
Submitted:
19 September 2024
Posted:
20 September 2024
You are already at the latest version
Abstract
Neurodegenerative diseases (NDDs) are a class of cognitive and motor disorders that include Alzheimer’s disease (AD), Parkinson’s disease (PD), Huntington's disease (HD), amyotrophic lateral sclerosis (ALS), and others. They are caused by lesions in cells and tissues of the central nervous system, resulting in corresponding dysfunctions and consequent decline in cognitive and motor functions. Neural tissues are extremely vulnerable to oxidative stress, which plays critical biological roles in NDDs. Aromatic compounds are found extensively in natural plants and have substantial effects of anti-oxidative stress damage, which not only have a wide range of research applications in cosmetics, foods, etc., but are also frequently utilized in the treatment of various central nervous system diseases. This review summarizes the relevant oxidative stress mechanisms in NDDs (AD, PD, HD, and ALS) and reviews aromatic compounds such as polyphenols, terpenoids, and flavonoids that can be used in the management of neurodegenerative diseases, as well as their specific mechanisms of antioxidant action. This review will serve as a reference for future experimental studies on neurodegenerative illnesses while also offering fresh insights into clinical therapy.
Keywords:
Neurodegenerative diseases
; Oxidative stress
; Antioxidants
; Aromatic plants extracts
1. Introduction
Neurodegenerative diseases (NDDs) are a class of diseases characterized by the gradual loss of nerve cells or a decline in their function with age. NDDs mainly include Alzheimer's disease (AD), Parkinson's disease (PD), Huntington's disease (HD), amyotrophic lateral sclerosis (ALS)[1,2]. With the aging population, NDDs not only reduce the quality of life of patients but also impose a huge burden on families and society. Due to the intricate pathologic causes of NDDs, their unclear molecular mechanisms, and their lack of therapeutic specificity, the search for effective preventive and therapeutic approaches is crucial for exploring the mechanisms of NDDs and potential treatment targets.
Oxidative stress is caused by an imbalance between reactive oxygen species (ROS) and reactive nitrogen species (RNS). Although ROS and RNS are important components of normal cellular function and the host’s defense against invading bacteria, their excesses can lead to the onset and progression of various pathological processes[3,4]. Due to high oxygen consumption, high glucose turnover rate, and elevated levels of redox-active iron in certain regions, along with relatively low levels of antioxidant enzymes, such as glutathione(GSH), glutathione peroxidase (GPx), catalase (CAT), and superoxide dismutase (SOD)[5], the brain is particularly susceptible to oxidative damage. Age-related bioenergetic damage, alterations in redox equilibrium, and a progressive decrease in endogenous antioxidant system efficiency all affect the brain parenchyma of the aged[6,7]; therefore, the elderly are more susceptible to oxidative stress. Moreover, as illustrated in Figure 1, oxidative stress is a major risk factor associated with aging and the onset and progression of age-related NDDs.
Antioxidants directly scavenge oxidizing free radicals and regenerate oxidized biomolecules in biological systems. In the biological antioxidant defense system, endogenous and exogenous antioxidants are classified as enzymatic and non-enzymatic antioxidants[8,9]. These two major antioxidant systems play an important role in maintaining the balance between pro-oxidants and antioxidants in the brain and reduce oxidative stress [10] through two main mechanisms. First, the antioxidant defense system is activated to counteract oxidative damage, prevent the production of ROS, and also block and trap free radicals that have already been produced[11]. The first line of defense has been identified as enzymatic antioxidant systems, including SOD, CAT, and GPx. The second line of defense comprise reduced thiols and non-enzymatic antioxidants, including water-soluble, fat-soluble, or metabolic compounds[8]. Secondly, damaged biomolecules are removed during antioxidant repair prior to aggregation, resulting in altered cellular metabolism. Interventions in repair systems include repair of oxidatively damaged nucleic acids by specific enzymes, removal of oxidized proteins by protein hydrolysis systems, and repair of oxidized lipids by phospholipases, peroxidases, or acryloyl transferases[10,11,12].
Aromatherapy is a natural form of treatment that employs natural plant spices or aromatic essential oils as a medium to administer bioactive substances through respiration, the skin or blood circulation using massage, inhalation, fumigation, and compression, to help harmonize the body's homeostasis and to achieve the effect of preventive health care and treatment of diseases[13]. Aromatic plants are considered the perfect source of natural antioxidants, such as polyphenols commonly found in plants; moreover, herbs are antioxidants with redox properties[14]. In addition to their antibacterial, antiviral, and antioxidant bioactivities, aromatic plant extracts can generate specific physiological responses for the prevention and treatment of a wide range of diseases, including cardiovascular disease, diabetes, Alzheimer's disease, and cancer[15,16], and are highly effective in the treatment of medical conditions that involve emotional and cognitive information processing as well as autonomic nervous system activity[17,18]. Therefore, this review focuses on the role of oxidative stress in the pathogenesis of AD, PD, HD, and ALS, and the antioxidant effects of aromatic plant extracts used in the treatment of these diseases.
2. Role of Oxidative Stress in Neurodegenerative Diseases
2.1. Alzheimer’s Disease (AD)
The main hypotheses of AD are the amyloid cascade hypothesis, the tau protein hypothesis, the inflammation hypothesis, the metal ion hypothesis, and the oxidative stress hypothesis. Amyloid-β (Aβ), the most important pathological feature in AD[19,20], is derived from the amyloidogenic pathway of Amyloid-β precursor protein (APP), which undergoes sequential enzymatic cleavage by β-secretase and γ-secretase to produce the neurotoxic Aβ1-40/ Aβ1-42[21]. In the pathogenesis of AD, there is a complex interaction between Aβ and oxidative stress that exacerbates the pathologic changes in AD[22]. On the one hand, Aβ generation directly induces excessive ROS production in neurons, leading to further oxidative stress. Aβ deposition causes a relative excess of Ca2+ in cytoplasmic lysate by reducing Ca2+ storage in the endoplasmic reticulum[23]. A reduction in GSH levels resulting from excess Ca2+ within the cytoplasm leads to an accumulation of ROS within neurons[24]. It has also been demonstrated that Aβ1-42 directly induces the generation of ROS in neurons through the activation of NADPH oxidase (NOX)[25]. On the other hand, oxidative stress can exacerbate the production of Aβ. Oxidative stress has been demonstrated to promote Aβ production by activating the JNK/c-Jun pathway and increasing the transcription of β-site amyloid precursor protein cleaving enzyme-1 (BACE1) and γ-secretase levels[26]. In AD, oxidative stress plays a pivotal role in cytopathic alterations and has a complex relationship with Aβ.
Another major pathological feature in AD is the formation of neuronal fiber tangles (NFTs), formed by hyperphosphorylated Tau protein (pTau). Studies have shown that oxidative stress can induce Tau protein phosphorylation in neuronal cells[27]. In pathological conditions, the normal function of Tau protein is disrupted by ROS, which then interacts with cyclin-dependent kinase 5 (CDK5) and glycogen synthase kinase-3β (GSK-3β), leading to their hyperphosphorylation[28]. Similar to Aβ, the process by which free pTau proteins first form oligomers and then aggregate to form NTFs leads to significant oxidative stress. Hyperphosphorylation of Tau proteins, microtubule disorganization, and accumulation of Tau proteins promote ROS generation[29].
In addition, oxidative stress has also been strongly associated with AD through its effects on neuroinflammation, mitochondrial function, and metal ions. In pathological conditions, the overproduction of ROS activates microglia, which in turn releases a range of inflammatory factors and leads to the activation of immune cells, resulting in a neuroinflammatory response[30]. Studies have shown that metal ion homeostasis is imbalanced in the brains of patients with AD, including the homeostasis of Fe, Cu, and Zn, which are closely related to Aβ and oxidative stress. Free Cu and Fe can form ligand complexes with Aβ, which are efficient catalysts for ROS generation[31]. A literature survey related to each hypothesis of AD reveals that each pathway appears independent but is implicitly linked to oxidative stress, meaning that oxidative stress is a key bridge that connects the various pathways.
2.2. Parkinson’s Disease (PD)
The main pathological hallmarks of PD are degeneration of dopaminergic neurons in the substantia nigra compacta, decreased striatal dopamine content, and intracellular aggregation of Lewy bodies[32,33,34]. Loss of dopaminergic neurons in the brains of patients with hereditary and sporadic Parkinson's disease may be associated with the causative factors of oxidative stress[35,36], mitochondrial dysfunction[37], protein aggregation[38], aberrant apoptotic signaling pathways[39], down-regulation of neurotrophic factors[40], and neuroinflammation[41,42].
Dopaminergic neurons in the dense portion of the substantia nigra require a significant amount of energy to transmit dopamine signals, and their high oxygen consumption and relatively low antioxidant defense capacity result in nigrostriatal neuronal cells being more susceptible to oxidative damage compared to other neuronal cells[43]. The disruption of redox homeostasis and the subsequent increase in oxygen free radicals may lead to mutations in PD-causing genes and abnormalities in the proteins they encode, thus contributing to the development of PD[44]. A decrease in GSH levels and an increase in lipid peroxidase (LPO) levels in the substantia nigra of the brain have been observed in PD mice following the administration of 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP)[45]. Decreased superoxide dismutase 2 (SOD2) and GSH levels and increased lipid peroxide malondialdehyde (MDA) levels have also been observed in the rotenone-induced PD rat model[46]. SOD2 and GSH can scavenge ROS, and their reduced levels indicate low antioxidant function in animal models of PD. The highly oxidized DNA and proteins in the substantia nigra of patients with PD who have high levels of lipid peroxidation and low levels of SOD2 and GSH, indicate a low level of functioning of the antioxidant defense system; hence, oxidative stress damage is pervasive in patients with PD[47,48,49] and suggests that oxidative stress may be a core component of the pathogenesis of PD.
Additionally, PD is associated with an increase in nigral iron content. This may result in oxidative stress damage to iron-sulfur cluster-containing proteins[50], possibly leading to cell death via reactive oxygen species-induced oxidative stress[51]. The increase in iron content may lead to an increase in melanin synthesized by dopamine in conjunction with iron and copper ions, transforming the original antioxidant properties of melanin into a pro-oxidant effect when the metal content is elevated[52]. Furthermore, in the context of elevated iron levels and oxidative stress, α-synuclein, which is a hallmark of PD, will undergo increased aggregation and deposition, thereby accelerating the progression of the disease[53,54].
Numerous preclinical and clinical studies have shown that PD is closely related to oxidative stress, which is involved in the inflammatory response of brain tissues, mitochondrial damage, iron-induced death, and other pathogenic mechanisms related to PD. Oxidative stress acts independently and simultaneously interacts with other pathogenic mechanisms to cause the loss of DA neurons in the substantia nigra, which obviously plays a key role in the development of PD.
2.3. Huntington's Disease (HD)
HD is an autosomal dominant, fully penetrant neurodegenerative disorder characterized by involuntary chorea-like movements, bradykinesia, dystonia, ankylosis, and dementia[55]. It is caused by an inherited CAG (polyglutamine) repeat amplification of the Huntington (HTT) gene on chromosome 4p16.3 that results in the production of a mutant Huntington (mHTT) protein[56]. The mutation leads to misfolding/aggregation of Huntington's protein with neurotoxic consequences.
A substantial body of evidence indicates that oxidative stress plays a pivotal role in the pathogenesis of HD[57,58]. It has been demonstrated that oxidative stress markers, including 8-hydroxy-2′-deoxyguanosine (8OHDG)[59], protein carbonylation products[57], 3-nitrotyrosine[60] are increased in patients with HD. Mutant Huntington proteins have been observed to mediate uncontrolled DNA damage by impairing the cellular DNA repair machinery, specifically by blocking the action of the DNA repair protein Ku70[61]. Furthermore, mutant Huntington proteins increase ROS levels in neurons[62], and the combination of uncontrolled DNA damage and oxidative stress further contributes to neuronal loss in HD. mHTT has also been shown to induce cytotoxicity through its action via the kynurenine pathway with concomitant production of quinolinic acid and ROS, both of which propagate oxidative stress, mitochondrial dysfunction, and neuronal death[63]. In a clinical study of 16 patients with HD and 36 age- and sex-matched controls, it was found that plasma malondialdehyde levels were elevated in patients with HD and 8-hydroxy-2-deoxyguanosine was elevated in their leukocytes as well. Furthermore, the activities of antioxidant enzymes, including Cu/Zn, SOD, and GSH-Px, were significantly decreased in the erythrocytes of these patients. Moreover, the levels of MDA in the plasma of these patients were associated with the severity of HD[64].
Mitochondrial dysfunction and abnormal energy metabolism caused by HD mutant HTT gene is a common feature of NDDs, of which oxidative stress is a key component. Mutant HTT genes can directly increase mitochondrial ROS and cause persistent DNA damage; furthermore, excessive ROS accumulation can exacerbate HTT-induced neuronal dysfunction. In addition to the obvious mutant proteins, oxidative stress may also damage various types of neuronal cells in the striatum, either alone or in conjunction with the mutant genes. Therefore, there is an ongoing search for appropriate antioxidant therapies that have neuroprotective effects and can thus inhibit the pathogenic role of ROS in patients with HD.
2.4. Amyotrophic Lateral Sclerosis (ALS)
ALS is a rapidly progressive neurological disorder that primarily affects the cerebral cortex, brainstem, and spinal cord motor neurons (MN), with progressive muscular atrophy, myelopathy, and conotruncal fasciculations as its main clinical manifestations[65]. The mechanism of MN apoptosis in ALS is still unclear. However, it involves several factors, including oxidative stress, mitochondrial dysfunction, axonal injury, microglia activation, inflammation, doping toxicity, and anomalous protein aggregation[66]. In the cerebrospinal fluid, cerebral cortex, and peripheral blood of patients with ALS, the levels of 8-hydroxyguanosine and MDA were significantly elevated, and the level of GSH was reduced, suggesting that oxidative stress may play an important role in the pathogenesis of ALS[67,68]. The first gene known to be associated with ALS is SOD1. Approximately 20% of familial ALS (FALS) and 4% of sporadic ALS (SALS) are associated with mutations in the SOD1 gene. In these patients, the unfolded mutant SOD1 protein forms toxic aggregates in the cytoplasm[69]. ALS was initially recognized as an oxidative stress disease because SOD1 converts superoxide to hydrogen peroxide (H2O2) [70]. Xiao et al. found that transgenic mice transfected with the human mutant SOD1 gene had clinical manifestations and pathological changes similar to those of patients with ALS and had severe motor impairment at approximately 120 days of age, and reached the end stage of the disease at approximately 160 days of age[71], which showed that the transient overexpression of SOD1 G93A directly promoted mitochondrial ROS production and could lead to mitochondrial dysfunction with intact motor neuron axons. This suggests that mitochondrial dysfunction and oxidative stress form a vicious circle that promotes neurodegeneration and muscle atrophy in ALS.
