness and fatigue in the former is well-defined. In most cases, antibodies to the postsynaptic acetylcholine receptor at the neuromuscular junction, as competitive inhibitors of acetylcholine, diminish the sarcolemmal activation and ensuing force of muscle contractions. Treatments increase the availability of acetylcholine and reduce antibody formation (Cantor 2010).
Muscle Strain Injury. Muscle strain is very common. Many frequently involved muscles cross two joints, act mainly in an eccentric fashion, and contain a high percentage of fast-twitch fibers. Muscle strain usually occurs during strenuous activity and causes acute pain (Noonan and Garrett 1999).
Cancer Cachexia is a complex multi-organ catabolic syndrome and features, in humans and animal models, a major decrease in skeletal muscle mass that depends on the severity and the localization of the tumor. It affects all types of muscle fibers. Animal studies indicate an impairment in mitochondrial oxidative metabolism resulting from a decrease in mitochondrial content, an alteration in mitochondrial morphology, and a reduction in mitochondrial metabolic fluxes. Other features are an increase in collagen deposit, an accumulation of fat depot outside and inside the muscle fiber, and a disrupted contractile machinery structure. The loss of muscle mass largely contributes to the loss of muscle function (Martin and Freyssenet 2021). In addition to wasting and ensuing weakness, muscle fatigue is a common symptom. The etiology is complex and involves peripheral and central mechanisms. Central factors may involve dysregulation of the HPT axis, 5-HT actions and circadian rhythm. Peripherally, muscle structure and function can be disturbed. Such factors include muscle-fiber atrophy, anemia, dehydration, and elevation of circulating cytokines. Cancer cells release interleukin 6 (IL-6), tumor-necrosis factor α (TNF-α), IL-1ß, and interferon γ, which drive inflammatory processes which in turn change metabolism. These factors also impair membrane integrity, which disrupts signaling pathways and leads to muscle damage (Powers et al. 2016).
Chronic (Inflammatory) Diseases include several types of cancer, rheumatoid arthritis among other rheumatic diseases, chronic heart failure and COPD. They appear to have direct deleterious effects of trunk and limb muscles, including atrophy with reduced muscle mass and fiber size and contractile dysfunction. Data suggest that pro-inflammatory cytokines are released by the diseased tissue into the circulation from where they exert endocrine effects on muscles. Primary candidates include IL-6, C-reactive protein, sphingomyelinase and, as primary driver, TNF-α. Reactive oxygen species (ROS) and NO may also play a role (Powers et al. 2016).
Systemic Autoimmune Myopathy. Patients with polymyositis and dermatomyositis suffer from reduced fatigue resistance (Yamada et al. 2022). A low mitochondrial oxidative capacity significantly contributes to this debilitating symptom.
3. Central Processes in Muscle Fatigue
The processes described above appear simply deleterious. The nervous system should find ways to at least mitigate them. Indeed, the nervous system has found quite a few.
3.1. Central Fatigue
3.1.1. Muscle Wisdom
In well-motivated human subjects executing sustained maximal voluntary contractions (MVC), the reduction in muscle force generating capacity is accompanied by a progressive decline of integrated EMG and the mean firing rate of individual MUs. “This, however, does not necessarily result in loss of force since the parallel slowing of muscle contractile speed reduces tetanic fusion frequency. It is suggested that the range of MN firing rates elicited by voluntary effort is regulated and limited for each muscle to the minimum required for maximum force generation, thus preventing neuromuscular transmission failure and optimizing motor control” (Bigland-Ritchie and Woods 1984). This optimization process has been dubbed `muscle wisdom’.
3.1.2. Potential Mechanisms
Muscle fatigue may arise not only because of peripheral changes at the muscle level, but also because the CNS fails to drive the MNs adequately. This goes under the rubric of `central fatigue’. Indeed, much evidence suggests that voluntary activation of human MNs and muscle fibers is sub-optimal, indicating that maximal voluntary force is commonly less than true maximal force. Voluntary activation usually diminishes during maximal voluntary isometric tasks, that is central fatigue develops, and MU firing rates decline. Transcranial magnetic stimulation (TMS) over the motor cortex during fatiguing exercise has revealed focal changes in cortical excitability and inhibitability based on EMG recordings, and a decline in supraspinal `drive’ based on force recordings. Central changes also occur at a spinal level due to the altered input from muscle spindles, Golgi tendon organs (GTOs), and group III (Aδ) and IV (C) muscle afferents innervating the fatiguing muscle. Some intrinsic adaptive properties of the MNs help to minimize fatigue. A number of other central changes occur during fatigue and affect, for example, proprioception, tremor, and postural control (Gandevia 2001; Kent-Braun et al. 2012; Ranieri and Di Lazzaro 2012).
3.2. Spinal Muscle Fatigue in Health
3.2.1. Motor Unit (MU) Properties as a First Line of Defense
As a first line of defense, intrinsic properties of MUs may mitigate the effects of muscle fatigue.
MU Types. One basis of force optimization during developing muscle fatigue resides in the differentiation of MUs into types of different fatigability. In cats (and humans), MUs are classified into three main types: large, strong, fast-contracting and fatiguable MUs (type FF); strong, fast-contracting and fatigue-resistant MUs (type FR); slowly contracting, fatigue-resistant (type S) MUs. During long-lasting weak muscle contractions, S-type MUs are active, while FR-type and FF-type are recruited at contractions of higher forces, where FF-type MUs fatigue relatively soon, with contraction and relaxation times increasing over time (Bączyk et al. 2022; Burke 2009).
Force Potentiation in FF-type MUs. When step-like depolarizing currents of 60 seconds duration were injected into different cat gastrocnemius MU types, S-type MUs showed only a small firing-rate adaptation and their isometric force declined hardly over time, while the FF-type MN discharge rate adapted more strongly, and so did MU force (Figure 1; see legend). Important in the latter case is the fact that FF-type MUs exhibited force potentiation between about 5 and 30 seconds after the start of current injection, compensating initially for force loss (Kernell and Monster 1982). This transitory relative force increase does not suffice to significantly prevent force loss, but must be complemented by sensory feedback from the fatiguing muscle.
Comments. The fact that skeletal muscles are built of assemblies of MUs with different mechanical and fatigue properties and the phenomenon of force potentiation in FF-type MNs are a first line of defense, but do not suffice for the CNS to control muscle fatigue. “You can only control what you sense” (McCloskey and Prochazka 1994). So sensors that monitor the events in fatiguing muscles are required. Which ones are suitable to do so?
Figure 1.
Schematic illustration of the typical adaptation behaviors of MN discharge rate (A) and MU isometric force (B) of a cat gastrocnemius S-type MU (“S unit”) and a gastrocnemius FF-type MU (“FF unit”) in response to step-like, long-lasting depolarizing currents injected into the respective MNs. The red dashed curves were fitted to data from Kernell and Monster (1982; Fig. 5). These curves represent relative values and, for better comparison, were normalized to the initial values set to 100%. As obvious, neither the discharge rate (A) nor the force (B) of the S-type MU adapted very much over time. By contrast, the respective values of the FF-MU did. The lower dashed curve for FF-MU force (B) was fitted to the data during the first five seconds and the data beyond 35 seconds, and extrapolated in between. As seen, the actual recorded MU force (green dashed curve) exceeded the extrapolated values, indicating that, for about 30 seconds, the FF MU developed a potentiated force. In fact, the time course of the red fit is an estimate of the unpotentiated MU force development, as supported by independent data. Kernell and Monster (1982; Fig. 6) measured and averaged the forces of five fresh, i.e., non-fatigued and non-potentiated, type FF MUs, produced by stimulation with 1-second trains at the time-related adapted discharge rates when the unit was developing fatigue and potentiation during long-lasting depolarization. Abbreviations: FF: fast-contracting, fast-fatiguing; S: slowly contracting, fatigue-resistant.
Figure 1.
Schematic illustration of the typical adaptation behaviors of MN discharge rate (A) and MU isometric force (B) of a cat gastrocnemius S-type MU (“S unit”) and a gastrocnemius FF-type MU (“FF unit”) in response to step-like, long-lasting depolarizing currents injected into the respective MNs. The red dashed curves were fitted to data from Kernell and Monster (1982; Fig. 5). These curves represent relative values and, for better comparison, were normalized to the initial values set to 100%. As obvious, neither the discharge rate (A) nor the force (B) of the S-type MU adapted very much over time. By contrast, the respective values of the FF-MU did. The lower dashed curve for FF-MU force (B) was fitted to the data during the first five seconds and the data beyond 35 seconds, and extrapolated in between. As seen, the actual recorded MU force (green dashed curve) exceeded the extrapolated values, indicating that, for about 30 seconds, the FF MU developed a potentiated force. In fact, the time course of the red fit is an estimate of the unpotentiated MU force development, as supported by independent data. Kernell and Monster (1982; Fig. 6) measured and averaged the forces of five fresh, i.e., non-fatigued and non-potentiated, type FF MUs, produced by stimulation with 1-second trains at the time-related adapted discharge rates when the unit was developing fatigue and potentiation during long-lasting depolarization. Abbreviations: FF: fast-contracting, fast-fatiguing; S: slowly contracting, fatigue-resistant.

3.2.2. Neuronal Sensors of Muscle Fatigue
Several classes of neuronal sensors and their afferent fibers could signal the muscle state during fatigue. Among them are the larger-diameter group Ia and II afferents from muscle spindles and group Ib afferents from GTOs, and smaller-diameter group II and IV afferents from free nerve endings. Muscle spindle afferents are commonly regarded as muscle length receptors, GTO as afferents muscle force receptors, and group II and IV afferents are more complex.
3.2.2.1. Fatigue-Related Changes in Firing of Muscle Spindle Afferents
In the course of fatiguing muscle contractions, human muscle spindle afferents reduced their average discharge rate, possibly due to a reduction in fusimotor drive to spindles (Macefield et al. 1991). Homonymous and synergistic MNs would thereby be disfacilitated, which could contribute to their firing rate decline. The differences in spindle discharge to the above cat data is probably due to the continuous fusimotor activation throughout the fatiguing stimulation.
Comparable results were obtained in the rat masseter muscle system. The masseter muscle was fatigued by prolonged tetanic masseter-nerve electrical stimulation. Muscle fatigue caused a decrease in the discharge frequency of masseter muscle spindle afferents in most of the examined units. Hence, fatigue signals from the fatiguing muscle, traveling through capsaicin-sensitive fibers, are able to diminish the proprioceptive input by a central presynaptic influence (Brunetti et al 2003).
The question arises whether beyond the average discharge rate of spindle afferents, their discharge fluctuation could provide valuable information on muscle state, in particular about fatigue.
Sensitivity of Spindle Afferents to Single MU Contractions. The precise temporal discharge patterns of muscle spindle afferents depend on their sensitivity to small-amplitude mechanical events in muscles, especially MU contractions Thus, in cats, muscle spindle afferents, in particular group Ia afferents, respond to single MU contractions. Hence, the contraction of a single MU can be an effective stimulus to a spindle receptor and may induce afferent firing pattern alterations similar to those observed with whole muscle contraction. There are quantitative differences in the response of a receptor to contraction of different MUs and to contraction of the same MU at different lengths. It has been suggested that, rather than simply acting as generalized force or length sensors for the muscle as a whole, each receptor’s spike train carries information about the state of a particular set of MUs (Binder et al. 1976). The quality of this information of course depends on the activity of many MUs on an afferent. Thus, by simultaneously stimulating multiple α-axons with different uncorrelated patterns and recording from multiple group Ia fibers, post-stimulus time histograms (PSTH) revealed widely graded influences of single MU contractions on different muscle spindles, as described above. But in most instances, the signal transmission from a single MU to a single Ia fiber was disturbed to varying degrees by concomitant activity from other MUs (Schwestka et al. 1981). Nonetheless, using the same stimulus paradigm, contractions of single MUs were reflected in membrane potential changes of homonymous α-MNs (Koehler et al. 1984). What is the relevance to muscle fatigue signaling? Using the same approach as Schwestka et al (1981), time-domain (PSTH) and frequency-domain (gain) computations were performed to study the effects of fatiguing muscle-unit contractions on the signal transmission from motor efferents to spindle afferents. In the course of muscle unit fatigue, during which the gain of the force-producing sub-system decreased, the gain of the sub-system transforming force to afferent discharge increased so that the overall gain between motor efferents and spindle afferents remained relatively high. This could be a mechanism that preserves a high quality of afferent information on MU contractions (Christakos and Windhorst 1986). Still, whether under natural conditions single-unit activity patterns have an influential effect on MNs during fatigue remains in doubt.
3.2.2.2. Fatigue-Related Changes in Firing of GTO Afferents
The firing of group Ib afferents from GTOs would be expected to decrease with declining muscle force. Indeed, alterations in the stretch sensitivity of group Ib afferents have been reported in fatigued gastrocnemius muscles of cats. Group Ib afferent responses to the velocities of Achilles tendon stretch were either completely abolished or depressed over several seconds compared to pre-fatigue firing frequencies. Post-excitation depression of the group Ib receptor potential appears to be one possible mechanism (Hutton and Nelson 1986). However, how these changes would influence the reflex effects is less clear. Excitatory and inhibitory interneurons intercalated in spinal reflex pathways of group Ib afferents from GTOs receive input from group III/IV afferents (Jankowska 1992; Schomburg 1990; Figure 2) and might thus reduce their discharge with fatigue. And since group Ia and Ib afferents in part converge on the same interneurons, support from muscle spindles would be expected to also decline. In fact, in cats, excitatory and inhibitory interneurons are shared to a large extent with group II afferents and in part even group Ia afferents from muscle spindles (Jankowska and Edgley 2010). The way the discharge of these interneurons behave during muscle fatigue would depend on a delicate balance between influences. – It has been suggested that GTOs are able to monitor muscle damage resulting from repeated eccentric contractions. Such contractions cause an increase in passive muscle tension and GTO discharge (Gregory et al. 2003).
Figure 2.
Rarified scheme of group III/IV afferent connections to some important spinal neurons in the cat. For simplicity, only connections from ankle extensor calf muscles are shown, while flexor muscle afferents are not. A muscle spindle is symbolized by a long black bar. For simplicity, secondary endings with their group II afferents are omitted. Spinal excitatory neurons are symbolized by open turquoise circles and their synaptic terminals by T-junctions. Inhibitory interneurons are symbolized by closed and filled purple circles, as are their synaptic terminals. Closed yellow circles symbolize either excitatory or inhibitory neurons, their synaptic terminals being symbolized by superimposed small closed yellow circles and T-junctions. Every circle symbolizes groups of neurons and synapses. Abbreviations: RC, Renshaw cell; RIaI, reciprocal Ia inhibitory interneuron (Own design; see also Windhorst 2003, Windhorst and Dibaj 2023 as well as Dibaj and Windhorst 2024).
Figure 2.
Rarified scheme of group III/IV afferent connections to some important spinal neurons in the cat. For simplicity, only connections from ankle extensor calf muscles are shown, while flexor muscle afferents are not. A muscle spindle is symbolized by a long black bar. For simplicity, secondary endings with their group II afferents are omitted. Spinal excitatory neurons are symbolized by open turquoise circles and their synaptic terminals by T-junctions. Inhibitory interneurons are symbolized by closed and filled purple circles, as are their synaptic terminals. Closed yellow circles symbolize either excitatory or inhibitory neurons, their synaptic terminals being symbolized by superimposed small closed yellow circles and T-junctions. Every circle symbolizes groups of neurons and synapses. Abbreviations: RC, Renshaw cell; RIaI, reciprocal Ia inhibitory interneuron (Own design; see also Windhorst 2003, Windhorst and Dibaj 2023 as well as Dibaj and Windhorst 2024).
