
Submitted:
28 September 2024
Posted:
30 September 2024
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
Amidst the ongoing global challenge of the SARS-CoV-2 pandemic, the quest for effective antiviral medications remains paramount. This comprehensive review delves into the dynamic landscape of FDA-approved medications repurposed for COVID-19, categorized as antiviral and non-antiviral agents. Our focus extends beyond conventional narratives, encompassing vaccination targets, repurposing efficacy, clinical studies, innovative treatment modalities, and future outlooks. Unveiling the genomic intricacies of SARS-CoV-2 variants, including the WHO-designated Omicron variant, we explore diverse antiviral categories such as Fusion inhibitors, Protease inhibitors, Transcription inhibitors, Neuraminidase inhibitors, Nucleoside reverse transcriptase, and non-antiviral interventions like Importin α/β1-mediated nuclear import inhibitors, Neutralizing antibodies and convalescent plasma. Notably, Molnupiravir emerges as a pivotal player, now licensed in the UK. This review offers a fresh perspective on the historical evolution of COVID-19 therapeutics, from repurposing endeavors to the latest developments in oral anti-SARS-CoV-2 treatments, ushering in a new era of hope in the battle against the pandemic.
Keywords:
Molnupiravir
; SARS-CoV-2
; Drug repurposing
; Vaccines
; Variant of concerns (VOC)
; Paxlovid
1. Introduction
Coronaviruses (CoVs) are enveloped RNA viruses and are members of the order Nidovirales' Coronaviridae family that cause respiratory, hepatic, neurological, and intestinal disease. [1] Four endemic coronaviruses in humans, known as CoV-229E, CoV-OC43, CoV-NL63, and CoV-HKU1, are frequently linked to mild respiratory illness in healthy people. [2] Coronaviruses, such as the Middle East Respiratory Syndrome (MERS-CoVs), Severe Acute Respiratory Syndrome (SARS-CoV), [3] and, eventually, the new Severe Acute Respiratory Syndrome (SARS-CoV-2) that cause COVID-19, are all examples of deadly outbreaks caused by coronaviruses. [4,5,6,7,8] COVID-19 was proposed to be detected in SARS-CoV-2 in Wuhan, China, by December 2019. [3,4,9,10,11,12] As a result, the World Health Organization (WHO) directed various tests about the recent onset of this outbreak. [12,13] The outbreak, which was caused by COVID-19's initial appearance, came from an unidentified animal source at market, [12] but evidence suggested that zoonotic species, [14] including intermediate hosts such as bats, were the main source of the infection. [10] SARS-CoV-2 is a positive single-RNA stranded virus that has the potential to infect humans or animals. SARS-CoV-2 belongs to the beta-CoV subfamily, one of the four -CoV subfamilies; gamma, delta, alpha, and beta. The beta and alpha CoV viruses attack mammals, but the delta and gamma COV viruses only affect birds. [8,15,16] SARS-CoV-2 exhibits persistent transmission from person to person via direct/indirect contact and through the environment as respiratory droplets and/or aerosols. [17,18,19] The research on SARS-CoV-2 suggests that the majority of cases could be caused by relatively low viral loads, with different symptoms ranging and lasting a variety of times. [20] The onset of more severe virus symptoms with a larger load can be seen in common symptoms such as fever and cough. [3,14] Some symptoms, including fever, inflammatory reactions, pneumonia, and hypoxemia, may manifest as the illness worsens. Most COVID-19 patients either show no symptoms or have mild illness, and respiratory patients should therefore visit hospitals. [5,20,21,22] Since December 2019, COVID-19 has been a significant public concern around the globe. On November 9th, 2021, over 250 million of COVID-19 patients were reported in 224 countries and territories. [23] The discovery of this novel virus has inspired researchers to create new vaccinations and test them to ensure their efficiency. [24] It is crucial to draw attention to the SARS-CoV-2 variations discovered in the genomes of SARS-CoV-2 virions. These variations are anticipated to have an advantageous effect on the phenotype of the virus in some circumstances. Such modifications may have an impact on the pathogenicity, transmissibility, infectiousness, as well as antigenicity of viruses. [25] On November 24, 2021, the World Health Organization classified the SARS-CoV-2 Omicron variant B.1.1.529, which appeared in South Africa, as a variation under monitoring (VUM). Two days later, the Omicron version was classified as a variant of concern (VOC). This variant has several mutations, with about 15 changes to the spike receptor-binding domain (RBD). [26]
There have been various proposed techniques to combat SARS-CoV-2. Among these, targeting ACE2 either directly through supplements or inadvertently through medications. [27] Additionally, the main protease (Mpro) has also been targeted to combat SARS-COV-2. [7,28] Based on the preceding, it is crucial to gather and comprehend the earlier articles written about the attempts and trials made to resist COVID-19. [29,30] This article evaluates the majority of previously repurposed FDA-approved medications, their clinical studies, and the most recent possibilities for fighting SARS-CoV-2. This article could help researchers from around the globe in developing a comprehensive understanding of this pandemic and potential therapeutical approaches.
2. SARS-CoV-2 Structure, Life Cycle and Mechanism of Action
In the replication cycle of SARS-CoV-2 (Figure 1), the virus enters the host cell either through the fusion of its viral envelope with the host cell membrane or via an endosomal route. To initiate viral entry, the receptor-binding domain (RBD) of the spike (S) protein binds to the human host cell receptor (ACE2). [31,32] Subsequent to the RBD interaction with the receptor, the S protein undergoes proteolytic cleavage, facilitated by various host proteases such as TMPRSS2, furin, and cathepsin B/L. Following entry, SARS-CoV-2 releases its genomic RNA into the cytoplasm, utilizing both host cell and intrinsic enzymatic machinery to replicate its genetic material and form new viral particles.
The viral RNA genome is initially translated into viral replicase polyproteins (pp1a and pp1ab), which are then cleaved into 16 non-structural proteins (nsps). [33] In the genome replication and transcription process, mediated by the replication transcription complex (RTC), negative-sense (- sense) genomic RNA is synthesized. This RNA serves as a template for generating positive-sense (+ sense) genomic RNA and subgenomic RNAs. The interaction of the viral genomic RNA with structural proteins located in the endoplasmic reticulum (ER) and ER-Golgi intermediate compartment facilitates viral formation (ERGIC). Eventually, these virions are exocytotically secreted from the infected cell and discharged to the plasma membrane through deacidified lysosomes.
