
Submitted:
20 September 2024
Posted:
23 September 2024
You are already at the latest version
Abstract
Bioactive like resveratrol, epigallocatechin gallate, curcumin, and other polyphenols often target various signalling pathways, including NFκB, TGFβ, and Wnt/β-catenin in osteoarthritis (OA) pathogenesis, by executing epigenetic-modifying activities. Epigenetic modulation by bioactive is gaining traction with the onset of nutrigenomics. Such modulations can target genes by histone modifications, promoter DNA methylation, and non-coding RNA expression, some of which are directly involved in disease pathophysiology but less explored in OA. OA patients often explore options that can improve the quality of their life in addition to existing treatment with nonsteroidal anti-inflammatory drugs (NSAIDs). Although bioactive and natural compounds exhibited therapeutic potential against OA, several disadvantages loom over them, like insolubility and poor bioavailability. Nanoformulated bioactives promise a better way to alleviate OA since they also control systemic events, including metabolic, immunological, and inflammatory responses, by modulating host gut microbiota that can regulate OA pathogenesis. Recent data suggest gut dysbiosis in OA. However, limited evidence is available on the role of bioactive as epigenetic and gut modulators in ameliorating OA. Moreover, the question of whether polyphenolic bioactive modulates gut microbial response that results in epigenetic-controlling activities of OA pathogenesis is not known. This narrative review explored the interplay of nanoformulated bioactive and host gut microbiota on chondrocyte growth, metabolism, and epigenetic alteration linked with OA progression and pathogenesis.
Keywords:
Osteoarthritis
; Epigenome
; Nanoformulation
; Bioactive
; Polyphenol
; Nanocurcumin
; Non-coding RNA
; Gut microbiota
; Chondrocyte
1. Introduction
Osteoarthritis (OA) is a progressive musculoskeletal disease resulting in pain, joint stiffness and loss of flexibility caused by cartilage degradation and subchondral bone remodelling [1]. The prevalence of OA is rapidly increasing from 4.8% (1990) to 7.6% (2020) among the age-normalized global population [2]. In addition to older people, OA also affects around 3.5% of working (30-60 years) adults due to increasing rates of obesity and exposure to various lifestyle and environmental factors like diet, exercise, and others [2]. The OA pathogenesis shows narrowed joint space, bone marrow lesions, cartilage erosion, synovitis, and osteophyte formation [3]. Dysregulated intercellular cytokine-mediated communication resulting in inflammation is also involved in OA pathophysiology. Since inflammation and arthralgia are predominant symptoms of OA, non-steroidal anti-inflammatory drugs (NSAIDs) are often prescribed despite their side effects and toxicity. Chronic use of NSAIDs causes adverse drug reactions that might affect the cardiovascular, central nervous, renal, and gastrointestinal systems [4]. Other disease-modifying OA drugs include monoclonal antibodies, peptides, nucleic acids, enzyme inhibitors, cytokine inhibitors, and various bone-active growth factors. These drugs have been focused on inhibiting the progression of OA by preventing cartilage loss [5]. However, these drugs demonstrate a short half-life, cause adverse reactions, and thus decrease therapeutic activity.
To date, OA is incurable, and joint replacement surgery is the last resort to provide symptomatic relief. Despite surgery, joint function may not be restored entirely, and patients may experience flare-ups [5]. In addition, various post-surgery risks and complications and a high chance of revision surgery [6] have opened the scope for novel biotherapeutics. It has been estimated that roughly 47% of patients suffering from OA use alternate therapy, including lifestyle changes like exercise, diet, and nutraceuticals to alleviate pain [7]. NSAIDs commonly provide symptomatic relief by inhibiting cyclooxygenase and thereby preventing the biosynthesis of prostaglandins. However, inhibiting prostaglandin production might elevate the risk of renal, gastrointestinal, and cardiovascular complications [8,9]. Further, it has been suggested to reduce NSAID dosage as they exert dose-dependent side effects. Several options, including nanoformulation and encapsulation, are emerging to enhance the therapeutic efficacy of the drug at lower dosages [10]. These formulations have reduced particle size, improved bioavailability, and better absorption at the tissue level. Moreover, drug encapsulation limits systemic exposure, enhances drug solubility, and exhibits controlled release over time.
Several bioactives, including polyphenols, have protective roles against inflammatory diseases, including OA [11]. Polyphenols play a role in modulating the immune system and are being studied from a therapeutic perspective [12]. Despite their potential, the therapeutic efficacies are limited due to poor solubility and bioavailability, which can be improved by introducing encapsulation or using a nanocarrier delivery system [13]. Nanoformulation comprises liposomes, micelles, polymers, and dendrimers, emerging as drug delivery systems carriers [14]. Based on the European Commission, particle or particle aggregates in the size range of 1-100nm are defined as nanomaterial. However, reference frameworks for their biosafety, quality, and efficacy are evolving to comply with the regulatory perspective of these formulations [15].
The use of nanotherapy in improving bone health, treating and preventing bone ailments, and maintaining a healthy bone status could have multiple benefits, but they need to be extensively reviewed. With a focus on various polyphenolic nanoformulations, including curcumin’s anti-inflammatory potential as a complement therapy in addition to NSAID, exploring various potential bioactives and the precise molecular mechanisms involved in delaying the progression of the disease is not known. Recently, epigenetic regulation of chondrocyte homeostasis during OA has been drawing attention [16]. Understanding their modalities in systemic responses involving gut microbiota and their possible effects on epigenetic controls of OA pathogenesis could harness a viable therapeutic option in delaying and treating OA.
An extensive literature search was conducted using Scopus, PubMed, and Science Direct databases to find relevant studies using search terms like osteoarthritis, chondrocytes, inflammation, nanoformulation, curcumin, polyphenols, bioactive, epigenetics, gut microbiome, and similar keywords. Articles published in English over the last 15 years, comprising both clinical trials and pre-clinical studies, were included. This narrative review aimed to provide current knowledge with potential mechanisms involved with the therapeutic management of OA with the help of nanoformulation.
2. Nanotherapeutics in Osteoarthritis
2.1. Nanoformulation and Nanoemulsion as Bioactive Carriers
Due to its avascular nature, the cartilage tissue has a major limitation in its ability to heal or repair, which poses a challenge for drug delivery. Systemic delivery of drugs does not reduce disease symptoms as the damaged tissue lacks vascularity. Hence, localized delivery of drugs via intra-articular administration could be a viable alternative [17]. The emergence of nanotherapeutics serves as a boon due to its various advantages in drug delivery for OA (Figure 1).
Compared to conventional therapy, nanoformulations provide the advantages of improving drug delivery and efficacy, reducing cartilage damage, and promoting M2 macrophage polarization and extracellular matrix protein synthesis. Nanoformulations used in drug delivery can be classified based on the type of encapsulation carrier, such as polymer-based (polymer micelles, polymer nanoparticles, dendrimers, and polymer vesicles) or lipid-based (nanostructured lipid carriers, lipid nanocarriers, solid-lipid nanocarriers, liposomes, and nanoemulsions) [18]. Certain other inorganic nanoformulations have also been reported in OA. For example, magnetic nanoparticles coupled with transient receptor potential vanilloid type 1 (TRPV1) antibodies were found to alleviate OA by preventing chondrocyte ferroptosis and macrophage inflammation via alternating magnetic field stimulation [19]. Further, the limitations associated with the bioavailability of polyphenols as an alternate therapy in OA have led to a shift in focus towards polyphenolic nanoformulations and drug delivery systems that are widely gaining interest in treating and slowing down disease progression [20].
Polymeric nanoformulations utilize natural or synthetic polymers that self-assemble to encapsulate drugs using emulsification, nanoprecipitation, and solvent evaporation techniques. These are emerging for their roles in drug delivery due to their inherent advantages like reduced toxicity, target site delivery, prolonged retention, controlled release, increased bioavailability, biodegradable properties, and increased drug solubility [21]. Polymers used in OA nanotherapy include chitosan, hyaluronic acid, polylactic acid (PLA), polylactic-co-glycolic acid (PLGA), polycaprolactone (PCL) and polyamidoamine (PAA). Chitosan is a cationic polymer that promotes transcellular and paracellular transport of drugs and exhibits pH-dependent drug release due to its ability to solubilize at acidic pH [22]. Hyaluronic acid is, in contrast, an anionic polymer present in the extracellular matrix of cartilage with a specific binding affinity with the CD44 receptor [23]. PLGA is a synthetic biodegradable polymer that hydrolyses to form metabolite monomers – glycolic acid and lactic acid [24]. These polymers are finding emerging applications both individually and synergistically in treating OA.
Lipid-based nanoformulation utilizes lipid biomolecules as a carrier and demonstrates superior targeted drug delivery via ligand-receptor interaction in OA treatment [25]. They comprise an aqueous region surrounded by a lipid bilayer where drugs are encapsulated and released gradually at the target site [18]. Liposomes are self-assembling spherical vesicles that elevate drug diffusion across the plasma membrane [26]. Celecoxib-loaded liposomes conjugated with hyaluronic acid hydrogels were found to be responsive to shear by restructuring, provided lubrication, increased retention, and exhibited targeted delivery of celecoxib (NSAID), thus attenuating OA [27]. Solid-lipid nanoparticles (SLNs) contain biocompatible lipids like biowaxes, fatty acids, or triglycerides that remain solid at room temperature. Solid-lipid nanoparticles conjugated with chondroitin sulfate and loaded with aceclofenac exhibited increased cellular uptake and extended drug release in an OA model [28]. Nanostructured lipid carriers (NLC) are different from SLNs due to the use of liquid lipids, which confers their unique properties for drug delivery [29]. NLC loaded with ibuprofen in the form of a topical gel was found to enhance drug permeation via skin in mice for the treatment of joint inflammation associated with OA [30]. Nanoemulsions are stable liquid-in-liquid dispersions with increased stability, bioavailability, and cellular uptake [31]. Nanoemulsion gel of chondroitin sulfate and glucosamine was found to alleviate symptoms of knee OA by attenuating cartilage damage and reducing pain [32].
2.2. Polyphenolic Nanoformulation and Osteoarthritis
The recent advancement of nanotechnology has significantly impacted disease treatment, particularly in the context of OA, by addressing the bioavailability challenges associated with bioactive polyphenols. Various nanoformulations, including nanoemulsion, lipid nanoparticles, polymeric micelles, polymeric nanoparticles, organic-based nanoparticles, inorganic-based nanoparticles, carbon-based nanoparticles, β-lactoglobulin assembled nanoparticles, and metal oxide nanoparticles are currently under research [33]. When combined with bioactive compounds, these formulations enhance solubility, improve bioavailability, enable controlled release, and reduce the quantity and frequency of doses, thereby enhancing their therapeutic potential.
Plant polyphenols comprise multiple phenol units that occur naturally as secondary metabolites and have potent anti-inflammatory properties as they act on pro-inflammatory mediators. In addition, polyphenols target oxidative stress by scavenging reactive-oxygen species (ROS) molecules [34]. Recent data suggests that these polyphenolic compounds inhibit matrix degradation caused during OA pathogenesis by promoting the expression of extracellular matrix components such as aggrecan and type II collagen [35]. Some polyphenols also directly bind to matrix metalloproteinases (MMPs) and inhibit their activities [36]. Polyphenols also exhibit their chondroprotective role by activating the Nrf2/ARE pathway [37]. This redox homeostasis signalling pathway attenuates chondrocyte apoptosis, oxidative stress, and extracellular matrix degradation. Further, polyphenol supplementation in knee OA patients improved physical function and alleviated pain and inflammation [38]. Various bioactive polyphenols such as methyl gallate, hydroxytyrosol, quercetin and rosmarinic acid showed chondroprotective effects to ameliorate OA, which are possible therapeutics [39,40]. Rosmarinic acid effectively suppressed MMP1, MMP3 and MMP13, subsequently upregulating type II collagen [41,42].
The beneficial effects of bioactives and their nanoformulation have only recently come to light. For instance, the poor bioavailability of hydroxytyrosol has led to the formulation of a hydrogel containing nanoparticles. These hydroxytyrosol-chitosan nanoparticles, in combination with hyaluronic acid and pluronic micelles, efficiently suppress pro-inflammatory effects and oxidative stress in chondrocytes [43]. Morin hydrate, a natural antioxidant and anti-inflammatory compound, has been used to formulate a copper-morin-based metal-organic framework (MOF) to mimic metalloenzymes [44]. These nanoenzymes suppress OA progression by repairing mitochondrial function and modifying the levels of pro-inflammatory markers. Nanofiber microspheres of tannic acid and strontium ions were found to reduce apoptosis, downregulate IL1β, TNFα expression, and inhibit cartilage degradation upon intra-articular treatment [45]. Quercetin, curcumin, dimethyl curcumin, resveratrol, and oxymatrine are some bioactives encapsulated into liposomes and shown to modulate pro-inflammatory cytokines involved in OA [46]. Recent data show the beneficial effects of polyphenol nanoformulations in osteoarthritis (Table 1).
2.3. Epigallocatechin in Osteoarthritis
Epigallocatechin-3 gallate (EGCG) is a polyphenol, predominant in green tea known for its antioxidant and anti-inflammatory effects [59]. Chondroprotective effects of EGCG were observed in primary osteoarthritic chondrocytes. Several proteins such as IL6, MCP1, IL8, GM-CSF, GROα, GRO, MCP3, IP10, GCP2, NFκB, and NAP2 that were upregulated upon IL1β stimulation were found to be downregulated by EGCG [60]. EGCG has been reported to exhibit antiarthritic properties by epigenetic modulation of global miRNA expression, thus reducing inflammation in IL1β-induced primary chondrocytes. Further, it was found that EGCG inhibited the expression of ADAMTS5 and COX2, which is upregulated upon IL1β stimulation via the modulation of hsa-miR-140-3p and hsa-miR-199a-3p [61,62]. Several nanoformulations used to improve the bioavailability of epigallocatechin-3 gallate have been researched. Copper EGCG nanosheets modulated ROS signaling and downregulated pro-inflammatory cytokines’ expression in primary chondrocytes by converting M1 macrophages to M2 macrophages [63]. EGCG nanodrug, in combination with selenomethionine, significantly ameliorated OA by reducing oxidative stress, accumulation of Fe2+, and promoting activation of glutathione peroxidase 4 [64]. Nanoparticles of EGCG and glucosamine mixture displayed improved anti-inflammatory potential and higher antiarthritic activity compared to the native mix [65]. A novel formulation with near-infrared nano-enzyme of EGCG with Au and Ag improves OA treatment by reducing cartilage damage [55]. Recent work on hydrogels of epigallocatechin-3 gallate seems to be a promising therapy for OA. EGCG, in combination with hyaluronic acid, showed chondroprotective effects by inhibiting IL1β, TNFα, and MMP13, scavenging ROS, and inducing M2 macrophage polarization in chondrocytes [66,67].
2.4. Resveratrol in Osteoarthritis
Resveratrol is a polyphenol known for its antioxidant and anti-inflammatory functions, which highlights its chondroprotective effects by improving joint function and reducing cartilage degradation [68,69,70]. The poor solubility and low bioavailability of resveratrol have led to the exploration of nanoformulation. Resveratrol nano-encapsulated in a self-assembling lipid core led to reduced nitric oxide levels in primary human chondrocytes [71]. Resveratrol nanoemulsions have been found to increase intracellular uptake compared to native resveratrol, minimize toxicity, and decrease oxidative stress [72]. Further, PLGA nanoparticles of resveratrol could inhibit chondrocyte apoptosis, induce autophagy and subsequently alleviate symptoms of OA [73]. Due to their sustained release, these nanoparticles showed improved bioavailability with quantifiable concentrations of resveratrol for 35 days. The differentially expressed genes such as CXCL1, IL6, NOX4 and MMP3 were upregulated in primary chondrocytes from OA patients, and those downregulated upon resveratrol treatment [74]. In addition, targeting MALAT1/miR-9/NFκB1 and caspase-3/MMP13 axis with resveratrol revealed its therapeutic potential in managing OA [75]. The combined efficacy of resveratrol with meloxicam (NSAID) significantly alleviated symptoms of knee pain in OA and had superior efficacy compared to meloxicam alone [76,77]. Hydrogel of resveratrol combined with oxidised hyaluronic acid upregulated SOX9, aggrecan, and type II collagen while suppressing expression matrix metalloproteinases in LPS-induced inflammation in chondrocytes [78], indicating its further scope in eliminating the disadvantages of bioaccessibility issue of the resveratrol.
2.5. Curcumin in Osteoarthritis
Curcumin, a dietary polyphenol and an active ingredient of Curcuma longa, has shown potent anti-inflammatory, anti-cancer, antioxidant, and anti-bacterial effects. Evidence suggests anti-arthritic, immunomodulatory and chondroprotective effects of curcumin [79]. Curcumin has emerged as a safer and more effective supplement for ameliorating OA [80]. Curcumin alone or co-supplemented with other polyphenols effectively reduced symptoms of OA, as evidenced by improved physical performance and reduced WOMAC (Western Ontario and McMaster Universities Osteoarthritis Index) and VAS (Visual Analog Scale) scores [81]. Curcumin supplementation reduces the expression and secretion of various serum inflammatory markers such as IL4, IL6, TNFα, and CRP/hs-CRP (high-sensitivity C-reactive protein) [82,83,84,85]. Despite its potential, poor solubility, limited distribution, restricted bioaccessibility, and inadequate bioavailability of curcumin restrict its therapeutic efficacy. Therefore, nanoformulation approaches are extensively adopted to improve the effectiveness of curcumin.
Curcumin nanoformulations have been designed using various methods to improve solubility, bioavailability, and targeted delivery. For example, self-assembling lipid-core curcumin nanocapsules showed improved solubility and were more effective in reducing nitric oxide in reducing apoptosis in the inflamed chondrocytes [71]. Further, to avoid any harmful effects of nanocarriers, a carrier-free assembly of curcumin and icariin nanoparticles by π-π stacking formulation exhibited improved cellular uptake, displayed prolonged drug release and synergistic anti-inflammatory effects, thus alleviating OA by protecting the cartilage [86]. Liposomal formulations of curcuminoids (curcumin and bisdemethoxycurcumin) promoted cellular uptake, improved osteoprotegerin/receptor activator nuclear factor κB (OPG/RANKL) ratio and downregulated IL1β, demonstrating its role in attenuating OA progression and preventing osteoclastogenesis [87]. Curcumin’s antioxidant potential has been proven to increase upon its encapsulation [88]. To further enhance its antioxidant potential, a self-assembling ROS-responsive polymeric micelle encapsulated with curcumin displayed extended drug release and ROS-scavenging when triggered with H2O2 in chondrocytes and in the OA rat model [89]. Polymeric micelles in self-assembling acid-activatable curcumin polymer showed chondroprotective effects by downregulating IL1β and TNFα in the acidic microenvironment of monoiodoacetate (MIA)-induced OA [90,91]. Similarly, curcumin encapsulated in a pH-responsive cyclic brush zwitterionic polymer exhibited controlled drug release, upregulated type II collagen, and aggrecan, suppressed pro-inflammatory markers such as MMP13, IL1β and improved lubrication in the synovial joints [92].
The efficacy of curcumin-loaded polylactic-co glycolic acid nanoparticles (PLGA NPs) in MIA-induced OA model exhibited a higher stability, bioavailability and greater chondroprotective effects over native molecules by downregulating MMP1, MMP3, MMP13, IL1β, TNFα and improving locomotor function [93]. In another study, Cur-PLGA nanoparticles alleviated knee-OA in rats by downregulating TNFα, IL6, IL1β, TGFβ and NFκB [94]. An analysis of the synergistic effects of hyaluronic acid/chitosan nanoparticles and curcuminoids in the knee-OA model revealed a decrease in chondrocyte apoptosis by upregulating the IκB and thereby inhibiting the NFκB pathway, downregulating the MMP1, MMP13 and promoting the expression of type II collagen [95]. A similar combination of hyaluronic acid and curcumin-loaded chitosan nanoparticles ameliorated OA due to their prolonged retention in the synovial cavity, inhibiting chondrocyte apoptosis, downregulating pro-inflammatory cytokines and upregulating RUNX2 and AP1 [96]. Curcumin microgel using poly (ethylene glycol) dimethacrylate efficiently attenuated IL1β-induced inflammatory response in chondrocytes and promoted cartilage repair in the OA model, thus displaying its pro-regenerative potential [97]. Curcumin was encapsulated into gelatin/silk fibroin microspheres to target localized delivery and studied for its effect on MIA-induced OA. Curcumin microspheres displayed extended anti-inflammatory effects, showed significant histological improvements in rat bone tissue, and reduced apoptosis and serum IL6 [98]. Another study involving silk fibroin nanoparticles (SFN) demonstrated how curcumin’s efficacy and anti-inflammatory potential are enhanced when encapsulated into SFNs. Curcumin SFNs could provide an environment for controlled drug release and cytocompatibility, improve anti-inflammatory effects by regulating RANTES and IL6, and display antioxidant effects by ROS-scavenging [99].
