
Submitted:
23 September 2024
Posted:
23 September 2024
You are already at the latest version
Abstract
Quinoa (Chenopodium quinoa, Amaranthaceae) is a pseudocereal native to the Andes of South America that contains high protein content and adequate nutrient levels. Peru possesses an abundant morphological diversity of quinoas and is among the top producers and exporters worldwide of this precious crop. However, knowledge about the genetic and population components of quinoa from the Peruvian Andes is still limited. We here used 13 microsatellite markers to determine the genetic diversity and population structure of 105 landraces of quinoa cultivated in 11 provinces of Ayacucho, the southern Peruvian Andes. A total of 285 bands were manually scored, generating a 105 x 285 presence/absence data set. Principal coordinate analysis, and similar to a dendrogram using UPGMA clustering algorithm, showed that quinoa from Ayacucho is grouped into three clusters without a clear geographic component. Estimation of the genetic diversity indices was conducted considering the three populations (C1: south 1, C2: south 2, C3: north) determined by STRUCTURE analysis, showing mean expected heterozygosity was 0.08. The highest population divergence (FST) was exhibited for C2 and C3 (0.03), whereas the lowest for C1 and C3 (0.02). Analysis of molecular variance revealed the greatest variation within populations (80.07%) and indicated that variability between populations is 19.93%. Microsatellite markers were effective; however, more studies of the genetic components of quinoa from other Peruvian Andean localities are still needed. We expect this work helps pave the way towards the development of modern breeding programs of quinoa in Peru with accurate strategies of conservation of this nutritious crop.
Keywords:
Andean crop
; molecular markers
; SSR
; germplasm
1. Introduction
Quinoa (Chenopodium quinoa Willd), pseudocereal of the Amaranthaceae family, is native crop of the South American Andes, it is an allotetraploid (2n = 4X=36) crop with disomic inheritance [1,2]. This crop originated by hybridization of an unknown female American Chenopodium diploid (AA genome) with an unknown male old world diploid species (BB genome), dating back 3.3 to 6.3 million years possibly in North America [3]. This was confirmed by molecular investigations of spacer regions in 5S rDNA showing two different non-transcribed sequence (NTS) clases [4], two homoeologous SOS1 loci (cqSOS1A and cqSOS1B) [4], two distinct homoeologs of GBSS1 gene (GBSS1A and GBSS1B) [5] and two distinct subgenomes by FTL intron markers [6]. Archaeological evidence of its domestication and use indicates that it was 5,000 years ago before Christ (B.C:) in Ayacucho [7,8,9], also in Puno 3,000 years ago (b.c.); from which quinoa evolved and adapted to particular agroecosystems. Currently cultivated from sea level to 4,000 meters above sea level (m.a.s.l.)[10].
The importance of quinoa is due to its nutritional balance. Its average protein content is 15.33%, fat 6.93%, ash 3.42%, carbohydrates 68.35% and fiber 4.78% [11]. Quinoa is appreciated also due to its value biologically determined by the high content of essential amino acids for human nutrition, including high lysine content compared to other cereals [12], omega-6 unsaturated fatty acids, dietary fiber and phytosterols [13], with a low glycemic index [12,13,14]. Due to its nutritional quality, it is considered a functional food [15]. Quinoa is a crop that contributes to the food security of humanity [16], and due to its versatility for industrialization its cultivation has spread from the centers of origin and domestication to the entire world. It was introduced in the 1970s in England, and thereafter in Denmark and in the European Union [17]. This crop was successfully tested in North America and Africa and has been cultivated in the USA since the early 1980s [18]. It was successfully introduced in India in the early 1990s [10] and was introduced in Pakistan in 2007 [19]. Field tests have been conducted in Japan in the climatic conditions of Southern Kanto District [20]. Since the year 2007, quinoa has been cultivated in the deserts of Southern Sinai for promoting this crop in Egypt [18]. Results also indicated quinoa showed a high potential of adaptation in Morocco [21].
The diversity of the Peruvian quinoa has been classified by its ecological adaptation. Thus, first, native highland varieties have been identified in regions of Titicaca adapted to ultraviolet radiation at altitudes higher than 3850 m.a.s.l called Kancolla, Blanca de July, Huariponcho, Witulla, and Kellu [17]; second, other native varieties adapted to soils with a high salt concentration, called Pandela, Utusaya, Toledo, and Achachino [22]; third, native varieties of inter-Andean valleys called Amarilla de Marangani, Blanca de Junín, Acostambo, Roja Coporaque, Nariño [23]; fourth, to arid zones, called Antahuara, Ucha, Ccoyto, and Roja Ayacuhana [9]; fifth, adapted to sea level, called Quinoa Blanca, Kinwamapuche, Lito, Faro and Islunga [24]; sixth, also native jungle varieties, adapted to conditions of high humidity and temperatures throughout the year, such as those grown in the Amazon [23].
Different molecular markers were used to characterize the genetic diversity of quinoa, RAPD [25,26,27], AFLP[28,29], ISSR [30], INDEL [31], SRAP [32,33], and SCoT [34]. However, SSR markers high reproducibility and co-dominance, has made them the preferred marker when studying the genetics of quinoa populations across Latin America [35,36,37,38,39,40,41,42,43] Christiansen et al. [35] reported the genetic diversity of international collections of quinoas conserved in greenhouses using microsatellite markers, which included accessions from Peru consisting of 20 accessions from Puno and three from Cusco, none from Ayacucho. Therefore, the present work is the first report of the genetic diversity of quinoa from Ayacucho using microsatellites.
Ayacucho, located in the central-southern Andean zone of Peru, is one of the microcenters of origin and domestication of quinoa [7,8,9]. It has varied ecological niches from 1,100 to 4,500 m.a.s.l. and is home to living germplasms of cultivated native quinoa identified with Quechua and/or Spanish names based on some particular morphological characteristic (often grain color). Studies of this genetic pool are very limited; most information is mainly based on morphological characterization [44], just one molecular study with SSR was reported [45]. On the other hand, Ayacucho suffered the impact of armed social violence in the 80's and 90's of the last century, causing massive migration of local communities to large cities mainly Huamanga and Lima [46,47], abandoning the cultivation of quinoa and stewardship of the native germplasm biodiversity.
In this study, the genetic diversity and population structure of a collection of 105 landraces of quinoa cultivated in all the 11 provinces of Ayacucho was assessed with 13 SSRs. The results encourage us to strengthen its conservation and give us baseline knowledge to design and start a modern breeding program for quinoa with molecular tools in favor of rural communities of the Andes.
2. Materials and Methods
2.1. Plant Material and DNA Extraction
One hundred and five landraces of quinoas from the Ayacucho geographical department were collected from farmer fields (on-farm) (Table S1). Samples of these landraces were obtained from the 11 provinces of Ayacucho: five from Huanta (H), six from La Mar (LM), 38 from Huamanga (Ha), seven from Cangallo (C), five from Vilcashuaman (V), four from Victor Fajardo (VF), two from Huancasancos (HS), 17 from Sucre (S), nine from Lucanas (L), six from Parinacochas (P) and seven from Paucar del Sara Sara (PS); including the impact area of the Camisea Gas Project (CGP).
Seeds of quinoa were germinated on filter paper in petri dishes soaked with distilled water. Genomic DNA was isolated from the cotyledons and radicles using CTAB method [48] with modifications. Quantity and quality of DNA were verified through absorbance ratio 260/280 using a BioSpectrometer (Eppendorf EP6137000015). DNA integrity was evaluated in 1% agarose gel electrophoresis.
2.2. DNA Amplification
Ten SSR markers [35,49,50] were used to fingerprint 105 quinua landraces. Label primers were synthesized by Integrated DNA Technologies (IDT). The forward primers included an additional sequence M13 (Li-Cor), 5'-CACGACGTTAAAACGAC-3', to binding of the fluorogen IRDye 700/800nm (Li-Cor). Amplification was performed using the M13-tailing method (Schuelke 2000) in a 10 μl volume for each reaction. The master mix was preparaded according to Schuelke (2000) with modifications, using 2 μl of Gotaq Buffer (1X), 0.1 μl of Taq Polymerase (0.05 U μl −1 ), 1.0 μl of MgCl2 (2.5 mM), 0.2 μl of dNTPs (0.2 mM) (Promega, USA), 0.6 μl of IR-Dye M13–700 or 800 (0.02 μM) (LI-COR Bioscience USA), 0.3 μl of primers forward and reverse (0.3 μM), 2 ul of the DNA sample (10 ng μl −1 ), and 3.5 ul NFW (Sigma, USA). For PCR reactions, we used the following temperature profile in Eppendorf Mastercycler Pr: 4 min at 94 °C, followed by 30 cycles of 45 seconds at 94 °C, one step of 4 min at 94°C, followed by 30 cycles of 45 s at 94°C, 30 s at TM °C (Table 2), 10 m at 72°C , and finilizing 4 min at 72°C.
