Submitted:
23 September 2024
Posted:
24 September 2024
You are already at the latest version
Abstract
Ox Tongue (Echium amoenum) is an important medicinal plant rich in secondary metabolites, including alkaloids, quinones, and unsaturated fatty acids. The present study examined the concentration of oil, fatty acid profiles, and antioxidant activity of six different Iranian Echium accessions. The oil concentration varied across these populations, ranging from 16.20% to 22.36% in the first year and from 21.00% to 25.50% in the second year. The major fatty acids identified in the oils were oleic acid (18.35% to 19.40%), linoleic acid (23.80% to 26.20%), and α-linolenic acid (32.30% to 34.20%) during the first year. In the second year, the predominant fatty acids were oleic acid (15.82% to 19.32%), linoleic acid (21.55% to 24.70%), and α-linolenic acid (33.95% to 37.40%). Principal component analysis revealed distinct profiles for each population: Sari was notable for its high α-linolenic acid and γ-linolenic acid content, Fasa was distinguished by its high stearidonic acid levels, Roodsar had a high concentration of stearic acid, Vazneh Sar was rich in linoleic acid and palmitic acid, and Leron had the highest oleic acid content. The Sari population had the highest oil content at 25.50%, while Fasa had the highest α-linolenic acid content at 37.40% during the establishment year. Leron exhibited the highest antioxidant attributes. These findings indicate that selecting elite Echium populations with high levels of unsaturated fatty acids and potent antioxidant properties could be advantageous for the food industry, as they offer enhanced nutritional and functional qualities.
Keywords:
Ox tongue
; Fatty acids
; Antioxidant
; α-linolenic acid
1. Introduction
Echium amoenum, commonly known as Iranian ox tongue, is a perennial herb in the Boraginaceae family. It's predominantly found in northern Iran and parts of the Caucasus region, thriving at elevations exceeding 2500 meters above sea level [1]. In traditional medicine, various parts of the plant including flowers, seeds, and branches have been utilized for multiple purposes. These include purifying blood, softening breast tissue, strengthening the heart, promoting urination, inducing calmness, reducing inflammation, and alleviating respiratory issues such as coughs [2]. Contemporary scientific investigations have revealed that E. amoenum possesses properties that enhance energy levels and boost immune function. These effects are attributed to the plant's rich composition of secondary metabolites, including alkaloids, quinones, and unsaturated fatty acids, which also contribute to its antimicrobial and disinfectant capabilities [3,2]. The plant's flowers are particularly notable for their diverse array of biologically active compounds. These include: essential oils [4], Fatty acids [5], Phytosterols [6], various minerals, anthocyanins, alavonoid aglycones, saponins, pyrrolizidine alkaloids, this extensive range of bioactive substances underlies the plant's potential for various therapeutic applications, supporting its traditional uses and suggesting ways for future pharmacological research [7] and pyrrolizidine alkaloids [8].
Research on the oil content and composition of Iranian ox tongue (Echium amoenum) seeds is limited. The seed oil extraction efficiency is approximately 45%, with linolenic acid being the primary fatty acid, constituting about 75% of the oil [9].
Earlier studies have identified seven fatty acids in Echium amoenum oil, with linolenic, linoleic, and oleic acids being the most prevalent [5]. Additionally, researchers have isolated and characterized four toxic alkaloids in the plant such as echimidine I, echimidine isomer II, 7-angeloyl retronecine III, and 7-tigloyl retronecine IV [8].
Plant oils are highly valued natural products that hold significant promise as sustainable bioenergy sources and as important components of the human diet [10]. These oils are often rich in unsaturated fatty acids, including monounsaturated and polyunsaturated types, which are known for their health benefits. Unsaturated fatty acids play a crucial role in promoting cardiovascular health by lowering cholesterol levels, thereby reducing the risk of heart disease and other related conditions. Oils that are high in monounsaturated fats, like oleic acid, can enhance the balance of good (HDL) cholesterol relative to bad (LDL) cholesterol, contributing to overall heart health. [10].
Moreover, plant oils rich in polyunsaturated fatty acids, such as linoleic acid and α-linolenic acid, provide essential nutrients that the human body cannot synthesize on its own, necessitating their inclusion in the diet [11]. These essential fatty acids are vital for various physiological functions, including cell membrane integrity, inflammatory response modulation, and brain and visual development. Linoleic acid, an omega-6 fatty acid, and α-linolenic acid, an omega-3 fatty acid, are particularly important for maintaining optimal health, supporting metabolic processes, and contributing to the prevention of chronic diseases. Given their dual role in nutrition and bioenergy, plant oils represent a key area of interest for both health-focused dietary applications and renewable energy solutions, underscoring their multifaceted value in promoting both human well-being and environmental sustainability [11,10].
Compared to animal-derived oils, plant oils generally have lower melting points, lower viscosity, and higher purity, making them easier to extract. Consequently, research into the accumulation and quality of fatty acids in plant oils has become a significant focus in plant metabolomics [12].
The current study is aimed to provide an in-depth evaluation of the natural variability in oil concentration, fatty acid composition, yield-related traits, and antioxidant properties among six distinct Iranian Echium accessions. The primary goal is to thoroughly characterize these populations to understand the extent of their genetic and phenotypic diversity in these key attributes.
In addition, the research findings are expected to guide future breeding efforts by providing a scientific basis for the selection of populations with the most desirable traits. Populations with high unsaturated fatty acid content are especially valuable due to their health benefits and relevance in food and cosmetic industries. Accessions with increased oil yield and enhanced antioxidant properties will contribute to more efficient and sustainable production processes.
Ultimately, this comprehensive assessment aims to facilitate the breeding of Echium varieties that not only meet industry standards but also offer improved nutritional and functional benefits, thereby expanding their commercial potential. The insights gained from this study will be instrumental in advancing the breeding program and optimizing the use of Echium in various applications, enhancing the economic and ecological value of these plants.
2. Materials and Methods
2.1. Chemicals and Reagents
The study employed a range of chemicals and reagents to conduct various analyses, including Folin & Ciocalteu's phenol reagent, 2,2-diphenyl-1-picrylhydrazyl (DPPH), gallic acid, rutin, aluminum chloride (AlCl₃), hydrochloric acid (HCl), sodium bicarbonate (NaHCO₃), methanol, and n-hexane sourced from Sigma-Aldrich (USA) and Merck (Germany).
2.2. Plant Material
Seeds from six wild population of Echium amoenum L., sourced from diverse locations across Iran were used in this study (Table 1). The seeds were cultivated in a controlled greenhouse environment, with planting occurring on September 11, 2022. The temperature of the greenhouse maintained between 10°C and 15°C daily. After a month of growth, the seedlings were thinned to three healthiest per container. The selected plants were further relocated to the Shahid Fozveh research facility in Iran (coordinates: 32°36′N, 51°26′E; elevation: 1612 meters) in mid-October 2021. The experiment site is categorized as mild semi-arid under Ghosen's system. The area experiences an annual mean temperature of 16°C, with extremes ranging from -17°C to 40°C. Annual precipitation in the area averages 140 mm, as reported by [13].
2.3. Soil Chemical and Physical Analysis
Before transplanting, soil samples were taken from the top 30 cm layer to determine the key chemical and physical characteristics. Various analytical methods were employed:
Total nitrogen: measured using the Kjeldahl method [14]. Available phosphorus extracted with sodium bicarbonate and quantified via the molybdo-phosphate blue color technique [15], potassium: extracted using ammonium acetate and analyzed through flame photometry [16]. The soil was classified as silty clay in texture. Its physicochemical profile was as follows: pH: 7.6, organic matter content: 0.42%, electrical conductivity (EC): 2.8 dS m−1. Total nitrogen: 0.14%, available phosphorus: 10 mg kg−1, available potassium: 280 mg kg−1.
2.4. Experimental Design and Treatments
The six wild populations of Echium amoenum L., were arranged in a randomized complete block design (RCBD) with three repetitions. Plot dimensions were 4×2 m and plants were grown in four equidistant rows in each plot. The row spacing was 50 cm with 50 cm between individual plants within each row. The study adhered to natural growing conditions, avoiding the use of fertilizers or pesticides throughout the experimental period. Weed control was performed manually as needed to maintain plot integrity.
2.5. Morphological Characteristics
At peak bloom, several plant attributes, including plant height, canopy diameter, number of flowers , leaf dimensions (length and width) were recorded. In post-harvesting, samples were dehydrated in an oven at 40°C to constant weight. For each distinct population, two additional measurements, including 1000-seeds weight and seed yield per individual plant were determined.
