
Submitted:
25 September 2024
Posted:
29 September 2024
You are already at the latest version
Abstract
Circular RNA (circRNA) vaccines are emerging as a revolutionary strategy in cancer immunotherapy, offering novel mechanisms for inducing robust and durable immune responses. Unlike traditional linear mRNA vaccines, circRNAs exhibit exceptional stability, enhanced translational efficiency, and resistance to exonuclease degradation, making them ideal candidates for vaccine development. This review delves into the fundamental principles underlying circRNA biology, highlighting their unique structural advantages and translational potential. We examine recent advancements in circRNA vaccine design, focusing on their application in oncology. As the circRNA-based cancer vaccine is a relatively novel approach, accounts of all the major studies describing the efficacy have been discussed. To maximize the anti-cancer effects of circRNA vaccines, we also investigate their combination with other immunotherapeutic modalities such as immune checkpoint inhibitors and adoptive cell therapies. Large-scale manufacturing, immunogenicity optimization, delivery systems, and other challenges and future directions in the sector are also discussed. This study aims to thoroughly analyse the state-of-the-art and potential future applications of circRNA vaccines in cancer immunotherapy, highlighting them as exciting possibilities for next-generation cancer therapies.
Keywords:
circular RNA vaccines
; Cancer immunotherapy
; cancer vaccine
; vaccine delivery
; mRNA vaccine
1. Introduction
In the field of cancer therapeutics, the continuous effort to develop effective immunotherapeutic strategies persists, motivated by the need to overcome the limitations of current treatment methods. Despite the considerable advancements made in the field of cancer treatment, conventional therapies such as chemotherapy, radiation therapy, and immune checkpoint inhibitors (ICIs) continue to face challenges in terms of limited effectiveness, adverse effects, and the development of resistance. Immune checkpoint inhibitors (ICIs) are the essential components of cancer vaccines because they enhance the effectiveness of vaccine-induced anti-tumor immunity, strengthen the immune system's response, and assist in countering cancers' ability to evade the immune system. Although ICIs have transformed cancer treatment for some patients, a significant number show resistance or do not respond, emphasizing the necessity for alternative strategies to enhance immune activation and overcome tumor immune evasion mechanisms (Wang et al., 2023).
CircRNA-based cancer vaccines present a hopeful resolution to these obstacles by utilizing the stability and extended antigen expression found in their covalently closed-loop structures. (Niu et al., 2023) These vaccines have the potential to enhance patient outcomes and open up possibilities for personalized cancer immunotherapy by overcoming the drawbacks of current immunotherapy approaches.
Although possessing favorable characteristics, the domain of circRNA-based cancer vaccines encounters various research deficiencies and obstacles that necessitate resolution to unlock their complete capabilities (Xie et al., 2023). Key inquiries persist concerning the most effective configuration of circRNA structures, the processes driving immune stimulation, and the creation of effective distribution mechanisms. Furthermore, thorough preclinical and clinical verification is crucial to confirm the safety, effectiveness, and viability of circRNA-based vaccines for cancer immunotherapy (Chen et al., 2022).
The review provides an overview of circRNA vaccines in the context of cancer immunotherapy. It meticulously explores various facets, including the intricate design principles, diverse delivery modes, and the underlying mechanisms of immune activation. The current research findings will summarize the current situation of cancer immunotherapy and also reveal prospective pathways for future developments in using circRNA vaccines to battle cancer effectively.
2. Cancer Immunotherapy Landscape: Advances and Hurdles
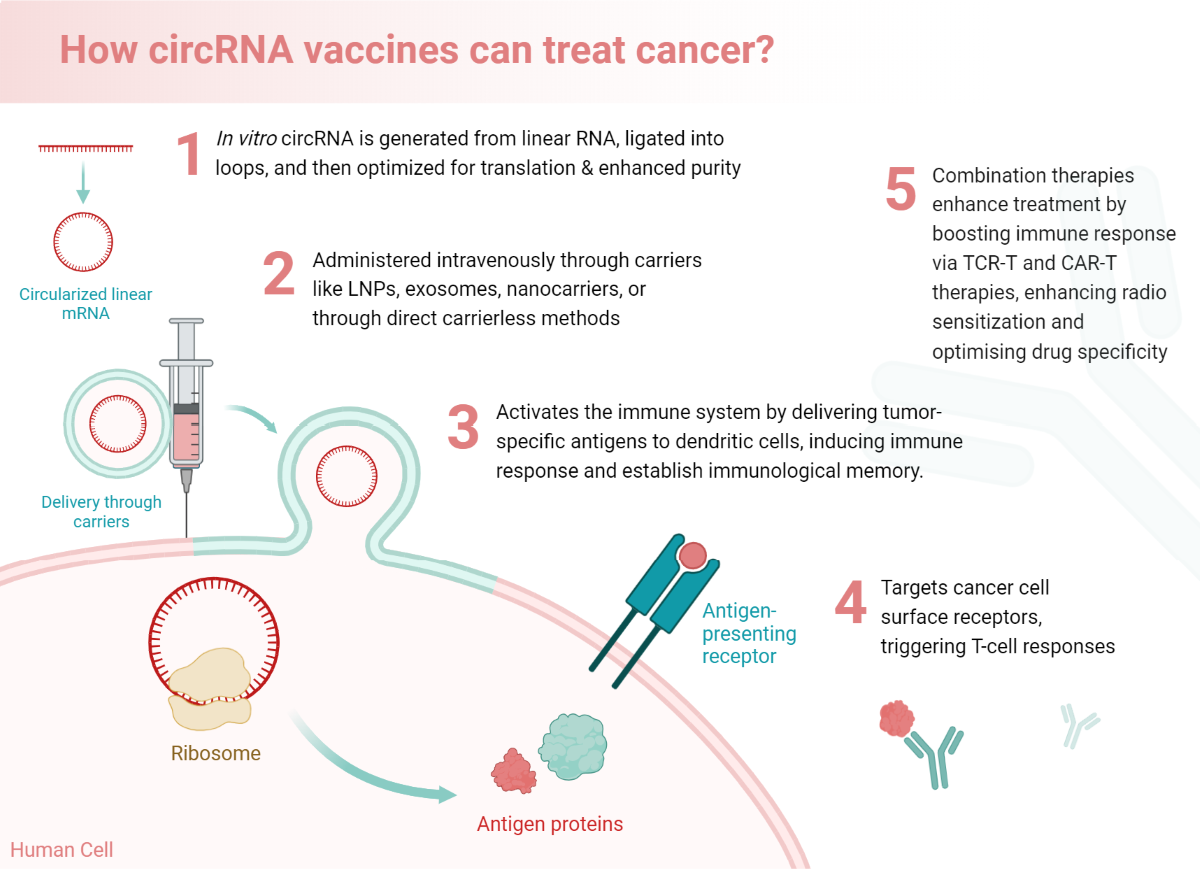
Throughout the timeline, of cancer immunotherapy development, we have seen remarkable progress, particularly with the advent of immune checkpoint inhibitors (ICIs) and CAR-T cell therapies. ICIs have transformed cancer treatment by enabling the immune system to target cancer cells more effectively, while CAR-T therapies have shown significant success in treating certain blood cancers. Additionally, cancer vaccines and adoptive cell transfer (ACT) techniques, such as tumor-infiltrating lymphocytes (TIL) therapy, are promising areas of development. A bried history of the history of advancements in cancer immunotherapy especially wrt circRNA vaccines is shown in Figure 1. However, the field faces several hurdles, including the immunosuppressive tumor microenvironment, manufacturing complexities, resistance mechanisms, and severe side effects. Overcoming these hurdles requires continuous research and innovation to enhance the efficacy and accessibility of immunotherapies, ultimately improving patient outcomes.
3. Role of Circ-RNAS in Cancer Progression and Therapeutic Resistance
In the field of cancer therapeutics, there are four main types of therapy - immune cell or tumor-derived, peptide-based, viral vector-based, and nucleic acid-based vaccines (Miao et al., 2021a).Several neoantigen vaccines, including DNA, mRNA, and synthetic long peptide (SLP)-based vaccines, are developing (Guo et al., 2018). However, on a broader translational approach, every other type has a few flaws that need to be fixed for medical implementation. To address the hurdles, the field has taken turns toward offering a more targeted, safe, and effective immunotherapy alternative, when circRNAs come into play as a potential cancer therapeutics approach.
CircRNAs contribute to therapeutic resistance by modulating stress responses and promoting cancer cell survival. Mechanisms of resistance include activating pro-survival signaling, altering drug metabolism, and evading apoptosis (Lei et al., 2023). Targeting circRNAs with RNA interference techniques, such as siRNAs or shRNAs, can sensitize cancer cells to chemotherapy and targeted therapies. Additionally, antisense oligonucleotides (AONs) and nanoparticle-based delivery systems enable precise modulation of circRNAs, potentially overcoming resistance (Xin et al., 2017; Mendes et al., 2022).
CircRNAs also interact with neoantigen presentation pathways, modulating immune recognition in cancer. However, their dysregulation is implicated in cancer progression, influencing key processes like cell proliferation, apoptosis evasion, angiogenesis, and epithelial-mesenchymal transition (EMT). CircPVT1, for instance, drives tumorigenesis in several cancer types by sponging microRNAs and modulating oncogenic pathways (Derderian et al., 2019). CircRNAs also play crucial roles in metastasis, enhancing migration, invasion, and colonization of distant organs by cancer cells (Yuan et al., 2020; Zhang et al., 2020b).
CircRNAs have emerged as valuable biomarkers for cancer diagnosis and prognosis. Their stable and tissue-specific expression and presence in bodily fluids make them ideal candidates for non-invasive cancer screening and monitoring (Su et al., 2019). High-throughput sequencing has identified circRNA biomarkers across various cancer types, with their integration into clinical assessments improving diagnostic accuracy (Dong et al., 2023b). A list of diverse types of circRNAs, their functions, design, delivery and examples is provided in Table S2.
CircRNAs also influence immunotherapy resistance. For example, targeting circUHRF1 enhances responses to anti-PD-1 immunotherapy in hepatocellular carcinoma by restoring T cell-mediated cytotoxicity and preventing immune escape (Zhang et al., 2020a). To accumulate the pros and cons, this is probable that CircRNA-targeted therapies can offer the potential to improve cancer immunotherapy outcomes soon.
4. Unlocking the Power of Circular RNAS over Mrnas: Advantages in Cancer Vaccines
The distinctive properties of circular RNAs (circRNAs) position them as particularly advantageous compared to commonly known long-coding RNAs, such as mRNAs, in the emerging field of therapeutics. Unlike mRNAs, circRNAs exhibit enhanced stability, resistance to exonuclease-mediated degradation, and unique regulatory functions, which underscore their potential as superior therapeutic agents. (Figure 1). First off, unlike eukaryotic mRNAs, which typically have five regions—the 5'-cap, 5'-UTR, open reading frame, 3'-UTR, and 3'-poly(A) tail—circRNAs lack the 5'-cap, 3'-poly (A) tail and have a closed loop structure which is devoid of the normal polarity (directionally). Second, most mRNAs in eukaryotic cells are translated by ribosome recruitment to the 5'-cap followed by ribosome scanning in the direction of a start codon known as cap-dependent translation. MRNAs can also attract ribosomes via IRES to facilitate translation in certain cases. However, circRNAs translate without the need for a cap. manner (Legnini et al., 2017; Pamudurti et al., 2017; Wang et al., 2015a; Y. Yang et al., 2018).
