
Submitted:
28 September 2024
Posted:
29 September 2024
You are already at the latest version
Abstract
During recent archeological excavations in the Roman city of Carnuntum three spots with residues of streets, houses and side buildings from the time span between 1st and 4th century have been investigated. 40 soil samples taken from different layers gave the possibility to investigate the local soil bacterial communities by Next Generation Sequencing (NGS) of 16S rRNA. The background of this study was to check similarities and differences in the composition of microbial communities and to check of special types of proved bacteria or groups of them could be related to the archaeological findings. In result, the 16S rRNA data shows similarities between some samples, but also important differences in the compositions of soil bacterial communities of single samples. On the one hand, some samples are distinguished by the dominance of single major components from other samples. On the other hand, specific differences between soil samples are reflected by the patterns of mediate and less abundant bacterial groups. This supports the hypothesis, that differences in soil bacteria composition are strongly influenced by differences of the deposed soil material and, therefore, dependent on ancient human activities which created the deposits and local ancient environmental conditions many hundred years ago. This “echo” of ancient human impact on local soil bacteria composition is reflected, for example, by manure- and oil-related OTUs originating from two different street surfaces as well as by contaminated and mixed soil fills and ancient hearths and fire places. The findings support the concept of “ecological memory of soil” meaning that early impacts impress traces in the composition of soil bacterial communities which are detectable up to the present.
Keywords:
Soil bacteria
; Bacterial communities
; NGS
; Archaeology
; Excavation
; Roman Empire
; Archaeo-ecology
1. Introduction
Molecular sources are increasingly considered relevant in archaeological research. Besides the classical typology of human artefacts − pottery, tools, weapons and so on − organic residues and biomolecular markers are able to supply important information about early human populations or on the history or prehistory of ancient settlements or other places related to former human activities. Among other molecular markers, ancient DNA is particularly well suited for the identification and characterization of ancient human residues or residues of plants and animals [1,2,3,4]. The analysis of ancient DNA made it possible, for example, to reconstruct prehistoric migration processes [5], to identify the ancient cultivation of plants [6], or to detect the role of parasites in medieval cities [7].
In addition to the analysis of DNA originating from multicellular organisms, the characteristic DNA of microorganisms can help to illuminate ancient life conditions and the effect of diseases on the development of human populations. Thus, the importance of pathogenic bacteria as the Yersinia pestis which is known for the dramatic role they played during the middle ages could be understood for prehistoric populations, too [8].
Besides genetic data originating from humans or from pathogens, the genetic composition of soil bacterial communities can have been affected by human activities and, therefore, carry information on the ancient situation of a prehistoric or historic settlement or work place [9,10]. This information is of high interest for archaeologists, for reconstruction of use of places and ancient local environmental conditions and the interactions between human activities as land use, settlements, handcraft and livestock farming with soil, on the one hand. On the other hand, the understanding of ancient human impact on soil bacterial community could open new insights into long-term ecological processes in soils as well as into the possible consequences of human activities on local ecological situations after centuries and millennia.
The composition of communities of soil bacteria can vary very strongly and is often marked by a very high diversity [11]. It depends on the soil pH, salt content, humidity, nutrients and other chemical conditions and is influenced by the fate of a place in the past [12]. Changing environmental conditions mean, in general, a change in the dominance of growing bacteria in soil. But, such changes do not mean a total and immediate disappearing of all bacteria growing before the environment changes. In opposite, some of the formerly growing bacteria might switch into a dormant state, to form spores and are reduced in their concentration, but can remain be present to a certain extend for a long time. Therefore, the composition of soil bacteria has not only to be understood as a mirror of the recent ecological situation of a place, but can also be interpreted as an “ecological memory” conserving information on former local ecological conditions [13,14]. This point of view makes analyses of local soil bacteria communities interesting for archaeology. Recent studies suggest the effect of former local human impact on soil bacteria situation [15,16], for example, for an iron age settlement [17] and iron age burials [18], for prehistoric hillforts [19], for pre-industrial mining activities [20] and a historical tannery site [21]. From an archaeological point of view, a “biological memory of soil” was recognized in 2008 during the investigation of mycobiota in cultural layers of a medieval urban city and the term “soil mycological memory” was proposed [22]. The concept of ecological memory was applied for describing [23] and understanding the dynamics of interactions in communities over longer timescales [24]. Paleoecological investigations suggest ecological memory effects at millennial time scales [25]. By means of memory effects in simulations it seems to be possible to estimate the effect of perturbations on ecosystems and their resilence [26]. Memory effects in soil are in the focus of interest due to their practical consequences for agriculture [27]. Therefore, the composition of soil bacterial communities is particular important [28]. Thus, short-term memory effects in top soil related to ecological dynamics in humus and soil fertility [29,30] as well as long-term biogeocenotic functions of microorganisms and the response behavior of soil microbiota on perturbations are important factors reflecting the role of ecological memory effects on different time scales which is interesting for archaeology, but also important for future measures in response to changing ecosystems and under climate and other anthropogenically caused stress [31].
Beside the above mentioned recent investigations, published results of investigations on soil bacteria of archaeological soil samples are comparatively rare. But, data of soil bacteria compositions can include a lot of information concerning the state and the history of soil. This concerns recent chemical and nutritional conditions which are still influenced by changing composition of non-biological soil components in the past, on the one hand. They can include considerable effects as salt content, soil pH, gas permeability and oxygen availability, organic residues or presence of different nitrogen and sulfur species. On the other hand, it has to be taken in mind that formerly active bacteria could be conserved in soil pores in a less active or completely dormant state. This way, genetic information of soil bacteria components which had been active hundreds or thousands years ago could be stored in the covered soil and mirroring aspects of formerly local environmental conditions. This idea is based on the fact that changing environmental conditions cause changes of bacterial growth and shift the compositions of soil bacterial communities, but does not have to be responsible for an immediate disappearing of bacteria disadvantaged by change. In contrast, it is assumed, that dormant states of bacteria are stabilized and stored after re-locations of soil material and after covering of surface soil layers by other soil material, residues of buildings or deposition of other material on top during the changing use of a place in the past.
The present study is based on 16S rRNA analyses and shows considerable differences in soil bacteria composition at minor spatial distances. The series of soil samples from an archaeological excavation in the ancient city of Carnuntum (Austria) [32] offered the possibility to investigate soil bacteria communities from different functional and temporal archaeological contexts from one place. 40 samples taken from different horizons and well-defined specific locations with different ancient functions gave the unique opportunity to compare soil bacterial communities, to analyzing them for similarities and searching for specific types. These samples seemed to be very interesting for looking for indications contradicting or confirming the concept of “ecological memory of soil”. In addition, such an analysis was motivated by the idea of a model study for checking the potential of the genetic data of soil bacteria for supplementing of archaeological findings and conclusions. Here, the results of the analysis of the 16S rRNA of this sample set are reported.
2. Experimental
2.1. Archaeological Setting
In 2021 a 110 kV high voltage powerline was constructed which follows a trace from Bad Deutsch Altenburg (province of Lower Austria) northwards across the river Danube. It traverses the archaeological site of Carnuntum, which is situated at the southern bank of the Danube, at the site wherein antiquity the amber road forded the river. Ancient Carnuntum, the capital city of the Roman province of Upper Pannonia, consisted of a fortified legionary camp, the canabae legionis (the typical civilian settlement in the immediate vicinity of a Roman military installation) around it, a fortified cavalry camp and a separate civilian town of considerable size [33,34,35]. In its heydays, the agglomeration accommodated close to 20.000 inhabitants and occupied an area of several square kilometers. Although near to nothing of the ancient settlement and its buildings is preserved above the current surface, the complete extent of its known vestiges below ground is protected heritage. Nevertheless, it was unavoidable that three of the powerline’s pylons were situated in the known area of the canabae, east of the legionary fortress proper. To make the best out of this situation, the pits for the pylons‘ concrete fundations were not dug by machine but in the course of regular archaeological excavations. With the aim of minimizing the inevitable damage to the monument, the resulting trenches M 06, M 07 and M 08 had been purposefully situated in places, for which previous geophysical research had provided evidence that the ancient build-up was less dense than in other areas of the canabae. An overview of the archaeological excavation sites is given by combined georadar plot - aerial image in Figure 1.
In all three trenches ancient stratigraphy and architecture were exposed. Excavations followed a context oriented/stratigraphical protocol in which every discernible component of the excavated ground was treated as a distinctive and separated context (also called a stratigraphical unit), isolating it during excavation in order not to mix up different contexts and their materials. The surface layer of a road was, for instance, treated as a context, the fill-stratum underneath this surface as a separate context, the fill of a ditch dug along the road was treated as a third, separate context, the fill of a pit was yet another context, as was, for instance, a hearth, a wall, a foundation trench and its content and so on. Finds and samples collected and taken from each context were processed and archived separately, strictly avoiding any co-processual or post-processual mixing/contamination [36,37]. Soil sampling, whereas, had not been part of the original excavation strategy and was taken up only while work was proceeding. Nevertheless, even while not every single context was soil-sampled separately, every main horizon was sampled at least once, and most often twice, thrice or more often.
Due to limitation in the analysis capacity it was only possible to include a restricted numbers of samples in the investigation program. We decided to renounce of duplicates or triplicates in the investigations, but instead include 40 single separate samples in order to integrate the largest possible size of set of archaeologically well-characterized samples from the same area but originating from different horizons and different functional situations. These samples are partially taken in close neighborhood to each other. This strategy means that the reported result has a preliminary character, on the one hand. On the other hand, there was the hope to become able to demonstrate that the analysis of bacterial communities can supply variable information for supporting the interpretation of archaeological findings and to give indications to the increase of bacterial diversity due to different specific ancient human impacts.
A detailed description of the three excavation sites is given in the supplementary material. An overview on all investigated samples is given in supplementary Table S1.
All soil samples (water-dispersed soil material) show rather similar pH values in the range between about 8.4 and 9.1, which represents a weak alkaline character of soil, in general. The values of the electric conductivities the of water-dispersed soil samples are in the range between 63 and 150 µS/cm which mean moderate electrical conductivity and moderate differences between the single samples as well. The slightly enhanced conductivity values of the samples C178, C231, C232 and C266 might be caused by residual enhanced ion strength due to antique salt residues from fire-related activities. The highest conductivity was observed in the soil sample of the earth quake collapse horizon sample C295 (horizon A). This finds its most obvious cause in modern fertilizing practiced during the last six or seven decades.
2.2. Isolation of DNA, DNA Amplification, Molecular Labelling
The kits DNeasy® PowerSoil® Pro kits (Qiagen, Hilden (GE)) were applied in order to extract DNA from the single soil samples. For the procedures the suppliers’ instructions were strictly followed. For DNA amplification by PCR, a commercial laboratory thermocycler Edvocycler (Edvotek, Washington D.C.) was used. The result of each PCR process was evaluated by gel electrophoresis applying agarose gels with a solid content of 1%. Both the primary obtained PCR products and the final DNA libraries after pooling of labelled samples were treated in a pro ProNex® Size-Selective Purification System (Promega, Madison (USA)) for purification following the protocol supplied by the manufacturer.
The adapator primers have been obtained from Eurofins Genomics (Ebersberg, Germany; Amplicon PCR A519F-Ad (5’ TCGTCGG-CAGCGTCAGATGTGTATAAGAGACAGCAGCMGCCGCGGTAA 3’) and Bact_805R-Ad (5’-GTCTCGTGGGCTCGGAGATGTGTATAAGAGACAGGACTACHVGGGTATCTAATC 3’)). They were applied in the PCR with a concentration of 100 pmol/µl.
The PCR mixtures (25 µl in total per reaction) were composed as following: 0,5 µl of DNA iso-lation eluate, 2 mM MgCl2, 200 µM dNTP mix, 0,65 Units GoTaq® G2 Hot Start DNA Polymerase, nuclease-free water (all reagents from Promega, Madison (USA)) and 1 µM of each primer. The PCR process consisted of the following steps: first denaturation for 5 minutes at 94 °C, followed by 30 amplification cycles involving 30 s denaturation at 94 °C, 30 s primer annealing at 50 °C and 30 s extension at 72 °C. The temperature cycles were finished with a final extension reaction at 72 °C for 5 min.
For molecular labelling an index primer PCR was applied. The required forward and reverse indexing primers were supplied by Eurofins Genomics (Ebersberg, Germany), too. These primers were used at a concentration of 1.25 pmol/µl. For index PCR with a total volume of 25 µl per reaction, the following composition was applied: 2.5 µl amplicon PCR product, 2.5 mM MgCl2, 300 µM dNTP mix, 0.5 units GoTaq® Mdx Hot Start DNA Polymerase, nuclease-free water (all reagents from Promega, Madison (USA)), and 125 nM of each of the two primers of the respective indexing primer pair.
Following process steps were applied for the DNA amplification with index primers: initial denaturation for 3 minutes at 95 °C; eight amplification cycles involving 30 s denaturation at 95 °C, 30 s primer annealing at 55 °C and 30 s extension at 72 °C. The temperature cycles were finished with a final extension at 72 °C for 5 min.
Blank samples have been processed for the DNA extraction as well as in both PCR steps. For the control experiments the complete mastermix (buffer, nucleoties, primers, polymerase) was applied. In all steps the quality of sample DNA was checked in parallel with the amplificates of blanks in a common gel electrophoresis using ethidiumbromide as double strand DNA indicator.
2.3. Next-Generation Sequencing (NGS): Data Processing
The amplified and labelled 16S rRNA-related sample DNA was sequenced by an Illumina process by Eurofins Genomics (Ebersberg, Germany).
The NGS supplied fastq files of forward and reverse aligned 16S rRNA. In the first step of digital processing, these data were converted to fasta format including contig files and quality files (mothur (version 1.39.5)). For this, the open source platform Galaxy was used (https://usegalaxy.org/). The quality of all investigated datasets was checked by a median quality score. The value was always found to be high, indicating a very high quality of data.
In the next step, the contig files were aligned to rRNA databases based on the NCBI cloud using the SILVAngs data analysis service (https://ngs.arb-silva.de/silvangs). This procedure allowed a detailed analysis on the basis of the previously obtained sequencing data, supplying information about the bacterial community of the related sample [38,39,40]. For all analyses, the preset parameter configurations of the SILVAngs database version 138.1 were applied [40]. For identification of OTUs an identity index of 0.98 (threshold of 98%) was used. For the taxonomical alignment of NGS data, a classification similarity index of 0.93 was applied. In the overwhelming part of classifications, this index is about 0.99.
The alignment of sequencing data supplies a taxonomic assignment on phylum, class, order, family and genus level. In most cases, identification down to the genus level is feasible. In some other cases, the assignment is only possible for higher taxonomic levels. The lowest identified level for each distinguished bacterial type is referred as the Operational Taxonomic Unit (OTU). In the following, the compositions of soil bacterial communities are always related to these OTUs, that means on the genera level as far as possible.
2.4. Data Analyses
The data analyses are based on three main approaches: At first, the composition of soil bacterial community of each single sample could be described, discussed and compared with each other. At second, data of groups of samples should be regarded in order to identify possible patterns in the appearance of selected bacterial types. At third, it should be tried to find specific bacterial types or patterns with the potential to support the interpretation of archaeological findings and to give hints for checking the concept of “archaeological memory of soil.
Different methods are applied to evaluate the taxonomic data and sample-specific abundances of OTUs. First, the percentages of the most abundant phyla are simply compared in a bar chart. The associated plot includes an automatic normalization of the read counts. Bar charts are also used to plot absolute read counts reflecting the different abundances of specific OTUs in various samples.
On the one hand, the correlation of abundances when considering the complete data sets was represented by binary logarithmic correlation plots. They allow comparison of sample pairs based on the individual abundances of all detected OTUs by plotting these abundances with one sample as the x-axis and the other as the y-axis. On the other hand, correlation coefficients were calculated for all sample pairs.
The operational tools of Microsoft Excel was applied for simple quantitative analysis of soil bacteria abundances in the communities of single samples, as well as for looking for sample-specific bacterial types and characteristic pattern in abundances of small groups of bacteria. The same program was used for calculation of correlation coefficients and for graphical plots of abundance data.
Since double logarithmic correlation plots give a better impression of the relationship between pairs of samples than linear plots, normalized logarithmic abundance values were included in some analyses. These normalized abundance r-values are expressed as: the ratio of the individual read counts N to the total read count of a sample Nsum:
r = log10 (1+106 * N / Nsum)
The idea behind this log transformation was the intention to enhance the awareness against less abundant OTUs and to include them with a similar weight into comparisons between OTU patterns of different samples. This strategy seems to be interesting under the assumption that slowly growing or dormant bacteria in lower concentrations might represent the long-term microbial memory of soil, in particular. In graphs based on the r-values, the complete populations in the bacterial communities are compared according to the magnitude of abundances. In this way, the correlations between higher and lower abundant OTUs are presented in a common picture.
The distribution of OTUs in the different samples can also be investigated by principle component analyses (PCA). This analysis was performed by the PCA function of Mathlab. The corresponding two- dimensional plots illustrate similarities and dissimilarities between samples and groups of them very clearly.
The motivation for this study was to get an idea how information on local soil bacterial communities can be used for supporting interpretations of archaeological findings. Therefore the sample set from Carnuntum gave us an excellent opportunity because it consists of 40 samples from different sampling points and eight different temporal horizons spanning a time frame of about 4 centuries and are related to different local human impact as substreet construction, street surface, building, courts, different fillings, hearths and fire places. Due to the relative high number of samples and limited resources we decide to process all samples instead of making duplicates or triplicates of only a part of them. Besides an overview about the presence of phyla in the single samples, we add data and graphs illustrating the Alpha diversities in single samples as well as the Beta diversity of soil bacterial community. In addition, the manuscript and the supplementary material contain beam diagrams reflecting specificities of samples and relations between them.
3. Results and Discussions
3.1. Composition of Soil Bacteria Communities by Phyla
The sample set is generally characterized by high abundances of Proteobacteria, Actinobacteria and Firmicutes, in most cases (Figure 2). The strongest dominance of Proteobacteria was found in samples C137, C150 and C299. These samples were taken from horizons of a 2nd century building and adjacent courtyard. In sample C232 − taken from the ashlayer on a hearth dating to the late 1st century − more than 60% of reads belong to Firmicutes. A particularly high content of Firmicutes was also found in sample C300, which originates from the floor of a 3rd century courtyard. Sample C165 – taken from the fill underneath the surface of a 2nd century street – is dominated by more than 60% of reads of Actinobacteria. The samples show the abundance of Archaea, too, among them Nanoarchaea and − in most cases in comparatively high abundances − Crenarchaea.
3.2. Composition of Soil Bacterial Communities by OTUs
3.2.1. Alpha Diversity Characterized by Logarithmic Rank Functions
The NGS of the investigated samples supplied between about 20,000 and 80,000 reads per sample (with exception of samples C282, which showed very low read numbers). The number of OTUs with at least 1 read per sample was in the range between 142 and 791. The number of OTUs with at least 10 reads per sample was in the range between 39 and 382. This means that the sample group is marked by a comparatively large spectrum of alpha diversities.
The differences in the alpha diversities are well reflected by logarithmic rank order functions (Figure 3). Each sample is marked by a characteristic line reflecting the distribution of read numbers on OTUs. The logarithmic plot illustrates particular well the differences in the distribution of reads for OTUs with low and mediate abundances.
It is remarkable that a part of samples shows rather similar rank functions with a simple exponential character (approximately linear range in the logarithmic plot) for lower and mediate abundant OTUs (up to about 100 reads per OUT). In contrast, the other part of samples is marked by lacks of lower abundant OTUs which results in strong shoulders in the logarithmic rank functions. This concerns particular strong the samples C119, C150, C169, C298, C299 and C300 which are marked by low total numbers of proved OTUs. It seems that the shoulder in the rank function of these samples are preferred related to samples from filling material. In contrast, some samples from inside buildings, from hearth places and street surfaces are marked by a high number of proved OTUs resulting in straight lines in the lower parts of logarithmic rank orders.
