
Submitted:
18 September 2024
Posted:
20 September 2024
You are already at the latest version
Abstract
Background/Aims: Obstructive sleep apnea syndrome (OSAS) affects a large part of the aging population. It is characterized by chronic intermittent hypoxia and associated with neurocognitive dysfunction. One hypothesis is that the blood-brain barrier (BBB) functions could be altered by exosomes. Exosomes are nanovesicles found in biological fluids. Through the study of exosomes and their content in tau and amyloid beta (Ab), the aim of this study was to show how exosomes could be used as biomarkers of OSAS and of their cognitive disorders. Methods: Two groups of 15 volunteers from the PROOF cohort were selected: severe apnea (AHI >30) and control (AHI <5). After exosome isolation from blood serum, we characterized and quantified them (CD81, CD9, CD63) by western blot and ELISAs and put them 5h in contact with an in vitro BBB model. The apparent permeability of the BBB was measured using sodium-fluorescein and TEER. Cell ELISAs were performed on tight junctions (ZO-1, claudin-5, occludin). The amount of tau and Ab proteins found in the exosomes was quantified using ELISAs. Results: Compared to controls, OSAS patients had a greater quantity of exosomes, tau and Ab proteins in their blood sera, which induced an increase in BBB permeability in the model and was reflected by a loss of tight junction’expression. Conclusion: Elderly patients suffering severe OSAS released more exosomes in serum from the brain compartment than controls. Such exosomes increased BBB permeability. The impact of such alterations on risk of developing cognitive dysfunction and/or neurodegenerative diseases is questioned.
Keywords:
elderly
; obstructive sleep apnea
; exosomes
; biomarkers
; blood-brain-barrier
; tau
; amyloid
; tight junction proteins
1. Introduction
Obstructive sleep apnea syndrome (OSAS) is characterized by repeated partial or complete upper airway’obstruction during sleep. These obstructions alter cyclically alveolar ventilation leading to episodic hypoxemia and hypercapnia, as well as sleep fragmentation. OSAS is a very common syndrome among the elderly with an increase in prevalence from the age of 60 years, but it remains a syndrome that is not very often diagnosed. Between 37.5-62% of people over 60 years may suffer from this syndrome [1]. OSAS is associated with an increased risk of diseases including cardiovascular, cerebrovascular and cognitive impairment [2,3], potentially leading to memory loss and loss of autonomy, particularly in the vulnerable elderly [4,5]. At the heart of consideration, is the definition of OSAS, commonly defined according to an apnea-aypopnea-index (AHI) > 5. The AHI is currently used as reference measurement tool, with reference values enabling us to distinguish between non-apneic subjects with an AHI < 5, subjects with mild apnea whose AHI is between 5-15, subjects with moderate apnea whose AHI is between 15-30, and finally severe apneics whose AHI > 30 [3]. Also, OSAS is defined according to the oxygen-desaturation-index (ODI), which is the number of episodes of oxygen desaturation per hour of sleep during which blood oxygen has fallen by ≥ 3% [6]. According to the American Academy of Sleep Medicine (AASM), the diagnosis of OSAS is based on AHI. An AHI ≥ 5 with symptoms indicates mild OSAS, while an AHI ≥ 15 is diagnostic of moderate to severe OSAS, even in the absence of symptoms or comorbidities [7]. European recommendations for the diagnosis of OSAS are also based on AHI. AHI ≥ 5 is diagnostic of mild OSAS, but diagnosis may be more focused on the presence of clinically relevant symptoms. An AHI ≥ 15 allows the diagnosis of moderate to severe OSAS, with greater consideration for associated comorbidities and impact on quality of life compared with the AASM recommendations [8].
On the basis of a cutoff at 65-95 years, prevalence rates are 62% for an AHI >10, 44% for an AHI > 20, and 24% for an AHI >40 [3,9]. In the elderly population, this OSAS pathology is still rarely diagnosed and its consequences underestimated, such as cognitive decline. The reference treatment is continuous positive airway pressure (CPAP). One of the main obstacles to treatment with CPAP is patient compliance, as this is a long-term, restrictive treatment that patients often stop. When patients do not use CPAP enough, clinical effects are compromised. Furthermore, the efficacy of CPAP on certain neurological disorders remains to be demonstrated; although there is evidence to suggest that CPAP interventions may improve cognitive function [10]. Identification of patients with a high risk of cognitive decline remains difficult and sometimes after a long time. Thus, improved understanding of the cellular and molecular mechanisms that lead to central nervous system (CNS) dysfunctions is needed in order to develop therapeutic strategies. The hypothesis that a disruption of the blood-brain-barrier (BBB) has emerged and may contribute to cognitive impairments in OSAS associated with intermittent hypoxia and sleep fragmentation [11,12].
The BBB is a multicellular vascular structure which acts as a diffusion barrier to prevent the entry of most compounds forming blood to the brain, thereby allowing the maintenance of brain homeostasis. Endothelial cells form the walls of the brain capillaries represent the anatomical basis of the BBB [13]. The BBB is mainly composed of endothelial cells linked together by tight junctions (TJs). It is also composed of astrocytes which play a major role in the maintenance of the junctions, as well of pericytes and neurons (Daneman and Prat 2015).
The mechanism connecting sleep disturbances and memory seems to be the nightly elimination of toxins or misfolded proteins such as amyloid beta peptides (Aβ), tubulin-associated unit protein (tau) accumulated in the brain during the previous day. More recently, it has been proposed that exosomal cargo containing Aβ and tau provides ues to mechanistic pathways while also serving as biomarkers of neurocognitive risks in patients [15]. It has been demonstrated that Alzheimer’s disease (AD) and sleep apnea share common features like inducing tau and Aβ protein accumulation [16] (Sun and al. 2022),(Pan and Kastin 2014),(Baril and al. 2018).
Exosomes are a class of endosome-derived membrane vesicles shed by cells, which contain proteins and other constituents of their cellular origin. They are known for their potential involvement in neurodegenerative diseases via their passage through the BBB.[19,20,21]. Their main role is inter-cellular communication. According to their origin, their composition differs [22].
Based on the leading hypothesis that OSAS produces altering exosomal cargo in serum, and that these altering exosomes increase BBB permeability and promote pathophysiological mechanisms that have been implicated in cognitive deficits, we proposed to explore this major hypothesis.
Therefore, in this study we were interested in exosomes carrying the tau and Aβ proteins, both from the cerebral compartment. These two proteins have also been shown to be involved in neurodegenerative diseases such as AD [10] and; as providing cues to mechanistic pathways, while also serving as biomarkers of neurocognitive risks in patients. It has also been proposed that exosomal cargo may serve as biomarkers of neurocognitive risks in patients suffering from OSAS. If exosomes cargo contain tau and Aβ proteins, the detection of these exosome signature proteins supports the possibility of their role in the generation of cognitive decline.
Our project was to establish serum biomarkers and prevention methods to determine the phenotype at risk of OSAS by establishing a score of risk with transdisciplinary approaches. The objective of this study was to show how exosomes could be used as biomarkers of OSAS and neurocognitive disorders following OSAS by showing their presence in high quantity in the blood sera of elderly patients with apnea, as well as their origin from the brain compartment by the presence of tau and Aβ proteins [15,23].
In addition, this study evaluated the effect of exosomes from a group of elderly apneic patients versus controls patients on an in vitro model of the BBB using permeability measurements. According to the results of this study, we could consider use of exosomes in the diagnosis of OSAS in neurocognitive diseases, with exosomes being secreted well before the onset of symptoms. This is in line with the idea of preventing cognitive disorders and loss of autonomy in the elderly and to promote aging well.
2. Results
2.1. PROOF Subjects Characteristics
Our selected population comprised of elderly patients (Table 1). Two groups: OSAS with severe apnea (AHI > 30) and non-apneic subjects considered as control (AHI < 5). These two groups were similar in terms of age but different in terms of gender, AHI, Oxygen desaturation index (ODI) which is the number of times per hour of sleep that the blood oxygen level drops by a certain degree from baseline, and their hypoxemic load. Each of the participants underwent polysomnography, which allowed the calculation of the AHI index: the number of apneas/hypopneas made per hour of sleep, describing the severity of the apnea. Blood samples were then collected in the same time frame for all participants.
2.2. Characterization of Exosomes - Western Blot and Homemade ELISA
Following precipitation, the exosomes were further purified using a chromatography column. Analysis with the Zeta Sizer confirmed the purification, revealing the presence of exosomes with a size of 108.7 nm and a concentration of 2.3E+7 particles/mL. (Figure 1).
Two methods were used to characterize and then perform comparative quantification on exosome samples obtained by precipitation and chromatography for each patient. Our results showed a higher amount of tetraspanins (CD81, CD9 and CD63) characteristic of exosomes (Figure 2 and Figure 3); ELISA’s of CD81 (** p=0.0026<0.01) in OSAS patients (AHI >30) vs. controls. Similarly, a higher amount of Aβ protein was observed in OSAS patients (** p=0.0014<0.01) vs. controls. On the other hand, there was no significant difference in tau protein abundance in OSAS vs. controls (p=0.1453>0.01); although there was a slight increase in tau in OSAS patients (Figure 3). Five points stand out (Figure 3 red points), reflecting a high level of CD81, Aβ and tau proteins in patients with OSAS, patients with an AHI ranging from 30.3-63.6 (AHI mean: 48.46).
