
Submitted:
20 September 2024
Posted:
23 September 2024
You are already at the latest version
Abstract
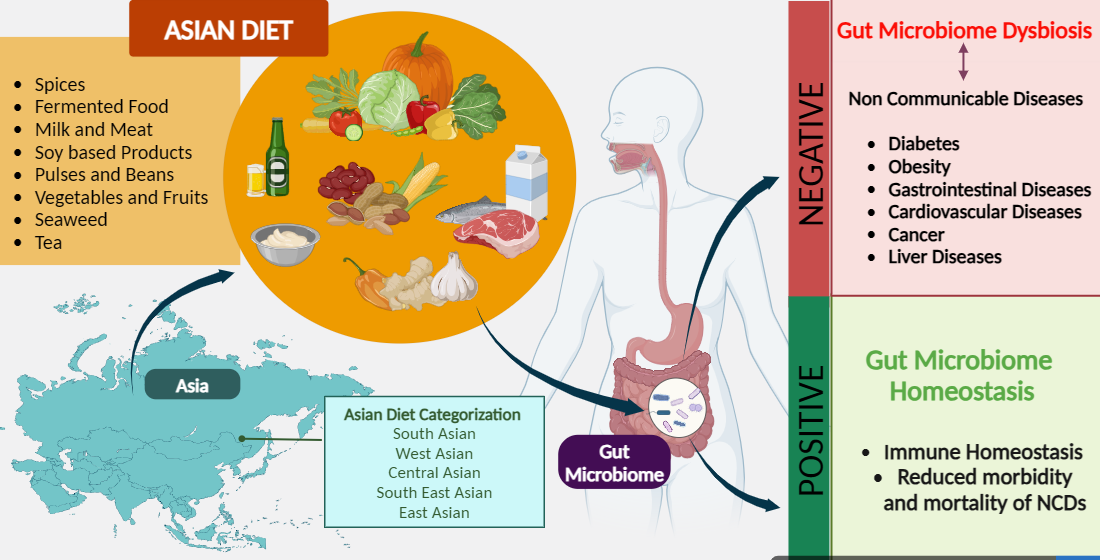
The gut microbiome known as the “hidden organ” in humans has received wide attention due to its pivotal role in human health, influencing various physiological functions and metabolic processes. Multiple factors influence the gut microbiome and diet emerging as a prominent determinant. Increasing evidence verifies different dietary patterns modulate gut microbial composition and its functionality. Asia, home to the world's largest population, showcases immense diversity, reflected in its complex dietary patterns influenced by tradition and modernization. A plethora of research evidence proves Asian diet confers beneficial effects on maintaining and improving health through favorable alteration of gut microbiome. Yet up to date, no comprehensive review exists on the influence of the Asian diet on gut microbiome. This review critically examines published data on dietary components from Eastern, Southeastern, Southern, Western and Central Asian diet and their interaction with the human gut microbiome across healthy and diseased states. It identifies strengths and limitations in Asian gut microbiome research. Asia experiences rapid dietary transitions demanding further investigation into their effects on the gut microbiota. However, evidence proves traditional Asian diets hold promise for developing personalized dietary interventions and therapeutics to promote gut health and prevent chronic diseases by modulating gut microbiome.
Keywords:
Asian diet
; diet
; gut microbiome
; gut health
; health promotion
; non-communicable diseases
1. Introduction
The traditional view of humans being a singular biological species is now subjected to a paradigm shift with the advancement of science, as humans are discovered to be hosting trillions of microorganisms in different sites of their bodies. These microorganisms containing bacteria, archaea, viruses, and eukaryotic microbes coexist with humans as a communal group benefiting each other[1]. Hence humans are considered a superorganism. Around a decade ago, most knowledge about the adult human microbiota stemmed from labor and time-consuming culture-based methods which limited the knowledge available on the microbiome. But later with the introduction of projects such as the 2001 Human Genome Project, MetaHIT (Metagenomics of the Human Intestinal Tract) project, and DNA and RNA-based microbial identification techniques (16S rRNA) supported the comprehensive phylogenetic assessment of human microbiome [2]. Based on the information that has been currently generated, the composition and density of microorganisms vary among the body sites [3]. Unlike the other organs, the gastrointestinal tract (GI tract) has a highly complex and interactive system with the host and the intestinal microbial inhabitants [2,4]. The number of microorganisms inhabiting the GI tract has been estimated to exceed 1014, which contains 10 times more bacterial cells than total no. of body cells in humans [2,5]. This community of gut microbes and their interrelationship with each other and the human has been found to impact on human wellbeing, including metabolism, physiology, nutrition, and immune function [6,7,8]. Several intrinsic and extrinsic parameters are found to be influencing the development and maintenance of the gut microbiome, where perturbation from any of those factors is indicated to be disrupting the gut microbiota (dysbiosis) [9]. Out of the above factors, diet is considered one of the major factors influencing gut microbiome [10,11]. Based on research conducted using animal models, 50% of the variation of microbial population in mice can be explained using dietary variations [12]. Diet can regulate the crosstalk between the gut microbiome and the host by which beneficial effects are conferred upon both [10]. The recent advancements of science related to microbial genomics, has made it easier to understand the effect of each dietary component including macronutrients, micronutrients, and other bioactive compounds on the gut microbiome and the outcomes of these effects on human health. A plethora of studies are available on the effect of individual dietary components on gut microbial modulation. However, from the point of view of consumption, a diet is not attributed to the ingestion of a single component but rather a complex mixture of multiple components. Therefore, finding the cumulative effect of different dietary components is essential in understanding the effect of a diet on the gut microbial population.
Asia, the home to 60% of the world's population has a huge geographic, socioeconomic, biological, and cultural diversity which is also reflected in their diet [13]. The dietary patterns of Asia are complex and are influenced by both tradition and modernization [14]. Compared to other continents, Asian diets vary remarkably within the continent. Owing to this huge variation, the Asian diet is difficult to describe under a single structure. Nevertheless, the Asian diet, in general, is characterized by high dietary fiber, vitamins, and antioxidants due to the incorporation of a diverse array of fresh plant-based foods, low concentrated fat and high carbohydrate content as rice is the staple food in many countries [15,16]. Traditional Asian diets are considered to exert beneficial effects against non-communicable diseases in preventing and reducing the risk of susceptibility or in some instances as therapeutics [17,18,19]. But the rapid economic development followed by industrialization and globalization has incurred drastic lifestyle and dietary changes in Asia characterized by increased consumption of highly processed, calorie-dense foods with high salt concentration, low fiber, and low micronutrient content [19,20,21]. This nutrient transition has resulted in a high prevalence of non-communicable diseases, notably diet-related non-communicable diseases [22,23]. Diet significantly influences the composition of the gut microbiome, and the intricate interrelation among dietary components and gut microbiota plays a crucial role in the pathogenesis of various health conditions. Therefore, dietary changes and nutrient transitions can impact on altering the gut microbiome and thereby may have an impact on occurrence of non-communicable diseases. While the full extent of the impact of gut microbiome alterations on pathogenesis remains unclear, dietary shifts in Asia have demonstrated observable alterations in the gut microbiome [24,25,26]. In this context, Asia is an ideal destination to study the interplay of the gut microbial community and diet together with their effects on host health. Yet so far, no compilation of data is available regarding the effect of the Asian diet on modulating the gut microbiome. This review critically analyses the available data on Asian dietary components and their interplay with the human gut microbiome both under healthy and disease conditions.
2. Human Gut Microbiome
The human gut microbiota has drawn attention due to its clinical significance as well as its constant and dynamic changes in the environment due to frequent interaction with substrates during food consumption. The human GI tract represents one of the largest interfaces between the host, environmental factors and microbes in the human body [2]. The human gut having an area of 200–300 m2 mucosa is found to host approximately 1011 - 1012 cells/g of luminal contents where the majority are anaerobic bacteria belonging to 50 bacterial phyla and about 100–1000 bacterial species [27,28]. All residing microorganisms in a host are collectively known as the ‘microbiota’. The collection of genes from microbiota are known as the ‘microbiome’ which in the GI tract is 150 times larger than the human genome [29,30]. Hence the gut microbiome is considered as a “hidden organ” in humans [31].
Humans are born sterile and microbial colonization begins immediately at birth and the intestinal tract is colonized by the microorganisms shortly after birth and undergo various compositional changes before stabilizing at the end of the first year of life. The development of the gut microbiome in the first year of life is influenced by several factors as the delivery mode of the baby, breast milk vs. formula feeding and cessation of milk feeding, timing of the introduction of solid foods and antibiotic usage [32]. Hereafter, up until three years of age a child’s gut microbiome develops becoming similar in profile to that of an adult and this development is based on a combination of various extrinsic and intrinsic factors such as living environment, host genetics and physiology, lifestyle, diet and consumption of drugs (antibiotics). This developed microbial profile becomes the core gut microbiome profile of a person throughout their life and is less likely to be subjected to permanent alterations unless there are long-term changes in any of the above-mentioned factor or factors [33]. Based on the identified data collected through extensive research, the dominant and the most studied gut microbial population is bacteria. For the convenience of studying, taxonomically they are classified into phyla, classes, orders, families, genera, and species [34]. So far, around 160 species of gut bacteria have been identified and they represent only few phyla out of which Firmicutes, Bacteroidetes, Actinobacteria, Proteobacteria, Fusobacteria, and Verrucomicrobia are dominating) [35] The two phyla of Firmicutes and Bacteroidetes represent 90% of the gut microbiota out of which the most abundant is Firmicutes (65%) [34]. Most of the species under the phylum Bacteroidetes belong to the genus of Bacteroides, and Prevotella. Bacterial species under the phylum Firmicutes such as Clostridium clusters IV and XIVa include the genus Clostridium, Eubacterium and Ruminococcus which are abundant in the gut [36].
Based on metagenomic data collected through extensive research, it is concluded that the composition and diversity of gut microbiome varies from one individual to another. Although microbial genes are being shared among the world population, gut microbiome indicates substantial variation among individuals making it difficult to define a healthy gut microbiota [37]. Apart from variations among individuals the microbial profile changes along the gastrointestinal tract of a human. The density and population of gut microorganisms are altered based on the chemical, nutritional and immunological gradients along the gut. This can be both longitudinal and transverse [2]. To simplify the study of the human gut microbiome, GI bacteria are classified into three enterotypes which condenses the wide variability among the population into a few categories. They are, enterotype 1 characterized by dominance of Bacteroides, Enterotype 2 Prevotella dominant group, and enterotype 3 dominated by Ruminococcus. However, there are certain debates regarding this classification of enterotypes as Liang et al. have identified a new enterotype dominated by the family Enterobacteriaceae, in a study of metagenomic analysis in Taiwan which suggested that there can be a new enterotype among Asians [39]. Apart from enterotyping for the ease of studying the structure of microbial community, is divided into alpha and beta diversity. Alpha diversity is a measure of average species diversity as richness and evenness within a habitat type at a local scale, while beta-diversity indicates the differentiation between microbial communities from different environments [40].
3. Asian Gut Microbiome
Studying the gut microbiome in Asians poses a significant challenge owing not only to the larger population but also to the considerable variability among its diverse populations. The region encompasses a vast array of ethnicities, cultures, diets, lifestyles, environmental and economic conditions contributing to substantial differences in gut microbial composition within populations. Many numbers of research, indicates that both enterotype 1 and enterotype 2 are identified among Asian populations. At the same time there are certain debates regarding this classification of enterotypes based on research conducted in Asia as a new enterotype dominated by the family Enterobacteriaceae, in a study of metagenomic analysis in Taiwan which suggested that there can be a new enterotype among Asians [39]. Based on data from Asian Microbiome project East Asian has predominantly Bacteroides enterotype and towards Southeast, Southern and Central Asia it skews towards Prevotella enterotype [41,42]. However, it should be noted that the abundance of these enterotypes can differ among the population within a country. For an example in India the prevalent enterotype among North Indian regions are Prevotella while in Southern states it’s Bacteroides and Ruminococcus [42]. In China towards Southern regions Prevotella enterotype is prominent despite other areas having Bacteroides [43]. These differences are believed to be mainly due to geographical variations in the diet and lifestyle. The bacterial genera which were abundantly found in the gut microbiome of Asians are indicated in Table 1. Many of these genera are found in Asian gut in various proportions. They are not equally distributed among the populations and the relative abundance of certain genera are used as indicators to distinguish between populations.
4. Functions of Gut Microbiome In Human
Apart from the recent technological advances, the main reason for gut microbiome getting the spotlight is due to the newfound knowledge on its complex series of functions performed in human, both in relation to health promotion and disease prevention. With encoding over 150 times more genetic information than that of the entire human genome, gut microbiome is now identified as one of the key factors in maintaining human health [51]. Hence, the gut microbiota is able to code for biochemical pathways that aren’t present in humans. The most studied association is nutrient metabolism by gut microbiome fulfilling nutrient and energy metabolism of the host. Dietary fiber, proteins and peptides which cannot be digested by humans, are digested and metabolized by gut microbial population through fermentation and anaerobic degradation producing energy and valuable metabolites [8]. These metabolites include beneficial compounds as short chain fatty acids, phenolic and indolic compounds [8,52]. Moreover, they can biosynthesize B group vitamins and vitamin K and essential amino acids[53,54]. These compounds while contributing to the nutritional needs, confer other beneficial effects such as maintaining gut physiology and contributing to host immune functions [55]. Short chain fatty acids and vitamins produced by gut bacteria are considered protective elements against the infiltration of gut pathogens and the development of pathologies [56]. Other than their produced metabolites, gut microbes themselves act as a protective barrier preventing colonization, overgrowth and persistence of pathogenic microorganisms in the gut [57]. It is achieved by commensal microorganisms via competing with pathogenic microbes for resource availability or niche opportunity which is collectively known as colonization resistance [57,58]. This mechanism of action mainly prevents infections localized in the gut. But simultaneously gut microbiota prevents infections in the other organs of the body by developing the host's innate and adaptive systemic immune system and improving the efficiency of immune responses. The role of the gut microbiome in developing the immune system is proven through several studies done on germ-free mice and gnotobiotic mice models where they both indicated high susceptibility to infections caused by pathogenic microorganisms [59,60]. Germ-free mice have shown the absence of tolerogenic dendritic cells and regulatory T cells (Tregs) that are essential in mediating immune responses and development of the immune system [61,62]. In humans, commensal microorganisms aid the differentiation of T cells which is essential for the development of the adaptive immune system [61].
The above-mentioned protective mechanisms of gut microbiome are focused on the prevention of infectious diseases. However, recent discoveries of the multi-directional interaction of gut microbiota with the organs and systems of the host have sparked a new interest in discovering the role of gut microbiome in the pathogenesis of non-communicable diseases [58]. It has been identified that gut microbiome can influence the occurrence of non-communicable gastrointestinal diseases, cardiovascular diseases (CVD), cancer, liver diseases, diabetes, liver diseases, chronic kidney diseases, mental disorders and neurological disorders [63,64,65]. However, it has not been fully understood whether gut microbes or their metabolic action leads to the occurrence of the diseases, or the disease condition creates an environment supporting dysbiosis of the gut microbiome. But in certain disease conditions such as in cardiovascular diseases, gut microbiome plays a role in the disease propensity and the presence of cardiovascular disease cause alterations in the gut microbiome as well [66]. Gut bacteria convert choline and lecithin ingested from the diet into trimethylamine (TMA) which is oxidized into trimethylamine N-oxide (TMAO) in the liver increases the risk of occurrence of cardiovascular diseases [67]. At the same time, a study conducted employing patients with atherosclerotic CVD revealed an increased abundance of Enterobacteriaceae and Streptococcus spp. in the fecal matter compared to healthy subjects. Comparatively, the abundance of common bacterial species of the gut microbiome Bacteroides spp., Prevotella copri, and Alistipes shahii were relatively depleted compared to the healthy population further indicating CVD can lead to dysbiosis of the gut microbiome [68]. Considering the role played by the gut microbiome on human physiology, having a healthy and balanced gut microbiome becomes essential in maintaining the human health. Even though the exact definition of a healthy microbiome has not been discovered, any perturbation leading to dysbiosis of the microbial profile can increase the pathogenesis and dysregulation of the body’s physiological processes [69]. Hence, it is important to identify which factors have the potency to develop and maintain the gut microbiome profile.
5. Factors Affecting on Gut Microbiome
After extensive research, it has been found that a person’s gut microbiome is unique to an individual although there are shared similarities among people belonging to the same family, geographical location, and culture [70,71,72,73]. Yet so far, no identical gut microbiota among two people has been discovered [74,75]. Even two identical twins who share closely related genetics and environment do not indicate an identical gut microbiome profile. A study conducted among 1126 twins, where the data revealed that there was no significant similarity between the gut microbiome profile of monozygotic nor dizygotic twins [3]. These results can be interpreted in two aspects. One is that even between monozygotic twins the gut microbial profiles aren’t similar, which means the likelihood of having the same gut microbial profile between two individuals is less. Secondly, genetic inheritance is not a major factor affecting shaping the gut microbial profile. Hence researchers highlight that gut microbiome composition is determined not just by a single factor but a complex interplay of a variety of factors which are host genetics, early environment, and immediate environment [33]. These factors affect a whole to development and shaping of the microbiome profile of adult humans. Although it is accepted that once developed, permanent alteration of the gut microbiome of an adult is difficult, there is evidence that certain perturbations can alter the dynamics of the microbiome profile [76].
5.1. Effect of Diet on the Gut Microbiome
Out of the factors discussed, diet is considered to be one of the major factors influencing the development and maintenance of gut microbiome [11,77]. Certain studies indicate that gut microbiota shares similarities in people belonging to the same nationality, geographical location, and race [49,70,78]. However, the similarities shown in gut microbiome are due to similar lifestyles and similar dietary preferences among people sharing the same geographical or sociocultural context [49]. A study conducted on 214 Malaysian citizens belonging to three different ethnicities indicated a variation in gut microbiome composition between different ethnic groups while similarities were shown within the same ethnic group. Considering the fact all the participants were chosen from the same geographical location, it is the ethnicity that has played a role in determining the gut microbiome composition. As per the above study, diet and lifestyle are determining factors for variation across ethnicities [79]. This was further strengthened by research done on human and mammalian gut microbiomes. A group of scientists conducted an analysis on the gut microbiome of humans and 59 mammalian species were based on the microbial composition species were able to cluster into groups. These clusters were correlated to their dietary patterns, where the clusters represented carnivores, omnivores and herbivores [80]. This further established the fact that diet is a determining factor for microbial composition not only in humans but in other mammalian species as well [81,82]. Another example is found in Asia where a higher prevalence of porphyranase and agarase-encoding genes are found in the gut microbiome of Japanese and Chinese populations than in North American and European populations. The reason for this difference is, consumption of Nori a red alga by the Japanese and Chinese as an essential part of their diet. Zobellia galactanivorans, a bacteria inhabiting in Nori has transferred these protein breakdown genes to Bacteroides plebeius, a resident bacterium in the human gut whilst passing through the gastrointestinal tract [83]. Hence diet is considered a major influencing factor that affects on modulation of gut microbiome as well as maintaining the gut homeostasis in adults [1].