On a structural basis, circRNAs have a covalently closed loop structure that enhances their stability and longevity. This structure protects them from exonuclease-mediated degradation, as the absence of free ends prevents enzymatic cleavage. Additionally, circRNAs are resistant to other decay mechanisms, such as endonucleolytic cleavage, allowing them to persist longer in cells. Interactions with RNA-binding proteins (RBPs) further stabilize circRNAs, either through direct binding or secondary structure interactions.
Furthermore, circRNAs' translation may vary depending on the unique components (such as IRESs) in the circRNAs. Third, the three exonucleases that break down most mRNAs promote hydrolysis at the 3’ end, and 5’ exonucleases, which break down RNAs from the 5’ ends. Instead, only a few endonucleases are capable of breaking down circRNAs (Ren et al., 2022). Because of its circular form, circRNA is not easily broken down by RNase R enzymes or RNA exonucleases. Because of this, circRNA typically exhibits greater stability and a longer half-life (around 80-116 hours as opposed to 43-49 hours). Because of its enhanced stability, circRNA is a good option for several uses, especially in developing vaccinations against neoantigens (Chen et al., 2019b; Chen et al., 2020a; Ge et al., 2021; Li et al., 2021; Mo et al., 2022; Qu et al., 2022a).
CircRNA vaccines have the potential to produce larger amounts of antigens. Target antigen expression rises and persists longer as a result, which is essential for a successful immune response (Qu et al., 2022b). Since circRNA vaccinations do not integrate into the genome, there is no chance of gene disruption, making them safer than DNA vaccines. Because of circRNAs distinct structure, a particular translation mechanism is possible, allowing for the synthesis of bigger proteins with less genetic material (Chen et al., 2020a; X. Liu et al., 2022; Yue et al., 2021).
Exogenous circular RNAs (circRNAs) originate from external sources like pathogens and can affect cellular processes, including immune responses. When introduced into cells, exogenous circRNAs can trigger innate immunity, aiding in the elimination of pathogens and abnormal cells, such as cancer. Unlike traditional vaccines, circRNA vaccines can activate multiple immune pathways simultaneously, potentially making them more effective (Yang et al., 2021b).
CircRNAs have a covalently closed loop structure that enhances their stability and longevity. This structure protects them from exonuclease-mediated degradation, as the absence of free ends prevents enzymatic cleavage. Additionally, circRNAs are resistant to other decay mechanisms, such as endonucleolytic cleavage, allowing them to persist longer in cells. Interactions with RNA-binding proteins (RBPs) further stabilize circRNAs, either through direct binding or secondary structure interactions. The competitive structural advantage of circRNA vaccines over mRNA vaccines is illustrated in Figure 2.

Table 1.
Advantages and drawbacks of different types of vaccines.
| Vaccines | Advantages | Disadvantages | References |
|---|---|---|---|
| SLP (Synthetic long peptide) Vaccine | Stable The long peptide can stimulate the response of CD4+ and CD8+ T cells |
Complex synthesis in vitro, high cost Weak anti-tumor immune response Potential adverse reactions. |
Chen et al., 2021b; Guo et al., 2018; Jou et al., 2021; Supabphol et al., 2021; Wei et al., 2021 |
| DNA Vaccine (Plasmid DNA encoding a new tumor antigen) |
Production process is simple, rapid, and low-cost Virus-like infection mechanism Allows the modification of coding antigens |
Risk of genomic integration | Chen et al., 2019b; Li et al., 2021; Supabphol et al., 2021; X. Yang et al., 2021b |
| mRNA Vaccine (mRNA transcribed in vitro encoding neoantigens) |
Production process is simple and rapid, and low-cost Translation without entering the nucleus No risk of genome integration Self-adjuvant effect. |
Poor stability Lack of effective transfection method for nuclease susceptibility. |
Barbier et al., 2022; Beck et al., 2021; Cafri et al., 2020; Chakraborty et al., 2021; Guo et al., 2018; Huang et al., 2021; Huff et al., 2022; Jain et al., 2021; Miao et al., 2021a; Qu et al., 2022b; Xu et al., 2020 |
| circRNA Vaccine (CircRNA encoding new tumor antigen constructed in vitro) | Good stability Simple and rapid production process with low-cost Translating without entering the nucleus High antigen expression efficiency No risk of genomic integration Self-adjuvant effect. |
Lack of large standardized production and purification methods Lack of effective transfection methods. |
Liu et al., 2019a; Prats et al., 2020; Wei et al., 2021; Zhao et al., 2022 |
Again, circRNAs in the cytoplasm form complexes with RBPs and microRNAs, acting as competing endogenous RNAs (ceRNAs) to regulate gene expression while protecting against degradation. In the nucleus, circRNAs can influence processes like alternative splicing and gene regulation, interacting with chromatin-modifying complexes and transcription factors to shield themselves from degradation. Pattern recognition receptors (PRRs) in immune cells recognize specific pathogen-associated patterns, and circRNAs play a role in evading immune detection while providing stability and regulatory functions within cells (Wesselhoeft et al., 2019).
4. Exploring circRNA Vaccines in Cancer: Immunological Perspective
The immunological aspect is critical in circRNA-based cancer vaccine research because the success of these vaccines relies on their ability to trigger a robust and specific immune response against tumor cells. Circular RNA (circRNA) is a type of genetic material capable of activating and regulating immune cells, making it a promising tool in cancer immunotherapy. Generally, tumor vaccines introduce tumor-specific antigens to the body, stimulating the immune system or generating CD8+ cytotoxic T lymphocytes (CTL) (Morse et al., 2021). These vaccines target tumor-associated antigens (TAAs), proteins expressed at low levels in healthy cells but elevated in tumor cells. Genomic instability during carcinogenesis causes mutations in coding regions, leading to neoantigens, which T cells can recognize when displayed by major histocompatibility complexes (MHC), triggering an immune response (Zhang et al., 2021).
CircRNA vaccines not only enhance the immune system’s ability to combat cancer more effectively than traditional vaccines due to their resistance to degradation and immune evasion (Yang et al., 2021a; Wesselhoeft et al., 2019) but also hold the potential to modulate the activity of key immune players, such as natural killer (NK) cells. CircRNAs, like circARSP91, have been shown to enhance NK cell cytotoxicity by upregulating ULBP1, thereby augmenting the immune surveillance and tumor-killing functions of NK cells (Piątkiewicz et al., 2013). This dual capability of circRNAs to evade immune detection while simultaneously boosting innate immune responses highlights their promise as potent agents in cancer immunotherapy.
This dual capability of circRNAs to evade immune detection while simultaneously boosting innate immune responses highlights their promise as potent agents in cancer immunotherapy, potentially overcoming the immunosuppressive effects exerted by tumor-associated macrophages (TAMs), which suppress cytotoxic T-cell responses through immune checkpoint activation, metabolite depletion, and anti-inflammatory cytokine production (Zhao et al., 2021). CircRNAs are believed to play a role in macrophage polarization, with three circRNAs showing distinct expression patterns in M1 cells and macrophages. CircRNAs may be crucial for macrophage polarization, as evidenced by the varying expression levels of particular circRNAs in various macrophage polarization states (Garrido-Martin et al., 2020) (Raskov et al., 2021).
Additionally, chemokines, known for attracting immune cells, are often implicated in cancer progression. In colorectal cancer (CRC), chemokines CCL20 and CXCL8 are downregulated, promoting cell migration, invasion, and epithelial-mesenchymal transition (EMT). CircFGFR1 is associated with CXCR4 expression in non-small cell lung cancer (NSCLC), potentially enhancing anti-PD-1 therapy. Similarly, the downregulation of circ_0067934 in thyroid carcinoma reduces CXCR1 expression, leading to apoptosis and reduced proliferation, migration, and invasion (Shen et al., 2019; Zhang et al., 2019). Tumor cytokines, produced by immune-suppressive or cancerous cells, contribute to an inflammatory tumor microenvironment. Inflammation is driven by caspase-1 activation, releasing proinflammatory cytokines like IL-1β and IL-18. CircRNAs such as hsa_circRNA_002178 in breast cancer and circ-0016347 in osteosarcoma have been shown to modulate inflammation-related pathways, suggesting a role for circRNAs in regulating tumor inflammation and progression (Jin et al., 2017; Liu et al., 2020)
Figure 3 illustrates the diverse functions of circRNAs in cellular processes, including the regulation of RNA-binding proteins (RBPs), transcription modulation, acting as miRNA sponges, influencing apoptosis, controlling the tumor microenvironment, modulating signaling pathways, and impacting metastasis.
5. Innovations in Designing circRNA-Based Cancer Vaccines
In vitro, circRNA synthesis begins with linear RNA precursor generation, followed by end ligation to form a closed-loop structure. Precursor design involves incorporating essential vaccine elements. CircRNA vaccines utilize host translation machinery, requiring antigen-encoding ORFs in precursors. Optimization involves selecting efficient coding sequences, considering codon bias and mRNA stability. Translation efficiency is enhanced by strategically including elements like Kozak sequences and IRES. Specific hexamers have been identified to trigger circRNA translation (He et al., 2024).
Figure S1 illustrates the key processes of circRNA formation during mRNA transcription, highlighting intron pairing, RNA-binding protein involvement, and lariat-driven circularization. Understanding these mechanisms is essential for optimizing circRNA-based cancer vaccine design.
Figure S2 illustrates the splicing events involved in the generation of diverse circRNAs, highlighting key mechanisms such as intron pairing, RNA-binding protein involvement, and lariat-driven circularization. Understanding these processes is crucial for optimizing circRNA-based cancer vaccine design.
Chemical approaches, such as phosphoramidite and solid-based synthesis, utilize derivatives of natural nucleoside triphosphates with inactive protective groups replacing reactive amino and hydroxyl groups. However, these methods have limited circularization capacity. IVT yields large numbers of linear precursors, with optimal reactions requiring guanosine in the transcripts. Further, ribozymes, RNA enzymes catalyzing precursor ligation, offer another method for circRNA synthesis (Obi & Chen, 2021).
Circularization of linear RNA involves innovative chemical and enzymatic methods to create stable, functional RNA loops essential for vaccine development. Chemical strategies, using cyanogen bromide (BrCN) or water-soluble carbodiimides like EDC/EDAC/EDCI, link the 5'-end phosphate to the 3'-end hydroxyl on linear RNA, facilitating circularization through self-templating driven by intrinsic secondary structure. (He et al., 2023).
Enzymatic RNA circularization, typically more efficient, is catalyzed by enzymes like T4 DNA ligase, which repairs nicks in double-stranded RNA substrates but requires cytidine and adenosine at ligation sites, requiring significant enzyme amounts due to low turnover efficiency on RNA. T4 RNA ligase II and the use of complementary DNA or RNA splints can improve intramolecular ligation, in some cases leading to RNA concatemers (Nandakumar et al., 2004) (Wesselhoeft et al., 2018).
Ribozymes, RNA enzymes catalyzing precursor RNAs' ligation can also create circRNAs by directing specific operations using antisense sequences. Techniques like the preparation of Internally Expressed (PIE) circRNA involve adding internal ribosome entry sites (IRES) and optimized accessory sequences to enhance stability and protein expression in eukaryotic cells (Wesselhoeft et al., 2018). Group I intron catalysis, using components like pre-tRNAleu and thymidylate synthase (td) genes, along with GTP and Mg2+, facilitates high-yield circRNA synthesis (Viollet et al., 2011).
