
Submitted:
28 September 2024
Posted:
29 September 2024
You are already at the latest version
Abstract
Snakebite envenoming represents a critical global health challenge, particularly prevalent in regions with limited access to healthcare resources, where venomous snakes pose a significant threat to human populations. Antivenom therapy which mainly rely on antibody production by immunization of large animals with venom components is labour intensive, time consuming and associated with various ethical concerns. Consequently, access to quality and affordable antivenom remains limited in many affected regions with high mortality associated with snakebites. In traditional medicine, many plant species have been ethnobotanically reported for their antivenom properties and are used to neutralize animal toxins. Ribosome-inactivating proteins (RIPs) are a diverse group of toxins found in various organisms, including plants that possess the ability to inhibit protein synthesis by irreversibly damaging the ribosomes. Even though considered to be harmful, the biological role of RIPs has gained increasing attention in recent years due to their potential therapeutic implications. With these insights, this review underscores the potential of RIPs as promising candidates for adjunct treatments in snakebite management strategies. In silico analysis by molecular docking of RIPs with major snake venom proteins resulted in effective binding and shows the interface residues involved in the interaction. This integrative approach enhances our understanding of snakebite pathophysiology, accelerating the development of novel next-generation antivenom therapies that are safer and more effective.
Keywords:
Ribosome inactivating proteins
; major snake venom proteins
; next-generation antivenom
; neutralizing snake venom
; snakebite envenoming
; in silico analysis
; molecular docking
; interface residues
1. Introduction
Certain species in the animal kingdom have evolved with production of toxic compounds in various parts of their body for the purpose of defence, predation or competition [1]. When humans are attacked by these animal species, they inject venoms or poisons that cause various inflammatory reactions leading to short-term and long-term effects on human health [2]. Since these venomous bites are considered as a medical emergency, allopathic treatments are often employed for the recovery. In traditional medicine, many plants were ethnobotanically reported in neutralizing these animal toxins. But, the bioactive compound present in these plants acting against the animal toxins were not completely characterized. By understanding those compounds, we would be able to produce an alternative medicine.
Nature has always blessed human beings with numerous plant species. Plants, being used as food, feed, medicine, fuel and even as poison are subjected to various stresses from the surrounding environment. In order to overcome stress and survive, they undergo several defence mechanisms and one such interesting mechanism adopted is producing toxic bioactive compounds (Figure 1) [3].
These toxic compounds range from low molecular weight phytochemicals to high molecular weight proteins [4] such as ribosome inactivating proteins, plant protease inhibitors, lectins, α-amylase inhibitors, canatoxin like proteins, ureases, arcelin, pore-forming toxins, and antimicrobial peptides undergo specific activities in plants to give resistance against several biotic and abiotic stresses aiming to a persistent life [5]. These phytotoxins when consumed by humans may lead to numerous side effects. However, when the intake is restricted to a certain level these biomolecules have highly significant therapeutic properties in human life [6]. Why not use these toxic proteins to serve as a potential antidote? Here in this review, preliminary discussions were made regarding the role of a phytotoxic protein in neutralizing the animal toxin.
2. An overview of Ribosome Inactivating Proteins (RIPs)
One among the toxic protein family is the Ribosome inactivating proteins (RIPs) that are mostly present in plants and studies have revealed that they are also present in some bacteria, fungi, mushrooms, algae and insects. The discovery of ribosome-inactivating proteins in the nineteenth century occurred when the scientist Stillmark isolated ricin from Ricinus communis and observed its toxic effects. [7]. However, the characterization of these proteins came into the spotlight only from the 1970s [8]. In the group of eminent researchers who contributed to the discovery of RIPs from plants, Professor Fiorenzo Stirpe and his team had detected, isolated and characterized many RIPs including the purification and characterization of immunotoxins from RIPs [9]. He is the pioneer to have documented about the various biological roles of RIPs for human welfare [10]. Based on the previous studies, N-glycosylation in 60S ribosomes is the enzymatic mechanism by which ribosome inactivating proteins remove a particular adenine at position A4324 from the rat 28S rRNA [11]. This adenine is located within the α-sarcin/ricin loop (SRL), a conserved 14 nucleotide sequence found in major rRNAs across diverse organisms, from bacteria to humans. The core of this loop features a GAGA sequence, where the first adenine residue acts as RIP's substrate (Figure 2).
This irreversible modification of the targeted adenine residue inhibits the GTPase activities of elongation factors EF-1 and EF-2 subsequently impeding the process of translation and leading to cell death in eukaryotes [12]. This mechanism of targeted cell death may be employed for the detoxification of animal toxins in the human body.
2.1. Origin and Types of RIPs
In nature, RIP genes were more widely distributed among plant species. Many different types of plants produce numerous isoforms of RIPs, which have evolved through a range of molecular mechanisms such as gene duplication, gene deletion, gene fusion with other genes, and horizontal gene transfer between species [13]. According to the available data, the RIP domain about 30 kDa in size made up of a single polypeptide chain with N-glycosidase activity was formed at least 300 million years ago are currently classified as type I RIPs [14]. Pokeweed antiviral protein (PAP) was the first type I RIP which was isolated from Phytolacca americana (American pokeweed) [15]. Moreover, there exists a single domain protein exhibiting N-glycosidase activity with arginine/glutamate rich polypeptides (AGRPs) at their N-terminal sequences that have a lower molecular weight than that of normal type I RIPs are termed as small type I RIPs. These small type I RIPs are most commonly present in the Cucurbitaceae family [16]. Later on, it is said that a plant managed to combine the RIP domain with a lectin domain duplication led to all current type II RIPs. The A domain with N-glycosidase activity is approximately 30 kDa which is structurally similar to type I RIPs, while the B domain with lectin characteristics is approximately 35 kDa and has the capability to bind specifically to galactosyl terminated structures on the surface of the cells, aiding the entry of A domain into the cell (Figure 3).
For example, ricin and abrin belong to the category of type II RIPs which are highly toxic and not all type II RIPs are toxic. The absence of the B domain in type I RIPs accounts for their comparatively lower toxicity than type II RIPs. Type III RIPs being less prevalent were descended from the resulting ancestor type II RIPs through domain fusion and deletion containing a N-terminal domain similar to the A domain of type I RIPs, fused to an unidentified functional C-terminal domain [14]. Proteins belonging to this type are produced as single domain proenzymes such as barley JIP60 and maize b-32 [16,17]. Plants tightly express these RIP genes only in specific tissues or in response to stress leading to the synthesis of RIPs and are localised extracellularly as well as in subcellular compartments such as vacuoles and endoplasmic reticulum undergoing various post translational modifications allowing the RIPs to function effectively in host cells and also ensure to prevent the self-toxicity of native proteins [12,18,19,20].
2.2. Biological Role of RIPs
Despite having the highly toxic effect of RIPs on pathogens and pests to overcome stress in plants [16], their N-β-glycosylase activity inhibiting translation seems to trigger the pathways activating caspases leading to the induction of apoptosis in mammalian cells [21]. The high toxicity of type II RIPs to humans and animals through ricin poisoning [22], abrin poisoning [23,24] and other toxicity analysis were studied previously [8]. The Type I RIPs being less toxic than Type II RIPs when administered at higher doses may induce toxic symptoms and even cell death in rats [25]. Consequently, many RIPs were isolated and used by criminals in assassination plots or in the event of murder leading to mass destruction [26,27]. Considering this deadly activity, the US government in 1997 classified several toxic biological compounds including some RIPs as category B biological warfare agents [26,28].
Besides being a toxic weapon, RIPs also have many other biological roles in the field of agriculture as anti-plant viral activity [29,30], antifungal activity and insecticidal activity and in the field of medicine as anticancer activity, embryotoxic activity, abortifacient activity, antimicrobial activity, antiviral activity, anti-HIV activity and various other applications in the area of neuroscience research and developing immunotoxins as a therapy against cancer (Figure 4) [10,31]. Here, in this review, the effect of RIPs on snake venom proteins was studied using in silico approaches and their role against snake venom envenoming was discussed.
3. Severity of Snakebite and Need for Effective Antivenom
Snakebite, the most serious Neglected Tropical Disease (NTD) is a potentially life threatening lethal medical emergency that is responsible for enormous suffering and disability in every continent caused by the toxins released by the venomous snakes [32]. According to the World Health Organization (WHO), 1.8 to 2.7 million cases of envenomation occur annually among 5.4 million estimated snakebites worldwide. Every year there are around 81,410 to 1,37,880 deaths due to snakebites and in some cases, they also result in permanent impairments [33]. Being a cold blooded reptile [34] most of the snake species rely on tropical climatic regions to survive and almost half of all snakebite deaths worldwide are said to occur in India, a tropical country where an estimated average of 50,000 to 60,000 deaths are reported each year [35,36]. Mainly rural and peri-urban communities involved in agricultural and other allied activities are highly vulnerable to snakebites [37]. Moreover, venomous snake encounters resulting in the death of cattle, poultry and other livestock causes severe economic loss to farming communities [38]. Among 3,783 species of snakes which are distributed worldwide only around 600 species are most dangerous and highly venomous in nature [39,40]. The venomous species mainly come under the families like Elapidae (401 species), Viperidae (383 species), Atractaspididae (69 species) and some in Colubridae (2105 species) [41,42]. In India, the medically important snakes which causes severe damage to the livelihood are termed as “The Big Four” and they are Indian cobra (Naja naja), Common krait (Bungarus caeruleus), Russell’s viper (Daboia russelii) and Saw-scaled viper (Echis carinatus) [43]. These venomous snakes have a special structure in their mouth region called fangs connected to the venom gland through venom duct. The venom gland which secretes venom is a modified salivary gland surrounded by the compressor muscles which aid in the delivery of the secreted venom fluid while biting. Generally, the evolution of these features was adapted by the snakes in order to capture, immobilise and digest their preys easily or attack their predators [44,45].
3.1. Composition of Snake Venom
Experimental studies in identifying the composition of snake venom revealed that it is a complex mixture of organic and inorganic substances including proteins, carbohydrates, lipids and other metal ions that elicit a wide range of pathophysiological effects in humans [46]. Composition of venom varies based on factors including age, nutrition, climate, temperature and geographic location among various species and also within the members of the same species [47]. Around 90 to 95 percent of the dry weight of snake venom is made up of proteins and peptides, which account for nearly all of the biological effects of the venom [48]. According to the studies based on the occurrence and abundance of snake venom proteins by Theo Tasoulis and Geoffrey K. Isbister, the venom proteins are classified into 59 different protein families and are categorised into five different groups (Table 1).
The deadliest composition of snake venom makes it so effective by causing various physiological changes in the organism leading to death. These toxic changes can range from local tissue damage at the site of envenomation to systemic effects affecting multiple organ systems. The type of mechanism of toxin activity may be:
- Neurotoxin—Flaccid paralysis;
- Myotoxin—Systemic skeletal muscle damage;
- Haematotoxin—Interfere in haemostasis, causing either bleeding or thrombosis;
- Necrotoxin—Death of tissues at the bite site;
- Cardiotoxin—Direct damage to cardiovascular tissues;
- Nephrotoxin—Direct damage to renal tissues.