2.6. Photosynthetic Pigments
The concentrations of chlorophyll a, chlorophyll b, and total carotenoids were measured using the method developed by [17]. To begin the process, 0.5 gram of fresh plant material, specifically young and fully developed leaves, were carefully weighed and placed into a mortar. Liquid nitrogen was added to the mortar to freeze the plant material, making it brittle and easier to grind thoroughly into a fine powder. Once the plant material was ground, 20 mL of 80% acetone was added to the powdered sample. This mixture facilitated the extraction of the pigments. The resulting solution was subjected to centrifugation at 6,000 rpm for 10 minutes. After centrifugation, the clear upper layer (supernatant) containing the extracted pigments was carefully decanted and transferred into a glass flask. For the spectrophotometric analysis, a portion of the extracted solution was poured into a spectrophotometer cuvette. Absorbance measurements were taken at three specific wavelengths: 663 nm, 645 nm, and 470 nm using a UNICO UV-2100 spectrophotometer. These wavelengths correspond to the absorbance peaks of chlorophyll a, chlorophyll b, and carotenoids, respectively.
The concentrations of chlorophyll a and chlorophyll b were calculated using specific formulas, which relate the absorbance values to the pigment concentrations. The results were expressed in milligram per gram of fresh sample weight, providing a precise quantification of the chlorophyll and carotenoid content in the plant material.
Chlorophyll a (Chl a) = 12.25 A663.2 − 2.79 A646.8
Chlorophyll b (Chl b) = 21.5 A646.8 − 5.1 A663.2
Carotenoids = (1000A470 − 1.82Chla − 85.02Chl b) ∕198
2.7. Plant Extract
For the preparation of each plant extract, 5 gram of ground dried samples were mixed with 50 mL of absolute methanol and shaken for 72 hours. The mixture was then filtered through Whatman filter paper (No. 4) and left in the laboratory for 48 hours at an appropriate temperature.
2.8. Determination of Total Phenol
To assess the total phenol content in fully expanded young petals, the Folin-Ciocalteu method was employed, following the procedure outlined by [18]. The process began by preparing extracts from the petals at various concentrations, derived from different treatments. For each sample, 10 μL of the extract was combined with 490 μL of distilled water and 500 μL of Folin-Ciocalteu reagent. This mixture was thoroughly mixed using a vortex mixer for 5 minutes to ensure proper reaction. Subsequently, 3 mL of a 1% sodium carbonate solution was added to the mixture. This addition caused the formation of a blue-colored solution, indicating the presence of phenolic compounds. The reaction mixture was then allowed to stand at room temperature for two hours to complete the development of color. After the incubation period, the absorbance of the blue solution was measured at a wavelength of 765 nm using a UNICO UV-2100 spectrophotometer. To quantify the phenolic content, a calibration curve was prepared using gallic acid, with a correlation coefficient (R²) of 0.998. The total phenolic content in the extracts was then calculated and expressed as milligram of gallic acid equivalents per gram of extract (mg GAE/g extract).
2.9. Determination of Total Flavonoids
The total flavonoid content in fully expanded young petals was assessed using the methodology described by [19]. The process began with the preparation of the petal’s samples: 0.1 gram of fresh, fully developed petals were weighed accurately. The petal samples were then subjected to extraction by mixing with 10 mL of 96% ethanol. This solvent was used to dissolve and extract the flavonoids present in the petals tissue. After the extraction, a portion of the extract was prepared for analysis. Specifically, 1 mL of the extracted solution was mixed with 2 mL of a 2% aluminum chloride solution and 6 mL of a 5% potassium acetate solution. These reagents react with flavonoids, leading to the formation of a colored complex. The reaction mixture was incubated at room temperature, (37°C), for a duration of 40 minutes. This incubation period allowed sufficient time for the color development, which is indicative of the flavonoid content. Following the incubation, the absorbance of the resulting solution was measured at a wavelength of 450 nm using a UNICO UV-2100 spectrophotometer. This measurement provides an indication of the flavonoid concentration based on the intensity of the color formed. To ensure the reliability of the results, each sample was analyzed in triplicate, meaning that three independent measurements were taken for each sample to verify accuracy and consistency. To quantify the flavonoid content, a calibration curve was constructed using rutin as a standard. Rutin solutions at concentrations of 0.011 and 0.370 mg mL⁻¹ were used to create this calibration curve, which had a high correlation coefficient (R²) of 0.999. The flavonoid concentration in each sample extract was then determined by comparing the absorbance values to this calibration curve, and the results were reported as milligrams of rutin per gram of dry plant extract.
2.10. Total Anthocyanin Content
The concentration of total anthocyanins was measured using the method established by [20]. For this analysis, 0.1 gram of dried petals was immersed in 10 mL of acidified methanol, prepared by mixing methanol with hydrochloric acid in a 99:1 ratio (v/v). This acidified methanol solution aids in the extraction of anthocyanins from the petals. The mixture of petals and acidified methanol was then stored in a dark environment at a temperature of 4°C for a period of 12 hours. This storage period allowed sufficient time for the anthocyanins to be extracted into the methanol solution. After extraction, the mixture was centrifuged at 10,000 rpm for 10 minutes to separate the liquid extract (supernatant) from any remaining solid residue. The supernatant, which contains the anthocyanins, was then analyzed. The absorbance of the supernatant was measured at a wavelength of 550 nm using a spectrophotometer. This measurement is critical for determining the concentration of anthocyanins, as the absorbance at this wavelength corresponds to the presence of these pigments. The total anthocyanin content was calculated using the extinction coefficient of cyanidin-3-glucoside, a standard reference compound. The results were expressed as milligram of cyanidin equivalents per gram of dry weight (mg CyE/g DW) of the Echium petals.
2.11. Determination of Antioxidant Properties of Extracts
The antioxidant properties of the extracts were assessed using the DPPH radical scavenging activity method outlined by [21]. To begin, extracts were prepared at a range of concentrations from 2.5 to 300 μg mL⁻¹. Each extract was then mixed with 3.9 mL of freshly prepared solution of DPPH (2,2-diphenyl-1-picrylhydrazyl) in methanol. DPPH is a stable free radical that reacts with antioxidants. The mixtures of extract and DPPH solution were incubated in a dark environment at room temperature for 30 minutes. This dark incubation prevents the degradation of the DPPH radical by light and allows time for any antioxidants present in the extracts to react with the DPPH radicals. After the incubation period, the degree of scavenging of the DPPH radicals by the extracts was quantified by measuring the absorbance of the solution at a wavelength of 515 nm using a UNICO UV-2100 spectrophotometer. The reduction in absorbance indicates the extent to which the DPPH radicals were neutralized by the antioxidants in the extracts. The antioxidant capacity of each extract was expressed as the percentage inhibition of DPPH free radicals. This percentage reflects the effectiveness of the extracts in neutralizing free radicals compared to a control sample with no antioxidant activity.
2.12. Extraction of Fixed Oils
Fixed oils were extracted using the Soxhlet method as outlined by [22] with the procedure performed in triplicate for accuracy. Initially, 10 gram of dried seed samples was finely ground using a heavy-duty grinder to increase the surface area for extraction. The ground samples were then combined with 100 mL of petroleum ether as the solvent. This mixture was agitated on a shaker set at 150 rpm for four hours to ensure thorough extraction of the oils. Following the shaking process, the samples were centrifuged at 4°C for 10 minutes to separate the oil-containing supernatant from the solid residues. The supernatant was carefully filtered through Whatman filter paper with a pore size of 0.45 μm to remove any remaining solid particles. The filtered oil solution was collected into clean glass vials.
To remove the petroleum ether and isolate the oil, the solution was subjected to evaporation using a rotary evaporator set at 40°C. This process gently removed the solvent without affecting the oil's quality. The extracted oil was then quantified, and its proportion was calculated using a specific formula to determine the percentage of oil content in the seeds.
2.13. Fatty Acid Methyl Ester (Preparation)
To prepare samples for gas chromatography analysis, free fatty acids were converted into fatty acid methyl esters (FAMEs) through a process of volatilization, as outlined in [23]. The procedure began by combining 0.1 gram of oil with 2 mL of isooctane and 0.1 mL of 2 N methanolic potassium hydroxide (2 Molar). This mixture was then agitated for one minute to facilitate the conversion of fatty acids into their methyl ester forms.
After shaking, the solution was allowed to settle for approximately two minutes. Following this, 2 mL of sodium chloride solution was added to the mixture, and the container was gently shaken to ensure thorough mixing. This step helps to separate the layers and facilitates the extraction of FAMEs into the isooctane layer.
The isooctane layer, which now contains the fatty acid methyl esters, was carefully separated and transferred to a new vial containing about 1 gram of sodium sulfate. The sodium sulfate serves as a drying agent to remove any residual moisture from the isooctane layer.
