
Submitted:
22 September 2024
Posted:
24 September 2024
You are already at the latest version
Abstract
Cancer immunobiology is one of the hot topics of discussion amongst researchers today, and immunotherapeutic modalities are among the selected few emerging approaches to cancer treatment that have exhibited a promising outlook. However, immunotherapy is not a new kid on the block; it has been around for centuries. The origin of cancer immunotherapy in modern medicine can be traced back to the initial reports of spontaneous regression of malignant tumors in some patients following an acute febrile infection, at the turn of the twentieth century. This review briefly revisits the historical accounts of immunotherapy, highlighting some of the significant developments in the field of cancer immunobiology, that have been instrumental in bringing back the immunotherapeutic approaches to the forefront of cancer research. Some of the topics covered are: Coley's toxin - the first immunotherapeutic; the genesis of the theory of immune surveillance; discovery of T-lymphocytes and dendritic cells and their roles; the role of tumor antigens; relevance of tumor microenvironment; anti-tumor (therapeutic) ability of Bacillus Calmette Guerin; Melacine - the first therapeutic vaccine engineered; theories of immunoediting and immunophenotyping of cancer; and Provenge – the first FDA approved therapeutic vaccine. In this review, head and neck cancer has been taken as the reference tumor, for narrating the progression of cancer immunobiology, particularly, for highlighting the advent of immunotherapuetic agents.
Keywords:
Coley`s toxin
; T-lymphocytes
; Dendritic cells
; Immunosurveillance
; Immunoediting
; Tumor microenvironment
; Head and Neck neoplasm
; Immunotherapy
; Therapeutic vaccines
; Checkpoint inhibitors
; Immunobiology
1. Introduction
Cancer Immunobiology is one of the hot topics of discussion amongst researchers today, and immunotherapeutic modalities are among the selected few emerging management approaches to have exhibited a promising outlook [1,2]. Some of the immunotherapeutics, such as Pembrolizumab and Nivolumab, have already received approval from the Food and Drug Administration (FDA) for their clinical use in head and neck cancer (HNC), and more than 40 candidate vaccines are currently under scrutiny for their potential clinical utility [3,4]. However, immunotherapy is not a new kid on the block; instead, it has been around for centuries, like a frivolous next-door neighbor, only to turn the table in its favor lately, and this transition is aided by the remarkable progress in the field of biotechnology in the recent times [5,6].
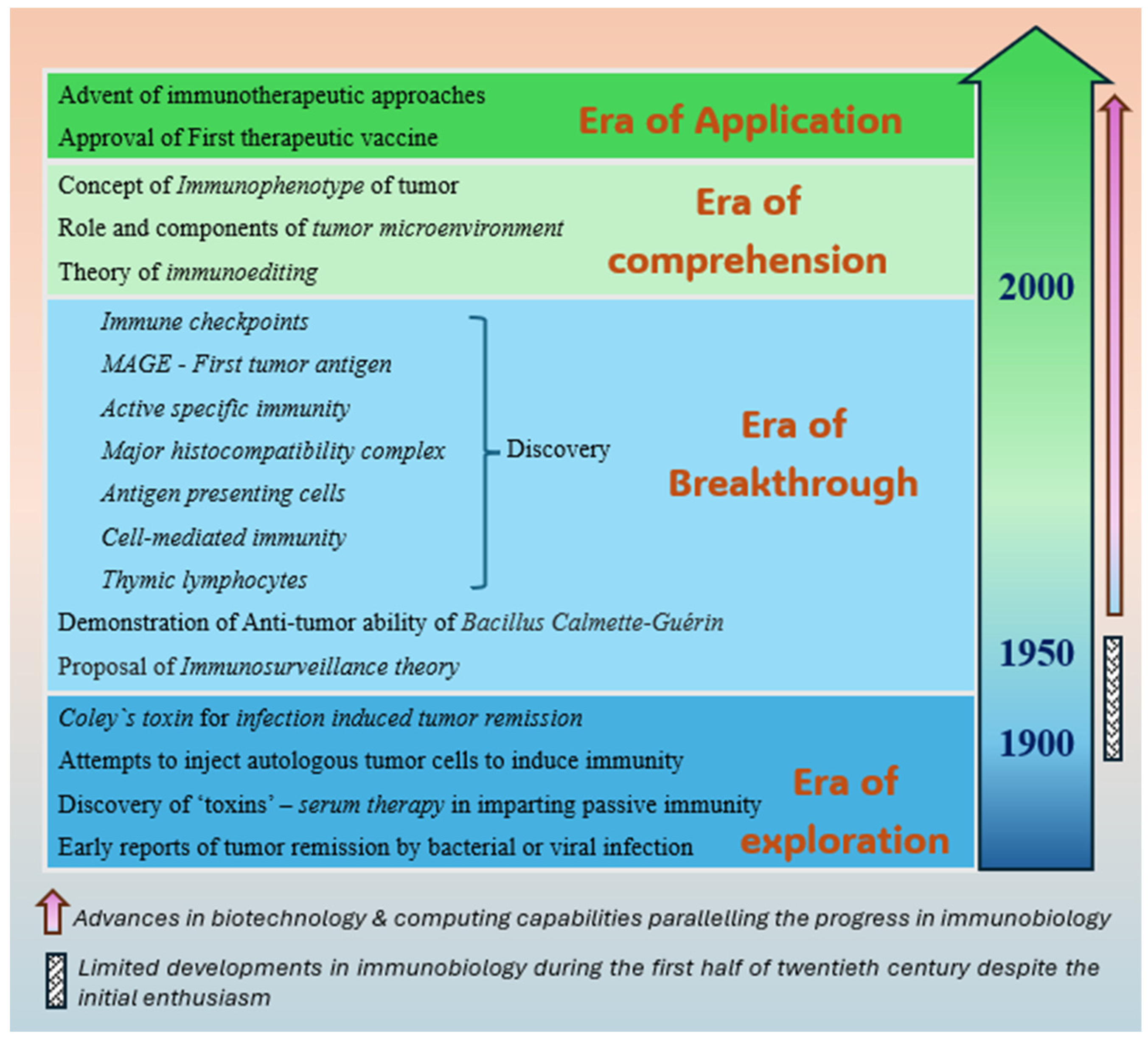
In the modern medical literature, the supposed origin of cancer immunotherapy can be traced back to the published accounts of spontaneous regression of malignant tumors following acute febrile infection [6,7]. Although sporadic reports of infection-induced-remission of cancers existed for centuries, it was the seminal work of an American surgeon, William Bradley Coley, just at the turn of the twentieth century, that eventually paved the way to this novel approach of treating the tumors non-surgically with the aid of activated immune system [5,6]. Coley's hypothesis, which states that ‘an acute infection can induce an anti-tumor immune response and eradicate a proliferative disease’ stands valid even today and explains a rare and sporadic phenomenon of the spontaneous regression of a diagnosed neoplasm [8]. However, Coley's hypothesis was disregarded entirely by majority of the scientific community for almost half a century until the advancing technology unveiled the molecular characteristics of cancer and related immunobiological interactions, layer by layer. Greatly benefitted from these pieces of emerging evidence, the renowned scientists of the 1950s, Frank Macfarlane Burnet and Lewis Thomas, described the theory of immune surveillance, which not only validated Coley's postulation about the immune activation, but also rejuvenated the field of immunotherapy [9,10]. Since then, immunotherapy has come a long way from being a speculative and inconsequential approach to becoming one of the most promising treatment modalities for cancer. The eventual realization of Coley's remarkable contribution to cancer immunobiology has earned him the title of ‘Father of cancer immunotherapy’ [11]. This review revisits some of the major developments in the field of cancer immunobiology since Coley's description and discusses the implications of those milestones, which have been instrumental in bringing back the immunotherapeutic approaches to the forefront of cancer research. Figure 1 summarizes the strides made by the immunobiology of cancer that has culminated in its expanding clinical applications in recent times.
2. Origin of the Concepts of Immunology at the Turn of Twentieth Century
2.1. First Reports of Infection-Induced Cancer Remission
Although Coley described and propagated the idea of treating a malignant disease by inducing a bacterial infection, the credit for the first experimental induction of infection to treat a tumor in the era of modern medicine goes to a German physician Wilhelm Busch [6,12]. As per a report published in 1868, Busch was fascinated by incidents of spontaneous resolution of sarcoma in some of the patients who had a severe form of bacterial skin infection called erysipelas, and himself induced a severe form of erysipelas in a 19-year-female patient with sarcoma of the neck [12]. Although the tumor did shrink considerably in Busch`s patient within two weeks of infection, the patient developed severe systemic issues prompting aggressive measures to control infection, which in turn led to the relapse of the neoplastic disease. Subsequently, a few other German clinicians too reported the remission of malignant tumors in a few of their patients after ‘induced erysipelas’, which they did by injecting the patients with cultured Streptococcus pyogenes [6,13,14]. However, these random reports largely went unnoticed, owing to the lack of scientific explanation of the underlying mechanism of anti-tumor activity.
2.2. Discovery of Antibodies
In the later quarter of nineteenth century, Robert Koch and his proteges, Emil Behring and Paul Ehrlich, both Germens, and Shibasaburo Kitasato, a microbiologist from Japan, were instrumental in discovering the antibodies and their active role in imparting protection against specific infection bacterial toxins.