2.3. Impact of Exosomes on the BBB Model - Monolayer of HBEC-5i Endothelial Cells in Conditioned Medium of HA Astrocytes
The protocol was optimized according to the literature and cytotoxicity tests. A time of 5h and a concentration of 10 μg enabled us to observe the effects of exosomes in the model, while maintaining optimal culture conditions. This condition was applied for all permeability tests.
2.3.1. Permeability Tests - Na-F and TEER
After 5h of incubation with 10 μg exosomes on the luminal side of the model, permeability tests were carried out. Firstly, using Na-F, we observed a significant increase in permeability in OSAS (AHI > 30) vs. controls (**p=0.0049<0.01) (Figure 4A). The same observation was made when permeability was measured using the TEER method (***p=0.0009<0.01) (Figure 4B). Five points stand out (Figure 4 red points), reflecting a sharp increase in permeability in OSAS patients, being patients with an AHI between 30.3-63.6 (AHI mean: 48.46).
2.4. TJs Expressions: ZO-1, Claudin-5 and Occludin
The expression of junctions (ZO-1, claudin-5 and occludin) was studied after 5h incubation of HBEC-5i cells cultured in 96-well plates with 10 μg of exosomes. Claudin-5 expression decreased significantly (*** p=0.0008<0.01) in OSAS patients (AHI> 30) compared to controls (Figure 5B). ZO-1 expression (**p=0.001<0.01) decreased less than claudin-5 expression but significantly in OSAS patients (AHI> 30) compared to controls (Figure 5A). Concerning occludin expression, despite a slight decrease in expression in OSAS patients (AHI> 30) (Figure 5C), there were no significant differences compared to controls (p=0.3126>0.01).
2.5. Correlation Tests
The following data were correlated using Pearson's coefficient: AHI, ODI, CD81 expression, tau expression, Aβ expression, apparent permeability: NaF, ZO-1 expression, claudin-5 expression and occluding expression. We performed correlation significance tests and discussed Pearson correlation coefficients with a p value < 0.01 (Figure 6A).
There was a strong positive correlation between AHI and ODI (R2 = 0.92), reflecting the fact that the higher the AHI, the higher the ODI and therefore the greater the oxygen saturation. AHI was also positively correlated with CD81 expression (R2 = 0.71), showing that the more severe the patient's apnea, and therefore the higher the AHI, the greater the quantity of exosomes found in the patient's blood serum. Since AHI was also positively correlated with permeability (Na-F) (R2 = 0.66) (Figure 6B), we could understand that a high AHI induced a high secretion of exosomes as well as an alteration of the BBB, also seen through the negative correlation of junctions: claudin-5 (R2 = -0.65). ODI is therefore positively correlated with CD81 expression (R2 = 0.67), and with permeability (Na-F) (R2 = 0.51). ODI is also correlated with Aβ expression (R2 = 0.51) which is a fairly low coefficient but with a p value < 0.01. As AHI, ODI is negatively correlated with claudin-5 (R2 = -0.65).
CD81 expression was also strongly and positively correlated (R2 = 0.89) with permeability (Na-F) (Figure 6C), allowing us to assert that a high AHI induced greater CD81 expression, which in turn is at the origin of BBB alteration.
Finally, we note that the expression of Aβ correlated with a decrease in the expression of the junctional protein ZO-1 (R2 = -0.69). No correlation was observed for tau protein expression or for occludin junction protein expression.
3. Discussion
Our study aimed to investigate the link between circulating exosomes and their cargos in brain endothelial dysfunction in severe OSAS elderlies. The integrity of the BBB is crucial for the maintenance of efficient cerebral homeostasis. Disruption of this integrity has already been shown in aging and associated with various neurological disorders as well as OSAS [24,25,26].
Indeed, recent studies have demonstrated that specific diseases or intermittent chronic stress as sleep apnea/hypopnea can disrupt brain endothelial TJs [26] and affect cognition [12]. It has been shown in our laboratory that blood serum from elderly apneic patients in contact with an in vitro model of the BBB induced an alteration of the BBB resulting in an increase in permeability of the model, and opened in vitro BBB barrier (Voirin and al. 2020). Previous work suggested that OSAS may play a key role in the emergence of cognitive disorders, notably through increased BBB permeability via modulation of Nrf2 expression and dysregulation of ABC transporters. (Zolotoff and al. 2020).
Our study highlighted the potential of serum exosomes as biomarkers of OSAS, revealing significantly higher levels of CD81, CD9 and CD63 proteins in elderly OSAS patients compared to elderly healthy subjects. Elderly patients with an AHI > 30 and a mean of 48.5 had much higher quantities of exosomes in their blood serum, making our study original since we understand that there is a potential correlation threshold between AHI, ODI (i.e. the severity of the sleep related breathing disorders) and the quantity of exosomes crossing the BBB and found in blood serum. In fact, AHI and ODI have a Pearson coefficient of 0.909, allowing us to correlate the amount of exosomes with ODI as marker of intermittent hypoxemic load.
The study of exosomes and their cargos could provide significant valuable insight into the mechanism behind cell-to-cell communication and disease development and progression in OSAS. Exosomes are extracellular vesicles involved in intercellular communication. Their implications could be crucial in the pathogenesis of neurological disorders by facilitating a pro-inflammatory phenotype and compromising the integrity of the BBB [15] . Our in vitro experimentation revealed that exposure of BBB models to similar amounts of exosomes from apneic vs. control elderly subjects significantly increased BBB permeability, suggesting a specific OSAS-related detrimental interaction of exosomes on the barrier. The present study showed that exosomes from elderly patients with severe OSAS induced significant increases in permeability of the BBB model, reflecting the high BBB toxicity of exosomes from patients with such diseases.
Processes of BBB alteration could be mediated via exosomal-related biological activities directly impacting the BBB permeability and functionality (Khalyfa, Kheirandish-Gozal, and Gozal 2018)(Chen and al. 2016). The opening of the BBB and subsequent infiltration of serum components to the deep brain can lead to a host of processes resulting in progressive synaptic, dendritic neural soma, dysfunction, and detrimental neuroinflammation environmental changes as microglial pre-activation previously demonstrated in a mouse OSAS model [28]. Such processes have been implicated in different diseases, including vascular dementia, AD, stroke, Parkinson’s disease, multiple sclerosis, amyotrophic lateral sclerosis, severe hypoxia, ischemia, and diabetes mellitus (Alkhalifa and al. 2023)(Knox and al. 2022)(Michalicova, Majerova, and Kovac 2020).
The BBB is an essential structure protecting the CNS by regulating blood flow to the brain, and paracellular transport is notably regulated by junction proteins such as ZO-1, claudin-5 and occludin. A decrease in expression of these junctions in contact with exosomes, in subjects with an AHI > 30, leads to BBB embrittlement [31,32].
ZO-1 and claudin-5 proteins play a major role in BBB development [33]. ZO-1 and claudin-5 show a significant decrease in expression when endothelial cells encounter (direct contact exposure) exosomes in elderly apneic patients, compared to controls, reflecting BBB dysfunction. Alteration of ZO-1 expression has already been shown to be associated with Aβ protein expression in AD [34]. On the other hand, there was no significant difference concerning occludin expression, as is often the case in the literature [35], despite the finding of an alteration in these TJs involved in maintaining BBB integrity.
Studies have established possible correlations between OSAS and neurodegenerative diseases such as AD [10], notably by investigating levels of the proteins Aβ and tau, marked by their accumulation in the brains of patients with AD [36,37]. These proteins, usually measured in cerebrospinal fluid, can also be detected in blood via exosome analysis, enabling the use of a less invasive and equally significant method [21,38]. Indeed, as well as being less invasive for the patient, increasingly research is looking into the use of blood biomarkers, notably in AD and other neurodegenerative diseases, to enable better and often earlier diagnosis [39]. Fiandaca and al [16] supports the fact that detection in individuals of elevated amyloid and tau exosomal proteins in the blood could reflect a current or future neurodegenerative disease.
Markedly elevated levels of Aβ protein in exosomes from elderly apneic subjects suggest a possible direct correlation between OSAS and neurological changes, corroborating the findings of Sun et al. [10]. These authors demonstrated that intermittent hypoxia causes a decrease in alpha secretase activity and an increase in beta secretase, thereby increasing the production of Aβ in the brain and its secretion into exosomes as they form in the brain. Although we observed an increase in tau protein we observed in the exosomes of apneic subjects, it did not reach statistical significance in comparison with healthy non-apneic subjects, possibly due to the high prevalence of increased plasmatic level this protein in our elderly cohort, including in the control group; and indeed this hypothesis was proposed in the literature [10,40].