Finally, all prepared samples were stored at 4°C until analysis.
2.14. Quantification of Fatty Acids
The analysis of oils for fatty acid content was performed using gas chromatography (GC) with an Agilent model 7890A gas chromatograph. This instrument was fitted with a DB-WAX column, which is a fused polyethylene glycol (PEG) column with dimensions of 60 meters in length, 0.25 mm in diameter, and a film thickness of 0.25 μm.
The temperature program for the GC oven was as follows: it began at an initial temperature of 170°C, which was maintained for 5 minutes. Following this, the temperature was increased gradually at a rate of 1°C per minute until it reached 220°C.
Both the detector (Flame Ionization Detector, FID) and the injector were set to a temperature of 220°C. Nitrogen gas was used as the carrier gas, with a flow rate of 1 mL per minute, ensuring a consistent linear velocity throughout the analysis.
Quantification of the fatty acids was based on the area percentage obtained from the GC-FID (Gas Chromatography-Flame Ionization Detection) analysis. No correction factors were applied to the data during this quantification process [24].
2.15. Statistical Analysis
Statistical analyses were carried out to evaluate the data using various methods and software tools. One-way analysis of variance (ANOVA) was employed to determine significant differences among groups, followed by the least significant difference (LSD) test to identify specific group differences. These analyses were performed using SAS software version 9.4 (SAS Institute, USA). To explore the relationships and distinctions among the studied populations based on their biochemical and yield characteristics, cluster analysis was conducted using Euclidean distance metrics. This analysis was executed with R Studio software version 4.3.1. Additionally, Principal Component Analysis (PCA) was performed using R Studio to categorize the Echium populations. This analysis focused on the predominant fatty acids, oil content, yield traits, and antioxidant properties of the populations, providing a comprehensive overview of their distinct characteristics.
3. Results and Discussion
3.1. Morphological and Yield Characteristics
In both growing seasons, the impact of different accessions on various plant characteristics—such as the number of flowers, number of lateral branches, leaf length, 1000-seed weight, and seed weight per plant—was found to be statistically significant (p<0.01) (Table 2).
In the second year, the effect of accession was notably significant for plant height, canopy diameter, and leaf length. During the first season of the study, the Fasa accession produced a visually impressive display with the highest numbers of flowers and lateral branches, recording 311.70 flowers and 88.30 lateral branches, respectively.
Regarding leaf size, the Leron accession with 12.8 cm demonstrated the largest leaf lengthbut had the lowest flower count among the studied accessions. The Fasa accession had the highest 1000-seed weight at 1.33 grams, while the Sari out yielded other accessions and produced 6.00 grams of seeds per plant.
In the second year of the experiment, the Fasa accession led other accessions in plant height, canopy diameter, flower count, and number of lateral branches. The Leron and Vazneh Sar accessions had the greatest leaf length and width, measuring 13.70 cm and 3.50 cm, respectively. Additionally, the Roodsar accession produced the highest 1000-seed weight, which was 10.30 grams, whereas the Sari accession had the highest seed weight per plant at 6.5 grams. The findings of this study revealed that the upper range of 1000-seed weight of E. amoenium populations in the second year surpassed the values reported in earlier studies in various regions across Iran [5].
3.2. Photosynthetic Pigments
Table 3 presents the chlorophyll a, chlorophyll b, and carotenoids levels of various accessions.
In the first year of the study (stablishment year), the Ghazvin accession yielded the highest amounts of photosynthetic pigments compared to the other accessions. The carotenoid content in the Ghazvin accession was 68% greater than in the Fasa accession, which had the lowest carotenoid levels among the accessions tested (Table 3).
During the initial establishment year, the Ghazvin accession also exhibited the highest concentrations of chlorophyll a and chlorophyll b, recording levels of 7.20 mg/g and 6.30 mg/g, respectively.
Earlier report by [25] also indicated that there were significant variations in chlorophyll concentrations among various cultivars of Echium plantagineum during flowering stage.
As plants matured, the Fasa population exhibited the highest concentrations of chlorophyll a and b, measuring 15.60 mg/g and 7.6 mg/g, respectively in the second year. Conversely, the Leron population contained less chlorophyll compared to the other accessions, with concentrations of 3.20 mg/g and 1.40 mg/g for chlorophyll a and b, respectively.
Overall, the variations in chlorophyll content among different populations of Echium plantagineum can be attributed to their geographical locations and environmental conditions, as noted by [26]. For example, populations from Vazneh Sar and Leron, located in the northern region with a climate characterized by heavy rainfall and high humidity, exhibited poor establishment and the lowest concentrations of photosynthetic pigments.
3.3. Total Phenol Content
In this study, significant variation was observed in the total polyphenol content among Echium populations across both growing seasons. Each population displayed distinct levels of total phenols. According to (Table 3), the Ghazvin population recorded the highest total polyphenol contents, with values of 32.90 mg GAE/g dry weight (DW) in 2022 and 34.10 mg GAE/g DW in 2023.
Previous studies have shown that the composition and concentration of polyphenols in petals can vary widely due to several factors, including cultivar, season, and the geographical location [27,28]. Phenolic compounds are known for their antioxidant properties, attributed to their hydroxyl (OH) groups, which help in neutralizing free radicals [29]. In particular, the predominant phenolic acids found in all Borago officinalis extracts are rosmarinic acid, 3,4-dihydroxyphenylacetic acid, and ferulic acid [30].
Drought stress in plants often leads to the increased production of certain metabolites, including polyphenols, flavonoids, and essential oils [31]. The differences in total polyphenol content (TPC) detected across the populations in this study can be attributed to the distinct soil types and climatic conditions from which the populations originated.
Furthermore, [32] identified several key factors that influence the production of secondary metabolites. These include genetic factors, environmental conditions such as sunlight exposure, soil properties, and temperature, as well as conditions before and after the harvest. Additionally, gene expression patterns and the choice of solvent used for extraction may also impact the metabolite level. Collectively, these factors account for the variability in metabolite concentrations observed in the current study.
3.4. Total Flavonoids Content (TFC)
Flavonoids are crucial for various plant functions, including enhancing stress resilience, shielding against ultraviolet radiation, participating in signaling pathways, and defending against pathogens. Moreover, flavonoids, particularly in health-related products, are recognized for their potent antioxidant properties [33].
In the first year of the experiment, the total flavonoid content (TFC) across the studied populations varied significantly, ranging from 3.00 to 11.80 mg rutin per gram of DW, with statistical diference (p-value < 0.01). The Ghazvin and Roodsar populations exhibited the highest and lowest flavonoid concentration,respectively.
In the second year of experiment when he palnts were well-stablished, there was a notable increase in flavonoid content, with the Roodsar population reaching the highest level at 19.50 mg rutin per gram of DW, and the Leron population recorded the lowest at 17.20 mg rutin per gram of DW (Table 3). This observation suggests that flavonoid content tends to increase as plants mature. Supporting these findings, [7] reported elevated flavonoid concentrations in the petals of Echium amoenum with the age of the plants.
Among the primary flavonoids in Borago officinalis, compounds such as oleuropein, quercetin, apigenin, luteolin, and naringenin were prominently identified using LC-MS/MS [34].
3.5. Total Anthocyanins Content (TAC)
In both growing seasons, there were significant variations in the total anthocyanin content among the populations examined (p-value < 0.01).
In the initial year of the study, the Fasa population exhibited the highest level of anthocyanins, measuring 30.5 mg cyanidin equivalents per gram of dry weight (mg CyE g⁻¹ DW). This population was sourced from a location that was geographically close to the experimental site, which likely contributed to its high anthocyanin levels (Table 3).
Conversely, the Leron population, which was collected from the northern region of Iran, charactyerized for its high humidity, showed the lowest anthocyanin concentration at 15.30 mg CyE g⁻¹ DW (Table 3). This disparity underscores the influence of geographical and environmental factors on the anthocyanin content in different populations.
In the second year of the study, the Roodsar population displayed the highest concentration of anthocyanins, reaching 31.25 mg CyE g⁻¹ DWwhich was considerably higher compared to the first (esyablishing) year. On the other hand, the Leron population maintained the lowest anthocyanin concentration, which was 27.90 mg CyE g⁻¹ DW, reflecting a decrease from the first year.
Overall, the results suggest that Echium populations from similar geographic regions tend to have comparable anthocyanin concentrations. This implies that environmental factors specific to a region may contribute to the consistency of anthocyanin level within that area. Conversely, Echium plants cultivated across different geographical settings—characterized by variations in factors such as sunlight exposure, rainfall, temperature, soil moisture, and soil fertility—exhibit diverse anthocyanin profiles. This indicates that geographic and environmental conditions play a significant role in shaping the anthocyanin content in these plants.