In 1884, Friedrich Loeffler, another mentee of Robert Koch, reviewed the works of Edwin Klebs, and acknowledged the aetiologic agent of diphtheria as bacteria, as described by Klebs [15,16]. In fact, Klebs had already realized the role of ‘a circulating toxin in blood stream’ as the reason for manifestations of the disease, and Loeffler suggested that neutralizing this toxin could be effective in curing the disease. In 1888, Emile Roux and Alexandre Yersin from the Institute Pasteur isolated this toxin [16]. The same year, Behring also published his report on anthrax and noted that the serum of the rats that were able to kill anthrax bacteria was responsible for the immunity of these rats against anthrax [16,17]. Subsequently, in another experiment, he injected untreated guinea pigs with diphtheria toxin and treated them afterward with sera of animals that had survived the disease, thereby leading to the discovery of the principle of the antitoxic effect of serum therapy [16]. Kitasato, in a separate report, also noted similar findings with mice and rabbits immunized with cell-free fluid against tetanus [16]. In 1890, Behring and Kitasato together published an article on their success on ability to cure animals infected with both diphtheria and tetanus and on how they could protect the uninfected animals through the passive immunization with antibodies against these bacterial toxins [16,18]. Shortly after, Kitasato returned to Japan and worked towards revolutionizing immunological research in Japan [19]. Further, Behring teamed up with Ehrlich, courtesy of Koch, and produced a high-quality, standardized anti-diphtheria serum from dairy cattle, which they distributed to pediatric clinics across Berlin [20]. This serum therapy was hugely successful and reduced the mortality rate of diphtheria drastically between 1892 and 1894, as a result of which, during the Internal Medicine Congress of 1895, the serum therapy was hailed unanimously as the best method of cure ever devised [16]. Interestingly, Ehrlich had already noted that exposure to a toxin in small incremental quantities could render the animals immune to subsequent lethal doses [20,21]. He proposed the concepts of active and passive immunization and explained their mechanisms with his side-chain theory published in 1901 [21]. Though not accurate in description, Ehrilch`s lock-and-key principle provided a critical explanation of the mechanism of immunity [20]. Behring was awarded a Nobel prize in 1901, the first for medicine and physiology, for his work on serum therapy [16]. Later, Ehrlich also received the Nobel prize in 1908, together with Elie Metchnikoff, for his work in immunology [20]. Section may be divided by subheadings. It should provide a concise and precise description of the experimental results, their interpretation, as well as the experimental conclusions that can be drawn.
2.3. Coley's Toxin—The First Immunotherapeutic
At about the same time, Coley had just started his practice after graduating from the New York cancer hospital when he operated upon a 17-year-old female patient with sarcoma, who succumbed to the aggressive disease despite a radical treatment [5,11]. This patient, named Bessie Dashiell, was a close friend of John D. Rockefeller Jr, who would eventually contribute greatly to Coley`s work in the later years [5]. Devastated by the loss of a young patient early in his surgical career, Coley pledged his time and dedication to finding an effective treatment for the malignant disease [5,11]. He reviewed numerous case studies and hospital data of his senior colleagues, during which he found an intriguing case of inoperable recurrent sarcoma that had vanished following erysipelas, and on tracing the patient himself, learned about the patient's disease-free status at seven years following surgery [5,11]. Influenced by this and outcomes of similar reports from the past, in 1891, Coley injected a broth culture of Streptococcus erysipelas into one of his other patients, a 35-year male, who had recurrent spindle cell sarcoma of tonsil and neck [5,11,22,23]. He noted a complete regression of the tumor within two weeks of developing severe erysipelas [22]. Notably, Coley had undertaken this experiment within a year of starting this quest for better therapeutic options for sarcoma. Nevertheless, the first recipient of this unique treatment by Coley reportedly did well for eight years until the fatal relapse of the disease [22]. Coley attributed the therapeutic response seen in this case, and in previous reports, to the reactivation of the immune response [22,23]. In the next few years, Coley published favorable results in several other patients with sarcoma and carcinoma; however, the good therapeutic responses were apparent only in cases that got induced with frank bacterial infection by his toxin [22]. Ehrlich himself found merit in Coley`s postulation, as he also reported similar outcomes in his patients in 1908 [24].
By 1910, Coley had used this strategy successfully in numerous sarcoma cases, by injecting the mixed toxins of the Streptococcus erysipelas (now known as Streptococcus pyogenes) and the Bacillus prodigiosus (now known as Serratia marcescens), both as a primary therapeutic as well as an adjuvant after surgery to prevent the recurrence [22]. Unfortunately, despite the breakthrough oncological outcomes of mixed toxins, not many physicians and pathologists of that era accepted Coley's postulations, and a few even criticized his methods [25,26,27,28,29]. Coley himself had recognized that the infection-remission saga works best in sarcoma, but he was not able to explain this selective activity against sarcoma and the lack of predictability of therapeutic response even amongst sarcoma patients [6]. These drawbacks and the advent of radiotherapy techniques and chemotherapy diverted attention away from the therapeutic efficacy of infection-induced-immune-response until the middle of the twentieth century [11]. However, these factors did not deter Coley's tenacity, as he continued to use the so-called Coley's toxin, a combination of heat-killed Streptococcus pyogenes and Serratia marcescens, to cure hundreds of patients with bone and soft-tissue sarcoma over the years [11,30].
2.4. Other Attempts to Induce Anti-Tumor Immunity
On the other hand, in those years, many other researchers also made attempts to cure neoplastic disease by injecting autologous tumor cells [31]. Von E.v. Leyden and F. Blumenthal were the first ones to attempt this in 1902, by using the autologous tumor cell suspension as the vaccine to treat patients suffering from advanced metastatic disease [31,32]. Although initial attempts were barely successful, some of the subsequent studies reported a measurable clinical response with preparations of autologous tumor cells [31,32,33,34]. Nevertheless, these attempts technically constitute the first attempts at developing therapeutic vaccines for cancer [31,35]. However, the concept itself was not appealing to the scientific community, for until a few decades later, when renowned immunologists and pathologists found incredible evidence supporting Coley's hypothesis and mechanisms of tumor immunobiology [11]. The turn of the twentieth century had also seen few reports of tumor remission with viral infection or immunization. In 1904, George Dock of Michigan University reported a case of transient remission of acute leukemia after an infection with influenza, suggesting the role of viruses in anti-tumor treatment [36,37]. Although many studies over the years had reported the spontaneous clinical remissions of neoplastic diseases after natural viral infections such as measles or after immunization with rabies vaccines, Dock`s report was probably one of the first to have discussed the virus induced anti-tumor activity in early 1900 [37].
These works of the 1900s seem to have inspired several other scientists of that era, who, through the experiments on animal models, hypothesized the presence of immune mechanisms responsible for the rejection of transplanted tumors in mice, and provided genetic rationale for the same [7,38,39]. Shortly thereafter, James B. Murphy and John J. Morton from the Rockefeller Institute of Medical Research, a dedicated research center in New York, identified the crucial role of circulating lymphocytes in both natural and induced immunity against transplanted tumors in mice, in 1915 [40]. They demonstrated that the experimental prevention of lymphoid reaction failed to offer protection against transplanted tumors [40]. Despite these compelling works, the research on tumor-immunology seems to have hit a roadblock in the early part of the century, as not many published studies provided any credible evidence on the immunobiology of cancer, until the middle of the twentieth century.
3. Advancement of Immunobiology in the Latter Half of the Twentieth Century
3.1. Establishment of a Dedicated Institute for Research on Cancer Immunology
In 1891, Coley met Rockefeller Jr. and his father, John D. Rockefeller Sr. during the funeral of Dashiell, the patient who Coley had operated upon early in his career and lost to the progression of disease. Rockefeller and his family, owing to the bond shared with Rockefeller`s ‘adopted sister’ Dashiell, and having been impressed with the early work of Coley, supported Coley`s work in the initial years [5,11]. In 1916, Rockefeller Jr. donated land in New York for the relocation of the only dedicated cancer hospital then, which eventually paved the way for the genesis of the now-iconic Memorial Sloan-Kettering Cancer Center (MSKCC) in 1960 [5]. After the demise of Coley in 1936, his daughter Helen Coley Nauts tried reviving interest in Coley`s toxin, by reviewing the results of 800+ proven cases [5]. With a grant of 2000 USD from Nelson Rockefeller, son of Rockefeller Jr, and the support of her friend, Oliver R. Grace, she founded the Cancer Research Institute (CRI) in New York in the year 1953 [5,41]. The CRI, by continuing the research on Coley`s postulations, and by supporting works of numerous stalwarts, such as Lloyd J. Old, Rolf Kiessling, Ralph Steinman, Susumu Tonegawa, etc., has been instrumental in several discoveries that have changed the landscape of cancer immunobiology. Over the years, CRI has evolved to become a global leader in research on cancer immunobiology and immunotherapy, and has contributed to the development and launch of therapeutics, such as Gardasil, the first preventive human papilloma virus (HPV) vaccine, and more [41].
