
Submitted:
24 September 2024
Posted:
26 September 2024
You are already at the latest version
Abstract
Cereal crops are affected by one of the most devastating diseases worldwide, known as Fusarium head blight (FHB), with Fusarium graminearum being the most isolated causal pathogen. Another species associated to this disease is Fusarium poae. This species has been considered a relatively weak pathogen compared to F. graminearum, but its importance has increased for its occurrence in cereal grains worldwide. Considering the advantages of using B. distachyon as a plant model and the importance of F. poae in crops, our study aimed to evaluate the potential use of Brachypodium as a plant model to evaluate the compatible interaction with F. poae. For this, 12 Brachypodium spp. accessions from different countries were inoculated with a selected F. poae set of isolates. Disease severity, conidial quantification, fungal DNA biomass, and nivalenol quantification were assessed. The results showed a compatible interaction between Brachypodium accessions and Fusarium poae, which allowed the use of the model plant for future plant-pathogen interaction studies.
Keywords:
plant-pathogen interactions
; Fusarium poae
; disease severity
; Brachypodium spp
1. Introduction
In the past several years, shifting climates have increased the number of diseases caused by pathogens, which are the major contributors to worldwide health insecurity [1]. Crops with agronomically relevant traits, such as barley, wheat, oats, and maize, are affected by multiple phytopathogenic fungi [2,3,4,5]. One of the most harmful fungi in crops globally belongs to the genus Fusarium [6]. These species of fungi are globally distributed but are prevalent in particular localities depending on the climate patterns and cropping systems [7]. In Argentina, wheat and barley are the main large-area winter crops and are affected by one of the most devastating diseases worldwide, known as Fusarium head blight (FHB) [8,9]. Globally, among the most relevant species of the Fusarium genus, those belonging to the Fusarium sambucinum species complex (FSSC) are the most frequent in small grain cereals [10]. At the molecular level, this complex consists of at least 35 species, but Fusarium graminearum is the main disease responsible worldwide [11,12,13]. Fusarium spp. can infect several parts of the plants in different developmental stages [14]. Moreover, Fusarium graminearum is also responsible for other diseases such as crown rot, Fusarium root rot (FRR), and seedling blight [15,16]. Other Fusarium species associated with FHB worldwide are F. cerealis (syn. F. crookwellense), F. culmorum, F. poae, and F. pseudograminearum [17,18,19]. In wheat, symptoms are associated with the heads (ears) of the infected plants which develop bleaching spikes and often do not produce grains or, in some cases, produce shrivelled kernels with lightweight [20,21].
At present, F. poae isolates has become relevant for their occurrence in cereal grains [22]. This fungus has been considered a relatively weak pathogen compared to F. graminearum [23,24]. F. poae affects glumes or grains, but interest has increased because of the lack of visible symptoms on spikelet after infection [25]. Most recent studies show that this fungus is frequently isolated from barley grain in different parts of the world, with a high incidence in Argentina [25,26]. In addition, this pathogen can produce a large number of mycotoxins potentially harmful to the health of humans and animals [27]. These secondary metabolites are present in food and feed prepared from contaminated cereal grains [28]. Nivalenol (NIV) is one of the most important mycotoxins belonging to type B trichothecenes that can be produced by F. poae, making this pathogen even more relevant [29]. Other mycotoxins are type A trichothecenes, such as diacetoxiscirpenol (DAS) and neosolaniol (NEO) [21]. Moreover, several works reported the ability of F. poae to produce toxin T-2 and toxin HT-2. However, it has been recently demonstrated by Witte et al. [24] that the TRI16 gene responsible for T-2 and HT-2 production is not intact in F. poae isolates therefore concluding that this species cannot produce these harmful mycotoxins.
Plant–pathogen interaction studies are crucial to understanding how the plant pathogen accesses the resources necessary for its growth, reproduction, and survival within the host [30]. Therefore, hosts have developed different mechanisms to control their resistance/susceptibility to infection for pathogens [31]. In this field of research, cereal crops pose difficulty in studying interactions for their large complex genomes [32]. Furthermore, the size of these crops makes it difficult to work in laboratories [33]. Therefore, Brachypodium distachyon (L. P. Beauv.) has been proposed as a model pathosystem for research cereal crop diseases [34]. Genetic surveys have revealed that B. distachyon is more phylogenetically related to wheat and barley than other grasses such as rice or maize. Genomically, this synteny allows the transfer of Brachypodium information to Triticaceae genotypes [35]. B. distachyon is a monocot member of the Poaceae (Gramineae) family with characteristics of biological interest, including a small genome (approximately 271 Mbp), self-fertility, small stature, and a short generation time [36]. In the last decade, B. distachyon has emerged as a model plant for its capacity to host many cereal pathogens and develop symptoms of infection such as FHB [37,38]. It is known as the sequenced genome of accession Bd 21, a line sequence used as a reference [39,40]. Several researchers have shown these plant physiological and genetic advantages as a comparative and functional genetic model for agricultural studies [3,15,32,34].
Therefore, considering the advantages of using B. distachyon as a plant model and the importance of F. poae in crops, the present study aimed to evaluate the potential use of Brachypodium as a plant model to evaluate the compatible interaction with F. poae.
2. Materials and Methods
2.1. Fusarium poae Isolates Selection.
A total of eight isolates of F. poae were obtained from the fungal collection of the Instituto de Biología Funcional y Biotecnología (BIOLAB-Azul, Buenos Aires province, Argentina). These fungal isolates were first identified macro and microscopically [41] and were molecularly confirmed by using PCR species-specific primers [42]. Four F. poae isolates from barley grains were identified as 15-22.2-25.5-46.2 [26], and four isolates from wheat grains were identified as 37-40-43-47 [28]. Two accessions of Brachypodium were selected based on the susceptibility of roots to different Fusarium species, showing Bd-21 and Pakistan accessions more tolerance and susceptibility to F. poae, respectively [3]. Seeds were pre-germinated (ten seeds per accession) in square plastic Petri dishes containing filter paper on a damp surface of 0.8% water agar for approximately five days at 4°C. Subsequently, they were incubated vertically for ten days at 22°C under a photoperiod of 16 hrs light/8 hrs dark. A total of 30 roots with a similar length (three replicates per isolate per Brachypodium accession) under sterile conditions were inoculated. Inoculum was prepared using the protocol of Goddard et al. [43]. The Fusarium slurry was deposited 1 cm below of the roots using a small sterile syringe (0.1 a 5 mL) (Figure 1A). At two days post-inoculation (dpi), seedling roots were washed with sterile distilled water to remove the inoculum residues. Roots were photographed at 2, 4, 6, and 8 dpi (Figure 1B) to measure necrosis using ImageJ software [44]. The area under the disease curve (AUDPC) was calculated using the trapezoidal integration method [45], and the necrosis area on the root was expressed as a percentage of the necrosis area under the total root area. ANOVA analyses were performed, and the significance levels were established using Tukey tests at P<0.05.