OSAS induces sleep fragmentation, which disrupts sleep and the recycling of metabolic waste products. Reduced glymphatic activity hampers the process of clearing Aβ and tau proteins, enabling us to make a direct link between OSAS and AD [41]. In addition, intermittent hypoxia causes oxidative stress and inflammation, potentially contributing to the accumulation of these neurotoxic proteins [42,43]. As before, exosomes from elderly patients with an AHI > 30 and a mean of 48.46 induced significant increases in Aβ within exosomes. This suggests that there is an AHI or hypoxemic load threshold above which the patient is at a high risk of developing cognitive impairment, and that exosomes alter the BBB in a way that is clearly more toxic in elderlies or adults above this threshold. In fact, the more exosome concentration the patients have, the more likely they are to alter the BBB, which will no longer play its filtration role, and the greater the quantities of Aβ found inside the exosomes. Moreover, the high levels of Aβ are also associated with poor recycling during sleep, as mentioned above. With regard to tau protein, elderly patients with severe OSAS showed a marked but non-significant increase in the presence of such biomarker.
This study highlights the possible role of exosomes in mediating negative OSAS impacts on the BBB. This has significant implications for understanding how the risk of developing cognitive disorders might be assessed and anticipated. The use of exosomal biomarkers from blood serum in combination with AHI is particularly important for detecting high-risk individuals. These findings suggest that exosomes could be used for early neurological risk assessment, with the ability to detect changes up to 10 years before the emergence of clinical symptoms. [16].
It is essential to show the importance of early detection of markers of BBB dysfunction, such as exosomes carrying Aβ and tau proteins, for the introduction of therapeutic strategies aimed to limit or stop progression to cognitive impairment. Early treatment of OSAS, notably with CPAP, has demonstrated a reduction in Aβ and tau levels, highlighting the reversible potential of these biomarkers in response to targeted therapeutic intervention [10].
However, our study is limited by the fact that our target population is the elderly. It might be relevant to compare our results with a cohort of younger OSAS patients, as we hypothesize that the amount of tau protein recovered is age-related, so we might be able to observe differences in younger subjects. Furthermore, a study of the amount of phosphorylated tau could have provided us with additional information, but this protein is more difficult to assay. Elderly subjects are exposed to more comorbidities and therefore naturally have a greater quantity of serum exosomes. The gender factor also comes into play, with a higher proportion of women than men. It would also be useful to supplement our study with an analysis of synchronized neuropsychological tests and brain MRI or PET imaging [44]. It should be noted that our tests were carried out on in vitro models of the BBB made up of an hBEC-5i cell type and conditioned astrocyte medium; however, to get as close as possible to reality, we would like to develop a tri-culture model involving pericytes which are known to play an important role in aging, which is our target population [45]. According to the literature, we have chosen to expose in vitro BBB models to 10 ug of exosome concentration, which does not necessarily reflect the amount each individual possesses. Exosomes contain a wide variety of molecules, including miRNAs (e.g.; miR-132), and the study of these miRNAs could enable us to make even more precise studies with a view to developing new diagnostics and treatments.
4. Materials and Methods
4.1. Subjects
The blood sera used was from the PROOF cohort study in Saint-Etienne (France). This cohort was set up in 2001 and recruited 1011 subjects aged 65 years at the time of inclusion. The aim of this study was to evaluate the role and the alteration of the autonomic nervous system in the occurrence of cardiovascular or cerebrovascular events, and of dementia over 20 years of follow-up [46]. The PROOF study was made possible by an association with the SYNAPSE study, which focused on approximatively the same sample group on sleep-disordered breathing and CNS incident disorders. Thirty sera were selected for our study: 15 from elderly patients with severe apnea OSAS (AHI > 30) and 15 from elderly patients without apnea and no OSAS (AHI < 5). Several studies have already been performed using the PROOF cohort, approved by the CCPRB Rhône Alpes Loire IRB N532016 and IRB2002/22/CHUSETE and agreement was given by the CNIL (national committee for information and liberty) for data collection. The participants gave their written consent to participate in the PROOF studies.
4.2. Biological Samples
Blood samples were collected in the morning after recording by polysomnography during the night. Then after centrifugation and recovery of the serum, it was immediately frozen at -80 °C after having made each sample anonymous according to good practices. Clinical trial numbers of SYNAPSE and PROOF studies are NCT00766584 and NCT 00759304.
4.3. Chemicals and Reagents
4.3.1. Cell Culture
Endothelial cell growth supplement (ECGS) was obtained from Sigma-Aldrich; foetal bovine serum (FBS) was from Eurobio Scientific; trypsin, amphotericin B and penicillin-streptomycin solutions were from Corning; human brain endothelial cells HBEC-5i were obtained from ATCC; human astrocyte cells HA and astrocyte medium AM were from CliniSciences; GIBCO Dulbecco’s Modified Eagle Medium/Nutriment Mixture (DMEM-F12) was from Fisher Scientific; 24-well inserts (transparent PET membrane, 0.45-μm pore diameter size) and all consumable for cell culture came from D.Dutscher.
4.3.2. Permeability Measurement
EVOM volt ohmmeter system was purchased from World Precision Instruments; all compounds for Ringer HEPES buffer and sodium-fluorescein sodium (Na-F) were from Sigma-Aldrich.
4.3.3. Exosome Isolation & Characterization
EX02-50-Exospin blood with 48 columns were from Cell Guidance. Bicinchoninic acid (BCA) protein assay kit for low concentrations were from Abcam. MiniPROTEN Tetracell was from Biorad, Pierce ECL Western Blotting Substrate, PageRuler Plus Prestained Lader, Phosphatase and proteases inhibitors and 0.2-μm PVDF membrane were from Thermo Fisher Scientific, RIPA buffer, blocking buffer (with BSA and PBS Tween) and all compounds for Laemeli solution were from Sigma-Aldrich.
4.3.4. ELISA
Recombinant human tau441 protein from Abcam, Aβ and CD81 proteins came from kits: the anti-β-Amyloid kit was from Thermo Fisher Scientific and the ExoELISA-ULTRA CD81 kit was from Systeme Biosciences. Tetramethylbenzidine (TMB) was from Sigma-Aldrich. ELISA buffer kit was purchased from Thermo Fisher Scientific.
For all antibodies, see Table 2.
4.4. Blood Serum Exosome Isolation and Purification
Exosomes were isolated from human sera according to the manufacturer's protocol by Cell Guidance Systems (EX02-50). The end step was purification by passage to the steric size exclusion chromatography column. The column is composed of pores about 30 nm in resin according to the kit recommendations. The exosomes could be used immediately; for long term storage the exosomes were frozen at -80 °C.
4.5. Protein Measurements
Isolated exosomes samples were dispersed in 96-well plates, to which a BCA solution was added. The realization of a standard range from a solution of BSA at 2 mg/mL provided in the kit, to which a solution of BCA was also added in equivalent quantity to what was added to the samples of exosomes. This will made it possible to establish a standard linear curve in order to calculate the total protein concentration of each sample. After incubation for 2 h at 37 °C, a reading was carried out at a wavelength of 560-nm.
4.6. Exosomes Characterization
4.6.1. Western Blot
The characterization of exosomes was done using the western blot method: a semi-quantitative technique allowing the detection of specific proteins within a sample. 30 μg of exosomes were used per migration well. The freshly isolated exosome samples were sonicated and mixed with 1X RIPA lysis buffer and 2X Laemmli solution before being introduced into 1.5-mm SDS-PAGE gel wells (15% separation gel and 4% concentration gel). Migration was performed for 30 min at 0.04 A and 1h at 0.07 A. The next transfer was done on a 0.2-μm PVDF membrane for 1h at 100 V. Subsequently the membrane was saturated for 1h in blocking buffer made from BSA powder and PBS Tween20. The PVDF membrane was then incubated overnight at 4 °C with antibodies to CD81, CD63, CD9, tau and GAPDH at 1/100 dilution and to Aβ at 1/200 dilution. After washing, the PVDF membrane was incubated for 1h at room temperature with the secondary antibody m-IgGκBP-HRP at 1/500 dilution.
Revelation was performed by chemiluminescence using an ECL kit.
4.6.2. Homemade ELISA
The western blot characterization method was completed with an ELISA method via the implementation of an inhouse anti-CD81, Aβ and tau ELISAs (results compared with the anti-CD81ExoELISA-ULTRA CD81 kit (System Biosciences: EXEL-ULTRA-CD81-1) and the anti-β-Amyloid 42 kit (Thermo Fisher scientific: KHB3441).
A standard range was performed using CD81, Aβ and tau standard proteins, giving us standard curves. In 96-well plates, the wells were coated with a mixture of 50 μg of exosomes and phosphate coating buffer for 2 nights at 37 °C. Exosomes were incubated for 1h with antibody anti-CD81 at 1/500 dilution and antibodies to Aβ and tau at 1/1000 dilution. Then HRP secondary antibody diluted to 1/10000 was added to each well and incubated for 1h at room temperature. Revelation was done with TMB 10 minutes. Then a stop solution was added to stop the reaction. The plate was read with an absorption spectrophotometer at a wavelength of 450-nm. Coating buffer, wash buffer, assay buffer (for antibody dilution), TMB and stop solution were supplied in the ELISA buffer kit (Thermo Fisher Scientific).
