
Submitted:
29 September 2024
Posted:
30 September 2024
You are already at the latest version
Abstract
Bee colonies that belong to the biocenosis are constantly threatened by the bacterial larvae disease called American foulbrood, caused by Paenibacillus larvae. The current state of the law prevents the inclusion of pharmacotherapy in the treatment and the only control is based on physical eradication of infected colonies by order of the Vet. The purpose of this study was to investigate the antimicrobial activity of selected bacteria with proven probiotic potential that could reduce American foulbrood pressure by promoting the development of the larvae microbiome that competes with and blocks the excessive proliferation and production of P. larvae endospores. The antimicrobial activity of inoculants of selected strains was studied: B. pumilus, B. licheniformis, S. narbonensis, L. fusiformis, L. brevis, B. megaterium against P. larvae ATCC 9545 and P. larvae CCUG 48973. Analyses were performed by the well-diffusion method according to EUCAST standards with modifications due to the specificity of the bacteria used. The largest zone of growth inhibition of P. larvae was confirmed with: S. narbonensis, B. licheniformis, B. megaterium, medium activity was observed with L. brevis and B. pumilus. Negligible activity was shown by L. fusiformis. Differences were noticed in the resistance of indicator strains of P. larvae and between the media and carriers used.
Keywords:
AFB
; Paenibacillus larvae
; bees
; probiotics
; biocontrol
; biosecurity
; beneficial microorganisms
; bioasecuration
; American foulbrood
1. Introduction
The work of honeybee workers for humans and the ecosystem is invaluable. EFSA reports that the annual pollination by bees has been valued at around 22 billion Euros, and in Poland alone, this value exceeds 964 million Euros [1,2]. Worldwide, the economic value of pollination with honeybee workers has been estimated at 265 billion Euros per year [3,4]. Other sources report that the global value of pollination involving honeybee workers is estimated to be between US$ 235-577 billion and up to half a trillion dollars [5,6,7,8]. Researchers have been searching for many years for factors that reduce losses in apiary management and increase the security of the honeybee as the main pollinator in the food production chain [9,10,11]. Bee colonies are constantly threatened by brood and adult bee diseases, susceptibility to xenobiotics (including pesticides), an impoverished forage base, lack of diverse protein food resulting in massive worker malnutrition (nutritional stress), environmental stress, and newer and newer pests arriving from different parts of the world (populations of the small hive beetle or Asian hornet) [12,13]. The honey bee is also accompanied by a microflora whose biodiversity is modified by many environmental factors, including stress factors such as pesticides, forage shortages, pressure from the pathogenic microbiome, and immune impairment [14,15,16]. According to various literature sources, there are between 6,000 and 8,000 different microbial strains in the microbiome of bee colonies [17].
One microbial threat to the balance of the colony microbiome is Paenibacillus larvae—a Gram-positive, peridomestic, mobile, virulent, spore-forming larvae that cause a brood disease called American foulbrood [18]. The bacteria Paenibacillus larvae, which produces approximately one billion spores per infected larva, is the main vector of this disease [19]. Only spores can cause infection. Spores can survive for up to 50 years under favorable conditions and exhibit infectious properties for 35 years. A temperature of 100°C destroys spores after 5 days, while at 140-170°C they die after only 2 hours. Spores are also killed in 5-10% formalin within 6 hours, while soda lye kills spores at a concentration of 2% within 4 minutes. 5% sodium hypochlorite is also effective. Vegetative forms die at 60°C [20,21,22]. An infected and disease-dead larva can contain up to 2.5 billion endospores, but only 10 to 35 spores are needed to infect another larva, indicating high virulence and a high rate of disease spread [23,24]. Spores can reside in honey, wax, royal jelly, propolis, pollen, and bee feathers. Once in the digestive tract of the larvae, they germinate after 24 hours, leading to bacterial superinfection manifested by rapid growth of vegetative forms. The vegetative cells damage the epithelium and intestinal walls and later attack all internal organs leading to cytolysis and histolysis [25,26,27]. Nine days after infection, spores begin to form. The maggots die after 2-3 days after the cell is sealed [28]. The dead larvae become flaccid, changing color first to yellow, then to a brown, sticky, and malleable mass. Five serotypes of American foulbrood are currently recognized worldwide: Eric I, Eric II, Eric III, Eric IV, and Eric V [29,30].
The epizootiological agent for all varieties remains the same bacterial species: Paenibacillus larvae. Sick bee colonies are physically eradicated on the recommendation of veterinarians, legislation in the EU does not allow the use of any form of pharmacotherapy, and in Poland, American foulbrood is eradicated ex officio. In the past, antibiotics and sulphonamides were used. Currently, only administrative procedures are available for American foulbrood control [31]. Once American foulbrood is confirmed, the district veterinarian may decide to eradicate the colonies or, in justified cases, order a double relocation procedure to a new or decontaminated hive on frames with hoses [32]. Most cases with confirmed cases of foulbrood are classified as a high-risk group and end up with the burning of infected colonies. Prevention consists primarily of maintaining appropriate sanitary standards involving, for example, frequent changes of combs, and disinfection of hives and tools (e.g., with Virkon S). Other prevention methods include descriptions of the use of essential oils: cinnamon, rosemary, thyme, lemon or aniseed. The effect of extracts of various herbal species on P. larvae cells is also known from in vitro laboratory studies [33,34]. Solutions described as effective in reducing P. larvae abundance include products to be applied as food additives (syrup or cake) and products to be applied as sprays or sprinkled on the inter-frame streets [35].
The Institute of Veterinary Medicine in Poland reports that P. larvae spores are present in between 23 and 45% of the cases analyzed. Infected colonies weaken very quickly, the number of spring decreases dramatically leading to so-called colony unsealing, the number of bees decreases, which leads to a reduction in the economy of the farm or the total liquidation of the apiary. Statistical data from the Veterinary Inspectorate in Poland indicates several hundred cases of detected outbreaks, but these data are unreliable and underestimated and differ from the actual state. In previous years, i.e., 2009-2011, screening for the occurrence of Paenibacillus larvae in bee colonies was carried out in 162 districts belonging to 9 voivodships. 6 510 honey samples were taken from 32 550 bee colonies from 2 294 apiaries. The disease was diagnosed in 45% of all apiaries examined, and the greatest intensity occurred in the Małopolskie voivodeship. In the Warmińsko-Mazurskie voivodeship, the presence of the bacteria was confirmed in 58% of the apiaries examined. Between 26% and 47% of apiaries were contaminated with P. larvae spores [36]. An even earlier history of the occurrence of American foulbrood in Poland is described in research studies conducted in 2005 and 2007. In 2005, out of 142 samples, 34 tested positive for P. larvae, meaning that the bacteria were present in 23% of all samples. Similar statistics were found in 2007, where 23 out of 100 tests were positive. The number of Aedospores in 1 gram of honey ranged from 10 to more than 1,000. Such a huge scale of infections, still increasing from year to year, shows that the Polish beekeeping environment requires and needs the immediate implementation of a solution to reduce the uncontrolled growing number of infections in apiaries by supporting the bees’ natural protective barriers and stimulating increased nest hygiene [37]. The data obtained from the above-mentioned monitoring indicates a growing problem with P. larvae in bee colonies, which promotes the increasing incidence of American foulbrood. A high microbial burden on bee colonies is also observed in other countries, with a high number of infections also recorded in Austria [38].
Currently, the number of bee colonies in Poland is 2.35 million, so this is a threefold increase since 2005 (827.4 thousand bee families). Bee families are looked after by around 97,000 beekeepers. The average bee colony per km2 is 7.50. Unfortunately, the ever-increasing degree of apiculture in Poland favors the circulation of diseases between bee colonies [39]. As of today, there are only 61 Veterinarians in Poland with a specialization in diseases of commercial insects [40]. In Poland, there are no specific legal restrictions on the movement of migratory apiaries, which exacerbates the epidemiological situation.
The phenomenon of competition between microorganisms is well-known. Bacterial metabolites, also known as metabiotics, have become an object of scientific interest in recent years. Micro-organisms isolated from the digestive tracts of bees with in vitro antimicrobial activity against P. larvae have been described in the literature [41,42]. Such activity may be called bioassurance. The definition given by Zygmunt Pejsak includes specific indications to be implemented to reduce or eliminate the risk of pathogen transmission into the flock. The aim of bioassurance is, among other things, to reduce the risk of introducing an infectious agent into the herd [43]. In the definition of another researcher Thomas Gillespie, bioassurance must reduce the risk of pathogen intrusion and must consist of bioexclusion, which will control the transmission of pathogens. This element of bioassurance has been referred to as biocontainment [44]. Bioassurance is the so-called biological protection consisting of protecting the animals through preventive and sanitary measures using biocontainment on the farm as well as in the immediate surroundings [45,46]. Adherence to bio-assurance aims, among other things, to reduce the incidence of pathogens and to improve animal health. Bioassurance is a set of measures aimed at maintaining or improving the health status of a herd through the use of specific organizational methods [47,48]. In other words, it is the biological protection of the farm. Effective bioassurance gives protection to the herd against the transmission of infectious agents [49,50,51,52]. Also, the Cambridge English Corpus states that these are the methods that are used to stop a disease or infection from spreading from one person, animal, or place to others or actions that are taken to prevent damage from biological threats using biological methods [53]. Biotisation as a tool for the introduction of bioassurance may be a new biotechnological approach to animal husbandry. It consists of inoculating the animal environment with beneficial micro-organisms such as fungi or bacteria to increase their dominance in the environmental microbiome by increasing the tolerance of animals to biotic and abiotic stresses (in practice this is spraying or fogging). Biotisation is the process of inoculating with symbiotic native bacteria (bacterization) or involves the introduction of beneficial fungi-yeasts (microzation) [54,55]. Beneficial micro-organisms also known as beneficial or effective microorganisms (EM) have an important function in restoring the microbial balance in the native microbiome of animals and also supply animals with valuable metabolites and protect them from pathogens. Biosanitisation is a process that seeks to eliminate as many pathogenic microorganisms as possible, most commonly on flat surfaces and in the animal environment. Sanitization is carried out using common cleaning agents or biopreparations. This process does not guarantee the sterility of the cleaned object but allows the degraded and sterilized animal microbiome to be inoculated and rebuilt [56,57]. In the case of bee colonies, this can include inoculation of nests and hives with probiotics as a permanent integral part of good beekeeping practice. Bacteriophage therapies appear to be some kind of innovation in reducing P. larvae infections, but phages are not officially authorized for use in Europe [58].
The lack of effective product solutions on the market to prevent the development of superinfection, the unstable administrative and legal status of American foulbrood, the risky practices of beekeepers and growers exacerbating the American foulbrood problem, the increased interest on the market in natural products like targeted microbial biopreparations with increased metabolite content, the untapped potential of biotechnology and microbiology in the control and prevention of livestock diseases and animal welfare are just some elements of why this topic of work was undertaken.
2. Materials and Methods
2.1. Research Problem
The complexity of the research problem concerns: (1) the reduction of American foulbrood pressure in bee colonies by supporting the development of the physiological microbiome of honeybee larvae, competing with and blocking the excessive proliferation and production of P. larvae endospores, and (2) testing the sensitivity of P. larvae vegetative cells to biologically active substances, constituted by biotechnological processes aimed at obtaining a high content of bioactive compounds in the final product using bacterial probiotic components.
2.2. Purpose of the Study
The aim of the study was: (1) to test the biological activity of selected bacterial components with confirmed probiotic potential that could constitute the composition of a pilot pre-formulation of a future metabolic preparation, and (2) to compare the antimicrobial activity of inoculants and post-culture (post-fermentation) fluids of selected, functional bacterial strains against Paenibacillus larvae ser. Eric I and Eric II.
2.3. Scope of Work
The overall scope of the work involves making inoculants of functional bacterial strains by revitalizing the bacterial strains by hydration of lyophilizes according to in-house procedures, then carrying out the actual multiplication (fermentation) in glass bioreactors and obtaining the first half pre-formulations on a laboratory scale. This consisted of transferring the functional bacteria cultures to selected media and multiplying by a technological process aimed at obtaining bioactive metabolites. The final step was to determine the antimicrobial activity of the culture fluids by determining the inhibition zones of the indicator strain. This was done by inoculating bloody agar with a suspension obtained from the selective multiplication of biological material on selective nutrient slants for P. larvae. Results were presented after 48-72 hours of incubation at 37°C.