2.2. Brachypodium Accessions
Brachypodium accessions used come from different parts of the world: tree diploid B. distachyon from United States of America (USA) (WA 36678), Turkey Adi-7 (WA 39240), and Bd-21 from Iraq (PI 254867) and nine hexaploid B. hybridum from Afghanistan (PI 219968), Australia (PI 533015), Iran (PI 239714), Iraq (PI 254868), Israel (PI 233228), Pakistan (PI 250647), South Africa (PI 208216), Spain (PI 287783), and Uruguay (PI 372187). For each trial, seeds of each Brachypodium accession were disinfected with sodium hypochlorite (0.6% v/v) for ten minutes with continuous shaking and three washes with sterile distilled water for ten minutes.
2.3. Pathogenicity Assay and F. poae Inoculation
A total of 5 seeds per 5 L pots filled with clay loam soil were sowed, irrigated, and drained to maintain humidity under greenhouse conditions where the minimum and maximum temperatures were registered daily. For inoculation, the selected F. poae isolate was cultured in Petri dishes containing 2% PDA and was grown at 25°C ± 2°C with 12h light/darkness. For the conidial harvest, 5 mL of distilled water was added on the PDA medium, and the conidia were taken with a bent glass rod. The suspension was filtered using cheesecloth, and the conidial suspension was adjusted to 1 x 105 conidia per mL [46], using a haemacytometer (Neubauer) and a binocular microscope. Tween® 20 (0.05%) (Biopack) was added as a surfactant. Floral point inoculation was made when spikelets reached anthesis (around 30-35 days after sowing). For this purpose, 3 µL of F. poae inoculum was applied in the central part of the floral cavity in the second spikelet starting from below. The control treatment was applied using water plus Tween® 20 (0.05%) (Biopack). After inoculation, plants were covered with polypropylene bags to improve fungal growth and humidity conditions. The design was completely randomised with eight replicates.
2.4. Disease Severity and Quantification of Fungal Sporulation
Disease severity (DS) was evaluated at 7, 14, and 21 days post inoculation (dpi) considering the following symptom score: 0 (no visible symptom), 1 (presence of symptom in the inoculated cavity of the spikelet), 2 (presence of symptom not only in the inoculated cavity but also in 1 or 2 neighbouring cavities of the spikelet), 3 (presence of symptom in 3 or more cavities continuous to the inoculated one), and 4 (the whole spikelet with symptoms and symptoms in adjacent spikelets) [47].
For conidial quantification, spikes were collected (five spikes per accession) at 7, 10, 14, and 21 days post inoculation (dpi) and were transferred in sterile Erlenmeyers containing 15 mL of sterile distilled water. Spikes were vigorously shaken for 30 min at 180 rpm in sterile distilled water to resuspend conidia. Finally, the obtaining suspension was filtered, and conidia were quantified (conidia/mL) using a Neubauer haemacytometer and a binocular microscope (Olympus CX 31®).
For DS and conidial quantification, ANOVA analyses were performed, and the significance levels were established using Tukey tests [48].
2.5. Fungal Genomic DNA Quantification
A total of 5 spikes with three replicates were collected at 21 dpi and placed in liquid nitrogen until DNA extraction [49]. The DNA quantity was calculated using a Fluorometer (Qubit Fluorometer, Invitrogen). A standard curve with pure F. poae DNA was built with known concentrations of 10 ng/µL to 0.001 ng/µL. Quantification of F. poae DNA was made by qPCR on 10 ng of total DNA using F. poae specific primers: FpoaeA51 fwd (5’-ACC GAA TCT CAA CTC CGC TTT-3’) and FpoaeA98 rev (5’-GTC TGT CAA GCA TGT TAG CAC AAG T-3’) [50]. PCR reactions were performed twice on an Applied Biosystems 7500 real-time PCR system (Thermo Fisher Scientific) using the following cycling protocol: 2 min at 50°C; 95°C 10 min; 40 cycles of 95°C for 15 s and 60°C for 1 min following by dissociation curve analysis at 60 to 95°C. SsoAdvancedTM Universal SYBR® Green Supermix (BIO-RAD) was used to reveal amplifications.
2.6. NIV Quantification
Brachypodium spikes at BBCH 97 were taken for NIV quantification and finely ground in a laboratory grinder (Arcano Fw-100, High-speed universal disintegrator) [51]. A volume of 20 mL of extraction solvent (CH3CN/H2O/HAc 79 + 20 + 1) was added to 1 g of ground Brachypodium spp. spikes. The samples were homogenised with the Ultraturrax for 3 min, sonicated for 60 min, and finally centrifuged for 5 min at 3000 rpm. A volume of 10 mL of extract was transferred into glass vials and evaporated to dryness at 45°C under a stream of N2. Samples were resuspended in methanol/water (70:30) and filtered through a 0.22-mm nylon filter before analysis. Nivalenol was identified and quantified using high-performance liquid chromatography coupled with tandem mass spectrometry (HPLC MS/MS) with a detection limit of 10 ng/g and a quantification limit of 30 ng/g [52]. The analyses were conducted in a Thermo Scientific ™ system consisting of a degasser, quaternary pump, column oven, and an LTQ XL™ ion trap mass spectrometer. Chromatographic separations were performed with a C18 100 x 2.1 mm Hypersil™ ODS (5 mm particle size) column. A solution of ammonium formate in acetonitrile (10 mM) was used as a mobile phase. Samples (10 mL) were analysed at a 0.2 mL/min flow rate at 45°C.
3. Results
3.1. Fusarium poae Selection
The selection of F. poae isolates was made based on Brachypodium root aggressiveness. The results showed different percentages of necrosis root area of all isolates on the average of the two Bd-21 and Pakistan Brachypodium spp. accessions (Figure 2). The isolate N° 47 from wheat was selected due to the higher aggressiveness in Brachypodium spp. root (4.30% ± 1.25%). The less aggressive isolate was N° 25.5 from barley (3.22% ± 1.23%). Although the differences among Bd-21 and Pakistan accessions were not statistically significant, Bd-21 (3.52% ± 1.67%) showed less necrotic root area caused by F. poae than Pakistan (3.66% ± 1.58%). Interestingly, although the differences were not statistically significant, the isolates from wheat (3.78% ± 1.65%) tended to be more aggressive than those obtained from barley (3.4% ± 1.51%).
