Submitted:
23 September 2024
Posted:
24 September 2024
You are already at the latest version
Abstract
Organisms from the genus Streptomyces feature actinobacteria with complex developmental cycle and great ability to produce a variety of natural products, which is possible due to complicated crosstalk between primary and secondary metabolism. These soil bacteria produce more than 2/3 of antibiotics used in medicine, and a large variety of bioactive compounds for industry and agricultural use. Although Streptomyces spp. have been studied for decades, the engineering of these bacteria remains challenging, and available genetic tools rather limited. Recent advancements in genetic manipulation of Streptomyces involving proposal of CRISPR/Cas9-based workflows as well as synthetic components (e.g. promoters, ribosome-binding sites, terminators, reporter genes) allowed to facilitate the turnaround time of strain engineering, but still has strain-specific limitations. However, a new perspective offered by synthetic biology to exploit the potential of existing and novel pathways in primary and secondary metabolism allows combining of different biosynthetic steps originating from diverse bacteria using a limited toolbox. Manipulation of interplay between primary and secondary metabolism proposes strategies to overcome strain-specific difficulties in engineering, leveraging insights in Streptomyces-specific physiological features. In this review, developments of these approaches for Streptomyces engineering are discussed and an overview of the synthetic biology developments is provided.
Keywords:
synthetic biology
; Streptomyces
; biotechnology
; primary metabolism
; secondary metabolism
1. Streptomyces Complex Metabolism—A Great Source of Valuable Targets for Engineering
1.1. Microbial Diversity as a Basis for Metabolic Diversity
Prokaryotes form the basis of the biosphere and many metabolic physiological technical processes (e.g., chemolithotrophic growth, specific fermentations, nitrogen fixation, methane formation, anoxygenic photosynthesis, secondary metabolite production as antibiotics or toxins, etc.) are limited to bacteria. Without prokaryotic organisms it would not be possible to completely break down organic substances into inorganic ones. Prokaryotes occur wherever physical and chemical conditions allow the existence of life; this includes even extreme conditions such as hot springs, the deep sea and Antarctic lakes, which can function under unusual conditions. For example, a heat-stable DNA polymerase from the thermophilic bacterium Thermus aquaticus [1]. Furthermore, prokaryotes possess a much larger genetic potential than any other group of organisms on Earth – as an example the genome of Mycobacterium tuberculosis features 376 unique proteins out of a total of 3995 putative ones (approx. 9.4%) [2], which do not show similarities to any known proteins in other organisms. In combination with the above-mentioned advantages, prokaryotes became an important source in the search for new biocatalysts and natural products. The search for certain metabolic performance usually first leads to the natural habitat of the organism involved. The occurrence of certain nutrients represents ecological niches for organisms that have specialized in the utilization of these molecules in the course of evolution. It determines nutrient supply for cells in the respective ecological niche and therefore colonization by certain species of microorganisms that can utilize these nutrients [3].
1.2. Environment of Streptomyces, Biological Natural Product Producers
In the environment, bacteria live together in complex communities, which comprise other microorganisms such as bacteria and fungi, but also higher organisms such as animals and plants that need to be interacted with [4]. This interaction essentially leads to competition, symbiosis, pathogenicity, or commensalism, involving the absorption of nutrients and survival. It depends on the type of interaction, whether it is one-sided or mutual benefit [5]. Microbial community formation is influenced by environmental conditions, physiological stress and the interaction of various species [6]. With 104 to 107 bacteria/g, organisms from the genus Streptomyces, phylum Actinobacteia, are one of main representatives in soil [7].
Actinomycetes have adapted to a large variety of ecological niches including terrestrial, intracellular, aquatic, artificial, and others. Many species, especially from the genus Streptomyces, are capable of polymer decomposition of dead plant, fungal and animal material. This biological function is a contribution to the recycling of biomaterials, global carbon, nitrogen and phosphate cycle as well as soil organic matter formation. Furthermore, representatives of Actinomycetes (e.g., from the genus Frankia) have nitrogen-fixing functions in symbiotic associations with plants or other organisms like insects [8].
In general, due to the high abundance and diversity of their metabolites, bacteria are the most common terrestrial and marine symbionts of eukaryotes [9]. One of such well-studied symbioses is between streptomycetes and fungi. For instance, interaction between Streptomyces hygroscopicus and Aspergillus nidulans is an inducer of secondary metabolite production on the part of the fungus. Such interaction requires direct physical contact between the two partners, which is a signal for communication [6].
One of the best-known symbioses between streptomycetes and higher organisms is between the leaf-cutting ant Acromyrmex echinatior and various Streptomyces spp. [10]. The leaf-cutter ant cultivates the forage fungus Leucoagaricus gongylophorus in a so-called mushroom garden. To protect it from infection by pathogenic fungi from genera Syncephalastrum, Escarvopsis, Fusarium and Trichoderma, the ant carries the antibiotic producing streptomycetes as a biofilm on their integument. Ants distribute it in the nest and thus prevent the spread of the harmful fungus. The streptomycete in return, presumably receives nutrients [9]. The solitary digger wasp of the genus Philanthus takes a similar precaution. It carries streptomycetes as endosymbionts in its antenna glands and protects their larvae by storing Streptomyces spp. in the brood cells infestation with a pathogenic fungus [11]. Another example is a complex interaction between plants, bacteria, and fungi in the rhizosphere. The rhizosphere refers to area narrow zone surrounding plant roots influenced by root exudates and associated microorganisms [12]. Root exudates consist of amino acids, plant secondary metabolites, organic acids, phenols, sugars, proteins and/or oligosaccharides, leading to increased number of bacterial species due to the increased supply of nutrients in the rhizosphere. For example, 7% of the total bacterial number in the wheat rhizosphere are representatives of the phylum Actinobacteria [13]. Over 70% of land plants form symbiotic interactions with arbuscular mycorrhizal fungi. Mycorrhizal fungi are key components of the plant microbiota and are important organisms in terrestrial ecosystems. There they facilitate the exchange of carbon (C) and minerals. Arbuscular mycorrhizal fungi can interact with microorganisms in the rhizosphere and are influenced by them [14]. Such interaction can be synergistic or antagonistic. In the synergistic interaction Streptomyces spp. may stimulate mycorrhiza formation [15]. Both fungus and bacteria can work equally positively on the plant by providing water and nutrient uptake. In return, the fungus and streptomycete receive carbohydrates from the plant and/or from hyphal exudates. Antagonistic interaction is provided by either the streptomycete or the mycorrhizal fungus for better plant growth, but both can inhibit each other’s development. Streptomyces has been reported to be a keystone shaping the bacterial community structure at the surface of arbuscular mycorrhizal fungal hyphae [16]. Other rhizobacteria can positively influence mycorrhiza formation too, by promoting the susceptibility of the roots, root-fungus recognition, or fungal germination [15].
Streptomyces spp. can influence other interaction partners in the rhizosphere by producing antibiotics, for example inhibiting plant pathogens or breaking down harmful substances. Streptomycetes can secrete the plant growth hormone auxin having a direct effect on plant growth [9]. Streptomyces are known to influence complex systems like plants and fungi. For instance, Streptomyces sp. AcM29 inhibits gram-positive bacteria, producing the siderophores ferulic acid and desferrioxamine B and showing a negative influence on the mycorrhizal fungi Amanita muscaria, Hebeloma cylindrosporum and Laccaria bicolor, while plant pathogenic fungi are not inhibited. Streptomyces sp. GB4-2 promotes the growth of the plant-pathogenic fungus Heterobasidion abietinum, but at the same time reduces the plant’s defense mechanisms. Furthermore, produced by the strain piceamycin is effective against gram-positive bacteria, and can detoxify the phytotoxin fomannoxin [17].
In addition to positive interactions that promote plant health, there is also a few negative influences that Streptomyces can cause on plants. A small number of streptomycetes are known to cause damage to potatoes, sweet potatoes, carrots, radishes, tomatoes, and eggplants [18,19]. An example of a pathogen among Streptomyces spp. is Streptomyces scabies, which targets potato tubers and can cause lesions on the tuber surface and subsequent potato blight solves with the help of the phytotoxin thaxtomin [19].
The interaction of Streptomyces with other bacteria can, like interactions with fungi also, lead to the induction or increase of secondary metabolite biosynthesis. A predatory bacterium that lives on Streptomyces - Myxococcus xanthus - can induce actinorhodin production during aerial mycelium development in Streptomyces coelicolor. Such inductions could also be caused by other bacteria, such as Bacillus subtilis [20]. Other bacteria, such as Amycolatopsis spp. AA4 or Streptomyces venezuelae, can influence the prodigiosin or siderophore biosynthesis in Streptomyces coelicolor [20].
1.3. Primary Metabolism in Streptomyces as an Important Part of Secondary Metabolism
Streptomyces belong to the bacteria group of actinomycetes, phylum Actinobacteria, and are immobile, strictly aerobic, gram-positive eubacteria, which feature a mycelium-like growth and intensive secondary metabolites production. Streptomyces spp. are ubiquitous in the soil. Only very few Streptomyces species are pathogenic to humans and animals, some can cause disease in plants. Bacteria of the genus Streptomyces are characterized by a very complex life cycle with different stages of development. Starting from a spore these bacteria form first the substrate mycelium consisting of fine, partially septate hyphae with a diameter of approx. 0.5-1.5 µm. These hyphae branch out into individual compartments usually containing several copies of the chromosome. Triggered by physiological stress, such as lack of nutrition, but also through chemical signaling substances, the aerial mycelium is created. During the growth of the aerial mycelium, many cells of the substrate mycelium are broken down, following the build-up of the aerial mycelium, in which afterwards spore formation and secondary metabolites biosynthesis takes place under strict regulatory control [21].
Streptomycete genomes have a very high GC content of over 70% [22]. The linear chromosome usually has the size of over 8 Mb and is among the largest known genomes in the bacterial kingdom [22]. The size of the genome reflects the metabolic diversity of Streptomyces spp. In addition to a wide variety of common substrates such as sugar, alcohols, amino acids, Streptomycetes can utilize many natural substances that are difficult to decompose such as cellulose and chitin. This is possible due to production of hydrolytic enzymes such as amylases, lipases, proteases, cellulases, ligninases and chitinases [23].
Actinobacteria from the genus Streptomyces live in their natural habitat soil under varied nutrient conditions. They can utilize diverse C-sources, such as monosaccharides, polyols, disaccharides, amino acids, sugar alcohols, deoxy sugars, glycosides, dicarboxylic acids, ketoacids [23,24] as well as amines like ethanolamine and polyamines [25]. The optimal carbon source for Streptomyces is glucose. Selection of a preferred carbon source is a major determining factor in the microbial growth rate and competition with other microorganisms in environment [26]. Under high concentrations glucose interferes with the formation of diverse compounds in the primary and secondary metabolism. The mechanism of carbon catabolite repression allows cells to regulate levels of carbon sources in cells, repress protein synthesis under presence of a catabolite generated from an exogenous carbon source, and to protect themselves against wasting the protein-synthesizing machinery [27,28]. Carbon catabolite repression allows regulation of genes for utilization of glycerol, arabinose, xylose, fructose and galactose, as well as extracellular polysaccharide-degrading enzymes like chitinase and amylase [27]. Overall, availability of carbon sources greatly impacts morphological development and production of secondary metabolites in Streptomyces [23,24,25,26,27,28].
Streptomyces can utilize various compounds as N-sources, which usually requires use of specialized enzymes for utilization of such nitrogen-containing compounds as ammonium, nitrate urea, amino acids, amino sugars as well as monoamines and polyamines [23,25]. Key enzymes in nitrogen assimilation are the glutamine synthetase (GS) and glutamine-2-oxoglutarate-aminotransferase (GOGAT) that fulfill the same function in ammonium assimilation as GSs and GOGAT in E. coli: their function is to convert ammonium and α–Ketoglutarate to glutamate using ATP and NADPH. GS is active under conditions of nitrogen deficiency (<0.1 mM) having a high substrate affinity and catalyzes condensation of ammonium and glutamate synthesizing glutamine under ATP consumption. GOGAT catalyzes conversion of glutamine to glutamate from glutamine and 2-oxyglutarate. At high ammonium concentration, glutamate dehydrogenase (GDH) catalyzes the conversion of ammonium and α-ketoglutarate to glutamate (reductive amination) and the reverse reaction having low substrate affinity [29].