3.2.2. Highly Abundant Special Bacteria
A great part of samples is marked by relatively high abundances of single OTUs. 14 samples contain more than 5% Nocardioides (Figure 4a). This genus generally has comparatively high abundances. It is mesophilic and aerob. Aeromicrobium was found in 12 samples in high abundances (Figure 4b). Aeromicrobium is closely related to Nocardioides and belongs to the family Nocardioidaceae, too. It is remarkable that this genus appears in 5 samples of excavation spot C, among them four times in fire-related contexts. With more than 20% of reads, it is particularly dominant in the context of the debris of a 2nd century cupola oven. The soil samples with high Aeromicrobium read numbers from other excavated contexts seem to confirm the relation of this genus to fire-related matter. High abundances of Limnobacter were detected in the lowest investigated layer originating from the first century, for example in soil fill (Figure 4c). Limnobacter was first described in a German fresh water lake sediment in the species Limnobacter thiooxidans which has the ability to oxidize sulphur-containing compounds [41]. This ability could hint to sulphur-containing substrates which might have originated from sulphur-rich organic materials, for example proteins like keratin from hair, horn or other animal residues. Pseudonocardia (Figure 4d) are particular abundant (more than 10% of reads) in samples C119, C162 and C165 originating from the same spot (B) and deeper horizons D and E (late 2nd century). Such a high abundance of Pseudonocardia was also observed in the sandy alluvium (”Marchsand”, C263, sampled in trench M 07). Pseudonocardia is, in general, a widely distributed genus of Actinobacteria. The abundance of Acinetobacter is interesting as well. It was observed with over 20% of the reads in samples C150 and C299, in about 50% of the reads in C137 and about 10% of the reads in sample C263 (sandy Alluvium), whereas all other samples contain low or very low contents of Acinetobacter. In analogy to Pseudonocardia, high abundances of Acinetobacter appeared in horizons D and E (late 2nd century), but at the sampling sites C and D (trench M08). Acinetobacter generally is a highly abundant soil bacterium too. It belongs to Gammaproteobacteria. The genus is known for strains which are able to degrade aromatic hydrocarbons. Four general highly abundant and frequently dominating genera are also the most abundant OTUs in sample of sandy alluvium C263 which can be regarded as a reference sample. This concerns Nocardioides, Pseudonocardia, Bacillus and Acinetobacter. Their presence in all samples and their massive occurence in C263 and in many of the other samples allows to interpreting them as main standard components of bacterial communities with higher relevance for the general and recent soil conditions than for a spot-specific ancient situation or ancient human impact. The soil-fill samples C119 and C283 show a similar abundance pattern as the marchsand sample C263 concerning these four main components. But all other samples deviate stronger from the abundance ratios of these OTUs in C263. This can be taken as a general hint to ancient human impact.
In contrast to the above mentioned OTUs, there are few groups of soil bacteria which appear in a single sample much higher abundant than in all others (Figure 4g-j). Thus, Lysinibacterium and Psychrobacillus are highly abundant in sample C232 (ash of hearth, horizon H, 2nd half 1st century), Exiguobacterium in C300 (floor filling in court area, horizon C, 3rd century). In particular, in sample C232, the high read numbers seem to indicate a special ecological situation in the hearth/ash-determined soil material. Lysinibacterium, Exiguobacterium as well as Psychrobacillus belong to the gram positive family Bacillaceae. Exiguobacterium is reported to be able to metabolize highly resistant synthetic polymers as polystyrene [42].
Besides the appearance of the dominant OTUs in special samples, the complete absence or very low presence of generally highly abundant OTUs in one or few samples shed a light on specific recent situations in these samples. The findings support the hypothesis that formerly human impact can causes a considerable increase in Beta diversity [21]. An overview on the OTU patterns of single samples is given in the supplementary material.
3.2.3. Relations between Samples Reflected by Principle Component Analyses (PCA)
For PCA, the logarithm-related r-values of abundances have been used. This procedure should help to consider the less abundant types adequately besides the higher abundant OTUs. The correlation for first and second principle component over all OTUs reflects certain distribution, but not clear clustering of samples.
In addition, PCA was performed separately for different abundance classes. Therefore, the total abundance over all samples was chosen as criterion. The groups of lower abundant OTUs (10-100 and 100-1000 reads in total) showed certain structured patterns. It seemed that the first PC for low and mediate abundant OTUs was better suited for distinguishing of samples then second PC. Therefore, we chosen the first principle component of the both lower abundant groups and plotted these data in a 2D diagram (Supplementary Figure S14). This plot support the impression that the total of lower abundant OTUs is better suited than the highly abundant types for distinguishing specificities of samples in memory effects in their soil bacterial communities.
In addition, this possibility of distinguishing sample groups shows for our opinion clearly that – despite all noise and uncertainty in interpretation of quantitative data of single OTUs – the common reconsideration of all OTUs of one abundance class can help to distinguish samples by quantification in PCA. This interpretation meets the assumption that OTUs related to ancient perturbation and human impact on soil are preferably found among the mediate and lower abundant bacteria. In our case, the assigning of the cluster types to the soil profile supplies neither a clear relation to the specific locations nor to the horizons (Supplementary Figure S14). Thus it can only be speculated that other differences in formerly human impacts are reflected by the sample clustering when lower abundant OTUs are reconsidered. The clustering seems to separate a group of samples from divers fills from a sample group including furnaces, hearths and the barbecue pit beside other fills.
For low read numbers (10-100 reads in total) four clusters of samples can be distinguished which are mainly separated by PC1 (Figure 5a). For high read numbers (>10,000 reads) two groups of samples are identified, distinguished from each other slightly in the first and the second principal component PC1 and PC2. It is remarkable that the fire-related samples are mostly found in one of the both cluster (low values for PC1), what speaks for a memory effect to fire reflected in the group of highly abundant bacteria (Figure 5b). By PCA data it can only be speculated about specific differences in the local recent and ancient ecological situation for selected samples groups. Thus, samples C190, C218 and C219 (trench M08, sampling area A) are found in the same cluster, in all cases reflecting the strong local relation and, probably, a certain ecological similarity, too. The same is true for C295 (horizon A) and C297 (horizon C) both originating from trench M08 sampling area D, whereas the intermediary sample C296 (horizon B) from the same sampling area is in all ways quite different from the both other samples. The last mentioned conspicuousness is confirmed by looking at highly abundant OTUs, as well (see below).
A PCA analysis for OTUs with more than 10 000 reads in total allowed to identifying a set of five OTUs, which form a separate cluster in PC1/PC2 correlation diagram (green triangles in supplementary Figure S15). To this cluster belong Kryptoniales BSV26, Polaromonas, Limnobacter, a uncultivated genus of Solimonadaceae and strains of Nitrosotaleaceae. Their abundance distribution in the samples shows for all five OTUs a clear subdivision in two abundance groups: samples with mediate to high abundances (above about 40 reads) and samples with no or a few reads, only. The last mentioned bacteria group (low abundance) concerns mainly fill material from horizons A-E, including C119, C150, C162, C165, C296, C299 and C300. Higher abundances of the five mentioned OTUs were proved in samples from horizon F-H, but in addition in samples C295 (horizon A) and C297 (horizon C). These samples are related to court and building. Remarkably, Polaromonas and Limnobacter are described by thiosulfate and sulphur oxidizing strains [43,44]. The Kryptonia group – found in geothermal springs - seems to be related to iron respiration and might by related to sulphur redox processes, too [45]. Sulphur-related organic residues could hint to organic material from animals like horn and hair and can be expected from building and court. These features correspond to the lower stratigraphic situation of the horizons F – H and give a hint on translocation of deeper material in the higher levels of the sampling sites of C295 and C297.
Obviously, the formation of clusters by analysing the PCA is suited for a raw evaluation of sample groups and OTU groups. But it has to keep in mind that it is not suited to be applied for a getting a detailed picture of sample relations, in our case. For a comparative analysis in the following small groups of OTUs will be analysed in order to identifying relations between pairs or small groups of samples.
An estimation of similarities between pairs of samples was possible by calculation of Bray-Curtis Dissimilarity Indices. For these calculations, the r-values (normalized logarithmic values, eq. (1)) have been used in order to reconsider higher and lower abundant OTUs. The results for the 10 OTUs shown reads in all 40 samples are shown in Supplementary Table S2. For comparison, a second set of Bray-Curtis Dissimilarity Indices was calculated reconsidering a set of mediate abundant OTUs present in 24 of the 40 samples (supplementary Table S3). Low dissimilarity values have been found, for example, for sample pairs C124 and C169 and for C341 and C347 in the highly abundant OTU group (supplementary Table S2). In the group of OTUs present in 24 samples the three sample pairs C119/C263, C248/C347 and C297/C347 showed particular low dissimilarity values (supplementary Table S3).
3.3. Finger Print-like Abundance Patterns (FPP) and Relations between Samples
3.3.1. Close Neighbourhood
During the analysis of the complete data sheet of OTU abundances it was observed that in several cases groups of OTUs appear at a few or two samples, only, but are mostly absent in the majority of other samples. These OTU groups seem to represent finger print-like patterns (FPP). Besides, the sample-exclusive OTUs, the OTUs in the FPPs could be interpreted for getting sample-specific or group-specific information.
Some of the FPPs were found in samples which had been taken from sampling points situated closely to another. In these cases, specific local conditions are responsible, obviously, for the formation of these patterns. Such a situation is clearly indicated by the abundance pattern for a group of 19 OTUs which are proved in sample C218 as well as in C219 (Figure 6). Both samples originate from the fill of a barbecue pit. Besides this, it is remarkable that the pattern is mainly formed by OTUs with few dozen reads, only. This indicates that sets of small read numbers as found in the FPPs might be useful for identification of relations, in general.
In the majority of other samples, only a few, one or no of the 19 FPP-OTUs of samples C218 and C219 are present. Exceptions to this are the samples C147, C248, C258, C288, C295, C297, C330, C341 and C347, which are marked by parts of the mentioned OTU-group. These might speak for a certain relation of the samples to samples C218 and C219. It is remarkable that C147 and C178 originate from hearths. The samples C330, C341 and C347 originate from a heating system − they are fire-related as well. The group of the above mentioned 19 OTUs includes nine well described genera, which match all aerobic environmental conditions. Several of them are also present in samples C190 (street surface) and, in particular, in C288 (street, substructure). This concerns Solimonas [46], Bacteriovorax [47], the ammonia-oxidizing Archaeon Nitrosarchaeum [48], the starch-hydrolizing bacterium Flaviaestuariibacter [49], Pontibacter [50], the oxalate-decomposing Oxalicibacterium [51], Flavisolibacter [52], Ahniella [53] and Sphingorhabdus [54].
A second example of a remarkable FPP formed by 11 OTUs concerns the samples C295 and C297. The single involved OTUs are represented by read numbers in the order of magnitude of 10, only. The samples originate from the same areas, but different horizons. They are separated by the horizon of sample C296, but surprisingly only one of the 11 OTUs was detected in this sample (Figure 7a). The characteristic group of OTUs includes the methanogenic bacterium Methanobrevibacter [55], alkaliphilic and moderately halophilic Anaerobacillus [56], Labrys [57], the rumen bacterium Acetitomaculum [58], the moderately thermotolerant Sinibacillus [59] and Pseudogracilibacillus [60]. The majority of these bacteria indicate obviously a formerly more or less anaerobic environment.
A similar situation was observed for samples C162 and C288, which also come from the same area, but are separated by several intermediate horizons (Figure 7b). They are distinguished from most other samples by seven OTUs, including the genera Cystobacter, Planctomicrobium, Olivibacter, Cryptanaerobacter and Pseudoclostridium. Olivibacter was found in alkaline viscous oil mill waste [61] and could indicate formerly oil-rich and probably alkaline local environment. Cryptanaerobacter was first isolated from mixed waste with sewage sludge and swine waste and found to be able to transform phenols [62]. Pseudoclostridium was found in sludge from an oil field and is able to degrade Xylan [63]. This group of bacteria could indicate a surface contaminated with oil and manure. This could match well with the expected state of a street surface distinguishing it from a street substructure below its surface as corresponding to sample C165, originating from a intermediate horizon between C162 and C288. The interpretation of a formerly oil- and manure-contaminated surface for C162 corresponds very well with the profile of the less abundant exclusive types.
A local relation could also be reflected by the FPP connecting the samples C124 and C296. Whereas the specific bacterial compositions of these both samples are very different to C295 and C297, they both show a group of four bacterial types which was rarely found in other samples, among them the genera Pseudolabrys, Halodurantibacterium and Phytohabitans (Suppl.-Figure S16). Pseudolabrys was identified as a soil bacterium being able to metabolize organic acids [64]. Halodurantibacterium was isolated from an oil production mixture of a northeastern chinese oilfield [65], Phytohabitans was first isolated from orchid roots [66]. The first two OTUs mentioned might indicate a local environment contaminated to some degree with oil.
3.3.2. Stratigraphic Relations
Besides local relations, some pairs of samples from different sampling spots seem to be related by their stratigraphic situation. Several of these relations concern the low neighbourhood horizons G and H. Thus samples C218 and C219 (horizon G) on the one hand and C231 and C288 (neighbourhood horizon H) on the other hand are marked by a set of five OTUs from different taxonomic groups, which are rarer in other samples. In addition, sample C218 and C288 are related by a FPP of the five OTUs Hirschia, the sulphate-reducer Desulfosporosinus [67], Cand. Ryanbacteria, Lutispora and the type numbered as CG1-02-57-44.
The samples C124 and C162 belonging to the neighbourhood horizons E and D are distinguished from all other samples by a signifcant abundance pattern of a group of six bacteria (Suppl. Figure S17a) among them the halophilic Salinococcus [68], Actinoplanes, Enhydrobacter and OTUs from the families Thermoactinomycetaceae and Streptosporangiaceae. Sample C162 shows also a common FPP with sample C266. This bacteria group concerns Prauserella – first isolated from alkaline soil [69], Aerococcus and both the genetically determined OTUs Cand. Peregrinibacteria and the group Lachnospiraceae NK4A136. The samples C295 and C296, which are different in most parts of the abundance pattern show a common FPP of four types which are found together only in sample C178. It consists of the sulphur-oxidizer Sulfuriferula [70], Nakamurella, the phosphate-accumulating bacterium Microlunatus [71] and Nibribacter.
The pairing of samples C111 and C124 both originating from horizon D (first half of 3rd century) is marked by a rather similar abundance pattern (Suppl. Figure S17b) involving the ammonia-producing Curtobacterium [72], Facklamia, Knoellia, Ruminococcus and Terrisporobacter. Knoellia was found, for example, in pig manure [73], starch-decomposing Ruminococcus in bile [74]. Strains of the genus Terrisporobacter were isolated from bovine manure [75] as well as from a petrol reservoir of an oil field [76]. The existence of this OTU group suggests a relation to the common horizon D. All of these five bacteria are found with low read numbers in C248, too, four of them in samples C147, C169 and C295, which are related to different horizons. The appearance of these bacteria in C248 might be caused by the relocation of soil material during the excavation of 1905. A common feature of all samples could be the character of clayey fills contaminated by dung and other kinds of organic waste, which matches the interpretation of the sets of low abundant exclusive OTUs (see Section 3.2.2.) of both sampling areas.
3.3.3. Fill-Related Patterns
Sample C248 represents thoroughly mixed material of ancient and recent origin. It contains a group of 12 bacteria, most of which are observed in samples C295, C306 and C315 as well, but occur only partially or are completely absent in most other samples (Suppl. Figure S18a). It is remarkable, that the afore mentioned four samples all originate from mixed deposits – C248 from the archaeological excavation at the beginning of 20th century, C306 and C315 from debris left over from the earth quake of 360 AD, and C295 was taken from earch quake debris most probably conatminated by the plow. In addition, C248 has a common FPP of five bacteria – Lentimonas, Saccharotrix, Ahniella, JdFR-76 plus a genus not further characterized of Xanthobacteraceae – with the earth quake material from sample C315 (Suppl. Figure S18b). It seems justified to assume that this specific similarity between C248 and C315 is the result of the mixing of ancient and recent soil material – in one case (C248) by the old excavation.
A FPP of three bacteria connects the fill material of samples C124, C172, C286 and C306, but distinguishes them from the mixed soil material of C248 and most other samples. It concerns Fonticella, Aneurinibacillus and Ureibacillus. Following literature data, Ureibacillus [77] was found in compost and prefers higher temperatures for growth, as does the thermophilic bacterium Fonticella [78].
Samples C162 and C296 contain material from ancient surfaces, but originate from different horizons (E and A). They are marked by FPP of four bacteria, documented in sample C306 as well – an ancient planum after the 4th century earth quake.. This bacteria group includes Jiangella, Jeotgalibacillus, Micromonospora and an uncultivated genus of Beijerinckiaceae. Samples C169 (horizon F) and C283 (horizon H) are marked by a similar FPP with six bacteria consisting of thermophilic Herbinix [79], halophilic Haloplasma [80], and methylotrophic Methylobacillus, Methylamin-metabolizing Methylotenera [81] and uncultivated genera of the families Nocardioidaceae and Syntrophomonadaceae.
The FPP of three OTUs - Afipia, the Arsen-tolerant bacterium Arsenicitalea [82] and the halophilic and cellulolytic bacterium Halocella [83] - seems to be related to soil fills, too. It was found in samples C111, C162 and C223. Combinations of Arsenicitalea and Halocella were also found in samples C169 (fill in courtyard, 2nd century), C286 (fill of horizon H) and C288 (same horizon), combinations with Afipia in C283 (soil filling besides hearth, horizon H), C295 (horizon A), C297 (horizon B) and C341 (horizon of early 4th century).
3.3.4. Fire- or Heat-Related Abundance Patterns
The bacteria-quadrupel Solimonas, the thermophilic Fontimonas, Alicyclophilus – a nitrate-reducer able to degrade cyclohexanol [84] – and OTU PHOS-HE36 are forming a FPP which was found in the coal material of sample C219, in the sample from the hearth floor C231 as well as – from trench M 06 situated approximately 600 m distant – in the sample of the praefurnium floor C341 (Suppl. Figure S19a). Three of the bacteria (exception: PHOS-HE36) occur in praefurnium floor sample C347, taken from the lowest/earliest floor of the same praefurnium of which C341 represents a later horizon of use as well.
The community compositions of the conflagration debris sample C266 and sample C330 (praefurnium floor superimposed above sample floor C341) are marked by a common quadruple of lower abundant OTUs made up of the thermophilic bacterium Limnochorda [85], the ammonia-oxidizer Nitrosarchaeum, the thermophilic bacterium Planifilum [86] and Rhizobiales group A0839 (Suppl.-Figure S19b).
Samples from hearths are marked by OTUs which are reported to be related to enhanced salt contents. The measurements of electrical conductivity do not speak for a particular high salt content in the recent soil material. Therefore, it can be concluded that the rRNA-profiling indicates not a recent, but a historically enhanced salt content – probably due to patterns of use of the places. This would well agree with the character of fire places and accumulation of salts from ashes in antiquity.
3.4. Local Distribution of OTUs on Excavation Site M08
The extensive sampling carried out in trench M08 offers the possibility to analyse and interpret differences of closely located sampling points of different horizons. Beside some high abundant OTUs, especially lower abundant but specific OTUs are considered here.
Some of the identifiedOTUs are marked by some kind of special spatial distribution. Besides local concentrations, depth seems to be an important criterion. There are bacteria which are preferably found in upper horizons as Ralstonia (Figure 8a) and – in contrast − others which are higher abundant in lower layers as, for example, Hydrogenophaga (Figure 8b), Limnobacter and Zixibacteria (Suppl.-Figure S20). Ralstonia is, for example, a very abundant genus of soil microorganisms and includes phytopathogens. Hydrogenophaga is a genus of hydrogen-oxidizing bacteria [87]. Limnobacter is known for its ability to oxidize thiosulfate [41]. Zixibacteria have been identified in aquifer sediment samples, but not cultivated and studied in detail up to now [88]. Obviously, the three last mentioned OTUs are typically found in nutrient-rich and wet soil.