3.2. Genesis of the Theory of Immune Surveillance
In 1958, Tilden C. Everson and Warren H. Cole from Chicago, reported a series of 176 cases with spontaneous malignant tumor regression, including a few metastatic foci [42]. Although these authors could not find any scientific explanation for spontaneous regression, they identified and implicated febrile infection (suggesting activation of the immune response) as a possible reason in several cases. Meanwhile, the animal experiments continued to appraise the numerous plausible interactions between the tumor cells and the immune system [9]. The following year, Lloyd J. Old and his co-workers from MSKCC, New York, successfully demonstrated the anti-tumor ability of Bacillus Calmette-Guérin (BCG), a vaccine used primarily against tuberculosis, in a mouse model [43]. It was around the same time that Burnet and Thomas proposed the much-deliberated concept of that era- the theory of immune surveillance, which was inspired by the results of their work on immunological tolerance and drew support from studies by others [9,10]. Immune surveillance is the mechanism by which the host immune system can identify the new antigens associated with precursors of cancer and, in most cases, destroy them even before they become clinically apparent. They equated this mechanism to provoking immunological tolerance in homograft failure [9,10]. Burnet received the Nobel prize in 1960 for the discovery of acquired immunological tolerance, which he shared, although controversially, with Peter Medawar, who has contributed immensely to the science of transplantation biology [44,45]. Nevertheless, by virtue of these critical breakthroughs in the field of immunology witnessed in the middle part of the twentieth century, the scientific community could finally comprehend the complexity of immune mechanisms that underlie the anti-tumor activity of infection or febrile illness induced immune activation [25,26,27,28,29,46].
3.3. Discovery of T-Lymphocytes—The Dawn of the Era of Immunotherapy
Between 1959 and 1962, Jacques Miller published the results of a series of experiments that he had conducted as part of his Ph.D. research topic - pathogenesis of mouse leukemia induced by the Gross virus [25,47,48,49,50]. By studying the effects of thymectomy in neonatal mice on various situations, such as virus inoculation, thymus tissue grafting, skin grafting, etc., Miller discovered the novel thymic lymphocytes (now called T-lymphocytes or T-cells) and found evidence of their active involvement in the cell-mediated immunity against viruses as well as in countering the immune tolerance proposed by Burnet and Medawar [25,49,50] Miller, who had been inspired by the renowned immunologists like Burnet and Medawar, and Jim Gowans, another prominent immunologist of those days, was lauded by the same stalwarts as his commendable discovery helped them establish the evidence supporting their respective hypotheses on the immune mechanisms. Arguably, this is the most crucial milestone in the evolution of immunobiology that is said to have changed the face and pace of immunological research and its clinical implications forever [27,46,49]. As far as cancer immunobiology is concerned, Miller and his colleagues identified the critical role of T-lymphocytes in carinogenesis and its progression, thus supporting the hypothesis of immunological surveillance of Burnet and Thomas [47,48]. The discovery of T-lymphocytes and the realization of the role of cytolytic T-lymphocytes (CTL) in the systemic immune response not only set the wheels of immunology (and immunotherapy) in motion but also opened the floodgates for immunologists across the world to expand the horizons of immunology, including that of cancer immunobiology.
3.4. Discovery of Other Key Apparatus of Immune Mechanisms
In 1970, J C Cerottini, A A Nordin, and K T Brunner demonstrated that the action of CTL can be made specific to a particular antigen-bearing cell by pre-sensitizing them with that target antigen [51,52]. In 1973, Ralph M. Steinman and Zanvil A. Cohn discovered the dendritic cell (DC) in peripheral lymphoid organs of mice, which turned out to be one of the most potent antigen-presenting cells (APC) [53,54,55]. Further, Rolf Zinkernagel and Peter Doherty published their reports in 1974, in which they proposed the major histocompatibility complex (MHC)-I-restricted CTL recognition mechanism to explain the antigen-specificity of cell-mediated immunity [56,57]. This crucial discovery by Zinkernagel and Doherty won them a Nobel Prize in Physiology or Medicine in 1996 [58,59]. Similarly, Steinman's description of the morphology and tissue distribution of DC, and its crucial role in adaptive immunity, also got him the Nobel Prize in Physiology or Medicine in 2011 [53,60]. Interestingly, Steinman shared the Nobel prize with Bruce A. Beutler and Jules A. Hoffmann, awarded for their work on Toll-like receptors (TLR), which happens to be one of the critical components of the innate immune system responsible for recognizing pathogens [60]. Nevertheless, since these discoveries of the 1970s, the ability to induce an antigen-specific action of CTL has come a long way to become one of the commonly used strategies of therapeutic vaccines, which generally target a specific antigen expressed on tumor cells to execute anti-tumor immunity [4].
3.5. BCG—The First Therapeutic Vaccine with Definitive Indication
BCG, an attenuated strain of Mycobacterium bovis, was isolated in 1908 by Albert Calmette and Camille Guéri from a glycerol bile potato medium at the Pasteur Institute in Lille and was first used in humans in 1921, as vaccination against tuberculosis [61]. Shortly after, Raymond Pearl of John Hopkins University hypothesized antagonism between cancer and tuberculosis, when he observed active tuberculosis lesions in 6.6% of those with malignant lesions as compared to 16.3% in those without malignancy, during an autopsy study with a sample size of 816 [62]. Nevertheless, the attentention towards the utility of BCG as an anti-tumor agent only intensified after the first successful attempts of inducing the anti-tumor immunity with the BCG in a mouse model by LJ. Old and his colleagues in 1959 [43]. By the 1970s, several studies, both animal and clinical, documented the activation of the immune system with the BCG vaccine, which paralleled its anti-tumor abilities in the form of measurable regression of several types of solid tumors such as superficial cancer of the bladder, hepatocellular carcinoma, adenocarcinoma of kidney, malignant melanoma, and HNC [63,64,65,66,67]. Particularly, the clinical works of urologists from Canada, Alvaro Morales and David Eidinger, in patients with kidney and bladder cancers, provided the necessary momentum for the advancement of immunotherapeutic research with BCG [63,64]. Meanwhile, LJ Old and colleagues from MSKCC, New York, identified an endotoxin-like substance called tumor necrosis factor (TNF) in 1975, which was responsible for necrosis of transplanted tumors in mice [68]. In fact, LJ Old was another pioneer in cancer immunology, and his contributions via his mice studies with BCG, consequently, the discovery of TNF (later realized to be a key cytokine involved in cytolysis of cancer cells), have earned him the shared title of father of immunotherapy, with Coley [7,69]. Encouraged by these results and then the newly-discovered concept of sensitizing CTLs, some investigators of that era mixed the irradiated autologous tumor cells with the BCG, as an attempt to enhance and direct the resultant immune response towards the tumor cells, and found a desired immunological and clinical activity [70,71,72,73]. Although BCG monotherapy, by this time, was known for its potent immune stimulation mechanisms locally (via direct suppression of tumor growth as well as by effective recruitment of immune cells and increased expression of interferon-gamma (IFN-γ), its role as an adjuvant to other active agents caught the attention of the researchers, probably for the increased anti-tumor ability of the combination [74,75,76]. Nevertheless, BCG monotherapy, supposedly the first therapeutic vaccine to find a definitive clinical application and with predictable outcomes, continues to be a reliable topical agent for treating superficial bladder cancer even after 50 years of its introduction [74,75,76].
3.6. Central Mechanism of Therapeutic Vaccines
These reports on the anti-tumor ability of BCG marked the beginning of a systematic transition of immunotherapeutic approaches from being considered as an inducer of non-specific immune response that could kill tumor cells towards becoming a specific immune mechanism that exhibits cell-mediated cytotoxicity against tumor cells by producing antigen-specific CTL. The researchers of those days used the term Active specific immunotherapy (ASI) to define the specific anti-tumor immunological response observed in their studies after the injection of an active agent (inactivated tumor cells) and adjuvant (BCG) [70,71,72,73]. ASI, also known as antigen-specific immune response (ASIR), tumor-specific immune response, or specific immune-mediated tumor rejection, can be defined as a process in which an exogenous agent is administered to induce an active cell-mediated immunity against the specific antigen expressed by a particular tumor cell [70,71,72]. Most of these earlier studies used BCG-cell wall or Newcastle disease virus (NDV) based vaccines to deliver either the live or irradiated autologous tumor cells to induce the ASI [70,71,72]. In one of the studies, although the vaccine containing living tumor cells cured most of the guinea pigs, it induced tumors at the vaccine sites in a few animals. Contrastingly, the vaccines with irradiated tumor cells were not tumorigenic but required more tumor cells for successful therapy [70].
Nevertheless, by this time, it had become apparent that the APC recognized the specific antigens expressed on the tumor cells and presented them to the immune system, eventually activating the appropriate T-cell responses. It had also been realized by then that the production and release of the antigen- (tumor-) specific CTL is essential for anti-tumor immunity, as this antigen-specific cell-mediated cytotoxicity is the central process involved in the elimination of neoplastic cells [77,78]. It is interesting to note that the authors of this era had appreciated the role of 'then unknown' innumerable antigens that could get activated by a bacterial infection or a traumatic event and, in turn, stimulate the host immune response, at times even resulting in spontaneous regression of an invasive tumor [79]. Nevertheless, despite these glaring evidence and sound hypotheses, there were a few missing links, particularly regarding how invasive tumors evolve despite immunosurveillance. These critical questions, along with some of the contradicting reports and the technical hurdles of those days, culminated in doubts about immunosurveillance's veracity [80,81,82]. On the other hand, those promising responses of the earlier works with BCG encouraged the scientists to look for safer and more effective immunotherapeutic agents. Several key developments at the turn of the twenty-first century provided the needed impetus in this regard.