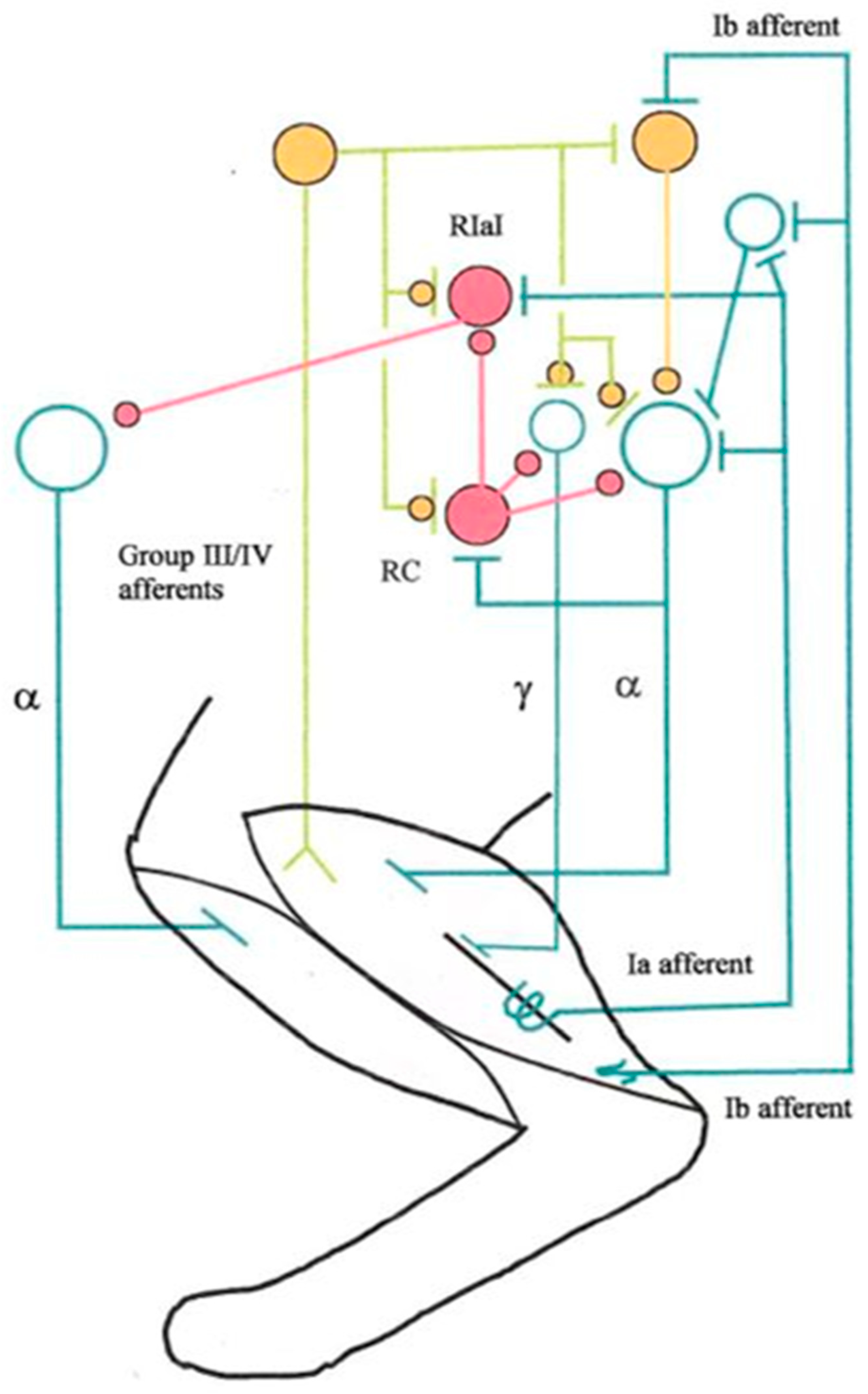
The same question as above for spindle afferents arises whether beyond the average discharge rate of GTO afferents, their discharge fluctuation could provide valuable information on muscle state, in particular about fatigue.
Sensitivity of GTO Afferents to Single MU Contractions. The responses of cat soleus GTOs to repetitive twitch contractions of: (i) different MUs within the muscle, (ii) single MUs at different muscle lengths, and (iii) single MUs when the pulse-train pattern of stimulation delivered to the MU axon was altered. Group Ib afferents responded to each of several hundred successive MU twitches with identical numbers of spikes and with relatively invariant latencies. GTOs are sensitive to subtle alterations in MU twitch waveform and amplitude, and this sensitivity was reflected in the precise timings of their afferent discharge. Considering the number of GTOs in a muscle, and the likelihood that every MU is connected with at least one receptor, the sensitivity of GTOs insures that every twitch of every MU will be reflected in the population of afferent signals (Binder et al. 1977).
3.2.2.3. Group III/IV Muscle Afferents
Group III/IV afferents from muscle free nerve endings are sensitive to both mechanical stimuli and metabolic and thermal changes in muscle. During fatiguing muscle contractions, they are excited by mechanical changes (Hayward et al. 1991), as well as by intra-muscular metabolic and chemical changes (Decherchi and Dousset 2003).
Mechanically Sensitive Afferents. Group III muscle afferents are more mechano-sensitive than group IV afferents during skeletal muscle contraction, force production, dynamic/static muscle stretch and local intramuscular pressure. But their response to a mechanical stimulus may be potentiated by chemical stimuli. Thus, muscle fatigue both increases spontaneous discharge in these mechanically sensitive afferents and sensitizes their response to muscle stretch, surface pressure, and, in a few instances, muscle contraction. These fatigue-induced changes occur after 5-10 min of sub-maximal fatiguing stimulation. However, the discharge of many slowly conducting mechano-receptor afferents declines during the initial phase of fatigue, which may argue against a primary role in mediating the initial decline in MN rate that is so prominent in fatiguing maximum voluntary muscular contraction (Hayward et al. 1991).
Metabolically Sensitive Afferents have been suggested to be of two sub-types. One sub-type, the so-called metabo- or ergoreceptors, only responds to innocuous concentrations of intramuscular metabolites (protons, lactate, ATP) associated with ‘normal’ (i.e. freely perfused and predominantly aerobic) exercise up to strenuous intensities. In contrast, the other subtype, the so-called metabo-nociceptors, only respond to higher (and concurrently noxious) levels of metabolites present in muscle during ischemic contractions or following hypertonic saline infusions – but not to non-noxious metabolite concentrations associated with normal exercise. The sub-types show molecular differences including the differential expression of purinergic receptors (P2X2,3,4), transient receptor potential vanilloid type 1 and/or 2 (TRPV1/2), and acid-sensing ion channels 1, 2, and 3 (ASIC 1-3) (Amann et al. 2015).
Muscle group IV afferents are more sensitive to metabolites released into the interstitium by muscle activity and their activation usually starts after a delay during prolonged muscle contraction and continues to discharge until the withdrawal of muscle metabolites (Amann et al. 2015; Decherchi and Dousset 2003; Gandevia 2001; Laurin et al. 2015; Mense 1993; Rotto and Kaufman 1988; Taylor et al. 2016).
Group III and IV muscle afferents originating in exercising limb muscles play a significant role in the development of fatigue during exercise in humans. Their activation causes effects all along the CNS. Due to their wide central distribution, they have diverse roles. They adjust heart rate, ventilation, blood pressure and vascular resistance during physical exercise, modulate spinal reflexes, and may contribute to improve muscle performance by regulating the peripheral fatigue development and by avoiding excessive muscle impairments. Feedback from group III/IV muscle afferents to the CNS reflexively increases ventilation and central (cardiac output) and peripheral (limb blood flow) hemodynamic responses during exercise and thereby assures adequate muscle blood flow and O2 delivery. This response is a key factor in minimizing the rate of development of peripheral fatigue and in optimizing aerobic exercise capacity. On the other hand, the central projection of group III/IV muscle afferents impairs performance and limits the exercising human via its diminishing effect on the output from spinal MNs which decreases voluntary muscle activation (i.e. facilitates central fatigue).
Tetrodotoxin-resistant (TTX-R) afferent fibers from rat muscles exhibit a multi-sensitive profile, including nociception. TTX-R afferent fibers play an important role in motor control, via spinal and supraspinal loops. To quantify the TTX-R afferent input, the cord-dorsum potential (CDP) was recorded, which results from the electrical fields set up within the spinal cord by the depolarization of dorsal horn (DH) interneurons, activated by an incoming volley of TTX-R muscle afferents. The changes in TTX-R CDP size before, during and after fatiguing electrical stimulation of the gastrocnemius-soleus (GS) muscle were taken as a measure of TTX-R C-unit activation. At the end of the fatiguing protocol, following an exponential drop in force, TTX-R CDP area decreased in the majority of trials (9/14) to 0.75 ± 0.03% (mean ± SEM) of the pre-fatigue value. Recovery to the control size of the TTX-R CDP was incomplete after 10 min. Furthermore, fatiguing trials could sensitize a fraction of the TTX-R C-fibers responding to muscle pinch. This suggests a long-lasting activation of the TTX-R muscle afferents after fatiguing stimulation (Kalezic and Steffens 2013).
Fatigue-induced Group III/IV Effects in the Spinal Cord. In the spinal DH, small-diameter afferents (group III or Aδ and group IV or C) contact a large variety of excitatory and inhibitory interneurons that provide for complex signal processing at spinal levels, and also connect with a minority of projection neurons that send axons rostrally. Projection neurons also produce axon collaterals that are widely distributed within and between spinal segments, whose functions are hardly known, however.
DH neurons also receive modulating inputs from higher centers.
Spinal Distributions of Group III/IV Afferent Connections. Group III/IV afferents distribute their signals in the spinal cord, as tested by Fos-immunoreactive (Fos-ir) expression and nicotinamide adenine dinucleotide phosphate-diaphorase (NADPH-d)-reactive neurons upon fatiguing contractions of the cat triceps surae muscle, caused by direct intermittent high-rate (100 s-1) electrical stimulation of the triceps surae muscle or the ventral root L5 (VRL5) for 30 min. The fatigue-related c-fos gene expression was more extensive in the L2-L5 segments on the stimulated side, and the majority of Fos-ir neurons were concentrated in the DH. After direct electrical muscle stimulation, the highest number of Fos-ir neurons were detected in two regions: layer 5, and superficial layers (1 and 2(o)), although many labeled cells were also found in layers 3, 4, 6, and 7. In response to VRL5 stimulation, the maximal density of Fos-ir neurons was detected in the middle and lateral parts of layers 1 and 2(o), the zone of termination of high-threshold muscle afferents. Fos-ir cells were also found in layers 5 and 7 on the stimulated side. A few Fos-ir neurons were found in the ventral horn (layer 8 and area 10) on both sides. The lamellar distribution of NADPH-d-reactive neurons was similar over all experimental groups of animals. In the L3-L6 segments, such reactive cells were arranged in two distinct regions: DH (layers 2(i), 3, and 5) and area 10; in the L1 and L2 segments, an additional cluster of NADPH-d positive cells was found in the intermediolateral cell column (IML). Double-labeled cells were not detected. We suggest that c-fos expression in response to muscle fatigue reveals activity of functionally different types of spinal neurons which could operate together with NOS-containing cells in pre-MN networks to modulate the MN output (Pilyavskii et al. 2001).
These data raise the possibility that muscle group III/IV afferents influence many, if not all neurons in the spinal cord (Figure 2; Windhorst 2003, 2007).
Comments. All the above sensory afferent systems have widespread distributions in the CNS, which in part are difficult to comprehend, depending on the complex biomechanics of the skeleto-muscular system (Windhorst 2021).
3.2.3. Spinal Fatigue-Related Reflexes
At spinal level, mechanically or metabolically activated group III/IV muscle afferents in cats have polysynaptic effects on α-MNs, γ-MNs and interneurons, such as reciprocal Ia inhibitory interneurons (mediating reciprocal inhibition between antagonist α-MNs), Renshaw cells (RCs) (mediating recurrent inhibition) (Figure 2). Prior to this, they are subjected to the scrutiny of presynaptic inhibition, which itself receives inputs from group III/IV muscle afferents.
3.2.3.1. Presynaptic Inhibition (PSI)
In humans, the stretch-reflex function was investigated after long-lasting stretch-shortening cycle exercise, which resulted in a clear deterioration in muscle function immediately after fatigue, which was accompanied by a clear reduction in active and passive reflex sensitivity. For the ratio of the electrically induced maximal H-reflex to the maximal mass compound AP, only one significant reduction occurred immediately after fatigue (71.2%, P < 0.01). A similar significant decrease in the stretch-resisting force of the muscle also occurred. Indirect markers of muscle damage (serum creatine kinase activity and skeletal troponin I) were increased, which could result from ultrastructural muscle damage. It was suggested that the acute reduction in reflex sensitivity was of reflex origin and due to two active mechanisms, disfacilitation and presynaptic inhibition and that the delayed second decline in the sensitivity of some reflex parameters may be attributable to the secondary injury due to some inflammatory response to the muscle damage. This might emphasize the role of presynaptic inhibition via group III and IV muscle afferents (Avela et al. 1999).
In humans, presynaptic modulation of group Ia afferents converging onto the MN pool of the extensor carpi radialis was compared during contractions (20% of maximal force) sustained to failure as subjects controlled either the angular position of the wrist while supporting an inertial load (position task) or exerted an equivalent force against a rigid restraint (force task). The results suggest a task- and time-dependent modulation of presynaptic inhibition of group Ia afferents during fatiguing contractions (Baudry et al. 2011). Also, there is evidence for effects of muscle fatigue in plantar flexors on presynaptic inhibition (Grosprêtre et al. 2018).
In adult rats, cholinergic (ACh) interneurons in DH laminae III/IV co-localized gamma-aminobutyric acid (GABA) and frequently form axo-axonic synapses with terminals of primary afferents. Therefore, they are probably last-order interneurons involved in presynaptic inhibition. Indeed, ACh interneurons receive contacts from both myelinated and unmyelinated primary afferents. Hence, ACh interneurons likely are components of an inhibitory feedback pathway that is monosynaptically activated by primary afferents (Olave et al. 2002).
In cats, the potential involvement of presynaptic inhibition was tested by conditioning the monosynaptic H-reflex by stimulation of the nerve to the posterior biceps and semitendinosus (PBSt) muscles. The intensity of presynaptic inhibition as measured by the reduction of the normalized value of monosynaptic reflex amplitude during conditioning, increased as compared to the pre-fatigue state, followed by a recovery. Both primary afferent depolarization (PAD) and the intensity of antidromic discharges in primary afferents increased with the presynaptic inhibition intensity. This demonstrated a fatigue-related suppression of group Ia excitation of synergistic MNs, probably arising from the activation of group III/IV afferents (Kalezic et al. 2004).
Rather than indirectly measuring H-reflexes, more direct monosynaptic reflex effects can be measured by intracellular recordings from MNs. In cats, responses of GS MNs to stretches of the homonymous muscles were recorded intracellularly before, during and after fatiguing stimulation (FST) of GS muscles. VRs L7 and S1 were cut, and FST was applied to VR S1, a single FST session including 4 to 5 repetitions of 12s periods of regular 40/s stimulation. Muscle stretches consisted of several phases of slow sinusoidal shortening-lengthening cycles and intermediate constant lengths. The maximal stretch of the muscles was 8.8 mm above the rest length. Effects of FST on excitatory postsynaptic potentials (EPSPs) and APs evoked by the muscle stretches were studied in 12 MNs. Stretch-evoked EPSPs and firing were prominently suppressed after FST, with the exception of a post-contraction increase of the first EPSP after FST, which was most likely due to after-effects in the activity of muscle spindle afferents. The post-fatigue suppression of EPSPs and spike activity was followed by restoration within 60-100 s. Additional bouts of FST augmented the intensity of post-fatigue suppression of EPSPs, with the AP activity sometimes disappearing completely. FST itself elicited EPSPs at latencies suggesting activation of muscle spindle group Ia afferents via stimulation of ß-fibers. The suppression of the stretch-evoked responses most likely resulted from fatigue-evoked activity of group III and IV muscle afferents. Presynaptic inhibition could be one of the mechanisms involved, but homosynaptic depression in the FST-activated group Ia afferents may also have contributed (Kostyukov et al. 2005).
In cats, the magnitude of reflex inhibition, induced in the soleus muscle by contraction or stretch of the medial gastrocnemius (MG) muscle, was substantially increased during and after electrically induced fatigue of the MG (Hayward et al. 1988).
Presynaptic inhibition could thus contribute to the reduction of proprioceptive input to MNs because it is subject to group III/IV afferents from fatiguing muscles.
Comments. Different motor tasks may require different patterns of proprioceptive feedback to regulate spinal motor control functions. Presynaptic inhibition (PSI) as a guard at the spinal input gate is an adaptable mechanism to control the inflow of sensory inputs, in particular from groups Ia, II, Ib, cutaneous and other afferents, which may presynaptically inhibit each other. The interneurons mediating PSI may be excited or inhibited in various proportions by group II to IV afferents, cutaneous and joint afferents, by signals descending in cortico-spinal (CST), rubro-spinal (RST), reticulo-spinal (ReST) and vestibulo-spinal (VST) tracts, nucleus reticularis gigantocellularis, as well as by serotonergic (5-HT) raphé-spinal and noradrenergic (NA) locus coeruleus (LC) systems (Jankowska 1992; Quevedo 2009; Rudomin 2009; Rudomin and Schmidt 1999).). In animals and humans, PSI can be set to different mean levels and modulated dynamically during rest, locomotion and voluntary movements. For example, the monosynaptic transmission from group Ia afferents to -MN is presynaptically inhibited more strongly during stance than rest (lying), and more strongly during running than walking (Katz et al. 1988; Nardone and Schieppati 2004).
3.2.3.2. Monosynaptic Group Ia Afferent Excitation
H-reflex. In humans, the soleus muscles were fatigued under ischemic conditions by intermittent stimulation at 15 Hz. The excitability of the soleus MNs, tested using the H-reflex, was significantly reduced during fatigue. Control experiments with ischemia or electrical stimulation, but without fatigue, failed to demonstrate any significant effects on reflex excitability. This favored the view of reflex inhibition of α-MNs during fatigue (Garland and McComas 1990). H-reflex depression also occurred during low-force fatigue. Eleven healthy subjects performed fatiguing contractions of low force (25% MVC) or high force (42-66% MVC). H-reflexes, muscle compound APs (M-waves), twitch contractile properties, and MU discharges were recorded from the soleus muscle. In the low-force fatigue task, MU firing rate increased gradually over time, whereas the resting H-reflex was significantly depressed at 15% of endurance time and remained quasi-constant for the rest of the task. This suggests that, during a low-force fatigue task, the processes mediating the resting H-reflex depression are relatively independent of those modulating the MU firing rate. In the high-force fatigue task, a decline in the average MU discharge rate was accompanied by a decrease in the resting H-reflex amplitude and a prolongation of the twitch half-relaxation time (HRT) at the completion of the fatigue task (Kuchinad et al. 2004).