The human structure of COVID-19 consists of the innermost layer of SARS-CoV-2 and nucleocapsid (N) protein, which makes up the viral genome. As previously mentioned, SARS-CoV-2 is an RNA-positive virus that uses a ribosomal host to translate its genome as soon as it enters the cell. There are 29 different proteins encoded in the genome as a result. The envelope (E) protein is present in the nucleocapsid, while the membrane (M) protein is present across the entire virus envelope. In addition, the virus uses the spike (S) protein in the enclosed membrane on its surface when attached to host cells. Additionally, oesophagal cells, alveolar cells, cardiac cells, proximal kidney cells, and absorbent enterocytes exhibit angiotensin-converting enzyme-2 (ACE2) receptors as the other cell types do. [34] As a result, the (S) protein primes transmembrane serine protease 2 (TMPRSS2), prevents SARS-CoV-2 infection in lung cells. In TMPRSS2-negative cells, [35] the cysteine proteases cathepsin B/L can facilitate the cleavage of the S-protein. There are 7 non-structural proteins; (ORF)7a, PLpro, Nsp12, Nsp3c, Mpro, Nsp1, and Nsp13, and 4 structural proteins that SARS-CoV-2 encodes; M, S, N, and E. [36] A crucial goal for the development of novel antiviral drugs is the understanding of the fundamental components and biochemical mechanisms involved in the coronavirus replication cycle, including RNA-dependent RNA polymerases and proteolytic enzymes.Respiratory droplets mostly spread [37] SARS-CoV-2 in people, but it has also the potential to spread into the air. [38] The virus enters the host cells either by the endosomes or through plasma membrane. It interacts with ACE2 as an entry sensor, and the viral S protein, which facilitates attachment to the host cell membrane through both pathways. [35,39] According to a recent study, the host protease TMPRSS2 activates the connection between the S protein and the ACE2 protein. In order to easily connect to the host receptors, the virus employs the S protein to deactivate antibodies. [40] Beta coronaviruses typically employ hemagglutinin esterase (HE) to bind to sialic acid present on the glycoprotein surface, although the exact fusion mechanism of SARS-CoV-2 is not fully understood. To impede these fusion stages. [41] fusion inhibitors can be employed. Following the completion of fusion, the envelope is shed, allowing the SARS-CoV-2 genome, along with its nucleocapsid, to enter the host cell's cytoplasm. [35] Within its genome are open reading frames ORF1a and ORF1b genes, generating two polyproteins (pp), pp1a and pp1b. These polyproteins, assisted by host ribosomes. [42] play a role in viral translation. Subsequently, papain-like protease (Ppro) and Mpro cleave the polyproteins, yielding various non-structural proteins. Based on a three-dimensional analytical model, it is suggested that, in addition to Ppro and Mpro, the presence of the 3C-like cysteine protease (3CLPro) in SARS-CoV-2 is likely, given its similarity to SARS-CoV, approximately 96%. [43] These proteases perform critical roles in transcription and replication of viruses. Furthermore, protease inhibitors that block these proteases have the potential to be antivirals against SARS-CoV-2. [44] The replication mechanism of SARS-CoV-2 based on models of MERS-CoV and SARS-CoV due to the similarity of their non-structural and structural proteins. RNA-dependent RNA polymerases are transcription and replication complexes formed by a non-structural protein known as nsp12 (RdRp). [45] In SARS-CoV, Nsp12 is coupled with its cofactors; nsp7 and nsp8. Using the initial positive RNA as a guide, this protein complex produces a complementary RNA with the opposite sense. [46] Subsequently, the virus utilizes the negative-strand RNA to generate new positive RNA molecules, facilitating the subsequent phases of translation and replication to form the genomes of the latest viral particles. In SARS-CoV, topoisomerase III-beta plays a role in controlling this process. The progression of these stages can be impeded by the budding and assembly of the encapsulated virus through the use of reverse transcription inhibitors. [47] Since post-translational modification is required, the structural protein complexes M, N, E, and S, produce sub-genomic RNA before joining the endoplasmic reticulum. [48] N- structural protein produces a nucleoprotein complex and positive-strand RNA in the cytoplasm. Both complexes work together to replicate the virus in the endoplasmic reticulum-Golgi apparatus compartment (ERGIC). [49] They are released from cells and discharged into the extracellular zone via the vesicles and Golgi apparatus as mature viruses further infecting other cells. [50]
3. Vaccines Targets for SARS-CoV2
Like other viral diseases, vaccination is the leading way to avoid COVID-19. Several vaccine platforms have been created since the SARS-CoV-2 emergency, and as on July, 2022, about 40 vaccines received global approval. Mainly, 196 vaccines are in preclinical trials, while 153 vaccines reached the clinical trials. The currently licenced vaccines are based on protein subunits (n = 16), inactivated virus (n = 11), nonreplicating viral vectors (n = 7), RNA (n = 4), DNA (n = 1), or viruslike particles (VLPs) (n = 1), Figure 2. The WHO issued Emergency Use Listing (EUL) for ten of these vaccinations, which are mentioned. [51,52] Vaccines utilizing protein subunits consist of antigenic fragments from pathogens, effectively preventing human viral infection. [53] However, they lack the full antigenic complexity of the virus, limiting their efficacy as protection may be reduced due to a limited number of viral fragments. [54] Examples of protein subunit vaccines, such as COVOVAX (produced by Serum Institute of India), Novavax formulation, and Nuvaxovid (Novavax), involve the recombinant nanoparticle S protein linked to the Matrix-M adjuvant. The S protein has undergone stabilising modifications designed to address the underlying issue of its conformational instability. [55] The inactivated vaccines such as Covilo (Sinopharm), CoronaVac (Sinovac) and Covaxin (Bharat Biotech), based on the whole virus in cells, followed by chemical inactivation, purification, and then mixing with particular substances that act as immune cell stimulants and immune response amplifiers, like aluminium hydroxide adjuvant. [56] It is known that pathogens which have been radioactively, chemically, or thermally inactivated occasionally lose their immunogenicity, making the platform less effective than those that use live attenuated pathogens. [57] Nonreplicated viral vector vaccines approved for human use rely on animal or human replication-defective adenovirus vectors. Notably, Vaxzevria (Oxford/AstraZeneca) and Covishield, produced through the Oxford and AstraZeneca formulation by Serum Institute of India and Fiocruz-Brazil, are licensed vaccines based on the chimpanzee adenovirus expressing the SARS-CoV-2 S glycoprotein. Additionally, Ad26.COV2.S, licensed by Janssen/Johnson & Johnson, utilizes a recombinant human adenovirus type 26 vector to express the S protein in a stable form. [58] Since RNA-based vaccines have been licenced for use in humans for the first time and have shown excellent safety and effectiveness profiles, this platform is leading the way in the rapid development of vaccinations against emerging cases. [59,60,61] Spikevax (Moderna) and Comirnaty (Pfizer/BioNTech) are nucleoside-modified RNA vaccines formulated with lipid nanoparticles. They encode the full-length SARS-CoV-2 S protein, modified by two proline mutations to maintain the pre-fusion conformation. Despite variances in their engineering processes, both vaccines share this key feature. As per a recent meta-analysis report, out of the majority of vaccines, 81% had an effect against severe disease, still higher than 70% after getting complete vaccination associated with a 10% minimal reduction six months after immunisation. [62] As most of these vaccines were developed using the prototype Wuhan-Hu-1 strain, they are less effective against the variant of concerns (VOCs) that have surfaced since the pandemic's inception. Therefore, to offer the best defence against these SARS-CoV-2 variations, modifications to vaccine composition to reflect the most common variant(s) of SARS-CoV-2 must be considered. Because COVID-19 vaccine-induced immunity is transient, new preventive measures that result in long-term protection are necessary.