Metal ion-mediated oxidative stress has received increasing attention[72]. For instance, hydroxyl radicals produced by the Fenton reaction involving the reduction of iron and copper ions with hydrogen peroxide can trigger lipid peroxidation[73]. Whereas SOD1 proteotoxicity is caused by misfolding of the copper/zinc active site of the antioxidant enzyme, misfolding of copper, zinc, or SOD1 may affect ROS levels, as well as oxidative stress within motor neurons[73]. In a previous study involving SOD1G93A transgenic mice, copper and zinc concentrations and metallothionein expression were measured, and elevated levels of copper ions and lipid peroxidation products, as well as increased expression of metallothionein I/II were detected in spinal cord tissues when the transgenic mice did not show motor dysfunction (at 8 weeks of age). They were further elevated when motor dysfunction was evident (at 16 weeks of age)[74,75]. Iron plays a crucial role in energy, blood, and oxidative stress metabolism, and elevated iron levels indicate higher levels of oxidative stress in an organism. In addition, elevated iron levels interfere with the body's calcium, copper, and zinc levels. Increased levels of iron ions were detected in the nuclei and cytoplasm of spinal motoneurons in patients with ALS[76]. Moreover, the levels of Zn, Fe, Na, Mg, and K were impaired in the blood of SOD1 G93A rats compared to those of SOD1WT rats, further highlighting the roles of impaired oxidative stress metabolism, neuronal homeostasis, and mineral metabolism[77].
Oxidative stress leads to excessive accumulation of ROS and the deposition of abnormal proteins and toxic metabolites in the brain, leading to mitochondrial damage and directly or indirectly triggering the apoptotic pathway of motor neurons, which is involved in and contributes to the pathogenesis of ALS. Although the mechanism of the interaction between oxidative stress and motor neuron damage in ALS is still unclear, future in-depth research would aid in clearly identifying the specific mechanism and therapeutic targets of oxidative stress in ALS, leading to the development of effective therapeutic solutions.
Oxidative stress has a vital role in the development of NDDs. Currently, there are very limited methods for the treatment of NDDs; therefore, the rational application of natural antioxidants in preventing and treating NDDs to alleviate the burden of oxidative stress on the nervous system may provide new ideas for neuroscience research.
3. Antioxidant Effects of Aromatic Plant Extracts in Neurodegenerative Diseases
We have established the important role that oxidative stress plays in the pathophysiology of NDDs. Therefore, antioxidant substances can be considered beneficial in preventing and treating these diseases. Whereas aromatic plants are considered to be a perfect source of natural antioxidants, aromatic plant extracts, such as phenols, terpenoids, ketones, alcohols, etc., have been shown to reduce oxidative stress markers when administered to patients with different diseases and animal models[14].
3.1. Polyphenol Antioxidants
Polyphenols are widely found in plants and are considered the largest group of plant secondary metabolites, with a wide variety of structures ranging from simple hydroxyl groups attached to aromatic rings to highly complex polymeric compounds in tannins and lignin[78].
Curcumin is a polyphenolic compound derived from turmeric. Its chemical structure is characterized by two methoxylated phenols linked via α,β-unsaturated carbonyl groups[79], which have been shown to exert anti-inflammatory, antioxidant, antitumor, and immunomodulatory effects in a variety of diseases[80,81,82]. Several studies have suggested that curcumin may have some therapeutic potential in NDDs[83,84,85]. curcumin can neutralize reactive oxygen species, such as H2O2, nitrite, and superoxide anion, to reduce oxidative damage in the brain, and can also activate the nuclear factor E2-related factor 2 (Nrf2) pathway to enhance the expression of antioxidant response elements (AREs) to counteract oxidative stress at the cellular level[86]. In vivo studies have shown that curcumin can increase SOD and GSH levels, inhibit α-synuclein aggregation, as well as elevate dopamine, serotonin, and norepinephrine levels, and decrease acetylcholinesterase, C-reactive protein (CRP), IL-6, angiotensin II (Ang II), and caspase 3 levels in the brain of PD mice, further reducing inflammation[87]. Additionally, curcumin has been shown to protect against oxidative stress-induced neurodegeneration in the brain tissues of rats with 6-hydroxyanisole (6-OHDA)-induced PD and to increase the viability and survival of primary neurons in rats[88]. Similar results have been found in AD models, and in the Aβ-induced SHSY5Y cell model, where curcumin intervention restored the normalization of SOD and hydrogen peroxide expression and activity, implying that curcumin can alleviate cellular Aβ-induced oxidative stress[89]. Curcumin may play crucial roles as an antioxidant and an anti-inflammatory agent in the treatment of NDDs.
Chlorogenic acid, a natural phenolic acid compound derived mainly from plants, such as eucommia and honeysuckle, has antibacterial, anti-inflammatory, antioxidant, and anti-tumor effects[90,91]. It has been found that chlorogenic acid can chelate free metal ions in animals and reduce the free radicals generated in the body through the Fenton reaction[92]. Additionally, chlorogenic acid activates the Nrf2/ARE signaling pathway and promotes the expression of antioxidant enzymes, such as heme oxygenase-1 (HO-1), NADPH quinone oxidoreductase-1 (NQO-1), GPx, SOD, and γ-glutamyl cysteine synthetase (γ-GCS). Furthermore, it facilitates the expression of other antioxidant enzymes that inhibit intracellular ROS production and ameliorate cellular oxidative damage[93]. In a mouse model of PD induced by MPTP, it was found that the activity of SOD can be increased and mitochondrial membrane potential can be restored in brain tissues following chlorogenic acid intervention[94]. Similar results were found in AD-related studies, where chlorogenic acid combined with aerobic exercise reversed elevated levels of ROS, H2O2, and MDA in the brain tissues of APP/PS1 mice while increasing the levels of CAT, SOD, and GPx to restore the antioxidant capacity of AD mice and to maintain oxidative/antioxidant balance in vivo[95].
Eugenol, also known as 4-allyl guaiacol, is a weakly acidic phenolic compound mainly derived from the dried flower buds of cloves in the myrtle family[96,97]. The antioxidant potential of eugenol is mainly based on free radical scavenging and reduced GSH, which reduces the formation of ROS and thus exerts a neuroprotective effect in NDDs[98,99]. It has been shown that eugenol combined with acupuncture treatment can reduce MDA content and increase SOD and GPx activities in the hippocampus, which in turn improves the learning and memory abilities of AD rats, and this effect was found to be superior to that of acupuncture treatment alone[100].
Ellagic acid, a natural polyphenol compound with antioxidant and anti-tumor effects, has four phenolic hydroxyl structures, which are the basis of its reaction with various ROS. The phenolic hydroxyl group in the structure can provide H atoms and oxygen radicals in vivo, which react with oxygen radicals to reduce the content of oxygen radicals in the body[101]; furthermore, it has been shown to affect a variety of signaling pathways that enhance the antioxidant capacity of the body[102,103]. In the quinolinic acid-induced HD rat model, ellagic acid restored the expression of endogenous antioxidant enzymes in brain tissues, inhibited lipid peroxidation, and scavenged free radicals[104].
Honokiol, a polyphenolic bioactive substance present in magnoliaceae such as thujaplicin, magnolia grandiflora, and magnolia officinalis, can increase antioxidant enzyme activity and decrease oxidase activity by scavenging free radicals modulating signaling pathways such as Nrf2, NF-κB, AMPK. It has been found that honokiol can increase antioxidant enzymes and decrease oxidative enzymes by scavenging free radicals, regulating signaling pathways such as Nrf2, NF-κB, AMPK, etc., and thus protect the body or cells from oxidative stress[105,106,107]. It was demonstrated that honokiol could alleviate oxidative stress by enhancing GSH synthesis and activating the NRF2-ARE pathway, which not only improved motor function and prolonged the lifespan of SOD1-G93A transgenic ALS mice but also increased the viability of NSC- 34 motor neuron-like cell expressing mutant G93A SOD1 proteins in vitro[108]. The same effect can also be observed in vitro and in vivo models of AD and PD.
Polyphenolic compounds are now widely used in different neurological disorders and can play an important role in the treatment of these diseases; therefore, better utilization of the antioxidant effects of polyphenolic compounds in aromatic plants, combined with aromatherapy, may provide new ideas for combating NDDs.

Table 1.
Antioxidant effects of polyphenolic compounds in neurodegenerative disorders.
| Compounds | Types of study | Cell line(s)/animal model(s) | Type of Disease | Mechanism of action/metabolic effects | References |
|---|---|---|---|---|---|
| Curcumin | In vitro | Aβ-induced SH-SY5Y cells | AD | Restore mitochondrial membrane potential; Reduce the expression of mitochondrial apoptotic proteins such as cytochrome c, caspase-3 and Bax; Restore normalized activity and expression of SOD1, SOD2 and catalase; Reduce the expression of total GSK-3β and phospho-Ser9-GSK-3β |
[89] |
| In vivo | Rotenone-induced swiss albino mice | PD | Reduce alpha-synaptic nucleoprotein aggregation; Elevate levels of DA and NA in brain tissue; Reduce MDA levels and increasing SOD, GSH activity |
[87] | |
| In vivo | 6-OHDA-induced SD rat | PD | Elevate dopamine levels; Increase SOD and GSH activity |
[109] | |
| In vitro | Aβ1-42 induction in primary cortical neurons of Wistar albino rats | AD | Decrease MDA level, inhibit lipid peroxidation; Elevate GSH, CAT levels and increase SOD activity; Increase neurotrophic factor levels |
[110] | |
| In vivo | 6-OHDA-induced SD rats | PD | Reduce MDA content, inhibit lipid peroxidation; Elevate SOD and GPx levels and increases Ach activity. |
[111] | |
| In vivo+ In vitro | 6-OHDA-induced SD rats+ SD rat primary neurons | PD | Increase SOD and GPx levels and decreased MDA levels; Increase Wnt3a, β-catenin protein and mRNA expression and c-myc and cyclinD1 mRNA expression |
[88] | |
| In vitro | AβO-induced SH-SY5Y cells | AD | Reduce ROS generation and attenuates oxidative stress Inhibit Tau hyperphosphorylation; |
[112] | |
| Chlorogenic Acid | In vivo | MPTP-induced Swiss albino mice | PD | Increase mtGSH Mn-SOD levels inhibited the activation of pro-apoptotic proteins (including Bax and caspase-3) while increasing the expression of anti-apoptotic proteins (e.g. Bcl-2) | [94] |
| In vitro | Aβ-induced primary hippocampal neurons of SD rats | AD | Elevate SOD and GPx levels | [113] | |
| In vivo | APP/PS1 transgenic mice | AD | Reduce the expression of IL-1β, IL-6 and TNF-α; Reduce the levels of MDA and H2O2, elevate the levels of SOD, CAT and GPx, and inhibit ROS generation; Reduce Aβ deposition and attenuate neuronal damage; Activate the SIRT1/PGC-1α signaling pathway |
[95] | |
| In vivo+ In vitro | Rotenone-induced C57BL/6 mice+ GLUTag cells | PD | Reduce MDA content in the striatum and cortex, elevate GSH levels, and attenuate oxidative damage; Restore the expression of colonic GPR-40 and GPR-43; Up-regulate the expression of GLP-1 receptor in colon, striatum and cortex; Reduce the accumulation of α-synuclein; Reduce dopaminergic neuron loss |
[114] | |
| Ellagic Acid | In vitro | Aβ25-35-induced PC12 cells | AD | Inhibit ROS production; Reduce calcium production. |
[115] |
| In vivo | AlCl3-induced Wistar rat | AD | Elevate SOD and GSH levels; Restore the normal structure of neurons; Down-regulate APP and caspase expression |
[116] | |
| In vivo | QA-induced Wistar rat | HD | Attenuate AchE activity; Increase the level of CAT in the cortex, restore the level of SOD in the cerebral cortex and the level of GSH in the striatum; Decrease the levels of IL-6, TNF-α; Decrease the level of caspase-3 |
[104] | |
| Honokiol | In vivo+ In vitro | hSOD1-G93A transgenic mice + NSC-34 cells transfected with SOD1 G93A | ALS | Activate NRF2-ARE pathway, increases GSH, CAT, GSR activities, and decreases MDA content; Restore mitochondrial function and morphology |
[108] |
| In vivo+ In vitro | Aβ1-42-induced C57BL/6 mice + AβO-induced primary hippocampal neurons of SD rats | AD | Decrease the production of Bax and caspase-9, increases the expression of Bcl2, and reduce neuronal apoptosis; Inhibit ROS generation, attenuate oxidative stress |
[117] |
Abbreviations: GSK-3β: glycogen synthase kinase-3β; 6-OHDA: 6-hydroxyanisole; MPTP: 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine; DA: dopamine; NA: norepinephrine; IL-1β: interleukin-1β; IL-6: interleukin-6; TNF-α: tumor necrosis factor-α; GPR: G Protein-Coupled Receptors; Bcl2: B-cell lymphoma-2.
3.2. Terpenoid Antioxidants
Terpenoids are a class of compounds with the general formula (C5H8)n as well as their oxygenated and variously saturated derivatives. Widespread in nature, higher plants, fungi, microorganisms, insects, and marine organisms, all have terpene constituents. Terpenoids are an important component of aromatic plant extracts, with antioxidant[118], anti-inflammatory[119,120], and antiviral effects[121], and it has been found that terpenoids also play an important role in the treatment of NDDs[122].