3.2. Conidial Quantification and Disease Severity
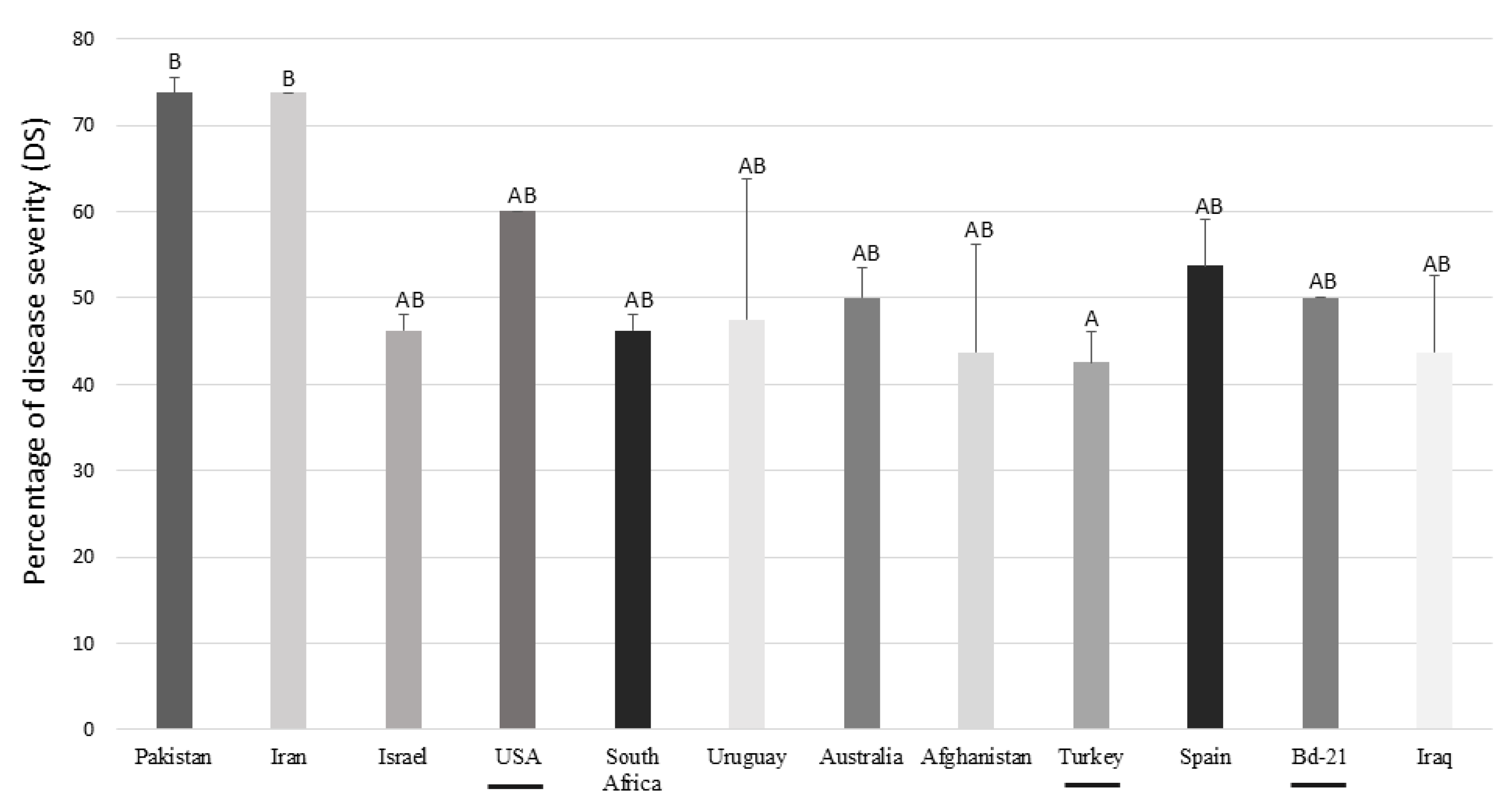
As regards DS, F. poae developed symptoms in all Brachypodium spp. evaluated (Figure 3). The accessions showed different behaviour against F. poae, showing the accession from Turkey the lowest DS values (42.5% ± 3.54%). On the other hand, the B. hybridum from Pakistan and Iran were the most affected lines (73.75% ± 1.77% and 73.75% ± 8.84%, respectively) (Figure 4). For the quantification of conidia, all suspensions from infected spikelets of Brachypodium spp. accessions presented conidia of F. poae independently of the day collected. However, statistical differences were observed in the days evaluated. At 7 dpi, the number of conidia was 4.83 ± 3.71 conidia/mL, at 14 dpi, it was 13.18 ± 7.13 conidia/mL, and finally, at 21 dpi, was 18.58 ± 12.71 conidia/mL.
3.3. Fusarium poae DNA and NIV Quantification
The results obtained from the analysis showed a significant difference between the Brachypodium accessions. Two groups were separated based on statistical differences: Pakistan and the remaining Brachypodium accessions. Pakistan showed the most important F. poae DNA quantity with a mean score of 2.82 ng ± 0.76 ng of F. poae DNA/10 ng of total DNA. Among the other group, Turkey was the accession that showed more F. poae DNA content (0.012 ng ± 0.01 ng), while USA accessions did not show F. poae DNA content. The analysis of mycotoxins shows that all the Brachypodium accessions showed no detectable NIV quantities (>10 ng/g).
4. Discussion
In simple practice, Brachypodium spp. has been recognized as an emerging system model. It is a small plant that is easy to maintain, but several important cereal diseases can develop, resulting in useful aspects for research [53]. Conversely, F. poae has been isolated with a high frequency in cereal grains worldwide [23,26,54,55]. Likewise, another study revealed that F. poae was the major fungal from wheat samples originating from Poland [56]. Currently, no information is available on the impact of F. poae on the Brachypodium spp. Therefore, our work aimed to determine the interaction of Brachypodium-F. poae to be useful for future plant-pathogen interaction studies.
For these studies, F. poae isolates from different crops (barley and wheat) were used. Our results demonstrate that the F. poae isolates were aggressive against Bd-21 and Pakistan Brachypodium roots. However, the isolates obtained from wheat showed more aggressiveness compared with those obtained from barley. Moreover, the isolate N°47 from wheat presented significant differences in the necrosis of the roots of the two accessions of Brachypodium, being selected for pathogenicity assay. Several works have shown that F. poae is an FHB pathogen affecting both crops, although barley is less frequent than wheat [22,26]. Differences in temporal and spatial flowering patterns among crops could be responsible for the differences observed [23]. In wheat, conidia of Fusarium spp. are deposited on or inside wheat spikes, germinating and initiating infection [57,58]. However, in barley, the fungal spreads from the exterior of the spikes under wet conditions, and internal spread in the rachis is more limited [59]. These could explain the differences in aggressiveness observed among isolates.