4. SARS-CoV2 Variants
Once the SARS-CoV-2 virus was identified in December 2019, clinical samples from around the world have had their viral genomes sequenced daily. As a result, thousands of whole genomes have been uploaded to databanks. Due to the RdRp's limited ability to rectify errors made during genome replication, RNA viruses, including SARSCoV-2, are known to exhibit significant mutation rates. [63] The Coronaviridae family is an outlier, though, as these viruses have exoribonuclease moiety that may perform proofreading as part of their machinery replication. [64] Additionally, SARS-CoV-2 was shown to have this moiety as part of its nsp14 nonstructural protein. [65] Published studies of several genomes revealed that SARS-CoV-2 had a low nucleotide diversity throughout the world. As long as the viral incidence rises, the nucleotide diversity also tends to rise. [66] The variants of SARS-CoV-2, which are genetically distinct from the initial SARS-CoV-2 lineage, have emerged as a result of virus mutation over time. These mutations may benefit the variations carrying them, boosting viral adaptation, or they may be phenotypically neutral, having little to no significant effects on viral biology. [67] The D614G mutation on the S protein was present in the first significant variation of the identified Wuhan reference strain. After being discovered in March 2020, this mutation quickly spread throughout the world and was found in most of the current viral lineages in circulation. D614G mutation benefits the virus during replication, which could account for its quick global spread. [68,69] SARS-CoV-2 variants have been divided into three categories by the CDC: variants of interest (VOIs), variants of concern (VOCs), and variants of high consequence (VOHCs). Variants of interest (VOIs), are mutations that are believed to alter the diagnostics, treatments, transmission, or sensitivity to antibodies created after prior exposure or immunisation. They are variants with low prevalence or dispersal. In order to qualify as variants of concern (VOCs), a substance must show increased incidence or transmission, diagnostic or therapeutic failure, or diminished neutralisation by antibodies developed following prior exposure or immunisation. Variants of high consequence (VOHCs) are those for which medical treatments or preventative measures are proved less effective. Since the pandemic began, SARS-CoV-2 mutation has been shown by the recurrent discovery of several variations. [70] There are five SARSCoV-2 variants, alpha, beta, gamma, delta, and omicron, categorised as VOCs. [70,71,72] CDC has not yet designated any variant as a VOHC. The D614G mutation is present in these variations, contributing to increased virus replication in the upper respiratory tract and higher transmissibility. [69,72] The World Health Organization (WHO) established these variants. [73] The variants of concern (VOCs) are outlined as follows (Figure 3):
The emergence and evolution of SARS-CoV-2 variants have significantly impacted the trajectory of the COVID-19 pandemic. The alpha variant, initially identified in the UK as VUI-202012/01, designated B.1.1.7, and subsequently referred to as alpha by the WHO, demonstrated increased transmissibility, higher viral loads, and a longer infectious period. Associated with elevated mortality, the alpha variant posed challenges for detection due to S gene target failure (SGTF). Notably, it exhibited resistance to certain vaccines and therapeutic monoclonal antibodies. The beta variant, B.1.351, originating in South Africa, showcased enhanced transmissibility and decreased neutralization by both convalescent sera and vaccines. The gamma variant, P.1, identified in Brazil, presented heightened transmissibility, increased viral loads, and potential impacts on herd immunity. The delta variant, B.1.617.2, contributed significantly to global transmission, displaying higher transmissibility and resistance to neutralization by certain antibodies and sera. Lastly, the omicron variant, B.1.1., emerged in Botswana and South Africa, marking the fourth wave of the pandemic. Characterized by distinct biological traits, including strong ACE2 receptor binding, exceptional transmissibility, environmental stability, and resistance to authorized monoclonal antibodies, the omicron variant poses new challenges to global efforts in combating COVID-19. The continuous monitoring of these variants, their interactions with existing treatments and vaccines, and the development of targeted interventions remain imperative for effective pandemic management.
JN.1, which first appeared in Denmark at the end of July, has quickly crossed international borders and been detected in a number of nations, including the United Kingdom, Canada, the United States, South Africa, Portugal and Sweden. Numerous mutations inside the spike gene set this version apart and further complicate our knowledge of the virus's activity. The WHO has made a noteworthy advancement in the fight against the virus by classifying the unique strain JN.1 as a "variant of interest." This designation highlights the strain's potential importance. [74]
4.1. Alpha SARS-CoV-2 Variant
After epidemiological and genomic surveillance, the UK revealed a SARS-CoV-2 variant in December 2020. It was initially identified as VUI-202012/01 as the first variant under investigation. [75] This variant was dubbed as B.1.1.7 according to Pango lineages; however, the WHO refers to this as alpha. It genetically differs from other variants by 23 nucleotide modifications, including 14 nonsynonymous substitutions, 6 synonymous substitutions, and 3 deletions. [76] The alpha variant, which over time became the most common in the UK and was linked to higher mortality, was believed to be more transmissible than earlier variants. [77,78] Depending on the model used to calculate the statistics, the alpha variation replication has moved from 43% to 90% more than prior variants in the United Kingdom and others. Recently, research has suggested that either a longer infectious period or more viral loads may be responsible for increased transmissibility. [77] Patients who have the alpha variant infection have shown high viral RNA levels and longer-lasting virus positive. [79,80] Due to nucleotide deletion inside the S gene of the viral genome, several commercial RT-qPCR kits targeting the S gene were unable to detect infection with the alpha version, known as S gene target failure (S) SGTF). [81] Funk et al. had observed an increased risk of hospitalisation for European patients, classified age groups as 20–39 and 40–59 years, as well as a higher risk of ICU in the age group of 40–59 years, using the SGTF, which is a method to distinguish the alpha variant from others. [82] Recent research has revealed that SARS-CoV-2, the alpha variant to a significant probability of death of about 60% in the infected patients. [78,83] The alpha variant's S protein mutation has also been associated with decreased neutralisation through the monoclonal antibodies. [84,85] They appear to have mild to no effect on the viral neutralisation from vaccine recipients. [84,85,86,87] It was demonstrated that the efficiency of the mRNA-based vaccines mRNA-1273 (Moderna) and BNT162b2 (BioNTech, Pfizer) against the alpha VOC were comparable to that of the prior variety. [88,89] Alpha variant was shown to be resistant to the inactivated-virus-based vaccines BBV152/COVAXIN (Bharat Biotech), and BBIBP-CorV (Sinopharm) [90,91] whereas CoronaVac (Sinovac) significantly decreased the neutralisation capability against this variation by a 0.5 factor. [91] While the general effectiveness of AZD1222 in both symptomatic and asymptomatic cases remained at 61.7% against the alpha variant and 77.3% against other variants, the nonreplicative-viral-vector-based vaccines, ChAdOx1 nCoV-19/AZD1222 (Oxford, AstraZeneca) and Ad26-COV2.S (Janssen), exhibited diminished neutralization capability against the alpha variant. [92] Ad26-COV2.S has not yet been tested for its effectiveness in avoiding COVID-19 brought on by the alpha version. The Gamaleya Institute's Sputnik V Ad26/Ad5 can still neutralise the alpha form. [93]
4.2. Beta SARS-CoV-2 Variant
Researchers from South Africa described another variation of SARS-CoV-2 that appeared following the initial epidemic wave in the same month of the initially detected alpha variant in the UK, [94] initially known as S501.V2, was referred as B.1.351 by Pango lineages and beta by the WHO. When the beta VOCs was initially identified, it had 31 mutations, four of which were also present in the B.1 variant. There are 21 nonsynonymous mutations among the 27 unique variations reported in this lineage, while 12 have been fixed in the variant population over time. [94] The N501Y alteration on the S protein, critical for viral phenotype, is shared by this developing variation and the alpha VOC. Beta VOC was exhibited to be 50% more transmissible than previously circulating versions. [95] When compared to non-VOCs, beta VOCs were associated with increased risk of hospitalisation in European patients in the categorized aged 40-59 and 60-79 years, as well as ICU with 40-59 year age group, although this did not increase deaths. [82] Beta VOC's decreased sensitivity to neutralisation by recuperating and vaccine-elicited sera appears to be its most significant trait to date. The ability of the mRNA-1273, BNT162b2, BBIBP-CorV, CoronaVac, ChAdOx1, Sputnik V Ad26/Ad5 and nCoV-19/AZD1222 vaccines to neutralise this variation was less effective. [85,86,91,93,96,97,98,99,100] For example, ChAdOx1 nCoV-19/AZD1222, [100] BNT162b2 appears to preserve its efficiency to prevent severe forms of the disease, despite a considerable decline in vaccination efficacy being seen in a population-based investigation. [88] According to assessment report EMA/158424/2021, the vaccine BBV152/COVAXIN and Ad26.COV2.S were evaluated to be effective against beta VOC. [101] This variant decreased neutralising by therapeutic monoclonal antibodies. [86,99] Therefore, the beta VOC needs to be continuously monitored by genetic monitoring as it may be linked to an increase in the frequency of reinfections and the failure of vaccines or treatments.
