
Submitted:
20 September 2024
Posted:
23 September 2024
You are already at the latest version
Abstract
Dietary intake of Wistar rats of biofortified with 8-hydroxy-7-iodo-5-quinolinesulfonic acid (8-OH-7-I-5QSA) kale of both cultivars ‘Oldenbor F1’ and ‘Redbor F1’ for 8 weeks resulted in sig-nificantly higher iodine concentrations in the urine and kidneys of rats, which proves iodine bio-availability. Rats diets with 'Oldenbor F1' and 'Redbor F1' kale non- and -biofortified with 8-OH-7-I-5QSA had a significantly lower or a tendency for lower concentration of TSH, triglyc-eride, total and direct bilirubin, TBARs, uric acid, aspartate aminotransferase (AST) and alanine aminotransferase (ALT) concentrations in serum. Dietary intake of 'Oldenbor F1' and 'Redbor F1' kale biofortified with 8-OH-7-I-5QSA significantly increased the total antioxidant status (TAS). The study confirms that kale biofortified with iodine in organic form iodoquinoline 8-OH-7-I-5QSA is bioavailable and well absorbed by the Wistar rat and has a positive effect on selected biochemical parameters.
Keywords:
iodoquinoline
; 8-hydroxy-7-iodo-5-quinolinesulfonic acid
; iodine
; kale
; biofortification
; biochemical parameters
; iodine bioavailability
1. Introduction
Iodine is a key trace element required for the production of thyroid gland hormones (3,5,3’-triiodo-L-thyronine, T3, 3,5,3’,5'-tetraiodo-L-thyronine, T4), which are essential for mammalian life [1]. The iodine-replete healthy adult has about 15–20 mg of iodine, 70%–80% of which is contained in the thyroid [2].
Its bioavailability depends on oral intake, and the recommended daily intake varies according to the age and physiological state of the individual. Iodine can also be absorbed from the air, through the mucous membranes of the respiratory system, and by the skin [3].
The most common method of addressing iodine deficiency is the iodisation of table salt, although critics point out that this does not always cover the full need for the element. Salt iodisation is not sufficient, due to the volatility of iodine and its loss during storage, transport and cooking. In addition, it is not recommended for people with cardiovascular disease, although the impact is still debated [4]. In most iodine-deficient countries, the iodisation of table salt is widely recognised as the most effective way to eliminate iodine-deficiency diseases (IDD). Despite this, one-third of the world's population is still not protected against iodine deficiency [5].
Mineral deficiencies can be addressed through dietary diversity and supplementation. An alternative and often complementary, method is the of biofortified foods, which can help to address micronutrient deficiencies. Biofortification as a strategy, is well documented and economically and environmentally effective in combating mineral malnutrition in various populations [6, 7]. Studies on appropriate application methods and dosage of micronutrients in plant crops have shown that this benefits both humans and plants [8, 9, 10, 11, 12, 13, 14, 15]. Biofortification aims to enrich plants with nutrients during their growth, instead of adding micronutrients after food processing [16]. The most common methods of biofortification include the use of nutrient fertilisers, plant crossing and genetic manipulation [17, 18, 19].
Previous research has focused on the enrichment of vegetables with mineral forms of iodine, such as KI and KIO3 [15, 20, 21, 22]. Organic iodine compounds such as iodosalicylates and iodobenzoates have been used to enrich lettuce [23, 24] and tomatoes [25, 26] and gave satisfactory results for iodine accumulation vs. mineral forms. Furthermore, the use of the organic iodine compounds, such as iodosalicylates, fits in with the ecological strategy of improving soil quality and iodine-enriched plants. In our study, we used iodoquinolines as organic forms of iodine for the first time. This was dictated by the fact that quinoline derivatives have many health-promoting properties, including antioxidant, anti-inflammatory, anticancer, antibacterial, antiviral and antifungal activities [27]. Moreover, we were the first to attempt to biofortify kale with iodoquinolines for several reasons. Kale is a rich source of nutrients such as: vitamins (C, A, K), folic acid and minerals (calcium, iron, magnesium) and non-nutrients i.e. polyphenols and glucosinolates [28]. It is indicated that these bioactive substances enhance its anticancer and anti-inflammatory potential [29]. On the other hand, kale as a cruciferous vegetable, contains substances that impede iodine absorption, therefore we assumed that iodoquinolines combined with kale - a vegetable with proven health-promoting properties - would bring tangible health benefits: 1) additional iodine in the diet and 2) contribute to increased of kale, which will translate into health benefits for consumers.
In our study, to our knowledge, we were the first to show that that kale biofortified with iodine in organic form 8-OH-7-I-5QSA is bioavailable and well absorbed by the Wistar rat and has a positive effect on selected thyroid-regulated biochemical parameters.
2. Materials and Methods
2.1. Kale ‘Oldenbor F1’ and ‘Redbor F1’ Cultivation and Biofortification with 8-Hydroxy-7-Iodo-5-Quinolinesulfonic Acid
Hydroponic cultivation of two cultivars of kale (Brassica oleracea var. sabellica L.) –'Oldenbor F1' (green kale) and 'Redbor F1' (red kale) was carried out in spring 2022. The experiments took place in a greenhouse on the campus of the Faculty of Biotechnology and Horticulture of the University of Agriculture in Kraków (50°05'00.3 "N 19°57'00.1 "E). The seeds were sown into propagation trays with 96 chambers, each measuring 42x42x65 mm, filled with vermiculite. Seedlings with three true leaves were transplanted into a hydroponic Nutrient Film Technique (NFT) system. A no-till growing method (NFT) was used, where plants were placed in the tray openings at 30 cm intervals. Plants were irrigated in day and night cycles, 5:00-19:00 and 1:00-2:00, respectively, for one minute every 5 minutes. The nutrient solution contained micro- and macronutrients at concentrations (mg·L-1) of: N-130, P-50, K-220, Ca-120, Mg-45, Fe-2, Mn-0.55, B-0.33, Zn-0.33, Cu-0.15, Mo-0.05 and pH corrected to 5.70 with 38% nitric acid. The final electrical conductivity (EC) was 2.0 mS·cm-1.
The experiment consisted of two treatments: a control one and an application of 8-hydroxy-7-iodo-5-quinolinosulphonic acid at a concentration of 10 µM, which started at the 4-5 leaf stage and continued until harvest. Four replicates were carried out with 6 plants in each, yielding 24 plants per combination of each cultivar and a total of 120 plants for each cultivar. Plants were harvested after 63 days of cultivation, at the 13-14 leaf stage.
2.2. Analysis in Plant Material
Fresh kale samples were frozen and freeze-dried using a Christ Alpha Gamma 1-16 LSCplus lyophiliser (Martin Christ Gefriertrocknungsanlagen GmbH, Harz, Germany) and ground in a laboratory hurricane mill (WZ-1, Sadkiewicz Instruments, ZDPP, Poland). The kale samples were stored in sealed polyethylene bags (at 2-8 °C) until the analysis. The basic chemical composition was measured in the samples thus prepared. Crude fat (method No. 935.38), total protein (method No. 950.36), total dietary fibre (method No. 985.29) [30] and ash (method No. 930.05) were measured according to AOAC [31, 32]. Digestible carbohydrates were calculated using the formula:
Digestible Carbohydrates = 100 - (protein + crude fat + dietary fibre + ash); [33].
The samples were analysed to determine the concentrations of iodine, using the ICP-MS/MS technique (Inductively coupled plasma–triple quadrupole mass spectrometry; iCAP TQ ICP-MS; ThermoFisher Scientific, Bremen, Germany). The analysis was based on research published by Smoleń et al. (PN-EN, 15111:2008) [34, 35]. The iodine content in freeze-dried kale leaves samples was analysed via the alkaline extraction (0.2 g of the samples) using tetramethylammonium hydroxide (TMAH), during the extraction process, amylase was used to digest starch. Next, inductively coupled plasma mass spectrometry (ICP-MS/MS) with a triple quadruple spectrometer (iCAP TQ ICP-MS), will be used. For the analysis, 0.2 g of freeze-dried leaves kale samples, 10 mL of double distilled water, and 1 mL of 25% TMAH (Sigma-Aldrich) were put into 30 mL falcon tubes. After thorough mixing, the samples were incubated for 3 hours at 90 °C. Following incubation, the samples were cooled to approximately 20 °C and made up to 30 mL with double distilled water. After mixing, the samples were centrifuged for 15 min at 4500 rpm. Iodine content was analysed in the supernatants. The same method was used to determine the iodine content of experimental diets for rats.
2.3. Animal Study
Five week old Wistar rats (n=40, males) were obtained from the Animal Husbandry at the Animal Jagiellonian University Medical College, Krakow, Poland. The first Local Ethical Committee approved the experimental procedures for the Animal Experiments in Krakow (Poland, res.no. 568/2021). Before the experiment, the animals were acclimatised for 7 days on standard laboratory chow. At the end of the acclimatisation period, the animals were randomly divided into five experimental groups (n=8). The mean body weight of the rats at the beginning of the experiment was 132 ± 10 g. The experimental diets were prepared based on AIN-93G [36]. The detailed composition of the diets can be found in Table 1, below. The research on iodine bioavailability was carried out only on males rats due to the limited number of animals in the study (according to the 3R rule).
Group 1 was fed an AIN-93G (C) diet, consisting of a mineral mixture containing iodine as recommended by [36]. For Group 2 (CO) with control 'Oldenbor F1' kale and for Group 4 (CR) with control 'Redbor F1' kale, both diets were prepared with a mineral mixture with iodine. In the diets containing biofortified kale (Group 3, BO diet with biofortified 8-OH-7-I-5QSA 'Oldenbor F1' kale and Group 5, BR diet with biofortified 8-OH-7-I-5QSA 'Redbor F1' kale), the only source of iodine was kale (the mineral mixture was devoid of iodine – Table 1).
Rodents were individually housed in steel metabolic cages during the first and eighth week of the experiment. The temperature was maintained at 21°C and the light/dark cycle was 12/12 hours. The animals had unlimited access to deionised distilled water throughout the experiment. The intake of the experimental diets was recorded daily. During the remaining weeks, rats were housed in conventional open-air cages with two or three individuals per cage. Body weight gain was measured weekly for the entire duration of the experiment, which lasted 8 weeks. Results were expressed as a fresh weight (µg·L-1) in urine and a dry weight (g·kg-1) in faeces.