We used LI-COR Bioscience Model 4300 DNA Analyser protocol to amplify the products. Samples were diluted with a BlueSTOP solution 4300 (1:1), denatured for 5 min at 94 °C and snap chilled on ice; a 1 μl sample was loaded into each well of a polyacrylamide gel composed of 20 ml of KB Matrix Gel Plus 6.5%, 150 μl of 10% ammonium persulfate and 15 μl of TEMED (tetramethylethylenediamine) with the Tris borate-EDTA buffer. The pre-run was programmed at 1500 V and 40 W per 20 min to focus the laser at 700 and 800 nm. To visualize the amplified SSR DNA fragments and determine their molecular weight, the Li-Cor DNA Analyzer 4300 system was employed with polyacrylamide gel to detect differences of up to 1 bp. Band size was automatically determined using the SAGA Generation V 3.2 software (Li-Cor, Lincoln, NE).
2.3. Data Analysis
Microsatellite amplification band patterns were scored in a data matrix, considering presence (1) or absence (0). Since the quinoa samples is an allotetraploid species that exhibits disomic inheritance [3], this crop possesses two copies of each of these two ancestral species, which makes its complete genome equal to 4 copies (2n=4x). On the other hand, by exhibiting disomic segregation, recombination only occurs between chromosomes of the same ancestral species, achieving that only two copies of a locus are inherited at most [51].
Allelic diversity for each polymorphic marker was estimated by heterozygocity (H) of each SSR, which was calculated according to Nei [52]:
H = 1 − ∑pi2
where pi is the allele frequency of the i-th allele of a certain SSR locus.
The polymorphism information content (PIC) of each population cluster with SSR marker was calculated according to:
PIC = 1 – [f i2 + (1 - f i)2 ]
where, fi is the frequency of the amplified band (1) and (1 - fi) is frequency of absence of band (0) [53].
Genetic distances based on Provesti’s coefficient [54] was calculated with R software v.4.2.2 [55], then a dendogram with the UPGMA clustering algorithm with 1000 bootstap replicates was generated using poppr package v2.9.2 [56]. Principal coordinate analysis (PCoA) was conducted using the ade4 v.1.7–16 [57] and adegenet v.2.1.3 [58] packages in R. Number of populations (K) were set from 1 to 10 by k-means clustering with 100,000 iterations. Population structure was estimated by STRUCTURE program v.2.3 [59] with ten runs for each number of populations (K value) ranging from 1 to 10 with a burnin length of 50,000 Monte Carlos iterations, which was followed by 150,000 iterations. An admixture model with no previous population information was considered; all other parameters were set to default values. Estimation of the most likely number of clusters was calculated by the Evanno method [60]. Membership probabilities ≥ 0.8 or the maximum membership probability was adopted to divide the quinua samples into different clusters. Population struture plots were generated with pophelper v.2.3.1 package [61] in R.
Clusters determined by STRUCTURE were used to conduct an analysis of molecular variance (AMOVA) with poppr R package. In addition, using the same package we calculated: (i) Shannon–Wiener index, (ii) Simpson’s index, and (iii) Nei’s gene diversity (expected heterozygosity). The degree of gene differentiation among clusters in terms of allele frequencies (FST) was estimated using the following formula:
Fst = 1 - (Hs/Ht)
where HS is the average expected heterozygosity estimated from each cluster and Ht is expected heterozygosity in the total cluster as estimated from the pooled allele frequencies.
3. Results
3.1. SSR Analysis
All 13 SSR primers generated well-defined bands with fragment sizes, according to previous studies [35,49,50]. SSR alleles ranged from 6 (accession Ha194) to 25 (accession L117) with an average of 15.2 per accession, with a total of 1597 alleles for all 105 quinoa individuals (Table S1). The 13 SSRs primers used in this study yielded 285 bands, generating a 105 x 285 presence/absence (PA) data set. Primer KAAT007 presented the highest number of alleles (54), whereas QAAT081 the lowest (6) with an average of 21.9. In addition, average of polymorphic alleles was 15.8 per primer. The gene diversity ranged from 0.7 (primer QAAT081) to 0.96 (primer KAAT007) (Table S2). Three non-polymorphic fragments were discarded by adegenet package in R, resulting in a final matrix of 105 x 282.

Table 1.
List of unique alleles with simple sequence repeat (SSR) markers in each sampling site in Ayacucho.
Table 1.
List of unique alleles with simple sequence repeat (SSR) markers in each sampling site in Ayacucho.
| Province | Accession | Common name | Unique alleles (primer name and base pair) | N° unique alleles |
|---|---|---|---|---|
| Sucre | S5 | Quinua | KAAT007_315 | 17 |
| S6 | Quinua | QGAA1_143 | ||
| S7 | Quinua | KAAT007_261, KAAT007_276, KGA16_194, QGA028_196 | ||
| S8 | Atoqpa Quinua | QAAT012_189, QGA028_214, QGAA1_236 | ||
| S9 | Quinua | KAAT007_294, QAAT84_167 | ||
| S22 | Quinua | KCAA011_205 | ||
| S39 | Quinua | QCA14_195 | ||
| S71 | Quinua | KCAA011_245 | ||
| S75 | Quinua | QGAA1_275 | ||
| S83 | Quinua | QGA17_193 | ||
| S92 | Quinua | KAAT007_335 | ||
| La Mar | LM26 | Quinua | KAAT007_300, KAAT007_345 | 3 |
| LM64 | Quinua | KAAT007_350 | ||
| Vilcashuaman | V60 | Quinua | KAAT007_368 | 2 |
| V61 | Quinua | KAAT007_329 | ||
| Victor Fajardo | VF41 | Quinua | KGA20_230 | 3 |
| VF73 | Quinua | KAAT007_326 | ||
| VF79 | Quinua | KCAA011_224 | ||
| Huanca Sancos | HS16 | Quinua | QGA17_182 | 2 |
| HS98 | Quinua | QAAT84_188 | ||
| Lucanas | L115 | Quinua | KAAT007_293 | 7 |
| L116 | Quinua | KGA20_182 | ||
| L117 | Quinua | QGAA1_230 | ||
| L119 | Quinua | KAAT007_299, QAAT84_206 | ||
| L120 | Quinua | QAAT012_240, QGA028_230 | ||
| Parinacochas | P124 | Quinua | KCAA011_230 | 5 |
| P125 | Quinua | QAAT84_215 | ||
| P128 | Quinua | KCAA011_263, QAAT081_ 216, QGAA1_317 | ||
| Cangallo | C34 | Quinua | KAAT007_282 | 3 |
| C186 | Utuloñawi | QAAT050_206 | ||
| C187 | yuraq quinua | QGAA1_430 | ||
| Huamanga | Ha10 | Quinua | QGAA1_245, QGAA1_266 | 28 |
| Ha14 | Quinua | KAAT007_222 | ||
| Ha20 | Quinua | KGA20_226, QGA028_208 | ||
| Ha21 | Quinua | KAAT007_210, KCAA011_238 | ||
| Ha24 | Quinua | QAAT012_216 | ||
| Ha47 | Quinua | KAAT007_297, QAAT84_182 | ||
| Ha50 | Quinua | KAAT007_231, KAAT007_303 | ||
| Ha56 | Quinua | QGA17_206 | ||
| Ha58 | Quinua | KGA16_192 | ||
| Ha62 | Quinua | KAAT007_371 | ||
| Ha67 | Quinua | KGA16_193 | ||
| Ha70 | Quinua | KAAT007_374 | ||
| Ha76 | Quinua | QGA028_212 | ||
| Ha81 | Quinua | QGAA1_239 | ||
| Ha82 | Quinua | QAAT071_188 | ||
| Ha85 | Quinua | QAAT071_146 | ||
| Ha94 | Quinua | KAAT007_296, KAAT007_344 | ||
| Ha96 | Quinua | KAAT007_227 | ||
| Ha189 | yuraq quinua | QGA17_150 | ||
| Ha190 | qello quinua | QAAT050_210 | ||
| Ha194 | Realquinua | QAAT050_256 | ||
| Ha197 | Yuraq salcedo | QGAA1_380 | ||
| Huanta | H28 | Quinua | KAAT007_336, QGAA1_362 | 4 |
| H36 | Quinua | KAAT007_252 | ||
| H203 | yuraq quinua | QCA14_245 | ||
| Total | 74 |
3.2. Genetic Diversity and Population Structure
Landraces of quinoa from the 11 provinces of Ayacucho exhibited low genetic diversity values. Shannon-Wiener index of diversity ranged from 0.69 (Huancasancos) to Huamanga (3.64). Similarly, quinoas from Huancasancos presented the lowest Simpson´s index of diversity (lambda) (0.5), while quinoas from Huamanga the highest (0.97). Genetic diversity (i.e. expected heterozygosity) varied from 0.08 (Paucar del Sara Sara) to 0.11 (Victor Fajardo) (Table S3). Our PCoA, based on the pairwise genetic distance matrix of 105 quinoas accessions depicted these are not grouped based on their geographic province of origin. A total of three groups were observed. The first axis explained 9.11% and second 8.18% of the variance (Figure 1A). Similarly, the UPGMA dendrogram based on Provesti’s genetic distance also revealed a lack of clustering based on the geographic province of origin of quinoas. Instead, this germplasm is contained in three major clades (Figure S1).