4.3. Gamma SARS-CoV-2 Variants
Another SARS-CoV-2 variation, known as P.1 (gamma) was discovered in Manaus, Brazil, in December 2020, which may have contributed to a significant rise in COVID-19 prevalence. Initially, this gamma variant was identified by 35 mutations dispersed throughout the entire genome. The S gene contains 10 nonsynonymous mutations, of which 3; K417T, E484 K, and N501Y, are shared with the variant B.1.351 and one, N501Y is shared with both B.1.1.7 and B.1.351 variants. [102] The gamma VOCs were the predominant variety in the city in January 2021 due to their estimated transmissibility, which was 1.7 to 2.5 times more than those of non-gamma variants circulating in Manaus. [102,103] The increased in viral loads was also observed in gamma variant-infected individuals, which may play a role in the more infectious behaviour of this variant. [103] Gamma variant infection was linked to a significant probability of hospitalisation and ICU admission. [82] The emergence of this variant may also be a factor in the reinfection patients [104,105] and the recurrence of disease in regions where previous variants likely contributed to herd immunity. [106] When neutralised by convalescent plasma and therapeutic monoclonal antibodies, the gamma variant is only partially to entirely susceptible. The vaccines mRNA-1273and BNT162b2 fared the best, with slight to moderate declines in their ability to neutralise this variation. [104,107,108,109] A patient case who had the entire BNT162b2 vaccination and experienced modest symptoms following gamma infection was documented. [110] CoronaVac's effectiveness against gamma was estimated, and AZD1222's ability to destroy this virus was diminished. [109]
4.4. Delta SARS-CoV-2 Variant
It is reported in India an upsurge in COVID-19 cases in December 2020 and the first few months of 2021, attributed to the appearance of the SARS-CoV-2 variants B.1.617.1, B.1.617.2, and B.1.617.3 which were transferred to other countries. [111] Delta variant B.1.617.2 created a community transmission chain in many countries around the world and quickly increased its share, becoming the dominant lineage there. [111,112] The delta VOC was predicted to become the main circulating lineage until the development of the omicron variation, with a reproduction number 97% higher than the observed number for non-VOCs and at least 30% more than those for other VOCs. [112] This variant has 12 nonsynonymous mutations, 5 of which are in the S gene. The enhanced transmissibility of the delta variant may be attributed to increased viral loads, presumably due to a faster replication rate than other variants. [113,114,115] The two spike mutations T478K and L452R, present in this variation, are projected to enhance its interaction with ACE2 and may increase the virus's capacity to penetrate human cells, although this prediction has to be verified. A higher likelihood of hospitalisation and disease severity were connected to the delta VOC. [113,116] The delta VOC is resistant to neutralisation by several therapeutic monoclonal antibodies and convalescent sera, as reported for other VOCs. Additionally, The BNT162b2-vaccinated sera showed a diminished ability to neutralise the delta variant, particularly after partial immunisation. Complete BNT162b2 vaccination appears to produce immunity against delta vs. other SARS-CoV-2 variants. [117,118,119,120] In reality, based on studies, [120,121] complete immunisation was equally effective against the delta VOC and the variant B.1.1.7. The protection provided by the mRNA-1273 immunisation against the emergence of symptomatic cases resulting from delta infection was inferior to that provided by B.1.1.7 infection. However, mRNA-based vaccines, BNT162b2 and mRNA-1273 examined collectively effectively prevented the onset of mild to severe disease in vaccinated subjects. [122] In a Chinese population, The effectiveness of several inactivated-virus-based vaccinations, such as CoronaVac's inactivated COVID-19 vaccine, Sinopharm's HB02 and WIV04, and Biokangtai's inactivated COVID-19 vaccine, were assessed collectively. They achieved 100% efficacy against severe diseases and 69.5% efficacy against pneumonia associated with COVID-19. [123] The amount of the delta variation that BBV152/COVAXIN-elicited sera were able to neutralise shows that these sera's potency against this strain was only slightly diminished. [124] Nonreplicative-viral-vector-based vaccinations impaired the ability of sera to neutralise the delta VOC. [86,117,125,126] There is need for more research because the effectiveness of AZD122 as determined by population-based studies varies. [120,121]
4.5. Omicron SARS-CoV-2 Variant
The SARS-CoV-2 omicron variant B.1.1., originally appeared in Botswana and South Africa and has been linked to a significant increase in the COVID-19 pandemic; on November 26, 2021, the WHO categorised it as a VOC. Its appearance has caused the COVID-19 pandemic's 4th wave to spread globally. [127,128] The omicron VOC exhibits distinctive biological traits, such as strong binding to the human ACE2 receptor and great transmissibility. The omicron VOC also exhibits excellent environmental stability, high resistance to monoclonal antibodies that clinical trials have authorised, and immunity produced by natural infection or vaccination. The SARS-CoV-2 omicron variant and a number of its traits have been updated. [127,128]
5. Antiviral Drugs against COVID-19
5.1. Fusion Inhibitors Targeting Spike Protein
5.1.1. Umifenovir (Arbidol)
A short indole derivative called umifenovir, Figure 4, has a variety of anti-RNA and anti-DNA viral effects. They inhibits viral internalisation or attachment and blocks viral penetration into the host cell. [129] It is authorised to prevent and cure influenza A and B infections, in China and Russia. [130,131] However, in vitro studies have shown activity against viral illnesses, including Hepatitis B and C viruses (HBV and HCV) and Ebola. Umifenovir was first authorised for influenza treatment in Russia in 1993. The drug is available in China and Russia for the treatment of upper respiratory influenza A and B infections. [132] Umifenovir is advised for COVID-19 patients in the "Treatment Scheme and New Coronavirus Pneumonia Diagnosis." Adults receive 0.2 g of drug three times per day for ten days. [129] In clinical trials, umifenovir, orally administrated, was found to reduce mortality and viral load in contrast to other unnamed antiviral medications or the interferon-only control group. [133] It includes mild general adverse effects such as, nausea, headaches, raised bilirubin, leukopenia, and high alkaline phosphatase (AKP) and other symptoms like abdominal pain. No notable side effects were observed during treatment, according to retrospective studies to assess the safety and effectiveness of umifenovir therapy in COVID-19 patients. [129]
5.1.2. Camostat Mesylate
Camostat mesylate, figure 4, another drug that targets virus fusion, is a serine protease inhibitor. [134] For target cells entry; SARS-CoV-2 binds to TMPRSS2 and/or ACE-2 receptors within the targeted host cells. [135,136] Camostat mesylate works by inhibiting TMPRSS2. [137] The SARS-CoV-2 spike (S) protein is downregulated to prevent the virus from entering the cell and thereby preventing surface fusion. [138] An earlier study found that camostat mesylate prevented SARS-CoV from entering human epithelial bronchial cells. [139] In vitro studies revealed that camostat mesylate and E-64d, a cysteine protease inhibitor, effectively inhibit SARS-CoV-2 TMPRSS2. [35] In Denmark and Germany, the effectiveness of hydroxychloroquine and camostat mesylate combination therapy has been evaluated. [50] Serine protease inhibitor, nafamostat mesylate, was found to have a 15-fold higher effectiveness for SARS-CoV-2 viral entrance into the host cells. Because nafamostat has more effective antiviral action and a good safety profile, it may be considered a safer alternative to camostat mesylate. Additionally, nafamostat mesylate is used to treat disseminated intravascular coagulation (DIC) with enhanced fibrinolysis seen in COVID-19 patients. [50]
5.2. Protease Inhibitors
5.2.1. Lopinavir