According to [84] the interventions on diet that lead to alterations of gut microbiome are of three types. One is short-term dietary changes. Core bacterial taxa are resilient to most temporary outside influences. However, short-term, dramatic dietary interventions have the ability to rapidly alter microbiota diversity in humans, though these alterations are transient and do not persist for more than a few days [33,85]. For long-lasting bacterial implantation and proliferation, continuous substrate availability with a habitual diet is required [82], which is confirmed by the fact that enterotypes mentioned earlier are highly correlated to long-term dietary patterns. The second method is long-term dietary interventions. Long-term dietary interventions can alter both the gut microbiome and the host physiology. Long-term dietary interventions are gaining popularity as they are used as dietary therapies in correcting metabolism during metabolic diseases and in the prevention of such diseases. These dietary interventions include caloric restrictions as in consumption of low carbohydrate and low-fat diets, alternate-day fasting (ADF), Buchinger fasting programs and water-only fasting or time-restricted interventions as Intermittent fasting or Ramadan fasting [86,87]. Many studies conducted on both healthy and diseased human subjects have proven that caloric-restricted diets have the ability to improve gut microbial profile and increase the abundance of commensal bacteria [87,88,89]. A study conducted on obese mice indicated results that feeding low calorie diet for 8 weeks could improve other physiological functions in mice as reducing fat accumulation, improving glucose tolerance and delay immune senescence other than altering the gut microbiome [89,90]. The effect of intermittent and Ramadan fasting on gut microbiome are discussed under the South Asian dietary patter as fasting is practiced as a religious practice in these regions. The third method of diet intervention is the incorporation of special dietary compounds to influence changes in microbes. Simply put it is the introduction of probiotics or prebiotics to the diet. The habitually consumed diet compounds with dietary supplements with prebiotics or/and probiotic effects can be introduced to strengthen the intestinal microbiome. The two terms ‘prebiotics’ and ‘probiotics’ are synonyms for gut health. The prebiotic concept was introduced by Gibson and Roberfroid which is defined as a “non-digestible food ingredient that beneficially affects the host by selectively stimulating the growth and/or activity of one or a limited number of bacteria in the colon”[91]. They can reach the large intestine without being absorbed nor digested in the upper gastrointestinal tract and undergo fermentation within the colon that will confer beneficial effects on gut microbiome [36]. Prebiotics identified so far consist of soluble fibers such as resistant starch, pectin, gums and oligosaccharide carbohydrates, including fructooligosaccharides, galactooligosaccharides, xylooligosaccharides, and glucooligosaccharides [92]. To be considered a prebiotic the selected compound has to be neither hydrolysed nor absorbed by the small intestine, needs to be a substrate for a limited number of beneficial bacteria, to be capable of altering the microbiota of the colon for a beneficial microbiome and need to induce beneficial effects in the host intestine [93,94,95]. For example, consumption of resistant starch-rich diet increases the abundance of Ruminococcus bromii an amylolytic bacteria in the gut of obese men [96]. During the fermentation of prebiotics in the gut metabolites as short chain fatty acids can make beneficial functional and structural changes in intestine and other organs [94,97,98]. Probiotics on the other hand as defined by Food and Agricultural Organization of the United Nations and the World Health Organization, are ‘living microorganisms, which when administered in adequate amounts confer health benefits on the host’ [99]. The beneficial action of probiotics includes manipulation of intestinal microbial communities by proliferation of beneficial microbes and suppression of pathogens, immunomodulation, stimulation of epithelial cell proliferation and differentiation and fortification of the intestinal barrier [100,101]. Specifically, certain probiotics can have beneficial effects against metabolic syndrome. When both prebiotics and probiotics are present as a mixture it is known as synbiotics that synergistically work on improving the gut microbiota [102].
6. Asian Diet
Asia the largest and the most populous continent on earth comprises 48 countries and spreads across 44 million square kilometers of land while being home to 60 % of the total world population [13]. Asia spans a vast area, encompassing diverse landscapes, climates, and geographical features. From the arid deserts of the Middle East to the lush rainforests of Southeast Asia, the continent's geography plays a crucial role in shaping its ecosystems and influencing human societies. It is a mosaic of culture, traditions, and languages. As well as inhibit a wide array of socioeconomic diversity. Asia, with its vast and varied climatic conditions, has given rise to a unique and sophisticated system of cultivation and farming practices. Intertwined with the rich tapestry of Asian cultures and varying socioeconomic conditions this agricultural diversity has, in turn, shaped a wide range of dietary patterns across the continent. The influences on diets are complex and include person-level considerations such as income, taste, preference, and culture. They also include external, or “food environment” influences, including prices, marketing, retail availability, and food safety [14]. Hence it is unjustifiable to use one common diet to represent the whole region in scientific studies. Even within the same country, there are instances where dietary patterns greatly vary among different communities. According to a study done on the dietary patterns of India which has the world’s largest population, it has been identified that altogether there are 41 patterns of diets existing only within India where the variances are mainly based on the geographical region of the country [103]. Moreover, with the recent economic boom in Asia followed by urbanization and globalization, a rapid change in diets is observed within the region. Still, a significant part of the population is following the traditional Asian diet and with the volatility of the nutritional transition existing trend can vary over a short period [103].
Hence, in this study the dietary patterns are identified based on the geographical regions of Asia as divided by geographical literature. Being situated in the same geographical conditions leading to similar agricultural and farming practices, together with cultural influences exerted by colonization and economic power play has contributed to a certain degree of uniformity in dietary patterns within these geographical regions [104]. In this review, the dietary pattern in Asia is being studied based on the geographical sub regions developed by The United Nations Statistics Division, as indicated in Table 2 [105].
7. East Asian and Southeast Asian diet
East Asia and Southeast Asia are two distinctive regions. However, there are major similarities in diet between the two regions, except for certain differences in some ingredients such as spices and condiments which vary based on the type of cuisine. These similarities exist as a result due to the influence of China through marketing and labor migration to the Southeast Asian countries. Vietnam, which is geographically categorized as a Southeast Asian country, is known as “Little China” which emphasizes the influence of China on the
East Asian countries [11]. Moreover, Vietnam is even considered part of East Asian countries which are altogether known as “the chopstick sphere” due to their distinctive culture in food intake [12]. Hence under this review, both the East Asian and Southeast Asian countries are discussed under one section and the major disparities are discussed separately. Similar to South Asia their staple food is rice. Besides that, these two regions are characterized by the consumption of high number of vegetables and fruits, fermented food, soya and soy-based food, seaweed and marine food and tea compared to other regions of the world.
7.1. Fermented Food in the Diet
Southeast Asia and East Asia are uniquely characterized by the inclusion of fermented food in their diet. In contrast to only limiting to fermented alcohol or fermented milk products in the other parts of the world, East and Southeast Asian regions showcase a wide range of fermented food products from fermented vegetables used as a side dish in South Korea and fermented fish sauce used as a condiment in Thailand.
Fermented foods have been identified to have beneficial effects on human health for a long period of time. However, the interest in the effect of fermented food on gut microbiome has been gaining popularity in recent times. Fermentation is a food processing technique, that has an origin of immemorial times. The definition of fermentation according to the International Scientific Probiotic and Prebiotic Association (ISAPP) is “foods made through desired microbial growth and enzymatic conversions of food components”[109]. Almost all categories of foods vegetables, fruits, cereals, pulses, legumes, meat, fish and milk are subjected to fermentation to produce a wide array of products. The variability among the products is determined by the substrate used for fermentation, the type of microorganism used for fermentation and the fermenting conditions [110]. Historically fermentation was used as a mode of preservation. But over time fermented food gained popularity owing to its altered organoleptic profile than the original product, improved nutritional content and beneficial effect on human health [111]. Most of East and Southeast Asian cuisines that include fermented food products are traditionally produced and consumed on a small scale at household levels, but in recent years certain products have been commercially produced. Fermented foods affect the gut microbiome through different pathways. They contain live microorganisms which are probiotics that were either purposefully added during the fermentation process or can grow in fermented food products [112]. These microorganisms are able to either restore a disturbed gut microbiome or prevent the proliferation of pathogenic bacteria. Moreover, fermented food products include metabolites produced from microbial fermentation. Such as, short chain fatty acids and organic acids can modulate gut immune function and thereby affect host-microbiota interactions. [113,114]
Asian traditional fermented foods are generally fermented by lactic acid bacteria such as Lactobacillus plantarum, L. pentosus, L. brevis, L. fermentum, L. casei, Leuconostoc mesenteroides, L. fallax, Weissella confusa, W. koreenis, W. cibaria, and Pediococcus pentosaceus, that are considered as the probiotic source of the food practice [115]. Lactic acid fermentation can occur in vegetables when conditions are suitable (anaerobic conditions, suitable temperature, and humidity and salt concentrations) and Southeast and East Asia are popular for their range of fermented vegetable products. Kimchi one of the fermented vegetable products made prominently by fermentation of Napa cabbages (Brassica rapa) or radishes (Raphanus raphanistrum) is considered one of the most popular and daily consumed fermented dishes in East Asia, especially in South Korea [116]. Studies conducted on the health benefits of kimchi are mounting hence kimchi is even considered as a medicinal food [117]. Having medicinal effects associated with the presence of beneficial live microorganisms has brought Kimchi into the arena of gut microbiome studies. A plethora of studies bring evidence that kimchi can positively alternate gut microbiome under diseases conditions and promote a health microbial profile in healthy adults. The effect of Kimchi consumption on modulating the gut microbial profile has been studied using both humans and animal models [116,118].
In a healthy adult consumption of kimchi increases beneficial bacteria such as Faecalibacterium and Bifidobacterium and a decrease in harmful bacteria such as Clostridium and Escherichia coli. It also increases the abundance of short-chain fatty acid production-related genera (Faecalibacterium, Roseburia, and Phascolactobacterium) [119]. In obese patients, consumption of kimchi has altered gut microbiome profile by increasing the relative abundance of Bacteroides and Prevotella while decreasing of Blautia. This alteration of the gut microbiome relates to the enterotype of lean subjects [120]. In the same study a significant negative correlation between Bifidobacterium longum a lactic acid bacteria present in fermented kimchi and waist circumference was observed. Even though there are a considerable amount of data available on popular products such as Kimchi, certain products such as fermented fish sauce, and kombucha lack information regarding their effect on gut microbiome. One study conducted using Kombucha on mice who were induced with non-alcoholic fatty liver disease (NFLD) through feeding methionine and choline-deficient diet indicated, after consumption of Kombucha, abundance of Allobaculum and Turicibacter in the gut microbiome were reduced. These two genera are related to pathogenesis of NFLD. Moreover, initially absent Lactobacillus were emerged in the gut after feeding of Kombutcha [121]. Durian a native fruit found in Southeast Asia, is fermented to produce tempoyak paste which has been studied for the presence of probiotic strains and have shown that several strains of Lactobacillus bacteria isolated from tempoyak have strong probiotic characteristics [122,123,124]
Fermented fish products are a significant source of protein, forming an integral part of the staple diet, and possessing great cultural significance in many Southeast and East Asian countries compared to other parts of the world which is also known by researchers as the central point of fermented fish products origin [125]. Fermented fish in these regions can greatly vary based on the method of processing, the substrate used, salt concentration and the form of the final product. Based on the form of the final product the products are mainly divided into three categories. Fish products retaining their original solid form, products reduced to a liquid (fish sauces) and products reduced to a paste (fish paste) are the three categories [126]. Despite the fact the microbial profile of the fermented fish products indicates the presence of probiotic bacteria, directed studies on the effect of fermented fish products on the human gut microbiome are lacking. Fermented fish products in Asia have a wide array of microorganisms residing in the end product and based on a study done on 18 types of fermented fish products in Asia, the predominant microbial species reported were Bacillus, Lactobacillus, Micrococcus and Staphylococcus. The Lactobacillus species isolated from the fish products are identified as probiotics but then again, their mode of action when consumed as a part of diet has not been identified so far [127].
Even though the consumption of fermented food is widespread and popular in East and Southeast Asia, a wide variety of region-specific fermented foods and beverages are consumed all over Asia. For instance, India is known for the consumption of a wide variety of fermented products ranging from yogurt, and curd to fermented Lentil-based dishes as Idly. Specifically, the Northeast Indian tribal population residing in the Eastern Himalayas, Northeast hills and the Brahmaputra and Barak Valley plains are known for the production and consumption of traditional fermented products [128]. The substrates and production technologies have been passed down from immemorial times and greatly vary among different tribes which are known to be around 225 [129]. For example, fermented bamboo shoots are a regional specialty in north India, and with slight alterations, there are many different variations of this product among different tribes. Soibum/ Soidon in the state of Manipur, Mesu among indigenous people in Darjeeling hills and Sikkim, Lung-seij in among Khasi people in state of Meghalaya, Bas-tenga made by people in Nagaland, Ekung/ Hirring made in Arunachal Pradesh and Miya mikhri produced by the Dimasa tribe of Assam are all varieties of fermented bamboo shoots belonging to different tribes in Northern India [130].
These fermented bamboo shoots are indicated to be prepared from Lactobacillus sp. They are indicated to be home to a plethora of microorganisms mainly belonging to the same species L. brevis, and L. plantarum, as the abundantly found lactic acid bacteria in fermented bamboo shoots and they are widely accepted as probiotics [131]. Other than residing probiotics, the bamboo shoots can act as an excellent source of prebiotics. In vitro studies have shown that bamboo shoot polysaccharides have an excellent ability to act as a prebiotic [132,133]. It is also shown that with the fermentation, phenolic content and flavonoid content of the bamboo shoots are increased. The gut microbiome modulation ability of fermented bamboo shoots also has been proven through studies which indicated that fermented bamboo shoots are able to increase the short chain fatty acids producing bacteria and reduce the abundance of harmful gut microbiomes while increasing the production of beneficial microbial metabolites in obese rats. This study also discovered that bamboo shoots have the ability to improve gut dysbiosis rather than only promoting a healthy gut microbiome [134].
7.2. Seaweed in the Diet
Seaweed or biologically known as macroalgae are a diverse group of organisms that spontaneously grow in suitable marine environments around the world. Seaweed is considered as one of the oldest foods in the world and have been identified to be linked with brain development of the humans over evolution. It is mainly hypothesized because macroalgae having all the brain-essential nutrients and microalgae being part of the human diet since prehistoric times [13]. Seaweed has been an essential part of Southeast and East Asian countries since ancient times as a main dish, a delicacy, as a flavor enhancer or a condiment [14]. This is well proven by the fact that Asia accounts for more than 97% of the world seaweed production out of which China and Indonesia dominate 80% of the total world production [15]. In recent times, the widespread gain of popularity of seaweed based on their potential health benefits has paved a way for closer collaboration between scientists, chefs, and gastronomical entrepreneurs to conduct research on them [14].
World seaweed cultivation production tonnage increased 1,000-fold during the past 50 years from 34.7 thousand tons to 34.7 million tons. However, it should be noted that the produced algae are not limited for direct human consumption [16]. Seaweed is currently consumed as a direct sea vegetable, or used to extract hydrocolloids, to extract food colorants, as animal feed, as a biofertilizer or to extract bioactive material [17,18]. Seaweeds are mainly categorized into three groups based on the pigment they contain which are brown (Phaeophyta), red (Rhodophyta) and green algae (Chlorophyta) [19]. There are more than 1000 species of seaweed, of which about 50 are available for human food. However, according to FAO statistics in 2019, the commercial production of seaweed is concentrated around five species groups namely, Laminaria, Undaria, carrageenan seaweeds, agar seaweeds (Primarily Gracilaria) and Porphyra [15].
Algae are directly consumed as a vegetable in countries like China, Japan and South Korea. As a quantity a Japanese person consumes on average 10.4 g of seaweed per day, a South Korean 8.5 g per day and a Chinese adult 5.2 g per day [20]. Some of the mainly consumed seaweed in Japan, and South Korea and the common name of the food product prepared from them are listed in Table 3.
Chemical and nutritional composition of seaweed vary among species and within species based on the time of harvest and season of growth. But overall, they contain high amount of indigestible carbohydrates, high content of minerals, low fat and high amount of polyphenols, flavonoids and other bioactive compounds [24]. The protein content however varies among the species where red seaweed contains up to 47% of the dry weight of protein and brown seaweed contains less than 15% [25]. As explained by Cornish and colleagues, the development of the brain of humans are interrelated with seaweed consumption through two mechanisms [13]. Since seaweed has brain-essential nutrients the brain was expected to develop over time amongst the populations who resided along the coastal areas in prehistoric times. On the other hand, seaweed can modulate gut microbiota which as a result affected on developing the brain through the brain-gut-axis [13].
The ability of seaweeds to modulate gut microbiota has been proven through a plethora of studies. And long-term consumption of seaweed as part of the diet does have a mandatory effect on modulating the gut microbiome. A study conducted on Japanese individuals indicated the presence of Bacteriodes plebeius in the gut, which contains genes that encode porphyranases and agaroses enzymes that digest porphyran and agarose which are polysaccharides found in marine algae. And importantly their ability to be used to extract compounds with prebiotic capacity has also been extensively studied. Considering the composition of seaweed high dietary fiber content, presence of essential amino acids, low-fat content where the majority is Poly Unsaturated Fatty Acids or Mono Unsaturated Fatty Acids, high mineral content and presence of vitamins make them ideal dietary sources for the improvement of a healthy gut microbiota [26,27,28].
The most prominent group of compounds in seaweed that promote a healthy gut microbiome is algal polysaccharides. These polysaccharides have the ability to act as dietary fiber and even surpass the fiber content in fruits and vegetables [29]. A study conducted by Dawczynski and collegues on 34 species of marine algae products indicates that dietary fiber content can go up to 46± 8% dry weight. Moreover, these fibers being indigestible in the upper gastrointestinal tract but being utilized in the colon by gut bacteria, qualify marine algal polysaccharides as prebiotics as well [30]. The type of polysaccharides contained in algae differ among algal groups. Brown algae are mainly composed of three polysaccharides, which are fucoidans, alginate, and laminarin [31]. Green algae contain sulphated polysaccharides, sulphated galactans and xylans [32]. And red algae consist of agars, carrageenan, xylans, floridean starch (amylopectin-like glucan), water-soluble sulphated galactan, as well as porphyrin [31]. A plethora of studies suggest that polysaccharides derived from marine algae have the ability to confer a positive impact on metabolic syndromes such as obesity, diabetes, immune disorders, cancer and atherosclerosis through modulation of the gut microbiome [33]. Multiple studies conducted on Fucoidan and alginates prove that they can increase the abundance of Lactobacillus, Faecalibacterium, Blautia, Bacteroides, Alistipes, Ruminococcus, and Alloprevotella, which are beneficial bacteria negatively correlated with obesity in high-fat diet administered mice [34,35]. And fucoidans and alginate both are able to decrease the Firmicutes/Bacteroidetes ratio which is negatively correlated with obesity. While having anti-obesity effect fucoidans and alginates have a therapeutic effect on alleviating the high fat diet induced gut dysbiosis and intestinal structural damage [35,36,37,38,39]. Other compounds such as carrageenan, laminarin, porphyrin and rhamnan sulfate have also been indicated to have an anti-obesity effect by modulation of gut bacteria proportions [34]. Fucoidans have been investigated for their anti-diabetic effects. Compared to other algal polysaccharides fucoidans have ability to reduce fasting blood glucose levels and improve glucose intolerance which is imparted through modulation of gut microbiome. Benign bacteria such as Bacteroides, Faecalibacterium, and Blautia are increased in abundance and Proteobacteria especially Desulfovibrio which is negatively affecting on glucose metabolism is indicated to decreases in abundance in mice with type 2 diabetes when fed with fucoidans [40,41].