3.6. Antioxidant Capacity (DPPH Radical Scavenging Activity)
Recent years have seen a growing interest from researchers and food manufacturers in the health benefits of functional foods that are rich in antioxidant polyphenols. These compounds are well-regarded for their potential in enhancing health and combat oxidative stress. Studies have highlighted the importance of these antioxidants, driving both academic research and commercial interest [35].
In this context, Echium has been recognized for its antioxidant properties. Many studies have documented its ability to neutralize free radicals, which contributes to its overall antioxidant activity. Research conducted by [36,37] has validated the efficacy of Echium in scavenging DPPH radicals, affirming its role as a valuable source of antioxidants in functional foods.
As illustrated in Table 3, during the initial year of the experiment, the Fasa population exhibited the highest antioxidant capacity, measured at 69.20%. This indicates that, in the first year, the Fasa population had the most significant ability to neutralize free radicals compared to other populations. However, by the second year, the Leron population was dominant in achieving the highest antioxidant capacity (70.90%).
These findings are consistent with previous research conducted by [36] which concluded that elevated antioxidant activity is closely linked to higher levels of total polyphenols, flavonoid content, and essential oils. Thus, the study reinforces the idea that the antioxidant effectiveness of a plant can be enhanced by increasing the concentrations of these specific compounds.
3.7. Multivariate Analysis of Antioxidants Attributes
In the analysis of antioxidant attributes, a comprehensive data matrix was created, comprising 18 data points derived from 6 different variables, each observed 3 times. This matrix was subjected to both cluster analysis and principal component analysis (PCA) to visualize and differentiate the various Echium wild populations based on their antioxidant properties and biochemical characteristics, as depicted in Figure 1.
The PCA revealed that the first two principal components were responsible for explaining 71.20% of the total variance observed across the data set. Specifically, the first principal component (F1) alone accounted for 48.40% of this variance, indicating its substantial role in differentiating the populations. In contrast, the second principal component contributed a smaller proportion, representing 22.80% of the total variance. This division of variance highlights the primary factors influencing the variability in antioxidant attributes among the studied Echium populations.
The cluster analysis divided the Echium wild populations into three distinct groups based on their antioxidant attributes and biochemical characteristics (Figure 1). The first group included the Ghazvin, Fasa, and Roodsar populations. The second group consisted solely of the Leron population. The third group was made up of the Sari and Vazneh Sar populations.
Principal Component Analysis (PCA) further clarified these groupings, indicating that the Ghazvin, Fasa, and Roodsar populations exhibited the highest levels of antioxidant properties, including phenols, anthocyanins, and flavonoids. Given their superior antioxidant attributes, these populations are recommended for use as natural sources of antioxidants and preservatives in the food industry, as suggested by [38].
3.8. Oil Content and Fatty Acids Profile
The oil percentage and fatty acid profiles among various Echium seed accessions showed considerable variability (p≤0.01). In the first year of the study, the oil content ranged from 16.50% to 22.40% (Table 3). However, this range increased in the following year, with oil content varied from 21.00% to 25.50%. Generally, the oil percentage recorded in this study was lower than those reported in previous research. For example [6] documented a wide range of oil content in seeds, from 6.2% in the E. italicum Kaleybar population to 33.8% in the E. amoenum Ramsar population. Similarly, [5] noted that the Rahim Abad population had the highest oil content at 36.14%, while the Roodbar population contained the lowest at 32.2%. These results, combined with the findings of the current study, underscore the significant variability in oil content among Echium seed accessions, highlighting the importance of this variation for selecting appropriate candidates for industrial purposes.
The combined use of Gas Chromatography-Mass Spectrometry (GC-MS) and validation with authentic standards via a Gas Chromatography-Flame Ionization Detection (GC-FID) device enabled the precise identification of seven fatty acids in the oil extracted from Echium seeds. The identified fatty acids and their respective concentrations were as follows: oleic acid, ranged from 18.35% to 19.40%; palmitic acid, ranging from 6.30% to 7.30%; linoleic acid, ranging from 23.80% to 26.20%; stearic acid, ranging from 4.10% to 4.80%; γ-linolenic acid, ranging from 4.70% to 5.00%; α-linolenic acid, ranging from 32.30% to 34.20%; and stearidonic acid, ranging from 4.60% to 5.40%. These fatty acids were characterized as the primary constituents in the oil extracted from different Echium populations (Table 4).
In the second year of the study, the fatty acid composition of Echium seed oil was observed with the following percentages: oleic acid 15.80% to 19.30%; palmitic acid 5.90% to 7.32%; linoleic acid 21.55% to 24.70%; stearic acid 4.50% to 4.97%; γ-linolenic acid 4.20% to 5.70%; α-linolenic acid 33.95% to 37.40%; and stearidonic acid 4.80% to 6.60%. These findings suggest that the fatty acid percentages remained relatively consistent between the first and second year of the experiment. Nevertheless, there was a slight increase in seed yield in the second year compared to the first. This indicates that as the plants matured, the oil percentage of the Echium seeds improved.
In both years of the study, α-linolenic acid was identified as the predominant fatty acid in the Echium seed oil profile, followed closely by linoleic acid. Specifically, the average concentration of α-linolenic acid across all six Echium accessions was 33.35% in the first year which increased to 36.30% in the second year (Table 4). The variation in α-linolenic acid content between the highest (Sari) and the lowest (Vazneh Sar) accessions was 1.84%. Linoleic acid was the second most prevalent fatty acid, with average percentages of 24.84% in the first year and 22.99% in the second year (Table 4). Notably, the Vazneh Sar accession exhibited the highest percentage of linoleic acid in both years.
[6] documented a notable presence of stearidonic acid in the seeds of Echium italicum, with the Boumehen and Kaleybar populations containing 10.47% and 12.45% stearidonic acid, respectively. In contrast, the current study found significantly lower levels of stearidonic acid, ranging from 4.59% to 6.59% across all six Echium accessions in both years. Additionally, they identified α-linolenic acid (ALA, 18:3ω3) and linoleic acid (LA, C18:2ω6) as the predominant unsaturated fatty acids in the oils, with the highest concentrations of 44.15% and 26.94%, respectively, observed in Echium amoenum seeds from the Hezar Jarib and Behshahr populations.
[5] reported that the highest concentrations of fatty acids in their study were linoleic and linolenic acids. Specifically, the Rahim Abad population contained 19.27% linoleic acid, while the Roodbar population had a notably higher concentration of linolenic acid at 40.53%.
The greatest proportion of gamma-linolenic acid (GLA) within the total fatty acids was detected in the species Borago morisiana and Borago pygmaea, with GLA constituting 23.1% and 22.9% of their total fatty acid content, respectively. [39] In contrast, other indigenous species of the Borago genus, specifically Borago trabutii and Borago longifolia, exhibited lower levels of GLA, comprising 18.9% and 19.0% of their total fatty acids, respectively [39].
The total unsaturated fatty acids (ΣUSFA) was more abundant than both the total saturated fatty acids (ΣSFA) and the polyunsaturated fatty acids (PUFAs) across all studied populations (Table 4). Specifically, Echium oil exhibited a higher concentration of polyunsaturated fatty acids, ranging from 4.20% to 37.40%, compared to monounsaturated fatty acids (ranging from 16.44% to 19.40%) in both years of the study.
Based on the values, Echium oil is considered a highly nutritious edible oil. Overall, it can be concluded that choosing superior genotypes from the Fasa population would be advantageous for industrial applications, as it has a higher total unsaturated fatty acid (ΣUSFA) level compared to other populations.
The principal component analysis (PCA) and cluster plots indicated considerable variations exist among Echium wild populations, regarding their fatty acid profiles (Figure 2). The analysis revealed that the first two principal components together explained 87% of the overall variance. Specifically, the first principal component (F1) accounted for 75.60% of the variance, while the second principal component (F2) contributed 11.40% to the total variance.
The cluster analysis identified two separate groups of Echium populations (Figure 2). The first group included the Ghazvin, Fasa, Roodsar, and Sari populations, which were clustered together based on their fatty acid profiles. The second group was comprised of the Leron and Vazneh Sar populations.
Further examination using principal component analysis (PCA) demonstrated that the Fasa and Sari populations were notably had higher levels of α-linolenic acid and γ-linolenic acid.