Regarding Brachypodium-Fusarium poae pathogenicity assays, the inoculation in floral tissues as a classical method for evaluation of the aggressiveness of Fusarium spp. was used [60]. In our study, this technique allowed the development of symptoms in all the accessions inoculated with F. poae. Moreover, the conidial quantification and fungal DNA biomass showed different results among accessions, which could indicate that some accessions respond differently to the pathogen presence. It would be interesting to evaluate the expression of defense genes among accessions inoculated with the pathogen to know the differences in responses observed. Compatible interaction among Brachypodium and different fungal pathogenic species has been confirmed. For example, the potential use of Brachypodium as a plant model for discovering genetic variation in resistance to Rhizoctonia solani was demonstrated [61]. Also, Brachypodium was used as a model plant against Puccinia graminis [62]. Moreover, various reported studies showed compatible interactions between Brachypodium and Claviceps purpurea, Ramularia collo-cygni, Oculimacula spp., Magnaporthe grisea, Cochliobolus sativus, Gaeumannomyces graminis, Pyrenophora teres, Stagonospora nodorum, and Colletotrichum cereale [34,63,64].
Regarding Fusarium species, F. graminearum, F. cerealis, F. pseudograminearum, and F. poae showed interaction with Brachypodium roots [3]. Similarly, the interaction between Brachypodium and the most prevalent species of Fusarium in Europe: F. graminearum, and F. culmorum, has been evaluated [15]. Their results showed not only the capacity of these species to develop symptoms on root, coleoptile, and foliar tissues but also that the plant model exhibited characteristics of susceptibility similar to those of wheat. Likewise, transcriptomic and metabolomic assays to evaluate the interaction between Brachypodium and two isolates of F. graminearum: a wild-type producing Deoxynivalenol (DON) mycotoxin and another impaired in DON production was developed [14]. The results showed an extensive colonisation of the pathogen over Brachypodium. Moreover, researchers demonstrated the capacity of DON to act as a virulence factor, as the isolate producing the mycotoxin was more aggressive than the isolate impaired in the production of DON. Fusarium poae cannot produce DON, but this species can produce NIV. In our study, the samples evaluated did not have detectable NIV quantities. The temperature range favouring the production of NIV by F. poae was between 25°C and 35°C (the estimated optimum being 27.5°C), which is different from the growth temperature of the fungus [65]. The mean minimum and maximum temperatures registered in our assay were 15.46 and 34.80°C, respectively. Moreover, previous studies revealed that stress responses, including abiotic and biotic factors such as pH, temperature, light, and interaction with other microorganisms, can cause oxidative bursts [66]. As a result, fungi release secondary metabolites called mycotoxin as defence agents. These factors could explain the absence of NIV in the samples evaluated despite the development of the fungus. Moreover, its role as a virulence factor has been discussed, assessing the effect of trichothecenes in the virulence of the pathogen [67]. For this objective, the gene coding for the initial enzyme of trichothecenes biosynthesis (Tri5 trichodiene synthase) was disrupted in three F. graminearum isolates with different mycotoxin production profiles. These isolates were used as inoculum to pathogenicity assays in wheat, barley, and maize. The results showed that the role of the trichothecenes varied depending on the crop evaluated. In wheat, the Tri5 disruption significantly reduced the F. graminearum virulence, while in barley, the decrease was not statistically significant. Interestingly, NIV is a virulence factor in maize, where the absence of NIV production reduces disease severity. In Brachypodium, as described before, DON acts as a virulence factor, but the role of NIV in this crop has not been described yet [14].
5. Conclusion
Our results demonstrated that the model plant proposed Brachypodium spp. could be useful for plant-F. poae interactions. Moreover, Brachypodium accessions responded differently against F. poae. It would be interesting for future work to elucidate if the Brachypodium responses correspond to some defence genes that activate in pathogen presence in some accessions that could explain better the differential behaviour against F. poae.
Funding
This research was funded by Fondo para la Investigación Científica y Tecnológica (FONCYT) - Proyectos de Investigación Científica y Tecnológica (PICT), grant number 00731/2020 and Fundación Williams. .
Data Availability Statement
The original contributions presented in the study are included in the article, further inquiries can be directed to the corresponding authors.
Acknowledgments
We thank Cristian Rodriguez for technical assistance in the plant-pathogen assays.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Catalan, P.; Chalhoub, B.; Chochois, V.; Garvin, D.F.; Hasterok, R.; Manzaneda, A.J.; et al. Update on the genomics and basic biology of Brachypodium: International Brachypodium Initiative (IBI). Trends Plant Sci. 2014, 19, 414–418. [Google Scholar] [CrossRef] [PubMed]