The different types of ligation methods, their advantages and drawbacks are summarised in the table S1.
Further, circRNA synthesis utilizes genomes of subviral agents like viroids and hepatitis delta virus, which contain hairpin ribozymes (HPRs) and hammerhead ribozymes with intrinsic cleavage and ligation capabilities. These elements allow circRNA production from circular, single-stranded DNA templates and potentially broaden RNA circularization through "Twister" aptamers, recognized by tRNA endonuclease to produce 2', 3'-cyclic phosphate groups (Obi & Chen, 2021).
A schematic depiction of various RNA circularization methods is presented in Figure 4
The purity of circular RNA (circRNA) is crucial because impurities like DNA or double-stranded RNA (dsRNA) can trigger adverse immunological reactions. Cytoplasmic receptors like RIG-I and endosomal Toll-like receptors (TLRs) detect foreign RNA and induce inflammation. Additionally, proteins such as protein kinase R (PKR) react with dsRNA to trigger immune responses (Chen et al., 2019a).
Large-scale circRNA production can lead to unintended side effects, reduced yields, and complex purification processes. To address this, scientists propose using microreactors, which offer precise reaction control and minimize adverse effects while enhancing circRNA production (Lee et al., 2022).
6. Translational Milestones: from Bench to Bedside with circRNA Vaccines
Translational research combines clinical insights with laboratory research, driving advancements in human trials and diagnostics, with an emphasis on community acceptance, cost-effectiveness, and treatment strategies (Rubio et al., 2010). Among the promising cancer vaccines, mRNA vaccines have shown significant potential, though they are prone to RNase degradation, as discussed in “Unlocking the Power of Circular RNAs over mRNAs: Advantages in Cancer Vaccines.” In contrast, circular-RNA (circRNA) vaccines have demonstrated encouraging outcomes (H. Li et al., 2022).
Li et al. (2022) highlighted circRNA vaccines' superior results in SARS-CoV-2 studies. Though still nascent in cancer treatment, circRNA vaccines hold great potential. Yang et al. (2021a) introduced chemically modified RNA (cmRNA), which supports substantial, sustained protein expression. When encoding cytokines, cmRNA injected into tumors improved systemic antitumor immune responses, enhancing tumor suppression in syngeneic mouse models treated with PD-1 antibodies. This formulation triggers significant CD4+ and CD8+ T cell infiltration, boosting intratumoral antitumor activity. cmRNA’s capacity to express diverse proteins further underscores its potential for developing vaccines and treatments (Yang et al., 2022).
Amaya et al. (2023) demonstrated that circRNA vaccines induce robust adaptive immune responses, inhibiting tumor growth in mice. Similarly, Li et al. (2022) showed that circRNA delivered via lipid nanoparticles elicits a strong innate immune response, activating antigen-specific cytotoxic T cells to eliminate difficult tumors. Combining circRNA vaccination with adoptive cell transfer therapy was particularly effective in late-stage mouse tumor models, with potential applications as primary or adjuvant cancer therapy.
CircRNAs are deregulated in multiple cancers, such as colorectal (Ji et al., 2018), hepatocellular (El Sharkawi et al., 2021; Yao et al., 2018), and lung adenocarcinomas (Chen et al., 2019a), correlating with cancer progression and metastasis. High-throughput sequencing identified distinct circRNA expression patterns in lung adenocarcinoma plasma exosomes, revealing 105 upregulated and 78 downregulated circRNAs compared to controls (Chen et al., 2019a). In gastric cancer, circRNAs drive tumor behavior via the PI3K/Akt/mTOR pathway (Peng et al., 2020). These findings highlight circRNAs’ potential as diagnostic and prognostic biomarkers, providing a foundation for early detection and targeted therapies (Chen et al., 2019a).
CircRNA vaccines also function as important cancer immunotherapies and biomarkers. Due to their resistance to degradation, circRNAs persist in body fluids, making them ideal for noninvasive liquid biopsies and diagnostic purposes (Fang et al., 2019). Recent studies demonstrate that circRNA vaccines encode tumor-specific neoantigens, training the immune system to target these cancers, thus opening pathways for innovative cancer therapies (Sahin & Türeci, 2018).
7. Emerging Trends and Challenges
In the field of vaccine development, a significant shift towards harnessing the potential of circular RNA (circRNA) vaccines has emerged, marked by their unique "self-adjuvant" effect akin to linear mRNA vaccines, yet distinguished by distinct characteristics necessitating careful consideration in research and development (Xie et al., 2023). Current efforts are concentrated on optimizing circRNA vaccine design and formulation, with circularization efficiency serving as a pivotal stage influencing vaccine capacity and subsequent purification processes (Bai et al., 2023). Unlike mRNA vaccines, circRNA vaccines possess a loop structure, demanding tailored approaches for efficient circularization of different target sequences, wherein intron sequences and exon length also play critical roles (Xie et al. 2023).
7.1. Personalized Treatment
The growing field of personalized therapy aims to choose the most appropriate therapy regime that is optimum for each patient suffering from cancer (Kumar et al., 2022). In cancer therapy, personalized treatment involves focusing on particular mutations found in a patient's tumor. CircRNA vaccines can be designed in such a way that it encodes neoantigens that result from tumor-specific mutations (Aldous & Dong, 2018), (Sahin & Türeci, 2018). The individual's immune system can be trained to identify and fight against tumor cells expressing these antigens by developing a vaccination that encodes these neoantigens. CircRNA vaccines can be engineered in such a way that particular changes or sequences can be incorporated that improve the immunogenicity of the vaccination (Poboży et al., 2023). This targeted treatment opens avenues for a more effective and precise immune response against patients' unique cancer types and characteristics (Wu et al., 2018, Khansari et al., 2019).
7.2. Challenges
RNAs with circular shapes are special molecules that work as effective microRNA sponges and support the development of immune cells, control of signaling, and anticancer activity. However, there is a problem with the paucity of comprehensive study on their effects on linear mRNA expression and epigenetic alterations, as well as their role in innate immunity and cancer progression. To further improve the therapeutic potential of circRNA-based vaccines, concerns like biosafety, yield optimization, and modification procedures need to be addressed. Cutting-edge techniques like CRISPR-Cas9 for modifying gene expression and enhanced detection strategies are essential for propelling circRNA research forward.
Table 2.
Challenges associated with circRNA constructs.
| Challenges | Description |
| Detection and Quantification | The circular nature makes circRNAs elusive to accurate detection and quantification as this can be omitted by traditional RNA sequencing approaches or underestimated in terms of abundance (Dong et al., 2023; Kristensen et al., 2018; Nguyen et al., 2022; Szabo & Salzman, 2016). |
| Functional Annotation | It becomes difficult to understand the roles of circRNA because they may act as miRNA sponges, bind to RBPs, regulate gene transcription, and their mechanisms of action are diverse (Lee et al., 2022). |
| Design and Engineering | This is challenging since there will be a need to design circRNA constructs for experiments such as overexpression or knockdown which require inherent circularity necessitating specialized techniques such as antisense oligonucleotides or CRISPR-based strategies (Feng et al., 2023; Gao et al., 2022; Li et al., 2018; Obi & Chen, 2021; Szabo & Salzman, 2016). |
| Stability | CircRNAs possess varying stability across several different subcellular localizations (He et al., 2021; Kristensen et al., 2018; Ren et al., 2022; Wu et al., 2022). |
| Delivery systems | CircRNA-based constructs from specific cells or tissues require efficient delivery systems that include viral vectors or nanoparticles (Feng et al., 2023; He et al., 2021; Loan Young et al., 2023; Zhao et al., 2022). |
| Bioinformatics tools | Bioinformatics tools require circRNA sequencing data to differentiate between circular and linear RNA reads to determine back-splicing regions (Cheng et al., 2021; Feng et al., 2023; Miao et al., 2021; Niu et al., 2024). |
8. Conclusions
Circular RNAs (circRNAs) are unique molecules with significant roles in the immune system, including acting as microRNA sponges and contributing to cell differentiation and immune cell development. They also regulate cell signaling pathways during activation and exhibit antitumor functions, particularly in breast cancer by modulating tumor progression. However, the mechanisms through which circRNAs influence immune responses and cancer progression remain underexplored, presenting a challenge in fully understanding their potential. The translation of circRNAs into functional small proteins that may act as molecular inhibitors in cancer is promising but requires further evidence-based research. Additionally, circRNA-based vaccines face challenges related to biosafety, drug responses, and their potential interference with linear mRNA expression, which could lead to epigenetic mutations during cancer metastasis. Further innovations, such as CRISPR-Cas9 techniques and improved reaction conditions, are needed to overcome these obstacles and enhance the utility of circRNAs in therapeutic settings.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
U. Das: Conceptualization, Validation, Writing - Original Draft, Writing - Review & Editing, Visualization, Supervision, Project administration. S. Banerjee: Writing - Original Draft, Writing - Review & Editing, Visualization, Supervision. M. Sarkar: Writing - Original Draft, Writing - Review & Editing, Visualization. F. Muhammad L: Writing - Original Draft, Writing - Review & Editing, Visualization. T. K. Soni: Writing - Original Draft, Writing - Review & Editing, Visualization. M. Saha: Writing - Review & Editing, Visualization. G. Pradhan: Writing - Review & Editing, Visualization. B. Chatterjee: Writing - Original Draft.
Data Availability Statement
No data was used for the research described in the article.
Acknowledgments
We would like to express their heartfelt gratitude to our parents and friends whose support, encouragement, and understanding made this review possible.
Conflicts of Interest
The authors report no conflict of interest.