Among all the snake venom protein families, the dominant protein family group causes significant pathophysiological changes in animals, human and other organisms [51,52].
3.1.1. Phospholipase A2 (PLA2)
Phospholipase A2 is one of the most abundant enzyme components in snake venom which are structurally characterized by a single polypeptide chain that is folded into a tight globular form by disulfide bonds with a conserved catalytic site containing a calcium ion [53]. The main activity of PLA2 is to disrupt the membrane integrity by hydrolysing the phospholipids in the cell membrane. This causes pain, inflammation and localized tissue damage at the envenomation site. As a result, other venom components can easily be disseminated into the bloodstream facilitating quick spread of toxicity [54]. Certain PLA2 have the direct ability to activate the elements of the blood coagulation cascade like prothrombin and factor X, resulting in procoagulant effects leading to systemic haemostatic disruptions and intravascular thrombosis [55]. PLA2 can also cause damage to muscles by inducing calcium influx into the muscle cells resulting in the rupture of sarcolemma membranes leading to pain, weakness and muscle necrosis aiding in the immobilization of the prey [56].
3.1.2. Three-Finger Toxins (3FTx)
Three-finger toxins are typically small and highly stable proteins which are characterized by their unique three-finger fold consisting of loops stabilized by disulfide bonds involved in specific target binding and are commonly present in cobras and kraits [57]. These proteins are primarily neurotoxic interfering with neurotransmission by binding to various types of receptors of the nervous system, such as nicotinic acetylcholine receptors (nAChRs) and muscarinic acetylcholine receptors (mAChRs). By binding to these receptors, they can block nerve impulse leading to paralysis, respiratory failure and ultimately the prey gets immobilized [58]. Some 3FTxs interfere with blood coagulation by inhibiting factors involved in the coagulation cascade leading to systemic haemorrhage and facilitating prey capture and digestion [59].
3.1.3. Snake Venom Metalloproteases (SVMP)
Snake venom metalloproteases are zinc-dependent enzymes characterized by a catalytic domain containing a zinc ion coordinated by histidine residues [60]. SVMPs are commonly present in viperid snakes where these proteins target the extracellular proteins like collagen, elastin and fibronectin leading to local tissue destruction, haemorrhage and inflammation which can even cause organ damage to the envenomed organism [61,62]. SVMPs can have various structural domains like metalloproteinase, disintegrin, cysteine-rich and lectin-like domains which aid in diverse biological activities. SVMPs with disintegrin-like domains can attach to integrin receptors on platelet surfaces limiting platelet aggregation which contributes to coagulopathic effects and produces antiplatelet activity [61].
3.1.4. Snake Venom Serine Proteases (SVSP)
Snake venom serine proteases are enzymes that belong to the serine protease family typically consisting of one or more catalytic domains containing the conserved catalytic triad (histidine, aspartate and serine). These proteins often contain additional domains such as exosites or disulfide-rich loops that contribute to substrate specificity and inhibitor binding [63]. They modulate haemostasis either by activating or inhibiting the components of the blood clotting cascade mechanism ultimately resulting in blood coagulation and fibrinolysis [64]. Some SVSPs cause intravascular thrombosis and consumption coagulopathy by activating the clotting factors such as factor X and prothrombin. While others inhibit platelet aggregation or fibrin formation, resulting in bleeding disorders [65]. SVSPs also play a crucial role in degrading the components of the extracellular matrix and disrupt cell to cell adhesion facilitating the easy digestion of prey [63].
3.2. Production of Antivenom for Curing Snakebites
In India, antivenom for snakebite commercially produced as Anti Snake Venom (ASV) is a life-saving medication used to treat patients with snakebite envenomation. ASVs are mostly polyvalent antibodies that try to neutralize the toxic effects of any venomous snakebite, therefore preventing serious medical complications and death [66]. The production of ASV involves a serotherapy process of immunizing the animals mainly equines and ovines by injecting the mixture of venom collected from venomous snakes. Through repeated exposure to venom, these animals develop prominent IgG antibodies in their blood, which are harvested and purified to create ASVs. The purified antibodies are then tested to ensure their quality, efficiency and safety for human use [67]. Once a person is bitten by a venomous snake, upon severity of the symptoms the ASV is continuously administered intravenously with complete medical surveillance. The antibodies in ASV bind to the venom toxins in the bloodstream and neutralize their effects by reducing symptoms such as tissue damage, bleeding, paralysis and organ failure [68]. However, antivenom does not reverse damage that has already occurred but can stop the progression of envenomation. Despites its life saving capacity, ASVs may also cause some allergic reactions ranging from local itching to severe anaphylaxis [69]. The major challenge in the production of ASVs is the variability in venom composition between the species which is highly specific [70,71]. ASVs may not be readily available in the regions of envenomation, even the limited availability has been reported previously in India. Currently there are eight authorized manufacturers of polyvalent antibodies in India raised against the big four venomous snake species with a total installed capacity of 6.75 million vials per annum. However, only 1.5–2 million vials are produced in India every year [72]. Considering these constraints, various other studies with designing unique aptamers, employing phage display techniques, using small molecule inhibitors, isolating natural phytochemicals in neutralizing the snake venom were undergone previously [73]. But when numerous clinical trials report the failure of ASVs even against the Indian geographical snakes due to the complex variable nature of venom proteins, it is an alarming threat to each and every citizen of India [74]. Hence the need for next generation antivenom is the spotlight area for researchers to develop a sustainable system to treat snakebite envenoming.
4. Plants with Antivenom Properties in Traditional Medicine
The utilization of plants in the Indian traditional healing medicine systems such as Ayurveda, Homoeopathy, Naturopathy, Unani and Siddha has been integral to human health and wellbeing for centuries. Medicinal plants have various therapeutic values highlighting their cultural significance along with contributions to modern health care [75]. Since ancient periods, plants have been used for the treatment of snakebites traditionally by many people in remote areas all over the world. Phytochemical extracts of various parts of the plants exhibit diverse mechanisms of action against snake venom toxins [76,77,78,79]. Plant families like Fabaceae, Asteraceae, Zingiberaceae, Salicaceae, Amaranthaceae, Mimosaceae, Euphorbiaceae, Cucurbitaceae, Piperaceae, Musaceae, Rutaceae, Liliaceae, Malvaceae and many other families were previously studied for the presence of bioactive substances with neutralizing properties of snake venom [80,81,82,83,84,85,86,87]. Generally, the secondary metabolites such as phenolics, terpenoids, alkaloids and saponins present in different plants under these families were reported as natural inhibitors of snake venom proteins [88,89]. The list of these compounds studied for antivenom properties were clearly mentioned in the review written by Adrião et al. in 2022. Some modified glycosides like homaloside D from Homalium ceylanicum, itoside from Itoa orientalis, scoloposide from Schisandra chinensis and salireposide from Symplocos racemosa had been isolated and studied for the inhibition of venom compounds. Even proteins like Withania somnifera glycoprotein (WSG) and turmerin from Curcuma longa also showed inhibition activity of phospholipase A2 proteins in snake venom [90]. In the view of these research insights and preclinical studies, plant products offer a vast opportunity to delve in the area of snakebite envenomation. While screening the literature, there were many plants which are ethnopharmacologically reported with antivenom properties have yet to be thoroughly investigated to identify the specific compound responsible for this effect [91]. Further, by employing in silico analysis using molecular docking and simulation to identify the potential bioactive compounds that bind to the snake venom proteins effectively with better interactions could help in designing modified natural products with high capacity to neutralize the toxicity caused by snake venom in the human body [73,92].
5. In Silico Analysis of Plant Derived RIPs against Snake Venom Proteins
For the in silico study, the major proteins from highly venomous “The Big Four” snake species namely Indian cobra, Indian common krait, Russell’s viper and Saw-scaled viper were selected as the target (Figure 5).
The plants that have antivenom property which have also reported for the presence of RIPs were chosen as the ligand protein for protein-protein interaction studies using molecular docking (Table 2). Since many of the plants with antivenom property are not characterized with the particular compound responsible for the effect, choosing plant derived RIPs as a ligand protein could enable us to hypothesize the antivenom activity of RIPs against major snake venom proteins.
The available 3D structures of the snake venom proteins and the plant derived RIPs were retrieved from protein data bank (PDB) [93]. The target proteins (Naja naja phospholipase A2, Alpha-delta bungarotoxin, Echistatin, Russell's viper venom serine protease) were prepared for the docking analysis using the BIOVIA Discovery Studio (DS4.5, Accelrys, Inc., San Diego, CA, USA). The energy minimization of the proteins was done using CHARMm force field to ensure the structural stability and all the other parameters were set to their default values, allowing for accurate and efficient docking. The ligand proteins (Abrin, APA-1, amaranthin, luffaculin, momordin, β-momorcharin, Nicotiana tabacum RIP, ebulin, Viscum articulatum RIP) were virtually analysed for toxicity prediction using ToxinPred 2.0 web server [94]. Since all the ligand proteins were RIPs, the prediction of the server resulted as toxic compounds. The molecular docking was performed in Accelrys Discovery Studio by ZDOCK protocol with angular step size as six and all other parameters were set to default. The docking of four target proteins individually with all nine ligand proteins resulted in 2000 docked poses each. Then, the Process DOCK protocol was performed only for the top poses in the largest clusters. From the results of Process DOCK, refine docking (RDOCK) is performed to get the best interaction. After processing the poses, the best pose with high ZDOCK score and ZRANK score among the protein-protein interactions was chosen for analysing the protein interface to find the interface residues involved in the interaction (Table 3 and Table 4). The step-by-step process of analysis is shown in the Figure 6. The three-dimensional representation is shown in the Figure 7 to visualize the interaction between the proteins.
6. Future Prospects
The RIPs with high binding affinities to snake venom proteins could be involved as an innovative approach in building a successive candidate biomolecule for the inhibition of snake venom components. Not only performing in silico studies is enough to prove that RIPs are the effective candidate molecules but also in vitro and in vivo assays should be conducted for the standard results. Alternative binding scaffolds like nanobodies, affimers, adnectins, affibodies, affitins, anticalins, avimers, armadillo repeat proteins, β-hairpin mimetics, bicyclic peptides, designed ankyrin repeat proteins (DARPins) and fynomers are already underway in the progression of development of promising therapeutic modalities leading to an effective next-generation envenoming therapies [101]. Entailing RIPs along with these scaffolds or any other effective ligand with advanced technologies could be a revolutionary breakthrough for snakebite treatment. Research works in designing RIP based immunotoxins for the targeted cancer therapy is an upcoming technology where the RIPs with N-glycosylation domain inducing apoptotic pathway are bound with the antibodies targeting the oncogenic cells leading to the death of cancer cells [102,103,104]. Such methodology can be utilized in designing immunotoxins that are specific to snake venom protein complexes for the treatment of snakebite envenomation [105]. However, the non-specific broad spectrum cytotoxic effect of RIPs is a challenging part to overcome the untargeted lethal effect on healthy cells. In order to overcome the unintended effects of RIPs, protein engineering is a prominent biotechnological approach to create an effective pharmacokinetic agent against snake venom [106]. And also, the native immunogenicity of the host organism may mislead the immune response towards the RIP candidates developing resistance to its effect [104]. Moreover, RIPs which are macromolecules face barriers in systemic delivery into the cells that can be sorted out by the effective encapsulation with nanoparticle-based carriers and various other ligand based delivery systems [107,108].