- Alberione, E.J. Sanitary update on wheat in Argentina. J. Plant Sci. Phytopathol. 2022, 6, 087–090. [Google Scholar]
- Dinolfo, M.I.; Martínez, M.; Nogueira, M.S.; Nicholson, P.; Stenglein, S.A. Evaluation of interaction between Brachypodium distachyon roots and Fusarium species. Eur. J. Plant Pathol. 2021, 159, 269–278. [Google Scholar] [CrossRef]
- Ng, C.A.; Poštulková, M.; Matoulková, D.; Psota, V.; Hartman, I.; Branyik, T. Methods for suppressing Fusarium infection during malting and their effect on malt quality. Czech J. Food Sci. 2021, 39, 340–359. [Google Scholar] [CrossRef]
- Pereyra, S.; Dill-Macky, R. Fusarium species recovered from wheat and barley grains in Uruguay, pathogenicity and deoxynivalenol content. Agrociencia Uruguay 2010, 14, 33–44. [Google Scholar] [CrossRef]
- Imathiu, S.M.; Edwards, S.G.; Ray, R.V.; Back, M.A. Fusarium langsethiae–a HT-2 and T-2 toxins producer that needs more attention. J. Phytopathol. 2013, 161, 1–10. [Google Scholar] [CrossRef]
- Ma, Z.; Xie, Q.; Li, G.; Jia, H.; Zhou, J.; Kong, Z.; et al. Germplasms, genetics and genomics for better control of disastrous wheat Fusarium head blight. Theor. Appl. Genet. 2020, 133, 1541–1568. [Google Scholar] [CrossRef] [PubMed]
- Albuquerque, D.R.; Patriarca, A.; Pinto, V.F. Can discrepancies between Fusarium graminearum trichothecene genotype and chemotype be explained by the influence of temperature in the relative production of 3-ADON and 15-ADON. Fungal Biol. 2021, 125, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Castañares, E.; Albuquerque, D.R.; Dinolfo, M.I.; Pinto, V.F.; Patriarca, A.; Stenglein, S.A. Trichothecene genotypes and production profiles of Fusarium graminearum isolates obtained from barley cultivated in Argentina. Int. J. Food Microbiol. 2014, 179, 57–63. [Google Scholar] [CrossRef]
- Crous, P.W.; Lombard, L.; Sandoval-Denis, M.; Seifert, K.A.; Schroers, H.J.; Chaverri, P.; et al. Fusarium: more than a node or a foot-shaped basal cell. Stud. Mycol. 2021, 98, 100–116. [Google Scholar] [CrossRef]
- Goswami, R.S.; Kistler, H.C. Heading for disaster: Fusarium graminearum on cereal crops. Mol. Plant Pathol. 2004, 5, 515–525. [Google Scholar] [CrossRef]
- Kazan, K.; Gardiner, D.M. Transcriptomics of cereal–Fusarium graminearum interactions: what we have learned so far. Mol. Plant Pathol. 2018, 19, 764–778. [Google Scholar] [CrossRef] [PubMed]
- Kikot, G.E.; Moschini, R.; Consolo, V.F.; Rojo, R.; Salerno, G.; Hours, R.A.; et al. Occurrence of different species of Fusarium from wheat in relation to disease levels predicted by a weather-based model in Argentina pampas region. Mycopathologia 2011, 171, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Pasquet, J.C.; Chaouch, S.; Macadré, C.; Balzergue, S.; Huguet, S.; Martin-Magniette, M.L.; Bellvert, F.; Deguercy, X.; Thareau, V.; Heintz, D.; Saindrenan, P.; et al. Differential gene expression and metabolomics analyses of Brachypodium distachyon infected by deoxynivalenol producing and non-producing strains of Fusarium graminearum. BMC Genom. 2014, 15, 629. [Google Scholar] [CrossRef] [PubMed]
- Peraldi, A.; Beccari, G.; Steed, A.; Nicholson, P. Brachypodium distachyon: A new pathosystem to study Fusarium head blight and other Fusarium diseases on wheat. BMC Plant Biol. 2011, 11, 100. [Google Scholar] [CrossRef]
- Snijders, C.H.A. Resistance in wheat to Fusarium infection and trichothecene formation. Toxicology Letters 2004, 153(1), 37–46. [Google Scholar] [CrossRef]
- Boughalleb, N.; Souli, M.; Ouled Sghaier, H. Occurrence and geographic distribution of wheat Fusarium head blight and Fusarium root rot in Jendouba’s areas of Tunisia. African J. Plant Sci. Biotechnol. 2008, 2, 23–26. [Google Scholar]
- Stenglein, S.A. Fusarium poae a pathogen that needs more attention. J. Plant Pathol. 2009, 91, 25–36. [Google Scholar]
- Van Coller, G.J.; Rose, L.J.; Boutigny, A.L.; Ward, T.J.; Lamprecht, S.C.; Viljoen, A. The distribution and type B trichothecene chemotype of Fusarium species associated with head blight of wheat in South Africa during 2008 and 2009. PLoS ONE 2022, 17, e0275084. [Google Scholar] [CrossRef]
- Birr, T.; Hasler, M.; Verreet, J.A.; Klink, H. Composition and predominance of Fusarium species causing Fusarium head blight in winter wheat grain depending on cultivar susceptibility and meteorological factors. Microorganisms 2020, 8, 617. [Google Scholar] [CrossRef]
- Martínez, M.; Arata, A.F.; Lázaro, L.; Stenglein, S.A.; Dinolfo, M.I. Effects of waterlogging stress on plant-pathogen interaction between Fusarium poae and wheat/barley. Acta Sci.-Agron. 2019, 41, e42629. [Google Scholar] [CrossRef]
- Martínez, M.; Ramírez Albuquerque, L.D.; Arata, A.F.; Biganzoli, F.; Fernández Pinto, V.; Stenglein, S.A. Effects of Fusarium graminearum and Fusarium poae on disease parameters, grain quality and mycotoxins contamination in bread wheat (part I). J. Sci. Food Agric. 2020, 100, 863–873. [Google Scholar] [CrossRef] [PubMed]
- Stenglein, S.A.; Dinolfo, M.I.; Barros, G.; Bongiorno, F.; Chulze, S.; Moreno, M.V. Fusarium poae pathogenicity and mycotoxin accumulation on selected wheat and barley genotypes at a single location in Argentina. Plant Dis. 2014, 98, 1733–1738. [Google Scholar] [CrossRef]