Several types of formulations are being developed to improve curcumin’s bioavailability further. For example, next-generation ultrasol curcumin (NGUC) efficiently alleviated OA pathophysiology by downregulating CRP, TNFα, IL6, IL1β, NFκB, COX2 and MMP3 and upregulating GPX, CAT and SOD [100]. Another formulation, palmitoyl-glucosamine (PGA), co-micronized with curcumin, results in improved bioavailability and reduces pain, tissue damage and paw edema in OA pathophysiology. The PGA-cur downregulated MIA-induced pro-inflammatory cytokines such as IL1β, TNFα, MMP1, MMP3, and MMP9 when integrated into the diet, was found to maintain meloxicam-based pain relief in dogs with OA pain [101,102]. A formulation of curcumin in water-dispersible form, was found to facilitate MIA-induced OA by inhibiting chondrocyte apoptosis and cartilage damage, in addition to improving weight-bearing imbalance and downregulating caspase-3, nitrotyrosine, phospho-NFκB, and TNFα [103]. Several curcumin nanoformulations are reported in clinical trials as a therapeutic against OA, tabulated in Table 2.
Despite evidence of curcumin’s safety and efficacy, the mechanism of curcumin’s action in ameliorating OA is still unclear. Several mechanisms have been postulated, and curcumin might regulate more than one pathway simultaneously. Curcumin was found to modulate key amino acid metabolism like threonine, serine and glycine, histidine, cysteine and methionine, glycerolipid, and inositol phosphate metabolism [111]. Curcumin also acts on the NFκB pathway, thereby inhibiting the expression of matrix metalloproteinases and reducing cartilage degradation [112]. Curcumin may act like a natural COX inhibitor and regulate inflammatory pathways [113]. Clinical trials with comparative supplementation of curcumin and a NSAID have evidenced similar efficacy [82,83,114]. Further, curcumin has been shown to inhibit the production of pro-inflammatory mediators in an OA model in chondrocytes as well as in the articular cartilage of rats in vivo [115,116]. Despite the current knowledge gap of curcumin’s mode of action, nanoencapsulation and its use as a nutritional intervention exhibited great potential.
3. Epigenetic Modulations in Osteoarthritis Pathogenesis
Gene expression modulates OA pathogenesis via epigenetic variables, including changes in chromatin structure, histone modifications, DNA methylation, transcriptional regulation, and the role of non-coding RNA. These mechanisms, in turn, modulate gene expression of transcription factors, specific extracellular matrix proteins, cytokines, and matrix-degrading proteinases [117]. Epigenetic regulation is also involved in dysregulated gene expression in the articular cartilage via the suppression of NFAT1 (nuclear factor of activated T cell 1) and SOX9 in an age-dependent manner [118].
3.1. Role of Histone Modifications in Osteoarthritis Pathology
Histone modifications and histone-modifying proteins play a significant role in cartilage development and, subsequently, OA disease pathology. Histone hyperacetylation has been linked to pathogenesis in rheumatoid arthritis patients [119]. p300/CBP, a histone acetyltransferase is involved in the hyperacetylation of COL2A1 at H3K9 (lysine 9 on histone 3) and H4K8 (lysine 8 on histone 4) via SOX9 [120]. p300/CBP associated with transcriptional factors, such as SOX5, SOX6 and SOX9, upregulated cartilage oligomeric matrix protein [121]. Histone deacetylase (HDAC) enzymes have been indicated to play a role in regulating chondrocyte differentiation [122]. Zinc finger protein 521 (Zfp521) exerted chondroprotective functions by upregulating nuclear HDAC4 and promoting cartilage growth [123]. In osteoarthritic chondrocytes, upregulated HDAC1, HDAC2, and HDAC3 expression was observed [124,125]. Further, HDAC7 overexpression upregulates MMP13 and impairs β-catenin and Wnt/β-catenin signalling in OA [126,127]. In this context, HDAC inhibitors (HDACi) that target Nrf2, MAPK, and NFκB can be potentially used as a therapeutic for the management of OA [128]. EZH2 (enhancer of zeste homolog 2), a histone-lysine N-methyltransferase upregulated SDC1 by inhibiting miR-138 via histone methylation in the promoter region and induced cartilage damage in IL1β induced OA [129]. In osteoarthritic chondrocytes, EZH2 is significantly overexpressed and upregulated Indian hedgehog (IHH), COL10A1, ADAMTS5, and MMP13, downregulated upon EZH2 inhibition [130]. Further analysis revealed that EZH2 deficiency upregulated osteogenic gene expression in chondrocytes [131]. EZH2 inhibition could slow down OA progression by reducing cartilage degradation and downregulating the expression of genes associated with the inflammatory response [132]. In addition, NSD1, which is a H3K36 (36th lysine on histone 3) methyltransferase, was found to regulate cartilage homeostasis via upregulating OSR2 expression and H3K36 methylation [133]. Analysis of chromatin immunoprecipitation sequencing (ChIP-seq) data, RNA-seq data, and genome-wide association studies (GWAS) of OA using 3D chromatin structure mapped onto chondrocyte-specific network revealed possible risks in gene expression associated with OA [134]. GWAS data combined with Hi-C mapping led to the identification of genetic variants within chromatin loops, including the enhancer-promoter loops, thus identifying putative effector genes such as PAPPA (pregnancy-associated plasma protein A) and SPRY4 (sprout RTK signalling antagonist 4) in OA [135].
3.2. Modulation of Promoter DNA Methylation in Osteoarthritis
Methylomes of healthy and OA cartilage show significant differences with increased DNA methylation in the latter [136,137]. Increased promoter methylation has been negatively correlated with gene expression and vice versa [138]. Promoter-specific hypermethylation of COL9A1 and SOX9 has been evidenced in osteoarthritic chondrocytes [139,140]. In addition, promoter-specific hypomethylation of C-terminal binding protein (CtBP) was found to upregulate CtBP-dependent NLRP3 expression and downstream pathways and aggravate OA pathogenesis [141]. Studies have concluded that CpG sites on several genes such as MMP3, MMP9, MMP13, FGFR2, GDF5, SOST, RUNX2, IL1β, IL8, iNOS, COL11A2, COL9A3, and ADAMTS4 which play diverse roles in OA undergo differential methylation [138]. Hypomethylation of leptin has been seen to upregulate leptin levels and MMP13, its downstream inhibitor contributing to OA progression [142]. DNA methylation studies to understand OA pathogenesis that was initially limited to chondrocytes are also being studied in synovial tissue and subchondral bone where significant epigenetic changes have been noted [143,144]. Differentially methylated regions (DMRs) found in OA patients were characterized by increased inflammatory responses [145]. The majority of differentially methylated sites, around 50% seen in the enhancer region, could denote the critical role of enhancer methylation in OA [146]. DNA hypermethylation also suppresses miR146a and miR140-5p in osteoarthritic synoviocytes and chondrocytes, respectively [147].
The role of transcriptional regulation in the OA microenvironment and its involvement in disease pathogenesis has become increasingly apparent in recent years. Global gene expression analysis has revealed that certain transcription factors, like FOS, FOSL2, JUN, RELA, MYC, and EGR1 are suppressed in OA pathogenesis and dysregulating other genes involved in OA [148]. Further, FoxO1 (forkhead o family transcription factor) has been elucidated to play a major role in OA pathophysiology by modulating autophagy, and promoting proteoglycan expression. It may contribute to the upregulation of antioxidant genes [149,150,151]. As a therapeutic approach, a recent study found that histone deacetylase inhibitor Panobinostat targets FoxO transcription factors, downregulates IL1β induced pro-inflammatory cytokines, ameliorates OA by promoting autophagy, and reduces cartilage damage [152]. Another transcription factor, SOX4, which is upregulated in OA, could be a potential therapeutic target in addition to its use as a diagnostic biomarker [153]. Growth differentiation factor 5 (GDF5) is vital in joint homeostasis and development. A functional polymorphism of GDF5 (rs143383) at 5’UTR has emerged as a significant risk factor for OA [154,155]. Overexpression of the transcription factor HOXA10 promoted GDF5 expression in a Gdf5-HiBiT knock-in mice model [156]. This data suggests modulation of promoter DNA methylation could play an intricate role in transcriptional regulation in OA pathogenesis.
3.3. Role of Non-Coding RNAs in Osteoarthritis Pathogenesis
Evidence elucidating the role of non-coding RNAs (ncRNAs) such as long non-coding RNA (lncRNA), microRNA (miRNA), circular RNA (circRNA), and small-interfering RNA (siRNA) in chondrogenesis, chondrocyte differentiation and cartilage development in OA is expanding [157]. Further, evidence suggests that crosstalk exists between mRNA, lncRNA, circRNA, and miRNA that regulate OA progression [158]. Their specific roles in OA pathogenesis and associated therapeutic applications are currently being explored. Many studies have concluded that lncRNAs are involved in OA-related cartilage homeostasis [159]. LncRNAs such as MALAT1, HOTAIR, H19, XIST, and GAS5 have been extensively studied for their role in OA [160]. MALAT1 has been shown to promote OA-related matrix degradation and chondrocyte apoptosis by downregulating miR150-5p [161]. LncRNA-ATB downregulation in patients serves as a diagnostic marker for OA [162]. LncRNA SNHG1 overexpression in the OA model activates the PI3K/Akt pathway and reduces chondrocyte apoptosis [163]. LncRNA AC005165.1 mitigates inflammation and apoptosis in osteoarthritic chondrocytes by interacting with miR199a-3p and TXNIP [164]. Further, L-glutamine was found to upregulate lncRNA NKILA, downregulate MMP13, NOS, COX2, NFκB and TNFα and subsequently reduced extracellular matrix degradation [165]. LncRNAs have also been linked to obesity, which is a significant risk factor for OA progression [166]. Further, some circRNAs such as circSOD2, circ_0044235, and cir_0037658 also regulate OA pathogenesis through modulating the growth-related signalling axis [167,168,169,170].
Aberrant expression of miRNAs has been associated with OA pathogenesis and progression. Increased expression of miRNA128a has been associated with cartilage degradation via repressing autophagy in OA chondrocytes [171]. Downregulation of miR214-3p expression activated the NFκB signalling and aggravated OA [172]. Upregulation of miR103 expression in osteoarthritic cartilage also led to OA progression by inducing chondrocyte apoptosis [173]. Additionally, miR9-5p, miR146a-5p, miR138-5p, miR335-5p and miR98 expression were also upregulated significantly in the osteoarthritic cartilage or serum as compared to healthy control and could serve as novel biomarkers [174,175]. Downregulation of miR222 expression led to the activation of HDAC4 and MMP13, and promoted cartilage degradation [176]. In addition to their involvement in the progression of OA, miRNA expression is connected to several signalling pathways, including NFκB, IGF, TGFβ, and BMP. It regulates various proteins, and cytokines involved in OA [177]. A recent meta-analysis of miRNA, SNP datasets, and differentially expressed mRNA in the osteoarthritic cartilage has revealed novel OA-related genes such as LHDB, PPIB, ARCP4-TTLL3, TPI1, and ASS1, which could serve as therapeutic targets [178]. Integrated analysis of RNA-sequencing data obtained from healthy and OA subjects, microarray, and gene expression dataset revealed novel differentially expressed microRNAs (DEMs) such as miR4435-SOS1, miR4435-PIK3R3, miR584-5p-KRAS and miR183-5p-NRAS and their target genes are involved in primary OA regulatory network [179].
3.4. Polyphenol as a Potential Epigenetic Modulator
Polyphenols such as curcumin, resveratrol, EGCG, gallic acid, and genistein have been reported to modulate epigenetic changes in disease pathogenesis [180]. It has been found that autophagy could be dysregulated by altered epigenetic regulations where several miRNAs expression is abnormally up or downregulated in OA [181]. Autophagy, a mechanism to maintain cell homeostasis by intracellular degradation, is often dysregulated in OA. The ability of polyphenols to modulate epigenetic mechanisms can be exploited in this scenario to regulate autophagy and delay OA progression [182]. Among polyphenols, more data is emerging with curcumin that can regulate inflammation and oxidative stress via epigenetic controlled transcriptional regulation and gene expression, as evidenced by pharmacokinetic and pharmacodynamic studies [183,184]. Curcumin inhibits p300 histone acetyl transferase (HAT) via proteosome-dependent degradation and displays anti-inflammatory effects by regulating TREM1, an amplifier of TLR-mediated responses [185,186]. Recent studies support curcumin-induced epigenetic changes by modulating miRNA expression, histone acetylation, deacetylation, and transcription factors [180,187].
RNA sequencing and differential gene expression analysis of Curcuma longa in primary chondrocytes have demonstrated significant modification of gene expression, leading to the downregulation of pro-inflammatory cytokines and upregulation of antioxidant and cytoprotective genes [188]. Several phytochemicals like berberine, resveratrol, curcumin and EGCG have been shown to modulate the expression of lncRNA of which curcumin has been evidenced to modulate – HOTAIR, PVT1, H19, LINC00623 and H2BFXP which are also involved in OA [160,189]. Despite this, studies showing the direct effect of curcumin on OA by modulating long-non coding RNAs are limited. Curcumin has been studied for its role in modulating transcription factors – EGR1, NRF2, STATS, and PPARγ [190,191]. Curcumin was found to reverse diabetic nephropathy by activating FoxO3a transcription factor and NRF2 while inhibiting NFκB [192]. Curcumin was found to restore autophagy in foam cells by promoting the nuclear translocation of transcription factor EB (TFEB) and reducing inflammation [193]. Further, tetrahydrocurcumin, a curcumin metabolite, could activate FoxO4, inhibit the PKB/Akt pathway, and attenuate oxidative stress [194] However, its potential mechanism in OA-related transcriptional regulation still needs to be established. Curcumin is also a modulator of DNA methyl transferases, but evidence concerning OA is lacking [195].
Concerning OA, curcumin demonstrated chondroprotective effects in high-fat diet-fed rats by reducing miR34a levels, which were associated with inhibition of apoptosis, upregulation of autophagy, upregulation of E2F1/PITX1 pathway, and amelioration of OA-like lesions [196]. The administration of oral curcumin in addition to physical exercise, such as swimming, in the MIA-induced rat model led to decreased serum CRP and downregulated inflammatory cytokines. It restored HDAC3 and miR130a expression, easing joint stiffness and pain [197]. Curcumin primed onto extracellular vesicles (Cur-EVs) derived from bone marrow stem cells (BMSC) was tested on IL1β stimulated osteoarthritic chondrocytes. Cur-EVs could modulate inflammatory signalling by reducing apoptosis, PI3K/Akt phosphorylation, and alleviate IL1β induced effects by upregulating miR126-3p in osteoarthritic chondrocytes which was initially downregulated in the absence of Cur-EVs [198]. Further, curcumin-treated mesenchymal stem cells could modulate the upregulation of miR124 and miR143 by reducing their promoter-specific DNA methylation, thus attenuating OA progression [199]. Curcumin nanomicelles were found to supress the expression of specific microRNAs such as miR16, miR138, and miR155 [108]. These miRNAs play crucial functions in immune responses during OA. Further, this nanoformulation depicted immunomodulatory effects by reducing the frequency of CD4+ T cells, CD8+ T cells, Th17 cells and B cells while increasing the frequency of Treg cells [84]. RNA interference via siRNA-based therapy for OA is also steadily growing. For the delivery of curcumin and siRNA of hypoxia-inducible factor (HIF2α) into OA joints, a pH-responsive metal-organic framework was used for encapsulation. Synergistic efficacy of curcumin and HIF2α gene silencing led to reduced cartilage degradation and downregulated inflammatory response in IL1β induced inflammatory chondrocytes [200]. Curcumin pre-treatment that targeted p300 HAT and STAT3 siRNA could synergistically effectively inhibit leptin-mediated IL8 in OA synovial fibroblasts [201].
4. Interplay of Gut Microbiome, Epigenetics, and Osteoarthritis: Mechanism of Bioactive Actions
4.1. Target Osteoarthritis by Epigenetic Modulation
Recent studies have elucidated the role of epigenetic regulation on gene expression involved in OA pathogenesis (Figure 2).
These studies not only aid in the understanding of OA pathogenesis but also serve as preceding evidence for future epigenetic-based therapeutic approaches [202,203]. Understanding the interactive perspective of gut microbiome and epigenetics in OA pathophysiology provides precise direction targeting pathogenesis. While some miRNAs have been evidenced to be involved in disease progression of OA as discussed in Section 3.3, others have exhibited their roles as therapeutic target for OA. MicroRNA1 (miR1) was found to ameliorate cartilage damage by downregulating Indian hedgehog (IHH) signalling in a transgenic mouse model [204]. Overexpression of miR193b-5p downregulated HDAC7 and decreased MMP3, MMP13 levels in IL1β-induced OA model exhibiting its protective role [205]. miR92a-3p and miR193b-3p expression increased promoter-specific histone acetylation on COL2A1, COMP, AGGRECAN, and inhibited HDAC2, HDAC3 expression in promoting chondrogenesis [125,206]. miR16-5p expression inhibited MAPK pathway and ameliorated IL1β-induced OA in chondrocytes [207]. Overexpression of miR93 effectively suppressed TLR4 and NFκB signalling, thus downregulating the activation of various pro-inflammatory cytokines in chondrocytes [208]. MicroRNA31 overexpression led to upregulation of aggrecan and type I collagen and promoted chondrocyte proliferation [209]. Inhibiting miRNA126 led to decreased apoptosis and inflammation via upregulation of Bcl2 expression [210]. Again, inhibition of miR486-5p promoted aggrecan and type II collagen via the SMAD2 pathway in OA [211]. Recent studies are focusing on nanoparticle-based delivery of miRNA therapeutics. For example, intra-articular delivery of miR141/200c antagomir in PEGylated polyamidoamine nanoparticles demonstrated increased retention and provided chondroprotection in transgenic mice [212]. Antagomir of miR365 encapsulated into nanotubes combined with yeast cell wall particle (YCWP) quickly crossed the gastrointestinal tract post oral administration, showed no toxicity, inhibited target miRNA, downregulated several pro-inflammatory markers, decreased cartilage injury and alleviated symptoms in an OA model [213].
Small interfering RNA (siRNA) utilizes the RNA interference (RNAi) mechanism to suppress target gene expression. siRNA-based therapeutics are being studied for their use in OA by targeting NFκB, mTORC1, TGFβ/SMAD, and Wnt/β-catenin pathways [214,215]. Epigenetic regulation of leptin gene via siRNA inhibited MMP13 expression in OA chondrocytes, thus exhibiting its therapeutic potential [142]. However, one major strength of siRNA therapy is its localized delivery at the target site. For this reason, siRNA-based nanotherapy is slowly gaining traction. siRNA nanotherapy can specifically target site-specific inflammatory genes resulting in lesser pro-inflammatory effects and further improve tissue homeostasis and cartilage regeneration [216]. Nanocomplexes with suitable carriers are being developed to confer stability to siRNA and recognize avascular target cell in chondrocytes. For example, siRNA targeting MMP2 in complex with positively charged nanoparticles that counteract OA by preventing ECM degradation and cartilage damage is demonstrated [217]. siRNA nanocomplexes targeting MMP13 in the osteoarthritic model led to sustained release and reduced cartilage fibrillation and synovial inflammation [218,219]. Another combination of nitric oxide scavenger and siRNA targeting carbonic anhydrase 9 (CA-9)-NAHA-CaP/siCA9 nanoparticles alleviated OA progression by reducing pro-inflammatory effects and cartilage protection [220]. Further, suppressing LPCAT3 by siRNA-lipid nanoparticles decreased OA-related cartilage erosion and pro-inflammatory cytokines via the MALAT1-LPCAT3 pathway [221].