Interestingly, Sargassum fusiforme fucoidan have anti-diabetic effects where the mechanism of action is not improving the gut microbiome but inhibiting the nuclear receptors farnesoid X receptor (FXR)- small heterodimer partner (SHP) signaling pathway to reduce colon-derived biosynthesis of ceramide [42]. The bile-acid-activated nuclear receptor or farnesoid X receptor (FXR), is the master regulator of bile acid synthesis and secretion and glucose and lipid metabolism in the liver and intestine. It is also considered to maintain the barrier function of intestinal epithelial cells and prevent bacterial translocation in the intestinal tract [43]. And ceramides, a class of bioactive sphingolipids with cell signaling and second messenger capabilities, are contributing to insulin resistance [44]. Hence the effect of algal polysaccharides on gut health is not limited to alteration of gut microbiome but also moves towards regulating intestinal cell signaling pathways as well. This was proven in another study conducted using Porphyra yezoensis a red algal species consumed in China, Japan and Korea. P. yezoensis peptides induced epithelial cell proliferation in rats which associated with the activation of insulin-like growth factor cell signaling pathways [45].
The gut-friendly components in marine algae are yet again not limited to their polysaccharides. Presence of polyphenols as catechins, flavonols, bromophenols and phlorotannin that may contain upto 5% – 30% of the dry algal mass are also studied under in vitro and in vivo studies [46,47]. Out of these polyphenols, phlorophenols which are made of repeating units phloroglucinol and bromophenol molecules composed of one to five phenol groups, bound to one or more bromine are unique to algae [34,46]. Only 5–10% of polyphenols are absorbed in the upper gastrointestinal tract which is beneficial for the lower GI tract microbes as they can convert them into beneficial bioactive metabolites and inhibit pathogenic microorganisms [35]. Polyphenols extracted from Lessonia trabeculate was fed to diabetic-induced mice and the results indicates that the polyphenol extracts could increase the abundance of Odoribacter, Chnospiraceae and Alistipes, which are short chain fatty acid producing bacteria. The increase of abundance was supported by the amount of short chain fatty acids identified in the mice faecal matter. Moreover, the study resulted that other than improving the gut microbiome of a healthy mice the polyphenols are able to regulate microbial dysbiosis [48]. This was re-confirmed in another study where the flavonoids present in Enteromorpha prolifera affected the balance of the gut microbiota in diabetic mice [49]. The effect of algal bioactive compounds has been tested under both in vitro and in vivo conditions. But in vivo studies are only performed on rats, pigs and mice and human studies are relatively limited which restricts the interpretation of the effects of seaweed on gut microbiome in terms of humans.
7.3. Consumption of Soybeans
Soybeans (Glycine max) are an oilseed belonging to the family of Leguminosae which is high consumed in Southeast and East Asian regions. It is considered one of the best plant-based proteins as they have all the essential amino acids that are required for the functioning of humans. Generally, soybeans are consumed either as fermented or non-fermented beans. In Eastern Asia soybeans are consumed in the fermented form where as in the other parts of the world it is popular in the non-fermented form [50]. Natto, miso, tofu from Japan, douchi, sufu from China, cheonggukjang, doenjang, kanjang and meju in Korea, tempeh from Indonesia, Thua Nao in Thailand, and kinema, hawaijar, tungrymbai from India are majorly consumed soy based fermented products in Asia (do Prado et al. 2022). Whereas soymilk, tofu and soy protein concentrated products are widely available non fermented products. Soy proteins are popular among vegans and vegetarians as it is an excellent source of protein that can replace animal protein sources. Due to the same reason and the functional properties, soy protein is isolated from the beans to produce meat alternatives.
Composition wise soybean has similar dietary fiber content as other legumes and pulses but contain non-starch polysaccharides that include pectic polysaccharides, xyloglucan and cellulose which have been shown to promote intestinal fermentation [52]. Apart from high protein content soybeans have a high amount of oil around 15–25% which is the reason for them being categorized under oilseeds. And their fatty acid profile consists of five free fatty acids which are palmitic acid (16:0) - 10%, stearic acid (18:0)- 4%, oleic acid (18:1)- 18%, linoleic acid (18:2)- 55%, and linolenic acid (18:3)- 13% [53]. The inclusion of both omega-6 fatty acids, linoleic acid and alpha-linolenic acid than other legume crops are a speciality of soybeans’ fatty acid profile. Similar to legumes soybeans also contain anti-nutritional factors such as trypsin inhibitor, lectin, α-amylase inhibiting factor, goitrin, phytic acids etc. which limits the consumption of raw soybeans [54]. But contrary to the traditional belief of anti-nutritional factors being futile compounds that interfere in the absorption and metabolism of micronutrients, studies have shown that they possess beneficial effects as antioxidant, anti-inflammatory, anticarcinogenic and immunomodulation [55].
The fermentation process results in the chemical modification and reduction of soy components especially the presence of anti-nutritional components [55]. At the same time, soy fermentation improves the availability of end-metabolites that are required to promote human health [56,57]. Fermented soybean products in Asia are produced with either only with bacteria, others using only filamentous fungi, and, in many cases, a mixture of both are used. Some of commonly consumed fermented soy products in Asia and the fermentative microorganism/microorganisms used for the process of production are listed in Table 4.
7.3.1. Soy Isoflavones in the Diet.
Soybeans and soy protein products are an abundant source of isoflavones. Isoflavones are class of phenolic compounds, which in soybeans are present in three forms daidzein, glycitein, and genistein. Soy isoflavones are plant-derived phytoestrogens that have structural similarity to mammalian-synthesized estrogen which can bind with the estrogen receptors ER-α and ER-β to act as estrogen agonists or antagonists. Hence soybean isoflavones are also known as phytoestrogens [63]. The gut microbiota is involved in a prominent and the most favourable chemical transformation in isoflavones of soybeans. The inherent bioavailability of native isoflavones is limited by their glycosylation, which hinders absorption in the human gastrointestinal tract. Isoflavones can be categorized as glycosides, featuring sugar moieties, and aglycones. Notably, the aglycone forms exhibit enhanced absorption and exert more pronounced health benefits. However, aglycones constitute only 2–3 % of the total isoflavone content in unfermented soybeans, primarily occurring as β-glucoside conjugates. Consequently, biotransformation of glycosides into aglycones through fermentation represents a desirable strategy to increase the availability and bioactivity of isoflavones, with fermented soy products exhibiting significantly higher aglycone levels (40–100%) [64].
Similar fermentation action is exerted on isoflavones present in the soybeans by gut bacteria to form their aglycone forms which have better estrogenic and antioxidant activities than their glycoside forms. Soybean isoflavone daidzin is converted to daidzein and equol by Bifidobacterium spp., Eubacterium spp., Blautia spp., and Adlercreutzia spp in gut. Genistin, another soybean glycosidic isoflavone, is converted into genistein by Lactobacillus spp., Bacteroides spp., and Bifidobacterium spp.. Daidzein, equol, and genistein exhibited greater estrogenic and antioxidant activities than those of daidzin and genistin [65]. An individual’s ability to produce equol, has been hypothesized to be critical for obtaining the health benefits from a soy-rich diet as it exerts high estrogenic potency. This hypothesis supports the role of the gut microbiome in isoflavone metabolism. This is also yet again determined by the composition of gut microbiota where 55 % of individuals in Asian populations are equol-producers, compared to only 20–35 % of individuals in Western populations which is expected to be shaped by the total dietary intake [63,66]. There are studies conducted on effect of the isoflavone-based equols on gut microbiome using rodents. But rodents are efficient equol producers which limits the translation of available data into human studies. The interpersonal variations in the gut microbiome complicate the interpretation of data collected from humans. Furthermore, because rodents are efficient equol-producers, translatability between rodent models and humans is challenging. A human based study conducted on the impact of soy genistein on obesity concluded that genistein can reduce insulin resistance and improve fatty acid metabolism through modulation of gut microbiome. It increased in low abundant phyla, including Cyanobacteria, Deferribacteres and Tenericutes in obese subjects and decreased the Firmicutes: Bacteroidetes ratio which is attributed to weight loss in humans. Moreover, the characteristic of genistein to increase the abundance of phyla Verrucomicrobia was yet again confirmed through this study as well. Also the administration of genistein indicated an increase in fatty acid metabolism along with the biosynthesis of secondary bile acids [67]. Another study based on humanized mice induced with breast tumour proved that the ability of genistein to modulate the gut microbiome affects on increasing the latency of breast tumour and reducing tumour growth [68]. This study gave evidence to the hypothesis of Asian women’s less susceptibility to developing breast cancer compared to their western counterparts due to the Asians consume soy-based products an average of 20–50 times than the Westerners [69]. Even though the focus is greatly on isoflavone metabolism in soybeans during fermentation several other bioactive compounds are formed during fermentation by the breakdown of macromolecules by fermentative bacteria (do Prado et al. 2022). As an example, due to the metabolic activity of starter cultures, the levels of vitamin B complexes are increased during fermentation. In tempeh, Rhizopus oligosporus and the bacterial species Klebsiella pneumoniae and Citrobacter freundii produce Vitamin B12 which is a favorable compound for the proliferation of gut microbiomes [71]. Evidence from in vitro studies suggests that vitamin B12 may be associated with changes in bacterial abundance specifically increasing alpha-diversity and shifting gut microbiome composition (beta-diversity) and increasing production of short chain fatty acids [72].
7.3.2. Soy Oligosaccharides in the Diet.
Another component in soybeans, oligosaccharides, which are stachyose and raffinose have undergone several research to determine their potential as prebiotics. Both of the compounds have shown prebiotic effects in in vitro and in vivo assessments as they are able to reach the colon without microbial fermentation and once, they reach the colon are metabolized by Bifidobacteria and lactobacilli [73,74]. Ma et al. (2017) also reported that soybean oligosaccharides have been shown to increase microbial diversity in the gut, including the abundance in short chain fatty acid producing bacterial taxa such as Bifidobacterium and Lactobacillus. While aiding the proliferation of the commensal bacteria, soybean oligosaccharides promote competitive exclusion of pathogenic microbes in the gut [76]. By modulating the gut microbiome, soy oligosaccharides benefit immune function by promoting the metabolism of beneficial commensal gut bacteria; and increase levels of superoxide dismutase and IgG [77]. Moreover, soy oligosaccharides are able to safeguard the intestinal gut health through the production of short chain fatty acids and butyric acids, which was confirmed in one human based study that compared soy oligosaccharides with soy polysaccharides and oat fiber [55].
7.3.3. Fermented Soy Products in the Diet
Apart from considering effect of individual components, fermented soy products have been also studied as a whole on their effect on gut microbiome. Natto a traditional Japanese fermented soy product, has indicated therapeutic effects on gut microbiota in alleviating pathogenesis. One such study conducted on mice with low LDL receptors who were prone to develop atherosclerosis plaque indicated that natto consumption inhibited atherosclerosis through suppression of intestinal inflammation and decreasing expression of chemokines CCL2, a key regulator of macrophage recruitment in atherosclerosis [78]. The same study also indicated that inhabiting bacteria in natto have the ability to proliferate in cecum. Miso a Japanese fermented dish made of soy has been indicated to having an inhibitory effect on tyrosine kinase and particularly exhibited a potent anti-H. pylori activity. Helicobacter pylori a bacteria involved in stomach inflammation and peptic ulcers has been identified as related to stomach cancer [79]. Fermented soy milk manufactured using Lactobacillus and Bifidobacterium affects populations of human faecal microbiota in a desirable way, inducing effects that include especially alleviation of menopausal symptoms, control of hypercholesterolemia and modulation of mitogen-stimulated splenocyte proliferation [80].
7.4. Tea, a Widely Consumed Beverage
Tea is one of the most consumed beverages in the world. It is an inherent part of the Asian culture and in the diet. Originating in China it has now spread all around the world mainly as a result of the western colonisation of Asia and through the trade route of silk [81]. Tea is a beverage made from the infusion of the leaves of the Camellia sinensis plant. Based on the processed method of the leaves tea can be divided into several categories which are black tea, dark tea, yellow tea, oolong tea, green tea, and white tea. Over the years, the discovered health benefits of tea are mounting. Along with its antioxidant and anti-inflammatory properties tea has been associated to multiple health benefits, namely in the treatment of obesity, diabetes, cancer, kidney, liver, brain and bone diseases [82]. In certain disease treatments the beneficial effects are found to be imparted as a result of interplay between tea bioactive compounds and gut microbes [83]. The main variants of tea are green and black tea which both are popularly consumed in Asia. Black tea consumption is abundant in Western Asia including Turkey and South Asia. Whereas Green tea consumption is the highest in East Asian region [84]. Under this topic the considered tea types will be mainly green and black tea as they both have significantly higher consumption than other tea types in the Asian region.
7.4.1. Effect of Black Tea on Gut Microbiome.
Black tea differs from green tea as it undergoes a fermentation step during processing where catechins present in Camelia sinensis are undergone extensive oxidation and oligomerization giving rise to colour and flavour compounds. Black tea polyphenols out of which the majority include theaflavin and thearubigins. They have a higher molecular weight which makes them less bioavailable for metabolism at the upper gastrointestinal tract and are bio converted by colonic microbiota in the large intestine [85]. The produced secondary metabolites during biotransformation are absorbed to the body and are also used within the gut itself to maintain the gut epithelial cell layer[86]. Due to practical and ethical limitations in vivo studies of the effect of black tea polyphenols on gut microbiome are lacking. Studies have shown that black tea inhibits intestinal pathogens (e.g., Vibrio cholera and Salmonella enterica serovar Typhi) in in vitro studies [87]. Both from in vivo and in vitro studies black tea has proven to exert beneficial effects on gut health through various mechanisms not limiting to the modulation of gut microbiota.
In an in vivo study conducted on 72 Japanese men and women, it has been found out that black tea consumption can increase the abundance of Prevotella and Flavonifractor plautii which is a butyrate-producing bacteria and were able to improve mucosal immunity of the gut [88]. Several other in vitro study conducted on rats have shown that black tea enriched several short chain fatty acid producers who were related to increased luminal butyric acid levels and enhanced intestinal barrier function [89,90]. In another study where theaflavins of tea were subjected to in vitro anaerobic gut microbiota fermentation which resulted in increasing the abundance of Flavonifractor plautii, Bacteroides uniformis and Eubacterium ramulus who were involved in TF catabolism. Along with that a total of 17 metabolites formed after metabolism by the microbes [91].
7.4.2. Effect of Green Tea on Gut Microbiome.
In green tea, phenolic compounds constitute 24–36% of the dry weight. As green tea does not undergo the fermentation process catechins are the prime polyphenolic compound present which is present in several forms namely, epicatechin (EC), epigallocatechin (EGC), epicatechin-3- gallate (ECG), and epigallocatechin-3-O-gallate (EGCG) [92]. Polyphenols are already known to be having prebiotic effect therefore research have conducted multiple studies to determine effect of green tea polyphenols as prebiotics. They indicated tea catechins favor the growth of potentially beneficial bacteria, while hindering growth of some potentially detrimental microbes such as Bacillus cereus, Campylobacter jejuni, Clostridium perfringens, Escherichia coli, Helicobacter pylori, Legionella pneumophila, and Mycobacterium spp. [92]. Whilst hindering the growth of pathogenic microorganisms, green tea extracts attributed by the presence of polyphenols have been demonstrated to increase the abundance of commensal bacteria. A systemic review performed by Liu and colleagues indicates that green tea extract-fed mice gut exhibited reduction in Firmicutes/Bacteroidetes ratios which is positively correlated with obesity and cardiovascular disease [93]. Another study conducted on humans who did not usually drink green tea, tended to increase the proportion of Bifidobacteria abundance during consumption of green tea. However, effects were temporary as short-term dietary interventions are not adequate to do significant alterations in the gut microbiome [94]. Besides altering the gut microbiota of healthy species, green tea extracts has demonstrated ability to ameliorate microbiome dysbiosis in obese or high -fat-fed animal and human subjects [83,95]. Whilst exerting beneficial effect on gut microbiome, certain bacteria present in the colon such as Eubacterium sp. strain SDG-2, Flavonifractor plautii aK2, Flavonifractor plautii DSM 6740, Eggerthella lenta rK3, Klebsiella pneumoniae, Bifidobacterium longum sp. infantis, Enterobacter aerogenes, Raoultella planticola, Clostridium coccoides, and Bifidobacterium infantis are able to metabolize catechins into secondary metabolites [86]. In the same review, it has been listed that these microbial-derived metabolites impart antioxidative, anti-inflammatory and anti-proliferative activity. Therefore, the gut microbiome acts as a mediator in health-promoting effects of green tea where the gut microbiome itself yields beneficial effects from green tea polyphenols whilst facilitating green tea to exert their health benefits on the host.
8. South Asian Diet
South Asia also known as the Indian sub-continent has remarkable demographic importance as it is the most populated region in the world with 1.92 billion people living by the year 2022 [96]. Being the homeland to four major religions; Buddhism, Sikhism, Hinduism and Jainism and with its large population South Asia has a highly diverse culture as well as a socio-economic aspect both by which according to a study are found to contribute factors to determine diet of South Asians. Nevertheless, the traditional South Asian diet is rich in fresh fruits and vegetables, beans, legumes, rice, whole wheat, nuts, herbs, and aromatic spices [97]. The staple food of more than 70% South Asians are rice which is consumed with dishes made with fresh vegetables with addition of a lot of spices[98]. Vegetarianism and lower consumption of meat is a common dietary characteristic seen in South Asia especially due to religious influences. However, it should be also noted that South Asia has undergone and currently undergoing a dietary shift and the diet is being influenced by western dietary practices [99]. As a result the diet is now consisting of high proportions of high sugar and caloric food and larger amount of processed food [100]. But even prior to westernization South Asian diet has been characterized by a high intake of carbohydrates with less intake of dietary fiber. And this dietary pattern is considered a reason for the high prevalence of metabolic syndrome among South Asians compared to other population groups [97,101].
8.1. Spices
Spices are known as the dried part of a plant used to season or flavour food, typically seeds, fruits, berries, roots, rhizomes, bark, flowers or buds, as opposed to the green leaves and stems [102]. Spices have been an inseparable part of Asian culture that ignited Western empires to embark on voyages across the great seas in search for this hidden fortune. Asia crowned with the name of “Land of Spices” still is the largest continent of spice production and consumption out of which South Asia centred around India is known to be the world’s largest spice producer. The use of spices in especially South Asia is expanded from kitchen to clinic since ancient times under indigenous and Ayurveda medicine. In the past therapeutic use of spices were based on traditional knowledge passed down from generation to generation, but with the modern advancement of science a plethora of studies have been conducted to generate scientific evidence to back therapeutic claims related to the spices both in the form of drugs or as part of the daily diet [103]. Out of the 109 spices listed by the ISO that are grown around the world, India grows 52 spices, 31 spices are grown in Nepal, Bangladesh grows 20, Bhutan 20, Pakistan 52, Maldives 8 and Sri-Lanka more than 10 types [104].
A countless number of research has been done to prove the therapeutic potential and the role of spices on reducing the risk of disease where results partially or fully support the claims. This beneficial health effects have been claimed to be exerted by antioxidant and anti-inflammatory potential, digestive stimulant effects, hypolipidemic actions, anti-lithogenic properties, antidiabetic influence, antimutagenic, and anticarcinogenic potential of spices due to the rich repository of bioactive compounds as terpenes and terpenoid components, phenolic compounds alkaloids, saponins, glycosides, phenolic compounds, and organic acids [105,106]. The bioactive present are sometimes very specific to a certain spice while belonging to above mentioned main categories of compounds.