- Witte, T.E.; Hicks, C.; Hermans, A.; Shields, S.; Overy, D.P. Debunking the myth of Fusarium poae T-2/HT-2 toxin production. J. Agric. Food Chem. 2024, 72, 3949–3957. [Google Scholar] [CrossRef] [PubMed]
- Barreto, D.; Carmona, M.; Ferrazini, M.; Zanelli, M.; Perez, B.A. Occurrence and pathogenicity of Fusarium poae in barley in Argentina. Cereal Res. Commun. 2004, 32, 53–60. [Google Scholar] [CrossRef]
- Nogueira, M.S.; Decundo, J.; Martínez, M.; Dieguez, S.N.; Moreyra, F.; Moreno, M.V.; et al. Natural contamination with mycotoxins produced by Fusarium graminearum and Fusarium poae in malting barley in Argentina. Toxins 2018, 10, 78. [Google Scholar] [CrossRef] [PubMed]
- Dinolfo, M.I.; Castañares, E.; Stenglein, S.A. Characterization of a Fusarium poae world-wide collection by using molecular markers. Eur. J. Plant Pathol. 2014, 140, 119–132. [Google Scholar] [CrossRef]
- Dinolfo, M.I.; Stenglein, S.A.; Moreno, M.V.; Nicholson, P.; Jennings, P.; Salerno, G.L. ISSR markers detect high genetic variation among Fusarium poae isolates from Argentina and England. Eur. J. Plant Pathol. 2010, 127, 483–491. [Google Scholar] [CrossRef]
- Stenglein, S.A.; Dinolfo, M.I.; Bongiorno, F.; Moreno, M.V. Response of wheat (Triticum spp.) and barley (Hordeum vulgare) to Fusarium poae. Agrociencia 2012, 46, 46–299. [Google Scholar]
- Chhabra, B.; Tiwari, V.; Gill, B.S.; Dong, Y.; Rawat, N. Discovery of a promising susceptibility factor for Fusarium head blight in wheat. bioRxiv 2020. [Google Scholar]
- Campos, M.D.; Patanita, M.; Varanda, C.; Materatski, P.; Felix, M.D.R. Plant-pathogen interaction. Biology 2021, 10, 444. [Google Scholar] [CrossRef] [PubMed]
- Zhong, S.; Ali, S.; Leng, Y.; Wang, R.; Garvin, D.F. Brachypodium distachyon-Cochliobolus sativus pathosystem is a new model for studying plant-pathogen interactions in cereal crops. Phytopathology 2015, 105, 482–489. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, A.; Donn, S.; Ryan, P.R.; Mathesius, U.; Devilla, R.; Jones, A.; et al. Microbiome and exudates of the root and rhizosphere of Brachypodium distachyon, a model for wheat. PLoS ONE 2016, 11, e0164533. [Google Scholar] [CrossRef] [PubMed]
- Peraldi, A.; Griffe, L.L.; Burt, C.; McGrann, G.R.D.; Nicholson, P. Brachypodium distachyon exhibits compatible interactions with Oculimacula spp. and Ramularia collo-cygni, providing the first pathosystem model to study eyespot and ramularia leaf spot diseases. Plant Pathol. 2014, 63, 554–562. [Google Scholar] [CrossRef] [PubMed]
- Verelst, W.; Bertolini, E.; De Bodt, S.; Vandepoele, K.; Demeulenaere, M.; Pè, M.E.; et al. Molecular and physiological analysis of growth-limiting drought stress in Brachypodium distachyon leaves. Mol. Plant 2013, 6, 311–322. [Google Scholar] [CrossRef]
- Luo, N.; Liu, J.; Yu, X.; Jiang, Y. Natural variation of drought response in Brachypodium distachyon. Physiol. Plant. 2011, 141, 19–29. [Google Scholar] [CrossRef]
- Fitzgerald, T.L.; Powell, J.J.; Schneebeli, K.; Hsia, M.M.; Gardiner, D.M.; Bragg, J.N.; et al. Brachypodium as an emerging model for cereal–pathogen interactions. Ann. Bot. 2015, 115, 717–731. [Google Scholar] [CrossRef]
- Pasquet, J.C.; Changenet, V.; Macadré, C.; Boex-Fontvieille, E.; Soulhat, C.; Bouchabké-Coussa, O.; et al. A Brachypodium UDP-glycosyltransferase confers root tolerance to deoxynivalenol and resistance to Fusarium infection. Plant Physiol. 2016, 172, 559–574. [Google Scholar] [CrossRef]
- Des Marais, D.L.; Razzaque, S.; Hernandez, K.M.; Garvin, D.F.; Juenger, T.E. Quantitative trait loci associated with natural diversity in water-use efficiency and response to soil drying in Brachypodium distachyon. Plant Sci. 2016, 251, 2–11. [Google Scholar] [CrossRef]
- Rancour, D.M.; Marita, J.M.; Hatfield, R.D. Cell wall composition throughout development for the model grass Brachypodium distachyon. Front. Plant Sci. 2012, 3, 266. [Google Scholar] [CrossRef]
- Leslie, J.F.; Summerell, B.A. (Eds.) The Fusarium Laboratory Manual; Blackwell Publishing, Oxford, UK, 2006.
- Parry, D.W.; Nicholson, P. Development of a PCR assay to detect Fusarium poae in wheat. Plant Pathol. 1996, 45, 383–391. [Google Scholar] [CrossRef]
- Goddard, R.; Peraldi, A.; Ridout, C.; Nicholson, P. Enhanced disease resistance caused by BRI1 mutation is conserved between Brachypodium distachyon and barley (Hordeum vulgare). Mol. Plant-Microbe Interact. 2014, 27, 1095–1106. [Google Scholar] [CrossRef] [PubMed]
- Abramoff, M.D.; Magalhães, P.J.; Ram, S.J. Image processing with ImageJ. Biophotonics Int. 2004, 11, 36–42. [Google Scholar]
- Introduction to Plant Disease Epidemiology; Jhon Willey & Sons. New York, USA, 1990. Campbell, C.L.; Madden, L.V. (Eds.).
- Brennan, J.M.; Leonard, G.; Fagan, B.; Cooke, B.M.; Ritieni, A.; Ferracane, R.; Nicholson, P.; Simpson, D.; Thomsett, M.; et al. Comparison of commercial European wheat cultivars to Fusarium infection of head and seedling tissue. Plant Pathol. 2007, 56, 55–64. [Google Scholar] [CrossRef]
- Chen, J.; Griffey, C.A.; Saghai Maroof, M.A.; Stromberg, E.L.; Biyashev, R.M.; Zhao, W.; et al. Validation of two major quantitative trait loci for Fusarium head blight resistance in Chinese wheat line W14. Plant Breed. 2006, 125, 99–101. [Google Scholar] [CrossRef]
- Di Rienzo, J.; Balzarini, M.; Gonzalez, L.; Casanoves, F.; Tablada, M.; Robledo, C. W. Infostat: software para análisis estadístico; Universidad Nacional de Córdoba, Argentina, 2012.