Interestingly, when native quinoa accessions were labelled according to their geographic zone of origin (north, and south) in Ayacucho, our PCoA revealed the following three clusters: (C1: south 1, C2: south 2, C3: north) (Figure 1B). However, these groups presented admixture with few samples from different origin but showing a prevalence of samples from the corresponding geographic zone of origin (Figure S2). Similarly, our dendrogram represented also three clusters (Figure 2). The best K value (number of populations) was three for our data set (Figure S3): i) cluster 1 included 39 accessions, ii) cluster 2 with 44 accessions and iii) cluster 3 contained 22 accessions (Figure 3, Table 2). STRUCTURE analysis showed no admixture for quinoa germplasm, except very low for accession Ha67 (Figure 3).
Genetic diversity index estimates for these three clusters (C) was limited, ranging from 0.06 (C3) to 0.1 (C2). The Shannon–Wiener index (H) ranged from 3.09 (C3) to 3.78 (C2) and Simpson’s index from 0.96 (C3) to 0.98 (C2). Polymorphic information content ranged from 0.09 to 0.1. Moreover, cluster 3 exhibited the lowest percentage of polymorphic loci (52.48%), whereas cluster 1 the highest (73.05%) (Table 3).
Table 2.
Origin of the 105 accessions of quinua among the three populations (clusters) determined by the STRUCTURE analysis.
Table 2.
Origin of the 105 accessions of quinua among the three populations (clusters) determined by the STRUCTURE analysis.
| Province | Cluster 1 | Cluster 2 | Cluster 3 |
|---|---|---|---|
| Cangallo | 1 | 4 | 2 |
| Huamanga | 11 | 11 | 16 |
| La Mar | 4 | 1 | 0 |
| Huanta | 2 | 0 | 3 |
| Vilcashuaman | 0 | 4 | 1 |
| Huancasancos | 1 | 1 | 0 |
| Victor Fajardo | 1 | 3 | 0 |
| Sucre | 12 | 5 | 0 |
| Lucanas | 0 | 9 | 0 |
| Parinacochas | 0 | 6 | 0 |
| Paucar del Sara Sara | 7 | 0 | 0 |
Population divergence (FST) between clusters 2 and 3 revealed the highest genetic difference (0.03), and the lowest Fst was observed between clusters 1 and 3 (0.02) (Table 4). Analysis of molecular variance showed that 19.93% of the total variation was found between populations of quinoa while 80.07% was within clusters (Table 5).
4. Discussion
4.1. Microsatellite Diversity
A total of 1597 different SSR alleles were obtained using 105 quinoa accessions from 11 provinces of Ayacucho with 13 SSR markers. The number of different alleles obtained is much greater than those reported (420 alleles) by Christensen et al. [35], despite having used 36 SSR marker loci and 152 quinoa accessions (samples from Peru-Puno, Bolivia, Argentina, Chile and USA). This significant difference is possibly a consequence of the type of sample used, which in large percentage were personal collections donated for the research of Christensen et al. [35], while we used samples obtained in the field in each province of Ayacucho - Peru, geographically close to the Pikimachay cave, where archaeological remains of quinoa from seven thousand years ago were found [7,8,9]. Nine of the microsatellite markers employed in this study are named with the initial letter Q (QAAT, QCA, QGA) developed by Mason et al [49], whose PCR product sizes (pb) and number of alleles observed do not differ much from those. Four other markers correspond to the names that begin with the letter K (KAAT, KCAA and KGA) developed by Jarvis [50], in which there are differences in the sizes of the PCR products and number of alleles observed, probably due to the difference of the biological material used.
The expected heterozygosity (HE) values of the 13 microsatellite loci used in this study are in the range of 0.7 to 0.96 with a mean of 0.83. These values are higher than those reported by Mason et al. [49] and Christensen et al. [35] whose values were 0.57 and 0.75, respectively. Markers with H ≥ 0.1 are polymorphic, while markers with H ≥ 0.7 are highly polymorphic according to Ott [62]. According to this definition, these 13 SSR markers are considered highly polymorphic, possibly as a consequence of the random sample obtained in Ayacucho and the use of the Saga Microsatellite software technology of the LI-COR biosciences sequencer.
The number of unique alleles was determined in each province of Ayacucho, adding up to 72. When the provinces were labelled as north, center and south of Ayacucho, a greater number of unique alleles were found in northern Ayacucho (Huamanga and Huanta) with 32 unique alleles (48.6%). Also, there is an important group of unique alleles in the south of Ayacucho (Lucanas, Parinacohas, Sucre and especially Paucar del Sara Sara) that presented 29 unique alleles (42.3%). The little residual difference corresponds to the center of Ayacucho. The greater number of unique alleles and heterozygosity in northern Ayacucho could be related to the early domestication of quinoa by the Pikimachay inhabitants, whose archaeological findings are located in the province of Huamanga [63] dated in 13 thousand years BP, which Lumbreras [8] considers evidence of plant domestication (quinoa, amatantus, chili, squash dated to 5500 to 4200 years BP). This information suggests that quinoas from Ayacucho may possess ancestral genotypes and possible hybridization with wild relatives that coexist in the countryside that would be generating the appearance of unique alleles.
4.2. Population Structure
Based on the clustering result, the best K value was three for our data set. It is noteworthy that the majority of the accessions maintain genetic purity in each cluster, while some quinoa accessions showed a level of admixture that varies from less than 3% (LM 55, Ps49, H28, LM26, S39, Ha77, LM64, Ha202, Ha198, Ha191, Ha194, Ha190) up to 5% (LM54, H203) and only a sample with 25% (Ha67). This result indicates the quinoa germplasm employed in this work corresponds to three different gene pools or genetic groups that retain their genetic purity. The identification of three populations of quinoa may be explained due to the low levels of allogamy of quinoa.
Similarly, our UPGMA dendrogram demonstrated quinoa from Ayacucho is contained within three major groups. Salazar et al. [42] reported similar results when 84 quinoa genotypes from Ecuador were evaluated with SSR markers. They also indicated quinoas from Ecuador are not grouped based on their geographic origin. Ayacucho is located in the southern Andes of Peru, with its capital Huamanga as part of the commercial, political and touristic attraction where inhabitants of the surrounding provinces (Huanta, Cangallo and Vilcashauaman) converge. While the provinces of Lucanas, Parinacochas and Paucar del Sara-Sara have less flow to Huamanga, mainly due to greater distance and difficult direct access only by land, still under construction. Consequently, this affects the grouping due to movement of quinoa genotypes between nearby provinces in north or south Ayacucho. Therefore, one group to northem Ayacucho (cluster 3) and two groups existing in southern Ayacucho (clusters 1 and 2).