The comparison of correlations of highly abundant with OTUs of low abundance creates a different picture. An impression of the differences in the general community composition is given by the correlation of the abundances of 38 highly abundant bacteria in single samples and on average. It is obvious, that there are samples with comparatively high similarity to the average as C124, and surprisingly also C218 and C219 (Suppl. Figure 8a-c) and others which show no correlation, as, for example, soil fill horizons C119, C 150 and C165 (Suppl. Figure S21d−f). Low correlation is also observed between the mean r-values of C232 (hearth ash, 1st century, Suppl. Figure S21g), C300 (soil fill in courtyard, 3rd century, Suppl. Figure S21h) and C263 (undisturbed alluvium, reference sample, Suppl. Figure S21i).This picture is confirmed also by the comparison of other samples with sample C263 (alluvium). The PCA of r-values of the 40 bacteria representing the highest similarity with C263 (procedure described in supplementary material) displays a clear distinction between C263-similar (blue diamonds in Figure 9a) and C263-dissimilar samples (red squares in Figure 10a). Whereas the C263-similar samples form a rather closed cloud of points, the C263-strange samples also shows high distances between each other indicating a high diversity in the matrix of abundances. In Figure 9b, these both groups of samples are marked by blue (C263-similar) and orange fields (C263-dissimilar) in the schematic soil profile of excavation trench M08. Besides C232 (ash from a small hearth) and C137 (burned clay of a hearth), the other C263-dissimilar samples are related to different soil fills - mostly from the higher part of the stratification.
This picture is confirmed grosso modo if the correlation coefficients of r-values (eq. 1) for the abundances of all OTUs with more than 10,000 reads (in total) are regarded. The filling material samples C111, C119, C150, C282, C298, C299 and C300 (grey labelled in Figure 9c) show no correlation-coefficients above 0.7, neither with each other nor with any other sample. In contrast, the group of C263-similar samples (blue labelled in Figure 9a and 9b) form groups of correlated samples with correlation coefficients around 0.7 and higher (colour-labelled in Figure 9c). Thus samples C295 and C297, C165 and C296, C162 and C124, C190 and C147 as well as C 177 and C231 represent pairs of samples correlated by highly abundant OTUs. In addition, C169, C218, C223, C286 and C288 form a group of correlated samples. Obviously these correlations are less related to the direct local situation of sampling sites, but seem to indicate similarities in the character of the soil material deposits in the related pairs and in the sample group.
Besides the general similarities and differences of samples characterized by PCAs and by similarity to mean values and to the reference sample, OTUs appearing exclusively or preferably in one or a few samples carry additional information about the past of soils and sampling sites. Taking the specific lower abundant OTUs and the OTUs of FPPs, it seems to be possible to speculate about specific environments in single sampling sites which are reflected by these bacteria. A possible interpretation based on reported ecological properties of involved OTUs is presented in Figure 10. This hypothetic picture shows similarities and differences which probably result from specific situations in each horizon and each sampling place. Thus, most hearths or fire places as well as the surfaces of streets are significantly different from simple soil fills or fills underneath street surfaces.
It seems remarkable that literature-reported environment-related features of OTUs in the FPPs (Section 3.3.1.) match well the character of low-abundant sample-specific OTUs (see Section 3.2.2.) in several cases. This concerns, for example, the hints to a mixture of manure and oil for the street surfaces of the 2nd half of the first and the 2nd half of the second century (samples C288 and C162) by an OTU profile which is significantly different from the layer of sample C165 lying in between. Other examples are the similar profiles of C295 and C297 which are separated by the differently appearing layer of C296 or the court fill C111 and C124, taken from the fill inside a building, which are marked by lower abundant bacteria indicating a combination of pig and bovine manure, starch, oil and nitrate. This matches nicely with the archaeological interpretation of the structure as a stable.
A synoptic consideration of the results discussed above suggests a hypothesis on the possible interpretation of highly abundant and low abundant components of bacteria communities reflected in the 16S rRNA data: All bacteria compositions reflect differences in soil composition of single spots and horizons which are specifically influenced by ancient human activities. In this memory-effect, the highly abundant bacteria seem to reflect mostly differences in the soil material composition affecting the recent soil microbial activity. In contrast, the lower abundant OTUs are partially reflecting specific soil compositions and human impacts related to former soil microbial activities. These activities are related to specific material compositions and soil contaminations, too, but could date back into the past – probably back to specific bacterial activities in antiquity.
4. OTU Patterns of Single Samples
In this section, the sequencing results of the forty investigated single soil samples are discussed by the appearance of dominant OTUs on the one hand and exclusive OTUs on the other hand. It is assumed that the group of dominant OTUs might be related to antique human impact, but reflects mostly the recent ecological situation in the soil sample, which results from a superposition of ancient soil formation and deposition, recent processes in the soil and the development of soil situation in the intermediate time. In contrast, among the minor components of soil bacterial community OTUs are expected which are stronger related to the ancient ecological situation. With respect to the fact, that the soils of the single sampling spots of the whole excavation area experienced nearly the same fate since the late Roman Empire period it is supposed, that in particular types which are found exclusively or nearly exclusively in one sample can reflect the specific ecological situation in the phase before or during the deposition of soil material in the Antiquity. Thus, the list below includes OTUs with lower read numbers despite the fact that some remaining uncertainty has to be taken in mind.
C111; Hz D, first half 3rd century, soil fill in courtyard area: The soil bacterial community represented in the 16S rRNA data (SBC) is dominated by Nocardioides (17.8 %), Paenibacillus (5.1 %), Bacillus (4.7 %) and Burkholderiales TRA3-20 (4.4 %). Among a few exclusive strains, there were registered 27 reads of uncultivated Chlamydiaceae and 8 reads of Yuhusiella, a thermotolerant actinobacterium first isolated from a desert region in western China [89]. Four other genera (Oceaniovalibus, Gelidibacter, Proteinophilum and Limosilactobacillus) have been found (33 – 88 read) nearly exclusive in C111. Oceaniovalibus belongs to Alphaproteobacteria and was first isolated from surface sea water near Taiwan [90]. Gelidibacter was described as a seawater bacterium, too [91]. Proteiniphilum is a strictly anaerobic proteinolytic bacterium found in the sludge of brewery waste water [92]. Limosilactobacillus is a thermophilic bacterium. Strains of these genera have been isolated, for example, from human urine [93]. The appearance of these OTUs could be interpreted as a signature related to an environment with enhanced salt content and protein-rich wastes.
C119: Hz D, soil fill under street surface, shortly after 200 AD.: Pseudonocardia is with more than 15% of all reads the clearly dominant genus of the bacterial community. The sample supplied five exclusive OTUs with considerable read numbers between 110 and 173. Among them are Pseudoglutamicibacter, the carotinoid dye-producing Bizionia and Saccharofermentans, an anaerobic bacterium first isolated from brewery wastewater [94].
C124: Hz D, clayey fill inside building, deposited around 200 AD.: The bacterial community is dominated by Clostridium sensu stricto 5 (8,7%) and Bacillus (4,3%). Remarkably, the sample supplied 10 exclusive or nearly exclusive OTUs with more than 10 reads, among them Dethiosulfatibacter, Providencia, Meiothermus, Myroides, Nitrosococcus, Paenalcaligenes and Alkalibacter. Dethiosulfatibacter is a sulfate and thiosulfate-reducing organism found in coastal marine sediment [95]. Meiothermus is known as thermophilic bacterium. Several strains have been isolated from hot springs [96]. Alkalibacter is a anaerobic, alkaliphilic carbohydrates decomposing bacterium [97]. With respect to the fact that the pH value and the electrical conductivity in the recent soil material are similar to the values of other samples, it could be assumed that the reads of these bacteria hint to a nutrient-rich and possibly alkaline and salt-containing environment in the ancient situation.
C137: Hz E, hearth with burned soil and clay, 2nd half of 2nd century: The sample is strongly dominated by Acinetobacter (49%). 15 specific OTUs with more than 10 reads have been found in this sample. This set includes the sulphate-reducing genera Desulfobulbus [98] and the pigment-producing genera Janthionbacterium [99] and Vogesella [100]. Other OTUs as Peptoniphilus, Haemophilus and Treponema are known for metabolization of amino acids and proteins or are pathogens for plants or animals [101].
C139: Hz E, hearth, 2nd half 2nd century: The bacterial community is very strongly dominated by Nocardioides (31%). Exclusively found OTUs are Sporomusa, Granulicatella and Thermodesulfovibrio. Species of the genus Granulicatella are known for metabolizing different amines and alkohols [102]. Thermodesulfovibrio is a sulphat-reducing thermophilic bacterium [103].
C147: Hz E, hearth, red burned soil and clay from hearth floor, 2nd half 2nd century: Nocardioides (19,5 %) is the dominating OTU. Only three specific OTUs have been observed in the sample. One of them, Garicola was first isolated from a highly salted fermented shrimp food [104] and could indicate residues of salt and fish – possibly in the context of garum, a fish sauce which was very popular in antiquity.
C150: Hz E, soil fill in a courtyard, 2nd half of 2nd century: The bacterial community of this sample is strongly dominated by Acinetobacter (25.5%). In addition, Ralstonia (12.4%) and Aquabacterium (10.7%) are also highly abundant. In contrast, some OTUs frequently highly abundant in most other samples as Nocardioides, Myxococcota, Bacteriap25, Burkholderia TRA3-20 and Chloroflexi KD4-96 show only low abundances in C150. With 0.4% Bacillus shows also a comparatively low presence. The ratio of reads to OTUs with ten or more reads (544) is the highest of all samples, which means that the sample shows a comparatively low diversity (the average of this ratio over all samples is 234). Only two specific OTUs have been found. One of them is Kinneretia, a strictly aerobic bacterium originally isolated from a fresh water lake [105].
C162: Hz E, Street, gravel fill and street surface, 2nd half of 2nd century: The bacterial community is dominated by Pseudonocardia (11%). The soil material supplied 13 specific OTUs, among them Flexilina, Polycladomyces, Actinotalea und Cloacibacterium. These OTUs seem to be an echo of a soil situation with a high load of seawage or faeces, organic waste as starch, and enhanced ion concentration. Flexilinea was isolated from methanogenic sludge from treating wastewater with a high starch-content [106] Polycladomyces is thermophilic bacterium firstly isolated from sea water [107]. Actinotalea was isolated from seawater, too [108]. Cloacibacterium is an anaerobic bacterium isolated from seawage [109].
C165: Hz E, Road substructure (excluding the surface of use) directly below C162, 2nd half of 2nd century: The sample is marked by Nocardioides (28.5%) and Pseudonocardia (20.6%) as strongly dominating genera. In strong contrast to the street horizon directly above (C162), two specific OTUs with more than 10 reads – Acidicapsa and Asanoa - were found, only. This observation agrees with the fact that the total number of reads between sample C162 and C165 are similar (about 35,000) whereas the number of types is rather different (308 in sample C162, but only 157 different OTUs in C165). Obviously, the different sizes of the spectra of “sample-specific” types indicate a strong difference between the character of the fill directly underneath the street and the street surface itself in the ancient situation.
C169: Hz F, Clayey fill in a courtyard, deposited around 120 AD.: The bacterial community is dominated by Nocardioides (8 %), but there is a comparatively high abundance of Zixibacteria (4.1 %), too. In contrast to other samples, the analysis supplied more than 20 exclusive OTUs with less than 10 reads, but no exclusive types with ten reads or more.
C172: Hz F, clayey fill above a hearth, deposed after the hearth’s abandonment around 120 AD.: The bacterial community is dominated by Aeromicrobium (12.6 %). In addition Bacillus (6.1 %) and Nocardioides (5.1 %) are highly abundant, too. Sample C172 was taken in the vicinity of the hearth-related samples C137, C147, and C178. It was marked by three specific OTUs, Thermosediminibacter, Glaciihabitans and Saliterribacillus. Two of them are particularly interesting because they hint to formerly enhanced soil salinity. Thermosediminibacter is an anaerobic and thermophilic organism isolated from deep sea sediment [110]. Saliterribacillus was first described as a halophilic bacterium isolated from a hypersaline lake [111]. The mentioned bacteria might reflect the nearness of hearth and enhanced salt concentration by ashes.
C177: Hz F, residues of a cupola oven and its destruction debris, around 120 AD.: As C172 it is dominated by Aeromicrobium (21.8 %). In addition, Nocardioides (6.5 %) and Pseudonocardia (5.5 %) are highly abundant, too. 11 reads of the Spirochaetae bacterium Leptospiraceae are the only hint of exclusivity of this sample. Species of this family are known as pathogens for mammals and man.
C178: Hz F, from the immediate vicinity of the cupola oven C 177, around 120 AD.: The bacterial community is dominated by Nitrososphaeraceae (9.1 %). 15 reads of Actinomarinales were observed, an OTU which is also present in sample C303, whereas clear exclusive OTUs are missing.
C190: Hz F: from street fill and street surface, about middle of the 2nd century AD.: The bacterial community is dominated by Nocardioides (10.4 %). The Crenarchaeota bacterium Cand. Nitrososphaera (4.9 %) and Myxococcota bacteriap25 (4.2 %) are highly abundant, too. The sample is marked by a few exclusive types, for example the two alphaproteobacteria Shinella and Ancylobacter. Shinella was isolated from anaerobic sludge [112]. Ancyclobacter is able to decompose kerosene and other hydrocarbons [113]. The last one might be interpreted to indicate an exposition to oil.
C218: Hz G: fill of a barbecue pit, around 120 AD.: Cand Nitrosphaera (6.3 %), Kryptonales BSV 26 (6.5 %) and Myxococcota bacterap25 (5.6%) are the three dominating OTUs. The 16S rRNA-reflected bacterial communities of samples C218 and C219 have a lot of similarities distinguishing them from all other samples. Besides these OTUs, there are some “sample-specific” OTUs, too. Erythrobacter is the only OTU of the barbecue pit sample C218 with more than 10 reads which appears neither in sample C219 nor in any other sample.
C219: Hz G: Coal sample from fill in barbecue pit (C218), around 120 AD.: Myxococcota bacteriap25 with 5.2 % and uncultivated Solimonadaceae are the dominating OTUs of the coal sample. This sample supplied eight “exclusive” OTUs with more than 10 reads among them Colwellia, a deep sea bacterium [114] and a strain of the sulphate-reducing order Desulfobacterales [115]. Both samples C218 and C219 contain the genus Desulfitibacterium which was first described by a strain from lake sediment and is able to metabolize chloraromates [116].
C223: Hz G, soil fill in courtyard, around 100 AD.: The bacterial community is dominated by Bacillus (9.4 %) and to a lower amount by Paenibacillus (4.8 %) and Cand. Nitrosphaera (4.4 %). Fluviicola and Oceanisphaera are specific OTUs. Fluviicula was found first in a freshwater environment [117]), whereas the halophilic bacterium Oceanisphaera was originally isolated from the sea bottom [118].
C231: Hz H, small hearth, red burned, fire-hardened hearth floor, 70−90 AD.: The bacterial community is dominated by Nocardioides (10.6 %) and Aeromicrobium (11.5 %). Besides 10 reads of the iron-oxidizing thermophilic and acidophilic bacterium Acidimicrobium, there are no further hints to specific OTUs. The features of iron oxidation and thermophilicity obviously match the characteristics of a fire place.
C232: Hz H, ash from hearth C231, 70−90 AD.: The bacterial community is strongly dominated by Lysinibacillus (25.1 %). In addition, Aeromicrobium (9.8 %) and Bacillus (13.3%) are also highly abundant genera. The ratio of reads to OTUs with ten or more reads (451) is the second highest of all samples, what means the sample shows a comparatively low diversity. The place shows two exclusive OTUs of Proteobacteria with 20 and 37 reads, resp.
C248: Trench M07, context 01, disturbed material, refill of the archaeological excavation of 1905: The sample is dominated by Nitrososphaeraceae (8.5 %), Bacillus (6.8 %) and Paenibacillus (5.6 %). The sample showed a comparatively high number of OTUs in total (783; 313 with ten or more reads) at a moderate total number of reads (43,777), which can be described as a high alpha-diversity. This sample shows the highest number of relations with other samples by signature-like patterns of lower abundant OTUs (see below). This observation can be interpreted as a strong indicator for the fact, that a considerable part of lower abundant OTUs reflects the local conditions in the ancient situation and conserved this information after mixing during the excavation of 1905. The sample supplied 37 exclusive OTUs indicated from sequencing with only one or a few reads. One OTU, only, (Cand. Abawacabacteria) is represented by 10 reads. Thus, this sample is marked by negligible exclusivity for OTUs with clear representation, but seems to show a high background noise of very low abundant types. This feature corresponds well to the above expressed assumption, that the sample reflects a mixing of different soils conserving important aspects of their ancient specific environmental information.
C263: Trench M07 context 06, fine grained, sandy alluvium underneath the deepest anthropogenic stratum, reference sample, no human interference: The bacterial community is dominated by Pseudonocardia (19.4 %) and Bacillus (17.8 %). Acinetobacter (8.5 %), Exiguobacterium (6 %) and Nocardioides (5.9 %) are also well represented, as well as Ralstonia (3.9 %) and Pseudomonas (2.9 %). It should be remarked here, that some other OTUs which are dominating or highly abundant in several other samples are comparatively low in C263. This concerns Aeromicrobium (0.05 %), Paenibacillus (0.8 %) and Myxococcota bacteriap 25 (0.5 %). C263 is marked by a comparatively low number of total reads (27,432) and OTUs (164; 101 with ten or more reads). Occallatibacter – a genus originally isolated from samples from the Namibian savannah and a river bank there [119] − is the only exclusively proved genus of C263, indicating that this sample is not distinguishable by special low abundant types from the other samples. Vice versa, this could be interpreted in the way to assuming that higher numbers of exclusively found OTUs in other samples indicate their specificity reflecting the ancient environmental situation and distinguish them from the marchsand.
C266: Trench M07 context 07, burnt debris from al large refuse pit, probably first half 2nd century: The bacterial community is strongly dominated by Aeromicrobium (22.2 %) and Limnobacter (16.3 %). The sample shows low indication of specificity: Desmospora was found by 54 reads and is present in C303 as the only other sample. Desmospora was first described from the sputum of a patient with suspected tuberculosis [120].
C282: Hz H, 70-90 AD.: soil fill directly underneath the hearth C232, 70−90 AD.: The bacterial community is dominated by Pseudomonas (9.2 %) and Nocardioides (6.4 %). The sample showed no exclusive OTUs, but it has to be remarked that the sequencing result for this sample can hardly be compared with the other samples because very low numbers of only 109 reads in total and 67 OTUs had been proved.
C283: Hz H, soil fill west of hearth (C232), 70−90 AD.: The bacterial community is dominated by Limnobacter (9.3 %), Nitrososphaeraceae (6.1 %), Paenibacillus (5.6 %) and uncultivated Anaerolineaceae (5.3 %). Persicitalea (11 reads) and Lachnospiraceae UCG-009 (13 reads) are present exclusively in this sample. Persicitalea is a aerobic organism and was first isolated from seawater [121].
C286: soil fill, Hz H, 70−90 AD.: The bacterial community is dominated by Nitrososphaeraceae (9 %) and Nocardioides (8.2 %). In addition, Bacillus (7.4 %) and Cand. Nitrosphaera (6.5 %) are also well represented. The sample supplied some exclusively found OTUs represented by less than ten reads, but showed no exclusive OTUs with ten reads or more.