In combination with lopinavir (Figure 5), Ritonavir is used for HIV infection treatment. Additionally, it has been shown that lopinavir inhibits SARSCoV-2 replication with an EC50 of 2.660/1.671 µM ratio. [140] Half-maximal effective concentration (EC50). The number of eosinophils increased among COVID-19 cases after it was administered in China as an emergency medication. [141] Ritonavir and lopinavir, as HIV protease inhibitors, have shown encouraging results as SARS-CoV-2 major protease (Mpro) inhibitors in an in vitro investigation. [11] An earlier investigation revealed that the drug kaletra®, a specific lopinavir/ritonavir combination, has antiviral activity against the SARS-CoV in vitro and clinical studies. [142] As a result, this combination is also employed as a backup plan for COVID-19 patients. An earlier investigation showed that while lopinavir/ritonavir therapy was associated with better outcomes, it did not improve the patient's clinical recovery from COVID-19 infection. [143] Although lopinavir's efficacy for treating COVID-19 has not yet evaluated, but USA, Japan, and Singapore treat COVID-19 patients with such ritonavir/lopinavir combination. The lopinavir/ritonavir efficacy for COVID-19 is now being studied in clinical trials in many countires such as France, Spain, Thailand, China, Hong Kong, Canada, and the United States. [50] Furthermore, according to the WHO, a "solidarity" clinical trial for coronavirus was conducted using an inflammation-regulating molecule, interferon (INF)-β alone or with lopinavir/ritonavir. [140] Additionally, patients with COVID-19 can experience improved clinical symptoms and a lower viral loads when using the lopinavir/ritonavir combination. [133]
5.2.2. Lopinavir/Ritonavir + Ribavirin
Ritonavir (RTV) and lopinavir (LPV), (figure 5) are the protease inhibitors suggested in a combined form (Kaletra®) which is a HIV medication. [20,129] Because lopinavir inhibits the HIV protease, [144] may reduce the virus infection rate and interfere with the development of mature virus particles. Lopinavir has a short half-life and a limited bioavailability. [129] Lopinavir, an HIV protease inhibitor, can affect the maturation of viral particles and reduce the virus's ability to spread. Conversely, Ritonavir blocks the cytochrome CYP3A4 enzyme, slows down the cytochrome P450 and reduces the lopinavir metabolism. The bioavailability of lopinavir in vivo can be increased by administering ritonavir and lopinavir together. [16,129,145] Ribavirin is a guanosine analogue that inhibits the RdRp-mediated elongation of viral RNA chains. [20] The bioavailability and in vivo antiviral activity of lopinavir are markedly increased by the combination of lopinavir and ritonavir. Compared to Remdesivir, prophylactic ritonavir/lopinavir-interferon was somewhat effective at reducing the viral load for MERS-CoV infection in mice. However, there were no significant post-infection effects on acute lung injury, viral load, or lung haemorrhage. [146] 400 mg/100 mg of lopinavir/ritonavir each can be taken every 12 hours for a minimum of 10 days and a maximum of 14 days. The entire dose of 400/100 mg is prescribed for children at a rate of 10 mg/kg for children weighing 15–40 kg and 12 mg/kg for children weighing 7–15 kg. Interferon-β is administered in conjunction with ritonavir/lopinavir on a 44 g for three doses every six days if the history of symptoms is less than seven days. [147]
5.2.3. Lopinavir/Ritonavir + Umifenovir
Compared to lopinavir/ritonavir alone, patients who got the triple combination therapy of lopinavir/ritonavir along with umifenovir demonstrated an advantage in preventing the onset of illness. In addition, the progression of lung injury was dramatically halted by the ritonavir/lopinavir and umifenovir combination. [50]
5.2.4. Danoprevir
Hepatitis C protease activity is effectively inhibited by danoprevir (figure 5). [148,149] By 2018, it had been authorized in China as an oral antiviral drug to treat hepatitis C. As a result, it was called a repurposed drug COVID-19 treatment. A triple combination of pegylated-interferon-α, ritonavir-boosted danoprevir, and ribavirin demonstrated a sustained virologic response within 12 weeks (SVR12) in Chinese patients infected with non-cirrhotic hepatitis C virus. [150] In both phase II and III clinical trials, the SVR12 rate for infected cases approached 99% when utilizing this triple combination of ribavirin, ritonavir-boosted danoprevir, and ravidasvir, an HCV NS5A inhibitor, as part of the entire oral administered therapy for non-cirrhotic HCV. [151]
5.2.5. Darunavir
The treatment of COVID-19 in Italy has been suggested using the anti-HIV medication darunavir. In a regimen, it may be used with cytochrome P-450 inhibitors like cobicistat or ritonavir. Additionally, good anti-proliferative results against SARS-CoVs have been observed in in vitro studies. [152] In Thailand, a clinical investigation is underway to evaluate the effectiveness of darunavir in combination with other agents like hydroxychloroquine and antivirals for coronavirus patients. Additionally, a clinical trial in China explored the combination of cobicistat and darunavir. Consequently, prezcobix®, a fixed-dose combination of cobicistat and darunavir, could potentially be utilized in the treatment of COVID-19 patients. [50] Recently, HIV-positive individuals already on darunavir medication were exposed to COVID-19, prompting interest in the efficacy of this HIV protease inhibitor (146). However, darunavir may not be effective in preventing SARS-CoV-2 infection at the currently recommended dose of 800 mg. [50]
5.2.6. Atazanavir
According to an in-silico investigation, atazanavir is more potent than lopinavir at the binding site in SARS-CoV-2 Mpro. Furthermore, an in vitro investigation demonstrated that atazanavir suppressed SARS-CoV-2 replication. [153] Ritonavir/atazanavir use is linked to higher lipid parameters and glucose absorption when compared to lopinavir/ritonavir use, according to research on HIV-infected individuals. [154] Studies show that Atazanavir could substitute for lopinavir, as Atazanavir when combined with ritonavir, will have the same effect as lopinavir alone. [155]
5.2.7. Saquinavir and Other Protease Inhibitors
Saquinavir (figure 5) and other protease inhibitors, including nelfinavir, amprenavir, indinavir, and nelfinavir, serve similarly against COVID-19 as protease inhibitors due to the similarity between their chemical structures. [156] Saquinavir and indinavir were found to inhibit 3CLpro activity in SARS-CoV-2 in silico investigation. Another study discovered that indinavir, saquinavir, nelfinavir, and amprenavir suppressed SARS-CoV-2 in vitro. [157] Saquinavir, however, provides the most potent inhibition when compared to the other drugs. The drug saquinavir has been used to treat COVID-19 patients in Singapore. In a separate in silico investigation, two more possibilities, raltegravir, and paritaprevir, were investigated and showed inhibitory effects on the SARS-CoV-2 3CLpro. [158] Through current screening of medicinal plant libraries, potent anti-viral phytochemicals that may function as inhibitors against SARS-CoV-2 3CLpro have been discovered. [159,160] As a result, further in-vitro investigations could be used to examine the reported antiviral agents. [50,158]
5.2.8. Niclosamide
Niclosamide, an FDA-approved anthelmintic drug, is employed for the treatment of various viral infections spanning from nanomolar to micromolar concentrations. These infections include Zika virus (ZIKV), hepatitis C virus (HCV), Middle East respiratory syndrome coronavirus (MERS-CoV), severe acute respiratory syndrome coronavirus (SARS-CoV), Ebola virus (EBOV), Chikungunya virus (CHIKV), Japanese encephalitis virus (JEV), and human adenovirus (HAdV). Due to its cost-effectiveness and minimal in vivo toxicity profiles as an FDA-approved drug, niclosamide holds the potential to be a valuable therapeutic agent in combating the escalating COVID-19 pandemic. [161] Screening studies of a small commercialised pharmacological library show that niclosamide might stop the replication of SARS-CoV and viral antigen. [162] Niclosamide prevented the SARS-cytopathic CoV's effect (CPE), which in turn prevented the virus from replicating in Vero E6 cells. The SARS-CoV 3CLpro is a key tool for identifying anti-SARS drugs and plays a significant role in synthesizing polyproteins. [163,164,165] 2-chloro-4-nitroaniline compounds were discovered to be effective SARS-CoV 3CLpro inhibitors. Up to 50 M, niclosamide had no overt inhibitory activity against SARS-CoV 3CLpro. Additionally, after 48 hours post-infection, studies have shown that niclosamide reduces MERS-CoV replication.