4.2. Gut Microbiome-Mediated Epigenetic Modulation
The importance of diet and nutrition in health and disease and their possible modulation at the epigenetic level has led to the emergence of nutriepigenetics. It is notable in this regard that the gut microbiome that is influenced by diet also modulates epigenetics right from early life [222]. Gut microbiota controls several systemic events, including metabolic, immunological, and inflammatory responses. The hypothesis of the gut-joint axis and its interaction with various factors like diet, physical exercise, age, and gender has been explored recently [223,224]. Recent studies have reported gut microbiome dysbiosis in OA [225,226]. Analysis of large-scale GWAS identified three major microbial taxa such as Methanobacteriaceae, Ruminiclostridium 5, and Desulfovibrionales, that were associated with knee-OA and showed protective effects by their prevalence [227]. Further, an abundance of Streptococcus spp, results in an increased ratio of Firmicutes/Bacteroides and promoted local inflammation in OA [228]. Local inflammation due to this factor can lead to gut dysbiosis and reduce short chain fatty acids (SCFA) levels, thus posing a risk to the development of both obesity and OA. Obesity, metabolic syndrome and their impact on the immune system are also a major risk factor for OA progression [229]. In obesity, dietary factors and habits lead to aberrant epigenetic programming, resulting in hyperglycaemia and increased adiposity [230]. Obesity and gut dysbiosis cause chronic low-grade inflammation in chondrocytes, triggering inflammatory pathways that lead to cartilage degradation involving epigenetic alterations [231]. Prebiotics, probiotics and synbiotics are beneficial in obesity-related OA [232]. These supplements might regulate gut metabolite synthesis and reduce OA disease progression. Further, gut microbiota has also been reported to influence the bioavailability and metabolism of OA drugs, as highlighted by the European Society for Clinical and Economic aspects of Osteoporosis, Osteoarthritis and Musculoskeletal diseases (ESCEO) [233].
The importance of bioactive from the diet has also been elucidated, where gut metabolites derived from diet, majorly SCFAs, cause epigenetic changes [234]. Concerning obesity and related comorbidities, gut metabolites affect host metabolism by epigenetic modulation of gluconeogenesis, lipogenesis, appetite, and inflammation, suggesting that diet can either positively or negatively affect gut microbiota and associated metabolites [235]. Obesity-related gut dysbiosis also modulates inflammatory response via the epigenetic regulation of free fatty acid receptors (FFARs) [236]. In addition to SCFAs, other metabolites like vitamins, polyphenols and polyamines cause epigenetic reprogramming [237,238]. Potential mechanisms for this epigenetic regulation include – the action of gut metabolites on epigenetic enzyme regulators and substrates influencing enzyme activity of HDACs and DNMTs, involvement in DNA methylation, chromatin remodeling, histone modifications, and lncRNA, and miRNA regulation [239,240]. Gut metabolites like butyrate, an SCFA that is commonly derived from Roseburia spp. or Faecalibacterium prausnitzii, have been shown to contribute to histone modifications [241]. Histone deacetylases modulated by butyrate and certain commensal bacteria are involved in histone crotonylation and maintain intestinal homeostasis [242,243]. The role of butyrate has recently been reported its association with disease-related gut dysbiosis. Butyric acid was evidenced to lower inflammation by regulating FOSL2 modifications [244]. Further, a decrease in butyrate levels has been linked to overexpression of HDAC3, causing gut dysbiosis associated with diabetes [245]. A pilot study of whole-genome methylation analysis from blood and visceral adipose tissue revealed differential methylation levels in subjects with Firmicute-dominant bacteria, thus linking it with obesity and metabolic syndrome [246,247]. Further, increased abundance of Fusobacterium is potentially associated with DNA hypermethylation [248]. Commensal gut microbiota induces TET2/3-dependent localized DNA methylation during an inflammatory state and affect chromatin accessibility [249].
Inflammation is the major characteristic feature in OA pathogenesis. Gut microbiota as well as their derived metabolites have functions in immune modulation via epigenetic alterations. Lactic acid promoted acetylation of histone H3K27 inhibiting pro-inflammatory function of macrophages and resulting in immunosuppression [250]. DNA methylation at CpG islands of toll-like receptors such as TLR2 and TLR4 are linked to gut dysbiosis in terms of higher Firmicutes/Bacteroidetes ratio and abundance of lactic acid bacteria [251]. A randomized clinical trial reported that guided lifestyle changes and dietary interventions could decrease DNA methylation age (DNAmAge) by 3.23 years and potentially reverse epigenetic age in humans [252]. Indole-3-lactic acid derived from Lactobacillus plantarum is reported to modulate chromatin accessibility in CD8+T cells [253]. Further, β-glucan was found to inhibit the deposition of active histone marks upon LPS exposure [254]. Gut-derived inositol phosphate upregulates the activity of histone deacetylase-3 and promotes epithelial repair [255]. Capsiate, another gut metabolite, was found to inhibit ferroptosis involved in OA via the activation of SLC2A1 and HIF1α inhibition [256]. Thus, dietary modifications could reverse epigenetic changes, promote bone health, and slow disease progression by modulating gut microbiota.
5. Future Perspectives and Conclusions
Nutriepigenetics encodes the modulation of the epigenome by nutrients or bioactive, which is an emerging area of research. Bioactive polyphenols can modify or reverse aberrant gene expression via epigenetics in OA pathologies [181]. Emerging studies that dwell on the mechanisms of osteoarthritic and normal cartilage have reported that epigenetic deregulation is indeed involved in OA pathogenesis [257,258]. In synergy with pathogenesis, it is unravelling how bioactives might act on the gut microbiome and alleviate OA via epigenetic modification. We recently gathered evidence that curcumin, one of the most studied bioactive, potentially modulates the gut microbiota and alleviates OA via the gut-bone axis [11]. Several studies confirmed the increased efficacy of nanocurcumin over native, with acceptable toxicity profile, biosafety, and chondroprotective effects [93,259,260]. Despite extensive research on this golden nutraceutical, the precise mechanisms involved in curcumin’s mode of action remain elusive. One reason may be its ability to produce pleiotropic effects by simultaneously modulating several signal transduction pathways and enzymatic activities and its newly established role in epigenetic regulation. Curcumin and its metabolites may positively modulate the gut microbiome and alleviate OA by altering intestinal barrier integrity [261], mucosal inflammation [261], gut microbial profile [262], gut metabolites [263,264] and promoting chondroprotective and anti-inflammatory properties. Changes occurring at the gut-immune promote the ameliorating effects of curcumin against OA (Figure 3).
OA, while currently incurable, is witnessing a surge of potential treatments in the form of additional approaches including formulated dietary bioactives. These emerging approaches, facilitated by improved packaging and delivery to the target tissue, enhance bioavailability of bioactive or drugs. Nanoparticle-based therapy containing bioactive are showing promising benefits than native molecules, leading to higher therapeutic efficiency. These nanoformulations, which include diverse types such as polymer-based micelles, dendrimers, vesicles, liposomes, and emulsions, are demonstrating better solubility, stability, intracellular uptake, accessibility, bioavailability, and efficacy compared to native molecules in the treatment of OA.
Despite several advantages of polyphenolic nanoformulations, certain limitations still exist that must be noted. The stability of the polyphenols is guaranteed in a nanoformulation as they are shielded; however, aggregation of these nanoformulations could result in loss of functionality and might pose a threat in terms of toxicity. The right combination of bioactive based on their compatibility on various attributes, including solubility, efficacy, and toxicity, should be considered. Despite their potential in applications for drug delivery in OA, polymeric nanoformulations suffer from batch-to-batch variation, cytotoxicity, and biosafety related to cationic polymers and phagocytosis by macrophages [21]. Therefore, nanoformulation should be optimized as an efficient nanocarrier in the bioactive or drug delivery systems, ensuring stability, sustained release, prolonged bioavailability, increased cellular uptake, targeted delivery, and reduced adverse reactions in treating OA.
Author Contributions
KSNH wrote and reviewed the original draft; AKD reviewed and edited the draft; SB conceptualized, reviewed, edited, and finalized the draft.
Funding
The work is part of a research project sponsored by the ICMR-National Institute of Nutrition (grant number 20-BS07).
Conflicts of Interest
Authors declare no conflict of interest.
Acknowledgement
KSNH was supported by the DST-Inspire Fellowship, Dep. of Science & Technology, Gov. of India.
References
- Olansen, J.; Dyke, J.P.; Aaron, R.K. Is Osteoarthritis a Vascular Disease? Front. Biosci. (Landmark Ed. ) 2024, 29, 113. [Google Scholar] [CrossRef] [PubMed]
- Steinmetz, J.D.; Culbreth, G.T.; Haile, L.M.; Rafferty, Q.; Lo, J.; Fukutaki, K.G.; Cruz, J.A.; Smith, A.E.; Vollset, S.E.; Brooks, P.M.; et al. Global, regional, and national burden of osteoarthritis, 1990–2020 and projections to 2050: a systematic analysis for the Global Burden of Disease Study 2021. Lancet Rheumatol. 2023, 5, e508–e522. [Google Scholar] [CrossRef]
- Yao, Q.; Wu, X.; Tao, C.; Gong, W.; Chen, M.; Qu, M.; Zhong, Y.; He, T.; Chen, S.; Xiao, G. Osteoarthritis: pathogenic signaling pathways and therapeutic targets. Signal Transduct. Target. Ther. 2023, 8, 56. [Google Scholar] [CrossRef]
- Marcum, Z.A.; Hanlon, J.T. Recognizing the Risks of Chronic Nonsteroidal Anti-Inflammatory Drug Use in Older Adults. Ann. Long-Term Care Off. J. Am. Med. Dir. Assoc. 2010, 18, 24–27. [Google Scholar]
- Rodriguez-Merchan, E.C. The Current Role of Disease-modifying Osteoarthritis Drugs. Arch. Bone Jt. Surg. 2023, 11, 11–22. [Google Scholar] [CrossRef]
- Pulido, L.; Parvizi, J.; Macgibeny, M.; Sharkey, P.F.; Purtill, J.J.; Rothman, R.H.; Hozack, W.J. In hospital complications after total joint arthroplasty. J. Arthroplast. 2008, 23, 139–145. [Google Scholar] [CrossRef]
- Sukhikh, S.; Noskova, S.; Ivanova, S.; Ulrikh, E.; Izgaryshev, A.; Babich, O. Chondroprotection and Molecular Mechanism of Action of Phytonutraceuticals on Osteoarthritis. Molecules 2021, 26. [Google Scholar] [CrossRef]
- Crofford, L.J. Use of NSAIDs in treating patients with arthritis. Arthritis Res Ther 2013, 15 (Suppl. S3), S2. [Google Scholar] [CrossRef]
- Anwar, A.; Anwar, I.J.; Delafontaine, P. Elevation of cardiovascular risk by non-steroidal anti-inflammatory drugs. Trends Cardiovasc. Med. 2015, 25, 726–735. [Google Scholar] [CrossRef]
- Maniar, K.H.; Jones, I.A.; Gopalakrishna, R.; Vangsness, C.T., Jr. Lowering side effects of NSAID usage in osteoarthritis: recent attempts at minimizing dosage. Expert Opin. Pharmacother. 2018, 19, 93–102. [Google Scholar] [CrossRef]
- Basak, S.; Hridayanka, K.S.N.; Duttaroy, A.K. Bioactives and their roles in bone metabolism of osteoarthritis: evidence and mechanisms on gut-bone axis. Front Immunol 2024, 14, 1323233. [Google Scholar] [CrossRef] [PubMed]
- Mileo, A.M.; Nisticò, P.; Miccadei, S. Polyphenols: Immunomodulatory and Therapeutic Implication in Colorectal Cancer. Front Immunol 2019, 10, 729. [Google Scholar] [CrossRef] [PubMed]
- Tomar, R.; Das, S.S.; Balaga, V.K.R.; Tambe, S.; Sahoo, J.; Rath, S.K.; Ruokolainen, J.; Kesari, K.K. Therapeutic Implications of Dietary Polyphenols-Loaded Nanoemulsions in Cancer Therapy. ACS Appl. Bio Mater. 2024, 7, 2036–2053. [Google Scholar] [CrossRef] [PubMed]
- Yusuf, A.; Almotairy, A.R.Z.; Henidi, H.; Alshehri, O.Y.; Aldughaim, M.S. Nanoparticles as Drug Delivery Systems: A Review of the Implication of Nanoparticles’ Physicochemical Properties on Responses in Biological Systems. Polymers 2023, 15. [Google Scholar] [CrossRef] [PubMed]
- Soares, S.; Sousa, J.; Pais, A.; Vitorino, C. Nanomedicine: Principles, Properties, and Regulatory Issues. Front. Chem. 2018, 6. [Google Scholar] [CrossRef] [PubMed]
- Tong, L.; Yu, H.; Huang, X.; Shen, J.; Xiao, G.; Chen, L.; Wang, H.; Xing, L.; Chen, D. Current understanding of osteoarthritis pathogenesis and relevant new approaches. Bone Res. 2022, 10, 60. [Google Scholar] [CrossRef]
- Krishnan, Y.; Grodzinsky, A.J. Cartilage diseases. Matrix Biol. : J. Int. Soc. Matrix Biol. 2018, 71-72, 51–69. [Google Scholar] [CrossRef]
- Lu, H.; Zhang, S.; Wang, J.; Chen, Q. A Review on Polymer and Lipid-Based Nanocarriers and Its Application to Nano-Pharmaceutical and Food-Based Systems. Front. Nutr. 2021, 8, 783831. [Google Scholar] [CrossRef]
- Lv, Z.; Wang, P.; Li, W.; Xie, Y.; Sun, W.; Jin, X.; Jiang, R.; Fei, Y.; Liu, Y.; Shi, T.; et al. Bifunctional TRPV1 Targeted Magnetothermal Switch to Attenuate Osteoarthritis Progression. Res. (Wash. D.C.) 2024, 7, 0316. [Google Scholar] [CrossRef]
- Wang, H.; Wang, C.; Zou, Y.; Hu, J.; Li, Y.; Cheng, Y. Natural polyphenols in drug delivery systems: Current status and future challenges. Giant 2020, 3, 100022. [Google Scholar] [CrossRef]
- Pontes, A.P.; Welting, T.J.M.; Rip, J.; Creemers, L.B. Polymeric Nanoparticles for Drug Delivery in Osteoarthritis. Pharmaceutics 2022, 14. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, M.A.; Syeda, J.T.M.; Wasan, K.M.; Wasan, E.K. An Overview of Chitosan Nanoparticles and Its Application in Non-Parenteral Drug Delivery. Pharmaceutics 2017, 9. [Google Scholar] [CrossRef] [PubMed]
- Rao, N.V.; Rho, J.G.; Um, W.; Ek, P.K.; Nguyen, V.Q.; Oh, B.H.; Kim, W.; Park, J.H. Hyaluronic Acid Nanoparticles as Nanomedicine for Treatment of Inflammatory Diseases. Pharmaceutics 2020, 12. [Google Scholar] [CrossRef]
- Danhier, F.; Ansorena, E.; Silva, J.M.; Coco, R.; Le Breton, A.; Préat, V. PLGA-based nanoparticles: An overview of biomedical applications. J. Control. Release 2012, 161, 505–522. [Google Scholar] [CrossRef]
- Puri, A.; Loomis, K.; Smith, B.; Lee, J.H.; Yavlovich, A.; Heldman, E.; Blumenthal, R. Lipid-based nanoparticles as pharmaceutical drug carriers: from concepts to clinic. Crit. Rev. Ther. Drug Carr. Syst. 2009, 26, 523–580. [Google Scholar] [CrossRef] [PubMed]
- Guimarães, D.; Cavaco-Paulo, A.; Nogueira, E. Design of liposomes as drug delivery system for therapeutic applications. Int. J. Pharm. 2021, 601, 120571. [Google Scholar] [CrossRef]
- Lei, Y.; Wang, X.; Liao, J.; Shen, J.; Li, Y.; Cai, Z.; Hu, N.; Luo, X.; Cui, W.; Huang, W. Shear-responsive boundary-lubricated hydrogels attenuate osteoarthritis. Bioact. Mater. 2022, 16, 472–484. [Google Scholar] [CrossRef]
- Bishnoi, M.; Jain, A.; Hurkat, P.; Jain, S.K. Aceclofenac-loaded chondroitin sulfate conjugated SLNs for effective management of osteoarthritis. J. Drug Target. 2014, 22, 805–812. [Google Scholar] [CrossRef]
- Müller, R.H.; Shegokar, R.; Keck, C.M. 20 years of lipid nanoparticles (SLN and NLC): present state of development and industrial applications. Curr. Drug Discov. Technol. 2011, 8, 207–227. [Google Scholar] [CrossRef]
- Sütő, B.; Berkó, S.; Kozma, G.; Kukovecz, Á.; Budai-Szűcs, M.; Erős, G.; Kemény, L.; Sztojkov-Ivanov, A.; Gáspár, R.; Csányi, E. Development of ibuprofen-loaded nanostructured lipid carrier-based gels: characterization and investigation of in vitro and in vivo penetration through the skin. Int J Nanomed. 2016, 11, 1201–1212. [Google Scholar] [CrossRef]
- Jaiswal, M.; Dudhe, R.; Sharma, P.K. Nanoemulsion: an advanced mode of drug delivery system. 3 Biotech 2015, 5, 123–127. [Google Scholar] [CrossRef]
- Leite, C.B.; Coelho, J.M.; Ferreira-Nunes, R.; Gelfuso, G.M.; Durigan, J.L.; Azevedo, R.B.; Muehlmann, L.A.; Sousa, M.H. Phonophoretic application of a glucosamine and chondroitin nanoemulsion for treatment of knee chondropathies. Nanomed. (Lond. Engl. ) 2020, 15, 647–659. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.R.; Desu, P.K.; Nakkala, R.K.; Kondi, V.; Devi, S.; Alam, M.S.; Hamid, H.; Athawale, R.B.; Kesharwani, P. Nanotechnology-based approaches applied to nutraceuticals. Drug Delivery and Translational Research 2021 12:3 2021, 12, 485–499. [Google Scholar] [CrossRef] [PubMed]
- Ansari, M.Y.; Ahmad, N.; Haqqi, T.M. Oxidative stress and inflammation in osteoarthritis pathogenesis: Role of polyphenols. Biomed Pharmacother 2020, 129, 110452. [Google Scholar] [CrossRef]
- Ashruf, O.S.; Ansari, M.Y. Natural Compounds: Potential Therapeutics for the Inhibition of Cartilage Matrix Degradation in Osteoarthritis. Life (Basel Switz. ) 2022, 13. [Google Scholar] [CrossRef]
- Suzuki, T.; Ohishi, T.; Tanabe, H.; Miyoshi, N.; Nakamura, Y. Anti-Inflammatory Effects of Dietary Polyphenols through Inhibitory Activity against Metalloproteinases. Molecules 2023, 28. [Google Scholar] [CrossRef]
- Wu, Z.; Yang, Z.; Liu, L.; Xiao, Y. Natural compounds protect against the pathogenesis of osteoarthritis by mediating the NRF2/ARE signaling. Front Pharmacol 2023, 14, 1188215. [Google Scholar] [CrossRef]