C288: levelling fill underneath street surface layer, Hz, H, 70-90 AD.: The bacterial community is dominated by Nocardioides (6.8 %), Cand. Nitrosphaera (6.7 %) and Paenibacillus (5.1 %). The sample supplied six exclusive or nearly exclusive OTUs, among them Kaistia (18 Reads), Ehrlichia (10 reads) and Belnapia (14 reads). Ehrlichia is known as intracellular parasite and pathogen for animals and humans [122]. Kaistia is an aerobic bacterium first found in a stream with industrial waste in Korea [123]. Belnapia was found in a biological soil crust and is an aerobic bacterium, too [124].
C295: Hz A, collapse horizon, partly disturbed by modern ploughing, early 2nd half 4th century: The bacterial community is moderately dominated by Nocardioides (6.5 %) and Bacillus (6.9 %). The sample supplied 37 exclusively found OTUs represented by less than ten reads, but showed no exclusive OTUs with ten reads or more. The high number of exclusive OTUs with very low read numbers is an interesting analogon to sample C248 and could have to do with the origin of the sample soil from a former mixing of different ancient soil components.
C296: Hz A, fill and ancient surface of use, 2nd half of 4th century: The bacterial community is moderately dominated by Nocardioides (7.8 %) and Bacillus (5.1 %), too. The sample is marked by four specific OTUs, among them the Gammaproteobacterium Herminiimonas and both Actinobacteria, Plantactinospora and Krasilnikoviella. The latter one was described from a mural painting in the stone chamber of a Japanese tomb tumulus from about 700 AD [125].
C297: Hz B, fill and surface of use in the courtyard, about 1st half of 4th century: The bacterial community is stronger dominated by Nocardioides (12%) and Bacillus (15%). C297 supplied no exclusive OTU with ten or more reads.
C298: Hz C, fill and surface of use in the building south of courtyard C297, about 2nd half of 3rd century: The bacterial community is dominated by Nocardioides (25.5 %). Bacillus (9.8 %) is the second highest abundant genus. In addition, Pseudoarthrobacter showed a high abundance (6.2 %). C298 supplied Rhodobacter HMB 11 as only exclusive OTU with ten or more reads.
C299: Hz C, fill and surface of use in the building south of the courtyard, about 2nd half of 3rd century: The bacterial community is very strongly dominated by Acinetobacter (29.3 %). Bacillus (10 %) is the second highest abundant genus, too. C299 supplied no exclusive OTU with ten or more reads.
C300: Hz C, fill and surface of use in the courtyard north of building C299, about 2nd half of 3rd century: The bacterial community is overwhelmingly dominated by Exiguobacteria (32 %) and Bacillus (16.4 %). C300 supplied no exclusive OTU with ten or more reads.
C303: Trench M06 context 06, Soil filland surface of use, post earthquake 360 AD.: The bacterial community is dominated by Aeromicrobium (9.7 %). The sample supplied no exclusive OTU with ten or more reads.
C306: trench M06 context 08, levelled rubble fill, post earthquake 360 AD.: The bacterial community is dominated by Nocardioides (7.3 %). In addition, Bacillus (5.8 %), Paenibacillus (4.1 %) and Aeromicrobium (4.4 %) are present in significant amounts as well. C306 supplied no exclusive OTU with ten or more reads.
C307: trench M06 context 10, earth fill beneath caementitium floor, probably 3rd century: The bacterial community is dominated by Bacillus (14.4 %). The hydrogen producing anaerobic bacterium Acetobacteroides [126] is the only exclusive OTU with more than ten reads present in this sample. Nearly as rarely documented in the samples presented here are Anaerococcus [127] and Micrococcus (13 reads in C139).
C315: trench M06 context 13, levelled rubble fill, post earthquake 360 AD.: The bacterial community is dominated by Aeromicrobium (7.4 %). The sample supplied no exclusive OTU with ten or more reads.
C325: trench M06 context 16, clayey surface of use, post earthquake 360 AD.: The bacterial community is dominated by Bacillus (8.8 %). The sample supplied two nearly exclusive OTUs: Pectinophilus group of Lachnospiraceae (35 reads) and Algoriphagus (60 reads). The latter was isolated from from sea ice and from saline lake cyanobacterial mats [128].
C330: trench M06 context 20, furnace floor surface in praefurnium of hypocaustum, around 360 AD.: The bacterial community is dominated by Nocardioides (6.2 %). C330 showed eight exclusive or nearly exclusive OTUs with ten or more reads, among them uncultured Arcobacteraceae (18 reads), Chitinophagaceae UTBCD1 (15 reads), Kordiimonas (14 reads), uncultured Proteinivoracales (13 reads), Cand. Neoehrlichia (13 reads), uncultured Cellvibrionaceae (12 reads), Roseiarcus (12 reads) and Porphyrobacter (10 reads). Kordiimonas was isolated from marine sediment and was found to be able to degrade polycyclic aromatic hydrocarbons [129]. Roseiarcus is a pigmented microaerophilic bacterium first isolated from a methanotrophic microorganismic consortium [130]. Its appearance would fit to conditions in residues of wood or other organic material and low oxygen content in the local environment inside the hypocaustum. Porphyrobacter was found in freshwater and is a bacteriochlorphyll synthezising microorganism [131].
C341: trench M06 context 24, lower furnace floor beneath C330, early 4th century: The bacterial community is dominated by (11.5 %). In addition, Nocardioides (8.2 %), Polycyclovorans (4 %) and Polaromonas (5.2 %) are particularly abundant. - Renibacterium, a sub-species of Actinobacteria, is nearly exclusive in sample C341. Renibacterium was first described as a microorganism causing kidney diseases in fish [132].
C347: trench M06 context 30, lowest furnace floor underneath C341, 3rd century: The bacterial community is dominated by Aeromicrobium (11.6 %) and Limnobacter (10 %). A strain of Vampirovibrionaceae was exclusively present in sample C347.
5. Archaeological Interpretation
Can the bacteria data presented above find any meanigful archaeological interpretation? Quite obviously, the answer is positive. First, it comes as a surprise that even after nearly two millennia of physical and chemical degrading there still is sufficient residual bacterial DNA present for a successful analysis. Exposition to precipitation or oxygen alone seems not to be a deciding factor for the preservation of ancient ground bacteria DNA, as layers very near to the recent surface and from depths of up to two meters below ground both yielded rich ancient bacteria signatures. Nor does, in this part of Carnuntum, spilling and deluting by ground moisture seem to be of much concern to ancient bacteria genomes.
Concerning specific bacteria species, Limosilactobacillus, a milk acid bacteria present in human or animal intestines , should not surprise in a courtyard (sample C 111 originates from the fill under its surface), into which animal and, probably, human execra will have found their way aplenty. The quite frequent presence of ammonia oxidizing Nitrosphaeraceae in samples from many contexts and horizons should probably be interpreted in a way that urine was a more or less ubiquitous component of the ancient environment at Carnuntum. The presence of Desmospora, which was first isolated from the sputum of a patient suspected to suffer from tuberculosis , should not surprise as tuberculosis must have been endemic in antiquity as is has been endemic all over the world until the advent of antibiotics. One should probably as well not disregard the possibility of bovine tuberculosis. Also not as a surprise comes the occurrence of Cloacibacterium on the surface of a road which was permanently trodden by animals (sample C 162) and will at least occasionally have been the final destination of the contents of night pots as well. What surprises more is the fact that the fill directly underneath this road surface yielded no trace of similar bacteria (sample C 165). Obviously the surface was compressed to such a density that feces and waste from its surface did not seep into the fill before being washed away by the occasional rain shower.
The most welcome surprise, whereas, is the definition of Garicola in the reddish burned clayey surface of a hearth in horizon E, dating to the mid 2nd cent. AD. This bacteria was first isolated from a Korean fermented fish dish. It is common knowledge that one of the most favorite seasonings ancient Romans delicated on was garum, a sauce produced from fermented fish. It must have tasted very much like modern Thai fish sauce and was traded in amphorae from production centers along the coast of the Mediterranean and Adriatic Seas to the Rhine- and Danube provinces and was ubiquitous in Roman cooking. So it should of course have happened more than once that either some garum was spilled while being added to a dish simmering on the fire, or that a cooking pot was damaged and its contets, alreadey seasoned with the sauce, spilled over the hearth.. Nevertheless, this likely proof of garum in the residues of an ancient hearth at Carnuntum is, to the authors’ knowledge, the first one achieved through DNA analysis.
The attempt presented here leads to some sample-specific interpretations. For this, several examples were given above. One opportunity presenting itself is the comparison of different horizons at one sampling spot suggesting that an ancient translocation of soil material is reflected by bacterial composition within these horizons. Thus, sample C296 (horizon A) shows relations to other samples of upper horizons. It is located between C295 and C297 (from horizons A and B, both dating to the 4th century), but shows significant differences to both these samples, which have features related to lower layers (horizons F – H, second half of 1st and 2nd century). This may be interpreted as evidence for ancient translocation of material from lower layers into the fill material of C295 and C297. This observation matches the description of C295 as a collapse horizon and a possible material mixing by the plough. It might suggest that the fill material of C295 was also – at least partially – originating from deeper layers. The foundation trench of Wall 36 of horizon B was dug through strata C and underlying right into stratum F, and a well which was dug into horizon B cut through the complete stratification of the area, through the underlying geology and obviously deep enough to reach the ground water table. Both ancient excavation activities will have resulted in the deposition of soil material from all earlier strata present in the building and the courtyard of horizon B and explain the phenomenon just noted.
6. Conclusions
The analysis of sequencing data and the comparison between the samples offers the possibility to interpret the history of soil material deposited or translocated in the past. Whereas the comparison of phyla abundances and highly abundant OTUs supply a raw picture of sample groups, small sets of low abundant OTUs – mostly with read numbers between about 10 and 100 for the single samples – obviously reflect the ancient situation in more detail. The differences in the distribution of dominant as well as lower abundant OTUs speak for the fact that the formerly human impact on soil can lead to an important increase in Beta diversity of soil bacterial communities.
In result, it was possible to show, that the samples are distinguished by majority (highly abundant) as well as by minority and by exclusive bacterial types. A Principle Component Analysis (PCA) of samples showed that a certain clustering in sample groups can be observed for the highest abundant part as well as for lower abundant bacteria part of bacterial communities, whereby these clusters are not identical for the both abundance groups. Thus, it can be hypothesized that highly and lower abundant bacteria carry different information about the formerly human impact on the recent bacterial community. It is assumed that the higher abundant bacteria reflect changes in soil by specific human impacts in the past which are preserved in the recent state of soil up to now. In contrast, at least a part of the lower abundant bacteria – possibly present in dormant states, only – reports about formerly local states of soil which are reflected, by recently by some minority components of bacterial communities, only.
Whereas the PCAs give a raw relation between sample groups by inclusion of a large set of bacterial types (OTUs), it was decided to look for fingerprint-like abundance patterns for identifying specific relations between sample pairs and small groups of samples. This approach was impressively confirmed by a rather large set of OTUs preferentially present in the barbecue site-related samples C218 and C219 (Figure 6). Other interesting examples are the relations between two street horizons (C162 and C288) and between the horizons C295 and C295 which are separated from each other by a less-related intermediate layer (Figure 7). Further examples given in the supplementary part support the concept of reconstruction of sample relations by searching for fingerprints in abundance patterns.
In general, it can be said that the study of soil bacteria communities by 16S rRNA data can contribute to the investigation of archaeological sites and to deeper insights into local environmental situations related to different horizons. Besides the interest to describing historical and prehistorical ecological situations, this type of analyses can promote a better understanding of relations between soil-related human impacts on cultural-embossed ecosystem and soil microbial biodiversity in the present and in the past. Thus, such investigations contribute to closing the recently identified research gap between soil-related cultural ecosystem services and soil microbial biodiversity [132].
On the one hand the NGS-based analyses allow to identifying specific characteristic similarities between soil samples. On the other hand, it promotes interpretation of earlier environmental conditions by known metabolic features and chemical tolerances found by recent investigations of related taxonomical units – here mainly on the genus level. It might even be rewarding to search sites of specific ancient human impact for unknown microorganisms and new ecological functions and metabolic features which could in future become interesting for biotechnological applications.
It has to be remarked here, that the 16S rRNA used in this study is well suited to establish a raw taxonomic profile of bacterial communities, but that this represents only a very small section of the whole bacterial genome present. In future, wider DNA investigations, metagenomics studies, functional genomics and full genome sequencings of mixed bacterial populations using advanced bioinformatics methods [133] will most probably be applied as a very powerful and universal toolbox. It will supply much more data and will undoubtedly enable archaeologists and archaeo-ecologists to draw far-reaching conclusions on historical ecological situations at different excavation sites and horizons which will have a high impact on the understanding of human activities and ancient living conditions and their importance for the present.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. The following statements should be used “Conceptualization, J.M.K. and J.C.; methodology, J.M.K.; validation, X.X., P.M.G..; formal analysis, P.M.G., J.M.K.; investigation, A.K., J.M.K, L.E..; data curation, P.M.G..; writing—original draft preparation, J.M.K., and A.K..; writing—review and editing J.M.K., A.K. J.C., L.E. ; visualization, J.M.K..; supervision, J.M.K..; All authors have read and agreed to the published version of the manuscript.” Please turn to the CRediT taxonomy for the term explanation. Authorship must be limited to those who have contributed substantially to the work reported.
Funding
This research was partially funded by Landesgraduiertenförderung Thüringen. L.E. is gratefully for a scholarship of Landesgraduiertenförderung.
Institutional Review Board Statement
not applicable
Informed Consent Statement
not applicable.
Data Availability Statement
All data are stored by TU Ilmenau
Acknowledgments
The support of S. Schneider in data conversion and handling is gratefully acknowledged. We thank N. Wolf for support in the preparation of manuscript. L.E. thanks for a grant of Thüringer LandesgraduiertenförderungReferences
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Warinner, Ch.; Herbig, A.; Mann, A.; Yates, J.A.F.; Weiß, C.L.; Burbano, H.A.; Orlando, L.; Krause, J. A robust framework for microbial archaeology. Ann. Rev. Genom. Hum. Genet. 2017, 18, 321–356. [Google Scholar] [CrossRef] [PubMed]
- Philips, A.; Stolarek, I.; Kuczkowska, B.; et al. Comprehensive analysis of microorganisms accompanying human archaeological remains. GigaScience 2017, 6, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Horsburgh, K.A.; Gosling, A.L.; Cochrane, E.E.; Kirch, P.V.; Swift, J.A.; McCoy, M.D. Origins of Polynesian pigs revealed by mitochrondrial whole genome ancient DNA. Animals 2022, 12, 2469. [Google Scholar] [CrossRef] [PubMed]
- Chernysheva, E.; Korobov, D.; Borisov, A. Thermophilic microorganisms in arable land around medieval archaeological sites in Northern Caucasus, Russia: Novel evidence of past manuring practices. Geoarchaeol. Intern. J. 2017, 32, 494–501. [Google Scholar] [CrossRef]
- Mallory, J.; Dybo, A.; Balanovsky, O. The impact of genetic research on archaeology and linguistics in Eurasia. Russian J. Genetics 2019, 55, 1472–1487. [Google Scholar] [CrossRef]
- Jones, M.; Brown, T. Agricultural origins: the evidence of modern and ancient DNA. Holocene 2000, 10, 769–776. [Google Scholar] [CrossRef]
- Sabin, S.; Yeh, H.Y.; Pluskowski, A.; Clamer, C.; Mitchell, P.D. Bos, K.I. Estimating molecular preservation of the intestinal microbiome via metagenomics analyses of latrine sediments from two medieval cities. Phil. Trans. Royal Soc. B 2020, 375, 20190576. [Google Scholar] [CrossRef] [PubMed]
- Warinner, C. An archaeology of microbes. J. Anthropol. Res. 2022. [CrossRef]
- Romanowicz, K.J.; Freedman, Z.B.; Upchurch, R.A.; Argiroff, W.A.; Zak, D.R. Active microorganisms in forest soils differ from total community yet are shaped by the same environmental factors: the influence of pH and soil moisture. FEMS Microbiol. Ecol. 2016, 92, 149. [Google Scholar] [CrossRef]
- Wolinska, A.; Wlodarczyk, K.; Kuzniar, A.; Marzec-Grzadziel, A.; Grzadziel, J.; Galazka, A.; Uzarowicz, L. Soil microbial community profiling and bacterial metabolic activity of technosols as an effect of soil properties following land reclamation: a case study from abandoned iron sulphide and uranium mine in Rudki (south-central Poland). Agronomy 2020, 10, 1795. [Google Scholar] [CrossRef]
- Torsvik, V.; Sorheim, R.; Gokskoyr, J. Total bacterial diversity in soil and sediment – a review. J. Indust. Microbiol. 1996, 17, 170–178. [Google Scholar] [CrossRef]
- Schmid, M.W.; VanMoorsel, S.J.; Hahkl, T.; DeLuca, E.; DeDeyn, G.B.; Wagg, C.; Niklaus, P.A.; Schmid, B. Effects of plant community history, soil legacy and plant diversity on soil microbial communities. J. Ecology 2021, 109, 3007–3023. [Google Scholar] [CrossRef]
- Jones, S.E.; Lennon, J.T. Dormancy contributes to the maintenance of microbial diversity. PNAS 2010, 107, 5881–5886. [Google Scholar] [CrossRef] [PubMed]
- Aanderud, Z.T.; Jones, S.E.; Fierer, N. 2015. Resuscitation of the rare biosphere contributes to pulses of ecosystem activity. Frontiers in Microbiology 2015, 6, 24. [Google Scholar] [CrossRef]
- C. E Wegner, W. Liesack. Unexpected dominance of elusive acidobacteria in early industrial soft coal slags. Frontiers Microbiol. 2017, 8, 1023. [Google Scholar]
- Singer, D.; Herndon, E.; Zemanek, L.; Kortney, C.; Sander, T.; Senko, J.; Perdrial, N. Biogeochemical controls on the potential for long-term contaminated leaching from soils developing on historical coal mine spoil. Soil Syst. 2021, 5, 3. [Google Scholar] [CrossRef]
- Margesin, R.; Siles, J.A.; Cajthaml, T.; Ohlinger, B.; Kistler, E. Microbiology meets archaeology: soil microbial communities reveal different human activities at archaic Monte Iato (Sixth century BC). Microbial Ecol. 2017, 73, 925–938. [Google Scholar] [CrossRef]
- Köhler, J.M; Kalensee, F.; Günther, P.M.; Schüler, T.; Cao, J. The local ecological memory of soil: majority and minority components of bacterial communities in prehistoric urns from Schöps (Germany). Int. J. Environ. Res. 2018, 12, 575–684. [Google Scholar] [CrossRef]
- Köhler, J.M.; Beetz, N.; Günther, P.M.; Möller, F.; Schüler, T.; Cao, J. Microbial community types and signature-like soil bacterial patterns from fortified prehistoric hills of Thuringia (Germany). Community Ecology 2020, 21, 107–120. [Google Scholar] [CrossRef]
- Köhler, J.M.; Kalensee, F.; Cao, J.; Günther, P.M. Hadesarchaea and other extremophile bacteria from ancient mining areas of the East Harz region (Germany) suggest an ecological long-term memory of soil SN Appl. Sci. 2019, 1, 839. [Google Scholar] [CrossRef]
- J.M. Köhler, L. Ehrhardt, J. Cao, F. Möller, T. Schüler, P.M. Günther. Beta-Diversity Enhancement by archaeological structures: bacterial communities of an historical tannery area of the city of Jena (Germany) reflected the ancient human impact. Ecologies 2023, 4, 325–343. [Google Scholar] [CrossRef]
- Marfennia, O.E.; Ivanova, A.E.; Sacharov, D.S. The mycologicl properties of medieval cultur layers as a form of ‘soil biological memory’ about urbanization. J. Soils Sediments 2008, 8, 340–348. [Google Scholar] [CrossRef]
- Dergacheva, M. Ecological function of soil humus. Eurasian Soil Sci. 2001, 34, 100–105. [Google Scholar]
- Khalighi, M; Gonze, D.; Faust, K.; Sommeria-Klein, G.; Lathi, L. Quantifiying the impact of ecological memory on the dynamics of interacting communities. PLOS Comput. Biol. 2022, 1009396.
