
Submitted:
26 September 2024
Posted:
30 September 2024
You are already at the latest version
Abstract
Colorectal cancer (CRC) is the third most common neoplasm in the world and the second with the highest mortality rate. Single nucleotide polymorphisms (SNPs) in miRNA genes (mirSNPs) may be related to dysregulated miRNA expression in several neoplasms. This systematic review aims to investigate studies that investigate SNPs located in regions of miRNA genes that influence their expression and are associated with CRC, as well as their potential as biomarkers for the disease, based on the available literature. For this, searches were performed in public databases, including MEDLINE/PubMed, Embase, Web of Science and Scopus. The rigorous review of the PRISMA 2020 guidelines and the methodological quality of the studies were assessed sing the Newcastle-Ottawa scale and the Mixed Methods Assessment Tool. Of the 175 studies identified, 26 were considered eligible: 18 of them highlighted mirSNPs as potential biomarkers of risk and prognosis for CRC; 4 studies suggested a protective role; 1 study linked mirSNPs to treatment; and 3 studies found no relevant evidence. These results highlight the need for further research on this topic, since these variants have the potential to serve as biomarkers and may be useful in assessing risk, prognosis, and developing therapeutic strategies for patients with CRC.
Keywords:
miRNA
; SNP
; MirSNP
; colorectal cancer
; biomarkers
Introduction
Colorectal cancer (CRC) is a malignancy affecting the large intestine and rectum and is notable for being the third most common cancer and the second leading cause of cancer-related deaths worldwide [1]. Most cases of CRC arise due to a combination of genetic and environmental factors, with several elements contributing to an increased risk of development. Modifiable risk factors include diet, obesity, physical inactivity, smoking, and alcohol consumption, while non-modifiable factors include gender, ethnicity, personal history of inflammatory bowel disease, personal or family history of polyps or cancer, inherited conditions such as Lynch syndrome, and genes involved in carcinogenesis [2]. Current methods of diagnosis and prognostic monitoring of CRC include colonoscopy and flexible sigmoidoscopy; however, these techniques are invasive, uncomfortable, and often suffer from poor patient compliance [3]. Alternative methods such as fecal occult blood tests, fecal immunochemical tests, carcinoembryonic antigen (CEA), and carbohydrate antigen 19–9 (CA19–9) are plagued by high false-positive rates and low sensitivity. Each method has distinct advantages and limitations that impact clinical decision-making. Consequently, there is an increasing focus on research into individualized genetic risk profiles to improve the diagnosis and prognosis of CRC [4,5].
Numerous studies aim to identify molecular markers that may contribute to the development of personalized medicine by improving disease prediction and patient outcomes [6]. Advances in molecular biology techniques are increasingly aimed at developing less invasive methods to obtain accurate information about tumors [7]. Among the widely studied markers, microRNAs (miRNAs) hold significant promise due to their differential expression in tumors and their presence in various body fluids, along with their stability under different storage and handling conditions [8,9,10,11].
MiRNAs are small non-coding RNAs, typically approximately 22 nucleotides in length, that regulate gene expression by binding to complementary sites within the 3′-untranslated regions (3′-UTRs) of target mRNAs, thereby inhibiting mRNA translation [12]. These molecules play a crucial role in cancer biology, influencing processes such as cell proliferation, differentiation, migration, angiogenesis, and apoptosis [13]. Research indicates that dysregulated miRNA expression is essential in the development and metastasis of CRC, with miRNAs acting as tumor suppressors or oncogenes by regulating the expression of specific mRNA targets [14]. Single nucleotide polymorphisms (SNPs) in the 3′-UTR, known as mirSNPs, can affect the binding affinity between miRNAs and target mRNAs, leading to alterations in miRNA expression and maturation, as well as changes in the expression levels of target genes [15,16]. These genetic variants can occur in regions such as pre-miRNAs, mature miRNAs, seed region or miRNA binding sites, significantly impacting miRNA function and contributing to susceptibility to diseases, including cancer [17,18]. In this systematic review, we specifically focused on genetic variants in miRNA binding sites associated with CRC, exploring their potential as biomarkers for this disease, in alignment with the existing literature.
Methods
- Protocol and registration
This systematic review follows the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) guidelines (Table S1) [19]. The review protocol was registered in the International Prospective Register of Systematic Reviews (PROSPERO) under number CRD42024574543. Following registration, changes to the protocol were made and recorded and can be accessed at https://www.crd.york.ac.uk/prospero/display_record.php?ID=CRD42024574543.
- Inclusion and exclusion criteria
Eligibility criteria were carefully formulated to identify studies addressing polymorphisms in miRNA binding binding sites in target genes associated with CRC. This review included observational and clinical studies published without date restrictions, peer-reviewed in English and published in scientific journals. Review studies, systematic reviews, books, case reports, short communications, editorials, letters to the editor, preprints, conference proceedings, dissertations, theses, preclinical studies and studies addressing polymorphisms in lcnRNAs were excluded.
- Literature search strategy
We conducted a systematic search for original articles in the literature in July 2024, using the MEDLINE/PubMed, Embase, Scopus, and Web of Science databases. The search strategy employed a wide range of keywords, aligned with the study objectives, and Boolean operators (AND, OR, NOT) were strategically employed to refine the search terms. The following terms were used: “colorectal cancer mirSNP” OR “neoplasm colorectal mirSNP” AND “miRNA SNP colorectal cancer” OR “miRNA SNP colorectal neoplasm”. After the search, the articles were imported into the RAYYAN software, accessed at https://www.rayyan.ai, for duplicate removal. Two authors (KAS and LMCAS) independently screened the studies by evaluating titles and abstracts based on predefined eligibility criteria. After the initial screening, the full texts of the articles were reviewed by both researchers independently for final inclusion. In case of discrepancy between researchers, the opinion of a third author (KSCS) was requested.
- Data
- collection
The bibliographic search in the databases resulted in 175 records. After eliminating 66 duplicates, 109 titles and abstracts were evaluated by two independent reviewers. Of these, 47 studies were excluded for not meeting the inclusion and exclusion criteria, leaving 62 studies for full reading. Among these, 36 studies were discarded for not meeting the eligibility criteria. As a result, 26 articles were considered eligible and included in the systematic review, as illustrated in the article screening process flowchart (Figure 1). Information about authors, year of publication, study type, study population, miRNA SNP location, altered miRNA, SNP rsID, target gene, and miRNA SNP effect on CRC were extracted from each study when available (Table 1).
- Quality assessment and risk of bias
Two reviewers (KAS and LMCAS) independently assessed the methodological quality and risk of bias of the studies included in the systematic review. The Mixed Methods Assessment Tool (MMAT) [20] was used for this assessment, as detailed in Table S2. The adapted Newcastle-Ottawa scale [21] was used to assess the risk of bias in case-control and cohort studies, with scores classified as very high risk of bias (0–3), high risk (4–6), or high methodological quality (7–9) (Table S3). Discrepancies in assessment were resolved through discussion between the reviewers or, if necessary, by consulting a third researcher.
Results
This review analyzed 26 studies conducted between 2008 and 2022 on SNPs that affect miRNA expression and their influence on the occurrence and progression of CRC in different populations. Most studies adopted a non-randomized quantitative design (96.15%), with emphasis on research conducted predominantly in the Chinese population [23,24,25,26,32,33,38,39,40,41,43,45]. The most frequently cited miRNA was miR-618 [22,40,42]. Of note, the SNP rs1051690 located in the miR-618 target gene region was associated with an increased risk of CRC in one study. In contrast, the SNP rs2682818 in mature miR-618 was correlated with a reduced risk. These discrepancies highlight that, in the case of miR-618, different SNPs (rs1051690 and rs2682818) may affect its expression and activity in distinct ways, resulting in opposite effects on CRC risk, as evidenced by the different results of the studies analyzed.
Eighteen studies highlighted mirSNPs as potential biomarkers of CRC risk and prognosis. Among the mirSNPs associated with CRC risk, the following were identified: SNP rs17281995 (miR-337, miR-582, miR-200a, miR-184, and miR-212) and rs1051690 (miR-618 and miR-612) [22], rs141178472 (miR-520a) [23], rs3814058 (hsa-miR-129-5p) [24], rs6504593 (miR-21) [25], rs1062044 (miR-423-5p) [26], rs2279398 (miR-370) [27], rs7930 (miR-4273-5p) [28], rs868 (let-7) [29], rs8176318 (hsa-miR-525-5p) and rs8905 (hsa-miR-214-3p) [30], rs2292832 (hsa-mir-149) and rs2910164 (hsa-mir-146a) [31] and rs895819 (miR-27a) [32].