7. Conclusions
Time is a critical factor in managing snakebite victims due to the swift onset of pathophysiological effects. Biomolecules offer not only an alternative therapeutic avenue but also serve as an auxiliary approach to minimize preclinical effects before antivenom administration. The intricate and diverse composition of snake venoms poses a significant challenge, making it impractical for a single molecule to neutralize all venom proteins. In response to this complexity, a multifaceted strategy is proposed, encompassing the integration of ethnobotanical insights from traditional medicine.
This multifaceted strategy involves leveraging traditional medicinal knowledge to uncover novel natural products, such as Ribosome-Inactivating Proteins (RIPs). The integration of these compounds with conventional pharmaceuticals holds promise for developing a comprehensive treatment approach that addresses the varied effects of envenomation. By exploring potential synergies between traditional remedies and modern pharmacology, it is possible to identify compounds that not only neutralize venom components but also provide a more tailored and effective response to the intricate biochemistry of snake venoms. This combined strategy, with its emphasis on diverse therapeutic treatments, presents a promising path forward in the quest to improve outcomes for snakebite victims.
Author Contributions
PS did the molecular docking and tabulation of results. BN guided the bioinformatics data acquisition. RP, BR and SVP reviewed the work and suggested improvements in the results and subsequently the manuscript. BR conceived the idea and mentored the study and guided the writing of the manuscript.
Funding and Acknowledgments
PS thanks the MEDHA fellowship offered by Bayer foundation (Fellowship ID: MAST-2023-11160). PS and BR thanks the Department of Plant Molecular Biology and Bioinformatics, TNAU, Coimbatore and BTIS network for allowing to utilize the Discovery Studio and other softwares.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Mackessy, S.P. Venom production and secretion in reptiles. Journal of Experimental Biology 2022, 225, jeb227348. [Google Scholar] [CrossRef]
- Warrell, D.A. Venomous animals. Medicine 2016, 44, 120–124. [Google Scholar] [CrossRef]
- Maag, D.; Erb, M.; Köllner, T.G.; Gershenzon, J. Defensive weapons and defense signals in plants: some metabolites serve both roles. BioEssays 2015, 37, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Mithöfer, A.; Boland, W. Plant defense against herbivores: chemical aspects. Annual review of plant biology 2012, 63, 431–450. [Google Scholar] [CrossRef] [PubMed]
- Kocyigit, E.; Kocaadam-Bozkurt, B.; Bozkurt, O.; Ağagündüz, D.; Capasso, R. Plant Toxic Proteins: Their Biological Activities, Mechanism of Action and Removal Strategies. Toxins 2023, 15, 356. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Singh, H.; Bhardwaj, N.; Bhardwaj, S.K.; Khatri, M.; Kim, K.-H.; Peng, W. An exploration on the toxicity mechanisms of phytotoxins and their potential utilities. Critical Reviews in Environmental Science and Technology 2022, 52, 395–435. [Google Scholar] [CrossRef]
- Stirpe, F.; Gilabert-Oriol, R. Ribosome-inactivating proteins: An overview. Plant Toxins; Carlini, CR, Ligabue-Braun, R., Gopalakrishnakone, P., Eds 2015, 153-182.
- Bolognesi, A.; Bortolotti, M.; Maiello, S.; Battelli, M.G.; Polito, L. Ribosome-inactivating proteins from plants: A historical overview. Molecules 2016, 21, 1627. [Google Scholar] [CrossRef] [PubMed]
- Bolognesi, A.; Bortolotti, M.; Battelli, M.G.; Polito, L. Hyperuricaemia, xanthine oxidoreductase and ribosome-inactivating proteins from plants: the contributions of Fiorenzo Stirpe to frontline research. Molecules 2017, 22, 206. [Google Scholar] [CrossRef]
- Stirpe, F. Ribosome-inactivating proteins: From toxins to useful proteins. Toxicon 2013, 67, 12–16. [Google Scholar] [CrossRef]
- Barbieri, L.; Valbonesi, P.; Righi, F.; Zuccheri, G.; Monti, F.; Gorini, P.; Samori, B.; Stirpe, F. Polynucleotide: adenosine glycosidase is the sole activity of ribosome-inactivating proteins on DNA. The journal of biochemistry 2000, 128, 883–889. [Google Scholar] [CrossRef]
- Nielsen, K.; Boston, R.S. Ribosome-inactivating proteins: a plant perspective. Annual review of plant biology 2001, 52, 785–816. [Google Scholar] [CrossRef] [PubMed]
- Lapadula, W.J.; Ayub, M.J. Ribosome Inactivating Proteins from an evolutionary perspective. Toxicon 2017, 136, 6–14. [Google Scholar] [CrossRef] [PubMed]
- Peumans, W.J.; Van Damme, E.J. Evolution of plant ribosome-inactivating proteins. Toxic plant proteins 2010, 1–26. [Google Scholar]
- Dallal, J.A.; Irvin, J.D. Enzymatic inactivation of eukaryotic ribosomes by the pokeweed antiviral protein. FEBS letters 1978, 89, 257–259. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.; Zhou, Y.-K.; Ji, Z.-L.; Chen, X.-R. The plant ribosome-inactivating proteins play important roles in defense against pathogens and insect pest attacks. Frontiers in Plant Science 2018, 9, 146. [Google Scholar] [CrossRef]
- Sharma, A.; Gupta, S.; Sharma, N.R.; Paul, K. Expanding role of ribosome-inactivating proteins: From toxins to therapeutics. IUBMB life 2023, 75, 82–96. [Google Scholar] [CrossRef]
- Frigerio, L.; Vitale, A.; Lord, J.M.; Ceriotti, A.; Roberts, L.M. Free ricin A chain, proricin, and native toxin have different cellular fates when expressed in tobacco protoplasts. Journal of Biological Chemistry 1998, 273, 14194–14199. [Google Scholar] [CrossRef]
- Zhu, R.H.; Ng, T.B.; Yeung, H.W.; Shaw, P.C. High level synthesis of biologically active recombinant trichosanthin in Escherichia coli. International Journal of Peptide and Protein Research 1992, 39, 77–81. [Google Scholar] [CrossRef]
- Carzaniga, R.; Sinclair, L.; Fordham-Skelton, A.P.; Harris, N.; Croy, R.R. Cellular and subcellular distribution of saporins, type-1 ribosome-inactivating proteins, in soapwort (Saponaria officinalis L.). Planta 1994, 194, 461–470. [Google Scholar] [CrossRef]
- Das, M.K.; Sharma, R.S.; Mishra, V. Induction of apoptosis by ribosome inactivating proteins: importance of N-glycosidase activity. Applied biochemistry and biotechnology 2012, 166, 1552–1561. [Google Scholar] [CrossRef]
- Moshiri, M.; Hamid, F.; Etemad, L. Ricin toxicity: Clinical and molecular aspects. Reports of biochemistry & molecular biology 2016, 4, 60. [Google Scholar]
- Jang, D.H.; Hoffman, R.S.; Nelson, L.S. Attempted suicide, by mail order: Abrus precatorius. Journal of medical toxicology 2010, 6, 427–430. [Google Scholar] [CrossRef] [PubMed]
- Dickers, K.J.; Bradberry, S.M.; Rice, P.; Griffiths, G.D.; Vale, J.A. Abrin poisoning. Toxicological reviews 2003, 22, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Meng, Y.; Liu, B.; Lei, N.; Zheng, J.; He, Q.; Li, D.; Zhao, X.; Shen, F. Alpha-momorcharin possessing high immunogenicity, immunotoxicity and hepatotoxicity in SD rats. Journal of ethnopharmacology 2012, 139, 590–598. [Google Scholar] [CrossRef] [PubMed]
- Walsh, M.J.; Dodd, J.E.; Hautbergue, G.M. Ribosome-inactivating proteins: Potent poisons and molecular tools. Virulence 2013, 4, 774–784. [Google Scholar] [CrossRef]
- Schep, L.J.; Temple, W.A.; Butt, G.A.; Beasley, M.D. Ricin as a weapon of mass terror—Separating fact from fiction. Environment international 2009, 35, 1267–1271. [Google Scholar] [CrossRef] [PubMed]
- Patočka, J.; Středa, L. Plant toxic proteins and their current significance for warfare and medicine. J Appl Biomed 2003, 1, 141–147. [Google Scholar] [CrossRef]
- Divakar, M.N.; Rajesh, S.; Renukadevi, P.; Rajagopal, B. Cloning, Expression and in silico Characterization of a Truncated Antiviral Protein Gene from Bougainvillea spectabilis Willd. Int. J. Curr. Microbiol. App. Sci 2019, 8, 2828–2836. [Google Scholar] [CrossRef]
- Biology, C.f.P.M. A cDNA (BAPc DNA L1) from Bougainvillea spectabilis willd leaves encoding a protein with antiviral & ribosome inactivating activity and a method for obtaining the same. 24 8606, 2006. [Google Scholar]
- Akkouh, O.; Ng, T.B.; Cheung, R.C.F.; Wong, J.H.; Pan, W.; Ng, C.C.W.; Sha, O.; Shaw, P.C.; Chan, W.Y. Biological activities of ribosome-inactivating proteins and their possible applications as antimicrobial, anticancer, and anti-pest agents and in neuroscience research. Applied microbiology and biotechnology 2015, 99, 9847–9863. [Google Scholar] [CrossRef]
- Organization, W.H. Snakebite envenoming: a strategy for prevention and control. 2019.
- Organization, W.H. Snakebite Envenoming. Available online: https://www.who.int/news-room/fact-sheets/detail/snakebite-envenoming (accessed on 01 February 2024).