2.4. Bacterial Isolates
2.4.1. Work Material
The working material consisted of the functional, bacterial strains which were used in an experiment to check their antimicrobial activity: Bacillus pumilus R5, Bacillus licheniformis M/542/M/18, Streptomyces narbonensis R71, Lysinibacillus fusiformis E23, Levilactobacillus brevis M/495/M/17, Bacillus megaterium R7. The strains were deposited at the Institute of Microbiological Technology in Turek, Poland, and subjected to deep-freezing at -80°C in glycerol. Isolation of the strains and species identification using 16S gene sequencing were carried out between 2014 and 2019. The morphology, physiology, and biochemical characteristics of the strains were very well described before the bacteria were banked using confocal microscopy, an API strip kit (BioMerieux), and the Biolog system (BioMaxima). The strains were from the wild environment from Poland and did not show virulence genes. The test strains had confirmed probiotic potential.
2.4.2. Biological Material
The biological material consisted of the bacterial, pathogenic, indicator strains belonging to Paenibacillus larvae ATCC 9545 (Labiol) and Paenibacillus larvae CCUG 48973 (Labiol). The strains were deposited at the Institute of Microbiological Technology in Turek, Poland, and subjected to deep-freezing at -80°C in glycerol. The strains were purchased for the experiment from an external supplier (Labiol) recommended by the district veterinary inspections. The supplier provided a protocol confirming the identification of the strains and a quality certificate with instructions for reviving the strains and the composition of the media. After revival, the species of the strains were control-confirmed by MALDI TOF/MS.
2.4.3. Cultivation of Microbial Cultures—Bacterial Inocula
The research and development work was divided into 2 stages. In stage 1, banked bacterial strains were revived. Colonies were fixed on appropriate media. Strains belonging to Bacillus pumilus R5, Bacillus licheniformis M/542/M/18, Lysinibacillus fusiformis E23, and Bacillus megaterium R7 were seeded on nutrient agar (Argenta) and incubation took place at 37°C under aerobic conditions. A strain belonging to Streptomyces narbonensis R71 was seeded on Streptomyces medium (Sigma-Aldrich) and incubation took place at 28°C under aerobic conditions. The strain belonging to Levilactobacillus brevis M/495/M/17 was seeded on MRS agar (Argenta) and incubation took place at 30°C under anaerobic conditions. All incubations were conducted for 24-48 hours (incubator BE400, Memmert, ZALMED, Poland). On reading the cultures, colony samples were control directed to MALDI-TOF/MS analyses to confirm species affiliation. Functional bacterial colonies were then transplanted onto agar-nutrient slants to derive young 24-hour colonies, incubated at specific temperature ranges for 24 hours under conditions appropriate to each strain (as above). At the end of the incubation, so-called microbial washes were made from the 24-hour cultures of pure bacterial cultures and inocula with an optical density of OD = 0.5° McFarland (approximately 1.0 x 108 CFU/cm3 (densitometer DEN-1B, Biosan, Poland) were obtained.
2.4.4. Preparation of Microorganism Cultures in Bioreactors
The prepared inoculants were used to inoculate in the bioreactors (Donserv, LPP Equipment) an industrial medium (autoclaved 4% aqueous solution of organic cane molasses on distilled water) at 4% by volume and a second, liquid tryptone-pepton with glucose and yeast extract medium (BTL) also at 4%. Cultures were conducted in 18 litre glass bioreactors at 30-38°C for 10 days under anaerobic conditions for L. brevis and for other bacteria under aerobic conditions all the time monitoring biotechnological parameters such as analysis of kinetics of pH value changes, ORP-Redox conductivity [µs/cm], sugar content [°Brix], asepsis, general contamination (culture in buffered peptone water), aerobic microorganisms (culture on PCA medium before and after application of buffered peptone water, Argenta), fungi (DRBC before and after buffered peptone water, Argenta), lactic fermentation bacteria (MRS and MRS with 2% fructose before and after buffered peptone water, Argenta), Clostridium sp. (culture on ISA medium before and after treatment with buffered peptone water, Argenta). The culture process was considered complete with a marked decrease in pH, an increase in conductivity (ORP-Redox), a decrease in Brix, and an increase in culture density [CFU/cm3].
2.5. Working Methodology
2.5.1. Agar Diffusion Assay
The second stage focused exclusively on screening and determination of antimicrobial activity against P. larvae. For the determination of antimicrobial activity, the well-diffusion method was used according to EUCAST standards with modifications due to the specificity of the bacteria used [59,60,61]. For this purpose, the P. larvae Eric I and Eric II strains were revived by transplanting colonies onto blood agar medium (Argenta) and then onto agar slants, respectively, incubated at 37°C for 24-48 h in a CO2 atmosphere (incubator BE400, Memmert, ZALMED, Poland). A so-called microbial wash was then performed with an optical density of OD = 0.5° McFarland (approximately 1.0 x 108 CFU/cm3, densitometer DEN-1B, Biosan, Poland). Each of the functional inoculants was combined respectively with the media used in beekeeping to feed the bee colonies (sterile water and sterile sucrose solutions of 50% and 60%, Merck), and mixtures of 3, 4, 5, 10% v/v were obtained. The culture fluid in undiluted form was treated to a concentration of 100%. Media without bacteria were used as control groups, i.e., sterile water and sterile sucrose solutions of 50% and 60% (Merck). Subsequently, surface cultures (1 ml) of P. larvae were inoculated using a microbial spatula on plates with blood agar and cut wells in the medium. 20 µl of the appropriate inoculant with carrier was transferred to the wells, and media not inoculated with inoculants was transferred to the control plates. All Petri dishes (Thermo Scientific) were incubated for 48-72 hours at 37°C in a CO2 atmosphere (incubator BE400, Memmert, ZALMED, Poland). After the time had elapsed, readings were taken, and ‘halo’ zones were measured [62,63,64,65].
2.5.2. Antimicrobial Activity of Bacterial Suspensions against P. larvae
The tests were carried out in 3 replicate series considering each functional strain (6 strains) and a mixture of strains (1), different media (2 media), percentage concentration of culture fluid (5 concentrations), media of appropriate concentration (3 media), and each replicate series consisted of a total of 210 Petri dishes (Thermo Scientific) for all strains and mixtures tested.
2.6. Statistical Analysis
Statistical analysis of the results obtained was performed using one-way ANOVA (Tukey’s parametric post-hoc test) at a significance level of p-value < 0.05. This analysis allows the means of any number of groups to be compared with each other, showing statistically significant similarities or statistically significant differences. The univariate scheme (one-way analysis of variance) tested whether one independent variable (factor) influenced the results of one dependent variable. The statistical analysis included results from the evaluation of antimicrobial potential (v/v concentrations of the inoculant—4 groups versus the carriers: water, sucrose 50%, and sucrose 60%—3 groups). In the scientific literature, results are also analyzed using Student’s t-test, Mann-Whitney U-test, Katz’s approximation, Pearson’s Chi-square test, or Fisher’s exact test [66,67,68,69].
3. Results
3.1. Preparation of Microorganism Cultures in Bioreactors
Strains belonging to the species: B. pumilus, B. licheniformis, S. narbonensis, L. fusiformis, L. brevis, and B. megaterium were successfully multiplied in a volume of 18 liters on target media. The culture lasted for 10 days. The release parameters met the final specifications of the process:
- Bacillus pumilus R5
Strain to belong to the species Bacillus pumilus successfully multiplied in a volume of 18 liters on target media. The culture lasted for 10 days. The final culture density was 4.5 x 105 CFU/cm3 in cane molasses medium to 4.7 x 105 CFU/cm3 in tryptone-peptone medium. There is a marked decrease in pH values, a slight decrease in ORP-Redox, and a slight consumption of sugars manifested by a weak decrease in Brix values. No infections with foreign microflora are observed [Table 1].
- Bacillus licheniformis M/542/M/18
Strain belonging to the species Bacillus licheniformis successfully multiplied in a volume of 18 liters on target media. The culture lasted for 11 days. The final culture density was 3.5 x 107 CFU/cm3 in cane molasses medium to 3.7 x 107 CFU/cm3 in tryptone-peptone medium. There is a marked decrease in pH values, a significant increase in ORP-Redox, and little consumption of sugars manifested by a weak decrease in Brix values. No infections with foreign microflora are observed [Table 2].
- Streptomyces narbonensis R71
Strain belonging to the species Streptomyces narbonensis successfully multiplied in a volume of 18 liters on target media. The culture lasted for 12 days. The final culture density was 4.5 x 106 CFU/cm3 in reed molasses medium to 4.7 x 106 CFU/cm3 in tryptone-peptone medium. There is a marked decrease in pH values, a significant increase in ORP-Redox, and little consumption of sugars manifested by a weak decrease in Brix values. No infections with foreign microflora are observed [Table 3].
- Lysinibacillus fusiformis E23
Strain belonging to the species Lysinibacillus fusiformis successfully multiplied in a volume of 18 liters on target media. The culture lasted for 10 days. The final culture density was 3.5 x 107 CFU/cm3 in cane molasses medium to 3.7 x 107 CFU/cm3 in tryptone-peptone medium. There is a marked decrease in pH values, a significant increase in ORP-Redox, and little consumption of sugars manifested by a weak decrease in Brix values. No infections with foreign microflora are observed [Table 4].
- Levilactobacillus brevis M/495/M/17
Strain belonging to the species Levilactobacillus brevis successfully multiplied in a volume of 18 litres on target media. The culture lasted for 8 days. The final culture density was 4.5 x 108 CFU/cm3 in cane molasses medium to 3.4 x 105 CFU/cm3 in tryptone-peptone medium. There is a marked decrease in pH values, a significant increase in ORP-Redox and significant consumption of sugars manifested by a significant decrease in Brix values. No infections with foreign microflora are observed [Table 5].
- Bacillus megaterium R7
Strain belonging to the species Bacillus megaterium successfully multiplied in a volume of 18 litres on target media. The culture lasted for 10 days. The final culture density was 2.7 x 108 CFU/cm3 in cane molasses medium to 2.6 x 108 CFU/cm3 in tryptone-peptone medium. There is a marked decrease in pH values, a slight increase in ORP-Redox for the cane molasses medium and a decrease in ORP-Redox for the tryptone-pepton medium, as well as a slight consumption of sugars manifested by a slight decrease in Brix values. No infections with foreign microflora are observed [Table 6].
3.2. Results Based on Agar Diffusion Assay
3.2.1. Antimicrobial Activity of Bacterial Suspensions against P. larvae ATCC 9545
An attempt to inhibit the growth of Paenibacillus larvae ser. Eric I—a factor threatening the health of honeybee larvae—using inoculants of selected bacteria with probiotic potential under in-vitro conditions was carried out successfully. Factors capable of reducing American foulbrood pressure in bee colonies by promoting the development of a physiological colony microbiome that competes with P. larvae and blocks excessive proliferation and production of P. larvae endospores were found, and strains were selected for further selection, biocontrol of American foulbrood and development of a bioactive composition. Comparative analysis of the antimicrobial activity of culture fluids of selected bacterial species against Paenibacillus larvae ser. Eric I under in-vitro conditions showed differences between the inoculants tested and thus between the bacterial strains used, as well as between the media, and culture media used. Under in-vitro conditions, 9 out of 12 single inoculants tested showed suppressive effects against indicator strains belonging to the P. larvae ser. Eric I species and additionally bacterial consortia labeled as Mix [Table 7].
The greatest zone of growth inhibition of P. larvae ser. Eric I was confirmed for culture fluids of bacterial species such as B. pumilus (suspended in water, molasses culture), Mix (suspended in 60% sucrose, molasses culture), L. brevis (suspended in 50% sucrose, molasses culture), and S. narbonensis (suspended in 60% sucrose, molasses culture) qualifying these microorganisms into the group with the greatest antimicrobial activity.
The group with high antimicrobial activity was assigned to strains belonging to the species: B. megaterium (suspended in 60% sucrose, molasses culture), L. brevis (suspended in 60% sucrose and 60% sucrose, molasses culture), L. brevis (suspended in 50% sucrose and 60% sucrose, tryptone-peptone culture), L. fusiformis (suspended in 60% sucrose, tryptone-peptone culture), S. narbonensis (suspended in 50% sucrose, molasses culture), B. licheniformis (suspended in 50% sucrose, molasses culture) and B. pumilus (suspended in 50% sucrose and 60% sucrose, molasses culture).
The group with lower antimicrobial activity was assigned to strains belonging to the species: B. licheniformis (suspended in water, molasses culture), S. narbonensis (suspended in 50% sucrose, tryptone-peptone culture), S. narbonensis (suspended in water, molasses culture), L. fusiformis (suspended in 50% sucrose, tryptone-peptone culture), L. brevis (suspended in water, molasses culture), B. megaterium (suspended in all carriers, tryptone-peptone culture), B. megaterium (suspended in water and 50% sucrose, molasses culture), Mix (suspended in water, tryptone-peptone culture) and Mix (suspended in water, molasses culture).