- Benito, B.M.; Gil-Romera, G.; Birks, H.J.B. Ecological memory at millennial time-scales: the importance of data constraints, species longevity and niche features. Ecography 2019, 43, 04772. [Google Scholar] [CrossRef]
- Gonze, D.; Coyte, K.; Lahti, L.; Faust, K. Microbial communities as dynamical system. Curr. Opin. Microbiol. 2018, 44, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Ogle, K.; Barber, J.J.; Baron-Gafford, G.A.; Benthley, L.P.; Young, J.; Huxman, T.E.; Loik, M.E.; Tissue, D.T. Quantifying ecological memory in plant and ecosystem processes. Ecol. Lett. 2015, 18, 221–235. [Google Scholar] [CrossRef]
- Canarini, A.; Schmidt, H.; Fuchslueger, L.; Martin, V.; Herbold, C.W.; Zezula, D.; Gundler. Ph.; Hasibeder, R.; Jecmenia, M.; Bahn, M. et al. Ecological memory of recurrent drought modifies soil processes via changes in soil micribial community. Nat. Commun. 2021, 12, 25675–4. [Google Scholar] [CrossRef]
- Zvyaintsev, D.G.; Dobrovolskaya, T.G.; Babeya, I.P.; Zenova, G.M.; Lysak, L.V.; Marfennia, O.Y. Role of microorganisms in the biogeocenotic functions of soil. Eurasian Soil Sci. 1992, 24, 30–46. [Google Scholar]
- Borisov, A.V.; Demkina, T.S.; Kashirskaya, N.N.; Khomutova, T.E.; Chernysheva, E.V. Changes in the past soil-forming conditions and human activity in soil biological memory: microbial and enzyme components. Eurasian Soil Sci. 2021, 54, 1078–1088. [Google Scholar] [CrossRef]
- Schaefer, V. Alien invasiens, eclogical restoration in cities and the loss of ecological memory. Restoration Ecology 2009, 17, 171–176. [Google Scholar] [CrossRef]
- Konecny, A.; Fuchshuber, N.; Pollhammer, E. Grabungen in den canabae legions von Carnuntum. Acta Carnuntina 2022, 12/2, 22 –41. [Google Scholar]
- Humer, F (ed.). Carnuntum. Wiedergeborene Stadt der Kaiser, Darmstadt 2014: Philip von Zabern.
- Gassner, V.; Jilek, S.; Ladstätter, S. Am Rande des Reichs. Die Römer in Österreich, Vienna: Ueberreuter 2002.
- Kandler, M; Humer, F. Carnuntum, in: Šašel Kos, M.; Scherrer, P. (eds.), The autonomous towns of Noricum and Pannonia II, Noricum,2004, Situla 42, Ljubljana: Narodni Muzej Slovenije,11−66.
- Gersbach, E. Ausgrabung heute. Methoden und Techniken der Feldgrabung, 3rd ed., 1999 Stuttgart,Theiss.
- Harris, E. Principles of Archaeological Stratigraphy, 2nd ed., London: Academic Press 1989.
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glockner, F.O. The SILVA ribosomal RNA gene database project: improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Klindworth, A.; Pruesse, E.; Schwee, T.; Peplies, J.; Quast, C.; Horn, M.; Glockner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, P.; Parfrey, L.-W.; Yarza, P.; Gerken, J.; Pruesse, E.; Quast, C.; Schweer, T.; Peplies, J.; Ludwig, W.; Glockner, F.O. The SILVA and “All-species Living Tree Project (LTP)” taxonomic frameworks. Nucleic Acid Res. 2014, 42, D643–D648. [Google Scholar] [CrossRef]
- Spring, S.; Kämpfer, P.; Schleifer, K. H. Limnobacter thiooxidans gen. nov., sp. nov., a novel thiosulfate-oxidizing bacterium isolated from freshwater lake sediment. Int. J. Syst. Evol. Microbiol. 2001, 51, 1463–1470. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Yang, J.; Wu, W.-M.; Zhao, J.; Song, Y.; Gao, L.; Yang, R.; Jiang, L. Biodegradation and Mineralization of Polystyrene by Plastic-Eating Mealworms: Part 2. Role of Gut Microorganisms. Environmental science & technology 2015, 49, 12087–12093. [Google Scholar]
- Irgens, R. L.; Gosink, J. J.; Staley, J. T. Polaromonas vacuolata gen. nov., sp. nov., a psychrophilic, marine, gas vacuolate bacterium from Antarctica. International journal of systematic bacteriology 1996, 46, 822–826. [Google Scholar] [CrossRef]
- Nguyen, T. M.; Kim, J. Limnobacter humi sp. nov., a thiosulfate-oxidizing, heterotrophic bacterium isolated from humus soil, and emended description of the genus Limnobacter Spring et al. 2001. Journal of microbiology (Seoul, Korea) 2017, 55, 508–513. [Google Scholar] [CrossRef] [PubMed]
- Eloe-Fadrosh, E. A.; Paez-Espino, D.; Jarett, J.; Dunfield, P. F.; Hedlund, B. P.; Dekas, A. E.; Grasby, S. E.; Brady, A. L.; Dong, H.; Briggs, B. R.; et al. Global metagenomic survey reveals a new bacterial candidate phylum in geothermal springs. Nature communications 2016, 7, 10476. [Google Scholar] [CrossRef]
- Kim, M. K.; Kim, Y.-J.; Cho, D.-H.; Yi, T.-H.; Soung, N.-K.; Yang, D.-C. Solimonas soli gen. nov., sp. nov., isolated from soil of a ginseng field. Int. J. Syst. Evol. Microbiol. 2007, 57, 2591–2594. [Google Scholar] [CrossRef] [PubMed]
- Baer, M. L.; Ravel, J.; Chun, J.; Hill, R. T.; Williams, H. N. A proposal for the reclassification of Bdellovibrio stolpii and Bdellovibrio starrii into a new genus, Bacteriovorax gen. nov. as Bacteriovorax stolpii comb. nov. and Bacteriovorax starrii comb. nov., respectively. Int. J. Syst. Evol. Microbiol. 2000, 50 Pt 1, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Jung, M.-Y.; Islam, M. A.; Gwak, J.-H.; Kim, J.-G.; Rhee, S.-K. Nitrosarchaeum koreense gen. nov., sp. nov., an aerobic and mesophilic, ammonia-oxidizing archaeon member of the phylum Thaumarchaeota isolated from agricultural soil. Int. J. Syst. Evol. Microbiol. 2018, 68, 3084–3095. [Google Scholar] [PubMed]
- Kang, J. Y.; Chun, J.; Seo, J.-W.; Kim, C. H.; Jahng, K. Y. Flaviaesturariibacter amylovorans gen. nov., sp. nov., a starch-hydrolysing bacterium, isolated from estuarine water. Int. J. Syst. Evol. Microbiol. 2015, 65, 2209–2214. [Google Scholar] [CrossRef] [PubMed]
- Nedashkovskaya, O. I.; Kim, S. B.; Suzuki, M.; Shevchenko, L. S.; Lee, M. S.; Lee, K. H.; Park, M. S.; Frolova, G. M.; Oh, H. W.; Bae, K. S.; et al. Pontibacter actiniarum gen. nov., sp. nov., a novel member of the phylum ‘Bacteroidetes’, and proposal of Reichenbachiella gen. nov. as a replacement for the illegitimate prokaryotic generic name Reichenbachia Nedashkovskaya et al. 2003. Int. J. Syst. Evol. Microbiol. 2005, 55, 2583–2588. [Google Scholar] [CrossRef] [PubMed]
- Tamer, A. U.; Aragno, M.; Sahin, N. Isolation and characterization of a new type of aerobic, oxalic acid utilizing bacteria, and proposal of Oxalicibacterium flavum gen. nov., sp. nov. Systematic and applied microbiology 2002, 25, 513–519. [Google Scholar] [CrossRef]
- Yoon, M.-H.; Im, W.T. Flavisolibacter ginsengterrae gen nov., sp. nov. And Flavisolibacter ginsengisoli sp. nov., isolated from ginseng cultivating soil. Int. J. Syst. Evol. Microbiol. 2007, 57, 1834–1839. [Google Scholar] [CrossRef]
- Hwang, W. M.; Ko, Y.; Kim, J.-H.; Kang, K. Ahniella affigens gen. nov., sp. nov., a gammaproteobacterium isolated from sandy soil near a stream. Int. J. Syst. Evol. Microbiol. 2018, 68, 2478–2484. [Google Scholar] [CrossRef]
- Jogler, M.; Chen, H.; Simon, J.; Rohde, M.; Busse, H.-J.; Klenk, H.-P.; Tindall, B. J.; Overmann, J. Description of Sphingorhabdus planktonica gen. nov., sp. nov. and reclassification of three related members of the genus Sphingopyxis in the genus Sphingorhabdus gen. nov. Int. J. Syst. Evol. Microbiol. 2013, 63, 1342–1349. [Google Scholar] [CrossRef]
- Balch, W. E.; Fox, G. E.; Magrum, L. J.; Woese, C. R.; Wolfe, R. S. Methanogens: reevaluation of a unique biological group. Microbiological reviews 1979, 43, 260–296. [Google Scholar] [CrossRef]
- Borsodi, A. K.; Aszalós, J. M.; Bihari, P.; Nagy, I.; Schumann, P.; Spröer, C.; Kovács, A. L.; Bóka, K.; Dobosy, P.; Óvári, M.; et al. Anaerobacillus alkaliphilus sp. nov., a novel alkaliphilic and moderately halophilic bacterium. Int. J. Syst. Evol. Microbiol. 2019, 69, 631–637. [Google Scholar] [CrossRef] [PubMed]
- Vasilyeva, L.V.; Semenov, A.M. Labrys monahos, a new buddng prosthecate bacterium with radial symmetry. Mikrobiologiya 1984, 53, 85–92. [Google Scholar]
- Greening, R. C.; Leedle, J. A. Enrichment and isolation of Acetitomaculum ruminis, gen. nov., sp. nov.: acetogenic bacteria from the bovine rumen. Archives of microbiology 1989, 151, 399–406. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Zhou, S. Sinibacillus soli gen. nov., sp. nov., a moderately thermotolerant member of the family Bacillaceae. Int. J. Syst. Evol. Microbiol. 2014, 64, 1647–1653. [Google Scholar] [CrossRef] [PubMed]
- Glaeser, S. P.; McInroy, J. A.; Busse, H.-J.; Kämpfer, P. Pseudogracilibacillus auburnensis gen. nov., sp. nov., isolated from the rhizosphere of Zea mays. Int. J. Syst. Evol. Microbiol. 2014, 64, 2442–2448. [Google Scholar] [CrossRef] [PubMed]
- Ntougias, S.; Fasseas, C.; Zervakis, G. I. Olivibacter sitiensis gen. nov., sp. nov., isolated from alkaline olive-oil mill wastes in the region of Sitia, Crete. Int. J. Syst. Evol. Microbiol. 2007, 57, 398–404. [Google Scholar] [CrossRef]
- Juteau, P.; Côté, V.; Duckett, M.-F.; Beaudet, R.; Lépine, F.; Villemur, R.; Bisaillon, J.-G. Cryptanaerobacter phenolicus gen. nov., sp. nov., an anaerobe that transforms phenol into benzoate via 4-hydroxybenzoate. Int. J. Syst. Evol. Microbiol. 2005, 55, 245–250. [Google Scholar] [CrossRef]
- Zhang, X.; Tu, B.; Dai, L.-R.; Lawson, P. A.; Zheng, Z.-Z.; Liu, L.-Y.; Deng, Y.; Zhang, H.; Cheng, L. Petroclostridium xylanilyticum gen. nov., sp. nov., a xylan-degrading bacterium isolated from an oilfield, and reclassification of clostridial cluster III members into four novel genera in a new Hungateiclostridiaceae fam. nov. Int. J. Syst. Evol. Microbiol. 2018, 68, 3197–3211. [Google Scholar] [CrossRef]
- Kämpfer, P.; Young, C.-C.; Arun, A. B.; Shen, F.-T.; Jäckel, U.; Rosselló-Mora, R.; Lai, W.-A.; Rekha, P. D. Pseudolabrys taiwanensis gen. nov., sp. nov., an alphaproteobacterium isolated from soil. Int. J. Syst. Evol. Microbiol. 2006, 56, 2469–2472. [Google Scholar] [CrossRef]
- Lv, X.-L.; Xie, B.-S.; Cai, M.; Tang, Y.-Q.; Wang, Y.-N.; Cui, H.-L.; Liu, X.-Y.; Tan, Y.; Wu, X.-L. Halodurantibacterium flavum gen. nov., sp. nov., a non-phototrophic bacterium isolated from an oil production mixture. Current microbiology 2015, 70, 141–148. [Google Scholar] [CrossRef]
- Inahashi, Y.; Matsumoto, A.; Danbara, H.; Ōmura, S.; Takahashi, Y. Phytohabitans suffuscus gen. nov., sp. nov., an actinomycete of the family Micromonosporaceae isolated from plant roots. Int. J. Syst. Evol. Microbiol. 2010, 60, 2652–2658. [Google Scholar] [CrossRef] [PubMed]
- Vandieken, V.; Niemann, H.; Engelen, B.; Cypionka, H. Marinisporobacter balticus gen. nov., sp. nov., Desulfosporosinus nitroreducens sp. nov. and Desulfosporosinus fructosivorans sp. nov., new spore-forming bacteria isolated from subsurface sediments of the Baltic Sea. Int. J. Syst. Evol. Microbiol. 2017, 67, 1887–1893. [Google Scholar] [CrossRef] [PubMed]
- Ventosa, A.; Marquez, M. C.; Ruiz-Berraquero, F.; Kocur, M. Salinicoccus roseus gen. nov., sp. nov., a new moderately halophilic gram-positive coccus. Syst. Appl. Microbiol. 1990, 13, 29–33. [Google Scholar] [CrossRef]
- Li, Q.; Rückert, C.; Li, G.; Huang, P.; Schneider, O.; Kalinowski, J.; Jiang, Y.; Zotchev, S. B.; Jiang, C. Prauserella flavalba sp. nov., a novel species of the genus Prauserella, isolated from alkaline soil. Int. J. Syst. Evol. Microbiol. 2020, 70, 380–387. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Kojima, H.; Fukui, M. Sulfuriferula multivorans gen. nov., sp. nov., isolated from a freshwater lake, reclassification of ‘Thiobacillus plumbophilus’ as Sulfuriferula plumbophilus sp. nov., and description of Sulfuricellaceae fam. nov. and Sulfuricellales ord. nov. Int. J. Syst. Evol. Microbiol. 2015, 65, 1504–1508. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; Hiraishi, A.; Yoshimi, Y.; Kawaharasaki, M.; Masuda, K.; Kamagata, Y. Microlunatus phosphovorus gen. nov., sp. nov., a new gram-positive polyphosphate-accumulating bacterium isolated from activated sludge. International journal of systematic bacteriology 1995, 45, 17–22. [Google Scholar] [CrossRef]
- Aizawa, T.; Ve, N. B.; Kimoto, K.-I.; Iwabuchi, N.; Sumida, H.; Hasegawa, I.; Sasaki, S.; Tamura, T.; Kudo, T.; Suzuki, K.-I.; et al. Curtobacterium ammoniigenes sp. nov., an ammonia-producing bacterium isolated from plants inhabiting acidic swamps in actual acid sulfate soil areas of Vietnam. Int. J. Syst. Evol. Microbiol. 2007, 57, 1447–1452. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Du, Y.; Wang, G. Knoellia flava sp. nov., isolated from pig manure. Int. J. Syst. Evol. Microbiol. 2012, 62, 384–389. [Google Scholar] [CrossRef]
- Molinero, N.; Conti, E.; Sánchez, B.; Walker, A. W.; Margolles, A.; Duncan, S. H.; Delgado, S. Ruminococcoides bili gen. nov., sp. nov., a bile-resistant bacterium from human bile with autolytic behavior. Int. J. Syst. Evol. Microbiol. 2021, 71. [Google Scholar] [CrossRef]
- Mitchell, M.; Nguyen, S. V.; Connor, M.; Fairley, D. J.; Donoghue, O.; Marshall, H.; Koolman, L.; McMullan, G.; Schaffer, K. E.; McGrath, J. W.; et al. Terrisporobacter hibernicus sp. nov., isolated from bovine faeces in Northern Ireland. Int. J. Syst. Evol. Microbiol. 2023, 73. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Guo, X.; Wang, Y.; He, M.; Ma, K.; Wang, H.; Chen, X.; Kong, D.; Yang, Z.; Ruan, Z. Terrisporobacter petrolearius sp. nov., isolated from an oilfield petroleum reservoir. Int. J. Syst. Evol. Microbiol. 2015, 65, 3522–3526. [Google Scholar] [CrossRef] [PubMed]
- Fortina, M. G.; Pukall, R.; Schumann, P.; Mora, D.; Parini, C.; Manachini, P. L.; Stackebrandt, E. Ureibacillus gen. nov., a new genus to accommodate Bacillus thermosphaericus (Andersson et al. 1995), emendation of Ureibacillus thermosphaericus and description of Ureibacillus terrenus sp. nov. Int. J. Syst. Evol. Microbiol. 2001, 51, 447–455. [Google Scholar] [CrossRef] [PubMed]
- Fraj, B.; Ben Hania, W.; Postec, A.; Hamdi, M.; Ollivier, B.; Fardeau, M.-L. Fonticella tunisiensis gen. nov., sp. nov., isolated from a hot spring. Int. J. Syst. Evol. Microbiol. 2013, 63, 1947–1950. [Google Scholar] [CrossRef] [PubMed]
- Koeck, D. E.; Ludwig, W.; Wanner, G.; Zverlov, V. V.; Liebl, W.; Schwarz, W. H. Herbinix hemicellulosilytica gen. nov., sp. nov., a thermophilic cellulose-degrading bacterium isolated from a thermophilic biogas reactor. Int. J. Syst. Evol. Microbiol. 2015, 65, 2365–2371. [Google Scholar] [CrossRef] [PubMed]
- Antunes, A.; Rainey, F. A.; Wanner, G.; Taborda, M.; Pätzold, J.; Nobre, M. F.; da Costa, M. S.; Huber, R. A new lineage of halophilic, wall-less, contractile bacteria from a brine-filled deep of the Red Sea. Journal of bacteriology 2008, 190, 3580–3587. [Google Scholar] [CrossRef] [PubMed]
- Kalyuzhnaya, M. G.; Bowerman, S.; Lara, J. C.; Lidstrom, M. E.; Chistoserdova, L. Methylotenera mobilis gen. nov., sp. nov., an obligately methylamine-utilizing bacterium within the family Methylophilaceae. Int. J. Syst. Evol. Microbiol. 2006, 56, 2819–2823. [Google Scholar] [CrossRef]
- Mu, Y.; Zhou, L.; Zeng, X.-C.; Liu, L.; Pan, Y.; Chen, X.; Wang, J.; Li, S.; Li, W.-J.; Wang, Y. Arsenicitalea aurantiaca gen. nov., sp. nov., a new member of the family Hyphomicrobiaceae, isolated from high-arsenic sediment. Int. J. Syst. Evol. Microbiol. 2016, 66, 5478–5484. [Google Scholar] [CrossRef]
- M.V. Simankova, M.V.; Chernych, N.A.; Osipov, G.A.; Zavarzin, G.A. Halocella cellulolytica gen. nov., sp. nov., a New Obligately Anaerobic, Halophilic, Cellulolytic bacterium Systematic and Applied Microbiology. 1993, 16, 385–389. [Google Scholar]
- Mechichi, T.; Stackebrandt, E.; Fuchs, G. Alicycliphilus denitrificans gen. nov., sp. nov., a cyclohexanol-degrading, nitrate-reducing beta-proteobacterium. Int. J. Syst. Evol. Microbiol. 2003, 53, 147–152. [Google Scholar] [CrossRef]