When investigating the underlying mechanism of the spices on alleviating or reducing the diseases occurrence, one possible proposed mechanism of action has been the modulation of gut microbiome and the gut environment [107]. It can be further proven by the presence of bioactive compounds such as polyphenols in spices which are continuously proven to confer beneficial effects on human gut microbiome [108,109]. Several specific South Asian spices have been studied for their ability to modulate gut microbiome and gut immunity, however, there is a limited amount of information regarding the activity of spice extracts and spices against intestinal bacteria, and a limited number of bacterial strains have been assessed for their susceptibility or antimicrobial activity against spices [110]. Many of the research identified are based on the major active constituents of turmeric (Curcuma longa) which is curcuminoids including curcumin. This can be possibly due to the fact that the consumption of turmeric in South Asian is relatively higher than that of other spices as part of the daily diet [107]. Curcuminoids is a lipophilic polyphenol belonging to the category of curcuminoids. It is exerting beneficial effects in two mechanisms. One is which curcumin is directly regulating the gut microbes and second in which yields active metabolites by biotransformation of curcumin by gut microbiome [111,112]. The main reason for ability of curcumin to exert beneficial effect on human gut is their resistance to low pH and poor absorption at the upper intestinal tract by which curcumin, without any chemical modifications, reaches the large intestine and undergoes metabolism by intestinal microbes. In summary, curcumin has been studied both under in vivo and in vitro conditions to test their potential on alleviating pathogenic conditions. When consumed by transgenic mice with Alzheimer disease, curcumin has shown to lower relative abundances of bacterial taxa such as Bacteroidaceae, Prevotellaceae, and Lactobacillaceae and increased the abundance of Rikenellaceae at family level who are positively related to Alzheimer’s disease. Moreover, curcumin decreased the relative abundance of Escherichia/Shigella ratio which is again another indicator associated with a peripheral inflammatory state in patients with cognitive impairment and brain amyloidosis [112]. Other studies confirmed that oral curcumin administration was able to increase the abundance of beneficial bacterial strains, such as Bifidobacteria, Lactobacilli, and butyrate-producing bacteria, and reduces the abundance of the pathogenic ones, such as Prevotellaceae, Coriobacterales, Enterobacteria and Rikenellaceae, often associated to the onset of systemic diseases (Dacrema et al. 2022; Di Cerbo et al. 2015; Peterson et al. 2018). Many of these research has been related to effect of curcumin on colorectal cancer. Other than improving the gut dysbiosis related to pathogenic conditions curcumin have the preventive ability against inflammatory disease by reducing pro-inflammatory Enterobacteria and Enterococci, but also increased the abundance of anti-inflammatory Lactobacilli and Bifidobacteria [116]. Another study done on healthy humans while administering extracts from turmeric and curcumin indicated that there was an increase by 7 % and 69 %, respectively in variety of bacterial species after 8 weeks of the treatment [114].
Chili pepper has long been recognized to have a beneficial effect on the gut microbiota in humans. Capsaicin, a pungent compound found in pepper is receiving higher attention due to the fact that in recent years, rapidly emerging evidence has demonstrated to be having a broad potent biological characteristic as antioxidant, anti-obesity, pain-alleviating, and anti-inflammation effects while conferring beneficial effects on human gut [117,118]. At the same time capsaicin extend those effect at lower doses as high doses cause gastrointestinal discomfort. It is estimated the daily mean capsaicin intake is 30–150 mg per person to prevent gastrointestinal discomfort [119]. A study conducted on mice treated with capsaicin by intragastric perfusion for one week, indicated the increase of Faecalibacterium that was initially absent in the gut. Faecalibacterium is considered the most important symbiotic component of the human gut microbiome and is considered a bioindicator of human health, being negatively associated with several non-communicable diseases such as inflammatory bowel disease (IBD), immunity, obesity, diabetes, asthma, major depressive disorder, and colorectal cancer [120]. A human based study conducted on healthy human subjects indicated an increase the abundance of Faecalibacterium. And also, the effect of capsaicin is depended on host gut enterotype, with more benefits obtained for enterotype 1 (Bacteroides enterotype) than for enterotype 2 (Prevotella enterotype) [120]. Consumption of capsaicin is proven to regulate glucose homeostasis through aa complex mechanism involved with modulating the gut microbiome and also indicated that they can reverse the gut dysbiosis resulting from type 2- diabetes [121].
Ginger rhizome (Zingiber officinale) another commonly consumed spice in Asia are rich in phenolic compounds, terpenes, polysaccharides, organic acids, and raw fiber. It is reported that oral administration of ginger extract modulates the gut microbiota, where it reduces the population of pathogenic bacteria such as Lactobacillus murinus, Lachnospiraceae bacterium 615, and Ruminiclostridium sp. KB18 [122]. Also it has shown to improve the bacterial diversity and altered the abundance of Helicobacter and Peptococcaceae species, in a gut after administration of antibiotics [123].
Few studies have been conducted on the effects of cinnamon (Cinnamomum zeylanicum, and Cinnamon cassia) on gut microbiome especially due to its widespread popularity of therapeutic effects against diabetes. Study has been conducted on effect of cinnamaldehyde; which is a bioactive compound in cinnamon, on alleviating and prevention of type 1 diabetes mellitus in rats. As per results the abundance of Lactobacillus johnsonii which is a specie that can delay onset of type I diabetes [124]. Cinnamon extracts are indicated to be improving gut microbial profile as well as the gut epithelial function when in mice that were fed with high fat diets. Ingestion of cinnamon extracts have improved the microbial population that could stimulate antimicrobial compounds such as Muc2, RegIIIγ and pIgR in gut epithelial cells which is an indication of improved gut barrier properties [125]. And it decreases the abundance of Proteobacteria which is increased upon administration of antibiotics and in metabolic disorders, inflammation, and cancer [124]. Specifically, Gammaproteobacteria belonging to phyla of Proteobacteria is associated with the occurrence of Inflammatory Bowel Disease (IBD). A study conducted on determining the effect of cinnamon extracts against IBD further proved that consumption of cinnamon extracts reduces the level abundance of Proteobacteria specifically Helicobacter pylori and in IBD patients [126].
Nevertheless, spices are not consumed as individual components in Asia but rather as mixture added to dishes. Therefore, collective effect of spices on gut microbiome has also been studied which is comparatively lacking compared to studies based on individual spice extracts. A study based on effect of spice mixture containing cinnamon, oregano, ginger, rosemary, black pepper and cayenne pepper, in dietary doses, on gut microbiome found that up to 26 operational taxonomic units (OTUs) were modulated as a result of the mixed-spice treatment compared with placebo consumed group. However significant differences in microbial population have not been identified by this study [127]. Another similar study done with 7 dried spice powders of turmeric, cumin, coriander, amla (Indian gooseberry), cinnamon, clove, and cayenne pepper identified that the overall alpha-diversity of gut microbiome in healthy Chinese adults did not change as a result of this dietary intervention which only lasted 24–48-hour time frame. This study again was limited by the time frame the study conducted and number of participants participated [128]. Looking into the results obtained and these two being the only published research based on intervention of spice mixtures on human gut microbiota, there’s a gap in scientific data in the area of spice mixtures which can be bridged by further studies.
8.2. Legumes in the Diet
Legumes are plant material that belong to the botanical family of Fabaceae (Leguminosae). But as food material, legumes are interchangeably known by terms pulses, legumes and beans although three terms are distinctively different. Even under country-specific guidelines, the term legume has been interpreted in various ways. However, according to the definition given by FAO pulses are only legumes with dry, edible seeds, with low fat content that excludes legume species used as vegetables (e.g., green peas, green beans), for oil extraction (e.g., soybean, groundnut) and for sowing purposes (e.g., clover, alfalfa). Which means pulses are a subcategory of legumes. Hence legumes are a collective term that can be used to define pulses, oilseed and legumes itself. Under this section the consumption of legumes in South Asia is discussed excluding oil seed legumes namely peanut and soy beans as they are being widely consumed in the region of East Asia. A study conducted by Hughes et al., 2022 on legume consumption in 94 countries of the world recorded that highest consumption of legumes are recorded in South Asian region specifically Afghanistan, Sri Lanka and Nepal [129]. Another report indicates that India as the world’s largest producer of pulses as a country and they account for 12.7% of the protein intake of an individual where global average lies lesser than 5% [130].
The world primarily consumes nine types of legume crops as whole foods: five of these are pulses [dry beans (Phaseolus vulgaris), chickpeas (Cicer arietinum), lentils (Lens culinaris), and dry peas (Pisum sativum)] and two are undried legumes (snap beans and snap peas) (Didinger and Thompson 2021). The nutrient composition of legumes vary between different types of legumes but as a whole compared to other plant-based food as cereals, yams and vegetables legumes contain higher protein content which can range between an average of 20%-26% [132]. Hence legumes are considered to be an excellent source of protein in vegan and vegetarian diets. Moreover, legumes have low concentrations of sulphur amino acids and Trypsine and are relatively high in essential amino acid Lysine which is limited in cereals. Therefore, usually, it is recommended to consume cereal along with legumes to have a balanced protein intake. Apart from oil seeds, legumes have a lower fat content, compared to carbohydrates and proteins (Didinger and Thompson 2021). Legumes are high in carbohydrates out of which a higher proportion is present as starch whereas a considerable amount of carbohydrates are in the form of a-galactosides which include raffinose, stachyose, and verbascose. In terms of gut health, a-galactosides are important as they are considered excellent candidates for prebiotics. The reason is they are not digested in the upper intestinal tract of human due to the absence of galactosidase enzymes but are fermented in the colon by residing bacteria which produces which produce CO2 and methane as by-products causing flatulence [134]. Other than the non-digestible oligosaccharides, legumes contain dietary fibre which are mainly cellulose and hemicellulose [135]. Apart from those starch which is already present undergoes structural and compositional changes during the legume processing and form resistant starch which also functions as dietary fibre [132]. Common beans have higher total dietary fibre (23–32 g/100 g) compared to chickpeas, lentils, and dry peas (range of 18–26 g/100 g)[136]. Both the carbohydrate types, soluble oligosaccharides and dietary fibre are non-digestible in the upper gastrointestinal tract and are fermented by colonic bacteria in the colon which produces fermentation products including gases and short chain fatty acids. Although the gases may cause digestive discomfort due to flatulence, the short chain fatty acids support the health of the intestinal mucosa [136]. Especially legume oligosaccharides are known as potential prebiotics [137,138,139]. The produced microbial metabolites are beneficial in proliferation of commensal bacteria in the gut and modulating a healthy gut microbiome. Another mechanism of action by which the dietary fiber affects gastrointestinal tract is through increasing water-holding capacity, viscosity, bulk, fermentability and the ability to bind bile acids. In terms of modulating the gut microbiome, α-galactoside oligosaccharides are generally proven to be increasing the genera of Bifidobacterium and Lactobacillus who prevent the proliferation of exogenous and native pathogenic microorganisms [140].
Apart from studying the individual effect of non-digestible components, many studies are conducted to find the effect of pulses. Chickpea diet fed obese mice have indicated to increase the abundance of genera of Coprococcus, saccharolyticum, Butyricicoccus, and Pullicaecorum who are depleted in the gut microbiome of obese subjects [141]. Other than conferring beneficial effects on gut microbes under pathogenic conditions chickpea has exhibited the ability to improve the gut function of a healthy subject as well. chickpea flour-based diet was fed to healthy C57BL/6 mice which indicated the diet enhanced taxa richness and abundance of Prevotella a short chain fatty acid producing species and Ruminococcus flavefaciens whose abundance has been shown to decrease when obese. The effect of increasing short chain fatty acid producing bacteria was further proven through the increased caecal short chain fatty acid concentration. Moreover, the changes in the gut microbiome increased flavonoid biosynthesis and butanoate metabolism [142]. Study conducted to identify the effect of both chickpea and chickpea oligosaccharide on human gut resulted that both oligosaccharide fraction and the chickpea extracts have similar effects on improving gut microbiome without indicating significant differences among the effects [143]. Chickpea proteins and peptides have been studied as well under in vitro conditions where chickpea peptides promoted Bifidobacterium growth, levels of lactic acid bacteria (i.e., Pediococcus, Weissella) and acetate and propionate-producing bacteria Veillonella [144]. Other than chickpea, lentils which is an overlooked, yet potential source of prebiotics has also been studied for their prebiotic potential which indicates per 100 g of lentils it contains 13 g of prebiotics [145]. Lentils can alleviate gut dysbiosis resulting by obesity and can positively affect on weight reduction by reducing triglyceride concentration and mean percent body fat [146]. Moreover, cooked lentils have the ability to increase fecal microbiota α-diversity and abundance of short-chain fatty acid producing bacteria as Prevotella, Roseburia and Dorea spp. [147].
8.3. Vegetarianism and Gut Microbiota
Vegetarianism is not just a dietary pattern but rather a way of life which in terms of diet characterized by partial or strict exclusion of food of animal origin and inclusion of higher portion of plant-based food. The vegetarian diet dates back to 3200 BC, where evidence is found in Egyptian civilizations where Egyptians were abstaining from meat consumption to facilitate reincarnation [148]. Throughout history, vegetarianism has been spreading around the world associated with religious beliefs and practices as respecting all living beings and following the path of nonviolence. These principles are bound with religions as Hinduism, Jainism, Sikhism, Buddhism, the Hare Krishna movement, and the Seventh-day Adventist Church. India being the cradle of many of the world religions Hinduism, Jainism, Sikhism and Buddhism is having world’s largest vegetarian population where despite the debates around the accuracy of the statistics mention nearly 37% total population follow vegetarianism [149]. Being influenced by the religious philosophies in India, other countries in the South Asian region do follow vegetarianism as well. Vegetarian diet has certain subcategories based on how restrictive they are in the inclusion and exclusion of foods of animal origin which is described in Table 5. The choice of each pattern by a person can vary based on their motif on following the particular diet [150].
Vegetarianism has wide gained popularity due to the recent development of cruelty free diets and the widespread awareness on its health benefits. According to the American Dietetic Association, a well-planned vegetarian diet can not only lead to significant health benefits, but also meet the nutritional needs of all age groups [151]. Considering the dietary composition of vegetarian and omnivore diets, a vegetarian diet is high in dietary fiber, unsaturated fats, vitamins such as vitamin C, vitamin A and folate and minerals as calcium and magnesium. On the other hand, consumption of protein, total fat and sodium is low in vegetarians due to low consumption of meat [150,152]. But at the same time non-nutrient phytochemical consumption such as isoflavone and phytosterols are higher in vegetarians due to their high inclusion of vegetables and fruits in the diet [150].
There is accumulating epidemiological evidence that a vegetarian diet has preventive and therapeutic effects on non-communicable diseases such as cardiovascular disease (CVD), type-2 diabetes, high systolic and diastolic blood pressure, bone-related osteoporosis, gastroesophageal reflux disease, asthma, degenerative diseases including cancers, ischemic heart disease and autoimmune disease as rheumatoid arthritis [153,154,155,156,157,158,159,160,161,162,163,164]. These beneficial effects are exerted due to the high vegetable and fruit consumption in vegetarian diet which contributes to consumption of high dietary fiber, presence of high amounts of micronutrients as vitamins and minerals and phytochemicals such as polyphenols that possess the antioxidant properties [165,166,167]. And it can also be due to absence of meat consumption specifically red meat and processed meat that provides high amount of fat which are majorly saturated fat with low amount of fiber [168]. This difference in the nutritional and non-nutritional composition between vegetarian and non-vegetarian diets have affected on gut microbiome composition as well. Based on studies vegetarian diets can strongly affect the relative abundance of certain genera, and significantly increase gut microbial diversity which is attributed by an increase in the α-diversity index in vegetarians [169]. This is explained by the same reason of having favorable nutrients for gut microflora from vegetables and fruits such as dietary fiber, and phenolic compounds that easily reach the colon and are utilized by the colonic microorganisms and produce favorable microbial metabolites and will create a favorable environment for commensal microorganisms within the intestinal tract. Due to the presence of these non-digestible components, a vegetarian diet requires a more diverse set of microorganisms for the digestion [170]. In summary it is found to be increasing the abundance of bacteria that ferment dietary fiber, such as Clostridium, Lactobacillus, Ruminococcus, Eubacterium rectale and Faecalibacterium prausnitzii [150,171]. Despite following a vegetarian diet in communities where consumption of plant-based food is high, the gut microbial ratio of Firmicutes/Bacteroidetes is less which is correlated to lean body types[172]. Many studies have shown Firmicutes/Bacteroidetes is related to obesity [169]. They also have a lower abundance of Bifidobacterium [173]. On the other hand, long-term vegetarianism was related to a less diverse T-cell repertoire, reduced levels of IgE which is a crucial allergy-related immunological indicator [174].
Apart from gut microbiome changes taken due to long-term vegetarian diet certain studies have shown that short term dietary changes can also improve the gut microbial profile. A study based on a 3-month consumption of lacto-ovo-vegetarian diet by healthy omnivorous volunteers indicated that the diversity of gut microbiota was changed after the vegetarian diet. Specifically, abundance of microbes in Alistipes, a bile-tolerant microorganism which is characteristically increased during meat-based diets were reduced in tested volunteers. It is also shown that a short-term vegetarian diet does not have a significant effect on changing the gut microbiome or immune system [175]. This temporary dietary intervention can have a beneficial effect on modifying the gut of healthy subjects as well. A study conducted on six obese human subjects with type 2 diabetes/or hypertension has been assigned to a strict vegetarian diet for one month. The participants at the end of one month has indicated reduced body weight and reduced concentrations of triglycerides, total cholesterol, low-density lipoprotein cholesterol. In terms of gut the microbes it reduced the Firmicutes/Bacteroidetes ratio which is an indicator of obesity when high. Furthermore, after one month of diet, pathogenic bacteria such as Enterobacteriacea were reduced and commensal microbes such as Bacteroides fragilis and Clostridium species were increased [176].
9. Diet in Central Asia
Central Asia has a quite versatile dietary pattern than the rest of the Asian region which is mainly due to different agricultural farming practices shaped by, dry, mountainous, extreme and harsh weather conditions which makes it stressful for farmers to engage in the cultivation of crops. Limitation of water due to lack of rainfall is the major difference that is observed in Central Asia compared to other Asian regions that limits them from engaging in agricultural farming [172]. For 7,000 years, agriculture in the better-provided areas of Central Asia has meant primarily barley and wheat. For centuries they have been living under a nomadic way of life, raring cattle, sheep, goats, camel, and horse known as “five muzzles” [172]. Still the diet of Central Asians is mainly composed of dairy and meat from goat, sheep and horses. Although it is less prevalent at present, hunting has played a huge role in Central Asian food pattern. Present Central Asian food culture is on one part influenced by the West due to its location in the middle of the Eurasian border and several other parts of the region are influenced by cultures of countries like China, India, Russia and Iran [177]. As a whole, Central Asian diet is now characterized by the consumption of wheat, along with minor amounts of millet, barley, meat from sheep which are supplemented by goats, cattle and horses and the consumption of milk in both fermented and fresh form. Nevertheless, based on recent data Central Asia has one of the highest rates of premature mortality from non-communicable diseases (NCDs), such as cardiovascular diseases, diabetes, and certain types of cancer. Dietary habits are one of the major factors contributing to the prevalence of NCDs [178]. In fact, a recent study of about 200 countries showed that the burden of diet-related deaths in Central Asia is among the highest in the world [179].