5.2.9. Nelfinavir
Nelfinavir is an antiviral drug that uses different mechanisms to target HIV. It is an inhibitor of HIV-1 protease. The mechanism of action of nelfinavir involves binding to the HIV-1 protease active site and inhibiting the processing of functioning proteins required for HIV. According to the in vitro studies conducted during the SARS pandemic, Nelfinavir was identified as a potential agent with SARS-CoV inhibitory effects. [166] Out of the 30 examined drugs, nelfinavir provides the most potent protective effect against SARS-CoV-2. Its efficacy against SARS-CoV or SARS-CoV-2 in humans has not yet been investigated. The recommened dose for HIV is 1250 mg or 750 mg orally twice a day. However, the required dose for COVID-19 therapy is uncertain. [167] Pharmacokinetics of nelfinavir in chronic hepatic disease patients can be varied. [168,169] It causes gastrointestinal intolerance, including nausea, and diarrhoea. [167]
5.2.10. Ebselen
Serving as peroxiredoxin mimic enzyme and glutathione peroxidase, ebselen, (figure 5) is an organoselenium substance that reduces peroxynitrite and hydroperoxides. [170] Ebselen interacts with certain thiols protein via a selenosulfide link, leading to anti-inflammatory, pleiotropic, and antibacterial effects in addition to its antiviral activity. Regarding the SARS-CoV-2 Mpro is a potential therapeutic target, ebselen had been identified as over 10,000 chemicals panel as a prospective inhibitor of Mpro. The role of Ebselen is to identify cysteine proteases in viral and other microbial infections and influences lung inflammation. Ebselen can also be used as a lead target to suppress SARS-CoV-2 Mpro effectively, and in vivo adherent cell studies have shown that it has inhibitory effects. Therefore, COVID-19 attributes possible benefits to the critical pathogenetic goals of modulating cytokines and inflammatory oxidants. [140] Diverse biological pathway targets, such as enzymes linked with inflammatory reactions, were found in early studies of ebselen for clinical diseases. [171,172,173] No adverse effects have been observed in scientific investigations. [174] Eliminating selenium is then excreted and degraded as methylated conjugates and glucuronidated. [140,175] Ebselen also possesses anti-inflammatory activity in vivo, suggesting that it may have a vital role in host responses to viral infections. [129] Furthermore, it prevents the formation of oxidised substances in brain tissue caused by encephalitis HIV and Simian immune deficiency virus encephalitis in macaques. [135] The idea is to use ebselen's therapeutic properties to stop the signs of a neurological HIV-1 infection in the brain.
5.3. RNA-Dependent RNA Polymerase Target, Reverse Transcriptase Inhibitors
5.3.1. Remdesivir (GS-5734, Veklury) (Gilead Sciences)
Remdesivir, depicted in Figure 6, stands as the first authorized drug specifically developed for the treatment of Ebola. [176,197] Ebola, being a single-strand RNA virus, faces inhibition from Remdesivir, which acts as an adenosine analogue. This inhibition targets the viral RNA-dependent RNA polymerase (RdRp), leading to either premature or delayed RNA chain termination. [20,177,178,179] Notably, Remdesivir has demonstrated antiviral activity against respiratory viral infections, including SARS-CoV-2, in in vitro settings. [180] In vivo experiments involving SARS-CoV and MERS-CoV-infected animals revealed reduced airway inflammation and improved lung function, showcasing similar protective effects. The efficacy of post-exposure therapy is contingent on the timing of Remdesivir administration. [20] Furthermore, the licensing of nucleoside analogues for treating both DNA and RNA viruses is crucial in comprehending the mechanism of action underlying Remdesivir. However, several nucleoside analogue inhibitors have been observed to be ineffective against CoVs. [181,182] Remdesivir, a nucleoside analogue, works as an RdRp inhibitor by concentrating on the viral genome involved in viral replication. In the RdRp process, it thereby inhibits the protein complex of CoVs. The host breaks down Remdesivir to its active metabolite nortriptyline (NTP), which is then conjugated to ATP and incorporated into the developing RNA strand. Inclusion of a new strand stops the RNA synthesis and the expansion of the RNA strand once nucleotides are added. All CoVs include a proofreading process that detects and removes other nucleoside analoge activity, keeping antiviral activity. Surprisingly, it has been discovered that the mutant Murine hepatitis virus (MHV) lacked proofreading capabilities and was hence highly vulnerable to Remdesivir. It is also likely that mutations that enhance proofreading or base-pairing precision will result in Remdesivir resistance. Some data suggested that Remdesivir might work via a different mechanism, permitting partial antiviral vitality to endure despite viral changes. [182] WHO has authorized/approved the emergency use of Remdesivir. WHO revised its conditional advice against Remdesivir in hospitalised patients in November 2020 and is not recommended in this situation under any circumstances. [183] Phase III clinical research on Remdesivir is crucial for obtaining the more potent antiviral drug to combat this outbreak. [184] Clinical trials in 36 of 53 patients show appropriate data in 61 hospitalised patients taking Remdesivir off-label. However, without a placebo group, these findings are difficult to comprehend. An initial randomised controlled trial was flawed, favouring Remdesivir with a non-significant trend toward shorter time to clinical changes. This trial was insufficient, however it did show that patients treated with Remdesivir had better healing as an average recovery time of 11 days vs. placebo of 15 days. In addition, there were improvements for better survival on day 14. The research indicated disadvantages in individuals with high-flow oxygen, and invasive or non-invasive ventilation, indicating that antivirals like Remdesivir might have a poor impact in late diseases where the phenotype is likely to be inflammatory. The analysis was published before the full results could be obtained via follow-up research. [20] Remdesivir is now the subject of several clinical research studies regarding COVID-19 prevention. An initial dose of 200 mg of intravenous Remdesivir is given to participants in this double-blinded, placebo-controlled study on the first day, followed by a controlled dose of 100 mg per day and up to a maximum of 10 cumulative days of treatment before release. The initial trial result is expressed as the percentage of patients in each group, employing a seven-category clinical severity scale, up to the fifteenth day following the initiation of therapy, as indicated by the United States National Library of Medicine clinical trials registry.Gilead Sciences is also supporting a Remdesivir study in patients with severe COVID-19 that will combine a primary outcome test of fever with an outcome test of oxygen normalization. In Hubei Province, China, two double-blind placebo-controlled trials included patients: one for hospitalized individuals with mild-to-moderate COVID-19 and the other for severe cases. [178] In the mild-to-moderate study, key success criteria include the normalization of body temperature, oxygen consumption, breathing rate, and cough recovery for a minimum of 72 hours. Timing for health advancement is the key outcome in the extreme case study, which is presented using a six-category ratio scale from discharge to fatality. [178] It is described that Remdesivir was also found to be effective against MERS-CoV, reducing viral loads in the infected mice and restoring normal lung-based function. [185] Additionally, it is regarded as a treatment-assist agent for SARS-CoV-2. [176] The viral load in oropharyngeal and nasopharyngeal swabs could be reduced by Remdesivir treatment for about 12 days, according to preliminary studies. [87] The combination of Chloroquine, an anti-malarial drug, and Remdesivir, can successfully stop the growth of SARS-CoV-2 in Vero E6.86 cell lines in an in vitro studies. The potential effect of Remdesivir in