The highlighted mirSNPs associated with prognosis were: rs2910164 (miR-146a) [33], rs12373 (miR-571) [34], rs12904 (miR-200C and miR-429) [35], KRAS-LCS6 (let-7) [36], rs4919510 (hsa-miR-608) [37], rs11614913 (miR-196a2) [38], rs2273626 (hsa-miR-4707-3p) and rs202195689 (hsa-miR-4274) [39]. Furthermore, four studies identified mirSNPs as potential protective biomarkers: rs2682818 (miR-618) [40], rs187960998 (miR-211) [41], rs2682818 (miR-618) [42], and rs35301225 (miR-34a) [43]. One study reported a mirSNP associated with the treatment of CRC patients: rs4919510 (miR-608) [44]. Finally, three studies found nonsignificant associations between mirSNPs and CRC: rs11614913 (miR-196a2) [45,46] and rs12904 (miR-200c) [47] (Table 1).
- mirSNPs associated with the risk and prognosis of CRC
Ten studies highlighted CRC risk-associated polymorphisms located in miRNA target gene regions [22,23,24,25,26,27,28,29,30,31] while one study investigated polymorphisms located in mature miRNA regions [32]. The earliest study, conducted by Landi et al. [22], included 697 CRC cases and 624 controls in the Czech Republic. This study identified two relevant SNPs: rs17281995 in the Cluster of Differentiation 86 (CD86) gene and rs1051690 in the insulin receptor (INSR). Both SNPs affect miRNA binding sites and are associated with an increased CRC risk. Furthermore, the G/A genotype in the 3′ UTR of INSR demonstrated a significant biological impact on the target miRNA. In a study conducted by Ding et al. [23] with 386 cases and 394 controls in China, it was identified that the SNP rs141178472, located in the miR-520a binding site of the phosphatidylinositol-4,5-bisphosphate 3-kinase catalytic subunit alpha gene (PIK3CA), is associated with the risk of CRC. Individuals with the CC genotype had a significantly higher risk of CRC compared with those with the TT genotype. In the same year, Ni et al. [24] performed an in vitro experiment involving 1033 CRC cases and 1147 controls from the same Chinese population. The results suggested that the SNP rs3814058 C>T in the target site of hsa-miR-129-5p may influence the expression of the pregnane X receptor (PXR) gene, increasing the susceptibility to the development of CRC. Another study conducted in the Chinese population by Xie et al. [25] analyzed 1147 individuals with CRC and 1203 controls in a case-control study. The authors found that the SNP rs6504593, located in the miR-21 binding site in insulin-like growth factor 2 (IGF2BP1) mRNA-binding protein, may modify the expression of this gene and contribute to the risk of CRC. In vitro analysis revealed that the T allele of rs6504593 negatively regulates the post-transcriptional expression of IGF2BP1 by altering the binding affinity of miR-21. Inhibition of miR-21 led to decreased cell growth, while miR-21 mimetics promoted cell proliferation and suppressed apoptosis. In a study conducted by Ke et al. [26] with 2,347 cases and 3,390 Chinese controls, it was demonstrated that the rs1062044 SNP influences the interaction between miR-423-5p and Laminin Subunit Gamma 1 (LAMC1). Carriers of the A allele had a significantly higher risk of CRC. Additionally, a negative correlation between the expression levels of miR-423-5p and LAMC1 was observed in patients with colon adenocarcinomas. Another in vitro study by Kang et al. [27] in Korea suggested that the SNP rs2279398 G>A, located in the miRNA binding site, may impact the expression of the docking protein 3 (DOK3) gene. This influence is due to the direct binding of miR-370 to the 3′-UTR of the gene. These findings indicate that genetic variations in the DOK3 gene may play a significant role in the risk of CRC. In addition, another study conducted in Korea involving 325 CRC patients and 977 controls identified the SNP rs7930 in miR-4273-5p as a potential risk factor for CRC. These results highlight the importance of genetic variations in the predisposition to the development of this disease. The results of the study by Lee et al. [28] indicated that carriers of the rs7930 AG genotype had an increased risk of CRC compared to those with the AA genotype. This increased vulnerability is related to the interaction of miR-4273-5p specifically with the A allele of the rs7930 SNP, resulting in the suppression of the expression of the TOM20 mitochondrial import receptor subunit (TOMM20) gene. Thus, the A allele is considered protective, while carriers of the G allele have an increased risk of developing CRC. Additionally, a study conducted in Spain investigated the association between the rs868 polymorphism, located in the miRNA let-7 binding site in the 3′UTR of the transforming growth factor beta receptor 1 gene (TGFBR1). The results demonstrated that the presence of this genetic variant increases the binding of TGFBR1 to the miRNA let-7b-5p, resulting in decreased TGFBR1 expression and, consequently, increasing the risk of CRC. Xicola et al. [30] also explored the influence of altered miRNA expression in SNPs located in target gene regions and their impact on cancer risk. The study revealed that two specific SNPs, rs8176318 AA, located in the 3′ UTR of the Breast Cancer 1 (BRCA1) gene, predicted to bind to miR-525-5p, and rs8905 GG, which targets the cAMP-dependent protein kinase type I regulatory subunit alpha (PRKAR1A) in miR-214-3p, were significantly associated with the risk of colon cancer in the United States population. Chayeb et al. [31] sought to clarify whether the genetic variants rs2292832 C/T in hsa-mir-149, rs2910164 G/C in hsa-mir-146a, and rs11614913 C/T in hsa-mir-196a2 are associated with the risk and prognosis of CRC. The study included 152 CRC patients and 161 Tunisian controls, and the results revealed a significant association between the SNPs rs2292832 C/T in hsa-mir-149 and rs2910164 G/C in hsa-mir-146a with the risk of CRC, considering ethnicity. However, no significant association was found for rs11614913C/T in hsa-mir-196a2. In a complementary study, carried out by Cao et al. [32] with 254 CRC patients and 238 healthy controls from the Han Chinese population, the relationship between the expression of the SNP rs895819 in miR-27a and the risk of CRC was examined. The results showed that patients with AG+GG genotypes had a significantly higher risk of CRC compared with those with the AA genotype. Furthermore, further analysis indicated that miR-27a was more highly expressed in the tumor tissue samples of patients with the GG genotype relative to those with the AA genotype.
Three studies have identified polymorphisms associated with CRC prognosis in miRNA target gene regions [33,34,35]. One study focused on complementary regions [36], while two analyzed mature miRNA regions [37,38], and another studied a SNP located in the seed region [39]. In a study conducted in China, Hu et al. [33] investigated the effects of rs2910164 and rs3746444 polymorphisms, belonging to miR-146a and miR-499, respectively, in 276 cases and 373 healthy Chinese individuals. The results demonstrated that the rs2910164 G>C SNP in miR-146a is associated with CRC susceptibility and differentiation, while the rs3746444 SNP in miR-499 showed no significant association with the disease. The authors highlighted the rs2910164 variant in miR-146a as a potential biomarker for CRC initiation and progression. In the same year, Kim et al. [34] conducted a study with 831 Korean individuals diagnosed with CRC. The analyses demonstrated that the SNPs rs3757417 T>G and rs12373 A>C in the 3′-UTR of Tyrosylprotein Sulfotransferase 1 (TPST1) and pancreatic adenocarcinoma upregulated factor (PAUF), respectively, are considered potential prognostic markers for patients undergoing surgical resection. The authors also reported that the genetic variant rs12373A>C increases the binding affinity of miR-571, resulting in the suppression of PAUF gene expression, which may impact the survival of CRC patients. In a case-control study conducted by Salem et al. [35] involving 146 CRC patients and 132 healthy individuals from Iran, it was identified that the rs12904 AA genotype plays a crucial role in the pathogenesis and metastasis of CRC. In contrast, patients carrying the rs12904 A>G genotype showed significantly reduced disease progression. The authors also observed that the rs12904 SNP in the ephrin A1 ligand gene (EFNA1) contains binding sites for the miRNAs miR-200C and miR-429, which may be important in regulating the expression of this gene. Another study, conducted by Smits et al. [36] with a significant number of Dutch individuals, evaluated a SNP located in the complementary site let-7 in the 3′-UTR of the KRAS gene (KRAS-LCS6). The analyses indicated that the T>G allele in the 3′ UTR region is associated with improved survival in patients with early-stage CRC. Thus, the KRAS-LCS6 G variant may be considered a potential prognostic biomarker for early-stage CRC. The study by Ryan et al. [37] revealed that a SNP in the mature sequence of hsa-miR-608 may have a significant impact on the prognosis of CRC patients, analyzing 245 cases and 446 controls from Caucasian and African-American populations. Although the results did not show a significant association between the rs4919510 SNP and CRC risk, a correlation with survival was identified. The GG genotype was associated with an increased risk of mortality among Caucasians, while African-Americans had a reduced risk of death, suggesting that the effect of this SNP may vary by ethnicity. Another study conducted by Zhan et al. [38], which involved 252 CRC patients and 543 healthy individuals from the Chinese population, sought to determine whether the rs11614913 SNP in miR-196a2 is related to CRC progression and susceptibility. The results indicated that the CC genotype was more frequent among CRC patients compared with controls, and that this genotype, or any variant containing at least one C allele, was associated with higher levels of mature miR-196a and increased susceptibility to CRC. However, no association with disease progression was observed. The authors emphasized that these results were evident only in patients who received chemotherapy treatment. Huang et al. [39] observed that the SNP rs2273626 A>C in hsa-miR-4707-3p was significantly associated with a decreased risk of grade 2 leukopenia and the SNP rs202195689 hsa-miR-4274 with overall survival and free survival of disease in patients who received postoperative chemoradiotherapy. These results suggest these mirSNPs as potential predictive and prognostic biomarkers in chemoradiotherapy and clinical care of patients with rectal cancer.