- 8Watanabe, M.; Kojima, H.; Fukui, M. Limnochorda pilosa gen. nov., sp. nov., a moderately thermophilic, facultatively anaerobic, pleomorphic bacterium and proposal of Limnochordaceae fam. nov., Limnochordales ord. nov. and Limnochordia classis nov. in the phylum Firmicutes. Int. J. Syst. Evol. Microbiol. 2015, 65, 2378–2384. [Google Scholar] [CrossRef]
- Hatayama, K.; Shoun, H.; Ueda, Y.; Nakamura, A. Planifilum fimeticola gen. nov., sp. nov. and Planifilum fulgidum sp. nov., novel members of the family ‘Thermoactinomycetaceae’ isolated from compost. Int. J. Syst. Evol. Microbiol. 2005, 55, 2101–2104. [Google Scholar] [CrossRef] [PubMed]
- Willems, A.; Busse, J.; Goor, M.; Pot, B.; Falsen, E.; Jantzen, E.; Hoste, B. ; Gillis,, M.;Kersters, K.; Auling, G.; De Ley, J. Hydrogenophaga, a New Genus of Hydrogen-Oxidizing Bacteria That Includes Hydrogenophaga flava comb. nov. (Formerly Pseudomonas flava), Hydrogenophaga palleronii (Formerly Pseudomonas palleronii), Hydrogenophaga pseudoflava (Formerly Pseudomonas pseudoflava and “Pseudomonas carboxy do flava”), and Hydrogenophaga taeniospiralis (Formerly Pseudomonas taeniospiralis). Int. J. Syst. Evol. Microbiol. 1989, 39, 319–332. [Google Scholar]
- Castelle, C.J.; Hug, L.A.; Wrighton, K.C.; Thomas, B.C.; Williams, K.H.; Wu, D.; Tringe, S.G.; Singer, S.W.; Eisen, J.A.; Banfield, J.F. Extraordinary phylogenetic diversity and metabolic versatility in aquifer sediment. Nat. Commun. 2013, 4, 2120. [Google Scholar] [CrossRef] [PubMed]
- Mao, J.; Wang, J.; Dai, H.-Q.; Zhang, Z.-D.; Tang, Q.-Y.; Ren, B.; Yang, N.; Goodfellow, M.; Zhang, L.-X.; Liu, Z.-H. Yuhushiella deserti gen. nov., sp. nov., a new member of the suborder Pseudonocardineae. Int. J. Syst. Evol. Microbiol. 2011, 61, 621–630. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Zong, R.; Li, Q.; Fu, Y.; Xu, Y.; Wang, Y.; Jiao, N. Oceaniovalibus guishaninsula gen. nov., sp. nov., a marine bacterium of the family Rhodobacteraceae. Current microbiology 2012, 64, 385–391. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.-C.; Margesin, R. Gelidibacter sediminis sp. nov., isolated from a sediment sample of the Yellow Sea. Int. J. Syst. Evol. Microbiol. 2015, 65, 2304–2309. [Google Scholar] [CrossRef]
- Chen, S.; Dong, X. Proteiniphilum acetatigenes gen. nov., sp. nov., from a UASB reactor treating brewery wastewater. Int. J. Syst. Evol. Microbiol. 2005, 55, 2257–2261. [Google Scholar] [CrossRef]
- Ksiezarek, M.; Ribeiro, T. G.; Rocha, J.; Grosso, F.; Perovic, S. U.; Peixe, L. Limosilactobacillus urinaemulieris sp. nov. and Limosilactobacillus portuensis sp. nov. isolated from urine of healthy women. Int. J. Syst. Evol. Microbiol. 2019, 71. [Google Scholar] [CrossRef]
- Chen, S.; Niu, L.; Zhang, Y. Saccharofermentans acetigenes gen. nov., sp. nov., an anaerobic bacterium isolated from sludge treating brewery wastewater. Int. J. Syst. Evol. Microbiol. 2010, 60, 2735–2738. [Google Scholar] [CrossRef]
- Takii, S.; Hanada, S.; Tamaki, H.; Ueno, Y.; Sekiguchi, Y.; Ibe, A.; Matsuura, K. Dethiosulfatibacter aminovorans gen. nov., sp. nov., a novel thiosulfate-reducing bacterium isolated from coastal marine sediment via sulfate-reducing enrichment with Casamino acids. Int. J. Syst. Evol. Microbiol. 2007, 57, 2320–2326. [Google Scholar] [CrossRef]
- Pires, A. L.; Albuquerque, L.; Tiago, I.; Nobre, M. F.; Empadinhas, N.; Veríssimo, A.; da Costa, M. S. Meiothermus timidus sp. nov., a new slightly thermophilic yellow-pigmented species. FEMS microbiology letters 2005, 245, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Garnova, E.S.; Zhilina, T.N.; Tourova, T.P.; Kostrikina, N.A.; Zavarzin, G.A. Anaerobic, alkaliphilic, saccharolytic bacterium Alkalibacter saccharofermentans gen. nov., sp. nov. from a soda lake in the Transbaikal region of Russia. Extremophiles: life under extreme conditions. 2004, 8, 309–316. [Google Scholar] [PubMed]
- Suzuki, D.; Ueki, A.; Katsuji, A.; Ueki, K. Desulfobulbus japonicus sp. nov., a novel Gram-negative propionate-oxidizing, sulfate-reducing bacterium isolated from an estuarine sediment in Japan. Int J Syst Evol Microbiol. 2007, 57(Issue 4). [Google Scholar] [CrossRef]
- De Ley, J.; Segers, P.; Gillis, M. Intra- and intergeneric similarities of Chromobacterium and Janthinobacterium ribosomal ribonucleic acid cistrons. International Journal of Systematic Bacteriology 1978, 28, 154–168. [Google Scholar] [CrossRef]
- Grimes, D. J.; Woese, C. R.; MacDonell, M. T.; Colwell, R. R. Systematic study of the genus Vogesella gen. nov. and its type species, Vogesella indigofera comb. nov. International journal of systematic bacteriology 1997, 47, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Mang, S. M.; Frisullo, S.; Elshafie, H. S.; Camele, I. Diversity Evaluation of Xylella fastidiosa from Infected Olive Trees in Apulia (Southern Italy). The Plant Pathology Journal 2016, 32, 102–111. [Google Scholar] [CrossRef]
- Collins, M. D.; Lawson, P. A. The genus Abiotrophia (Kawamura et al.) is not monophyletic: proposal of Granulicatella gen. nov., Granulicatella adiacens comb. nov., Granulicatella elegans comb. nov. and Granulicatella balaenopterae comb. nov. Int. J. Syst. Evol. Microbiol. 2000, 50, 365–369. [Google Scholar] [CrossRef]
- Henry, E. A.; Devereux, R.; Maki, J. S.; Gilmour, C. C.; Woese, C. R.; Mandelco, L.; Schauder, R.; Remsen, C. C.; Mitchell, R. Characterization of a new thermophilic sulfate-reducing bacterium Thermodesulfovibrio yellowstonii, gen. nov. and sp. nov.: its phylogenetic relationship to Thermodesulfobacterium commune and their origins deep within the bacterial domain. Archives of microbiology 1994, 161, 62–69. [Google Scholar] [CrossRef]
- Lo, N.; Lee, S. H.; Jin, H. M.; Jung, J. Y.; Schumann, P.; Jeon, C. O. Garicola koreensis gen. nov., sp. nov., isolated from saeu-jeot, traditional Korean fermented shrimp. Int. J. Syst. Evol. Microbiol. 2015, 65, 1015–1021. [Google Scholar] [CrossRef]
- Gomila, M.; Pinhassi, J.; Falsen, E.; Moore, E. R. B.; Lalucat, J. Kinneretia asaccharophila gen. nov., sp. nov., isolated from a freshwater lake, a member of the Rubrivivax branch of the family Comamonadaceae. Int. J. Syst. Evol. Microbiol. 2010, 60, 809–814. [Google Scholar] [CrossRef]
- Sun, L.; Toyonaga, M.; Ohashi, A.; Matsuura, N.; Tourlousse, D. M.; Meng, X.-Y.; Tamaki, H.; Hanada, S.; Cruz, R.; Yamaguchi, T.; et al. Isolation and characterization of Flexilinea flocculi gen. nov., sp. nov., a filamentous, anaerobic bacterium belonging to the class Anaerolineae in the phylum Chloroflexi. Int. J. Syst. Evol. Microbiol. 2016, 66, 988–996. [Google Scholar] [CrossRef] [PubMed]
- Tsubouchi, T.; Shimane, Y.; Mori, K.; Usui, K.; Hiraki, T.; Tame, A.; Uematsu, K.; Maruyama, T.; Hatada, Y. Polycladomyces abyssicola gen. nov., sp. nov., a thermophilic filamentous bacterium isolated from hemipelagic sediment. Int. J. Syst. Evol. Microbiol. 2013, 63, 1972–1981. [Google Scholar] [CrossRef]
- Yi, H.; Schumann, P.; Chun, J. Demequina aestuarii gen. nov., sp. nov., a novel actinomycete of the suborder Micrococcineae, and reclassification of Cellulomonas fermentans Bagnara et al. 1985 as Actinotalea fermentans gen. nov., comb. nov. Int. J. Syst. Evol. Microbiol. 2007, 57, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Allen, T. D.; Lawson, P. A.; Collins, M. D.; Falsen, E.; Tanner, R. S. Cloacibacterium normanense gen. nov., sp. nov., a novel bacterium in the family Flavobacteriaceae isolated from municipal wastewater. Int. J. Syst. Evol. Microbiol. 2006, 56, 1311–1316. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-J.; Wagner, I. D.; Brice, M. E.; Kevbrin, V. V.; Mills, G. L.; Romanek, C. S.; Wiegel, J. Thermosediminibacter oceani gen. nov., sp. nov. and Thermosediminibacter litoriperuensis sp. nov., new anaerobic thermophilic bacteria isolated from Peru Margin. Extremophiles : life under extreme conditions 2005, 9, 375–383. [Google Scholar] [CrossRef] [PubMed]
- Amoozegar, M. A.; Bagheri, M.; Didari, M.; Shahzedeh Fazeli, S. A.; Schumann, P.; Sánchez-Porro, C.; Ventosa, A. Saliterribacillus persicus gen. nov., sp. nov., a moderately halophilic bacterium isolated from a hypersaline lake. Int. J. Syst. Evol. Microbiol. 2013, 63, 345–351. [Google Scholar] [CrossRef]
- An, D.-S.; Im, W.-T.; Yang, H.-C.; Lee, S.-T. Shinella granuli gen. nov., sp. nov., and proposal of the reclassification of Zoogloea ramigera ATCC 19623 as Shinella zoogloeoides sp. nov. Int. J. Syst. Evol. Microbiol. 2006, 56, 443–448. [Google Scholar] [CrossRef]
- Raj, H.D. Proposal of Ancylobacter gen. nov. as a Substitute for the Bacterial Genus Microcyclus Ørskov 1928, Int. J. Syst. Evol. Microbiol 1983, 33, 397–398. [Google Scholar] [CrossRef]
- Deming, J. W.; Somers, L. K.; Straube, W. L.; Swartz, D. G.; MacDonald, M. T. Isolation of an obligately barophilic bacterium and description of a new genus, Colwellia gen. nov. Syst. Appl. Microbiol. 1988, 1, 1,152–160. [Google Scholar] [CrossRef]
- Baumgartner, L.K.; Reid, R.P.; Dupraz, C.; Decho, A.W; Buckley, D.H. J.R.; Spear, K.M. Przekop, P.T. Visscher: Sulfate reducing bacteria in microbial mats: Changing paradigms, new discoveries. Sedimentary Geology 2006, 185, 131–145. [Google Scholar] [CrossRef]
- Utkin, I.; Woese, C.; Wiegel, J. Isolation and characterization of Desulfitobacterium dehalogenans gen. nov., sp. nov., an anaerobic bacterium which reductively dechlorinates chlorophenolic compounds. International journal of systematic bacteriology 1994, 44, 612–619. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, L. A.; Rinna, J.; Humphreys, G.; Weightman, A. J.; Fry, J. C. Fluviicola taffensis gen. nov., sp. nov., a novel freshwater bacterium of the family Cryomorphaceae in the phylum ‘Bacteroidetes’. Int. J. Syst. Evol. Microbiol. 2005, 55, 2189–2194. [Google Scholar] [CrossRef] [PubMed]
- Romanenko, L. A.; Schumann, P.; Zhukova, N. V.; Rohde, M.; Mikhailov, V. V.; Stackebrandt, E. Oceanisphaera litoralis gen. nov., sp. nov., a novel halophilic bacterium from marine bottom sediments. Int. J. Syst. Evol. Microbiol. 2003, 53, 1885–1888. [Google Scholar] [CrossRef] [PubMed]
- Foesel, B. U.; Mayer, S.; Luckner, M.; Wanner, G.; Rohde, M.; Overmann, J. Occallatibacter riparius gen. nov., sp. nov. and Occallatibacter savannae sp. nov., acidobacteria isolated from Namibian soils, and emended description of the family Acidobacteriaceae. Int. J. Syst. Evol. Microbiol. 2016, 66, 219–229. [Google Scholar] [CrossRef]
- Yassin, A. F.; Hupfer, H.; Klenk, H.-P.; Siering, C. Desmospora activa gen. nov., sp. nov., a thermoactinomycete isolated from sputum of a patient with suspected pulmonary tuberculosis, and emended description of the family Thermoactinomycetaceae Matsuo et al. 2006. Int. J. Syst. Evol. Microbiol. 2009, 59, 454–459. [Google Scholar] [CrossRef]
- Yoon, J.; Ishikawa, S.; Kasai, H.; Yokota, A. Persicitalea jodogahamensis gen. nov., sp. nov., a marine bacterium of the family ‘Flexibacteraceae’, isolated from seawater in Japan. Int. J. Syst. Evol. Microbiol. 2007, 57, 1014–1017.77. [Google Scholar] [CrossRef]
- Walker, D.H.; Ismail, N.; Olano, J.P.; McBride, J.W.; Yu, X.-J.; Feng, H.-M. Ehrlichia chaffeensis: a prevalent, life-threatening, emerging pathogen. Trans. Am. Clin. Climatol. Assoc. 2004, 115, 375–82, discussion 382-4. [Google Scholar]
- Im, W.-T.; Yokota, A.; Kim, M.-K.; Lee, S.-T. Kaistia adipata gen. nov., sp. nov., a novel alpha-proteobacterium. The Journal of general and applied microbiology 2004, 50, 249–254. [Google Scholar] [CrossRef]
- Reddy, G. S. N.; Nagy, M.; Garcia-Pichel, F. Belnapia moabensis gen. nov., sp. nov., an alphaproteobacterium from biological soil crusts in the Colorado Plateau, USA. Int. J. Syst. Evol. Microbiol. 2006, 56, 51–58. [Google Scholar] [CrossRef]
- Nishijima, M.; Tazato, N.; Handa, Y.; Umekawa, N.; Kigawa, R.; Sano, C.; Sugiyama, J. Krasilnikoviella muralis gen. nov., sp. nov., a member of the family Promicromonosporaceae, isolated from the Takamatsuzuka Tumulus stone chamber interior and reclassification of Promicromonospora flava as Krasilnikoviella flava comb. nov. Int. J. Syst. Evol. Microbiol. 2017, 67, 294–300. [Google Scholar] [CrossRef]
- Su, X.-L.; Tian, Q.; Zhang, J.; Yuan, X.-Z.; Shi, X.-S.; Guo, R.-B.; Qiu, Y.-L. Acetobacteroides hydrogenigenes gen. nov., sp. nov., an anaerobic hydrogen-producing bacterium in the family Rikenellaceae isolated from a reed swamp. Int. J. Syst. Evol. Microbiol. 2014, 64, 2986–2991. [Google Scholar] [CrossRef] [PubMed]
- Ezaki, T.; Kawamura, Y.; Li, N.; Li, Z. Y.; Zhao, L.; Shu, S. Proposal of the genera Anaerococcus gen. nov., Peptoniphilus gen. nov. and Gallicola gen. nov. for members of the genus Peptostreptococcus. Int. J. Syst. Evol. Microbiol. 2001, 51, 1521–1528. [Google Scholar] [CrossRef] [PubMed]
- Bowman, J. P.; Nichols, C. M.; Gibson, J. A. E. Algoriphagus ratkowskyi gen. nov., sp. nov., Brumimicrobium glaciale gen. nov., sp. nov., Cryomorpha ignava gen. nov., sp. nov. and Crocinitomix catalasitica gen. nov., sp. nov., novel flavobacteria isolated from various polar habitats. Int. J. Syst. Evol. Microbiol. 2003, 53, 1343–1355. [Google Scholar] [CrossRef] [PubMed]
- Kwon, K. K.; Lee, H.-S.; Yang, S. H.; Kim, S.-J. Kordiimonas gwangyangensis gen. nov., sp. nov., a marine bacterium isolated from marine sediments that forms a distinct phyletic lineage (Kordiimonadales ord. nov.) in the ‘Alphaproteobacteria’. Int. J. Syst. Evol. Microbiol. 2005, 55, 2033–2037. [Google Scholar] [CrossRef]
- Kulichevskaya, I. S.; Danilova, O. V.; Tereshina, V. M.; Kevbrin, V. V.; Dedysh, S. N. Descriptions of Roseiarcus fermentans gen. nov., sp. nov., a bacteriochlorophyll a-containing fermentative bacterium related phylogenetically to alphaproteobacterial methanotrophs, and of the family Roseiarcaceae fam. nov. Int. J. Syst. Evol. Microbiol. 2014, 64, 2558–2565. [Google Scholar] [CrossRef]
- Fuerst, J. A.; Hawkins, J. A.; Holmes, A.; Sly, L. I.; Moore, C. J.; Stackebrandt, E. Porphyrobacter neustonensis gen. nov., sp. nov., an aerobic bacteriochlorophyll-synthesizing budding bacterium from fresh water. International Journal of Systematic Bacteriology 1993, 43, 125–134. [Google Scholar] [CrossRef]
- Oberreich, M.; Steinhoff-Knopp, B.; Burkhard, B.; Kleemann, J. The research gap between soil biodiversity and soil-related cultural ecosystem services. Soil Systems 2024, 8, 97. [Google Scholar] [CrossRef]
- Liu, Y.-X.; Qin, Y.; Chen, T.; Lu, M.; Qian, X.; Guo, X.; Bai, Y. A practical guide to amplicon and metagenomic analysis of microbiome data. Protein and Cell 2021, 12, 315–330. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The canabae legionis east and southeast of the legionary camp of Carnuntum. Georadar plot with sites of the pylons M 06, M 07 and M 08 (Konecny – Kulturabteilung des Landes Niederösterreich− LBI ArchPro).
Figure 1.
The canabae legionis east and southeast of the legionary camp of Carnuntum. Georadar plot with sites of the pylons M 06, M 07 and M 08 (Konecny – Kulturabteilung des Landes Niederösterreich− LBI ArchPro).

Figure 2.
Composition of bacterial communities of all samples by phyla (16S rRNA data); trench 6: samples C303-C347; trench 7: C248, C263, C266, trench 8: all other samples; sample names and short description of the data of their soil bacterial communities are given in Section 3.2.2. A short overview on sample numbers, soil pH and electrical conductivity data is given in supplementary Table S1.
Figure 2.
Composition of bacterial communities of all samples by phyla (16S rRNA data); trench 6: samples C303-C347; trench 7: C248, C263, C266, trench 8: all other samples; sample names and short description of the data of their soil bacterial communities are given in Section 3.2.2. A short overview on sample numbers, soil pH and electrical conductivity data is given in supplementary Table S1.