4.6.3. Nanoparticle Tracking Analysis (NTA)
A few millimeters of exosomes diluted in PBS were analyzed using a Zeta Sizer instrument (ZetaView, particle metrix Twin colocalization, EXCILONE, Elancourt FRANCE). The exosomes were derived from the blood serum of elderly apneic patient. During the analysis, the temperature was 29.99°C, pH 7.0 and conductivity was 15,000 μS/cm. The instrument settings included a laser wavelength of 488 nm, a diffusion filter, a sensitivity of sensitivity 80.0 and a shutter speed of 100.
4.7. In Vitro BBB Model
The BBB model was composed of endothelial cells HBEC-5i cultivated in PET transwells with 0.45-μm pores, in contact with HA conditioned medium as previously described [13,26]. Initially, HBEC-5i were cultivated in DMEM/F12 HAM, containing 1% antibiotic solutions (amphotericin B and penicillin-streptomycin), 10% FBS and 40 ug/mL of ECGS. HBEC-5i must be used before passage number 20 to maintain all BBB properties necessary to our model. Regarding HA, cells were initially cultured in AM supplemented with 1% penicillin-streptomycin solution, 1% AGS and 2% FBS. Hereafter, HA were cultivated with the same medium as HBEC-5i. HA and HBEC-5i were incubated (5% CO2 and at 37 °C). The BBB model was composed of 2x105 HBEC-5i per insert, in contact with conditioned medium for 14 days: the time necessary to see an optimal permeability (Papp) of the model, which was stable for 5 days. The HA conditioned medium corresponded to the medium that has remained in contact with HA for 48h. We noted that the medium was renewed every 2 days and this was interesting because of all factors and chemicals compounds released by HA which may have effects on endothelial cells and on our BBB model permeability [13].
4.8. Permeability Measurements – Barrier Properties
4.8.1. Trans Endothelial Electrical Resistance (TEER)
To analyze the effects of exosomes on the BBB, first we tested the tightness of our in vitro BBB model under 2 conditions: inserting it in contact with exosomes from blood serum of elderly apneic vs. elderly control. 10 μg of exosomes were added to the luminal upper side endothelial cells. The exosomes were mixed with DMEM-F12 medium without FBS and incubated for 5h. Then we tested the TEER using an EVOM resistance meter on the endothelial cell monolayer. This method consists of calculating the potential difference between the two poles using electrodes: one electrode on the luminal side and a second on the abluminal side. For each well measurement, three values were noted. After measurement, the following formula was applied:
(TEER – control TEER) x 0.33
With 0.33 being the membrane area and the control TEER as a blank filter without cells.
Ideally, the TEER should be around 40 Ω/cm2, as previously described [13].
4.8.2. Sodium-Fluorescein
After measuring the TEER, we performed a new method to test the apparent permeability Papp of our model, using Na-F, which is a hydrophilic fluorescent molecule (MW: 376 Da). A solution of Ringer HEPES buffer diluted with 10 μg/mL of Na-F were placed onto the luminal upper side for 1h and incubated, while the abluminal side contained only Ringer HEPES solution.
Then the solution was removed from the abluminal side, and the fluorescence was measured using a fluorescence spectrophotometer (Fluoroskan AscentTM, Thermo Fisher Scientific), with 485-nm excitation and 530-nm emission wavelengths. Papp is measured in cm/s and is calculated using the formula used in our previous laboratory study [26].
Papp is the apparent permeability, Vr is the volume of medium in the abluminal side, S is the monolayer’s area, and C0 and C1 are the concentration of fluorescent compound in the luminal chamber at t and in the abluminal side after t time -h of incubation, respectively.
4.9. Protein Expression and Quantification
Endothelial cells were seeded at a density of 2x104 HBEC-5i per well, into 96-well permeable plates and incubated with DMEM-F12. At confluence, 10 μg of exosomes were added to each well during 5h. Then, cells were fixed with 4% paraformaldehyde and permeabilized with a solution of H2O2 and methanol. ZO-1, claudin-5 and occludin expressions were analyzed with antibodies of diluted ZO-1(4 μg/ml), claudin-5 (2 μg/ml) and occludin (1 μg/ml) respectively, at room temperature for 2h. Secondary IgG antibody diluted 1/2500 for anti-rabbit and 1/500 for anti-mouse was incubated at room temperature for 2h. Then, TMB substrate was added into each well for 15 minutes. To finish, hydrochloric acid was added and stopped the reaction before doing measurements at 450-nm.
4.10. Statistics Analysis
Statistical analysis was realized using t test unpaired by GraphPad software. The differences between means were considered to be significant when p values were < 0.05. Results are presented as sample mean +/- SEM.
5. Conclusions
Our results establish a strong link between sever OSAS and BBB dysfunction in the elderly. Exosomes could represent an interesting biomarker of the dysfunction of BBB in such patients, opening the way to innovative diagnostic and therapeutic approaches based on serum exosome analysis. Further research is needed to elucidate the precise mechanisms by which exosomes alter BBB permeability and to develop clinical methods for the routine isolation and characterization of these vesicles, in order to better personalize the management of OSAS patients. An initial idea for thresholding severe apneic subjects could help predict the alteration of the BBB and possibly the consecutive cognitive dysfunctions.
Author Contributions
P.G. designed the study, wrote the main manuscript text and prepared and analyzed the Figures. N.P. F.R. and N.B. designed the study and read and corrected the article. F.R. analyzed the records. S.C. prepared and analyzed the Table. All authors have read and agreed to the published version of the manuscript.
Funding
This research was carried out as part of a CIFRE (Convention Industrielle de Formation par la Recherche) thesis, funded by Gérontopôle AURA in partnership with the Inserm U1059 Sainbiose laboratory, and supported by the Association Nationale de la Recherche et de la Technologie (ANRT).
Institutional Review Board Statement
Clinical trial numbers of SYNAPSE and PROOF studies are NCT00766584 and NCT 00759304. Studies has performed using the PROOF cohort, approved by the CCPRB Rhône Alpes Loire IRB N532016 and IRB2002/22/CHUSETE and agreement was given by the CNIL (national committee for information and liberty) for data collection. The participants gave their written consent to participate in the PROOF studies.
Informed Consent Statement
All participants in the SYNAPSE and PROOF studies gave their informed consent to the publication of anonymized data and study results. Informed consent was obtained from all subjects involved in the study.
Acknowledgments
This work was supported by grants from Jean Monnet University of Saint Etienne and grants from DRCI of Saint-Etienne University Hospital (PHRC National and PHRC Regional), France. The authors thank Christian Moritz (SYNATAC), Cindy Zolotoff (PhD), Chloé Mounichetty (PhD), the Association Synapse, Saint-Etienne, France (Martine Borel: current President), and all of the participants in the study.
Conflicts of Interest
The authors declare no conflict of interest (financial or nonfinancial).
References
- Norman D, Loredo JS. Obstructive Sleep Apnea in Older Adults. Clin Geriatr Med. févr 2008;24(1):151-65. [CrossRef]
- Beaudin AE, Waltz X, Hanly PJ, Poulin MJ. Impact of obstructive sleep apnoea and intermittent hypoxia on cardiovascular and cerebrovascular regulation: Obstructive sleep apnoea, intermittent hypoxia and vascular regulation. Exp Physiol. 1 juill 2017;102(7):743-63.
- Sforza E, Roche F. Sleep Apnea Syndrome and Cognition. Front Neurol [Internet]. 2012 [cité 13 juin 2023];3. Disponible sur: http://journal.frontiersin.org/article/10.3389/fneur.2012.00087/abstract. 10.3389/fneur.2012.00087.
- Launois SH, Pépin JL, Lévy P. Sleep apnea in the elderly: A specific entity? Sleep Med Rev. avr 2007;11(2):87-97.
- Bucks RS, Olaithe M, Eastwood P. Neurocognitive function in obstructive sleep apnoea: A meta-review: Cognitive function in OSA: a meta-review. Respirology. janv 2013;18(1):61-70.
- Sforza E, Roche F, Thomas-Anterion C, Kerleroux J, Beauchet O, Celle S, et al. Cognitive Function and Sleep Related Breathing Disorders in a Healthy Elderly Population: the Synapse Study. Sleep. avr 2010;33(4):515-21. [CrossRef]
- Berry RB, Budhiraja R, Gottlieb DJ, Gozal D, Iber C, Kapur VK, et al. Rules for Scoring Respiratory Events in Sleep: Update of the 2007 AASM Manual for the Scoring of Sleep and Associated Events: Deliberations of the Sleep Apnea Definitions Task Force of the American Academy of Sleep Medicine. J Clin Sleep Med. 15 oct 2012;08(05):597-619.