Regulation of the nitrogen metabolism in Streptomyces is carried out at the transcriptional and post-translational levels, being controlled by proteins GlnR, GlnRII, AmtR, NnaR, Crp, AsfR. The transcription of glnA and glnII as well as the glnA2 gene in S. coelicolor is regulated by GlnR or GlnRII [25,30]. The post-translational regulation involves the signaling protein PII and modulation of the activity of key enzymes GDH and GS by adenylation/uridylation. GSI is regulated via adenylation and deadenylation by GlnE [31]. Another type of glutamine synthetase in Streptomyces is GSII (GlnII) that was first described in Rhizobium – it is a heat-labile octamer and has sequence similarity to eukaryotic GSs. According to recent studies, GSII occurred at an early stage of evolution from gamma-Proteobacteria via horizontal gene transfer into the photosynthesizing eukaryotes Chloroplastida [32]. GSII was found in many members of Actinomycetales, but it is missing in Amycolatopsis, Mycobacterium and Corynebacterium [33]. GSII lacks post-translational regulation through adenylation [34]. In addition to glutamine synthetases, Streptomyces spp. possess a set of GS-like proteins: GlnA2, GlnA3 and GlnA4 have been described in Actinomycetales, e.g., in S. coelicolor [25] and in M. tuberculosis [35].
Another essential nutrient element for metabolism in Actinobacteria is phosphate. It triggers the production of secondary metabolites [36]. Phosphorous is present in nature in the form of phosphate salts, organophosphates, and phosphonates. In the Streptomyces spp. the phosphate binding protein PstS is attached to the outer side of the cell membrane and glycosylated. Phosphate can be transported by the PstSCAB system, which is mediated by binding phosphorylated PhoP to the promoter region of the PstSCAB operon [36,37].
In many Streptomyces sp. gene clusters pitH1-pstSCAB-ppk, are linked to phosphate metabolism. The ppk gene encodes an enzyme catalyzing the reversible polymerization of the gamma phosphate of ATP into polyphosphate. It was demonstrated to play a negative role in the control of antibiotic biosynthesis in Streptomyces [38]. Streptomyces were reported to be able to degrade simple phosphonates. Phosphate control in Streptomyces is mediated by the two-component system (TCS) PhoR-PhoP, which orthologs have been found in most Actinobacteria [36,37]. It belongs to class IIIA of TCSs with the PhoR protein as a sensor kinase. PhoP belongs to the OmpR family of DNA-binding response regulators PhoR-PhoP control the primary metabolism and secondary metabolism, which has been in S. coelicolor, S. lividans, S. tsukubaensis, S. natalensis, S. avermitilis and some others [36,37,38].
Sulfur is a common compound in the environment and is one of essential elements for bacterial metabolism. It is used in the biosynthesis of the amino acids, such as cysteine and methionine, which are incorporated into other biomolecules. Sulfur plays an essential role in a variety of enzyme cofactors, such as coenzyme A, coenzyme M, thiamine, biotin and lipoic acid. Sulfur is and is critical in redox processes. For biosynthetic processes, sulfur is derived from assimilation of inorganic sulfate. In bacteria, sulfur amino acid (SAA) metabolism is adaptive and diversified [39]. In Streptomyces spp., biosynthesis of cysteine and methionine via homocysteine (Hcy) is linked through transsulfuration pathways, whereas cysteine is the key intermediate in most pathways of sulfur metabolism. After the inorganic sulfate is transported into the cell, it is incorporated into the sulfate assimilation pathway, and sulfur is converted to 3′-phosphoadenosine-5′-phosphosulfate, subsequently being reduced to sulfite and to sulfide. Afterwards, sulfide is transferred onto an organic moiety to generate molecules of cysteine. Inter-conversion between homocysteine and methionine occurs in the activated methyl cycle (AMC), which involves ATP and additional cofactors cobalamin and 5-methyl-tetrahydrofolate. In Streptomyces spp. a direct sulfhydrylation pathway involving metX and metY has been found (Figure 1) [40].
1.4. Streptomyces as Producers of Secondary Metabolites
Actinomycetes have gained attention mainly because of their biosynthetic potential to produce diverse medically and industrially relevant secondary metabolites. The discovery of the first antibiotic derived from Actinomycetes, actinomycin, let to isolation of Actinobacteria from different habitats as well as development of methods to identify and test the wild type isolates by screening, cultivation, extraction of metabolites, chromatographic analysis and activity assays. With the beginning of the “Golden Age” of antibiotic discovery in 1940s–1960s, a lot of valuable actinomycetes-derived compounds were discovered, being afterwards in many cases developed to commercial agrochemical and pharmaceuticals products [41].
Streptomyces are capable of production of a wide variety of secondary metabolites, which are not essential primarily for survival, but enable them to protect themselves against certain environmental influences. These include chemically diverse compounds relevant for agriculture, medicine and industry, such as saccharides (e.g., aminoglycosides, lincosamides), nucleologists (aminonucleosides, peptidyl nucleosides), polyketides (e.g., tetracyclines, macrolides), non-ribosomal synthesized peptides (e.g., actinomycin), polypeptides, glycosides, triterpenoids (hopanoids), lipoproteins, alkaloids, polyethers, lantibiotics [42]. Despite the great diversity, many natural products can rely on similar biosynthesis principles to be led back. Basic units or preliminary stages are developed through a systematic chemical, regulated production process to form a biologically active molecule. mixed together. These underlying biosynthetic principles can be found in many different production strains [41,42,43].
Examples of secondary metabolites produced by Streptomyces include siderophores, which serve to improve the absorption of iron from the environment, pigments (e.g., melanin, carotenoids), antibiotics (e.g., chloramphenicol, lincomycin, neomycin, streptomycin, tetracycline, albaflavenone), immunosuppressants (FK-506, FK-520), cytostatics, antimycotics/fungicides (e.g., nystatin, natamycin, amphotericin B), herbicides (phosphinothricin), anti-tumor substances (doxorubicin), anthelmintic substances (avermectin), growth promoters in ruminant animal feed (monensin), as anticholesterol and coccidiostatic substances (e.g., lasalocid) and as insecticides (milbemycin), osmoprotectants (e.g., ectoin) [41,42,43,44]. Specifically, antibiotics feature different modes of action having impact on cell wall biosynthesis, membrane transport, RNA synthesis, DNA replication, translation, nucleotide and fatty acid build-up etc. About 2/3 of all known antibiotics are synthesized by Streptomyces [18], but the resources are far from exhausted [44]. The search for new natural substances involves a wide variety of paths taken in addition to the classic, complex biological and chemical screening methods, the numerous genetic and bioinformatics-based methods are preferred nowadays. Genomic approaches are used to search, identify, and investigate secondary metabolite clusters in isolated strains. This reveals the entire potential, which resources can be used much more efficiently [45]. Further efforts are required to identify the biosynthetic gene clusters (BGCs) encoded in the genome responsible for the biosynthesis of newly produced secondary metabolites. Targeted activation of biosynthetic gene clusters is the preferred approach for the discovery of novel secondary metabolites, which involves engineering cluster-situated regulators, refactoring the target clusters, and heterologous expression. Efficient genetic manipulation tools are required for targeted activation of biosynthetic gene clusters [44,45].
Advancements in whole-genome sequencing revealed that the genomes encode many (GCs to produce secondary metabolites. At the same time, only a few of the BGCs are expressed under standard conditions and the biosynthetic potential of a majority of actinomycetes remains unexploited. To address this challenge, methods of natural product discovery, such as isolation and strain cultivation methods, metabolomics and (meta)genomics as well as new molecular tools for the genetic engineering of Actinobacteria have been introduced [46].
1.5. Hosts for Heterologous Expression
Streptomyces have been repeatedly reported to be successfully used as hosts for the heterologous expression of genes. However, during cultivation and genetic handling, challenges can occur arising from complex metabolism and genome of Streptomyces. Another challenge is the long generation times of typical streptomycetes. In both liquid culture and solid media, it takes several days until cell growth has reached the stationary phase. In addition, Streptomyces are generally difficult to transform due to one or more efficient restriction/modification systems [47]. Also, after successful transformation efficient expression often fails because the host’s own strongly active proteases and synthesized heterologous proteins may immediately degrade. However, for some strains such as Streptomyces venezuelae and Streptomyces albus, high-throughput screening and high-frequency transformation protocols could be established along with optimized handling under laboratory conditions [48].
For improvement of yields of secondary metabolites, genetically modified Streptomyces hosts (also referred to as “super-hosts”) were generated. In such strains endogenous BGCs and nonessential genes or genomic regions are removed, which resulted in strains that can conserve energy and precursor building blocks having specific precursor pool. Examples include engineered strains of S. coelicolor, S. lividans, S. albus, S. avermitilis, S. chattanoogensis [49], S. tsukubaensis [50], S. venezuelae [51] and multiple others that have shown improved target secondary metabolite production.
2. Genetic Tools for Streptomyces
Genetic manipulation of Streptomyces is crucial for many academic and industrial goals and applications. The commonly used strategies of genetic engineering in Streptomyces include expression of multiple copies of the whole BGC, refactoring of the BGC by substitution or modification of native regulatory elements like promoters, expression of the BGC in optimized native or heterologous hosts, expression of regulator genes, deletion of genes encoding repressors. Such engineering approaches require genetic manipulation, which depends on available molecular toolboxes for assembling genetic constructs, as well as procedures for their introduction into the host. In the past decades, several bottlenecks were identified that retarded the process of natural product discovery from Streptomyces [18].
Lack of compatible molecular tools, limited cloning and DNA transfer methods, genetic instability, DNA degradation, high GC-content, and different codon usage of the newly introduced foreign DNA are obstacles that often prevent the genetic manipulation of actinobacterial strains [48]. Classical approaches include DNA overexpression, deletion, disruption, and replacement as well as employment of diverse plasmids (e.g., suicide vectors, with temperature-sensitive plasmids). Threse approaches require the selection and screening of single- and double-crossover recombination events, making these strategies time-consuming and of rather low efficiency, since double-crossover mutants are rarely obtained.
Advances in sequencing techniques and bioinformatics tools facilitated fast identification of new BGCs and other promising targets for genetic engineering. Enzymes for molecular biology applications, cloning strategies and approaches of synthetic biology involving new vectors and genetic parts, such as selection markers, attachment sites, replicons, promoters and terminators have been optimized for introduction and maintenance in actinobacterial strains. New methods for transfer of the generated constructs into the cell were established too [52,53].
Key challenges in development of genetic methods for actinomycetes have been the lack of suitable vehicles for DNA transfer, difficulties in introduction of DNA into the cell by passing the thick cell-wall barrier, restriction of the exogenous introduced DNA, inefficient gene expression due to high GC-content [54].
To challenge these limitations, different genome editing technologies have been developed: a) techniques to optimize expression of the BGCs in the heterologous host with acquisition of the target gene cluster from the native host genome (e.g., using a genomic library of cosmids, fosmids, BAC/PAC); b) techniques for ligation and assembly of the BGC to the vector by sticky/blunt end ligation, Gibson cloning or recombination in different hosts; c) techniques to transfer the BGC-encoded vector to the heterologous host for expression by conjugation or protoplast transformation; d) techniques to target secondary metabolite production by expression of an integrative or replicative BGC vector. Furthermore, during the last couple decades novel techniques have been proposed, such as PCR-targeting [55], Cre-loxP recombination system [56], I-SceI promoted recombination system [57], CRISPR/Cas-based approaches [58]. Discoveries on the clustered regularly interspaced short palindromic repeat (CRISPR)/CRISPR-associated protein (Cas)-based tools have further improved the genetic toolbox of Streptomyces. Furthermore, application of genetics parts, such as synthetic promoters (e.g., constitutive ermE, SF14P, kasOP, gapdh, rpsL promoters; inducible tipA nitA, xylA promoters), ribosome-binding sites (AAAGGAGG; native or synthetic RBSs), terminators (e.g., Fd, TD1), reporter genes (luxAB, amy, xylE, gusA; eGFP, sfGFP, mRFP, mCherry) expanded the toolbox for Streptomyces engineering (Figure 2) [59].