4. Explosion of Immunological Concepts—At the Turn of the Twenty-First Century
4.1. Discovery of Immune Tolerance Mechanisms—Checkpoints
By 1990, it was discovered that the activation of CTL by tumor antigen-led ASI is also associated with the co-induction of a specific class of immunoglobulins that could counter the activity of CTL itself [83,84]. These controls are primarily meant to serve as checkpoints to prevent collateral damage due to immune responses against the self-cells, and their activation leads to suppression or induced dysfunction of CTLs, typically in the form of T-cell exhaustion, eventually hindering their anti-tumor activity [7]. These findings substantiated the hypothesis of immune tolerance and provided early evidence of mechanisms on how the tumor cells can effectively negate the host immune response. Particularly, of the several identified immune checkpoints (IC) that attenuate the activity of T cells, the two prominent ones, named cytotoxic T-lymphocyte protein 4 (CTLA4) and programmed cell death protein 1 (PD-1), have attracted a lot of attention, and are being instrumental in the conceptualization of immune-tolerance or immune-escape mechanisms [85,86,87]. The impact of the discoveries of PD-1 and CTLA-4 on cancer immunobiology and immunotherapy was so disruptive that, Professor Tasuku Honjo and Professor James Allison, who were respectively responsible for these discoveries, have been bestowed with multiple laurels, including a Nobel prize in 2018 under the category of Physiology or Medicine [88,89]. Also, Pembrolizumab and Nivolumab, the only two immunotherapeutic medications to have got FDA approval for use in HNC to date, act by inhibiting PD-1 [3].
4.2. Melacine—The First Therapeutic Vaccine to be Engineered
In a phase-I trial, Mitchell et al. from the University of Southern California School of Medicine, Los Angeles, injected the mechanical lysates (homogenates) of two allogeneic melanoma cell lines with a novel adjuvant called DETOX (detoxified Freund’s adjuvant, Ribi ImmunoChem Research, Inc, Hamilton, MT) in 22 patients of melanoma, and found complete remission of the disease in two patients, partial response in three patients and minor responses in another three patients [90]. In their results, published in 1988, they noted elevated levels of CTL precursors in all eight patients who had a clinical response, and none of the seven patients with no increase in their CTL precursors had any clinical response. Subsequently, the authors repeated the same experiment in another set of 25 patients with metastatic melanoma and found that the ASIR induced by these allogeneic lysates of melanoma with adjuvant DETOX could induce regression of the disease in a proportion of patients with little toxicity [91]. Here also, an increase in precursors CTLs correlated well with the clinical outcome. These results led to the engineering of the first therapeutic vaccine in 1998, called Melacine (Ribi ImmunoChem Research, Hamilton, MT), developed as a lyophilized preparation from the same two allogeneic melanoma cell lysates used by Mitchell et al. [92]. The trials from other centers too reported tumor regression with Melacine in around 10% of the patients, with long-term stabilization of disease in 10% to 20% [92]. Notably, in these studies, the benefit of Melacine was best appreciated in an adjuvant-treatment setting [93]. Interestingly, at around the same time, researchers from John Wayne Center Institute, Santa Monica, CA, also reported promising results in advanced cases of melanoma, with another therapeutic vaccine called Canvaxin, an autologous whole tumor cell-based polyvalent vaccine admixed with BCG [73]. However, subsequent phase III randomized controlled trials on Melacine and Canvaxin failed to demonstrate any advantage of these vaccines in the adjuvant-treatment settings of melanoma patients [94,95]. Also, the adjuvants used in these vaccines, DEXOT and BCG, though immunogenic, had a few safety concerns [96,97]. Nevertheless, since Melacine and Canvaxin, our understanding of tumor immunobiology has progressed in leaps and bounds, and consequently, its applications, including the development of therapeutic vaccines, have also come a long way. Interestingly, among the several transforming changes or inventions seen over the years, the three major developments that can be credited for the metamorphosis of therapeutic vaccines are all from around the same timeline as the designing and evaluation of Melacine.
4.3. Enhanced Ability to Measure the Elicited ASIR
In 1983, Cecil C. Czerkinsky and colleagues from Sweden introduced a new high-throughput technology called Enzyme-linked immunosorbent spot (ELISpot) assay, a versatile and sensitive alternative to the conventional plaque-forming cell assays that were used to enumerate cells involved in immune response till then [98]. The high accuracy, sensitivity, reproducibility, and robustness of ELISpot led to its subsequent widespread use for measuring cell-mediated ASIR in both mice and humans quantitatively. Owing to its ability to measure a broad range of magnitudes of response, including the critical ASIR activities (for measuring cytotoxicity) such as IFN-γ secretion and granzyme B release, the CTL monitoring by ELISpot is one of the current gold standard methods for the evaluation of immune responses in clinical trials and vaccine candidature [99,100].
4.4. Isolation of DC
DC, dubbed nature's adjuvant, is known for its exceptional capacity to initiate both innate as well as adaptive immune responses [101]. The isolation of DC from the bone marrow precursors in the 1990s eventually led to the development of DC-based therapeutic vaccines that quickly became the preferred vehicles and adjuvant for the development of vaccines [102,103]. As the DCs are critically involved in (tumor-) antigen presentation to the host immune system, conjugating DC with a tumor antigen potentiates the induced ASIR against the tumor cells. The realization of the role played by the adjuvant in increasing the therapeutic response of a vaccine has led to the discovery of several other potent adjuvants in recent times, such as Montanide ISA, based on detoxified or Freund’s incomplete adjuvant (IFA), TLR-based adjuvants, granulocyte-macrophage colony-stimulating factor (GM-CSF), polylactic acid, polylactide-coglycolide, and imiquimod [4,96,97].
4.5. MAGE—The First Tumor Antigen to be Recognized
Tumor antigens are nothing but antigens expressed by tumor cells that could be exploited as specific targets for inducing ASIR. Around the same time as the development of Melacine by Mitchel et al., several other investigators identified a few novel tumor antigens and documented their efficiency in inducing the ASIR as well as their ability to hinder the growth of tumors in experimental models [77,104,105]. A tumor antigen identified by Pierre van der Bruggen et al. in 1991, called melanoma antigen encoding gene (MAGE), encoding a tumor antigen of melanoma, was the first real molecular target identified for the induction of ASIR [105]. Since then, batteries of new and potent tumor antigens have been recognized in various tumors, paving the way for developing therapeutic vaccines of different classes [106,107]. As for HNC, the envelope proteins E6 and E7 of HPV 16 and 18, are the most attractive and widely targeted tumor antigen for developing therapeutic vaccines [4]. The involvement of these oncogenic viruses in the carcinogenesis of HPV-driven HNC offers a peculiar advantage by providing an opportunity to selectively target those HPV antigens harboring tumor cells and sparing the normal cells [4,108]. The tumor antigens can be broadly classified as shared antigens- expressed in tumor cells as well as in normal tissues and tumor-specific antigens- expressed only in tumor cells. Alternatively, they could be classified based on their peculiar expression characteristics as cancer-testis antigens, oncoviral antigens, overexpressed /differentiation antigens, neoantigens, and stealth antigens [109,110,111,112]. In 2009, the Translational Research Working Group of the National Cancer Institute (NCI) published a report on its pilot project, whose objective was to prioritize and rank the tumor antigens based on their immunogenicity (17%), oncogenicity (15%), specificity (15%), and five other criteria [113]. Using the analytic hierarchy process, the expert committee of NCI found that none of the 75 evaluated antigens could satisfy the requirements to be called an "Ideal" tumor antigen [113].
4.6. Theory of Immunoediting
In the early 2000s, overwhelmed by the growing body of evidence supporting the temporal and spatial dynamicity of the interactions between the immunogenic and the tolerogenic mechanisms, Dunn et al. proposed the peculiar hypothesis of cancer immunoediting, explaining the dual role played by the immune system, via host-protective mechanisms as well as tumor-sculpting functions, throughout the process of carcinogenesis [114,115]. According to this theory, cancer immunoediting represents a dynamic process encompassing three phases - elimination, equilibrium, and escape. When an oncogenic stimulus sets in the process of carcinogenesis, the precursor cells express distinct tumor-specific markers and generate proinflammatory immune responses that take part in the process of immunoediting. In the first phase of the process, elimination, which represents the classical concept of cancer immunosurveillance, the cells and the molecules of innate and adaptive immunity work to eradicate a developing tumor. However, if this process is not successful in destroying the tumor cell, the second phase of immunoediting - equilibrium phase sets in, wherein the viable tumor cells are either maintained chronically or immunologically sculpted by immune "editors" to produce new variants of tumor cells selectively. When these newly produced variants may eventually evade the immunological restraints of the equilibrium phase, they manifest as clinically detectable tumors in the third and final phase of immunoediting, the escape phase [115]. At the outset, the proposed model of immunoediting and these underlying mechanisms answer most of the earlier questions related to immune tolerance and have also found evidential support in the latest studies [115]. Moreover, the acknowledgment of these immunoediting processes, particularly the comprehension of the dynamic changes at the level of the tumor microenvironment (TME), has been playing a critical role in bringing immunotherapeutics closer to clinical application than ever before [115].