The group with negligible antimicrobial activity included a strain belonging to the species: B. pumilus (suspended in all carriers, tryptone-peptone culture), B. licheniformis (suspended in all carriers, tryptone-peptone culture), S. narbonensis (suspended in water and 60% sucrose, tryptone-peptone culture), L. fusiformis (suspended in water, tryptone-peptone culture), L. fusiformis (suspended in all carriers, molasses culture), L. brevis (suspended in water, tryptone-peptone culture) [Table 7, Figure 1, Figure 3].
The study also showed the superiority of the ‘Mix’ bacterial consortium over single inoculants for biocontrol of the development of P. larvae ser. Eric I infection [Table 7, Figure 1].
For some samples, bacterial overgrowth or infection of the plates marked with ‘P’ and ‘Z’ was noticed. The results of the interaction of the control samples (with water, 50% sucrose and 60% sucrose) always indicated a level of 0 mm. This confirms that the sucrose solution alone does not have an antimicrobial effect [Table 7, Figure 1].
3.2.2. Antimicrobial Activity of Bacterial Suspensions against P. larvae CCUG 48973
An attempt to inhibit the growth of Paenibacillus larvae ser. Eric I—a factor threatening the health of honeybee larvae—using inoculants of selected bacteria with probiotic potential under in-vitro conditions was carried out successfully. Factors capable of reducing American foulbrood pressure in bee colonies by promoting the development of a physiological colony microbiome that competes with P. larvae and blocks excessive proliferation and production of P. larvae endospores were found, and strains were selected for further selection, biocontrol of American foulbrood and development of a bioactive composition. Comparative analysis of the antimicrobial activity of culture fluids of selected bacterial species against Paenibacillus larvae ser. Eric II under in-vitro conditions showed differences between the inoculants tested and thus between the bacterial strains used, as well as between the media, and culture media used. Under in-vitro conditions, 9 out of 12 single inoculants tested showed suppressive effects against indicator strains belonging to the P. larvae ser. Eric II species and additionally bacterial consortia labeled as Mix [Table 8, Figure 3].
The greatest zone of growth inhibition of P. larvae ser. Eric II was confirmed for culture fluids of bacterial species such as B. licheniformis (suspended in water, molasses culture), Mix (suspended in water, 60% sucrose and 50% sucrose, tryptone-peptone culture), B. megaterium (suspended in water, tryptone-peptone culture), S. narbonensis (suspended in water, tryptone-peptone culture) qualifying these microorganisms into the group with the greatest antimicrobial activity.
The group with high antimicrobial activity was assigned to strains belonging to the species: L. brevis (suspended in 50% sucrose and 60% sucrose, tryptone-peptone culture and suspended in water, 50% sucrose, 60% sucrose, molasses culture), B. licheniformis (suspended in 50% sucrose and 60% sucrose, molasses culture), S. narbonensis (suspended in 50% sucrose and 60% sucrose, tryptone-peptone culture), S. narbonensis (suspended in water, molasses culture), B. megaterium (suspended in 50% sucrose, tryptone-peptone culture), B. megaterium (suspended in all carriers, molasses culture) and B. pumilus (suspended in water, 50% sucrose and 60% sucrose, molasses culture).
The group with lower antimicrobial activity was assigned to strains belonging to the species: S. narbonensis (suspended in 50% sucrose and 60% sucrose, molasses culture), L. fusiformis (suspended in all carriers, tryptone-peptone culture), L. fusiformis (suspended in 60% sucrose, molasses culture), B. megaterium (suspended in 60% sucrose, tryptone-peptone culture), Mix (suspended in all carriers, molasses culture).
The group with negligible antimicrobial activity included a strain belonging to the species L. fusiformis (suspended in water and 50% sucrose, molasses culture), L. brevis (suspended in water, tryptone-peptone culture), B. licheniformis (suspended in all carriers, tryptone-peptone culture) and B. pumilus (suspended in all carriers, tryptone-peptone culture) [Table 8, Figure 2, Figure 3].
The study also showed the superiority of the ‘Mix’ bacterial consortium over single inoculants for biocontrol of the development of P. larvae ser. Eric II infection [Table 8, Figure 2].
For some samples, bacterial overgrowth or infection of the plates marked with ‘P’ and ‘Z’ was noticed. The results of the interaction of the control samples (with water, 50% sucrose, and 60% sucrose) always indicated a level of 0 mm. This confirms that the sucrose solution alone does not have an antimicrobial effect [Table 8, Figure 2].
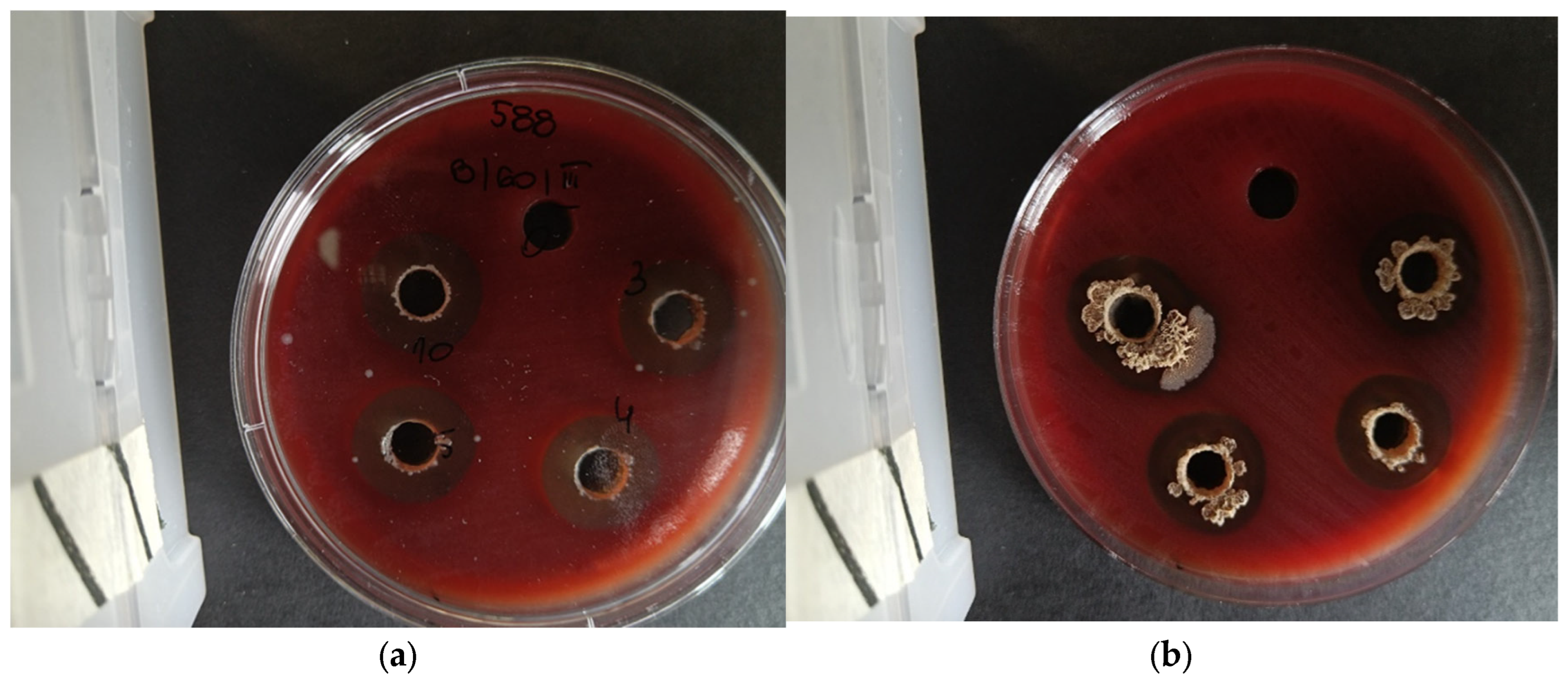
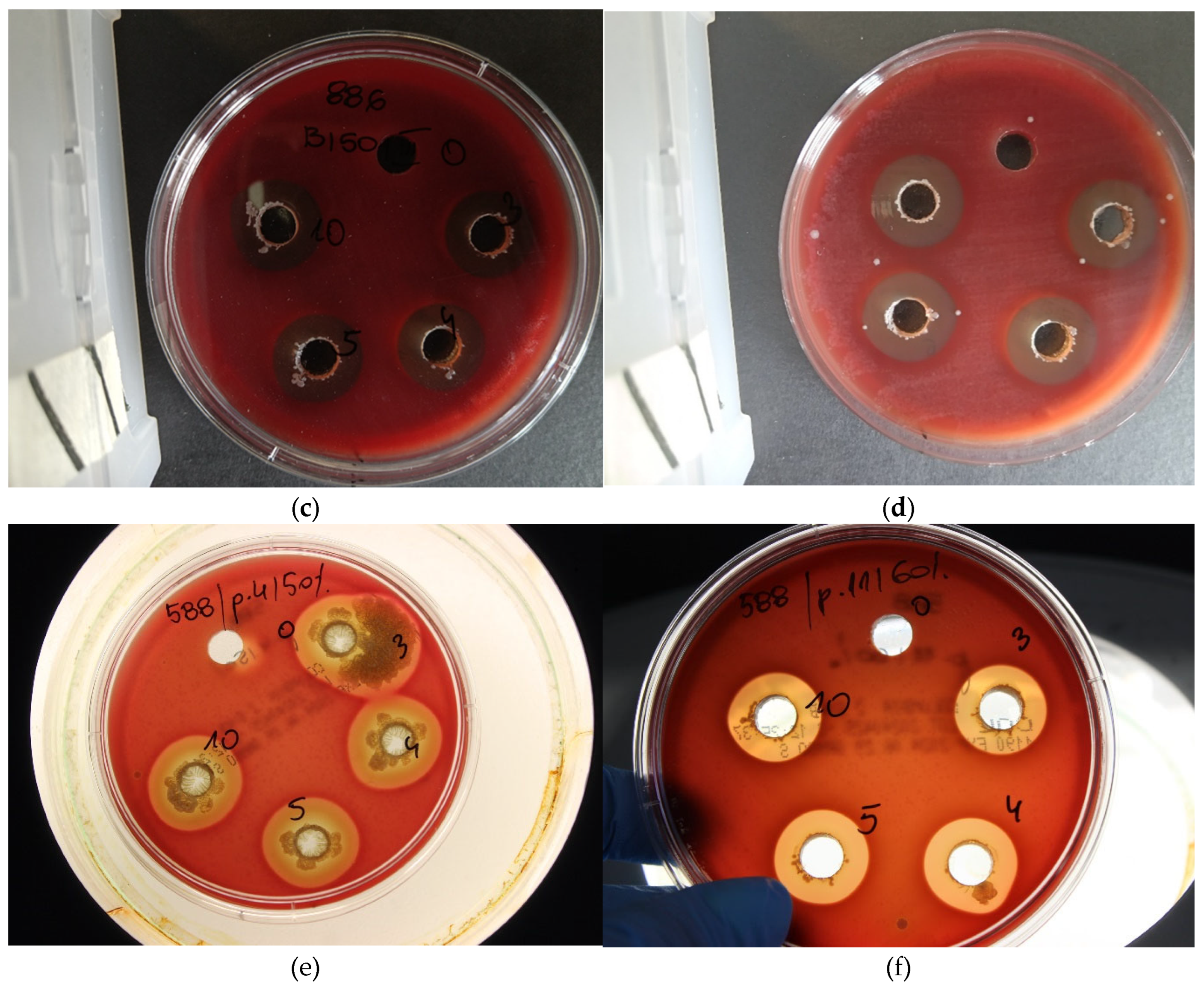
Figure 3.
Observed ‘halo’ effects with a visible zone of growth inhibition against strains belonging to the P. larvae species. [a-f] Tests were performed using the well-diffusion method according to EUCAST guidelines with modifications due to the specific bacteria used. [a, c, d, f] Antimicrobial mechanisms are of interest, showing antagonism between the microorganisms used, which may include, for example, the secretion of metabiotics, i.e., metabolites and bacteriocins, called antimicrobial substances or competition for food and growth conditions. [b, e] Impressive bacterial overgrowth of beneficial microorganisms is also observed, which takes away the growth capacity of pathogenic cells and even leads to lysis of pathogenic cells.
Figure 3.