References
- Aldous, A. R., & Dong, J. Z. (2018). Personalized neoantigen vaccines: A new approach to cancer immunotherapy. Bioorganic & Medicinal Chemistry, 26(10), 2842–2849. [CrossRef]
- Amaya, L., Grigoryan, L., Li, Z., Lee, A., Wender, P. A., Pulendran, B., & Chang, H. Y. (2023). Circular RNA vaccine induces potent T-cell responses. Proceedings of the National Academy of Sciences, 120(20). [CrossRef]
- Ashwal-Fluss, R., Meyer, M., Pamudurti, N. R., Ivanov, A., Bartok, O., Hanan, M., Evantal, N., Memczak, S., Rajewsky, N., & Kadener, S. (2014). CircRNA biogenesis competes with pre-mRNA splicing. Molecular Cell, 56(1), 55–66. [CrossRef]
- Barbier, A. J., Jiang, A. Y., Zhang, P., Wooster, R., & Anderson, D. G. (2022, May 9). The clinical progress of mRNA vaccines and immunotherapies. Nature Biotechnology, 40(6), 840–854. [CrossRef]
- Baxevanis, C. N., Fortis, S. P., Ardavanis, A., & Perez, S. A. (2020). Exploring essential issues for improving therapeutic cancer vaccine trial design. Cancers, 12(10), 2908. [CrossRef]
- Beck, J. D., Reidenbach, D., Salomon, N., & et al. (2021). mRNA therapeutics in cancer immunotherapy. Molecular Cancer, 20(69). [CrossRef]
- Cafri, G., Gartner, J. J., Zaks, T., Hopson, K., Levin, N., Paria, B. C., Parkhurst, M. R., Yossef, R., Lowery, F. J., Jafferji, M. S., Prickett, T. D., Goff, S. L., McGowan, C. T., Seitter, S., Shindorf, M. L., Parikh, A., Chatani, P. D., Robbins, P. F., & Rosenberg, S. A. (2020, October 5). mRNA vaccine–induced neoantigen-specific T cell immunity in patients with gastrointestinal cancer. Journal of Clinical Investigation, 130(11), 5976–5988. [CrossRef]
- Cardillo, F., Bonfim, M., Da Silva Vasconcelos Sousa, P., Mengel, J., Ribeiro Castello-Branco, L. R., & Pinho, R. T. (2021). Bacillus Calmette–Guérin Immunotherapy for Cancer. Vaccines, 9(5), 439. [CrossRef]
- Cech, T. R., Zaug, A. J., & Grabowski, P. J. (1981). In vitro splicing of the ribosomal RNA precursor of tetrahymena: Involvement of a guanosine nucleotide in the excision of the intervening sequence. Cell, 27(3), 487–496. [CrossRef]
- Chakraborty, C., Sharma, A. R., Bhattacharya, M., & Lee, S. S. (2021, July 7). From COVID-19 to Cancer mRNA Vaccines: Moving from Bench to Clinic in the Vaccine Landscape. Frontiers in Immunology, 12. [CrossRef]
- Chen, C., & Sarnow, P. (1995). Initiation of Protein Synthesis by the Eukaryotic Translational Apparatus on Circular RNAs. Science, 268(5209), 415–417. [CrossRef]
- Chen, C., Chen, X., Tang, H., Jiang, Y., Wang, S., Zhang, X., Huang, T., Yuan, X., Wang, J., & Peng, L. (2022). Circular RNAs involve in immunity of digestive cancers From bench to bedside: a review. Frontiers in Immunology, 13. [CrossRef]
- Chen, F., Huang, C., Wu, Q., Jiang, L., Chen, S., & Chen, L. (2019a). Circular RNA expression profiles in plasma exosomes from early-stage lung adenocarcinoma and the potential biomarkers. Journal of Cellular Biochemistry, 121(3), 2525–2533. [CrossRef]
- Chen, F., Zou, Z., Du, J., Su, S., Shao, J., Meng, F., Yang, J., Xu, Q., Ding, N., Yang, Y., Liu, Q., Wang, Q., Sun, Z., Zhou, S., Du, S., Wei, J., & Liu, B. (2019b, April 8). Neoantigen identification strategies enable personalized immunotherapy in refractory solid tumors. Journal of Clinical Investigation, 129(5), 2056–2070. [CrossRef]
- Chen, L. L. (2020a, May 4). The expanding regulatory mechanisms and cellular functions of circular RNAs. Nature Reviews Molecular Cell Biology, 21(8), 475–490. [CrossRef]
- Chen, Z., Zhang, S., Han, N., Jiang, J., Xu, Y., Ma, D., Lu, L., Guo, X., Qiu, M., Huang, Q., Wang, H., Mo, F., Chen, S., & Yang, L. (2021b, August 13). A Neoantigen-Based Peptide Vaccine for Patients with Advanced Pancreatic Cancer Refractory to Standard Treatment. Frontiers in Immunology, 12. [CrossRef]
- ClinicalTrials.gov. (n.d.). Phase 1, open-label, dose-ranging study of the safety and immunogenicity of 2019-nCoV vaccine (mRNA-1273) in healthy adults. Retrieved May 23, 2024, from https://clinicaltrials.gov/ct2/show/NCT04283461.
- Cong, J., Wang, X., Zheng, X., Wang, D., Fu, B., Sun, R., Tian, Z., & Wei, H. (2018, August). Dysfunction of Natural Killer Cells by FBP1-Induced Inhibition of Glycolysis during Lung Cancer Progression. Cell Metabolism, 28(2), 243-255.e5. [CrossRef]
- Derderian, C., Orunmuyi, A. T., Olapade-Olaopa, E. O., & Ogunwobi, O. O. (2019). PVT1 signaling is a mediator of cancer progression. Frontiers in Oncology, 9. [CrossRef]
- Dong, Y., Gao, Q., Chen, Y., Zhang, Z., Du, Y., Liu, Y., Zhang, G., Li, S., Wang, G., Chen, X., Liu, H., Han, L., & Ye, Y. (2023b). Identification of CircRNA signature associated with tumor immune infiltration to predict the therapeutic efficacy of immunotherapy. Nature communications, 14(1), 2540. [CrossRef]
- El Sharkawi, F. Z., Awad, M. S., Elagawy, W., Al Sawaf, H., & Taha, H. (2021). Circular RNAs 0064286 and 0000475: Potential Diagnostic Biomarkers in Hepatocellular Carcinoma. Asian Pacific Journal of Cancer Prevention, 22(9), 3039–3044. [CrossRef]
- Fan, Y.-N., Li, M., Luo, Y.-L., Chen, Q., Wang, L., Zhang, H.-B., Shen, S., Gu, Z., & Wang, J. (2018). Cationic lipid-assisted nanoparticles for delivery of mRNA cancer vaccine. Biomaterials Science, 6(11), 3009–3018. [CrossRef]
- Feng, X.-Y., Zhu, S.-X., Pu, K.-J., Huang, H.-J., Chen, Y.-Q., & Wang, W.-T. (2023). New insight into circRNAs: Characterization, strategies, and biomedical applications. Experimental Hematology & Oncology, 12(1), 91. [CrossRef]
- Garrido-Martin, E. M., Mellows, T. W. P., Clarke, J., Ganesan, A. P., Wood, O., Cazaly, A., Seumois, G., Chee, S. J., Alzetani, A., King, E. V., Hedrick, C. C., Thomas, G., Friedmann, P. S., Ottensmeier, C. H., Vijayanand, P., & Sanchez-Elsner, T. (2020, July). M1hottumor-associated macrophages boost tissue-resident memory T cells infiltration and survival in human lung cancer. Journal for ImmunoTherapy of Cancer, 8(2), e000778. [CrossRef]
- Ge, J., Wang, J., Xiong, F., Jiang, X., Zhu, K., Wang, Y., Mo, Y., Gong, Z., Zhang, S., He, Y., Li, X., Shi, L., Guo, C., Wang, F., Zhou, M., Xiang, B., Li, Y., Li, G., Xiong, W., & Zeng, Z. (2021, July 28). Epstein-–Barr Virus–Encoded Circular RNA CircBART2.2 Promotes Immune Escape of Nasopharyngeal Carcinoma by Regulating PD-L1. Cancer Research, 81(19), 5074–5088. [CrossRef]
- Guo, Y., Lei, K., & Tang, L. (2018, July 2). Neoantigen Vaccine Delivery for Personalized Anticancer Immunotherapy. Frontiers in Immunology, 9. [CrossRef]
- Haabeth, O. A. W., Blake, T. R., McKinlay, C. J., Waymouth, R. M., Wender, P. A., & Levy, R. (2018). mRNA vaccination with charge-altering releasable transporters elicits human T cell responses and cures established tumors in mice. Proceedings of the National Academy of Sciences of the United States of America, 115(39). [CrossRef]
- Hassett, K. J., Benenato, K. E., Jacquinet, E., Lee, A., Woods, A., Yuzhakov, O., Himansu, S., Deterling, J., Geilich, B. M., Ketova, T., Mihai, C., Lynn, A., McFadyen, I., Moore, M. J., Senn, J. J., Stanton, M. G., Almarsson, Ö., Ciaramella, G., & Brito, L. A. (2019). Optimization of lipid nanoparticles for intramuscular administration of mRNA vaccines. Molecular Therapy. Nucleic Acids, 15, 1–11. [CrossRef]
- He, T., Xu, Y., Li, X., Wang, X., Li, J., Ou-Yang, D., Cheng, H., Li, H., Qin, J., Huang, Y., & Wang, H. (2023). A linear and circular dual-conformation noncoding RNA involved in oxidative stress tolerance in Bacillus altitudinis. Nature Communications, 14(1). [CrossRef]
- He, W., Zhang, X., Zou, Y., Li, J., Chang, L., He, Y., Jin, Q., & Ye, J. (2024). Effective synthesis of circRNA via a thermostable T7 RNA polymerase variant as the catalyst. Frontiers in Bioengineering and Biotechnology, 12. [CrossRef]
- Hewitt, S. L., Bai, A., Bailey, D., Ichikawa, K., Zielinski, J., Karp, R., Apte, A., Arnold, K., Zacharek, S. J., Iliou, M. S., Bhatt, K., Garnaas, M., Musenge, F., Davis, A., Khatwani, N., Su, S. V., MacLean, G., Farlow, S. J., Burke, K., & Frederick, J. P. (2019). Durable anticancer immunity from intratumoral administration of IL-23, IL-36γ, and OX40L mRNAs. Science Translational Medicine, 11(477). [CrossRef]
- Hiam-Galvez, K. J., Allen, B. M., & Spitzer, M. H. (2021). Systemic immunity in cancer. Nature Reviews. Cancer, 21(6), 345–359. [CrossRef]
- Hoover, H. C., Jr, Brandhorst, J. S., Peters, L. C., Surdyke, M. G., Takeshita, Y., Madariaga, J., Muenz, L. R., & Hanna, M. G., Jr. (1993). Adjuvant active specific immunotherapy for human colorectal cancer: 6.5-year median follow-up of a phase III prospectively randomized trial. Journal of Clinical Oncology: Official Journal of the American Society of Clinical Oncology, 11(3), 390–399. [CrossRef]