- Valsamidou, E.; Amerikanou, C.; Tzavara, C.; Skarpas, G.; Mariolis-Sapsakos, T.D.; Zoumpoulakis, P.; Kaliora, A.C. A standardized nutraceutical supplement contributes to pain relief, improves quality of life and regulates inflammation in knee osteoarthritis patients; A randomized clinical trial. Heliyon 2023, 9, e20143. [Google Scholar] [CrossRef]
- Li, Y.; Shen, B.; Lv, C.; Zhu, X.; Naren, Q.; Xu, D.; Chen, H.; Wu, F. Methyl gallate prevents oxidative stress induced apoptosis and ECM degradation in chondrocytes via restoring Sirt3 mediated autophagy and ameliorates osteoarthritis progression. Int Immunopharmacol 2023, 114, 109489. [Google Scholar] [CrossRef]
- Permatasari, D.A.; Karliana, D.; Iskandarsyah, I.; Arsianti, A.; Bahtiar, A. Quercetin prevent proteoglycan destruction by inhibits matrix metalloproteinase-9, matrix metalloproteinase-13, a disintegrin and metalloproteinase with thrombospondin motifs-5 expressions on osteoarthritis model rats. J. Adv. Pharm. Technol. Res. 2019, 10, 2–8. [Google Scholar] [CrossRef]
- Eo, S.H.; Kim, S.J. Rosmarinic acid induces rabbit articular chondrocyte differentiation by decreases matrix metalloproteinase-13 and inflammation by upregulating cyclooxygenase-2 expression. J. Biomed. Sci. 2017, 24, 75. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.P.; Jin, G.J.; Xiong, Y.; Hu, P.F.; Bao, J.P.; Wu, L.D. Rosmarinic acid down-regulates NO and PGE(2) expression via MAPK pathway in rat chondrocytes. J Cell Mol Med 2018, 22, 346–353. [Google Scholar] [CrossRef] [PubMed]
- Valentino, A.; Conte, R.; De Luca, I.; Di Cristo, F.; Peluso, G.; Bosetti, M.; Calarco, A. Thermo-Responsive Gel Containing Hydroxytyrosol-Chitosan Nanoparticles (Hyt@tgel) Counteracts the Increase of Osteoarthritis Biomarkers in Human Chondrocytes. Antioxid. (Basel Switz. ) 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Liu, L.F.; Qin, Z.; Liu, S.; Wang, Y.; Chen, Z.; Yao, Y.; Zheng, L.; Zhao, J.; Gao, M. Natural Morin-Based Metal Organic Framework Nanoenzymes Modulate Articular Cavity Microenvironment to Alleviate Osteoarthritis. Res. (Wash. D.C.) 2023, 6, 0068. [Google Scholar] [CrossRef]
- Chen, Y.; Xu, W.; Shafiq, M.; Song, D.; Wang, T.; Yuan, Z.; Xie, X.; Yu, X.; Shen, Y.; Sun, B.; et al. Injectable nanofiber microspheres modified with metal phenolic networks for effective osteoarthritis treatment. Acta Biomater. 2023, 157, 593–608. [Google Scholar] [CrossRef]
- Craciunescu, O.; Icriverzi, M.; Florian, P.E.; Roseanu, A.; Trif, M. Mechanisms and Pharmaceutical Action of Lipid Nanoformulation of Natural Bioactive Compounds as Efficient Delivery Systems in the Therapy of Osteoarthritis. Pharmaceutics 2021, 13. [Google Scholar] [CrossRef]
- Li, Y.; Chen, M.; Yan, J.; Zhou, W.; Gao, S.; Liu, S.; Li, Q.; Zheng, Y.; Cheng, Y.; Guo, Q. Tannic acid/Sr(2+)-coated silk/graphene oxide-based meniscus scaffold with anti-inflammatory and anti-ROS functions for cartilage protection and delaying osteoarthritis. Acta Biomater. 2021, 126, 119–131. [Google Scholar] [CrossRef]
- Xiong, F.; Qin, Z.; Chen, H.; Lan, Q.; Wang, Z.; Lan, N.; Yang, Y.; Zheng, L.; Zhao, J.; Kai, D. pH-responsive and hyaluronic acid-functionalized metal-organic frameworks for therapy of osteoarthritis. J Nanobiotechnology 2020, 18, 139. [Google Scholar] [CrossRef]
- Li, G.; Liu, S.; Chen, Y.; Zhao, J.; Xu, H.; Weng, J.; Yu, F.; Xiong, A.; Udduttula, A.; Wang, D. , et al. An injectable liposome-anchored teriparatide incorporated gallic acid-grafted gelatin hydrogel for osteoarthritis treatment. Nat. Commun. 2023, 14, 3159. [Google Scholar] [CrossRef]
- Yang, H.; Yan, R.; Chen, Q.; Wang, Y.; Zhong, X.; Liu, S.; Xie, R.; Ren, L. Functional nano drug delivery system with dual lubrication and immune escape for treating osteoarthritis. J. Colloid Interface Sci. 2023, 652, 2167–2179. [Google Scholar] [CrossRef]
- Yan, R.; Yang, H.; Liu, Y.; Wang, Y.; Liu, S.; Xie, R.; Ren, L. A Dual Functional Bioinspired Lubricant for Osteoarthritis Treatment and Potential Prevention. ACS Appl. Mater. Interfaces, 2024. [Google Scholar] [CrossRef]
- Ouyang, Z.; Tan, T.; Liu, C.; Duan, J.; Wang, W.; Guo, X.; Zhang, Q.; Li, Z.; Huang, Q.; Dou, P.; et al. Targeted delivery of hesperetin to cartilage attenuates osteoarthritis by bimodal imaging with Gd(2)(CO(3))(3)@PDA nanoparticles via TLR-2/NF-κB/Akt signaling. Biomaterials 2019, 205, 50–63. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhong, Y.; He, S.; Liang, R.; Liao, C.; Zheng, L.; Zhao, J. Application of the pH-Responsive PCL/PEG-Nar Nanofiber Membrane in the Treatment of Osteoarthritis. Front Bioeng Biotechnol 2022, 10, 859442. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chen, X.; Zhou, Z.; Fang, B.; Chen, Z.; Huang, Y.; Hu, Y.; Liu, H. Berberine oleanolic acid complex salt grafted hyaluronic acid/silk fibroin (BOA-g-HA/SF) composite scaffold promotes cartilage tissue regeneration under IL-1β caused stress. Int. J. Biol. Macromol. 2023, 250, 126104. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Chang, L.; Zhao, X.; Hu, Y.; Lin, Y.; Chen, Z.; Ren, X.; Mei, X. Preparation of epigallocatechin gallate decorated Au-Ag nano-heterostructures as NIR-sensitive nano-enzymes for the treatment of osteoarthritis through mitochondrial repair and cartilage protection. Acta Biomater. 2022, 144, 168–182. [Google Scholar] [CrossRef]
- Lee, C.Y.; Chang, Y.C.; Yang, K.C.; Lin, Y.F.; Wu, A.T.H.; Tseng, C.L. Development and functional evaluation of a hyaluronic acid coated nano-formulation with kaempferol as a novel intra-articular agent for Knee Osteoarthritis treatment. Biomed Pharmacother 2024, 175, 116717. [Google Scholar] [CrossRef]
- Guo, J.; Su, K.; Wang, L.; Feng, B.; You, X.; Deng, M.; Toh, W.S.; Wu, J.; Cheng, B.; Xia, J. Poly(p-coumaric acid) nanoparticles alleviate temporomandibular joint osteoarthritis by inhibiting chondrocyte ferroptosis. Bioact. Mater. 2024, 40, 212–226. [Google Scholar] [CrossRef]
- Shaban, N.S.; Radi, A.M.; Abdelgawad, M.A.; Ghoneim, M.M.; Al-Serwi, R.H.; Hassan, R.M.; Mohammed, E.T.; Radi, R.A.; Halfaya, F.M. Targeting Some Key Metalloproteinases by Nano-Naringenin and Amphora coffeaeformis as a Novel Strategy for Treatment of Osteoarthritis in Rats. Pharm. (Basel Switz. ) 2023, 16. [Google Scholar] [CrossRef]
- Nagle, D.G.; Ferreira, D.; Zhou, Y.D. Epigallocatechin-3-gallate (EGCG): chemical and biomedical perspectives. Phytochemistry 2006, 67, 1849–1855. [Google Scholar] [CrossRef]
- Akhtar, N.; Haqqi, T.M. Epigallocatechin-3-gallate suppresses the global interleukin-1beta-induced inflammatory response in human chondrocytes. Arthritis Res Ther 2011, 13, R93. [Google Scholar] [CrossRef]
- Rasheed, Z.; Rasheed, N.; Al-Shaya, O. Epigallocatechin-3-O-gallate modulates global microRNA expression in interleukin-1β-stimulated human osteoarthritis chondrocytes: potential role of EGCG on negative co-regulation of microRNA-140-3p and ADAMTS5. Eur. J. Nutr. 2018, 57, 917–928. [Google Scholar] [CrossRef]
- Rasheed, Z.; Rasheed, N.; Al-Shobaili, H.A. Epigallocatechin-3-O-gallate up-regulates microRNA-199a-3p expression by down-regulating the expression of cyclooxygenase-2 in stimulated human osteoarthritis chondrocytes. J Cell Mol Med 2016, 20, 2241–2248. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Qin, J.; Huang, Q.; Jin, Z.; Zheng, L.; Zhao, J.; Qin, Z. Epigallocatechin-3-gallate (EGCG) based metal-polyphenol nanoformulations alleviates chondrocytes inflammation by modulating synovial macrophages polarization. Biomed Pharmacother 2023, 161, 114366. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Song, Z.; Yu, J.; Ren, B.; Dong, Y.; You, Y.; Zhang, Z.; Jia, C.; Zhao, Y.; Zhou, X.; et al. Supramolecular self-assembly of EGCG-selenomethionine nanodrug for treating osteoarthritis. Bioact. Mater. 2024, 32, 164–176. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Xiao, L.; Yu, C.; Jin, P.; Qin, D.; Xu, Y.; Yin, J.; Liu, Z.; Du, Q. Enhanced Antiarthritic Efficacy by Nanoparticles of (-)-Epigallocatechin Gallate-Glucosamine-Casein. J. Agric. Food Chem. 2019, 67, 6476–6486. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Koh, R.H.; Kim, S.H.; Kim, K.M.; Park, G.K.; Hwang, N.S. Injectable anti-inflammatory hyaluronic acid hydrogel for osteoarthritic cartilage repair. Mater. Sci. Eng.. C Mater. Biol. Appl. 2020, 115, 111096. [Google Scholar] [CrossRef]
- Li, H.; Xiang, D.; Gong, C.; Wang, X.; Liu, L. Naturally derived injectable hydrogels with ROS-scavenging property to protect transplanted stem cell bioactivity for osteoarthritic cartilage repair. Front Bioeng Biotechnol 2022, 10, 1109074. [Google Scholar] [CrossRef]
- Dave, M.; Attur, M.; Palmer, G.; Al-Mussawir, H.E.; Kennish, L.; Patel, J.; Abramson, S.B. The antioxidant resveratrol protects against chondrocyte apoptosis via effects on mitochondrial polarization and ATP production. Arthritis Rheum 2008, 58, 2786–2797. [Google Scholar] [CrossRef]
- Wang, J.; Gao, J.S.; Chen, J.W.; Li, F.; Tian, J. Effect of resveratrol on cartilage protection and apoptosis inhibition in experimental osteoarthritis of rabbit. Rheumatol. Int. 2012, 32, 1541–1548. [Google Scholar] [CrossRef]
- Singh, K.; Gupta, J.K.; Kumar, S. The Pharmacological Potential of Resveratrol in Reducing Soft Tissue Damage in Osteoarthritis Patients. Curr Rheumatol Rev 2024, 20, 27–38. [Google Scholar] [CrossRef]
- Kann, B.; Spengler, C.; Coradini, K.; Rigo, L.A.; Bennink, M.L.; Jacobs, K.; Offerhaus, H.L.; Beck, R.C.; Windbergs, M. Intracellular Delivery of Poorly Soluble Polyphenols: Elucidating the Interplay of Self-Assembling Nanocarriers and Human Chondrocytes. Anal. Chem. 2016, 88, 7014–7022. [Google Scholar] [CrossRef]
- Le Clanche, S.; Cheminel, T.; Rannou, F.; Bonnefont-Rousselot, D.; Borderie, D.; Charrueau, C. Use of Resveratrol Self-Emulsifying Systems in T/C28a2 Cell Line as Beneficial Effectors in Cellular Uptake and Protection Against Oxidative Stress-Mediated Death. Front. Pharmacol. 2018, 9. [Google Scholar] [CrossRef]
- Wei, L.; Pan, Q.; Teng, J.; Zhang, H.; Qin, N. Intra-articular administration of PLGA resveratrol sustained-release nanoparticles attenuates the development of rat osteoarthritis. Mater. Today. Bio 2024, 24, 100884. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Wang, D.; Tang, J. Identification of the Resveratrol Potential Targets in the Treatment of Osteoarthritis. Evid Based Complement Altern. Med 2021, 2021, 9911286. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Zhang, H.; You, W.; Tang, X.; Li, X.; Gong, Z. Therapeutic effect of Resveratrol in the treatment of osteoarthritis via the MALAT1/miR-9/NF-κB signaling pathway. Exp Ther Med 2020, 19, 2343–2352. [Google Scholar] [CrossRef]
- Hussain, S.A.; Marouf, B.H.; Ali, Z.S.; Ahmmad, R.S. Efficacy and safety of co-administration of resveratrol with meloxicam in patients with knee osteoarthritis: a pilot interventional study. Clin. Interv. Aging 2018, 13, 1621–1630. [Google Scholar] [CrossRef] [PubMed]
- Marouf, B.H.; Hussain, S.A.; Ali, Z.S. Correlation between serum pro inflammatory cytokines and clinical scores of knee osteoarthritic patients using resveratrol as a supplementary therapy with meloxicam. Indian J. Pharmacol. 2021, 53, 270–277. [Google Scholar] [CrossRef]
- Sheu, S.Y.; Chen, W.S.; Sun, J.S.; Lin, F.H.; Wu, T. Biological characterization of oxidized hyaluronic acid/resveratrol hydrogel for cartilage tissue engineering. J. Biomed. Mater. Res.. Part A 2013, 101, 3457–3466. [Google Scholar] [CrossRef]
- Zhao, J.; Liang, G.; Zhou, G.; Hong, K.; Yang, W.; Liu, J.; Zeng, L. Efficacy and safety of curcumin therapy for knee osteoarthritis: A Bayesian network meta-analysis. J. Ethnopharmacol. 2024, 321, 117493. [Google Scholar] [CrossRef]
- Zeng, L.; Yu, G.; Hao, W.; Yang, K.; Chen, H. The efficacy and safety of Curcuma longa extract and curcumin supplements on osteoarthritis: a systematic review and meta-analysis. Biosci. Rep. 2021, 41. [Google Scholar] [CrossRef]
- Basak, S.; Hridayanka, K.S.N.; Duttaroy, A.K. Bioactives and their roles in bone metabolism of osteoarthritis: evidence and mechanisms on gut-bone axis. Front. Immunol. 2024, 14. [Google Scholar] [CrossRef]
- Heidari-Beni, M.; Moravejolahkami, A.R.; Gorgian, P.; Askari, G.; Tarrahi, M.J.; Bahreini-Esfahani, N. Herbal formulation “turmeric extract, black pepper, and ginger” versus Naproxen for chronic knee osteoarthritis: A randomized, double-blind, controlled clinical trial. Phytother. Res. PTR 2020, 34, 2067–2073. [Google Scholar] [CrossRef] [PubMed]
- Yuan, T.; Cai, D.; Hu, B.; Zhu, Y.; Qin, J. Therapeutic Effects of Curcumin on Osteoarthritis and Its Protection of Chondrocytes Through the Wnt/Β-Catenin Signaling Pathway. Altern. Ther. Health Med. 2022, 28, 28–37. [Google Scholar] [PubMed]
- Atabaki, M.; Shariati-Sarabi, Z.; Tavakkol-Afshari, J.; Mohammadi, M. Significant immunomodulatory properties of curcumin in patients with osteoarthritis; a successful clinical trial in Iran. Int Immunopharmacol 2020, 85, 106607. [Google Scholar] [CrossRef] [PubMed]
- Panahi, Y.; Alishiri, G.H.; Parvin, S.; Sahebkar, A. Mitigation of Systemic Oxidative Stress by Curcuminoids in Osteoarthritis: Results of a Randomized Controlled Trial. J. Diet. Suppl. 2016, 13, 209–220. [Google Scholar] [CrossRef]
- Dai, W.; Jin, P.; Li, X.; Zhao, J.; Lan, Y.; Li, H.; Zheng, L. A carrier-free nano-drug assembled via π-π stacking interaction for the treatment of osteoarthritis. Biomed Pharmacother 2023, 164, 114881. [Google Scholar] [CrossRef]
- Yeh, C.C.; Su, Y.H.; Lin, Y.J.; Chen, P.J.; Shi, C.S.; Chen, C.N.; Chang, H.I. Evaluation of the protective effects of curcuminoid (curcumin and bisdemethoxycurcumin)-loaded liposomes against bone turnover in a cell-based model of osteoarthritis. Drug Des. Dev. Ther. 2015, 9, 2285–2300. [Google Scholar] [CrossRef]
- Ghumman, S.A.; Ijaz, A.; Noreen, S.; Aslam, A.; Kausar, R.; Irfan, A.; Latif, S.; Shazly, G.A.; Shah, P.A.; Rana, M. , et al. Formulation and Characterization of Curcumin Niosomes: Antioxidant and Cytotoxicity Studies. Pharm. (Basel Switz. ) 2023, 16. [Google Scholar] [CrossRef]
- Tang, S.; Gao, Y.; Wang, W.; Wang, Y.; Liu, P.; Shou, Z.; Yang, R.; Jin, C.; Zan, X.; Wang, C. , et al. Self-Report Amphiphilic Polymer-Based Drug Delivery System with ROS-Triggered Drug Release for Osteoarthritis Therapy. ACS Macro Lett. 2024, 13, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Kang, C.; Jung, E.; Hyeon, H.; Seon, S.; Lee, D. Acid-activatable polymeric curcumin nanoparticles as therapeutic agents for osteoarthritis. Nanomedicine 2020, 23, 102104. [Google Scholar] [CrossRef]
- Lombardi, A.F.; Ma, Y.; Jang, H.; Jerban, S.; Tang, Q.; Searleman, A.C.; Meyer, R.S.; Du, J.; Chang, E.Y. AcidoCEST-UTE MRI Reveals an Acidic Microenvironment in Knee Osteoarthritis. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- Zhang, M.; Peng, X.; Ding, Y.; Ke, X.; Ren, K.; Xin, Q.; Qin, M.; Xie, J.; Li, J. A cyclic brush zwitterionic polymer based pH-responsive nanocarrier-mediated dual drug delivery system with lubrication maintenance for osteoarthritis treatment. Mater. Horiz. 2023, 10, 2554–2567. [Google Scholar] [CrossRef] [PubMed]
- Niazvand, F.; Khorsandi, L.; Abbaspour, M.; Orazizadeh, M.; Varaa, N.; Maghzi, M.; Ahmadi, K. Curcumin-loaded poly lactic-co-glycolic acid nanoparticles effects on mono-iodoacetate -induced osteoarthritis in rats. Vet Res Forum 2017, 8, 155–161. [Google Scholar]
- Hamdalla, H.M.; Ahmed, R.R.; Galaly, S.R.; Naguib, I.A.; Alghamdi, B.S.; Ahmed, O.M.; Farghali, A.; Abdul-Hamid, M. Ameliorative Effect of Curcumin Nanoparticles against Monosodium Iodoacetate-Induced Knee Osteoarthritis in Rats. Mediat. Inflamm. 2022, 2022, 8353472. [Google Scholar] [CrossRef]
- Wang, J.; Wang, X.; Cao, Y.; Huang, T.; Song, D.X.; Tao, H.R. Therapeutic potential of hyaluronic acid/chitosan nanoparticles for the delivery of curcuminoid in knee osteoarthritis and an in vitro evaluation in chondrocytes. Int J Mol Med 2018, 42, 2604–2614. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, L.; Zhu, J.; Gu, J.; Wang, X.; Tao, H. Hyaluronic Acid Modified Curcumin-Loaded Chitosan Nanoparticles Inhibit Chondrocyte Apoptosis to Attenuate Osteoarthritis via Upregulation of Activator Protein 1 and RUNX Family Transcription Factor 2. J. Biomed. Nanotechnol. 2022, 18, 144–157. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Yin, W.; Ru, X.; Liu, C.; Song, B.; Qian, Z. Dual role of injectable curcumin-loaded microgels for efficient repair of osteoarthritic cartilage injury. Front Bioeng Biotechnol 2022, 10, 994816–994816. [Google Scholar] [CrossRef]
- Ratanavaraporn, J.; Soontornvipart, K.; Shuangshoti, S.; Shuangshoti, S.; Damrongsakkul, S. Localized delivery of curcumin from injectable gelatin/Thai silk fibroin microspheres for anti-inflammatory treatment of osteoarthritis in a rat model. 2017.
- Crivelli, B.; Bari, E.; Perteghella, S.; Catenacci, L.; Sorrenti, M.; Mocchi, M.; Faragò, S.; Tripodo, G.; Prina-Mello, A.; Torre, M.L. Silk fibroin nanoparticles for celecoxib and curcumin delivery: ROS-scavenging and anti-inflammatory activities in an in vitro model of osteoarthritis. Eur. J. Pharm. Biopharm. : Off. J. Arbeitsgemeinschaft Fur Pharm. Verfahrenstechnik E.V 2019, 137, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Yabas, M.; Orhan, C.; Er, B.; Tuzcu, M.; Durmus, A.S.; Ozercan, I.H.; Sahin, N.; Bhanuse, P.; Morde, A.A.; Padigaru, M.; et al. A Next Generation Formulation of Curcumin Ameliorates Experimentally Induced Osteoarthritis in Rats via Regulation of Inflammatory Mediators. Front Immunol 2021, 12, 609629. [Google Scholar] [CrossRef] [PubMed]
- Gugliandolo, E.A.-O.; Peritore, A.A.-O.X.; Impellizzeri, D.; Cordaro, M.A.-O.; Siracusa, R.A.-O.; Fusco, R.A.-O.; D’Amico, R.A.-O.; Paola, R.A.-O.; Schievano, C.; Cuzzocrea, S.A.-O.; et al. Dietary Supplementation with Palmitoyl-Glucosamine Co-Micronized with Curcumin Relieves Osteoarthritis Pain and Benefits Joint Mobility. LID - 10.3390/ani10101827 [doi] LID - 1827. 2020.