Observed ‘halo’ effects with a visible zone of growth inhibition against strains belonging to the P. larvae species. [a-f] Tests were performed using the well-diffusion method according to EUCAST guidelines with modifications due to the specific bacteria used. [a, c, d, f] Antimicrobial mechanisms are of interest, showing antagonism between the microorganisms used, which may include, for example, the secretion of metabiotics, i.e., metabolites and bacteriocins, called antimicrobial substances or competition for food and growth conditions. [b, e] Impressive bacterial overgrowth of beneficial microorganisms is also observed, which takes away the growth capacity of pathogenic cells and even leads to lysis of pathogenic cells.
3.4. Results Based on Statistical Analyses
Statistical analysis of the results obtained was performed using one-way ANOVA (Tukey’s parametric post-hoc test) at a significance level of p-value < 0.05. This analysis allows the means of any number of groups to be compared with each other, showing statistically significant similarities or statistically significant differences.
It has been proven that there is a correlation and a statistically significant correlation between the microorganism used multiplied in the biotechnological fermentation process and the subsequent antimicrobial activity of the metabolites and bacteria remaining in the culture fluid. In some cases (experimental systems), there is a correlation and a statistically significant correlation between the inoculant concentration used and its subsequent antimicrobial activity. The analysis of antimicrobial activity showed statistically significant differences between the inoculants tested and therefore between the bacterial strains used and between the media, media, and indicator strains used. A one-way ANOVA analysis indicated a superior efficacy of the Mix against the pathogen analyzed. Statistical analysis (Tukey’s parametric post-hoc test) indicates increased resistance of the Paenibacillus larvae strain ATCC 9545 Eric I to the tested agents for American foulbrood biocontrol.
4. Discussion
In nature, the phenomena of competition and induction of antagonism between microorganisms are well known. Targeted microbial biopreparations for agriculture and wastewater treatment plants, metabolic preparations with targeted stimulatory effects in veterinary medicine, or targeted probiotics in supplementation and therapeutics with a strict therapeutic purpose and effect are of increasing interest. The approach to the gut microbiota and the environmental microbiome has changed dramatically in the last 40 years. Unfortunately, as a result of various factors, this microflora is constantly being depleted, which justifies the regular use of probiotics to restore the degraded microflora. In line with the provisions of the European Field-to-Table Strategy and the Green Deal, a reduction in the use of antibiotics in the ecosystem cycle is recommended. This is also indirectly related to increasing antibiotic resistance [70,71].
Also in the environment of the honey bee population, there is a continuous reduction of beneficial microbes. Despite prohibitions, beekeepers quite often use prohibited antimicrobial substances, including antibiotics, which then easily pass into bee products and become a real threat to consumers in the food safety chain. To overcome these problems, steps should be taken to develop a leak-proof prevention system and to implement natural product solutions, which could contain proven functional microorganisms, metabolites, or plant extracts to displace and reduce the pressure of pathogens from the bee colonies’ physiological microbiome. The research undertaken in this study has achieved TRL Technology Readiness Level IV and will certainly continue to develop an effective composition to reduce bacterial superinfections caused by Paenibacillus larvae, together with a methodology for the application of such selected care products in beekeeping practice.
The development of American foulbrood and its milder variant, European foulbrood, caused by the bacterium Melissococcus plutonius, mainly occurs in stressful situations, when malnutrition and starvation occur in the bee colony. Endospores of Paenibacillus larvae and M. plutonius begin to germinate and transform into vegetative forms of the bacteria when the glucose concentration in the simple intestine of the honeybee larva falls below 2.5 %. Certainly, the conditions for the development of any bacterial or fungal brood disease are linked to a disruption of the hive microflora, which, as a result of undetermined factors, has been reduced to such an extent that the spread of American foulbrood bacteria occurs at an extremely fast rate, leading to an exponential logarithmic growth of vegetative forms. American and European foulbrood bacteria are proteolytic pathogens, meaning that they rapidly lead to cytolysis and histolysis of the bee larvae. When the hive has a shaky, impoverished microbiome, even the smallest number of spores delivered to the hive with frames or through bee trophallaxis can lead to the expansion and domination of the hive environment by P. larvae [72,73,74].
Beekeepers’ use of mite killers to reduce the population of the bee tormentor Varroa destructor also has a large impact on the significant reduction of the beneficial hive microflora. Such agents include acaricides, pyrethroids, organic acids, thymol, or organophosphate insecticides [75].
The problem is global and also affects regions where antibiotic treatment is allowed. The pharmacokinetics of an antibiotic usually ends when one or, less frequently, two of the active substances are depleted or degraded, causing bacteria to rapidly acquire resistance to the active substance. In the case of probiotics, there is a specific fusion of many different biologically active substances with a complex structure and diverse mechanisms of action, which means that the bacterial cell is not able to produce as many defense mechanisms and provide resistance. Probiotic bacteria secrete a variety of phenol acids, flavonoid compounds, phenolic compounds, bacteriocins, antibiotics, organic acids, bioflavonoids, and biologically active compounds [76,77,78].
Researchers searching for microbial antagonistic agents for American foulbrood biocontrol are most often limited to testing strains from the genera Lactobacillus and Bifidobacterium. Studies in which bacteria obtained from healthy bee colonies are isolated are also encountered. These are specific microorganisms derived from the microbiome of bee colonies and usually involve the genera Bacillus sp., Streptococcus, Candida sp., Saccharomyces, Pseudobacterium, Fructobacillus, Parasaccharibacter, Ochrabactum, Acinetobacter, Stenotrophomonas, Gilliamella, Snodgrassella, Actinomadura, Streptomyces, Apilactobacillus, Bombilactobacillus and many others. As there are between 6,000 and 8,000 different microbial strains in the bee microbiome it seems nothing new or harmful to administer specific probiotics and supplement the microflora. In human medicine, the effect of so-called probiotic interventions and the administration of specific strains to support, for example, diabetes, cancer, or obesity, thus modifying the diseased microbiome, is increasingly being studied [79,80,81,82].
The results from the comparative analysis of the antimicrobial activity of culture fluids of selected probiotic bacterial species against Paenibacillus larvae ser. Eric I and II under in-vitro conditions are consistent with literature data and tests evaluated by E. Forsgren, S. Lamei, and T. Olofsson [83,84,85]
High efficacy in antimicrobial activity was also achieved in a 2006 study conducted by Alippi and Reynaldi, confirming high antimicrobial activity for Bacillus pumilus, Bacillus licheniformis, and Bacillus megaterium against P. larvae ATCC 9545 [86].
Audisio et al. described the effective combination of a Bacillus subtilis strain with a plant extract against P. larvae highlighting the high utility of Bacillus spp. [87].
Also Bartel and Alippi in 2018 identified natural antagonists of P. larvae by selecting strains of Bacillus species and related genera producing a broad range of antimicrobial compounds, with activity against bacteria and fungi that include peptides, lipopeptides, bacteriocins, and bacteriocin-like inhibitory substances. By using biological tools, they evaluated the antagonistic activity of 34 bacterial strains against Paenibacillus larvae and Ascosphaera apis, the causal agents of American Foulbrood and Chalkbrood diseases of honey bee larvae, respectively. In the work in question, a strong bactericidal effect attributed to a strain of Bacillus megaterium and Bacillus licheniformis was demonstrated. Similarly, in studies by other researchers, B. megaterium showed strong antimicrobial activity due to the production of megacins by Bacillus megaterium and also lichenin and lichenicidin produced by Bacillus licheniformis [88].
Studies by other authors have documented that Bacillus licheniformis and another strain tested in the experiment reduced the mortality of American foulbrood-infected larvae. Treatments showed positive effects and reduced mortality [89].
Many studies by other authors confirm the efficacy of the bacterial species analyzed in reducing American foulbrood. This demonstrates the pertinent and successful selection of bacterial strains that can find application in the prevention of this disease. In other studies, B. licheniformis and B. subtilis also showed antimicrobial activity against P. larvae isolated from the bee environment in Saudi Arabia [90].
There are no sources that report what antagonistic effect Streptomyces narbonensis species may have. Korean researchers demonstrated that isolates extracted from forest soil, also identified as human pathogens, containing mainly strains of the genus Streptomyces showed strong pressure and antimicrobial activity against Paenibacillus larvae isolates supporting the theory that the source of the search for microbial agents need not only be the healthy bee microbiome but also other sources and natural resources [91].
Bacterial metabolites are reported in great detail by Adrian A. Pinto-Tomas and co-workers, who isolated Actinobacteria in the genus Streptomyces from foraging bees, and especially common in pollen stores. One strain, isolated from pollen stores, exhibited pronounced inhibitory activity against Paenibacillus larvae [92].
Q. Jamal, on the other hand, describes the widespread use of bacteria of the genus Lysinibacillus. Moreover, some Lysinibacillus species have antimicrobial potential due to bacteriocins, peptide antibiotics, and other therapeutic molecules. Therefore, this review will explore the biological disease-fighting, food preservation, therapeutic, plant growth-promoting, bioremediation, and entomopathogenic potentials of the genus Lysinibacillus [93].
This shows a well-chosen vector of search and selection of strains for the work in question. Good probiotic properties, high survival rates, and origin from the hive microbiome are reported by Polish researchers in the context of B. pumilus, and B. licheniformis which again shows the correct selection in microbial agents for biocontrol of American foulbrood. It is therefore confirmed that B. pumilus and B. licheniformis species also occur in the hive environment [94]
Levilactobacillus brevis also shows the ability to reduce the biofilm formation of P. larvae isolated from honey bee guts or fresh pollen samples, as indicated by the mechanism of action of this species [95].
In a study by Polish scientists, Levilactobacillus brevis B50 increased the expression of pattern recognition receptors and genes encoding antimicrobial peptides (defensin-1 and abaecin [96]. The LAB isolated from the honey bee gut has been demonstrated to be helpful for the inhibition of P. larvae. The antibiotic action of L. brevis is also based on its secretion of organic acids [97,98].
5. Conclusions
Using lactic acid bacteria (LAB) as an alternative to chemical treatments is a promising novel technique for tackling honey bee diseases and improving their immunity. The concept of reducing bacterial superinfections should move towards strict prevention using biosecurity to improve the welfare of honeybee colonies. It is highly advisable that in the case of developing bacterial superinfections with P. larvae, the legislative authorities, through veterinary entities, authorize specific measures consisting of the implementation of probiotic interventions using inoculants of tested bacteria with proven beneficial effects on bees. Such a strategy is illustrated by results from numerous scientific reports.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table 7: Data show antimicrobial activity against Paenibacillus larvae ATCC 9545 (ser. Eric I), using well-diffusion method according to EUCAST standards [mm] with heat map effect (red color means low antimicrobial activity or lack of antimicrobial properties, green color means high activity); Figure 1: Radar plot illustrating the distribution of tested inoculants with antimicrobial potential against Paenibacillus larvae ATCC 9545 ser. Eric I using the well-diffusion method [mm] according to EUCAST standards. Volumetric concentrations of 3%, 4%, 5%, and 10% were considered, the symbol ‘TP’ means tryptone-peptone medium, the symbol ‘M’ means reed molasses medium, the symbol ‘50% S’ means a 50% sucrose solution, the symbol ‘60% S’ means a 60% sucrose solution, the symbol ‘Mix’ means a consortium of all functional strains for a given experimental system; Table 8: Data show antimicrobial activity against Paenibacillus larvae CCUG 48973 (ser. Eric II), using well-diffusion method according to EUCAST standards [mm] with heat map effect (red color means low antimicrobial activity or lack of antimicrobial properties, green color means high activity); Figure 2: Radar plot illustrating the distribution of tested inoculants with antimicrobial potential against Paenibacillus larvae CCUG 48973 ser. Eric II uses the well-diffusion method [mm] according to EUCAST standards. Volumetric concentrations of 3%, 4%, 5%, and 10% were considered, the symbol ‘TP’ means tryptone-peptone medium, the symbol ‘M’ means reed molasses medium, the symbol ‘50% S’ means a 50% sucrose solution, the symbol ‘60% S’ means a 60% sucrose solution, the symbol ‘Mix’ means a consortium of all functional strains for a given experimental system; Figure 3: Observed ‘halo’ effects with a visible zone of growth inhibition against strains belonging to the P. larvae species. [a-f] Tests were performed using the well-diffusion method according to EUCAST guidelines with modifications due to the specific bacteria used. [a, c, d, f] Antimicrobial mechanisms are of interest, showing antagonism between the microorganisms used, which may include, for example, the secretion of metabiotics, i.e., metabolites and bacteriocins, called anti-microbial substances or competition for food and growth conditions. [b, e] Impressive bacterial overgrowth of beneficial microorganisms is also observed, which takes away the growth capacity of pathogenic cells and even leads to lysis of pathogenic cells.