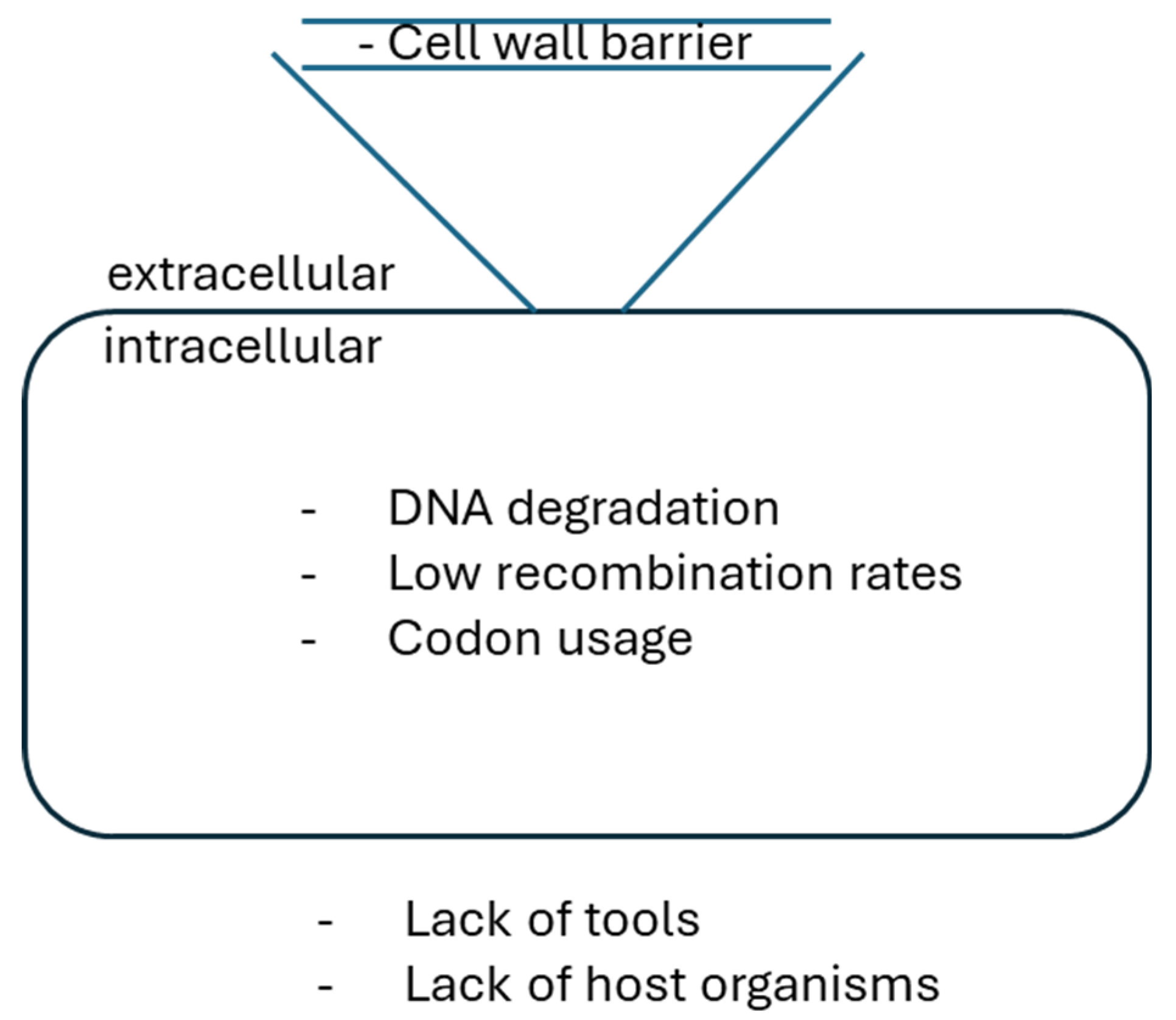
Figure 2.
Schematic representation of a cell with potential barriers to genetic manipulation, such as: lack of tools for DNA delivery and genome engineering, cell-wall barrier, DNA degradation, and codon usage (modified after [54]).
Figure 2.
Schematic representation of a cell with potential barriers to genetic manipulation, such as: lack of tools for DNA delivery and genome engineering, cell-wall barrier, DNA degradation, and codon usage (modified after [54]).
2.1. Methods for Heterologous DNA Transfer into Filamentous Actinobacteria
The process of DNA transfer into a cell involves its uptake across the complex cell wall. To overcome this barrier, multiple methods for DNA delivery into actinobacterial cells have been developed, including protoplast transformation, electroporation and conjugation. Protoplast transformation requires preparation of competent cells as unicellular protoplasts or as mycelium. Electroporation is based on the use of high-voltage pulse to a suspension of mycelium to induce the formation of membrane pores. Conjugation allows the DNA transfer by cell–cell contact [54].
Besides a few actinobacterial model organisms from the genus Streptomyces, such as S. coelicolor, S. lividans, S. griseus, S. avermitilis, S. venezuelae and S. albus, the majority of Actinobacteria are under investigation due to a production of a particular metabolite. Thus, DNA delivery methods are required have to be specifically adapted to a particular strain. There are several obstacles that challenge DNA transfer, e.g., thick cell walls and their composition. The DNA transfer protocols developed for Actinobacteria implemented different strategies to penetrate this barrier. Another obstacle is restriction modification systems that can interfere with the introduction of exogenous plasmid DNA, e.g., site-specifc endodeoxyribonucleases, restriction enzymes, nuclease-dependent systems. For example, the neomycin producer strain S. fradiae possess multiple restriction systems. Non-type II restriction and oxidative cleavage were reported as additional challenges to introducing foreign DNA. In troubleshooting efforts, DNA delivery methods have been widely optimized for non-model actinomycetes [18,54].
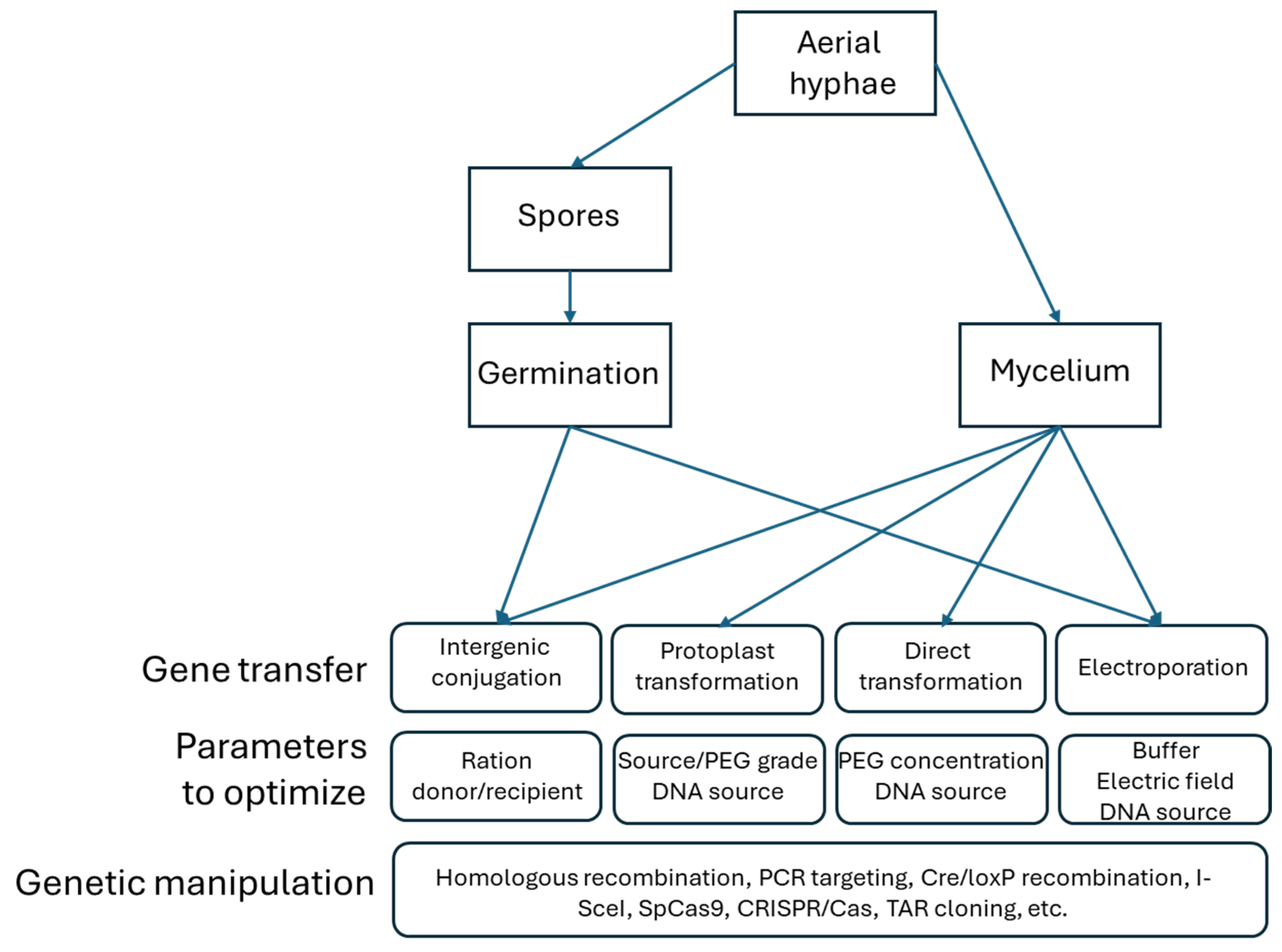
Figure 3.
Overview of the actinomycete DNA transfer systems including the parameters to be evaluated when applied to genetically manipulating strains (modified after [54]).
Figure 3.
Overview of the actinomycete DNA transfer systems including the parameters to be evaluated when applied to genetically manipulating strains (modified after [54]).
2.2. Synthetic Parts
2.2.1. Integrative and Replicative Plasmid Expression Systems for Actinomycetes-Cloning Vectors for the Genetic Manipulation of Filamentous Actinomycetes
Genetic manipulation of actinomycetes heavily depends on the availability of suitable vectors, such as plasmids, fosmids, cosmids as well as bacterial or P1-derived artificial chromosomes (BACs, PACs). Such genetic elements are required to support the molecular cloning of DNA fragments, introduction of DNA into the host cells, stable integration in the host cell and expression of the cloned genes. Such mobile genetic elements can either autonomously replicate in the cell cytoplasm or integrate into defined sites of the host chromosome. Although many naturally occurring mobile genetic elements like bacteriophages, plasmids and transposons have been found in Actinobacteria, only a few of them were applied for the genetic manipulation of actinomycetes [54,60].
The commonly used vectors can be grouped into two different types. First are autonomic replicative vectors featuring different copy number and including linear plasmids as well as suicidal (temperature-sensitive) vectors. Second type are integrative vectors that can carry out integration of DNA into the host genome via site-specific recombination. Such vectors are either derived from actinophages (e.g., FBT1, FC31, VWB) or actinobacterial integrative/conjugative elements (e.g., AICE). Various resistance markers have been applied for applications of plasmids for Streptomyces engineering. Some of these markers were sources from existing biosynthetic gene clusters. The commonly used marker genes provide resistance against thiostrepton (tsr), chloramphenicol (cat), hygromycin (hph), erythromycin (mls), viomycin (vio). Resistance genes originating from transposons of Gram-negative bacteria can also be applied for actinobacterial engineering, such as neomycin/kanamycin (aphII) and phleomycin (ble) from Tn5, gentamicin (aacC1 79) and apramycin (aac(3)IV). These resistance genes are expressed in Streptomyces from their own transposon promoters and seem to be functional in a wide range of actinobacterial strains [54,60,61].
Streptomycetes naturally have a wide variety of conjugative plasmids, however only very large linear plasmids encode biosynthetic pathways for antibiotics. There are “low copy” plasmids, small “multi copy” plasmids as well as integrative and conjugative elements of actinomycetes (AICE) [61]. Generally, plasmids are distinguished by genetic information for replication, maintenance, and conjugative transfer, but they vary in size, number of copies, form and replication mechanism. Some are exclusively extrachromosomal; others can integrate into the chromosome through specific recombination mechanisms. Low copy plasmids, small “multi-copy” plasmids and AICEs can also facilitate the transfer of chromosomal markers (chromosome mobilization ability) with the probability of 0.1%-1% [18,61,62].
Plasmid vectors are needed to allow cloning of foreign genes with high speed and frequency. Vectors with the possibility of positive/negative selection optimized workflows, e.g., SacB or RpsL. An alternative to these systems is the use of inversely repetitive sequences that flank the MIC (multiple cloning site) of the vector. In the initial state (“empty” vector), these complementary regions of the plasmid are separated by a region of DNA that acts as a spacer. During the cloning process the spacer area is cut out and replaced by a new DNA fragment. In the case of failure of insertion by re-ligation, the free DNA ends do not have a spacer between the inversely repetitive sequence areas no longer. This can happen due to their complementary interaction with each other, e.g., forming a hairpin structure. This hinders replication of the vector, so that synthesis of the DNA strand at this point is going to break off in the event of a termination loop during transcription [18,62]. It is of importance for practical use, as all successful transformants should carry a plasmid with a cloned insert. An example of such classical system is the plasmid pIJ699 [63].
Furthermore, the use of strong promoters in expression vectors is necessary to ensure sufficiently high expression rates. Although a regulatable promoter can offer additional advantages, it must be considered that regulated promoters usually have the problem of lower activity, while strong promoters are often unregulated. This has a great genetic potential, especially in soil or humus habitat, where most degradation processes of organic and inorganic material take place, which requires corresponding enzymatic performance [64]. Soil also provides the habitat of most active Actinomycetes, including Streptomyces, which possess many enzymes involved in biochemical reactions for degradation of organic and inorganic material. For example, the S. avermitilis genome possesses genes encoding 148 hydrolases, 59 esterases, 10 lipases, 128 proteases and peptidases and 78 oxygenases annotated [65]. Comparable figures apply also for the S. coelicolor genome [22].