- Huang, Q., Zeng, J., & Yan, J. (2021, February). COVID-19 mRNA vaccines. Journal of Genetics and Genomics, 48(2), 107–114. [CrossRef]
- Huff, A. L., Jaffee, E. M., & Zaidi, N. (2022, March 15). Messenger RNA vaccines for cancer immunotherapy: progress promotes promise. Journal of Clinical Investigation, 132(6). [CrossRef]
- Islam, M. A., Xu, Y., Tao, W., Ubellacker, J. M., Lim, M., Aum, D., Lee, G. Y., Zhou, K., Zope, H., Yu, M., Cao, W., Oswald, J. T., Dinarvand, M., Mahmoudi, M., Langer, R., Kantoff, P. W., Farokhzad, O. C., Zetter, B. R., & Shi, J. (2018). Restoration of tumour-growth suppression in vivo via systemic nanoparticle-mediated delivery of PTEN mRNA. Nature Biomedical Engineering, 2(11), 850–864. [CrossRef]
- Jain, S., Venkataraman, A., Wechsler, M. E., & Peppas, N. A. (2021, December). Messenger RNA-based vaccines: Past, present, and future directions in the context of the COVID-19 pandemic. Advanced Drug Delivery Reviews, 179, 114000. [CrossRef]
- Ji, W., Qiu, C., Wang, M., Mao, N., Wu, S., & Dai, Y. (2018). Hsa_circ_0001649: A circular RNA and potential novel biomarker for colorectal cancer. Biochemical and Biophysical Research Communications, 497(1), 122–126. [CrossRef]
- Jin, H., Jin, X., Zhang, H., & Wang, W. (2017, March 10). Circular RNA hsa-circ-0016347 promotes proliferation, invasion, and metastasis of osteosarcoma cells. Oncotarget, 8(15), 25571–25581. [CrossRef]
- Jou, J., Harrington, K. J., Zocca, M. B., Ehrnrooth, E., & Cohen, E. E. (2021, February 1). The Changing Landscape of Therapeutic Cancer Vaccines—Novel Platforms and Neoantigen Identification. Clinical Cancer Research, 27(3), 689–703. [CrossRef]
- Kantoff, P. W., Higano, C. S., Shore, N. D., Berger, E. R., Small, E. J., Penson, D. F., Redfern, C. H., Ferrari, A. C., Dreicer, R., Sims, R. B., Xu, Y., Frohlich, M. W., & Schellhammer, P. F. (2010). Sipuleucel-T immunotherapy for castration-resistant prostate cancer. The New England Journal of Medicine, 363(5), 411–422. [CrossRef]
- Karikó, K., Muramatsu, H., Ludwig, J., & Weissman, D. (2011). Generating the optimal mRNA for therapy: HPLC purification eliminates immune activation and improves translation of nucleoside-modified, protein-encoding mRNA. Nucleic Acids Research, 39(21), e142–e142. [CrossRef]
- Kazakov, S. A., Balatskaya, S. V., & Johnston, B. H. (2006). Ligation of the hairpin ribozyme in cis induced by freezing and dehydration. RNA, 12(3), 446–456. [CrossRef]
- Khansari, N. (2019). Neoantigen: A New Hope for Effective Cancer Immunotherapy. Vaccination Research – Open Journal, 4(1), 19–20. [CrossRef]
- Kleaveland, B., Shi, C. Y., Stefano, J., & Bartel, D. P. (2018). A network of noncoding regulatory RNAs acts in the mammalian brain. Cell, 174(2), 350-362.e17. [CrossRef]
- Kristensen, L. S., Hansen, T. B., Venø, M. T., & Kjems, J. (2018). Circular RNAs in cancer: Opportunities and challenges in the field. Oncogene, 37(5), 555–565. [CrossRef]
- Kumar, A., Weller, K. P., & Vilgelm, A. E. (2022). Personalized cancer immunotherapy. Engineering Technologies and Clinical Translation, 399–426. [CrossRef]
- Le Moignic, A., Malard, V., Benvegnu, T., Lemiègre, L., Berchel, M., Jaffrès, P.-A., Baillou, C., Delost, M., Macedo, R., Rochefort, J., Lescaille, G., Pichon, C., Lemoine, F. M., Midoux, P., & Mateo, V. (2018). Preclinical evaluation of mRNA trimannosylated lipopolyplexes as therapeutic cancer vaccines targeting dendritic cells. Journal of Controlled Release: Official Journal of the Controlled Release Society, 278, 110–121. [CrossRef]
- Lee, K. H., Kim, S., & Lee, S. (2022). Pros and cons of in vitro methods for circular RNA preparation. International Journal of Molecular Sciences, 23(21), 13247. [CrossRef]
- Legnini, I., Di Timoteo, G., Rossi, F., Morlando, M., Briganti, F., Sthandier, O., Fatica, A., Santini, T., Andronache, A., Wade, M., Laneve, P., Rajewsky, N., & Bozzoni, I. (2017). Circ-ZNF609 Is a Circular RNA that Can Be Translated and Functions in Myogenesis. Molecular Cell, 66(1), 22-37. e9. [CrossRef]
- Lei, Z., Tian, Q., Teng, Q., Wurpel, J. N., Zeng, L., Pan, Y., & Chen, Z. (2023). Understanding and targeting resistance mechanisms in cancer. MedComm, 4(3). [CrossRef]
- Li, H., Peng, K., Yang, K., Ma, W., Qi, S., Yu, X., He, J., Lin, X., & Yu, G. (2022). Circular RNA cancer vaccines drive immunity in hard-to-treat malignancies. Theranostics, 12(14), 6422–6436. [CrossRef]
- Li, J., Xu, Q., Huang, Z., Mao, N., Lin, Z., Cheng, L., Sun, B., & Wang, G. (2021a). CircRNAs: A new target for the diagnosis and treatment of digestive system neoplasms. Cell Death & Disease, 12(2), 1–13. [CrossRef]
- Li, W., Liu, J., Chen, M., Xu, J., & Zhu, D. (2020). Circular RNA in cancer development and immune regulation. Journal of Cellular and Molecular Medicine, 26(6), 1785–1798. [CrossRef]
- Lisiecka, U., & Kostro, K. (2016). Mechanisms of tumor escape from immune surveillance. Journal of Veterinary Research, 60(4), 453–460. [CrossRef]
- Liu, J., Song, S., Lin, S., Zhang, M., Du, Y., Zhang, D., Xu, W., & Wang, H. (2019a, June 14). Circ-SERPINE2 promotes the development of gastric carcinoma by sponging miR-375 and modulating YWHAZ. Cell Proliferation, 52(4). [CrossRef]
- Liu, L., Wang, Y., Miao, L., Liu, Q., Musetti, S., Li, J., & Huang, L. (2018). Combination immunotherapy of MUC1 mRNA nano-vaccine and CTLA-4 blockade effectively inhibits growth of triple negative breast cancer. Molecular Therapy: The Journal of the American Society of Gene Therapy, 26(1), 45–55. [CrossRef]
- Liu, T., Ye, P., Ye, Y., Lu, S., & Han, B. (2020, January 19). Circular RNA hsa_circRNA_002178 silencing retards breast cancer progression via microRNA-328-3p-mediated inhibition of COL1A1. Journal of Cellular and Molecular Medicine, 24(3), 2189–2201. [CrossRef]
- Liu, X., Zhang, Y., Zhou, S., Dain, L., Mei, L., & Zhu, G. (2022, August). Circular RNA: An emerging frontier in RNA therapeutic targets, RNA therapeutics, and mRNA vaccines. Journal of Controlled Release, 348, 84–94. [CrossRef]
- Ma, Y., Wang, T., Zhang, X., Wang, P., & Long, F. (2024). The role of circular RNAs in regulating resistance to cancer immunotherapy: mechanisms and implications. Cell Death and Disease, 15(5). [CrossRef]
- Mai, Y., Guo, J., Zhao, Y., Ma, S., Hou, Y., & Yang, J. (2020). Intranasal delivery of cationicT liposome-protamine complex mRNA vaccine elicits effective anti-tumor immunity. Cellular Immunology, 354(104143), 104143. [CrossRef]
- Mendes, B. B., Conniot, J., Avital, A., Yao, D., Jiang, X., Zhou, X., Sharf-Pauker, N., Xiao, Y., Adir, O., Liang, H., Shi, J., Schroeder, A., & Conde, J. (2022). Nanodelivery of nucleic acids. Nature Reviews Methods Primers, 2(1). [CrossRef]
- Miao, L., Zhang, Y., & Huang, L. (2021a, February 25). mRNA vaccine for cancer immunotherapy. Molecular Cancer, 20(1). [CrossRef]
- Mo, Y., Wang, Y., Wang, Y., Deng, X., Yan, Q., Fan, C., Zhang, S., Zhang, S., Gong, Z., Shi, L., Liao, Q., Guo, C., Li, Y., Li, G., Zeng, Z., Jiang, W., Xiong, W., & Xiang, B. (2022, October 5). Circular RNA circPVT1 promotes nasopharyngeal carcinoma metastasis via the β-TrCP/c-Myc/SRSF1 positive feedback loop. Molecular Cancer, 21(1). [CrossRef]
- Morse, M. A., Gwin, W. R., & Mitchell, D. A. (2021, January 29). Vaccine Therapies for Cancer: Then and Now. Targeted Oncology, 16(2), 121–152. [CrossRef]
- Nandakumar, J., & Shuman, S. (2004). How an RNA Ligase Discriminates RNA versus DNA Damage. Molecular Cell, 16(2), 211–221. [CrossRef]
- Nigro, J. M., Cho, K. R., Fearon, E. R., Kern, S. E., Ruppert, J. M., Oliner, J. D., Kinzler, K. W., & Vogelstein, B. (1991). Scrambled exons. Cell, 64(3), 607–613. [CrossRef]
- Niu, D., Wu, Y., & Lian, J. (2023). Circular RNA vaccine in disease prevention and treatment. Signal Transduction and Targeted Therapy, 8(1). [CrossRef]
- Obi, P., & Chen, Y. G. (2021). The design and synthesis of circular RNAs. Methods, 196, 85–103. [CrossRef]
- Pamudurti, N. R., Bartok, O., Jens, M., Ashwal-Fluss, R., Stottmeister, C., Ruhe, L., Hanan, M., Wyler, E., Perez-Hernandez, D., Ramberger, E., Shenzis, S., Samson, M., Dittmar, G., Landthaler, M., Chekulaeva, M., Rajewsky, N., & Kadener, S. (2017). Translation of CircRNAs. Molecular Cell, 66(1), 9-21. e7. [CrossRef]
- Pasman Z, Been MD & Garcia-Blanco MA (1996). Exon circularization in mammalian nuclear extracts. RNA (New York, N.Y.);2(6):603-10.