- mirSNPs associated with reduced risk and protection of CRC
Three studies highlighted polymorphisms associated with reduced risk and protection against CRC, located in mature miRNA regions [40,41,42], and one study focused on target gene regions [43]. In 2018, Chen et al. [40] analyzed 878 CRC patients and 884 controls from the Chinese Han population of China, investigating the relationship of the SNP rs2682818 in miR-618 with CRC. The results showed that the AA and AC/AA genotypes of rs2682818 were associated with a reduced risk of CRC compared to the CC genotype. The following year, Zhu et al. [41] studied 685 patients with colon cancer (CC) and 618 healthy controls in the same population, evaluating the SNP rs187960998, located in miR-211, and its possible relationship with the occurrence of CC. The SNP was identified as a tumor suppressor, acting in the regulation of the target gene chromodomain helicase DNA binding protein 5 (CHD5). The authors found a significant association between the rs187960998 C>T genotypes and the presence of the T allele, which correlated with smaller tumor size, better differentiation and lower probability of metastasis. Thus, the results suggest that the SNP rs187960998 in miR-211 may be related to a reduced risk of CC, acting as a protective factor by inhibiting binding to the 3′UTR of the tumor suppressor gene CHD5. In Bulgaria, Radanova et al. [42] investigated the association of the rs2682818 SNP in miR-618 with susceptibility to metastatic colon cancer (mCC) in 104 patients before initiation of chemotherapy. The results showed that the risk of developing mCC was significantly lower in patients with the heterozygous AC genotype compared with those who had the homozygous CC and AA genotypes. On the other hand, Jiang et al. [43] conducted a study with 685 newly diagnosed CRC patients and 618 controls from the Chinese population, aiming to evaluate the association of the rs35301225 SNP in miR-34a with the risk of CRC. The authors found that the rs35301225 SNP in miR-34a may act as a tumor suppressor by downregulating the tumor-promoting gene E2F Transcription Factor 1 (E2F1). Thus, this variant can be considered a protective factor against CRC. However, the authors reported that the presence of the C/A genotype results in an increased expression of E2F1, which is associated with worse survival in patients with CRC.
- mirSNPs associated with CRC treatment
Only one study has identified polymorphisms associated with CRC treatment located in mature miRNA regions. Sclafani et al. [44] investigated the role of the rs4919510 polymorphism in mature miR-608 through a retrospective study. The research evaluated the impact of perioperative treatment with neoadjuvant Capecitabine and Oxaliplatin (CAPOX), followed by chemoradiotherapy, surgery and adjuvant treatment with CAPOX and Cetuximab, in 155 patients with locally advanced rectal cancer (LARC). The results indicated that the CC genotype in miR-608 was associated with a worse prognosis compared with the CG/GG genotypes, in patients undergoing systemic chemotherapy, chemoradiotherapy, surgery and neoadjuvant chemotherapy.
- mirSNPs not associated with CRC
Several studies have analyzed the association of mirSNPs with CRC and found no statistical significance in their results. Among them, two focused on polymorphisms located in miRNA target gene regions [45,47], while one study focused on regions of mature miRNAs [46]. Chen et al. [45] investigated the relationship between SNP rs11614913 in miR-196a2 and CRC susceptibility and progression. The research involved 126 CRC patients and 407 healthy individuals from the Chinese population, but no significant association with increased risk or disease progression was identified. In 2020, Ayadilord et al. [46] conducted a case-control study with 52 CRC patients and 120 healthy Iranian individuals to evaluate the correlation between SNP rs11614913 in miR-196a2 and CRC. Genotyping results showed no statistically significant association between this SNP and CRC. Another study conducted in Iran involving 152 CRC patients and 160 controls investigated the influence of the rs12904 SNP located in the 3′UTR region of the EFNA1 gene on the regulation of miR-200c expression and CRC risk. The results also did not demonstrate a significant association between the rs12904 SNP and CRC risk in the analyzed groups [47].
- Quality assessment and risk of bias
The methodological quality of the studies included in the systematic review was carefully assessed and the results can be found in the Supplementary Material (Tables S3 and S4). Of the 22 case-control studies and 3 cohort studies analyzed, 17 case-control studies and 1 cohort study had scores between 7 and 9, reflecting high methodological quality. In contrast, 5 case-control studies and 2 cohort studies had scores of 4 to 6, raising concerns regarding the risk of bias and the validity of the results.
Discussion
In this systematic review, we performed a detailed analysis of miRNA genes (SNPs) associated with CRC, aiming to identify promising markers for early detection, risk prediction and prognosis of the disease. To achieve this objective, we reviewed 26 studies that met the established inclusion criteria.
Polymorphisms in miRNAs represent a promising class of genetic variations that deserve detailed exploration as markers for individual susceptibility to complex diseases, prognosis, and clinical decision-making [48]. These genetic variants can influence miRNA-mediated post-transcriptional regulation, resulting in changes in the expression of important genes and, consequently, contributing to interindividual variability in cancer risk [49,50,51]. Furthermore, SNPs located in miRNA target regions in genes relevant to CRC may play a significant role in its etiopathogenesis [52]. SNPs in the pri-miRNA and pre-miRNA regions can affect miRNA processing and maturation, impacting primary transcription and miRNA-target interactions, which may lead to altered expression of mature miRNA [40,53]. Similarly, SNPs in the mature sequence or seed region of the miRNA can modify interactions with target sites, altering the binding affinity between miRNAs and their target mRNA [37].
- rs4919510
- in miR-608
Two studies [37,44] found significant associations between the rs4919510 SNP in mature miR-608 and the prognosis of CRC patients. The study by Xing et al. [53] demonstrated that rs4919510 in pre-miR-608 is associated with changes in recurrence-free survival in patients undergoing chemotherapy. In contrast, another study indicated that the G allele and GG genotype are associated with decreased susceptibility to CRC, observed in codominant (CG vs. CC; GG vs. CC), dominant (GC/GG vs. CC), and recessive (GG vs. CC/CG) analyses, suggesting a protective effect among stage 0-II cases in the Chinese population [54]. This evidence corroborates what was observed in the study by Pardini et al. [55], which revealed that individuals carrying the rs4919510G variant in miR-608 had a reduced risk of recurrence compared to those carrying the wild-type genotype. However, further studies are needed to investigate the role of the rs4919510 SNP in miR-608 in relation to CRC risk and prognosis.
- rs2682818
- and rs1051690 in miR-618
Two studies have associated the rs2682818 SNP in mature miR-618 with reduced risk of CRC, especially in patients with the AC genotype [40,42]. In contrast, in the study by Landi et al. [22], the rs1051690 SNP in the miR-618 binding site was associated with an increased risk of CRC, especially in patients with the G/A genotype. Furthermore, Shao et al. [34] demonstrated that the rs2682818 SNP is associated with CRC risk, observing that individuals with the GG/TG genotypes had worse overall survival and reduced expression of mature miR-618 compared with those with the homozygous TT genotype. To date, no other studies have been identified that explored the rs1051690 SNP in the context of miR-618. Therefore, further studies are needed to investigate the functional effects of these genetic variants on miR-618 and their relationship with CRC susceptibility, aiming to improve risk stratification and develop better disease screening and prevention strategies.
- rs11614913
- in miR-196a2
Three studies found no significant association between the rs11614913 SNP in mature miR-196a2 and CRC risk, especially in individuals with the TC genotype, in two distinct populations (31,45,46). In contrast, Zhan et al. [38] reported that Chinese individuals with the CC genotype or carrying at least one C allele had higher levels of mature miR-196a and were associated with a higher risk of CRC, although no association with cancer progression was observed. Furthermore, another study indicated that the CT genotype of the rs11614913 SNP, located in the hsa-mir-196a2 pre-miRNA, was associated with an increased risk of CRC compared with the CC genotype [56]. On the other hand, a meta-analysis suggested that the TT genotype of the rs11614913 SNP might be associated with a reduced cancer risk, especially for CRC and lung cancer, in Korean and Indian populations [57]. These contradictory findings highlight the need for further investigation to better understand the impact of rs11614913 on miR-196a2 and its relationship with CRC risk.