- Della Rocca, G.; Schievano, C.; Di Salvo, A.; Conti, M.B.; Della Valle, M.F. Palmitoyl-glucosamine co-micronized with curcumin for maintenance of meloxicam-induced pain relief in dogs with osteoarthritis pain. BMC Vet. Res. 2023, 19, 37. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Lee, C.K.; Song, S.H.; Yun, J.H.; Lee, A.; Park, H.J. Highly bioavailable curcumin powder suppresses articular cartilage damage in rats with mono-iodoacetate (MIA)-induced osteoarthritis. Food Sci Biotechnol 2020, 29, 251–263. [Google Scholar] [CrossRef]
- Nakagawa, Y.; Mukai, S.; Yamada, S.; Matsuoka, M.; Tarumi, E.; Hashimoto, T.; Tamura, C.; Imaizumi, A.; Nishihira, J.; Nakamura, T. Short-term effects of highly-bioavailable curcumin for treating knee osteoarthritis: a randomized, double-blind, placebo-controlled prospective study. J. Orthop. Sci. : Off. J. Jpn. Orthop. Assoc. 2014, 19, 933–939. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, Y.; Mukai, S.; Yamada, S.; Murata, S.; Yabumoto, H.; Maeda, T.; Akamatsu, S. The Efficacy and Safety of Highly-Bioavailable Curcumin for Treating Knee Osteoarthritis: A 6-Month Open-Labeled Prospective Study. Clin. Med. Insights. Arthritis Musculoskelet. Disord. 2020, 13, 1179544120948471. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, Y.; Mori, K.; Yamada, S.; Mukai, S.; Hirose, A.; Nakamura, R. The Oral Administration of Highly-Bioavailable Curcumin for One Year Has Clinical and Chondro-Protective Effects: A Randomized, Double-Blinded, Placebo-Controlled Prospective Study. Arthrosc. Sports Med. Rehabil. 2022, 4, e393–e402. [Google Scholar] [CrossRef]
- Hashemzadeh, K.; Davoudian, N.; Jaafari, R.M.; Mirfeizi, Z. The Effect of Nanocurcumin in Improvement of Knee Osteoarthritis: A Randomized Clinical Trial. Curr. Rheumatol. Rev. 2020, 16, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Atabaki, M.; Shariati-Sarabi, Z.; Tavakkol-Afshari, J.; Taghipour, A.; Jafari, M.R.; Nikpoor, A.R.; Mohammadi, M. Curcumin as an effective suppressor of miRNA expression in patients with knee osteoarthritis. Avicenna J. Phytomedicine 2022, 12, 346–356. [Google Scholar] [CrossRef]
- Gupte, P.A.; Giramkar, S.A.; Harke, S.M.; Kulkarni, S.K.; Deshmukh, A.P.; Hingorani, L.L.; Mahajan, M.P.; Bhalerao, S.S. Evaluation of the efficacy and safety of Capsule Longvida(®) Optimized Curcumin (solid lipid curcumin particles) in knee osteoarthritis: a pilot clinical study. J Inflamm Res 2019, 12, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Baharizade, M.; Ghetmiri, S.I.; Mohammady, M.; Mohammadi-Samani, S.; Yousefi, G. Revolutionizing Knee Osteoarthritis Treatment: Innovative Self-Nano-Emulsifying Polyethylene Glycol Organogel of Curcumin for Effective Topical Delivery. AAPS PharmSciTech 2024, 25, 80. [Google Scholar] [CrossRef]
- Deng, W.; He, Q.; Zhang, W. Analysis of the mechanism of curcumin against osteoarthritis using metabolomics and transcriptomics. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2024, 397, 3313–3329. [Google Scholar] [CrossRef]
- Swallow, J.; Seidler, K.; Barrow, M. The mechanistic role of curcumin on matrix metalloproteinases in osteoarthritis. Fitoterapia 2024, 174, 105870. [Google Scholar] [CrossRef]
- Rao, C.V. Regulation of COX and LOX by curcumin. Adv Exp Med Biol 2007, 595, 213–226. [Google Scholar] [CrossRef]
- Shep, D.; Khanwelkar, C.; Gade, P.; Karad, S. Safety and efficacy of curcumin versus diclofenac in knee osteoarthritis: a randomized open-label parallel-arm study. Trials 2019, 20, 214. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Jung, J.; Lee, M.; Kim, M.; Kang, N.; Kim, O.K.; Lee, J. Curcuma longa L. extract exhibits anti-inflammatory and cytoprotective functions in the articular cartilage of monoiodoacetate-injected rats. Food Nutr. Res. 2024, 68. [Google Scholar] [CrossRef] [PubMed]
- Mathy-Hartert, M.; Jacquemond-Collet, I.; Priem, F.; Sanchez, C.; Lambert, C.; Henrotin, Y. Curcumin inhibits pro-inflammatory mediators and metalloproteinase-3 production by chondrocytes. Inflammation research : official journal of the European Histamine Research Society... [et al. 2009, 58, 899–908. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Egan, B.; Wang, J. Epigenetic mechanisms underlying the aberrant catabolic and anabolic activities of osteoarthritic chondrocytes. Int. J. Biochem. Cell Biol. 2015, 67, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Theleman, J.L.; Lygrisse, K.A.; Wang, J. Epigenetic Mechanisms Underlying the Aging of Articular Cartilage and Osteoarthritis. Gerontology 2019, 65, 387–396. [Google Scholar] [CrossRef]
- Huber, L.C.; Brock, M.; Hemmatazad, H.; Giger, O.T.; Moritz, F.; Trenkmann, M.; Distler, J.H.; Gay, R.E.; Kolling, C.; Moch, H.; et al. Histone deacetylase/acetylase activity in total synovial tissue derived from rheumatoid arthritis and osteoarthritis patients. Arthritis Rheum 2007, 56, 1087–1093. [Google Scholar] [CrossRef] [PubMed]
- Furumatsu, T.; Tsuda, M.; Yoshida, K.; Taniguchi, N.; Ito, T.; Hashimoto, M.; Ito, T.; Asahara, H. Sox9 and p300 cooperatively regulate chromatin-mediated transcription. J. Biol. Chem. 2005, 280, 35203–35208. [Google Scholar] [CrossRef]
- Liu, C.J.; Zhang, Y.; Xu, K.; Parsons, D.; Alfonso, D.; Di Cesare, P.E. Transcriptional activation of cartilage oligomeric matrix protein by Sox9, Sox5, and Sox6 transcription factors and CBP/p300 coactivators. Front. Biosci. : A J. Virtual Libr. 2007, 12, 3899–3910. [Google Scholar] [CrossRef]
- Wan, C.; Zhang, F.; Yao, H.; Li, H.; Tuan, R.S. Histone Modifications and Chondrocyte Fate: Regulation and Therapeutic Implications. Front. Cell Dev. Biol. 2021, 9, 626708. [Google Scholar] [CrossRef]
- Huang, L.; Li, P.; Guo, L.; Li, L.; Yuan, J.; Zhao, R.; Li, H.; Wei, X. Zinc finger protein 521 attenuates osteoarthritis via the histone deacetylases 4 in the nucleus. Bioengineered 2022, 13, 14489–14502. [Google Scholar] [CrossRef]
- Hong, S.; Derfoul, A.; Pereira-Mouries, L.; Hall, D.J. A novel domain in histone deacetylase 1 and 2 mediates repression of cartilage-specific genes in human chondrocytes. Faseb J 2009, 23, 3539–3552. [Google Scholar] [CrossRef]
- Meng, F.; Li, Z.; Zhang, Z.; Yang, Z.; Kang, Y.; Zhao, X.; Long, D.; Hu, S.; Gu, M.; He, S.; et al. MicroRNA-193b-3p regulates chondrogenesis and chondrocyte metabolism by targeting HDAC3. Theranostics 2018, 8, 2862–2883. [Google Scholar] [CrossRef]
- Higashiyama, R.; Miyaki, S.; Yamashita, S.; Yoshitaka, T.; Lindman, G.; Ito, Y.; Sasho, T.; Takahashi, K.; Lotz, M.; Asahara, H. Correlation between MMP-13 and HDAC7 expression in human knee osteoarthritis. Mod. Rheumatol. 2010, 20, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Bradley, E.W.; Carpio, L.R.; Olson, E.N.; Westendorf, J.J. Histone deacetylase 7 (Hdac7) suppresses chondrocyte proliferation and β-catenin activity during endochondral ossification. J. Biol. Chem. 2015, 290, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.M.; Haqqi, T.M. Epigenetics in osteoarthritis: Potential of HDAC inhibitors as therapeutics. Pharmacol Res 2018, 128, 73–79. [Google Scholar] [CrossRef]
- Wang, J.; Wang, X.; Ding, X.; Huang, T.; Song, D.; Tao, H. EZH2 is associated with cartilage degeneration in osteoarthritis by promoting SDC1 expression via histone methylation of the microRNA-138 promoter. Lab. Investig. ; A J. Tech. Methods Pathol. 2021, 101, 600–611. [Google Scholar] [CrossRef]
- Chen, L.; Wu, Y.; Wu, Y.; Wang, Y.; Sun, L.; Li, F. The inhibition of EZH2 ameliorates osteoarthritis development through the Wnt/β-catenin pathway. Sci Rep 2016, 6, 29176. [Google Scholar] [CrossRef] [PubMed]
- Camilleri, E.T.; Dudakovic, A.; Riester, S.M.; Galeano-Garces, C.; Paradise, C.R.; Bradley, E.W.; McGee-Lawrence, M.E.; Im, H.J.; Karperien, M.; Krych, A.J. , et al. Loss of histone methyltransferase Ezh2 stimulates an osteogenic transcriptional program in chondrocytes but does not affect cartilage development. J. Biol. Chem. 2018, 293, 19001–19011. [Google Scholar] [CrossRef]
- Allas, L.; Brochard, S.; Rochoux, Q.; Ribet, J.; Dujarrier, C.; Veyssiere, A.; Aury-Landas, J.; Grard, O.; Leclercq, S.; Vivien, D. , et al. EZH2 inhibition reduces cartilage loss and functional impairment related to osteoarthritis. Sci Rep 2020, 10, 19577. [Google Scholar] [CrossRef]
- Shao, R.; Suo, J.; Zhang, Z.; Kong, M.; Ma, Y.; Wen, Y.; Liu, M.; Zhuang, L.; Ge, K.; Bi, Q. , et al. H3K36 methyltransferase NSD1 protects against osteoarthritis through regulating chondrocyte differentiation and cartilage homeostasis. Cell Death Differ. 2024, 31, 106–118. [Google Scholar] [CrossRef]
- Thulson, E.; Davis, E.S.; D’Costa, S.; Coryell, P.R.; Kramer, N.E.; Mohlke, K.L.; Loeser, R.F.; Diekman, B.O.; Phanstiel, D.H. 3D chromatin structure in chondrocytes identifies putative osteoarthritis risk genes. Genetics 2022, 222. [Google Scholar] [CrossRef] [PubMed]
- Bittner, N.; Shi, C.; Zhao, D.; Ding, J.; Southam, L.; Swift, D.; Kreitmaier, P.; Tutino, M.; Stergiou, O.; Cheung, J.T.S. , et al. Primary osteoarthritis chondrocyte map of chromatin conformation reveals novel candidate effector genes. Ann. Rheum. Dis. 2024. [Google Scholar] [CrossRef]
- Alvarez-Garcia, O.; Fisch, K.M.; Wineinger, N.E.; Akagi, R.; Saito, M.; Sasho, T.; Su, A.I.; Lotz, M.K. Increased DNA Methylation and Reduced Expression of Transcription Factors in Human Osteoarthritis Cartilage. Arthritis Rheumatol. (Hoboken N.J.) 2016, 68, 1876–1886. [Google Scholar] [CrossRef] [PubMed]
- Rushton, M.D.; Reynard, L.N.; Barter, M.J.; Refaie, R.; Rankin, K.S.; Young, D.A.; Loughlin, J. Characterization of the cartilage DNA methylome in knee and hip osteoarthritis. Arthritis Rheumatol. (Hoboken N.J.) 2014, 66, 2450–2460. [Google Scholar] [CrossRef] [PubMed]
- Lafont, J.E.; Moustaghfir, S.; Durand, A.-L.; Mallein-Gerin, F. The epigenetic players and the chromatin marks involved in the articular cartilage during osteoarthritis. Front. Physiol. 2023, 14. [Google Scholar] [CrossRef]
- Imagawa, K.; de Andrés, M.C.; Hashimoto, K.; Itoi, E.; Otero, M.; Roach, H.I.; Goldring, M.B.; Oreffo, R.O. Association of reduced type IX collagen gene expression in human osteoarthritic chondrocytes with epigenetic silencing by DNA hypermethylation. Arthritis Rheumatol. (Hoboken N.J.) 2014, 66, 3040–3051. [Google Scholar] [CrossRef]
- Kim, K.I.; Park, Y.S.; Im, G.I. Changes in the epigenetic status of the SOX-9 promoter in human osteoarthritic cartilage. J. Bone Miner. Res. : Off. J. Am. Soc. Bone Miner. Res. 2013, 28, 1050–1060. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Xiao, L.; Chen, J.; Chen, X.; Chen, X.; Yao, S.; Li, H.; Zhao, G.; Ma, J. DNA methylation is involved in the pathogenesis of osteoarthritis by regulating CtBP expression and CtBP-mediated signaling. Int. J. Biol. Sci. 2020, 16, 994–1009. [Google Scholar] [CrossRef]
- Iliopoulos, D.; Malizos, K.N.; Tsezou, A. Epigenetic regulation of leptin affects MMP-13 expression in osteoarthritic chondrocytes: possible molecular target for osteoarthritis therapeutic intervention. Ann. Rheum. Dis. 2007, 66, 1616–1621. [Google Scholar] [CrossRef]
- Jeffries, M.A.; Donica, M.; Baker, L.W.; Stevenson, M.E.; Annan, A.C.; Beth Humphrey, M.; James, J.A.; Sawalha, A.H. Genome-Wide DNA Methylation Study Identifies Significant Epigenomic Changes in Osteoarthritic Subchondral Bone and Similarity to Overlying Cartilage. Arthritis Rheumatol. (Hoboken N.J.) 2016, 68, 1403–1414. [Google Scholar] [CrossRef]
- Yang, J.; Wang, N. Genome-wide expression and methylation profiles reveal candidate genes and biological processes underlying synovial inflammatory tissue of patients with osteoarthritis. Int. J. Rheum. Dis. 2015, 18, 783–790. [Google Scholar] [CrossRef]
- Fernández-Tajes, J.; Soto-Hermida, A.; Vázquez-Mosquera, M.E.; Cortés-Pereira, E.; Mosquera, A.; Fernández-Moreno, M.; Oreiro, N.; Fernández-López, C.; Fernández, J.L.; Rego-Pérez, I. , et al. Genome-wide DNA methylation analysis of articular chondrocytes reveals a cluster of osteoarthritic patients. Ann. Rheum. Dis. 2014, 73, 668–677. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Li, L.; Liu, X.; Tian, J.; Zheng, W.; Li, J.; Wang, L. Genome-wide analysis of aberrant methylation of enhancer DNA in human osteoarthritis. BMC Med. Genom. 2020, 13, 1. [Google Scholar] [CrossRef] [PubMed]
- Papathanasiou, I.; Trachana, V.; Mourmoura, E.; Tsezou, A. DNA methylation regulates miR-140-5p and miR-146a expression in osteoarthritis. Life Sci 2019, 228, 274–284. [Google Scholar] [CrossRef]
- Fisch, K.M.; Gamini, R.; Alvarez-Garcia, O.; Akagi, R.; Saito, M.; Muramatsu, Y.; Sasho, T.; Koziol, J.A.; Su, A.I.; Lotz, M.K. Identification of transcription factors responsible for dysregulated networks in human osteoarthritis cartilage by global gene expression analysis. Osteoarthr. Cartil. 2018, 26, 1531–1538. [Google Scholar] [CrossRef]
- Yue, J.; Aobulikasimu, A.; Sun, W.; Liu, S.; Xie, W.; Sun, W. Targeted regulation of FoxO1 in chondrocytes prevents age-related osteoarthritis via autophagy mechanism. J Cell Mol Med 2022, 26, 3075–3082. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Zhang, S.; Previn, R.; Chen, D.; Jin, Y.; Zhou, G. Role of Forkhead Box O Transcription Factors in Oxidative Stress-Induced Chondrocyte Dysfunction: Possible Therapeutic Target for Osteoarthritis? Int J Mol Sci 2018, 19. [Google Scholar] [CrossRef]
- Matsuzaki, T.; Alvarez-Garcia, O.; Mokuda, S.; Nagira, K.; Olmer, M.; Gamini, R.; Miyata, K.; Akasaki, Y.; Su, A.I.; Asahara, H. , et al. FoxO transcription factors modulate autophagy and proteoglycan 4 in cartilage homeostasis and osteoarthritis. Sci. Transl. Med. 2018, 10. [Google Scholar] [CrossRef]
- Ohzono, H.; Hu, Y.; Nagira, K.; Kanaya, H.; Okubo, N.; Olmer, M.; Gotoh, M.; Kurakazu, I.; Akasaki, Y.; Kawata, M. , et al. Targeting FoxO transcription factors with HDAC inhibitors for the treatment of osteoarthritis. Ann. Rheum. Dis. 2023, 82, 262–271. [Google Scholar] [CrossRef]
- Ahmed, E.A.; Alzahrani, A.M. SOXC Transcription Factors as Diagnostic Biomarkers and Therapeutic Targets for Arthritis. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef]
- Wang, Y.P.; Di, W.J.; Yang, S.; Qin, S.L.; Xu, Y.F.; Han, P.F.; Hou, K.D. The association of growth differentiation factor 5 rs143383 gene polymorphism with osteoarthritis: a systematic review and meta-analysis. J. Orthop. Surg. Res. 2023, 18, 763. [Google Scholar] [CrossRef]
- Zhang, R.; Yao, J.; Xu, P.; Ji, B.; Luck, J.V.; Chin, B.; Lu, S.; Kelsoe, J.R.; Ma, J. A comprehensive meta-analysis of association between genetic variants of GDF5 and osteoarthritis of the knee, hip and hand. Inflammation research : official journal of the European Histamine Research Society... [et al.] 2015, 64, 405–414. [Google Scholar] [CrossRef] [PubMed]
- Murakami, T.; Ruengsinpinya, L.; Takahata, Y.; Nakaminami, Y.; Hata, K.; Nishimura, R. HOXA10 promotes Gdf5 expression in articular chondrocytes. Sci Rep 2023, 13, 22778. [Google Scholar] [CrossRef] [PubMed]