- Atoui, A.; Khoury, A.E.; Kallassy, M.; Lebrihi, A. Quantification of Fusarium graminearum and Fusarium culmorum by real-time PCR system and zearalenone assessment in maize. Int. J. Food Microbiol. 2012, 154, 59–65. [Google Scholar] [CrossRef]
- Nicolaisen, M.; Suproniene, S.; Nielsen, L.K.; Lazzaro, I.; Spliid, N.H.; Justesen, A.F. Real-time PCR for quantification of eleven individual Fusarium species in cereals. J. Microbiol. Meth. 2009, 76, 234–240. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.Y.; Park, J.H.; Cho, S.H.; Yang, M.S.; Park, C.M. Phenological growth stages of Brachypodium distachyon: codification and description. Weed Res. 2011, 51, 612–620. [Google Scholar] [CrossRef]
- Arroyo-Manzanares, N.; Huertas-Pérez, J.F.; Gámiz-Gracia, L.; García-Campaña, A. M. Simple and efficient methodology to determine mycotoxins in cereal syrups. Food Chem. 2015, 177, 274–279. [Google Scholar] [CrossRef]
- Draper, J.; Mur, L.A.; Jenkins, G.; Ghosh-Biswas, G.C.; Bablak, P.; Hasterok, R.; et al. Brachypodium distachyon. A new model system for functional genomics in grasses. Plant Physiol. 2001, 127, 1539–1555. [Google Scholar] [CrossRef]
- Beccari, G.; Arellano, C.; Covarelli, L.; Tini, F.; Sulyok, M.; Cowger, C. Effect of wheat infection timing on Fusarium head blight causal agents and secondary metabolites in grain. Int. J. Food Microbiol. 2019, 290, 214–225. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.M.; Parry, D.W.; Nicholson, P.; Thomsett, M.A.; Simpson, D.; Edwards, S.G.; et al. Predominance and association of pathogenic fungi causing Fusarium ear blight in wheat in four European countries. Eur. J. Plant Pathol. 2005, 112, 143–154. [Google Scholar] [CrossRef]
- Kulik, T.; Jestoi, M. Quantification of Fusarium poae DNA and associated mycotoxins in asymptomatically contaminated wheat. Int. J. Food Microbiol. 2009, 130, 233–237. [Google Scholar] [CrossRef]
- Bai, G.; Shaner, G. Management and resistance in wheat and barley to Fusarium head blight. Annu. Rev. Phytopathol. 2004, 42, 135–161. [Google Scholar] [CrossRef] [PubMed]
- Tini, F.; Covarelli, L.; Cowger, C.; Sulyok, M.; Benincasa, P.; Beccari, G. Infection timing affects Fusarium poae colonization of bread wheat spikes and mycotoxin accumulation in the grain. J. Sci. Food Agric. 2022, 102, 6358–6372. [Google Scholar] [CrossRef] [PubMed]
- Janssen, E.M.; Liu, C.; Van der Fels-Klerx, H.J. Fusarium infection and trichothecenes in barley and its comparison with wheat. World Mycotoxin J. 2018, 11, 33–46. [Google Scholar] [CrossRef]
- Malbrán, I.; Mourelos, C.A.; Girotti, J.R.; Aulicino, M.B.; Balatti, P.A.; Lori, G.A. Aggressiveness variation of Fusarium graminearum isolates from Argentina following point inoculation of field grown wheat spikes. Crop Protec. 2012, 42, 234–243. [Google Scholar] [CrossRef]
- Schneebeli, K.; Mathesius, U.; Matt, M. , Brachypodium distachyon is a pathosystem model for the study of the wheat disease rhizoctonia root rot. Plant Pathol. 2015, 64, 91–100. [Google Scholar] [CrossRef]
- Figueroa, M.; Alderman, S.; Garvin, D.F.; Pfender, W.F. Infection of Brachypodium distachyon by formae speciales of Puccinia graminis: early infection events and host-pathogen incompatibility. PLoS ONE 2013, 8, e56857. [Google Scholar] [CrossRef]
- Routledge, A.P.; Shelley, G.; Smith, J.V.; Talbot, N.J.; Draper, J.; Mur, L.A. Magnaporthe grisea interactions with the model grass Brachypodium distachyon closely resemble those with rice (Oryza sativa). Mol. Plant Pathol. 2004, 5, 253–265. [Google Scholar] [CrossRef]
- Sandoya, G.V.; de Oliveira Buanafina, M.M. Differential responses of Brachypodium distachyon genotypes to insect and fungal pathogens. Physiol. Mol. Plant Pathol. 2014, 85, 53–64. [Google Scholar] [CrossRef]
- Nazari, L.; Pattori, E.; Manstretta, V.; Terzi, V.; Morcia, C.; Somma, S.; et al. Effect of temperature on growth, wheat head infection, and nivalenol production by Fusarium poae. Food Microbiol. 2018, 76, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Ponts, N. Mycotoxins are a component of Fusarium graminearum stress-response system. Front. Microbiol. 2015, 6, 133574. [Google Scholar] [CrossRef] [PubMed]
- Maier, F.J.; Miedaner, T.; Hadeler, B.; Felk, A.; Salomon, S.; Lemmens, M.; et al. Involvement of trichothecenes in fusarioses of wheat, barley and maize evaluated by gene disruption of the trichodiene synthase (Tri5) gene in three field isolates of different chemotype and virulence. Mol. Plant Pathol. 2006, 7, 449–461. [Google Scholar] [CrossRef]
Figure 1.
Selection of F. poae isolation based on the susceptibility of Brachypodium spp. roots. (A) Slurry of Fusarium poae on Pakistan accessions root to measure the necrotic area. (B) B. distachyon inoculated with F. poae showing necrosis symptoms.
Figure 1.
Selection of F. poae isolation based on the susceptibility of Brachypodium spp. roots. (A) Slurry of Fusarium poae on Pakistan accessions root to measure the necrotic area. (B) B. distachyon inoculated with F. poae showing necrosis symptoms.
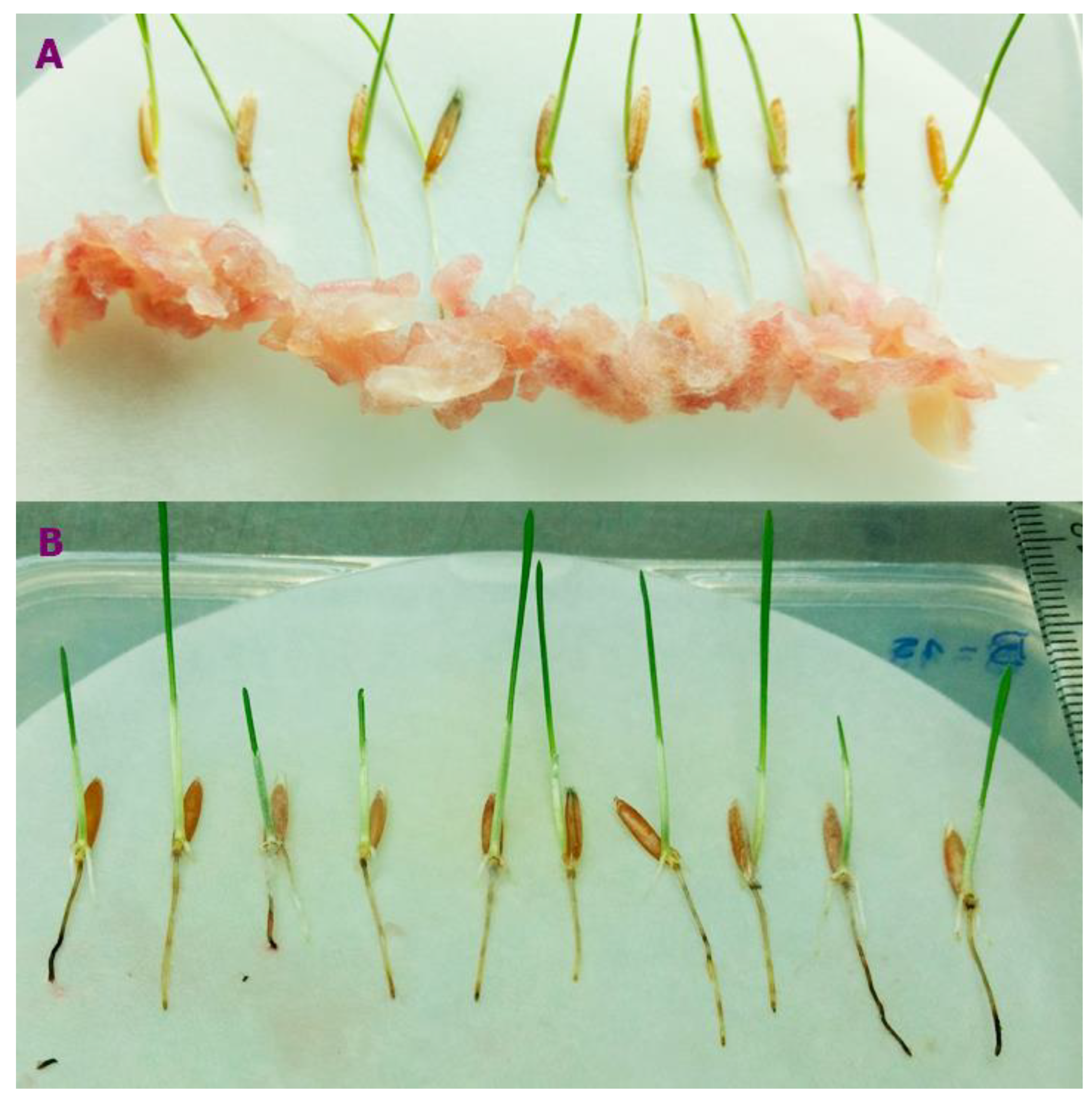
Figure 2.