Linalool, an aromatic plant-derived monoterpene, is found primarily in the essential oils of plants such as coriander, basil and peppermint. Linalool is known to possess anticancer, antihyperlipidemic, antimicrobial and neuroprotective properties and has a wide range of biological activities, including antioxidant and anti-inflammatory effects[123,124]. Linalool was found to scavenge mitochondrial ROS and restore mitochondrial membrane potential in a glutamate-induced oxidative stress cell model, demonstrating the potential neuroprotective effects of linalool under oxidative stress conditions[125]. In Aβ-induced Drosophila and SD rat models, linalool increased the survival rate of Drosophila, decreased the level of ROS in the optic discs of Drosophila larvae, and also decreased the content of the lipid peroxidation product, 4-hydroxynonenal (4-HNE) in the brain tissues of AD rats, thus preventing the neurotoxicity of Aβ through antioxidant and anti-inflammatory[126]. In an in vitro model of PD established after MPP+ induction in SH-SY5Y cells, the protein expression of Nrf2 in the nucleus and HO-1 in the cytoplasm was elevated by linalool treatment, whereas the protein expression of gp91phox (also known as NOX2), one of the catalytic subunits of nicotinamide adenine dinucleotide phosphate (NADPH) oxidase, which induces the production of ROS, was downregulated. It is sufficiently demonstrated that linalool can enhance the antioxidant defense of MPP+-treated SH-SY5Y cells[127].
Safranal, a monoterpene aldehyde, is the main volatile component of saffron and the source of its distinctive odor. Several studies have shown that safranal has antioxidant[128], anti-inflammatory[129], cardioprotective[130] and other pharmacological activities. In recent years, safranal has been found to have potential in NDDs associated with oxidative stress, inflammation, and impaired mitochondrial function[131], and is particularly excellent in combating oxidative stress[132]. For instance, safranal reduced MDA, ROS, protein carbonyls, interleukin-1β (IL-1β), IL-6, tumor necrosis factor-α (TNF-α), nuclear factor-κB (NF-κB), and apoptotic biomarkers including cystatin 3 and acetylcholinesterase (AChE) activities, and improved the SOD activity and mitochondrial membrane potential (MMP) levels in the hippocampus of Aβ-induced AD rats, improving the cognitive deficits in rats by ameliorating the effects of apoptosis, oxidative stress, inflammation, and cholinesterase activity[133]. Aromatic plant-derived compounds have been less studied in the treatment of HD. However, one study one study found that safranal prevented the 3-NP-induced elevation of nitrite and MDA levels as well as the reduction of SOD, catalase activity, and GSH in the cortex and striatum of HD rats, thereby improving their motor dysfunction[134]. Thus, safranal could be more intensively investigated as a potential therapeutic agent for HD.
In addition, many terpenoids play an essential role in treating of NDDs. For example, β-caryophyllene is a natural plant oil derived from clove and cinnamon and belongs to the class of bicyclic sesquiterpenes[135]. It was found that β-caryophyllene could effectively restore the activities of SOD and CAT antioxidant enzymes and inhibit lipid peroxidation in rotenone-induced Wistar rats, further exerting an anti-oxidative stress effect[136]. 1,8-Cineole, also known as eucalyptol or caffeol, is a monoterpene ether found in many plants' essential oils. Tan et al. found that 1,8-Cineole was effective in enhancing C. elegans survival under conditions of acute oxidative stress and significantly reduced ROS levels, while exerting anti-AD and antioxidant effects through activation of the SKN-1/Nrf-2 pathway, using the Aβ-induced model of the C. elegans[137]. Limonene is a monoterpene compound with a lemon-like flavor widely in nature, mainly found in the aromatic volatile oils of Citrus spp. and some aquatic plants[138]. Eddin et al. found that limonene significantly reduced MDA levels and increased the activities/concentrations of SOD, CAT, and GSH, as well as reduced the levels of TNF-α, IL-1β, and IL-6, and the levels of iNOS, COX-2, P-NF-κB, and P-IOS in the striatum, in the caviar-induced rat model of PD. Limonene can play multiple roles, including antioxidant, anti-inflammatory, and others, in treating PD[139]. Artemisinin, a compound derived from Artemisia annua, has been used clinically for decades mainly in the treatment of malaria[140], and a large number of studies have now demonstrated that artemisinin and its derivatives have significant neuroprotective effects[141,142]. Lim et al. found that artemisinin increased the levels of Nrf2 DNA-binding activity and its regulated proteins HO-1 and NQO1 in differentiated PC12 cells and mouse brain, attenuated the toxicity of MPP+ to PC12 cells, and inhibited the generation of ROS, suggesting that artemisinin can protect dopaminergic neurons by activating the Nrf2 pathway to generate antioxidant enzymes and thereby ameliorating motor behavioral deficits in mice[143].
Terpenoids are now widely used in the treatment of AD and PD, and they can also be fully utilized in the prevention and treatment of other NDDs to explore their mechanisms of action further and maximize their neuroprotective effects.
Table 2.
Antioxidant effects of terpenoids in neurodegenerative diseases.
| Compounds | Types of study | Cell line(s)/animal model(s) | Type of Disease | Mechanism of action/metabolic effects | References |
|---|---|---|---|---|---|
| Linalool | In vivo | Aβ-induced C57BL/6 mice | AD | Elevate SOD and GPx activities; Decrease malondialdehyde levels; Decrease the expression of caspase-9 and caspase-3 |
[144] |
| In vivo | AD Drosophila model + Aβ-induced SD rats | AD | Reduce the level of ROS in the optic disc of Drosophila larvae; Reduce the level of lipid peroxidation product 4-HNE in brain tissue of AD rats |
[126] | |
| In vivo | AlCl3-induced C57BL/6 mouse | AD | Reduce MDA content and inhibits lipid peroxidation; Restore normal levels of SOD and GPx; Activate the Nrf2/HO-1 signaling pathway to reduce oxidative stress; Upregulate CaMKII protein level, improving synaptic plasticity; Increase the level of BDNF |
[145] | |
| In vivo+ In vitro | MPP+-induced SHSY5Y cells + MPTP-induced C57BL/6 mice | PD | Increase protein expression of nuclear Nrf2 and cytoplasmic HO-1 and down-regulatedgp91phox expression in SHSY5Y cells; Inhibite ROS generation and attenuate oxidative stress; |
[127] | |
| Safranal | In vitro | rotenone-induced dopaminergic neurons | PD | Inhibition of ROS generation in dopaminergic neurons by Nrf2-induced downstream antioxidant enzyme genes including GST, GCLs, NQO1 and HO-1-inducible | [146] |
| In vivo | Aβ1-40-induced Wistar rat | AD | Reduce MDA, ROS, protein carbonyl, IL-1β, IL-6, TNF α, NF-kB, and apoptotic biomarkers including cystatinase 3 and acetylcholinesterase activities in hippocampus, and improve SOD activity and mitochondrial membrane potential levels | [133] | |
| In vivo | 3-NP-induced Wistar rats | HD | Prevent 3-NP-induced increasing in nitrite and MDA levels, as well as decreasing in SOD, catalase activity, and GSH. | [134] | |
| In vivo | QA-induced Wistar rat | HD | Reduction of QA-induced lipid peroxidation and oxidative DNA damage prevents QA-generated reduction of hippocampal thiol redox and antioxidant status. | [147] | |
| β-Caryophyllene | In vivo | Rotenone- induced Wistar rats | PD | Prevent the loss of dopaminergic neurons and striatal nerve fibers; Reduce MDA level bar and prevent GSH depletion; Restore SOD and CAT activity; Reduce IL-1β, IL-6, and TNF-a levels; Reduce COX-2 and iNOS expression. |
[136] |
| In vitro | MPP+-treated SH-SY5Y cells | PD | Inhibit MPP+-induced lactate dehydrogenase release and ROS production and increase intracellular GSH and GPx activity; Decrease caspase-3 and Bax levels and increase Bcl-2 expression; |
[148] | |
| In vivo | MPTP-induced C57BL/6 mice | PD | Enhancement of NQO1 expression and enzyme activity inhibits oxidative stress-induced cell death in MPTP-exposed dopaminergic neurons | [149] | |
| Limonene | In vivo | Rotenone- induced Wistar rats | PD | Decrease dopaminergic neuron loss; Increase levels of BDNF and decreased accumulation of alpha-synuclein; Decrease MDA levels and increase activity/concentration of SOD, catalase and GSH; Reduce levels of TNF-α, IL-1β and IL-6; Reduce expression of iNOS, COX-2, P-NF-κ B and P-I κ B in the striatum; Reduce ROT-induced phosphorylation of MAPK signaling proteins in the striatum |
[139] |
| 1,8-cineole | In vitro | Aβ1-42-induced C.elegans | AD | Significantly reduce ROS levels; Activate the SKN-1/Nrf-2 pathway and upregulates the expression of SKN-1, GCS-1 and GST-4" |
[137] |
| In vitro | Aβ25-35-induced PC12 cells | AD | CIN significantly reduced ROS levels in AB25-35 cells in a dose-dependent relationship | [150] | |
| Arteannuin | In vivo+ In vitro | MPTP-induced C57BL/6 mice + (MPP+)-induced PC12 cells | PD | Increase the level of Nrf2 DNA binding activity and its regulated proteins HO-1 and NQO1 in PC12 cells and mouse brain tissue Attenuate the cytotoxicity of MPP+ and decreased the level of ROS; Reduce mitochondrial membrane potential and cleaved cysteine-3 activity Reduce dopaminergic neuron loss in mice |
[143] |
Abbreviations: CaMKII: Ca2+/Calmodulin-Dependent Protein Kinase II; BDNF: brain-derived neurotrophic factor; GCLs: glutamate-cysteine ligase catalytic subunit; GST: glutathione S-transferase; NQO1: NADPH-quinone oxidoreductase 1; HO-1: heme oxygenase 1; 3-NP: 3-Nitropropionic acid; 4-HNE: 4-hydroxynonenal; Nrf2: nuclear factor E2-related factor 2; QA: quinolinic acid.
3.3. Flavonoid Antioxidants
Flavonoid is a natural compound widely found in various green plants; its parent structure is by two benzene rings, a C3 phase linkage formation, containing flavonoids, flavanones, flavonols, etc[151]. A large body of research literature suggests that flavonoids such as quercetin, naringin and glycyrrhizin can contribute to the reduction of major neuronal lesions present in the brain of patients with NDDs by modulating the processes of oxidative stress, inflammation and apoptosis, providing an adequate neuroprotective effect[152]. It is also recognized that flavonoids can cross the blood-brain barrier, thus forming chelates that exert powerful antioxidant properties against ROS and neuroprotective effects on the brain[153].
Quercetin belongs to an important group of flavonoids, widely found in versatile vegetables and fruits, with good antioxidant stress response, strong inhibition and scavenging of ROS, which can play an important role in a variety of diseases and is also an excellent neuroprotective agent[154]. Studies have shown that oral administration of quercetin can reduce the expression of pro-inflammatory cytokines (IL-6, TNF-α, IL-1β, and COX-2) and prevent neurological damage[155]. In rodent studies, oral administration of quercetin was found to protect against oxidative stress and neurotoxicity induced by various neurotoxicities[156]. In studies of AD, quercetin was found to reduce ROS production, apoptosis, β-site amyloid precursor protein cleavage enzyme 1 expression, and Aβ accumulation in H2O2-induced SHSY5Y cells[157], as well as increasing the expression levels of the antioxidant enzymes SOD1, CAT, and GPx1, and decreases the expression level of iNOS in the brain tissues of AlCl3-induced Wistar rats and reduces ROS generation[158]. Napatr Sriraksa et al. conducted an oral quercetin experiment in Parkinsonian rats and concluded that quercetin reduced oxidative damage, increased neuronal density, and attenuated cognitive deficits by decreasing the level of MDA and AChE, thereby increasing the activities of SOD, CAT, and GPx[159]. Similarly, Sandhir's laboratory found restorations in SOD and CAT activities after treatment with quercetin supplementation in rat models of 3-NP-induced HD[160]. It is also evident that quercetin is now widely used in the management of NDDs.
Naringin also belongs to the bioflavonoids class, which has various pharmacological effects, especially positive effects on a wide range of neurological disorders. A Meta-analytical study showed that naringin inhibits neurological disorders in the brain of rodents induced by a wide range of physical and chemical stimuli, mainly relying on its effect on the antioxidant stress response; it has also been shown to restore oxidative stress markers to varying degrees[161]. In a mouse AD model induced by AlCl3, control treatment with naringenin gavage revealed that naringenin reduced hippocampal MDA and NO levels, increased GSH levels, and decreased cerebellar iNOS expression, and ameliorated lipid peroxidation and oxidative stress due to AlCl3 by virtue of its antioxidant properties[162]. Kumar et al., in Huntington's experimental model of 3-NP, found that the naringenin-treated 3-NP treatment group significantly attenuated lipid peroxidation and nitrite concentration, restored SOD and catalase activity, and improved mitochondrial function, which resulted in improved motor dysfunction in mice[163].
In addition to the two flavonoids mentioned above that have significant antioxidant effects in treating NDDs, other flavonoid compounds are essential in treating NDDs. Hesperidin is one of the most abundant phytoflavonoids in citrus fruits. It possesses various biological properties, including antioxidant and anti-inflammatory properties[164]. Kuppusamy Tamilselvam et al. pretreated a Parkinson's cell model established by rotenone-induced SK-N-SH cells using hesperidin, and the results indicated that hesperidin significantly increased the cellular activities of SOD, CAT, and GPx, and that its favorable antioxidant effects resulted in neuroprotection[165]. Glycyrrhizin, a major constituent of licorice, a common herb, has been reported to have antioxidant activity in a variety of studies, and its potent antioxidant properties can play an important role in the treatment of a wide range of diseases[166]. By applying glycyrrhizic acid gavage treatment to a scopolamine-induced mouse model of cognitive impairment, Ju Yeon Ban et al. found that in the glycyrrhizic acid-treated group there was a significant reversal of scopolamine-induced superoxide dismutase and catalase activities, and had significant neuroprotective effects against neurological deficits[167]. In the HD model established after 3-NP stimulation in Wistar rats, glycyrrhizin significantly ameliorated oxidative stress by restoring GSH, SOD, and Nrf2, while suppressing malondialdehyde levels[168].
Flavonoids are now widely used to treat tumors, cardiovascular diseases, and diseases in other organ systems; furthermore, their anti-inflammatory and antioxidant activities make them perfect for anti-tumor and cardioprotection. Therefore, active research on the mechanisms of action of flavonoids in the management of NDDs may bring new hope for the prevention and treatment of NDDs.
Table 3.