The findings of this study align with previous research suggesting a strong connection between fatty acid profiles and climatic conditions [40,10]. The notable variation in fatty acid content across different Echium populations can be attributed to a range of factors. These include genetic differences, variations in soil and climate conditions, environmental influences, as well as cultural practices such as harvesting, postharvest processing, and agricultural methods [5]. (Daneshfar et al., 2013). However, the influence of genetic factors on fatty acid profiles should be emphasized more than other contributing elements. Recent research utilizing genotyping by sequencing (GBS), a method within genome-wide association studies (GWAS), identified 53 single nucleotide polymorphisms (SNPs) linked to acyl-lipid biosynthesis pathways and 24 fatty acid traits in Brassica napus L. [41]. Additionally, in cotton, genes involved in fatty acid elongation and biosynthesis processes showed high expression levels from 20 days post-anthesis (DPA) to 30 DPA (Zhu et al., 2021). These findings suggest that genetic factors play a more significant role in shaping fatty acid profiles compared to climatic conditions.
The emphasis on genetic factors in determining fatty acid profiles is crucial, as they appear to have a more significant impact than other contributing elements. Recent advancements in genome-wide association studies (GWAS), specifically through genotyping by sequencing (GBS), have shed light on the genetic underpinnings of fatty acid composition. For instance, a study on Brassica napus L. identified 53 single nucleotide polymorphisms (SNPs) that are directly associated with acyl-lipid biosynthesis pathways and 24 distinct fatty acid traits [41]. This discovery underscores the substantial influence of genetic variation on the metabolic pathways that govern fatty acid synthesis. Moreover, research in cotton has provided additional evidence of the genetic regulation of fatty acid profiles. Genes involved in fatty acid elongation and biosynthesis exhibited significantly high expression levels during a critical developmental, specifically from 20 to 30 days post-anthesis (DPA) [42]. This period is pivotal for seed development and oil accumulation, highlighting the timing and specificity of genetic influence on fatty acid production.
Collectively, these findings suggest that genetic factors play a more decisive role in shaping fatty acid profiles compared to environmental influences such as climate. While environmental conditions can affect fatty acid composition, the strong association between specific genetic markers and fatty acid traits points to the predominance of genetic control. Thus, the genetic architecture of an organism is the key determinant of its fatty acid profile, overshadowing the role of external factors.
4. Conclusions
Echium is renowned for its medicinal properties and is extensively used across Middle Eastern and Mediterranean regions. This study highlights the considerable variations in both antioxidant properties and fatty acid profiles among different Echium populations collected from Iran. Analysis using GC-FID revealed that three primary fatty acids—α-linolenic acid, linoleic acid, and oleic acid—comprise over 70% of the oil's composition. Notably, α-linolenic acid emerged as the predominant fatty acid across Echium seed accessions.
The study classified the Echium germplasm into two distinct groups based on their predominant fatty acid compositions and oil content. Among these, the Fasa population exhibited the highest concentration of polyunsaturated fatty acids (ΣPUSFA), reaching up to 37.40%. This high level of polyunsaturated fatty acids underscores the potential of Fasa's oil as a valuable source of unsaturated fatty acids for the food industry. Additionally, the study reaffirmed that genetic factors play a more significant role in determining the fatty acid profiles and antioxidant properties of Echium than environmental conditions.
Authorship Contribution
Zohreh Emami Bistgani was responsible for the conceptualization, methodology, software, validation, formal analysis, writing original drafts and project administration. Vahid Rowshan Sarvestani handled the investigation, visualization and supervision. Masoud Hashemi participated in writing original drafts and project administration.
Acknowledgments
The authors would like to appreciate the cooperation of Shahid Fozveh Station staffs in Isfahan Agricultural and Natural Resources Research and Education,.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Mozaffarian, V. A pictorial dictionary of botany botanical taxonomy (Latin English French–Germany–Persian/complied). Farahang Moaser. 2008. [Google Scholar]
- Sayyah, M.; Boostani, H.; Pakseresht, S.; Malaieri, A. Efficacy of aqueous extract of Echium amoenum in treatment of obsessive-compulsive disorder. Prog Neuropsychopharmacol Biol Psychiatry. 2009, 33, 1513–1516. [Google Scholar] [CrossRef] [PubMed]
- Mehrabani, M.; Ghassemi, N.; Sajjadi, S.E.; Ghannadi, A.; Shams-Ardakani, M. Main phenolic compounds of petals of Echium amoenum fisch. and C.A. Mey., a famous medicinal plant of Iran. Daru. 2005, 13, 65–69. [Google Scholar]
- Ghasemi Pirbalouti, A. Medicinal plants used in Chaharmahal and Bakhtyari districts, Iran. Herba Pol. 2009, 55, 69–75. [Google Scholar]
- Daneshfar, E.; Alirezalu, K.; Ahmadi Hoseini, M.; Naghavi, M.R.; Omidbaigi, R. Evaluation of oil content, fatty acid composition and physicochemical characteristics of some of Echium amoenum Fisch. Accessions. Iranian Journal of Medicinal and Aromatic Plants (IJMAPR). 2013, 28, 700–708.
- Abbaszadeh, S.; Radjabian, T.; Taghizadeh, M. Identification and determination of phytosterols in oilseeds of some populations from two Iranian Echium species. Iranian Journal of Medicinal and Aromatic Plants (IJMAPR). 2013, 28, 742–755. [Google Scholar]
- Naderi Hagy Bagher Candy, M.; Rezaee, M.B. Primory phytochemical investigation of Echium amoenium. Iranian Journal of Medicinal and Aromatic Plants (IJMAPR). 2004, 20, 377–383. [Google Scholar]
- Mehrabani, M.; Ghannadi, A.; Sajadi, E.; Ghasemi, N.; Ardakani, M.R. Toxic pyrrolozidine alkaloids of Echium amoenum fisch. & C.A. Mey, DARU. 2006, 14, 122–127.
- Mojab, F.; Behfar, A.; Kobarfard, F.; Nick avar, B.; Jafari, B. Investigating the composition of fatty acids in the seeds of Echium amoenum. Mey.et Fisch. J. Med. Plant Res. 2008, 29, 80–87. [Google Scholar]
- Sun, C.; Jia, L.; Xi, B.; Wang, L.; Weng, X. Natural variation in fatty acid composition of Sapindus spp. seed oils. Ind. Crops Prod. 2017, 102, 97–104. [Google Scholar] [CrossRef]
- Nayebpour, N.; Asadi-Gharneh, H.A. Variability of fatty acids composition of wild sumac (Rhus coriaria L). fruit. J. Med. Plants 2019, 3, 118–129. [Google Scholar] [CrossRef]
- Fei, X.; Qi, Y.; Lei, Y.; Wang, S.; Hu, H.; Wei, A. Transcriptome and metabolite analysis reveals key genes for melanin synthesis during the development of Zanthoxylum bungeanum seeds. Ind. Crops Prod. 2021, 165, 113419. [Google Scholar] [CrossRef]
- Yaghmaei, L.; Soltani, S.; Khodagholi, M. Bioclimatic classification of Isfahan province using multivariate statistical methods. Int. J. Climatol. 2009, 29, 1850–1861. [Google Scholar] [CrossRef]
- Page, A.L. (Eds.),. Methods of Soil Analysis. Part 2-Chemical and Microbiological Properties, Second edition. 1982.
- Olsen, S.R.; Cole, C.V.; Watanabe, F.S.; Dean, L.A. Estimation of available phosphorus in soils by extraction with sodium bicarbonate. In: Banderis, A.D., Barter, D.H., Anderson, K. (Eds.), Agricultural and Advisor. U.S. Department of Agriculture Circular No. 1954, 939.
- Black, C.A. Methods of Soil Analysis, Part I-Physical and Mineralogical Properties Including Statistics of Measurement and Sampling. 1965.
- Lichtenthaler, H.K. Chlorophylls and carotenoids: pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar]
- Spanos, G.A.; Wrolstad, R.E. Influence of processing and storage on the phenolic composition of Thompson seedless grape juice. J. Agric. Food Chem. 1990, 38, 1565–1571. [Google Scholar] [CrossRef]
- Quettier-Deleu, C.; Gressier, B.; Vasseur, J.; Dine, T.; Brunet, J.; Luyck, M.; Cazin, M. , Cazin, J. C, Bailleul, F., Trotin, F. Phenolic compounds and antioxidant activities of buckwheat (Fagopyrum esculentum Moench). hulls and flour. J. Ethnopharmacol. 2000, 72, 35–40. [Google Scholar] [CrossRef]
- Wagner, G.J. Content and vacuole/extra vacuole distribution of neutral sugars free amino acids, and anthocyanins in protoplast. Plant. Physiol. 1979, 64, 88–93. [Google Scholar] [CrossRef]
- Wang, M.; Rangarajan, L.J.; Shao, M.; La Voie, E.J.; Huang, C.T.; Ho, C.T. Antioxidative phenolic compounds from sage (Salvia officinalis). J. Agric. Food Chem. 1998, 46, 4869–4873. [Google Scholar] [CrossRef]
- Rezig, L.; Chouaibi, M.; Msaada, K.; Hamdi, S. Chemical composition and profile characterisation of pumpkin (Cucurbita maxima) seed oil. Ind. Crops Prod. 2012, 37, 82–87. [Google Scholar] [CrossRef]
- ISO, 12966-4. Fatty acid methyl esters (FAME) in oil samples. Merck KGaA, Darmstadt, Germany. 2015.