- rs2292832
- in miR-149
Chayeb et al. [31] reported that the rs2292832 C/T variant in miR-149 and rs2910164 G/C variant in miR-146a are associated with CRC risk. A meta-analysis indicated that individuals with the TT genotype have a higher risk of CRC compared with those who carry at least one C allele [58]. Furthermore, another study suggested that individuals with the rs2292832 variant and the TT genotype have an increased risk of breast cancer. This polymorphism in pre-mir-149 may affect miR-149 processing, resulting in changes in the abundance of the mature form of the miRNA. These changes may influence both tumor progression and metastasis [59].
- rs2910164
- in miR-146a
The rs2910164 G>C SNP in the miR-146a region has been associated with CRC risk and progression in two studies [31,33]. Iguchi et al. [60] reported a significant association between the CC/CG genotypes in pre-miR-146a and liver metastasis. Another study analyzing four SNPs in the pre-miRNA region demonstrated that the rs2910164 GC genotype was associated with an increased risk of CRC, while the CC genotype was associated with a decreased risk [56]. Furthermore, a meta-analysis revealed that the C allele of rs2910164 is associated with a decreased risk of cancer in the Chinese population [57]. Another study in the same population indicated that the rs2910164 GG genotype or the G allele was associated with an increased risk of CRC, especially in men, when compared with the CC genotype or the C allele [52]. These results suggest that miR-146a may play distinct roles depending on an individual’s genotype, highlighting the need for further research to fully understand these variations.
- rs895819
- in miR-27a
The SNP rs895819 is located in the terminal loop of pre-miR-27a and would affect the secondary structure of pre-miR-27a resulting in the abnormal expression of hsa-miR-27a [61]. Cao et al. [32] demonstrated in their study that the GG genotype of rs895819 in miR-27a was significantly associated with the risk of CRC and metastasis compared to the AA genotype in the Chinese population. A similar finding in an independent study performed in the same population suggested that the GG genotype of rs895819 was significantly associated with the risk of CRC and that the G allele was associated with a greater risk of progression. In contrast, patients with the A allele (AA/GA) were significantly associated with decreased risk of CRC [62]. Wang et al. [63] demonstrated that individuals with the GG genotype and G allele exhibited a significantly increased risk of CRC in the same population. On the other hand, a study carried out in the European population showed no association between the rs895819T>C polymorphism in miR-27a and the risk of CRC [64], demonstrating the need for further studies including other populations of different ethnic origins.
- rs187960998
- in miR-211
Only one study identified the SNP rs187960998 in mature miR-211 as a tumor suppressor, demonstrating that it inhibits the proliferation and invasion of CC cells through the upregulation of CHD5 [41]. Other studies have focused on the expression of miR-211 and its role in CRC, without establishing a clear association with the presence of the genetic variant [65,66]. Therefore, further investigations are needed to clarify the impact of the polymorphism in miR-211 in the context of cancer.
- rs2273626
- in miR-4707 and rs202195689 in miR-4274
The current literature on the rs2273626 variant in miR-4707 and rs202195689 in miR-4274 in the context of cancer is limited. Huang et al. [39] associated these variants, located in miRNA seed regions, with a reduced risk of grade 2 leukopenia, as well as improved overall survival and disease-free survival. Ghanbari et al. [67] demonstrated that the rs2273626 SNP in miR-4707 influences the interaction with the glaucoma-related target gene Caspase Recruitment Domain Family Member 10 (CARD10), resulting in elevated levels of this gene. Additionally, Natalia et al. [68] suggested that the rs1553867776 variant in miR-4274 is associated with susceptibility to intestinal gastric cancer.
- rs8176318
- in miR-525-5p
The study by Mullany et al. [30] highlighted that the SNP rs8176318 AA, located in the 3′ UTR of BRCA1 and predicted to bind to miR-525-5p, is associated with an increased risk of CC. In contrast, a study conducted in Pakistan revealed that women with the GG genotype in this same region had an increased risk of developing breast cancer, while those with the TT genotype had a reduced risk compared with those carrying the GG genotype [69]. However, the results of Cao et al. [70] demonstrated that the GT and GT + TT genotypes of rs8176318 in the 3′-UTR of BRCA1 could be associated with an increased risk of breast cancer compared with the GG genotype. These divergent results suggest that further research is needed to clarify the role of miR-525-5p and the rs8176318 variant in the 3′-UTR region of BRCA1, as well as their impact on different cancer types.
- rs12904
- in miR-200c
A study in the Iranian population revealed that the rs12904 AA genotype is associated with CRC pathogenesis and metastasis, while the rs12904 A>G genotype is related to reduced disease progression. Furthermore, the rs12904 SNP in the EFNA1 gene influences the expression of this gene, mediated by the microRNAs miR-200c and miR-429a [35]. In contrast, Simonian et al. [47] found no association between the rs12904 SNP and the risk of CRC in the same population. On the other hand, a study carried out in the Chinese population identified that the rs12904 SNP located in the miR-200c binding site of the EFNA1 gene is associated with an increased risk of developing gastric cancer, with the AA genotype correlating with a higher expression of EFNA1 in gastric tissues, although it has no effect on the expression of the miRNA [71].
- KRAS-LCS6
- and rs868 in let-7
Studies by Smits et al. [36] and Xicola et al. [29] investigated the genetic variants KRAS-LCS6 of the KRAS gene and rs868 of the TGFBR1 gene in the miRNA let-7 binding site. The results showed that the KRAS-LCS6 variant is associated with the prognosis of CRC, while the rs868 SNP is related to the risk of the disease. Furthermore, another study indicated that patients with metastatic colorectal cancer (mCRC) harboring the LCS6 variant had a better response to therapy with anti-EGFR monoclonal antibodies (moAbs) [72]. On the other hand, Christensen et al. [73] demonstrated that patients with head and neck squamous cell carcinoma harboring the KRAS-LCS6 variant had significantly reduced survival time, suggesting that this variant may alter the phenotype or therapeutic response in this condition. The rs868 SNP, located in the 3′UTR of the TGFBR1 gene, has been shown to have a regulatory effect on miRNA/mRNA interaction by influencing negative RNA correlation in patients with hepatitis C virus-associated end-stage liver disease. Furthermore, this variant affects the expression of miRNAs let-7 and miR98 after liver transplantation, with a particularly significant effect in carriers of the rs868 AG genotype. Despite the associations identified between KRAS-LCS6 and rs868 variants in different aspects of CRC and other diseases, several open questions remain. Additional studies are needed to better understand the mechanisms by which these variants influence risk, prognosis, and response to treatment [74].
This review presents some limitations that should be considered when interpreting the results. In addition to the scarcity of studies focused on genetic variants in specific miRNA regions for CRC, we observed significant heterogeneity among the included studies. Most participants were predominantly Caucasian or Asian, which may introduce genetic and geographic variations that influence the findings. Despite these limitations, a notable advantage of this systematic review is the ability to evaluate, based on the existing literature and studies of high methodological quality, the hypothetical associations of SNPs in miRNA regulatory regions and their potential as biomarkers for CRC.
Conclusion
In recent years, the study of polymorphisms in miRNA regions has aroused great interest in the search for new promising, less invasive, sensitive and specific biomarkers for the early detection of CRC. This systematic review provides valuable insights into studies related to the diagnosis, prognosis and treatment of CRC, focusing on mirSNPs and contributing to a better understanding of the pathways involved in the development of the disease. Our findings highlight the potential of polymorphisms in miRNAs as important tools in the early identification and management of CRC.
Declarations
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Funding
The authors received no specific funding for this work.
Authors’ contributions
K.A.S. was responsible for the concept and design of the original study. K.A.S, L.M.C.A.S, K.S.C.S and O.M.A performed a systematic search in the literature of original articles. A.D.L. and V.N.S. were responsible for the critical review of the manuscript. All authors read and approved the final version of the manuscript.
Ethics approval and consent to participate
Not applicable
Availability of data and materials
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Acknowledgements
Foundation for the Coordination of Improvement of Higher Education Personnel (CAPES), Brazil; National Council for Scientific and Technological Development (CNPq), Brazil.
Competing interests
We declare that there is no conflict of interest between the authors.
Consent for publication
Not applicable
References
- Age standardized (World) incidence rates, colorectal cancer, males, all ages. Available from: https://gco.iarc.