- Patop, I. L., & Kadener, S. (2018). CircRNAs in cancer. Current Opinion in Genetics & Development, 48, 121–127. [CrossRef]
- Petkovic, S., & Müller, S. (2015). RNA circularization strategies in vivo and in vitro. Nucleic Acids Research, 43(4), 2454–2465. [CrossRef]
- Piątkiewicz, P., Miłek, T., Bernat-Karpińska, M., Ohams, M., Czech, A., & Ciostek, P. (2013, March 2). The Dysfunction of NK Cells in Patients with Type 2 Diabetes and Colon Cancer. Archivum Immunologiae Et Therapiae Experimentalis, 61(3), 245–253. [CrossRef]
- Piwecka, M., Glažar, P., Hernandez-Miranda, L. R., Memczak, S., Wolf, S. A., Rybak-Wolf, A., Filipchyk, A., Klironomos, F., Cerda Jara, C. A., Fenske, P., Trimbuch, T., Zywitza, V., Plass, M., Schreyer, L., Ayoub, S., Kocks, C., Kühn, R., Rosenmund, C., Birchmeier, C., & Rajewsky, N. (2017). Loss of a mammalian circular RNA locus causes miRNA deregulation and affects brain function. Science (New York, N.Y.), 357(6357). [CrossRef]
- Poboży, K., Domańska, J., & Domański, P. (2023). Neoantigen therapeutic cancer vaccines: a promising approach to personalized immunotherapy. OncoReview. [CrossRef]
- Prats, A. C., David, F., Diallo, L. H., Roussel, E., Tatin, F., Garmy-Susini, B., & Lacazette, E. (2020, November 14). Circular RNA, the Key for Translation. International Journal of Molecular Sciences, 21(22), 8591. [CrossRef]
- Propper, D. J., & Balkwill, F. R. (2022). Harnessing cytokines and chemokines for cancer therapy. Nature Reviews. Clinical Oncology, 19(4), 237–253. [CrossRef]
- Puttaraju, M., & Been, M. (1992). Group I permuted intron-exon (PIE) sequences self-splice to produce circular exons. Nucleic Acids Research, 20(20), 5357–5364. [CrossRef]
- Qu, H., Chen, M., Ge, J., Zhang, X., He, S., Xiong, F., Yan, Q., Zhang, S., Gong, Z., Guo, C., Wang, F., Zeng, Z., Li, X., Li, G., Xiong, W., & Wu, X. (2022a, January). A fluorescence strategy for circRNA quantification in tumor cells based on T7 nuclease-assisted cycling enzymatic amplification. Analytica Chimica Acta, 1189, 339210. [CrossRef]
- Qu, L., Yi, Z., Shen, Y., Lin, L., Chen, F., Xu, Y., Wu, Z., Tang, H., Zhang, X., Tian, F., Wang, C., Xiao, X., Dong, X., Guo, L., Lu, S., Yang, C., Tang, C., Yang, Y., Yu, W., . . . Wei, W. (2022b, May). Circular RNA vaccines against SARS-CoV-2 and emerging variants. Cell, 185(10), 1728-1744.e16. [CrossRef]
- Raskov, H., Orhan, A., Gaggar, S., & Gögenur, I. (2021). Cancer-Associated fibroblasts and Tumor-Associated macrophages in cancer and cancer immunotherapy. Frontiers in Oncology, 11. [CrossRef]
- Ren, D., Hua, Y., Yu, B., Ye, X., He, Z., Li, C., Wang, J., Mo, Y., Wei, X., Chen, Y., Zhou, Y., Liao, Q., Wang, H., Xiang, B., Zhou, M., Li, X., Li, G., Li, Y., Zeng, Z., & Xiong, W. (2020). Predictive biomarkers and mechanisms underlying resistance to PD1/PD-L1 blockade cancer immunotherapy. Molecular Cancer, 19(1). [CrossRef]
- Romero, P., & Coukos, G. (2014). Successful engineering cancer immunotherapy. European Journal of Immunology, 44(2), 318–320. [CrossRef]
- Rubio, D. M., Schoenbaum, E. E., Lee, L. S., Schteingart, D. E., Marantz, P. R., Anderson, K. E., Platt, L. D., Baez, A., & Esposito, K. (2010). Defining Translational Research: Implications for Training. Academic Medicine, 85(3), 470–475. [CrossRef]
- Sahin, U., & Türeci, Z. (2018). Personalized vaccines for cancer immunotherapy. Science, 359(6382), 1355–1360. [CrossRef]
- Sanger, H. L., Klotz, G., Riesner, D., Gross, H. J., & Kleinschmidt, A. K. (1976). Viroids are single-stranded covalently closed circular RNA molecules existing as highly base-paired rod-like structures. Proceedings of the National Academy of Sciences of the United States of America, 73(11), 3852–3856. [CrossRef]
- Schindewolf, C., Braun, S., & Domdey, H. (1996). In vitro Generation of a Circular Exon from a Linear Pre-mRNA Transcript. Nucleic Acids Research, 24(7), 1260–1266. [CrossRef]
- Sebastian, M., Schröder, A., Scheel, B. et al. (2019). A phase I/IIa study of the mRNA-based cancer immunotherapy CV9201 in patients with stage IIIB/IV non-small cell lung cancer. Cancer Immunol Immunother, 68, 799–812. [CrossRef]
- Shen, T., Cheng, X., Liu, X., Xia, C., Zhang, H., Pan, D., Zhang, X., & Li, Y. (2019, October 14). Circ_0026344 restrains metastasis of human colorectal cancer cells via miR-183. Artificial Cells, Nanomedicine, and Biotechnology, 47(1), 4038–4045. [CrossRef]
- Son, S., Nam, J., Zenkov, I., Ochyl, L. J., Xu, Y., Scheetz, L., Shi, J., Farokhzad, O. C., & Moon, J. J. (2020). Sugar-nanocapsules imprinted with microbial molecular patterns for mRNA vaccination. Nano Letters, 20(3), 1499–1509. [CrossRef]
- Španielová, H., & Brdička, R. (2023). Oncolytic viruses and cancer treatment. Klinická Onkologie, 36(1). [CrossRef]
- Su, M., Xiao, Y., Ma, J., Tang, Y., Tian, B., Zhang, Y., Xu, L., Wu, Z., Yang, D., Zhou, Y., Wang, H., Liao, Q., & Wang, W. (2019). Circular RNAs in Cancer: emerging functions in hallmarks, stemness, resistance and roles as potential biomarkers. Molecular Cancer, 18(1). [CrossRef]
- Sun, C., Sun, H. Y., Xiao, W. H., Zhang, C., & Tian, Z. G. (2015, June 15). Natural killer cell dysfunction in hepatocellular carcinoma and NK cell-based immunotherapy. Acta Pharmacologica Sinica, 36(10), 1191–1199. [CrossRef]
- Supabphol, S., Li, L., Goedegebuure, S. P., & Gillanders, W. E. (2021, March 31). Neoantigen vaccine platforms in clinical development: understanding the future of personalized immunotherapy. Expert Opinion on Investigational Drugs, 30(5), 529–541. [CrossRef]
- Tateshita, N., Miura, N., Tanaka, H., Masuda, T., Ohtsuki, S., Tange, K., Nakai, Y., Yoshioka, H., & Akita, H. (2019). Development of a lipoplex-type mRNA carrier composed of an ionizable lipid with a vitamin E scaffold and the KALA peptide for use as an ex vivo dendritic cell-based cancer vaccine. Journal of Controlled Release: Official Journal of the Controlled Release Society, 310, 36–46. [CrossRef]
- Verbeke, R., Lentacker, I., Breckpot, K., Janssens, J., Van Calenbergh, S., De Smedt, S. C., & Dewitte, H. (2019). Broadening the message: A nanovaccine co-loaded with messenger RNA and α-GalCer induces antitumor immunity through conventional and natural killer T cells. ACS Nano. [CrossRef]
- Viollet, S., Fuchs, R. T., Munafo, D. B., Zhuang, F., & Robb, G. B. (2011). T4 RNA Ligase 2 truncated active site mutants: improved tools for RNA analysis. BMC Biotechnology, 11(1). [CrossRef]
- Wang, B., Yin, H., Zhang, Y., Zhao, Q., Wang, H., Wei, J., Meng, L., Xin, Y., & Jiang, X. (2023). Overcoming acquired resistance to cancer immune checkpoint therapy: potential strategies based on molecular mechanisms. Cell & Bioscience, 13(1). [CrossRef]
- Wang, W. (2015a). NK cell-mediated antibody-dependent cellular cytotoxicity in cancer immunotherapy. Frontiers in Immunology, 6. [CrossRef]
- Wei, G., Zhu, J., Hu, H. B., & Liu, J. Q. (2021, March). Circular RNAs: Promising biomarkers for cancer diagnosis and prognosis. Gene, 771, 145365. [CrossRef]
- Wesselhoeft, R. A., Kowalski, P. S., & Anderson, D. G. (2018). Engineering circular RNA for potent and stable translation in eukaryotic cells. Nature Communications, 9(1). [CrossRef]
- Wesselhoeft, R. A., Kowalski, P. S., Parker-Hale, F. C., Huang, Y., Bisaria, N., & Anderson, D. G. (2019, May). RNA Circularization Diminishes Immunogenicity and Can Extend Translation Duration In vivo. Molecular Cell, 74(3), 508-520.e4. [CrossRef]
- Wu, C. J. (2018). Abstract IA11: Developing and improving personalized neoantigen-targeting cancer vaccines. Cancer Immunology Research, 6(9_Supplement), IA11. [CrossRef]
- Xie, J., Ye, F., Deng, X., Tang, Y., Liang, J.-Y., Huang, X., Sun, Y., Tang, H., Lei, J., Zheng, S., & Zou, Y. (2023). Circular RNA: A promising new star of vaccine. Journal of Translational Internal Medicine, 11(4), 372–381. [CrossRef]
- Xin, Y., Huang, M., Guo, W., Huang, Q., Zhang, L. Z., & Jiang, G. (2017). Nano-based delivery of RNAi in cancer therapy. Molecular Cancer, 16(1). [CrossRef]
- Xu, S., Yang, K., Li, R., & Zhang, L. (2020, September 9). mRNA Vaccine Era—Mechanisms, Drug Platform and Clinical Prospection. International Journal of Molecular Sciences, 21(18), 6582. [CrossRef]
- Yang, H., Wang, W., Romano, K. A., Gu, M., Sanidad, K. Z., Kim, D., Yang, J., Schmidt, B., Panigrahy, D., Pei, R., Martin, D. A., Ozay, E. I., Wang, Y., Song, M., Bolling, B. W., Xiao, H., Minter, L. M., Yang, G. Y., Liu, Z., . . . Zhang, G. (2018). A common antimicrobial additive increases colonic inflammation and colitis-associated colon tumorigenesis in mice. Science Translational Medicine, 10(443). [CrossRef]
- Yang, Q., Li, F., He, A. T., & Yang, B. B. (2021a, May). Circular RNAs: Expression, localization, and therapeutic potentials. Molecular Therapy, 29(5), 1683–1702. [CrossRef]
- Yang, X., Fan, J., Wu, Y., Ma, Z., Huang, J., Zhang, Y., Zhou, Z., Mo, F., Liu, X., Yuan, H., Xu, Y., Pan, L., & Chen, S. (2021b, October). Synthetic multiepitope neoantigen DNA vaccine for personalized cancer immunotherapy. Nanomedicine: Nanotechnology, Biology and Medicine, 37, 102443. [CrossRef]
- Yao, T., Chen, Q., Shao, Z., Song, Z., Fu, L., & Xiao, B. (2018). Circular RNA 0068669 as a new biomarker for hepatocellular carcinoma metastasis. Journal of Clinical Laboratory Analysis, 32(8). [CrossRef]
- Yuan, X., Diao, J., Du, A., Wen, S., Zhou, L., & Pan, Y. (2020). Circular RNA expression profiles and features in NAFLD mice: a study using RNA-seq data. Journal of Translational Medicine, 18(1). [CrossRef]
- Yue, S., Li, Y., Qiao, Z., Song, W., & Bi, S. (2021, November). Rolling Circle Replication for Biosensing, Bioimaging, and Biomedicine. Trends in Biotechnology, 39(11), 1160–1172. [CrossRef]
- Zhang, P., Gao, C., Huang, X., Lu, J., Guo, X., Shi, G., Cai, J., & Ke, A. (2020a). Cancer cell-derived exosomal circUHRF1 induces natural killer cell exhaustion and may cause resistance to anti-PD1 therapy in hepatocellular carcinoma. Molecular Cancer, 19(1). [CrossRef]
- Zhang, Q., Wang, W., Zhou, Q., Chen, C., Yuan, W., Liu, J., Li, X., & Sun, Z. (2020b, January 23). Roles of circRNAs in the tumour microenvironment. Molecular Cancer, 19(1). [CrossRef]
- Zhang, Z. G., Buller, B., & Chopp, M. (2019). Exosomes—Beyond stem cells for restorative therapy in stroke and neurological injury. Nature Reviews Neurology, 15(4), 193–203. [CrossRef]
- Zhang, Z., Lu, M., Qin, Y., Gao, W., Tao, L., Su, W., & Zhong, J. (2021, April 16). Neoantigen: A New Breakthrough in Tumor Immunotherapy. Frontiers in Immunology, 12. [CrossRef]
- Zhao, X., Zhong, Y., Wang, X., Shen, J., & An, W. (2022). Advances in Circular RNA and Its Applications. International Journal of Medical Sciences, 19(6), 975–985. [CrossRef]
Figure 1.