9.1. Red meat in the Central Asian Diet
A distinctive characteristic of the Central Asian diet is the inclusion of a high amount of red meat. Meat has been an integral part of their lifestyle originating from the earlier nomadic lifestyle of their ancestors. Hence it is not a simple dietary choice they have made. Although the consumption pattern has fluctuated due to several factors as urbanization and socio-economical transformations still red meat is a significant dietary component in the Central Asian diet [180,181]. The average per capita consumption of meat ranges from 50 kg to 70 kg per year in the Central Asian region which as a region records highest in the world. And unlike in the other parts of the world the preferred meat types are sheep, horse, cattle, camels and goats [172]. From the region, Mongolia has the highest meat consumption owing to being a landlocked country having less arable land for crop cultivation [172]. Considerably higher proportions of lamb meat and a lesser number of poultry and pork are consumed compared to the meat consumption patterns in neighboring regions of East Asia and Europe. Contrary to the popular norms horse meat is abundantly consumed in Kazakhstan and Kyrgyzstan as a part of daily diet [182]. This category of red meat has gained global attention due to claims that they are associated with diet-related non-communicable diseases. Many studies have indicated that red meat relates to higher risk of cardiovascular disease, coronary heart disease, stroke and type 2 diabetes mellitus [183]. Central Asia already showing high morbidity and mortality rate of these diseases it’s worth observing the gut microbial composition of the population. At the enterotype level, it has been found out that both in Mongolia and Kazakhstan the prominent genus being Enterotype 2 which is characterized by the high intake of carbohydrate rich diet. But however, changes in the genus level in Central Asian gut be characterized by the intake of red meat. A study conducted on Mongolians indicated high abundance of Collinsella in the gut who have a positive correlation with high protein intake as well as with symptomatic atherosclerosis [184].
Although Central Asia has a different dietary pattern compared to other parts of the world which sets them apart even from the rest of the other Asian countries, the focus given on studying changes in gut microbiota composition related to diet pattern is less. Studies have been conducted on determining the composition of the gut microbiome, however they are not comparative and are highly based on individual communities in countries. And the few research available have contrasting results and do not clearly clarify the reasons for the differences in these results [185].
9.2. Mare’s milk in Central Asian Diet
For centuries, dairy products in Central Asia regarded as the “white food” of the steppe were processed according to different procedures as fermentation, distillation, drying, and clotting to ensure a longer shelf-life. As the milk is produced from a variety of species, the diversity of the products was very high, especially after the lactic fermentation process. Out of other types of food products consumed in Central Asia, consumption of mare’s milk is unique to their diet as mare’s milk consumption is endemic and centralized in Central Asian Countries. Historically the first domestication of horses and consumption of mare milk has taken place in present-day Akmola province of Kazakhstan which is part of Central Asia approximately in 3500 BCE [186]. Mare’s milk is mainly consumed in its fermented form which is known as koumiss or kumis. It is an acidic and slightly alcoholic drink, the fermentation of which involves lactic bacteria and yeasts. Composition-wise mare milk is more similar to human milk than they are to cow’s milk. It has a higher amount of whey protein and lesser casein protein content similar to human milk. And the whey protein proportion is abundant with antibacterial compounds as lysozymes, immunoglobulins, lactoperoxidases, and lactoferrin [187,188]. Due to similar composition of mare milk to human milk it is assumed and also proven through several research that mare milk has the similar ability of human milk to recover the intestinal microbiota after antibiotic therapy and confer immunomodulatory effects. Mares’ milk contains prebiotic properties which can restore the gut microbiota disrupted by consumption of antibiotics. An in vivo study conducted using six children aged 4 to 5 years diagnosed with bilateral bronchopneumonia who were prescribed cephalosporin (cefuroxime) antibiotics indicated consumption of mare milk restored the disturbed 11 genera of bacteria [189]. Another similar study done on children indicated that consumption of Mare milk while administering antibiotics prevented the bloom of Mollicutes which is a pathogenic class of bacteria in humans and animals, while preventing the loss of Coriobacteriales. Coriobacteriales, who are in the phylum of Actinobacteria and belong to the commensal flora and typically inhabit the oral cavity, gastrointestinal tract, and genital tract who which perform important functions, such as the conversion of bile salts and steroids [187]. A study directly conducted to assess the suitability of cow, goat and mare milk as a replacement for human milk concluded that out of three types of milk mare milk can be a possible replacement for human milk [190].
10. Western Asia
Western Asian countries being situated in the middle of Asia, Africa and Europe has a highly variable dietary style across countries. Even some studies indicate that due to the influence of Indian, African, and European cuisine on Western Asia dietary pattern determination is quite difficult as food selection and consumption practices greatly vary from country to country [185]. Nevertheless, there are certain commonalities among the countries based on religious practices. The majority of the population in Western Asian countries follow Islam and hence refrain from consuming pork and alcohol as part of their diet. In general, Western Asian diets are characterized by the consumption of wheat and wheat-based products as the staple, with meat, egg and sea food along with pulses, and fruits and vegetables which have lesser consumption quantity than the daily recommended intake [191]. One of the distinctive characteristics of the Western Asian diet is the consumption of dates. Date fruit (Phoenix dactylifera L.) contains high levels of both dietary fiber and polyphenols which are beneficial for a proliferation of a healthy gut microbiome. Few studies have been conducted on the effect of date consumption on gut microbiome under both in vivo and in vitro conditions. Under in vitro conditions both the date fruit extract and date fruit polyphenol extract have been indicated to increase production of short chain fatty acids and abundance of beneficial bacteria [192]. However, under in vivo conditions the date consumption has not indicated a considerable change in the gut microbiome profile or the microbial metabolite production, yet increased the bowel movement and has reduced stool ammonia concentration [193]. In both the studies date extracts and consumption od dates has indicated to inhibit proliferation of colon cancer cells.
Similar to that of Central Asia, gut microbiome related studies are limitedly available in Western Asian countries especially on their interplay with the dietary components [194]. However, even though diet component related data are lacking, “fasting” a dietary regimen has been studied with related to other parts of the world due to their potential health benefits. Western Asians since the majority of Muslims and Jews practice fasting as a part of their religious practices. Hence “Fasting” is discussed under the section but not limiting to the Western Asia but consider the data available from other parts of Asia on effect of fasting on the modulation of gut microbiome profile.
10.1. Fasting as a Dietary Regimen
Intermittent fasting a is a widely popularizing dietary regimen in the world due to their positive effect on weight loss or health improvement. It’s a method of fasting where there are prolonged periods of abstinence from food and drink between meals. Even though it has now gained popularity around the health-conscious communities this has been in practice for thousands of years as a part of religious practice followed by Muslims during the Ramadan period. During Ramadan, Muslims abstain from ingesting food and liquids between sunrise and sunset which usually last 12–18 h/day, throughout a month-long period [195]. There are several forms of intermittent fasting (IF), all using fasting periods that extend well beyond the duration of an overnight fast and implicating limited feeding time-windows, with or without caloric restriction [196]. In Asia, fasting is practiced mandatorily by the Muslim population during the month of Ramadan. Out of 1.57 billion Muslims in the world 60% of are residing in Asia with the highest density in Southeastern Asia. Being the cradle of Islam, the Arabian Peninsula which is part of Western Asia consider Islam as the major religion as well [197]. Apart from Islamism, other religions in the Asian region as Jainism, Judaism and Buddhism also practice fasting as part of their religious practice [197]. Apart from religious practices, non-religious practices such as “Beego” in China also recommend fasting for mental and physical health. Hence fasting is an important dietary regimen in Asia that should be focused on in terms of its effect on human health.
The major reason for the widespread popularity of fasting is due to increased evidence of health benefits related to the practice. Fasting more specifically intermittent fasting is known for reducing body weight, positively affecting glucose regulation, reducing systemic inflammation, reducing high blood pressure through improving the lipid profile, ameliorating some cardiometabolic parameters, healing of thrombophlebitis healing of refractory dermal ulcers and tolerance of elective surgery [198,199,200]. One method by which these beneficial effects are conferred is through modulating the metabolism in the human body. The main identified metabolic change is switching from glucose metabolism to ketogenesis due to a lack of glucose intake for energy metabolism. During fasting triglycerides from adipose tissue are converted to fatty acids and glycerol, which are subsequently metabolized for energy. This process has certain beneficial metabolic effects; it decreases blood sugar, and insulin levels as well as IGF-1 levels, stimulates glycogen release, and increases lipolysis and ketogenesis [197]. Fasting also reduces oxidative stress through decreasing reactive oxygen species (ROS), increasing antioxidant enzyme activities or increasing the turnover rate of oxidized macromolecules. This is likely the reason by which prevention of cancer and inflammation occurs through fasting [201]. Considering the relationship between obesity and inflammatory conditions with gut microbiome alterations, studies have been conducted to identify the effect of fastening in the modulation of gut microbiome during alleviating the pathological conditions. Studies have specifically focused on gut microbiome alterations related to Ramadan fasting. A very recent study conducted on 12 healthy adult individuals who practiced fasting 17 hours per day for 29 consecutive days indicated after the period of fasting abundance of Firmicutes was decreased and the abundance of Proteobacteria was decreased. And also levels of seven genera of bacteria namely Blautia, Coprococcus, Dorea, Faecalicatena, Fusicatenibacter, Lachnoclostridium, and Mediterraneibacter were decreased after the period of Ramadan [202]. Another study by Ozkul and colleagues revealed that Butyricicoccus, Bacteroides, Faecalibacterium, Roseburia, Allobaculum, Eubacterium, Dialister and Erysipelotrichi were significantly enriched genera after the end of Ramadan fasting. This is contrary to the earlier study as Butyricicoccus, Faecalibacterium, and Roseburia are Firmicutes [203]. The results are further deviated from another study conducted with participants of Pakistani and Chinese origin who were part of Ramadan fasting. This study indicated different results among the two different populations where Dorea, Klebsiella and Faecalibacterium were more abundant in the Chinese group after fasting, while Sutterella, Parabacteroides and Alistipes were significantly enriched after fasting in the Pakistani group. Compared to the data derived from [202], levels of Dorea and Faecalicatena decreasing is contrary to the previous study. Considering these differences, researchers have concluded that even though fasting exerts a beneficial effect on the human gut microbiota of the individual, the variation in gut microbiota largely depends on dietary habits of the person during and previous to Ramadan fasting.
11. Recommendations and Way Forward
Studying and bringing the diet-based variability of gut microbiome in Asia under one umbrella is extremely difficult due to high variability across the continent in terms of economic, social and cultural aspects. Yet completely segregating them into different categories is also difficult as every dietary culture has been influenced and is influencing another up to a certain extent. This process will become even more complicated with the rapid dietary transition that Asia currently undergoes. With rapid economic growth, globalization, and industrialization, the Asian diet transition is in a very volatile state and based on current research the dietary patterns are shifting more towards Western diets. Compared to a few decades ago food availability and purchasing power of the citizens have increased reducing malnutrition in the region. However, parallelly micronutrient deficiencies, the percentage of obesity and overweight is on the rise in the region [204]. These dietary changes have already proven to have caused alterations in the gut microbiome and these alterations are favorable towards the occurrence of non-communicable diseases. This has been studied under a limited number of studies with related communities [205,206,207]. To mitigate any possible adverse effects of dietary transitions on gut microbiome furthermore inclusive studies have to be conducted in the Asian region. This can also be beneficial in identifying specific gut microbes that are already present in the gut microbiome of Asians which can be used as therapeutics for the non-communicable diseases.
In terms of the availability of research on the association of diet components and the gut microbiome, there’s a huge disparity across regions. Many of the research available are conducted based on East and Southeast Asian regions while only a few have been conducted in the Central and Western Asian region. Almost all the novel and traditional foods from East and Southeast Asia are being studied which gives a comprehensive overview of the gut microbiome and their behavior when these foods are consumed which has further evolved into studies on the development of already consumed food into better probiotics. South Asian regions also do have a considerable amount of research conducted on this area, but the traditional and indigenous dietary patterns are yet to be further studied. Apart from mainstream food components, community-specific food products consumed by tribal populations have not gained adequate attention. However, in Central and Western Asian regions not even the major dietary components lack publicized data. Apart from the generation of information the already acquired data can be integrated into fruitful research in developing therapeutics for NCDs to reduce their morbidity and mortality. At the same time, this generated knowledge can be transferred into food related national policies to ensure quality of life of the population through a healthy diet.
Author Contributions
Conceptualization, E.M., N.R and V.S.; Investigation, V.N.; resources, X.X.; data curation, X.X.; writing—original draft preparation, V.N.; writing—review and editing, E.M. and N.R.; supervision, E.M. and N.R.; All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Ren, Y.; Wu, J.; Wang, Y.; Zhang, L.; Ren, J.; Zhang, Z.; Chen, B.; Zhang, K.; Zhu, B.; Liu, W.; et al. Lifestyle Patterns Influence the Composition of the Gut Microbiome in a Healthy Chinese Population. Sci Rep 2023, 13, 1–16. [Google Scholar] [CrossRef]
- Nakayama, J.; Watanabe, K.; Jiang, J.; Matsuda, K.; Chao, S.H.; Haryono, P.; La-Ongkham, O.; Sarwoko, M.A.; Sujaya, I.N.; Zhao, L.; et al. Diversity in Gut Bacterial Community of School-Age Children in Asia. Sci Rep 2015, 5, 1–11. [Google Scholar] [CrossRef]
- Das, B.; Ghosh, T.S.; Kedia, S.; Rampal, R.; Saxena, S.; Bag, S.; Mitra, R.; Dayal, M.; Mehta, O.; Surendranath, A.; et al. Analysis of the Gut Microbiome of Rural and Urban Healthy Indians Living in Sea Level and High Altitude Areas. Sci Rep 2018, 8, 1–15. [Google Scholar] [CrossRef]
- Ang, Q.Y.; Alba, D.L.; Upadhyay, V.; Bisanz, J.E.; Cai, J.; Lee, H.L.; Barajas, E.; Wei, G.; Noecker, C.; Patterson, A.D.; et al. The East Asian Gut Microbiome Is Distinct from Colocalized White Subjects and Connected to Metabolic Health. Elife 2021, 10, 1–28. [Google Scholar] [CrossRef]
- Nakayama, J.; Zhang, H.; Lee, Y.-K. Asian Gut Microbiome. Sci Bull (Beijing) 2017, 62, 816–817. [Google Scholar] [CrossRef]
- Lim, M.Y.; Hong, S.; Bang, S.-J.; Chung, W.-H.; Shin, J.-H.; Kim, J.-H.; Nam, Y.-D. Gut Microbiome Structure and Association with Host Factors in a Korean Population. mSystems 2021, 6. [Google Scholar] [CrossRef]
- Shinoda, A.; Shirchin, D.; Jamiyan, D.; Lkhagvajav, T.; Purevdorj, C.; Sonomtseren, S.; Chimiddorj, B.; Namdag, B.; Therdtatha, P.; Nakayama, J. Comparative Study of Gut Microbiota Mongolian and Asian People. Mongolian Journal of Agricultural Sciences 2021, 33, 1–7. [Google Scholar] [CrossRef]
- Syromyatnikov, M.; Nesterova, E.; Gladkikh, M.; Smirnova, Y.; Gryaznova, M.; Popov, V. Characteristics of the Gut Bacterial Composition in People of Different Nationalities and Religions. Microorganisms 2022, 10. [Google Scholar] [CrossRef]
- Yang, B.; Yan, S.; Chen, Y.; Ross, R.P.; Stanton, C.; Zhao, J.; Zhang, H.; Chen, W. Diversity of Gut Microbiota and Bifidobacterial Community of Chinese Subjects of Different Ages and from Different Regions. Microorganisms 2020, 8, 1108. [Google Scholar] [CrossRef]
- United Nations Classification and Definition of Regions.
- Holcombe, C. A History of East Asia. A History of East Asia 2017, 907. [Google Scholar] [CrossRef]
- Phan, U.T.X. Meals and Snacks in Southeast and East Asia. Handbook of Eating and Drinking: Interdisciplinary Perspectives. [CrossRef]
- Cornish, M.L.; Critchley, A.T.; Mouritsen, O.G. Consumption of Seaweeds and the Human Brain. J Appl Phycol 2017, 29, 2377–2398. [Google Scholar] [CrossRef]
- Mouritsen, O.G.; Rhatigan, P.; Pérez-Lloréns, J.L. World Cuisine of Seaweeds: Science Meets Gastronomy. Int J Gastron Food Sci 2018, 14, 55–65. [Google Scholar] [CrossRef]
- Cai, J. Global Status of Seaweed Production, Trade and Utilization; 2021.
- Fleurence, J. Seaweeds as Food; Elsevier Inc., 2016; ISBN 9780128027936.
- Rogel-Castillo, C.; Latorre-Castañeda, M.; Muñoz-Muñoz, C.; Agurto-Muñoz, C. Seaweeds in Food: Current Trends. Plants 2023, 12. [Google Scholar] [CrossRef]
- Pati, M.P.; Das Sharma, S.; Nayak, L.; Panda, C.R. Uses of Seaweed and Its Application to Human Welfare: A Review. Int J Pharm Pharm Sci 2016, 8, 12–20. [Google Scholar]
- Buschmann, A.H.; Camus, C.; Infante, J.; Neori, A.; Israel, Á.; Hernández-González, M.C.; Pereda, S.V.; Gomez-Pinchetti, J.L.; Golberg, A.; Tadmor-Shalev, N.; et al. Seaweed Production: Overview of the Global State of Exploitation, Farming and Emerging Research Activity. Eur J Phycol 2017, 52, 391–406. [Google Scholar] [CrossRef]
- Ficheux, A.-S.; Pierre, O.; Le Garrec, R.; Roudot, A.-C. Seaweed Consumption in France: Key Data for Exposure and Risk Assessment. Food and Chemical Toxicology 2022, 159, 112757. [Google Scholar] [CrossRef] [PubMed]
- Mouritsen, O.G.; Rhatigan, P.; Pérez-Lloréns, J.L. World Cuisine of Seaweeds: Science Meets Gastronomy. Int J Gastron Food Sci 2018, 14, 55–65. [Google Scholar] [CrossRef]
- Hwang, E.K.; Park, C.S. Seaweed Cultivation and Utilization of Korea. Algae 2020, 35, 107–121. [Google Scholar] [CrossRef]
- Rioux, L.E.; Beaulieu, L.; Turgeon, S.L. Seaweeds: A Traditional Ingredients for New Gastronomic Sensation. Food Hydrocoll 2017, 68, 255–265. [Google Scholar]
- Olsson, J.; Toth, G.B.; Albers, E. Biochemical Composition of Red, Green and Brown Seaweeds on the Swedish West Coast. J Appl Phycol 2020, 32, 3305–3317. [Google Scholar] [CrossRef]
- Thiviya, P.; Gamage, A.; Gama-Arachchige, N.S.; Merah, O.; Madhujith, T. Seaweeds as a Source of Functional Proteins. Phycology 2022, 2, 216–243. [Google Scholar] [CrossRef]
- Lopez-Santamarina, A.; Miranda, J.M.; Mondragon, A.D.C.; Lamas, A.; Cardelle-Cobas, A.; Franco, C.M.; Cepeda, A. Potential Use of Marine Seaweeds as Prebiotics: A Review. Molecules 2020, 25. [Google Scholar] [CrossRef] [PubMed]
- Rocha, C.P.; Pacheco, D.; Cotas, J.; Marques, J.C.; Pereira, L.; Gonçalves, A.M.M. Seaweeds as Valuable Sources of Essential Fatty Acids for Human Nutrition. Int J Environ Res Public Health 2021, 18, 4968. [Google Scholar] [CrossRef]
- Siddik, M.A.B.; Francis, P.; Rohani, M.F.; Azam, M.S.; Mock, T.S.; Francis, D.S. Seaweed and Seaweed-Based Functional Metabolites as Potential Modulators of Growth, Immune and Antioxidant Responses, and Gut Microbiota in Fish. Antioxidants 2023, 12, 2066. [Google Scholar] [CrossRef]
- Dawczynski, C.; Schubert, R.; Jahreis, G. Amino Acids, Fatty Acids, and Dietary Fibre in Edible Seaweed Products. Food Chem 2007, 103, 891–899. [Google Scholar] [CrossRef]
- Zheng, L.X.; Chen, X.Q.; Cheong, K.L. Current Trends in Marine Algae Polysaccharides: The Digestive Tract, Microbial Catabolism, and Prebiotic Potential. Int J Biol Macromol 2020, 151, 344–354. [Google Scholar] [CrossRef] [PubMed]
- Flores-Contreras, E.A.; Araújo, R.G.; Rodríguez-Aguayo, A.A.; Guzmán-Román, M.; García-Venegas, J.C.; Nájera-Martínez, E.F.; Sosa-Hernández, J.E.; Iqbal, H.M.N.; Melchor-Martínez, E.M.; Parra-Saldivar, R. Polysaccharides from the Sargassum and Brown Algae Genus: Extraction, Purification, and Their Potential Therapeutic Applications. Plants 2023, 12, 2445. [Google Scholar] [CrossRef]
- Kraan, S. Algal Polysaccharides, Novel Applications and Outlook. In; Chang, C.-F., Ed.; IntechOpen: Rijeka, 2012; p. Ch. 22.