- Yu, X.; Wu, N.C.; Ge, L.; Li, L.; Zhang, Z.; Lei, J. Artificial shelters provide suitable thermal habitat for a cold-blooded animal. Scientific Reports 2022, 12, 5879. [Google Scholar] [CrossRef] [PubMed]
- Suraweera, W.; Warrell, D.; Whitaker, R.; Menon, G.; Rodrigues, R.; Fu, S.H.; Begum, R.; Sati, P.; Piyasena, K.; Bhatia, M. Trends in snakebite deaths in India from 2000 to 2019 in a nationally representative mortality study. Elife 2020, 9, e54076. [Google Scholar] [CrossRef] [PubMed]
- Collaborators, G.S.E. Global mortality of snakebite envenoming between 1990 and 2019. Nature Communications 2022, 13, 6160. [Google Scholar]
- Ralph, R.; Faiz, M.A.; Sharma, S.K.; Ribeiro, I.; Chappuis, F. Managing snakebite. BMJ 2022, 376. [Google Scholar] [CrossRef] [PubMed]
- Bolon, I.; Finat, M.; Herrera, M.; Nickerson, A.; Grace, D.; Schütte, S.; Martins, S.B.; de Castañeda, R.R. Snakebite in domestic animals: First global scoping review. Preventive Veterinary Medicine 2019, 170, 104729. [Google Scholar] [CrossRef] [PubMed]
- Wallach, V.; Williams, K.L.; Boundy, J. SnakeS of theWorld. A Catalogue of Living and Extinct Species. Boca Raton: CRC Press-Taylor & Francis Group 2014.
- Tednes, M.; Slesinger, T.L. Evaluation and treatment of snake envenomations. In StatPearls [Internet]; StatPearls Publishing: 2022.
- Goswami, P.K.; Samant, M.; Srivastava, R.S. Snake venom, anti-snake venom & potential of snake venom. International Journal of Pharmacy and Pharmaceutical Sciences 2014, 6, 4–7. [Google Scholar]
- Uetz, P. The Reptile Database. 2023.
- Simpson, I.D.; Norris, R.L. Snakes of medical importance in India: is the concept of the “Big 4” still relevant and useful? Wilderness & environmental medicine 2007, 18, 2–9. [Google Scholar]
- Kerkkamp, H.M.; Casewell, N.R.; Vonk, F.J. Evolution of the snake venom delivery system. Evolution of venomous animals and their toxins 2015, 1–11. [Google Scholar]
- Barlow, A.; Pook, C.E.; Harrison, R.A.; Wüster, W. Coevolution of diet and prey-specific venom activity supports the role of selection in snake venom evolution. Proceedings of the Royal Society B: Biological Sciences 2009, 276, 2443–2449. [Google Scholar] [CrossRef]
- Ojeda, P.G.; Ramírez, D.; Alzate-Morales, J.; Caballero, J.; Kaas, Q.; González, W. Computational studies of snake venom toxins. Toxins 2017, 10, 8. [Google Scholar] [CrossRef]
- Santhosh, M.S.; Hemshekhar, M.; Sunitha, K.; Thushara, R.; Jnaneshwari, S.; Kemparaju, K.; Girish, K. Snake venom induced local toxicities: plant secondary metabolites as an auxiliary therapy. Mini Rev Med Chem 2013, 13, 106–123. [Google Scholar] [CrossRef] [PubMed]
- Munawar, A.; Ali, S.A.; Akrem, A.; Betzel, C. Snake venom peptides: Tools of biodiscovery. Toxins 2018, 10, 474. [Google Scholar] [CrossRef] [PubMed]
- Tasoulis, T.; Isbister, G.K. A current perspective on snake venom composition and constituent protein families. Archives of Toxicology 2023, 97, 133–153. [Google Scholar] [CrossRef] [PubMed]
- Tasoulis, T.; Isbister, G.K. A review and database of snake venom proteomes. Toxins 2017, 9, 290. [Google Scholar] [CrossRef]
- Gutiérrez, J.M.; Calvete, J.J.; Habib, A.G.; Harrison, R.A.; Williams, D.J.; Warrell, D.A. Snakebite envenoming. Nature reviews Disease primers 2017, 3, 1–21. [Google Scholar] [CrossRef]
- Toxinology Department, W.s.C.s.H.W. Clinical Toxinology Resources Website. Available online: http://www.toxinology.com/ (accessed on 07 February 2024).
- Marcussi, S.; Sant'Ana, C.D.; Oliveira, C.Z.; Quintero Rueda, A.; Menaldo, D.L.; Beleboni, R.O.; Stabeli, R.G.; Giglio, J.R.; M Fontes, M.R.; Soares, A.M. Snake venom phospholipase A2 inhibitors: medicinal chemistry and therapeutic potential. Current topics in medicinal chemistry 2007, 7, 743–756. [Google Scholar] [CrossRef]
- Doley, R.; Zhou, X.; Kini, R.M. Snake venom phospholipase A2 enzymes. Handbook of venoms and toxins of reptiles 2010, 1, 173–205. [Google Scholar]
- Saikia, D.; Mukherjee, A.K. Anticoagulant and membrane damaging properties of snake venom phospholipase A 2 enzymes. Snake Venoms 2017, 87–104. [Google Scholar]
- Teixeira, C.d.F.P.; Landucci, E.; Antunes, E.; Chacur, M.; Cury, Y. Inflammatory effects of snake venom myotoxic phospholipases A2. Toxicon 2003, 42, 947–962. [Google Scholar] [CrossRef]
- Utkin, Y.N. Three-finger toxins, a deadly weapon of elapid venom–milestones of discovery. Toxicon 2013, 62, 50–55. [Google Scholar] [CrossRef]
- Nirthanan, S. Snake three-finger α-neurotoxins and nicotinic acetylcholine receptors: Molecules, mechanisms and medicine. Biochemical Pharmacology 2020, 181, 114168. [Google Scholar] [CrossRef]
- Das, D. Studies on the crude venom and purified three finger toxin of Naja kaouthia from north east India. 2015.
- Olaoba, O.T.; Dos Santos, P.K.; Selistre-de-Araujo, H.S.; de Souza, D.H.F. Snake venom metalloproteinases (SVMPs): a structure-function update. Toxicon: X 2020, 7, 100052. [Google Scholar] [CrossRef] [PubMed]
- Sonavane, M.; Almeida, J.R.; Rajan, E.; Williams, H.F.; Townsend, F.; Cornish, E.; Mitchell, R.D.; Patel, K.; Vaiyapuri, S. Intramuscular bleeding and formation of microthrombi during skeletal muscle damage caused by a snake venom metalloprotease and a cardiotoxin. Toxins 2023, 15, 530. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, C.d.F.P.; Fernandes, C.M.; Zuliani, J.P.; Zamuner, S.F. Inflammatory effects of snake venom metalloproteinases. Memórias do Instituto Oswaldo Cruz 2005, 100, 181–184. [Google Scholar] [CrossRef]
- Matsui, T.; Fujimura, Y.; Titani, K. Snake venom proteases affecting hemostasis and thrombosis. Biochimica et Biophysica Acta (BBA)-Protein Structure and Molecular Enzymology 2000, 1477, 146–156. [Google Scholar] [CrossRef]
- Kini, R.M. Serine proteases affecting blood coagulation and fibrinolysis from snake venoms. Pathophysiology of haemostasis and thrombosis 2006, 34, 200–204. [Google Scholar] [CrossRef]
- Latinović, Z.; Leonardi, A.; Koh, C.Y.; Kini, R.M.; Trampuš Bakija, A.; Pungerčar, J.; Križaj, I. The procoagulant snake venom serine protease potentially having a dual, blood coagulation factor V and X-Activating activity. Toxins 2020, 12, 358. [Google Scholar] [CrossRef]
- Dias da Silva, W.; De Andrade, S.A.; Megale Â, A.A.; De Souza, D.A.; Sant'Anna, O.A.; Magnoli, F.C.; Guidolin, F.R.; Godoi, K.S.; Saladini, L.Y.; Spencer, P.J.; et al. Antibodies as Snakebite Antivenoms: Past and Future. Toxins (Basel) 2022, 14. [Google Scholar] [CrossRef]
- Ratanabanangkoon, K. Polyvalent Snake Antivenoms: Production Strategy and Their Therapeutic Benefits. Toxins 2023, 15, 517. [Google Scholar] [CrossRef]
- Tan, C.H. Snake Venomics: Fundamentals, recent Updates, and a look to the next decade. Toxins 2022, 14, 247. [Google Scholar] [CrossRef]
- Alangode, A.; Rajan, K.; Nair, B.G. Snake antivenom: Challenges and alternate approaches. Biochemical Pharmacology 2020, 181, 114135. [Google Scholar] [CrossRef] [PubMed]
- Casewell, N.R.; Jackson, T.N.; Laustsen, A.H.; Sunagar, K. Causes and consequences of snake venom variation. Trends in pharmacological sciences 2020, 41, 570–581. [Google Scholar] [CrossRef] [PubMed]
- Modahl, C.M.; Brahma, R.K.; Koh, C.Y.; Shioi, N.; Kini, R.M. Omics technologies for profiling toxin diversity and evolution in snake venom: impacts on the discovery of therapeutic and diagnostic agents. Annual Review of Animal Biosciences 2020, 8, 91–116. [Google Scholar] [CrossRef] [PubMed]
- Gajbhiye, R.K.; Munshi, H.; Bawaskar, H.S. National programme for prevention & control of snakebite in India: Key challenges & recommendations. Indian J Med Res 2023, 157, 271–275. [Google Scholar] [CrossRef] [PubMed]
- Vanuopadath, M.; Rajan, K.; Alangode, A.; Nair, S.S.; Nair, B.G. The need for next-generation antivenom for snakebite envenomation in india. Toxins 2023, 15, 510. [Google Scholar] [CrossRef]
- Mukherjee, A.K.; Mackessy, S.P. Prevention and improvement of clinical management of snakebite in Southern Asian countries: a proposed road map. Toxicon 2021, 200, 140–152. [Google Scholar] [CrossRef]
- Poddar, S.; Sarkar, T.; Choudhury, S.; Chatterjee, S.; Ghosh, P. Indian traditional medicinal plants: A concise review. International Journal of Botany Studies 2020, 5, 174–190. [Google Scholar]
- MS, V.; More, V.S.; Zameer, F.; Muddapur, U.; More, S.S. Ethnomedicinal plants and isolated compounds against Snake venom activity: A review. 2021.
- Magowska, A. The natural history of the concept of antidote. Toxicology reports 2021, 8, 1305–1309. [Google Scholar] [CrossRef] [PubMed]
- Deshpande, A.M.; Sastry, K.V.; Bhise, S.B. A contemporary exploration of traditional Indian snake envenomation therapies. Tropical Medicine and Infectious Disease 2022, 7, 108. [Google Scholar] [CrossRef]
- Premendran, S.J.; Salwe, K.J.; Pathak, S.; Brahmane, R.; Manimekalai, K. Anti-cobra venom activity of plant Andrographis paniculata and its comparison with polyvalent anti-snake venom. J Nat Sci Biol Med 2011, 2, 198–204. [Google Scholar] [CrossRef]
- Singh, S.; Saxena, N. Traditional healing practices for treatment of animal bites among tribes of India: A systematic review. Indian Journal of Traditional Knowledge (IJTK) 2023, 22, 638–645. [Google Scholar]
- Bala, A.A.; Mohammed, M.; Umar, S.; Ungogo, M.A.; Hassan, M.A.-K.; Abdussalam, U.S.; Ahmad, M.H.; Ishaq, D.U.; Mana, D.; Sha'aban, A. Pre-clinical efficacy of African medicinal plants used in the treatment of snakebite envenoming: A systematic review. Toxicon 2023, 107035. [Google Scholar] [CrossRef] [PubMed]
- Mogha, N.G.; Kalokora, O.J.; Amir, H.M.; Kacholi, D.S. Ethnomedicinal plants used for treatment of snakebites in Tanzania–a systematic review. Pharmaceutical Biology 2022, 60, 1925–1934. [Google Scholar] [CrossRef] [PubMed]
- Liaqat, A.; Mallhi, T.H.; Khan, Y.H.; Khokhar, A.; Chaman, S.; Ali, M. Anti-snake venom properties of medicinal plants: a comprehensive systematic review of literature. Brazilian Journal of Pharmaceutical Sciences 2022, 58. [Google Scholar] [CrossRef]
- Upasani, S.V.; Upasani, M.S. Plants from northeast India utilize in snakebite treatment–an ethanobotanical review. 2021.