Population structure analysis was performed on the complete 129 quinoa accessions from USDA and private collection based on the 147 markers: 85 INDEL, 14 gSSR and 48 EST-SSR [31], although they only identified two groups for quinoa accessions, the mountain and coastal quinoa populations. Meanwhile, in the present study the 105 quinoa accessions are only from the mountains and inter Andean valley, although SSR alleles evidencing three groups genetic of this native quinoa conserved in the Ayacucho Andes.
4.3. Genetic Diversity
We used microsatellite or simple sequence repeats (SSRs) markers as they provide greater benefits in terms of identifying polymorphisms, multi-allelism, and codominant inheritance modes, accuracy, reproducibility and enable the detection of genetic diversity between individuals within species [50]. A high Shannon–Wiener index (H) value was determined for the quinoas, showing that although throughout Ayacucho there is high genetic diversity of quinoa, in the south of Ayacucho (Cluster 1, 2) there is greater genetic diversity than in northern Ayacucho (Cluster 3). Simpson’s index was also high, indicating high genetic variability. Although the expected heterozygosity (HE) were low in general, the highest HE indices were found in cluster 1 and 2. The same pattern was observed for polymorphic loci (PPL), confirming the genetic diversity difference of the south vs. northern Ayacucho.
Fuentes et al [64] reported Shanon indices of 2.58 and 3.71 for highland and coastal quinoa, respectively, using 20 SSRs and 59 accessions. Those values agree with our study as we obtained an average of 3.51, reflecting high genetic variability for the quinoas from Ayacucho. The very low expected heterozigosity may be explained by the presence of a high degree of homozygosity in the studied populations.
Our AMOVA indicated 19.93% of the variance was due to the difference between clusters and 80.07% corresponds to molecular differences within the clusters. A FST value greater than 0.15 is considered a significant indicator of genetic differentiation between subpopulations, as reported by Allende Ciballero [65]. However, the low population divergence (FST) among Ayacucho's clusters (0.022 – 0.034) indicated very low genetic differentiation, suggesting the presence of genetic drift that can be observed in the PCoA, due to the fixation of some alleles only in three clusters and the loss of the less frequent ones. Diversity analysis works such as those of Via y Rada Fernández [39] with genetic material mainly from Puno and Cusco (172 quinoa accessions) also obtained a low FST of 0.05. Similarly, Cárdenas Cordova [40] in 129 quinoa accessions from Puno found FST values between 0.028 to 0.046; On the other hand, Allende Ciballero [65], in his work with germplasm from Cusco (quinoa from the inter-Andean valley) and Puno (quinoa from the Altiplano), obtained a low FST with a value of 0.08, which could be indicating the presence of genetic drift due to the low differentiation value similar to that obtained in the present work with the Ayacucho quinoa germplasm. The low FST may be explained by the mating system of quinoa, and also due to the influence of the cultural practices of farmers who use the same accessions repeatedly for long periods, geographical isolation, as stated by Allende Ciballero [65] for the quinoas from Cusco and Puno.
Ayacucho was highly impacted by armed violence during the 80's and 90's of the last century. This violence was also experienced in the geographic departments of Huancavelica, Apurimac, Junín, Cusco, Puno and Arequipa, among others, but the greatest impact was in Ayacucho reported by The Truth and Reconciliation Commission [47]. This social problem caused massive migration of farmers, peasants, residents and rural authorities towards the city of Huamanga and/or towards the capital of Peru, Lima. The man-made disaster, high-impact socio-political factors such as the armed violence experienced in Ayacucho for two decades, could lead to a state of bottleneck-type genetic drift in the quinoa population of the Ayacucho region, due to the migration of rural populations to large cities in the 80's and 90's of the last century leaving crops adrift, abandoning in situ conservation of germplasm on Ayacucho region endured, leading to loss of allele diversity.
Social return programs, funded by the Peruvian government, promoted the return and repopulation of the peasant communities, districts and rural sectors of Ayacucho between years 1995 to 2000, provided bonuses and materials for the construction of their new homes [66]. The population of retornants were made up mainly of the descendants of those who migrated; few complete families returned or only fragmented families returned; many elderly residents no longer returned for different reasons, as they were the holders of the ancestral knowledge of in situ conservation of crop diversity [46]. The returnees resumed agricultural activity, sowing the abandoned fields with seeds from the quinoa accessions that they found available or those that survived abandonment for two decades, that were cultivated and formed the new genetic pool of quinoa in Ayacucho, contributing to the loss of diversity of the quinoa gene pool that is observed today with characteristics of a genetic drift of bottleneck effect. This phenomenon was also observed in the quinoa germplasm of Cusco and Puno [39,40,65].
Quinoa was recognized internationally due to its exceptional nutritional qualities of quinoa, its adaptability to different agroecological zones and its potential contribution in the fight against hunger and malnutrition and declared the year 2013 as the international year of quinoa [67]. Since 2013, the Peruvian government have been promoting the cultivation of quinoa. However, there quinoa cultivar resilient to climate change are still not available for farmers. We expect our work will promote the development of molecular tools for this precious Andean crop together with modern breeding activities. Moreover, the results of this work may stimulate decision-makers for precise conservation strategies for this crop genetic resource.
5. Conclusions
Here, we inferred the genetic diversity and population structure of quinoa germplasm from Ayacucho, an Andean geographical department of Peru. High genetic diversity values were reported, except for expected heterozigosity. In addition, a total of three population that are in agreement with the geographical zones of Ayacucho (north and south) were found. It is very likely that the armed violence that existed in Ayacucho caused genetic drift in the Ayacucho quinoa germplasm. More research on quinoa is needed considering additional samples from other Andean localities of Peru.
Supplementary Materials
The following supporting information can be downloaded at: Preprints.org, Table S1: Quinoa germplasm (105) examined in this study. Table S2: List of the primers used in the SSR analysis of quinoa from Ayacucho, their corresponding temperature of annealing, number of alleles, fragment size, polymorphic alleles and gene diversity (H). Table 3. Population genetic diversity for 105 quinoa accessions from Ayacucho based on 13 SSR markers among the 11 geographical provinces. Figure S1: Phylogenetic tree based on Provesti´s genetic distance and the UPGMA algorithm of 105 native quinoas from Ayacucho using 13 SSRs. Cluster assignation is based on geographical province of origin. Number above branches represent bootstrap values, showing only those higher than 70%. Figure S2: Principal coordinate analysis (PCoA) of 105 quinoa landraces from Ayacucho. Variance explained by each coordinate are noted in parentheses. Quinoa germplasm is labelled according to their geographic zone origin. Figure S3: Plot of K from 1 to 15 values obtained from STRUCTURE analysis.
Author Contributions
Conceptualization, G.D.C. and E.D.; methodology, G.D.C., E.D. and F.M.; software, E.N., F.M. and C.I.A.; validation, F.M., C.L.S. and C.I.A.; formal analysis, G.D.C., C.L.S. and C.I.A.; data curation, C.I.A. and C.L.S.; writing—original draft preparation, G.D.C.; writing—review and editing, G.D.C., E.N., F.M., C.L.S. and C.I.A.; funding acquisition, G.D.C. and C.I.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by CONCYTEC-PERU, Contract N°346-2012-CONCYTEC-OAJ and the Socioeconomic Development Fund of Camisea FOCAM (Law N° 28451-MEF-PERU) of the Universidad Nacional San Cristóbal de Huamanga. Institutional collaborative non-accounting financing of UPCH.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors on request.