4.7. Role of TME in the Process of Immunoediting
The balance between immune surveillance and immune escape is a result of a complex interplay between numerous cellular proteins and pathways, determined by several genetic, environmental, and tumor-related factors that vary drastically between individuals. The cancer-immune set point is the equilibrium between factors that promote and suppress anticancer immunity, and a delicate balance of this set point is essential for recognizing and eliminating tumor cells at one end and preventing autoimmunity on the other [87]. At the molecular level, induction of CTL-mediated ASI is one of the primary mechanisms of anti-tumor immunity, which is carried out by releasing cytokines such as IFN-γ/TNF-α, perforin, and granzyme that eradicate cancer cells [116,117]. On the other hand, the principal mechanism of hindering anti-tumor immunity includes countering the activation, differentiation, or release of T-cells or attenuating their function at TME, in which ICs play a crucial role. Some of the other mechanisms known to exhibit anti-immunity action include (a)activation of immunosuppressive cells such as tumor-associated macrophage (TAM), T/B-regulatory cell (T/B-reg), myeloid-derived suppressor cell (MDSC), (b)secretion of immunosuppressive cytokines, like transforming growth factor-β, (c)activation of immunosuppressive signaling pathways, like signal transducer and stimulator of transcription-3, and (d)formation of physical barriers and intricate vascular networks within the tumor, that hinder the drug delivery and render the cells hypoxic [4,118]. Though several of these molecular mechanisms were recognized later, it had already become apparent a few decades ago that immune surveillance represents only one dimension of the intricate interactions between the host immune system and factors responsible for carcinogenesis [114,119]. Cancer immunotherapeutics, including therapeutic vaccines, act either by producing a robust ASIR against the tumor cells or by negating the immunosuppressive conditions/factors to enable the reactivation of naturally existing anti-tumor immunity [4].
4.8. Immunophenotyping of Cancer
As per the current understanding of cancer immunology, malignant tumors can be broadly classified into three types of immunophenotypes, depending on the relative abundance and distribution of immune cells in TME [118]. Tumors with high T-cell infiltration, high PD-L1 expression, and exhibiting high tumor mutational load (TMB) are categorized as immunoinflammatory tumors. In immune-excluded or -immune-rejected tumors, the cytotoxic T cells are often blocked at the surface without being able to enter cells to exert cytotoxic effects. Lastly, immune-desert tumors are characterized by few or no CD8+ T cells, low PD-L1 expression, and low TMB [118]. Each of these cancer-immunophenotypes seems to have specific underlying molecular mechanisms of immune escape that prevent the host's immune response from eradicating cancer [87]. While hyper-exhaustion of T-cells by the ICs is attributed majorly to the immune escape in inflamed tumors, the presence of immunosuppressive cells such as TAM, T/B-reg, MDSC, and a hypoxic microenvironment are held responsible for the immunosuppressive TME associated with the non-inflamed tumors [87,118]. Interestingly, the profile of recruited immune cells that contribute greatly to the immunophenotype of a particular tumor also dictates its hallmark traits like biological behavior and therapeutic responsiveness [120,121,122]. For instance, the immunoinflammatory types of tumors respond efficiently to immunotherapeutics strategies like inhibition of ICs (with anti-PD-L1/PD-1 therapy), owing to the pre-existing anti-tumor immune response that gets rejuvenated by these agents. Contrastingly, the non-inflamed tumors respond poorly to this approach as they have few or no CD8-carrying T cells and exhibit low PD-L1 expression [87]. In other words, for therapeutic actionability, the tumors with immune-desert and immune-excluded phenotypes are considered 'cold tumors,’ as against the ‘hot’ and actionable inflamed tumors [123,124,125].
4.9. Provenge—The First FDA-Approved Therapeutic Vaccine
Sipuleucel-T (Provenge, Dendreon Pharmaceuticals LLC, California, US), an autologous immunotherapeutic agent designed to stimulate an ASIR against prostate cancer, showed a definitive clinical response and survival benefits among men with metastatic castration-resistant prostate cancer (m CRPC) in early clinical studies [126,127]. Sipuleucel-T contains autologous peripheral-blood mononuclear cells, including APCs, activated ex vivo with a recombinant fusion protein - PA2024. PA2024 consists of prostatic acid phosphatase (PAP), a tissue antigen over-expressed by prostate cancer cells, fused to GM-CSF as an adjuvant [128]. When injected intravenously, Sipuleucel-T stimulates an immune response to prostate cancer cells by targeting PAP and generating specific T cells capable of recognizing and killing prostate cancer cells that express PAP [129]. Favorable evidence from early clinical studies led to a double-blind, placebo-controlled, multicenter phase III trial (NCT00065442), called Immunotherapy Prostate Adenocarcinoma Treatment (IMPACT), which was carried out between July 2003 to January 2009, with 512 asymptomatic or minimally symptomatic patients of mCRPC. In this trial, three two-weekly intravenous infusions of sipuleucel-T led to a relative reduction of risk of death by 22%, representing a 4.1-month advantage in medial survival over the placebo [128]. At three years, sipuleucel-T increased the overall survival by 38% (31.7% in the sipuleucel-T group versus 23.0% in the placebo group) [128]. These results prompted the FDA to approve sipuleucel-T in 2010 for treating patients with asymptomatic or minimally symptomatic mCRPC [129]. Sipuleucel-T is the first and the only FDA-approved therapeutic vaccine for clinical use to date. However, several approaches and agents are currently being evaluated for their safety and efficacy as therapeutic vaccines in different cancer lines, including HNC. A few of these agents, particularly those targeting tumor antigens of HPV-16/18, have already received the FastTrack designation by the FDA for use in recurrent/metastatic/progressive oropharyngeal cancers [4]. Further details on the various approaches and candidates of therapeutic vaccines for HNC can be found in our recent comprehensive review [4]. Some of these agents can potentially change the landscape of cancer chemotherapy for the better. Moreover, with technological advancements and evolving research capabilities, the concept of precision medicine in cancer immunotherapy is not far from being practical.
5. Conclusions
Though immunobiology based avenues are re-emerging as potential game changers in cancer therapeutics, the principles behind these approaches are not new. From earlier reports of spontaneous regression of malignant tumors following acute febrile infection to the approval of the first immunotherapeutic by the FDA for clinical use, the approaches and modalities of immunotherapy have come a long way. Particularly, milestones like the discovery of T-lymphocytes and dendritic cells, and anti-tumor (therapeutic) ability of BCG and other vaccines, and the development of theories such as immunosurveillance and immunoediting, have all contributed to this extraordinary journey.
6. Patents
Not applicable.
Author Contributions
Conceptualization, K.D. and S.A.; methodology, K.D.; validation, S.A., K.S. and M.S.; resources, K.D. M.S. and K.S.; data curation, K.D.; writing—original draft preparation, K.D.; writing—review and editing, S.A. and K.S.; visualization, K.D. and M.S.; project administration, S.A. and M.S.; All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Nil.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Waldman, A.D.; Fritz, J.M.; Lenardo, M.J. A Guide to Cancer Immunotherapy: From T Cell Basic Science to Clinical Practice. Nat Rev Immunol 2020, 20, 651–668. [CrossRef]
- Blass, E.; Ott, P.A. Advances in the Development of Personalized Neoantigen-Based Therapeutic Cancer Vaccines. Nat Rev Clin Oncol 2021, 18, 215–229. [CrossRef]
- Devaraja, K. Current Prospects of Molecular Therapeutics in Head and Neck Squamous Cell Carcinoma. Pharmaceut Med 2019, 33, 269–289. [CrossRef]
- Devaraja, K.; Aggarwal, S.; Singh, M. Therapeutic Vaccination in Head and Neck Squamous Cell Carcinoma-A Review. Vaccines (Basel) 2023, 11, 634. [CrossRef]
- Levine, D.B. The Hospital for the Ruptured and Crippled: William Bradley Coley, Third Surgeon-in-Chief 1925-1933. HSS J 2008, 4, 1–9. [CrossRef]
- Hobohm, U. Fever and Cancer in Perspective. Cancer Immunol Immunother 2001, 50, 391–396. [CrossRef]
- Oiseth, S.J.; Aziz, M.S. Cancer Immunotherapy: A Brief Review of the History, Possibilities, and Challenges Ahead. Journal of Cancer Metastasis and Treatment 2017, 3, 250–261. [CrossRef]
- Suzuki, A.; Abe, S.; Koyama, K.; Suzuki, S.; Nagao, M.; Kobayashi, M.; Nomura, J.; Tsutsumi, T.; Takeda, T.; Oka, Y.; et al. Spontaneous Regression of Blastic Plasmacytoid Dendritic Cell Neoplasm Following Sepsis by Serratia Marcescens: A Case Report and Literature Review. Intern Med 2021, 60, 927–933. [CrossRef]
- Burnet, M. Cancer; a Biological Approach. I. The Processes of Control. Br Med J 1957, 1, 779–786. [CrossRef]
- Thomas, L.; Lawrence, H. Cellular and Humoral Aspects of the Hypersensitive States. New York: Hoeber-Harper 1959, 529–532.
- McCarthy, E.F. The Toxins of William B. Coley and the Treatment of Bone and Soft-Tissue Sarcomas. Iowa Orthop J 2006, 26, 154–158.
- Busch, W. Aus Der Sitzung Der Medicinischen Section Vom 13 November 1867. Berl Klin Wochenschr 1868, 5, 137.
- Fehleisen, N. Ueber die Züchtung der Erysipelkokken auf künstlichem Nährboden und ihre Uebertragbarkeit auf den Menschen. Dtsch Med Wochenschr 1882, 8, 553–554. [CrossRef]
- Bruns, P. von Die Heilwirkung Des Erysipels Auf Geschwulste. Beitr Klin Chir 1887, 3, 443–466.
- Loeffler, F. Untersuchungen Über Die Bedeutung Der Mikroorganismen Für Die Entstehung Der Diphtherie Beim Menschen, Bei Der Taube Und Beim Kalbe; 1884;
- Winau, F.; Winau, R. Emil von Behring and Serum Therapy. Microbes Infect 2002, 4, 185–188. [CrossRef]
- Behring, E. von Über Die Ursache Der Immunität von Ratten Gegen Milzbrand. Drucke 19. Jh. 1888.