COVID-19 is being studied in clinical trials in France, USA, and Norway. Remdesivir was used in treatment in Singapore and the USA and injected intravenously into the first patient who recovered there. [186] In a different research, 584 participants received Remdesivir or continued receiving conventional therapy, and 533 (91%) of these patients finished the experiment. Patients in the 5-day Remdesivir group received an average of 5 days of treatment, while those in the 10-day Remdesivir group received an average of 6 days. Compared to patients getting standard therapy, patients in the 5-day Remdesivir group showed statistically noticeably higher probabilities of a better clinical status distribution on day 11. [187] However, trial outcomes regarding safety, secondary outcomes, and viral load showed 22 of 158 Remdesivir patients died (14%), compared to ten of 78 placebo patients (13%), and there was no evidence that viral load declined differently over time in the placebo groups and Remdesivir. [188] Remdesivir's limited oral bioavailability often limits its preventive use. Additional pharmacological measures are required to make the drug available to the outpatient population. Remdesivir inhalation Phase 1 trial and FDA approval were recently reported by the manufacturer. [189] During clinical trials, Remdesivir was administered as a freeze-dried powder injection. The dosage technique employed is as follow; on the first day, an initial dose of 200 mg of Remdesivir is supplied via intravenous dripping. Then, for the next 9 days, 100 mg is supplied intravenously as a maintenance dose. [129]
5.3.2. Favipiravir
Favipiravir is classified as a purine analogue (Figure 6) and was licensed for influenza treatment in Japan. [190] In vitro, it also exhibits selectivity against several RNA viruses, including SARS-CoV-2. Favipiravir improved the survival of the influenza A virus in mice. [20] It has been shown, in vitro, to be effective against oseltamivir-resistant A, B, and C viruses. In many RNA viruses, favipiravir is regarded as a substrate of viral RNA polymerase once it has been transformed to the active phosphoribosylated form. [184] The enzyme, in the tissue, phosphoribosylated it to its active form, favipiravir-RTP. The mechanism of action of favipiravir could be described as follows; (a) The RNA-dependent RNA-polymerase (RdRp) enzyme, which serves as a substrate molecule, misinterprets it for a purine nucleotide. As a result, its activity is inhibited, effectively ending the viral protein replication. [191] (b) By blocking further extension and integrating into the RNA viral chain, this mode of action, together with the catalytic domain's ability to keep the RdRp enzyme active, demonstrates the wide range of activities this molecule. [192] (c) Favipiravir is considered as a virucidal drug. During the influenza infection, it is determined to cause lethal in vitro mutagenesis. However, it is unclear whether Favipiravir has a similar action against SARS-CoV-2. Favipiravir is an orally administered drug with a mode of action similar to Remdesivir. There is less evidence to support the use of favipiravir. However, it is still emerging as a helpful drug within mild-to-moderate circumstances. [193] The advantage of favipiravir is that it is administered orally. Therefore, it can be administered as part of hospital treatment for patients with symptoms that are not critical. This drug may be utilised in several cases because mild to moderate concomitant disorders are present in many COVID-19 cases and then care should be given at home. Favipiravir reduces viremia if given after the inception of COVID-19 symptoms. The effect of favipiravir on prophylaxis is also being studied in an ongoing study. [193] Heavy pill pressure, 18 tablets filled on the first day, and 8 tablets each day for the duration of the treatment term is its principal drawback. With the recent addition of a 400 mg dose, these concerns over the intense pill pressure are being partially allayed. A two-week prescription medication time is another drawback. Teratogenicity is the most significant side effect of favipiravir. Additionally, there were adverse outcomes such as neutropenia, elevated ALT and AST, diarrhoea, and increased uric acid throughout the phase III clinical investigation in the patients in Japan. The most frequent side effects of favipiravir with COVID-19 included hepatic enzyme abnormalities, psychiatric and gastrointestinal symptoms, and blood elevations of uric acid. Although pregnant women should not take favipiravir, the side effects are often mild. [129,193] In Japan, favipiravir is authorised to treat new or recurrent influenza. It was one of the first medications for COVID-19 management to be approved. [12,129,193] The recommended favipiravir dosage for influenza in China is 1600 mg given orally every 12 hours on the first day, followed by 600 mg given orally every 12 hours, and on the sixth day, 600 mg given orally every 24 hours. [129] Additionally, individuals with COVID-19 infections were enrolled in randomised trials with either interferon and favipiravir or baloxavir, marboxil, and favipiravir. [184] To determine whether the therapies work in concert or separately, umifenovir and other antivirals are frequently combined. [184] In an open-label, non-randomized study conducted in China, SARS-CoV-2 patients who received a double dose of interferon and favipiravir as opposed to a triple dose of interferon and ritonavir and lopinavir experienced viral clearance in 4 days vs. 11 days, and their chest X-rays significantly improved. [20]
5.3.3. Ribavirin
Ribavirin, is known as a guanosine analogue (Figure 6) with various RNA antiviral activities. [129,194] Many viral infections, including hepatitis C virus (HCV), respiratory syncytial virus (RSV), and other hemorrhagic viruses, are treated with Ribavirin. In vitro studies exhibited antiviral activity against SARS-CoV at 50 mg/mL concentrations. Reducing haemoglobin, which is is a negative side effect that is hazardous to those who are experiencing respiratory failure. [194] Hemolytic anaemia and reproductive damage are the most severe side effects of ribavirin. [195] The antiviral activities of ribavirin entail non-specific or specific chain end-up, lethal mutagenesis, and suppression of nucleotide biosynthesis. [135] For COVID-19 patients, reported by the Chinese government, ribavirin is utilised. Adults get a dose of 500 mg administered intravenously 2 to 3 times each day for a maximum of 10 days. Ribavirin should be used with interferon or both ritonavir/lopinavir. [129] Another retrospective research followed the adverse effects, of 126 patients using 2000 mg of ribavirin as part of their treatment. Increased hemolytic haemorrhage, transaminase (40%), and bradyarrhythmia (14%) were present in certain patients. [196] Therefore, it is essential to monitor the ribavirin dose when treating COVID-19 individuals carefully. [178] During previous MERS-CoV and SARS-CoV outbreaks, ribavirin was also examined, although the results were conflicting. To stop virus replication at the micromolar stages for SARS-CoV-2, a randomised Phase II clinical trial involving 127 patients with mild to moderate COVID-19 was carried out. [197] It is more advised to combine ribavirin/interferon-β with ritonavir/lopinavir than to use just those drugs. [198] Additionally, it has been demonstrated that anti-parasitic medications such as nitazoxanide and ivermectin enhance the effects of interferon-α/β and, consequently, the immunological responses. They were investigated for COVID-19 instances without comorbidities while using ribavirin. However, the clinical trials mentioned above are insufficient to explain how ribavirin has successfully slowed the progression of the disease. On the other hand, a Phase I evaluation showed inhaled ribavirin preparation as the sole drug in the hospitalisation of adult COVID-19 patients. [199] Tenofovir, a nuclear adenosine analogue, used to treat HIV or HBV infections that have persisted for a long time. Tenofovir's efficient integration can effectively stop the polymerase reaction into RNA-dependent RNA polymerase. Tenofovir is regarded as a first-line substitute for HIV prevention both before and after exposure and is thought to be a very effective component of HIV-contaminated antiretroviral therapy when combined with emtricitabine. Two-randomised phase III clinical studies utilising emtricitabine/tenofovir were conducted for prophylaxis prior to the exposure of healthcare workers to COVID-19, which is inconsistent with the aforesaid therapeutic approach. [200]