Acknowledgments
C.I.A. thanks Vicerrectorado de Investigación of UNTRM.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Heitkam, T.; Weber, B.; Walter, I.; Liedtke, S.; Ost, C.; Schmidt, T. Satellite DNA Landscapes after Allotetraploidization of Quinoa (Chenopodium Quinoa) Reveal Unique A and B Subgenomes. Plant Journal 2020, 103, 32–52. [Google Scholar] [CrossRef] [PubMed]
- Maughan, P.J.; Kolano, B.A.; Maluszynska, J.; Coles, N.D.; Bonifacio, A.; Rojas, J.; Coleman, C.E.; Stevens, M.R.; Fairbanks, D.J.; Parkinson, S.E.; et al. Molecular and Cytological Characterization of Ribosomal RNA Genes in Chenopodium Quinoa and Chenopodium Berlandieri. Genome 2006, 49, 825–839. [Google Scholar] [CrossRef] [PubMed]
- Jarvis, D.E.; Ho, Y.S.; Lightfoot, D.J.; Schmöckel, S.M.; Li, B.; Borm, T.J.A.; Ohyanagi, H.; Mineta, K.; Michell, C.T.; Saber, N.; et al. The Genome of Chenopodium Quinoa. Nature 2017, 542, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Maughan, P.J.; Turner, T.B.; Coleman, C.E.; Elzinga, D.B.; Jellen, E.N.; Morales, J.A.; Udall, J.A.; Fairbanks, D.J.; Bonifacio, A. Characterization of Salt Overly Sensitive 1 (SOS1) Gene Homoeologs in Quinoa (Chenopodium Quinoa Willd.). Genome 2009, 52, 647–657. [Google Scholar] [CrossRef]
- Brown, D.C.; Cepeda-Cornejo, V.; Maughan, P.J.; Jellen, E.N. Characterization of the Granule-Bound Starch Synthase I Gene in Chenopodium. Plant Genome 2015, 8. [Google Scholar] [CrossRef]
- Štorchová, H.; Drabešová, J.; Cháb, D.; Kolář, J.; Jellen, E.N. The Introns in FLOWERING LOCUS T-LIKE (FTL) Genes Are Useful Markers for Tracking Paternity in Tetraploid Chenopodium Quinoa Willd. Genet Resour Crop Evol 2015, 62, 913–925. [Google Scholar] [CrossRef]
- Uhle, M. Fundamentos Etnicos y Arqueologías de Arica y Tacna; Universidad Central: Quito, 1922. [Google Scholar]
- Lumbreras, L.G.; Kaulicke, P.; Santillana, J.I.; Espinoza, W. Economía Prehispanica. In Compendio de historia economica del Peru; Contreras, C., Ed.; Banco Central de Reserva del Perú: Lima, 2020. [Google Scholar]
- Bazile, D.; Fuentes, F.; Mujica, Á. Historical Perspectives and Domestication. In Quinoa: botany, production and uses; CABI: Wallingford, 2013; pp. 16–35. [Google Scholar]
- Bhargava, A.; Shukla, S.; Ohri, D. Chenopodium Quinoa - An Indian Perspective. Ind Crops Prod 2006, 23, 73–87. [Google Scholar] [CrossRef]
- Vargas Zambrano, P.; Arteaga Solorzano, R.; Cruz Viera, L. Análisis Bibliográfico Sobre El Potencial Nutricional de La Quinua (Chenopodium Quinoa) Como Alimento Funcional. Centro azúcar 2019, 46, 89–100. [Google Scholar]
- Melini, V.; Melini, F. Functional Components and Anti-Nutritional Factors in Gluten-Free Grains: A Focus on Quinoa Seeds. Foods 2021, 10, 351. [Google Scholar] [CrossRef]
- Abellán Ruiz, M.S.; Barnuevo Espinosa, M.D.; García Santamaría, C.; Contreras Fernández, C.J.; Aldeguer García, M.; Soto Méndez, F.; Guillén Guillén, I.; Luque Rubia, A.J.; Quinde Ràzuri, F.J.; Martínez Garrido, A.; et al. Efecto Del Consumo de Quinua (Chenopodium Quinoa) Como Coadyuvante En La Intervención Nutricional En Sujetos Prediabéticos. Nutr Hosp 2017, 34, 1163–1169. [Google Scholar] [CrossRef]
- Grados Torrez, R.E.; Trino, R.D.; Perez Gonzales, J.; Gonzales Dávalos, E. Determinación Del Índice Glucémico de Un Producto Elaborado a Base de Amaranto (Amaranthus Caudatus Linnaeus), Quinua (Chenopodium Quinoa Willd) y Tarwi (Lupinus Mutabilis Sweet) Para Tratamiento Coadyuvante de Diabetes Tipo 2 y Obesidad. Revista Con-Ciencia 2018, 6, 73–82. [Google Scholar]
- Abugoch, L.; Castro, E.; Tapia, C.; Añón, M.C.; Gajardo, P.; Villarroel, A. Stability of Quinoa Flour Proteins (Chenopodium Quinoa Willd.) during Storage. Int J Food Sci Technol 2009, 44, 2013–2020. [Google Scholar] [CrossRef]
- Bioversity International; FAO; PROINPA; INIAF; FICA Descriptores Para Quinua (Chenopodium Quinoa Willd.) y Sus Parientes Silvestres; Bioversity International: Rome, 2013;
- Jacobsen, S.-E. The Worldwide Potential for Quinoa ( Chenopodium Quinoa Willd.). Food Reviews International 2003, 19, 167–177. [Google Scholar] [CrossRef]
- Bhargava, A.; Srivastava, S. Quinoa: Botany, Production and Uses; CABI, 2013.
- Kumar, A.; Bhargava, A.; Shukla, S.; Singh, H.B.; Ohri, D. Screening of Exotic Chenopodium Quinoa Accessions for Downy Mildew Resistance under Mid-Eastern Conditions of India. Crop Protection 2006, 25, 879–889. [Google Scholar] [CrossRef]
- Isobe, K.; Sugiyama, H.; Okuda, D.; Murase, Y.; Harada, H.; Miyamoto, M.; Koide, S.; Higo, M.; Torigoe, Y. Effects of Sowing Time on the Seed Yield of Quinoa (Chenopodium Quinoa Willd) in South Kanto, Japan. Agricultural Sciences 2016, 07, 146–153. [Google Scholar] [CrossRef]
- Zohry, A.E.-H. Prospects of Quinoa Cultivation in Marginal Lands of Egypt. Moroccan Journal of Agricultural Sciences 2020, 1, 132–137. [Google Scholar]
- Bertero, H.D.; De La Vega, A.J.; Correa, G.; Jacobsen, S.E.; Mujica, A. Genotype and Genotype-by-Environment Interaction Effects for Grain Yield and Grain Size of Quinoa (Chenopodium Quinoa Willd.) as Revealed by Pattern Analysis of International Multi-Environment Trials. Field Crops Res 2004, 89, 299–318. [Google Scholar] [CrossRef]
- Bhargava, A.; Shukla, S.; Rajan, S.; Ohri, D. Genetic Diversity for Morphological and Quality Traits in Quinoa (Chenopodium Quinoa Willd.) Germplasm. Genet Resour Crop Evol 2007, 54, 167–173. [Google Scholar] [CrossRef]
- Nunez Carrasco, L.; Bazile, D.; Chia, E.; Hocdé, H.; Negrete Sepulveda, J.; Martinez, E.A. Representaciones Sociales Acerca de La Conservacion de La Biodiversidad En El Caso de Productores Tradicionales de Chenopodium Quinoa Willd Del Secano Costero En Las Regiones de O’Higgins y El Maule. In Proceedings of the XXX Congreso Nacional y XV Internacional de Geografia; Talca, 2009; p. 36.
- Morillo, E. Estudio de La Diversidad Genética Del Banco de Germoplasma de Quinua (Chenopodium Quinoa W.) Utilizando Marcadores Moleculares; Quito, 2002.