Brief timeline about the development of cancer immunotherapy. [1] - Romero and Coukos, 2014; [2] - Cardillo et al., 2021; [3] - Sanger et al., 1976; [4] - Cech et al., 1981; [5] - Hoover et al., 1993; [6] - Nigro et al., 1991; [7] - Puttaraju and Been, 1992; [8] - Kazakov et al., 2006; [9] - Petkovic and Müller, 2015; [10] - Chen and Sarnow, 1995; [11] - Pasman et al., 1996, Schindewolf et al., 1996 [12] - Liu et al., 2022; [13] - Beck et al., 2021; [14] - Kantoff et al., 2010; [15] - Karikó et al., 2011; [16] - Ashwal-Fluss et al., 2014; [17] - Kleaveland et al., 2018, Piwecka et al., 2017; [18] - Patop and Kadener, 2018, Kristensen et al., 2018; [19] - Fan et al., 2018, Liu et al., 2018, Le Moignic et al., 2018, Islam et al., 2018, Haabeth et al., 2018; [20] - Chen et al., 2019b; [21] - Tateshita et al., 2019, Hewitt et al., 2019, Hassett et al., 2019, Verbeke et al., 2019; [22] - Baxevanis et al., 2020; [23] - Zhang et al., 2020b, Son et al., 2020, Sebastian et al., 2019, Mai et al., 2020; [24] - ClinicalTrials.gov, n.d.; [25] - Feng et al., 2023. Abréviations : DCC - Deleted in Colorectal Carcinoma; TAA - Tumor-Associated Antigens; PIE - Permuted Intron-Exon; CLAN - Cationic Lipid-assisted Nanoparticle; LCP NPs - Lipid-Calcium-Phosphate Nanoparticles; triMN-LPR - Triply-modified Nanoparticle Liposome-Protamine-RNA; PGCP NPs - Polyethylene Glycol-coated Calcium Phosphate Nanoparticles; ssPalmE-KALA - Sessile Polymeric Micelle with E/K Peptides for Antigen Loading; Galsome - Galactose-conjugated Liposome; DOTAP/DP7-C/LL2 - Dioleoyltrimethylammonium Propane / DP7 Conjugate / LL2 Peptide; RNA-seq - Ribonucleic Acid Sequencing; COVID-19 - Coronavirus Disease 2019; NCT - National Clinical Trial; BNT111 - BioNTech's cancer vaccine.
Figure 1.
Brief timeline about the development of cancer immunotherapy. [1] - Romero and Coukos, 2014; [2] - Cardillo et al., 2021; [3] - Sanger et al., 1976; [4] - Cech et al., 1981; [5] - Hoover et al., 1993; [6] - Nigro et al., 1991; [7] - Puttaraju and Been, 1992; [8] - Kazakov et al., 2006; [9] - Petkovic and Müller, 2015; [10] - Chen and Sarnow, 1995; [11] - Pasman et al., 1996, Schindewolf et al., 1996 [12] - Liu et al., 2022; [13] - Beck et al., 2021; [14] - Kantoff et al., 2010; [15] - Karikó et al., 2011; [16] - Ashwal-Fluss et al., 2014; [17] - Kleaveland et al., 2018, Piwecka et al., 2017; [18] - Patop and Kadener, 2018, Kristensen et al., 2018; [19] - Fan et al., 2018, Liu et al., 2018, Le Moignic et al., 2018, Islam et al., 2018, Haabeth et al., 2018; [20] - Chen et al., 2019b; [21] - Tateshita et al., 2019, Hewitt et al., 2019, Hassett et al., 2019, Verbeke et al., 2019; [22] - Baxevanis et al., 2020; [23] - Zhang et al., 2020b, Son et al., 2020, Sebastian et al., 2019, Mai et al., 2020; [24] - ClinicalTrials.gov, n.d.; [25] - Feng et al., 2023. Abréviations : DCC - Deleted in Colorectal Carcinoma; TAA - Tumor-Associated Antigens; PIE - Permuted Intron-Exon; CLAN - Cationic Lipid-assisted Nanoparticle; LCP NPs - Lipid-Calcium-Phosphate Nanoparticles; triMN-LPR - Triply-modified Nanoparticle Liposome-Protamine-RNA; PGCP NPs - Polyethylene Glycol-coated Calcium Phosphate Nanoparticles; ssPalmE-KALA - Sessile Polymeric Micelle with E/K Peptides for Antigen Loading; Galsome - Galactose-conjugated Liposome; DOTAP/DP7-C/LL2 - Dioleoyltrimethylammonium Propane / DP7 Conjugate / LL2 Peptide; RNA-seq - Ribonucleic Acid Sequencing; COVID-19 - Coronavirus Disease 2019; NCT - National Clinical Trial; BNT111 - BioNTech's cancer vaccine.
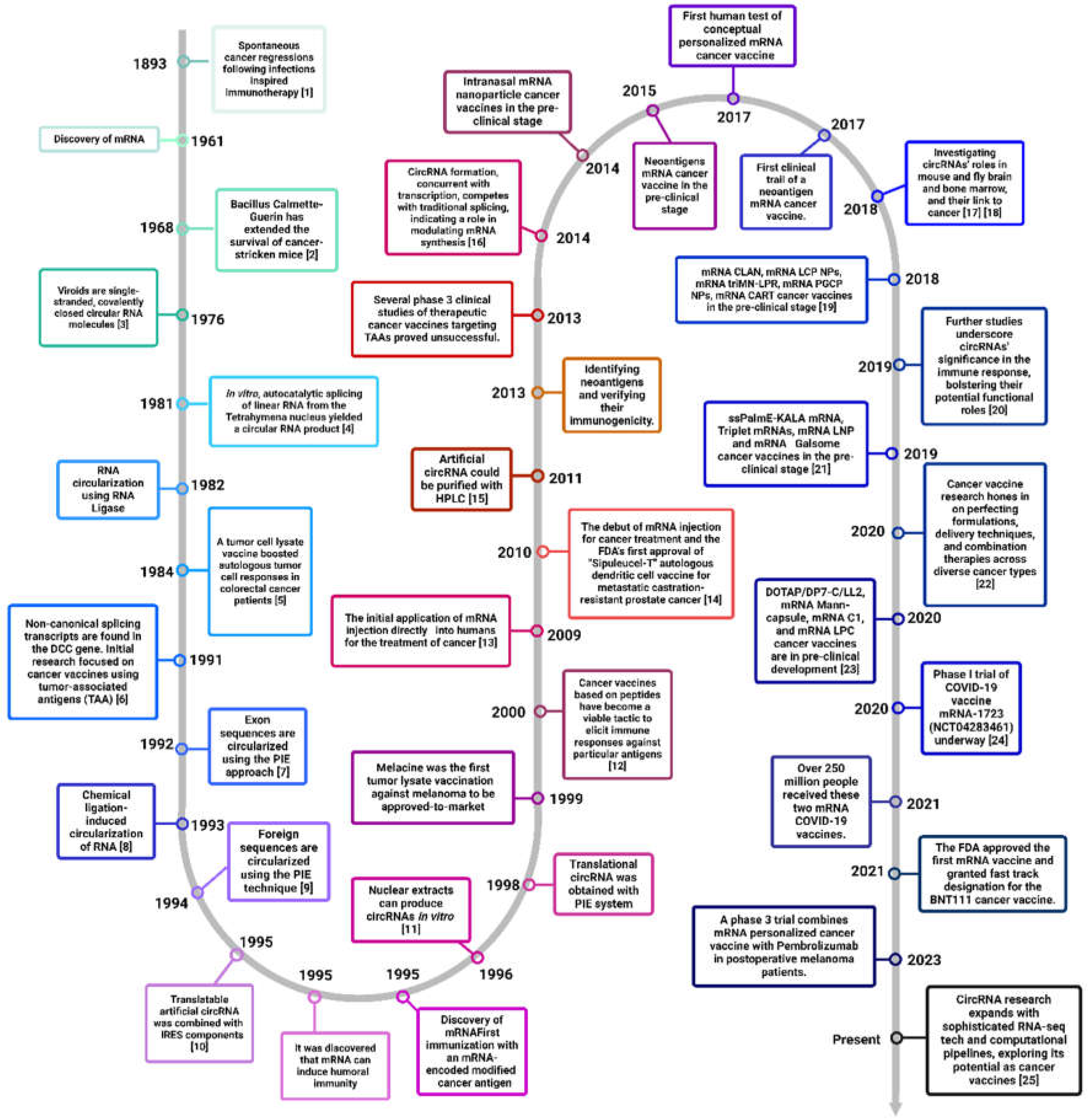
Figure 2.
Illustration of how circRNA is better than linear RNA w.r.t exonuclease activity: A) Since each mRNA has a 3 -poly (A) tail and a 5 -cap, cap-dependent translation can be used to translate it. Exonucleases from the 5 and/or 3 ends break down the majority of mRNAs. B) Because circRNAs don't have 5' and 3' ends, they can translate caps independently and are resistant to exonuclease destruction. (Sun & Yang, 2023).
Figure 2.
Illustration of how circRNA is better than linear RNA w.r.t exonuclease activity: A) Since each mRNA has a 3 -poly (A) tail and a 5 -cap, cap-dependent translation can be used to translate it. Exonucleases from the 5 and/or 3 ends break down the majority of mRNAs. B) Because circRNAs don't have 5' and 3' ends, they can translate caps independently and are resistant to exonuclease destruction. (Sun & Yang, 2023).
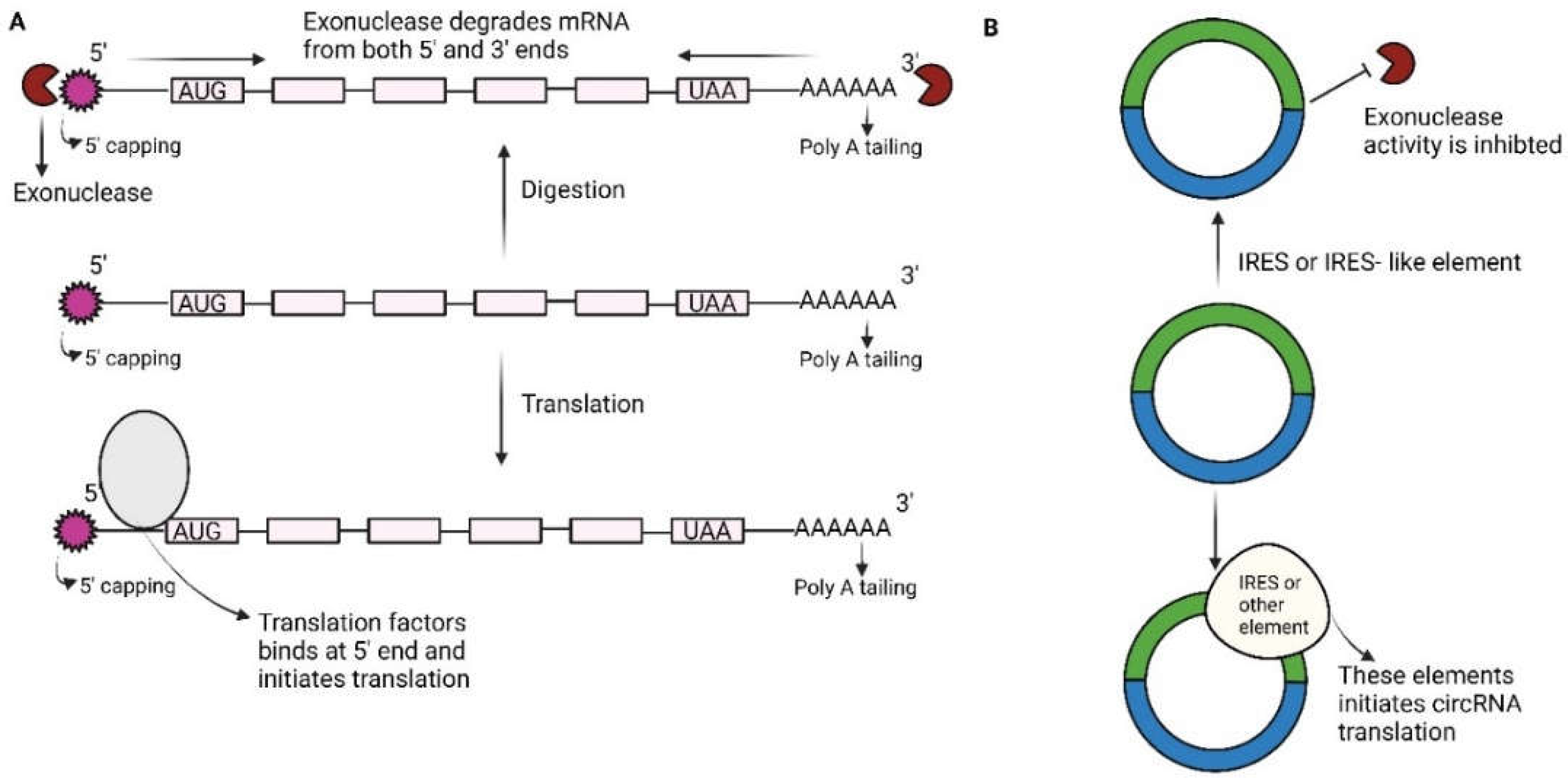
Figure 3.