Role of flavonoids in neurodegenerative diseases.
| Compounds | Types of study | Cell line(s)/animal model(s) | Type of Disease | Mechanism of action/metabolic effects | References |
|---|---|---|---|---|---|
| Quercetin | In vitro | H2O2-induced SH-SY5Y cells | AD | Reduce hydrogen peroxide-induced reactive oxygen species production, apoptosis, β-site amyloid precursor protein cleaving enzyme 1 expression and Aβ accumulation in SH-SY5Y cells | [157] |
| In vivo | Streptozotocin-induced Wistar rat | AD | Increase SOD and CAT activities; Elevate total antioxidant levels in hippocampus; Reduce MDA content and inhibit lipid peroxidation |
[169] | |
| In vivo | AlCl3-induced Wistar rats | AD | Decrease AchE activity Increase the expression level of antioxidant enzymes SOD1, CAT and GPx, decrease the expression level of iNOS, and reduce ROS generation; Increase the expression of anti-apoptotic gene Bcl2 and decrease the expression of pro-apoptotic gene BAX |
[158] | |
| In vivo | 6-OHDA-induced Wistar rat | PD | Reduce AchE activity; Decrease MDA content and inhibit lipid peroxidation; Increase the activity of SOD, GPx, CAT |
[159] | |
| In vitro | H2O2-induced PC12 cells | PD | Reduce ROS generation, lower MDA content and inhibit lipid peroxidation; Enhance the activities of CAT, SOD, and GPx; Increase Bcl2, decreased Bax expression, decrease expression of cleaved caspase-3 and p53, and decrease apoptosis |
[170] | |
| In vivo | 3-NP-induced Wistar rat | HD | Restoration of SOD and CAT activity; Restoration of mitochondrial function. |
[160] | |
| Naringin | In vivo | Aβ1-42 and manganese induction in Wistar rats | AD | Attenuate elevation of MDA and nitrite concentrations and restore CAT and GSH levels Restore mitochondrial enzyme complex (I, II and IV) activity and enhance the protective effect of AChE; Attenuate the elevation of TNF-α; Reduce the precipitation of Aβ |
[171] |
| In vivo | AlCl3-induced albino rat | AD | Reduce cerebellar iNOS expression and phosphorylation of Tau protein Decrease cerebellar iNOS expression and Tau protein phosphorylation |
[162] | |
| In vitro | 6-OHDA-induced Parkinsonian zebrafish + 6-OHDA-induced SH-SY5Y cells | PD | Increase GSH, SOD and CAT levels and attenuates oxidative stress; Decrease ROS production; Increase mitochondrial membrane potential; Downregulate the expression levels of lrrk2, polg and caspase9 genes |
[172] | |
| In vivo | Vanadium-induced Wistar rats | PD | Effective improvement of GPx, CAT | [173] | |
| In vivo | MPTP-induced C57BL/6J mice | PD | Increase the activity of glutathione reductase and catalase, reduce the content of LPO, and reverse the toxic effect of MPTP. | [174] | |
| In vivo | 3-NP-induced Wistar rats | HD | Significantly reduced lipid peroxidation, nitrite concentration, restored superoxide dismutase and catalase activity | [163] | |
| Hesperidin | In vivo | AlCl3-induced Albino Wistar rats | AD | Elevate GSH levels, increase SOD, CAT, GPx activities, and reduce oxidative stress; Decrease Bax levels, increase Bcl2 levels, and decrease cellular autophagy |
[175] |
| In vitro | Rotenone-induced SK-N-SH cells | PD | Increase GSH levels and activities of SOD, CAT, and GPx; Inhibite the generation of ROS; Increase intracellular ATP levels; Restore mitochondrial membrane potential; Increase Bcl-2 expression and decreased Bax expression |
[165] | |
| Glycyrrhizic acid | In vivo | 3-NP-induced Wistar albino rats | HD | Restore GSH, SOD, and Nrf2 activity, inhibits malondialdehyde activity Decrease TNF-α, IL-1β, and IL-6 levels and reduce inflammatory response, and Increase BDNF content, improve neuronal damage |
[168] |
| In vivo | Rotenone-induced Wistar rats | PD | Increase antioxidant enzyme activity, inhibit glutathione depletion, inhibit lipid peroxidation, and attenuate dopaminergic neuron loss | [46] |
Abbreviations: AchE: acetylcholinesterase; iNOS: inducible nitric oxide synthase; LPO: Lipid Hydroperoxide.
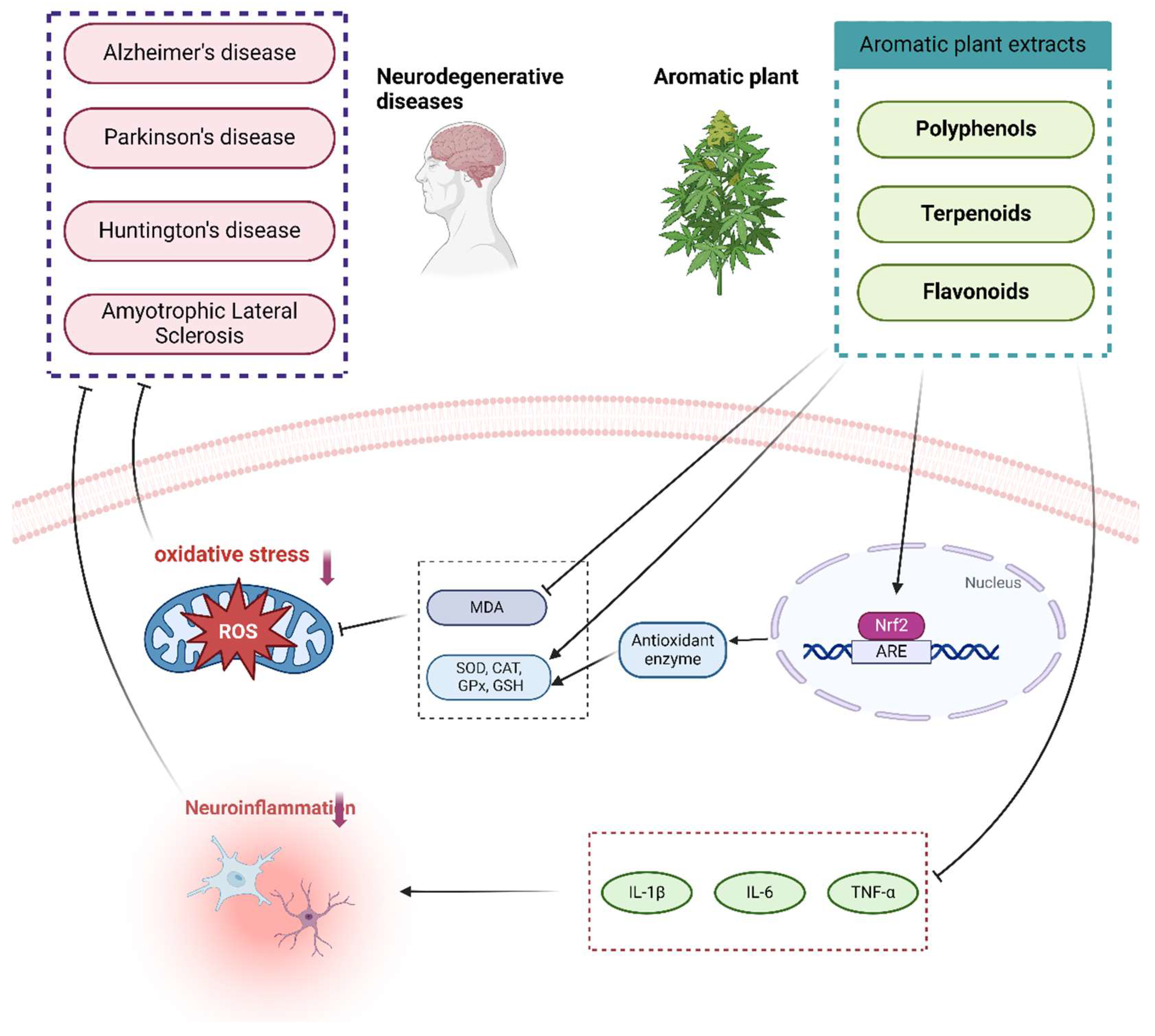
Figure 2.
Role of aromatic plant extracts in neurodegenerative diseases. Polyphenols, terpenoids, and flavonoids play different roles in different NDDs (AD, PD, HD, and ALS). However, in general, they mainly reflect the reduction of MDA content and the increase of SOD, CAT, GPx, and GSH activities, and also play an antioxidant role by affecting the Nrf2 signaling pathway to activate the gene expression of antioxidant enzymes and inhibit the production of ROS. At the same time, it can also inhibit the production of pro-inflammatory factors IL-1β, IL-6, and TNF-α and play an anti-inflammatory role in preventing and treating NDDs.
Figure 2.
Role of aromatic plant extracts in neurodegenerative diseases. Polyphenols, terpenoids, and flavonoids play different roles in different NDDs (AD, PD, HD, and ALS). However, in general, they mainly reflect the reduction of MDA content and the increase of SOD, CAT, GPx, and GSH activities, and also play an antioxidant role by affecting the Nrf2 signaling pathway to activate the gene expression of antioxidant enzymes and inhibit the production of ROS. At the same time, it can also inhibit the production of pro-inflammatory factors IL-1β, IL-6, and TNF-α and play an anti-inflammatory role in preventing and treating NDDs.
4. Conclusions
The central nervous system has a high degree of metabolic activity, making it particularly sensitive to ROS. A previous study showed that there were more than 190 types of neurological diseases related to the oxidative stress response[176]. The treatment of NDDs has even become a global challenge, and common neurological treatments are complex and expensive. In the above comprehensive study, we found that natural antioxidants are a large family of biomolecules with abundant antioxidant and anti-inflammatory effects that offer great opportunities and challenges. Furthermore, aromatic plant extracts, as a kind of natural antioxidant, play important roles in the treatment of neurological diseases. After repeated studies, these compounds have been shown to be potential treatment options for NDDs and need to be vigorously developed[177]. Furthermore, studies have shown that adequate supplementation with antioxidants through special dietary modifications and the intake of aromatic antioxidants from specific foods can also have a therapeutic effect on neurodegenerative disorders and can even have a better effect on preventing harmful activities and enhancing cell regeneration[177,178]. With the application of more antioxidant substances in the management of NDDs, the public perception of multiple antioxidant substances has gradually changed from treatment to prevention and enhancement.
Aromatic compounds are widely found in various natural plants, such as vegetables, fruits, grains, spices, and medicinal plants, and can be extracted and applied through different modalities such as distillation and ultrasonication[179]. This review comprehensively examines aromatic compounds, such as polyphenols, terpenoids, and flavonoids, which play major roles in neurological therapy and are involved in several in vivo and in vitro experiments. Some aromatic compounds have also been found to exert favorable antioxidant effects, inhibit inflammatory reactions, avoid apoptosis of nerve cells, and protect the nerves from neurotoxicity. Among the four types of NDDs studied in this research, aromatic compounds led to clear improvements in oxidative stress and precise neuroprotection; they are also safe and effective, have no obvious toxic side effects, and are suitable for long-term use in the treatment of a variety of NDDs. These compounds also provide an effective treatment option and a novel management philosophy for NDDs; moreover, these compounds have a wide range of application prospects. Therefore, its active application and further research is warranted.
Studies have shown that antioxidants are classified as either endogenous or exogenous[180], and although aromatic compounds are supplemental exogenous antioxidants, they can affect the levels of endogenous antioxidants, such as CAT, SOD, and GPx. However, the existence of any correlation between the effects of these antioxidants and the conditions under which they are affected is unclear and would need to be examined further. Moreover, since the therapeutic effects of these compounds on neurological diseases are based on their ability to cross the blood-brain barrier, the therapeutic effects of these compounds need to be further investigated experimentally to evaluate their effects[180].
Hence, it is imperative to conduct further experimental investigations into the therapeutic effects of these compounds. Such research will help further our understanding of the mechanisms of action of these compounds and determine essential factors such as drug efficacy, dosage, and duration of action.