- Razmara, E.; Bitaraf, A.; Yousefi, H.; Nguyen, T.H.; Garshasbi, M.; Cho, W.C.; Babashah, S. Non-Coding RNAs in Cartilage Development: An Updated Review. Int J Mol Sci 2019, 20. [Google Scholar] [CrossRef] [PubMed]
- Kong, H.; Sun, M.L.; Zhang, X.A.; Wang, X.Q. Crosstalk Among circRNA/lncRNA, miRNA, and mRNA in Osteoarthritis. Front. Cell Dev. Biol. 2021, 9, 774370. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhang, H.; Xie, Q.; Feng, H.; Li, H.; Li, Z.; Yang, K.; Ding, J.; Gao, G. LncRNA-mediated cartilage homeostasis in osteoarthritis: a narrative review. Front Med (Lausanne) 2024, 11, 1326843. [Google Scholar] [CrossRef]
- Wang, R.; Shiu, H.T.; Lee, W.Y.W. Emerging role of lncRNAs in osteoarthritis: An updated review. Front Immunol 2022, 13, 982773. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, F.; Chen, G.; He, R.; Yang, L. LncRNA MALAT1 promotes osteoarthritis by modulating miR-150-5p/AKT3 axis. Cell Biosci. 2019, 9, 54. [Google Scholar] [CrossRef]
- Dang, X.; Lian, L.; Wu, D. The diagnostic value and pathogenetic role of lncRNA-ATB in patients with osteoarthritis. Cell. Mol. Biol. Lett. 2018, 23, 55. [Google Scholar] [CrossRef]
- Wang, Q.; Yang, J.; Pan, R.; Zha, Z. LncRNA SNHG1 overexpression alleviates osteoarthritis via activating PI3K/Akt signal pathway and suppressing autophagy. Immunobiology 2024, 229, 152799. [Google Scholar] [CrossRef]
- Gu, X.; Xie, T. LncRNA AC005165.1 Alleviates IL-1β-Induced Osteoarthritis via miR-199a-3p/TXNIP Axis. Biochem. Genet. 2024. [Google Scholar] [CrossRef]
- Ma, X.; Cai, D.; Zhu, Y.; Zhao, Y.; Shang, X.; Wang, C.; Zhang, H.; Bian, A.; Yu, H.; Cheng, W. L-Glutamine alleviates osteoarthritis by regulating lncRNA-NKILA expression through the TGF-β1/SMAD2/3 signalling pathway. Clinical science (London, England : 1979) 2022, 136, 1053–1069. [Google Scholar] [CrossRef]
- Losko, M.; Kotlinowski, J.; Jura, J. Long Noncoding RNAs in Metabolic Syndrome Related Disorders. Mediat. Inflamm. 2016, 2016, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Cao, Y.; Chang, C.; Huang, W.; Su, S.; Peng, Z.; Zhang, J. Knockdown of circSOD2 ameliorates osteoarthritis progression via the miR-224-5p/PRDX3 axis. J. Orthop. Surg. Res. 2023, 18, 432. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Wang, Y.; Wu, X. Knockdown of Circ_0037658 Alleviates IL-1β-Induced Osteoarthritis Progression by Serving as a Sponge of miR-665 to Regulate ADAMTS5. Front. Genet. 2022, 13, 886898. [Google Scholar] [CrossRef] [PubMed]
- Qian, W.; Mei, K.; Zhu, L.; Chu, Y.; Lv, J.; Yun, C. Circ_0044235 regulates the development of osteoarthritis by the modulation of miR-375/PIK3R3 axis. J. Orthop. Surg. Res. 2024, 19, 241. [Google Scholar] [CrossRef] [PubMed]
- Li, H.Z.; Lin, Z.; Xu, X.H.; Lin, N.; Lu, H.D. The potential roles of circRNAs in osteoarthritis: a coming journey to find a treasure. Biosci. Rep. 2018, 38. [Google Scholar] [CrossRef]
- Lian, W.S.; Ko, J.Y.; Wu, R.W.; Sun, Y.C.; Chen, Y.S.; Wu, S.L.; Weng, L.H.; Jahr, H.; Wang, F.S. MicroRNA-128a represses chondrocyte autophagy and exacerbates knee osteoarthritis by disrupting Atg12. Cell Death Dis 2018, 9, 919. [Google Scholar] [CrossRef]
- Cao, Y.; Tang, S.; Nie, X.; Zhou, Z.; Ruan, G.; Han, W.; Zhu, Z.; Ding, C. Decreased miR-214-3p activates NF-κB pathway and aggravates osteoarthritis progression. EBioMedicine 2021, 65, 103283. [Google Scholar] [CrossRef]
- Li, F.; Yao, J.; Hao, Q.; Duan, Z. miRNA-103 promotes chondrocyte apoptosis by down-regulation of Sphingosine kinase-1 and ameliorates PI3K/AKT pathway in osteoarthritis. Biosci. Rep. 2019, 39. [Google Scholar] [CrossRef]
- Kopańska, M.; Szala, D.; Czech, J.; Gabło, N.; Gargasz, K.; Trzeciak, M.; Zawlik, I.; Snela, S. MiRNA expression in the cartilage of patients with osteoarthritis. J. Orthop. Surg. Res. 2017, 12, 51. [Google Scholar] [CrossRef]
- Zheng, W.D.; Zhou, F.L.; Lin, N.; Liu, J. Investigation for the role of CTX-III and microRNA-98 in diagnosis and treatment of osteoarthritis. Eur Rev Med Pharmacol Sci 2018, 22, 5424–5428. [Google Scholar] [CrossRef]
- Song, J.; Jin, E.H.; Kim, D.; Kim, K.Y.; Chun, C.H.; Jin, E.J. MicroRNA-222 regulates MMP-13 via targeting HDAC-4 during osteoarthritis pathogenesis. BBA Clin. 2015, 3, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Li, Y.Y.; Ma, J.; Pei, F.X. Roles of microRNA and signaling pathway in osteoarthritis pathogenesis. J. Zhejiang University. Science. B 2016, 17, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Shorter, E.; Avelar, R.; Zachariou, M.; Spyrou, G.M.; Raina, P.; Smagul, A.; Ashraf Kharaz, Y.; Peffers, M.; Goljanek-Whysall, K.; de Magalhães, J.P. , et al. Identifying Novel Osteoarthritis-Associated Genes in Human Cartilage Using a Systematic Meta-Analysis and a Multi-Source Integrated Network. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- Jiang, Y.; Shen, Y.; Ding, L.; Xia, S.; Jiang, L. Identification of transcription factors and construction of a novel miRNA regulatory network in primary osteoarthritis by integrated analysis. BMC Musculoskelet. Disord. 2021, 22, 1008. [Google Scholar] [CrossRef] [PubMed]
- Reuter, S.; Gupta, S.C.; Park, B.; Goel, A.; Aggarwal, B.B. Epigenetic changes induced by curcumin and other natural compounds. Genes Nutr. 2011, 6, 93–108. [Google Scholar] [CrossRef] [PubMed]
- Arias, C.; Salazar, L.A. Autophagy and Polyphenols in Osteoarthritis: A Focus on Epigenetic Regulation. Int J Mol Sci 2021, 23. [Google Scholar] [CrossRef]
- Joven, J.; Micol, V.; Segura-Carretero, A.; Alonso-Villaverde, C.; Menéndez, J.A. Polyphenols and the modulation of gene expression pathways: can we eat our way out of the danger of chronic disease? Crit. Rev. Food Sci. Nutr. 2014, 54, 985–1001. [Google Scholar] [CrossRef]
- Boyanapalli, S.S.S.; Huang, Y.; Su, Z.; Cheng, D.; Zhang, C.; Guo, Y.; Rao, R.; Androulakis, I.P.; Kong, A.N. Pharmacokinetics and Pharmacodynamics of Curcumin in regulating anti-inflammatory and epigenetic gene expression. Biopharm. Drug Dispos. 2018, 39, 289–297. [Google Scholar] [CrossRef]
- Cheng, D.; Li, W.; Wang, L.; Lin, T.; Poiani, G.; Wassef, A.; Hudlikar, R.; Ondar, P.; Brunetti, L.; Kong, A.N. Pharmacokinetics, Pharmacodynamics, and PKPD Modeling of Curcumin in Regulating Antioxidant and Epigenetic Gene Expression in Healthy Human Volunteers. Mol Pharm 2019, 16, 1881–1889. [Google Scholar] [CrossRef]
- Marcu, M.G.; Jung, Y.J.; Lee, S.; Chung, E.J.; Lee, M.J.; Trepel, J.; Neckers, L. Curcumin is an inhibitor of p300 histone acetylatransferase. Med. Chem. (Shariqah (United Arab Emir. )) 2006, 2, 169–174. [Google Scholar] [CrossRef]
- Yuan, Z.; Syed, M.A.; Panchal, D.; Rogers, D.; Joo, M.; Sadikot, R.T. Curcumin mediated epigenetic modulation inhibits TREM-1 expression in response to lipopolysaccharide. Int. J. Biochem. Cell Biol. 2012, 44, 2032–2043. [Google Scholar] [CrossRef]
- Hassan, F.U.; Rehman, M.S.; Khan, M.S.; Ali, M.A.; Javed, A.; Nawaz, A.; Yang, C. Curcumin as an Alternative Epigenetic Modulator: Mechanism of Action and Potential Effects. Front. Genet. 2019, 10, 514. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, C.; Zappia, J.; Lambert, C.; Foguenne, J.; Dierckxsens, Y.; Dubuc, J.-E.; Delcour, J.-P.; Gothot, A.; Henrotin, Y. Curcuma longa and Boswellia serrata Extracts Modulate Different and Complementary Pathways on Human Chondrocytes In Vitro: Deciphering of a Transcriptomic Study. Front. Pharmacol. 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.; Verma, S.S.; Rai, V.; Awasthee, N.; Chava, S.; Hui, K.M.; Kumar, A.P.; Challagundla, K.B.; Sethi, G.; Gupta, S.C. Long non-coding RNAs are emerging targets of phytochemicals for cancer and other chronic diseases. Cell Mol Life Sci 2019, 76, 1947–1966. [Google Scholar] [CrossRef] [PubMed]
- Shishodia, S.; Singh, T.; Chaturvedi, M.M. Modulation of transcription factors by curcumin. Adv Exp Med Biol 2007, 595, 127–148. [Google Scholar] [CrossRef]
- Nguyen, H.D.; Kim, M.S. The protective effects of curcumin on metabolic syndrome and its components: In-silico analysis for genes, transcription factors, and microRNAs involved. Arch. Biochem. Biophys. 2022, 727, 109326. [Google Scholar] [CrossRef]
- JZ, A.L.; AlFaris, N.A.; Al-Farga, A.M.; Alshammari, G.M.; BinMowyna, M.N.; Yahya, M.A. Curcumin reverses diabetic nephropathy in streptozotocin-induced diabetes in rats by inhibition of PKCβ/p(66)Shc axis and activation of FOXO-3a. J Nutr Biochem 2021, 87, 108515. [Google Scholar] [CrossRef]
- Li, X.; Zhu, R.; Jiang, H.; Yin, Q.; Gu, J.; Chen, J.; Ji, X.; Wu, X.; Fu, H.; Wang, H. , et al. Autophagy enhanced by curcumin ameliorates inflammation in atherogenesis via the TFEB-P300-BRD4 axis. Acta Pharm. Sinica. B 2022, 12, 2280–2299. [Google Scholar] [CrossRef]
- Xiang, L.; Nakamura, Y.; Lim, Y.M.; Yamasaki, Y.; Kurokawa-Nose, Y.; Maruyama, W.; Osawa, T.; Matsuura, A.; Motoyama, N.; Tsuda, L. Tetrahydrocurcumin extends life span and inhibits the oxidative stress response by regulating the FOXO forkhead transcription factor. Aging 2011, 3, 1098–1109. [Google Scholar] [CrossRef]
- Arora, I.; Sharma, M.; Tollefsbol, T.O. Combinatorial Epigenetics Impact of Polyphenols and Phytochemicals in Cancer Prevention and Therapy. Int J Mol Sci 2019, 20. [Google Scholar] [CrossRef]
- Yao, J.; Liu, X.; Sun, Y.; Dong, X.; Liu, L.; Gu, H. Curcumin-Alleviated Osteoarthritic Progression in Rats Fed a High-Fat Diet by Inhibiting Apoptosis and Activating Autophagy via Modulation of MicroRNA-34a. J Inflamm Res 2021, 14, 2317–2331. [Google Scholar] [CrossRef] [PubMed]
- Saber, M.M.; Mahmoud, M.M.; Amin, H.M.; Essam, R.M. Therapeutic effects of combining curcumin and swimming in osteoarthritis using a rat model. Biomed Pharmacother 2023, 166, 115309. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Stöckl, S.; Lukas, C.; Herrmann, M.; Brochhausen, C.; König, M.A.; Johnstone, B.; Grässel, S. Curcumin-primed human BMSC-derived extracellular vesicles reverse IL-1β-induced catabolic responses of OA chondrocytes by upregulating miR-126-3p. Stem Cell Res. Ther. 2021, 12, 252. [Google Scholar] [CrossRef] [PubMed]
- Qiu, B.; Xu, X.; Yi, P.; Hao, Y. Curcumin reinforces MSC-derived exosomes in attenuating osteoarthritis via modulating the miR-124/NF-kB and miR-143/ROCK1/TLR9 signalling pathways. J Cell Mol Med 2020, 24, 10855–10865. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.J.; Hou, Y.K.; Chen, M.W.; Yu, X.Z.; Chen, S.Y.; Yue, Y.R.; Guo, X.T.; Chen, J.X.; Zhou, Q. A pH-responsive metal-organic framework for the co-delivery of HIF-2α siRNA and curcumin for enhanced therapy of osteoarthritis. J Nanobiotechnology 2023, 21, 18. [Google Scholar] [CrossRef]
- Tong, K.M.; Shieh, D.C.; Chen, C.P.; Tzeng, C.Y.; Wang, S.P.; Huang, K.C.; Chiu, Y.C.; Fong, Y.C.; Tang, C.H. Leptin induces IL-8 expression via leptin receptor, IRS-1, PI3K, Akt cascade and promotion of NF-kappaB/p300 binding in human synovial fibroblasts. Cell. Signal. 2008, 20, 1478–1488. [Google Scholar] [CrossRef]
- Cai, Z.; Long, T.; Zhao, Y.; Lin, R.; Wang, Y. Epigenetic Regulation in Knee Osteoarthritis. Front. Genet. 2022, 13. [Google Scholar] [CrossRef]
- Caldo, D.; Massarini, E.; Rucci, M.; Deaglio, S.; Ferracini, R. Epigenetics in Knee Osteoarthritis: A 2020–2023 Update Systematic Review. Life 2024, 14, 269. [Google Scholar] [CrossRef]
- Che, X.; Chen, T.; Wei, L.; Gu, X.; Gao, Y.; Liang, S.; Li, P.; Shi, D.; Liang, B.; Wang, C. , et al. MicroRNA-1 regulates the development of osteoarthritis in a Col2a1-Cre-ERT2/GFPfl/fl-RFP-miR-1 mouse model of osteoarthritis through the downregulation of Indian hedgehog expression. Int J Mol Med 2020, 46, 360–370. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, Z.; Chang, Z.; Mao, G.; Hu, S.; Zeng, A.; Fu, M. miR-193b-5p regulates chondrocytes metabolism by directly targeting histone deacetylase 7 in interleukin-1β-induced osteoarthritis. J Cell Biochem 2019, 120, 12775–12784. [Google Scholar] [CrossRef]
- Mao, G.; Zhang, Z.; Huang, Z.; Chen, W.; Huang, G.; Meng, F.; Zhang, Z.; Kang, Y. MicroRNA-92a-3p regulates the expression of cartilage-specific genes by directly targeting histone deacetylase 2 in chondrogenesis and degradation. Osteoarthr. Cartil. 2017, 25, 521–532. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Zhang, X.; Li, Q. Modeling Osteoarthritis: MiR-16-5p Attenuates IL-1β Induced Chondrocyte Dysfunction by Targeting MAP2K1 through the MAPK Pathway. Ann. Clin. Lab. Sci. 2023, 53, 248–258. [Google Scholar] [PubMed]
- Ding, Y.; Wang, L.; Zhao, Q.; Wu, Z.; Kong, L. MicroRNA-93 inhibits chondrocyte apoptosis and inflammation in osteoarthritis by targeting the TLR4/NF-κB signaling pathway. Int J Mol Med 2019, 43, 779–790. [Google Scholar] [CrossRef]
- Dai, Y.; Liu, S.; Xie, X.; Ding, M.; Zhou, Q.; Zhou, X. MicroRNA-31 promotes chondrocyte proliferation by targeting C-X-C motif chemokine ligand 12. Mol Med Rep 2019, 19, 2231–2237. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.D.; Miao, W.H.; Zhang, Y.Y.; Zou, M.J.; Yan, X.F. Inhibition of miR-126 protects chondrocytes from IL-1β induced inflammation via upregulation of Bcl-2. Bone Jt. Res. 2018, 7, 414–421. [Google Scholar] [CrossRef]
- Shi, J.; Guo, K.; Su, S.; Li, J.; Li, C. miR-486-5p is upregulated in osteoarthritis and inhibits chondrocyte proliferation and migration by suppressing SMAD2. Mol Med Rep 2018, 18, 502–508. [Google Scholar] [CrossRef]
- Ji, M.L.; Jiang, H.; Wu, F.; Geng, R.; Ya, L.K.; Lin, Y.C.; Xu, J.H.; Wu, X.T.; Lu, J. Precise targeting of miR-141/200c cluster in chondrocytes attenuates osteoarthritis development. Ann. Rheum. Dis. 2021, 80, 356–366. [Google Scholar] [CrossRef]
- Zhang, L.; Peng, H.; Zhang, W.; Li, Y.; Liu, L.; Leng, T. Yeast Cell wall Particle mediated Nanotube-RNA delivery system loaded with miR365 Antagomir for Post-traumatic Osteoarthritis Therapy via Oral Route. Theranostics 2020, 10, 8479–8493. [Google Scholar] [CrossRef]
- Yan, H.; Duan, X.; Pan, H.; Holguin, N.; Rai, M.F.; Akk, A.; Springer, L.E.; Wickline, S.A.; Sandell, L.J.; Pham, C.T. Suppression of NF-κB activity via nanoparticle-based siRNA delivery alters early cartilage responses to injury. Proc Natl Acad Sci U S A 2016, 113, E6199–e6208. [Google Scholar] [CrossRef]
- Tang, X.; Muhammad, H.; McLean, C.; Miotla-Zarebska, J.; Fleming, J.; Didangelos, A.; Önnerfjord, P.; Leask, A.; Saklatvala, J.; Vincent, T.L. Connective tissue growth factor contributes to joint homeostasis and osteoarthritis severity by controlling the matrix sequestration and activation of latent TGFβ. Ann. Rheum. Dis. 2018, 77, 1372–1380. [Google Scholar] [CrossRef]
- Rai, M.F.; Pan, H.; Yan, H.; Sandell, L.J.; Pham, C.T.N.; Wickline, S.A. Applications of RNA interference in the treatment of arthritis. Transl. Res. : J. Lab. Clin. Med. 2019, 214, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Conte, R.; Finicelli, M.; Borrone, A.; Margarucci, S.; Peluso, G.; Calarco, A.; Bosetti, M. MMP-2 Silencing through siRNA Loaded Positively-Charged Nanoparticles (AcPEI-NPs) Counteracts Chondrocyte De-Differentiation. Polymers 2023, 15. [Google Scholar] [CrossRef]