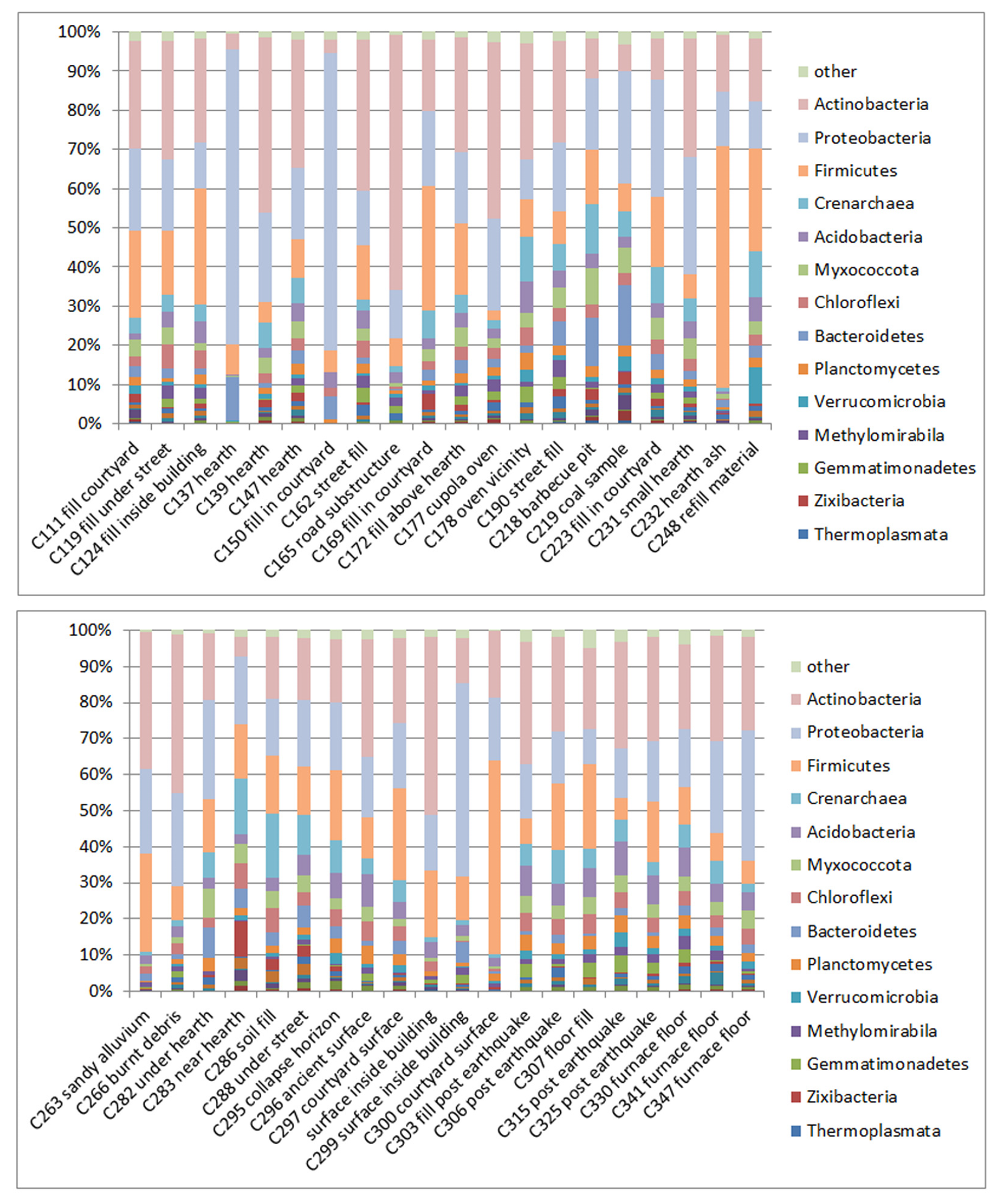
Figure 3.
Rank functions illustrating the differences in the alpha diversity for the different sampling points: a) samples C111 – C263, b) samples C266-C341.
Figure 3.
Rank functions illustrating the differences in the alpha diversity for the different sampling points: a) samples C111 – C263, b) samples C266-C341.
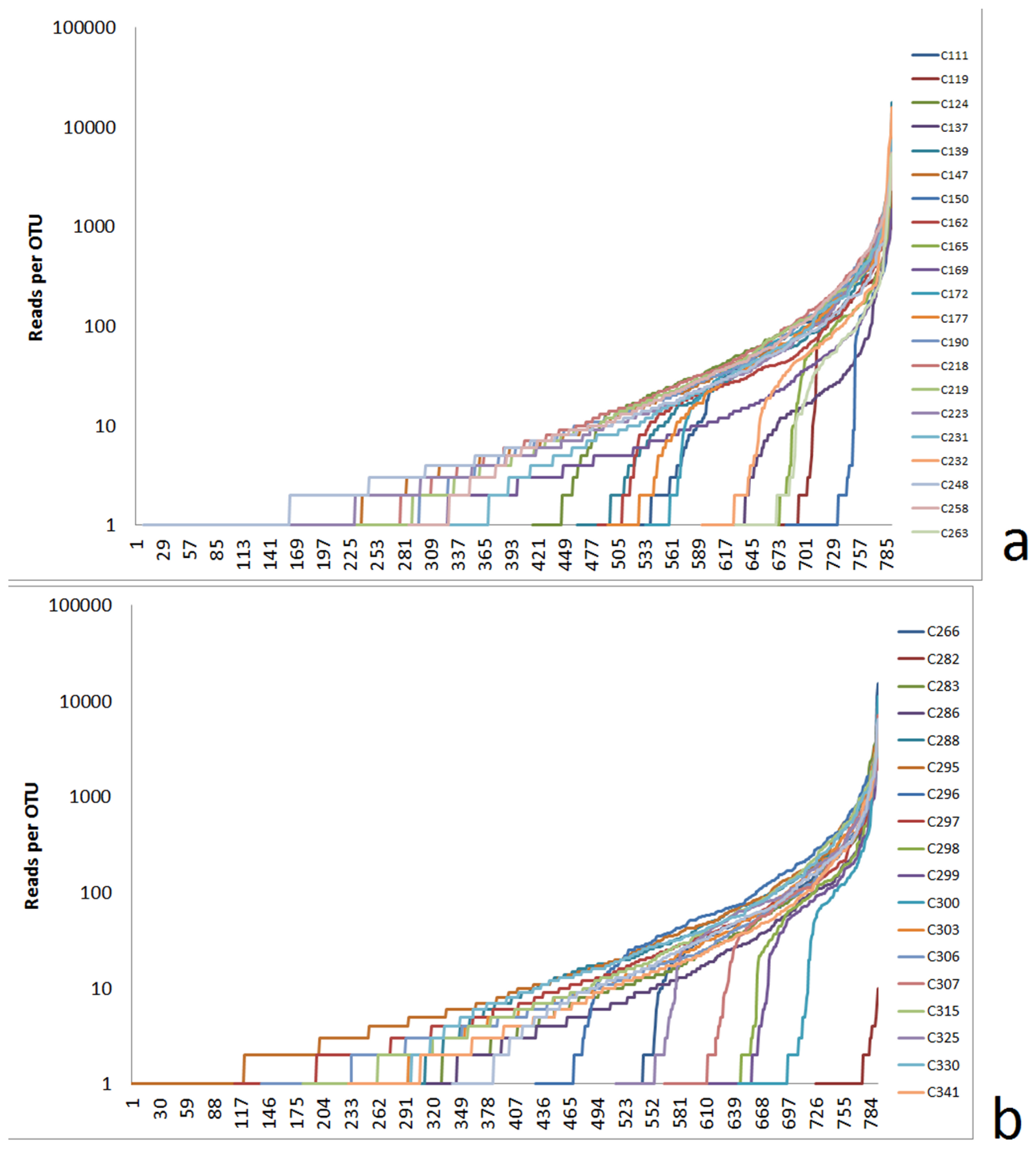
Figure 4.
Highly abundant OTUs: Samples sorted by the percentages of highly abundant bacteria; Sample names, meaning and short description of the data of their soil bacterial communities are given in chapter 4 (OTU patterns of single samples). A short overview on sample numbers, soil pH and electrical conductivity data is given in supplementary Table S1.
Figure 4.
Highly abundant OTUs: Samples sorted by the percentages of highly abundant bacteria; Sample names, meaning and short description of the data of their soil bacterial communities are given in chapter 4 (OTU patterns of single samples). A short overview on sample numbers, soil pH and electrical conductivity data is given in supplementary Table S1.
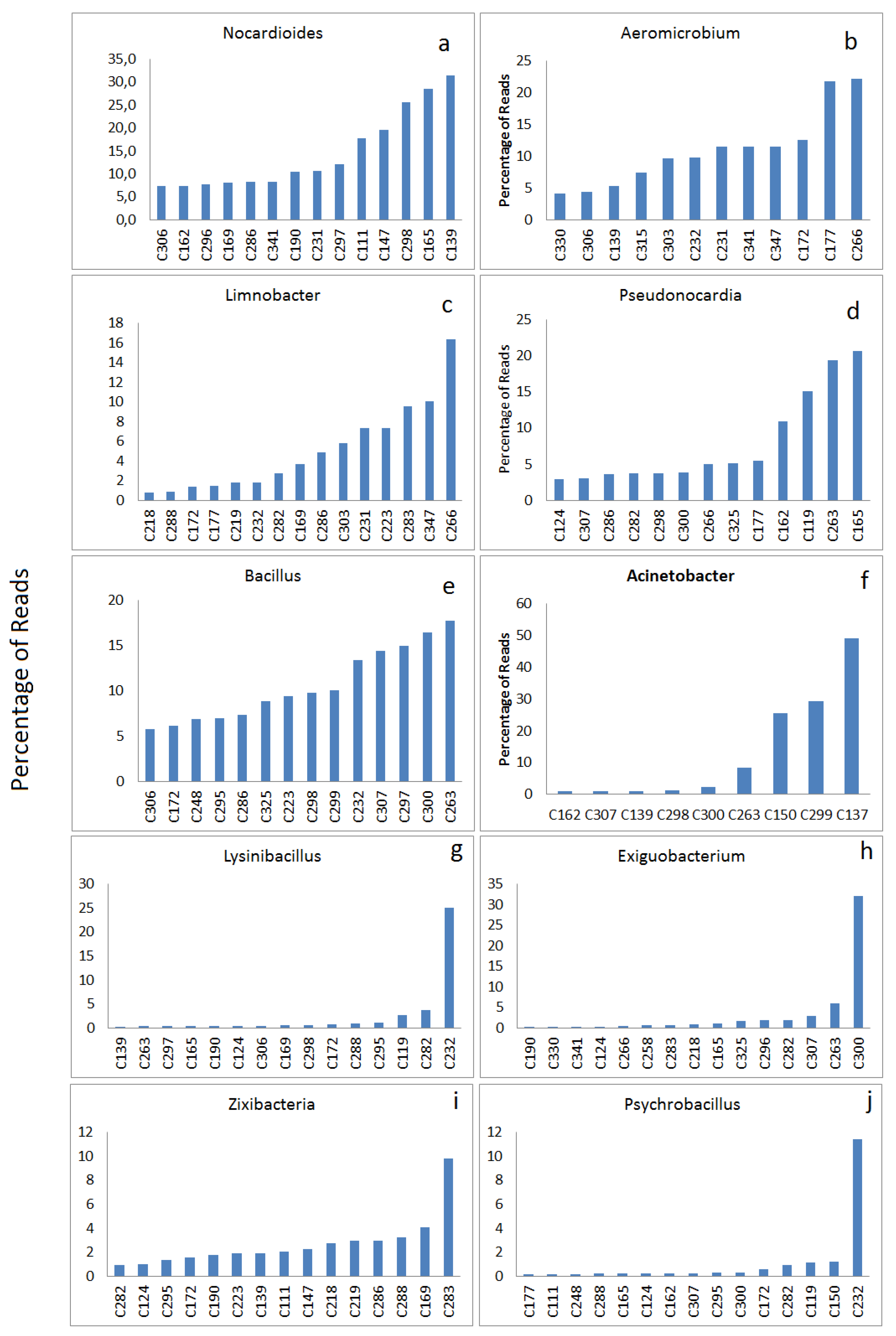
Figure 5.
PCA of different abundance classes: a) 10-99 reads in total over all samples (different symbols ate arbitrarily chosen for illustrating a certain clustering of samples), b) >10,000 reads in total (read symbols represent fire-related samples).
Figure 5.
PCA of different abundance classes: a) 10-99 reads in total over all samples (different symbols ate arbitrarily chosen for illustrating a certain clustering of samples), b) >10,000 reads in total (read symbols represent fire-related samples).
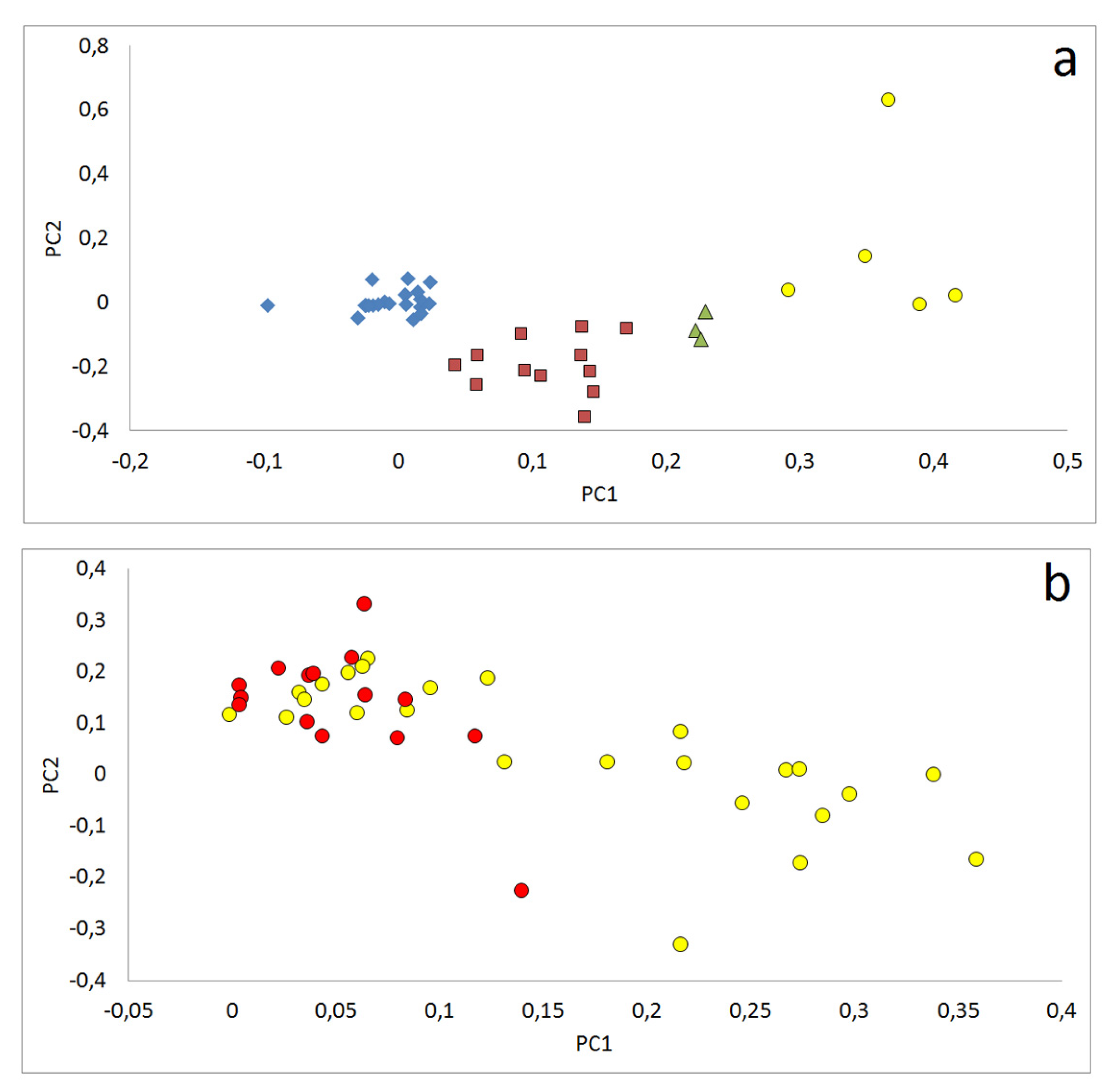
Figure 6.
Finger print-like abundance pattern (“FPP”) for the close-neighbourhood samples C218 (Trench M08 fill of a barbecue pit) and C219 (coal from the same fill pit) in comparison with all other samples; Sample names, meaning and short description of the data of their soil bacterial communities are given in Section 3.2.2. A short overview on sample numbers, soil pH and electrical conductivity data is given in supplementary Table S1.
Figure 6.
Finger print-like abundance pattern (“FPP”) for the close-neighbourhood samples C218 (Trench M08 fill of a barbecue pit) and C219 (coal from the same fill pit) in comparison with all other samples; Sample names, meaning and short description of the data of their soil bacterial communities are given in Section 3.2.2. A short overview on sample numbers, soil pH and electrical conductivity data is given in supplementary Table S1.
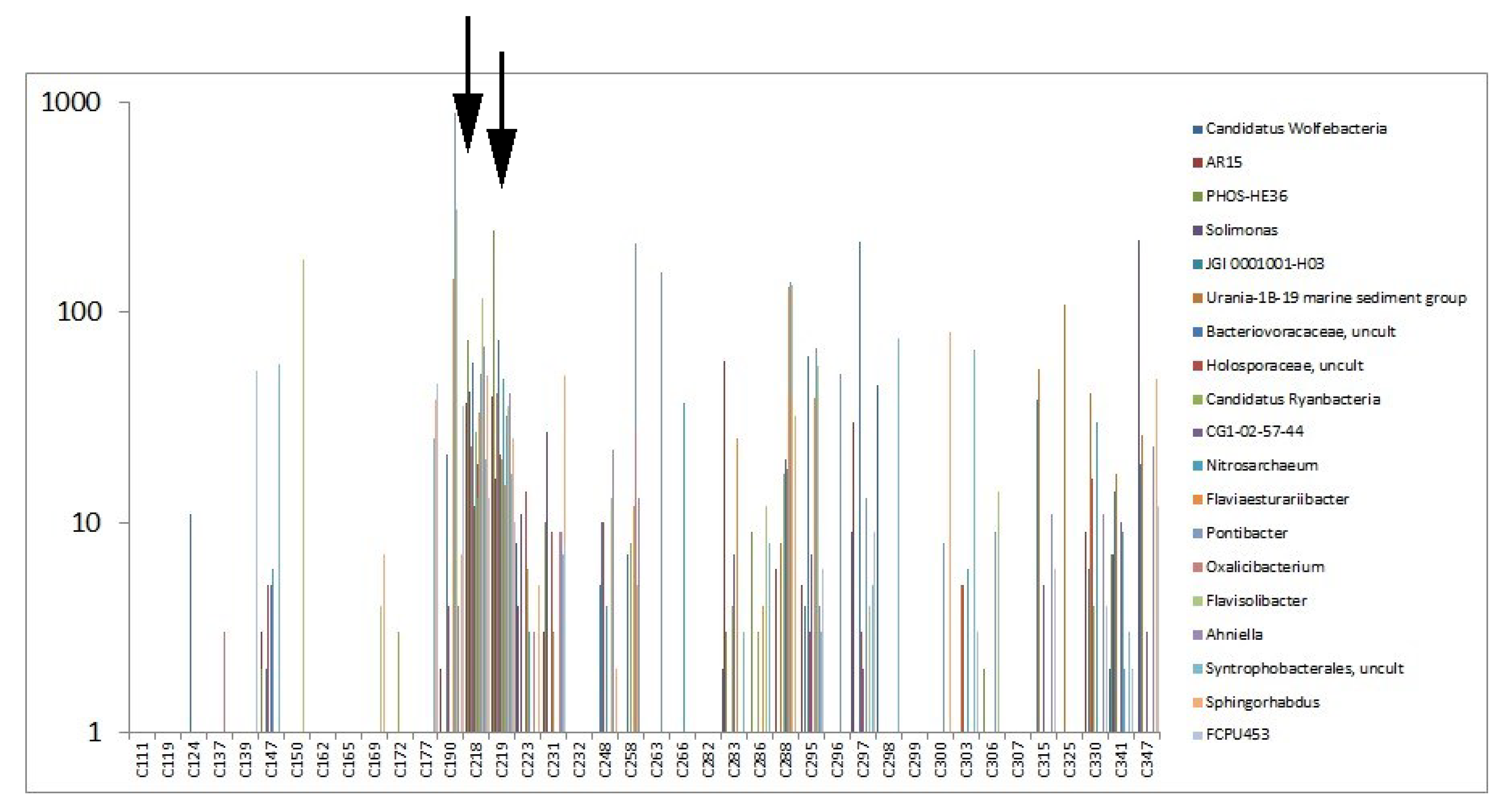
Figure 7.