Author Contributions
Y.Z. and M.T. collected the literature, performed the analysis and interpretation, and wrote the manuscript. Y.Q. and T.Z. contributed to the literature collection and the analysis and interpretation of the results. D.Q. and Z.L. revised the manuscript interpretation of the results and secured funding for the study. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the National Natural Science Foundation of China (82160923, 82374425, 82260929); Applied Basic Research Programs of Science and Technology Commission Foundation of Yunnan Province (202301AS070053); Key Laboratory of Traditional Chinese Medicine for Prevention and Treatment of Neuropsychiatric Diseases, Yunnan Provincial Department of Education; Scientific Research Projects for High-level Talents of Yunnan University of Chinese Medicine (2019YZG01); Young Top-Notch Talent in 10,000 Talent Program of Yunnan Province (YNWR-QNBJ-2019-235); Postdoctoral Fellowship Program of CPSF (GZB20230665); Scientific Research Fund Project of Yunnan Provincial Department of Education (2024Y395).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Xu, J.; Du, W.; Zhao, Y.; Lim, K.; Lu, L.; Zhang, C.; Li, L. Mitochondria targeting drugs for neurodegenerative diseases-Design, mechanism and application. Acta Pharm Sin B 2022, 12, 2778-2789. [CrossRef]
- Golpich, M.; Amini, E.; Mohamed, Z.; Azman Ali, R.; Mohamed Ibrahim, N.; Ahmadiani, A. Mitochondrial Dysfunction and Biogenesis in Neurodegenerative diseases: Pathogenesis and Treatment. CNS Neurosci Ther 2017, 23, 5-22. [CrossRef]
- Yoshikawa, T.; You, F. Oxidative Stress and Bio-Regulation. Int J Mol Sci 2024, 25. [CrossRef]
- Reddy, V.P. Oxidative Stress in Health and Disease. Biomedicines 2023, 11. [CrossRef]
- Cobley, J.N.; Fiorello, M.L.; Bailey, D.M. 13 reasons why the brain is susceptible to oxidative stress. Redox Biol 2018, 15, 490-503. [CrossRef]
- Korovesis, D.; Rubio-Tomás, T.; Tavernarakis, N. Oxidative Stress in Age-Related Neurodegenerative Diseases: An Overview of Recent Tools and Findings. Antioxidants (Basel) 2023, 12. [CrossRef]
- Islam, M.T. Oxidative stress and mitochondrial dysfunction-linked neurodegenerative disorders. Neurol Res 2017, 39, 73-82. [CrossRef]
- Pisoschi, A.M.; Pop, A. The role of antioxidants in the chemistry of oxidative stress: A review. Eur J Med Chem 2015, 97, 55-74. [CrossRef]
- Poljsak, B.; Šuput, D.; Milisav, I. Achieving the balance between ROS and antioxidants: when to use the synthetic antioxidants. Oxid Med Cell Longev 2013, 2013, 956792. [CrossRef]
- Jiang, T.; Sun, Q.; Chen, S. Oxidative stress: A major pathogenesis and potential therapeutic target of antioxidative agents in Parkinson's disease and Alzheimer's disease. Prog Neurobiol 2016, 147, 1-19. [CrossRef]
- Lobo, V.; Patil, A.; Phatak, A.; Chandra, N. Free radicals, antioxidants and functional foods: Impact on human health. Pharmacogn Rev 2010, 4, 118-126. [CrossRef]
- Cadet, J.; Davies, K.J.A. Oxidative DNA damage & repair: An introduction. Free Radic Biol Med 2017, 107, 2-12. [CrossRef]
- Perry, N.; Perry, E. Aromatherapy in the management of psychiatric disorders: clinical and neuropharmacological perspectives. CNS Drugs 2006, 20, 257-280. [CrossRef]
- Proestos, C.; Varzakas, T. Aromatic Plants: Antioxidant Capacity and Polyphenol Characterisation. Foods 2017, 6. [CrossRef]
- Edris, A.E. Pharmaceutical and therapeutic potentials of essential oils and their individual volatile constituents: a review. Phytother Res 2007, 21, 308-323. [CrossRef]
- Ma, Y.; Li, Y.; Yin, R.; Guo, P.; Lei, N.; Li, G.; Xiong, L.; Xie, Y. Therapeutic potential of aromatic plant extracts in Alzheimer's disease: Comprehensive review of their underlying mechanisms. CNS Neurosci Ther 2023, 29, 2045-2059. [CrossRef]
- Villemure, C.; Bushnell, M.C. Mood influences supraspinal pain processing separately from attention. J Neurosci 2009, 29, 705-715. [CrossRef]
- Haze, S.; Sakai, K.; Gozu, Y. Effects of fragrance inhalation on sympathetic activity in normal adults. Jpn J Pharmacol 2002, 90, 247-253. [CrossRef]
- Sun, Z.T.; Ma, C.; Li, G.J.; Zheng, X.Y.; Hao, Y.T.; Yang, Y.; Wang, X. Application of Antibody Fragments Against Aβ With Emphasis on Combined Application With Nanoparticles in Alzheimer's Disease. Front Pharmacol 2021, 12, 654611. [CrossRef]
- Behl, T.; Kaur, I.; Fratila, O.; Brata, R.; Bungau, S. Exploring the Potential of Therapeutic Agents Targeted towards Mitigating the Events Associated with Amyloid-β Cascade in Alzheimer's Disease. Int J Mol Sci 2020, 21. [CrossRef]
- Coronel, R.; Bernabeu-Zornoza, A.; Palmer, C.; Muñiz-Moreno, M.; Zambrano, A.; Cano, E.; Liste, I. Role of Amyloid Precursor Protein (APP) and Its Derivatives in the Biology and Cell Fate Specification of Neural Stem Cells. Mol Neurobiol 2018, 55, 7107-7117. [CrossRef]
- Tamagno, E.; Guglielmotto, M.; Vasciaveo, V.; Tabaton, M. Oxidative Stress and Beta Amyloid in Alzheimer's Disease. Which Comes First: The Chicken or the Egg? Antioxidants (Basel) 2021, 10. [CrossRef]
- Liu, Z.; Zhou, T.; Ziegler, A.C.; Dimitrion, P.; Zuo, L. Oxidative Stress in Neurodegenerative Diseases: From Molecular Mechanisms to Clinical Applications. Oxid Med Cell Longev 2017, 2017, 2525967. [CrossRef]
- Ferreiro, E.; Oliveira, C.R.; Pereira, C.M.F. The release of calcium from the endoplasmic reticulum induced by amyloid-beta and prion peptides activates the mitochondrial apoptotic pathway. Neurobiol Dis 2008, 30, 331-342. [CrossRef]
- Hwang, S.; Kim, J.K. Effects of NADPH Oxidase Inhibitors and Mitochondria-Targeted Antioxidants on Amyloid β(1-42)-Induced Neuronal Deaths in Mouse Mixed Cortical Cultures. Chonnam Med J 2018, 54, 159-166. [CrossRef]
- Tamagno, E.; Guglielmotto, M.; Aragno, M.; Borghi, R.; Autelli, R.; Giliberto, L.; Muraca, G.; Danni, O.; Zhu, X.; Smith, M.A.; et al. Oxidative stress activates a positive feedback between the gamma- and beta-secretase cleavages of the beta-amyloid precursor protein. J Neurochem 2008, 104, 683-695. [CrossRef]
- Ibáñez-Salazar, A.; Bañuelos-Hernández, B.; Rodríguez-Leyva, I.; Chi-Ahumada, E.; Monreal-Escalante, E.; Jiménez-Capdeville, M.E.; Rosales-Mendoza, S. Oxidative Stress Modifies the Levels and Phosphorylation State of Tau Protein in Human Fibroblasts. Front Neurosci 2017, 11, 495. [CrossRef]
- Bai, R.; Guo, J.; Ye, X.Y.; Xie, Y.; Xie, T. Oxidative stress: The core pathogenesis and mechanism of Alzheimer's disease. Ageing Res Rev 2022, 77, 101619. [CrossRef]
- Dias-Santagata, D.; Fulga, T.A.; Duttaroy, A.; Feany, M.B. Oxidative stress mediates tau-induced neurodegeneration in Drosophila. J Clin Invest 2007, 117, 236-245. [CrossRef]
- Leng, F.; Edison, P. Neuroinflammation and microglial activation in Alzheimer disease: where do we go from here? Nat Rev Neurol 2021, 17, 157-172. [CrossRef]
- Faller, P.; Hureau, C. A bioinorganic view of Alzheimer's disease: when misplaced metal ions (re)direct the electrons to the wrong target. Chemistry 2012, 18, 15910-15920. [CrossRef]
- Dawson, T.M.; Dawson, V.L. Molecular pathways of neurodegeneration in Parkinson's disease. Science 2003, 302, 819-822. [CrossRef]
- Ye, H.; Robak, L.A.; Yu, M.; Cykowski, M.; Shulman, J.M. Genetics and Pathogenesis of Parkinson's Syndrome. Annu Rev Pathol 2023, 18, 95-121. [CrossRef]
- Morris, H.R.; Spillantini, M.G.; Sue, C.M.; Williams-Gray, C.H. The pathogenesis of Parkinson's disease. Lancet 2024, 403, 293-304. [CrossRef]
- Robea, M.A.; Balmus, I.M.; Ciobica, A.; Strungaru, S.; Plavan, G.; Gorgan, L.D.; Savuca, A.; Nicoara, M. Parkinson's Disease-Induced Zebrafish Models: Focussing on Oxidative Stress Implications and Sleep Processes. Oxid Med Cell Longev 2020, 2020, 1370837. [CrossRef]
- Imbriani, P.; Martella, G.; Bonsi, P.; Pisani, A. Oxidative stress and synaptic dysfunction in rodent models of Parkinson's disease. Neurobiol Dis 2022, 173, 105851. [CrossRef]
- Ammal Kaidery, N.; Ahuja, M.; Thomas, B. Crosstalk between Nrf2 signaling and mitochondrial function in Parkinson's disease. Mol Cell Neurosci 2019, 101, 103413. [CrossRef]
- Leem, E.; Kim, S.R. Limited therapeutic potential of astrocyte elevated gene-1 transduction in an animal model of Parkinson's disease. Neural Regen Res 2020, 15, 1850-1851. [CrossRef]
- Leem, E.; Kim, H.J.; Choi, M.; Kim, S.; Oh, Y.S.; Lee, K.J.; Choe, Y.S.; Um, J.Y.; Shin, W.H.; Jeong, J.Y.; et al. Upregulation of neuronal astrocyte elevated gene-1 protects nigral dopaminergic neurons in vivo. Cell Death Dis 2018, 9, 449. [CrossRef]
- Nam, J.H.; Leem, E.; Jeon, M.T.; Jeong, K.H.; Park, J.W.; Jung, U.J.; Kholodilov, N.; Burke, R.E.; Jin, B.K.; Kim, S.R. Induction of GDNF and BDNF by hRheb(S16H) transduction of SNpc neurons: neuroprotective mechanisms of hRheb(S16H) in a model of Parkinson's disease. Mol Neurobiol 2015, 51, 487-499. [CrossRef]
- Araújo, B.; Caridade-Silva, R.; Soares-Guedes, C.; Martins-Macedo, J.; Gomes, E.D.; Monteiro, S.; Teixeira, F.G. Neuroinflammation and Parkinson's Disease-From Neurodegeneration to Therapeutic Opportunities. Cells 2022, 11. [CrossRef]
- Isik, S.; Yeman Kiyak, B.; Akbayir, R.; Seyhali, R.; Arpaci, T. Microglia Mediated Neuroinflammation in Parkinson's Disease. Cells 2023, 12. [CrossRef]
- Ren, X.; Zou, L.; Zhang, X.; Branco, V.; Wang, J.; Carvalho, C.; Holmgren, A.; Lu, J. Redox Signaling Mediated by Thioredoxin and Glutathione Systems in the Central Nervous System. Antioxid Redox Signal 2017, 27, 989-1010. [CrossRef]
- Dorszewska, J.; Kowalska, M.; Prendecki, M.; Piekut, T.; Kozłowska, J.; Kozubski, W. Oxidative stress factors in Parkinson's disease. Neural Regen Res 2021, 16, 1383-1391. [CrossRef]
- Krishnamoorthy, A.; Sevanan, M.; Mani, S.; Balu, M.; Balaji, S.; P, R. Chrysin restores MPTP induced neuroinflammation, oxidative stress and neurotrophic factors in an acute Parkinson's disease mouse model. Neurosci Lett 2019, 709, 134382. [CrossRef]
- Ojha, S.; Javed, H.; Azimullah, S.; Abul Khair, S.B.; Haque, M.E. Glycyrrhizic acid Attenuates Neuroinflammation and Oxidative Stress in Rotenone Model of Parkinson's Disease. Neurotox Res 2016, 29, 275-287. [CrossRef]
- Nakabeppu, Y.; Tsuchimoto, D.; Yamaguchi, H.; Sakumi, K. Oxidative damage in nucleic acids and Parkinson's disease. J Neurosci Res 2007, 85, 919-934. [CrossRef]
- Leathem, A.; Ortiz-Cerda, T.; Dennis, J.M.; Witting, P.K. Evidence for Oxidative Pathways in the Pathogenesis of PD: Are Antioxidants Candidate Drugs to Ameliorate Disease Progression? Int J Mol Sci 2022, 23. [CrossRef]
- Al-Zaid, F.S.; Hurley, M.J.; Dexter, D.T.; Gillies, G.E. Neuroprotective role for RORA in Parkinson's disease revealed by analysis of post-mortem brain and a dopaminergic cell line. NPJ Parkinsons Dis 2023, 9, 119. [CrossRef]
- Ayton, S.; Lei, P.; McLean, C.; Bush, A.I.; Finkelstein, D.I. Transferrin protects against Parkinsonian neurotoxicity and is deficient in Parkinson's substantia nigra. Signal Transduct Target Ther 2016, 1, 16015. [CrossRef]
- Sian-Hülsmann, J.; Mandel, S.; Youdim, M.B.; Riederer, P. The relevance of iron in the pathogenesis of Parkinson's disease. J Neurochem 2011, 118, 939-957. [CrossRef]
- Wakamatsu, K.; Fujikawa, K.; Zucca, F.A.; Zecca, L.; Ito, S. The structure of neuromelanin as studied by chemical degradative methods. J Neurochem 2003, 86, 1015-1023. [CrossRef]