G+C content of organisms found in soil is often very high, which makes cloning of their DNA difficult. In addition, there are also complex purification steps involved to remove all impurities from the DNA samples isolated from soil (e.g., humic acids) may lead to additional drop of cloning efficiency [66]. Thus, a cloning vector that has a positive selection is advantageous. This ensures that during cloning steps transformation of respective host organism works properly. Classical positive selection is based on the SacB system and RpsL system. SacB from Bacillus subtilis is a secreted levansucrase that catalyzes from sucrose in the presence of sucrose in the nutrient medium the synthesis of levan, which is lethal to many bacteria [67]. However, the SacB system is not suitable for use in Streptomyces hosts since it is not lethal for transformants with insert-free vectors so that they cannot be distinguished from transformants that carry vectors with an insert [68].
The RpsL system is based on the effect of the antibiotic streptomycin, whose site of action is the ribosomal protein 30S subunit. If this site of action is mutated, it gives the organism resistance to streptomycin. The vectors of the RpsL system carry the wild-type gene of the S12 protein and are designed in such a way that if cloning is successful, the gene is replaced by the cloned insert. The S12 protein of the host organism must be mutated before it is transformed with a cloning vector. During transformation, the host receives a vector molecule without an insert, e.g., with the intact S12 wild-type gene, the latter gives it back sensitivity to streptomycin because it is dominant over the mutated allele in the chromosome. Although the RpsL system in Streptomyces is functional, its major disadvantage lies in the laborious and time-consuming mutagenesis that is required for every host organism [69].
Inverse repetitive sequences have already been reported to be used as a possibility for positive selection, for example in the plasmids pIJ699, pJGSF14 and pJGSF15 [18]. These inversely repetitive sequences are separated by a spacer, which is replaced by the insert during cloning. The vector without spacer or insert forms directly adjacent complementary secondary structures that facilitate the replication of the vector, disrupt, or prevent it in the transformed host. A decisive advantage of this principle is its universal application in every bacterial host [61].
pUWL218, pUWL219, pUWL-SK and pUWL-KS shuttle vectors for E. coli and Streptomyces are used as expression vectors. The majority of integrative vectors are used in genetic engineering of Actinobacteria involve phage-derived site-specific DNA recombination systems, such as ΦC31, ΦBT1, VWB, TG1, SV1, R4. Some are derived from the integrative plasmid pSAM2 (λ-integrase) [61].
The ΦC31-derived vectors are able to integrate into the host genome at the attB site, but also when the correct attB site is missing. At lower frequencies such integration occurs at multiple locations via pseudo attachment sites. Thus, the ΦC31-derived integrative plasmids might have a broader host range. To have the possibility to use two compatible integrating vectors in the same organism, other similar vectors have been constructed based on the integrase genes and attP sites of other actinophages, such as ΦBT1, SV1, VWB [52,54,61].
Vectors harboring the ΦBT1 site are able to integrate at the unique attB site, which is localized in the sco4848 gene of the S. coelicolor genome or its orthologues in other Streptomyces spp. Integration of a plasmid at this position into Streptomyces genomes inactivates sco4849 and the co-transcribed gene sco4848. The most common integrative vectors used in Streptomyces are derived from the S. coelicolor A3(2) phage ΦC31, such as pSET152, pIJ8600, pOJ436, pOJ444. Furthermore, alternative integration sites were proposed, e. g. from the actinophage ΦJoe, which encodes a serine integrase and belongs to the largest Streptomyces phage R4-like clusters. Applicability of the ΦJoe system was demonstrated in in vitro recombination assays as well as in vivo in S. venezuelae and E. coli [52,54,59,61].
Recently, several vectors were introduced based on the concept of modular construct assembly or “plug-and-play” strategy. For instance, SEVA (Standard European Vector Architecture) was introduced as a web-based resource developed for easier construction of plasmids based on a fixed architecture. The modular concept of SEVA as well as the public availability of sequences for exchanging the modules within different vectors are of advantage for high-throughput generation of plasmids for engineering of Streptpmyces [70].
2.2.2. Promoters
In Streptomyces, constitutive promoters are extensively used for research and industrial applications (e.g., expression of BGCs), since they generate constant gene expression levels regardless of growth phase. The most used such strong promoter is ermE*, which is a derivative of the ermE promoter and contains a trinucleotide deletion in the ermEp1 region of the erythromycin resistance gene from S. erythraeus. Other used promoters include KasO from the SARP family regulator in S. coelicolor A3 and SF14 from the S. ghanaensis phage I19 genome, as well as promoters gapdh and rpsL from S. griseus, which have higher activity than the ermE* promoter [59,60,71].
To develop strong constitutive promoters, screening of strong synthetic promoters from a randomized promoter library has been shown to be an effective approach. The synthetic kasO promoter was designed based on a library that was used to generate synthetic promoters with promoter strength of 0.95%-187.5%, compared to the native parental kasO promoter [72]. Also, on the ermE and actII-orf4 promoters of S. coelicolor such an approach has been conducted, but synthetic promoters were still weaker than the ermE* promoter [73]. Strong promoter can also be identified using gene expression data, for example in S. albus and in S. coelicolor promoter sequences of genes with strong expression were selected based on their transcriptional profile, resulting in the selection of promoters that were stronger than the ermE* promoter [74].
Furthermore, controllable gene expression systems were established in Streptomyces. The most widely used inducible promoter in Streptomyces is the tipA promoter induced by thiostrepton. The expression level of the tipA promoter is considerable. This limits precise regulation of the targeted gene expression, but its expression is sometimes used to maintain low expression levels of toxic genes [75]. Another example is the tetracycline-inducible promoter tcp830, which was constructed by combining the ermE promoter and Tn10 tetR/tetO systems [76]. Other inducible promoters include PA3-rolO and P21-cmt promoters. The PA3-rolO promoter is resorcinol-inducible and is a combination of the rolO operator and synthetic promoter PA3. The P21-cmt promoter is a cumate-inducible system synthesized by fusing the operator of the Pseudomonas putida F1 cumate degradation operon to the P21 synthetic promoter [77]. Further inducible systems in Streptomyces include nitA and xylA promoters. The nitA promoter originating from the nitrilase promoter of Rhodococcus rhodochrous, can be induced by ε-caprolactam and the transcription regulator NitR. The xylA promoter is regulated by xylose. There are also several glycerol-inducible systems that have been developed for Streptomyces [60].
2.2.3. Reporter Genes
Characterization of genetic parts in Streptomyces requires reporter systems, which allow to rapidly represent gene expression level with almost no influence on the cell physiology. Many antibiotic resistance genes have been used as conventional markers for gene expression. However, effects of such genes on cellular metabolism and specific dynamic range are pitfalls when using these systems. For quantification of gene expression, colorimetric methods are preferable since gene expression levels can be quantified. Various colorimetric methods have been developed for Streptomyces, e.g., based on luxAB, amy, xylE, and gusA [48,59]. gusA is the most widely used colorimetric reporter system in Streptomyces, but colorimetric reporter systems based on an enzymatic reaction require an additional substrate treatment, which may negatively affect cellular metabolism. For instance, catechol dioxygenase (encoded by xylE) produces hydroxymuconic semialdehyde from catechol. On the other hand, fluorescent proteins do not require supplemental reagents and are suitable for screening in Streptomyces, for example GFP derivatives, such as eGFP and sfGFP, and mRFP, mTagBFP, mCerulean, mTFP, sfGFP, mCherry, mKate, and mCardinal. Since Streptomyces have relatively high levels of background autofluorescence, several studies have argued that fluorescent proteins are not suitable for high-throughput screening of Streptomyces [59,78].
2.2.4. Ribosome-Binding Sites
Protein levels in cells do not directly correlate with mRNA abundance and depends on translational efficiency. Thus, transcriptional regulation alone is considered to be not sufficient to design an efficient gene expression system in Streptomyces [79]. Development of a translational regulatory genetic part is rather at the initial stage. Transcriptional and translational genetic parts with various strengths have to be combined to be utilized to design and control enzyme stoichiometry in gene clusters. Translational efficiency is primarily determined by the 5′ untranslated regions (5′-UTR), ribosomal binding site (RBS), and codon usage of target genes. The RBS contains the Shine-Dalgarno (SD) sequence, which includes a complementary sequence with the 3′ end of the 16s rRNA region of the 30S ribosomal subunit. Accessibility and diversity of the SD sequence influence binding affinity with the ribosome and determine translational efficiency [80].
An example of RBS search and design is a study conducted in S. venezuelae, in which sequences of the strongest RBS among 15 native RBSs were selected and randomized. Subsequently, 177 synthetic RBSs with activity over 200-fold were compared to their parental RBSs. Seven promoters were combined with nine RBSs in a pairwise manner for screening of the most optimal promoter-RBS set for gene expression. Another example is a selection of two promoters and four 5′-UTR sequences that was conducted in S. coelicolor based on TSS-seq, RNA-seq, and Ribo-seq. Pairwise sets of promoters and 5′-UTR sequences demonstrated strength in a range of 0.03-2.4-fold, in comparison to the ermE* promoter containing the SD sequence of the nitA gene [81].
2.2.5. Terminators
Only a limited number of terminator sequences is available for Streptomyces. One example is TD1 from Bacillus subtilis phage ϕ29. Other examples include Fd, which is a bidirectional transcription terminator originated from Escherichia coli phage fd. Lambda T0 and T7 terminators have been used in multiple Streptomyces vectors, but such terminators have not been systematically validated in Streptomyces for effects on gene expression levels. Transcription terminators play crucial roles in recycling transcription complexes and in gene expression levels. Recently, Term-seq, which is a RNA sequencing method enabling genome-wide determination of transcript 3′ end positions, was proposed as a suitable technique for screening terminator sequences in Streptomyces genomes [48,59,82].
2.2.6. Riboswitches for Biosensors
A possibility to regulate gene expression is to use natural or modified riboswitches. Riboswitches are non-protein coding RNAs. They can regulate diverse cellular processes, including transcription and translation. These regulatory parts at the 5′-region of mRNA control the gene expression based on allosteric alterations of the structure. Riboswitches are specific to different ligands. This feature was leveraged for the development of biosensors as a synthetic biology tool [83].
Biosensors are composed of a signal input module (transcription factors or riboswitches), a regulatory module like transcription factor dependent promoters, and a signal output module, such as reporter genes. Application of the theophylline-dependent riboswitches were reported for S. coelicolor. Furthermore, an antibiotic-specific whole-cell TetR repressor-based biosensor has been introduced for screening and optimization of some antibiotic producer strains, such as pamamycin producer [48,84].
2.3. Bioinformatics-Based Approaches for Natural Products Discovery
Multiple genome-mining tools have been developed for identification of secondary metabolite biosynthetic gene clusters, most remarkable of which are ClustSCAN, NP searcher, GNP/PRISM, and antiSMASH. Currently, AntiSMASH is considered to be the most widely used tool for genome mining. It allows prediction of a broad spectrum of secondary metabolite biosynthetic gene clusters [45].
Information about the composition of secondary metabolite BGCs gained with genome mining is crucial for discovery of new secondary metabolites. It is a resource for synthetic biology based rational design of BGCs and yield improvement strategies of target compounds. For example, polyketides and non-ribosomal peptides can be redesigned due to synthesis by connected modular enzymes that are able to recognize module-specific molecules [85].
2.4. Assembly Strategies for Generation of Constructs for Genetic Engineering of Streptomyces
Cloning of actinobacterial sequences is challenging due to the high GC content genomes, highly conserved and repetitive sequences, and large BGCs size that can span more than 100 kb. Thus, new strategies to assemble constructs for genetic engineering of Actinobacteria were introduced [48,54].
The iCatch is a strategy based on the BioBricks concept. This tool was designed to facilitate “catching” of large DNA regions like actinobacterial BGCs. The BioBricks concept is based on the creation of DNA fragments flanked by the XbaI and SpeI restriction sites generating compatible cohesive ends. This allows easy assembly and ligation of any desired DNA sequence [86].
Direct Pathway cloning (DiPaC) allows the assembly of complete biosynthetic pathways, which is conducted by covering full BGCs with long-amplicon PCR. Introduction of homologous nucleotide overhangs allows subsequent in vitro HiFi DNA assembly like Gibson assembly. Moreover, sequence- and ligation-independent cloning (SLIC) can also be used as a not time-consuming alternative, since the terminal homologous sequences of the fragments anneal in vitro, but stitching of the gaps takes place in vivo in the E. coli host. DiPaC is suitable for cloning short to midsized BGCs in strains with high-GC content [87].
Another “plug-and-play” method is the artificial gene operon assembly system (AGOS). It is suitable for reconstruction and assembly of gene operons. This tool provides a set of entry plasmids, which are designed for artificial gene operon manufacturing. This application of this strategy has been demonstrated for disassembly of the 20 kb large novobiocin BGC [88].
Gibson Assembly is a commonly used method in synthetic biology and genetic engineering of Streptomyces. This method allows assembly of multiple up to 900 kb large DNA fragments into a linearized vector of choice and required only overlapping DNA sequences. However, the classical Gibson reaction is not optimal for cloning of large high GC content DNA fragments, since the self-ligation rate can reach 80%. In further modifications, two universal terminal single-stranded overhangs with high AT contents were added to the ends of the plasmid vector, decreasing the level of self-ligation to 45%, Also, the introduction of the restriction enzyme sites NdeIa nd NheI into the respective side of the overhangs facilitated the hierarchical and seamless assembly of large DNA molecules. The applicability of these strategies has been demonstrated for the assembly of the PII (pristinamycin II) BGC from Streptomyces pristinaespiralis HCCB10218 increasing from 2.5% with the classical protocol to 20%–40% with the modified version [48,89].