- Simon, K. Clinical Interventions in Aging Dovepress Colorectal cancer development and advances in screening. Clin Interv Aging [Internet]. 2016 [cited 2024 Feb 6];11–967. [CrossRef]
- Rapado-González Ó, Álvarez-Castro A, López-López R, Iglesias-Canle J, Suárez-Cunqueiro MM, Muinelo-Romay L. Circulating microRNAs as Promising Biomarkers in Colorectal Cancer. Cancers (Basel) [Internet]. 2019 Jul 1 [cited 2024 Sep 16];11(7). [CrossRef]
- Chatterjee N, Shi J, García-Closas M. Developing and evaluating polygenic risk prediction models for stratified disease prevention. Nat Rev Genet [Internet]. 2016 Jul 1 [cited 2024 Feb 6];17(7):392. [CrossRef]
- Dunlop MG, Tenesa A, Farrington SM, Ballereau S, Brewster DH, Koessler T, et al. Cumulative impact of common genetic variants and other risk factors on colorectal cancer risk in 42,103 individuals. Gut [Internet]. 2013 Jun [cited 2024 Feb 6];62(6):871. [CrossRef]
- Zygulska AL, Pierzchalski P. Novel Diagnostic Biomarkers in Colorectal Cancer. Int J Mol Sci [Internet]. 2022 Jan 1 [cited 2024 Feb 6];23(2). [CrossRef]
- Andrei P, Battuello P, Grasso G, Rovera E, Tesio N, Bardelli A. Integrated approaches for precision oncology in colorectal cancer: The more you know, the better. Semin Cancer Biol [Internet]. 2022 Sep 1 [cited 2024 Feb 6];84:199–213. Available from: https://pubmed.ncbi.nlm.nih.gov/33848627/.
- Gallo A, Tandon M, Alevizos I, Illei GG. The Majority of MicroRNAs Detectable in Serum and Saliva Is Concentrated in Exosomes. PLoS One [Internet]. 2012 Mar 9 [cited 2024 Feb 6];7(3):e30679. Available from: https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0030679.
- Ortiz-Quintero, B. Cell-free microRNAs in blood and other body fluids, as cancer biomarkers. Cell Prolif [Internet]. 2016 Jun 1 [cited 2024 Feb 6];49(3):281. P: PMCID, 6496. [Google Scholar]
- Mitchell PS, Parkin RK, Kroh EM, Fritz BR, Wyman SK, Pogosova-Agadjanyan EL, et al. Circulating microRNAs as stable blood-based markers for cancer detection. Proc Natl Acad Sci U S A [Internet]. 2008 Jul 7 [cited 2024 Feb 6];105(30):10513. [CrossRef]
- Shi Y, Liu Z, Lin Q, Luo Q, Cen Y, Li J, et al. MiRNAs and Cancer: Key Link in Diagnosis and Therapy. Genes (Basel) [Internet]. 2021 Aug 1 [cited 2024 Feb 6];12(8). [CrossRef]
- Subramaniam S, Jeet V, Gunter JH, Clements JA, Batra J. Allele-Specific MicroRNA-Mediated Regulation of a Glycolysis Gatekeeper PDK1 in Cancer Metabolism. Cancers (Basel) [Internet]. 2021 Jul 2 [cited 2024 Sep 16];13(14). Available from: https://pubmed.ncbi.nlm.nih.gov/34298795/.
- Balacescu O, Sur D, Cainap C, Visan S, Cruceriu D, Manzat-Saplacan R, et al. The Impact of miRNA in Colorectal Cancer Progression and Its Liver Metastases. Int J Mol Sci [Internet]. 2018 Dec 1 [cited 2024 Sep 16];19(12). Available from: https://pubmed.ncbi.nlm.nih.gov/30469518/.
- Radanova M, Levkova M, Mihaylova G, Manev R, Maneva M, Hadgiev R, et al. Single Nucleotide Polymorphisms in microRNA Genes and Colorectal Cancer Risk and Prognosis. Biomedicines [Internet]. 2022 Jan 1 [cited 2024 Sep 16];10(1). Available from: https://pubmed.ncbi.nlm.nih.gov/35052835/.
- Wang Z, Sun X, Wang Y, Liu X, Xuan Y, Hu S. Association between miR-27a genetic variants and susceptibility to colorectal cancer. Diagn Pathol [Internet]. 2014 Jun 30 [cited 2024 Feb 7];9(1). [CrossRef]
- Kohler AF 1993-. The functional impact of a genetic variant in a microRNA binding site in MDM4, a breast cancer-associated gene. 2020 [cited 2024 Aug 11]; Available from: https://acervodigital.ufpr.br/handle/1884/76039.
- Dias F, Morais M, Teixeira AL, Medeiros R. Involving the microRNA Targetome in Esophageal-Cancer Development and Behavior. Cancers (Basel) [Internet]. 2018 Oct 12 [cited 2024 Feb 7];10(10). [CrossRef]
- Li YJ, Zhang ZY, Mao YY, Jin MJ, Jing FY, Ye ZH, et al. A genetic variant in MIR-146a modifies digestive system cancer risk: A meta-analysis. Asian Pacific Journal of Cancer Prevention. 2014;15(1):145–50. [CrossRef]
- Page MJ, McKenzie JE, Bossuyt PM, Boutron I, Hoffmann TC, Mulrow CD, et al. The PRISMA 2020 statement: an updated guideline for reporting systematic reviews. BMJ [Internet]. 2021 Mar 29 [cited 2024 Sep 12];372.
- Pace R, Pluye P, Bartlett G, Macaulay AC, Salsberg J, Jagosh J, et al. Testing the reliability and efficiency of the pilot Mixed Methods Appraisal Tool (MMAT) for systematic mixed studies review. Int J Nurs Stud [Internet]. 2012 Jan [cited 2024 Sep 12];49(1):47–53. Available from: https://pubmed.ncbi.nlm.nih.gov/21835406/.
- Stang, A. Stang A. Critical evaluation of the Newcastle-Ottawa scale for the assessment of the quality of nonrandomized studies in meta-analyses. Eur J Epidemiol [Internet]. 2010 Sep [cited 2024 Sep 12];25(9):603–5. Available from: https://pubmed.ncbi.nlm.nih.gov/20652370/.
- Landi D, Gemignani F, Naccarati A, Pardini B, Vodicka P, Vodickova L, et al. Polymorphisms within micro-RNA-binding sites and risk of sporadic colorectal cancer. Carcinogenesis [Internet]. 2008 Mar [cited 2024 Sep 12];29(3):579–84. Available from: https://pubmed.ncbi.nlm.nih.gov/18192692/.
- Ding L, Jiang Z, Chen Q, Qin R, Fang Y, Li H. A functional variant at miR-520a binding site in PIK3CA alters susceptibility to colorectal cancer in a Chinese Han population. Biomed Res Int [Internet]. 2015 [cited 2024 Sep 12];2015. Available from: https://pubmed.ncbi.nlm.nih.gov/25834816/.
- Ni H, Su B, Pan L, Li X, Zhu X, Chen X. Functional variants inPXRare associated with colorectal cancer susceptibility in Chinese populations. Cancer Epidemiol [Internet]. 2015 Dec 1 [cited 2024 Sep 12];39(6):972–7. Available from: https://pubmed.ncbi.nlm.nih.gov/26547791/.
- Xie L, Li S, Jin J, He L, Xu K, Zhu L, et al. Genetic variant in miR-21 binding sites is associated with colorectal cancer risk. J Cell Mol Med [Internet]. 2019 Mar 1 [cited 2024 Sep 12];23(3):2012–9. Available from: https://pubmed.ncbi.nlm.nih.gov/30569605/.
- Ke J, Tian J, Li J, Gong Y, Yang Y, Zhu Y, et al. Identification of a functional polymorphism affecting microRNA binding in the susceptibility locus 1q25.3 for colorectal cancer. Mol Carcinog [Internet]. 2017 Sep 1 [cited 2024 Sep 12];56(9):2014–21. Available from: https://pubmed.ncbi.nlm.nih.gov/28277607/.