- Yatsunenko, T.; Rey, F.E.; Manary, M.J.; Trehan, I.; Dominguez-Bello, M.G.; Contreras, M.; Magris, M.; Hidalgo, G.; Baldassano, R.N.; Anokhin, A.P.; et al. Human Gut Microbiome Viewed across Age and Geography. Nature 2012, 486, 222–227. [Google Scholar] [CrossRef]
- Zang, L.; Baharlooeian, M.; Terasawa, M.; Shimada, Y.; Nishimura, N. Beneficial Effects of Seaweed-Derived Components on Metabolic Syndrome via Gut Microbiota Modulation. Front Nutr 2023, 10. [Google Scholar] [CrossRef]
- Shannon, E.; Conlon, M.; Hayes, M. Seaweed Components as Potential Modulators of the Gut Microbiota. Mar Drugs 2021, 19, 1–50. [Google Scholar] [CrossRef]
- Jiang, P.; Zheng, W.; Sun, X.; Jiang, G.; Wu, S.; Xu, Y.; Song, S.; Ai, C. Sulfated Polysaccharides from Undaria Pinnatifida Improved High Fat Diet-Induced Metabolic Syndrome, Gut Microbiota Dysbiosis and Inflammation in BALB/c Mice. Int J Biol Macromol 2020, 167. [Google Scholar] [CrossRef]
- Zhang, Y.; Zuo, J.; Yan, L.; Cheng, Y.; Li, Q.; Wu, S.; Chen, L.; Thring, R.W.; Yang, Y.; Gao, Y.; et al. Sargassum Fusiforme Fucoidan Alleviates High-Fat Diet-Induced Obesity and Insulin Resistance Associated with the Improvement of Hepatic Oxidative Stress and Gut Microbiota Profile. J Agric Food Chem 2020, 68, 10626–10638. [Google Scholar] [CrossRef]
- Zheng, W.; Duan, M.; Jia, J.; Song, S.; Ai, C.; Li, S.; Wang, L.; Liu, B.; He, N.; Wang, Y.; et al. Sulfated Polysaccharides from Undaria Pinnatifida Improved High Fat Diet-Induced Metabolic Syndrome, Gut Microbiota Dysbiosis and Inflammation in BALB/c Mice. Int J Biol Macromol 2020, 167, 4773–4784. [Google Scholar] [CrossRef]
- Li, S.; Wang, L.; Liu, B.; He, N. Unsaturated Alginate Oligosaccharides Attenuated Obesity-Related Metabolic Abnormalities by Modulating Gut Microbiota in High-Fat-Diet Mice. Food Funct 2020, 11, 4773–4784. [Google Scholar] [CrossRef]
- Wu, Q.; Wu, S.; Cheng, Y.; Zhang, Z.; Mao, G.; Li, S.; Yang, Y.; Zhang, X.; Wu, M.; Tong, H. Sargassum Fusiforme Fucoidan Modifies Gut Microbiota and Intestinal Metabolites during Alleviation of Hyperglycemia in Type 2 Diabetic Mice. Food Funct 2021, 12, 3572–3585. [Google Scholar] [CrossRef]
- Cheng, Y.; Sibusiso, L.; Hou, L.; Jiang, H.; Chen, P.; Zhang, X.; Wu, M.; Tong, H. Sargassum Fusiforme Fucoidan Modifies the Gut Microbiota during Alleviation of Streptozotocin-Induced Hyperglycemia in Mice. Int J Biol Macromol 2019, 131, 1162–1170. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, J.; Mao, G.; Zuo, J.; Li, S.; Yang, Y.; Thring, R.W.; Wu, M.; Tong, H. Sargassum Fusiforme Fucoidan Alleviates Diet-Induced Insulin Resistance by Inhibiting Colon-Derived Ceramide Biosynthesis. Food Funct 2021, 12, 8440–8453. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Yang, L.; Wang, Z.; Huang, W. Bile Acid Nuclear Receptor FXR and Digestive System Diseases. Acta Pharm Sin B 2015, 5, 135–144. [Google Scholar] [CrossRef]
- Mandal, N.; Grambergs, R.; Mondal, K.; Basu, S.K.; Tahia, F.; Dagogo-Jack, S. Role of Ceramides in the Pathogenesis of Diabetes Mellitus and Its Complications. J Diabetes Complications 2021, 35, 107734. [Google Scholar] [CrossRef]
- Lee, M.-K.; Kim, I.-H.; Choi, Y.-H.; Nam, T.-J. A Peptide from Porphyra Yezoensis Stimulates the Proliferation of IEC-6 Cells by Activating the Insulin-like Growth Factor I Receptor Signaling Pathway. Int J Mol Med 2015, 35, 533–538. [Google Scholar] [CrossRef]
- Freile-Pelegrín, Y.; Robledo, D. Bioactive Phenolic Compounds from Algae. Bioactive Compounds from Marine Foods: Plant and Animal Sources. [CrossRef]
- Cotas, J.; Leandro, A.; Monteiro, P.; Pacheco, D.; Figueirinha, A.; Gonçalves, A.M.M.; da Silva, G.J.; Pereira, L. Seaweed Phenolics: From Extraction to Applications. Mar Drugs 2020, 18. [Google Scholar] [CrossRef]
- Yuan, Y.; Zheng, Y.; Zhou, J.; Geng, Y.; Zou, P.; Li, Y.; Zhang, C. Polyphenol-Rich Extracts from Brown Macroalgae Lessonia Trabeculate Attenuate Hyperglycemia and Modulate Gut Microbiota in High-Fat Diet and Streptozotocin-Induced Diabetic Rats. J Agric Food Chem 2019, 67, 12472–12480. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Yang, C.; Lin, G.; Chen, Y.; Miao, S.; Liu, B.; Zhao, C. Antidiabetic Potential of Green Seaweed Enteromorpha Prolifera Flavonoids Regulating Insulin Signaling Pathway and Gut Microbiota in Type 2 Diabetic Mice. J Food Sci 2019, 84, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Juturu, V.; Wu, A.H.; Ameer, K.; Messina, M.; Jr, E.J.; Duncan, A.; Messina, V.; Lynch, H.; Kiel, J.; Erdman Jr, J.W. The Health Effects of Soy: A Reference Guide for Health Professionals. Front Nutr 2022, 1–33. [Google Scholar]
- do Prado, F.G.; Pagnoncelli, M.G.B.; de Melo Pereira, G.V.; Karp, S.G.; Soccol, C.R. Fermented Soy Products and Their Potential Health Benefits: A Review. Microorganisms 2022, 10, 1–24. [Google Scholar] [CrossRef]
- Belobrajdic, D.P.; James-Martin, G.; Jones, D.; Tran, C.D. Soy and Gastrointestinal Health: A Review. Nutrients 2023, 15, 1959. [Google Scholar] [CrossRef]
- Clemente, T.E.; Cahoon, E.B. Soybean Oil: Genetic Approaches for Modification of Functionality and Total Content. Plant Physiol 2009, 151, 1030–1040. [Google Scholar] [CrossRef] [PubMed]
- Gu, C.; Pan, H.; Sun, Z.; Qin, G. Effect of Soybean Variety on Anti-Nutritional Factors Content, and Growth Performance and Nutrients Metabolism in Rat. Int J Mol Sci 2010, 11, 1048–1056. [Google Scholar] [CrossRef]
- Barac, M.; Stanojevic, S.; Pesic, M. Biologically Active Components of Soybeans and Soy Protein Products: A Review. Acta Periodica Technologica 2005, 266, 155–168. [Google Scholar] [CrossRef]
- Li, K.J.; Burton-Pimentel, K.J.; Vergères, G.; Feskens, E.J.M.; Brouwer-Brolsma, E.M. Fermented Foods and Cardiometabolic Health: Definitions, Current Evidence, and Future Perspectives. Front Nutr 2022, 9. [Google Scholar] [CrossRef]
- Kumari, M.; Kokkiligadda, A.; Dasriya, V.; Naithani, H. Functional Relevance and Health Benefits of Soymilk Fermented by Lactic Acid Bacteria. J Appl Microbiol 2022, 133, 104–119. [Google Scholar] [CrossRef] [PubMed]
- Elhalis, H.; Chin, X.H.; Chow, Y. Soybean Fermentation : Microbial Ecology and Starter Culture Technology. Crit Rev Food Sci Nutr 2023, 0, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Bahuguna, A.; Kumar, V.; Bodkhe, G.; Ramalingam, S.; Lim, S.M.; Joe, A.R.; Lee, J.S.; Kim, S.Y.; Kim, M. Safety Analysis of Korean Cottage Industries’ Doenjang, a Traditional Fermented Soybean Product: A Special Reference to Biogenic Amines. Foods 2023, 12. [Google Scholar] [CrossRef] [PubMed]
- Yi, S.H.; Hong, S.P. Characteristics of Bacterial Strains with Desirable Flavor Compounds from Korean Traditional Fermented Soybean Paste (Doenjang). Molecules 2021, 26. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, C.; Xiao, L.; Wang, S.; Wang, X.; Ma, K.; Ji, F.; Azarpazhooh, E.; Ajami, M.; Rui, X.; et al. Effects of Lactiplantibacillus Plantarum with Different Phenotypic Features on the Nutrition, Flavor, Gel Properties, and Digestion of Fermented Soymilk. Food Biosci 2023, 55, 103026. [Google Scholar] [CrossRef]
- Nguyen, N.-N.; Do, D.; Van, T.; Nguyen, V.; Tran, M.; Nguyen, Q.-D. Development of Dairy-free Soybean-based Yoghurt by Active Dry Starter Culture from Kombucha: An Investigation into Microencapsulation, Curd Formation, Protein and Texture Profiles during Storage. Int J Food Sci Technol 2024, 59. [Google Scholar] [CrossRef]
- Leonard, L.M.; Choi, M.S.; Cross, T.W.L. Maximizing the Estrogenic Potential of Soy Isoflavones Through the Gut Microbiome: Implication for Cardiometabolic Health in Postmenopausal Women. Nutrients 2022, 14. [Google Scholar] [CrossRef]
- do Prado, F.G.; Pagnoncelli, M.G.B.; de Melo Pereira, G.V.; Karp, S.G.; Soccol, C.R. Fermented Soy Products and Their Potential Health Benefits: A Review. Microorganisms 2022, 10. [Google Scholar] [CrossRef]
- Huang, L.; Zheng, T.; Hui, H.; Xie, G. Soybean Isoflavones Modulate Gut Microbiota to Benefit the Health Weight and Metabolism. Front Cell Infect Microbiol 2022, 12, 1–11. [Google Scholar] [CrossRef]
- Liu, B.; Qin, L.; Liu, A.; Uchiyama, S.; Ueno, T.; Li, X.; Wang, P. Prevalence of the Equol-Producer Phenotype and Its Relationship with Dietary Isoflavone and Serum Lipids in Healthy Chinese Adults. J Epidemiol 2010, 20, 377–384. [Google Scholar] [CrossRef]
- Paul, B.; Royston, K.J.; Li, Y.; Stoll, M.L.; Skibola, C.F.; Wilson, L.S.; Barnes, S.; Morrow, C.D.; Tollefsbol, T.O. Impact of Genistein on the Gut Microbiome of Humanized Mice and Its Role in Breast Tumor Inhibition. PLoS One 2017, 12, e0189756. [Google Scholar] [CrossRef] [PubMed]
- Hagan, M.; Fungwe, T. Determining the Effect of Seaweed Intake on the Microbiota: A Systematic Review. Functional Food Science 2023, 3. [Google Scholar] [CrossRef]
- Andres, S.; Abraham, K.; Appel, K.E.; Lampen, A. Risks and Benefits of Dietary Isoflavones for Cancer. Crit Rev Toxicol 2011, 41, 463–506. [Google Scholar]
- do Prado, F.G.; Pagnoncelli, M.G.B.; de Melo Pereira, G.V.; Karp, S.G.; Soccol, C.R. Fermented Soy Products and Their Potential Health Benefits: A Review. Microorganisms 2022, 10. [Google Scholar] [CrossRef]
- Teoh, S.Q.; Chin, N.L.; Chong, C.W.; Ripen, A.M.; How, S.; Lim, J.J.L. A Review on Health Benefits and Processing of Tempeh with Outlines on Its Functional Microbes. Future Foods 2024, 9, 100330. [Google Scholar] [CrossRef]
- Guetterman, H.M.; Huey, S.L.; Knight, R.; Fox, A.M.; Mehta, S.; Finkelstein, J.L. Vitamin B-12 and the Gastrointestinal Microbiome: A Systematic Review. Adv Nutr 2022, 13, 530–558. [Google Scholar] [CrossRef]
- Huang, H.; Krishnan, H.B.; Pham, Q.; Yu, L.L.; Wang, T.T.Y. Soy and Gut Microbiota: Interaction and Implication for Human Health. J Agric Food Chem 2016, 64, 8695–8709. [Google Scholar] [CrossRef] [PubMed]
- Mitsuoka, T. Prebiotics and Intestinal Flora. Biosci Microflora 2002, 21, 3–12. [Google Scholar] [CrossRef]
- Ma, Y.; Wu, X.; Giovanni, V.; Meng, X. Effects of Soybean Oligosaccharides on Intestinal Microbial Communities and Immune Modulation in Mice. Saudi J Biol Sci 2017, 24, 114–121. [Google Scholar] [CrossRef]
- Qiang, X.; YongLie, C.; QianBing, W. Health Benefit Application of Functional Oligosaccharides. Carbohydr Polym 2009, 77, 435–441. [Google Scholar] [CrossRef]
- Basson, A.R.; Ahmed, S.; Almutairi, R.; Seo, B.; Cominelli, F. Regulation of Intestinal Inflammation by Soybean and Soy-Derived Compounds. Foods 2021, 10, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Kono, K.; Murakami, Y.; Ebara, A.; Okuma, K.; Tokuno, H.; Odachi, A.; Ogasawara, K.; Hidaka, E.; Mori, T.; Satoh, K.; et al. Fluctuations in Intestinal Microbiota Following Ingestion of Natto Powder Containing Bacillus Subtilis Var. Natto SONOMONO Spores: Considerations Using a Large-Scale Intestinal Microflora Database. Nutrients 2022, 14, 3839. [Google Scholar] [CrossRef]
- Minamiyama, Y.; Okada, S. Miso: Production, Properties, And. Handbook of fermented functional foods 2003, 27.
- Jang, C.H.; Oh, J.; Lim, J.S.; Kim, H.J.; Kim, J.S. Fermented Soy Products: Beneficial Potential in Neurodegenerative Diseases. Foods 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Wambulwa, M.C.; Meegahakumbura, M.K.; Kamunya, S.; Wachira, F.N. From the Wild to the Cup: Tracking Footprints of the Tea Species in Time and Space. Front Nutr 2021, 8, 706770. [Google Scholar] [CrossRef]
- Stagg, G.V.; Millin, D.J. The Nutritional and Therapeutic Value of Tea—a Review. J Sci Food Agric 1975, 26, 1439–1459. [Google Scholar] [CrossRef]
- Bond, T.; Derbyshire, E. Tea Compounds and the Gut Microbiome: Findings from Trials and Mechanistic Studies. Nutrients 2019, 11, 1–13. [Google Scholar] [CrossRef]
- FAO International Tea Market : Market Situation, Prospects and Emerging Issues. food and Agriculture Organization of the United Nations 2022, 1–11.
- Gross, G.; Jacobs, D.M.; Peters, S.; Possemiers, S.; Van Duynhoven, J.; Vaughan, E.E.; Van De Wiele, T. In Vitro Bioconversion of Polyphenols from Black Tea and Red Wine/Grape Juice by Human Intestinal Microbiota Displays Strong Interindividual Variability. J Agric Food Chem 2010, 58, 10236–10246. [Google Scholar] [CrossRef]
- Chen, H.; Sang, S. Biotransformation of Tea Polyphenols by Gut Microbiota. J Funct Foods 2014, 7, 26–42. [Google Scholar] [CrossRef]
- Mandal, S.; DebMandal, M.; Pal, N.K.; Saha, K. Inhibitory and Killing Activities of Black Tea (Camellia Sinensis) Extract against Salmonella Enterica Serovar Typhi and Vibrio Cholerae O1 Biotype El Tor Serotype Ogawa Isolates. Jundishapur J Microbiol 2011, 4, 115–121. [Google Scholar]
- TOMIOKA, R.; TANAKA, Y.; SUZUKI, M.; EBIHARA, S. The Effects of Black Tea Consumption on Intestinal Microflora—A Randomized Single-Blind Parallel-Group, Placebo-Controlled Study. J Nutr Sci Vitaminol (Tokyo) 2023, 69, 326–339. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Xu, Y.; Yin, J. Black Tea Benefits Short-chain Fatty Acid Producers but Inhibits Genus Lactobacillus in the Gut of Healthy Sprague–Dawley Rats. J Sci Food Agric 2020, 100, 5466–5475. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Chen, Y.; Cheng, M.; Zhang, X.; Zheng, X.; Zhang, Z. The Modulatory Effect of Polyphenols from Green Tea, Oolong Tea and Black Tea on Human Intestinal Microbiota in Vitro. J Food Sci Technol 2018, 55, 399–407. [Google Scholar] [CrossRef]
- Sun, L.; Su, Y.; Hu, K.; Li, D.; Guo, H.; Xie, Z. Microbial-Transferred Metabolites of Black Tea Theaflavins by Human Gut Microbiota and Their Impact on Antioxidant Capacity. Molecules 2023, 28. [Google Scholar] [CrossRef] [PubMed]
- Szliszka, E.; Czuba, Z.P.; Domino, M.; Mazur, B.; Zydowicz, G.; Krol, W. Green Tea and Its Relation to Human Gut Microbiome. Molecules 2009, 14, 738–754. [Google Scholar]
- Liu, Y.C.; Li, X.Y.; Shen, L. Modulation Effect of Tea Consumption on Gut Microbiota. Appl Microbiol Biotechnol 2020, 104, 981–987. [Google Scholar] [CrossRef]
- Jin, J.-S.; Touyama, M.; Hisada, T.; Benno, Y. Effects of Green Tea Consumption on Human Fecal Microbiota with Special Reference to Bifidobacterium Species. Microbiol Immunol 2012, 56, 729–739. [Google Scholar] [CrossRef]
- Wang, L.; Zeng, B.; Liu, Z.; Liao, Z.; Zhong, Q.; Gu, L.; Wei, H.; Fang, X. Green Tea Polyphenols Modulate Colonic Microbiota Diversity and Lipid Metabolism in High-Fat Diet Treated HFA Mice. J Food Sci 2018, 83, 864–873. [Google Scholar] [CrossRef]
- United Nations, Department of Economic and Social Affairs, P.D. World Population Prospects: 2022 Revision. Available online: https://population.un.org/wpp/.