2.5. Genetic Approaches for Streptomyces Engineering
2.5.1. Transposon- and Homologous Recombination-Based Systems for Actinomycetes Engineering-I-SceI Meganuclease-Promoted Recombination System
Transposons (e.g., Tn5), homologous recombination and site-specific recombination mediated by Cre/loxP, Dre/rox, I-SceI systems belong to the most extensively used tools to generate mutants in Streptomyces strains. The engineering of other Actinomycetales, such as Planobispora sp., Kibdelosporangium sp., Amycolatopsis sp., remains difficult since most established methods fail in genetic manipulation of these strains [90].
For systematic studies of microorganisms, transposon-based genome mutagenesis is a widely applied technique. Meanwhile, a codon-optimized, hyperactive Tn5-based transposition system random mutagenesis in Streptomyces was introduced, which was utilized to investigate the regulation of prodiginine in S. coelicolor [91].
The I-SceI endonuclease together with the Cre recombinases have been used to generate large deletions in Streptomyces genomes. I-SceI meganuclease can recognize an 18-bp unique sequence (TAGGGATAACAGGGTAAT) and cause DNA double-strand breaks (DSBs), which promote double-crossover recombination events. This tool has been successfully validated in S. coelicolor: two BGCs that biosynthesize actinorhodin and undecylprodigiosin were successfully deleted by this technology [57].
2.5.2. PCR Targeting
In an attempt to overcome challenges of classical strain engineering in Streptomyces, a PCR targeting-based gene disruption approach was introduced. It permits deletions of entire gene clusters and allows rapid generation of nonpolar, in-frame deletions, avoiding polar effects on downstream genes. This procedure is based on the discovery that allelic exchanges on the E. coli chromosome can be induced by recombination with a PCR-amplified selectable marker, which is flanked at both ends by extensions of few tens nucleotides homologous to the desired region on the chromosome [48].
For this, the Redα (exo), Redβ (bet), and Redγ (gam) proteins of the phage λ should be present in the targeted strain. λ-Red can be used to promote recombination in E. coli between a PCR-amplified antibiotic resistance cassette selectable in E. coli and Streptomyces, and S. coelicolor DNA on a cosmid from the set used to sequence the S. coelicolor genome. An origin of transfer (oriT; RK2) included in the disruption cassette allows the transfer of the PCR-targeted cosmid into S. coelicolor by conjugation, readily yielding exconjugants with the desired gene replacement. This strategy also allows the elimination of the disruption cassette by FLP-recombinase-mediated site-specific recombination and was shown to be successfully applicable across Streptomyces strains [55].
2.5.3. Cre/loxP and Flp/FRT
Another tool for advanced Streptomyces engineering are the Cre/loxP system from the P1 phage and the Flp/FRT system from yeast. The Cre and Flp proteins are bidirectional tyrosine recombinases that catalyze reciprocal SSR of DNA at 34-bp target sites (loxP and FRT, respectively). It results in either excision or inversion depending on whether the target sites are located as direct or inverted repeats. Both recombinases are context independent since they do not require any cofactors or accessory proteins from the host. Both Flp and Cre recombinases have been successfully expressed in Actinobacteria. Successful expression of the Cre recombinase in Streptomyces coelicolor A3(2) was reported. Successful expression of two synthetic genes encoding the Cre and Flp recombinases were reported subsequently as well to delete resistance markers in members of the Streptomyces and Saccharothrix genera. The native Flp-encoding gene was also expressed in Streptomyces [92]. Recently, the Cre/loxP system was utilized for excision of a large genomic region - 1.3-Mb and 0.7-Mb were deleted in the natamycin producer strain Streptomyces chattanoogensis L10 [56,93].
2.5.4. CRISPR/Cas9-Based Editing Tools in Streptomyces
CRISPR/Cas9 (clustered regularly interspaced short palindromic repeats and CRISPR-associated protein 9) has recently emerged as a novel promising tool for genome engineering in Streptomyces. The CRISPR/Cas system usually consists of two components: a Cas protein with endonuclease activity and a guide RNA, which contains a spacer sequence that directs the endonuclease to its complementary protospacer sequence. A scaffold facilitates this interaction [48,58]. The Cas9 enzyme is an endonuclease that forms a complex with guide RNA (gRNA). It is guided to a protospacer sequence, which is complementary to a spacer sequence of guide RNA (gRNA). The guided Cas9 protein induces a double-strand DNA breaks (DSB) in the genome through two nuclease domains, HNH (complementary strand) and RuvC-like (non-complementary strand), when the correct protospacer adjacent motif (PAM) is present. That is followed by repairing the DSB via native non-homologous end joining repair (NHEJ) or homology directed repair (HDR) mechanisms. During DNA repair via the NHEJ pathway, small deletions or insertions can occur randomly at the DNA cleavage site [58,94].
Precise genome editing is enabled through DNA repair via the HDR pathway, which requires the donor DNA to be recombined into the cleavage site. CRISPR/Cas9 systems can facilitate unmarked genome engineering with reduced time and labor. Although other genome engineering strategies like zinc finger nuclease or transcription activator-like effector nuclease have been developed, protein-mediated recognition of target DNA sequence requires design of respective proteins for individual target sequences. This limits their applications. For this reason, the CRISPR/Cas9 system has become a dominant genome engineering tool nowadays. Genes involved in the bacterial NHEJ pathway are absent in most Streptomyces. Thus, genome editing mediated by the HDR pathway is preferred for CRISPR/Cas100 based genetic engineering [95]. Achieving precise and scarless genome editing in Streptomyces was usually mostly dependent on double-crossover events that required time-consuming and labor-intensive screening steps [18]. Given the lethal DSB induced by the CRISPR/Cas system in bacteria unless repaired, the surviving cells after transformation of the CRISPR/Cas plasmid are more likely to be successfully engineered, which reduced the time and labor required for the screening steps [94,95,96].
The first CRISPR/Cas system applied to Streptomyces was based on the class 2–type II CRISPR system. It uses Cas9 from Streptococcus pyogenes as a nuclease. Cas9 requires 5′-NGG-3′ as the PAM, which is frequently found in the GC-rich genomes of Streptomyces. Using the CRISPR/Cas9 system, genomic deletions of up to 31.4 kbp have been performed in S. lividans, S. viridochromogenes, and S. albus [94]. Additional CRISPR/Cas9 systems were explored subsequently leading to additional applications and advancements [95,96,97,98]. To enhance genome editing efficiency via the NHEJ approach, a core component of the NHEJ pathway DNA ligase D was co-expressed with the CRISPR/Cas system. Enhanced editing efficiency can facilitate rapid gene disruption as there is no requirement for donor DNAs for homologous recombination. Although most CRISPR/Cas9 systems rely on a temperature-sensitive origin to cure the plasmid, an alternative counterselection method for screening plasmid-cured strains has been described as well [98], based on a mutant cytosine deaminase that can convert 5-fluorocytosine into the highly toxic 5-fluorouracil. It was integrated into the CRISPR/Cas9 plasmid, and treatment with 5-fluorocytosine effectively eliminated the strains retaining the plasmid [58,98].
The CRISPR/Cas system for Streptomyces was recently evolving by expanding its applicable hosts, including S. rimosus, S. ambofaciens, and S. roseosporus, and exploiting different types of CRISPR/Cas systems [58,99,100,101,102]. Especially, the class 2–type V CRISPR/Cas system, exploiting Cas12a from Francisella novicida, was applied to Streptomyces, expanding the number of editable DNA sequences with 5′-TTV-3′ PAM [103]. Although the 5′- NGG-3′ PAM of Cas9 may provide a wide range of potential editing targets, the AT-rich PAM of Cas12a provided advantages for target specificity in the GC-rich genome of Streptomyces. Cas12a is also more suited for multiplexing compared to Cas9, because it enables the maturation of CRISPR RNA (crRNA) [104].
In addition to expanding the PAM sequence, the establishment of different types of CRISPR/Cas systems for engineering in Streptomyces is important due to the exploration of secondary metabolites. The applicability of a specific type of CRISPR system may differ from species to species [101,103]. Both CRISPR/Cas9 and CRISPR/Cas12a are convenient to use because only a single Cas protein is required to induce DSB in the genome [105]. The introduced foreign CRISPR/Cas system may crosstalk with the native CRISPR/Cas system encoded within the genome, resulting in reduced engineering efficiency and defects in the host [106]. In particular, the class 1 type I CRISPR/Cas system, which requires a Cas3 nuclease and a ribonucleoprotein complex comprising Cas5, Cas7, Cas8 proteins, and crRNA, is the most widespread CRISPR/Cas system in Streptomyces. Class 1 type I CRISPR/Cas was shown to appear advantageous for a broad range of Streptomyces species [58,102].
CRISPR/Cas9 approach has been applied to identify secondary metabolites gene clusters for multiple secondary metabolites, including sceliphrolactam, valinomycin, and griseusin, by deleting a synthesis gene in the BGC or introducing a stop codon in the synthesis gene using the CRISPR/Cas system [107]. BGC disruption enables functional screening of BGCs. For example, in S. globisporus SP6C4, the biosynthetic genes of 15 BGCs were deleted using the CRISPR/Cas system, and the BGC-disrupted strains were tested for antifungal and antibacterial activities, revealing three bioactive lantipeptide- or lassopeptide-type secondary metabolites [108]. The CRISPR/Cas system is innovative for characterizing secondary metabolite BGCs due to its high precision and speed - it is likely to accelerate the discovery of novel secondary metabolites using base edition [58,59].
2.5.5. CRISPR/Cas9 TAR Cloning Approach
Transformation associated recombination (TAR)-cloning allows isolation of large chromosomal regions without the effort of constructing a random clone library. The method is based on homologous recombination of a specific genome target as well as on a linear vector containing anchor-like, specific terminal targeting sequences, which are homolog to the sequence of interest. The vector and genomic DNA are introduced into yeast, with subsequent homologous recombination of the flanking regions of the target sequences and the vector anchors. This results in Yeast Artificial Chromosomes (YAC) containing the target sequence. TAR allows cloning of DNA fragments up to 250 kb with rather low efficiency: the correct construct is ~1%–5%. Newer approaches attempted to optimize this - the specificity of TAR cloning was increased by using CRISPR/Cas9 to introduce the DSBs, which increase efficiency by 30%. An advantage is that almost any sequence of interest can be cloned into a YAC vector, without the need of specific and rare restriction enzymes, which cut near the target sequence. This method was applied for cloning the prodigiosin BGC from S. coelicolor as well as the pristinamycin BGC from S. pristinaespiralis (Figure 4) [109,110].
Figure 4.
Design and strategy of TAR-based cloning and expression (modified after [109]).
Figure 4.
Design and strategy of TAR-based cloning and expression (modified after [109]).


Table 1.
A combined list of synthetic biology technologies for genetic manipulation of Streptomyces.
Table 1.