- Bedingfield, S.K.; Colazo, J.M.; Yu, F.; Liu, D.D.; Jackson, M.A.; Himmel, L.E.; Cho, H.; Crofford, L.J.; Hasty, K.A.; Duvall, C.L. Amelioration of post-traumatic osteoarthritis via nanoparticle depots delivering small interfering RNA to damaged cartilage. Nat. Biomed. Eng. 2021, 5, 1069–1083. [Google Scholar] [CrossRef] [PubMed]
- Bedingfield, S.K.; Colazo, J.M.; Di Francesco, M.; Yu, F.; Liu, D.D.; Di Francesco, V.; Himmel, L.E.; Gupta, M.K.; Cho, H.; Hasty, K.A. , et al. Top-Down Fabricated microPlates for Prolonged, Intra-articular Matrix Metalloproteinase 13 siRNA Nanocarrier Delivery to Reduce Post-traumatic Osteoarthritis. ACS Nano 2021, 15, 14475–14491. [Google Scholar] [CrossRef]
- Yan, Y.; Lu, A.; Dou, Y.; Zhang, Z.; Wang, X.Y.; Zhai, L.; Ai, L.Y.; Du, M.Z.; Jiang, L.X.; Zhu, Y.J. , et al. Nanomedicines Reprogram Synovial Macrophages by Scavenging Nitric Oxide and Silencing CA9 in Progressive Osteoarthritis. Adv. Sci. (Weinh. Baden-Wurtt. Ger. ) 2023, 10, e2207490. [Google Scholar] [CrossRef]
- Kondreddy, V.; Banerjee, R.; Devi, B.; Muralidharan, K.; Piramanayagam, S. Inhibition of the MALT1-LPCAT3 axis protects cartilage degeneration and osteoarthritis. Cell Commun. Signal. : CCS 2024, 22, 189. [Google Scholar] [CrossRef]
- Gabbianelli, R.; Bordoni, L.; Morano, S.; Calleja-Agius, J.; Lalor, J.G. Nutri-Epigenetics and Gut Microbiota: How Birth Care, Bonding and Breastfeeding Can Influence and Be Influenced? Int J Mol Sci 2020, 21. [Google Scholar] [CrossRef] [PubMed]
- Hao, X.; Shang, X.; Liu, J.; Chi, R.; Zhang, J.; Xu, T. The gut microbiota in osteoarthritis: where do we stand and what can we do? Arthritis Res Ther 2021, 23, 42. [Google Scholar] [CrossRef]
- Jeyaraman, M.; Ram, P.R.; Jeyaraman, N.; Yadav, S. The Gut-Joint Axis in Osteoarthritis. Cureus 2023, 15, e48951. [Google Scholar] [CrossRef]
- Favazzo, L.J.; Hendesi, H.; Villani, D.A.; Soniwala, S.; Dar, Q.A.; Schott, E.M.; Gill, S.R.; Zuscik, M.J. The gut microbiome-joint connection: implications in osteoarthritis. Curr. Opin. Rheumatol. 2020, 32, 92–101. [Google Scholar] [CrossRef]
- Wei, Z.; Li, F.; Pi, G. Association Between Gut Microbiota and Osteoarthritis: A Review of Evidence for Potential Mechanisms and Therapeutics. Front. Cell. Infect. Microbiol. 2022, 12, 812596. [Google Scholar] [CrossRef]
- Yu, X.H.; Yang, Y.Q.; Cao, R.R.; Bo, L.; Lei, S.F. The causal role of gut microbiota in development of osteoarthritis. Osteoarthr. Cartil. 2021, 29, 1741–1750. [Google Scholar] [CrossRef] [PubMed]
- Gleason, B.; Chisari, E.; Parvizi, J. Osteoarthritis Can Also Start in the Gut: The Gut-Joint Axis. Indian J. Orthop. 2022, 56, 1150–1155. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Ding, W.; Wang, H.L.; Dai, L.L.; Zong, W.H.; Wang, Y.Z.; Bi, J.; Han, W.; Dong, G.J. Gut microbiota and obesity-associated osteoarthritis. Osteoarthr. Cartil. 2019, 27, 1257–1265. [Google Scholar] [CrossRef]
- Amabebe, E.; Robert, F.O.; Agbalalah, T.; Orubu, E.S.F. Microbial dysbiosis-induced obesity: role of gut microbiota in homoeostasis of energy metabolism. Br. J. Nutr. 2020, 123, 1127–1137. [Google Scholar] [CrossRef]
- Berthelot, J.M.; Sellam, J.; Maugars, Y.; Berenbaum, F. Cartilage-gut-microbiome axis: a new paradigm for novel therapeutic opportunities in osteoarthritis. RMD Open 2019, 5, e001037. [Google Scholar] [CrossRef]
- Arora, V.; Singh, G.; O-Sullivan, I.; Ma, K.; Natarajan Anbazhagan, A.; Votta-Velis, E.G.; Bruce, B.; Richard, R.; van Wijnen, A.J.; Im, H.-J. Gut-microbiota modulation: The impact of thegut-microbiotaon osteoarthritis. Gene 2021, 785, 145619. [Google Scholar] [CrossRef]
- Biver, E.; Berenbaum, F.; Valdes, A.M.; Araujo de Carvalho, I.; Bindels, L.B.; Brandi, M.L.; Calder, P.C.; Castronovo, V.; Cavalier, E.; Cherubini, A. , et al. Gut microbiota and osteoarthritis management: An expert consensus of the European society for clinical and economic aspects of osteoporosis, osteoarthritis and musculoskeletal diseases (ESCEO). Ageing Res. Rev. 2019, 55, 100946. [Google Scholar] [CrossRef]
- Li, D.; Li, Y.; Yang, S.; Lu, J.; Jin, X.; Wu, M. Diet-gut microbiota-epigenetics in metabolic diseases: From mechanisms to therapeutics. Biomed Pharmacother 2022, 153, 113290. [Google Scholar] [CrossRef]
- Cuevas-Sierra, A.; Ramos-Lopez, O.; Riezu-Boj, J.I.; Milagro, F.I.; Martinez, J.A. Diet, Gut Microbiota, and Obesity: Links with Host Genetics and Epigenetics and Potential Applications. Adv. Nutr. (Bethesda Md.) 2019, 10, S17–s30. [Google Scholar] [CrossRef]
- Remely, M.; Aumueller, E.; Merold, C.; Dworzak, S.; Hippe, B.; Zanner, J.; Pointner, A.; Brath, H.; Haslberger, A.G. Effects of short chain fatty acid producing bacteria on epigenetic regulation of FFAR3 in type 2 diabetes and obesity. Gene 2014, 537, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Bhat, M.I.; Kapila, R. Dietary metabolites derived from gut microbiota: critical modulators of epigenetic changes in mammals. Nutr Rev 2017, 75, 374–389. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, S.; Singh, A.; Sandeep, K.; Yadav, D. Epigenetic Regulation of Gut Microbial Dysbiosis. Indian J. Microbiol. 2021, 61, 125–129. [Google Scholar] [CrossRef] [PubMed]
- Woo, V.; Alenghat, T. Epigenetic regulation by gut microbiota. Gut Microbes 2022, 14, 2022407. [Google Scholar] [CrossRef] [PubMed]
- Marín-Tello, C.; Jintaridth, P.; Sanchez, F.; González, C.; Zelada-Castillo, L.; Vásquez-Arqueros, A.; Guevara-Vásquez, A.; Vieira, A. Epigenetic regulation by metabolites from the gut microbiome. Benef. Microbes 2022, 13, 437–444. [Google Scholar] [CrossRef]
- Berni Canani, R.; Di Costanzo, M.; Leone, L. The epigenetic effects of butyrate: potential therapeutic implications for clinical practice. Clin. Epigenetics 2012, 4, 4. [Google Scholar] [CrossRef]
- Fellows, R.; Denizot, J.; Stellato, C.; Cuomo, A.; Jain, P.; Stoyanova, E.; Balázsi, S.; Hajnády, Z.; Liebert, A.; Kazakevych, J. , et al. Microbiota derived short chain fatty acids promote histone crotonylation in the colon through histone deacetylases. Nat. Commun. 2018, 9, 105. [Google Scholar] [CrossRef]
- Alenghat, T.; Osborne, L.C.; Saenz, S.A.; Kobuley, D.; Ziegler, C.G.; Mullican, S.E.; Choi, I.; Grunberg, S.; Sinha, R.; Wynosky-Dolfi, M. , et al. Histone deacetylase 3 coordinates commensal-bacteria-dependent intestinal homeostasis. Nature 2013, 504, 153–157. [Google Scholar] [CrossRef]
- Liu, K.; He, X.; Huang, J.; Yu, S.; Cui, M.; Gao, M.; Liu, L.; Qian, Y.; Xie, Y.; Hui, M. , et al. Short-chain fatty acid-butyric acid ameliorates granulosa cells inflammation through regulating METTL3-mediated N6-methyladenosine modification of FOSL2 in polycystic ovarian syndrome. Clin. Epigenetics 2023, 15, 86. [Google Scholar] [CrossRef]
- Noureldein, M.H.; Bitar, S.; Youssef, N.; Azar, S.; Eid, A.A. Butyrate modulates diabetes-linked gut dysbiosis: epigenetic and mechanistic modifications. J. Mol. Endocrinol. 2020, 64, 29–42. [Google Scholar] [CrossRef]
- Kumar, H.; Lund, R.; Laiho, A.; Lundelin, K.; Ley, R.E.; Isolauri, E.; Salminen, S. Gut microbiota as an epigenetic regulator: pilot study based on whole-genome methylation analysis. mBio 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Molina, B.; Sánchez-Alcoholado, L.; Cabrera-Mulero, A.; Lopez-Dominguez, R.; Carmona-Saez, P.; Garcia-Fuentes, E.; Moreno-Indias, I.; Tinahones, F.J. Gut Microbiota Composition Is Associated With the Global DNA Methylation Pattern in Obesity. Front. Genet. 2019, 10, 613. [Google Scholar] [CrossRef] [PubMed]
- Tahara, T.; Hirata, I.; Nakano, N.; Tahara, S.; Horiguchi, N.; Kawamura, T.; Okubo, M.; Ishizuka, T.; Yamada, H.; Yoshida, D. , et al. Potential link between Fusobacterium enrichment and DNA methylation accumulation in the inflammatory colonic mucosa in ulcerative colitis. Oncotarget 2017, 8, 61917–61926. [Google Scholar] [CrossRef] [PubMed]
- Ansari, I.; Raddatz, G.; Gutekunst, J.; Ridnik, M.; Cohen, D.; Abu-Remaileh, M.; Tuganbaev, T.; Shapiro, H.; Pikarsky, E.; Elinav, E. , et al. The microbiota programs DNA methylation to control intestinal homeostasis and inflammation. Nat. Microbiol. 2020, 5, 610–619. [Google Scholar] [CrossRef]
- Shi, W.; Cassmann, T.J.; Bhagwate, A.V.; Hitosugi, T.; Ip, W.K.E. Lactic acid induces transcriptional repression of macrophage inflammatory response via histone acetylation. Cell Rep 2024, 43, 113746. [Google Scholar] [CrossRef]
- Remely, M.; Aumueller, E.; Jahn, D.; Hippe, B.; Brath, H.; Haslberger, A.G. Microbiota and epigenetic regulation of inflammatory mediators in type 2 diabetes and obesity. Benef. Microbes 2014, 5, 33–43. [Google Scholar] [CrossRef]
- Fitzgerald, K.N.; Hodges, R.; Hanes, D.; Stack, E.; Cheishvili, D.; Szyf, M.; Henkel, J.; Twedt, M.W.; Giannopoulou, D.; Herdell, J. , et al. Potential reversal of epigenetic age using a diet and lifestyle intervention: a pilot randomized clinical trial. Aging 2021, 13, 9419–9432. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhao, Q.; Li, T.; Lu, L.; Wang, F.; Zhang, H.; Liu, Z.; Ma, H.; Zhu, Q.; Wang, J. , et al. Lactobacillus plantarum-derived indole-3-lactic acid ameliorates colorectal tumorigenesis via epigenetic regulation of CD8(+) T cell immunity. Cell Metab 2023, 35, 943–960.e949. [Google Scholar] [CrossRef]
- Novakovic, B.; Habibi, E.; Wang, S.Y.; Arts, R.J.W.; Davar, R.; Megchelenbrink, W.; Kim, B.; Kuznetsova, T.; Kox, M.; Zwaag, J. , et al. β-Glucan Reverses the Epigenetic State of LPS-Induced Immunological Tolerance. Cell 2016, 167, 1354–1368.e1314. [Google Scholar] [CrossRef]
- Wu, S.E.; Hashimoto-Hill, S.; Woo, V.; Eshleman, E.M.; Whitt, J.; Engleman, L.; Karns, R.; Denson, L.A.; Haslam, D.B.; Alenghat, T. Microbiota-derived metabolite promotes HDAC3 activity in the gut. Nature 2020, 586, 108–112. [Google Scholar] [CrossRef]
- Guan, Z.; Jin, X.; Guan, Z.; Liu, S.; Tao, K.; Luo, L. The gut microbiota metabolite capsiate regulate SLC2A1 expression by targeting HIF-1α to inhibit knee osteoarthritis-induced ferroptosis. Aging Cell 2023, 22, e13807. [Google Scholar] [CrossRef] [PubMed]
- Rice, S.J.; Beier, F.; Young, D.A.; Loughlin, J. Interplay between genetics and epigenetics in osteoarthritis. Nat. Rev.. Rheumatol. 2020, 16, 268–281. [Google Scholar] [CrossRef] [PubMed]
- Coutinho de Almeida, R.; Ramos, Y.F.M.; Meulenbelt, I. Involvement of epigenetics in osteoarthritis. Best Pract. Res.. Clin. Rheumatol. 2017, 31, 634–648. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Zhuang, X.; Xiang, X.; Liu, Y.; Zhang, S.; Liu, C.; Barnes, S.; Grizzle, W.; Miller, D.; Zhang, H.G. A novel nanoparticle drug delivery system: the anti-inflammatory activity of curcumin is enhanced when encapsulated in exosomes. Mol. Ther. : J. Am. Soc. Gene Ther. 2010, 18, 1606–1614. [Google Scholar] [CrossRef]
- Trivedi, M.K.; Mondal, S.C.; Gangwar, M.; Jana, S. Immunomodulatory potential of nanocurcumin-based formulation. Inflammopharmacology 2017, 25, 609–619. [Google Scholar] [CrossRef]
- Ghosh, S.S.; He, H.; Wang, J.; Gehr, T.W.; Ghosh, S. Curcumin-mediated regulation of intestinal barrier function: The mechanism underlying its beneficial effects. Tissue Barriers 2018, 6, e1425085. [Google Scholar] [CrossRef] [PubMed]
- Bertoncini-Silva, C.; Fassini, P.G.; Carlos, D.; de Paula, N.A.; Ramalho, L.N.Z.; Rodrigues Giuliani, M.; Pereira Í, S.; Guimarães, J.B.; Suen, V.M.M. The Dose-Dependent Effect of Curcumin Supplementation on Inflammatory Response and Gut Microbiota Profile in High-Fat Fed C57BL/6 Mice. Mol Nutr Food Res 2023, 67, e2300378. [Google Scholar] [CrossRef]
- Pluta, R.; Januszewski, S.; Ułamek-Kozioł, M. Mutual Two-Way Interactions of Curcumin and Gut Microbiota. Int J Mol Sci 2020, 21. [Google Scholar] [CrossRef]
- Scazzocchio, B.; Minghetti, L.; D’Archivio, M. Interaction between Gut Microbiota and Curcumin: A New Key of Understanding for the Health Effects of Curcumin. Nutrients 2020, 12. [Google Scholar] [CrossRef]
Figure 1.
Nanotherapeutics in osteoarthritis. Nanoparticle-based therapy uses polymer, lipid, and inorganic materials, such as encapsulated carriers with drugs or bioactive, to ameliorate osteoarthritis. A representative micelle structure (central) displays functional attributes of these nanoparticles. The yellow circle depicts a lipid-based nanocarrier, while the pink and green circles show inorganic and polymeric nanoformulation carriers.
Figure 1.
Nanotherapeutics in osteoarthritis. Nanoparticle-based therapy uses polymer, lipid, and inorganic materials, such as encapsulated carriers with drugs or bioactive, to ameliorate osteoarthritis. A representative micelle structure (central) displays functional attributes of these nanoparticles. The yellow circle depicts a lipid-based nanocarrier, while the pink and green circles show inorganic and polymeric nanoformulation carriers.
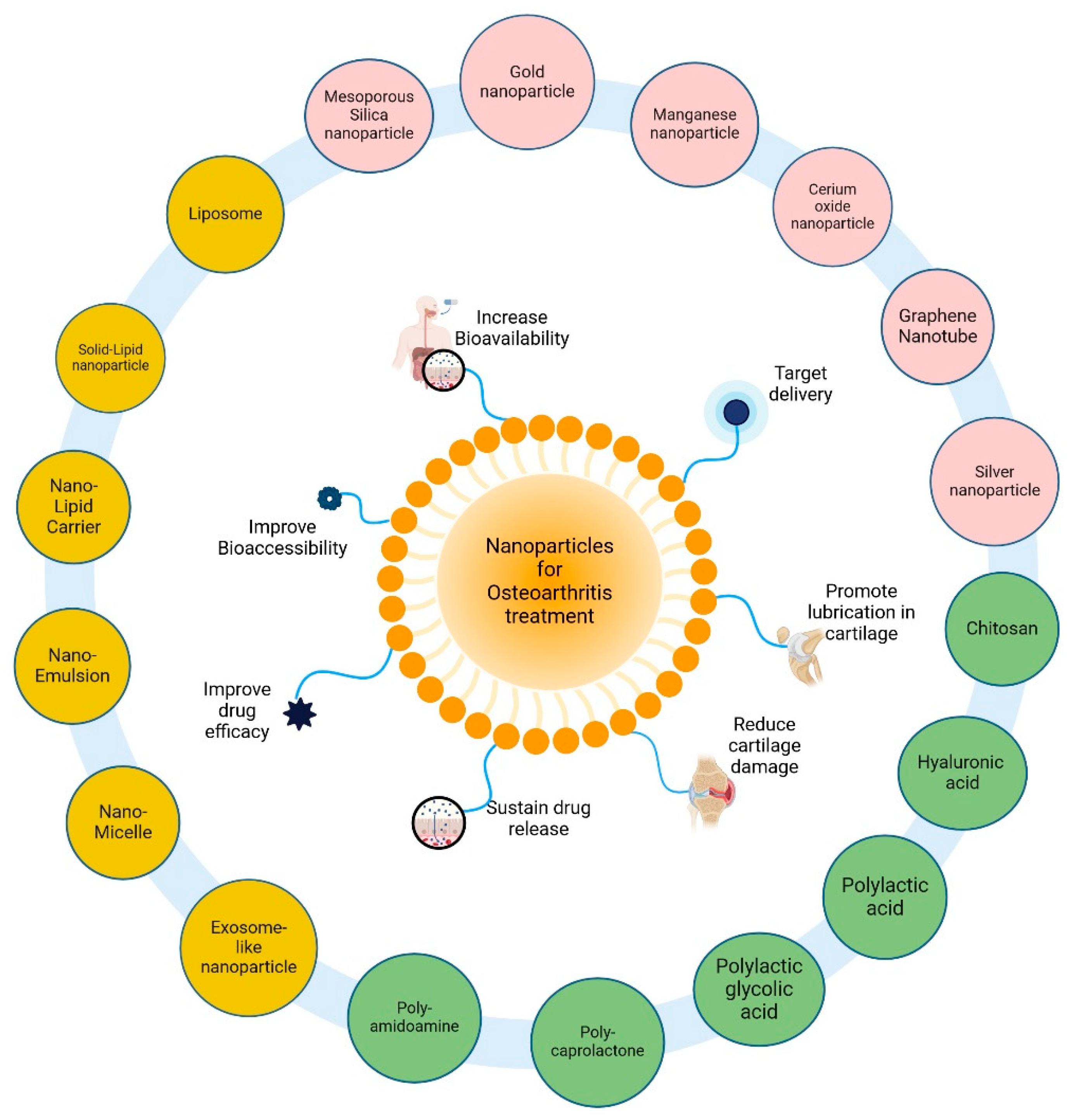
Figure 2.
Epigenetic regulation in osteoarthritis. The role of epigenetic modulation in osteoarthritis is emerging and paving the way to new epigenetic targets for disease treatment. During osteoarthritis progression, modifications in histone acetyltransferase, histone deacetylase, DNA hypomethylation, DNA hypermethylation and upregulated lncRNA collectively result in chondrocyte apoptosis, degradation of extracellular matrix, activation of inflammasomes and inflammatory cytokines. Similarly, during disease remission, lncRNAs, and DNA methyltransferases are downregulated and certain histone deacetylases, and histone acetyltransferases are inhibited which subsequently downregulate cytokine activation and reduce cartilage damage by modulation of ECM proteins via several pathways. OA – Osteoarthritis; ECM – Extracellular matrix; lncRNA – long non-coding RNA; PVT1 – Plasmacytoma variant translocation 1; MALAT1 – Metastasis-associated lung adenocarcinoma transcript 1; HOTAIR – HOX antisense intergenic RNA; HAT – Histone acetyltransferase; HDAC – Histone deacetylase; CBP – CREB binding protein; COL2A1 – Collagen type 2, alpha-1; COL9A1 – Collagen type 9, alpha-1; COL10/COLX – Collagen 10; SOX9 – SRY-box transcription factor 9; COMP – Cartilage oligomeric matrix protein; CtBP – C-terminal binding protein; NLRP3 - Nucleotide-binding domain (NOD) - like receptor protein 3; IL6 – Interleukin 6; IL10 – Interleukin 10; TNFα – Tumor necrosis factor alpha; IL8 – Interleukin 8; Bcl2 – B-cell leukemia/lymphoma 2; Bax – Bcl-2 associated X protein; LC3B – Light chain 3B; MMP – Matrix metalloproteinases; ACAN – Aggrecan; ADAMTS – A disintegrin and metalloproteinase with thrombospondin motif 5; COX2 – Cyclooxygenase-2; DNMT – DNA methyltransferase; Nrf2 – Nuclear factor erythroid 2 - related factor 2; IFN – Interferon; WIF1 – Wnt inhibitory factor 1.
Figure 2.