- Longhena, F.; Faustini, G.; Spillantini, M.G.; Bellucci, A. Living in Promiscuity: The Multiple Partners of Alpha-Synuclein at the Synapse in Physiology and Pathology. Int J Mol Sci 2019, 20. [CrossRef]
- Mehra, S.; Sahay, S.; Maji, S.K. α-Synuclein misfolding and aggregation: Implications in Parkinson's disease pathogenesis. Biochim Biophys Acta Proteins Proteom 2019, 1867, 890-908. [CrossRef]
- Vonsattel, J.P.G.; Difiglia, M. Huntington Disease. Journal of Neuropathology & Experimental Neurology 1998, 57, 369-384. [CrossRef]
- A novel gene containing a trinucleotide repeat that is expanded and unstable on Huntington's disease chromosomes. The Huntington's Disease Collaborative Research Group. Cell 1993, 72, 971-983. [CrossRef]
- Sorolla, M.A.; Reverter-Branchat, G.; Tamarit, J.; Ferrer, I.; Ros, J.; Cabiscol, E. Proteomic and oxidative stress analysis in human brain samples of Huntington disease. Free Radic Biol Med 2008, 45, 667-678. [CrossRef]
- Stack, E.C.; Matson, W.R.; Ferrante, R.J. Evidence of oxidant damage in Huntington's disease: translational strategies using antioxidants. Ann N Y Acad Sci 2008, 1147, 79-92. [CrossRef]
- Long, J.D.; Matson, W.R.; Juhl, A.R.; Leavitt, B.R.; Paulsen, J.S. 8OHdG as a marker for Huntington disease progression. Neurobiol Dis 2012, 46, 625-634. [CrossRef]
- Bandookwala, M.; Sengupta, P. 3-Nitrotyrosine: a versatile oxidative stress biomarker for major neurodegenerative diseases. Int J Neurosci 2020, 130, 1047-1062. [CrossRef]
- Enokido, Y.; Tamura, T.; Ito, H.; Arumughan, A.; Komuro, A.; Shiwaku, H.; Sone, M.; Foulle, R.; Sawada, H.; Ishiguro, H.; et al. Mutant huntingtin impairs Ku70-mediated DNA repair. J Cell Biol 2010, 189, 425-443. [CrossRef]
- Hands, S.; Sajjad, M.U.; Newton, M.J.; Wyttenbach, A. In vitro and in vivo aggregation of a fragment of huntingtin protein directly causes free radical production. J Biol Chem 2011, 286, 44512-44520. [CrossRef]
- Giorgini, F.; Guidetti, P.; Nguyen, Q.; Bennett, S.C.; Muchowski, P.J. A genomic screen in yeast implicates kynurenine 3-monooxygenase as a therapeutic target for Huntington disease. Nat Genet 2005, 37, 526-531. [CrossRef]
- Chen, C.M.; Wu, Y.R.; Cheng, M.L.; Liu, J.L.; Lee, Y.M.; Lee, P.W.; Soong, B.W.; Chiu, D.T. Increased oxidative damage and mitochondrial abnormalities in the peripheral blood of Huntington's disease patients. Biochem Biophys Res Commun 2007, 359, 335-340. [CrossRef]
- Feldman, E.L.; Goutman, S.A.; Petri, S.; Mazzini, L.; Savelieff, M.G.; Shaw, P.J.; Sobue, G. Amyotrophic lateral sclerosis. Lancet 2022, 400, 1363-1380. [CrossRef]
- Cappello, V.; Vezzoli, E.; Righi, M.; Fossati, M.; Mariotti, R.; Crespi, A.; Patruno, M.; Bentivoglio, M.; Pietrini, G.; Francolini, M. Analysis of neuromuscular junctions and effects of anabolic steroid administration in the SOD1G93A mouse model of ALS. Mol Cell Neurosci 2012, 51, 12-21. [CrossRef]
- Shibata, N.; Nagai, R.; Miyata, S.; Jono, T.; Horiuchi, S.; Hirano, A.; Kato, S.; Sasaki, S.; Asayama, K.; Kobayashi, M. Nonoxidative protein glycation is implicated in familial amyotrophic lateral sclerosis with superoxide dismutase-1 mutation. Acta Neuropathol 2000, 100, 275-284. [CrossRef]
- Ohta, Y.; Nomura, E.; Shang, J.; Feng, T.; Huang, Y.; Liu, X.; Shi, X.; Nakano, Y.; Hishikawa, N.; Sato, K.; et al. Enhanced oxidative stress and the treatment by edaravone in mice model of amyotrophic lateral sclerosis. J Neurosci Res 2019, 97, 607-619. [CrossRef]
- Bruijn, L.I.; Houseweart, M.K.; Kato, S.; Anderson, K.L.; Anderson, S.D.; Ohama, E.; Reaume, A.G.; Scott, R.W.; Cleveland, D.W. Aggregation and motor neuron toxicity of an ALS-linked SOD1 mutant independent from wild-type SOD1. Science 1998, 281, 1851-1854. [CrossRef]
- Barber, S.C.; Shaw, P.J. Oxidative stress in ALS: key role in motor neuron injury and therapeutic target. Free Radic Biol Med 2010, 48, 629-641. [CrossRef]
- Xiao, Y.; Karam, C.; Yi, J.; Zhang, L.; Li, X.; Yoon, D.; Wang, H.; Dhakal, K.; Ramlow, P.; Yu, T.; et al. ROS-related mitochondrial dysfunction in skeletal muscle of an ALS mouse model during the disease progression. Pharmacol Res 2018, 138, 25-36. [CrossRef]
- Sheykhansari, S.; Kozielski, K.; Bill, J.; Sitti, M.; Gemmati, D.; Zamboni, P.; Singh, A.V. Redox metals homeostasis in multiple sclerosis and amyotrophic lateral sclerosis: a review. Cell Death Dis 2018, 9, 348. [CrossRef]
- Carrí, M.T.; Ferri, A.; Cozzolino, M.; Calabrese, L.; Rotilio, G. Neurodegeneration in amyotrophic lateral sclerosis: the role of oxidative stress and altered homeostasis of metals. Brain Res Bull 2003, 61, 365-374. [CrossRef]
- Tokuda, E.; Watanabe, S.; Okawa, E.; Ono, S. Regulation of Intracellular Copper by Induction of Endogenous Metallothioneins Improves the Disease Course in a Mouse Model of Amyotrophic Lateral Sclerosis. Neurotherapeutics 2015, 12, 461-476. [CrossRef]
- Tokuda, E.; Okawa, E.; Watanabe, S.; Ono, S. Overexpression of metallothionein-I, a copper-regulating protein, attenuates intracellular copper dyshomeostasis and extends lifespan in a mouse model of amyotrophic lateral sclerosis caused by mutant superoxide dismutase-1. Hum Mol Genet 2014, 23, 1271-1285. [CrossRef]
- Qin, X.; Wu, P.; Wen, T.; Jia, R.; Zhang, R.; Jin, J.; Hu, F.; Chen, Q.Y.; Dang, J. Comparative assessment of blood Metal/metalloid levels, clinical heterogeneity, and disease severity in amyotrophic lateral sclerosis patients. Neurotoxicology 2022, 89, 12-19. [CrossRef]
- Aydemir, D.; Surucu, S.; Basak, A.N.; Ulusu, N.N. Evaluation of the Hematological and Serum Biochemistry Parameters in the Pre-Symptomatic and Symptomatic Stages of ALS Disease to Support Early Diagnosis and Prognosis. Cells 2022, 11. [CrossRef]
- Moradi, S.Z.; Jalili, F.; Farhadian, N.; Joshi, T.; Wang, M.; Zou, L.; Cao, H.; Farzaei, M.H.; Xiao, J. Polyphenols and neurodegenerative diseases: focus on neuronal regeneration. Crit Rev Food Sci Nutr 2022, 62, 3421-3436. [CrossRef]
- Chainoglou, E.; Hadjipavlou-Litina, D. Curcumin in Health and Diseases: Alzheimer's Disease and Curcumin Analogues, Derivatives, and Hybrids. Int J Mol Sci 2020, 21. [CrossRef]
- Abdollahi, E.; Momtazi, A.A.; Johnston, T.P.; Sahebkar, A. Therapeutic effects of curcumin in inflammatory and immune-mediated diseases: A nature-made jack-of-all-trades? J Cell Physiol 2018, 233, 830-848. [CrossRef]
- Liao, F.; Liu, L.; Luo, E.; Hu, J. Curcumin enhances anti-tumor immune response in tongue squamous cell carcinoma. Arch Oral Biol 2018, 92, 32-37. [CrossRef]
- Kotha, R.R.; Luthria, D.L. Curcumin: Biological, Pharmaceutical, Nutraceutical, and Analytical Aspects. Molecules 2019, 24. [CrossRef]
- Lou, S.; Gong, D.; Yang, M.; Qiu, Q.; Luo, J.; Chen, T. Curcumin Improves Neurogenesis in Alzheimer's Disease Mice via the Upregulation of Wnt/β-Catenin and BDNF. Int J Mol Sci 2024, 25. [CrossRef]
- Gomez-Sequeda, N.; Jimenez-Del-Rio, M.; Velez-Pardo, C. Combination of Tramiprosate, Curcumin, and SP600125 Reduces the Neuropathological Phenotype in Familial Alzheimer Disease PSEN1 I416T Cholinergic-like Neurons. Int J Mol Sci 2024, 25. [CrossRef]
- Tripathi, S.; Bhawana. Epigenetic Orchestration of Neurodegenerative Disorders: A Possible Target for Curcumin as a Therapeutic. Neurochem Res 2024. [CrossRef]
- Menon, V.P.; Sudheer, A.R. Antioxidant and anti-inflammatory properties of curcumin. Adv Exp Med Biol 2007, 595, 105-125. [CrossRef]
- Motawi, T.K.; Sadik, N.A.H.; Hamed, M.A.; Ali, S.A.; Khalil, W.K.B.; Ahmed, Y.R. Potential therapeutic effects of antagonizing adenosine A(2A) receptor, curcumin and niacin in rotenone-induced Parkinson's disease mice model. Mol Cell Biochem 2020, 465, 89-102. [CrossRef]
- Wang, Y.L.; Ju, B.; Zhang, Y.Z.; Yin, H.L.; Liu, Y.J.; Wang, S.S.; Zeng, Z.L.; Yang, X.P.; Wang, H.T.; Li, J.F. Protective Effect of Curcumin Against Oxidative Stress-Induced Injury in Rats with Parkinson's Disease Through the Wnt/ β-Catenin Signaling Pathway. Cell Physiol Biochem 2017, 43, 2226-2241. [CrossRef]
- Huang, H.C.; Xu, K.; Jiang, Z.F. Curcumin-mediated neuroprotection against amyloid-β-induced mitochondrial dysfunction involves the inhibition of GSK-3β. J Alzheimers Dis 2012, 32, 981-996. [CrossRef]
- Tošović, J.; Marković, S.; Dimitrić Marković, J.M.; Mojović, M.; Milenković, D. Antioxidative mechanisms in chlorogenic acid. Food Chem 2017, 237, 390-398. [CrossRef]
- Ye, J.; Gao, Y.; Ji, M.; Yang, Y.; Wang, Z.; Wang, B.; Jin, J.; Li, L.; Wang, H.; Xu, X.; et al. Oral SMEDDS promotes lymphatic transport and mesenteric lymph nodes target of chlorogenic acid for effective T-cell antitumor immunity. J Immunother Cancer 2021, 9. [CrossRef]
- Wang, X.; Fan, X.; Yuan, S.; Jiao, W.; Liu, B.; Cao, J.; Jiang, W. Chlorogenic acid protects against aluminium-induced cytotoxicity through chelation and antioxidant actions in primary hippocampal neuronal cells. Food Funct 2017, 8, 2924-2934. [CrossRef]
- Hussein, R.M.; Sawy, D.M.; Kandeil, M.A.; Farghaly, H.S. Chlorogenic acid, quercetin, coenzyme Q10 and silymarin modulate Keap1-Nrf2/heme oxygenase-1 signaling in thioacetamide-induced acute liver toxicity. Life Sci 2021, 277, 119460. [CrossRef]
- Singh, S.S.; Rai, S.N.; Birla, H.; Zahra, W.; Rathore, A.S.; Dilnashin, H.; Singh, R.; Singh, S.P. Neuroprotective Effect of Chlorogenic Acid on Mitochondrial Dysfunction-Mediated Apoptotic Death of DA Neurons in a Parkinsonian Mouse Model. Oxid Med Cell Longev 2020, 2020, 6571484. [CrossRef]
- Shi, D.; Hao, Z.; Qi, W.; Jiang, F.; Liu, K.; Shi, X. Aerobic exercise combined with chlorogenic acid exerts neuroprotective effects and reverses cognitive decline in Alzheimer's disease model mice (APP/PS1) via the SIRT1/ /PGC-1α/PPARγ signaling pathway. Front Aging Neurosci 2023, 15, 1269952. [CrossRef]
- Anees Ahmed, K.; Ubaid ur, R.; Moazzam Rafiq, K.; Amna, S.; Tariq, M.; Muneeb, K. Essential oil eugenol: sources, extraction techniques and nutraceutical perspectives. RSC Advances 2017. [CrossRef]
- Taleuzzaman, M.; Jain, P.; Verma, R.; Iqbal, Z.; Mirza, M.A. Eugenol as a Potential Drug Candidate: A Review. Curr Top Med Chem 2021, 21, 1804-1815. [CrossRef]
- Pontes, N.H.L.; Reis, T.; Vasconcelos, C.F.M.; Aragão, P.; Souza, R.B.; Catunda Junior, F.E.A.; Aguiar, L.M.V.; Cunha, R. Impact of eugenol on in vivo model of 6-hydroxydopamine-induced oxidative stress. Free Radic Res 2021, 55, 556-568. [CrossRef]
- Goyal, A.; Solanki, A.; Verma, A. Preclinical Evidence-based Review on Therapeutic Potential of Eugenol for the Treatment of Brain Disorders. Curr Mol Med 2023, 23, 390-400. [CrossRef]
- Liu, Z.; Niu, W.; Yang, X.; Wang, Y. Effects of combined acupuncture and eugenol on learning-memory ability and antioxidation system of hippocampus in Alzheimer disease rats via olfactory system stimulation. J Tradit Chin Med 2013, 33, 399-402. [CrossRef]
- Salem, A.M.; Mohammaden, T.F.; Ali, M.A.M.; Mohamed, E.A.; Hasan, H.F. Ellagic and ferulic acids alleviate gamma radiation and aluminium chloride-induced oxidative damage. Life Sci 2016, 160, 2-11. [CrossRef]
- Sanadgol, N.; Golab, F.; Tashakkor, Z.; Taki, N.; Moradi Kouchi, S.; Mostafaie, A.; Mehdizadeh, M.; Abdollahi, M.; Taghizadeh, G.; Sharifzadeh, M. Neuroprotective effects of ellagic acid on cuprizone-induced acute demyelination through limitation of microgliosis, adjustment of CXCL12/IL-17/IL-11 axis and restriction of mature oligodendrocytes apoptosis. Pharm Biol 2017, 55, 1679-1687. [CrossRef]