A combined list of synthetic biology technologies for genetic manipulation of Streptomyces.
| Technology | Category | Feature | Reference |
|---|---|---|---|
| Target BGC acquisition | Genetic manipulation strategy | Transfer into a target host using a genomic library of cosmids, fosmids, BAC, PAC | [60,61,62] |
| Ligation and BGC assembly to the vector | Genetic manipulation strategy | Sticky/blunt end ligation, Gibson cloning, recombination in different hosts, etc. | [48,54,59] |
| Transfer of the BGC-encoded vector to theheterologous host for expression | Genetic manipulation strategy | Biparental conjugation, protoplast transformation | [54] |
| Target secondary metabolite production by expression of the BGC vector | Genetic manipulation strategy | Expression of integrative (e.g., pSET152, pIJ8600, pOJ436, pOJ444) or replicative (e.g., pUWL218, pUWL219, pUWL-SK, pUWL-KS) vectors | [60,61,62] |
| iCatch | Assembly strategy | Facilitate “catching” of large DNA regions like actinobacterial BGCs | [86] |
| DiPAC | Assembly strat-egy | Assembly of complete biosynthetic pathways by covering full BGCs with long-amplicon PCR | [87] |
| AGOS | Assembly strat-egy | Reconstruction and assembly of gene operons | [88] |
| PCR-targetingsystem | Genetic manipulation strategy | Nonpolar/in-frame deletion of genes/gene clusters in Streptomyces | [55] |
| Cre-loxP/Flp-FRP recombination systems | Genetic manipulation strategy | Knock out large DNA fragments in Streptomyces | [56,92] |
| I-SceI promoted recombination system | Genetic manipulation strategy | DNA double-strand breaks (DSBs), which promote double-crossover recombination events | [90] |
| SpCas9-based genome editing | Genetic manipulation strategy | Transcribed synthetic guide RNA to direct Cas proteins to any site on the genome. Editing plasmids: pCRISPomyces-1/2, pKCas9dO, pCRISPR-Cas9-ScaligD, and pWHU2653 | [94,95,96,97] |
| CRISPRi-mediated gene repression for single cells | Genetic manipulation strategy | Gene repression tool based on dCas9 or ddCpf1 and the base editors (BEs) for targeted base mutagenesis based on dCas9 or Cas9n | [95] |
| FnCpf1-based genome editing and CRISPRi | Genetic manipulation strategy | Editing plasmids: pKCCpf1, pKCCpf1-MsmE, and pSETddCpf1, etc. | [103] |
| CRISPR/Cas-based base editing tools | Genetic manipulation strategy | Editing plasmids: pCRISPR-cBEST/-aBEST, and pKC-dCas9-CDA-ULstr, etc. | [58,94,95,96] |
| Alternative CRISPR/Cas-based genome editing | Genetic manipulation strategy | Editing plasmids: pCRISPomyces-FnCpf1, pCRISPomyces-Sth1Cas9, and pCRISPomyces-SaCas9, etc. | [101,102,103,104] |
| TAR-cloning | Genetic manipulation strategy | Isolation of large chromosomal regions without the constructing a random clone library | [109,110] |
| Synthetic promoters | Genetics parts | Constitutive ermE, SF14P, kasOP, gapdh, rpsL promoters as well as inducible tipA nitA and xylA promoters | [59,60,72] |
| Ribosome-binding sites | Genetics parts | AAAGGAGG, diverse native or synthetic RBSs | [80,81] |
| Terminators | Genetics parts | Fdand TD1 | [59,82] |
| Reporter genes | Genetics parts | luxAB, amy, xylE, and gusA / eGFP, sfGFP, mRFP, mCherry | [59] |
| Riboswitches/Biosensors | Genetics parts | Non-protein coding RNAs that can regulate cellular processes including transcription and translation | [83,84] |
3. Conclusions and Future Perspectives
The complex biosynthesis of secondary metabolites in Streptomyces wild type producers naturally results in rather low titers. Biosynthetic steps in primary and secondary metabolism as well as their regulation represent targets for engineering. The dependence of secondary metabolism in Streptomyces spp. on precursor supply coming from primary metabolism is crucial and can be optimized for biotechnological needs. However, genetic engineering of Streptomyces has historically been challenging. New tools proposed recently accelerated Streptomyces engineering, introduction a growing number of new approaches for BGC and metabolic engineering. The traditional method for the generation of deletion mutants is time-consuming. Novel approaches, such as CRISPR/Cas system, TAR-cloning, PCR-targeting or Cre/loxP recombination were adapted to Streptomyces for fast genome editing. For instance, several optimized CRISPR/Cas-based technologies were recently reported to improve the turnaround time for generation of deletion mutants even further. Implementation of new systems resulted in the efficient genome engineering of a broad range of Streptomyces strains and some other actinomycetes. Newly developed tools provide new opportunities for natural product discovery and metabolic engineering of Streptomyces. It is likely that this progress will continue, and additional cell factories can be developed for development and manufacturing of various valuable products.
Author Contributions
Conceptualization, writing—original draft preparation, visualization, S.K.
Funding
This research received no external funding.
Acknowledgments
I thank Mariko Matsuura, Mary Bosserman, April MacIntyre and Daniela Floss for proof-reading the paper.
Conflicts of Interest
Sergii Krysenko is employed by the Valent BioSciences. The author declares no conflict of interest, the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Saiki, R. K., Gelfand, D. H., Stoffel, S., Scharf, S. J., Higuchi, R., Horn, G. T., Mullis, K. B., & Erlich, H. A. (1988). Primer-directed enzymatic amplification of DNA with a thermostable DNA polymerase. Science (New York, N.Y.), 239(4839), 487–491. [CrossRef]
- Liu, F., Xiong, J., Kumar, S., Yang, C., Ge, S., Li, S., Xia, N., & Swaminathan, K. (2011). Structural and biophysical characterization of Mycobacterium tuberculosis dodecin Rv1498A. Journal of structural biology, 175(1), 31–38. [CrossRef]
- Hibbing, M. E., Fuqua, C., Parsek, M. R., & Peterson, S. B. (2010). Bacterial competition: surviving and thriving in the microbial jungle. Nature reviews. Microbiology, 8(1), 15–25. [CrossRef]
- Stubbendieck, R. M., Vargas-Bautista, C., & Straight, P. D. (2016). Bacterial Communities: Interactions to Scale. Frontiers in microbiology, 7, 1234. [CrossRef]
- Crawford, J. M., & Clardy, J. (2011). Bacterial symbionts and natural products. Chemical communications (Cambridge, England), 47(27), 7559–7566. [CrossRef]
- Netzker, T., Fischer, J., Weber, J., Mattern, D. J., König, C. C., Valiante, V., Schroeckh, V., & Brakhage, A. A. (2015). Microbial communication leading to the activation of silent fungal secondary metabolite gene clusters. Frontiers in microbiology, 6, 299. [CrossRef]
- Schrempf H, Cypionka H, Keller U (2016) Streptomyceten. Biologie in unserer Zeit 46:236-243. [CrossRef]
- Goodfellow, M.; Nouioui, I.; Sanderson, R.; Xie, F.; Bull, A.T. Rare Taxa and Dark Microbial Matter: Novel Bioactive Actinobacteria Abound in Atacama Desert Soils. Antonie Van Leeuwenhoek 2018, 111, 1315–1332.
- Seipke, R. F., Kaltenpoth, M., & Hutchings, M. I. (2012). Streptomyces as symbionts: an emerging and widespread theme?. FEMS microbiology reviews, 36(4), 862–876. [CrossRef]
- Haeder, S., Wirth, R., Herz, H., & Spiteller, D. (2009). Candicidin-producing Streptomyces support leaf-cutting ants to protect their fungus garden against the pathogenic fungus Escovopsis. Proceedings of the National Academy of Sciences of the United States of America, 106(12), 4742–4746. [CrossRef]
- Kaltenpoth, M., Yildirim, E., Gürbüz, M. F., Herzner, G., & Strohm, E. (2012). Refining the roots of the beewolf-Streptomyces symbiosis: antennal symbionts in the rare genus Philanthinus (Hymenoptera, Crabronidae). Applied and environmental microbiology, 78(3), 822–827. [CrossRef]
- Pantigoso, H. A., Newberger, D., & Vivanco, J. M. (2022). The rhizosphere microbiome: Plant-microbial interactions for resource acquisition. Journal of applied microbiology, 133(5), 2864–2876. [CrossRef]
- Upadhyay, S. K., Srivastava, A. K., Rajput, V. D., Chauhan, P. K., Bhojiya, A. A., Jain, D., Chaubey, G., Dwivedi, P., Sharma, B., & Minkina, T. (2022). Root Exudates: Mechanistic Insight of Plant Growth Promoting Rhizobacteria for Sustainable Crop Production. Frontiers in microbiology, 13, 916488. [CrossRef]
- Duan, S., Feng, G., Limpens, E., Bonfante, P., Xie, X., & Zhang, L. (2024). Cross-kingdom nutrient exchange in the plant-arbuscular mycorrhizal fungus-bacterium continuum. Nature reviews. Microbiology, 10.1038/s41579-024-01073-7. Advance online publication. [CrossRef]
- Zhang, L., Zhou, J., George, T. S., Limpens, E., & Feng, G. (2022). Arbuscular mycorrhizal fungi conducting the hyphosphere bacterial orchestra. Trends in plant science, 27(4), 402–411. [CrossRef]
- Jin, Z., Jiang, F., Wang, L., Declerck, S., Feng, G., & Zhang, L. (2024). Arbuscular mycorrhizal fungi and Streptomyces: brothers in arms to shape the structure and function of the hyphosphere microbiome in the early stage of interaction. Microbiome, 12(1), 83. [CrossRef]
- Le, K. D., Yu, N. H., Park, A. R., Park, D. J., Kim, C. J., & Kim, J. C. (2022). Streptomyces sp. AN090126 as a Biocontrol Agent against Bacterial and Fungal Plant Diseases. Microorganisms, 10(4), 791. [CrossRef]
- Kieser T., Bibb M. J., Buttner M. J., Chater K. F., Hopwood D. A. Practical Streptomyces Genetics, John Innes Foundation, Norwich, 2000.
- Ismail, S., Jiang, B., Nasimi, Z., Inam-Ul-Haq, M., Yamamoto, N., Danso Ofori, A., Khan, N., Arshad, M., Abbas, K., & Zheng, A. (2020). Investigation of Streptomycesscabies Causing Potato Scab by Various Detection Techniques, Its Pathogenicity and Determination of Host-Disease Resistance in Potato Germplasm. Pathogens (Basel, Switzerland), 9(9), 760. [CrossRef]
- Pérez, J., Muñoz-Dorado, J., Braña, A. F., Shimkets, L. J., Sevillano, L., & Santamaría, R. I. (2011). Myxococcus xanthus induces actinorhodin overproduction and aerial mycelium formation by Streptomyces coelicolor. Microbial biotechnology, 4(2), 175–183. [CrossRef]
- Yagüe, P., Lopez-Garcia, M. T., Rioseras, B., Sanchez, J., & Manteca, A. (2012). New insights on the development of Streptomyces and their relationships with secondary metabolite production. Current trends in microbiology, 8, 65–73.