Multifaceted Roles of circRNA in Cellular Processes: Regulation of RBPs, Transcription, miRNA Sponges, Apoptosis, Tumor Microenvironment Control, Signalling Pathway Modulation, and Metastasis.
Figure 3.
Multifaceted Roles of circRNA in Cellular Processes: Regulation of RBPs, Transcription, miRNA Sponges, Apoptosis, Tumor Microenvironment Control, Signalling Pathway Modulation, and Metastasis.
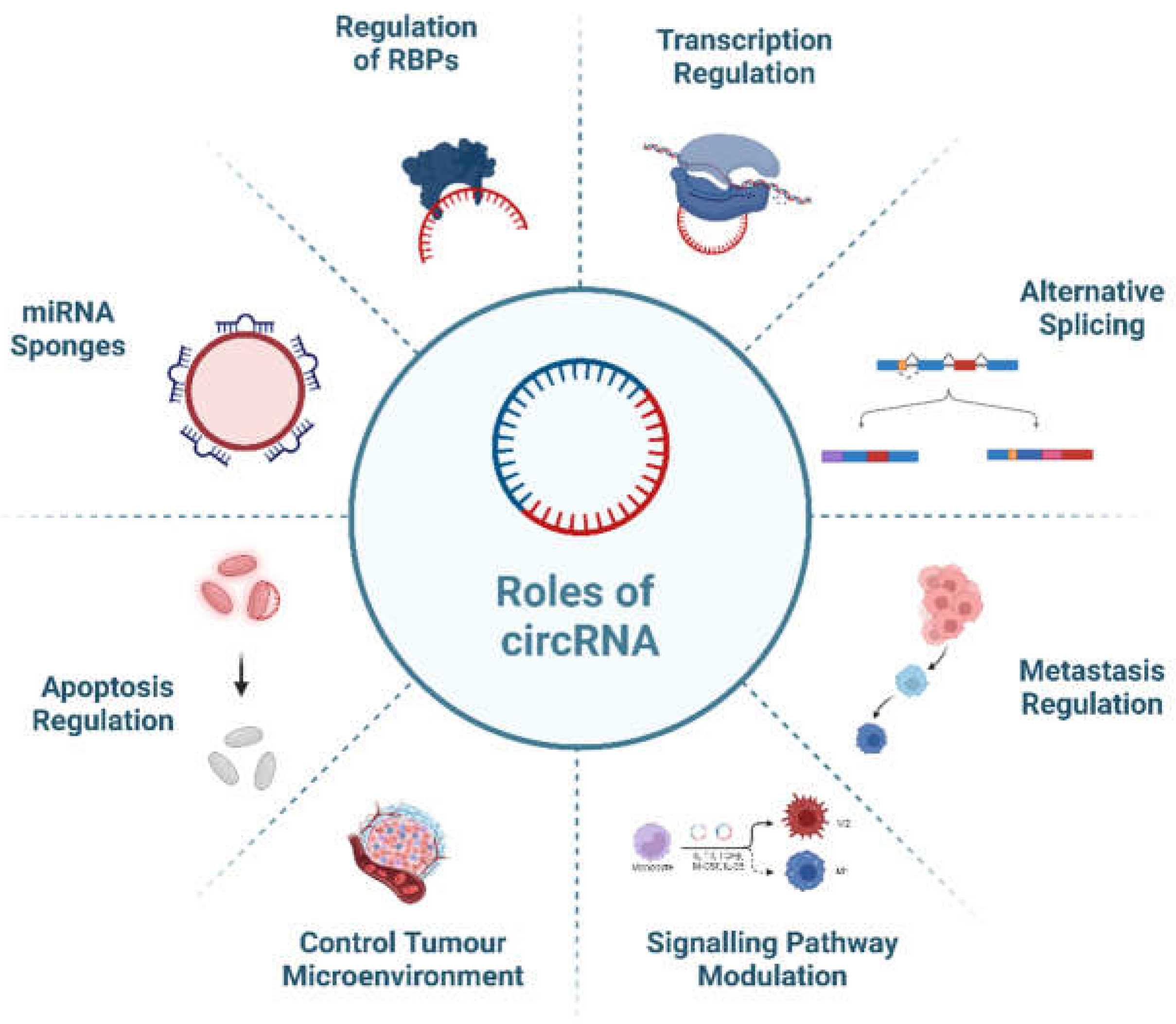
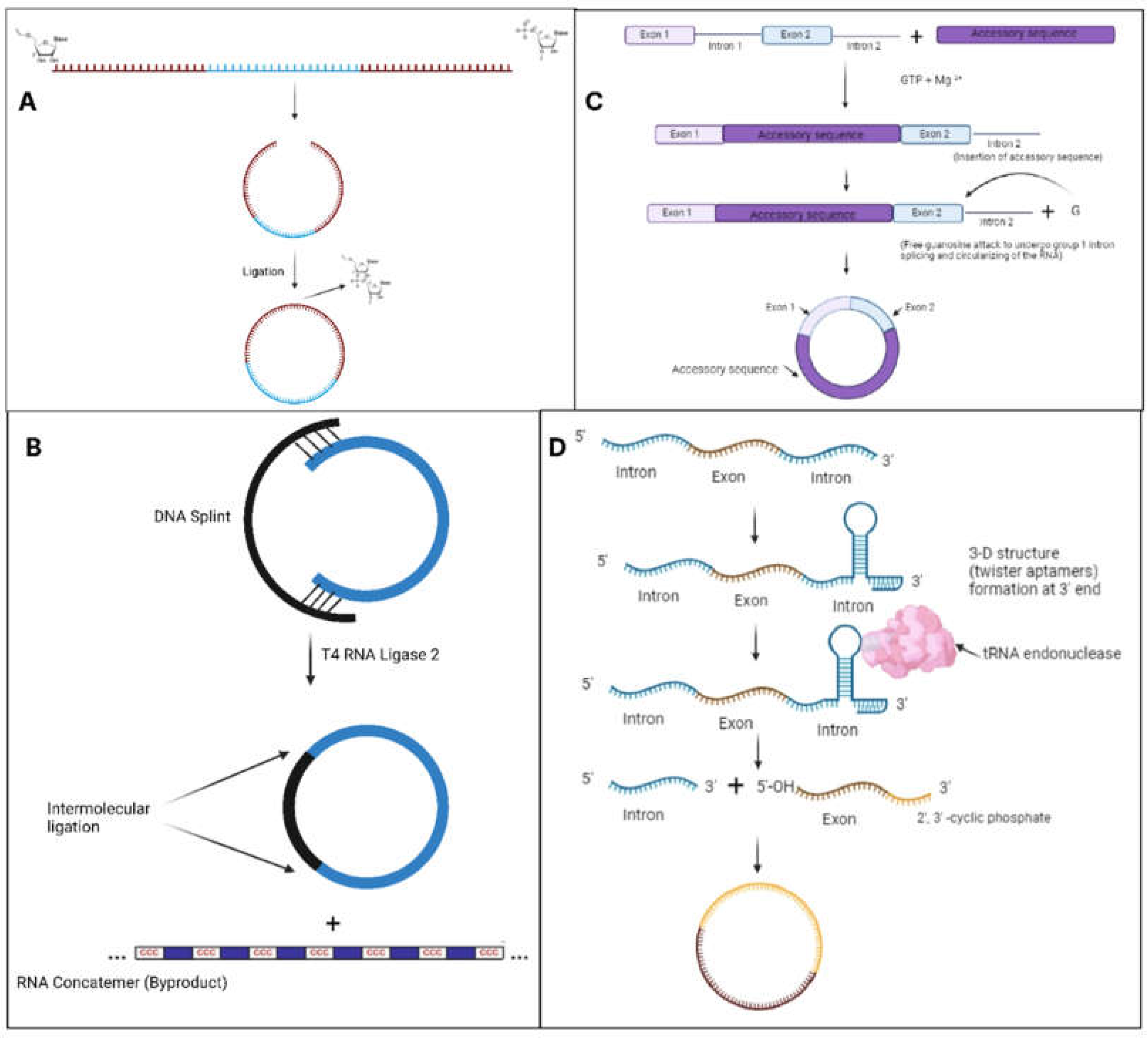
Figure 4.
Diagrammatic representation of several RNA circularization techniques: a) chemical ligation of RNA through the conjugation of 3'-end hydroxyl and 5'-end phosphate. b) DNA splints that bring the ends of RNA close together to enable site-specific ligation are used in T4 RNA ligase 2-catalyzed enzymatic RNA ligation. c) RNA ligation based on the PIE mechanism. A PIE construct is created by permuting a native group I intron and inserting a custom RNA (added to enhance circRNA stability and effectiveness) into the exonic region. In the presence of free guanosine, the PIE construct spontaneously circularizes to form the native exonic RNA's circRNA. d) Twister aptamers are special RNA sequences that can form specific three-dimensional structures.
Figure 4.
Diagrammatic representation of several RNA circularization techniques: a) chemical ligation of RNA through the conjugation of 3'-end hydroxyl and 5'-end phosphate. b) DNA splints that bring the ends of RNA close together to enable site-specific ligation are used in T4 RNA ligase 2-catalyzed enzymatic RNA ligation. c) RNA ligation based on the PIE mechanism. A PIE construct is created by permuting a native group I intron and inserting a custom RNA (added to enhance circRNA stability and effectiveness) into the exonic region. In the presence of free guanosine, the PIE construct spontaneously circularizes to form the native exonic RNA's circRNA. d) Twister aptamers are special RNA sequences that can form specific three-dimensional structures.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