Two examples of FPPs illustrating the relation between pairs of samples from the same spot, but different horizons separated by intermediate layers: a) abundances of a group of 11 OTUs present in in samples C295 and C297 (spot D of M08), separated by a floor horizon (sample C296), b) abundances of a group of 7 OTUs displaying an obvious relation of the both street horizons C162 (horizon E) and C288 (horizon H) of spot B of M08, separated by horizons F and G; Sample names, meaning and short description of the data of their soil bacterial communities are given in Section 3.2.2. A short overview on sample numbers, soil pH and electrical conductivity data is given in supplementary Table S1.
Figure 7.
Two examples of FPPs illustrating the relation between pairs of samples from the same spot, but different horizons separated by intermediate layers: a) abundances of a group of 11 OTUs present in in samples C295 and C297 (spot D of M08), separated by a floor horizon (sample C296), b) abundances of a group of 7 OTUs displaying an obvious relation of the both street horizons C162 (horizon E) and C288 (horizon H) of spot B of M08, separated by horizons F and G; Sample names, meaning and short description of the data of their soil bacterial communities are given in Section 3.2.2. A short overview on sample numbers, soil pH and electrical conductivity data is given in supplementary Table S1.
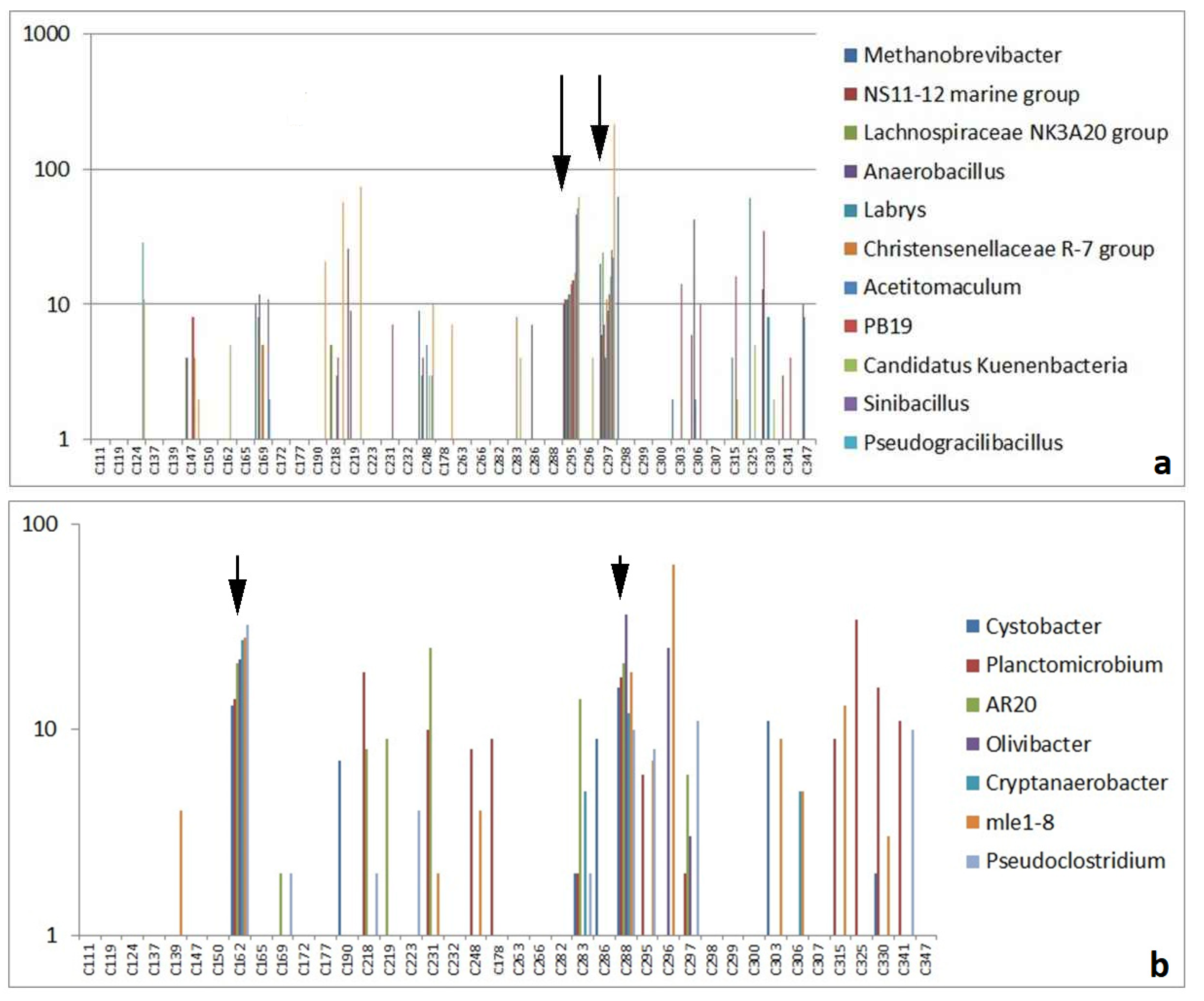
Figure 8.
Scheme of sampling sites in trench M08; Preferential localizations of high abundances of selected OTUs in different horizons: a) high concentration of Ralstonia in upper horizons (violet > 10%, dark blue: 2-4 %, light blue: 0.3 – 0.7%, yellow: < 0.3%), b) preferential concentration of Hydrogenophaga in lower horizons (blue: > 8 reads; dark blue: > 100 reads).
Figure 8.
Scheme of sampling sites in trench M08; Preferential localizations of high abundances of selected OTUs in different horizons: a) high concentration of Ralstonia in upper horizons (violet > 10%, dark blue: 2-4 %, light blue: 0.3 – 0.7%, yellow: < 0.3%), b) preferential concentration of Hydrogenophaga in lower horizons (blue: > 8 reads; dark blue: > 100 reads).
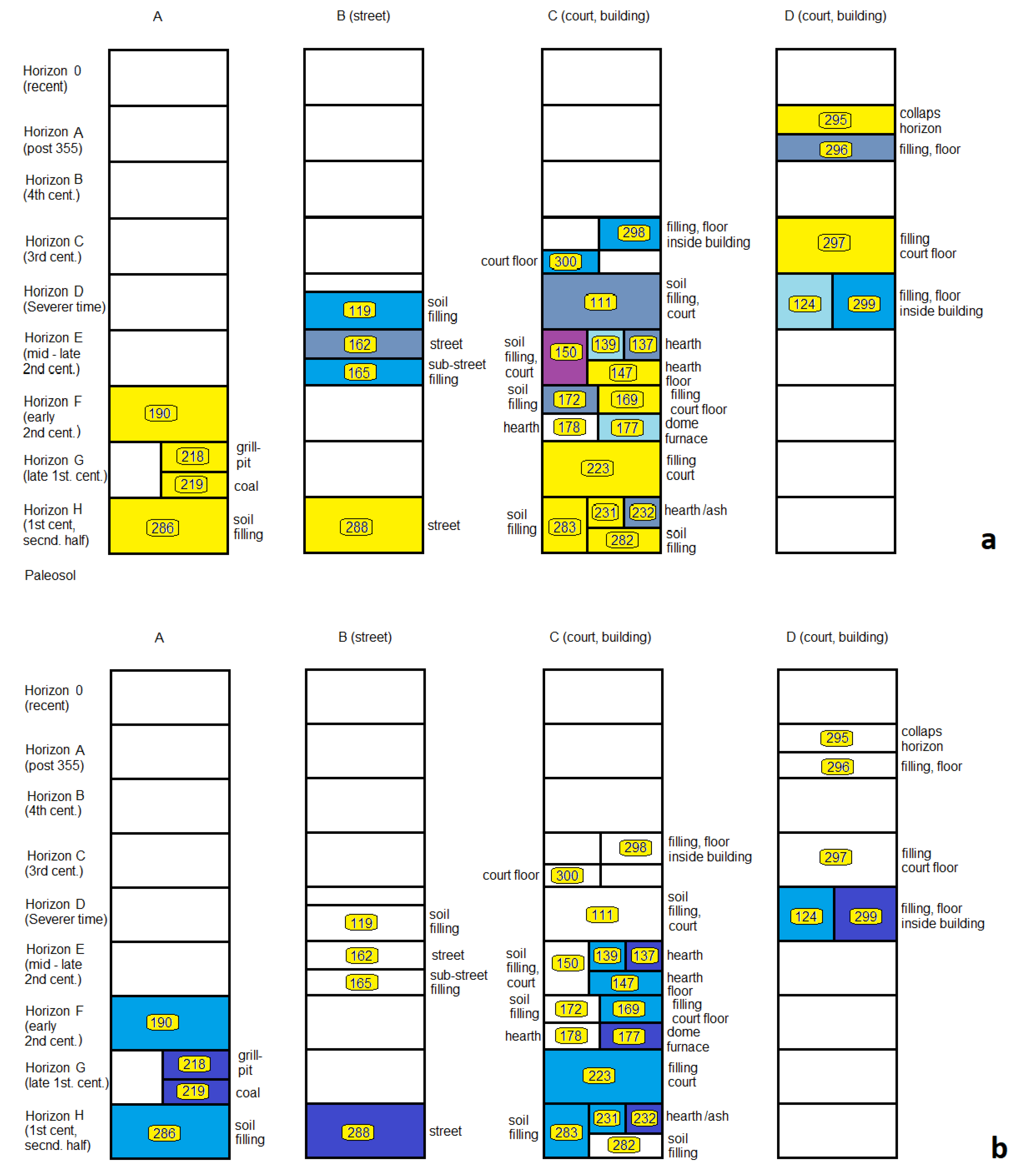
Figure 9.
Relations of single samples to the reference sample (C263, sandy alluvium): a) correlation of first and second principle component from a PCA reconsidering the 40 OTUs which are marked by the highest similarity of all samples (average) with sample C263 and indicating clearly the formation of two clusters of samples (marked, here, by blue diamonds and red squares), b) local distribution of samples of this both clusters at excavation trench M08 (blue fields corresponds to blue diamonds in Figure 9a, orange fields to red squares in Figure 9a, c) Color-coded pairs and groups of samples related by high correlation coefficients of r-values (logarithm of abundances) for OTUs with more than 10,000 reads in total (grey fields show no significant correlations with other samples).
Figure 9.
Relations of single samples to the reference sample (C263, sandy alluvium): a) correlation of first and second principle component from a PCA reconsidering the 40 OTUs which are marked by the highest similarity of all samples (average) with sample C263 and indicating clearly the formation of two clusters of samples (marked, here, by blue diamonds and red squares), b) local distribution of samples of this both clusters at excavation trench M08 (blue fields corresponds to blue diamonds in Figure 9a, orange fields to red squares in Figure 9a, c) Color-coded pairs and groups of samples related by high correlation coefficients of r-values (logarithm of abundances) for OTUs with more than 10,000 reads in total (grey fields show no significant correlations with other samples).
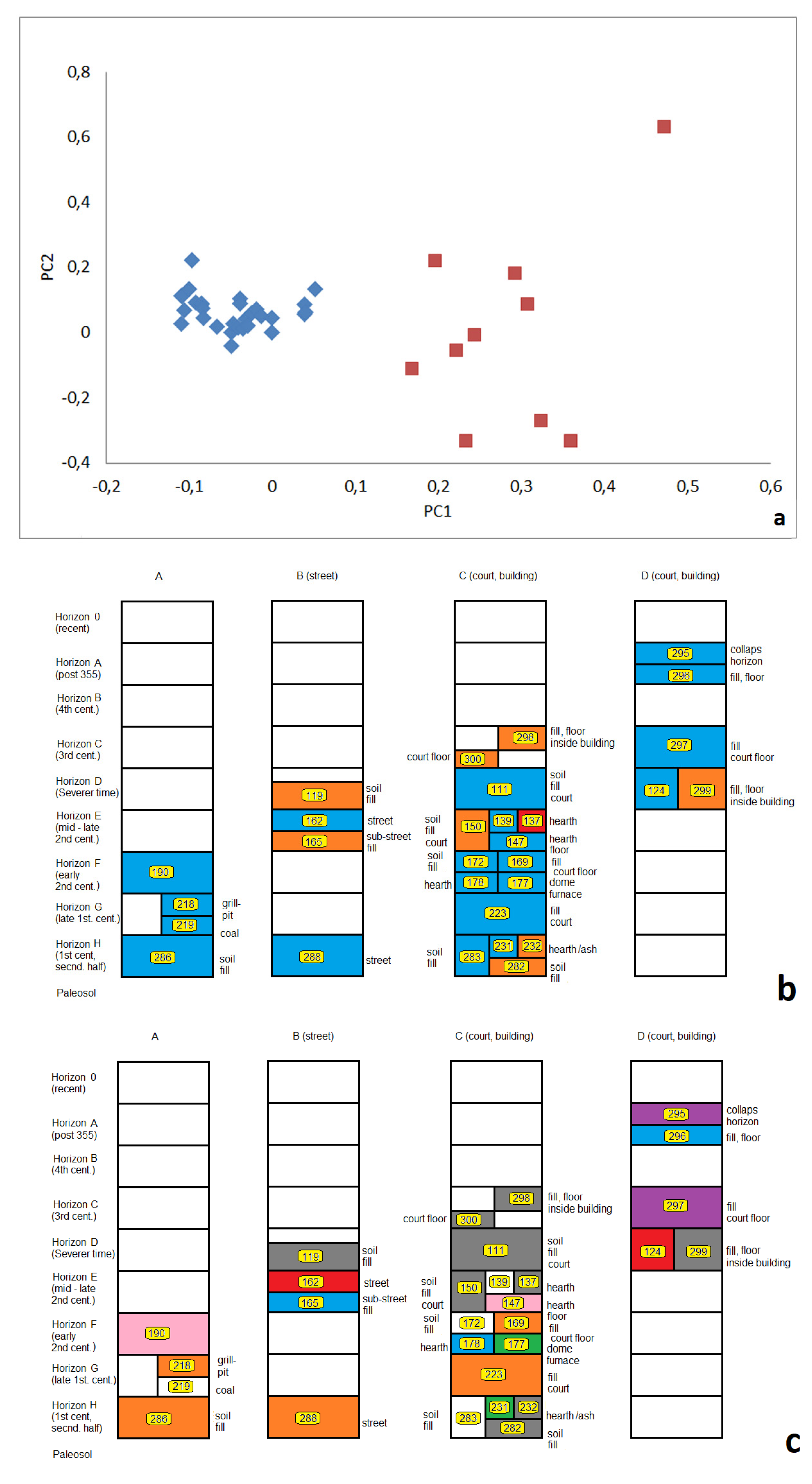
Figure 10.
Hypothetical ancient properties of soils at different sites and in different horizons of trench M08 proposed by interpretation of the composition of 16S rRNA data of local soil bacterial communities reconsidering mainly lower abundant and exclusive OTUs.
Figure 10.
Hypothetical ancient properties of soils at different sites and in different horizons of trench M08 proposed by interpretation of the composition of 16S rRNA data of local soil bacterial communities reconsidering mainly lower abundant and exclusive OTUs.
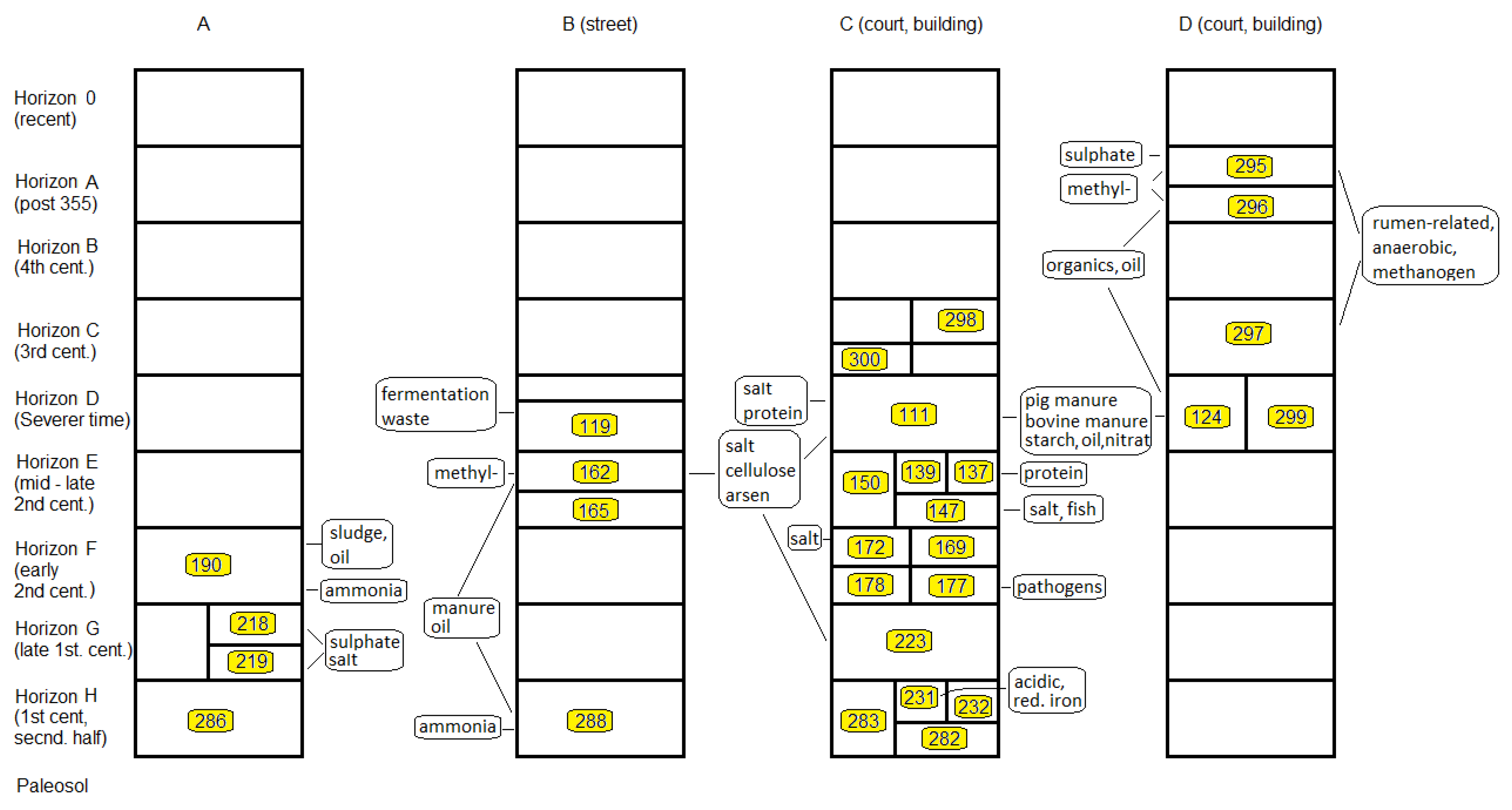
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