Author Contributions
Writing—original draft preparation, P.N.; conceptualization, P.N., B.G.; methodology, P.N., P.C., P.M., B.G.; investigation, P.N., P.C., P.M., B.G.; validation, P.C., P.M., B.G.; supervision and investigation. All authors contributed to writing the original draft, reviewing, and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Polish Ministry of Science and Higher Education, grant number N0CBR000.7117.UWD.6/CBR/2021.
Institutional Review Board Statement
Not applicable
Informed Consent Statement
Not applicable
Data Availability Statement
All data generated or analyzed during this study are included in this published article and are available from the authors upon reasonable request.
Acknowledgments
We thank the laboratory specialists at the Institute of Microbiological Technologies in Turek for excellent technical assistance.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Kumar, Ashok, Aman Shukla, and Shashank Kailkhura. Biodiversity Loss and Its Economic Costs: A Global Perspective. Research Journal of Recent Sciences 2024.
- Fikadu, Zekiros. The Contribution of Managed Honey Bees to Crop Pollination, Food Security, and Economic Stability: Case of Ethiopia. no. 1, 2019. [CrossRef]
- Potts, Simon G. The Assessment Report on Pollinators, Pollination and Food Production: Summary for Policymakers. Secretariat of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services, 2016.
- FAO. “Pollinators Vital to Our Food Supply under Threat.” Newsroom, https://www.fao.org/newsroom/detail/Pollinators-vital-to-our-food-supply-under-threat/en. Accessed 27 Sept. 2024.
- Khalifa, Shaden A. M., et al. “Overview of Bee Pollination and Its Economic Value for Crop Production.” Insects, vol. 12, no. 8, July 2021, p. 688. [CrossRef]
- European Commission. Nature and Biodiversity—European Commission. Available online: https://environment.ec.europa.eu/topics/nature-and-biodiversity_en (accessed on 18 September 2024).
- European Commission. Honey Bees—European Commission. Available online: https://food.ec.europa.eu/animals/live-animal-movements/honey-bees_en (accessed on 27 September 2024).
- Gallai, Nicola, et al. “Economic Valuation of the Vulnerability of World Agriculture Confronted with Pollinator Decline.” Ecological Economics, vol. 68, no. 3, Jan. 2009, pp. 810–21. [CrossRef]
- Genersch, E., Evans, J.D., Fries, I. Honey bee disease overview. Journal of Invertebrate Pathology. 2009. 103:S2-S4.
- Arbia, A., and B. Babbay. “Management Strategies of Honey Bee Diseases.” Journal of Entomology, vol. 8, no. 1, July 2010, pp. 1–15. [CrossRef]
- Jelena Bošković1,* and Nenad Trkulja. INFLUENCE OF INSECTICIDES IN PRODUCTION OF HONEY BEES. Available online: https://www.fimek.edu.rs/downloads/casopisi/jatem/issue/v2_3/02-.
- Cappa, Federico, et al. “Hornets and Honey Bees: A Coevolutionary Arms Race between Ancient Adaptations and New Invasive Threats.” Insects, vol. 12, no. 11, Nov. 2021, p. 1037. [CrossRef]
- Genersch, Elke. “Honey Bee Pathology: Current Threats to Honey Bees and Beekeeping.” Applied Microbiology and Biotechnology, vol. 87, no. 1, June 2010, pp. 87–97. [CrossRef]
- Shimanuki, Hachiro; Knox, David A. Diagnosis of Honey Bee. Available online: https://www.ars.usda.gov/is/np/honeybeediseases/honeybeediseases.pdf (accessed on 23 July 2024).
- Rúa, Pilar De la, et al. “Biodiversity, Conservation and Current Threats to European Honeybees.” Apidologie, vol. 40, no. 3, May 2009, pp. 263–84. [CrossRef]
- Johnson, Renée, and M. Corn. Bee Health: The Role of Pesticides. 2012. Available online: https://www.semanticscholar.org/paper/Bee-Health%3A-The-Role-of-Pesticides-Johnson-Corn/aeefdfda72ddceb6dac765cfa8b3c3dea6d5891a (accessed on 24 September 2024).
- Lipiński Z. Honey Bee Nutrition and Feeding: In the Temperate / Continental Climate of the Northern Hemisphere. Edited by Richard Jones, 1st. ed., Z. Lipiński, 2018.
- De Graaf, Dirk C., et al. “Standard Methods for American Foulbrood Research.” Journal of Apicultural Research, vol. 52, no. 1, 2013, pp. 1–28. [CrossRef]
- Ehtiati, Sajjad, et al. “Promising Influences of Caffeic Acid and Caffeic Acid Phenethyl Ester against Natural and Chemical Toxins: A Comprehensive and Mechanistic Review.” Journal of Functional Foods, vol. 107, Aug. 2023, p. 105637. [CrossRef]
- Stephan, J.G., de Miranda, J.R. & Forsgren, E. American foulbrood in a honeybee colony: spore-symptom relationship and feedbacks between disease and colony development. BMC Ecol 20, 15 (2020). [CrossRef]
- Ebeling, J., Knispel, H., Hertlein, G. et al. Biology of Paenibacillus larvae, a deadly pathogen of honey bee larvae. Appl Microbiol Biotechnol 100, 7387–7395 (2016). [CrossRef]
- Karina Antúnez, Matilde Anido, Jay D. Evans, Pablo Zunino. Secreted and immunogenic proteins produced by the honeybee bacterial pathogen, Paenibacillus larvae. Veterinary Microbiology, Volume 141, Issues 3–4, 2010, Pages 385-389, ISSN 0378-1135. [CrossRef]
- Neuendorf, Sandra, et al. “Biochemical Characterization of Different Genotypes of Paenibacillus Larvae Subsp. Larvae, a Honey Bee Bacterial Pathogen.” Microbiology, vol. 150, no. 7, 2004, pp. 2381–90. [CrossRef]
- Genersch EAshiralieva A, Fries I.2005.Strain- and Genotype-Specific Differences in Virulence of Paenibacillus larvae subsp. larvae, a Bacterial Pathogen Causing American Foulbrood Disease in Honeybees. Appl Environ Microbiol 71. [CrossRef]
- Poppinga, Lena, and Elke Genersch. “Molecular Pathogenesis of American Foulbrood: How Paenibacillus Larvae Kills Honey Bee Larvae.” Current Opinion in Insect Science, vol. 10, Aug. 2015, pp. 29–36. [CrossRef]
- Djukic, Marvin, et al. “How to Kill the Honey Bee Larva: Genomic Potential and Virulence Mechanisms of Paenibacillus Larvae.” PLOS ONE, vol. 9, no. 3, Mar. 2014, p. e90914. [CrossRef]
- Hornitzky, M. A. Z. (1998). The pathogenicity of Paenibacillus larvae subsp. larvae spores and vegetative cells to honey bee (Apis mellifera) colonies and their susceptibility to royal jelly. Journal of Apicultural Research, 37(4), 267–271. [CrossRef]
- Ashiralieva, Ainura, and Elke Genersch. “Reclassification, Genotypes and Virulence of Paenibacillus Larvae, the Etiological Agent of American Foulbrood in Honeybees—a Review.” Apidologie, vol. 37, no. 4, July 2006, pp. 411–20. [CrossRef]
- Antúnez, Karina, et al. “Paenibacillus Larvae Enolase as a Virulence Factor in Honeybee Larvae Infection.” Veterinary Microbiology, vol. 147, no. 1, Jan. 2011, pp. 83–89. [CrossRef]
- Beims, Hannes, et al. “Discovery of Paenibacillus Larvae ERIC V: Phenotypic and Genomic Comparison to Genotypes ERIC I-IV Reveal Different Inventories of Virulence Factors Which Correlate with Epidemiological Prevalences of American Foulbrood.” International Journal of Medical Microbiology, vol. 310, no. 2, Feb. 2020, p. 151394. [CrossRef]
- Hansen, Henrik, and Camilla Juul Brødsgaard. “American Foulbrood: A Review of Its Biology, Diagnosis and Control.” Bee World, vol. 80, no. 1, 1999, pp. 5–23. [CrossRef]
- Matović, Kazimir, et al. “American Foulbrood—Old and Always New Challenge.” Veterinary Sciences, vol. 10, no. 3, Mar. 2023, p. 180. [CrossRef]
- D.C. De Graaf, A.M. Alippi, M. Brown, J.D. Evans, M. Feldlaufer, A. Gregorc, M. Hornitzky, S.F. Pernal, D.M.T. Schuch, D. Titĕra, V. Tomkies, W. Ritter, Diagnosis of American foulbrood in honey bees: a synthesis and proposed analytical protocols, Letters in Applied Microbiology, Volume 43, Issue 6, 1 December 2006, Pages 583–590. [CrossRef]
- Spivak, Marla, and Gary S. Reuter. “Resistance to American Foulbrood Disease by Honey Bee Colonies Apis Mellifera Bred for Hygienic Behavior.” Apidologie, vol. 32, no. 6, Nov. 2001, pp. 555–65. [CrossRef]
- Datta, Samik, et al. “Modelling the Spread of American Foulbrood in Honeybees.” Journal of The Royal Society Interface, vol. 10, no. 88, Nov. 2013, p. 20130650. [CrossRef]
- Pohorecka, Krystyna, et al. “Screening of Paenibacillus Larvae Spores in Apiaries from Eastern Poland. Nationwide Survey. Part I.” Bulletin of the Veterinary Institute in Pulawy, vol. 56, no. 4, Dec. 2012, pp. 539–45. [CrossRef]
- Pohorecka, Krystyna & Bober, Andrzej. Occurrence of paenibacillus larvae spores in honey samples domestic apiaries. Journal of Apicultural Science. 2008. 52. 105-111.
- Wilhelm, Elfriede, et al. “Monitoring of Paenibacillus Larvae in Lower Austria through DNA-Based Detection without De-Sporulation: 2018 to 2022.” Veterinary Sciences, vol. 10, no. 3, Mar. 2023, p. 213. [CrossRef]
- Semkiw P. Stan pszczelarstwa w Polsce w 2023 roku. Przygotowanie raportu dotyczącego stanu pszczelarstwa w Polsce oraz analiza jakości (autentyczności/zanieczyszczeń i/lub zafałszowań) miodu importowanego z Ukrainy. Puławy. 2023. Available online: https://www.inhort.pl/wp-content/uploads/2024/01/11.3_Raport_stan_pszczelarstwa_2023.pdf (accessed on 12 September 2024).
- Krajowa Izba Lekarsko-Weterynaryjna. Available online: https://vetpol.org.pl/lista-lekarzy-weterynarii-specjalistow/#choroby-owadow-uzytkowych (accessed on 18 of August 2024).
- Moharrami, M., et al. “Role of Honey Bee Gut Microbiota in the Control of American Foulbrood and European Foulbrood Diseases.” Archives of Razi Institute, vol. 77, no. 4, Aug. 2022, pp. 1331–39. [CrossRef]
- Kuzyšinová, Katarína, et al. “The Use of Probiotics, Essential Oils and Fatty Acids in the Control of American Foulbrood and Other Bee Diseases.” Journal of Apicultural Research, vol. 55, no. 5, Dec. 2016, pp. 386–95. [CrossRef]
- Pejsak, Z., Truszczynski M. “Bioasekuracja—podstawowy sposób ochrony zwierząt przed chorobami zakaźnymi.” Życie Weterynaryjne, vol. 92, no. 06, 2017, http://agro.icm.edu.pl/agro/element/bwmeta1.element.agro-b12476c4-bab1-424e-91d5-62b44e7bbf0e.
- Gillespie T.: Herd health management through prevention and control of pathogens. Prevention starting with biosecurity. Proceedings of the 25 th International Pig Veterinary Society Congress, 11–14.06.2018, Chongqing, China.
- Lewis, D.G., Baker R.B.: Biosecurity of pigs and Farm security Board of Regents of the University of Nebraska, Extension, 2011, 1–31.
- Amass, S.F., Clark L.K.: Biosecurity considerations for pork production units., J. Swine Health Prod. 1999,7, 217–228.
- Dors, A., Czyżewska-Dors E., Pomorska-Mól M., Pejsak Z.: Biosecurity in Polish pig farms—a questionnaire survey. Berl Munch Tierarztl 2017. [CrossRef]
- Morley, Paul S. Biosecurity of veterinary practices. Veterinary Clinics: Food Animal Practice, Volume 18, Issue 1, 133—155.
- Miele, Mara, and John Lever. “Improving Animal Welfare in Europe: Cases of Comparative Bio-Sustainabilities.” Sustainable Food Systems, Routledge, 2014.
- Dewulf, Jeroen, and Filip Van Immerseel, editors. Biosecurity in Animal Production and Veterinary Medicine: From Principles to Practice. CABI, 2019.