- Kiani, M.; Alahdadi, I.; Soltani, E.; Boelt, B.; Benakashani, F. Variation of seed oil content, oil yield, and fatty acids profile in Iranian Nigella sativa L. landraces. Ind. Crops Prod. 2020, 2020. 149, 112367. [Google Scholar] [CrossRef]
- Wang, W.; Wei, L.; Li, H.; Xu, H.; Xu, Z.; Yan, C.; Wu, Y.; Ji, S.; Wang, T. Effects of sowing date on photosynthetic characteristics chlorophyll fluorescence and yield of different Echium plantagineum L. cultivars. Sci. Rep. 2023, 13, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Ghorbani, N.; Chamani, E.; Shokouhian, A.A.; Ramezanpour, S.S.; Soltanlou, H. Assessment of genetic diversity and photosynthetic pigments among wild populations of Yellow Flag (Iris pseudacorus). J. Plant Mol. Breed. 2021, 9, 1–11. [Google Scholar]
- Taamalli, A.; Arráez Román, D.; Gómez Caravaca, A.M.; Zarrouk, M.; Segura Carretero, A. Geographical characterization of Tunisian olive tree leaves (cv. Chemlali) using HPLC-ESI-TOF and IT/MS fingerprinting with hierarchical cluster analysis. J. Anal. Chem. 2018, 6789704, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Lukić, I.; Pasković, I.; Žurga, P.; Majetić Germek, V.; Brkljača, M.; Marcelić, Š.; Ban, D.; Grozić, K.; Lukić, M.; Užila, Z. Determination of the variability of biophenols and mineral nutrients in olive leaves with respect to cultivar, collection period and geographical location for their targeted and well-timed exploitation. Plants. 2020, 9, 1–20. [Google Scholar] [CrossRef]
- Granato, D.; Shahidi, F.; Wrolstad, R.; Kilmartin, P.; Melton, L.D.; Hidalgo, F.J.; … Finglas, P. Antioxidant activity, total phenolics and flavonoids contents: Should we ban in vitro screening methods? Food Chem. 2018, 264, 471–475. [Google Scholar] [CrossRef]
- Michalak, M.; Zagórska-Dziok, M.; Klimek-Szczykutowicz, M.; Szopa, A. , Phenolic profile and comparison of the antioxidant, anti-Ageing, anti-Inflammatory, and protective activities of Borago officinalis extracts on skin cells. Molecules. 2021, 28, 1–19. [Google Scholar] [CrossRef]
- Ahmadi, H.; Babalar, M.; Sarcheshmeh, M.A.A.; Morshedloo, M.R.; Shokrpour, M. Effects of exogenous application of citrulline on prolonged water stress damages in hyssop (Hyssopus officinalis L.): Antioxidant activity, biochemical indices, and essential oils profile. Food Chem 2020, 333, 127433. [Google Scholar] [CrossRef]
- Fereidoonfar, H.; Salehi-Arjmand, H.; Khadivi, A.; Akramian Safdari, L. Chemical variation and antioxidant capacity of sumac (Rhus coriaria L.). Ind. Crops Prod. 2019, 139, 111518. [Google Scholar] [CrossRef]
- Agati, G.; Brunetti, C.; Fini, A.; Gori, A.; Guidi, L.; Landi, M.; Tattini, M. Are flavonoids effective antioxidants in plants? Twenty years of our investigation. Antioxidants. 2020, 9, 1098. [Google Scholar]
- Zemmouri, H.; Ammar, S.; Boumendjel, A.; Messarah, M.; El Feki, A.; Bouaziz, M. Chemical composition and antioxidant activity of Borago officinalis L. leaf extract growing in Algeria. Arab. J. Chem. 2019, 12, 1954–1963. [Google Scholar] [CrossRef]
- Mumivand, H.; Babalar, M.; Tabrizi, L.; Craker, L.; Shokrpour, M.; Hadian, J. Antioxidant properties and principal phenolic phytochemicals of Iranian tarragon (Artemisia dracunculus L.) accessions. HORTIC ENVIRON BIOTE. 2017, 58, 414–422. [Google Scholar] [CrossRef]
- Zannou, O.; Pashazadeh, H.; Ghellam, M.; Ibrahim, S.A.; Koca, I. Extraction of Anthocyanins from Borage (Echium amoenum) flowers using choline chloride and a glycerol-based, deep eutectic solvent: Optimization, antioxidant activity, and in vitro bioavailability. Molecules. 2022, 27, 134. [Google Scholar] [CrossRef] [PubMed]
- Ranjbar, A.; Khorami, S.; Safarabadi, M.; Shahmoradi, A.; Malekirad, A.A.; Vakilian, K.; Mandegary, A.; Abdollahi, M. Antioxidant activity of Iranian Echium amoenum Fisch & C.A. Mey flower decoction in humans: A cross-sectional before/after clinical trial Evid. -Based Complement. J Evid Based Complementary Altern Med. 2006, 3, 469–473. [Google Scholar]
- Sakhr, K.; Sami, E. Physiochemical properties and medicinal, nutritional and industrial applications of Lebanese Sumac (Syrian Sumac-Rhus coriaria): A review. Heliyon. 2020, 6, e03207. [Google Scholar] [CrossRef]
- Guil-Guerreroa, J.L.; Gómez-Mercadob, F.; Ramos-Buenoai, R.P.; González-Fernándeza, M.J.; Urrestarazucii, J.B.; Bélair, G. Fatty acid profiles and sn-2 fatty acid distribution of γ-linolenic acid-rich Borago species. J. Food Compos. Anal. 2017, 66, 74–80. [Google Scholar] [CrossRef]
- Tsegay, G.; Redi-Abshiro, M.; Chandravanshi, B.S.; Ele, E.; Mohammed, A.M.; Mamo, H. Effect of altitude of coffee plants on the composition of fatty acids of green coffee beans. BMC Chemistry. 2020, 14, 1–11. [Google Scholar] [CrossRef]
- Gazave, E.; Tassone, E.E.; Baseggio, M.; Cyder, M.; Byriel, K.; Oblath, E.; … Pauli, D. Genome-wide association study identifies acyl lipid metabolism candidate genes involved in the genetic control of natural variation for seed fatty acid traits in Brassica napus L. Ind. Crops Prod. 2020, 145, 1–12. [Google Scholar] [CrossRef]
- Zhu, D.; Le, Y.; Zhang, R.; Li, X.; Lin, Z. . A global survey of the gene network and key genes for oil accumulation in cultivated tetraploid cottons. Plant Biotechnol. 2021, 19, 1170–1182. [Google Scholar] [CrossRef]
Figure 1.
Hierarchical cluster analysis (HCA)) and principal component analysis (PCA) classify Echium populations based on their total anthocyanin content, chlorophyll a, chlorophyll b, carotenoids levels, antioxidant characteristics, total phenol, and total flavonoids content.
Figure 1.
Hierarchical cluster analysis (HCA)) and principal component analysis (PCA) classify Echium populations based on their total anthocyanin content, chlorophyll a, chlorophyll b, carotenoids levels, antioxidant characteristics, total phenol, and total flavonoids content.