- Guerra S, Carsin AE, Keidel D, Sunyer J, Leynaert B, Janson C, et al. Health-related quality of life and risk factors associated with spirometric restriction. Eur Respir J. mai 2017;49(5):1602096. [CrossRef]
- Sforza E, Roche F, Chapelle C, Pichot V. Internight Variability of Apnea-Hypopnea Index in Obstructive Sleep Apnea Using Ambulatory Polysomnography. Front Physiol. 9 juill 2019;10:849. [CrossRef]
- Sun H, Gao Y, Li M, Zhang S, Shen T, Yuan X, et al. Altered amyloid-β and tau proteins in neural-derived plasma exosomes in obstructive sleep apnea. Sleep Med. juin 2022;94:76-83. [CrossRef]
- Kerner NA, Roose SP. Obstructive Sleep Apnea is Linked to Depression and Cognitive Impairment: Evidence and Potential Mechanisms. Am J Geriatr Psychiatry. juin 2016;24(6):496-508. [CrossRef]
- Lim DC, Pack AI. Obstructive sleep apnea and cognitive impairment: Addressing the blood–brain barrier. Sleep Med Rev. févr 2014;18(1):35-48. [CrossRef]
- Puech C, Hodin S, Forest V, He Z, Mismetti P, Delavenne X, et al. Assessment of HBEC-5i endothelial cell line cultivated in astrocyte conditioned medium as a human blood-brain barrier model for ABC drug transport studies. Int J Pharm. nov 2018;551(1-2):281-9. [CrossRef]
- Daneman R, Prat A. The Blood–Brain Barrier. Cold Spring Harb Perspect Biol. janv 2015;7(1):a020412.
- Khalyfa A, Kheirandish-Gozal L, Gozal D. Circulating exosomes in obstructive sleep apnea as phenotypic biomarkers and mechanistic messengers of end-organ morbidity. Respir Physiol Neurobiol. oct 2018;256:143-56. [CrossRef]
- Fiandaca MS, Kapogiannis D, Mapstone M, Boxer A, Eitan E, Schwartz JB, et al. Identification of preclinical Alzheimer’s disease by a profile of pathogenic proteins in neurally derived blood exosomes: A case-control study. Alzheimers Dement. juin 2015;11(6):600. [CrossRef]
- Pan W, Kastin AJ. Can sleep apnea cause Alzheimer’s disease? Neurosci Biobehav Rev. nov 2014;47:656-69.
- Baril AA, Carrier J, Lafrenière A, Warby S, Poirier J, Osorio RS, et al. Biomarkers of dementia in obstructive sleep apnea. Sleep Med Rev. déc 2018;42:139-48. [CrossRef]
- Ayloo S, Gu C. Transcytosis at the blood–brain barrier. Curr Opin Neurobiol. août 2019;57:32-8.
- Chen CC, Liu L, Ma F, Wong CW, Guo XE, Chacko JV, et al. Elucidation of Exosome Migration Across the Blood–Brain Barrier Model In Vitro. Cell Mol Bioeng. déc 2016;9(4):509-29. [CrossRef]
- Wood MJ, O’Loughlin AJ, Lakhal S. Exosomes and the blood–brain barrier: implications for neurological diseases. Ther Deliv. sept 2011;2(9):1095-9. [CrossRef]
- Gurung S, Perocheau D, Touramanidou L, Baruteau J. The exosome journey: from biogenesis to uptake and intracellular signalling. Cell Commun Signal. 23 avr 2021;19(1):47. [CrossRef]
- Zhang Y, Bi J, Huang J, Tang Y, Du S, Li P. Exosome: A Review of Its Classification, Isolation Techniques, Storage, Diagnostic and Targeted Therapy Applications. Int J Nanomedicine. sept 2020;Volume 15:6917-34. [CrossRef]
- Cao Y, Xu W, Liu Q. Alterations of the blood-brain barrier during aging. J Cereb Blood Flow Metab. 21 mars 2024;0271678X241240843. [CrossRef]
- Knox EG, Aburto MR, Clarke G, Cryan JF, O’Driscoll CM. The blood-brain barrier in aging and neurodegeneration. Mol Psychiatry. juin 2022;27(6):2659-73. [CrossRef]
- Voirin AC, Celle S, Perek N, Roche F. Sera of elderly obstructive sleep apnea patients alter blood–brain barrier integrity in vitro: a pilot study. Sci Rep. 9 juill 2020;10(1):11309. [CrossRef]
- Intermittent Hypoxia and Its Impact on Nrf2/HIF-1α Expression and ABC Transporters: An in Vitro Human Blood-Brain Barrier Model Study. Cell Physiol Biochem. 16 déc 2020;54(6):1231-48.
- Sapin E, Peyron C, Roche F, Gay N, Carcenac C, Savasta M, et al. Chronic Intermittent Hypoxia Induces Chronic Low-Grade Neuroinflammation in the Dorsal Hippocampus of Mice. Sleep. 1 oct 2015;38(10):1537-46. [CrossRef]
- Alkhalifa AE, Al-Ghraiybah NF, Odum J, Shunnarah JG, Austin N, Kaddoumi A. Blood–Brain Barrier Breakdown in Alzheimer’s Disease: Mechanisms and Targeted Strategies. Int J Mol Sci. 14 nov 2023;24(22):16288.
- Michalicova A, Majerova P, Kovac A. Tau Protein and Its Role in Blood–Brain Barrier Dysfunction. Front Mol Neurosci. 30 sept 2020;13:570045. [CrossRef]
- Ahmad AA, Taboada CB, Gassmann M, Ogunshola OO. Astrocytes and Pericytes Differentially Modulate Blood—Brain Barrier Characteristics during Development and Hypoxic Insult. J Cereb Blood Flow Metab. févr 2011;31(2):693-705. [CrossRef]
- Berezowski V, Landry C, Dehouck MP, Cecchelli R, Fenart L. Contribution of glial cells and pericytes to the mRNA profiles of P-glycoprotein and multidrug resistance-associated proteins in an in vitro model of the blood–brain barrier. Brain Res. août 2004;1018(1):1-9. [CrossRef]
- Katsuno T, Umeda K, Matsui T, Hata M, Tamura A, Itoh M, et al. Deficiency of Zonula Occludens-1 Causes Embryonic Lethal Phenotype Associated with Defected Yolk Sac Angiogenesis and Apoptosis of Embryonic Cells. Mostov K, éditeur. Mol Biol Cell. juin 2008;19(6):2465-75. [CrossRef]
- Zhou J, Camacho M, Tang X, Kushida CA. A review of neurocognitive function and obstructive sleep apnea with or without daytime sleepiness. Sleep Med. juill 2016;23:99-108. [CrossRef]
- Salminen A, Kauppinen A, Kaarniranta K. Hypoxia/ischemia activate processing of Amyloid Precursor Protein: impact of vascular dysfunction in the pathogenesis of Alzheimer’s disease. J Neurochem. févr 2017;140(4):536-49. [CrossRef]
- Rajendran L, Honsho M, Zahn TR, Keller P, Geiger KD, Verkade P, et al. Alzheimer’s disease β-amyloid peptides are released in association with exosomes. Proc Natl Acad Sci. 25 juill 2006;103(30):11172-7.
- Brunello CA, Merezhko M, Uronen RL, Huttunen HJ. Mechanisms of secretion and spreading of pathological tau protein. Cell Mol Life Sci. mai 2020;77(9):1721-44. [CrossRef]
- Jia L, Qiu Q, Zhang H, Chu L, Du Y, Zhang J, et al. Concordance between the assessment of Aβ42, T-tau, and P-T181-tau in peripheral blood neuronal-derived exosomes and cerebrospinal fluid. Alzheimers Dement. août 2019;15(8):1071-80.
- Teunissen CE, Verberk IMW, Thijssen EH, Vermunt L, Hansson O, Zetterberg H, et al. Blood-based biomarkers for Alzheimer’s disease: towards clinical implementation. Lancet Neurol. janv 2022;21(1):66-77.
- Carvalho DZ, St. Louis EK, Schwarz CG, Lowe VJ, Boeve BF, Przybelski SA, et al. Witnessed apneas are associated with elevated tau-PET levels in cognitively unimpaired elderly. Neurology [Internet]. 28 avr 2020 [cité 7 avr 2024];94(17). Disponible sur: https://www.neurology.org/doi/10.1212/WNL.0000000000009315. [CrossRef]
- Andrade AG, Bubu OM, Varga AW, Osorio RS. The Relationship between Obstructive Sleep Apnea and Alzheimer’s Disease. Perry G, Avila J, Moreira PI, Sorensen AA, Tabaton M, éditeurs. J Alzheimers Dis. 12 juin 2018;64(s1):S255-70.
- Ju YES, Lucey BP, Holtzman DM. Sleep and Alzheimer disease pathology—a bidirectional relationship. Nat Rev Neurol. févr 2014;10(2):115-9. [CrossRef]
- Xie L, Kang H, Xu Q, Chen MJ, Liao Y, Thiyagarajan M, et al. Sleep Drives Metabolite Clearance from the Adult Brain. Science. 18 oct 2013;342(6156):373-7.