- Garduño-Diaz, S.D.; Khokhar, S. South Asian Dietary Patterns and Their Association with Risk Factors for the Metabolic Syndrome. Journal of Human Nutrition and Dietetics 2013, 26, 145–155. [Google Scholar] [CrossRef]
- Bishwajit, G.; Sarker, S.; Kpoghomou, M.-A.; Gao, H.; Jun, L.; Yin, D.; Ghosh, S. Self-Sufficiency in Rice and Food Security: A South Asian Perspective. Agric Food Secur 2013, 2, 10. [Google Scholar] [CrossRef]
- Pingali, P. Westernization of Asian Diets and the Transformation of Food Systems: Implications for Research and Policy. Food Policy 2007, 32, 281–298. [Google Scholar] [CrossRef]
- Bishwajit, G. Nutrition Transition in South Asia: The Emergence of Non-Communicable Chronic Diseases. F1000Res 2015, 4. [Google Scholar] [CrossRef]
- Misra, A.; Khurana, L.; Isharwal, S.; Bhardwaj, S. South Asian Diets and Insulin Resistance. British Journal of Nutrition 2009, 101, 465–473. [Google Scholar] [PubMed]
- Bhathal, S.K.; Kaur, H.; Bains, K.; Mahal, A.K. Assessing Intake and Consumption Level of Spices among Urban and Rural Households of Ludhiana District of Punjab, India. Nutr J 2020, 19, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V. RETRACTED ARTICLE: Seven Spices of India—from Kitchen to Clinic. Journal of Ethnic Foods 2020, 7, 23. [Google Scholar] [CrossRef]
- Pandey, I.R.; Pandey, P.R. Challenges and Opportunities in Value Chain of Spices in South Asia (SAARC Agricultural Centre (SAC)); 2017; Vol. 73; ISBN 9789843441713.
- Shahidi, F.; Hossain, A. Bioactives in Spices, and Spice Oleoresins: Phytochemicals and Their Beneficial Effects in Food Preservation and Health Promotion. 2018, 8–75. [CrossRef]
- Dacrema, M.; Ali, A.; Ullah, H.; Khan, A.; Di Minno, A.; Xiao, J.; Martins, A.M.C.; Daglia, M. Spice-Derived Bioactive Compounds Confer Colorectal Cancer Prevention via Modulation of Gut Microbiota. Cancers (Basel) 2022, 14. [Google Scholar]
- Gidwani, B.; Bhattacharya, R.; Shukla, S.S.; Pandey, R.K. Indian Spices: Past, Present and Future Challenges as the Engine for Bio-Enhancement of Drugs: Impact of COVID-19. J Sci Food Agric 2022, 102, 3065–3077. [Google Scholar] [CrossRef]
- Duda-Chodak, A. The Inhibitory Effect of Polyphenols on Human Gut Microbiota. Journal of Physiology and Pharmacology 2012, 63, 497–503. [Google Scholar]
- Hervert-Hernández, D.; Goñi, I. Dietary Polyphenols and Human Gut Microbiota: A Review. Food Reviews International 2011, 27, 154–169. [Google Scholar] [CrossRef]
- Lu, Q.Y.; Summanen, P.H.; Lee, R.P.; Huang, J.; Henning, S.M.; Heber, D.; Finegold, S.M.; Li, Z. Prebiotic Potential and Chemical Composition of Seven Culinary Spice Extracts. J Food Sci 2017, 82, 1807–1813. [Google Scholar] [CrossRef]
- Scazzocchio, B.; Minghetti, L.; D’archivio, M. Interaction between Gut Microbiota and Curcumin: A New Key of Understanding for the Health Effects of Curcumin. Nutrients 2020, 12, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.-Z.; Li, X.-Y.; Wang, S.; Shen, L.; Ji, H.-F. Bidirectional Interactions between Curcumin and Gut Microbiota in Transgenic Mice with Alzheimer’s Disease. Appl Microbiol Biotechnol 2020, 104, 3507–3515. [Google Scholar] [CrossRef] [PubMed]
- Dacrema, M.; Ali, A.; Ullah, H.; Khan, A.; Di Minno, A.; Xiao, J.; Martins, A.M.C.; Daglia, M. Spice-Derived Bioactive Compounds Confer Colorectal Cancer Prevention via Modulation of Gut Microbiota. Cancers (Basel) 2022, 14. [Google Scholar] [CrossRef]
- Peterson, C.T.; Vaughn, A.R.; Sharma, V.; Chopra, D.; Mills, P.J.; Peterson, S.N.; Sivamani, R.K. Effects of Turmeric and Curcumin Dietary Supplementation on Human Gut Microbiota: A Double-Blind, Randomized, Placebo-Controlled Pilot Study. J Evid Based Integr Med 2018, 23. [Google Scholar] [CrossRef] [PubMed]
- Di Cerbo, A.; Palmieri, B.; Aponte, M.; Morales-Medina, J.C.; Iannitti, T. Mechanisms and Therapeutic Effectiveness of Lactobacilli. J Clin Pathol 2015. [Google Scholar] [CrossRef]
- Bereswill, S.; Muñoz, M.; Fischer, A.; Plickert, R.; Haag, L.-M.; Otto, B.; Kühl, A.A.; Loddenkemper, C.; Göbel, U.B.; Heimesaat, M.M. Anti-Inflammatory Effects of Resveratrol, Curcumin and Simvastatin in Acute Small Intestinal Inflammation. PLoS One 2010, 5, e15099. [Google Scholar] [CrossRef] [PubMed]
- Fattori, V.; Hohmann, M.S.N.; Rossaneis, A.C.; Pinho-Ribeiro, F.A.; Verri, W.A. Capsaicin: Current Understanding of Its Mechanisms and Therapy of Pain and Other Pre-Clinical and Clinical Uses. Molecules 2016, 21. [Google Scholar] [CrossRef]
- Peng, Z.; Zhang, W.; Zhang, X.; Mao, J.; Zhang, Q.; Zhao, W.; Zhang, S.; Xie, J. Recent Advances in Analysis of Capsaicin and Its Effects on Metabolic Pathways by Mass Spectrometry. Front Nutr 2023, 10, 1–8. [Google Scholar] [CrossRef]
- Xiang, Q.; Tang, X.; Cui, S.; Zhang, Q.; Liu, X.; Zhao, J.; Zhang, H.; Mao, B.; Chen, W. Capsaicin, the Spicy Ingredient of Chili Peppers: Effects on Gastrointestinal Tract and Composition of Gut Microbiota at Various Dosages. Foods 2022, 11, 686. [Google Scholar] [CrossRef]
- Wang, F.; Huang, X.; Chen, Y.; Zhang, D.; Chen, D.; Chen, L.; Lin, J. Study on the Effect of Capsaicin on the Intestinal Flora through High-Throughput Sequencing. ACS Omega 2020, 5, 1246–1253. [Google Scholar] [CrossRef]
- Hui, S.; Liu, Y.; Chen, M.; Wang, X.; Lang, H.; Zhou, M.; Yi, L.; Mi, M. Capsaicin Improves Glucose Tolerance and Insulin Sensitivity through Modulation of the Gut Microbiota-bile Acid-FXR Axis in Type 2 Diabetic Db/Db Mice. Mol Nutr Food Res 2019, 63, 1900608. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Geng, W.; Chen, S.; Wang, L.; Rong, X.; Wang, S.; Wang, T.; Xiong, L.; Huang, J.; Pang, X. Ginger Alleviates DSS-Induced Ulcerative Colitis Severity by Improving the Diversity and Function of Gut Microbiota. Front Pharmacol 2021, 12, 632569. [Google Scholar] [CrossRef]
- Zhou, X.; Liu, X.; He, Q.; Wang, M.; Lu, H.; You, Y.; Chen, L.; Cheng, J.; Li, F.; Fu, X. Ginger Extract Decreases Susceptibility to Dextran Sulfate Sodium-Induced Colitis in Mice Following Early Antibiotic Exposure. Front Med (Lausanne) 2022, 8, 755969. [Google Scholar] [CrossRef]
- Van Hul, M.; Geurts, L.; Plovier, H.; Druart, C.; Everard, A.; Ståhlman, M.; Rhimi, M.; Chira, K.; Teissedre, P.-L.; Delzenne, N.M.; et al. Reduced Obesity, Diabetes, and Steatosis upon Cinnamon and Grape Pomace Are Associated with Changes in Gut Microbiota and Markers of Gut Barrier. American Journal of Physiology-Endocrinology and Metabolism 2018, 314, E334–E352. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.I.; Lee, J.H.; Song, Y.; Kim, Y.T.; Lee, Y.H.; Kang, H. Oral Consumption of Cinnamon Enhances the Expression of Immunity and Lipid Absorption Genes in the Small Intestinal Epithelium and Alters the Gut Microbiota in Normal Mice. J Funct Foods 2018, 49, 96–104. [Google Scholar] [CrossRef]
- Li, A.L.; Ni, W.W.; Zhang, Q.M.; Li, Y.; Zhang, X.; Wu, H.Y.; Du, P.; Hou, J.C.; Zhang, Y. Effect of Cinnamon Essential Oil on Gut Microbiota in the Mouse Model of Dextran Sodium Sulfate-Induced Colitis. Microbiol Immunol 2020, 64, 23–32. [Google Scholar] [CrossRef]
- Lu, Q.-Y.; Rasmussen, A.M.; Yang, J.; Lee, R.-P.; Huang, J.; Shao, P.; Carpenter, C.L.; Gilbuena, I.; Thames, G.; Henning, S.M.; et al. Mixed Spices at Culinary Doses Have Prebiotic Effects in Healthy Adults: A Pilot Study. Nutrients 2019, 11. [Google Scholar] [CrossRef]
- Khine, W.W.T.; Haldar, S.; De Loi, S.; Lee, Y.-K. A Single Serving of Mixed Spices Alters Gut Microflora Composition: A Dose–Response Randomised Trial. Sci Rep 2021, 11, 11264. [Google Scholar] [CrossRef] [PubMed]
- Hughes, J.; Pearson, E.; Grafenauer, S. Legumes-A Comprehensive Exploration of Global Food-Based Dietary Guidelines and Consumption. Nutrients 2022, 14. [Google Scholar] [CrossRef]
- Joshi, P.K.; Rao, P.P. Global and Regional Pulse Economies: Current Trends and Outlook. 2016, 26–57.
- Didinger, C.; Thompson, H.J. Defining Nutritional and Functional Niches of Legumes: A Call for Clarity to Distinguish a Future Role for Pulses in the Dietary Guidelines for Americans. Nutrients 2021, 13. [Google Scholar] [CrossRef]
- Sánchez, X.; Jimenez Martinez, C.; Dávila Ortiz, G.; Álvarez-González, I.; Madrigal-Bujaidar, E. Nutrient and Nonnutrient Components of Legumes, and Its Chemopreventive Activity: A Review. Nutr Cancer 2015, 67, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Didinger, C.; Thompson, H.J. Defining Nutritional and Functional Niches of Legumes: A Call for Clarity to Distinguish a Future Role for Pulses in the Dietary Guidelines for Americans. Nutrients 2021, 13. [Google Scholar] [CrossRef] [PubMed]
- Afriana, riza devi Role of Probiotic α-Galactosidases in the Reduction of Flatulence Causing Raffinose Oligosaccharides (RFOs. Angewandte Chemie International Edition, 6(11), 951–952. 2017, 6, 5–24.
- Khan, A.R.; Alam, S.; Ali, S.; Bibi, S.; Khalil, I.A. Dietary Fiber Profile of Food Legumes. Sarhad J. Agric 2007, 23, 763–766. [Google Scholar]
- Tosh, S.M.; Yada, S. Dietary Fibres in Pulse Seeds and Fractions: Characterization, Functional Attributes, and Applications. Food research international 2010, 43, 450–460. [Google Scholar]
- Wu, D.; Wan, J.; Li, W.; Li, J.; Guo, W.; Zheng, X.; Gan, R.Y.; Hu, Y.; Zou, L. Comparison of Soluble Dietary Fibers Extracted from Ten Traditional Legumes: Physicochemical Properties and Biological Functions. Foods 2023, 12. [Google Scholar] [CrossRef] [PubMed]
- Krishna, H.; Suthari, S. Application of Legume Seed Galactomannan Polysaccharides. In; 2020; pp. 97–113 ISBN 978-3-030-53016-7.
- Wu, G.-J.; Liu, D.; Wan, Y.-J.; Huang, X.-J.; Nie, S.-P. Comparison of Hypoglycemic Effects of Polysaccharides from Four Legume Species. Food Hydrocoll 2019, 90, 299–304. [Google Scholar] [CrossRef]
- Cichońska, P.; Ziarno, M. Legumes and Legume-Based Beverages Fermented with Lactic Acid Bacteria as a Potential Carrier of Probiotics and Prebiotics. Microorganisms 2022, 10. [Google Scholar] [CrossRef]
- Chen, Y.; Huang, C. Effects of Chickpea ( Cicer Arietinum ) on Metabolic Dysfunction by Modulation of Gut Microbiota in Diet- Induced Obese Mice. 2020.
- Monk, J.M.; Lepp, D.; Wu, W.; Graf, D.; McGillis, L.H.; Hussain, A.; Carey, C.; Robinson, L.E.; Liu, R.; Tsao, R.; et al. Chickpea-Supplemented Diet Alters the Gut Microbiome and Enhances Gut Barrier Integrity in C57Bl/6 Male Mice. J Funct Foods 2017, 38, 663–674. [Google Scholar] [CrossRef]
- Fernando, W.M.U.; Hill, J.E.; Zello, G.A.; Tyler, R.T.; Dahl, W.J.; Van Kessel, A.G. Diets Supplemented with Chickpea or Its Main Oligosaccharide Component Raffinose Modify Faecal Microbial Composition in Healthy Adults. Benef Microbes 2010, 1, 197–207. [Google Scholar] [CrossRef]
- Kang, S.; Xu, Y.; Zhang, Y.; Gao, P.; Guan, Y.; Ku, S.; Xu, J.; Zhu, X.; Li, H. Modulation of Gut Microbiota by Chickpea-Derived Proteins and Peptides with Antioxidant Capabilities. LWT 2023, 187, 115341. [Google Scholar] [CrossRef]
- Johnson, C.R.; Thavarajah, D.; Combs, G.F.; Thavarajah, P. Lentil (Lens Culinaris L.): A Prebiotic-Rich Whole Food Legume. Food Research International 2013, 51, 107–113. [Google Scholar] [CrossRef]
- Siva, N.; Johnson, C.R.; Richard, V.; Jesch, E.D.; Whiteside, W.; Abood, A.A.; Thavarajah, P.; Duckett, S.; Thavarajah, D. Lentil (Lens Culinaris Medikus) Diet Affects the Gut Microbiome and Obesity Markers in Rat. J Agric Food Chem 2018, 66, 8805–8813. [Google Scholar] [CrossRef]
- Graf, D.; Monk, J.M.; Lepp, D.; Wu, W.; McGillis, L.; Roberton, K.; Brummer, Y.; Tosh, S.M.; Power, K.A. Cooked Red Lentils Dose-Dependently Modulate the Colonic Microenvironment in Healthy C57Bl,/6 Male Mice. Nutrients 2019, 11, 1–21. [Google Scholar] [CrossRef]
- Hargreaves, S.M.; Raposo, A.; Saraiva, A.; Zandonadi, R.P. Vegetarian Diet: An Overview through the Perspective of Quality of Life Domains. Int J Environ Res Public Health 2021, 18. [Google Scholar] [CrossRef]
- Natrajan, B.; Jacob, S. “Provincialising” Vegetarianism: Putting Indian Food Habits in Their Place. Econ Polit Wkly 2018, 53, 54–64. [Google Scholar]
- Xiao, W.; Zhang, Q.; Yu, L.; Tian, F.; Chen, W.; Zhai, Q. Effects of Vegetarian Diet-Associated Nutrients on Gut Microbiota and Intestinal Physiology. Food Science and Human Wellness 2022, 11, 208–217. [Google Scholar] [CrossRef]
- Melina, V.; Craig, W.; Levin, S. Position of the Academy of Nutrition and Dietetics: Vegetarian Diets. J Acad Nutr Diet 2016, 116, 1970–1980. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, N.S.; Jaceldo-Siegl, K.; Sabate, J.; Fraser, G.E. Nutrient Profiles of Vegetarian and Nonvegetarian Dietary Patterns. J Acad Nutr Diet 2013, 113, 1610–1619. [Google Scholar] [CrossRef]
- Johnston, C.S. 28 - Vegetarian Diet and Possible Mechanisms for Impact on Mood. In; Mariotti, F.B.T.-V. and P.-B.D. in H. and D.P., Ed.; Academic Press, 2017; pp. 493–509 ISBN 978-0-12-803968-7.
- Iikura, M. 27 - Plant-Based Diets and Asthma. In; Mariotti, F.B.T.-V. and P.-B.D. in H. and D.P., Ed.; Academic Press, 2017; pp. 483–491 ISBN 978-0-12-803968-7.
- Kahleova, H.; Pelikanova, T. 21 - Vegetarian Diets in People With Type 2 Diabetes. In; Mariotti, F.B.T.-V. and P.-B.D. in H. and D.P., Ed.; Academic Press, 2017; pp. 369–393 ISBN 978-0-12-803968-7.
- Mann, J. 23 - Ischemic Heart Disease in Vegetarians and Those Consuming a Predominantly Plant-Based Diet. In; Mariotti, F.B.T.-V. and P.-B.D. in H. and D.P., Ed.; Academic Press, 2017; pp. 415–427 ISBN 978-0-12-803968-7.
- Tonstad, S.; Clifton, P. 20 - Vegetarian Diets and the Risk of Type 2 Diabetes. In; Mariotti, F.B.T.-V. and P.-B.D. in H. and D.P., Ed.; Academic Press, 2017; pp. 355–367 ISBN 978-0-12-803968-7.
- Yokoyama, Y.; Nishimura, K.; Barnard, N.D.; Miyamoto, Y. 22 - Blood Pressure and Vegetarian Diets. In; Mariotti, F.B.T.-V. and P.-B.D. in H. and D.P., Ed.; Academic Press, 2017; pp. 395–413 ISBN 978-0-12-803968-7.