- Bentley, S. D., Chater, K. F., Cerdeño-Tárraga, A. M., Challis, G. L., Thomson, N. R., James, K. D., Harris, D. E., Quail, M. A., Kieser, H., Harper, D., Bateman, A., Brown, S., Chandra, G., Chen, C. W., Collins, M., Cronin, A., Fraser, A., Goble, A., Hidalgo, J., Hornsby, T., … Hopwood, D. A. (2002). Complete genome sequence of the model actinomycete Streptomyces coelicolor A3(2). Nature, 417(6885), 141–147. [CrossRef]
- Krysenko, S.; Wohlleben, W. Role of Carbon, Nitrogen, Phosphate and Sulfur Metabolism in Secondary Metabolism Precursor Supply in Streptomyces spp. Microorganisms 2024, 12, 1571. [CrossRef]
- Hodgson D. A. (2000). Primary metabolism and its control in streptomycetes: a most unusual group of bacteria. Advances in microbial physiology, 42, 47–238. [CrossRef]
- Krysenko S, Wohlleben W. (2022) Polyamine and Ethanolamine Metabolism in Bacteria as an Important Component of Nitrogen Assimilation for Survival and Pathogenicity. Medical Sciences; 10(3):40. [CrossRef]
- Görke, B., & Stülke, J. (2008). Carbon catabolite repression in bacteria: many ways to make the most out of nutrients. Nature reviews. Microbiology, 6(8), 613–624. [CrossRef]
- Romero-Rodríguez, A., Rocha, D., Ruiz-Villafán, B., Guzmán-Trampe, S., Maldonado-Carmona, N., Vázquez-Hernández, M., Zelarayán, A., Rodríguez-Sanoja, R., & Sánchez, S. (2017). Carbon catabolite regulation in Streptomyces: new insights and lessons learned. World journal of microbiology & biotechnology, 33(9), 162. [CrossRef]
- Jahreis, K., Pimentel-Schmitt, E. F., Brückner, R., & Titgemeyer, F. (2008). Ins and outs of glucose transport systems in eubacteria. FEMS microbiology reviews, 32(6), 891–907. [CrossRef]
- Merrick, M. J., & Edwards, R. A. (1995). Nitrogen control in bacteria. Microbiological reviews, 59(4), 604–622. [CrossRef]
- Fink, D., Weissschuh, N., Reuther, J., Wohlleben, W., & Engels, A. (2002). Two transcriptional regulators GlnR and GlnRII are involved in regulation of nitrogen metabolism in Streptomyces coelicolor A3(2). Molecular microbiology, 46(2), 331–347. [CrossRef]
- Fink, D., Falke, D., Wohlleben, W., & Engels, A. (1999). Nitrogen metabolism in Streptomyces coelicolor A3(2): modification of glutamine synthetase I by an adenylyltransferase. Microbiology (Reading, England), 145 ( Pt 9), 2313–2322. [CrossRef]
- Ghoshroy, S., Binder, M., Tartar, A., & Robertson, D. L. (2010). Molecular evolution of glutamine synthetase II: Phylogenetic evidence of a non-endosymbiotic gene transfer event early in plant evolution. BMC evolutionary biology, 10, 198. [CrossRef]
- Reuther, J., & Wohlleben, W. (2007). Nitrogen metabolism in Streptomyces coelicolor: transcriptional and post-translational regulation. Journal of molecular microbiology and biotechnology, 12(1-2), 139–146. [CrossRef]
- Hillemann, D., Dammann, T., Hillemann, A., & Wohlleben, W. (1993). Genetic and biochemical characterization of the two glutamine synthetases GSI and GSII of the phosphinothricyl-alanyl-alanine producer, streptomyces viridochromogenes Tü494. Journal of general microbiology, 139(8), 1773–1783. [CrossRef]
- Krysenko S, Emani CS, Bäuerle M, Oswald M, Kulik A, Meyners C, Hillemann D, Merker M, Wohlers I, Hausch F, Brötz-Oesterhelt H, Mitulski A, Reiling N, Wohlleben W. n.d. Glna3Mt is able to Glutamylate Spermine but it is not essential for the detoxification of Spermine in Mycobacterium tuberculosis. bioRxiv. [CrossRef]
- Martín, J. F., & Liras, P. (2020). The Balance Metabolism Safety Net: Integration of Stress Signals by Interacting Transcriptional Factors in Streptomyces and Related Actinobacteria. Frontiers in microbiology, 10, 3120. [CrossRef]
- Romero-Rodríguez, A., Maldonado-Carmona, N., Ruiz-Villafán, B., Koirala, N., Rocha, D., & Sánchez, S. (2018). Interplay between carbon, nitrogen and phosphate utilization in the control of secondary metabolite production in Streptomyces. Antonie van Leeuwenhoek, 111(5), 761–781. [CrossRef]
- Ghorbel, S., Smirnov, A., Chouayekh, H., Sperandio, B., Esnault, C., Kormanec, J., & Virolle, M. J. (2006). Regulation of ppk expression and in vivo function of Ppk in Streptomyces lividans TK24. Journal of bacteriology, 188(17), 6269–6276. [CrossRef]
- Ferla, M. P., & Patrick, W. M. (2014). Bacterial methionine biosynthesis. Microbiology (Reading, England), 160(Pt 8), 1571–1584. [CrossRef]
- Kulkarni, A., Zeng, Y., Zhou, W., Van Lanen, S., Zhang, W., & Chen, S. (2015). A Branch Point of Streptomyces Sulfur Amino Acid Metabolism Controls the Production of Albomycin. Applied and environmental microbiology, 82(2), 467–477. [CrossRef]
- De Simeis, D., & Serra, S. (2021). Actinomycetes: A Never-Ending Source of Bioactive Compounds-An Overview on Antibiotics Production. Antibiotics (Basel, Switzerland), 10(5), 483. [CrossRef]
- Alam, K., Mazumder, A., Sikdar, S., Zhao, Y. M., Hao, J., Song, C., Wang, Y., Sarkar, R., Islam, S., Zhang, Y., & Li, A. (2022). Streptomyces: The biofactory of secondary metabolites. Frontiers in microbiology, 13, 968053. [CrossRef]
- Krysenko, S. Impact of Nitrogen-Containing Compounds on Secondary Metabolism in Streptomyces spp.—A Source of Metabolic Engineering Strategies. SynBio 2023, 1, 204-225. [CrossRef]
- Alam, K., Mazumder, A., Sikdar, S., Zhao, Y. M., Hao, J., Song, C., Wang, Y., Sarkar, R., Islam, S., Zhang, Y., & Li, A. (2022). Streptomyces: The biofactory of secondary metabolites. Frontiers in microbiology, 13, 968053. [CrossRef]
- Albarano, L., Esposito, R., Ruocco, N., & Costantini, M. (2020). Genome Mining as New Challenge in Natural Products Discovery. Marine drugs, 18(4), 199. [CrossRef]
- Miethke, M., Pieroni, M., Weber, T., Brönstrup, M., Hammann, P., Halby, L., Arimondo, P. B., Glaser, P., Aigle, B., Bode, H. B., Moreira, R., Li, Y., Luzhetskyy, A., Medema, M. H., Pernodet, J. L., Stadler, M., Tormo, J. R., Genilloud, O., Truman, A. W., Weissman, K. J., … Müller, R. (2021). Towards the sustainable discovery and development of new antibiotics. Nature reviews. Chemistry, 5(10), 726–749. [CrossRef]
- Hwang, S., Lee, Y., Kim, J. H., Kim, G., Kim, H., Kim, W., Cho, S., Palsson, B. O., & Cho, B. K. (2021). Streptomyces as Microbial Chassis for Heterologous Protein Expression. Frontiers in bioengineering and biotechnology, 9, 804295. [CrossRef]
- Mitousis, L.; Thoma, Y.; Musiol-Kroll, E.M. An Update on Molecular Tools for Genetic Engineering of Actinomycetes—The Source of Important Antibiotics and Other Valuable Compounds. Antibiotics 2020, 9, 494. [CrossRef]
- Myronovskyi, M., Welle, E., Fedorenko, V., & Luzhetskyy, A. (2011). Beta-glucuronidase as a sensitive and versatile reporter in actinomycetes. Applied and environmental microbiology, 77(15), 5370–5383. [CrossRef]
- Schulz, S., Schall, C., Stehle, T., Breitmeyer, C., Krysenko, S., Mitulski, A., & Wohlleben, W. (2022). Optimization of the precursor supply for an enhanced FK506 production in Streptomyces tsukubaensis. Frontiers in bioengineering and biotechnology, 10, 1067467. [CrossRef]
- Kim, W., Hwang, S., Lee, N., Lee, Y., Cho, S., Palsson, B., & Cho, B. K. (2020). Transcriptome and translatome profiles of Streptomyces species in different growth phases. Scientific data, 7(1), 138. [CrossRef]
- Wohlleben, W., Hartmann, V., Hillemann, D., Krey, K., Muth, G., Nussbaumer, B., & Pelzer, S. (1994). Transfer and establishment of DNA in Streptomyces (a brief review). Acta microbiologica et immunologica Hungarica, 41(4), 381–389.
- Kormanec, J., Rezuchova, B., Homerova, D., Csolleiova, D., Sevcikova, B., Novakova, R., & Feckova, L. (2019). Recent achievements in the generation of stable genome alterations/mutations in species of the genus Streptomyces. Applied microbiology and biotechnology, 103(14), 5463–5482. [CrossRef]
- Musiol-Kroll, E. M., Tocchetti, A., Sosio, M., & Stegmann, E. (2019). Challenges and advances in genetic manipulation of filamentous actinomycetes - the remarkable producers of specialized metabolites. Natural product reports, 36(9), 1351–1369. [CrossRef]
- Gust, B., Challis, G. L., Fowler, K., Kieser, T., & Chater, K. F. (2003). PCR-targeted Streptomyces gene replacement identifies a protein domain needed for biosynthesis of the sesquiterpene soil odor geosmin. Proceedings of the National Academy of Sciences of the United States of America, 100(4), 1541–1546. [CrossRef]
- Komatsu, M., Uchiyama, T., Omura, S., Cane, D. E., & Ikeda, H. (2010). Genome-minimized Streptomyces host for the heterologous expression of secondary metabolism. Proceedings of the National Academy of Sciences of the United States of America, 107(6), 2646–2651. [CrossRef]
- Lu, Z., Xie, P., & Qin, Z. (2010). Promotion of markerless deletion of the actinorhodin biosynthetic gene cluster in Streptomyces coelicolor. Acta biochimica et biophysica Sinica, 42(10), 717–721. [CrossRef]
- Lee, Y., Hwang, S., Kim, W., Kim, J. H., Palsson, B. O., & Cho, B. K. (2024). CRISPR-aided genome engineering for secondary metabolite biosynthesis in Streptomyces. Journal of industrial microbiology & biotechnology, 51, kuae009. [CrossRef]
- Lee, N., Hwang, S., Lee, Y., Cho, S., Palsson, B., & Cho, B. K. (2019). Synthetic Biology Tools for Novel Secondary Metabolite Discovery in Streptomyces. Journal of microbiology and biotechnology, 29(5), 667–686. [CrossRef]
- Aubry, C., Pernodet, J. L., & Lautru, S. (2019). Modular and Integrative Vectors for Synthetic Biology Applications in Streptomyces spp. Applied and environmental microbiology, 85(16), e00485-19. [CrossRef]
- Thoma, L., & Muth, G. (2016). Conjugative DNA-transfer in Streptomyces, a mycelial organism. Plasmid, 87-88, 1–9. [CrossRef]
- Wang, W., Zheng, G., & Lu, Y. (2021). Recent Advances in Strategies for the Cloning of Natural Product Biosynthetic Gene Clusters. Frontiers in bioengineering and biotechnology, 9, 692797. [CrossRef]
- Kieser, T., & Melton, R. E. (1988). Plasmid pIJ699, a multi-copy positive-selection vector for Streptomyces. Gene, 65(1), 83–91. [CrossRef]
- Torsvik, V., & Øvreås, L. (2002). Microbial diversity and function in soil: from genes to ecosystems. Current opinion in microbiology, 5(3), 240–245. [CrossRef]
- Omura, S., Ikeda, H., Ishikawa, J., Hanamoto, A., Takahashi, C., Shinose, M., Takahashi, Y., Horikawa, H., Nakazawa, H., Osonoe, T., Kikuchi, H., Shiba, T., Sakaki, Y., & Hattori, M. (2001). Genome sequence of an industrial microorganism Streptomyces avermitilis: deducing the ability of producing secondary metabolites. Proceedings of the National Academy of Sciences of the United States of America, 98(21), 12215–12220. [CrossRef]
- Wnuk, E., Waśko, A., Walkiewicz, A., Bartmiński, P., Bejger, R., Mielnik, L., & Bieganowski, A. (2020). The effects of humic substances on DNA isolation from soils. PeerJ, 8, e9378. [CrossRef]
- Reyrat, J. M., Pelicic, V., Gicquel, B., & Rappuoli, R. (1998). Counterselectable markers: untapped tools for bacterial genetics and pathogenesis. Infection and immunity, 66(9), 4011–4017. [CrossRef]
- Jäger, W., Schäfer, A., Pühler, A., Labes, G., & Wohlleben, W. (1992). Expression of the Bacillus subtilis sacB gene leads to sucrose sensitivity in the gram-positive bacterium Corynebacterium glutamicum but not in Streptomyces lividans. Journal of bacteriology, 174(16), 5462–5465. [CrossRef]