- Macey PM, Prasad JP, Ogren JA, Moiyadi AS, Aysola RS, Kumar R, et al. Sex-specific hippocampus volume changes in obstructive sleep apnea. NeuroImage Clin. 2018;20:305-17. [CrossRef]
- Berthiaume AA, Schmid F, Stamenkovic S, Coelho-Santos V, Nielson CD, Weber B, et al. Pericyte remodeling is deficient in the aged brain and contributes to impaired capillary flow and structure. Nat Commun. 7 oct 2022;13(1):5912. [CrossRef]
- Barthélémy JC, Pichot V, Dauphinot V, Celle S, Laurent B, Garcin A, et al. Autonomic Nervous System Activity and Decline as Prognostic Indicators of Cardiovascular and Cerebrovascular Events: The ‘PROOF’ Study. Neuroepidemiology. 2007;29(1-2):18-28. [CrossRef]
Figure 1.
Characterization of exosomes using NTA zeta sizer instrument. (A) Particles in mL as a function of diameter in nm, (B) Visualization of exosomes.
Figure 1.
Characterization of exosomes using NTA zeta sizer instrument. (A) Particles in mL as a function of diameter in nm, (B) Visualization of exosomes.
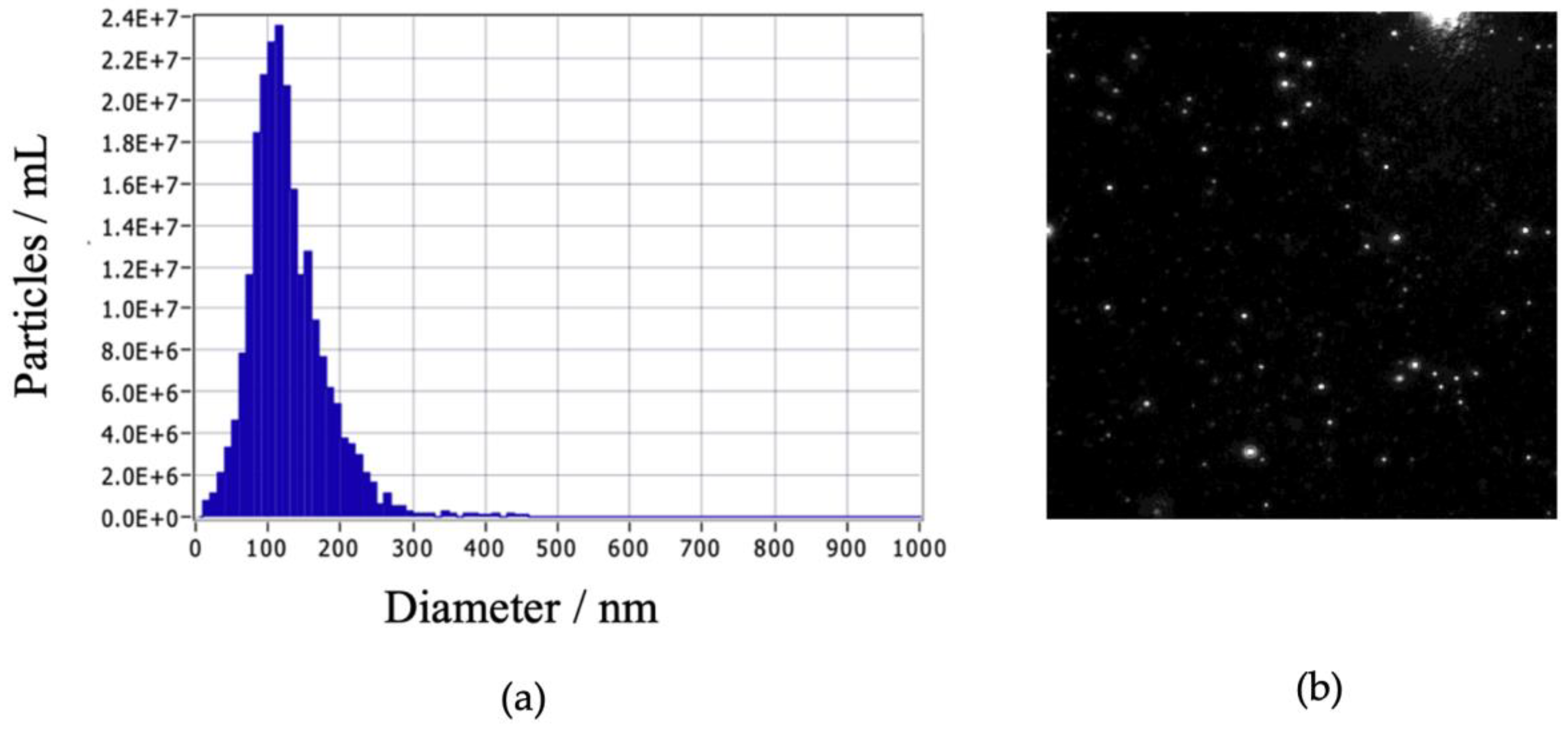
Figure 2.
Western blot analysis of the marker protein CD81 (A), CD63 (B), CD9 (C), Amyloid beta (D), Tau (E) and GAPDH as control (F), on exosomes isolated from patients without OSAS (AHI < 5) (G) vs. patients with OSAS (AHI >30) (H). Semi quantification normalized with GAPDH (G) (H), results obtained and analyzed with ImageJ.
Figure 2.
Western blot analysis of the marker protein CD81 (A), CD63 (B), CD9 (C), Amyloid beta (D), Tau (E) and GAPDH as control (F), on exosomes isolated from patients without OSAS (AHI < 5) (G) vs. patients with OSAS (AHI >30) (H). Semi quantification normalized with GAPDH (G) (H), results obtained and analyzed with ImageJ.
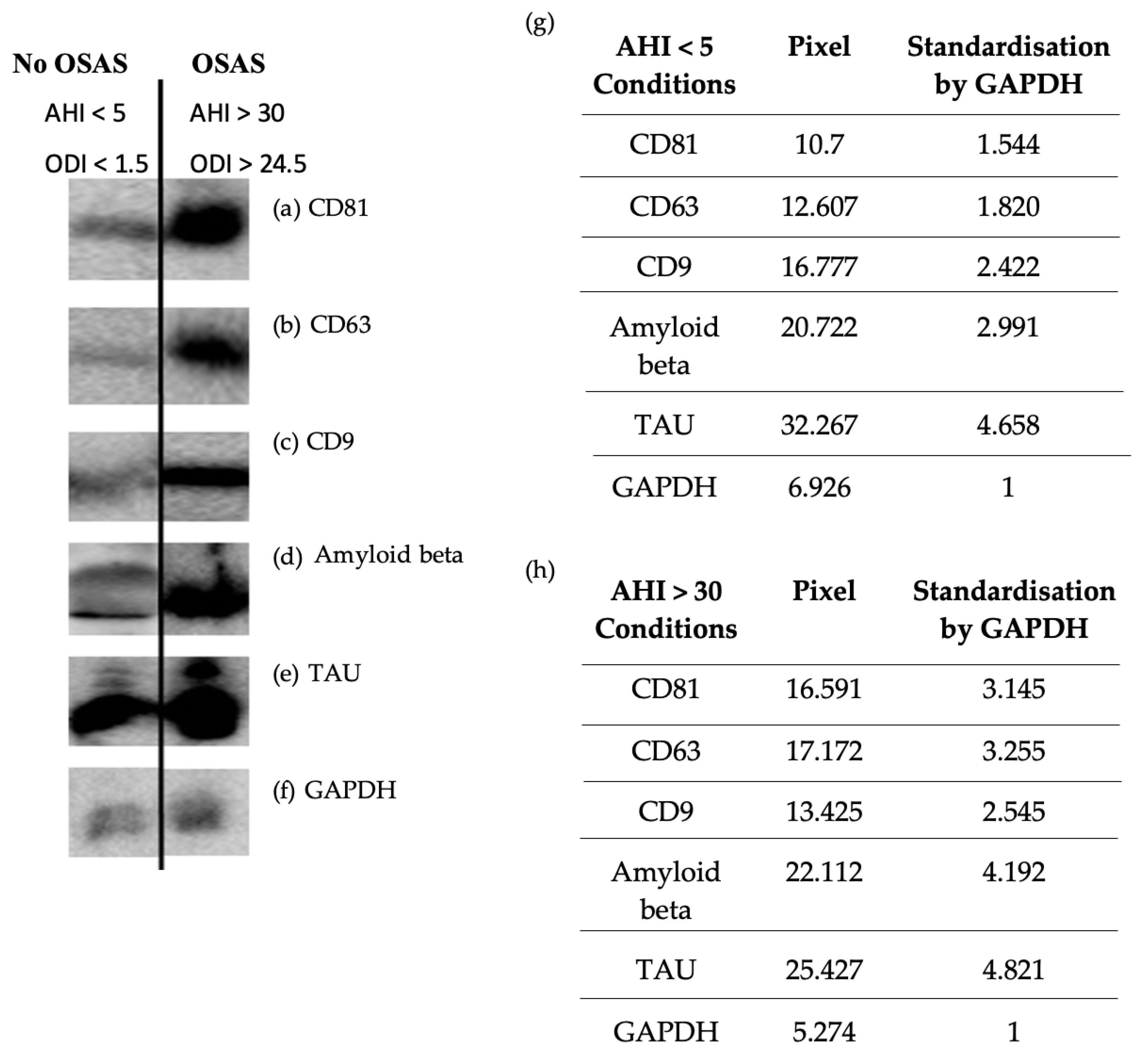
Figure 3.