In decerebrate cats, changes in the monosynaptic H-reflex of GS MNs were studied after fatiguing stimulation of the GS muscles. Monosynaptic reflexes were evoked by stimulation of Ia fibers in the GS nerve and recorded from a filament of ventral root (VR) L7. Intermittent 40/s stimulation for 10-12 minutes was applied to the distal part of the cut VR S1. The potential involvement of recurrent inhibition and presynaptic inhibition was tested by conditioning the monosynaptic reflex by stimulation a filament of VR L7 or of the nerve to the PBSt muscles, respectively. The intensity of presynaptic inhibition as measured by the reduction of the normalized value of monosynaptic reflex amplitude during conditioning, increased as compared to the prefatigue state, followed by a recovery. In contrast, the intensity of recurrent inhibition first diminished and then gradually recovered. Both PAD and the intensity of antidromic discharges in primary afferents increased with the presynaptic inhibition intensity. This demonstrated a fatigue-related suppression of group Ia excitation of synergistic MNs, probably arising from the activation of group III and IV afferents. The effects could in part be due to increased presynaptic inhibition, while recurrent inhibition plays a minor role (Kalezic et al. 2004).
3.2.3.3. Heteronymous Group Ib Afferent Inhibition
Heteronymous inhibition between lower limb muscles is primarily attributed to recurrent inhibitory circuits in humans but could also arise from Golgi tendon organs (GTOs). To distinguish, in humans, between recurrent inhibition and mechanical activation of GTOs, the unique influence of mechanically activated GTOs was examined by comparing the magnitude of heteronymous inhibition from quadriceps (Q) muscle stimulation onto ongoing soleus EMG at five Q stimulation intensities (1.5-2.5× motor threshold) before and after an acute bout of stimulation-induced Q fatigue. Fatigue was used to decrease Q stimulation evoked force (i.e., decreased GTO activation) despite using the same pre-fatigue stimulation currents (i.e., same antidromic recurrent inhibition input). Thus, a decrease in heteronymous inhibition after Q fatigue and a linear relation between stimulation-evoked torque and inhibition both before and after fatigue would support mechanical activation of GTOs as a cause of inhibition. A reduction in evoked torque but no change in inhibition would support recurrent inhibition. After fatigue, Q stimulation-evoked knee torque, heteronymous inhibition magnitude and inhibition duration were significantly decreased for all stimulation intensities. In addition, heteronymous inhibition magnitude was linearly related to twitch-evoked knee torque before and after fatigue. Hence, mechanical activation of GTOs may be a cause of heteronymous inhibition along with recurrent inhibition (Cuadra et al. 2024).
3.2.3.4. Recurrent Inhibition
Fatigue-activated group III/IV afferents might exert their inhibitory effects on MNs via Renshaw cells mediating recurrent inhibition (Figure 2), as suggested by Hayward et al (1991) who felt that this pathway might be involved in specifically regulating MN discharge rate. But at least in cats, Renshaw cells were suppressed by metabolically activated group III/IV afferents (Windhorst et al. 1997).
In cats, by recording changes in Renshaw cell spontaneous discharges and responses to antidromic electrical stimulation of motor axons when small-diameter calf muscle afferents were excited by intra-arterially injected bradykinin, 5-HT, lactic acid and KCI. Whenever such injections had an effect, it transiently raised or lowered the spontaneous firing rate and almost always decreased the antidromic response to motor axon stimulation. This suggests that the Renshaw cell-mediated effects of neurochemically excited afferents would predominantly disinhibit rather than inhibit MNs (Windhorst et al. 1997).
This was consistent with results from another experiment in cats. Using the approach described in Sect., the potential involvement of recurrent inhibition in fatigue conditions was tested by conditioning the monosynaptic H-reflex by stimulation of a filament of VR L7. The intensity of recurrent inhibition first diminished and then gradually recovered after GS fatigue. Recurrent inhibition was slightly decreased but probably plays a minor role (Kalezic et al. 2004).
By contrast, in humans, different results were obtained by recording changes in recurrent inhibition of soleus MNs when high-threshold, group III/IV fibers from the same muscle were tonically activated. At rest, such stimulation produced transient facilitation of both test H and reference H-reflexes. Under weak voluntary contraction, muscle group III/IV fiber stimulation produced long-lasting extra-inhibition and extra-facilitation of the test reflex and reference reflex respectively, the time course of which closely resembled that of the subjective muscle pain curve. It was concluded that the filtering property of recurrent inhibition may contribute to limit MN activity during muscle pain and/or adapt MN firing rate to the modified contractile properties of MUs as muscle fatigue developed (Rossi et al. 2003). This is more in line with the suggestion of Hayward et al (1991).
Comments. The lumped recurrent network depicted in Figure 2 is much simplified because Renshaw cells receive excitatory inputs not only from homonymous and synergistic MNs but also from antagonist MNs and `antagonist’ Renshaw cells mutually inhibit each other. Furthermore, Renshaw cell activity is modulated by sensory inputs and pathways descending from supraspinal sources (Katz and Pierrot-Deseilligny 1999; Windhorst 2021), which may explain the differences between cat and human results.
3.2.3.5. Reciprocal Inhibition
The effects of muscle fatigue on reciprocal Ia inhibition have not been intensively investigated. Here are two examples.
Ten healthy subjects performed intermittent isometric voluntary contraction of the ankle plantarflexion at 50% MVC as the fatiguing task. Reciprocal Ia inhibition was evaluated by the degree of H-reflex amplitude depression in the soleus muscle by the test stimulus following conditioning stimulus to the common peroneal nerve. The difference in H-reflex amplitude between before and after fatiguing task was also checked. There was no significant difference in the degree of H-reflex amplitude depression, although the H-reflex amplitude significantly decreased after the fatiguing task (p < 0.01).This has been interpreted to show that the decrease in H-reflex amplitude was caused by descending inhibitory input from supraspinal sources to MNs, while the excitability of the Ia inhibitory interneuron was not involved (Tanino et al.2004).
Reciprocal Ia inhibition from ankle flexors to extensors was investigated in 12 healthy subjects. The H-reflex in the soleus muscle was used to monitor changes in the amount of reciprocal Ia inhibition from common peroneal nerve as demonstrated during voluntary dorsi- or plantar-flexion and 50 Hz electrical stimulation induced dorsi- or plantar-flexion. The test soleus H-reflex was kept at 20-25% of maximum directly evoked motor response (M response) and the strength of the conditioning common peroneal nerve stimulation was kept at 1.0 x motor threshold. At rest, weak la inhibition was demonstrated in 12 subjects, maximal inhibition from the common peroneal nerve was 28.8%. During voluntary dorsiflexion and 50 Hz electrical stimulation induced dorsiflexion, the absolute amounts of inhibition increased as compared to at rest, and decreased or disappeared during voluntary plantar-flexion and 50 Hz electrical stimulation induced plantar-flexion as compared to at rest. During voluntary or electrical stimulation induced agonist muscle fatigue, the inhibition of the soleus H-reflex from the common peroneal nerve was greater during voluntary dorsi-flexion (maximal, 11.1%) and 50 Hz (maximal, 6.7%) electrical stimulation induced dorsi-flexion than at rest. The inhibition was decreased or disappeared during voluntary plantar-flexion 50 Hz electrical stimulation induced plantar-flexion. These results were considered to support the hypothesis that MNs and la inhibitory interneurons link to antagonist MNs in reciprocal inhibition (Sato et al. 1999).
Comments. If, as for simplicity depicted in Figure 2, only calf muscles were fatigued, selective effects from activated calf group III/IV afferents on reciprocal Ia inhibitory interneurons to antagonist MNs might be of little importance. But whether group III/IV afferents are so selective is by no means clear, calf afferents could also impact the antagonist-related reciprocal Ia interneurons. Furthermore, reciprocal Ia interneurons are inhibited by Renshaw cells (Figure 2). The matter becomes even more complicated by mutual inhibition between reciprocal Ia interneurons (not shown) and when antagonist muscles are active, e.g., during safety stance on a slippery surface. Finally, they receive modulatory inputs from sensory afferents (e.g., disynaptic inhibition from antagonist group Ia afferents, and excitatory and inhibitory effects from cutaneous, joint and groups II, III and IV muscle afferents), from propriospinal neurons, as well as from descending tracts [e.g., CST, VST, RST, ReST]. These influences may also be exerted indirectly via fusimotor neurons (Jankowska 1992; Schomburg 1990). The descending influences will become important below.
3.3. Supraspinal Muscle Fatigue in Health
Neuromuscular fatigue impairs exercise performance through central and peripheral mechanisms. Interactions between the two components can occur via feedback and feedforward processes. Feedback mechanisms, group III/IV afferents link limb muscles with the CNS. As to central mechanisms, corollary discharges copy the neural drive from the motor system to the working muscles and to sensory systems, which might influence fatigue. There is evidence for a previously proposed hypothesis based on the idea that a negative feedback loop operates to protect the exercising limb muscles from severe threats to homeostasis during whole-body exercise. The `sensory tolerance limit’ can be viewed as a negative feedback loop which accounts for the sum of all feedback (locomotor muscles, respiratory muscles, organs, and muscles not directly involved in exercise) and feedforward signals processed within the CNS with the purpose of regulating the intensity of exercise to ensure that voluntary activity remains tolerable (Hureau et al. 2018).
The following discussion will ascend from the spinal cord to the cerebral cortex.
3.3.1. Fatigue-Induced Effects in Supraspinal Structures
Nociceptive and other signals carried by group III/IV afferents are conveyed to supraspinal structures via several distributed ascending tracts. The main spinally ascending pathway is the spino-thalamic tract (STT), which projects to the posterior, medial and lateral thalamus, different nuclei of which project on to different cortical areas from the somatosensory cortices anteriorly to insular and cingulate cortices. STT collaterals target brainstem areas such as various nuclei in the reticular formation (RF), parabrachial nucleus (PBN), peri-aqueductal gray (PAG), hypothalamus and amygdala (Kuner and Kuner 2021).
In anaesthetized rats, muscle fatigue induced by intermittent high-rate electrical stimulation of dorsal neck muscles (mm. trapezius and splenius) produced significant increases in c-fos expression and NADPH-diaphorase reactivity ipsilaterally in the C1-C4 spinal segments as well as in ventro-lateral PAG, the contralateral central (Ce) and medial (Me) nuclei of the amygdala, and the paraventricular nucleus of hypothalamus. Hence, the PAG, limbic structures and the hypothalamus are involved in activation after neck muscle fatigue and might contribute to nociceptive processing and generation of the autonomic and affective components of muscle pain (Vlasenko et al. 1994).
3.3.1.1. Medulla Oblongata
Group III/IV afferents contact DH neurons, which transmit nociceptive, fatigue-related and other signals upwards. Some targets lie in the medulla oblongata.
In anesthetized rats, Fos-i and NADPH-d-positive neurons in the medulla were studied after fatiguing stimulation of hindlimb muscles. After both direct muscle stimulation and L5 VR stimulation, fatigue-related c-fos gene expression was most prominent within the ipsilateral nucleus tractus solitarii (NTS), the caudal ventro-lateral and rostral ventro-lateral reticular nuclei, and the intermediate reticular nucleus. Fos-ir neurons were co-distributed with NADPH-d-reactive cells within the dorso-medial medulla (DMM) and ventro-lateral medulla (VLM), and double-staining neurons were found in the NTS, intermediate reticular nucleus and lateral paragigantocellular nucleus (Maisky et al. 2002).
In response to fatiguing contractions of the triceps-surae muscles in cats, immune-reactive neurokinins were released in the lateral reticular nucleus, ventral regions of the NTS and the medial vestibular nucleus, suggesting that tachykinin neurons may be a component of the pathways regulating blood pressure during ergoreceptor activation (Williams et al. 1995). In response to fatiguing isometric contractions, immunoreactive substance P-like substances (irSP) were released from sites in the medial NTS (mNTS) and interacted with NK-1 receptors in the area to affect the cardio-vascular responses during the muscle pressor reflex (Williams and Fowler 1997).
3.3.1.2. Peri-Aqueductal Gray (PAG)
The PAG is a cell-dense region surrounding the midbrain aqueduct. It shows a high degree of anatomical and functional organization, which takes the form of longitudinal columns of afferent inputs, output neurons and intrinsic interneurons (Bandler and Shipley 1994; Koutsikou et al. 2017). The PAG receives ascending sensory inputs from nociceptive and thermo-sensitive fibers of the STT, the nociceptive inputs being relayed by the PBN (Kuner and Kuner 2021). The ventro-lateral (vlPAG) column receives direct input from the spinal DH and evokes an opioid-mediated analgesia together with a distinct set of coping behaviors including quiescence. Conversely, the lateral PAG (lPAG) receives direct inputs from the DH and SpV. This organization is consistent with the behavioral response when activated with rostral lPAG stimulation evoking forward defense and caudal lPAG stimulation evoking a fleeing response (Mills et al. 2021). The lPAG and dorso-lateral PAG (dlPAG) are involved in defense reactions associated with tachycardia, hypertension, and re-distribution of bloodflow by activation of sympathetic preganglionic fibers to relevant effectors. By contrast, the ventro-lateral column inhibits sympatho-excitatory neurons of the RVLM and activate the pre-ganglionic vagal neurons, thus initiating passive coping responses with immobility, bradycardia and hypotension. Active coping associated with sympatho-excitation goes along with opioid-independent analgesia and increased motor activity (locomotion), while passive coping associated with sympatho-inhibition concurs with opioid-mediated analgesia and motor freezing (Lamotte et al. 2021).
This raises the question whether muscle fatigue could also affect the PAG. Indirect signs exist. In response to fatiguing isometric contractions of the hind-limb muscles in cats, immunoreactive enkephalins were released. During fatiguing contractions, mean arterial blood pressure increased by 76 +/- 9 mmHg above resting and recovery levels. Levels of immuno-reactive enkephalins were elevated in the dlPAG during the isometric contraction when compared to resting levels. It is possible that isometric muscle contraction causes the release of Met-enkephalin-like substances in the PAG (Williams et al. 1992).
3.3.1.3. Amygdala
The amygdala is comprised of different nuclei; the lateral amygdala (LA), baso-lateral (BLA), medial (MeA), baso-medial (BMA) and central (CeA) nuclei and in between, the intercalated cells. The CeA receives direct and indirect nociceptive inputs from the STT and from the PBN (Allen et al. 2021; Kuner and Kuner 2021), and from several other regions. CeA output targets the PAG and the rostral ventro-medial medulla (RVMM), which are critical for mediating behavioral coping responses in the face of threat. Descending modulation of spinal nociceptive processing by CeA-PAG connections has been implicated in anti-nociceptive effects of opioids acting locally in the BLA (Kuner and Kuner 2021).
In anesthetized rats, c-fos gene expression in the cervical spinal cord and amygdala was examined after muscle fatigue caused by intermittent high-rate electrical stimulation of the dorsal neck muscles (m. trapezius and m. splenius). Fatigue-related increases in c-fos expression occurred on the stimulated muscle side in spinal segments C2-C4 (layers 1, 3-5, 7 and 10), bilaterally in the lumbar L4-L6 (layer 1) segments and in contralateral CeA, MeA, and BMA amygdaloid nuclei. A scarce number of staining cells were found within LA and BLA nuclei. The rostro-caudal extent of c-fos expression in the spinal cord supports functional coupling of the cervical and lumbar regions during neck-muscle fatigue development. The distinct c-fos expression in the CeA and MeA nuclei suggests that they may contribute to mediating the neck muscle fatigue-related nociception, autonomic and behavioral responses (Maznychenko et al. 2007).
3.3.1.4. Blood-Pressure and Respiratory Control in Medulla and Amygdala
It has been proposed that cardio-vascular and ventilatory responses to exercise are regulated by (i) a feedforward `central command’ and (ii) a feedback signal from contracting muscles (Amann 2012). Somatosensory feedback, specifically from group III and IV muscle afferents, influences cardio-ventilatory responses to leg rhythmic exercise in humans. Lumbar intrathecal fentanyl injections were used to impair the central projection of spinal opioid receptor-sensitive muscle afferents. With a number of controls, this approach allowed to demonstrate the essential contribution of group III and IV muscle afferent feedback to the cardio-vascular, ventilatory and perceptual responses to rhythmic exercise in humans, even in the presence of unaltered contributions from other major inputs to cardio-ventilatory control (Amann et al. 2010).
Cardio-vascular Regulation. In cats subjected to isometric exercise to fatigue, ergoreceptor signals may be processed through the PAG and the VLM to control arterial blood pressure (Williams et al. 1990). The rostral VLM contains premotor sympatho-excitatory neurons, which are involved in reflex control of blood pressure and responses to internal stressors. The caudal VLM plays a critical role in cardio-vascular regulation and relays reflex influences to medullary and hypothalamic effectors (Benarroch 2020; Sved et al. 2000). The nucleus paragigantocellularis in the VLM has been implicated in cardio-vascular, nociceptive and analgesic functions. It receives afferents from most laminae of the spinal cord, the caudo-lateral medulla, the medial reticular nuclei of the medulla and pons, the contralateral nucleus paragigantocellularis, the NTS, the medial vestibular nucleus, and issues major projections to the pontine LC (Andrezik et al. 1981; van Bockstaele et al. 1989).
Cardio-vascular regulation during high-intensity exercise has been hypothesized implicate the amygdala whose activation acts to limit maximum exercise performance. In Wistar rats subjected to maximum exercise, total running time and cardio-vascular responses were compared before and after bilateral CeA lesions, activated neurons were recorded in CeA and/or the hypothalamic paraventricular nucleus (PVN) that project to the nucleus tractus solitarii (NTS). (i) CeA lesions resulted in an increase in the total exercise time and the time at which an abrupt increase in arterial pressure appeared, indicating an apparent suppression of fatigue. (ii) High-intensity exercise activated both the PVN-NTS and CeA-NTS pathways. (iii) These suggests that cardio-vascular responses during high-intensity exercise are affected by CeA activation, which acts to limit maximum exercise performance, and may implicate autonomic control modulating the PVN-NTS pathway via the CeA (Tsukioka et al. 2022).