- Gbolade, A.A. Nigerian medicinal plants with anti-snake venom activity—A review. J Malar Res phytomedicine 2021, 4, 29–44. [Google Scholar]
- Omara, T.; Kagoya, S.; Openy, A.; Omute, T.; Ssebulime, S.; Kiplagat, K.M.; Bongomin, O. Antivenin plants used for treatment of snakebites in Uganda: ethnobotanical reports and pharmacological evidences. Tropical Medicine and Health 2020, 48, 1–16. [Google Scholar] [CrossRef]
- Dey, A.; De, J.N. Traditional use of plants against snakebite in Indian subcontinent: a review of the recent literature. Afr J Tradit Complement Altern Med 2012, 9, 153–174. [Google Scholar] [CrossRef]
- Gómez-Betancur, I.; Gogineni, V.; Salazar-Ospina, A.; León, F. Perspective on the Therapeutics of Anti-Snake Venom. Molecules 2019, 24. [Google Scholar] [CrossRef]
- Singh, P.; Yasir, M.; Hazarika, R.; Sugunan, S.; Shrivastava, R. A Review on Venom Enzymes Neutralizing Ability of Secondary Metabolites from Medicinal Plants. J Pharmacopuncture 2017, 20, 173–178. [Google Scholar] [CrossRef]
- Adrião, A.A.; Dos Santos, A.O.; de Lima, E.J.; Maciel, J.B.; Paz, W.H.; da Silva, F.; Pucca, M.B.; Moura-da-Silva, A.M.; Monteiro, W.M.; Sartim, M.A. Plant-derived toxin inhibitors as potential candidates to complement antivenom treatment in snakebite envenomations. Frontiers in Immunology 2022, 13, 842576. [Google Scholar] [CrossRef]
- Sofyantoro, F.; Yudha, D.S.; Lischer, K.; Nuringtyas, T.R.; Putri, W.A.; Kusuma, W.A.; Purwestri, Y.A.; Swasono, R.T. Bibliometric analysis of literature in Snake venom-related research worldwide (1933–2022). Animals 2022, 12, 2058. [Google Scholar] [CrossRef] [PubMed]
- Aruwa, C.E.; Mukaila, Y.O.; Ajao, A.A.-n.; Sabiu, S. An appraisal of antidotes’ effectiveness: Evidence of the use of phyto-antidotes and biotechnological advancements. Molecules 2020, 25, 1516. [Google Scholar] [CrossRef] [PubMed]
- H. M. Berman, J.W., Z. Feng, G. Gilliland, T.N. Bhat, H. Weissig, I.N. Shindyalov, P.E. Bourne. The Protein Data Bank. Nucleic Acids Research 2000, 28, 235–242. [Google Scholar] [CrossRef]
- Sharma, N.; Naorem, L.D.; Jain, S.; Raghava, G.P.S. ToxinPred2: an improved method for predicting toxicity of proteins. Briefings in Bioinformatics 2022, 23. [Google Scholar] [CrossRef] [PubMed]
- Polito, L.; Bortolotti, M.; Maiello, S.; Battelli, M.G.; Bolognesi, A. Plants Producing Ribosome-Inactivating Proteins in Traditional Medicine. Molecules 2016, 21. [Google Scholar] [CrossRef]
- Schrot, J.; Weng, A.; Melzig, M.F. Ribosome-inactivating and related proteins. Toxins 2015, 7, 1556–1615. [Google Scholar] [CrossRef]
- Gupta, Y.K.; Peshin, S.S. Do herbal medicines have potential for managing snake bite envenomation? Toxicol Int 2012, 19, 89–99. [Google Scholar] [CrossRef]
- Shendge, P.N.; Belemkar, S. Therapeutic Potential of Luffa acutangula: A Review on Its Traditional Uses, Phytochemistry, Pharmacology and Toxicological Aspects. Front Pharmacol 2018, 9, 1177. [Google Scholar] [CrossRef]
- Félix-Silva, J.; Silva-Junior, A.A.; Zucolotto, S.M.; Fernandes-Pedrosa, M.F. Medicinal Plants for the Treatment of Local Tissue Damage Induced by Snake Venoms: An Overview from Traditional Use to Pharmacological Evidence. Evid Based Complement Alternat Med 2017, 2017, 5748256. [Google Scholar] [CrossRef]
- Jabbari, M.; Daneshfard, B.; Emtiazy, M.; Khiveh, A.; Hashempur, M.H. Biological Effects and Clinical Applications of Dwarf Elder ( Sambucus ebulus L): A Review. J Evid Based Complementary Altern Med 2017, 22, 996–1001. [Google Scholar] [CrossRef]
- Jenkins, T.P.; Fryer, T.; Dehli, R.I.; Jürgensen, J.A.; Fuglsang-Madsen, A.; Føns, S.; Laustsen, A.H. Toxin neutralization using alternative binding proteins. Toxins 2019, 11, 53. [Google Scholar] [CrossRef] [PubMed]
- Landi, N.; Ciaramella, V.; Ragucci, S.; Chambery, A.; Ciardiello, F.; Pedone, P.V.; Troiani, T.; Di Maro, A. A Novel EGFR Targeted Immunotoxin Based on Cetuximab and Type 1 RIP Quinoin Overcomes the Cetuximab Resistance in Colorectal Cancer Cells. Toxins 2023, 15, 57. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.U. Therapeutic application of genetically engineered ribosome-inactivating toxin proteins for cancer. Journal ISSN 2021, 2766, 2276. [Google Scholar] [CrossRef]
- Flavell, D.J.; Flavell, S.U. Plant-Derived Type I Ribosome Inactivating Protein-Based Targeted Toxins: A Review of the Clinical Experience. Toxins 2022, 14, 563. [Google Scholar] [CrossRef] [PubMed]
- Knödler, M.; Buyel, J. Plant-made immunotoxin building blocks: A roadmap for producing therapeutic antibody-toxin fusions. Biotechnology Advances 2021, 47, 107683. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.-Q.; Zhu, Z.-N.; Zheng, Y.-T.; Shaw, P.-C. Engineering of ribosome-inactivating proteins for improving pharmacological properties. Toxins 2020, 12, 167. [Google Scholar] [CrossRef]
- Miranda, A.; Ismail, H.; Martien, R.; Ciptasari, U.H.; Kusniasari, A.; Arimurni, D.A.; Wahyudi, S.M.D.P.; Sismindari, S. Double-Coated Nanoparticle of Ribosome Inactivating Protein (RIP) from Mirabilis jalapa L. prepared from Chitosan-Sodium Tripolyphosphate and Alginate-Calcium Chloride: The New Strategy for Protein Drug in Oral Delivery. In Proceedings of the BIO Web of Conferences, 2023; p. 04001.
- Pizzo, E.; Di Maro, A. A new age for biomedical applications of Ribosome Inactivating Proteins (RIPs): from bioconjugate to nanoconstructs. J Biomed Sci 2016, 23, 54. [Google Scholar] [CrossRef]
Figure 1.
Synthesis of toxic bioactive compounds by plants to mitigate environmental stresses.
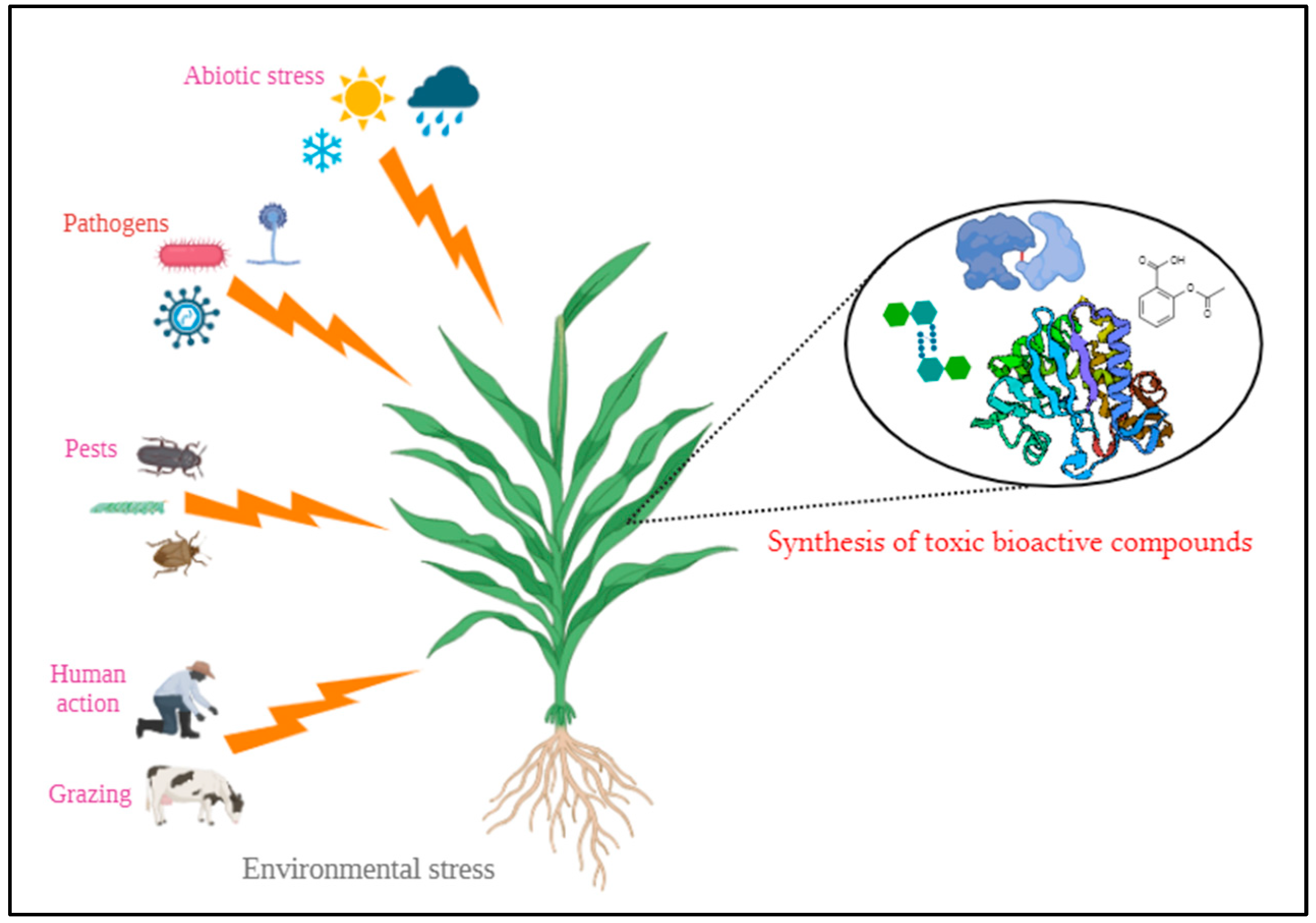
Figure 2.