Urine and faeces were collected from day 1 to day 5 and from day 49 to 54 of the experiment (week 1 and week 8, respectively) to assess iodine excretion. The collected samples were stored at -20°C until analysis. At the end of the 8-week experimental period, fasted rats (12 hours) were anaesthetised (isoflurane 4%, inhaled). Blood was collected by cardiac puncture into standard tubes. Blood samples were centrifuged (1500 x ƍ, 15 min) to obtain serum. The liver, kidney, thyroid gland and heart were excised, washed with 0.9% sodium chloride solution, dried with a paper towel and weighed. Serum and tissue samples were frozen at -80°C for analysis.
2.4. Iodine Content in Urine, Faeces and Selected Tissues
Collected samples of urine were adjusted to the same volume before analysis. The faeces, kidneys, livers were freeze-dried. After freeze-drying, the organs were weighed and ground in a laboratory hurricane mill (WZ-1. Sadkiewicz Instruments. ZDPP. Poland). Then the prepared samples (particle size about 1 mm) were used for measurements of iodine content. The content of iodine in these samples was analysed using the ICP-MS/MS technique (Inductively coupled plasma triple quadrupole mass spectrometry; iCAP TQ ICP-MS; ThermoFisher Scientific, Bremen, Germany, (PN-EN. 15111:2008) [35]. The procedure for the analysis of iodine in faeces, kidneys, livers was the same as for plant samples. Urinary iodine content was analysed according to the following procedure:
Urine samples were thawed and mixed. Then 4.8 mL of urine were collected into a PP tube, 0.2 mL of 25% TMAHu was added and incubated with this reagent. Subsequently, the samples were mixed and diluted 200x in water redistilled three times. Then the analysis was performed using the ICP-MS/MS technique.
2.5. Analysis of Serum
Serum levels of total cholesterol – TC; (cat. no. Liquick Cor-CHOL 60, 2-204, PZ Cormay S.A., Lublin, Poland), HDL cholesterol (cat. no. CORMAY HDL, 2-053, PZ Cormay S.A.) and triglycerides-TG (cat. no. Liquick Cor-TG 60, 2-253, PZ Cormay S.A.) were measured. Differences between TC and HDL were used to calculate LDL + VLDL levels [37]. The liver panel was examined including ALAT alanine aminotransferase activity; (cat. no. Liquick Cor-ALAT 60, 1-217, PZ Cormay S.A.), aspartate aminotransferase ASAT (cat. no. Liquick Cor-ASAT 60, 1-214, PZ Cormay S.A.). Total bilirubin – TB (cat. no. Liquick Cor-BIL TOTAL 60, 2-245, PZ Cormay S.A.), and direct bilirubin – DB (on cat. Liquick Cor-BIL DIRECT MALLOY-EVELYN 60, 2-348, PZ Cormay S.A.) were determined. Serum uric acid levels were also tested (cat. no. Liquick Cor-UA 60, 2-208, PZ Cormay S.A.).
Triiodothyronine (T3) levels were measured using ELISA Tests Enzyme-linked Immunosorbent Assay Kit for Triiodothyronine (cat. no. CEA453Ge, Cloud-Clone Corp, Katy, TX 77494, USA), and thyroxine (T4) levels were measured using ELISA Tests Enzyme-linked Immunosorbent Assay Kit for Thyroxine (cat. no. CEA452Ge, Cloud-Clone Corp, Katy, TX 77494, USA). Thyrotropic hormone (TSH) concentrations were determined with the Rat Thyroid Stimulating Hormone ELISA kit (cat. no. RTC007R, BioVendor R&D, Karásek, Brno, Czech Republic).
Glutathione reductase levels were determined using the Glutathione Reductase (GLUT RED) kit (cat. no. GR 2368, RANDOX, Crumlin, United Kingdom) and total oxidant status using the Total Antioxidant Status (TAS) kit (cat. no. NX 2332, RANDOX, Crumlin, United Kingdom). Determination of serum lipid peroxidation products was performed using the thiobarbituric acid - TBARs interaction according to Ohkawa, Ohishi & Yagi [38]. Results were expressed in terms of free malondialdehyde (MDA) a marker of lipid peroxidation, which is a marker of oxidative stress.
2.6. Statistical Analysis
The data were presented as mean ± SD (n=8). The experimental data were analysed using a one-factor analysis of variance (ANOVA), except for the content of iodine in urine and faeces (two-factor). Statistics were performed using the Statistica software v. 13.1 PL (Dell Inc., Tulsa, OK, USA). The level of significance considered was P ≤ 0.05. In the case of significant effects, homogeneous groups were distinguished by Duncan's post hoc test. In the two-factor experiment, the variables were: Factor No.1 type of diet: C – control diet (AIN-93G), CO – diet containing control curly kale 'Oldenbor F1', BO – diet containing biofortified curly kale 'Oldenbor F1', CR – diet containing control curly kale 'Redbor F1', BR – diet containing biofortified curly kale 'Redbor F1’, x Factor No.2 week: I, VIII.
3. Results
3.1. Iodine Content and Basic Chemical Composition in Plant Material
The control (non-biofortified) kale of both cultivars 'Oldenbor F1' and 'Redbor F1' was not significantly different in terms of iodine content (Table 2). Application of the biofortification process with 8-OH-7-I-5QSA significantly increased the iodine content of the plants of both cultivars, from 0.18 to 2.10 of 'Oldenbor F1' and from 0.20 to 2.43 of 'Redbor F1' mg·kg d.m.-1, respectively. Furthermore, the red cultivar 'Redbor F1' accumulated a significantly higher iodine content (P ≤ 0.05).
Analysing the results in Table 2 showed that green kale 'Oldenbor F1' (control and biofortified with 8-OH-7-I-5QSA) had more protein (P ≤ 0.05) than kale of the red cultivar 'Redbor F1' (control and biofortified with 8-OH-7-I-5QSA).
The total fat content of green kale of 'Oldenbor F1' cultivar in the control as well as the biofortified version was also higher (P ≤ 0.05) than that of red kale of 'Redbor F1' cultivar in the control and biofortified version.
For digestible carbohydrates, we notice significant changes (P ≤ 0.05) after biofortification with 8-OH-7-I-5QSA. For both cultivars of kale 'Oldenbor F1' and 'Redbor F1', the biofortification process with 8-OH-7-I-5QSA increased the digestible carbohydrate content compared to the control. Significant differences in digestible carbohydrates were observed between the two cultivars. The green cultivar 'Oldenbor F1' had more digestible carbohydrates than the red cultivar 'Redbor F1'.
The ash content of 'Redbor F1' kale was higher (P ≤ 0.05) compared to 'Oldenbor F1', for both control and biofortified with 8-OH-7-I-5QSA kale. A decreasing effect of the 8-OH-7-I-5QSA biofortification process, for both cultivars of kale 'Oldenbor F1' and 'Redbor F1', on the ash content was observed (P ≤ 0.05).
The kale of the red cultivar 'Redbor F1' had a higher dietary fibre content compared to the green cultivar 'Oldenbor F1' (P ≤ 0.05).
3.2. Body Weight Gain, Weight of Selected Organs
Table 3 shows the set of results on body gain, feed efficiency ratio (FER) and weight of selected organs. Dietary intake of Wistar rats of non-biofortified kale (CR) and kale biofortified with 8-OH-7-I-5QSA of both cultivars, ‘Oldenbor F1’ (BO) for 8 weeks resulted in significantly lower body weight gain compared to the AIN-93G (C) control. A trend towards lower body weight gain was shown by the CO, BR groups.
The feed efficiency ratio (FER) of rats fed diets with non-biofortified kale (CO, CR) and kale biofortified with 8-OH-7-I-5QSA (BO, BR) was lower compared to the fed control diet AIN-93G (C) without kale, but significantly (P ≤ 0.05) only for the BO and CR groups.
The liver weights of rats fed non-biofortified kale (CO, CR) and biofortified with 8-OH-7-I-5QSA kale (BO, BR) diet were the lowest (P ≤ 0.05) compared to the AIN-93G (C).
The sum of the weights of both kidneys in the group of rats fed the control diet AIN-93G (C) was similar (P ≥ 0.05) compared to the other experimental groups (CO, BO, BR). Kidneys weight was significantly lowest in the group of rats that fed non-biofortified 'Redbor F1' kale (CR).
Heart weight in the group of rats fed the AIN-93G (C) control diet was similar too (P ≥ 0.05) compared to the other experimental groups fed non-biofortified and biofortified with 8-OH-7-I-5QSA kale (CO, BO, BR). Significantly decrease of heart weight was only for CR group.
The thyroid gland weight of rats fed a diet enriched with non-biofortified kale (CO, CR) and kale biofortified with 8-OH-7-I-5QSA (BO) was similar (P ≥ 0.05) compared to control group (C), except for the BR group, where the weight of this organ increased significantly.
Visceral fat content was not significantly different in any group of rats fed non-biofortified kale (CO, CR) and kale biofortified with 8-OH-7-I-5QSA of both cultivars (BO, BR). In our study visceral fat content did not differ significantly in any rat study group (P ≥ 0.05).
3.3. Iodine Excretion in Urine, Faeces and Selected Organs
After 8 weeks on the diet, higher urinary iodine concentrations were determined in the BO study group. In addition, a trend towards higher urinary iodine concentrations after 8 weeks was also shown in the BR group. Thus, feeding diets with kale of both cultivars ('Oldenbor F1' and 'Redbor F1') biofortified with 8-OH-7-I-5QSA generally increased urinary iodine concentrations in time (Figure 1A).
A different situation was observed in the faecal iodine content of the Wistar rats tested. In general, no significant changes were shown in faecal iodine content at the beginning and end of the experiment (Figure 1B).