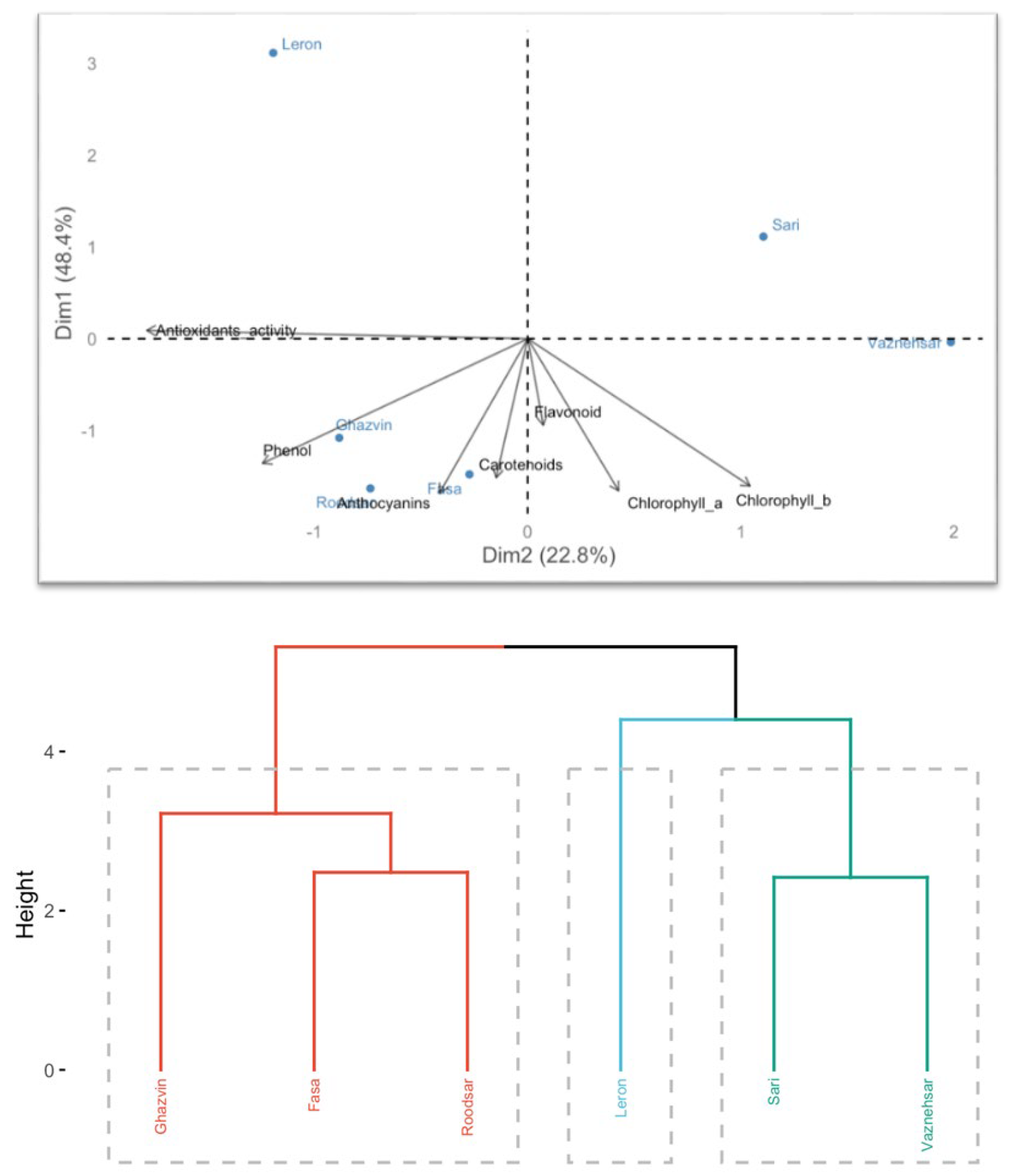
Figure 2.
Hierarchical clustering and principal component analysis (PCA) classify Echium seed populations based on their oil and fatty acids profile.
Figure 2.
Hierarchical clustering and principal component analysis (PCA) classify Echium seed populations based on their oil and fatty acids profile.
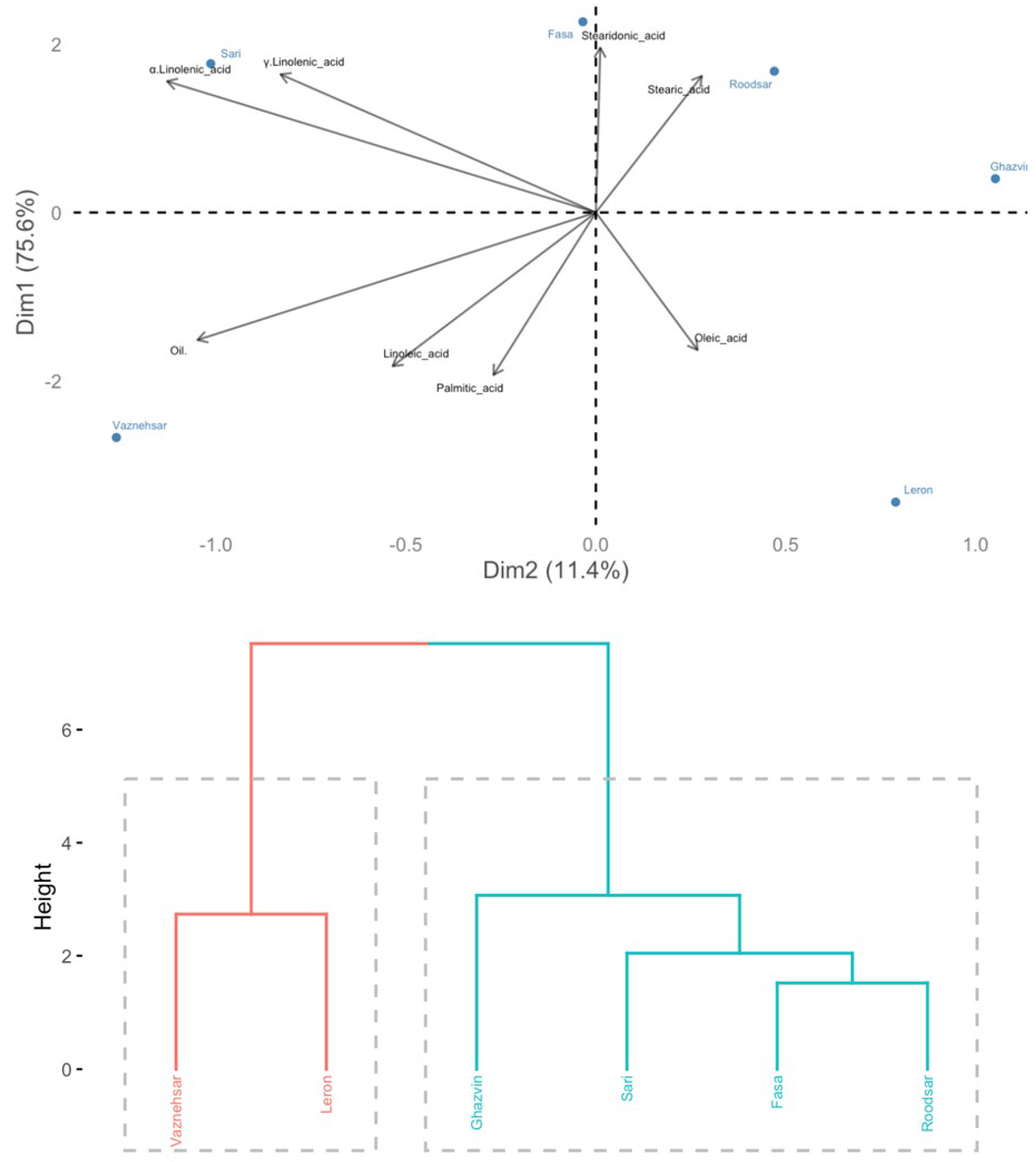

Table 1.
Origin, habitats and geographical characteristics of sumac population.
| N | Area of sampling collection | Populations | Longitude | Latitude | Altitude (m) |
Mean annual rainfall (mm) |
Mean annual temperature (◦C) |
Habitat |
| 1 | Fars Province | Fasa | 53°38′ | 28° 56′ | 1370 | 380 | 20 | Arid and Semi-arid |
| 2 | Gilan Province | Roodsar | ′ 28°50 | 37°13′ | -19 | 1178 | 15.8 | Mountain-Humid |
| 3 | Mazandaran Province | Sari | 5° 53′ | 36°39′ | 11 | 790 | 15.5 | Mountain-Humid |
| 4 | Ghazvin Province | Ghazvin | 57′ °49 | 36°18′ | 1314 | 318 | 14 | Mountain-Cold temperate |
| 5 | Gilan Province | Vazneh sar | 48°90′ | 37°80′ | 54 | 1300 | 16 | Mountain-Humid |
| 6 | Gilan Province | Leron | 48°80′ | 37°40′ | 60 | 1200 | 17 | Mountain-Humid |
Table 2.