Respiration. The mammalian medulla oblongata contains a network (central pattern generator) that generates the rhythmic respiratory activity, with the core circuitry residing in the VLM and a small nucleus in the pons. This network receives external inputs from pulmonary and chemo-receptive afferents, which elicit a number of respiratory reflexes which regulate ventilation and protect the airways (Lalley 2009; McCrimmon and Alheid 2009, Richter and Smith 2014).
This activity must be able to adapt to various conditions, particularly physical exercise. Evidence indicates that group III/IV muscle afferents indirectly affect locomotor performance by influencing neuromuscular fatigue. These neurons regulate the hemodynamic and ventilatory responses to exercise and, thus, assure appropriate locomotor muscle O2 delivery, which optimizes peripheral fatigue development and facilitates endurance performance (Amann et al. 2020).
3.3.1.5. Hypothalamus
The hypothalamus is a diencephalic structure in the basal forebrain, consisting of several nuclei that are critical for integrated autonomic and neuro-endocrine responses for homeostasis and adaptation to internal or external stimuli (Takayanagi and Onaka 2021). The hypothalamus receives converging nociceptive and visceral inputs from the spinal and trigeminal DHs, and direct and indirect (via PBN and NA A1 cells) nociceptive inputs from the STT (Kuner and Kuner 2021).
Ill-planned intense and exhaustive exercise may have deleterious effects. The effects of a short-duration exhaustive exercise on body chemical composition and hypothalamic-pituitary-adrenal (HPA) axis was investigated in C57Bl/6 mice. Exhaustive exercise consisted of a daily running session at 85 % of maximum speed until the animal reached exhaustion. Body weight as well as total body water, fat and protein content were determined from animal carcasses. HPA activation was assessed by plasma corticosterone levels measured by radioimmunoassay and the weight of both the adrenal glands and thymus were measured. Plasma corticosterone levels increased by 64%. The weight of the adrenal glands augmented by 74% and 45%, at 4 and 10 days of IE, respectively. The total carcass fat content decreased by 20% only at 4 days exhaustive exercise, whereas protein content decreased by 20%. Exhaustive exercise may be related to HPA-axis activation associated with remodeling of body chemical composition in C57BL/6 mice (Rosa et al. 2014).
3.3.1.6. Cerebellum
The cerebellum has been implicated in a number of functions, including oculomotor control, control of upright stance and locomotion, reaching and grasping and speech, timing and coordination of movement, control of motor-cortex excitability, prediction of sensory consequences of actions, error detection and correction, motor learning, classical conditioning (e.g. eyeblink conditioning), and even reward, language, and social behavior, emotional, motivational and cognitive functions. The cerebellum also has a role in pain processing and/or modulation, possibly due to its extensive connections with the prefrontal cortex (PFC) and brainstem regions involved in descending pain control (Adamaszek et al. 2017; Wang et al. 2022).
Nociceptive signals reach the cerebellum, among other structures (below),, and thus fatigue-related signals could also do so.
Alterations in sensory input due to neck muscle fatigue impact upper-limb sensory-motor processing, suggesting that neck fatigue may also impact cerebellum-to-motor cortex pathways in response to motor learning. Normally motor learning decreases cerebellar inhibition (CBI) to facilitate learning of a novel skill. Cervical extensor-muscle fatigue altered cerebellar inhibition in response to motor learning in that neck fatigue before motor skill acquisition led to less decrease in cerebellar inhibition and significantly less improvement in performance accuracy compared to a control group. Neck fatigue impacted the cerebellar-motor cortex interaction to distal hand muscles (Zabihhosseinianet al. 2020).
3.3.1.7. Cerebral Cortex
Group III/IV input reaches the cerebral cortex and has been suggested to inhibit descending drive to MNs by depressing motor-cortex (M1) excitability (Amann 2012; Amann et al. 2020). Another interpretation suggests that this inhibitory effect is counterbalanced by a motivational input that facilitates M1 output in order to overcome supraspinal fatigue (Tanaka and Watanace 2012).
Brain imaging and transcranial stimulation have been used to determine changes in brain areas after fatiguing muscle contractions of various sorts. Results have been somewhat variable.
Functional Magnetic Resonance Imaging (fMRI). Twelve healthy human subjects performed a submaximal (30%) intermittent fatiguing handgrip exercise (3 s grip, 2 s release, left hand) for approximately 9 min during fMRI scanning. Regression analysis was used to measure changes in fMRI signal from primary sensory-motor cortex (S1), premotor cortex (PM) and visual cortex (V1) in both hemispheres. Muscle force declined compared to pre-fatigue maximal force. The fMRI signal from S1 contralateral to the fatiguing hand increased compared to baseline. The fMRI signal from the ipsilateral S1 did not change significantly. The signal from V1 increased significantly for both hemispheres compared to baseline. This is in keeping with the notion of an increase in sensory processing and corticomotor drive during fatiguing exercise to maintain task performance as fatigue develops (Benwell et al. 2007).
Using fMRI in humans performing index-finger abduction tasks, several motor areas in the brain showed increased activity with increased muscle activity, both during force modulation and motor fatigue. The cerebellum showed a smaller increase in activation during compensatory activation due to fatigue, while additional activation was found in the pre-supplementary motor area and in a frontal area. During motor fatigue, force production decreased, force variability increased, and muscle activity increased. Brain areas comparable with the aforementioned areas also showed stronger activation over time. During fatigue, the subjects’ MVC force was reduced, accompanied by a decrease in activation of the supplementary motor area (sma). This suggests that especially the activity in the sma and frontal areas is affected by motor fatigue (van Duinen et al. 2007).
Human subjects showed differences in fMRI-measured cortical neuronal activation before and after different long-lasting contractions: right-hand movements with minimum, maximum, and post-fatigue maximum finger flexion. As compared to maximum movement, the total number of active voxels in primary sensory-motor (S1) areas, sma and cerebellum was strongly reduced in post-fatigue maximum movement (Storti et al. 2014).
Motor Cortex Stimulation. Single-pulse TMS in humans revealed a reduction in motor-cortex excitability when measured in relaxed muscle following sustained fatiguing contractions. The reduction occurred in motor-evoked potentials. During maximal and sub-maximal fatiguing contractions, voluntary activation measured by TMS decreased, suggesting the presence of supraspinal fatigue. This does not eliminate the possibility of spinal contributions. Comparing EMG responses to paired-pulse stimuli at the cortical and sub-cortical levels suggested that impaired MN responsiveness rather than intra-cortical inhibition may contribute to the development of central fatigue (Gruet et al. 2013).
There is evidence that the cortical inhibition is mediated by intra-cortical interneuron activity, which may contribute to central fatigue, and that spinal opioid receptor-sensitive muscle afferents might influence central fatigue by facilitating intra-cortical inhibition (Hilty et al. 2011). Indeed, feedback from group III/IV muscle afferents innervating locomotor muscles diminished the excitability of the motor cortex during fatiguing cycling exercise. This result is, at least in part, from the facilitating effect of these afferents on inhibitory GABAB intra-cortical interneurons (Sidhu et al. 2018).
In humans, whole body exercise (e.g., such as cycling or rowing) did not alter the net excitability of the cortico-spinal pathway during fatigue. This lack of an apparent effect does not mean that changes do not occur, but that there may be a counterbalance of excitatory and inhibitory influences on the two components of the cortico-spinal pathway, namely the motor cortex and the spinal MNs (Weavil and Amann 2018).
3.3.1.8. Cortico-Cerebello-Basal Ganglia-Thalamic System
The cerebral cortex, BG, cerebellum and thalamus are intimately related by being reciprocally connected with each other (Figure 3). It thus appears that the cortex works in concert with the thalamus, cerebellum and BG, constituting an integrated cerebello-BG-thalamo-cortical system and working in learning and control processes (Caligiori et al. 2017). The neocortex itself communicates bidirectionally and in simple and complex ways with the thalamus, forming cortico-thalamo-cortical loops, tightly interlinked with local cortical and cortico-cortical circuits and involving (massive) excitatory and inhibitory connections. The cerebellum and BG participate in multi-regional loops via their connections with the cortex and thalamus (Shepherd and Yamawaki 2021; Figure 3).
Two pathological examples may serve as support for the network operation.
Dystonia is a brain disorder characterized by sustained involuntary muscle contractions. It is typically inherited as an autosomal dominant trait with incomplete penetrance. Primary dystonia is considered to involve micro-structural and functional changes in neuronal circuitry (Argyelan et al. 2009). Dystonia is a good example of how abnormal activity in the cerebellum may propagate to the BG and cause dysfunction. In many forms of dystonia, abnormalities occur in the cerebello–thalamo–cortical pathways, cerebellar activity and functional connectivity. For example, as compared to controls, manifesting and non-manifesting carriers of genetic mutations that are associated with dystonia showed co-varying increases in metabolic activity in the putamen–GP and in the cerebellar cortex. Mouse models of dystonia also displayed structural and functional abnormalities in cerebello-thalamo-cortical pathways. In such models, abnormal (for example, bursting) cerebellar output evoked dystonic postures. Silencing, lesioning or normalizing cerebellar output abolished the dystonic postures. In keeping with these data, bursting cerebellar output has been recorded during surgery in a patient with dystonia (Bostan and Strick 2018). Magnetic resonance diffusion tensor imaging showed reduced integrity of cerebello-thalamo-cortical fiber tracts. In these subjects, reductions in cerebello-thalamic connectivity correlated with increased motor activation responses, consistent with loss of inhibition at the cortical level (Argyelan et al. 2009).
Figure 3.
Simplified block diagram of the cortico-cerebello-basal ganglia-thalamic system. The intricate circuitries within the cerebral cortex, BG, cerebellum and thalamus are omitted for simplicity. The cerebral cortex and thalamus interact in complex ways, involving the cerebral and thalamic networks with excitatory and inhibitory connections, where the purple double-arrowed lines symbolize excitatory projections which, however, most often also exert inhibitory actions by GABA interneurons in the target structure. The cortico-BG connection (the striatum being the BG input station) is direct and excitatory. The cortico-cerebellar connection is mediated via the pontine nuclei (PN). At least in part, the BG-to-cerebellum connection also runs through the PN, while the inverse cerebellum-to-BG connection travels via the thalamus. The pathways from BG and cerebellum to the cerebral cortex involve the thalamus. The red line symbolizes inhibitory effects exerted by the BG output station, the GPi/SNr. Green arrowed lines symbolize excitatory connections. Abbreviations: CL, centro-lateral nucleus; CM: centro-median nucleus; DCN: deep cerebellar nuclei; GPi: globus pallidus internus; PN: pontine nuclei; SNr: substantia nigra pars reticularis; STN: subthalamic nucleus; VA: ventro-anterior nucleus; Vim: ventro-intermediate nucleus; VL: ventro-lateral nucleus. Data from Bostan and Strick (2018); Shepherd and Yamawaki (2021); Wichmann and DeLong (2016).
Figure 3.
Simplified block diagram of the cortico-cerebello-basal ganglia-thalamic system. The intricate circuitries within the cerebral cortex, BG, cerebellum and thalamus are omitted for simplicity. The cerebral cortex and thalamus interact in complex ways, involving the cerebral and thalamic networks with excitatory and inhibitory connections, where the purple double-arrowed lines symbolize excitatory projections which, however, most often also exert inhibitory actions by GABA interneurons in the target structure. The cortico-BG connection (the striatum being the BG input station) is direct and excitatory. The cortico-cerebellar connection is mediated via the pontine nuclei (PN). At least in part, the BG-to-cerebellum connection also runs through the PN, while the inverse cerebellum-to-BG connection travels via the thalamus. The pathways from BG and cerebellum to the cerebral cortex involve the thalamus. The red line symbolizes inhibitory effects exerted by the BG output station, the GPi/SNr. Green arrowed lines symbolize excitatory connections. Abbreviations: CL, centro-lateral nucleus; CM: centro-median nucleus; DCN: deep cerebellar nuclei; GPi: globus pallidus internus; PN: pontine nuclei; SNr: substantia nigra pars reticularis; STN: subthalamic nucleus; VA: ventro-anterior nucleus; Vim: ventro-intermediate nucleus; VL: ventro-lateral nucleus. Data from Bostan and Strick (2018); Shepherd and Yamawaki (2021); Wichmann and DeLong (2016).
Tourette Syndrome (TS) and Obsessive–compulsive Disorder (OCD). Although the precise origin of tics in TS patients is still unclear, reduced GABA function in the striatum may be a primary cause of tic production. Microinjections of the GABA antagonist bicuculline into the striatum or into the globus pallidus externus of non-human primates can result in tics and stereotyped behaviors. Moreover, abnormal BG activity drives activity changes in both the cerebellum and the primary motor cortex (M1) that predict the onset of tics. Imaging studies in humans provided additional evidence for cerebellar involvement in TS, which revealed tic-related activation not only in the BG but also in the cerebellum. Brain imaging of both TS and OCD patients revealed changes in cerebellar activity. For example, relative to healthy controls, resting-state fMRI demonstrated that individuals with OCD exhibited increased whole-brain connectivity in regions of both the putamen and the cerebellar cortex. Also, in individuals with TS, brain metabolism was characterized by hypometabolism in the striatum co-varying with hypermetabolism in the cerebellar cortex. It has been proposed that the disynaptic pathway from the STN to the cerebellar cortex (Figure 3) may provide a link for such effects, although other pathways or independent structural changes in the cerebellum may also be involved. A recent computational model of BG–cerebellar–cerebral cortical network function in TS indicated that hyperactivity in the STN may propagate to the cerebellar cortex and contribute to tic generation (Bostan and Strick 2018).
Healthy Human Volunteers. Alterations of cerebral activity during prolonged static force exertion was determined by regional cerebral blood flow (rCBF) using H2(15)O positron emission tomography (PET) while six male healthy subjects pressed a morse-key with their right index finger with a constant force of 20% of MVC for different periods of time (1.5-4.5 min). Despite a considerable sense of fatigue and increased effort at the end of a 4.5 min. key press, no compensatory changes of activity were detected in motor or sensory related structures. In the right dorso-lateral PFC, the rCBF was significantly correlated with the duration of key-press, possibly reflecting processes overriding fatigue. In this static force task, the basal ganglia (BG) were strongly activated, but not in a previous force task involving repetitive dynamic force pulses, suggesting that sustained exertion of a static force is an active process modulated, at least in part, by the BG (Dettmers et al. 1996).
Healthy human volunteers performed a sustained handgrip contraction for 225 s and 320 intermittent handgrip contractions (approximately 960 s) at 30% maximal level, while fMRI was used to image their brain. For the sustained contraction, EMG signals of the finger flexor muscles increased linearly while the target force was maintained. The fMRI-measured cortical activities in the contralateral sensory-motor cortex increased sharply during the first 150 s, then plateaued during the last 75 s. For the intermittent contractions, the EMG signals increased during the first 660 s and then began to decline, while the handgrip force also showed a sign of decrease despite maximal effort to maintain the force. The fMRI signal of the contralateral sensory-motor area showed a linear rise for most part of the task and plateaued at the end. For both tasks, the fMRI signals in the ipsilateral sensory-motor cortex, PFC, cingulate gyrus, sma, and cerebellum exhibited steady increases. The brain thus increased its output to reinforce the muscle for the continuation of the performance and possibly to process additional sensory information (Liu et al. 2003). These results are somewhat at variance with those reported by Dettmers et al. (1996).
Healthy human subjects executed simple motor tasks before and under motor fatigue. fMRI showed that in both conditions, movements activated sensory-motor areas, sma, cerebellum, thalamus and BG. Importantly, the intensity and size of activation volumes in the sub-cortical areas, including thalamus and BG areas, were significantly decreased during motor fatigue, implying that fatigue disturbs the motor-control processing in a way that both sensory-motor areas and sub-cortical brain areas are less active (Hou et al. 2016).
Functional Connectivity in Muscle Fatigue. During muscle fatigue induced by intermittent handgrips at 50% of their MVC level, functional connectivity between fMRI signals of the M1 and the BG, cerebellum, and thalamus were determined. Widespread, statistically significant increases in functional connectivity occurred in bilateral BG, cerebellum, and thalamus with the left M1 during significant versus minimal fatigue stages. These sub-cortical nuclei are critical components in the motor control network and actively involved in modulating voluntary muscle fatigue, possibly, by working together with the M1 to strengthen the descending central command to prolong the motor performance (Jiang et al. 2016).
3.3.2. A Long Way Down: From Cerebral Cortex to Spinal Cord
3.3.2.1. Insufficient Drive from Motor Cortex
Even maximal voluntary effort to contract a muscle or group of muscles activates human MNs and muscle fibers only sub-optimally, maximal voluntary strength thus often being below true maximal muscle force. This could be demonstrated by TMS over the motor cortex. For example, when stimulus intensity is set appropriately, TMS during an isometric MVC of the elbow flexors commonly evoked a small twitch-like increment in flexion force, indicating that, despite the subject’s maximal effort, motor cortical output at the moment of stimulation was not maximal and was not sufficient to drive the MNs to produce maximal force from the muscle. TMS has uncovered focal changes in cortical excitability and inhibitability based on EMG recordings, and a decline in supraspinal `drive’ based on force recordings. The mechanisms of supraspinal fatigue are unclear. Although changes in the behavior of cortical neurons and spinal MNs occur during fatigue, they can be dissociated from supraspinal fatigue. Furthermore, in whole-body endurance performance, the brain monoaminergic neurotransmitter systems play a role, particularly with regard to exercise in hot environments (below) (Gandevia 2001; Taylor et al. 2016).