Homemade ELISAs. Results are represented as mean value +/- s.e.m (n=15 triplicate). Expression of CD81 protein (A)** p=0.0026<0.01 between AHI < 5 vs. AHI > 30. Expression of Amyloid beta protein (B)** p=0.0014<0.01 between AHI < 5 vs. AHI > 30. Expression of total TAU protein (C) p=0.1453>0.01 not significant between AHI < 5 vs. AHI > 30.
Figure 3.
Homemade ELISAs. Results are represented as mean value +/- s.e.m (n=15 triplicate). Expression of CD81 protein (A)** p=0.0026<0.01 between AHI < 5 vs. AHI > 30. Expression of Amyloid beta protein (B)** p=0.0014<0.01 between AHI < 5 vs. AHI > 30. Expression of total TAU protein (C) p=0.1453>0.01 not significant between AHI < 5 vs. AHI > 30.
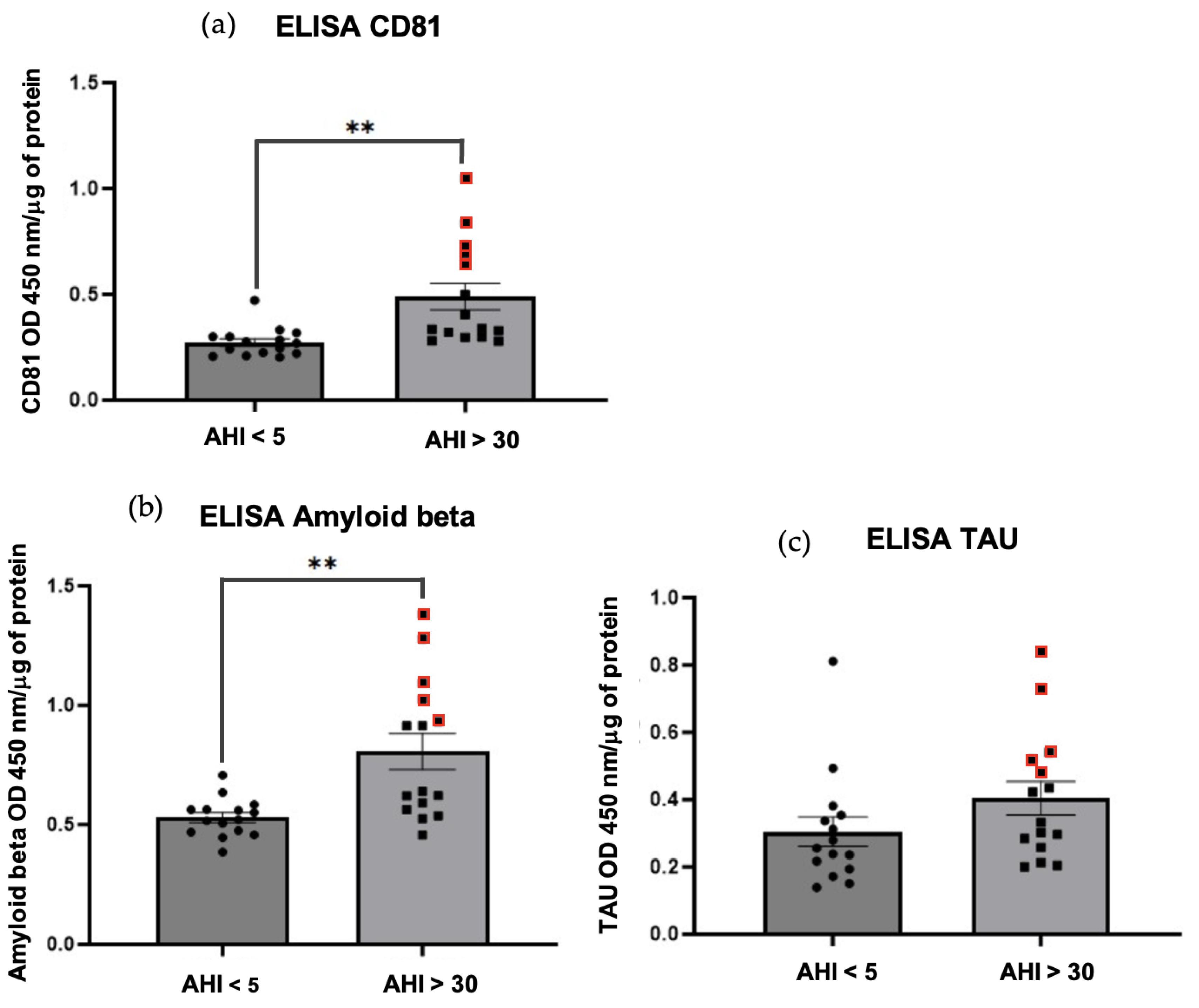
Figure 4.
Membrane permeability measurement of HBEC-5i after 5 hours of 10 μg exosomes from patients without OSAS AHI < 5 vs. patients with OSAS AHI >30. (A) NaF: sodium fluorescein - results are represented as mean value +/- s.e.m (n=15 triplicate) **p=0.0049<0.01 between AHI < 5 vs. AHI > 30. (B) TEER: transendothelial electrical resistance - results are represented as mean value +/- s.e.m (n=15 triplicate) ***p=0.0009<0.01 between AHI < 5 vs AHI > 30.
Figure 4.
Membrane permeability measurement of HBEC-5i after 5 hours of 10 μg exosomes from patients without OSAS AHI < 5 vs. patients with OSAS AHI >30. (A) NaF: sodium fluorescein - results are represented as mean value +/- s.e.m (n=15 triplicate) **p=0.0049<0.01 between AHI < 5 vs. AHI > 30. (B) TEER: transendothelial electrical resistance - results are represented as mean value +/- s.e.m (n=15 triplicate) ***p=0.0009<0.01 between AHI < 5 vs AHI > 30.
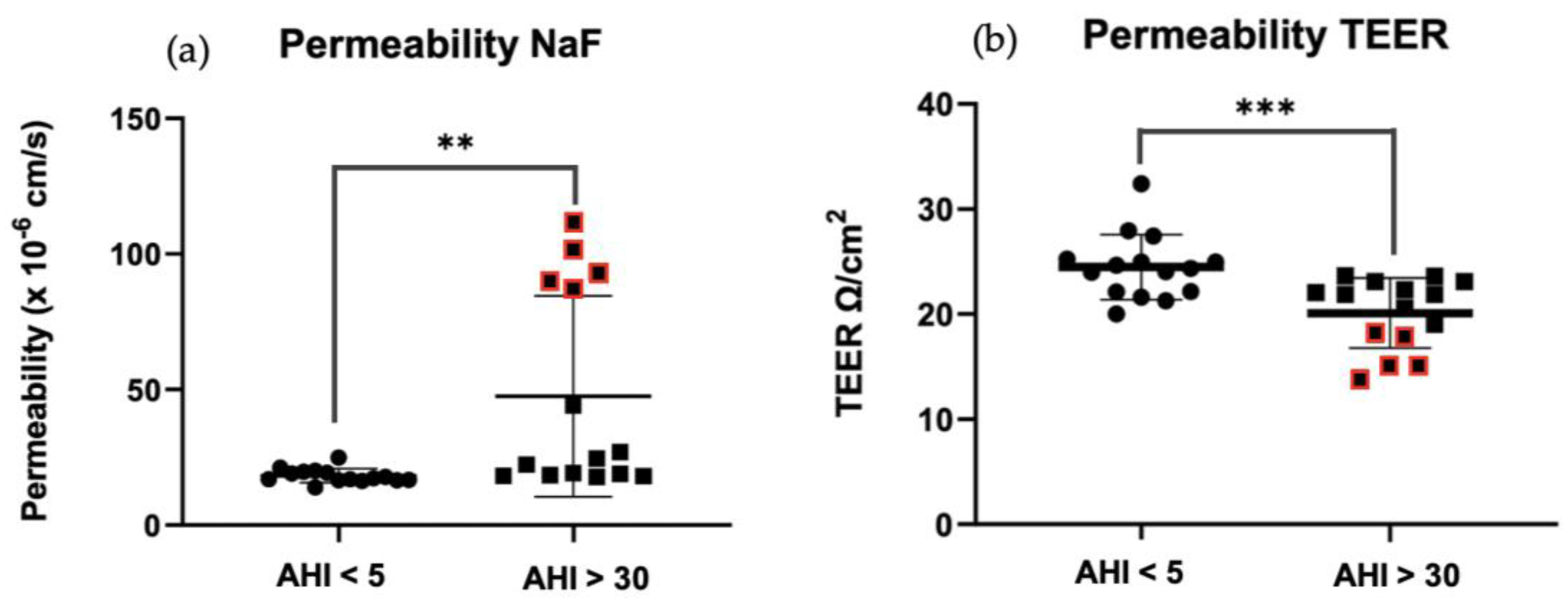
Figure 5.