The highest kidney iodine concentrations were determined in rodents fed diets of biofortified kale cultivars 'Oldenbor F1' (BO) and 'Redbor F1' (BR). Biofortified 'Oldenbor F1' kale (BO) had a higher iodine content in kidneys, than non-biofortified kale (CO). The same situation was for biofortified 'Redbor F1' kale (BR), where kidney iodine content was higher than non-biofortified control kale (CR). No similar results were found in the livers of the experimental rats (Table 4).
3.4. Selected Biochemical Parameters
The feeding of different types of diets by Wistar rats did affect the change (P ≥ 0.05) in the activity of the liver enzyme aspartate aminotransferase (AST) after 8 weeks of experimentation (Table 5). After feed of BR kale, a decrease (P ≥ 0.05) in AST enzyme activity or a tendency for BO to decrease (P ≥ 0.05) was observed. Feeding of the kale cultivar ‘Redbor F1’ reduces AST enzyme concentrations the most. In the liver enzyme alanine aminotransferase (ALT), where dietary use of 'Redbor F1' kale non-biofortified (CR) and 'Redbor F1' biofortified with 8-OH-7-I-5QSA (BR), resulted in a significant reduction in enzyme activity or only a tendency for it (P ≥ 0.05), in the case of ‘Oldenbor F1’ kale enriched in 8-OH-7-I-5QSA (BO). The addition of 'Redbor F1' non-biofortified and biofortified red kale to the diets significantly affected lower enzyme activity values, compared to the AIN-93G (C) diet and 'Oldenbor F1' control green kale (CO).
Total bilirubin values were the highest (P ≤ 0.05) in the group of rats fed the AIN-93G (C) control diet. The other experimental groups that were fed non-biofortified and biofortified kale had significantly lower total bilirubin values (Table 5). A similar situation was observed for direct bilirubin, where the experimental groups (CO, BO, BR), compared to the AIN-93G diet (C), showed lower concentrations (P ≤ 0.05) or a tendency (P ≥ 0.05) for a lower concentration values (CR).
The serum uric acid values of Wistar rats in our experiment were not affected by the diets (P ≥ 0.05). However, there was a trend towards lower uric acid concentrations in Wistar rats after feeding non- and biofortified with 8-OH-7-I-5QSA kale in both cultivars. Moreover, the lowest uric acid concentrations were in the groups where iodoquinoline was added.
Total cholesterol (TC) values in rats fed different diets did not differ significantly between groups (Table 5). HDL fraction cholesterol values were similar in all groups. Only a tendency for the highest HDL value was recorded in the AIN-93G (C) fed rats compared to the CO, BO and BR groups (P ≥ 0.05). But a significantly lower HDL fraction cholesterol value was recorded in the group of rats fed 'Redbor F1' control kale (CR) compared to the C group. LDL + VLDL cholesterol concentrations were not significantly different between all experimental groups. Triglyceride concentrations had only a tendency for the highest concentration in the group of rats fed AIN-93G (C), compared to the other experimental groups (CO, CR, BR), or were significantly higher compared to the BO group.
Antioxidant activity was determined in the blood serum of the test rats by glutathione reductase (RG), total antioxidant status (TAS) and products of lipid peroxidation by reaction with thiobarbituric acid (TBARs) – Table 5. After 8 weeks of the experiment, glutathione reductase levels did not differ significantly between the experimental groups (P ≥ 0.05).
Serum TAS determination in rats showed that dietary intake of 'Oldenbor F1' (BO) and 'Redbor F1' (BR) kale biofortified with 8-OH-7-I-5QSA significantly increased (or in one case resulted in a tendency) total antioxidant status relative to dietary intake of AIN-93G (C) and 'Oldenbor F1' green kale (CO).
The concentration of thiobarbituric acid reactive substances (TBARs) expressed in nmol malondialdehyde (MDA)·mL-1 was significantly highest in the AIN-93G control group (C) in comparison with the group fed 'Redbor F1' (CR, BR) or a tendency for the highest concentration was found in comparison with the CO and BO groups (P ≥ 0.05). MDA concentrations were dependent on the use of non-biofortified and biofortified 'Redbor F1' (CR, BR). It was observed that the addition to the diet of these two kinds of kale significantly reduced MDA concentrations in rats compared to rats fed with the control (C) diet (Table 5).
The highest thyrotropic hormone (TSH) concentrations were observed in the experimental group feeding the AIN-93G (C) diet compared to the groups fed the CO and BO diet (P ≤ 0.05) or a tendency for the highest concentration was found compared to the groups fed the CR and BR diet (P ≥ 0.05). Triiodothyronine (T3) and thyroxine (T4) hormone concentrations were not significantly different between all experimental groups of rats tested (P ≥ 0.05).
4. Discussion
Iodine is one of the trace elements essential for human life. The main objective of the biofortification of plants with iodine is to obtain foods with a higher content of this element compared to conventional foods. Biofortification of plants with iodine can increase the intake of this trace element by different populations. In addition, it may reduce the risk of iodine deficiency diseases [33].
In this study, the effects of kale (Brassica oleracea var. sabellica L.) of two cultivars, 'Oldenbor F1' and 'Redbor F1' biofortified with iodine in the form of 8-OH-7-I-5QSA, on iodine content in urine, faeces, selected tissues and various biochemical parameters of Wistar rats were evaluated for the first time.
Before moving on to the analysis of the in vivo experiment, we will first focus on the evaluation of iodine content and the basic chemical composition of the test kale Brassica oleracea var. sabellica L. of two cultivars, 'Oldenbor F1' and 'Redbor F1', which was added to the diet of Wistar rats. Kale is a long-known vegetable with high nutritional value due to its high content of bioactive compounds and macro- and micronutrients [39]. Improving the nutritional value of this vegetable by increasing the overall iodine content, through the application of a biofortification process, further enhances this crop. Application of iodine in the form of 8-OH-7-I-5QSA by nutrient solutions in hydroponic cultivation significantly increased the iodine content and did not affect the basic chemical composition of kale compared to control plants. However, differences in basic chemical composition were observed depending on the cultivar tested.
Application of the biofortification process with 8-OH-7-I-5QSA significantly increased the iodine content of the plants from 0.18 to 2.10 mg·kg s.m.-1 in the 'Oldenbor F1' cultivar and from 0.20 to 2.43 mg·kg s.m.-1 in the 'Redbor F1' cultivar, respectively. Furthermore, the red cultivar 'Redbor F1' accumulated a significantly higher iodine content (2.43 mg·kg d.m.-1) after biofortification compared to the green cultivar 'Oldenbor F1' (2.10 mg·kg d.m.-1). Previous studies by many authors confirm the effectiveness of the iodine biofortification process on increasing the content of this element in vegetables. This studies have mainly focused on the enrichment of vegetables with mineral forms of iodine (KI and KIO3) [15, 20, 21, 22]. Recently, our team has begun research on enriching plants, including vegetables, with organic forms of iodine. Organic forms of iodine i.e. iodosalicylates and iodobenzoates, have been used to enrich, for example lettuce [23, 24] or tomatoes [25, 26]. Moreover, the results of biofortification of vegetables by iodoquinolines, presented for the first time by our team, showed the effectiveness of these organic forms for the treatment of enriching plants, such as kale, lettuce and potatoes, with iodine [40, 41, 42].
Analysing the results of our study, it was observed that green kale 'Oldenbor F1' (control and biofortified with 8-OH-7-I-5QSA) contained more protein than kale (control and biofortified with 8-OH-7-I-5QSA) of the red cultivar 'Redbor F1'. Green 'Oldenbor F1' control kale had 32.98 g·100 g d.m. -1 of protein and biofortified 8-OH-7-I-5QSA had 30.74 g·100 g d.m. -1, respectively. In contrast, red 'Redbor F1' control kale had less protein at 26.03 g·100 g d.m. -1 and biofortified 8-OH-7-I-5QSA at 25.71 g·100 g d.m.-1, respectively. Slightly lower results were obtained in the work of Prade et al. [43] where the protein content of kale leaves of the cultivar Brassica oleracea var. sabellica L. was 150 g·kg d.m.-1. In the work, Pitura and Jarosz [44] showed that the total protein content in the dry matter of kale of another medium-high cultivar (Brassica oleracea var. acephala L.) ranged from 243.7 to 350.6 g·kg d.m-1. In all cases, the values did not differ significantly, but in general the protein content of the plant is dependent on the crop year, harvest date and cultivar [45].
The total fat content of green kale of the control and biofortified version of the 'Oldenbor F1' cultivar was higher than that of red kale of the control and biofortified version of the 'Redbor F1' cultivar. Kale, like most vegetables, has a low total fat content. In our study. the fat content was in the range of 3.16-5.46 g·100 g d.m.-1. According to Satheesh and Fanta [28], the total fat content of kale (Brassica oleracea var. acephala L.) was higher at 11.8 g·100 g d.m.-1. The differences in fat content, as with total protein content, may be a result of crop year, harvest date and cultivar [45].
Significant differences in digestible carbohydrates were observed between the two cultivars. The green cultivar 'Oldenbor F1' had more digestible carbohydrates than the red cultivar 'Redbor F1'. For digestible carbohydrates, we noticed significant changes after biofortification with 8-OH-7-I-5QSA. For both cultivars of kale, 'Oldenbor F1' and 'Redbor F1', the biofortification process with 8-OH-7-I-5QSA increased the digestible carbohydrate content compared to the control, while increasing the nutritional value. Overall, carbohydrate content ranged from 15.53-20.39 g·100 g d.m.-1.
Our study showed that kale of the red cultivar 'Redbor F1' had a higher dietary fibre content compared to the green cultivar 'Oldenbor F1'. Dietary fibre is important in the prevention of many diseases. Higher intake has a positive effect on improving insulin sensitivity, modulating the secretion of certain gut hormones and influencing various metabolic and inflammatory markers associated with the metabolic syndrome [46]. This makes the red cultivar 'Redbor F1' more attractive to consumers in this respect.
The ash content of 'Redbor F1' kale was higher than 'Oldenbor F1' kale. It was observed that the biofortification process with 8-OH-7-I-5QSA reduced the ash content of both cultivars of kale 'Oldendbor F1' and 'Redbor F1'.