- Von Behring, E.; Kitasato, S. Uber Das Zustandekommen Der Diphtherie-Immunitat Und Der Tetanus-Immunitat Bei Thieren. Deut. Med. Wochenschr. 1890, 16, 1113–1114.
- Triendl, R. Translational Research in Immunology: Japanese Perspectives. Nat Rev Immunol 2004, 4, 72–76. [CrossRef]
- Winau, F.; Westphal, O.; Winau, R. Paul Ehrlich--in Search of the Magic Bullet. Microbes Infect 2004, 6, 786–789. [CrossRef]
- Ehrlich, P. Die Seitenkettentheorie Und Ihre Gegner. Münchner Med Wochenschr 1901, 52, 2123–2124.
- Coley, W.B. The Treatment of Inoperable Sarcoma by Bacterial Toxins (the Mixed Toxins of the Streptococcus Erysipelas and the Bacillus Prodigiosus). Proc R Soc Med 1910, 3, 1–48.
- Coley, W.B. II. Contribution to the Knowledge of Sarcoma. Ann Surg 1891, 14, 199–220. [CrossRef]
- Ehrlich, P. Ueber Den Jetzigen Stand Der Karzinomforschung; 1908;
- Miller, J.F. Immunological Function of the Thymus. Lancet 1961, 2, 748–749. [CrossRef]
- Miller, J.F.; Mitchell, G.F.; Weiss, N.S. Cellular Basis of the Immunological Defects in Thymectomized Mice. Nature 1967, 214, 992–997. [CrossRef]
- Burnet, M. Role of the Thymus and Related Organs in Immunity. Br Med J 1962, 2, 807–811. [CrossRef]
- Claman, H.N.; Chaperon, E.A.; Triplett, R.F. Thymus-Marrow Cell Combinations. Synergism in Antibody Production. Proc Soc Exp Biol Med 1966, 122, 1167–1171. [CrossRef]
- Wagner, H.; Röllinghoff, M.; Nossal, G.J. T-Cell-Mediated Immune Responses Induced in Vitro: A Probe for Allograft and Tumor Immunity. Transplant Rev 1973, 17, 3–36. [CrossRef]
- Carlson, R.D.; Flickinger, J.C.; Snook, A.E. Talkin’ Toxins: From Coley’s to Modern Cancer Immunotherapy. Toxins (Basel) 2020, 12, 241. [CrossRef]
- Currie, G.A. Eighty Years of Immunotherapy: A Review of Immunological Methods Used for the Treatment of Human Cancer. Br J Cancer 1972, 26, 141–153. [CrossRef]
- Von Leyden, V.; Blumenthal, F. Vorläufige Mittheilungen Über Einige Ergebnisse Der Krebsforshung Auf Der 1. Medizinschen Klinik. Dtsch. Med. Wschr. 1902, 28, 637–638.
- RISLEY, E.H. The Gilman–Coca Vaccine Emulsion Treatment of Cancer. The Boston Medical and Surgical Journal 1911, 165, 784–788.
- VAUGHAN, J.W. Cancer Vaccine and Anticancer Globulins as an Aid in the Surgical Treatment of Malignancy. Journal of the American Medical Association 1914, 63, 1258–1265.
- Dobosz, P.; Dzieciątkowski, T. The Intriguing History of Cancer Immunotherapy. Front Immunol 2019, 10, 2965. [CrossRef]
- Dock, G. The Influence of Complicating Diseases upon Leukaemia. The American Journal of the Medical Sciences (1827-1924) 1904, 127, 563.
- Larson, C.; Oronsky, B.; Scicinski, J.; Fanger, G.R.; Stirn, M.; Oronsky, A.; Reid, T.R. Going Viral: A Review of Replication-Selective Oncolytic Adenoviruses. Oncotarget 2015, 6, 19976–19989. [CrossRef]
- Jensen, C. Experimental Studies on Cancer in Mice (from German). Zentralblatt für Bacteriologie, Parasitenkunde und Infektionskrankheiten 1903, 34, 28–34.
- Little, C.C. A POSSIBLE MENDELIAN EXPLANATION FOR A TYPE OF INHERITANCE APPARENTLY NON-MENDELIAN IN NATURE. Science 1914, 40, 904–906. [CrossRef]
- Murphy, J.B.; Morton, J.J. The Lymphocyte as a Factor in Natural and Induced Resistance to Transplanted Cancer. Proc Natl Acad Sci U S A 1915, 1, 435–437. [CrossRef]
- The CRI Timeline Available online: https://www.cancerresearch.org/timeline (accessed on 26 July 2023).
- Everson, T.C.; Cole, W.H. Spontaneous Regression of Malignant Disease. J Am Med Assoc 1959, 169, 1758–1759. [CrossRef]
- Old, L.J.; Clarke, D.A.; Benacerraf, B. Effect of Bacillus Calmette-Guerin Infection on Transplanted Tumours in the Mouse. Nature 1959, 184(Suppl 5), 291–292. [CrossRef]
- Silverstein, A.M. The Curious Case of the 1960 Nobel Prize to Burnet and Medawar. Immunology 2016, 147, 269–274. [CrossRef]
- Liston, A. Immunological Tolerance 50 Years after the Burnet Nobel Prize. Immunol Cell Biol 2011, 89, 14–15. [CrossRef]
- Gowans, J.L.; McGREGOR, D.D.; Cowen, D.M. Initiation of Immune Responses by Small Lymphocytes. Nature 1962, 196, 651–655. [CrossRef]
- Miller, J.F.; Grant, G.A.; Roe, F.J. EFFECT OF THYMECTOMY ON THE INDUCTION OF SKIN TUMOURS BY 3,4-BENZOPYRENE. Nature 1963, 199, 920–922. [CrossRef]
- Miller, J.F. INFLUENCE OF THYMECTOMY ON TUMOR INDUCTION BY POLYOMA VIRUS IN C57BL MICE. Proc Soc Exp Biol Med 1964, 116, 323–327. [CrossRef]
- Miller, J.F. a. P. Events That Led to the Discovery of T-Cell Development and Function--a Personal Recollection. Tissue Antigens 2004, 63, 509–517. [CrossRef]
- Miller, J. The Early Work on the Discovery of the Function of the Thymus, an Interview with Jacques Miller. Cell Death Differ 2020, 27, 396–401. [CrossRef]
- Cerottini, J.C.; Nordin, A.A.; Brunner, K.T. In Vitro Cytotoxic Activity of Thymus Cells Sensitized to Alloantigens. Nature 1970, 227, 72–73. [CrossRef]
- Cerottini, J.C.; Nordin, A.A.; Brunner, K.T. Specific in Vitro Cytotoxicity of Thymus-Derived Lymphocytes Sensitized to Alloantigens. Nature 1970, 228, 1308–1309. [CrossRef]
- Steinman, R.M.; Cohn, Z.A. Identification of a Novel Cell Type in Peripheral Lymphoid Organs of Mice. I. Morphology, Quantitation, Tissue Distribution. J Exp Med 1973, 137, 1142–1162. [CrossRef]
- Banchereau, J.; Steinman, R.M. Dendritic Cells and the Control of Immunity. Nature 1998, 392, 245–252. [CrossRef]
- Geissmann, F.; Gordon, S.; Hume, D.A.; Mowat, A.M.; Randolph, G.J. Unravelling Mononuclear Phagocyte Heterogeneity. Nat Rev Immunol 2010, 10, 453–460. [CrossRef]
- Zinkernagel, R.M.; Doherty, P.C. Restriction of in Vitro T Cell-Mediated Cytotoxicity in Lymphocytic Choriomeningitis within a Syngeneic or Semiallogeneic System. Nature 1974, 248, 701–702. [CrossRef]
- Zinkernagel, R.M.; Doherty, P.C. Immunological Surveillance against Altered Self Components by Sensitised T Lymphocytes in Lymphocytic Choriomeningitis. Nature 1974, 251, 547–548. [CrossRef]
- Doherty, P.C. Challenged by Complexity: My Twentieth Century in Immunology. Annu Rev Immunol 2007, 25, 1–19. [CrossRef]
- Raju, T.N. The Nobel Chronicles. 1996: Peter Charles Doherty (b 1940) and Rolf M Zinkernagel (b 1944). Lancet 2000, 356, 172. [CrossRef]
- Volchenkov, R.; Sprater, F.; Vogelsang, P.; Appel, S. The 2011 Nobel Prize in Physiology or Medicine. Scand J Immunol 2012, 75, 1–4. [CrossRef]
- Chen, J.; Gao, L.; Wu, X.; Fan, Y.; Liu, M.; Peng, L.; Song, J.; Li, B.; Liu, A.; Bao, F. BCG-Induced Trained Immunity: History, Mechanisms and Potential Applications. J Transl Med 2023, 21, 106. [CrossRef]
- Pearl, R. On the Pathological Relations Between Cancer and Tuberculosis. Proceedings of the Society for Experimental Biology and Medicine 1928, 26, 73–75. [CrossRef]
- Morales, A.; Eidinger, D. Bacillus Calmette-Guerin in the Treatment of Adenocarcinoma of the Kidney. J Urol 1976, 115, 377–380. [CrossRef]
- Morales, A.; Eidinger, D.; Bruce, A.W. Intracavitary Bacillus Calmette-Guerin in the Treatment of Superficial Bladder Tumors. J Urol 1976, 116, 180–183. [CrossRef]
- Hanna, M.G.; Peters, L.C. Specific Immunotherapy of Established Visceral Micrometastases by BCG-Tumor Cell Vaccine Alone or as an Adjunct to Surgery. Cancer 1978, 42, 2613–2625. [CrossRef]
- Grant, R.M.; Mackie, R.; Cochran, A.J.; Murray, E.L.; Hoyle, D.; Ross, C. Results of Administering B.C.G. to Patients with Melanoma. Lancet 1974, 2, 1096–1100. [CrossRef]
- Donaldson, R.C. Chemoimmunotherapy for Cancer of the Head and Neck. Am J Surg 1973, 126, 507–512. [CrossRef]
- Carswell, E.A.; Old, L.J.; Kassel, R.L.; Green, S.; Fiore, N.; Williamson, B. An Endotoxin-Induced Serum Factor That Causes Necrosis of Tumors. Proc Natl Acad Sci U S A 1975, 72, 3666–3670. [CrossRef]
- Old, L.J. Cancer Immunology. Sci Am 1977, 236, 62–70, 72–73, 76, 79. [CrossRef]
- Ashley, M.P.; Zbar, B.; Hunter, J.T.; Rapp, H.J.; Sugimoto, T. Adjuvant-Antigen Requirements for Active Specific Immunotherapy of Microscopic Metastases Remaining after Surgery. Cancer Res 1980, 40, 4197–4203.