- KANG BW, JEON H-S, CHAE YS, LEE SJ, PARK JS, CHOI GS, et al. Impact of Genetic Variation in MicroRNA-binding Site on Susceptibility to Colorectal Cancer. Anticancer Res. 2016;36(7). [CrossRef]
- Lee AR, Park J, Jung KJ, Jee SH, Kim-Yoon S. Genetic variation rs7930 in the miR-4273-5p target site is associated with a risk of colorectal cancer. Onco Targets Ther [Internet]. 2016 Nov 7 [cited 2024 Sep 14];9:6885. [CrossRef]
- Xicola RM, Bontu S, Doyle BJ, Rawson J, Garre P, Lee E, et al. Association of a let-7 miRNA binding region of TGFBR1 with hereditary mismatch repair proficient colorectal cancer (MSS HNPCC). Carcinogenesis [Internet]. 2016 Aug 1 [cited 2024 Aug 9];37(8):751. [CrossRef]
- Mullany LE, Wolff RK, Herrick JS, Buas MF, Slattery ML. SNP Regulation of microRNA Expression and Subsequent Colon Cancer Risk. PLoS One [Internet]. 2015 Dec 1 [cited 2024 Aug 9];10(12). [CrossRef]
- Chayeb V, Mahjoub S, Zitouni H, Jrah-Harzallah H, Zouari K, Letaief R, et al. Contribution of microRNA-149, microRNA-146a, and microRNA-196a2 SNPs in colorectal cancer risk and clinicopathological features in Tunisia. Gene [Internet]. 2018 Aug 5 [cited 2024 Sep 14];666:100–7. Available from: https://pubmed.ncbi.nlm.nih.gov/29715515/.
- Cao Y, Hu J, Fang Y, Chen Q, Li H. Association between a functional variant in microRNA-27a and susceptibility to colorectal cancer in a Chinese Han population. Genet Mol Res [Internet]. 2014 [cited 2024 Sep 14];13(3):7420–7. [CrossRef]
- Hu X, Li L, Shang M, Zhou J, Song X, Lu X, et al. Association between microRNA genetic variants and susceptibility to colorectal cancer in Chinese population. Tumour Biol [Internet]. 2014 Mar 1 [cited 2024 Sep 14];35(3):2151–6. Available from: https://pubmed.ncbi.nlm.nih.gov/24136745/.
- Kim JG, Chae YS, Lee SJ, Kang BW, Park JY, Lee EJ, et al. Genetic variation in microRNA-binding site and prognosis of patients with colorectal cancer. J Cancer Res Clin Oncol [Internet]. 2015 Jan 1 [cited 2024 Aug 9];141(1):35–41. Available from: https://pubmed.ncbi.nlm.nih.gov/25079514/.
- Salem E, Keshvari A, Mahdavinezhad A, Soltanian AR, Saidijam M, Afshar S. Role of EFNA1 SNP (rs12904) in Tumorigenesis and Metastasis of Colorectal Cancer: A Bioinformatic Analysis and HRM SNP Genotyping Verification. Asian Pac J Cancer Prev [Internet]. 2022 [cited 2024 Aug 9];23(10):3523. [CrossRef]
- Smits KM, Paranjape T, Nallur S, Wouters KAD, Weijenberg MP, Schouten LJ, et al. A Let-7 MicroRNA SNP in the KRAS 3′UTR Is Prognostic in Early-Stage Colorectal Cancer. Clin Cancer Res [Internet]. 2011 Dec 12 [cited 2024 Aug 9];17(24):7723.
- Ryan BM, McClary AC, Valeri N, Robinson D, Paone A, Bowman ED, et al. rs4919510 in hsa-mir-608 Is Associated with Outcome but Not Risk of Colorectal Cancer. PLoS One [Internet]. 2012 May 11 [cited 2024 Sep 14];7(5). [CrossRef]
- Zhan J fang, Chen L hua, Chen Z xian, Yuan Y wei, Xie G zhu, Sun A min, et al. A functional variant in microRNA-196a2 is associated with susceptibility of colorectal cancer in a Chinese population. Arch Med Res [Internet]. 2011 Feb [cited 2024 Sep 14];42(2):144–8. Available from: https://pubmed.ncbi.nlm.nih.gov/21565628/.
- Huang Y, Feng Y, Ren H, Zhang M, Li H, Qiao Y, et al. Associations of Genetic Variations in MicroRNA Seed Regions With Acute Adverse Events and Survival in Patients With Rectal Cancer Receiving Postoperative Chemoradiation Therapy. Int J Radiat Oncol Biol Phys [Internet]. 2018 Mar 15 [cited 2024 Sep 14];100(4):1026–33. Available from: https://pubmed.ncbi.nlm.nih.gov/29485044/.
- Chen Y, Du M, Chen W, Zhu L, Wu C, Zhang Z, et al. Polymorphism rs2682818 in miR-618 is associated with colorectal cancer susceptibility in a Han Chinese population. Cancer Med [Internet]. 2018 Apr 1 [cited 2024 Sep 14];7(4):1194–200. Available from: https://pubmed.ncbi.nlm.nih.gov/29533012/.
- Zhu L, Wang R, Zhang L, Zuo C, Zhang R, Zhao S. rs187960998 polymorphism in miR-211 prevents development of human colon cancer by deregulation of 3′UTR in CHD5. Onco Targets Ther [Internet]. 2019 [cited 2024 Sep 14];12:405–12. Available from: https://pubmed.ncbi.nlm.nih.gov/30655677/.
- Radanova M, Mihaylova G, Mihaylova Z, Ivanova D, Tasinov O, Nazifova-Tasinova N, et al. Circulating miR-618 Has Prognostic Significance in Patients with Metastatic Colon Cancer. Curr Oncol [Internet]. 2021 [cited 2024 Sep 14];28(2):1204–15. Available from: https://pubmed.ncbi.nlm.nih.gov/33804070/.
- Jiang H, Ge F, Hu B, Wu L, Yang H, Wang H. rs35301225 polymorphism in miR-34a promotes development of human colon cancer by deregulation of 3′UTR in E2F1 in Chinese population. Cancer Cell Int [Internet]. 2017 Mar 9 [cited 2024 Sep 14];17(1). Available from: https://pubmed.ncbi.nlm.nih.gov/28293146/.
- Sclafani F, Chau I, Cunningham D, Lampis A, Hahne JC, Ghidini M, et al. Sequence variation in mature microRNA-608 and benefit from neo-adjuvant treatment in locally advanced rectal cancer patients. Carcinogenesis [Internet]. 2016 Sep 1 [cited 2024 Sep 14];37(9):852–7. Available from: https://pubmed.ncbi.nlm.nih.gov/27381831/.
- Chen H, Sun LY, Chen LL, Zheng HQ, Zhang QF. A variant in microRNA-196a2 is not associated with susceptibility to and progression of colorectal cancer in Chinese. Intern Med J [Internet]. 2012 Jun [cited 2024 Sep 14];42(6). Available from: https://pubmed.ncbi.nlm.nih.gov/21241442/.
- Ayadilord M, Tavakoli T, Fakharian T, Soltaninejad E, Naseri M. Relationship analysis of the miR-196a2 polymorphism (rs11614913) with colorectal cancer risk in southern Khorasan, eastern Iran. Meta Gene. 2020 Dec 1;26:100813. [CrossRef]
- Simonian M, Mosallaei M, Khosravi S, Salehi R. rs12904 polymorphism in the 3′-untranslated region of ephrin A1 ligand and the risk of sporadic colorectal cancer in the Iranian population. J Cancer Res Ther [Internet]. 2019 Jan 1 [cited 2024 Sep 14];15(1):15–9. Available from: https://pubmed.ncbi.nlm.nih.gov/30880748/.
- Zhao Y, Du Y, Zhao S, Guo Z. Single-nucleotide polymorphisms of microRNA processing machinery genes and risk of colorectal cancer. Onco Targets Ther [Internet]. 2015 Feb 12 [cited 2024 Sep 16];8:421–5. Available from: https://pubmed.ncbi.nlm.nih.gov/25709475/.
- Cipollini M, Landi S, Gemignani F. MicroRNA binding site polymorphisms as biomarkers in cancer management and research. Pharmgenomics Pers Med [Internet]. 2014 Jul 23 [cited 2024 Sep 16];7(1):173–91. Available from: https://pubmed.ncbi.nlm.nih.gov/25114582/.
- Jiang Y, Cao Y, Wang Y, Li W, Liu X, Lv Y, et al. Cysteine transporter SLC3A1 promotes breast cancer tumorigenesis. Theranostics [Internet]. 2017 [cited 2024 Sep 16];7(4):1036–46. Available from: https://pubmed.ncbi.nlm.nih.gov/28382174/.
- Shen MH, Huang CJ, Ho TF, Liu CY, Shih YY, Huang CS, et al. Colorectal cancer concurrent gene signature based on coherent patterns between genomic and transcriptional alterations. BMC Cancer [Internet]. 2022 Dec 1 [cited 2024 Sep 16];22(1). Available from: https://pubmed.ncbi.nlm.nih.gov/35637462/.