- Rana, T.S.; Narzary, D.; Ohri, D. Genetic Diversity and Relationships among Some Wild and Cultivated Species of Chenopodium L. (Amaranthaceae) Using RAPD and DAMD Methods. Curr Sci 2010, 98, 840–846. [Google Scholar]
- Del Castillo, C.; Winkel, T.; Mahy, G.; Bizoux, J.P. Genetic Structure of Quinoa (Chenopodium Quinoa Willd.) from the Bolivian Altiplano as Revealed by RAPD Markers. Genet Resour Crop Evol 2007, 54, 897–905. [Google Scholar] [CrossRef]
- Anabalón Rodríguez, L.; Thomet Isla, M. Comparative Analysis of Genetic and Morphologic Diversity among Quinoa Accessions (Chenopodium Quinoa Willd.) of the South of Chile and Highland Accessions. African Journal of Crop Science 2018, 6, 001–007. [Google Scholar]
- García-Godos, P.; Cueva-Castillo, J.M. Genetic Variability of 29 Peruvian Quinoa (Chenopodium Quinoa Willd) Accessions Using AFLP Markers and Multivariate Analysis. Scientia Agropecuaria 2021, 12, 57–64. [Google Scholar] [CrossRef]
- Morillo Coronado, A.-C.; Manjarres, E.H.; Morillo Coronado, Y. Molecular Characterization of Chenopodium Quinoa Willd. Using Inter-Simple Sequence Repeat (ISSR) Markers. Afr J Biotechnol 2017, 16, 483–489. [Google Scholar] [CrossRef]
- Zhang, T.; Gu, M.; Liu, Y.; Lv, Y.; Zhou, L.; Lu, H.; Liang, S.; Bao, H.; Zhao, H. Development of Novel InDel Markers and Genetic Diversity in Chenopodium Quinoa through Whole-Genome Re-Sequencing. BMC Genomics 2017, 18. [Google Scholar] [CrossRef] [PubMed]
- Laosatit, K.; Taytragool, S.; Pimsaythong, K.; Somta, P.; Tanadul, O.U.M. Genetic Diversity of Quinoa (Chenopodium Quinoa Willd.) Germplasm as Revealed by Sequence-Related Amplified Polymorphism Markers. Agriculture and Natural Resources 2021, 55, 341–348. [Google Scholar] [CrossRef]
- El-Harty, E.H.; Ghazy, A.; Alateeq, T.K.; Al-Faifi, S.A.; Khan, M.A.; Afzal, M.; Alghamdi, S.S.; Migdadi, H.M. Morphological and Molecular Characterization of Quinoa Genotypes. 2021. [CrossRef]
- Abd El-Moneim, D.; ELsarag, E.; Aloufi, S.; El-Azraq, A.; ALshamrani, S.; Safhi, F.; Ibrahim, A. Quinoa (Chenopodium Quinoa Willd.): Genetic Diversity According to ISSR and SCoT Markers, Relative Gene Expression, and Morpho-Physiological Variation under Salinity Stress. Plants 2021, 10, 2802. [Google Scholar] [CrossRef]
- Christensen, S.A.; Pratt, D.B.; Pratt, C.; Nelson, P.T.; Stevens, M.R.; Jellen, E.N.; Coleman, C.E.; Fairbanks, D.J.; Bonifacio, A.; Maughan, P.J. Assessment of Genetic Diversity in the USDA and CIP-FAO International Nursery Collections of Quinoa (Chenopodium Quinoa Willd.) Using Microsatellite Markers. Plant Genetic Resources: Characterisation and Utilisation 2007, 5, 82–95. [Google Scholar] [CrossRef]
- Costa Tártara, S.M.; Manifesto, M.M.; Bramardi, S.J.; Bertero, H.D. Genetic Structure in Cultivated Quinoa (Chenopodium Quinoa Willd.), a Reflection of Landscape Structure in Northwest Argentina. Conservation Genetics 2012, 13, 1027–1038. [Google Scholar] [CrossRef]
- Manjarres-Hernández, E.H.; Morillo-Coronado, A.C. Genetic Diversity of Colombian Quinoa (Chenopodium Quinoa Willd.): Implications for Breeding Programs. Genet Resour Crop Evol 2022, 69, 2447–2458. [Google Scholar] [CrossRef]
- Kandy, A.; Choque, H.; Huanca Alanoca, N.; Mamani, F.M.; Torrico, S.V. Diversidad Fenotípica y Genotípica de 50 Nuevas Accesiones de Quinua (Chenopodium Quinoa Willd.), Del Banco de Germoplasma de Granos Altoandinos.
- Vía y Rada Fernández, R.N. Determinación de La Diversidad Genética de 172 Accesiones de La Colección Nacional de Chenopodium Quinoa Willd. “Quinua” Mediante Marcadores Microsatélites, Universidad Ricardo Palma: Lima, 2015.
- Cárdenas Córdova, R.G. Caracterización Molecular de 129 Accesiones de Quinua(Chenopodium Quinoa Willd.) de La Región Puno Mediante Marcadores Microsatélites, Universidad Nacional Mayor de San Marcos: Lima, 2017.
- Romero, M.; Mujica, A.; Pineda, E.; Ccamapaza, Y.; Zavalla, N.; de Juli, B.; Collana, N.; Inia, I.; Blanca de Juli, A.; Blanca de Arequipa, A.; et al. Genetic Identity Based on Simple Sequence Repeat (SSR) Markers for Quinoa (Chenopodium Quinoa Willd.) Chenopodium Petiolare from the CIP-Camacani. Cien. Inv. Agr 2019, 46, 166–178. [Google Scholar] [CrossRef]
- Salazar, J.; de Lourdes Torres, M.; Gutierrez, B.; Torres, A.F. Molecular Characterization of Ecuadorian Quinoa (Chenopodium Quinoa Willd.) Diversity: Implications for Conservation and Breeding. Euphytica 2019, 215. [Google Scholar] [CrossRef]
- Arenas Morales, V.E. Análisis de La Diversidad Genética y Estructura Poblacional En Genotipos de Quínoa Chilena (Chenopodium Quinoa Willd.) Usando Microsatélites, Universidad Andrés Bello: Santiago de Chile, 2018.
- Alfaro Mendivil, E. Establecimiento Del Banco Regional de Germoplasma y Caracterización Fenotípica de Accesiones de Quinua (Chenopodium Quinoa Willd.) Ayacucho, Universidad Nacional San Cristóbal de Huamanga: Huamanga, 2013.
- Menéndez Burns, F.M. Estudio Poblacional y Diversidad Genética de Los Cultivos Primarios de Chenopodium Quinoa En Ayacucho, Perú, Universidad Peruana Cayetano Heredia: Lima, 2016.
- Pedersen, D.; Tremblay, J.; Errázuriz, C.; Gamarra, J. The Sequelae of Political Violence: Assessing Trauma, Suffering and Dislocation in the Peruvian Highlands. Soc Sci Med 2008, 67, 205–217. [Google Scholar] [CrossRef] [PubMed]
- Truth and Reconciliation Commission Final Report.
- Doyle, J.J.; Doyle, J.L. A Rapid DNA Isolation Procedure for Small Quantities of Fresh Leaf Tissue. Phytochemical Bulletin 1987, 19, 11–15. [Google Scholar]
- Mason, S.L.; Stevens, M.R.; Jellen, E.N.; Bonifacio, A.; Fairbanks, D.J.; Coleman, C.E.; McCarty, R.R.; Rasmussen, A.G.; Maughan, P.J. Development and Use of Microsatellite Markers for Germplasm Characterization in Quinoa (Chenopodium Quinoa Willd.). Crop Sci 2005, 45, 1618–1630. [Google Scholar] [CrossRef]
- Jarvis, D.E.; Kopp, O.R.; Jellen, E.N.; Mallory, M.A.; Pattee, J.; Bonifacio, A.; Coleman, C.E.; Stevens, M.R.; Fairbanks, D.J.; Maughan, P.J. Simple Sequence Repeat Marker Development and Genetic Mapping in Quinoa (Chenopodium Quinoa Willd.). J Genet 2008, 87, 39–51. [Google Scholar] [CrossRef]
- Ward, S.M. Response to Selection for Reduced Grain Saponin Content in Quinoa (Chenopodium Quinoa Willd.). Field Crops Res 2000, 68, 157–163. [Google Scholar] [CrossRef]
- Nei, M.; Maruyama, T.; Chakraborty, R. The Bottleneck Effect and Genetic Variability in Populations. Evolution (N Y) 1975, 29, 1–10. [Google Scholar]
- Chesnokov, Yu.V.; Artemyeva, A.M. Evaluation of the Measure of Polymorphism Information of Genetic Diversity. Sel’skokhozyaistvennaya Biologiya 2015, 50, 571–578. [Google Scholar] [CrossRef]
- Prevosti, A.; Ocaña, J.; Alonso, G. Distances between Populations of Drosophila Subobscura, Based on Chromosome Arrangement Frequencies. Theoretical and Applied Genetics 1975, 45, 231–241. [Google Scholar] [CrossRef]
- R Core Team R: A Language and Environment for Statistical Computing 2022.
- Kamvar, Z.N.; Tabima, J.F.; Gr̈unwald, N.J. Poppr: An R Package for Genetic Analysis of Populations with Clonal, Partially Clonal, and/or Sexual Reproduction. PeerJ 2014, 2014, 1–14. [Google Scholar] [CrossRef]
- Dray, S.; Dufour, A.-B. The Ade4 Package: Implementing the Duality Diagram for Ecologists. J Stat Softw 2007, 22. [Google Scholar] [CrossRef]
- Jombart, T. Adegenet : A R Package for the Multivariate Analysis of Genetic Markers. Bioinformatics 2008, 24, 1403–1405. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of Population Structure Using Multilocus Genotype Data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef] [PubMed]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the Number of Clusters of Individuals Using the Software STRUCTURE: A Simulation Study. Mol Ecol 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed]
- Francis, R.M. Pophelper: An R Package and Web App to Analyse and Visualize Population Structure. In Proceedings of the Molecular Ecology Resources; Blackwell Publishing Ltd, January 1 2017; Vol. 17, pp. 27–32.