Epigenetic regulation in osteoarthritis. The role of epigenetic modulation in osteoarthritis is emerging and paving the way to new epigenetic targets for disease treatment. During osteoarthritis progression, modifications in histone acetyltransferase, histone deacetylase, DNA hypomethylation, DNA hypermethylation and upregulated lncRNA collectively result in chondrocyte apoptosis, degradation of extracellular matrix, activation of inflammasomes and inflammatory cytokines. Similarly, during disease remission, lncRNAs, and DNA methyltransferases are downregulated and certain histone deacetylases, and histone acetyltransferases are inhibited which subsequently downregulate cytokine activation and reduce cartilage damage by modulation of ECM proteins via several pathways. OA – Osteoarthritis; ECM – Extracellular matrix; lncRNA – long non-coding RNA; PVT1 – Plasmacytoma variant translocation 1; MALAT1 – Metastasis-associated lung adenocarcinoma transcript 1; HOTAIR – HOX antisense intergenic RNA; HAT – Histone acetyltransferase; HDAC – Histone deacetylase; CBP – CREB binding protein; COL2A1 – Collagen type 2, alpha-1; COL9A1 – Collagen type 9, alpha-1; COL10/COLX – Collagen 10; SOX9 – SRY-box transcription factor 9; COMP – Cartilage oligomeric matrix protein; CtBP – C-terminal binding protein; NLRP3 - Nucleotide-binding domain (NOD) - like receptor protein 3; IL6 – Interleukin 6; IL10 – Interleukin 10; TNFα – Tumor necrosis factor alpha; IL8 – Interleukin 8; Bcl2 – B-cell leukemia/lymphoma 2; Bax – Bcl-2 associated X protein; LC3B – Light chain 3B; MMP – Matrix metalloproteinases; ACAN – Aggrecan; ADAMTS – A disintegrin and metalloproteinase with thrombospondin motif 5; COX2 – Cyclooxygenase-2; DNMT – DNA methyltransferase; Nrf2 – Nuclear factor erythroid 2 - related factor 2; IFN – Interferon; WIF1 – Wnt inhibitory factor 1.
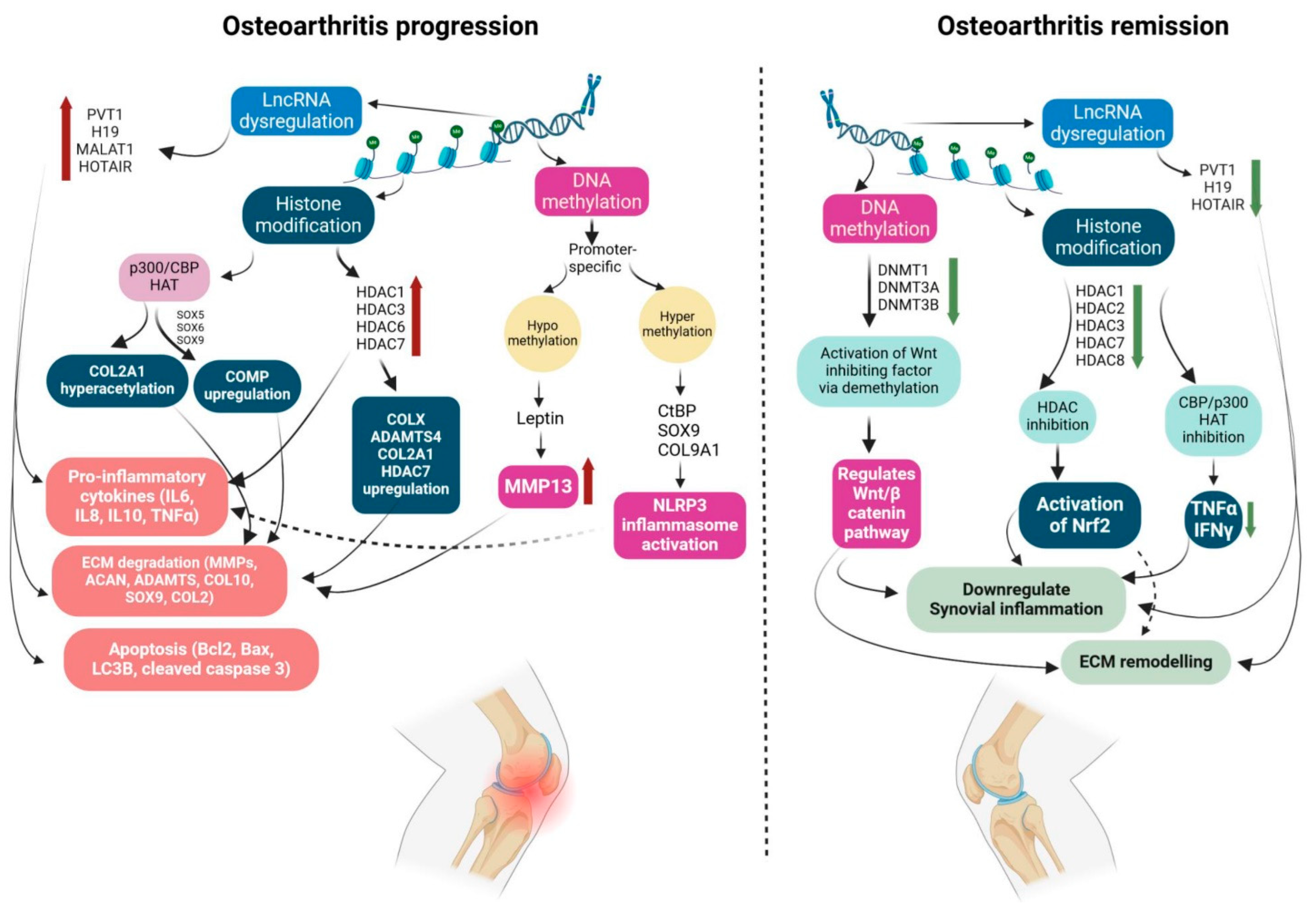
Figure 3.
Potential therapeutic roles of curcumin in osteoarthritis by anti-inflammatory and chondroprotective effects. Curcumin, upon ingestion, undergoes metabolic reduction and gets converted into various metabolites. Curcumin metabolites act on the gut-immune axis by promoting intestinal barrier integrity, immune function and gut microbial diversity. It upregulates tight junction proteins, and claudins and downregulates of cholesterol absorption. Curcumin reduces gut dysbiosis and thus promotes the production of microbial metabolites like lactic acid and SCFA. Curcumin also promotes M2 macrophage polarization and regulates T lymphocyte activity. These changes at the gut-immune axis promote the chondroprotective and anti-inflammatory effects of curcumin against osteoarthritis by inhibiting inflammasome activation, downregulating inflammatory cytokines and reducing chondrocyte apoptosis. ZO-1 - Zonula occludens-1; TJ- Tight junction proteins; LPS - lipopolysaccharide; SCFA - short chain fatty acids; IL2 - Interleukin 2; IFNγ - Interferon gamma; IL4 - Interleukin-4; IL10 - Interleukin-10; IL13 - Interleukin-13; TGFβ - Transforming growth factor beta; NFkB - Nuclear factor kappa B; TNFα - Tumor necrosis factor alpha; IL1β - Interleukin 1 beta; IL6 - Interleukin 6; MMP - Matrix metalloproteinases; NLRP3 - Nucleotide-binding domain (NOD) – like receptor protein 3.
Figure 3.
Potential therapeutic roles of curcumin in osteoarthritis by anti-inflammatory and chondroprotective effects. Curcumin, upon ingestion, undergoes metabolic reduction and gets converted into various metabolites. Curcumin metabolites act on the gut-immune axis by promoting intestinal barrier integrity, immune function and gut microbial diversity. It upregulates tight junction proteins, and claudins and downregulates of cholesterol absorption. Curcumin reduces gut dysbiosis and thus promotes the production of microbial metabolites like lactic acid and SCFA. Curcumin also promotes M2 macrophage polarization and regulates T lymphocyte activity. These changes at the gut-immune axis promote the chondroprotective and anti-inflammatory effects of curcumin against osteoarthritis by inhibiting inflammasome activation, downregulating inflammatory cytokines and reducing chondrocyte apoptosis. ZO-1 - Zonula occludens-1; TJ- Tight junction proteins; LPS - lipopolysaccharide; SCFA - short chain fatty acids; IL2 - Interleukin 2; IFNγ - Interferon gamma; IL4 - Interleukin-4; IL10 - Interleukin-10; IL13 - Interleukin-13; TGFβ - Transforming growth factor beta; NFkB - Nuclear factor kappa B; TNFα - Tumor necrosis factor alpha; IL1β - Interleukin 1 beta; IL6 - Interleukin 6; MMP - Matrix metalloproteinases; NLRP3 - Nucleotide-binding domain (NOD) – like receptor protein 3.
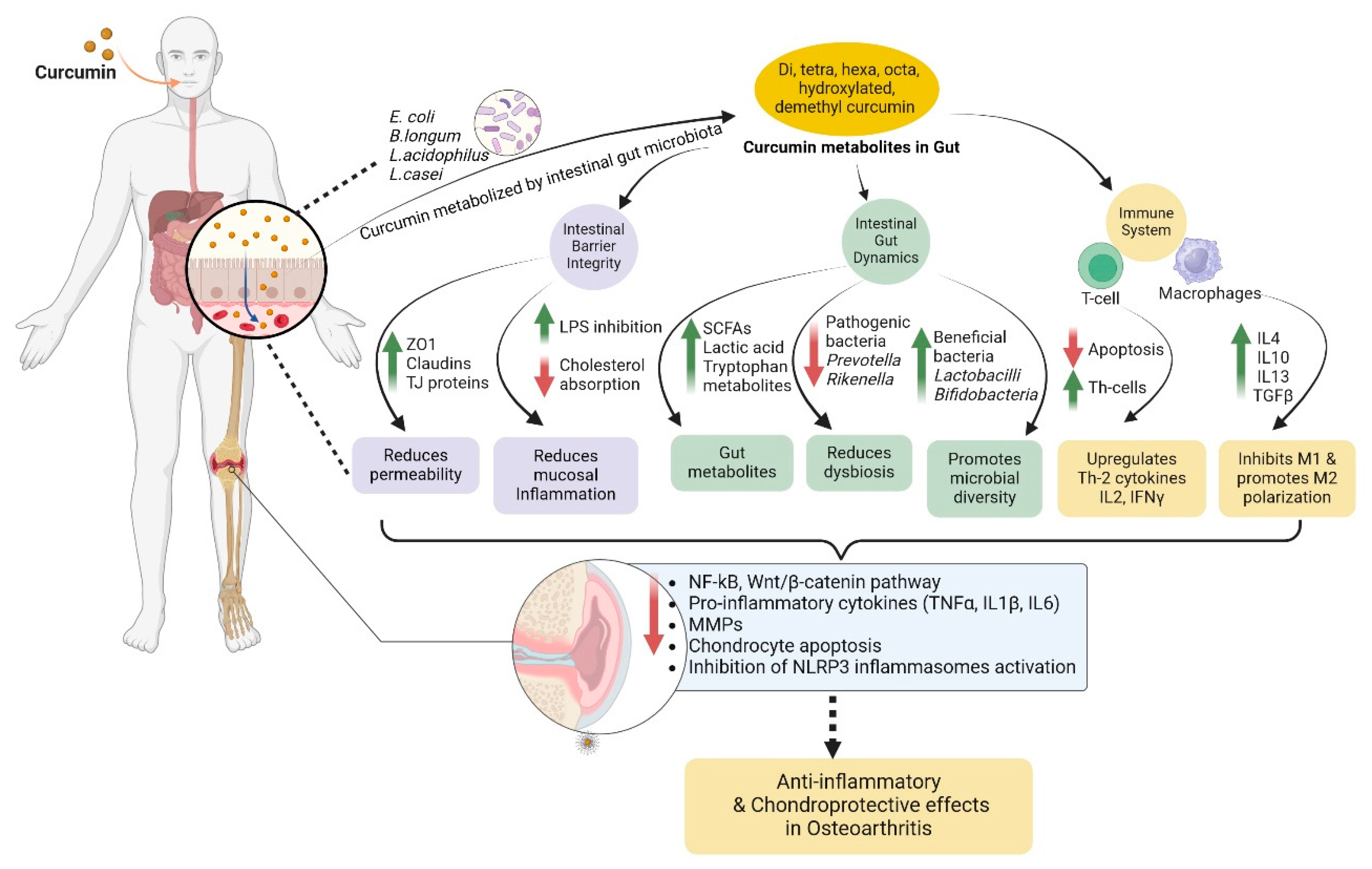

Table 1.
Evidence for the beneficial effects of polyphenol nanoformulations in osteoarthritis.
| Polyphenol nanoformulation |
Osteoarthritis model | Effects in vitro | Effects in vivo | Refs. |
|---|---|---|---|---|
| Tannic acid/Sr2+ coated Silk/graphene oxide-based meniscus scaffold | -In vitro – LPS-induced rabbit-synovial MSC -In vivo – Papain-induced OA rat model |
Increased extracellular matrix secretion and promoted cell migration | Reduced cartilage degeneration and OA damage by downregulating MMP, IL6, IL8 | [47] |
| pH responsive - metal organic framework of Hyaluronic acid loaded with protocatechuic acid |
- In vitro - IL1β-induced rat primary chondrocytes -In vivo - ALCT induced OA rat model |
-Downregulated IL6, COX2, MMP1, MMP3, MMP13, ADAMTS5 and iNOS; -Reduced synovial inflammation |
Promoted cartilage regeneration | [48] |
| Liposome-anchored teriparatide incorporated into Gallic acid-grafted gelatin hydrogel | -In vitro - IL1β-induced mouse chondrocyte cell line, ATDC5 -In vivo - DMM- induced OA mice model |
- Activated expression of p-PI3K and p-AKT - Promoted anti-apoptotic effect by upregulating Bcl-2 - Downregulated ADAMTS5 |
- Upregulated expression of ACAN, SOX9; -Promoted glycosaminoglycan secretion and ROS scavenging |
[49] |
| Gallic acid encapsulated polymeric nanoliposome | -In vitro - H2O2-induced human chondrocyte cell line, C28/I2 -In vivo - MIA- induced OA rat model |
-Promoted ROS scavenging; -Lowered cartilage damage via upregulation of aggrecan and collagen II |
-Mitigated joint wear by improving cartilage lubrication; -Lowered cartilage erosion; -Promoted ROS scavenging |
[50] |
| Gallic acid loaded liposome with hyaluronan grafted poly (2-acrylamide-2-methylpropanesulfonic acid sodium salt) | -In vitro - H2O2-induced human chondrocyte cell line, C28/I2 -In vivo - MIA- induced OA rat model |
-Upregulated expression of Col II and ACAN; -Reduced chondrocyte degeneration; -Promoted antioxidant effect |
-Lowered cartilage erosion and chondrocyte degeneration; -Promoted glycosaminoglycan deposition |
[51] |
| Polydopamine- coated Hesperetin-loaded Gd2 (CO3)3 nanoparticles |
-In vitro – IL1β – induced chondrocytes -In vivo – ACLT- induced OA mice model |
Downregulated TLR2, decreased inflammation, cellular apoptosis and promoted chondrocyte maturation by inactivating NFκB/Akt pathway | Displayed cartilage binding ability, mitigated cartilage degeneration | [52] |
| pH-responsive Polycaprolactone/polyethylene glycol naringenin nanofiber |
-In vitro – IL1β-induced primary rat chondrocytes -In vivo – ACLT-induced OA rat model |
Inhibited expression of IL6, IL1β, MMP3 and MMP13; Promoted COL2A1 | Reduced cartilage damage; increased proteoglycan retention and glycosaminoglycan content | [53] |
| Scaffold of Berberine oleanolic acid complex grafted onto Hyaluronic Acid/Silk fibroin composite |
In vitro – IL1β-induced OA model in primary rabbit articular chondrocytes |
Upregulated COL1, COL2 and SOX9; restored chondrocyte morphology |
Promoted cartilage tissue regeneration in nude mice post subcutaneous implantation | [54] |
| NIR-responsive Epigallocatechin gallate decorated Au-Ag nano-jars |
-In vitro – H2O2-induced primary rat chondrocytes -In vivo – ACLT-induced OA rat model |
Lowered chondrocyte apoptosis; downregulated p-NFκB, iNOS and COX2; promoted cell migration |
Reduced cartilage erosion; improved cartilage thickness; lowered chondrocyte apoptosis | [55] |
| Hyaluronic acid- coated gelatin nanoparticles loaded with kaempferol | -In vitro – IL1β- induced primary rat chondrocytes -In vivo – ACLT-induced OA rat model |
Downregulated expression of inflammatory cytokines – COX2, MMP9, MMP13, TNFα, IL1β; | Attenuated inflammation and matrix degradation; restored cartilage thickness | [56] |
| Poly p-Coumaric Acid Nanoparticles | In vivo – Temporomandibular joint OA rat model | -- | Inhibited chondrocyte ferroptosis; reduced ECM degradation; exhibited long-term efficacy and alleviated cartilage repair | [57] |
| Nano-naringenin |
In vivo – MIA-induced OA rat model | -- | Upregulated GSH and TIMP-3; downregulated MDA, ADAMTS5 and MMP3 | [58] |
Abbreviations: Sr2+ - Strontium ion; OA – Osteoarthritis; MMP – Matrix metalloproteinases; IL6 – Interleukin 6; IL8 – Interleukin 8; IL1β – Interleukin 1 beta; ACLT – Anterior Cruciate Ligament Transection; COX2 – Cyclooxygenase 2; ADAMTS5 – A disintegrin and metalloproteinase with thrombospondin motifs 5; iNOS – Inducible nitric oxide synthase; ATDC5 – Mouse chondrogenic cell line; DMM – Destabilization of medial meniscus; p-PI3K – Phosphorylated-Phosphoinositide 3-kinase; p-AKT – Phosphorylated protein kinase B; Bcl-2 – B cell lymphoma 2; ACAN – Aggrecan; SOX9 – SRY-box transcription factor 9; ROS – Reactive oxygen species; H2O2 – Hydrogen peroxide; C28/I2 – Human chondrocyte cell line; MIA – Monoiodoacetate; Col II – Collagen type II; Gd2(CO3)3 – Gadolinium carbonate; TLR2 – T cell receptor 2; NFκB – Nuclear factor kappa B; COL2A1 – Collagen type II alpha 1; COL – Collagen; NIR - Near infrared; Au – Gold; Ag – Silver; TNFα – Tumor necrosis factor alpha; ECM – Extracellular matrix; GSH – Glutathione; TIMP3 – Tissue inhibitor of metalloproteinase 3; MDA – Malondialdehyde.
Table 2.
Effects of curcumin nanoformulations on osteoarthritis subjects: consolidated clinical trials.
Table 2.
Effects of curcumin nanoformulations on osteoarthritis subjects: consolidated clinical trials.
| Composition | Dosage & Delivery | Duration/number of subjects (N) | Key outcomes | Refs. |
|---|---|---|---|---|
| Curcumin in water-dispersible form | 180mg/day, Oral | 8 weeks, N=50 | Reduced dependence on celecoxib vs placebo group; lower VAS scores of knee pain |
[104] |
| Curcumin in water-dispersible form | 180mg/day, Oral | 6 months, N=50 | Improved JOA, VAS and JKOM scores, observed no major side effects, 75.6% effective compared to placebo | [105] |
| Curcumin in water-dispersible form | 180mg/day, Oral | 12 months, N=50 | Reduced cartilage stiffness and time-dependent decrease in scores of JOA, VAS and JKOM | [106] |
| Curcumin nanomicelle | 80mg/day, Oral | 6 weeks, N=71 | Significant decrease in WOMAC score | [107] |
| Curcumin nanomicelle |
80mg/day, Oral | 3 months, N=30 | Decrease in VAS score, lower CRP levels, Immunomodulatory effects on T cells and B cells | [84] |
| Curcumin nanomicelle | 80mg/day, Oral | 3 months, N=30 | Suppressed expression of key miRNAs | [108] |
| Solid lipid curcumin particles | 160mg/day, Oral | 3 months, N=50 | Improved WOMAC and VAS scores comparable to ibuprofen. No significant change was observed in inflammatory markers | [109] |
| Curcumin self-nano-emulsifying-PEG organogel | 1.5g/twice per day, Topical | 8 weeks, N=75 | Significantly reduced WOMAC scores | [110] |
Abbreviations: VAS – Visual Analog Scale; JOA – Japanese Orthopaedic Association; JKOM – Japanese Knee Osteoarthritis Measure; WOMAC – Western Ontario and McMaster Universities Osteoarthritis Index; CRP – C-Reactive Protein; PEG – Polyethylene Glycol.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