- Renault, Véronique, et al. “Biosecurity Concept: Origins, Evolution and Perspectives.” Animals, vol. 12, no. 1, Jan. 2022, p. 63. [CrossRef]
- Huber, Nikolaus, et al. “What Is a Biosecurity Measure? A Definition Proposal for Animal Production and Linked Processing Operations.” One Health, vol. 15, Dec. 2022, p. 100433. [CrossRef]
- Dictionary Cambridge. Available online: https://dictionary.cambridge.org/dictionary/english/biosecurity (accessed on 21 August 2024).
- Kragh KN, Alhede M, Rybtke M, Stavnsberg C, Jensen PØTolker-Nielsen T, Whiteley M, Bjarnsholt T2018.The Inoculation Method Could Impact the Outcome of Microbiological Experiments. Appl Environ Microbiol 84: e02264-17. [CrossRef]
- Gianinazzi, S., et al. “BIOTIZATION OF MICROPLANTS FOR IMPROVED PERFORMANCE.” Acta Horticulturae, no. 625, 2003, pp. 165–72. [CrossRef]
- Barber, M. A. (1911). A Technic for the Inoculation of Bacteria and Other Substances into Living Cells. The Journal of Infectious Diseases, 8(3), 348–360.
- Lopez, M. E. S., et al. “Bio-Sanitization Using Specific Bacteriophages to Control Escherichia Coli O157:H7 in Cherry Tomatoes.” Advance Journal of Food Science and Technology, vol. 16, no. SPL, 2018, pp. 92–101. [CrossRef]
- Jończyk-Matysiak, E.; Owczarek, B.; Popiela, E.; Świtała-Jeleń, K.; Migdał, P.; Cieślik, M.; Łodej, N.; Kula, D.; Neuberg, J.; Hodyra-Stefaniak, K.; et al. Isolation and Characterization of Phages Active against Paenibacillus larvae Causing American Foulbrood in Honeybees in Poland. Viruses 2021, 13, 1217. [Google Scholar] [CrossRef] [PubMed]
- Balouiri, Mounyr, et al. “Methods for in Vitro Evaluating Antimicrobial Activity: A Review.” Journal of Pharmaceutical Analysis, vol. 6, no. 2, Apr. 2016, pp. 71–79. [CrossRef]
- EUCAST. Available online: https://www.eucast.org/ast_of_veterinary_pathogens (accessed on 24 August 2024).
- Hoover, DALLAS G., and SUSAN K. Harlander. “CHAPTER 2—Screening Methods for Detecting Bacteriocin Activity.” Bacteriocins of Lactic Acid Bacteria, edited by Dallas G. Hoover and Larry R. Steenson, Academic Press, 1993, pp. 23–39. [CrossRef]
- Antimicrobial activity of probiotic microorganisms: mechanisms of interaction and methods of examination—Scientific Figure on ResearchGate. Available online: https://www.researchgate.net/figure/Agar-well-diffusion-antimicrobial-assay_fig4_334535091 (accessed on 27 September 2024).
- Murray, Patrick R. “16—The Clinician and the Microbiology Laboratory.” Mandell, Douglas, and Bennett’s Principles and Practice of Infectious Diseases (Eighth Edition), edited by John E. Bennett et al., W.B. Saunders, 2015, pp. 191–223. [CrossRef]
- Hossain, Md Lokman, et al. “A Review of Commonly Used Methodologies for Assessing the Antibacterial Activity of Honey and Honey Products.” Antibiotics, vol. 11, no. 7, July 2022, p. 975. [CrossRef]
- Bubonja-Šonje M, Knežević S, Abram M. Challenges to antimicrobial susceptibility testing of plant-derived polyphenolic compounds. Arh Hig Rada Toksikol. 2020 Dec 31;71(4):300-311. [CrossRef] [PubMed] [PubMed Central]
- Cuenca-Estrella, Manuel, et al. “Comparative Evaluation of NCCLS M27-A and EUCAST Broth Microdilution Procedures for Antifungal Susceptibility Testing of Candida Species.” Antimicrobial Agents and Chemotherapy, vol. 46, no. 11, 2002, pp. 3644–47. [CrossRef]
- Arendrup, M. C., et al. “Comparison of EUCAST and CLSI Reference Microdilution MICs of Eight Antifungal Compounds for Candida Auris and Associated Tentative Epidemiological Cutoff Values.” Antimicrobial Agents and Chemotherapy, vol. 61, no. 6, 2017, pp. e00485-17. [CrossRef]
- Cuenca-Estrella, Manuel, et al. “Comparison of the Vitek 2 Antifungal Susceptibility System with the Clinical and Laboratory Standards Institute (CLSI) and European Committee on Antimicrobial Susceptibility Testing (EUCAST) Broth Microdilution Reference Methods and with the Sensititre YeastOne and Etest Techniques for In Vitro Detection of Antifungal Resistance in Yeast Isolates.” Journal of Clinical Microbiology, vol. 48, no. 5, 2010, pp. 1782–86. [CrossRef]
- Ceballos-Garzon, Andrés, et al. “Head-to-head Comparison of CLSI, EUCAST, Etest and VITEK®2 Results for Candida Auris Susceptibility Testing.” International Journal of Antimicrobial Agents, vol. 59, no. 4, Apr. 2022, p. 106558. [CrossRef]
- Ejdys, Joanna, and Danuta Szpilko. “European Green Deal — Research Directions. a Systematic Literature Review.” Ekonomia i Środowisko—Economics and Environment, vol. 81, no. 2, July 2022, pp. 8–38. [CrossRef]
- Cuadros-Casanova, Ivon, et al. “Opportunities and Challenges for Common Agricultural Policy Reform to Support the European Green Deal.” Conservation Biology, vol. 37, no. 3, 2023, p. e14052. [CrossRef]
- Belloy, Luc, et al. “Spatial Distribution of Melissococcus Plutonius in Adult Honey Bees Collected from Apiaries and Colonies with and without Symptoms of European Foulbrood.” Apidologie, vol. 38, no. 2, Mar. 2007, pp. 136–40. [CrossRef]
- Arai, Rie, et al. “Diversity of Melissococcus Plutonius from Honeybee Larvae in Japan and Experimental Reproduction of European Foulbrood with Cultured Atypical Isolates.” PLOS ONE, vol. 7, no. 3, Mar. 2012, p. e33708. [CrossRef]
- Lewkowski, Oleg, and Silvio Erler. “Virulence of Melissococcus Plutonius and Secondary Invaders Associated with European Foulbrood Disease of the Honey Bee.” Microbiology Open, vol. 8, no. 3, 2019, p. e00649. [CrossRef]
- Van Der Steen, Jozef, and Flemming Vejsnæs. “Varroa Control: A Brief Overview of Available Methods.” Bee World, vol. 98, no. 2, Apr. 2021, pp. 50–56. [CrossRef]
- Chávarri, María, et al. “Chapter 17—Secondary Metabolites From Probiotic Metabolism.” Advances in Probiotics, edited by Dharumadurai Dhanasekaran and Alwarappan Sankaranarayanan, Academic Press, 2021, pp. 259–76. [CrossRef]
- Tejero-Sariñena, Sandra, et al. “In Vitro Evaluation of the Antimicrobial Activity of a Range of Probiotics against Pathogens: Evidence for the Effects of Organic Acids.” Anaerobe, vol. 18, no. 5, Oct. 2012, pp. 530–38. [CrossRef]
- Licciardi, Paul V., et al. “Epigenome Targeting by Probiotic Metabolites.” Gut Pathogens, vol. 2, no. 1, Dec. 2010, p. 24. [CrossRef]
- Smutin, Daniil, et al. “Micro”bee”ota: Honey Bee Normal Microbiota as a Part of Superorganism.” Microorganisms, vol. 10, no. 12, Dec. 2022, p. 2359. [CrossRef]
- Motta, Erick V. S., and Nancy A. Moran. “The Honeybee Microbiota and Its Impact on Health and Disease.” Nature Reviews Microbiology, vol. 22, no. 3, Mar. 2024, pp. 122–37. [CrossRef]
- Moran, Nancy A. “Genomics of the Honey Bee Microbiome.” Current Opinion in Insect Science, vol. 10, Aug. 2015, pp. 22–28. [CrossRef]
- Panwar, Harsh, et al. “Probiotics as Potential Biotherapeutics in the Management of Type 2 Diabetes—Prospects and Perspectives.” Diabetes/Metabolism Research and Reviews, vol. 29, no. 2, Feb. 2013, pp. 103–12. [CrossRef]
- Lamei, Sepideh, et al. “The Secretome of Honey Bee-Specific Lactic Acid Bacteria Inhibits Paenibacillus Larvae Growth.” Journal of Apicultural Research, vol. 58, no. 3, May 2019, pp. 405–12. [CrossRef]
- Lamei, Sepideh, et al. “Improvement of Identification Methods for Honeybee Specific Lactic Acid Bacteria; Future Approaches.” PLOS ONE, vol. 12, no. 3, Mar. 2017, p. e0174614. [CrossRef]
- Forsgren, Eva, et al. “Novel Lactic Acid Bacteria Inhibiting Paenibacillus Larvae in Honey Bee Larvae.” Apidologie, vol. 41, no. 1, Jan. 2010, pp. 99–108. [CrossRef]
- Alippi, Adriana M., and Francisco J. Reynaldi. “Inhibition of the Growth of Paenibacillus Larvae, the Causal Agent of American Foulbrood of Honeybees, by Selected Strains of Aerobic Spore-Forming Bacteria Isolated from Apiarian Sources.” Journal of Invertebrate Pathology, vol. 91, no. 3, Mar. 2006, pp. 141–46. [CrossRef]
- Sabaté, D. C. , et al. “Synergistic Effect of Surfactin from Bacillus Subtilis C4 and Achyrocline Satureioides Extracts on the Viability of Paenibacillus Larvae.” World Journal of Microbiology and Biotechnology, vol. 28, no. 4, Apr. 2012, pp. 1415–22. [CrossRef]
- Bartel, Laura Cecilia, et al. “Bacillus and Brevibacillus Strains as Potential Antagonists of Paenibacillus Larvae and Ascosphaera Apis.” Journal of Apicultural Research, vol. 58, no. 1, Jan. 2019, pp. 117–32. [CrossRef]
- Al-Ghamdi, Ahmad, et al. “Effect of Gut Bacterial Isolates from Apis Mellifera Jemenitica on Paenibacillus Larvae Infected Bee Larvae.” Saudi Journal of Biological Sciences, vol. 25, no. 2, Feb. 2018, pp. 383–87. [CrossRef]
- Al-Ghamdi, Ahmad, et al. “In Vitro Antagonistic Potential of Gut Bacteria Isolated from Indigenous Honey Bee Race of Saudi Arabia against Paenibacillus Larvae.” Journal of Apicultural Research, vol. 59, no. 5, Oct. 2020, pp. 825–33. [CrossRef]
- Nguyen, Tuan Manh, et al. “Further Finding of Antimicribial and Antifungal Activities against Human Pathegens by Using the Isolates to Inhibit the Growth of Paenibacillus Lavae, an Agent of AFB.” Journal of Apiculture, vol. 30, no. 1, Apr. 2015, p. 37. [CrossRef]
- Grubbs, Kirk J., et al. “Pollen Streptomyces Produce Antibiotic That Inhibits the Honey Bee Pathogen Paenibacillus Larvae.” Frontiers in Microbiology, vol. 12, Feb. 2021. [CrossRef]
- Jamal, Qazi Mohammad Sajid, and Varish Ahmad. “Lysinibacilli: A Biological Factories Intended for Bio-Insecticidal, Bio-Control, and Bioremediation Activities.” Journal of Fungi, vol. 8, no. 12, Dec. 2022, p. 1288. [CrossRef]
- Grabek-Lejko, Dorota, and Mariusz Worek. “Honeydew Honey as a Reservoir of Bacteria with Antibacterial and Probiotic Properties.” Antibiotics, vol. 13, no. 9, Sept. 2024, p. 855. [CrossRef]
- Bielik, Bohumil, et al. “Biofilm-Forming Lactic Acid Bacteria of Honey Bee Origin Intended for Potential Probiotic Use.” Acta Veterinaria Hungarica, vol. 68, no. 4, Jan. 2021, pp. 345–53. [CrossRef]
- Leska, Aleksandra, et al. “Characterization and Protective Properties of Lactic Acid Bacteria Intended to Be Used in Probiotic Preparation for Honeybees (Apis Mellifera L.)—An In Vitro Study.” Animals, vol. 13, no. 6, Jan. 2023, p. 1059. [CrossRef]
- Tejerina, Marcos Raúl, et al. “Bacterial Strains Isolated from Stingless Bee Workers Inhibit the Growth of Apis Mellifera Pathogens.” Current Microbiology, vol. 81, no. 4, Feb. 2024, p. 106. [CrossRef]
- Mudroňová, D. , Toporčák, J., Nemcová, R., Gancarčíková, S., Hajdučková, V., & Rumanovská, K. Lactobacillus sp. as a potential probiotic for the prevention of Paenibacillus larvae infection in honey bees. Journal of Apicultural Research, 2011, 50, 323–324. [Google Scholar] [CrossRef]
Figure 1.