- Sukumar, S.; Hunter, J.T.; Terata, N.; Rapp, H.J. Eradication of Microscopic Hepatic Metastases by Active Specific Immunization. Cancer Immunol Immunother 1983, 14, 151–154. [CrossRef]
- Bier, H.; Armonat, G.; Bier, J.; Schirrmacher, V.; Ganzer, U. Postoperative Active-Specific Immunotherapy of Lymph Node Micrometastasis in a Guinea Pig Tumor Model. ORL J Otorhinolaryngol Relat Spec 1989, 51, 197–205. [CrossRef]
- Morton, D.L.; Foshag, L.J.; Hoon, D.S.; Nizze, J.A.; Famatiga, E.; Wanek, L.A.; Chang, C.; Davtyan, D.G.; Gupta, R.K.; Elashoff, R. Prolongation of Survival in Metastatic Melanoma after Active Specific Immunotherapy with a New Polyvalent Melanoma Vaccine. Ann Surg 1992, 216, 463–482. [CrossRef]
- Böhle, A.; Brandau, S. Immune Mechanisms in Bacillus Calmette-Guerin Immunotherapy for Superficial Bladder Cancer. J Urol 2003, 170, 964–969. [CrossRef]
- de Jong, W.H.; Teppema, J.S.; Wagenaar, S.S.; Paques, M.; Steerenberg, P.A.; Ruitenberg, E.J. Histological Evaluation of Immunologically Mediated Tumor Regression of the Line 10 Guinea Pig Hepatocarcinoma. Virchows Arch B Cell Pathol Incl Mol Pathol 1986, 50, 249–269. [CrossRef]
- Miyazaki, J.; Onozawa, M.; Takaoka, E.; Yano, I. Bacillus Calmette-Guérin Strain Differences as the Basis for Immunotherapies against Bladder Cancer. Int J Urol 2018, 25, 405–413. [CrossRef]
- Klein, E. Lymphocyte-Mediated Lysis of Tumor Cells in Vitro. Antigen-Restricted Clonal and Unrestricted Polyclonal Effects. Springer Semin Immunopathol 1982, 5, 147–159. [CrossRef]
- Stauss, H.J.; Van Waes, C.; Fink, M.A.; Starr, B.; Schreiber, H. Identification of a Unique Tumor Antigen as Rejection Antigen by Molecular Cloning and Gene Transfer. J Exp Med 1986, 164, 1516–1530. [CrossRef]
- Cole, W.H. Efforts to Explain Spontaneous Regression of Cancer. J Surg Oncol 1981, 17, 201–209. [CrossRef]
- Thomas, L. On Immunosurveillance in Human Cancer. Yale J Biol Med 1982, 55, 329–333.
- Burnet, M. Donn. Br Med Bull 1964, 20, 154–158. [CrossRef]
- Ribatti, D. The Concept of Immune Surveillance against Tumors. The First Theories. Oncotarget 2017, 8, 7175–7180. [CrossRef]
- Brunet, J.F.; Denizot, F.; Luciani, M.F.; Roux-Dosseto, M.; Suzan, M.; Mattei, M.G.; Golstein, P. A New Member of the Immunoglobulin Superfamily--CTLA-4. Nature 1987, 328, 267–270. [CrossRef]
- Linsley, P.S.; Wallace, P.M.; Johnson, J.; Gibson, M.G.; Greene, J.L.; Ledbetter, J.A.; Singh, C.; Tepper, M.A. Immunosuppression in Vivo by a Soluble Form of the CTLA-4 T Cell Activation Molecule. Science 1992, 257, 792–795. [CrossRef]
- Iwai, Y.; Ishida, M.; Tanaka, Y.; Okazaki, T.; Honjo, T.; Minato, N. Involvement of PD-L1 on Tumor Cells in the Escape from Host Immune System and Tumor Immunotherapy by PD-L1 Blockade. Proc Natl Acad Sci U S A 2002, 99, 12293–12297. [CrossRef]
- Dong, H.; Strome, S.E.; Salomao, D.R.; Tamura, H.; Hirano, F.; Flies, D.B.; Roche, P.C.; Lu, J.; Zhu, G.; Tamada, K.; et al. Tumor-Associated B7-H1 Promotes T-Cell Apoptosis: A Potential Mechanism of Immune Evasion. Nat Med 2002, 8, 793–800. [CrossRef]
- Chen, D.S.; Mellman, I. Elements of Cancer Immunity and the Cancer-Immune Set Point. Nature 2017, 541, 321–330. [CrossRef]
- Chen, Y.-S.; Shen, C.-R. Immune Checkpoint Blockade Therapy: The 2014 Tang Prize in Biopharmaceutical Science. Biomed J 2015, 38, 5–8. [CrossRef]
- Huang, P.-W.; Chang, J.W.-C. Immune Checkpoint Inhibitors Win the 2018 Nobel Prize. Biomed J 2019, 42, 299–306. [CrossRef]
- Mitchell, M.S.; Kan-Mitchell, J.; Kempf, R.A.; Harel, W.; Shau, H.Y.; Lind, S. Active Specific Immunotherapy for Melanoma: Phase I Trial of Allogeneic Lysates and a Novel Adjuvant. Cancer Res 1988, 48, 5883–5893.
- Mitchell, M.S.; Harel, W.; Kempf, R.A.; Hu, E.; Kan-Mitchell, J.; Boswell, W.D.; Dean, G.; Stevenson, L. Active-Specific Immunotherapy for Melanoma. J Clin Oncol 1990, 8, 856–869. [CrossRef]
- Mitchell, M.S. Perspective on Allogeneic Melanoma Lysates in Active Specific Immunotherapy. Semin Oncol 1998, 25, 623–635.