However, things are a bit more complicated. The properties of the cortico-spinal pathways change during fatiguing exercise in ways that could influence the development of central fatigue. Based on differences in motor cortical and MN excitability between exercise modalities (e.g. single-joint vs. locomotor exercise), the effect of these changes on functional impairments and performance limitations are not clear. There is though strong evidence for marked `inhibition’ of MNs as a direct result of voluntary drive (Amann et al. 2022).
The interaction between the cerebral cortex and the spinal cord also involves, besides `purely’ motor pathways, neuromodulatory systems, including monoaminergic ones.
3.3.2.2. Substantia Nigra Pars Compacta and Dopamine (DA)
In rodents, exercise increases the release of several neurotransmitters in different brain regions and the onset of fatigue can be manipulated when DA influx in the preoptic and anterior hypothalamus is increased, interfering with thermoregulation. In humans, things are not as straightforward: most studies manipulating brain neurotransmission failed to change the onset of fatigue in normal ambient temperatures. When the ambient temperature was increased, DA and combined DA and NA reuptake inhibition appeared to override a safety switch, allowing subjects to push harder and become much warmer, without changing their perception. In general, brain neurochemistry is clearly involved in the complex regulation of fatigue, but many other mediators also play a role (Meeusen and Roelands 2018). Adaptations in the DA systems may influence exercise capacity. For example, a reduction in DA neurotransmission in the substantia nigra pars compacta (SNpc) could impair activation of the BG and reduce stimulation of the motor cortex leading to central fatigue. Habitual wheel running produced changes in DA systems. Six weeks of wheel running sufficed to increase tyrosine hydroxylase mRNA expression and reduce D2 auto-receptor mRNA in the SNpc. The same exercise increased D2 postsynaptic receptor mRNA in the caudate putamen, a major projection site of the SNpc (Foley and Fleshner 2008).
3.3.2.3. Raphé Nuclei and Serotonin (5-HT)
The raphé nuclei and ventral tegmental filed (VTA) are a collection of functionally and anatomically diverse cell groups that span the brainstem and contain the majority of the 5-HT-producing neurons in the CNS (Brodal 1981). Noxious peripheral stimuli cause activity changes in neurons of the RVMM, which contains the nucleus raphé dorsalis (NRD). Raphé neurons are sensitive to fatigue, but differentially.
In adult male cats, single-unit activity of presumed 5-HT neurons in the mesencephalon (NRD) or medulla oblongata [nucleus raphé obscurus (NRO) and nucleus raphé pallidus (NRP)] was recorded during prolonged treadmill locomotion. Treadmill speed was set at a moderate level (0.4 m/s) in order to induce long-duration locomotion. The typical time to `fatigue’ (failure to keep pace, falling behind and reluctance to continue) was approximately 40 min in both groups, at which point cats typically displayed marked panting and vocalization. The activity of NRD neurons was unchanged from baseline during the locomotion trial and during the recovery phase. By contrast, the activity of NRO/NRP neurons decreased steadily across the locomotion trial, reaching a mean decrease of approximately 50% (during the first min after the treadmill was turned off). Full recovery of single unit activity to a level approximating the baseline discharge rate required 30-45 min (Fornal et al. 2006).
5-HT Effects on MNs. Endogenous forms of neuromodulation, e.g., by 5-HT, can directly affect MN output and central fatigue (Amann et al. 2022). 5-HT potently regulates neuronal firing rates, which can influence the force that can be generated by muscles during voluntary contractions. MN discharge can be facilitated or suppressed depending on the 5-HT receptor sub-type activated. The release of 5-HT from descending tracts into the spinal cord is linked to the level of motor activity being performed, where 5-HT can increase the discharge rate of MNs via excitatory 5-HT receptors on the soma and dendrites. This in turn can lead to increased voluntary muscle activation and maximal force generation. However, intense or long-lasting release of 5-HT onto MNs may lead to a spillover of 5-HT into extracellular compartments to activate inhibitory 5-HT receptors on the axon initial segment. This can cause a reduction in MNs discharge rate, thus decreasing voluntary muscle activation and maximal force generation. 5-HT may thus have different effects when neuromuscular fatigue induced by contractions of different intensities. Thus, enhanced levels of 5-HT reduced voluntary activation of muscle when fatigue was induced by strong but not weak contractions (Cotel et al. 2013; Henderson et al. 2022; Kavanagh and Taylor 2022).
3.3.2.4. Locus Coeruleus and Noradrenaline (NA)
The LC is a collection of NA cell groups A5 and A6 in the rostral pons of animals and humans. Noxious peripheral stimuli indirectly increase the activity in LC. In anesthetized rats, LC neurons were potently activated by foot shock (Chiang and Aston-Jones 1993). Whether LC receives fatigue-related signals is not clear.
It has been suggested that NA, but not DA re-uptake inhibition, contributes to the development of central/supraspinal fatigue after a prolonged cycling exercise performed in temperate conditions (Klass et al. 2012). Compared with placebo intake, the NA re-uptake inhibitor reboxetine (REB) reduced the endurance time and the MVC torque at task failure. The level of voluntary activation tested by TMS and electrical stimulation decreased at the end of the task. Although the motor-evoked potential did not change during fatigue, the H-reflex and electrically evoked torque decreased similarly in the placebo and REB conditions. After exercise, the reaction time increased in the REB condition but did not change in the placebo condition. This suggested that because of the NA re-uptake inhibition, the output from the motor cortex is decreased at a greater rate than that in the placebo condition, contributing thereby to shorten endurance time (Klass et al. 2016).
It is mentioned in passing that more neurotransmitters (e.g., NO, GABA) can also influence fatigue (Foley and Fleshner 2008).
3.4. Central Muscle Fatigue in Disease
Signals elicited by group III and IV afferents also reach somatosensory and frontal cortical areas.
3.4.1. Overview
Muscle fatigability is enhanced in various neurological diseases affecting the CNS, such as stroke, amyotrophic lateral sclerosis (ALS), PPS, MS, Parkinson’s disease, traumatic brain injury (TBI), spinal cord injury (SCI), cerebro-vascular disorders, GBS, and COVID-19. Fatigue occurs in diseases that affect the central, peripheral, and autonomic nervous systems, the main characteristics of central fatigue being enhanced perception of effort and limited endurance of sustained physical and mental activities. The pathophysiological process comprises metabolic and structural lesions that disrupt the activation in pathways interconnecting the BG, thalamus, limbic system, and higher cortical centers. A state of pre-existing relative hypo-cortisolemia might sensitize the HPA axis to development of persistent central fatigue after stress. In conclusion, fatigue appears to be a more or less universal symptom in central diseases, but there is a specific pathophysiology that leads to fatigue in different diseases (Chaudhuri and Behan 2004; Finsterer and Mahjoub 2014; Kent-Braun et al. 2012; Penner and Paul 2017; Zwarts et al. 2008).
3.4.2. Age
Fatigue induced by resistance exercises to lower limb and trunk muscles entails postural instability during quiet standing. Fatigue in hip, knee and ankle impairs functional reach, reduces the speed and power of sit-to-stand repetitions, and produces less stable and more variable walking patterns in older people (Helbostad et al. 2010). Accordingly, muscle fatigue deteriorates reactive postural control in the elderly (Papa et al. 2015).
3.4.3. Motoneuron Diseases
Fatigue is a common symptom in motoneuron Diseases, including ALS, spinal muscular atrophy (SMA), and post-polio syndrome (PPS). It has a significant impact on patients' quality of life.
3.4.3.1. Amyotrophic Lateral Sclerosis (ALS)
In ALS, both lower motoneurons (LMN) and upper motoneurons (UMN) are typically affected. However, muscle weakness is largely due to LMN loss. While the inability of MUs to maintain a given level of activity leads to the development of peripheral fatigue, dysfunction of the UMN leads to reduced voluntary drive and subsequent central fatigue. To address the question of the contribution of central muscle fatigue in ALS, measurements of isometric force, activation, and energy metabolism of the ankle dorsiflexor muscle were combined (Kent-Braun and Miller 2000). Compared to healthy controls, ALS patients experienced a significant failure of central motor activation during a fatiguing isometric exercise protocol. Only individuals in the ALS group had a remarkable increase in added force in response to a tetanic stimulus imposed during maximal voluntary contraction. TMS studies provide evidence that maladaptation of inhibitory cortical circuits contributes to the development of fatigue in ALS (Nardone et al. 2005; Vucic et al. 2011).
3.4.3.2. Spinal Muscular Atrophy (SMA)
The most common form of SMA is caused by a mutated survival motor neuron gene 1 (SMN1). Insufficient levels of the SMN protein leads to loss of LMN and causes weakness and wasting of various muscles. As with most patients suffering from neuromuscular diseases, fatigue is one of the common symptoms during SMA patients' daily activities and has a negative impact on the overall quality of life. There is evidence that fatigue occurs in SMA patients is associated with reduced muscle strength (Bartels et al. 2021). However, studies with prolonged exercise show that the abnormal performance fatigability is an additional aspect of attenuated motor function in SMA (Granger et al. 1999; Montes et al. 2010). Regarding the distinction between peripheral and central fatigue, there is no clear understanding of the possible impacts of SMN1 mutation-related peripheral cellular and metabolic changes on the central components of fatigue (Crisafulli et al. 2024).
3.4.3.3. Post-Polio Syndrome (PPS)
A significant proportion of poliomyelitis survivors may experience renewed deterioration in motor function decades after the acute illness, and this late deterioration is called PPS. In addition to an increase in weakness, patients with PPS often complain of fatigue and pain. A physiological study of the central and peripheral fatigue revealed the presence of impaired voluntary activation as a central deficit that likely contributes to attenuated motor function in PPS (Allen et al. 1994).
3.4.4. Spinal Cord Injury (SCI) and Traumatic Brain Injury (TBI)
Peripheral and central fatigue in patients with incomplete cervical cord injury was analyzed, among other parameters, in a neurophysiological study testing the voluntary activation level, MN excitability and fatigue index of an arm muscle (Lin et al. 2012). Patients with SCI showed a reduction in MVC and voluntary activation level and an increase in MN excitability compared to healthy controls. Only central fatigue was observed in patients with SCI, not peripheral fatigue. One of the study's key conclusions was that central drive deficit is an important cause of muscle weakness and fatigue in patients with spinal cord injury. Fatigue is also a common and debilitating phenomenon in patients with TBI. Some endocrine or inflammatory changes in the brain, such as growth hormone deficiency due to post-traumatic hypopituitarism (Mossberg et al. 2008; Zgaljardic et al. 2014) or reduced NO-cGMP pathway signaling (Zgaljardic et al. 2014), contribute to impaired muscle function after TBI.
3.4.5. Stroke and Spasticity
Fatigue is a common, debilitating symptom in patients who have suffered a stroke. Neuromuscular fatigue has central and peripheral causes, with stroke patients experiencing greater central fatigue and less peripheral fatigue during voluntary contractions of the paretic leg compared to healthy controls (Knorr et al. 2012). Etiological factors and infarct volume do not appear to be associated with poststroke fatigue (PSF), but involvement of the subcorticofrontal network seems to be involved in the development of PSF (Ponchel et al. 2015).
Spastic Paraparesis. In patients with spastic paraparesis and in normal controls, intermittent tetanic contractions and 31P magnetic resonance spectroscopy showed that, in the tibialis anterior (TA) muscle, the decline in TA tension was significantly greater, the half-relaxation time of the tetanus was more prolonged, and the decline in phospho-creatine and intracellular pH was significantly greater in patients than in controls. This suggests that biochemical changes in the muscles of patients with spastic paraparesis may contribute to their excessive fatigability (Miller et al. 1990). In patients with progressive paraparesis, there is evidence for decreased recurrent inhibition at rest, which may contribute to the exaggeration of the passively induced stretch reflex underlying spasticity (Katz and Pierrot-Deseilligny 1999).
3.4.6. Parkinson’s Disease (PD)
Fatigue is also very common in patients with PD. It may even precede the development of typical motor symptoms such as rigidity, tremor, and bradykinesia by several months (Chaudhuri and Behan 2000). Central fatigue in Parkinson's disease is related to DA deficiency. Accordingly, levodopa normalizes the increased excitability of UMN that accompanies motor fatigue (Lou et al. 2003). Regarding atypical Parkinson's syndromes, patients with multiple system atrophy present with marked central fatigue that is often more disabling than Parkinsonian or dysautonomic symptoms (Chaudhuri and Behan 2000). Apathy and fatigue are considered to be some of the earliest manifestations of progressive supranuclear palsy, another atypical Parkinson's syndrome (Chaudhuri and Behan 2000).
3.4.7. Multiple Sclerosis (MS)
MS is a chronic CNS disease characterized by inflammation, demyelination and neurodegeneration, with fatigue being an important symptom that can precede MS diagnosis by years. MS patients differ from healthy persons in motor variables (e.g., muscle force, walking ability, gait parameters, activity level) and neuropsychological variables (Powers et al. 2016). Patients with MS and SCI showed greater fatigability of their tibialis anterior (TA) muscles. During repetitive stimulation, there was a decrement in tension and an increase in half-relaxation time of tetanic contractions at 40 Hz in both subjects and patients, with the increase in half relaxation, being greater in MS and SCI patients than in healthy subjects. It has been suggested that the muscle fiber population changed toward that typical of fatiguable MUs and that the long-term inactivity affected the efficiency of the Ca2+ uptake mechanism of their muscle fibers (Lenman et al. 1989). A hypothesis propounds that fatigue might develop consequent to a DA imbalance. Both too much as too little DA would disrupt the meso-cortico-limbic pathway connecting the VTA, striatum, limbic area and prefrontal cortex (Powers et al. 2016).
Fatigue is not only one of the most common but also one of the most debilitating symptoms of MS. Fatigue affects the quality of life and ability to work. Central fatigue in MS is associated with dysfunction of the frontal cortex and basal ganglia as well as their adjacent white matter (Roelcke et al. 1997). A later imaging study confirmed and extended the findings, linking fatigue to disruption of brain networks involved in attention processes (Sepulcre et al. 2009).
Central fatigue in MS is partially mediated by the activation of proinflammatory cytokines (Heesen et al. 2006). Interestingly, MS patients with fatigue showed higher activity of the HPA axis than patients without fatigue, so it can be assumed that the HPA axis does not appear to play a role in the development of central fatigue in MS (Gottschalk et al. 2005). Accordingly, another study suggests that MS-related fatigue is not due to daily cortisol secretion (Malekzadeh et al. 2020). Some studies showed that fatigue as MS progresses may be related to low serum levels of dehydroepiandrosterone (Keller et al. 2021).
3.4.8. Guillain-Barré Syndrome (GBS)
Among neuromuscular diseases, fatigue is mainly described in patients with PPS, myasthenia gravis and immune-mediated neuropathies such as GBS (Zwarts et al. 2008). In neurologically well-recovered GBS patients examined in a physiological study, central fatigue occurred in the first minute of sustained MVC, while it could not be detected in healthy individuals (Garssen et al. 2007). This result differed from the findings on peripheral fatigue, which, interestingly, was even lower in the GBS patients mentioned than in the healthy individuals.
3.4.9. Fibromyalgia and Chronic Fatigue Syndrome (CFS)
As can be easily seen from the descriptions above, fatigue is a well-known manifestation of a number of somatic disorders. Fatigue also occurs as a side effect of pharmacological agents such as chemotherapy drugs and has also been described in various psychiatric disorders such as depression (Zwarts et al. 2008). In addition, there is a group of severely fatigued patients who have no somatically recognizable disease. These include fibromylagia patients and patients with CFS. Both diseases are characterized by severe central fatigue accompanied with various symptoms that affect daily life, such as pain, impaired memory, and depression. Similar to ALS patients, decreased central activation in the ankle dorsiflexor muscle was described in CFS patients (Kent-Braun et al. 1993). Similar results were obtained when examining the elbow flexors (Schillings et al. 2004).
There is no specific test to support the clinical diagnosis of fibromyalgia or CFS. To date, the search for biomarkers has rarely been successful and the biomarkers found often have low specificity. A recent fMRI study could interestingly show that increased activity in the anterior node of the default mode network can be considered a biomarker for post-exertional malaise in CFS patients (Rayhan and Baraniuk 2021).
3.4.10. COVID-19 Neuromuscular Symptoms
The manifestation of COVID-19 can range from mild to severe and can be limited in time (weeks) or persist for months in about 30-50% of patients. Long-COVID can affect the whole spectrum of people with COVID-19 with very mild acute disease to the most severe forms. Long-COVID can involve multiple organs and can affect many systems including the cardio-vascular, respiratory, gastro-intestinal, neurological and musculo-skeletal systems. The symptoms include muscle pain and fatigue, dyspnea, cardiac abnormalities, cognitive impairment, sleep disturbances, symptoms of post-traumatic stress disorder, concentration problems, and headache. Musculo-skeletal manifestations include loss of muscle mass, muscle weakness, fatigue or myalgia, and muscle injury (Crook et al. 2021; Meacci et al. 2022).