- Schneiderova M, Naccarati A, Pardini B, Rosa F, Gaetano C Di, Jiraskova K, et al. MicroRNA-binding site polymorphisms in genes involved in colorectal cancer etiopathogenesis and their impact on disease prognosis. Mutagenesis [Internet]. 2017 Oct 17 [cited 2024 Mar 7];32(5):533–42. [CrossRef]
- Xing J, Wan S, Zhou F, Qu F, Li B, Myers RE, et al. Genetic polymorphisms in pre-microRNA genes as prognostic markers of colorectal cancer. Cancer Epidemiol Biomarkers Prev [Internet]. 2012 Jan [cited 2024 Sep 16];21(1):217–27. Available from: https://pubmed.ncbi.nlm.nih.gov/22028396/.
- Ying HQ, Peng HX, He BS, Pan YQ, Wang F, Sun HL, et al. MiR-608, pre-miR-124-1 and pre-miR26a-1 polymorphisms modify susceptibility and recurrence-free survival in surgically resected CRC individuals. Oncotarget [Internet]. 2016 [cited 2024 Sep 16];7(46):75865–73. Available from: https://pubmed.ncbi.nlm.nih.gov/27713147/.
- Pardini B, Rosa F, Naccarati A, Vymetalkova V, Ye Y, Wu X, et al. Polymorphisms in microRNA genes as predictors of clinical outcomes in colorectal cancer patients. Carcinogenesis [Internet]. 2015 Jan 1 [cited 2024 Feb 29];36(1):82–6. [CrossRef]
- Lv M, Dong W, Li L, Zhang L, Su X, Wang L, et al. Association between genetic variants in pre-miRNA and colorectal cancer risk in a Chinese population. J Cancer Res Clin Oncol [Internet]. 2013 Aug [cited 2024 Sep 16];139(8):1405–10. Available from: https://pubmed.ncbi.nlm.nih.gov/23728616/.
- Xu Y, Gu L, Pan Y, Li R, Gao T, Song G, et al. Different effects of three polymorphisms in MicroRNAs on cancer risk in Asian population: evidence from published literatures. PLoS One [Internet]. 2013 Jun 4 [cited 2024 Sep 16];8(6). Available from: https://pubmed.ncbi.nlm.nih.gov/23750236/.
- Choupani J, Nariman-Saleh-Fam Z, Saadatian Z, Ouladsahebmadarek E, Masotti A, Bastami M. Association of mir-196a-2 rs11614913 and mir-149 rs2292832 Polymorphisms With Risk of Cancer: An Updated Meta-Analysis. Front Genet [Internet]. 2019 [cited 2024 Sep 15];10(MAR). Available from: https://pubmed.ncbi.nlm.nih.gov/30930933/.
- Fakhrezare F, Ebrahimi SO, Reiisi S. The association between genetic variation rs2292832 and the processing efficiency of pre-mir-149 affects the risk of breast cancer. Mol Biol Rep [Internet]. 2023 Jan 1 [cited 2024 Feb 29];50(1):679–85. Available from: https://link.springer.com/article/10.1007/s11033-022-08027-3.
- Iguchi T, Nambara S, Masuda T, Komatsu H, Ueda M, Kidogami S, et al. miR-146a Polymorphism (rs2910164) Predicts Colorectal Cancer Patients’ Susceptibility to Liver Metastasis. PLoS One [Internet]. 2016 Nov 1 [cited 2024 Sep 16];11(11):e0165912. Available from: https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0165912.
- Nguyen TTN, Tran MTH, Nguyen VTL, Nguyen UDP, Nguyen GDT, Huynh LH, et al. Single nucleotide polymorphisms in microRNAs action as biomarkers for breast cancer. Turkish J Biol [Internet]. 2020 [cited 2024 Sep 16];44(5):284. [CrossRef]
- Jiang Y, Lin DH, Xu JP, Chen WX, Zheng SJ, Song L. Genotype GG of rs895819 Functional Polymorphism Within miR-27a Might Increase Genetic Susceptibility to Colorectal Cancer in Han Chinese Population. J Clin Lab Anal [Internet]. 2016 Jul 1 [cited 2024 Sep 16];30(4):351–5. Available from: https://pubmed.ncbi.nlm.nih.gov/26302683/.
- Wang Z, Sun X, Wang Y, Liu X, Xuan Y, Hu S. Association between miR-27a genetic variants and susceptibility to colorectal cancer. Diagn Pathol [Internet]. 2014 Jun 30 [cited 2024 Sep 16];9(1).
- Kupcinskas J, Bruzaite I, Juzenas S, Gyvyte U, Jonaitis L, Kiudelis G, et al. Lack of association between miR-27a, miR-146a, miR-196a-2, miR-492 and miR-608 gene polymorphisms and colorectal cancer. Sci Reports 2014 41 [Internet]. 2014 Aug 8 [cited 2024 Sep 16];4(1):1–6. Available from: https://www.nature.com/articles/srep05993.
- Sümbül AT, Göğebakan B, Bayram S, Batmacı CY, Öztuzcu S. MicroRNA 211 expression is upregulated and associated with poor prognosis in colorectal cancer: a case–control study. Tumor Biol [Internet]. 2015 Dec 1 [cited 2024 Mar 21];36(12):9703–9. Available from: https://link.springer.com/article/10.1007/s13277-015-3708-4.
- Lotfi E, Kholghi A, Golab F, Mohammadi A, Barati M. Circulating miRNAs and lncRNAs serve as biomarkers for early colorectal cancer diagnosis. Pathol Res Pract [Internet]. 2024 Mar 1 [cited 2024 Sep 17];255. Available from: https://pubmed.ncbi.nlm.nih.gov/38377721/.
- Ghanbari M, Iglesias AI, Springelkamp H, van Duijn CM, Ikram MA, Dehghan A, et al. A Genome-Wide Scan for MicroRNA-Related Genetic Variants Associated With Primary Open-Angle Glaucoma. Invest Ophthalmol Vis Sci [Internet]. 2017 Oct 1 [cited 2024 Mar 5];58(12):5368.
- Landeros N, Corvalan AH, Musleh M, Quiñones LA, Varela NM, Gonzalez-Hormazabal P. Novel Risk Associations between microRNA Polymorphisms and Gastric Cancer in a Chilean Population. Int J Mol Sci [Internet]. 2021 Jan 1 [cited 2024 Mar 5];23(1). Available from: https://pubmed.ncbi.nlm.nih.gov/35008894/.
- Nguyen TTN, Tran MTH, Nguyen VTL, Nguyen UDP, Nguyen GDT, Huynh LH, et al. Single nucleotide polymorphisms in microRNAs action as biomarkers for breast cancer. Turkish J Biol [Internet]. 2020 [cited 2024 Sep 16];44(5):284.
- Cao J, Luo C, Yan R, Peng R, Wang K, Wang P, et al. rs15869 at miRNA binding site in BRCA2 is associated with breast cancer susceptibility. Med Oncol [Internet]. 2016 Dec 1 [cited 2024 Sep 16];33(12). Available from: https://pubmed.ncbi.nlm.nih.gov/27807724/.
- Li Y, Nie Y, Cao J, Tu S, Lin Y, Du Y, et al. G-A variant in miR-200c binding site of EFNA1 alters susceptibility to gastric cancer. Mol Carcinog [Internet]. 2014 Mar 1 [cited 2024 Sep 16];53(3):219–29. Available from: https://onlinelibrary.wiley.com/doi/full/10.1002/mc.21966.
- Saridaki Z, Weidhaas JB, Lenz HJ, Laurent-Puig P, Jacobs B, De Schutter J, et al. A let-7 microRNA-binding site polymorphism in KRAS predicts improved outcome in metastatic colorectal cancer (mCRC) patients treated with salvage cetuximab/panitumumab monotherapy. Clin Cancer Res [Internet]. 2014 Sep 9 [cited 2024 Sep 16];20(17):4499.
- Christensen BC, Moyer BJ, Avissar M, Ouellet LG, Plaza SL, Mcclean MD, et al. A let-7 microRNA-binding site polymorphism in the KRAS 3′ UTR is associated with reduced survival in oral cancers. Carcinogenesis [Internet]. 2009 [cited 2024 Sep 16];30(6):1003.
- Sajjad EA, Radkowski M, Perkowska-Ptasińska A, Pacholczyk M, Durlik M, Fedorowicz M, et al. Negative Correlation Between Hepatitis C Virus (HCV) and Let-7 MicroRNA Family in Transplanted Livers: The Role of rs868 Single-Nucleotide Polymorphism. Ann Transplant [Internet]. 2017 Oct 24 [cited 2024 Sep 16];22:638. [CrossRef]
Figure 1.
The flowchart illustrates the article selection methodology according to the recommendations of the PRISMA guidelines for systematic reviews [19].
Figure 1.
The flowchart illustrates the article selection methodology according to the recommendations of the PRISMA guidelines for systematic reviews [19].