Radar plot illustrating the distribution of tested inoculants with antimicrobial potential against Paenibacillus larvae ATCC 9545 ser. Eric I using the well-diffusion method [mm] according to EUCAST standards. Volumetric concentrations of 3%, 4%, 5%, and 10% were considered, the symbol ‘TP’ means tryptone-peptone medium, the symbol ‘M’ means reed molasses medium, the symbol ‘50% S’ means a 50% sucrose solution, the symbol ‘60% S’ means a 60% sucrose solution, the symbol ‘Mix’ means a consortium of all functional strains for a given experimental system.
Figure 1.
Radar plot illustrating the distribution of tested inoculants with antimicrobial potential against Paenibacillus larvae ATCC 9545 ser. Eric I using the well-diffusion method [mm] according to EUCAST standards. Volumetric concentrations of 3%, 4%, 5%, and 10% were considered, the symbol ‘TP’ means tryptone-peptone medium, the symbol ‘M’ means reed molasses medium, the symbol ‘50% S’ means a 50% sucrose solution, the symbol ‘60% S’ means a 60% sucrose solution, the symbol ‘Mix’ means a consortium of all functional strains for a given experimental system.
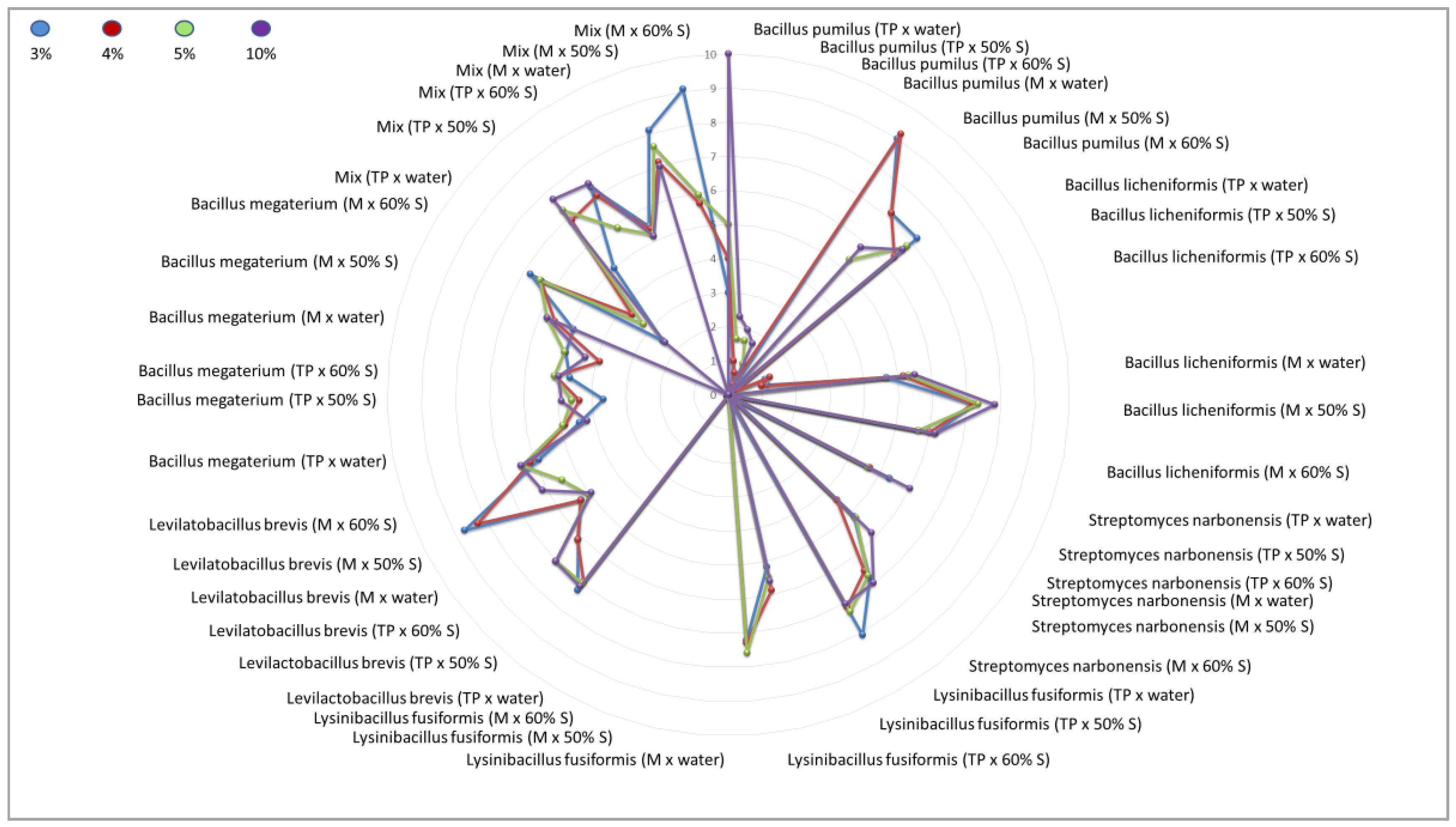
Figure 2.
Radar plot illustrating the distribution of tested inoculants with antimicrobial potential against Paenibacillus larvae CCUG 48973 ser. Eric II uses the well-diffusion method [mm] according to EUCAST standards. Volumetric concentrations of 3%, 4%, 5%, and 10% were considered, the symbol ‘TP’ means tryptone-peptone medium, the symbol ‘M’ means reed molasses medium, the symbol ‘50% S’ means a 50% sucrose solution, the symbol ‘60% S’ means a 60% sucrose solution, the symbol ‘Mix’ means a consortium of all functional strains for a given experimental system.
Figure 2.
Radar plot illustrating the distribution of tested inoculants with antimicrobial potential against Paenibacillus larvae CCUG 48973 ser. Eric II uses the well-diffusion method [mm] according to EUCAST standards. Volumetric concentrations of 3%, 4%, 5%, and 10% were considered, the symbol ‘TP’ means tryptone-peptone medium, the symbol ‘M’ means reed molasses medium, the symbol ‘50% S’ means a 50% sucrose solution, the symbol ‘60% S’ means a 60% sucrose solution, the symbol ‘Mix’ means a consortium of all functional strains for a given experimental system.
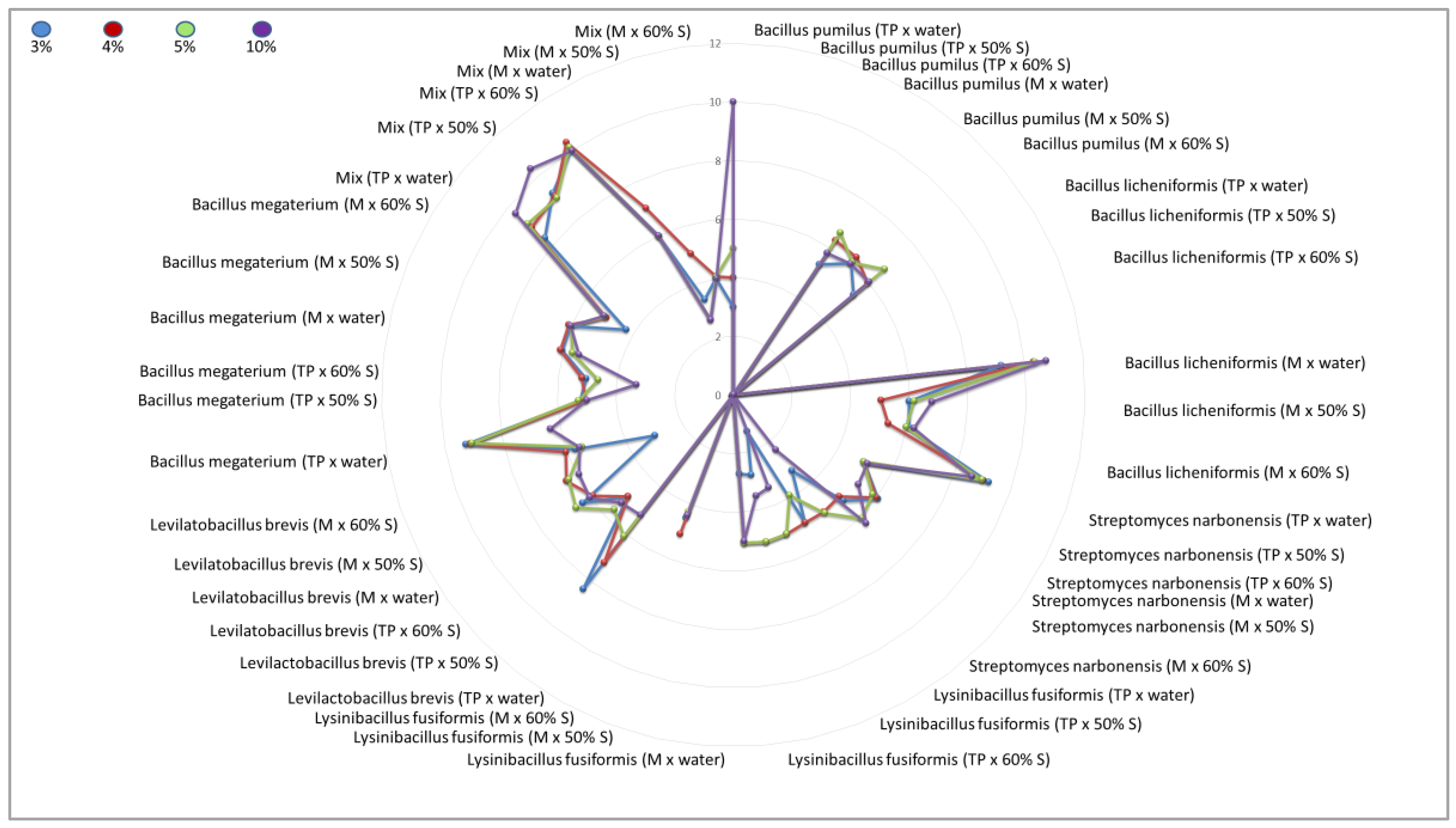

Table 1.
Reactor culture parameters for Bacillus pumilus R5 using: BPW (buffered peptone water), PCA (plate count agar), DRBC (dichloran rose bengal agar), MRS (De Man–Rogosa–Sharpe), ISA (iron sulfite agar), where symbol ‘G’ means growth and ‘NG’ means no growth.
Table 1.
Reactor culture parameters for Bacillus pumilus R5 using: BPW (buffered peptone water), PCA (plate count agar), DRBC (dichloran rose bengal agar), MRS (De Man–Rogosa–Sharpe), ISA (iron sulfite agar), where symbol ‘G’ means growth and ‘NG’ means no growth.
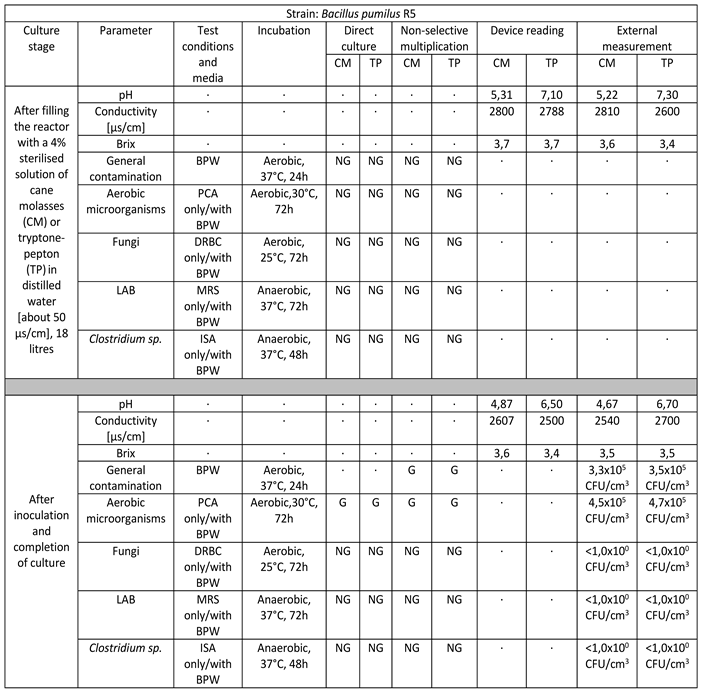 |
Table 2.