- Sondak, V.K.; Sosman, J.A. Results of Clinical Trials with an Allogenic Melanoma Tumor Cell Lysate Vaccine: Melacine. Semin Cancer Biol 2003, 13, 409–415. [CrossRef]
- Sondak, V.K.; Liu, P.-Y.; Tuthill, R.J.; Kempf, R.A.; Unger, J.M.; Sosman, J.A.; Thompson, J.A.; Weiss, G.R.; Redman, B.G.; Jakowatz, J.G.; et al. Adjuvant Immunotherapy of Resected, Intermediate-Thickness, Node-Negative Melanoma with an Allogeneic Tumor Vaccine: Overall Results of a Randomized Trial of the Southwest Oncology Group. J Clin Oncol 2002, 20, 2058–2066. [CrossRef]
- Morton, D.L.; Mozzillo, N.; Thompson, J.F.; Kelley, M.C.; Faries, M.; Wagner, J.; Schneebaum, S.; Schuchter, L.; Gammon, G.; Elashoff, R. An International, Randomized, Phase III Trial of Bacillus Calmette-Guerin (BCG) plus Allogeneic Melanoma Vaccine (MCV) or Placebo after Complete Resection of Melanoma Metastatic to Regional or Distant Sites. JCO 2007, 25, 8508–8508. [CrossRef]
- Apostólico, J. de S.; Lunardelli, V.A.S.; Coirada, F.C.; Boscardin, S.B.; Rosa, D.S. Adjuvants: Classification, Modus Operandi, and Licensing. J Immunol Res 2016, 2016, 1459394. [CrossRef]
- Firdaus, F.Z.; Skwarczynski, M.; Toth, I. Developments in Vaccine Adjuvants. Methods Mol Biol 2022, 2412, 145–178. [CrossRef]
- Czerkinsky, C.C.; Nilsson, L.A.; Nygren, H.; Ouchterlony, O.; Tarkowski, A. A Solid-Phase Enzyme-Linked Immunospot (ELISPOT) Assay for Enumeration of Specific Antibody-Secreting Cells. J Immunol Methods 1983, 65, 109–121. [CrossRef]
- Slota, M.; Lim, J.-B.; Dang, Y.; Disis, M.L. ELISpot for Measuring Human Immune Responses to Vaccines. Expert Rev Vaccines 2011, 10, 299–306. [CrossRef]
- Ranieri, E.; Netti, G.S.; Gigante, M. CTL ELISPOT Assay and T Cell Detection. Methods Mol Biol 2021, 2325, 65–77. [CrossRef]
- Saxena, M.; Bhardwaj, N. Turbocharging Vaccines: Emerging Adjuvants for Dendritic Cell Based Therapeutic Cancer Vaccines. Curr Opin Immunol 2017, 47, 35–43. [CrossRef]
- Inaba, K.; Inaba, M.; Romani, N.; Aya, H.; Deguchi, M.; Ikehara, S.; Muramatsu, S.; Steinman, R.M. Generation of Large Numbers of Dendritic Cells from Mouse Bone Marrow Cultures Supplemented with Granulocyte/Macrophage Colony-Stimulating Factor. J Exp Med 1992, 176, 1693–1702. [CrossRef]
- Young, J.W.; Szabolcs, P.; Moore, M.A. Identification of Dendritic Cell Colony-Forming Units among Normal Human CD34+ Bone Marrow Progenitors That Are Expanded by c-Kit-Ligand and Yield Pure Dendritic Cell Colonies in the Presence of Granulocyte/Macrophage Colony-Stimulating Factor and Tumor Necrosis Factor Alpha. J Exp Med 1995, 182, 1111–1119. [CrossRef]
- De Plaen, E.; Lurquin, C.; Van Pel, A.; Mariamé, B.; Szikora, J.P.; Wölfel, T.; Sibille, C.; Chomez, P.; Boon, T. Immunogenic (Tum-) Variants of Mouse Tumor P815: Cloning of the Gene of Tum- Antigen P91A and Identification of the Tum- Mutation. Proc Natl Acad Sci U S A 1988, 85, 2274–2278. [CrossRef]
- van der Bruggen, P.; Traversari, C.; Chomez, P.; Lurquin, C.; De Plaen, E.; Van den Eynde, B.; Knuth, A.; Boon, T. A Gene Encoding an Antigen Recognized by Cytolytic T Lymphocytes on a Human Melanoma. Science 1991, 254, 1643–1647. [CrossRef]
- Boon, T.; van der Bruggen, P. Human Tumor Antigens Recognized by T Lymphocytes. J Exp Med 1996, 183, 725–729. [CrossRef]
- Wang, R.F.; Rosenberg, S.A. Human Tumor Antigens for Cancer Vaccine Development. Immunol Rev 1999, 170, 85–100. [CrossRef]
- Devaraja, K.; Aggarwal, S.; Verma, S.S.; Gupta, S.C. Clinico-Pathological Peculiarities of Human Papilloma Virus Driven Head and Neck Squamous Cell Carcinoma: A Comprehensive Update. Life Sci 2020, 245, 117383. [CrossRef]
- Coulie, P.G.; Van den Eynde, B.J.; van der Bruggen, P.; Boon, T. Tumour Antigens Recognized by T Lymphocytes: At the Core of Cancer Immunotherapy. Nat Rev Cancer 2014, 14, 135–146. [CrossRef]
- Zarour, H.M.; DeLeo, A.; Finn, O.J.; Storkus, W.J. Categories of Tumor Antigens. Holland-Frei Cancer Medicine. 6th edition 2003.
- Kosaka, A.; Yajima, Y.; Hatayama, M.; Ikuta, K.; Sasaki, T.; Hirai, N.; Yasuda, S.; Nagata, M.; Hayashi, R.; Harabuchi, S.; et al. A Stealth Antigen SPESP1, Which Is Epigenetically Silenced in Tumors, Is a Suitable Target for Cancer Immunotherapy. Cancer Sci 2021, 112, 2705–2713. [CrossRef]
- Yajima, Y.; Kosaka, A.; Ishibashi, K.; Yasuda, S.; Komatsuda, H.; Nagato, T.; Oikawa, K.; Kitada, M.; Takekawa, M.; Kumai, T.; et al. A Tumor Metastasis-Associated Molecule TWIST1 Is a Favorable Target for Cancer Immunotherapy Due to Its Immunogenicity. Cancer Sci 2022, 113, 2526–2535. [CrossRef]
- Cheever, M.A.; Allison, J.P.; Ferris, A.S.; Finn, O.J.; Hastings, B.M.; Hecht, T.T.; Mellman, I.; Prindiville, S.A.; Viner, J.L.; Weiner, L.M.; et al. The Prioritization of Cancer Antigens: A National Cancer Institute Pilot Project for the Acceleration of Translational Research. Clin Cancer Res 2009, 15, 5323–5337. [CrossRef]
- Dunn, G.P.; Bruce, A.T.; Ikeda, H.; Old, L.J.; Schreiber, R.D. Cancer Immunoediting: From Immunosurveillance to Tumor Escape. Nat Immunol 2002, 3, 991–998. [CrossRef]
- Dunn, G.P.; Old, L.J.; Schreiber, R.D. The Immunobiology of Cancer Immunosurveillance and Immunoediting. Immunity 2004, 21, 137–148. [CrossRef]
- Song, Q.; Zhang, C.-D.; Wu, X.-H. Therapeutic Cancer Vaccines: From Initial Findings to Prospects. Immunol Lett 2018, 196, 11–21. [CrossRef]
- Shibata, H.; Xu, N.; Saito, S.; Zhou, L.; Ozgenc, I.; Webb, J.; Fu, C.; Zolkind, P.; Egloff, A.M.; Uppaluri, R. Integrating CD4+ T Cell Help for Therapeutic Cancer Vaccination in a Preclinical Head and Neck Cancer Model. Oncoimmunology 2021, 10, 1958589. [CrossRef]
- Wei, C.; Ma, Y.; Wang, F.; Liao, Y.; Chen, Y.; Zhao, B.; Zhao, Q.; Wang, D.; Tang, D. Igniting Hope for Tumor Immunotherapy: Promoting the “Hot and Cold” Tumor Transition. Clin Med Insights Oncol 2022, 16, 11795549221120708. [CrossRef]
- Smyth, M.J.; Godfrey, D.I.; Trapani, J.A. A Fresh Look at Tumor Immunosurveillance and Immunotherapy. Nat Immunol 2001, 2, 293–299. [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The next Generation. Cell 2011, 144, 646–674. [CrossRef]
- Hegde, P.S.; Karanikas, V.; Evers, S. The Where, the When, and the How of Immune Monitoring for Cancer Immunotherapies in the Era of Checkpoint Inhibition. Clin Cancer Res 2016, 22, 1865–1874. [CrossRef]
- de Visser, K.E.; Eichten, A.; Coussens, L.M. Paradoxical Roles of the Immune System during Cancer Development. Nat Rev Cancer 2006, 6, 24–37. [CrossRef]
- Kather, J.N.; Suarez-Carmona, M.; Charoentong, P.; Weis, C.-A.; Hirsch, D.; Bankhead, P.; Horning, M.; Ferber, D.; Kel, I.; Herpel, E.; et al. Topography of Cancer-Associated Immune Cells in Human Solid Tumors. Elife 2018, 7, e36967. [CrossRef]
- Troiano, G.; Rubini, C.; Togni, L.; Caponio, V.C.A.; Zhurakivska, K.; Santarelli, A.; Cirillo, N.; Lo Muzio, L.; Mascitti, M. The Immune Phenotype of Tongue Squamous Cell Carcinoma Predicts Early Relapse and Poor Prognosis. Cancer Med 2020, 9, 8333–8344. [CrossRef]
- Duan, Q.; Zhang, H.; Zheng, J.; Zhang, L. Turning Cold into Hot: Firing up the Tumor Microenvironment. Trends Cancer 2020, 6, 605–618. [CrossRef]
- Higano, C.S.; Schellhammer, P.F.; Small, E.J.; Burch, P.A.; Nemunaitis, J.; Yuh, L.; Provost, N.; Frohlich, M.W. Integrated Data from 2 Randomized, Double-Blind, Placebo-Controlled, Phase 3 Trials of Active Cellular Immunotherapy with Sipuleucel-T in Advanced Prostate Cancer. Cancer 2009, 115, 3670–3679. [CrossRef]
- Burch, P.A.; Croghan, G.A.; Gastineau, D.A.; Jones, L.A.; Kaur, J.S.; Kylstra, J.W.; Richardson, R.L.; Valone, F.H.; Vuk-Pavlović, S. Immunotherapy (APC8015, Provenge) Targeting Prostatic Acid Phosphatase Can Induce Durable Remission of Metastatic Androgen-Independent Prostate Cancer: A Phase 2 Trial. Prostate 2004, 60, 197–204. [CrossRef]
- Kantoff, P.W.; Higano, C.S.; Shore, N.D.; Berger, E.R.; Small, E.J.; Penson, D.F.; Redfern, C.H.; Ferrari, A.C.; Dreicer, R.; Sims, R.B.; et al. Sipuleucel-T Immunotherapy for Castration-Resistant Prostate Cancer. N Engl J Med 2010, 363, 411–422. [CrossRef]
- Cheever, M.A.; Higano, C.S. PROVENGE (Sipuleucel-T) in Prostate Cancer: The First FDA-Approved Therapeutic Cancer Vaccine. Clin Cancer Res 2011, 17, 3520–3526. [CrossRef]
Figure 1.
Illustration depicting the evolution of cancer immunobiology leading to introduction of several therapeutic avenues, such as tumor microenvironment, therapeutic vaccines and checkpoint inhibitors.
Figure 1.
Illustration depicting the evolution of cancer immunobiology leading to introduction of several therapeutic avenues, such as tumor microenvironment, therapeutic vaccines and checkpoint inhibitors.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