- Ott, J. Strategies for Characterizing Highly Polymorphic Markers in Human Gene Mapping. The American Society of Human Genetics 1992, 51, 283–290. [Google Scholar]
- Yataco Capcha, J.; Nami, H.G. A New View on the Late Pleistocene Lithic Remains from Pikimachay Cave, South Central Peru. Archaeological Discovery 2022, 10, 282–334. [Google Scholar] [CrossRef]
- Fuentes, F.F.; Martinez, E.A.; Hinrichsen, P. V.; Jellen, E.N.; Maughan, P.J. Assessment of Genetic Diversity Patterns in Chilean Quinoa (Chenopodium Quinoa Willd.) Germplasm Using Multiplex Fluorescent Microsatellite Markers. Conservation Genetics 2009, 10, 369–377. [Google Scholar] [CrossRef]
- Allende Ciballero, M.J. Caracterización Morfológica y Molecular de Accesiones de Quinua (Chenopodium Quinoa Willd.) Para Estimar Variabilidad Genética, Universidad Nacional Agraria la Molina: Lima, 2017.
- Stepputat, F.; Nyberg Sørensen, N. The Rise and Fall of ‘Internally Displaced People’ in the Central Peruvian Andes. Dev Change 2001, 32, 769–791. [Google Scholar] [CrossRef]
- FAO Evaluación Del Año Internacional de La Quinua (2013); Roma, 2014.
Figure 1.
Principal component analysis (PCoA) of 105 quinoa landraces from Ayacucho. Variance explained by each coordinate are noted in parentheses. (A) Quinoa germplasm labelled according to their geographic province origin. (B) Quinoa germplasm labelled according to the cluster assigned by STRUCTURE and geographic zone of origin.
Figure 1.
Principal component analysis (PCoA) of 105 quinoa landraces from Ayacucho. Variance explained by each coordinate are noted in parentheses. (A) Quinoa germplasm labelled according to their geographic province origin. (B) Quinoa germplasm labelled according to the cluster assigned by STRUCTURE and geographic zone of origin.
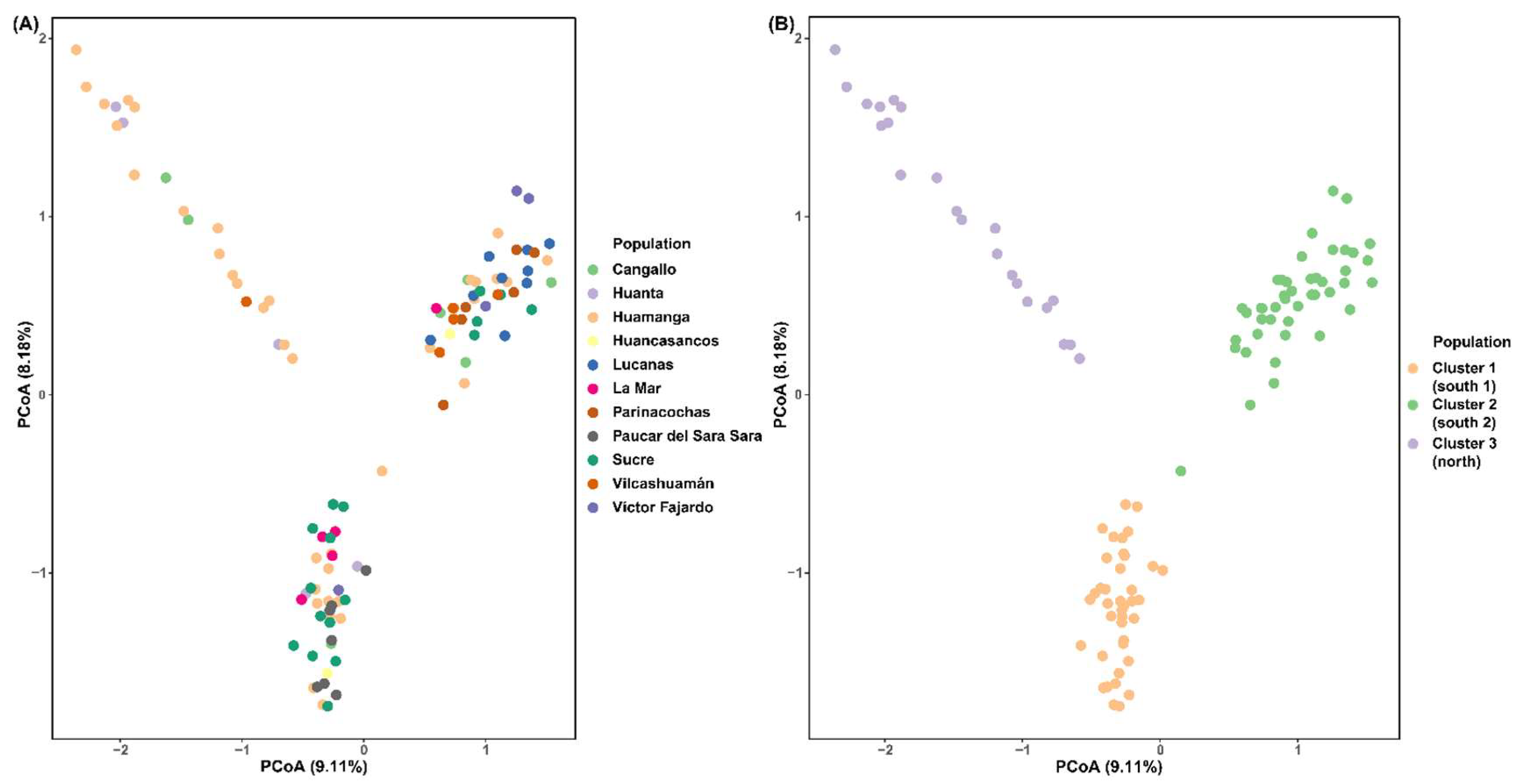
Figure 2.
Phylogenetic tree based on Provesti´s genetic distance and the UPGMA algorithm of 105 native quinoas from Ayacucho using 13 SSRs. Cluster assignation is based on STRUCTURE analysis. Number above branches represent bootstrap values, showing only those higher than 70%.
Figure 2.
Phylogenetic tree based on Provesti´s genetic distance and the UPGMA algorithm of 105 native quinoas from Ayacucho using 13 SSRs. Cluster assignation is based on STRUCTURE analysis. Number above branches represent bootstrap values, showing only those higher than 70%.