COVID-19 patients may also suffer from polyneuropathy, paresthesia, and GBS. Corticoid therapy is also considered important in muscle fatigue and functional incapacity. Recently, small-fiber neuropathy has been identified as a potential cause that, in some long-COVID patients, may underlie autonomic dysfunction and paresthesia (Abrams et al. 2023; Costa Silva et al. 2022; Pescaru et al. 2022).
Central fatigue is a major symptom and represents a long-term health impairment following a COVID-19 infection. Recent psychometric measurements found evidence of brain fog and central fatigue in COVID-19 survivors (Hsiao et al. 2024). A TMS study suggested that GABAergic impairments could be the basis for central fatigue in these patients (Ortelli et al. 2021), a fact that is reminiscent of the above-mentioned maladaptation of inhibitory cortical circuits that contributes to the development of central fatigue in ALS.
3.4.11. Yawning-Fatigue Syndrome
Yawning rarely occurs as a major feature in neurological diseases, although it has been observed in some neurological diseases, for example in the prodromal phase of migraine or together with parakinesia brachialis oscitans in stroke patients (Dibaj et al. 2021). Although not necessarily the case, yawning is often associated with central fatigue.
The recently described yawning-fatigue syndrome has a dopaminergic pathophysiology (Dibaj et al. 2020). When the DA agonist apomorphine was administered, the patient experienced yawning attacks, which were followed by severe fatigue. DA triggers yawning by activating D3-receptors in the diencephalon. In contrast to Parkinson's disease, in which DA deficiency in the nigrostriatal pathway leads to central fatigue that is treated by administration of levodopa (see above), dopaminergic stimulation in a correspondingly sensitive hypothalamic system leads to yawning-fatigue syndrome. The finding of a missense mutation in the conserved C2 domain of synaptotagmin 6 in the above-mentioned patient with yawning fatigue syndrome supports the assumption that an alteration in neurotransmitter release is the cause of the patient's symptoms (unpublished data).
3.4.12. Muscle Fatigue and Pain
During sustained muscle contractions, particularly at the limit of endurance, fatigue may occur together with pain, and both can restrict muscle performance. Patients with neuromuscular disorders may have chronic myofascial pain (e.g. fibromyalgia) or contraction-induced pain (e.g. in metabolic myopathies). In some patients, these two types of pain may coexist and both may inhibit central motor drive during exercise. Repetitive magnetic brain stimulation protocols can modulate the excitability of the motor system during exercise to delay the onset of peripheral fatigue, and to reduce chronic pain (Mastaglia 2012).
4. Concluding Remarks
Even if, in a simplistic view, muscle fatigue were considered as nothing more than an exercise-induced loss of force, everyone knows from own experience that this is not true. Muscle force production requires energy to be transformed by biochemical processes into force. Energy must be mobilized and transported to the exercising muscle(s) by means of increases in cardio-vascular and respiratory activities. Then precautions should be taken to limit muscle activities so as not to risk tissue damage. All this requires the organizational capacities of the nervous system, which cannot simply rely on feedforward mechanisms but also depends on feedback from the exercising muscle.
It has been proposed that, in both healthy and pathological populations, prolonged physical exercise leading to muscle fatigue would activate group III/IV muscle sensory afferents, which would then recruit CNS centers to increase heart rate and respiration, modulate spinal reflexes, decrease central motor drive, and thereby contribute to improve muscle performance by regulating peripheral fatigue development and by avoiding excessive muscle impairments. Hence, due to their sensitivity to intramuscular metabolites and mechanical events, group III/IV muscle afferents are multi-functional.
Group III/IV muscle afferents also modulate spinal reflexes fed by large-diameter afferents, i.e., group Ia and II afferents from muscle spindles and group Ib afferents from GTOs. Except for the former two, which connect monosynaptically to MNs, though subject to presynaptic inhibition, all of them also have oligosynaptic connections to MNs mediated by interneurons, onto which they converge in various patterns (Windhorst 2021).
Thus, what exactly group III/IV muscle afferents do at spinal level under natural or pathological conditions, is difficult to tell. “Can sense be made of spinal interneuron circuits?” (McCrea 1992). Only in part because many of these interneurons are “unknown” and integrated in complex networks whose actions remain mysterious, at least to a considerable extent (Windhorst 2021). These networks present an “embarrassment of riches” (Loeb 1987). Most probably, similar situations prevail at supraspinal levels.
These networks can easily be disrupted under pathological conditions, as shown by various neurological diseases. The exact mechanisms are not yet well understood and require much painstaking work.
Acknowledgements
UW is grateful to his wife Sigrid for occasional help and much patience.
Conflict of Interest
The authors declare no conflict of interest.
Author Contributions
Both authors wrote parts of the text, prepared the figures, searched and provided literature, revised and edited the manuscript, and approved the final manuscript.
Funding
This research received no external funding.
Ethics Approval and Consent to Participate
Not applicable.
Abbreviations
ACh: Acetylcholine; ADP: Adenosin-diphosphate; ALS: amyotrophic lateral sclerosis; AP: action potential; ATP: Adenosin-triphosphate; BLA: baso-lateral nucleus of amygdala; BMA: baso-medial nucleus of amygdala; BMD: Becker muscular dystrophy; CDP: cord-dorsum potential; CeA: central nucleus of amygdala; CM: centro-median nucleus of thalamus; CNS: central nervous system; COPD: chronic obstructive pulmonary disease; CrP: creatine phosphate; CST: cortico-spinal tract; DA: dopamine; DCN: deep cerebellar nuclei; DH: dorsal horn; dlPAG: dorso-lateral peri-aqueductal gray; DMD: Duchenne muscular dystrophy; DMM: dorso-medial medulla; ECC: excitation-contraction coupling; EMG: electromyographic, electromyography; EPSP: excitatory postsynaptic potential; fMRI: functional magnetic resonance imaging; Fos-ir: fos-immunoreactive; FST: fatiguing stimulation; GBS: Guillain-Barré Syndrome; GPi: globus pallidus internus; GS: gastrocnemius-soleus muscle; GTO: Golgi tendon organ; HPA: hypothalamic-pituitary-adrenal (axis); HPT: hypothalamic-pituitary-thyroid (axis); 5-HT: 5-hydroxytryptphan, serotonin; H-reflex: Hoffmann reflex; K+: potassium; LC: locus coeruleus; LMN: lower motoneuron; lPAG: lateral peri-aqueductal gray; M1: primary motor cortex; MeA: medial nucleus of amygdala; MELAS: Mitochondrial encephalopathy, lactic acidosis, and stroke-like episodes; MG: medial gastrocnemius muscle; MH: malignant hyperthermia; MN: motoneuron; MS: multiple sclerosis; MU: motor unit; MVC: maximal voluntary contraction; NA: noradrenaline, noradrenergic; NADPH-d: nicotinamide adenine dinucleotide; phosphate-diaphorase; NRD: nucleus raphé dorsalis; NRO: nucleus raphé obscurus; NRP: nucleus raphé pallidus; NTS: nucleus tractus solitarii; OCD: obsessive–compulsive disorder; PAD: primary afferent depolarization; PAG: peri-aqueductal gray; PBN: parabrachial nucleus; PBSt: posterior biceps and semitendinosus; PD: Parkinson’s disease; PET: positron emission tomography; Pf: parafascicul nucleus of thalamus; PFC: prefrontal cortex; Pi : inorganic phosphate; PN: pontine nuclei; PSF: poststroke fatigue; PSI: presynaptic inhibition; PSTH: post-stimulus time histograms; rCBF: regional cerebral blood flow; REB: reboxetine; ReST: reticulo-spinal tract; RF: reticular formation; ROS: reactive oxygen species; RVMM: rostral ventro-medial medulla; SCI: Spinal Cord Injury; S1: primary sensory-motor cortex; SNr: substantia nigra pars reticularis; SR: sarcoplasmic reticulum; sma: supplementary motor area; SMA: spinal muscular atrophy; SMN1: survival motor neuron gene 1; STN: subthalamic nucleus of BG; TBI: traumatic brain injury; TMS: transcranial magnetic stimulation; TS: Tourette syndrome; UMN: upper motoneuron; VA: ventro-anterior nucleus of thalamus; Vim: ventro-intermediate nucleus of thalamus; VL: ventro-lateral nucleus of thalamus; VLM: ventro-lateral medulla; vlPAG: ventro-lateral peri-aqueductal gray; VST: vestibulo-spinal tract; VTA: ventral tegmental filed
References
- Abrams RMC, Zhou L, Shin SC (2023) Persistent post-COVID-19 neuromuscular symptoms. Muscle Nerve 68(4):350-355. [CrossRef]
- Adamaszek M, D’Agata F, Ferrucci R, Habas C, Keulen S, Kirkby KC, Leggio M, Mariёn P, Molinari M, Moulton E, Orsi L, Van Overwalle F, Papadelis C, Priori A, Sacchetti L, Schutter DJ, Styliadis C, Verhoeven J (2017) Consensus Paper: Cerebellum and emotion. Cerebellum 16(2):552-576. [CrossRef]
- Allen DG (2004) Skeletal muscle function: role of ionic changes in fatigue, damage and disease. Clin Exp Pharmacol Physiol 31(8):485-493. [CrossRef]
- Allen DG, Lamb GD, Westerblad H (2008) Skeletal muscle fatigue: cellular mechanisms. Physiol Rev 88(1):287-332. [CrossRef]
- Allen DG, Trajanovska S (2012) The multiple roles of phosphate in muscle fatigue. Front Physiol 3:463. [CrossRef]
- Allen GM, Gandevia SC, Neering IR, Hickie I, Jones R, Middleton J (1994) Muscle performance, voluntary activation and perceived effort in normal subjects and patients with prior poliomyelitis. Brain 117:661-670. [CrossRef]
- Allen HN, Bobnar HJ, Kolber BJ (2021) Left and right hemispheric lateralization of the amygdala in pain. Prog Neurobiol 196:101891. [CrossRef]
- Amann M (2012) Significance of group III and IV muscle afferents for the endurance exercising human. Clin Exp Pharmacol Physiol 39(9):831-835. [CrossRef]
- Amann M, Blain GM, Proctor LT, Sebranek JJ, Pegelow DF, Dempsey JA (2010) Group III and IV muscle afferents contribute to ventilatory and cardiovascular response to rhythmic exercise in humans. J Appl Physiol (1985) 109(4):966-976. [CrossRef]
- Amann M, Sidhu SK, McNeil CJ, Gandevia SC (2022) Critical considerations of the contribution of the corticomotoneuronal pathway to central fatigue. J Physiol (Lond) 600(24):5203-5214. [CrossRef]
- Amann M, Sidhu SK, Weavil JC, Mangum TS, Venturelli M (2015) Autonomic responses to exercise: Group III/IV muscle afferents and fatigue. Auton Neurosci: Basic and Clin 188:19-233. [CrossRef]
- Amann M, Wan H-Y, Thurston TS, Georgescu VP, Weavil JC (2020) On the influence of group III/IV muscle afferent feedback on endurance exercise performance. Exerc Sport Sci Rev 48(4):209-216. [CrossRef]
- Ament W, Verkerke GJ (2009) Exercise and fatigue. Sports Med 39(5):389-422. [CrossRef]
- Andrezik JA, Chan-Palay V, Palay SL (1981) The nucleus paragigantocellularis lateralis in the rat. Demonstration of afferents by the retrograde transport of horseradish peroxidase. Anat Embryol (Berl) 161(4):373-390. [CrossRef]
- Argyelan M, Carbon M, Niethammer M, Ulug AM, Voss HU, Bressman SB, Dhawan V, Eidelberg D (2009) Cerebellothalamocortical connectivity regulates penetrance in dystonia. J Neurosci 29(31):9740-9747. [CrossRef]
- Avela J, Kyröläinen H, Komi PV, Rama D (1999) Reduced reflex sensitivity persists several days after long-lasting stretch-shortening cycle exercise. J Appl Physiol (1985) 86(4):1292-1300. [CrossRef]
- Ball D (2015) Metabolic and endocrine response to exercise: sympathoadrenal integration with skeletal muscle. J Endocrinol 224:R79-R95. [CrossRef]
- Bączyk M, Manuel M, Roselli F, Zytnicki D (2022) Diversity of mammalian motoneurons and motor units. Adv Neurobiol 28:131-150. [CrossRef]
- Bandler R, Shipley MT (1994) Columnar organization in the midbrain periaqueductal gray: modules for emotional expression? Trends Neurosci 7(9):379-389. [CrossRef]
- Bartels B, de Groot JF, Habets LE, Wadman RI, Asselman F, Nieuwenhuis EES, van Eijk RPA, Goedee HS, van der Pol WL (2021) Correlates of fatigability in patients with spinal muscular atrophy. Neurology 96(6):e845-e852. [CrossRef]
- Baudry S, Maerz AH, Gould JR, Enoka RM (2011) Task- and time-dependent modulation of Ia presynaptic inhibition during fatiguing contractions performed by humans. J Neurophysiol106(1):265-273. [CrossRef]
- Bellinger AM, Mongillo M, Marks AR (2008) Stressed out: the skeletal muscle ryanodine receptor as a target of stress. J Clin Invest 118(2):445-453. [CrossRef]
- Benarroch EE (2020) Physiology and pathophysiology of the autonomic nervous system. Continuum (Minneap Minn) 26(1):12-24. [CrossRef]
- Benwell NM, Mastaglia FL, Thickbroom GW (2007) Changes in the functional MR signal in motor and non-motor areas during intermittent fatiguing hand exercise. Exp Brain Res 182(1):93-97. [CrossRef]
- Bigland-Ritchie B (1984) Muscle fatigue and the influence of changing neural drive. Clin Chest Med 5:21-34.
- Bigland-Ritchie B, Woods JJ (1984) Changes in muscle contractile properties and neural control during human muscular fatigue. Muscle Nerve 7(9):691-699. [CrossRef]
- Binder MD, Kroin JS, Moore GP, Stauffer EK, Stuart DG (1976) Correlation analysis of muscle spindle responses to single motor unit contractions. J Physiol 257(2):325-336. [CrossRef]
- Binder MD, Kroin JS, Moore GP, Stuart DG (1977) The response of Golgi tendon organs to single motor unit contractions. J Physiol 271(2):337-349. [CrossRef]
- Bostan AC, Strick PL (2018) The basal ganglia and the cerebellum nodes in an integrated network. Nat Rev Neurosci 19:338-350. [CrossRef]
- Brodal A (1981) Neurological anatomy. In relation to clinical medicine, 3rd edn. Oxford University Press, New York.
- Brownstein CG, Millet GY, Thomas K (2021) Neuromuscular responses to fatiguing locomotor exercise. Acta Physiol (Oxf) 231(2):e13533. [CrossRef]
- Brunetti O, Della Torre G, Lucchi ML, Chiocchetti R, Bortolami R, Pettorossi VE (2003) Inhibition of muscle spindle afferent activity during masseter muscle fatigue in the rat. Exp Brain Res 152(2):251-262. [CrossRef]
- Burke RE (2009) Motor units. In: Binder MD, Hirokawa N, Windhorst U (eds) Encyclopedia of neuroscience. Springer-Verlag, Berlin Heidelberg, pp 2443-2446.