- Yeh, M.-C.; Glick-Bauer, M.; Katz, D.L. 18 - Weight Maintenance and Weight Loss: The Adoption of Diets Based on Predominantly Plants. In; Mariotti, F.B.T.-V. and P.-B.D. in H. and D.P., Ed.; Academic Press, 2017; pp. 333–344 ISBN 978-0-12-803968-7.
- Orlich, M.J.; Siapco, G.; Jung, S. 24 - Vegetarian Diets and the Microbiome. In; Mariotti, F.B.T.-V. and P.-B.D. in H. and D.P., Ed.; Academic Press, 2017; pp. 429–461 ISBN 978-0-12-803968-7.
- Mangano, K.M.; Tucker, K.L. 17 - Bone Health and Vegan Diets. In; Mariotti, F.B.T.-V. and P.-B.D. in H. and D.P., Ed.; Academic Press, 2017; pp. 315–331 ISBN 978-0-12-803968-7.
- Nath, P.; Singh, S.P. 26 - Defecation and Stools in Vegetarians: Implications in Health and Disease. In; Mariotti, F.B.T.-V. and P.-B.D. in H. and D.P., Ed.; Academic Press, 2017; pp. 473–481 ISBN 978-0-12-803968-7.
- Key, T.J. 19 - Cancer Risk and Vegetarian Diets. In; Mariotti, F.B.T.-V. and P.-B.D. in H. and D.P., Ed.; Academic Press, 2017; pp. 345–354 ISBN 978-0-12-803968-7.
- Jung, J.G.; Kang, H.W. 25 - Vegetarianism and the Risk of Gastroesophageal Reflux Disease. In; Mariotti, F.B.T.-V. and P.-B.D. in H. and D.P., Ed.; Academic Press, 2017; pp. 463–472 ISBN 978-0-12-803968-7.
- Pem, D.; Jeewon, R. Fruit and Vegetable Intake: Benefits and Progress of Nutrition Education Interventions-Narrative Review Article. Iran J Public Health 2015, 44, 1309–1321. [Google Scholar]
- Walsh, S.; Hebbelinck, M.; Deriemaeker, P.; Clarys, P. 11 - Dietary Patterns in Plant-Based, Vegetarian, and Omnivorous Diets. In; Mariotti, F.B.T.-V. and P.-B.D. in H. and D.P., Ed.; Academic Press, 2017; pp. 175–196 ISBN 978-0-12-803968-7.
- van Berleere, M.; Dauchet, L. 13 - Fruits, Vegetables, and Health: Evidence From Meta-Analyses of Prospective Epidemiological Studies. In; Mariotti, F.B.T.-V. and P.-B.D. in H. and D.P., Ed.; Academic Press, 2017; pp. 215–248 ISBN 978-0-12-803968-7.
- Boutron-Ruault, M.-C.; Mesrine, S.; Pierre, F. 12 - Meat Consumption and Health Outcomes. In; Mariotti, F.B.T.-V. and P.-B.D. in H. and D.P., Ed.; Academic Press, 2017; pp. 197–214 ISBN 978-0-12-803968-7.
- Sun, C.; Li, A.; Xu, C.; Ma, J.; Wang, H.; Jiang, Z.; Hou, J. Comparative Analysis of Fecal Microbiota in Vegetarians and Omnivores. Nutrients 2023, 15. [Google Scholar] [CrossRef]
- de Jonge, N.; Carlsen, B.; Christensen, M.H.; Pertoldi, C.; Nielsen, J.L. The Gut Microbiome of 54 Mammalian Species. Front Microbiol 2022, 13, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Tomova, A.; Bukovsky, I.; Rembert, E.; Yonas, W.; Alwarith, J.; Barnard, N.D.; Kahleova, H. The Effects of Vegetarian and Vegan Diets on Gut Microbiota. Front Nutr 2019, 6. [Google Scholar] [CrossRef] [PubMed]
- Buell, P.D.; Anderson, E.N.; de Pablo Moya, M.; Oskenbay, M. Crossroads of Cuisine. In Crossroads of Cuisine; BRILL, 2020 ISBN 9789004432109.
- Xiao, W.; Zhang, Q.; Yu, L.; Tian, F.; Chen, W.; Zhai, Q. Effects of Vegetarian Diet-Associated Nutrients on Gut Microbiota and Intestinal Physiology. Food Science and Human Wellness 2022, 11, 208–217. [Google Scholar] [CrossRef]
- Sidhu, S.R.; Kok, C.W.; Kunasegaran, T.; Ramadas, A. Effect of Plant-Based Diets on Gut Microbiota: A Systematic Review of Interventional Studies. Nutrients 2023, 15. [Google Scholar] [CrossRef]
- Zhang, C.; Björkman, A.; Cai, K.; Liu, G.; Wang, C.; Li, Y.; Xia, H.; Sun, L.; Kristiansen, K.; Wang, J.; et al. Impact of a 3-Months Vegetarian Diet on the Gut Microbiota and Immune Repertoire. Front Immunol 2018, 9, 1–13. [Google Scholar] [CrossRef]
- Kim, M.; Hwang, S.; Park, E.; Bae, J. Strict Vegetarian Diet Improves the Risk Factors Associated with Metabolic Diseases by Modulating Gut Microbiota and Reducing Intestinal Inflammation. Environ Microbiol Rep 2013, 5, 765–775. [Google Scholar] [CrossRef]
- Buell, P.D.; Anderson, E.N.; de Pablo Moya, M.; Oskenbay, M. Crossroads of Cuisine; 2020; ISBN 9789004432109.
- Muratalieva, E.; Nendaz, M.; Beran, D. Strategies to Address Non-Communicable Diseases in the Commonwealth of Independent States Countries: A Scoping Review. Prim Health Care Res Dev 2022, 23. [Google Scholar] [CrossRef]
- Karabay, A.; Bolatov, A.; Varol, H.A.; Chan, M.-Y. A Central Asian Food Dataset for Personalized Dietary Interventions. Nutrients 2023, 15, 1728. [Google Scholar] [CrossRef]
- Auyeskhan, U.; Azhbagambetov, A.; Sadykov, T.; Dairabayeva, D.; Talamona, D.; Chan, M.Y. Reducing Meat Consumption in Central Asia through 3D Printing of Plant-Based Protein—Enhanced Alternatives—a Mini Review. Front Nutr 2023, 10. [Google Scholar] [CrossRef]
- Otunchieva, A.; Borbodoev, J.; Ploeger, A. The Transformation of Food Culture on the Case of Kyrgyz Nomads—A Historical Overview. Sustainability (Switzerland) 2021, 13. [Google Scholar] [CrossRef]
- Belaunzaran, X.; Bessa, R.J.B.; Lavín, P.; Mantecón, A.R.; Kramer, J.K.G.; Aldai, N. Horse-Meat for Human Consumption - Current Research and Future Opportunities. Meat Sci 2015, 108, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.; Huang, X.; Schooling, C.M.; Zhao, J.V. Red Meat Consumption, Cardiovascular Diseases, and Diabetes: A Systematic Review and Meta-Analysis. Eur Heart J 2023, 44, 2626–2635. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Guo, Z.; Lim, A.A.Q.; Zheng, Y.; Koh, E.Y.; Ho, D.; Qiao, J.; Huo, D.; Hou, Q.; Huang, W.; et al. Mongolians Core Gut Microbiota and Its Correlation with Seasonal Dietary Changes. Sci Rep 2014, 4, 5001. [Google Scholar] [CrossRef] [PubMed]
- Kilmer, P.D. Review Article: Review Article. Journalism 2010, 11, 369–373. [Google Scholar] [CrossRef]
- Kondybayev, A.; Loiseau, G.; Achir, N.; Mestres, C.; Konuspayeva, G. Fermented Mare Milk Product (Qymyz, Koumiss). Int Dairy J 2021, 119, 105065. [Google Scholar] [CrossRef]
- Kushugulova, A.; Löber, U.; Akpanova, S.; Rysbekov, K.; Kozhakhmetov, S.; Khassenbekova, Z.; Essex, M.; Nurgozhina, A.; Nurgaziyev, M.; Babenko, D.; et al. Dynamic Changes in Microbiome Composition Following Mare’s Milk Intake for Prevention of Collateral Antibiotic Effect. Front Cell Infect Microbiol 2021, 11, 1–11. [Google Scholar] [CrossRef]
- Holmes, A.D.; Spelman, A.F.; Tyson Smith, C.; Kuzmeski, J.W. Composition of Mares’ Milk as Compared with That of Other Species. J Dairy Sci 1947, 30, 385–395. [Google Scholar] [CrossRef]
- Nurgaziyev, M.; Aitenov, Y.; Khassenbekova, Z.; Akpanova, S.; Rysbekov, K.; Kozhakhmetov, S.; Nurgozhina, A.; Sergazy, S.; Chulenbayeva, L.; Ospanova, Z.; et al. Effect of Mare’s Milk Prebiotic Supplementation on the Gut Microbiome and the Immune System Following Antibiotic Therapy. Biodiversitas 2020, 21, 5065–5071. [Google Scholar] [CrossRef]
- Li, N.; Xie, Q.; Chen, Q.; Evivie, S.E.; Liu, D.; Dong, J.; Huo, G.; Li, B. Cow, Goat, and Mare Milk Diets Differentially Modulated the Immune System and Gut Microbiota of Mice Colonized by Healthy Infant Feces. J Agric Food Chem 2020, 68, 15345–15357. [Google Scholar] [CrossRef]
- Golzarand, M.; Mirmiran, P.; Jessri, M.; Toolabi, K.; Mojarrad, M.; Azizi, F. Dietary Trends in the Middle East and North Africa: An Ecological Study (1961 to 2007). Public Health Nutr 2012, 15, 1835–1844. [Google Scholar] [CrossRef] [PubMed]
- Eid, N.; Enani, S.; Walton, G.; Corona, G.; Costabile, A.; Gibson, G.; Rowland, I.; Spencer, J.P.E. The Impact of Date Palm Fruits and Their Component Polyphenols, on Gut Microbial Ecology, Bacterial Metabolites and Colon Cancer Cell Proliferation. J Nutr Sci 2014, 3, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Eid, N.; Osmanova, H.; Natchez, C.; Walton, G.; Costabile, A.; Gibson, G.; Rowland, I.; Spencer, J.P.E. Impact of Palm Date Consumption on Microbiota Growth and Large Intestinal Health: A Randomised, Controlled, Cross-over, Human Intervention Study. British Journal of Nutrition 2015, 114, 1226–1236. [Google Scholar] [CrossRef]
- Plummer, E.; Bulach, D.; Carter, G.; Albert, M.J. Gut Microbiome of Native Arab Kuwaitis. Gut Pathog 2020, 12, 1–9. [Google Scholar] [CrossRef]
- Correia, J.M.; Santos, I.; Pezarat-Correia, P.; Silva, A.M.; Mendonca, G.V. Effects of Ramadan and Non-Ramadan Intermittent Fasting on Body Composition: A Systematic Review and Meta-Analysis. Front Nutr 2021, 7, 1–19. [Google Scholar] [CrossRef]
- Song, D.-K.; Kim, Y.-W. Beneficial Effects of Intermittent Fasting: A Narrative Review. Journal of Yeungnam Medical Science 2023, 40, 4–11. [Google Scholar] [CrossRef]
- Visioli, F.; Mucignat-Caretta, C.; Anile, F.; Panaite, S.-A. Traditional and Medical Applications of Fasting. Nutrients 2022, 14, 433. [Google Scholar] [CrossRef]
- Fanti, M.; Mishra, A.; Longo, V.D.; Brandhorst, S. Time-Restricted Eating, Intermittent Fasting, and Fasting-Mimicking Diets in Weight Loss. Curr Obes Rep 2021, 10, 70–80. [Google Scholar] [CrossRef]
- Longo, V.D.; Mattson, M.P. Fasting: Molecular Mechanisms and Clinical Applications. Cell Metab 2014, 19, 181–192. [Google Scholar] [CrossRef]
- Anton, S.D.; Moehl, K.; Donahoo, W.T.; Marosi, K.; Lee, S.A.; Mainous, A.G.; Leeuwenburgh, C.; Mattson, M.P. Flipping the Metabolic Switch: Understanding and Applying the Health Benefits of Fasting. Obesity 2018, 26, 254–268. [Google Scholar] [CrossRef]
- De Cabo, R.; Cabello, R.; Rios, M.; López-Lluch, G.; Ingram, D.K.; Lane, M.A.; Navas, P. Calorie Restriction Attenuates Age-Related Alterations in the Plasma Membrane Antioxidant System in Rat Liver. Exp Gerontol 2004, 39, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Saglam, D.; Colak, G.A.; Sahin, E.; Ekren, B.Y.; Sezerman, U.; Bas, M. Effects of Ramadan Intermittent Fasting on Gut Microbiome: Is the Diet Key? Front Microbiol 2023, 14. [Google Scholar] [CrossRef] [PubMed]
- Ozkul, C.; Yalinay, M.; Karakan, T. Structural Changes in Gut Microbiome after Ramadan Fasting: A Pilot Study. Benef Microbes 2020, 11, 227–233. [Google Scholar] [CrossRef] [PubMed]
- UNICEF UNICEF, WHO, World Bank Group Joint Malnutrition Estimates; 2023.
- Angelakis, E.; Yasir, M.; Bachar, D.; Azhar, E.I.; Lagier, J.C.; Bibi, F.; Jiman-Fatani, A.A.; Alawi, M.; Bakarman, M.A.; Robert, C.; et al. Gut Microbiome and Dietary Patterns in Different Saudi Populations and Monkeys. Sci Rep 2016, 6, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kisuse, J.; La-ongkham, O.; Nakphaichit, M.; Therdtatha, P.; Momoda, R.; Tanaka, M.; Fukuda, S.; Popluechai, S.; Kespechara, K.; Sonomoto, K.; et al. Urban Diets Linked to Gut Microbiome and Metabolome Alterations in Children: A Comparative Cross-Sectional Study in Thailand. Front Microbiol 2018, 9, 1–16. [Google Scholar] [CrossRef]
- Therdtatha, P.; Shinoda, A.; Nakayama, J. Crisis of the Asian Gut: Associations among Diet, Microbiota, and Metabolic Diseases. Biosci Microbiota Food Health 2022, 41, 83–93. [Google Scholar] [CrossRef]

Table 1.
Bacterial genera identified in the Asian gut microbiome.
| Phylum | Order | Family | Genus | Specie | Reference |
|---|---|---|---|---|---|
| Firmicutes | Clostridiales | Clostridiaceae |
Faecalibacterium Clostridium |
[1,2] | |
| Eubacteriaceae | Eubacterium | [3] | |||
| Lachnospiraceae |
Blautia Roseburia Dorea Butyrivibrio |
Blautia obeum | [2,3,4,5] | ||
| Ruminococcaceae |
Ruminococcus Oscillospira |
Ruminococcus gnavus | [3,5,6] | ||
| Lactobacillales | Lactobacillaceae |
Lactobacillus | [7] | ||
| Bacillales | Leuconostocaceae | Weissella | [5] | ||
| Erysipelotrichales | Erysipelatoclostridiaceae | Catenibacterium | [7] | ||
| Veillonellales -Selenomonadales | Veillonellaceae | Megasphaera | [8] | ||
| Bacteroidetes | Bacteroidales | Bacteroidaceae | Bateroides |
Eubacterium callanderi Bacteroides fragilis |
[1,4] |
| Rikenellaceae | Alistipes | [3] | |||
| Prevotellaceae |
Prevotella Alloprevotella |
Prevotella copri Prevotella stercorea |
[1,2] | ||
| Proteobacteria | Bifidobacteriales | Bifidobacteriaceae | Bifidobacterium | Bifidobacterum catenulatum | [2,9] |
Table 2.
Categorization of Sub-Regions of Asia.
| Region | Countries in the Region | Reference |
|---|---|---|
| Central Asia | Kazakhstan, Kyrgyzstan, Tajikistan, Turkmenistan, Uzbekistan | [10] |
| Eastern Asia | China, Hong Kong, Macao (China), Japan, Mongolia, North Korea, South Korea | |
| Southern Asia | Afghanistan, Bangladesh, Bhutan, India, Iran, Maldives, Nepal, Pakistan, Sri Lanka | |
| South-Eastern Asia | Brunei, Cambodia, Indonesia, Laos, Malaysia, Myanmar, Philippines, Singapore, Thailand, Timor-Leste, Vietnam | |
| Western Asia | Armenia, Azerbaijan, Bahrain, Cyprus, Georgia, Iraq, Israel, Jordan, Kuwait, Lebanon, Oman, Qatar, Saudi Arabia, Palestine, Syrian Arab Republic, Turkey, United Arab Emirates |
Table 3.
Commonly Consumed Seaweeds in Asia and Their Common Names.
| Country | Scientific name of seaweed | Food product | Reference |
|---|---|---|---|
| Japan |
Saccharina japonica S. longissima |
Kombu | [21] |
| Porphyra/Pyropia spp. | Nori | ||
| Undaria pinnatifida | Wakame | ||
| Cladosiphon okamuranus | Mozuku | ||
| Sargassum fusiforme | Hijiki | ||
| Ecklonia bicyclis | Arame | ||
| Gelidium amansii | Tengusa | ||
| South Korea | Pyropia spp. | Gim | [22] |
| Nemalion vermiculare | Chamguksunamul | ||
| Gelidium amansii | Umutkasari | ||
| Gloiopeltis furcata | Bultunggasari | ||
| G. tenax | Pulgasari | ||
| G. complanata | Aekipulgasari | ||
| Monostroma complex | Hotparae | ||
| Ulva complex | Parae | ||
| Capsosiphon fulvescens | Maesaengi | ||
| Codium fragile | Cheonggak | ||
| Scytosiphon lomentaria | Korimae | ||
| Ecklonia stolonifera | Kompi | ||
| Undaria pinnatifida | Miyok | ||
| Saccharina japonica | Dasim | ||
| Pelvetia siliquosa | Thumbugi | ||
| Sargassum fusiforme | Tot | ||
| S. fulvellum | Mojaban | ||
| S. Horneri | Kwaengsaegi-mojaban | ||
| World | Ulva lactuca | Sea lettuce | [23] |
| Chondrus crispus | Irish Moss | ||
| Himanthalia elongata | Sea Spaghetti |
Table 4.
Fermented Soy Products and their main starter culture organism.
| Fermented Product | Fermentative Microorganism | Reference |
|---|---|---|
| Douchi Tempeh Miso Tofu |
Aspergillus oryzae Mucor spp. Rhizopus spp. Fusarium spp. |
[58] |
| Natto Kinema Chungkookjang |
Bacillus | [58] |
| Doenjang |
Bacteria B. subtilis Fungi Rhizopus spp., Mucor spp., Geotrichum spp., and Aspergillus spp. |
[59,60] |
| Fermented soy milk |
Lactiplantibacillus plantarum |
[61] |
| Soya yogurt |
Levilactobacillus brevis L. plantarum |
[62] |
Table 5.
Characteristics of the Vegetarian Dietary Patterns.
| Dietary Pattern | Characteristics | Reference |
|---|---|---|
| Vegan | Excludes all products of animal origin | Adopted and modified from [150] |
| Lacto-Ovo Vegetarians | Exclude red meat and poultry include dairy and eggs | |
| Raw vegan | Based vegetables, fruits, nuts, seeds, legumes, and grains. | |
| Pescatarian | Exclude red meat and poultry include eggs, dairy and fish | |
| Lacto-vegetarian | Exclude eggs, fish, red meat and poultry include dairy | |
| Semi-vegetarian | Consume red meat, poultry and fish no more than once a week |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