(A) Different AUDPC of F. poae isolates from barley and wheat. (B) (C) Tissue necrosis (%) of F. poae isolates at 2, 4, 6, and 8 dpi isolates from barley and wheat, respectively. Different letters are statistically significant according to Tukey’s test at P < 0.05.
Figure 2.
(A) Different AUDPC of F. poae isolates from barley and wheat. (B) (C) Tissue necrosis (%) of F. poae isolates at 2, 4, 6, and 8 dpi isolates from barley and wheat, respectively. Different letters are statistically significant according to Tukey’s test at P < 0.05.
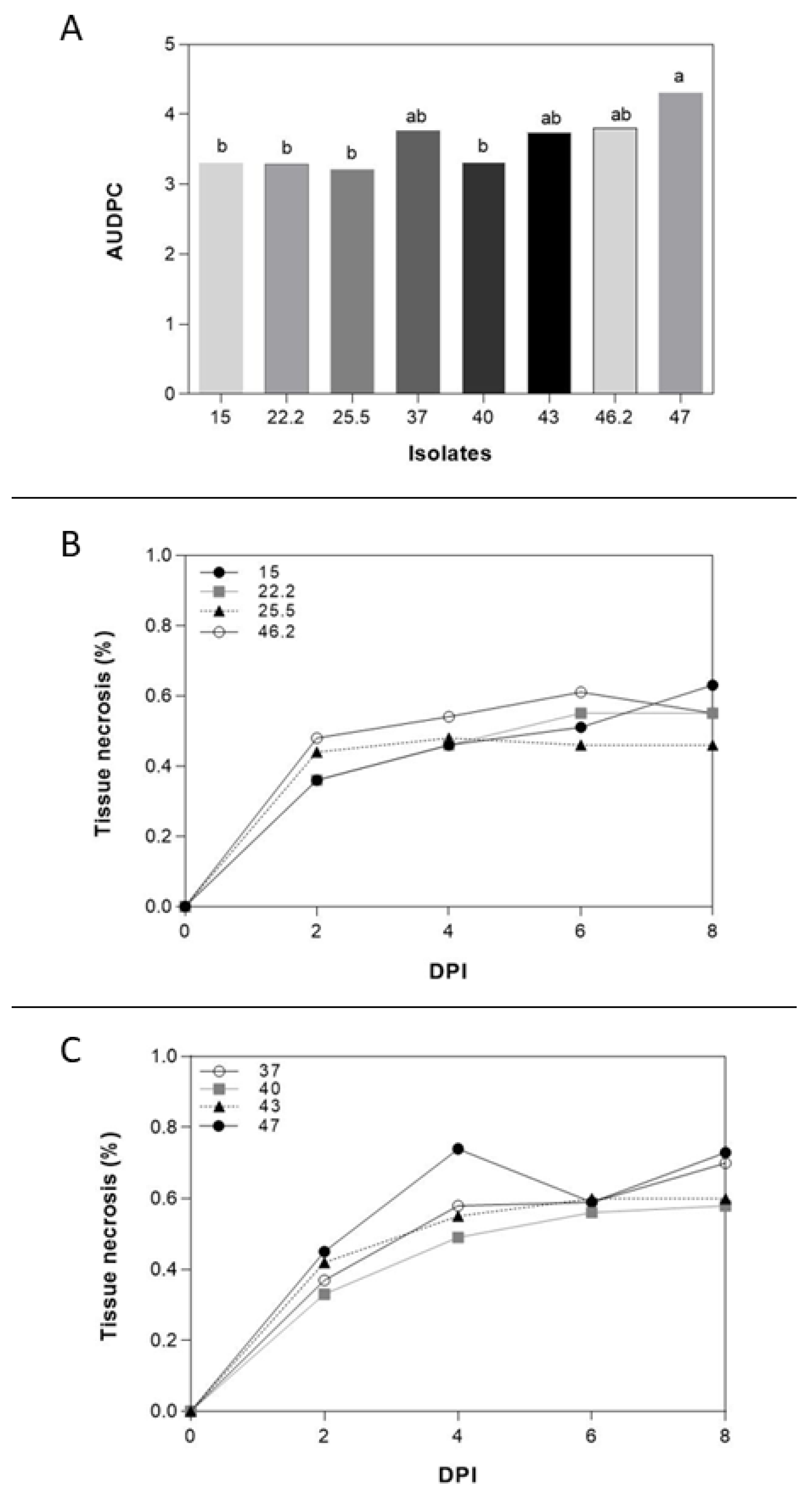
Figure 3.
Symptoms of F. poae developed on Brachypodium accessions. The diploid accessions were underlined.
Figure 3.
Symptoms of F. poae developed on Brachypodium accessions. The diploid accessions were underlined.

Figure 4.
Percentage of disease severity (DS) of Brachypodium accessions inoculated with F. poae. The diploid accessions were underlined. Different letters are statistically significant according to Tukey’s test at P < 0.10.
Figure 4.
Percentage of disease severity (DS) of Brachypodium accessions inoculated with F. poae. The diploid accessions were underlined. Different letters are statistically significant according to Tukey’s test at P < 0.10.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