- Byrne C, Twist C, Eston R (2004) Neuromuscular function after exercise-induced muscle damage: theoretical and applied implications. Sports Med 34(1):49-69. [CrossRef]
- Cairns SP (2006) Lactic acid and exercise performance: culprit or friend? Sports Med 36(4):279-291. [CrossRef]
- Caligiori D, Pezzulo G, Baldassare G, Bostan AC, Strick PL, Doya K, Helmich RC, Dirkx M, Houk J, Jörntell H, Lago-Rodriguez A, Galea JM, Miall RC, Popa R, Kishore A, Verschure PFMJ, Zucca R, Herreros I (2017) Consensus paper: towards a systems-level view of cerebellar function: the interplay between cerebellum, basal ganglia, and cortex. Cerebellum 16(1): 203–229. [CrossRef]
- Cantor F (2010) Central and peripheral fatigue: exemplified by multiple sclerosis and myasthenia gravis. PM R 2(5):399-405. [CrossRef]
- Carroll TJ, Taylor JL, Gandevia SC (2017) Recovery of central and peripheral neuromuscular fatigue after exercise. J Appl Physiol 122:1068-1076. [CrossRef]
- Chaudhuri A, Behan PO (2000) Fatigue and basal ganglia. J Neurol Sci 179:34–42. [CrossRef]
- Chaudhuri A, Behan PO (2004) Fatigue in neurological disorders. Lancet 363(9413):978-988. [CrossRef]
- Cheng AJ, Jude B, Lanner JT (2020) Intramuscular mechanisms of overtraining. Redox Biol 35:101480. [CrossRef]
- Cheng AJ, Place N, Westerblad H (2018) Molecular basis for exercise-induced fatigue: the importance of strictly controlled cellular Ca2+ handling. Cold Spring Harb Perspect Med. 2018 Feb 1;8(2):a029710. [CrossRef]
- Cheng AJ,Yamada T, Rassier DE, Andersson DC, Westerblad H, Lanner JT (2016) Reactive oxygen/nitrogen species and contractile function in skeletal muscle during fatigue and recovery. J Physiol 594(18):5149-5160. [CrossRef]
- Chiang C, Aston-Jones G (1993) Response of locus coeruleus neurons to foot shock stimulation is mediated by neurons in the rostral ventral medulla. Neuroscience 53(3):705-715. [CrossRef]
- Christakos CN, Windhorst U (1986) Spindle gain increase during muscle unit fatigue. Brain Res 365(2):388-392. [CrossRef]
- Clarkson PM, Hubal MJ (2002) Exercise-induced muscle damage in humans. Am J Phys Med Rehabil 81(11 Suppl):S52-69. [CrossRef]
- Clarkson PM, Newham DJ (1995) Associations between muscle soreness, damage, and fatigue. Adv Exp Med Biol 384:457-469. [CrossRef]
- Coletti C, Acosta GF, Keslacy S, Coletti D (2022) Exercise-mediated reinnervation of skeletal muscle in elderly people: An update. Eur J Transl Myol 32(1):10416. [CrossRef]
- Constantin-Teodosiu D, Constantin D (2021) Molecular mechanisms of muscle fatigue. Int J Mol Sci 22(21):11587. [CrossRef]
- Costa Silva C, Carneiro Bichara CN, Carneiro FRO, da Cunha Menezes Palacios VR, Van den Berg AVS, Quaresma JAS, Falcão LFM (2022) Muscle dysfunction in the long coronavirus disease 2019 syndrome: Pathogenesis and clinical approach. Rev Med Virol 32(6):e2355. [CrossRef]
- Cotel F, Exley R, Cragg SJ, Perrier J-F (2013) Serotonin spillover onto the axon initial segment of motoneurons induces central fatigue by inhibiting action potential initiation. Proc Natl Acad Sci U S A 110(12):4774-4779. [CrossRef]
- Crisafulli O, Berardinelli A, D'Antona G (2024) Fatigue in spinal muscular atrophy: a fundamental open issue. Acta Myol 43(1):1-7. [CrossRef]
- Crook H, Raza S, Nowell J, Young M, Edison P (2021) Long covid-mechanisms, risk factors, and management. BMJ 2021 374:n1648. [CrossRef]
- Cuadra C, De Boef A, Luong S, Wolf SL, Nichols TR, Lyle MA (2024) Reduced inhibition from quadriceps onto soleus after acute quadriceps fatigue suggests Golgi tendon organ contribution to heteronymous inhibition. Eur J Neurosci 60(3):4317-4331. [CrossRef]
- Debold EP (2015) Potential molecular mechanisms underlying muscle fatigue mediated by reactive oxygen and nitrogen species. Front Physiol 6:239. [CrossRef]
- Debold EP, Fitts RH, Sundberg CW, Nosek TM (2016) Muscle fatigue from the perspective of a single crossbridge. Med Sci Sports Exerc 48:2270-2280. [CrossRef]
- Decherchi P, Dousset E (2003) Le rôle joué par les fibres afférentes métabosensibles dans les mécanismes adaptifs neuromusculaires. Can J Neurol Sci 30:91-97. [CrossRef]
- Dettmers C, Lemon RN, Stephan KM, Fink GR, Frackowiak RS (1996) Cerebral activation during the exertion of sustained static force in man. Neuroreport 7(13):2103-2110 . [CrossRef]
- Dibaj P, Brockmann K, Gärtner J (2020) Dopamine-mediated yawning-fatigue syndrome with specific recurrent initiation and responsiveness to opioids. JAMA Neurol 77(2):254. [CrossRef]
- Dibaj P, Seeger D, Gärtner J, Petzke F (2021) Follow-up of a case of dopamine-mediated yawning-fatigue-syndrome responsive to opioids, successful desensitization via graded activity treatment. Neurol Int 13(1):79-84. [CrossRef]
- Dibaj P, Windhorst U (2024) Motor-control notions in health and disease (what controls motor control?). Preprints 2024, 2024031799. [CrossRef]
- Enoka RM, Duchateau J (2008) Muscle fatigue: what, why and how it influences muscle function. J Physiol 586(1):11-23. [CrossRef]
- Esposito F, Ce E, Rampichini S, Monti E, Limonta E, Fossati B, Meola G (2017) Electromechanical delays during a fatiguing exercise and recovery in patients with myotonic dystrophy type 1. Eur J Appl Physiol 117:551-566. [CrossRef]
- Finsterer J, Mahjoub SZ (2014) Fatigue in healthy and diseased individuals. Am J Hosp Palliat Care 31(5):562-755. [CrossRef]
- Fitts RH (1994) Cellular mechanisms of muscle fatigue. Physiol Rev 74:49-94. [CrossRef]
- Fitts RH (2008) The cross-bridge cycle and skeletal muscle fatigue. J Appl Physiol 104:551-558. [CrossRef]
- Foley TE, Fleshner M (2008) Neuroplasticity of dopamine circuits after exercise: implications for central fatigue. Neuromolecular Med 10(2):67-80. [CrossRef]
- Fornal CA, Martin-Cora FJ, Jacobs BL (2006) "Fatigue" of medullary but not mesencephalic raphe serotonergic neurons during locomotion in cats. Brain Res 1072(1):55-61. [CrossRef]
- Gandevia SC (2001) Spinal and supraspinal factors in human muscle fatigue. Physiol Rev 81:1725-1789. [CrossRef]
- Garland SJ, McComas AJ (1990) Reflex inhibition of human soleus muscle during fatigue. J Physiol 429:17-27. [CrossRef]
- Granger MW, Buschang PH, Throckmorton GS, Iannaccone ST (1999) Masticatory muscle function in patients with spinal muscular atrophy. Am J Orthod Dentofacial Orthop 115(6):697-702. [CrossRef]
- Garssen MPJ, Schillings ML, van Doorn PA, van Engelen BGM, Zwarts MJ (2007) Contribution of central and peripheral factors to residual fatigue in Guillain-Barré syndrome. Muscle Nerve 36(1):93-99. [CrossRef]
- Gottschalk, M, Kümpfel T, Flachenecker P, Uhr M, Trenkwalder C, Holsboer F, Weber F (2005) Fatigue and regulation of the hypothalamo-pituitary-adrenal axis in multiple sclerosis. Arch Neurol 62(2):277–280. [CrossRef]
- Green HJ (1997) Mechanisms of muscle fatigue in intense exercise. J Sports Sci 15(3):247-562. [CrossRef]
- Gregory JE, Morgan DL, Proske U (2003) Tendon organs as monitors of muscle damage from eccentric contractions. Exp Brain Res 151:346-355. [CrossRef]
- Grosprêtre S, Gueugneau N, Martin A, Lepers R (2018) Presynaptic inhibition mechanisms may subserve the spinal excitability modulation induced by neuromuscular electrical stimulation. J Electromyogr Kinesiol 40:95-101. [CrossRef]
- Gruet M, Temesi J, Rupp T, Lev P, Millet GY. Verges S (2013) Stimulation of the motor cortex and corticospinal tract to assess human muscle fatigue. Neuroscience 12:231:384-399. [CrossRef]
- Hall MM, Rajasekaran S, Thomsen TW, Peterson AR (2016) Lactate: Friend or Foe. PM R 8(3 Suppl):S8-S15. [CrossRef]
- Hayward L, Breitbach D, Rymer WZ (1988) Increased inhibitory effects on close synergists during muscle fatigue in the decerebrate cat. Brain Res 440(1):199-203. [CrossRef]
- Hayward L, Wesselmann U, Rymer WZ (1991) Effects of muscle fatigue on mechanically sensitive afferents of slow conduction velocity in the cat triceps surae. J Neurophysiol 65:360-370. [CrossRef]
- Heesen, C, Nawrath L, Reich C, Bauer N, Schulz K, Gold SM (2006) Fatigue in multiple sclerosis: an example of cytokine mediated sickness behaviour? J Neurol Neurosurg Psychiatry 77(1):34–39. [CrossRef]
- Helbostad JL, Sturnieks DL, Menant J, Delbaere K, Lord SR, Pijnappels M (2010) Consequences of lower extremity and trunk muscle fatigue on balance and functional tasks in older people: a systematic literature review. BMC Geriatr 17:10:56. [CrossRef]
- Henderson TT, Taylor JL, Thorstensen JR, Tucker MG, Kavanagh JJ (2022) Enhanced availability of serotonin limits muscle activation during high-intensity, but not low-intensity, fatiguing contractions. J Neurophysiol 128(4):751-762. [CrossRef]
- Hilty L, Lutz K, Maurer K, Rodenkirch T, Spengler CM, Boutellier U, Jäncke L, Amann M (2011) Spinal opioid receptor-sensitive muscle afferents contribute to the fatigue-induced increase in intracortical inhibition in healthy humans. Exp Physiol 96(5):505-517. [CrossRef]
- Hou LJ, Song Z, Pan ZJ, Cheng JL, Yu Y, Wang J (2016) Decreased activation of subcortical brain areas in the motor fatigue state: An fMRI study. Front Psychol 7:1154. [CrossRef]
- Hsiao Y, Elliot TR, Jaramillo J, Douglas ME, Powers MB, Warren AM (2024) The fatigue and altered cognition scale among SARS-CoV-2 survivors: psychometric properties and item correlations with depression and anxiety symptoms. J Clin Med 13(8):2186. [CrossRef]
- Hunter SK, Pereira HM, Keenan KG (2016) The aging neuromuscular system and motor performance. J Appl Physiol (1985) 121(4):982-995. [CrossRef]
- Hureau TJ, Romer LM, Amann M (2018) The 'sensory tolerance limit': A hypothetical construct determining exercise performance? Eur J Sport Sci 18(1):13-24. [CrossRef]
- Hutton RS, Nelson DL (1986) Stretch sensitivity of Golgi tendon organs in fatigued gastrocnemius muscle. Med Sci Sports Exerc 18(1):69-74.
- Jankowska E (1992) Interneuronal relay in spinal pathways from proprioceptors. Prog Neurobiol 38:335-378. [CrossRef]
- Jankowska E, Edgley SE (2010) Functional subdivision of feline spinal interneurons in reflex pathways from group Ib and II muscle afferents; an update. Eur J Neurosci 32: 881-893. [CrossRef]
- Jiang Z, Wang X-F, Yue GH (2016) Strengthened corticosubcortical functional connectivity during muscle fatigue. Neural Plast 2016:1726848. [CrossRef]
- Jones DA (1993) How far can experiments in the laboratory explain the fatigue of athletes in the field? In: Sargeant AJ, Kernell D (eds) Neuromuscular fatigue. Royal Netherlands Academy of Arts and Sciences, Amsterdam, pp 100-108.
- Kalezic I, Bugaychenko LA, Kostyukov AI, PilyavskiiAI, Ljubisavljevid M, Windhorst U, Johansson H (2004) Fatigue-related depression of the feline monosynaptic gastrocnemius-soleus reflex. J Physiol 556(Pt 1):283-296. [CrossRef]
- Kalezic I, Steffens H (2013) Changes in tetrodotoxin-resistant C-fibre activity during fatiguing isometric contractions in the rat.PLoS One 8(9):e73980. [CrossRef]
- Katz R, Meunier S, Pierrot-Deseilligny E (1988) Changes in presynaptic inhibition of Ia fibres in man while standing. Brain 111:417-437. [CrossRef]
- Katz R, Pierrot-Deseilligny E (1999) Recurrent inhibition in humans. Prog Neurobiol 57:325-355. [CrossRef]
- Kavanagh JJ, Taylor JL (2022) Voluntary activation of muscle in humans: does serotonergic neuromodulation matter? J Physiol 600(16):3657-3670. [CrossRef]
- Keller J, Zackowski K, Kim S, Chidobem I, Smith M, Farhardi F, Bhargava P (2021) Exercise leads to metabolic changes associated with imprived strength and fatigue in people with MS. Ann Clin Transl Neurol 8(6):1308-1317. [CrossRef]
- Kent JA,, Ørtenblad N, Hogan MC, Poole DC, Musch TI (2016) No muscle is an island: Integrative perspectives on muscle fatigue. Med Sci Sports Exerc 48(11):2281-2293. [CrossRef]
- Kent-Braun JA, Fitts RH, Christie A (2012) Skeletal muscle fatigue. Compr Physiol 2:997-1044. [CrossRef]
- Kent-Braun JA, Miller RG (2000) Central fatigue during isometric exercise in amyotrophic lateral sclerosis. Muscle Nerve 23:909-914. [CrossRef]
- Kent-Braun JA, Sharma KR, Weiner MW, Massie B, Miller RG (1993) Central basis of fatigue in chronic fatigue syndrome. Neurology 43(1):125-131. [CrossRef]
- Kernell D, Monster AW (1982) Motoneurone properties and motor fatigue. An intracellular study of gastrocnemius motoneurones of the cat. Exp Brain Res 46:197-204. [CrossRef]
- Klass M, Duchateau J, Rabec S, Meeusen R, Roelands B (2016) Noradrenaline reuptake inhibition impairs cortical output and limits endurance time. Med Sci Sports Exerc 48(6):1014-1023. [CrossRef]
- Klass M, Roelands B, Lévénez M, Fontenelle V, Pattyn N, Meeusen R, Duchateau J (2012) Effects of noradrenaline and dopamine on supraspinal fatigue in well-trained men. Med Sci Sports Exerc 44(12):2299-2308. [CrossRef]
- Knorr S, Rice CL, Garland SJ (2012) Perspective on neuromuscular factors in poststroke fatigue. Disabil Rehabil 34(26):2291-2299. [CrossRef]
- Koehler W, Hamm TM, Enoka RM, Stuart DG, Windhorst U (1984) Contractions of single motor units are reflected in membrane potential changes of homonymous alpha-motoneurons. Brain Res 296(2):379-834. [CrossRef]
- Kostyukov AI, Bugaychenko LA, Kalezic I, Pilyavskii AI, Windhorst U, Djupsjöbacka M (2005) Effects in feline gastrocnemius-soleus motoneurones induced by muscle fatigue. Exp Brain Res 163(3):284-294. [CrossRef]
- Kostyukov AI, Kalezic I, Serenko SG, Ljubisavljevic M, Windhorst U, Johansson H (2002) Spreading of fatigue-related effects from active to inactive parts in the medial gastrocnemius muscle of the cat. Eur J Appl Physiol 86:295-307. [CrossRef]
- Koutsikou S, Apps R, Lumb BM (2017) Top-down control of spinal sensorimotor circuits essential for survival. J Physiol (Lond) 595(13):4151-4158. [CrossRef]
- Kuchinad RA, Ivanova TD, Garland SJ (2004) Modulation of motor unit discharge rate and H-reflex amplitude during submaximal fatigue of the human soleus muscle. Exp Brain Res 158(3):345-355. [CrossRef]
- Kuner R, Kuner T (2021) Cellular circuits in the brain and their modulation in acute and chronic pain. Physiol Rev 101(1):213-258. [CrossRef]
- Lännergren J, Westerblad H, Allen DG (1993) Mechanisms of fatigue as studied in single muscle fibres. In: Sargeant AJ, Kernell D (eds) Neuromuscular fatigue. Royal Netherlands Academy of Arts and Sciences, Amsterdam, pp 3-11.
- Lalley PM (2009 Respiration – neural control. In: Binder MD, Hirokawa N, Windhorst U (eds) Encyclopedia of neuroscience. Springer-Verlag, Berlin Heidelberg, pp 3433-3441.