Feeding by Wistar rats of diets supplemented with non-biofortified (CO, CR) and biofortified 8-OH-7-I-5QSA (BO, BR) kale of both 'Oldenbor F1' and 'Redbor F1' cultivars for over 8 weeks resulted in significantly lower animal weight gain values compared to the control diet AIN-93G (C) – without added kale. Similarly, the feed efficiency ratio (FER) of rats fed diets with non-biofortified kale (CO, CR) and kale biofortified with 8-OH-7-I-5QSA (BO, BR) was lower compared to the AIN-93G (C) control diet without kale, but significantly (P ≤ 0.05) only for the BO and CR groups. Adding freeze-dried kale to the diets reduced the excessive weight gain of the test individuals. This may be due to the presence of high amounts of dietary fibre and many other bioactive compounds (polyphenols, glucosinolates) found in kale. Although we know that in our study the visceral fat content did not differ significantly in any rat study group, the highest value was observed in the AIN-93G (C) group at 4.39 g. Bioactive compounds reduce the absorption of lipids from the gastrointestinal tract and thus affect lower body weight gain in Wistar rats. In a study by Kopeć et al. [33], by Wistar rats of a diet enriched with – biofortified lettuce in the form of KI did not affect the weight gain values of individuals and the feed efficiency ratio FER. However, a study by Piątkowska et al. [47] showed that of KI-enriched cooked carrots by Wistar rats resulted in higher weight gain of individuals, compared to the AIN-93G diet. In a study by Rakoczy et al. [48], fed of lettuce biofortified with KI did not affect body weight gain or feed conversion ratio FER.
Diets with added kale non- and -biofortified with 8-OH-7-I-5QSA of both cultivars, ‘Oldenbor F1’ and ‘Redbor F1’, had a significant effect on lower liver weight. Significant reductions in kidney and heart weight were observed in a diet supplemented with non-biofortified 'Redbor F1' kale. The use of iodoquinolines influenced lower values of total body weight in the rats and thus the weight of individual organs. It can be suggested that the presence of various bioactive compounds in kale, especially polyphenolic compounds and dietary fibre, may result in lower absorption of crude fat from the gastrointestinal tract, which may have influenced the lower weight of the above organs. In a study by Kopeć et al. [33], heart and kidney weights in rats after feeding lettuce biofortified with KI, did not change. However, liver weight was significantly higher (P ≤ 0.05) in rats fed the AIN-93G control diet without added lettuce, compared to the liver of rats fed the diet with KI – biofortified lettuce. In a study by Rakoczy et al. [48], the addition of non- and -biofortified lettuce (KI) to the diet did not affect the kidney, liver, heart or femoral muscle weights of rats. In a study by Piątkowska et. al. [47], the feeding of KI-biofortified raw and cooked carrots influenced higher liver weight. In the same study, kidney weight was not affected by the different dietary treatments and the highest heart weight was found in groups of rats fed the AIN-93G (C) diet compared to the other experimental groups. It can be postulated that, depending on the experiment, the used dietary treatments affected the weight of individual organs differently. This is influenced by several factors. e.g. the type of vegetable used in the feed, the content of bioactive substances in the feed, the level and the form of iodine or the growing conditions.
The thyroid gland weight of rats fed diets with added kale (CO, BO, CR) was similar compared to the AIN-93G control diet - without added kale (C). Only the thyroid gland weight of rats with added kale BR was significantly higher compared to the AIN-93G control diet and other groups. The increase or decrease in organ weight depending on the provision of adequate iodine with the diet depends on several factors. For example, in a study by Sherrer et al. [49] rats were given 0, 1, 3, 10 and 100 mg·l-1 of iodine or iodide (in the form of Nal) in their drinking water for 100 days. Thyroid gland weight in male rats increased significantly with increasing iodide concentration in the water. In contrast, thyroid gland weight in female rats decreased after the highest dose of iodide was administered. The results of this study indicate that iodine and iodide affect thyroid gland weight in fundamentally different ways in individuals of different sexes. In our study, the iodine level was the same in each experimental group. Thus, it was not the iodine dose that influenced the increase in organ weight. Perhaps the differences were due to the origin of the iodine. The AIN-93G diet used iodine from a mineral mixture, the CO, CR diets used iodine found in green and red kale of natural origin and iodine of organic origin (BO, BR) after hydroponic application of 8-OH-7-I-5QSA. In addition, the increase in organ size may have been influenced by components found in kale e.g. goitrogens (goitrogenic substances). These are anti-nutritional compounds naturally occurring in foods (cabbage, broccoli, kale, etc.) that reduce the bioavailability of iodine from food or interfere with the absorption of iodine by the thyroid gland and consequently the production of its hormones. The pituitary gland, in response to a decrease in the concentration of hormones produced by the thyroid gland. releases thyroid-stimulating hormone (TSH), which causes excessive proliferation of thyroid gland tissue, ultimately leading to goitre [50].
One of the diagnostic methods for iodine deficiency (ID) used in this study was the determination of urinary iodine concentration. The assessment of human iodine nutrition is based on urinary iodine excretion [51] whereas faecal iodine concentration is not relevant for the assessment of iodine nutrition. However, iodine nutrition in rats can be assessed based on iodine content in urine, faeces and organs. Kirchgessner, He & Windisch [52] observed that iodine concentrations in urine, faeces and organs of rats increased with increasing iodine content in the diet.
The highest urinary iodine excretion was measured at week eight in groups fed diets supplemented with 8-OH-7-I-5QSA biofortified kale 'Oldenbor F1' (BO) and 'Redbor F1' (BR) compared to the other experimental groups (C, CO, CR). A different situation was observed in the faecal iodine content of the Wistar rats tested. In general, no significant changes were shown in faecal iodine content at the beginning and the end of the experiment. However, it is noteworthy that there was a trend towards higher faecal iodine concentrations after 8 weeks in the diets with kale biofortified with 8-OH-7-I-5QSA (BO, BR). It is known that iodine is absorbed in the stomach and duodenum and removed by the kidneys and thyroid gland. In our experiment, significantly higher iodine concentrations after 8 weeks of the experiment were found in the kidneys of rats fed a diet containing biofortified 8-OH-7-I-5QSA kale of the 'Oldenbor F1' as well as 'Redbor F1' cultivars compared to the AIN-93G (C) and CO and CR diets. Only for the liver of the BO group, there was a tendency for an increase in iodine concentration - (P ≥ 0.05). Increased iodine accumulation in the kidney and liver following diets with KI-biofortified vegetable additives is confirmed by studies by Kopeć et al. [33], Piatkowska et. al. [47], Rakoczy et al. [48]. When iodine in the diet is present as I- , it is rapidly and efficiently absorbed in the gastrointestinal tract [53]. Our study showed that iodine contained in biofortified kale was more bioavailable. Rats fed diets supplemented with biofortified 8-OH-7-I-5QSA kale (BO, BR) accumulated more iodine in selected organs e.g. kidneys. In our study, iodine doses were equal in each experimental group, confirming that the high bioavailability of the element in tissues is a result of the origin of the iodine, i.e. the organic form, and not the excessive amount of iodine supplied with the diet. Despite the increased excretion of iodine with urine after 8 weeks of the experiment, further high tissue saturation with iodine was achieved. It can, therefore, be suggested that the accumulation of iodine in the organs is probably to protect the animals from the potential occurrence of ID in food. Furthermore, in the groups with low dietary iodine bioavailability C and CO, CR urinary and faecal iodine excretion was lower after 8 weeks to protect the organism from iodine deficiency.
The diagnosis of ID is based on the analysis of urinary iodine concentration, TSH concentration and the development of thyroid gland goitre [51]. Two methods of ID diagnosis were used in this study. The first was the analysis of urinary iodine levels in Wistar rats, which has been discussed above. The second method was the examination of thyrotropic hormone, which is a biochemical marker of thyroid gland function and its hormones. TSH increases iodine uptake by the thyroid gland and stimulates the secretion of the hormones thyroxine (T4) and triiodothyronine (T3) into the blood [54]. Elevated TSH levels may be indicative of impaired T4 and T3 synthesis and insufficient dietary iodine supply. The highest thyrotropic hormone (TSH) concentrations were observed in the experimental group the AIN-93G (C) diet. The lowest TSH concentrations were observed in the experimental groups of rats that fed a diet with control green kale 'Oldenbor F1' (CO) and biofortified 8-OH-7-I-5QSA kale (BO). Feeding of control (CR) and biofortified (BR) 'Redbor F1' kale by Wistar rats resulted in slightly higher TSH hormone concentrations compared to 'Oldenbor F1', but not higher than AIN-93G (C). This indicates that iodine from non-biofortified and biofortified 8-OH-7-I-5QSA kale 'Oldenbor F1' and 'Redbor F1' cultivars was bioavailable to the rats and consequently was utilised for thyroid gland hormone synthesis. In contrast, elevated serum TSH levels in rats from the AIN-93G (C) – no kale supplementation groups, indicated insufficient iodine levels in this diet, compared to the rats' requirement for this trace element. Thyroxine (T4) and triiodothyronine (T3) hormone concentrations were not significantly different between all experimental groups of rats tested. T3 is produced by deiodination of T4. The enzyme hepatic deiodinase type 1 is involved in this metabolic pathway. Furthermore, hepatic deiodinase type 1 is responsible for the serum T3 content [55, 56, 57]. It can be suggested that serum T4 concentrations in all groups of rats were not increased because T4 was used to produce T3.
The feed of different types of diets by Wistar rats did affect the change (P ≥ 0.05) in the activity of the liver enzyme aspartate aminotransferase (AST) after 8 weeks of experimentation (Table 5). After feeding BR kale, a decrease in AST enzyme activity, or a tendency for BO to decrease, was observed. The feed of the kale cultivar ‘Redbor F1’, reduces AST enzyme concentrations the most. In the liver enzyme alanine aminotransferase (ALT), fed a diet with kale non-biofortified (CR) and -biofortified with 8-OH-7-I-5QSA (BR) 'Redbor F1', caused a significant reduction or a tendency for a lower, in the case of use of 'Oldenbor F1' kale enriched in 8-OH-7-I-5QSA (BO), activity of the enzyme. In a study by Kopeć et. al. [33] using lettuce biofortified with KI, enzyme concentrations did not change. However, in the study by Piątkowska et al. [47], ALT enzyme concentrations increased in the experimental groups where rats fed cooked and KI-biofortified cooked carrots. It may be suggested that inorganic forms of iodine may influence increased liver activity and thus increase ALT enzyme activity. In contrast, in our experiment, the organic form of iodine in kale appears to be safer, as it caused a decrease in ALT activity. In addition, diets with red kale reduced ALT enzyme activity; this may be related to the high content of bioactive compounds with liver-protective effects.