- Lin, W.; Liu, G.; Kang, X.; Guo, P.; Shang, Y.; Du, R.; Wang, X.; Chen, L.; Yue, R.; Kong, F.; et al. Ellagic acid inhibits high glucose-induced injury in rat mesangial cells via the PI3K/Akt/FOXO3a signaling pathway. Exp Ther Med 2021, 22, 1017. [CrossRef]
- Bains, M.; Kaur, J.; Akhtar, A.; Kuhad, A.; Sah, S.P. Anti-inflammatory effects of ellagic acid and vanillic acid against quinolinic acid-induced rat model of Huntington's disease by targeting IKK-NF-κB pathway. Eur J Pharmacol 2022, 934, 175316. [CrossRef]
- Guo, S.; Xu, J.J.; Wei, N.; Han, J.Y.; Xue, R.; Xu, P.S.; Gao, C.Y. Honokiol Attenuates the Memory Impairments, Oxidative Stress, Neuroinflammation, and GSK-3β Activation in Vascular Dementia Rats. J Alzheimers Dis 2019, 71, 97-108. [CrossRef]
- Li, C.G.; Ni, C.L.; Yang, M.; Tang, Y.Z.; Li, Z.; Zhu, Y.J.; Jiang, Z.H.; Sun, B.; Li, C.J. Honokiol protects pancreatic β cell against high glucose and intermittent hypoxia-induced injury by activating Nrf2/ARE pathway in vitro and in vivo. Biomed Pharmacother 2018, 97, 1229-1237. [CrossRef]
- Xia, S.; Lin, H.; Liu, H.; Lu, Z.; Wang, H.; Fan, S.; Li, N. Honokiol Attenuates Sepsis-Associated Acute Kidney Injury via the Inhibition of Oxidative Stress and Inflammation. Inflammation 2019, 42, 826-834. [CrossRef]
- Zhou, Y.; Tang, J.; Lan, J.; Zhang, Y.; Wang, H.; Chen, Q.; Kang, Y.; Sun, Y.; Feng, X.; Wu, L.; et al. Honokiol alleviated neurodegeneration by reducing oxidative stress and improving mitochondrial function in mutant SOD1 cellular and mouse models of amyotrophic lateral sclerosis. Acta Pharm Sin B 2023, 13, 577-597. [CrossRef]
- Lv, H.; Liu, J.; Wang, L.; Zhang, H.; Yu, S.; Li, Z.; Jiang, F.; Niu, Y.; Yuan, J.; Cui, X.; et al. Ameliorating effects of combined curcumin and desferrioxamine on 6-OHDA-induced rat mode of Parkinson's disease. Cell Biochem Biophys 2014, 70, 1433-1438. [CrossRef]
- Alamro, A.A.; Alsulami, E.A.; Almutlaq, M.; Alghamedi, A.; Alokail, M.; Haq, S.H. Therapeutic Potential of Vitamin D and Curcumin in an In Vitro Model of Alzheimer Disease. J Cent Nerv Syst Dis 2020, 12, 1179573520924311. [CrossRef]
- Song, S.; Nie, Q.; Li, Z.; Du, G. Curcumin improves neurofunctions of 6-OHDA-induced parkinsonian rats. Pathol Res Pract 2016, 212, 247-251. [CrossRef]
- Yu, H.; Yamashita, T.; Hu, X.; Bian, Z.; Hu, X.; Feng, T.; Tadokoro, K.; Morihara, R.; Abe, K. Protective and anti-oxidative effects of curcumin and resveratrol on Aβ-oligomer-induced damage in the SH-SY5Y cell line. J Neurol Sci 2022, 441, 120356. [CrossRef]
- Shi, M.; Sun, F.; Wang, Y.; Kang, J.; Zhang, S.; Li, H. CGA restrains the apoptosis of Aβ(25-35)-induced hippocampal neurons. Int J Neurosci 2020, 130, 700-707. [CrossRef]
- Sharma, N.; Soni, R.; Sharma, M.; Chatterjee, S.; Parihar, N.; Mukarram, M.; Kale, R.; Sayyed, A.A.; Behera, S.K.; Khairnar, A. Chlorogenic Acid: a Polyphenol from Coffee Rendered Neuroprotection Against Rotenone-Induced Parkinson's Disease by GLP-1 Secretion. Mol Neurobiol 2022, 59, 6834-6856. [CrossRef]
- Shen, Y.C.; Juan, C.W.; Lin, C.S.; Chen, C.C.; Chang, C.L. NEUROPROTECTIVE EFFECT OF TERMINALIA CHEBULA EXTRACTS AND ELLAGIC ACID IN PC12 CELLS. Afr J Tradit Complement Altern Med 2017, 14, 22-30. [CrossRef]
- Ramadan, W.S.; Alkarim, S. Ellagic Acid Modulates the Amyloid Precursor Protein Gene via Superoxide Dismutase Regulation in the Entorhinal Cortex in an Experimental Alzheimer's Model. Cells 2021, 10. [CrossRef]
- Wang, M.; Li, Y.; Ni, C.; Song, G. Honokiol Attenuates Oligomeric Amyloid β1-42-Induced Alzheimer's Disease in Mice Through Attenuating Mitochondrial Apoptosis and Inhibiting the Nuclear Factor Kappa-B Signaling Pathway. Cell Physiol Biochem 2017, 43, 69-81. [CrossRef]
- Chakraborty, K.; Krishnan, S.; Joy, M. Antioxidative oxygenated terpenoids with bioactivities against pro-inflammatory inducible enzymes from Indian squid, Uroteuthis (Photololigo) duvaucelii. Nat Prod Res 2021, 35, 909-920. [CrossRef]
- Tousif, M.I.; Nazir, M.; Riaz, N.; Saleem, M.; Tauseef, S.; Azam, S.M.; Arfan Yawer, M.; Zengin, G. Terpenoids as Human Neutrophil Elastase (HNE) Inhibitors: A Comprehensive Review of Natural Anti-inflammatory Isoprenoids. Chembiochem 2023, 24, e202300346. [CrossRef]
- Ge, J.; Liu, Z.; Zhong, Z.; Wang, L.; Zhuo, X.; Li, J.; Jiang, X.; Ye, X.Y.; Xie, T.; Bai, R. Natural terpenoids with anti-inflammatory activities: Potential leads for anti-inflammatory drug discovery. Bioorg Chem 2022, 124, 105817. [CrossRef]
- Abreu, L.S.; do Nascimento, Y.M.; Costa, R.D.S.; Guedes, M.L.S.; Souza, B.; Pena, L.J.; Costa, V.C.O.; Scotti, M.T.; Braz-Filho, R.; Barbosa-Filho, J.M.; et al. Tri- and Diterpenoids from Stillingia loranthacea as Inhibitors of Zika Virus Replication. J Nat Prod 2019, 82, 2721-2730. [CrossRef]
- Mony, T.J.; Elahi, F.; Choi, J.W.; Park, S.J. Neuropharmacological Effects of Terpenoids on Preclinical Animal Models of Psychiatric Disorders: A Review. Antioxidants (Basel) 2022, 11. [CrossRef]
- Letizia, C.S.; Cocchiara, J.; Lalko, J.; Api, A.M. Fragrance material review on linalool. Food Chem Toxicol 2003, 41, 943-964. [CrossRef]
- Pereira, I.; Severino, P.; Santos, A.C.; Silva, A.M.; Souto, E.B. Linalool bioactive properties and potential applicability in drug delivery systems. Colloids Surf B Biointerfaces 2018, 171, 566-578. [CrossRef]
- Sabogal-Guáqueta, A.M.; Hobbie, F.; Keerthi, A.; Oun, A.; Kortholt, A.; Boddeke, E.; Dolga, A. Linalool attenuates oxidative stress and mitochondrial dysfunction mediated by glutamate and NMDA toxicity. Biomed Pharmacother 2019, 118, 109295. [CrossRef]
- Yuan, C.; Shin, M.; Park, Y.; Choi, B.; Jang, S.; Lim, C.; Yun, H.S.; Lee, I.S.; Won, S.Y.; Cho, K.S. Linalool Alleviates Aβ42-Induced Neurodegeneration via Suppressing ROS Production and Inflammation in Fly and Rat Models of Alzheimer's Disease. Oxid Med Cell Longev 2021, 2021, 8887716. [CrossRef]
- Chang, W.H.; Hsu, H.T.; Lin, C.C.; An, L.M.; Lee, C.H.; Ko, H.H.; Lin, C.L.; Lo, Y.C. Linalool, a Fragrance Compound in Plants, Protects Dopaminergic Neurons and Improves Motor Function and Skeletal Muscle Strength in Experimental Models of Parkinson's Disease. Int J Mol Sci 2024, 25. [CrossRef]
- Cerdá-Bernad, D.; Valero-Cases, E.; Pastor, J.J.; Frutos, M.J. Saffron bioactives crocin, crocetin and safranal: effect on oxidative stress and mechanisms of action. Crit Rev Food Sci Nutr 2022, 62, 3232-3249. [CrossRef]
- Alayunt Ö, N.; Aksoy, L.; Karafakioğlu, Y.S.; Sevimli, S. Assessment of Anti-inflammatory and Antioxidant Properties of Safranal on CCI4-Induced Oxidative Stress and Inflammation in Rats. An Acad Bras Cienc 2019, 91, e20181235. [CrossRef]
- Xue, Y.; Jin, W.; Xue, Y.; Zhang, Y.; Wang, H.; Zhang, Y.; Guan, S.; Chu, X.; Zhang, J. Safranal, an active constituent of saffron, ameliorates myocardial ischemia via reduction of oxidative stress and regulation of Ca(2+) homeostasis. J Pharmacol Sci 2020, 143, 156-164. [CrossRef]
- Yang, W.; Qiu, X.; Wu, Q.; Chang, F.; Zhou, T.; Zhou, M.; Pei, J. Active constituents of saffron (Crocus sativus L.) and their prospects in treating neurodegenerative diseases (Review). Exp Ther Med 2023, 25, 235. [CrossRef]
- Fazeli, E.; Ghalibaf, M.H.E.; Forouzanfar, F. Neuroprotective Potency of Safranal Against Neurological Disorders. Curr Mol Med 2023, 23, 952-959. [CrossRef]
- Baluchnejadmojarad, T.; Mohamadi-Zarch, S.M.; Roghani, M. Safranal, an active ingredient of saffron, attenuates cognitive deficits in amyloid β-induced rat model of Alzheimer's disease: underlying mechanisms. Metab Brain Dis 2019, 34, 1747-1759. [CrossRef]
- Fotoohi, A.; Moloudi, M.R.; Hosseini, S.; Hassanzadeh, K.; Feligioni, M.; Izadpanah, E. A Novel Pharmacological Protective Role for Safranal in an Animal Model of Huntington's Disease. Neurochem Res 2021, 46, 1372-1379. [CrossRef]
- Ullah, H.; Di Minno, A.; Santarcangelo, C.; Khan, H.; Daglia, M. Improvement of Oxidative Stress and Mitochondrial Dysfunction by β-Caryophyllene: A Focus on the Nervous System. Antioxidants (Basel) 2021, 10. [CrossRef]
- Ojha, S.; Javed, H.; Azimullah, S.; Haque, M.E. β-Caryophyllene, a phytocannabinoid attenuates oxidative stress, neuroinflammation, glial activation, and salvages dopaminergic neurons in a rat model of Parkinson disease. Mol Cell Biochem 2016, 418, 59-70. [CrossRef]
- Tan, X.; Xu, R.; Li, A.P.; Li, D.; Wang, Y.; Zhao, Q.; Long, L.P.; Fan, Y.Z.; Zhao, C.X.; Liu, Y.; et al. Antioxidant and anti-Alzheimer's disease activities of 1,8-cineole and its cyclodextrin inclusion complex. Biomed Pharmacother 2024, 175, 116784. [CrossRef]
- Anandakumar, P.; Kamaraj, S.; Vanitha, M.K. D-limonene: A multifunctional compound with potent therapeutic effects. J Food Biochem 2021, 45, e13566. [CrossRef]
- Eddin, L.B.; Azimullah, S.; Jha, N.K.; Nagoor Meeran, M.F.; Beiram, R.; Ojha, S. Limonene, a Monoterpene, Mitigates Rotenone-Induced Dopaminergic Neurodegeneration by Modulating Neuroinflammation, Hippo Signaling and Apoptosis in Rats. Int J Mol Sci 2023, 24. [CrossRef]
- Collins, J.M.; King, A.E.; Woodhouse, A.; Kirkcaldie, M.T.; Vickers, J.C. The effect of focal brain injury on beta-amyloid plaque deposition, inflammation and synapses in the APP/PS1 mouse model of Alzheimer's disease. Exp Neurol 2015, 267, 219-229. [CrossRef]
- Kiss, E.; Kins, S.; Zöller, Y.; Schilling, S.; Gorgas, K.; Groß, D.; Schlicksupp, A.; Rosner, R.; Kirsch, J.; Kuhse, J. Artesunate restores the levels of inhibitory synapse proteins and reduces amyloid-β and C-terminal fragments (CTFs) of the amyloid precursor protein in an AD-mouse model. Mol Cell Neurosci 2021, 113, 103624. [CrossRef]
- Xia, L.; Pang, Y.; Li, J.; Wu, B.; Du, Y.; Chen, Y.; Luo, M.; Wang, Y.; Dong, Z. Dihydroartemisinin Induces O-GlcNAcylation and Improves Cognitive Function in a Mouse Model of Tauopathy. J Alzheimers Dis 2021, 84, 239-248. [CrossRef]
- Lim, H.S.; Park, G. Artemisinin protects dopaminergic neurons against 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-induced neurotoxicity in a mouse model of Parkinson's disease. Biomed Pharmacother 2024, 170, 115972. [CrossRef]
- Xu, P.; Wang, K.; Lu, C.; Dong, L.; Gao, L.; Yan, M.; Aibai, S.; Yang, Y.; Liu, X. Protective effects of linalool against amyloid beta-induced cognitive deficits and damages in mice. Life Sci 2017, 174, 21-27. [CrossRef]
- Xu, P.; Wang, K.; Lu, C.; Dong, L.; Gao, L.; Yan, M.; Aibai, S.; Yang, Y.; Liu, X. The Protective Effect of Lavender Essential Oil and Its Main Component Linalool against the Cognitive Deficits Induced by D-Galactose and Aluminum Trichloride in Mice. Evid Based Complement Alternat Med 2017, 2017, 7426538. [CrossRef]
- Pan, P.K.; Qiao, L.Y.; Wen, X.N. Safranal prevents rotenone-induced oxidative stress and apoptosis in an in vitro model of Parkinson's disease through regulating Keap1/Nrf2 signaling pathway. Cell Mol Biol (Noisy-le-grand) 2016, 62, 11-17.
- Sadeghnia, H.R.; Kamkar, M.; Assadpour, E.; Boroushaki, M.T.; Ghorbani, A. Protective Effect of Safranal, a Constituent of Crocus sativus, on Quinolinic Acid-induced Oxidative Damage in Rat Hippocampus. Iran J Basic Med Sci 2013, 16, 73-82.