- Lamotte G, Shouman K, Benarroch EE (2021) Stress and central autonomic network. Auton Neurosci 235:102870. [CrossRef]
- Laurin J, Pertici V, Doucet E, Marqueste T, Decherchi P (2015) Group III and IV muscle afferents: role on central motor drive and clinical implications. Neuroscience 290:543-551. [CrossRef]
- Lenman AJ, Tulley FM, Vrbova G, Dimitrijevic MR, Towle J (1989) Muscle fatigue in some neurological disorders. Muscle Nerve 12(11):938-942. [CrossRef]
- Lin K, Chen Y, Luh J, Wang C, Chang Y (2012) H-reflex, muscle voluntary activation level, and fatigue index of flexor carpi radialis in individuals with incomplete cervical cord injury. Neurorehabilitation and Neural Repair 26(1):68–75. [CrossRef]
- Lindinger MI, Cairns SP (2021) Regulation of muscle potassium: exercise performance, fatigue and health implications. Eur J Appl Physiol 121(3):721-748. [CrossRef]
- Liu JZ, Shan ZY, Zhang LD, Sahgal V, Brown RW, Yue GH (2003) Human brain activation during sustained and intermittent submaximal fatigue muscle contractions: an FMRI study. J Neurophysiol 90(1):300-312. [CrossRef]
- Lou JS, Benice T, Kearns G, Sexton G, Nutt J (2003) Levodopa normalizes exercise related cortico-motoneuron excitability abnormalities in Parkinson’s disease. Clin Neurophysiol 114(5):930–937. [CrossRef]
- Macefield G, Hagbarth KE, Gorman R, Gandevia SC, Burke D (1991) Decline in spindle support to alpha-motoneurones during sustained voluntary contractions. J Physiol (Lond) 440:497-512. [CrossRef]
- Maisky VA, Pilyavskii AI , Kalezic I, Ljubisavljevid M, Kostyukov AI, Windhorst U, Johansson H (2002) NADPH-diaphorase activity and c-fos expression in medullary neurons after fatiguing stimulation of hindlimb muscles in the rat. Auton Neurosci 101(1-2):1-12. [CrossRef]
- Malekzadeh A, Bader I, van Dieteren J, Heijboer AC, Beckerman H, Twisk JWR, de Groot V, Teunissen CE (2020) Diurnal cortisol secretion is not related to multiple sclerosis-related fatigue. Front Neurol 10:1363. [CrossRef]
- Martin A, Freyssenet D (2021) Phenotypic features of cancer cachexia-related loss of skeletal muscle mass and function: lessons from human and animal studies.J Cachexia Sarcopenia Muscle 12(2):252-273. [CrossRef]
- Mastaglia FL (2012) The relationship between muscle pain and fatigue. Neuromuscul Disord 22 Suppl 3:S178-180. [CrossRef]
- Maznychenko AV, Pilyavskii AI, Kostyukov AI, Lyskov E, Vlasenko OV, Maisky VA (2007) Coupling of c-fos expression in the spinal cord and amygdala induced by dorsal neck muscles fatigue. Histochem Cell Biol 28(1):85-90. [CrossRef]
- McCloskey DI, Prochazka A (1994) The role of sensory information in the guidance of voluntary movement. Somatosens Mot Res 11:21-37. [CrossRef]
- McCrimmon DR, Alheid GF (2009) Respiratory reflexes. In: Binder MD, Hirokawa N, Windhorst U (eds) Encyclopedia of neuroscience. Springer-Verlag, Berlin Heidelberg, pp 3474-3480. [CrossRef]
- McKenna MJ, Bangsbo J, Renaud JM (2008) Muscle K+, Na+, and Cl- disturbances and Na+- K+ pump inactivation; implications for fatigue. J Appl Physiol 104:288-295. [CrossRef]
- Meacci E, Pierucci F, Garcia-Gil M (2022) Skeletal muscle and COVID-19: The potential involvement of bioactive sphingolipids. Biomedicines 10(5):1068. [CrossRef]
- Meeusen R, Roelands B (2018) Fatigue: Is it all neurochemistry? Eur J Sport Sci 18(1):37-46. [CrossRef]
- Mense S (1993) Nociception from skeletal muscle in relation to clinical muscle pain. Pain 54(3):241-289. [CrossRef]
- Miller RG, Green AT, Moussavi RS, Carson PJ, Weiner MW (1990) Excessive muscular fatigue in patients with spastic paraparesis. Neurology 40(8):1271-1274. [CrossRef]
- Mills EP, Keay KA, Henderson LA (2021) Brainstem pain-modulation circuitry and its plasticity in neuropathic pain: Insights from human brain imaging investigations. Front Pain Res (Lausanne) 2:705345. [CrossRef]
- Montes J, McDermott MP, Martens WB, Dunaway S, Glanzman AM, Riley S, Quigley J, Montgomery MJ, Sproule D, Tawil R, Chung WK, Darras BT, De Vivo DC, Kaufmann P, Finkel RS; Muscle study group and the pediatric neuromuscular clinical research network (2010) Six-Minute Walk Test demonstrates motor fatigue in spinal muscular atrophy. Neurology 74(10):833-838. [CrossRef]
- Mossberg K, Masel B, Gilkison C, Urban R (2008) Aerobic capacity and growth hormone deficiency after traumatic brain injury. J Clin Endocrinol Metab 93:2581–2587. [CrossRef]
- Nardone R, Buffone E, Florio I, Tezzon F (2005) Changes in motor cortex excitability during muscle fatigue in amyotrophic lateral sclerosis. J Neurol Neurosurg Psychiatry 76:429–31. [CrossRef]
- Nardone A, Schieppati M (2004) Group II spindle fibres and afferent control of stance. Clues from diabetic neuropathy. Clin Neurophysiol 115:779-789. [CrossRef]
- Noonan TJ, Garrett WE (1999) Muscle strain injury: diagnosis and treatment. J Am Acad Orthop Surg 7(4):262-269. [CrossRef]
- Olave MJ, Puri N, Kerr R, Maxwell DJ (2002) Myelinated and unmyelinated primary afferent axons form contacts with cholinergic interneurons in the spinal dorsal horn. Exp Brain Res 45(4):448-456. [CrossRef]
- Ortelli P, Ferrazzoli D, Sebastianelli L, Engl M, Romanello R, Nardone R, Bonini I, Koch G, Saltuari L, Quartarone A, Oliviero A, Kofler M, Versace V (2021) Neuropsychological and neurophysiological correlates of fatigue in post-acute patients with neurological manifestations of COVID-19: Insights into a challenging symptom. J Neurol Sci 420:117271. [CrossRef]
- Ørtenblad N, Westerblad H, Nielsen J (2013) Muscle glycogen stores and fatigue. J Physiol (Lond) 591:4405-4413. [CrossRef]
- Papa EV, Garg H, Dibble LE (2015) Acute effects of muscle fatigue on anticipatory and reactive postural control in older individuals: a systematic review of the evidence. J Geriatr Phys Ther 38(1):40-48. [CrossRef]
- Pearson AM, Young RB (1993) Diseases and disorders of muscle. Adv Food Nutr Res 37:339-423. [CrossRef]
- Penner I, Paul F (2017) Fatigue as a symptom or comorbidity of neurological diseases. Nat Rev Neurol 13(11):662-675. [CrossRef]
- Pescaru CC, Mariţescu A, Costin EO, Trăilă D, Marc MS, Truşculescu AA, Pescaru A, Oancea CI (2022) The effects of COVID-19 on skeletal muscles, muscle fatigue and rehabilitation programs outcomes. Medicina (Kaunas) 58(9):1199. [CrossRef]
- Pilyavskii AI. Maisky VA, Kalezic I, Ljubisavljevic M, Kostyukov AI, Windhorst U, Johansson H (2001) C-fos expression and NADPH–d reactivity in spinal neurons after fatiguing stimulation of hindlimb muscles in the rat. Brain Research 293: 91-102. [CrossRef]
- Ponchel A, Bombois S, Bordet R, Hénon H (2015) Factors associated with poststroke fatigue: a systematic review. Stroke Res Treat 2015:347920. [CrossRef]
- Powers SK, Deminice R, Ozdemir M, Yoshihara T, Bomkamp MP, Hyatt H (2020) Exercise-induced oxidative stress: Friend or foe? J Sport Health Sci 9(5):415-425. [CrossRef]
- Powers SK, Jackson MJ (2008) Exercise-induced oxidative stress: cellular mechanisms and impact on muscle force production. Physiol Rev 88:1243-1276. [CrossRef]
- Powers SK, Lynch GS, Murphy KT, Reid MB, Zijdewind I (2016) Disease-induced skeletal muscle atrophy and fatigue. Med Sci Sports Exerc 48(11):2307-2319. [CrossRef]
- Proske U (2019) Exercise, fatigue and proprioception: a retrospective. Exp Brain Res 237(10):2447-2459. [CrossRef]
- Quevedo JN (2009) Presynaptic inhibition. In: Binder MD, Hirokawa N, Windhorst U (eds) Encyclopedia of neuroscience. Springer-Verlag, Berlin Heidelberg, pp 3266-3270.
- Ramazzini B (1713) De morbis artificum diatriba. In: Diseases of workers. New York Academy of Medicine, History of Medicine Series (1964) Hafner, N.Y.
- Ranieri F, Di Lazzaro V (2012) The role of motor neuron drive in muscle fatigue. Neuromuscul Disord 22 Suppl 3:S157-161. [CrossRef]
- Rayhan RU, Baraniuk JN (2021) Submaximal exercise provokes increased activation of the anterior default mode network during the resting state as a biomarker of postexertional malaise in myalgic encephalomyelitis/chronic fatigue syndrome. Front Neurosci 15:748426. [CrossRef]
- Renaud J-M, Ørtenblad N, McKenna MJ, Overgaard K (2023) Exercise and fatigue: integrating the role of K+, Na+ and Cl- in the regulation of sarcolemmal excitability of skeletal muscle. Eur J Appl Physiol 123(11):2345-2378. [CrossRef]
- Richter DM, Smith JC (2014) Respiratory rhythm generation in vivo. Physiology (Bethesda) 29(1):58-71. [CrossRef]
- Roelcke U, Kappos L, Lechner-Scott J, Brunnschweiler H, Huber S, Ammann W, Plohmann A, Dellas S, Maguire RP, Missimer J, Radü EW, Steck A, Leenders KL (1997) Reduced glucose metabolism in the frontal cortex and basal ganglia of multiple sclerosis patients with fatigue: a 18F-fluorodeoxyglucose positron emission tomography study. Neurology 48(6):1566-1571. [CrossRef]
- Rosa EF, Alves GA, Luz J, Silva SMA, Suchecki D, Pesquero JB, Aboulafia J, Nouailhetas VLA (2014) Activation of HPA axis and remodeling of body chemical composition in response to an intense and exhaustive exercise in C57BL/6 mice. Physiol Res 63(5):605-613. [CrossRef]
- Rossi A, Mazzocchio R, Decchi B (2003) Effect of chemically activated fine muscle afferents on spinal recurrent inhibition in humans. Clin Neurophysiol 114(2):279-287. [CrossRef]
- Rotto DM, Kaufmann MP (1988) Effect of metabolic products of muscular contraction on discharge of group III and IV afferents. J Appl Physiol 64:2306-2313. [CrossRef]
- Rudomin P (2009) In search of lost presynaptic inhibition. Exp Brain Res 196:139-151. [CrossRef]
- Rudomin P, Schmidt RF (1999) Presynaptic inhibition in the vertebrate spinal cord revisited. Exp Brain Res 129:1-37. [CrossRef]
- Sato T, Tsuboi T, Miyazaki M, Sakamoto K (1999) Post-tetanic potentiation of reciprocal Ia inhibition in human lower limb. J Electromyogr Kinesiol 9(1):59-66. [CrossRef]
- Schillings ML, Kalkman JS, van der Werf SP, van Engelen BGM, Bleijenberg G, Zwarts MJ (2004) Diminished central activation during maximal voluntary contraction in chronic fatigue syndrome. Clin Neurophysiol 115(11):2518-2524. [CrossRef]
- Schirinzi E, Ricci G, Torri F, Mancuso M, Siciliano G (2023) Biomolecules of muscle fatigue in metabolic myopathies: Biomolecules 14(1):50. [CrossRef]
- Schomburg ED (1990) Spinal sensorimotor systems and their supraspinal control. Neurosci Res 7: 265-340. [CrossRef]
- Schwestka R, Windhorst U, Schaumberg R (1981) Patterns of parallel signal transmission between multiple alpha efferents and multiple Ia afferents in the cat semitendinosus muscle. Exp Brain Res 43(1):34-46. [CrossRef]
- Semmler JG (2014) Motor unit activity after eccentric exercise and muscle damage in humans. Acta Physiol (Oxf) 210(4):754-767. [CrossRef]
- Sepulcre J, Masdeu JC, Goñi J, Arrondo G, Vélez de Mendizábal N, Bejarano B, Villoslada P (2009) Fatigue in multiple sclerosis is associated with the disruption of frontal and parietal pathways. Mult Scler 15(3):337-344. [CrossRef]
- Shepherd GMG, Yamawaki N (2021) Untangling the cortico-thalamo-cortical loop: cellular pieces of a knotty circuit puzzle. Nat Rev Neurosci 22(7):389-406. [CrossRef]
- Sidhu SK, Weavil JC, Thurston TS, Rosenberger D, Jessop JE, Wang E, Richardson RS, McNeil CJ, Amann M (2018) Fatigue-related group III/IV muscle afferent feedback facilitates intracortical inhibition during locomotor exercise. J Physiol 596(19):4789-4801. [CrossRef]
- Storti SF, Formaggio E, Motetto D, Bertoldo A, Pizzini FB, Beltramello A, Fiaschi A, Toffolo GM, Manganoptti P (2014) Effect of voluntary repetitive long-lasting muscle contraction activity on the BOLD signal as assessed by optimal hemodynamic response function. MAGMA 27(2):171-184. [CrossRef]
- Sved AF, Ito S, Madden CJ (2000) Baroreflex dependent and independent roles of the caudal ventrolateral medulla in cardiovascular regulation. Brain Res Bull 51(2):129-133. [CrossRef]
- Takayanagi Y, Onaka T (2021) Roles of oxytocin in stress responses, allostasis and resilience. Int J Mol Sci ;23(1):150. [CrossRef]
- Tanaka M, Watanace Y (2012) Supraspinal regulation of physical fatigue. Neurosci Biobehav Rev 36(1):727-734. [CrossRef]
- Tanino Y, Daikuya S, Nishimori T, Takasaki K, Kanei K, Suzuki T (2004) H-reflex and reciprocal Ia inhibition after fatiguing isometric voluntary contraction in soleus muscle. Electromyogr Clin Neurophysiol 44(8):473-476.
- Taylor JL, Amann M, Duchateau J, Meeusen R, Rice CL (2016) Neural contributions to muscle fatigue: from the brain to the muscle and back again. Med Sci Sports Exerc 48:2294-2306. [CrossRef]
- Tsukioka K, Yamanaka K, Waki H (2022) Implication of the central nucleus of the amygdala in cardiovascular regulation and limiting maximum exercise performance during high-intensity exercise in rats. Neuroscience 496:52-63. [CrossRef]
- Van Bockstaele EJ, Pieribone VA, Aston-Jones G (1989) Diverse afferents converge on the nucleus paragigantocellularis in the rat ventrolateral medulla: retrograde and anterograde tracing studies. J Comp Neurol 290(4):561-584. [CrossRef]
- Van Duinen H, Renken R, Maurits N, Zijdewind I (2007) Effects of motor fatigue on human brain activity, an fMRI study. Neuroimage 35(4):1438-1449. [CrossRef]
- Vlasenko OV, Maĭs'kyĭ VO, Maznychenko AV, Piliavs'kyĭ OI, Moroz VM (1994) [Investigation of c-fos expression and NADPH-diaphorase activity in the spine cord and brain in the development of the neck muscle weakness in rats]. Fiziol Zh (1994) 2006;52(6):3-14 [Article in Ukrainian].
- Vucic S, Cheah BC, Kiernan MC (2011) Maladaptation of cortical circuits underlies fatigue and weakness in ALS. Amyotroph Lateral Scler 12:414–20. [CrossRef]
- Wang M, Tutt JO, Dorricott NO, Parker KL, Russo AF, Sowers LP (2022) Involvement of the cerebellum in migraine. Front Syst Neurosci 16: 984406. [CrossRef]
- Weavil JC, Amann M (2018) Corticospinal excitability during fatiguing whole body exercise. Prog Brain Res 219-246. [CrossRef]
- Wichmann T, DeLong MR (2016) Deep brain stimulation for movement disorders of basal ganglia origin: restoring function or functionality? Neurotherapeutics 13(2):264-283. [CrossRef]
- Williams CA, Fowler WL (1997) Substance P released in the rostral brainstem of cats interacts with NK-1 receptors during muscle pressor response. Neuropeptides 31(6):589-600. [CrossRef]
- Williams CA, Gopalan R, Nichols PL, Brien PL (1995) Fatiguing isometric contraction of hind-limb muscles results in the release of immunoreactive neurokinins from sites in the rostral medulla in the anesthetized cat. Neuropeptides 28(4):209-218. [CrossRef]
- Williams CA, Holtsclaw LI, ChivertonJA (1992) Release of immunoreactive enkephalinergic substances in the periaqueductal grey of the cat during fatiguing isometric contractions. Neurosci Lett 139(1):19-23. [CrossRef]
- Williams CA, Roberts JR, Freels DB (1990) Changes in blood pressure during isometric contractions to fatigue in the cat after brain stem lesions: effects of clonidine. Cardiovasc Res . 1990 24(10):821-833. [CrossRef]
- Williams JH, Klug GA (1995) Calcium exchange hypothesis of skeletal muscle fatigue: a brief review. Muscle Nerve 18(4):421-344. [CrossRef]
- Windhorst U (2003) Short-term effects of group III-IV muscle afferent nerve fibers on bias and gain of spinal neurons. In: Johansson H, Windhorst U, Djupsjöbacka M, Passatore M (eds) Neuromuscular mechanisms behind work -related chronic muscle pain syndromes, Gävle University Press, Gävle Sweden.
- Windhorst U (2007) Muscle proprioceptive feedback and spinal networks. Brain Res Bull 73:155-202. [CrossRef]
- Windhorst U (2021) Spinal cord circuits: models and reality. Neurophysiology 53(3-6):142-222. [CrossRef]
- Windhorst U, Dibaj P (2023) Plastic spinal motor circuits in health and disease. J Integr Neurosci 22(6):167. [CrossRef]
- Windhorst U, Meyer-Lohmann J, Kirmayer D, Zochodne D (1997) Renshaw cell responses to intra-arterial injection of muscle metabolites into cat calf muscles. Neurosci Res 27(3):235-247. [CrossRef]
- Yamada T, Ashida Y, Tamai K, Kimura I, Yamauchi N, Naito A, Tokuda N, Westerblad H, Andersson DC, Himori K (2022) Improved skeletal muscle fatigue resistance in experimental autoimmune myositis mice following high-intensity interval training. Arthritis Res Ther 24:156. [CrossRef]
- Zabihhosseinian M, Yielder P, Berkers V, Ambalavanar U, Holmes M, Murphy B (2020) Neck muscle fatigue impacts plasticity and sensorimotor integration in cerebellum and motor cortex in response to novel motor skill acquisition. J Neurophysiol 124(3):844-855. [CrossRef]
- Zgaljardic DJ, Durham WJ, Mossberg KA, Foreman J, Joshipura K, Masel BE, Urban R, Sheffield-Moore M (2014) Neuropsychological and physiological correlates of fatigue following traumatic brain injury. Brain Inj 28(4):389–397. [CrossRef]
- Zwarts MJ, Bleijenberg G, van Engelen BGM (2008) Clinical neurophysiology of fatigue. Clin Neurophysiol 119(1):2-10. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