Total and direct bilirubin values were the highest in the group of rats fed the AIN-93G control diet (C). The other experimental groups fed non-biofortified (CO) and biofortified (BO, BR) kale had significantly lower or a tendency for lower, in the case of CR, total and direct bilirubin values. Higher total and direct bilirubin values in group C may be indicative of liver problems (e.g. hepatitis, cirrhosis), blood diseases leading to increased breakdown of red blood cells (haemolysis), or bile duct problems (e.g. cholelithiasis). The addition of kale to the other CO, CR, BO, BR diets showed a health-promoting effect, through the polyphenols, glucosinolates, dietary fibre present, which had a protective effect on liver function and thus reduced bilirubin absorption in the gastrointestinal tract [28].
The serum uric acid values of Wistar rats in our experiment were not affected by the diets. However, there was a trend towards lower uric acid concentrations in Wistar rats after feeding non- and biofortified with 8-OH-7-I-5QSA kale in both cultivars. Moreover, the lowest uric acid concentrations were in the groups where iodoquinoline was added. High levels of uric acid cause hyperuricaemia. Its higher levels sustained over a long period of time can result in the formation of sodium urate crystals and the development of diseases such as gout, kidney stones or cardiovascular conditions including hypertension [58].
Changes in the HDL fraction cholesterol were observed. The highest HDL value was observed in rats fed AIN-93G (C) and in the CO, BO and BR groups. Furthermore, it is noteworthy that feeding of CO, BO, BR diets showed a tendency for a decrease in HDL concentrations, relative to the AIN-93G control. Significantly, the lowest HDL cholesterol concentration value was obtained by the research group feeding non-biofortified 'Redbor F1' kale. This could be explained by the presence of fibre and other biologically active components. However, total cholesterol (TC) and LDL + VLDL were not affected by the different dietary treatments. Triglyceride concentrations had only a tendency for the highest concentration in the group of rats fed AIN-93G (C), compared to the other experimental groups (CO, BO, CR, BR). This may be explained by the presence of bioactive compounds in kale, which improve fat metabolism and thus faster excretion of fats with the gastrointestinal tract [59].
Antioxidant activity was determined in the serum of test rats by glutathione reductase (RG), total antioxidant status (TAS), and thiobarbituric acid reaction lipid peroxidation products (TBARs) – Table 5. After 8 weeks of the experiment, glutathione reductase levels were not significantly different between the experimental groups.
Determination of TAS in rat serum showed that feeding of 'Oldenbor F1' kale (BO) and ‘Redbor F1’ kale (BR) biofortified with 8-OH-7-I-5QSA significantly increased total antioxidant status. The increase in total antioxidant status may be due to the presence of numerous bioactive substances naturally present in kale. Furthermore, we can observe that the use of an iodoquinoline additive in the feed of rats also increased TAS concentrations. Iodoquinolines are a small number of quinoline derivatives that have been attributed with beneficial medical effects, including antibacterial, antifungal, antiviral, anti-inflammatory and anticancer effects [60]. The presence of these compounds may have a positive effect on increasing serum TAS concentrations in rats, but this requires further research.
Thiobarbituric acid reactive substances (TBARs) concentrations expressed in nmol malondialdehyde (MDA)·ml-1 were the highest in the AIN-93G control group (C). Research groups of rats which fed diets supplemented with red kale of the ‘Redbor F1’ cultivar (CR, BR) yielded the lowest values of serum MDA concentrations. Moreover, adding kale of the ‘Oldenbor F1’ cultivar to the feed (CO, BO) caused a tendency for a reduction in MDA concentrations. MDA, which is an end product of lipid peroxidation, is commonly used as a marker of oxidative stress [61]. Kale has naturally occurring polyphenols with antioxidant activity. Polyphenols have been shown to have the ability to inhibit the increase in lipid peroxidation levels in laboratory animals [62]. It can be assumed that the effect of reducing serum MDA levels in rats fed non- and biofortified kale was due to the presence of polyphenols and iodoquinolines. Kopeć et al. [33] obtained similar results to ours. The addition of iodine-fortified lettuce (KI) and control lettuce significantly reduced serum TBARs in rats. Different results were presented by Piątkowska et al. [47], where the addition of control and cooked KI-biofortified carrots increased MDA concentrations. In the study by Rakoczy et al. [48], the use of KI-biofortified lettuce did not affect serum and liver MDA levels in rats.
The iodoquinoline biofortified kale cultivars, 'Oldenbor F1' and 'Redbor F1', can be considered as potential safe sources of iodine in the daily diet, preventing deficiencies of this trace element in different populations.
5. Conclusions
In conclusion, rats fed diets containing kale biofortified with iodine had higher iodine content in urine and kidneys, this proves the effective availability of iodine from kale biofortified with iodoquinolines, despite the presence of substances that hinder the absorption of iodine (goitrogens).
In rats feeding diets supplemented with ‘Oldenbor F1’ and ‘Redbor F1’ kale non- and -biofortified with 8-OH-7-I-5QSA, there were numerous health benefits through a tendency for a lower concentration of TSH, triglycerides, TBARs, total bilirubin, direct bilirubin, uric acid, aspartate aminotransferase (AST) and alanine aminotransferase (ALT) in the Wistar rats. What’s more, dietary intake of 'Oldenbor F1' and 'Redbor F1' kale biofortified with 8-OH-7-I-5QSA significantly increased total antioxidant status (TAS).
Biofortified kale can be considered as a potential source of iodine in the daily diet, but more in vivo studies need to be conducted, and an initiative to study the effect of human of vegetables biofortified with iodoquinolines should be undertaken. The use of iodine-enriched kale diets has demonstrated effective bioavailability and absorption in the gastrointestinal tract of experimental rats. With the increasing popularity of kale as a superfood in recent years, there is a potential to enrich the daily diet with iodine by biofortified vegetables such as kale enriched with 8-OH-7-I-5QSA. The results of this research may lead to new health approaches that focus on increasing iodine availability, especially in regions with low levels of this element in the environment. Such initiatives may not only improve overall health but also reduce the risk of iodine deficiency-related diseases such as thyroid gland disorders, hypertension, obesity and type 2 diabetes.
6. Patents
The method of biofortification of vegetables in iodine cultivated using traditional, soilless and hydroponic method and the use of 8-hydroxy-7-iodo-5-quinolinesulfonic acid for biofortification of vegetables with iodine. These are patent application number P.443218 (Polish Patent Office; December 21. 2022).
Author Contributions
Conceptualization, A.Kp., and A.K.; validation, S.S., A.K.; formal analysis, J.K.; A.Kp.; A.K.; investigation, J.K.; E.P.; A.Kp.; S.S.; writing—original draft preparation, J.K.; writing—review and editing, E.P.; A.Kp.; S.S.; T.L.; A.K.; visualization, J.K.; supervision, A.K; S.S.; project administration, A.K.; funding acquisition, A.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was financed by the National Science Center, Poland (grant no. UMO-2020/37/B/NZ9/02710) “The use of iodoquinolines for biostimulation and biofortification of selected vegetables in iodine and investigation of their anticancer properties in the in vitro breast cancer model’’.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Grasberger, H.; Refetoff, S. Genetic causes of congenital hypothyroidism due to dyshormonogenesis. Current Opinion in Pediatrics, 2011, 23(4): 421-428. [CrossRef]
- Zimmermann, M.B.; Jooste, P.L.; Pandav, C.S. Iodine-deficiency disorders. Lancet, 2008, 4:372(9645):1251-1262.
- Andersson, M.; Takkouche, B.; Egli, I.; Allen, H.E.; Benoist, B. Current global iodine status and progress over the last decade towards the elimination of iodine deficiency. Bull World Health Organization, 2005, 83(7), 518-25. PMCID: PMC2626287.
- O’Donnell, M.; Mente, A.; Alderman, M.H.; Brady, A.J.B.; Diaz, R.; Gupta, R.; et al. Salt and cardiovascular disease: insufficient evidence to recommend low sodium intake. European Heart Journal, 2020, 41: 3363–73. [CrossRef]
- World Health Organization. People in the WHO European Region at greater risk of iodine deficiency due to changing diets. Available online: https://www.who.int/azerbaijan/news/item/28-06-2024-people-in-the-who-european-region-at-greater-risk-of-iodine-deficiency-due-to-changing-diets (accessed on 24 June 2024).