- Wang, G.; Ma, W.; Du, J. β-Caryophyllene (BCP) ameliorates MPP+ induced cytotoxicity. Biomed Pharmacother 2018, 103, 1086-1091. [CrossRef]
- Flores-Soto, M.E.; Corona-Angeles, J.A.; Tejeda-Martinez, A.R.; Flores-Guzman, P.A.; Luna-Mujica, I.; Chaparro-Huerta, V.; Viveros-Paredes, J.M. β-Caryophyllene exerts protective antioxidant effects through the activation of NQO1 in the MPTP model of Parkinson's disease. Neurosci Lett 2021, 742, 135534. [CrossRef]
- Khan, A.; Vaibhav, K.; Javed, H.; Tabassum, R.; Ahmed, M.E.; Khan, M.M.; Khan, M.B.; Shrivastava, P.; Islam, F.; Siddiqui, M.S.; et al. 1,8-cineole (eucalyptol) mitigates inflammation in amyloid Beta toxicated PC12 cells: relevance to Alzheimer's disease. Neurochem Res 2014, 39, 344-352. [CrossRef]
- Jeong, S.H.; Kim, H.H.; Park, M.Y.; Bhosale, P.B.; Abusaliya, A.; Won, C.K.; Park, K.I.; Kim, E.; Heo, J.D.; Kim, H.W.; et al. Flavones: The Apoptosis in Prostate Cancer of Three Flavones Selected as Therapeutic Candidate Models. Int J Mol Sci 2023, 24. [CrossRef]
- Calderaro, A.; Patanè, G.T.; Tellone, E.; Barreca, D.; Ficarra, S.; Misiti, F.; Laganà, G. The Neuroprotective Potentiality of Flavonoids on Alzheimer's Disease. Int J Mol Sci 2022, 23. [CrossRef]
- Basli, A.; Soulet, S.; Chaher, N.; Mérillon, J.M.; Chibane, M.; Monti, J.P.; Richard, T. Wine polyphenols: potential agents in neuroprotection. Oxid Med Cell Longev 2012, 2012, 805762. [CrossRef]
- Qi, W.; Qi, W.; Xiong, D.; Long, M. Quercetin: Its Antioxidant Mechanism, Antibacterial Properties and Potential Application in Prevention and Control of Toxipathy. Molecules 2022, 27. [CrossRef]
- Mehta, V.; Parashar, A.; Udayabanu, M. Quercetin prevents chronic unpredictable stress induced behavioral dysfunction in mice by alleviating hippocampal oxidative and inflammatory stress. Physiol Behav 2017, 171, 69-78. [CrossRef]
- Ishisaka, A.; Ichikawa, S.; Sakakibara, H.; Piskula, M.K.; Nakamura, T.; Kato, Y.; Ito, M.; Miyamoto, K.; Tsuji, A.; Kawai, Y.; et al. Accumulation of orally administered quercetin in brain tissue and its antioxidative effects in rats. Free Radic Biol Med 2011, 51, 1329-1336. [CrossRef]
- Ho, C.L.; Kao, N.J.; Lin, C.I.; Cross, T.L.; Lin, S.H. Quercetin Increases Mitochondrial Biogenesis and Reduces Free Radicals in Neuronal SH-SY5Y Cells. Nutrients 2022, 14. [CrossRef]
- Amanzadeh Jajin, E.; Esmaeili, A.; Rahgozar, S.; Noorbakhshnia, M. Quercetin-Conjugated Superparamagnetic Iron Oxide Nanoparticles Protect AlCl(3)-Induced Neurotoxicity in a Rat Model of Alzheimer's Disease via Antioxidant Genes, APP Gene, and miRNA-101. Front Neurosci 2020, 14, 598617. [CrossRef]
- Sriraksa, N.; Wattanathorn, J.; Muchimapura, S.; Tiamkao, S.; Brown, K.; Chaisiwamongkol, K. Cognitive-enhancing effect of quercetin in a rat model of Parkinson's disease induced by 6-hydroxydopamine. Evid Based Complement Alternat Med 2012, 2012, 823206. [CrossRef]
- Sandhir, R.; Mehrotra, A. Quercetin supplementation is effective in improving mitochondrial dysfunctions induced by 3-nitropropionic acid: implications in Huntington's disease. Biochim Biophys Acta 2013, 1832, 421-430. [CrossRef]
- Viswanatha, G.L.; Shylaja, H.; Moolemath, Y. The beneficial role of Naringin- a citrus bioflavonoid, against oxidative stress-induced neurobehavioral disorders and cognitive dysfunction in rodents: A systematic review and meta-analysis. Biomed Pharmacother 2017, 94, 909-929. [CrossRef]
- Hassan, H.M.; Elnagar, M.R.; Abdelrazik, E.; Mahdi, M.R.; Hamza, E.; Elattar, E.M.; ElNashar, E.M.; Alghamdi, M.A.; Al-Qahtani, Z.; Al-Khater, K.M.; et al. Neuroprotective effect of naringin against cerebellar changes in Alzheimer's disease through modulation of autophagy, oxidative stress and tau expression: An experimental study. Front Neuroanat 2022, 16, 1012422. [CrossRef]
- Kumar, P.; Kumar, A. Protective effect of hesperidin and naringin against 3-nitropropionic acid induced Huntington's like symptoms in rats: possible role of nitric oxide. Behav Brain Res 2010, 206, 38-46. [CrossRef]
- Parhiz, H.; Roohbakhsh, A.; Soltani, F.; Rezaee, R.; Iranshahi, M. Antioxidant and anti-inflammatory properties of the citrus flavonoids hesperidin and hesperetin: an updated review of their molecular mechanisms and experimental models. Phytother Res 2015, 29, 323-331. [CrossRef]
- Tamilselvam, K.; Braidy, N.; Manivasagam, T.; Essa, M.M.; Prasad, N.R.; Karthikeyan, S.; Thenmozhi, A.J.; Selvaraju, S.; Guillemin, G.J. Neuroprotective effects of hesperidin, a plant flavanone, on rotenone-induced oxidative stress and apoptosis in a cellular model for Parkinson's disease. Oxid Med Cell Longev 2013, 2013, 102741. [CrossRef]
- Bakr, A.F.; Shao, P.; Farag, M.A. Recent advances in glycyrrhizin metabolism, health benefits, clinical effects and drug delivery systems for efficacy improvement; a comprehensive review. Phytomedicine 2022, 99, 153999. [CrossRef]
- Ban, J.Y.; Park, H.K.; Kim, S.K. Effect of Glycyrrhizic Acid on Scopolamine-Induced Cognitive Impairment in Mice. Int Neurourol J 2020, 24, S48-55. [CrossRef]
- Gendy, A.M.; El-Sadek, H.M.; Amin, M.M.; Ahmed, K.A.; El-Sayed, M.K.; El-Haddad, A.E.; Soubh, A. Glycyrrhizin prevents 3-nitropropionic acid-induced neurotoxicity by downregulating HMGB1/TLR4/NF-κB p65 signaling, and attenuating oxidative stress, inflammation, and apoptosis in rats. Life Sci 2023, 314, 121317. [CrossRef]
- Molaei, A.; Hatami, H.; Dehghan, G.; Sadeghian, R.; Khajehnasiri, N. Synergistic effects of quercetin and regular exercise on the recovery of spatial memory and reduction of parameters of oxidative stress in animal model of Alzheimer's disease. Excli j 2020, 19, 596-612. [CrossRef]
- Bao, D.; Wang, J.; Pang, X.; Liu, H. Protective Effect of Quercetin against Oxidative Stress-Induced Cytotoxicity in Rat Pheochromocytoma (PC-12) Cells. Molecules 2017, 22. [CrossRef]
- Kaur, G.; Prakash, A. Involvement of the nitric oxide signaling in modulation of naringin against intranasal manganese and intracerbroventricular β-amyloid induced neurotoxicity in rats. J Nutr Biochem 2020, 76, 108255. [CrossRef]
- Kesh, S.; Kannan, R.R.; Balakrishnan, A. Naringenin alleviates 6-hydroxydopamine induced Parkinsonism in SHSY5Y cells and zebrafish model. Comp Biochem Physiol C Toxicol Pharmacol 2021, 239, 108893. [CrossRef]
- Adekeye, A.O.; Fafure, A.A.; Ogunsemowo, A.E.; Enye, L.A.; Saka, O.S.; Ogedengbe, O.O. Naringin ameliorates motor dysfunction and exerts neuroprotective role against vanadium-induced neurotoxicity. AIMS Neurosci 2022, 9, 536-550. [CrossRef]
- Sugumar, M.; Sevanan, M.; Sekar, S. Neuroprotective effect of naringenin against MPTP-induced oxidative stress. Int J Neurosci 2019, 129, 534-539. [CrossRef]
- Justin Thenmozhi, A.; William Raja, T.R.; Manivasagam, T.; Janakiraman, U.; Essa, M.M. Hesperidin ameliorates cognitive dysfunction, oxidative stress and apoptosis against aluminium chloride induced rat model of Alzheimer's disease. Nutr Neurosci 2017, 20, 360-368. [CrossRef]
- Hassan, W.; Noreen, H.; Rehman, S.; Kamal, M.A.; da Rocha, J.B.T. Association of Oxidative Stress with Neurological Disorders. Curr Neuropharmacol 2022, 20, 1046-1072. [CrossRef]
- Marino, A.; Battaglini, M.; Moles, N.; Ciofani, G. Natural Antioxidant Compounds as Potential Pharmaceutical Tools against Neurodegenerative Diseases. ACS Omega 2022, 7, 25974-25990. [CrossRef]
- Delgado, A.; Gonçalves, S.; Romano, A. Mediterranean Diet: The Role of Phenolic Compounds from Aromatic Plant Foods. Foods 2023, 12. [CrossRef]
- Vlčko, T.; Rathod, N.B.; Kulawik, P.; Ozogul, Y.; Ozogul, F. The impact of aromatic plant-derived bioactive compounds on seafood quality and safety. Adv Food Nutr Res 2022, 102, 275-339. [CrossRef]
- Korczowska-Łącka, I.; Słowikowski, B.; Piekut, T.; Hurła, M.; Banaszek, N.; Szymanowicz, O.; Jagodziński, P.P.; Kozubski, W.; Permoda-Pachuta, A.; Dorszewska, J. Disorders of Endogenous and Exogenous Antioxidants in Neurological Diseases. Antioxidants (Basel) 2023, 12. [CrossRef]
Figure 1.
Processes of oxidative stress in neurodegenerative diseases. ROS are produced from extracellular and intracellular sources. Extracellular sources of ROS are dominated by radiation exposure, microbial infection, and exposure to engineered nanoparticles. Intracellular ROS can be produced by mitochondria, ER stress, cellular metabolic enzymes and NOX family. With age, the increase in the levels of reactive oxygen radicals, reactive nitrogen radicals, and lipid peroxidation in the brain, coupled with the decrease in the levels of antioxidant enzymes, leads to a gradual decrease in the efficiency of the endogenous antioxidant system, resulting in oxidative damage to neuronal cells and leading to the development of neurodegenerative diseases. ER: endoplasmic reticulum; ROS: reactive oxygen species; RNS: reactive nitrogen species; MDA: malondialdehyde; GSH: glutathione; GPx: glutathione peroxidase; CAT: catalase; SOD: superoxide dismutase.
Figure 1.
Processes of oxidative stress in neurodegenerative diseases. ROS are produced from extracellular and intracellular sources. Extracellular sources of ROS are dominated by radiation exposure, microbial infection, and exposure to engineered nanoparticles. Intracellular ROS can be produced by mitochondria, ER stress, cellular metabolic enzymes and NOX family. With age, the increase in the levels of reactive oxygen radicals, reactive nitrogen radicals, and lipid peroxidation in the brain, coupled with the decrease in the levels of antioxidant enzymes, leads to a gradual decrease in the efficiency of the endogenous antioxidant system, resulting in oxidative damage to neuronal cells and leading to the development of neurodegenerative diseases. ER: endoplasmic reticulum; ROS: reactive oxygen species; RNS: reactive nitrogen species; MDA: malondialdehyde; GSH: glutathione; GPx: glutathione peroxidase; CAT: catalase; SOD: superoxide dismutase.
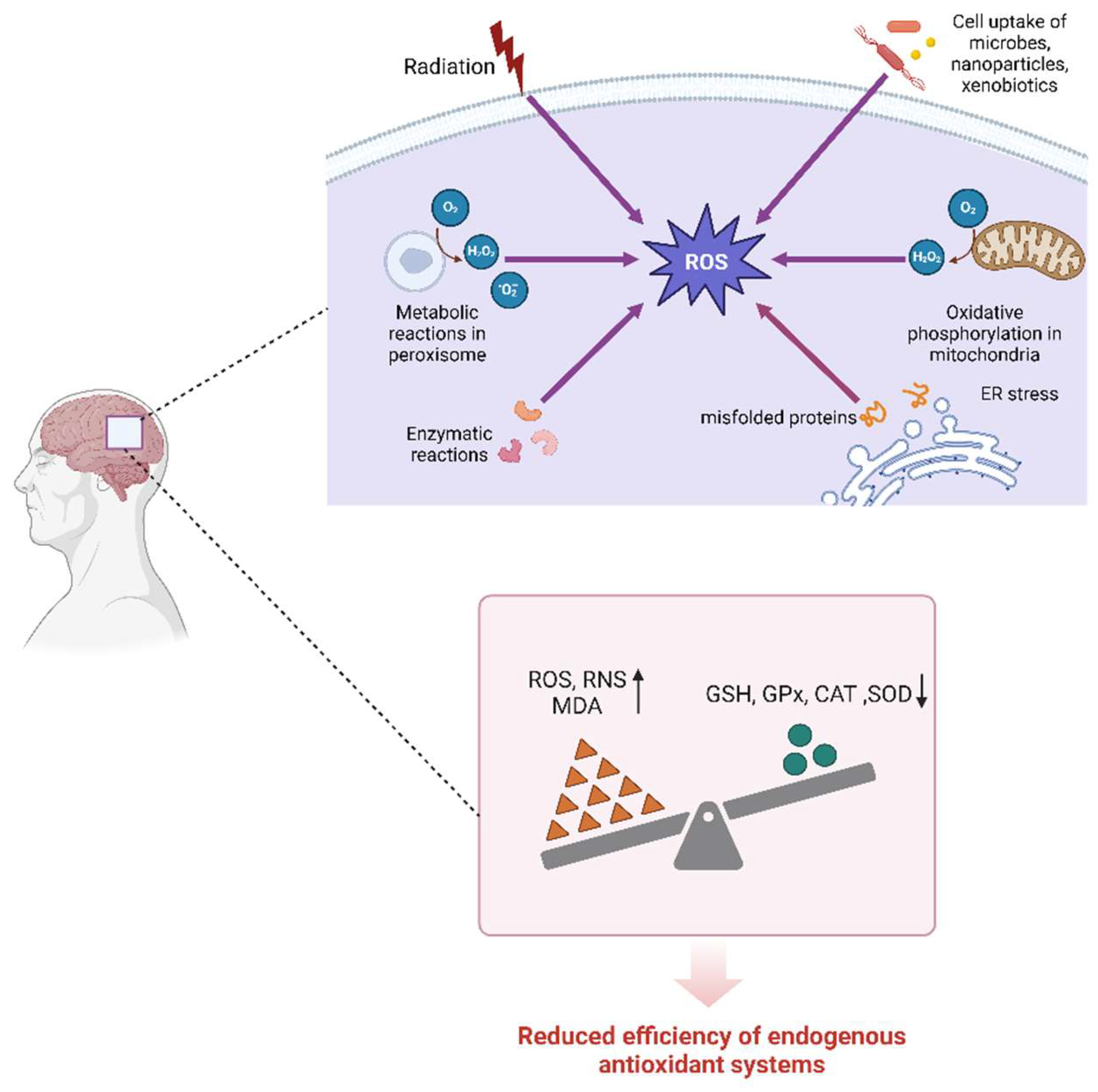
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