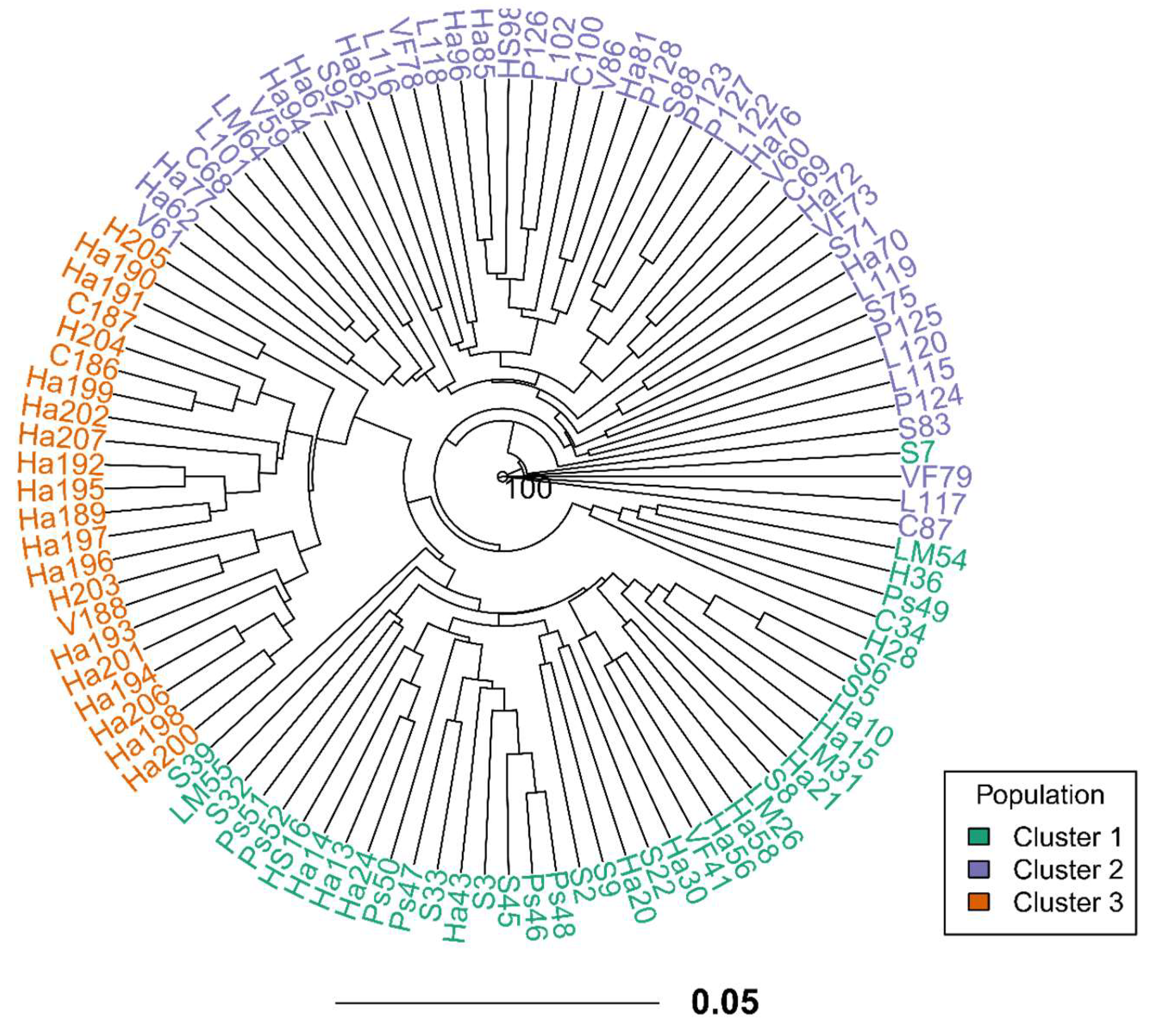
Figure 3.
Population structure of 105 accessions of quinua cultivated in Ayacucho inferred by the STRUCTURE analysis using 13 SSR markers. Each color represents a population.
Figure 3.
Population structure of 105 accessions of quinua cultivated in Ayacucho inferred by the STRUCTURE analysis using 13 SSR markers. Each color represents a population.
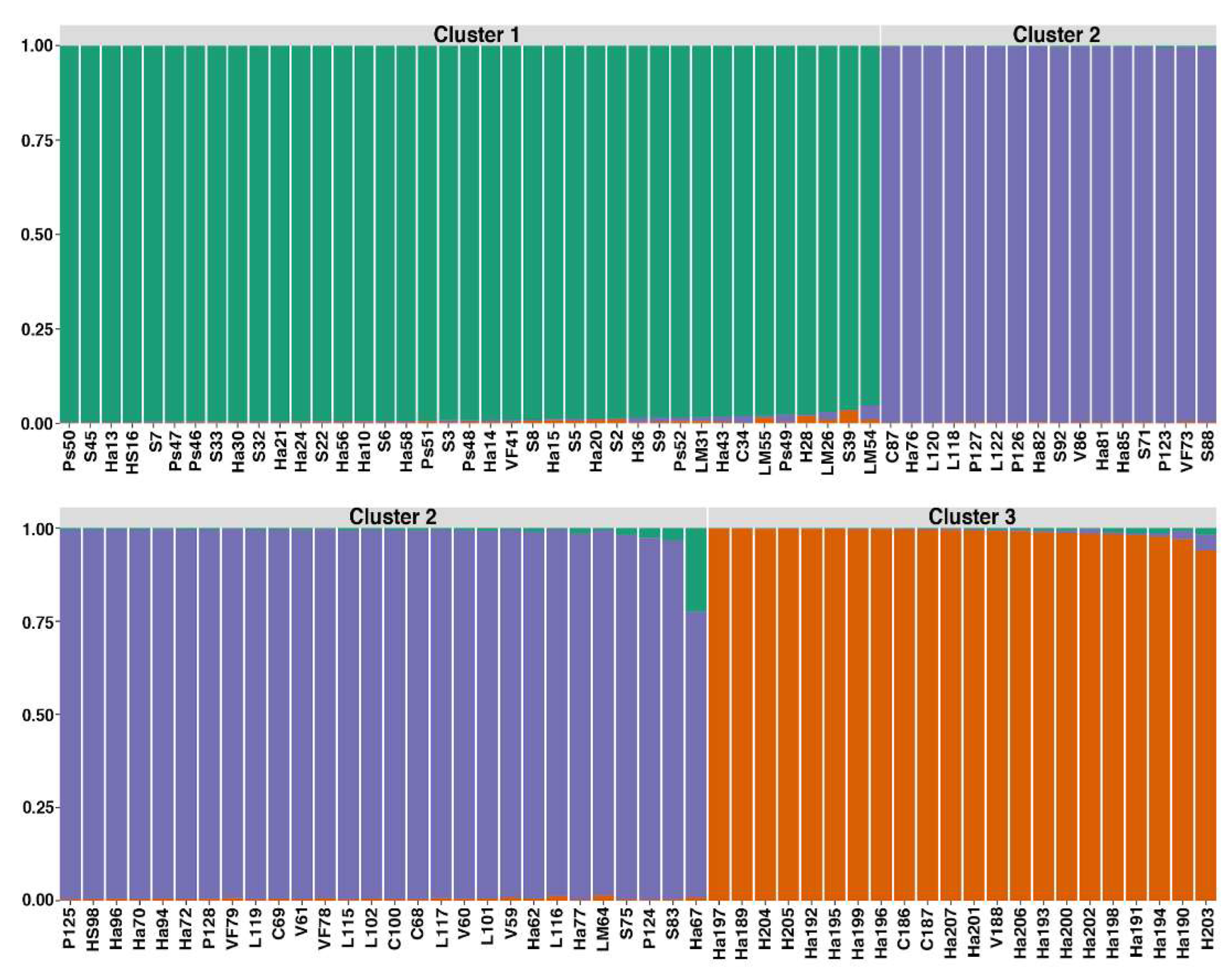
Table 3.
Population genetic diversity for 105 quinoa accessions from Ayacucho based on 13 SSR markers among three clusters inferred by the STRUCTURE analysis.
Table 3.
Population genetic diversity for 105 quinoa accessions from Ayacucho based on 13 SSR markers among three clusters inferred by the STRUCTURE analysis.
| Cluster | N | H | Lambda | HE | PIC | PPL |
|---|---|---|---|---|---|---|
| C1 (south 1) | 39 | 3.66 | 0.974 | 0.0838 | 0.095 | 73.05 |
| C2 (south 2) | 44 | 3.78 | 0.977 | 0.095 | 0.087 | 68.44 |
| C3 (north) | 22 | 3.09 | 0.955 | 0.0594 | 0.093 | 52.48 |
N: population size, H: Shannon–Wiener index, Lambda: Simpson’s index, HE: Nei’s 1978 expected heterozygosity, PPL: percentage of polymorphic loci.
Table 4.
The FST values among the four clusters inferred by STRUCTURE analysis.
| Cluster (C) | C1 | C2 | |
|---|---|---|---|
| C2 | 0.023 | ||
| C3 | 0.022 | 0.034 | |
Table 5.
Analysis of molecular variance (AMOVA) using 10 SSR markers of the genetic variation of 106 accessions of quinua among the three clusters inferred by STRUCTURE.
Table 5.
Analysis of molecular variance (AMOVA) using 10 SSR markers of the genetic variation of 106 accessions of quinua among the three clusters inferred by STRUCTURE.
| Source | Df | SS | MS | Est. Var. | % |
|---|---|---|---|---|---|
| Between clusters | 2 | 221.27 | 110.64 | 2.93 | 19.93 |
| Within clusters | 102 | 1200.77 | 11.77 | 11.77 | 80.07 |
| Total | 104 | 1422.00 | 13.67 | 14.70 | 100.00 |
df: degree of freedom, SS: sum of squares, MS: mean squares, Est. Var.: estimated variance, %: percentage of genetic variation.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