Reactor culture parameters for Bacillus licheniformis M/542/M/18 using: BPW (buffered peptone water), PCA (plate count agar), DRBC (dichloran rose bengal agar), MRS (De Man–Rogosa–Sharpe), ISA (iron sulfite agar), where symbol ‘G’ means growth and ‘NG’ means no growth.
Table 2.
Reactor culture parameters for Bacillus licheniformis M/542/M/18 using: BPW (buffered peptone water), PCA (plate count agar), DRBC (dichloran rose bengal agar), MRS (De Man–Rogosa–Sharpe), ISA (iron sulfite agar), where symbol ‘G’ means growth and ‘NG’ means no growth.
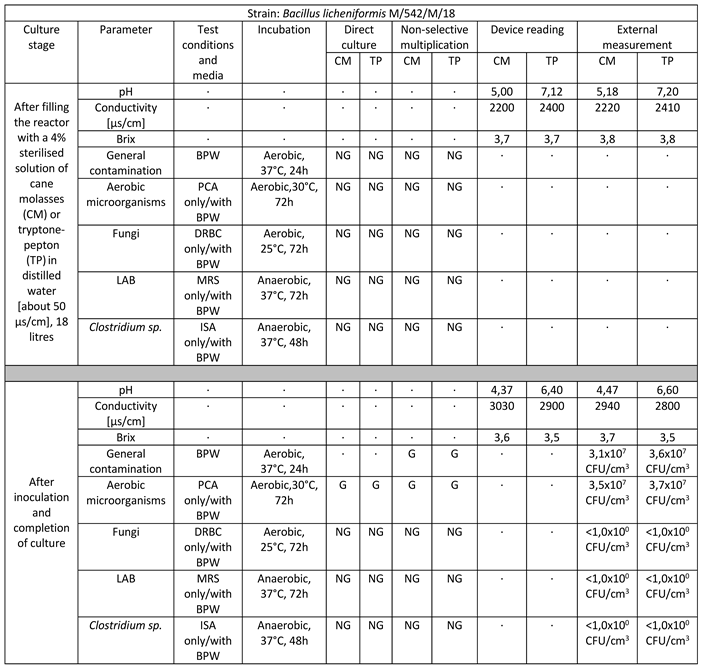 |
Table 3.
Reactor culture parameters for Streptomyces narbonensis R71 using: BPW (buffered peptone water), PCA (plate count agar), DRBC (dichloran rose bengal agar), MRS (De Man–Rogosa–Sharpe), ISA (iron sulfite agar), where symbol ‘G’ means growth and ‘NG’ means no growth.
Table 3.
Reactor culture parameters for Streptomyces narbonensis R71 using: BPW (buffered peptone water), PCA (plate count agar), DRBC (dichloran rose bengal agar), MRS (De Man–Rogosa–Sharpe), ISA (iron sulfite agar), where symbol ‘G’ means growth and ‘NG’ means no growth.
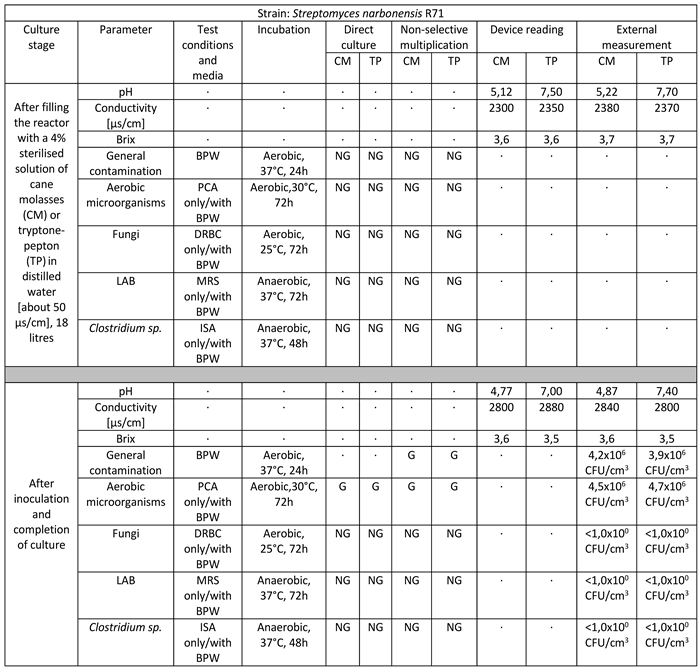 |
Table 4.
Reactor culture parameters for Lysinibacillus fusiformis E23 using: BPW (buffered peptone water), PCA (plate count agar), DRBC (dichloran rose bengal agar), MRS (De Man–Rogosa–Sharpe), ISA (iron sulfite agar), where symbol ‘G’ means growth and ‘NG’ means no growth.
Table 4.
Reactor culture parameters for Lysinibacillus fusiformis E23 using: BPW (buffered peptone water), PCA (plate count agar), DRBC (dichloran rose bengal agar), MRS (De Man–Rogosa–Sharpe), ISA (iron sulfite agar), where symbol ‘G’ means growth and ‘NG’ means no growth.
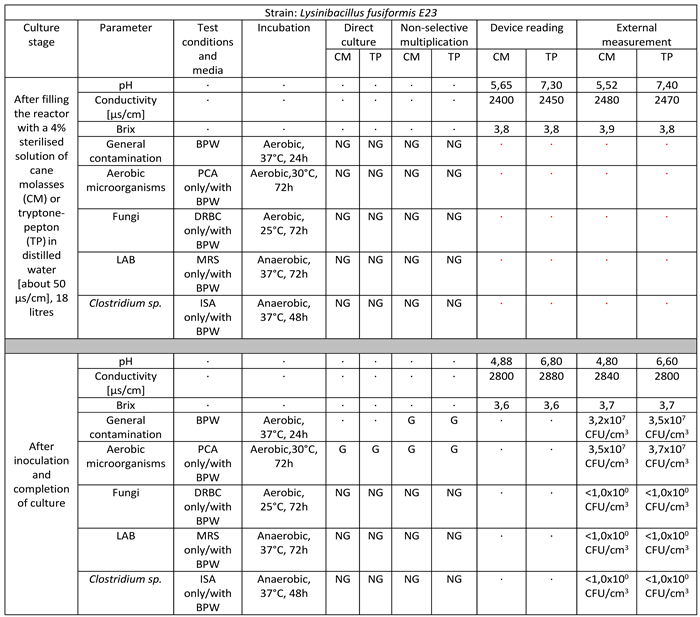 |
Table 5.
Reactor culture parameters for Levilactobacillus brevis M/495/M/17 using: BPW (buffered peptone water), PCA (plate count agar), DRBC (dichloran rose bengal agar), MRS (De Man–Rogosa–Sharpe), ISA (iron sulfite agar), where symbol ‘G’ means growth and ‘NG’ means no growth.
Table 5.
Reactor culture parameters for Levilactobacillus brevis M/495/M/17 using: BPW (buffered peptone water), PCA (plate count agar), DRBC (dichloran rose bengal agar), MRS (De Man–Rogosa–Sharpe), ISA (iron sulfite agar), where symbol ‘G’ means growth and ‘NG’ means no growth.
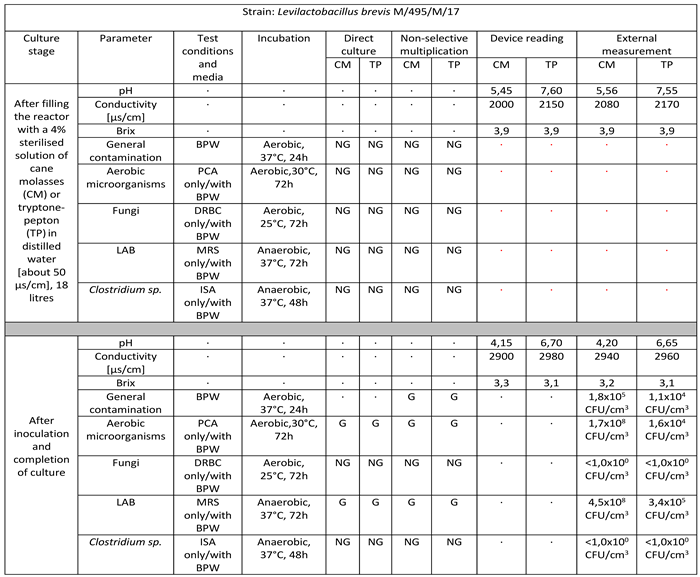 |
Table 6.
Reactor culture parameters for Bacillus megaterium R7 using: BPW (buffered peptone water), PCA (plate count agar), DRBC (dichloran rose bengal agar), MRS (De Man–Rogosa–Sharpe), ISA (iron sulfite agar), where symbol ‘G’ means growth and ‘NG’ means no growth.
Table 6.
Reactor culture parameters for Bacillus megaterium R7 using: BPW (buffered peptone water), PCA (plate count agar), DRBC (dichloran rose bengal agar), MRS (De Man–Rogosa–Sharpe), ISA (iron sulfite agar), where symbol ‘G’ means growth and ‘NG’ means no growth.
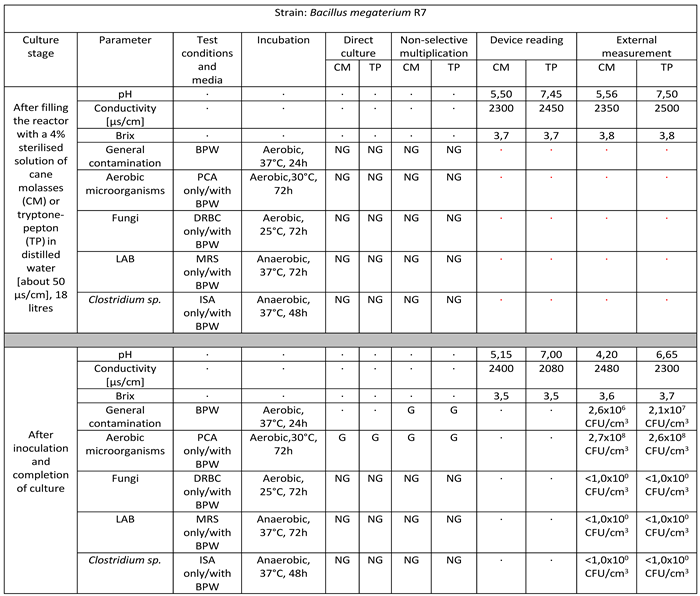 |
Table 7.
Data show antimicrobial activity against Paenibacillus larvae ATCC 9545 (ser. Eric I), using the well-diffusion method according to EUCAST standards [mm] with heat map effect (red color means low antimicrobial activity or lack of antimicrobial properties, green color means high activity).
Table 7.
Data show antimicrobial activity against Paenibacillus larvae ATCC 9545 (ser. Eric I), using the well-diffusion method according to EUCAST standards [mm] with heat map effect (red color means low antimicrobial activity or lack of antimicrobial properties, green color means high activity).
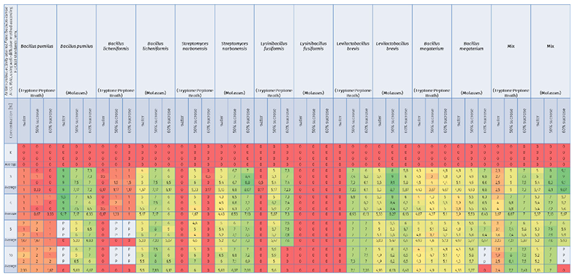 |
Table 8.
Data show antimicrobial activity against Paenibacillus larvae CCUG 48973 (ser. Eric II), using the well-diffusion method according to EUCAST standards [mm] with heat map effect (red color means low antimicrobial activity or lack of antimicrobial properties, green color means high activity).
Table 8.
Data show antimicrobial activity against Paenibacillus larvae CCUG 48973 (ser. Eric II), using the well-diffusion method according to EUCAST standards [mm] with heat map effect (red color means low antimicrobial activity or lack of antimicrobial properties, green color means high activity).
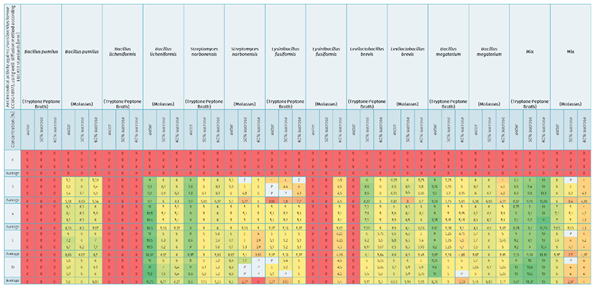 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