- Meharg, A. Marschner’s Mineral Nutrition of Higher Plants. 3rd edition. Edited by P. Marschner. Amsterdam, Netherlands: Elsevier/Academic Press, 2011, pp. 684. Experimental Agriculture, 2012, 48(2): 305-305. [CrossRef]
- Kiferle, C.; Gonzali, S.; Holwerda, H.T.; Real Ibaceta, R.; Perata, P. Tomato fruits: a good target for iodine biofortification. Frontiers in Plant Science, 2013, 4:205. [CrossRef]
- Incrocci, L.; Carmassi, G.; Maggini, R.; Poli, C.; Saidov, D.; Tamburini, C.; et al. Iodine accumulation and tolerance in sweet basil (Ocimum basilicum L.) with green or purple leaves grown in floating system technique. Frontiers in Plant Science, 2019, 10:1494. [CrossRef]
- Sabatino, L.; D’Anna, F.; Iapichino, G.; Moncada, A.; D’Anna, E.; De Pasquale, C. Interactive effects of genotype and molybdenum supply on yield and overall fruit quality of tomato. Frontiers in Plant Science, 2019, 9: 1922. [CrossRef]
- Sabatino, L.; Ntatsi, G.; Iapichino, G.; D’Anna, F.; De Pasquale, C. Effect of selenium enrichment and type of application on yield, functional quality and mineral composition of curly endive grown in a hydroponic system. Agronomy, 2019, 9:207. [CrossRef]
- Sabatino, L.; Consentino, B.B.; Rouphael, Y.; De Pasquale, C.; Iapichino, G.; D’Anna, F, et al. Protein hydrolysates and mo-biofortification interactively modulate plant performance and quality of ‘canasta’ lettuce grown in a protected environment. Agronomy, 2021, 11:1023. [CrossRef]
- Sabatino, L.; La Bella, S.; Ntatsi, G.; Iapichino, G.; D’Anna, F.; De Pasquale, C.; et al. Selenium biofortification and grafting modulate plant performance and functional fures of cherry tomato grown in a soilless system. Scientia Horticulturae, 2021, 285: 110095. [CrossRef]
- Sabatino. L.; Di Gaudio, F.; Consentino, B.B.; Rouphael, Y.; El-Nakhel, C.; La Bella, S.; et al. Iodine biofortification counters micronutrient deficiency and improve functional quality of open field grown curly endive. Horticulturae, 2021, 7(3): 58. [CrossRef]
- La Bella, S.; Consentino, B.B.; Rouphael, Y.; Ntatsi, G.; De Pasquale, C.; Iapichino, G.; et al. Impact of ecklonia maxima seaweed extract and mo foliar trments on biofortification, spinach yield, quality and NUE. Plants, 2021, 10(6): 1139. [CrossRef]
- Puccinelli, M.; Landi, M.; Maggini, R.; Pardossi, A.; Incrocci, L. Iodine biofortification of sweet basil and lettuce grown in two hydroponic systems. Scientia Horticulturae, 2021, 276: 109783. [CrossRef]
- Garg, M. et al. Biofortified crops generated by breeding, agronomy, and transgenic approaches are improving lives of millions of people around the world. Frontiers in Nutrition, 2018, 5:12. [CrossRef]
- Bouis, H.E. et al. Biofortification - a sustainable agricultural strategy for reducing micronutrient malnutrition in the global south. Crop Science, 2010, 50:20-32. [CrossRef]
- Gómez-Galera, S.; Rojas, E.; Sudhakar, D. et al. Critical evaluation of strategies for mineral fortification of staple food crops. Transgenic Research, 2010, 19, 165–180. [CrossRef]
- Basharat, A.; Asghar, A.; Muhammad, T.; Shafaqat, A. Growth, seed yield and quality of mungbean as influenced by foliar application of iron sulphate. Pakistan Journal of Life and Social Sciences, 2014, 12(1): 20-25.
- Smoleń, S.; Sady, W. Influence of iodine form and application method on the effectiveness of iodine biofortification, nitrogen metabolism as well as the content of mineral nutrients and heavy metals in spinach plants (Spinacia oleracea L.). Scientia Horticulturae, 2012, 143, 176–183. [CrossRef]
- Medrano-Macias, J.; Leija-Martínez, P.; González-Morales, S.; Juárez-Maldonado, A.; & Benavides Mendoza, A. Use of iodine to biofortify and promote growth and stress tolerance in crops. Frontiers in Plant Science, 2016, 7:1146. [CrossRef]
- Golob, A.; Novak, T.; Maršić, K. N.; Šircelj, H.; Stibilj, V.; Jerše, A., & et al. Biofortification with selenium and iodine changes morphological properties of Brassica oleracea L. var. gongylodes) and increases their contents in tubers. Plant Physiology and Biochemistry, 2020, 150, 234–243. [CrossRef]
- Sularz, O.; Smoleń, S.; Koronowicz, A.; Kowalska, I.; Leszczyńska, T. Chemical composition of lettuce (Lactuca sativa L.) biofortified with iodine by KIO3 , 5-Iodo-, 3.5-Diiodosalicylic acid in a Hydrophonic Cultivation. Agronomy, 2020, 10 (7), 1022. [CrossRef]
- Smoleń, S.; Kowalska, I.; Skoczylas, Ł.; Tabaszewska, M.; Pitala, J.; Mrożek, J.; Kovacik, P. Effectiveness of enriching lettuce with iodine using 5-iodosalicylic and 3,5-diiodosalicylic acids and the chemical composition of plants depending on the type of soil in a pot experiment. Food Chemistry, 2022, 382, 132347. [CrossRef]
- Halka, M.; Smoleń, S.; Czernicka, M.; Klimek-Chodacka, M.; Pitala, J.; Tutaj, K. Iodine biofortification through expression of HMT, SAMT and S3H genes in Solanum lycopersicum L. Plant physiology and biochemistry, 2019, 144, 35–48. [CrossRef]
- Halka, M.; Smoleń, S.; Ledwożyw-Smoleń, I.; Sady, W. Iodosalicylates and iodobenzoates supplied to tomato plants affect the antioxidative and sugar metabolism differently than potassium iodide. Folia Horticulturae, 2019, 31 (2), 385–400. [CrossRef]
- Matada, B.S.; Pattanashettar, R.; Yernale, N.G. A comprehensive review on the biological interest of quinoline and its derivatives. Bioorganic & Medicinal Chemistry, 2021, 32, 115973. [CrossRef]
- Satheesh, N. & Fanta, S. Kale: Review on nutritional composition, bio-active compounds, anti-nutritional factors, health beneficial properties and value-added products. Cogent Food & Agriculture, 2020, 6. 1-32. [CrossRef]
- Šamec, D.; Urlić, B.; Salopek-Sondi, B. Kale (Brassica oleracea var. acephala) as a superfood: review of the scientific dvidence behind the statement. Critical Reviews in Food Science and Nutrition, 2019, 59 (15), 2411–2422. [CrossRef]
- Total Dietary Fiber—Assay Procedure—Megazyme. Available online: https://www.megazyme.com/documents/Assay (accessed on 1 February 2024).
- AOAC. Official methods of analysis (18th ed.). Gaithersburg: Association of Official Analytical Chemists International, 2006.
- Standard Operation Procedure. Ref. Ares. 2020; p. 1848056-31/03/2020. Available online: https://ec.europa.eu/research/participants/documents/downloadPublicdocumentIds=080166e5cd8d669b&appId=PPGMS (accessed on 1 February 2024).
- Kopeć, A.; Piątkowska, E.; Bieżanowska-Kopeć, R.; Pysz, M.; Koronowicz, A.; Kapusta-Duch, J.; Smoleń, S.; Rakoczy, R.; Skoczylas, Ł.; Leszczyńska, T.; Ledwożyw-Smoleń, I. Effect of lettuce biofortified with iodine by soil fertilization on iodine concentration in various tissues and selected biochemical parameters in serum of Wistar rats. Journal of Functional Foods, 2015, 14: 479-486. [CrossRef]
- Smoleń, S.; Kowalska, I.; Kovácik, P.; Sady, W.; Grzanka, M.; Kutman, U.B. Changes in the Chemical Composition of Six Lettuce Cultivars (Lactuca sativa L.) in Response to Biofortification with Iodine and Selenium Combined with Salicylic Acid Application. Agronomy, 2019, 9, 660. [CrossRef]
- PN-EN: 15111:2008. Foodstuffs—Determination of Trace Elements—Determination of Iodine Content by ICP-MS (Inductively Coupled Plasma Mass Spectrometry). Polish Committee for Standardisation: Warsaw, Poland, 2008. Available online: https://sklep.pkn.pl/pn-en-15111-2008p.html (accessed on 22 January 2024).
- Reeves, P. G. Components of the AIN-93 diets as improvements in the AIN-76A diet. Journal of Nutrition, 1997, 127, 838–841.
- Friedewald, W. T.; Fredrick, D. S.; & Levy, R. I. Estimation of concentration of low-density lipoprotein cholesterol in plasma, without use of preparative ultracentrifuge. Clinical Chemistry, 1972, 18(6), 499–502.
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Analytical Biochemistry, 1979, 95(2): 351-8. [CrossRef]
- Ayaz, F.A.; Glew, R.H.; Millson, M.; Huang, H.S.; Chuang, L.T.; Sanz, C.; Hayirlioglu-Ayaz, S. Nutrient contents of kale (Brassica oleraceae L. var. acephala DC). Food Chemistry, 2006, 96(4), 572–579. [CrossRef]
- Dyląg, A.; Smoleń, S.; Wisła-Świder, A.; Kowalska, I.; Sularz, O.; Krzemińska, J.; Pitala, J.; & Koronowicz, A. Evaluation of the chemical composition and nutritional value of lettuce (Lactuca sativa L.) biofortified in hydroponics with iodine in the form of iodoquinolines. Frontiers in plant science, 2023, 14, 1288773. [CrossRef]
- Krzemińska, J.; Smoleń, S.; Kowalska, I.; Pitala, J.; Sularz, O.; Koronowicz, A. Effect of Biofortification with Iodine by 8-Hydroxy-7-iodo-5-quinolinesulfonic Acid and 5-Chloro-7-iodo-8-quinolinol on the Chemical Composition and Antioxidant Properties of Potato Tubers (Solanum tuberosum L.) in a Pot Experiment. Applied Science, 2023, 13, 4659. [CrossRef]
- Krawczyk, K.; Smoleń, S.; Wisła-Świder, A.; Kowalska, I.; Kiełbasa, D.; Pitala, J.; Krzemińska, J.; Waśniowska, J.; Koronowicz, A. Kale (Brassica oleracea L. var. sabellica) biofortified with iodoquinolines: Effectiveness of enriching with iodine and influence on chemical composition. Scientia Horticulturae, 2024, 323, 112519. [CrossRef]
- Prade, T.; Muneer, F.; Berndtsson, E.; Nynäs, A.L.S.; Svensson. S.E.; Newson, W.R.; Johansson, E. Protein fractionation of broccoli (Brassica oleracea, var. Italica) and kale (Brassica oleracea, var. Sabellica) residual leaves — A pre-feasibility assessment and evaluation of fraction phenol and fibre content. Food and Bioproducts Processing, 2021, 130, 229-243. [CrossRef]
- Pitura, K.; Jarosz, Z. Chemical composition and biological value of kale depending on the varied mineral fertilization. Agronomy Science, 2020, 78 (4), 97-107. https://orcid.org/0000-0002-1561-4457.