Whole cell ELISA assay. Results are represented as mean value +/- s.e.m (n=10 triplicate). Expressions of ZO-1(A)**p=0.001<0.01, claudin-5 (B)*** p=0.0008<0.01, and occludin (C) p=0.3126> 0.01 proteins after exposure of HBEC-5i 5 hours of 10 μg exosomes from patients without OSAS AHI < 5 vs. patients with OSAS AHI >30.
Figure 5.
Whole cell ELISA assay. Results are represented as mean value +/- s.e.m (n=10 triplicate). Expressions of ZO-1(A)**p=0.001<0.01, claudin-5 (B)*** p=0.0008<0.01, and occludin (C) p=0.3126> 0.01 proteins after exposure of HBEC-5i 5 hours of 10 μg exosomes from patients without OSAS AHI < 5 vs. patients with OSAS AHI >30.

Figure 6.
Pearson correlation coefficient R2 between parameters : AHI (Apnea Hypopnea Index) ODI (Oxygen Desaturation Index), CD81 tetraspanin, Tau protein, Aβ protein, Na-F permeability, ZO-1 protein, claudin-5 protein, occludin protein. (A): Correlation significance tests with a p value < 0.01. Pearson correlation coefficient between Na-F and AHI (B). Pearson correlation coefficient between Na-F and CD81 (C). .
Figure 6.
Pearson correlation coefficient R2 between parameters : AHI (Apnea Hypopnea Index) ODI (Oxygen Desaturation Index), CD81 tetraspanin, Tau protein, Aβ protein, Na-F permeability, ZO-1 protein, claudin-5 protein, occludin protein. (A): Correlation significance tests with a p value < 0.01. Pearson correlation coefficient between Na-F and AHI (B). Pearson correlation coefficient between Na-F and CD81 (C). .
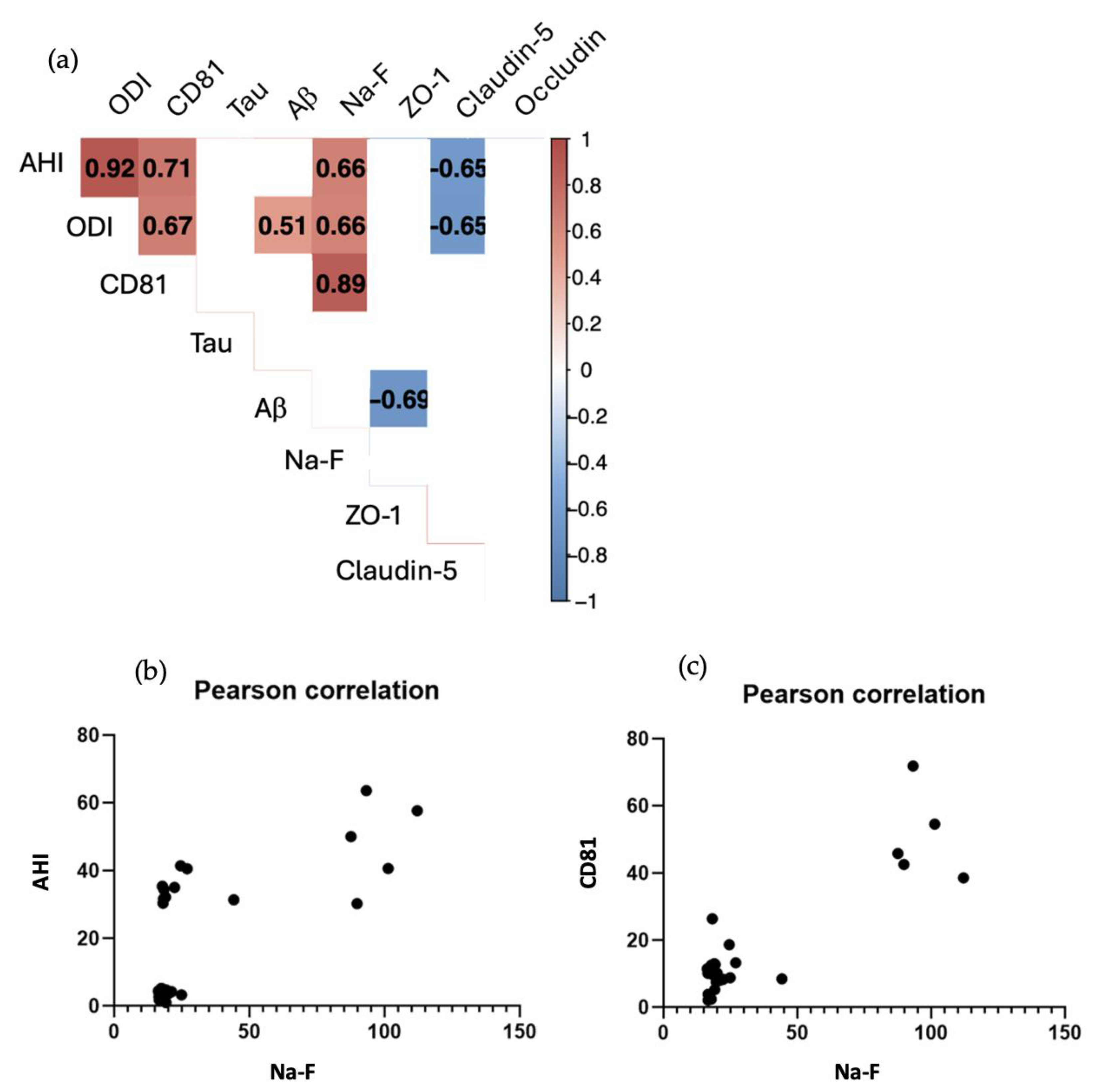

Table 1.
Descriptive characteristics of the population studies from the PROOF cohort. F: female, M: male, AHI: apnea hypopnea index, ODI: oxyhemoglobin desaturation index, SaO2: oxyhemoglobin saturation, HDL: high lipoprotein.
Table 1.
Descriptive characteristics of the population studies from the PROOF cohort. F: female, M: male, AHI: apnea hypopnea index, ODI: oxyhemoglobin desaturation index, SaO2: oxyhemoglobin saturation, HDL: high lipoprotein.
| Variables | Whole population | OSAS | No OSAS | p |
|---|---|---|---|---|
| Age (y) | 75.8 ± 0.9 | 75.7 ± 0.9 | 75.9 ± 0.9 | 0.621 |
| Sex (F/M) | 19/11 | 6/9 | 13/2 | 0.008 |
| AHI (h-1) | 22.5 ± 21.0 | 41.2 ± 12.6 | 3.8 ± 1.5 | < 0.001 |
| ODI (h-1) | 14.7 ± 15.1 | 28.1 ± 9.3 | 1.4 ± 0.6 | < 0.001 |
| SaO2 min (%) | 88.0 ± 5.2 | 84.3 ± 4.6 | 91.7 ± 2.1 | < 0.001 |
| SaO2moy (%) | 94.4 ± 1.7 | 93.4 ± 2.0 | 95.3 ± 0.7 | < 0.01 |
| %time SaO2 < 90 | 4.8 ± 11.5 | 9.6 ± 15.0 | 0 | < 0.05 |
| Total cholesterol (g.L-1) | 2.3 ± 0.4 | 2.1 ± 0.4 | 2.4 ± .4 | 0.057 |
Table 2.
Summary table of the antibodies used.
| Antibody | Name | Anti | Reference | Supplier |
|---|---|---|---|---|
| Primary | CD81 (B-11) |
Anti-mouse | SC-166029 | Santa Cruz Biotechnology |
| Primary | CD63 (MX-49.129.5) |
Anti-mouse | SC-5275 | Santa Cruz Biotechnology |
| Primary | CD9 (ALB 6) |
Anti-mouse | SC-59140 | Santa Cruz Biotechnology |
| Primary | TAU (Tau-13) |
Anti-mouse | SC-21796 | Santa Cruz Biotechnology |
| Primary | Amyloid beta (B-4), |
Anti-mouse | SC-28365 | Santa Cruz Biotechnology |
| Primary | GAPDH (0411) |
Anti-mouse | SC-47724 | Santa Cruz Biotechnology |
| Primary | ZO-1 | Anti-rabbit | 40-2200 | Thermo Fisher |
| Primary | Claudin-5 | Anti-mouse | SC-374221 | Santa Cruz Biotechnology |
| Primary | Occludin | Anti-rabbit | 711500 | Life tech |
| Secondary | m-IgGκBP-HRP | Anti-mouse | SC-516102 | Santa Cruz Biotechnology |
| Secondary | Goat anti-Rabbit IgG, HRP conjugate | Anti-rabbit | 12-348 | Millipore |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