Mechanism of action of Ribosome Inactivating Proteins (RIPs).
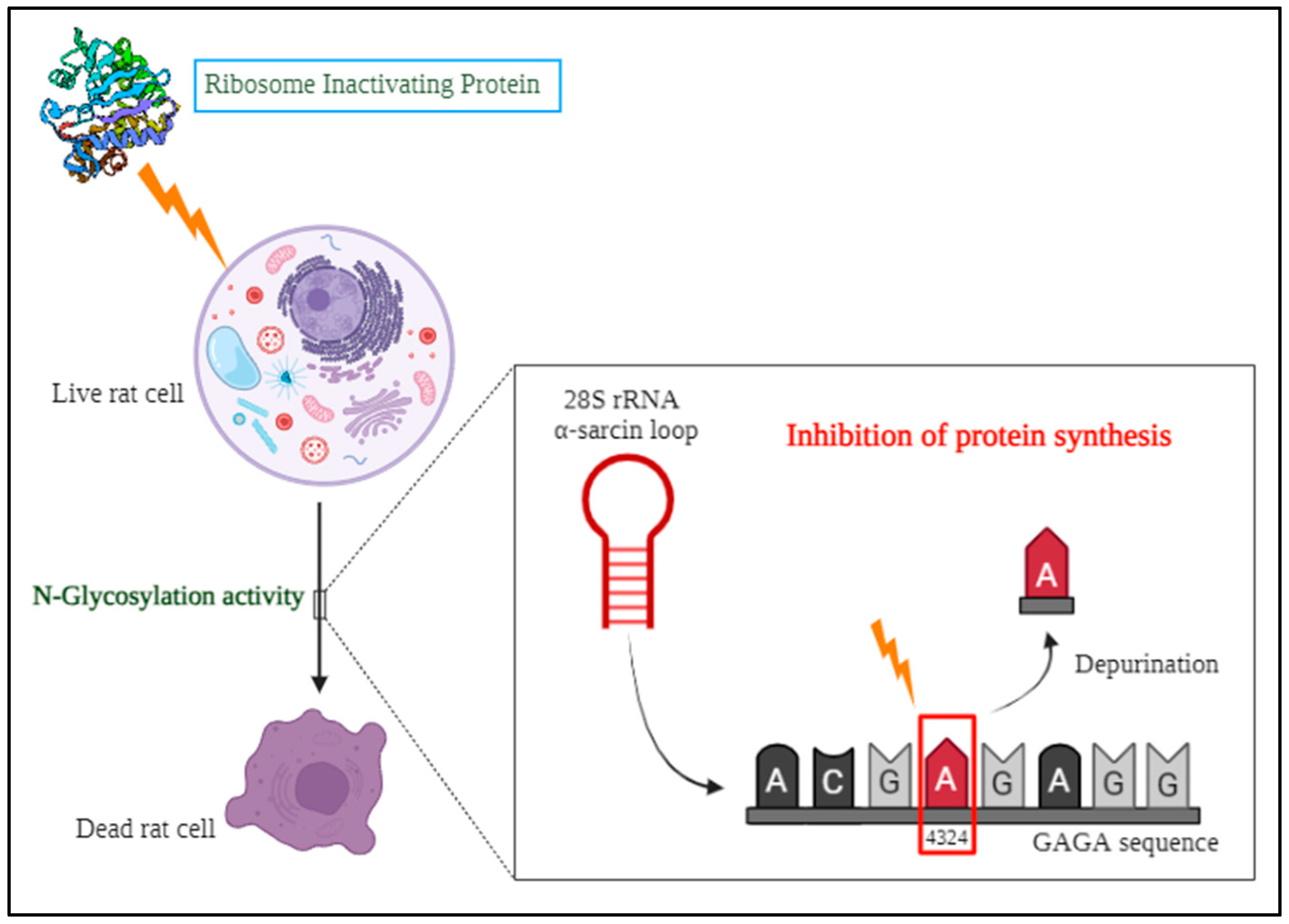
Figure 3.
Mechanism of entry of type I and type II RIPs into the host cell.
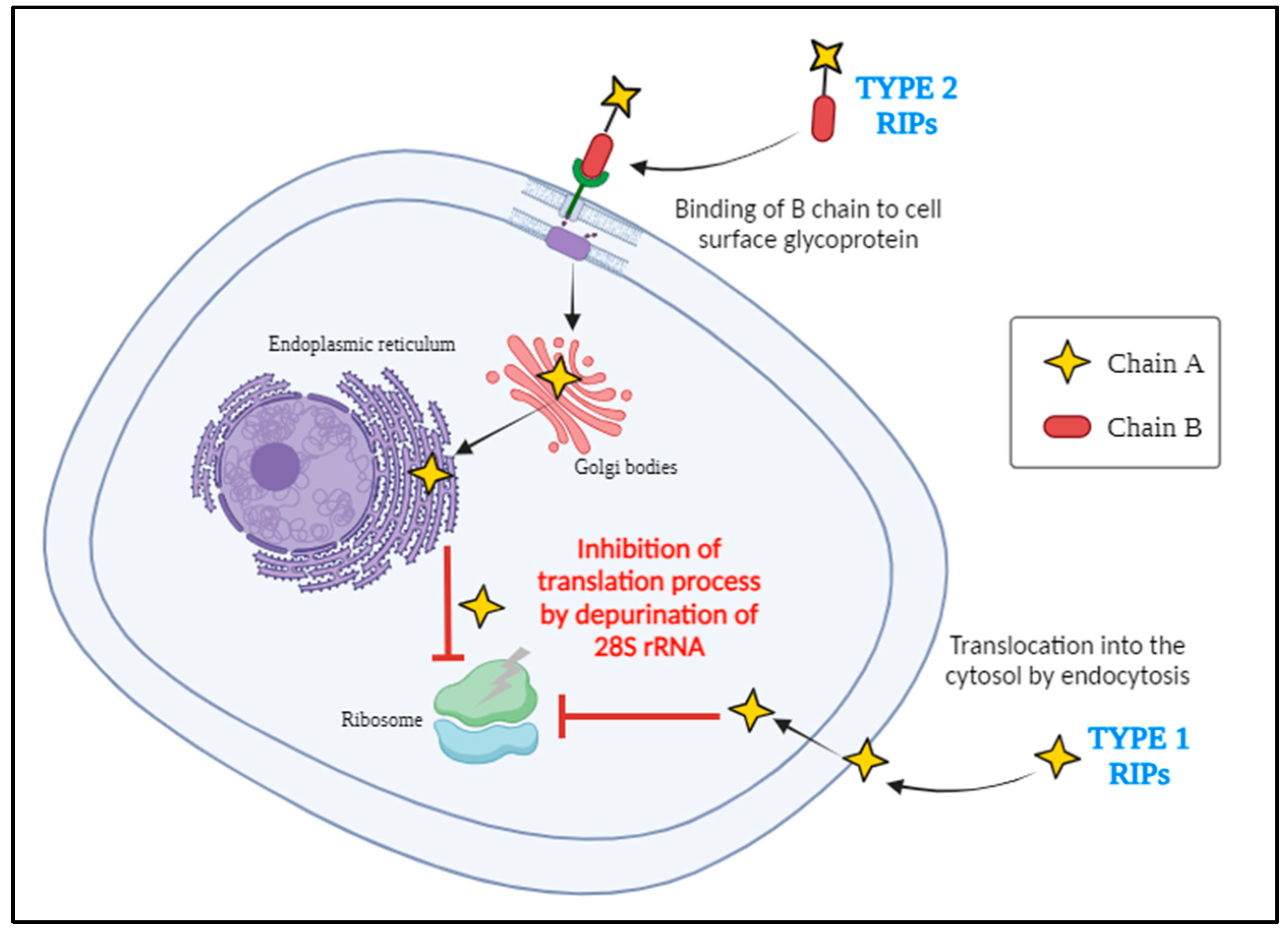
Figure 4.
Various biological roles of Plant RIPs.
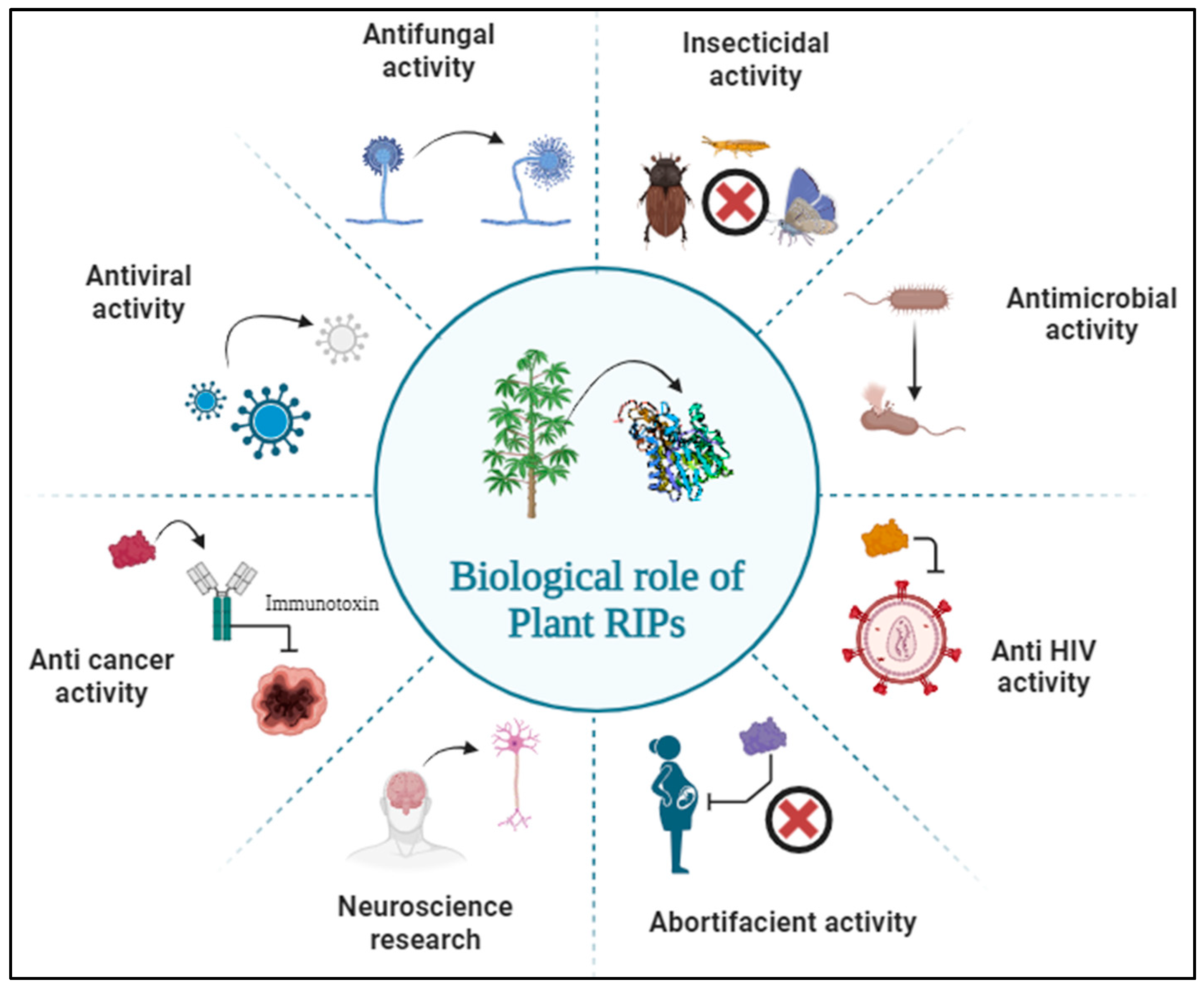
Figure 5.