- Korus, A. Kale: a Valuable Brassica Vegetable. Part II. The nutritional Value of kale. Fermentation and Fruit and Vegetable Industry, 2015, 2. [CrossRef]
- Weickert, M.O.; Pfeiffer, A.F.H. Metabolic Effects of Dietary Fiber and Prevention of Diabetes. The Journal of Nutrition, 2008, 138(3): 439–442. [CrossRef]
- Piątkowska, E.; Kopeć, A.; Bieżanowska-Kopeć, R.; Pysz, M.; Kapusta-Duch, J.; Koronowicz, A., et al. The Impact of Carrot Enriched in Iodine through Soil Fertilization on Iodine Concentration and Selected Biochemical Parameters in Wistar Rats. PLoS ONE, 2016, 11(4): e0152680. [CrossRef]
- Rakoczy, R.; Kopeć, A.; Piątkowska, E.; Smoleń, S.; Skoczylas, Ł.; Leszczyńska, T.; Sady, W. The Iodine Content in Urine, Faeces and Selected Organs of Rats Fed Lettuce Biofortified with Iodine Through Foliar Application. Biological Trace Element Research, 2016, 174, 347–355. [CrossRef]
- Sherer, T.T.; Thrall, K.D.; Bull, R.J. Comparison of toxicity induced by iodine and iodide in male and female rats. Journal of Toxicology and Environmental Health, 1991, 32 (1), 89–101. [CrossRef]
- Kanno, J.; Matsuoka, C.; Furuta, K. et al. Tumor Promoting Effect of Goitrogens on the Rat Thyroid. Toxicologic Pathology, 1990, 18(2): 239-246. [CrossRef]
- WHO/UNICEF. Iodine deficiency in Europe: a continuing public health problem. Geneva, Switzerland, 2007. ISBN ISBN: 9789241593960.
- Kirchgessner, M.; He, J.; Windisch, W. Homeostatic adjustments of iodine metabolism and tissue iodine to widely varying iodine supply in 125I labeled rats. Journal of Animal Physiology and Animal Nutrition, 1999, 82(5): 238–250. [CrossRef]
- Winger, R.J.; König, J.; House, D.A. Technological issues related to food enrichment with iodine. Trendy Food Sci Tech, 2008, 19:94–101.
- Cai, J.; Fang, Y.; Jing, D.; Xu, S.; Ming, J.; Gau, B.; Shen, H.; Zhang, R.; Ji, Q. Reference intervals of thyroid hormones in a previously iodine-deficient but presently more than adequate area of Western China: a population-based survey. Endocrine Journal, 2016, 63 (4), 381-388. [CrossRef]
- Konturek, S. Human physiology. Gastrointestinal tract and endocrine glands (Vol. 5). Kraków, Poland: Scientific Publishing House: DWN (in Polish), 1994.
- Schneider, M. J.; Fiering, S. N.; Thai, B.; Wu, S. Y.; St Germain, E.; Parlow, A. F.; St Germain, D. L.; & Galton, V. A. Targeted disruption of the type 1 selenodeiodinase Gene (Dio1) results in marked changes in thyroid hormone economy in mice. Endocrinology, 2006, 147(1), 580–589. [CrossRef]
- Bianco, A. C.; Kim, B. W. Deiodinases: Implications of the local control of thyroid hormone action. The Journal of Clinical Investigation, 2014, 116,(10), 2571–2579. [CrossRef]
- George, C.; Leslie, S.W.; Minter, D.A. Hyperuricemia. In: Stat Pearls, Treasure Island (FL): Stat Pearls Publishing, 2023. Available online: https://www.ncbi.nlm.nih.gov/books/NBK459218/ (accessed on 25 May 2024).
- Pal, S.; Khossousi, A.; Binns, C.; Dhaliwal, S.; Ellis, V. The effect of a fibre supplement compared to a healthy diet on body composition, lipids, glucose, insulin and other metabolic syndrome risk factors in overweight and obese individuals. British Journal of Nutrition, 2011, 105(1): 90-100. [CrossRef]
- Pippi, B.; Reginatto, P.; Monte, M.; Rosa, G.; Zafaneli Bergamo, V.; Flores Dalla Lana, D.; Lettieri Teixeira, M.; Lopardi Franco, L.; Alves Ricardo, J.; Andrade Saulo, F.; Meneghello Fuentefria, A. Evaluation of 8-Hydroxyquinoline Derivatives as Hits for Antifungal Drug Design. Medical Mycology, 2011, 55(7): 763–773. [CrossRef]
- Tkaczewska, J.; Jamróz, E.; Piątkowska, E.; Borczak, B.; Kapusta-Duch, J.; Morawska, M. Furcellaran-Coated Microcapsules as Carriers of Cyprinus carpio Skin-Derived Antioxidant Hydrolysate: An In Vitro and In Vivo Study. Nutrients, 2019, 11, 2502. [CrossRef]
- Iwai, K. Antidiabetic and antioxidant effects of polyphenols in brown alga Ecklonia stolonifera in genetically diabetic KK-Ay mice. Plant Foods for Human Nutrition, 2008, 63, 163-169. [CrossRef]
Figure 1.
Concentrations of iodine in urine and feces of Wistar rats. Contents of I in urine (A), contents of I in faeces (B); C - control diet (AIN-93G), CO - diet containing control curly kale 'Oldenbor F1', BO - diet containing biofortified curly kale 'Oldenbor F1', CR - diet containing control curly kale 'Redbor F1', BR - diet containing biofortified curly kale 'Redbor F1’. Means followed by the same letters are not significantly different at P ≥ 0.05 (Duncan’s post-hoc test); bars indicate standard error (n=8). Homogeneous groups refer to two-factor analysis of variance: Factor No.1 type of diet: C, CO, BO, CR, BR; , x Factor No.2 week: I, VIII.
Figure 1.
Concentrations of iodine in urine and feces of Wistar rats. Contents of I in urine (A), contents of I in faeces (B); C - control diet (AIN-93G), CO - diet containing control curly kale 'Oldenbor F1', BO - diet containing biofortified curly kale 'Oldenbor F1', CR - diet containing control curly kale 'Redbor F1', BR - diet containing biofortified curly kale 'Redbor F1’. Means followed by the same letters are not significantly different at P ≥ 0.05 (Duncan’s post-hoc test); bars indicate standard error (n=8). Homogeneous groups refer to two-factor analysis of variance: Factor No.1 type of diet: C, CO, BO, CR, BR; , x Factor No.2 week: I, VIII.
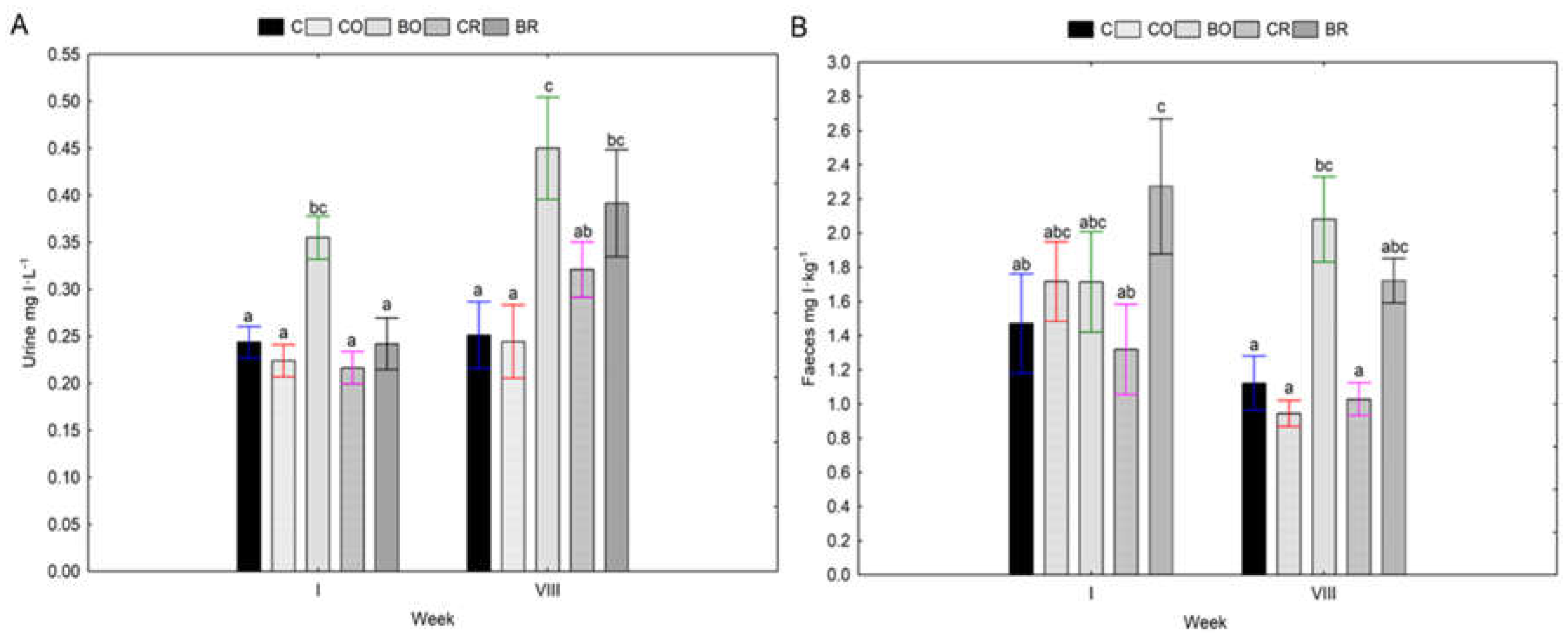

Table 1.