- Hosted, T. J., & Baltz, R. H. (1997). Use of rpsL for dominance selection and gene replacement in Streptomyces roseosporus. Journal of bacteriology, 179(1), 180–186. [CrossRef]
- Martínez-García, E., Goñi-Moreno, A., Bartley, B., McLaughlin, J., Sánchez-Sampedro, L., Pascual Del Pozo, H., Prieto Hernández, C., Marletta, A. S., De Lucrezia, D., Sánchez-Fernández, G., Fraile, S., & de Lorenzo, V. (2020). SEVA 3.0: an update of the Standard European Vector Architecture for enabling portability of genetic constructs among diverse bacterial hosts. Nucleic acids research, 48(D1), D1164–D1170. [CrossRef]
- Fayed, B., Ashford, D. A., Hashem, A. M., Amin, M. A., El Gazayerly, O. N., Gregory, M. A., & Smith, M. C. (2015). Multiplexed integrating plasmids for engineering of the erythromycin gene cluster for expression in Streptomyces spp. and combinatorial biosynthesis. Applied and environmental microbiology, 81(24), 8402–8413. [CrossRef]
- Wang, W., Li, X., Wang, J., Xiang, S., Feng, X., & Yang, K. (2013). An engineered strong promoter for streptomycetes. Applied and environmental microbiology, 79(14), 4484–4492. [CrossRef]
- Seghezzi, N., Amar, P., Koebmann, B., Jensen, P. R., & Virolle, M. J. (2011). The construction of a library of synthetic promoters revealed some specific features of strong Streptomyces promoters. Applied microbiology and biotechnology, 90(2), 615–623. [CrossRef]
- Li, S., Wang, J., Li, X., Yin, S., Wang, W., & Yang, K. (2015). Genome-wide identification and evaluation of constitutive promoters in streptomycetes. Microbial cell factories, 14, 172. [CrossRef]
- Takano, E., White, J., Thompson, C. J., & Bibb, M. J. (1995). Construction of thiostrepton-inducible, high-copy-number expression vectors for use in Streptomyces spp. Gene, 166(1), 133–137. [CrossRef]
- Rodríguez-García, A., Combes, P., Pérez-Redondo, R., Smith, M. C., & Smith, M. C. (2005). Natural and synthetic tetracycline-inducible promoters for use in the antibiotic-producing bacteria Streptomyces. Nucleic acids research, 33(9), e87. [CrossRef]
- Horbal, L., Fedorenko, V., & Luzhetskyy, A. (2014). Novel and tightly regulated resorcinol and cumate-inducible expression systems for Streptomyces and other actinobacteria. Applied microbiology and biotechnology, 98(20), 8641–8655. [CrossRef]
- Phelan, R. M., Sachs, D., Petkiewicz, S. J., Barajas, J. F., Blake-Hedges, J. M., Thompson, M. G., Reider Apel, A., Rasor, B. J., Katz, L., & Keasling, J. D. (2017). Development of Next Generation Synthetic Biology Tools for Use in Streptomyces venezuelae. ACS synthetic biology, 6(1), 159–166. [CrossRef]
- Szafran, M. J., Gongerowska, M., Małecki, T., Elliot, M., & Jakimowicz, D. (2019). Transcriptional Response of Streptomyces coelicolor to Rapid Chromosome Relaxation or Long-Term Supercoiling Imbalance. Frontiers in microbiology, 10, 1605. [CrossRef]
- Na, D., & Lee, D. (2010). RBSDesigner: software for designing synthetic ribosome binding sites that yields a desired level of protein expression. Bioinformatics (Oxford, England), 26(20), 2633–2634. [CrossRef]
- Yi, J. S., Kim, M. W., Kim, M., Jeong, Y., Kim, E. J., Cho, B. K., & Kim, B. G. (2017). A Novel Approach for Gene Expression Optimization through Native Promoter and 5′ UTR Combinations Based on RNA-seq, Ribo-seq, and TSS-seq of Streptomyces coelicolor. ACS synthetic biology, 6(3), 555–565. [CrossRef]
- Hwang, S., Lee, N., Choe, D., Lee, Y., Kim, W., Kim, J. H., Kim, G., Kim, H., Ahn, N. H., Lee, B. H., Palsson, B. O., & Cho, B. K. (2022). System-Level Analysis of Transcriptional and Translational Regulatory Elements in Streptomyces griseus. Frontiers in bioengineering and biotechnology, 10, 844200. [CrossRef]
- Winkler W. C. (2005). Riboswitches and the role of noncoding RNAs in bacterial metabolic control. Current opinion in chemical biology, 9(6), 594–602. [CrossRef]
- Findeiß, S., Etzel, M., Will, S., Mörl, M., & Stadler, P. F. (2017). Design of Artificial Riboswitches as Biosensors. Sensors (Basel, Switzerland), 17(9), 1990. [CrossRef]
- Blin, K., Wolf, T., Chevrette, M. G., Lu, X., Schwalen, C. J., Kautsar, S. A., Suarez Duran, H. G., de Los Santos, E. L. C., Kim, H. U., Nave, M., Dickschat, J. S., Mitchell, D. A., Shelest, E., Breitling, R., Takano, E., Lee, S. Y., Weber, T., & Medema, M. H. (2017). antiSMASH 4.0-improvements in chemistry prediction and gene cluster boundary identification. Nucleic acids research, 45(W1), W36–W41. [CrossRef]
- Wang, J., Lu, A., Liu, J., Huang, W., Wang, J., Cai, Z., & Zhao, G. (2019). iCatch: a new strategy for capturing large DNA fragments using homing endonucleases. Acta biochimica et biophysica Sinica, 51(1), 97–103. [CrossRef]
- Greunke, C., Duell, E. R., D’Agostino, P. M., Glöckle, A., Lamm, K., & Gulder, T. A. M. (2018). Direct Pathway Cloning (DiPaC) to unlock natural product biosynthetic potential. Metabolic engineering, 47, 334–345. [CrossRef]
- Basitta, P., Westrich, L., Rösch, M., Kulik, A., Gust, B., & Apel, A. K. (2017). AGOS: A Plug-and-Play Method for the Assembly of Artificial Gene Operons into Functional Biosynthetic Gene Clusters. ACS synthetic biology, 6(5), 817–825. [CrossRef]
- Li, L., Zhao, Y., Ruan, L., Yang, S., Ge, M., Jiang, W., & Lu, Y. (2015). A stepwise increase in pristinamycin II biosynthesis by Streptomyces pristinaespiralis through combinatorial metabolic engineering. Metabolic engineering, 29, 12–25. [CrossRef]
- Goryshin, I. Y., Naumann, T. A., Apodaca, J., & Reznikoff, W. S. (2003). Chromosomal deletion formation system based on Tn5 double transposition: use for making minimal genomes and essential gene analysis. Genome research, 13(4), 644–653. [CrossRef]
- Xu, J., Zhang, J., Zhuo, J., Li, Y., Tian, Y., & Tan, H. (2017). Activation and mechanism of a cryptic oviedomycin gene cluster via the disruption of a global regulatory gene, adpA, in Streptomyces ansochromogenes. The Journal of biological chemistry, 292(48), 19708–19720. [CrossRef]
- Herrmann, S., Siegl, T., Luzhetska, M., Petzke, L., Jilg, C., Welle, E., Erb, A., Leadlay, P. F., Bechthold, A., & Luzhetskyy, A. (2012). Site-specific recombination strategies for engineering actinomycete genomes. Applied and environmental microbiology, 78(6), 1804–1812. [CrossRef]
- Bu, Q. T., Li, Y. P., Xie, H., Wang, J., Li, Z. Y., Chen, X. A., Mao, X. M., & Li, Y. Q. (2020). Comprehensive dissection of dispensable genomic regions in Streptomyces based on comparative analysis approach. Microbial cell factories, 19(1), 99. [CrossRef]
- Cobb, R. E., Wang, Y., & Zhao, H. (2015). High-efficiency multiplex genome editing of Streptomyces species using an engineered CRISPR/Cas system. ACS synthetic biology, 4(6), 723–728. [CrossRef]
- Tong, Y., Charusanti, P., Zhang, L., Weber, T., & Lee, S. Y. (2015). CRISPR-Cas9 Based Engineering of Actinomycetal Genomes. ACS synthetic biology, 4(9), 1020–1029. [CrossRef]
- Tong, Y., Whitford, C. M., Robertsen, H. L., Blin, K., Jørgensen, T. S., Klitgaard, A. K., Gren, T., Jiang, X., Weber, T., & Lee, S. Y. (2019). Highly efficient DSB-free base editing for streptomycetes with CRISPR-BEST. Proceedings of the National Academy of Sciences of the United States of America, 116(41), 20366–20375. [CrossRef]
- Huang, H., Zheng, G., Jiang, W., Hu, H., & Lu, Y. (2015). One-step high-efficiency CRISPR/Cas9-mediated genome editing in Streptomyces. Acta biochimica et biophysica Sinica, 47(4), 231–243. [CrossRef]
- Zeng, H., Wen, S., Xu, W., He, Z., Zhai, G., Liu, Y., Deng, Z., & Sun, Y. (2015). Highly efficient editing of the actinorhodin polyketide chain length factor gene in Streptomyces coelicolor M145 using CRISPR/Cas9-CodA(sm) combined system. Applied microbiology and biotechnology, 99(24), 10575–10585. [CrossRef]
- Jia, H., Zhang, L., Wang, T., Han, J., Tang, H., & Zhang, L. (2017). Development of a CRISPR/Cas9-mediated gene-editing tool in Streptomyces rimosus. Microbiology (Reading, England), 163(8), 1148–1155. [CrossRef]
- Nidhi, S., Anand, U., Oleksak, P., Tripathi, P., Lal, J. A., Thomas, G., Kuca, K., & Tripathi, V. (2021). Novel CRISPR-Cas Systems: An Updated Review of the Current Achievements, Applications, and Future Research Perspectives. International journal of molecular sciences, 22(7), 3327. [CrossRef]
- Yeo, W. L., Heng, E., Tan, L. L., Lim, Y. W., Lim, Y. H., Hoon, S., Zhao, H., Zhang, M. M., & Wong, F. T. (2019). Characterization of Cas proteins for CRISPR-Cas editing in streptomycetes. Biotechnology and bioengineering, 116(9), 2330–2338. [CrossRef]
- Whitford, C. M., Gren, T., Palazzotto, E., Lee, S. Y., Tong, Y., & Weber, T. (2023). Systems Analysis of Highly Multiplexed CRISPR-Base Editing in Streptomycetes. ACS synthetic biology, 12(8), 2353–2366. [CrossRef]
- Li, K., Cai, D., Wang, Z., He, Z., & Chen, S. (2018). Development of an Efficient Genome Editing Tool in Bacillus licheniformis Using CRISPR-Cas9 Nickase. Applied and environmental microbiology, 84(6), e02608-17. [CrossRef]
- Zetsche, B., Heidenreich, M., Mohanraju, P., Fedorova, I., Kneppers, J., DeGennaro, E. M., Winblad, N., Choudhury, S. R., Abudayyeh, O. O., Gootenberg, J. S., Wu, W. Y., Scott, D. A., Severinov, K., van der Oost, J., & Zhang, F. (2017). Multiplex gene editing by CRISPR-Cpf1 using a single crRNA array. Nature biotechnology, 35(1), 31–34. [CrossRef]
- Pickar-Oliver, A., & Gersbach, C. A. (2019). The next generation of CRISPR-Cas technologies and applications. Nature reviews. Molecular cell biology, 20(8), 490–507. [CrossRef]
- Qiu, M., Li, Y., Ye, W., Zheng, X., & Wang, Y. (2021). A CRISPR/Cas9-mediated in situ complementation method for Phytophthora sojae mutants. Molecular plant pathology, 22(3), 373–381. [CrossRef]
- Beck, C., Gren, T., Ortiz-López, F. J., Jørgensen, T. S., Carretero-Molina, D., Martín Serrano, J., Tormo, J. R., Oves-Costales, D., Kontou, E. E., Mohite, O. S., Mingyar, E., Stegmann, E., Genilloud, O., & Weber, T. (2021). Activation and Identification of a Griseusin Cluster in Streptomyces sp. CA-256286 by Employing Transcriptional Regulators and Multi-Omics Methods. Molecules (Basel, Switzerland), 26(21), 6580. [CrossRef]
- Kim, D. R., & Kwak, Y. S. (2021). A Genome-Wide Analysis of Antibiotic Producing Genes in Streptomyces globisporus SP6C4. The plant pathology journal, 37(5), 503. [CrossRef]
- Lee, N. C., Larionov, V., & Kouprina, N. (2015). Highly efficient CRISPR/Cas9-mediated TAR cloning of genes and chromosomal loci from complex genomes in yeast. Nucleic acids research, 43(8), e55. [CrossRef]
Figure 1.
A scheme of the main interconnections of primary metabolic pathways in Streptomyces. Arrows indicate positive effects, while dashed arrows indicate negative effects. The dotted lines indicate an uncharacterized or indirect regulation (modified after [23]).
Figure 1.
A scheme of the main interconnections of primary metabolic pathways in Streptomyces. Arrows indicate positive effects, while dashed arrows indicate negative effects. The dotted lines indicate an uncharacterized or indirect regulation (modified after [23]).
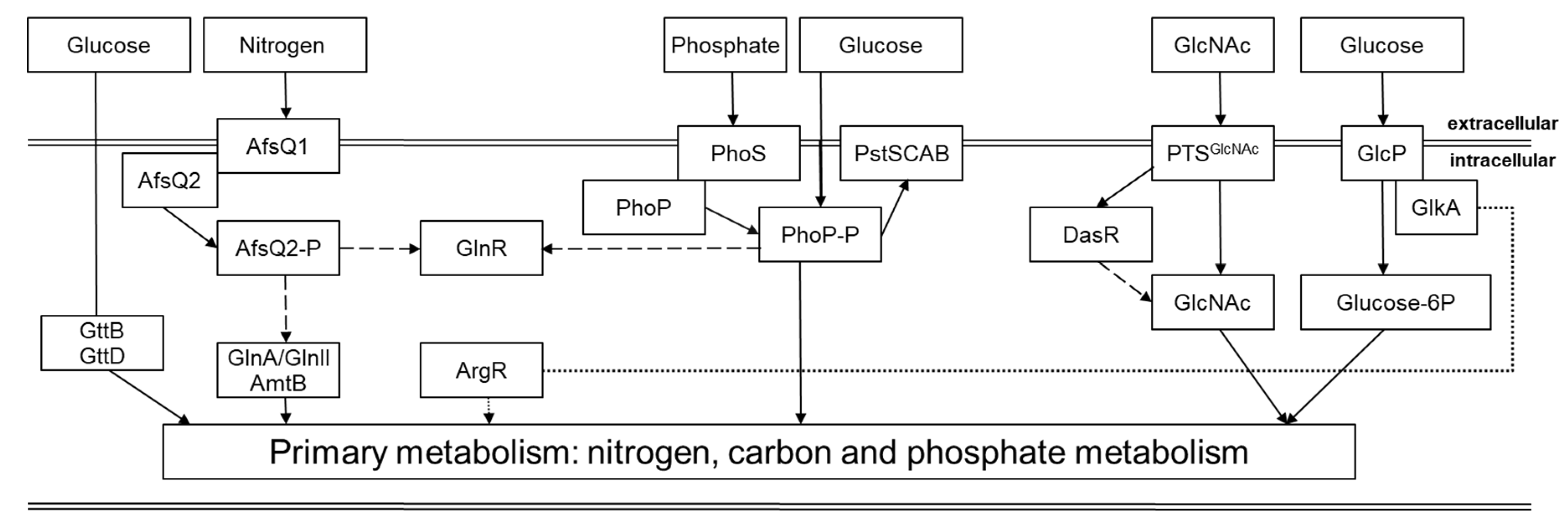
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