Selected snake venom protein targets for molecular docking.
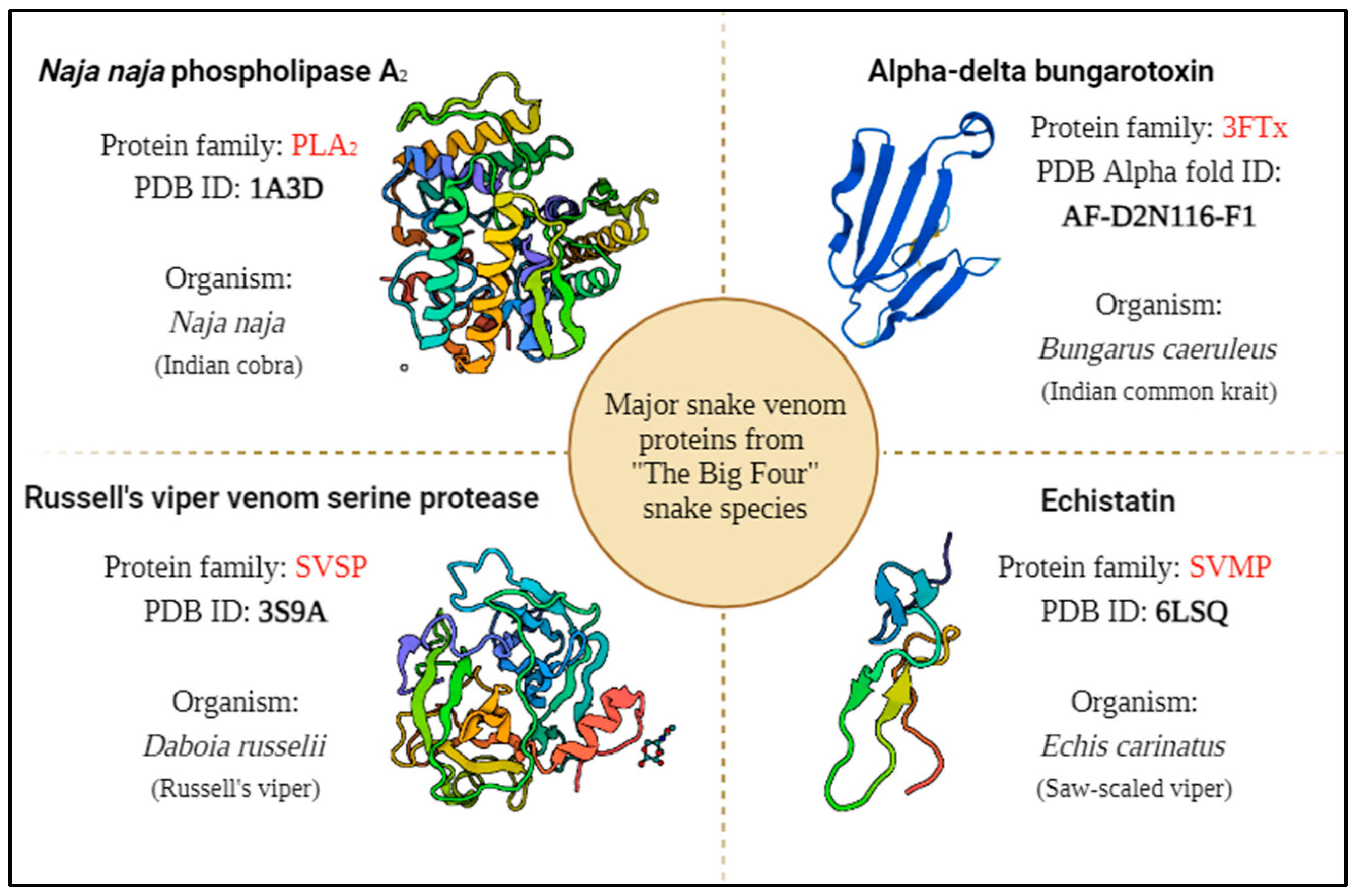
Figure 6.
Step-by-step procedure for protein-protein docking.
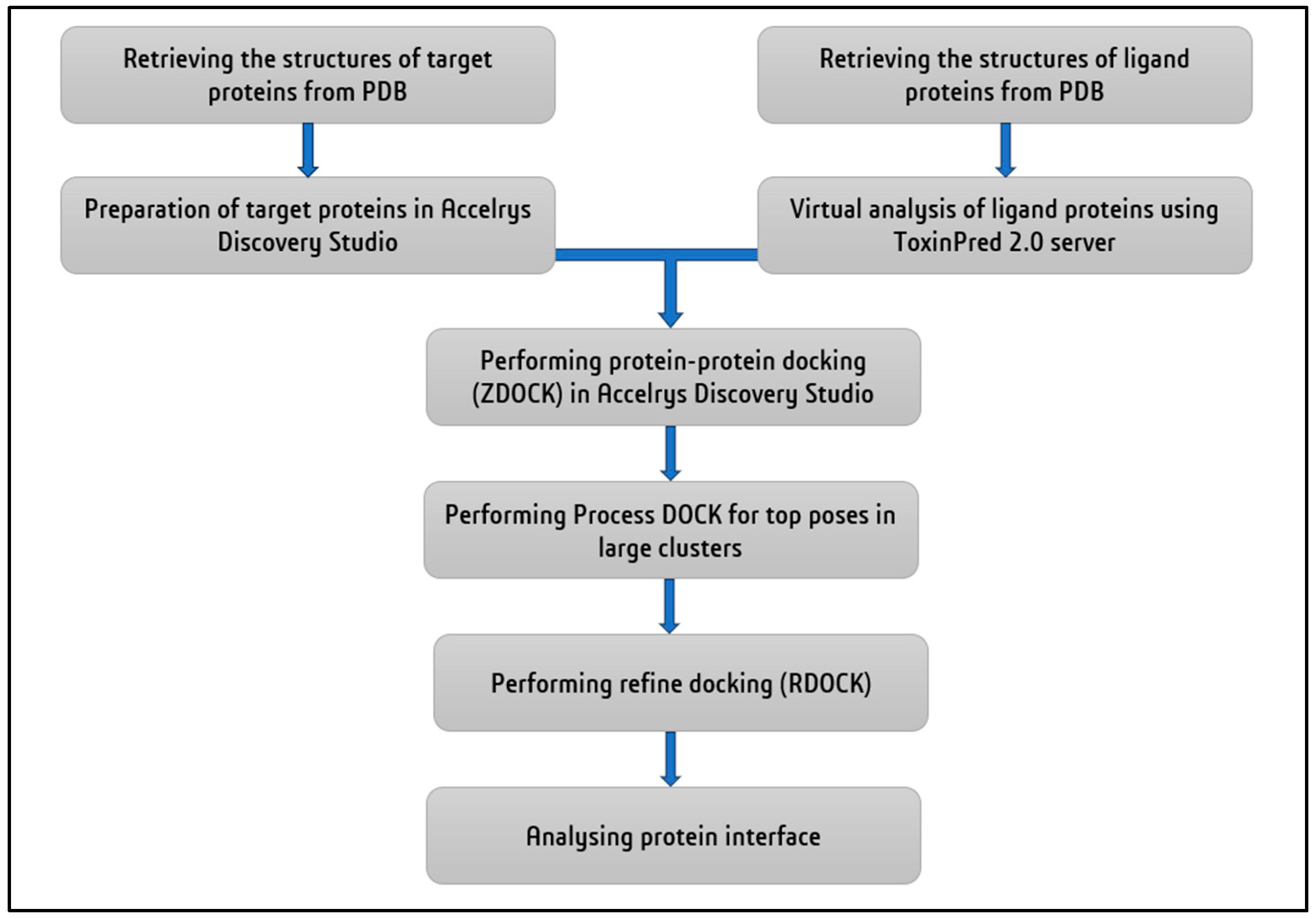
Figure 7.
Three-dimensional representation of best docked poses.