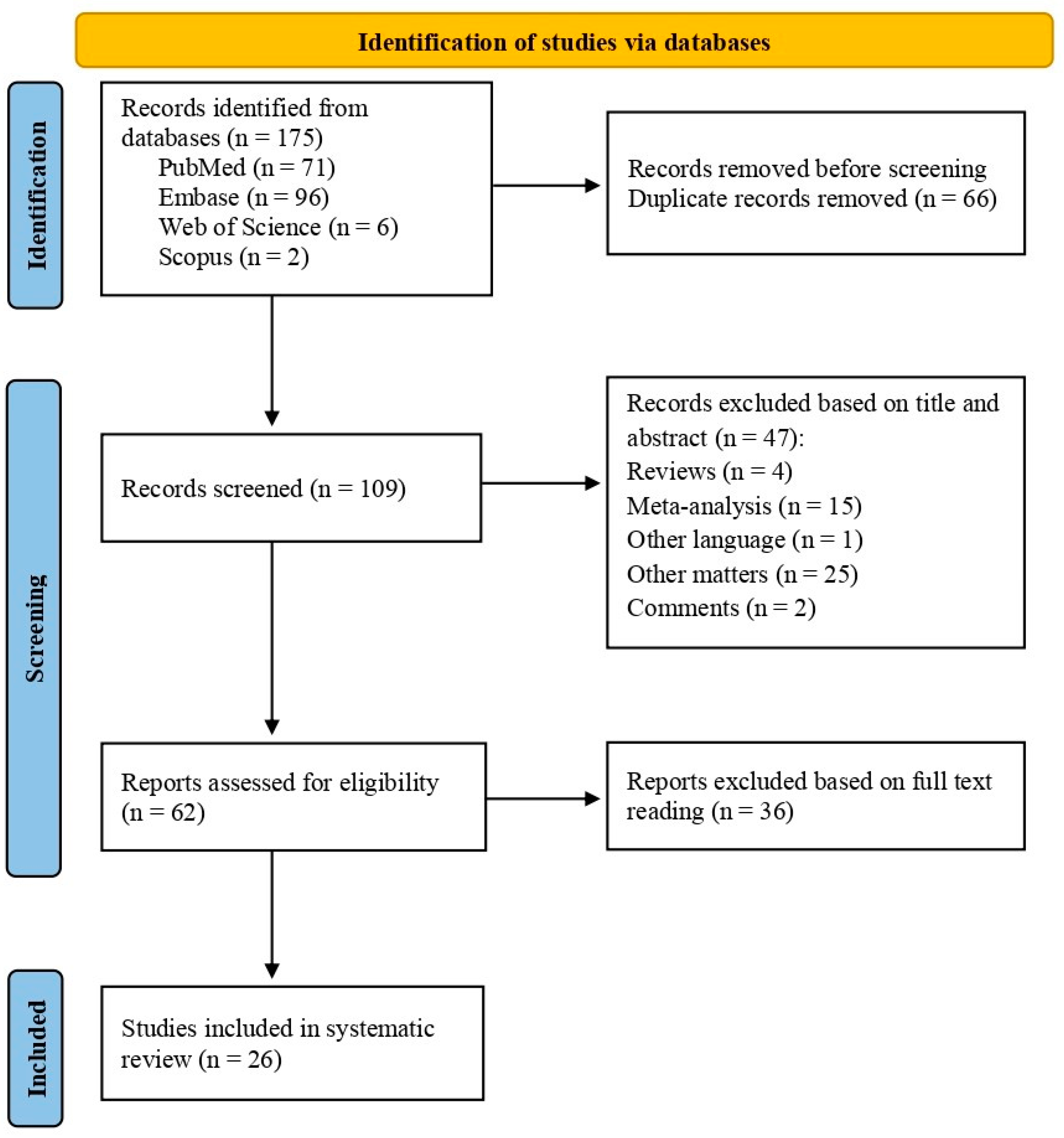

Table 1.
General characterization of included studies.
| STUDY DESIGN | AUTHOR | POPULATION | SNP ID | ALLELE/ GENOTYPE |
GENE ID | miR ID | ASSOCIATION OF mirSNPS WITH CRC |
|---|---|---|---|---|---|---|---|
| Case-control study | Landi et al. | Czech Republic | rs17281995 rs1051690 | G/A | CD86 INSR |
miR-337, miR-582, miR-200a, miR-184, and miR-212 miR-618 and miR-612 |
Increased risk |
| Case-control study | Ding et al. | Chinese | rs141178472 | CC | PIK3CA | miR-520a | Increased risk |
| Case-control study | Ni et al. | Chinese | rs3814058 | C>T | PXR | hsa-miR-129-5p | Increased risk |
| Case-control study | Xie et al. | Chinese | rs6504593 | T | IGF2BP1 | miR-21 | Increased risk; cell proliferation and apoptosis |
| Case-control study | Ke et al. | Chinese | rs1062044 | A | LAMC1 | miR-423-5p | Increased risk; colon adenocarcinomas |
| Case-control study | Kang et al. | Korea | rs2279398 | G>A | DOK3 | miR-370 | Increased risk |
| Case-control study | Lee et al. | Korea | rs7930 | AG | TOMM20 | miR-4273-5p | rs7930 AG: Increased risk; Suppression of TOMM20 gene expression |
| Case-control study | Xicola et al. | Spain | rs868 | TGFBR1 | let-7 | Increased risk | |
| Case-control study | Mullany et al. | United States | rs8176318 rs8905 | AA GG |
BRCA1 PRKAR1A | miR-525-5p miR-miR-214-3p | Increased risk |
| Case-control study | Chayeb et al. | Tunisian | rs2292832 rs2910164 | C/T G/C |
hsa-mir-149 hsa-mir-146a |
Increased risk | |
| Case-control study | Cao et al. | Chinese | rs895819 | GG | miR-27a | Increased risk | |
| STUDY DESIGN | AUTHOR | POPULATION | SNP ID |
ALLELE/ GENOTYPE |
GENE ID | miR ID | ASSOCIATION OF mirSNPS WITH CRC |
| Case-control study | Hu et al. | Chinese | rs2910164 | G>C | miR-146a | Susceptibility and differentiation | |
| Cohort Studies | Kim et al. | Korea | rs3757417 rs12373 | T>G A>C |
TPST1 PAUF | miR-571 | Prognostic markers |
| Case-control study | Salem et al. | Iran | rs12904 | AA/ A>G | EFNA1 | miR-200C and miR-429 | rs12904 AA: Pathogenesis and metástasis; rs12904 A>G: reduced disease progression; regulation of EFNA1 gene expression. |
| Cohort Studies | Smits et al. | Dutch | KRAS-LCS6 | T>G | KRAS | let-7 | Prognostic biomarker |
| Case-control study | Ryan et al. | Caucasian and African-American | rs4919510 | GG | hsa-miR-608 | Increased risk of death in Caucasians and a reduced risk in African Americans | |
| Case-control study | Zhan et al. | Chinese | rs11614913 | CC | miR-196a2 | Increased susceptibility in patients who have undergone chemotherapy treatment | |
| Cohort Studies | Huang et al. | Chinese | rs2273626 rs202195689 | A>C CCCCA>del |
hsa-miR-4707-3p hsa-miR-4274 | Predictive and prognostic biomarkers after chemoradiotherapy | |
| Case-control study | Chen et al. | Chinese | rs2682818 | AA and AC/AA | miR-618 | Reduction of risk | |
| Case-control study | Zhu et al. | Chinese | rs187960998 | C>T | CHD5 | miR-211 | Decreased risk and protective factor for CC by preventing binding of the tumor suppressor gene CHD5 |
| Case-control study | Radanova et al. | Bulgaria | rs2682818 | AC | miR-618 | Decreased risk of susceptibility to mCC | |
| Case-control study | Jiang eta al. | Chinese | rs35301225 | C/A | E2F1 | miR-34a | Tumor suppression and protection by downregulating the E2F1 gene; rs35301225 C/A: upregulation of E2F1 expression, resulting in worse survival. |
| STUDY DESIGN | AUTHOR | POPULATION | SNP ID |
ALLELE/ GENOTYPE |
GENE ID | miR ID | ASSOCIATION OF mirSNPS WITH CRC |
| Randomized study | Sclafani et al. | Multicenter study | rs4919510 | CC | miR-608 | Prognostic biomarker | |
| Case-control study | Chen et al. | Chinese | rs11614913 | T>C | miR-196a2 | No significant association with increased risk or disease progression was identified | |
| Case-control study | Ayadilord et al. | Iran | rs11614913 | C/T | miR-196a2 | No significant association was identified between the SNP rs11614913 in miR-196a2 and CRC | |
| Case-control study | Simonian et al. | Iran | rs12904 | EFNA1 | miR-200c | No significant association was demonstrated between rs12904 and CRC risk |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