The effect of accessions on morphological characteristics in 2022 and 2023.
| First Year | |||||||||||
| Seed weight per plant | 1000-seed weight | Leaf width | Leaf length | Number of Lateral Branch | Flower number | Canopy diameter | Plant Height | Accession names | Accessions number | ||
| 3.00±0.72b | 1.30±0.17a | 2.10±0.76ab | 10.60±0.60b | 88.30±31.75a | 311.70±155.4a | 53.30±5.03a | 38.30±7.57a | Fasa | 1 | ||
| 4.37±0.94ab | 1.20±0.25ab | 2.60±0.12ab | 6.90±0.40c | 55.70±13.00ab | 196.70±56.19c | 48.30±7.63ab | 44.30±9.07a | Roodsar | 2 | ||
| 3.24±1.30b | 1.20±0.09ab | 3.00±0.05a | 4.80±0.15d | 64.30±35.70ab | 211.70±73.20b | 46.00±6.92abc | 48.00±18.35a | Ghazvin | 3 | ||
| 6.00±1.86a | 1.00±0.09bc | 2.40±1.24ab | 6.00d±0.25c | 37.70±26.80bc | 175.70±111.36d | 50.00±0.01ab | 44.30±6.02a | Sari | 4 | ||
| 0.87±0.04c | 0.70±0.04c | 2.95±1.13a | 9.506±1.28b | 10.70±3.54c | 31.70±18.92c | 32.00±3.78c | 31.70±2.88a | Vazneh | 5 | ||
| 0.52±0.08c | 0.70±0.06c | 1.80±0.47b | 12.80±1.25a | 13.70±5.85c | 29.00±7.93f | 29.00±9cd | 30.50±0.57a | Leron | 6 | ||
| Second year | |||||||||||
| 3.83±0.27b | 4.50±0.50d | 2.70±1.07a | 11.50±0.50b | 181.30±7.63a | 373.70±25.10a | 55.15±0.53a | 67.70±2.51a | Fasa | 1 | ||
| 4.80±1.00ab | 10.30±1.52a | 2.95±0.05a | 7.60±0.52c | 79.00±11.01c | 251.70±20.81c | 53.00±0.68a | 50.00±0.87c | Roodsar | 2 | ||
| 3.70±1.82b | 3.60±0.52d | 3.43±0.51a | 5.30±0.57d | 152.00±7.54b | 315.30±13.61b | 48.34±0.68b | 49.80±4.64c | Ghazvin | 3 | ||
| 6.50±2.36a | 6.90±1.12b | 2.90±1.15a | 6.50±0.51cd | 161.00±15.09b | 321.30±20.00b | 51.54±0.41ab | 59.00±7.00b | Sari | 4 | ||
| 0.86±0.11c | 4.83±0.76cd | 3.50±1.57a | 10.20±1.58b | 23.16±10.61d | 53.30±15.27d | 33.00±0.59c | 41.60±0.34d | Vazneh sar | 5 | ||
| 0.64±0.20c | 6.50±0.50bc | 2.23±0.49a | 13.70±1.52a | 27.33±15.37d | 26.70±2.88d | 23.00±5.56d | 28.90±0.62e | Leron | 6 | ||
Values with different letters are statistically different at p≤0.05.
Table 3.
The effect of accessions on biochemical characteristics in 2022 and 2023.
| First Year | |||||||||
| Oil (%) |
Antioxidant activity (%) | Anthocyanins (mg g-1) |
Flavonoids (mg g-1) |
Phenol (mg g-1) |
Carotenoids (mg g-1) |
Chb (mg g-1) |
Cha (mg g-1) |
Accession names | Accessions number |
| 17.60±0.17 | 69.20±0.76a | 30.50±1.32a | 4.90±0.60c | 18.90±0.31b | 1.50±0.07b | 2.80±0.13d | 3.40±0.20c | Fasa | 1 |
| 16.20±0.06 | 58.20±1.04d | 30.10±0.76a | 3.00±1.19d | 31.25±1.91a | 2.30±0.10b | 3.50±0.37c | 4.40±0.26bc | Roodsar | 2 |
| 17.70±0.05 | 66.80±2.23b | 24.70±2.08b | 11.80±0.66a | 32.90±1.71a | 4.80±1.49a | 6.30±0.64a | 7.20±1.71a | Ghazvin | 3 |
| 16.50±0.1 | 61.00±1.11c | 24.30±2.08b | 5.80±0.58c | 8.00±0.58d | 3.80±1.48a | 2.70±0.23d | 4.00±1.25c | Sari | 4 |
| 20.60±0.09 | 51.10±1.31e | 19.00±1.00c | 7.60±0.44b | 8.80±0.38d | 4.60±0.46a | 2.60±0.41d | 4.80±1.38bc | Vazneh sar | 5 |
| 22.40±0.10 | 60.60d±1.80c | 15.30±1.52d | 3.90±0.45d | 11.80±0.68c | 4.60±0.36a | 4.50±0.40b | 5.77±1.23b | Leron | 6 |
| Second year | |||||||||
| 22.65±0.17 | 61.60±0.24c | 30.80±0.11b | 19.30±0.26a | 33.40±0.17b | 5.80±0.22b | 7.60±0.20a | 15.60±1.62a | Fasa | 1 |
| 21.50±0.13 | 63.60±0.56b | 31.25±0.17a | 19.50±0.20a | 33.00±0.13bc | 6.80±0.13a | 5.90±0.41b | 10.70±0.59b | Roodsar | 2 |
| 22.20±0.12 | 52.00±0.28f | 28.40±0.24d | 18.20±0.08c | 34.10±0.21a | 2.20±0.12e | 6.80±0.11ab | 7.50±0.25 c | Ghazvin | 3 |
| 25.50±0.12 | 58.10±0.63d | 29.30±0.24c | 18.60±0.11b | 32.70±0.46c | 3.20±0.17d | 2.80±0.09c | 6.00±0.18d | Sari | 4 |
| 24.00±0.21 | 54.90±2.25e | 28.20±0.21de | 17.30±0.21d | 32.87±0.53bc | 3.60±0.03c | 7.40±2.08ab | 10.84±0.26b | Vazneh sar | 5 |
| 21.00±1.0 | 70.90±0.59a | 27.90±0.04e | 17.20±0.18d | 33.62±0.18c | 1.80±0.21f | 1.40±0.14c | 3.20±0.11e | Leron | 6 |
Values with different letters are statistically different at p≤0.05.
Table 4.
The effect of accessions on fatty acid profile in 2022 and 2023.
| Stearidonic acid. (C18:4) |
α-Linolenic acid (C18:3) |
γ-Linolenic acid (C18:3) |
Linoleic acid (C18:2) |
Oleic acid (C18:0) |
Stearic acid (C18:0) |
Palmitic acid (C16:0) |
Accession names | Accessions number | ||
| 5.40±1.02a | 33.92±1.04b | 5.00±0.10a | 23.80±0.15d | 19.30±0.16ab | 4.60±0.13b | 6.30±0.10cde | Fasa | 1 | ||
| 5.00±1.04b | 33.90±0.97b | 4.80±0.07c | 24.30±0.11cd | 18.80±0.13c | 4.70±0.06ab | 6.60±0.05cd | Roodsar | 2 | ||
| 5.00±0.89b | 33.20±1.50c | 4.94±0.06b | 24.40±0.20cd | 19.40±0.13a | 4.80±0.07a | 6.80±0.06c | Ghazvin | 3 | ||
| 5.00±1.03b | 34.20±1.00a | 4.90±0.13b | 24.90±0.07c | 18.35±0.13cd | 4.70±0.07ab | 6.60±0.02cd | Sari | 4 | ||
| 4.60±0.95d | 32.30±1.10de | 4.70±0.07cd | 26.20±0.09a | 19.10±0.13ab | 4.25±0.02bc | 7.30±0.08a | Vazneh sar | 5 | ||
| 4.90±0.74c | 32.70±1.09d | 4.90±0.11b | 25.50±0.09b | 19.20±0.08ab | 4.10±0.09bcd | 7.00±0.10ab | Leron | 6 | ||
| 6.80±0.21a | 37.40±0.11a | 5.70±0.23a | 21.55±0.54e | 16.44±0.12d | 4.60±0.05ab | 5.70±0.18cd | Fasa | 1 | ||
| 6.40±0.15ab | 36.95±0.05b | 5.50±0.08bc | 22.20±0.17de | 16.55±0.11d | 4.50±0.13b | 5.90±0.10c | Roodsar | 2 | ||
| 5.60±0.05c | 34.92±0.05c | 5.40±0.12bcd | 22.69±0.13c | 17.94±0.10c | 4.97±0.34a | 6.10±0.15b | Ghazvin | 3 | ||
| 6.60±0.33ab | 37.30±0.06ab | 5.65±0.07ab | 22.30±0.07d | 15.80±0.07e | 4.52±0.30b | 5.90±0.08c | Sari | 4 | ||
| 5.00±0.03cd | 37.30±0.03ab | 5.65±0.20ab | 24.50±0.15ab | 19.30±0.16a | 4.64±0.10ab | 7.30±0.20a | Vazneh sar | 5 | ||
| 4.80±0.14de | 33.95±0.10d | 4.20±0.06de | 24.70±0.32a | 18.30±0.19b | 4.48±0.14bc | 7.30±0.07a | Leron | 6 | ||
Values with different letters are statistically different at p≤0.05.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