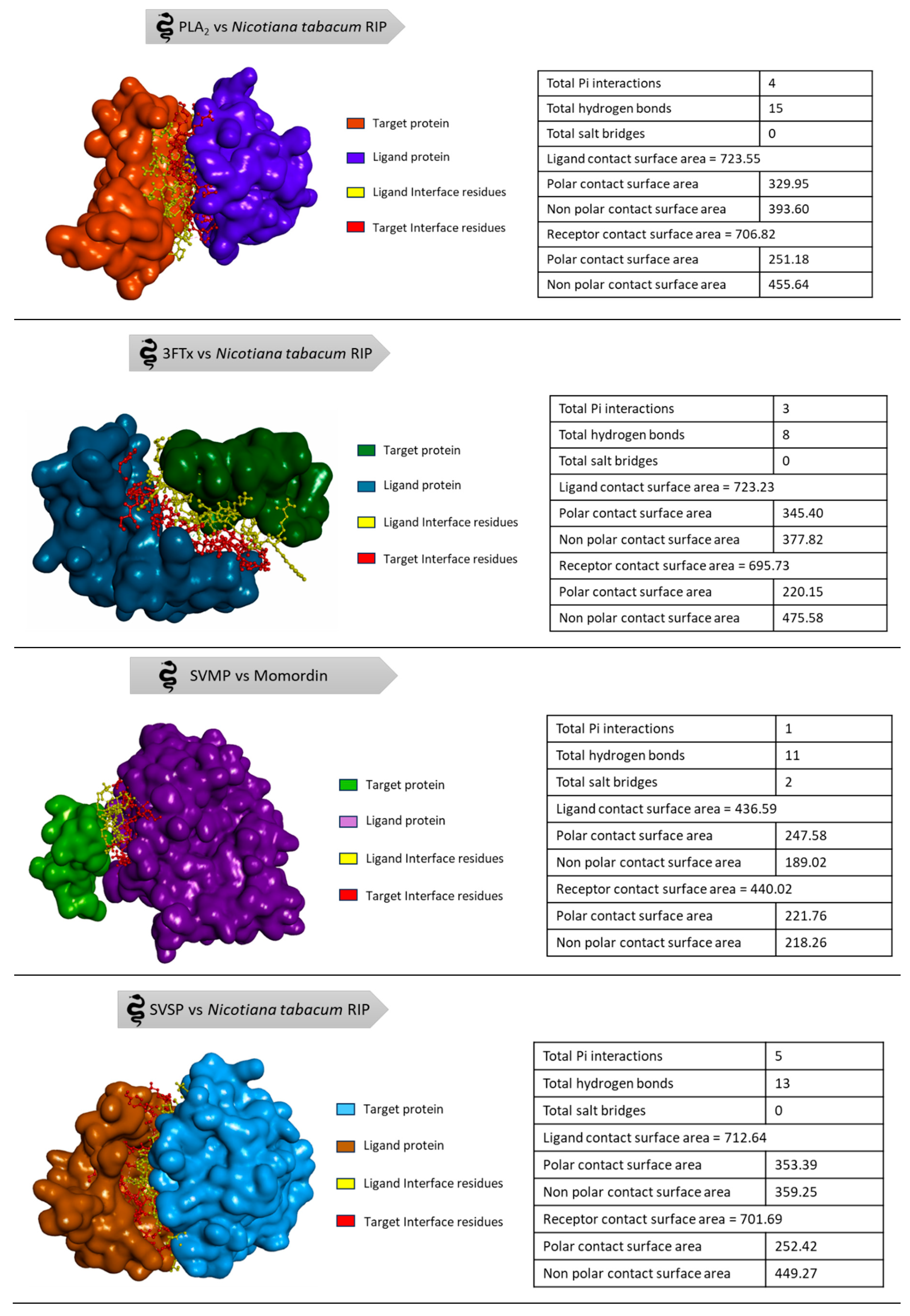

Table 1.
Group of snake venom protein families.
| S.No. | Groups | No. of Families | Family Names | References |
|---|---|---|---|---|
| 1. | Dominant protein family | 4 | Phospholipase A2 (PLA2), Three-finger toxins (3FTx), Snake venom metalloproteases (SVMP) and Snake venom serine proteases (SVSP) |
[49,50] |
| 2. | Secondary protein family | 6 | Kunitz peptides (KUN), L-amino acid oxidases (LAAO), Cysteine-rich secretory proteins (CRiSP), C-type lectins (CTL), Disintegrins (DIS) and Natriuretic peptides (NP) |
|
| 3. | Minor protein family | 9 | Acetylcholinesterase, Hyaluronidase, 5’nucleotidase, Phosphodiesterase, Phospholipase B, Nerve growth factor, Vascular endothelial growth factor, Vespryn/Ohanin and Snake venom metalloprotease inhibitor | |
| 4. | Rare protein family | 36 | Glutaminyl cyclase, Aminopeptidase, Endonuclease, Cobra venom factor, Transferrin, Waprin, Endopeptidase, Glutothione peroxidase, Kazal-type inhibitor, Galactose-binding protein, Trypsinogen, Albumin, Prokineticin, Selectin, Peroxiredoxin, Protein C activator, Cholinesterase, Polyglycine peptides, Glycine-histidine rich peptide, Flavine monoamine oxidase, Lysosomal acid lipase A, Fibrinogenases, Haemoglobins, Neurotrophin, Aspartic protease, Type-B carbyoxylesterase, Cytotoxin, Neuronal membrane glycoprotein, Insulin-like growth factor, Sulfhydryl oxidase, Aminotransferase, Complement decay accelerating factor, Kinesin-like protein, Ribosomal protein, Multiple inositol polyphosphate phosphatase and Phospholipase A2 inhibitor | |
| 5. | Unique protein family | 4 | Defensins, Waglerin, Maticotoxin and Cystatins |
Table 2.
List of plants with antivenom properties that express RIPs.
| S.No. | Scientific Name of the Plant | Common Name | Family | Plant Parts Used for Curing Snakebite | Name of the RIPs Isolated | Type | Source of RIPs | PDB/ AlphaFold ID | Molecular Weight (kDa) | References |
|---|---|---|---|---|---|---|---|---|---|---|
| 1. | Abrus precatorius | Rosary pea | Fabaceae | Roots | Abrin | Type II | Seeds | 1ABR | 60.06 | [7,87,95,96] |
| APA-1 (Abrus precatorius Agglutinin 1) |
Type II | Seeds | 2ZR1 | 118.01 | ||||||
| 2. | Amaranthus viridis | Slender amaranth | Amaranthaceae | Leaves, stem | Amaranthin | Type I | Leaves | AF-Q9SAQ5-F1 | 30.4 | [7,95,96,97] |
| 3. | Luffa acutangula | Ridge gourd | Cucurbitaceae | Fruit, seed tendrils | Luffaculin | Type I | Seeds | 2OQA | 53.11 | [7,96,98] |
| 4. | Momordica charantia | Bitter gourd | Cucurbitaceae | Flower | Momordin | Type I | Seeds | 1AHA | 27.53 | [7,95,96,99] |
| β-momorcharin | Type I | Seeds | 1CF5 | 58.3 | ||||||
| 5. | Nicotiana tabacum | Tobacco | Solanaceae | Leaves | Nicotiana tabacum RIP | Type I | Leaves | AF-B0EVM3-F1 | 14.48 | [7,78,86,96] |
| 6. | Sambucus ebulus | Danewort | Adoxaceae | Leaves | Ebulin | Type II | Leaves, green fruits, rhizomes, barks | 1HWM | 58.95 | [7,95,96,100] |
| 7. | Viscum articulatum | Flat mistletoe | Santalaceae | Whole plant | Viscum articulatum RIP | Type II | Whole plant | AF-B3F5I6-F1 | 61.96 | [7,87,96] |
Table 3.
ZDOCK and ZRANK scores of top protein poses.
| ZDOCK Scores | ZRANK Scores | |||||||
|---|---|---|---|---|---|---|---|---|
| PLA2 | 3FTx | SVMP | SVSP | PLA2 | 3FTx | SVMP | SVSP | |
| Abrin | 55.99 | 58.22 | 52.88 | 61.72 | −97.538 | −92.226 | −93.556 | −95.095 |
| APA-1 | 54.61 | 48.96 | 49.53 | 54.5 | −93.592 | −90.773 | −96.193 | −102.322 |
| Amaranthin | 60.59 | 48.27 | 51.17 | 55.59 | −109.673 | −81.871 | −87.46 | −89.727 |
| Luffaculin | 55.35 | 52.5 | 45.41 | 51.27 | −118.52 | −99.576 | −85.885 | −114.996 |
| Momordin | 62.84 | 58.19 | 46.26 | 52.51 | −123.173 | −94.896 | −98.891 | −96.195 |
| β-momorcharin | 58.19 | 62.33 | 50.12 | 55.02 | −118.085 | −109.959 | −87.866 | −99.142 |
| Nicotiana tabacum RIP | 56.06 | 62.51 | 46.34 | 54.59 | −125.688 | −113.133 | −93.512 | −127.68 |
| Ebulin | 50.89 | 52.49 | 46.42 | 58.09 | −108.197 | −102.067 | −73.95 | −101.827 |
| Viscum articulatum RIP | 74.06 | 64.39 | 66.08 | 69.31 | −105.068 | −98.496 | −97.277 | −113.711 |
Table 4.
Interface residues involved in best docked protein-protein interaction.
| Best PPI. | ZRANK Scores | Interface Residues of Target Protein Involved in the Interaction | Interface Residues of Ligand Protein Involved in the Interaction | |
|---|---|---|---|---|
| Target Protein | Ligand Protein | |||
| PLA2 | Nicotiana tabacum RIP | −125.688 | ASP39, ARG42, CYS43, GLN45, VAL46, ASN49, CYS50, GLU53, ALA54, GLU70, SER72, GLN73, GLY74, THR75, LEU76, THR77, CYS78, LYS79, GLY80, ASN82, SER88, ASP91, CYS92, ARG94, LEU95, ALA96, ILE98, CYS99, ALA101, GLY102, ALA103, PRO104 | THR1, ASN2, VAL3, VAL5, MET6, GLY7, TYR8, LEU9, VAL10, ASN11, SER12, ALA25, GLN27, TYR28, VAL29, PHE30, LYS31, GLY32, SER33, THR34, PHE62, PHE65, ILE69, PHE73, ILE87, THR90, THR91, ALA92, ALA94, SER95, ILE104 |
| 3FTx | Nicotiana tabacum RIP | −113.133 | TYR1, LEU3, SER20, GLY21, ASN23, LEU24, THR27, MET29, LYS40, ALA48, THR49, CYS50, PRO51, GLN52, PRO53, GLU58, THR60, CYS61, CYS62, SER63, THR64, ASP65, LYS66, CYS67, ASN68, PRO69, PRO71, GLN73, ARG74, PRO75, GLY76 | THR1, ASN2, VAL3, TYR4, VAL5, MET6, GLY7, TYR8, LEU9, VAL10, ASN11, SER12, PHE16, ALA25, TYR28, VAL29, PHE30, LYS31, GLY32, SER33, PHE62, PHE65, ASP66, SER67, ILE69, THR70, LEU72, PHE73, ILE87, THR90, THR91, ALA92, SER95, TRP125 |
| SVMP | Momordin | −98.891 | GLU1, CYS2, GLU3, SER4, GLY5, PRO6, CYS8, ARG9, ASN10, CYS11, LYS12 | ILE116, ALA117, ALA118, GLY119, LYS120, LYS124, ILE125, PRO126, PRO130, ALA131, ASP133, SER134, SER137, THR138, HIS141, ASP143, THR145, ALA146, ALA147, GLY149, VAL153, ASP178 |
| SVSP | Nicotiana tabacum RIP | −127.68 | ALA56, HIS57, ASP59, ARG60, ARG62, LYS88, TYR89, PHE90, CYS91, LEU92, ASN93, THR94, LYS95, PRO96, ASN97, GLY98, LEU99, PRO170, LEU171, TYR172, TRP173, HIS192, SER217, GLU218, LYS224, ARG245 | VAL3, TYR4, VAL5, MET6, GLY7, TYR8, ASN11, TYR28, VAL29, PHE30, LYS31, GLY32, PHE62, PHE65, ASP66, ILE69, THR70, PHE73, ILE87, ILE88, THR90, THR91, ALA94, SER95, GLU101, ILE104, VAL105, ILE108 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