Ingredients of experimental diets composition.
| Ingredient (g·kg-1) | C | CO | BO | CR | BR |
|---|---|---|---|---|---|
| Corn starch | 532.49 | 524.72 | 525.00 | 526.04 | 526.62 |
| Saccharose | 100 | 100 | 100 | 100 | 100 |
| Casein | 200 | 200 | 200 | 200 | 200 |
| Soybean oil | 70 | 70 | 70 | 70 | 70 |
| Fibre | 50 | 47.02 | 47.19 | 46.47 | 47 |
| Vitamin mix a | 10 | 10 | 10 | 10 | 10 |
| Mineral mix a | 35 | 35 | 35 b | 35 | 35 b |
| Choline | 2.5 | 2.5 | 2.5 | 2.5 | 2.5 |
| TBHQ c | 0.014 | 0.014 | 0.014 | 0.014 | 0.014 |
| Biofortified kale d | - | - | 10.3 | - | 8.87 |
| Control kale d | - | 10.75 | - | 9.98 | - |
C - control diet (AIN-93G), CO - diet containing control curly kale 'Oldenbor F1', BO - diet containing biofortified curly kale 'Oldenbor F1', CR - diet containing control curly kale 'Redbor F1', BR - diet containing biofortified curly kale 'Redbor F1’. a according to AIN-93G. b mineral mix without iodine; in these diets the source of iodine was biofortified curly kale in amounts providing an equivalent amount of iodine to the amount contained in AIN-93G. c tert-butylhydroquinone. d freeze-dried curly kale ‘Oldenbor F1’ or ‘Redbor F1’.
Table 2.
Iodine content and basic chemical composition of kale both cultivars ‘Oldenbor F1’ and ‘Redbor F1’ used for preparation of experimental diets (g·100 g d.m.-1).
Table 2.
Iodine content and basic chemical composition of kale both cultivars ‘Oldenbor F1’ and ‘Redbor F1’ used for preparation of experimental diets (g·100 g d.m.-1).
| Control raw kale 'Oldenbor F1’ | Raw biofortified with 8-OH-7-I-5QSA kale 'Oldenbor F1’ | Control raw kale‘Redbor F1’ | Raw biofortified with 8-OH-7-I-5QSA kale ‘Redbor F1’ | |
|---|---|---|---|---|
| Iodine [mg·kg d.m.-1] | 0.18 ± 0.02 a | 2.10 ± 0.04 b | 0.20 ± 0.01 a | 2.43 ± 0.08 c |
| Basic chemical composition | ||||
| Protein | 32.98 ± 0.06 b | 30.74 ± 1.06 b | 26.03 ± 2.31 a | 25.71 ± 0.88 a |
| Crude fat | 5.46 ± 0.13 b | 5.46 ± 0.26 b | 3.24 ± 0.13 a | 3.16 ± 0.15 a |
| Digestible carbohydrates | 17.39 ± 0.77 a | 20.39 ± 0.16 c | 15.53 ± 0.43 b | 18.35 ± 0.86 a |
| Dietary fiber | 29.06 ± 0.82 a | 28.45 ± 0.11 a | 36.39 ± 0.33 c | 34.71 ± 0.81 b |
| Ash | 20.06 ± 0.23 b | 19.21 ± 0.19 a | 21.33 ± 0.09 d | 20.76 ± 0.03 c |
Values in rows with different letters (a, b, c, d) are significantly different, P ≤ 0.05 (Duncan’s post-hoc test); bars indicate standard error (n = 4).
Table 3.
Body gain, fed efficiency ratio (FER) and weight of selected organs in fresh mass.
| Type of diet | C | CO | BO | CR | BR |
|---|---|---|---|---|---|
| Body gain (g) | 302.38 ± 27.43 b | 280.88 ± 26.62 ab | 277.50 ± 17.65 a | 273.38 ± 15.32 a | 293.25 ± 20.10 ab |
| FER* | 0.202 ± 0.02 b | 0.187 ± 0.02 ab | 0.185 ± 0.01 a | 0.182 ± 0.01 a | 0.195 ± 0.01 ab |
| Liver (g) | 16.14 ± 2.17 c | 14.14 ± 1.52 b | 13.04 ± 1.35 ab | 12.32 ± 0.98 a | 13.54 ± 1.80 ab |
| Kidney** (g) | 2.68 ± 0.14 a | 2.47 ± 0.14 ab | 2.55 ± 0.24 ab | 2.29 ± 0.48 b | 2.65 ± 0.24 a |
| Heart (g) | 1.27 ± 0.09 b | 1.18 ± 0.08 ab | 1.22 ± 0.08 ab | 1.14 ± 0.07 a | 1.22 ± 0.08 ab |
| Thyroid gland (g) | 0.23 ± 0.06 a | 0.26 ± 0.03 ab | 0.24 ± 0.04 ab | 0.27 ± 0.03 ab | 0.28 ± 0.03 b |
| Visceral fat (g) | 4.39 ± 0.80 a | 3.88 ± 0.68 a | 4.18 ± 0.77 a | 3.52 ± 1.06 a | 3.77 ± 0.58 a |
Values in rows with different letters (a, b, c) are significantly different, P ≤ 0.05 (Duncan’s post-hoc test); bars indicate standard error (n = 8). * FER - feed efficiency ratio - (body weight gain (g) /diet consumed (g)). ** weight of both kidneys. C - control diet (AIN-93G), CO - diet containing control curly kale 'Oldenbor F1', BO - diet containing biofortified curly kale 'Oldenbor F1', CR - diet containing control curly kale 'Redbor F1', BR - diet containing biofortified curly kale 'Redbor F1’.
Table 4.
Concentration of iodine in selected organs.
| Type of diet | C | CO | BO | CR | BR |
|---|---|---|---|---|---|
| Kidney [mg I·kg d.m.-1] |
0.14 ± 0.01 b | 0.13 ± 0.02 ab | 0.17 ± 0.03 c | 0.11 ± 0.02 a | 0.18 ± 0.04 c |
| Liver [mg I·kg d.m.-1] |
0.12 ± 0.02 ab | 0.11 ± 0.02 a | 0.13 ± 0.01 b | 0.11 ± 0.01 a | 0.12 ± 0.01 ab |
Values in rows with different letters (a, b, c) are significantly different, P ≤ 0.05 (Duncan’s post-hoc test); bars indicate standard error (n = 8). C - control diet (AIN-93G), CO - diet containing control curly kale 'Oldenbor F1', BO - diet containing biofortified curly kale 'Oldenbor F1', CR - diet containing control curly kale 'Redbor F1', BR - diet containing biofortified curly kale 'Redbor F1’.
Table 5.
Selected biochemical parameters in serum of experimental rats.
| Type of diet | C | CO | BO | CR | BR |
|---|---|---|---|---|---|
| Liver panel | |||||
| AST [U·L-1] | 36.11 ± 9.11 a | 33.53 ± 12.39 a | 30.54 ± 10.49 ab | 31.11 ± 5.03 ab | 19.79 ± 8.35 b |
| ALT [U·L-1] | 26.55 ± 7.84 a | 25.09 ± 6.81 a | 23.74 ± 6.72 a | 15.85 ± 4.84 b | 13.93 ± 4.8 b |
| Bilirubin | |||||
| Total bilirubin [µmol·L-1] | 10.38 ± 6.08 b | 3.73 ± 1.84 a | 3.15 ± 3.17 a | 5.34 ± 3.47 a | 4.28 ± 2.79 a |
| Direct bilirubin [µmol·L-1] | 7.59 ± 3.85 b | 3.35 ± 2.40 a | 2.13 ± 1.27 a | 4.53 ± 4.56 ab | 4.27 ± 1.87 a |
| Uric acid | |||||
| Uric acid [µmol·L-1] | 166.04 ± 62.01 a | 132.35 ± 69.70 a | 116.11 ± 67.30 a | 112.50 ± 48.74 a | 108.29 ± 19.76 a |
| Lipid profile | |||||
| TC [mmol·L-1] | 3.01 ± 0.53 a | 2.94 ± 0.22 a | 2.77 ± 0.30 a | 2.64 ± 0.34 a | 2.55 ± 0.65 a |
| HDL [mmol·L-1] | 1.89 ± 0.18 b | 1.71 ± 0.22 ab | 1.76 ± 0.34 ab | 1.54 ± 0.27 a | 1.64 ± 0.17 ab |
| LDL + VLDL [mmol·L-1] | 1.12 ± 0.37 a | 1.23 ± 0.31 a | 1.01 ± 0.42 a | 1.10 ± 0.20 a | 0.91 ± 0.55 a |
| TG [mmol·L-1] | 1.45 ± 0.62 b | 1.35 ± 0.36 ab | 0.99 ± 0.16 a | 1.15 ± 0.44 ab | 1.05 ± 0.22 ab |
| Antioxidant activity | |||||
| Glutathione reductase [U·L-1] | 375.72 ± 205.75 a | 348.31 ± 174.47 a | 444.23 ± 83.52 a | 378.96 ± 148.61 a | 438.01 ± 68.74 a |
| TAS [mmol·L-1] | 0.89 ± 0.20 ab | 0.87 ± 0.08 a | 1.04 ± 0.08 c | 0.99 ± 0.10 abc | 1.02 ± 0.10 bc |
| TBARs [nmol MDA·mL-1] | 554.52 ±27.36 c | 535.31 ± 88.82 bc | 513.88 ± 49.46 abc | 474.93 ± 63.19 ab | 459.47 ± 66.32 a |
| Hormones | |||||
| TSH [ng·mL-1] | 2.18 ± 0.21 b | 2.02 ± 0.13 a | 1.99 ± 0.08 a | 2.10 ± 0.17 ab | 2.07 ± 0.11 ab |
| T3[pg·mL-1] | 3.81 ± 0.19 ab | 3.92 ± 0.15 b | 3.73 ± 0.06 a | 3.68 ± 0.09 a | 3.84 ± 0.07 ab |
| T4 [ng·mL-1] | 2.75 ± 0.59 a | 2.87 ± 0.44 a | 2.51 ± 0.53 a | 2.59 ± 1.10 a | 2.24 ± 0.84 a |
Values in rows with different letters (a, b, c) are significantly different, P ≤ 0.05 (Duncan’s post-hoc test); bars indicate standard error (n = 8). C - control diet (AIN-93G), CO - diet containing control curly kale 'Oldenbor F1', BO - diet containing biofortified curly kale 'Oldenbor F1', CR - diet containing control curly kale 'Redbor F1', BR - diet containing biofortified.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