5.3.4. Galidesivir
Galidesivir, an analogue of adenosine, [129,194] swaps nitrogen for carbon at position 7 of a base and nitrogen for oxygen at position 1 of the ribose ring (Figure 6). [191] The structural modification alters the electrostatic interaction with the viral RNA polymerase and causes the expanded RNA strand to end prematurely. [201] Galidesivir is now being used as an antiviral treatment for Ebola and has demonstrated a wide range of antiviral activities against Flavivirus, Arenavirus, Togavirus, Bunyavirus, Paramyxovirus, Coronavirus, one Picornavirus Filovirus, and one Orthomyxovirus. [201,202,203] In primary human cell cultures, Galidesivir inhibits SARS-CoV, SARS-CoV-2, and MERS-CoV infection, enhancing respiratory function and reducing viral load. The antiviral studies demonstrated its capacity to integrate numerous deadly mutations via viral RNA, but not via host RNA, resulting in a significant barrier to drug resistance development. [202] Galidesivir was also shown in a pilot study to partially protect against Rift Valley Fever Virus (RVFV) infection in mice. [105] Recently, models for tick-borne Flavivirus and Zika infection have shown effectiveness. Galidesivir's phase I human research showed excellent tolerability and promising pharmacokinetics features when administered intramuscularly to healthy individuals compared to placebo. [201,204]
6. Other Nucleoside/Nucleotide Analogs (Transcription Inhibitors)
It is possible to consider other nucleoside and nucleotide analogue medications. They either focus on treating various viral infections (such as those treated with ribavirin, sofosbuvir, tenofovir, and telbivudine) or are being professionally researched (such as galidesivir, and EIDD–2801). [202] They are anticipated to have antiviral effect against SARS-CoV-2 because of their structural similarities to either ribavirin or Remdesivir. The FDA has granted authorization for certain drugs, including abacavir, alafenamide, tenofovir, didanosine, adefovir, ganciclovir, disoproxil, and tenofovir, as Nucleoside Analog Reverse Transcriptase Inhibitors (NtRtIs). Other inhibitors include delavirdine, efavirenz, rilpivirine, nevirapine, and nucleoside reverse transcriptase inhibitors (NRTIs), such as zalcitabine, lamivudine, azvudine, stavudine, and emtricitabine can also be used to show the antiviral activity against SARS-CoV-2. More Preclinical and clinical trials should be conducted to evaluate the clinical trial progress in silico trials, even though some have previously been evaluated by molecular docking. [50] As a result of interfering with the protein activity, ribavirin and sofosbuvir can be tightly bonded to the newly evolved RdRp coronavirus and eradicate the virus. It is important to note that sofosbuvir functions as a strong inhibitor of the recently discovered HCoV COVID-19 type.
6.1. Neuraminidase Inhibitors
6.1.1. Oseltamivir
A neuraminidase inhibitor is oseltamivir (Figure 7). [144] It is authorized for the prevention of influenza and the treatment of paediatric influenza. [205] Due to the unidentified presence of SARS-CoV-2 neuraminidase, drugs such as oseltamivir, peramivir, and zanamivir, which are neuraminidase inhibitors, are not anticipated to be effective in treating COVID-19 patients. [206] According to studies, people in Wuhan who have COVID-19 are treated with ganciclovir with oseltamivir or ritonavir/lopinavir with oseltamivir. Computational studies further supported the synergistic effects of ritonavir/lopinavir and oseltamivir in SARSCoV-2. [207,208] Oseltamivir was utilised in Afghanistan along with ceftriaxone and terbutaline to treat COVID-19 patients. It is revealed that three days of oseltamivir therapy significantly improved the patients' lungs on the CT scan. In Singapore and Indonesia, oseltamivir is utilised as the COVID-19 treatment of choice. [50] Oseltamivir is administered orally for the treatment of COVID-19 and suspected patients in Chinese hospitals, however, there is currently no solid proof that it has a tangible impact on the recovery of COVID-19 patients. [207]
6.1.2. Zanamivir and Peramivir
Another neuraminidase inhibitor that can be used for ventilated COVID-19 patients who are resistant to oseltamivir treatment is the zanamivir solution. Peramivir, as an antiviral medication, is given intravenously. Peramivir has a certain response for patients who do not respond to zanamivir or oseltamivir. [206,207] In Chinese hospitals, oseltamivir was administered orally to patients with 2019-nCoV confirmed infections. Oseltamivir may be helpful for treating COVID-19 patients, although there is currently no concrete evidence to support this. It has recently been suggested that neuraminidase inhibitors like oseltamivir, peramivir and zanamivir (Figure 7) are ineffective against COVID-19 and are not advised to be utilised for treatment procedures. [206]
6.2. M2 Ion-Channel Protein Target
Adamantane, Amantadine, and Rimantadine
The pH of the viral sheath must be kept constant through the M2 channel protein on the sheath. The channel, in order for steward cells to enter and pass through the trans-Golgi membrane prior to viral maturation, is essential, as well as in combating influenza viruses. A previous study demonstrated that amantadine could inhibit the HCV p7 protein, which is crucial for producing ion channels in the host cell membranes. Amantadine (Figure 8) reportedly has a potent in vitro action against coronavirus, according to a 1973 publication. [208] A recent study demonstrated that amantadinem (Figure 8) could inhibit SARS-CoV protein-membrane channel function. [209] Even though there is growing evidence that amantadine possesses antiviral potency appropriate for COVID-19 treatment, further investigation is necessary to determine its effectiveness. [50,210]
6.3. Non-Antiviral Drugs against SARS-CoV-2
6.3.1. Baricitinib
Baricitinib (Figure 9) has a high affinity against the Janus kinase (JAK) inhibitor by binding to and inhibiting adaptor-associated protein kinase 1 (AAK1). As a result, it may decrease both the inflammatory response and viral penetration brought on by SARS-CoV-2 infection. JAK inhibitors are used to treat rheumatoid arthritis and inflammatory diseases, including cancer. [211] Similar to baricitinib, JAK inhibitors such as ruxolitinib and fedratinib raise the degree of clathrin-mediated endocytosis, which may make them less efficient at lowering viral infectivity to tolerable levels. Lymphocytopenia, neutropenia, and viral reactivation are linked to its medical use. [50] Recent research has shown that the immunomodulatory drug Baricitinib and the antiviral Remdesivir have a good therapeutic effect against COVID-19. [212] By inhibiting JAK1/JAK2, baricitinib is anticipated to alleviate the cytokine storm brought on by COVID-19. Many clinical trials have been conducted worldwide, and one of them, in which baricitinib (2-4 mg) was universally provided for 1-2 weeks, showed encouraging results. Baricitinib should only be administered with extreme caution in patients with risk factors. For its use in pregnant women, human data is insufficient. [213] The patients with renal insufficiency should therefore be cautiously examined, along with termination or dose modification. [214]
6.3.2. Ivermectin; Importin α/β1-Mediated Nuclear Import Inhibitors
Ivermectin , is an anti-parasitic medication, approved by FDA. It has also been demonstrated to be an effective antiviral for both the Dengue virus and HIV in humans. [202,215] A single dose of the drug can reduce the viral RNA by around 5000 times. [216] The importin α/β heterodimer pre-formed that delivers the viral protein charge nuclearly may also be isolated. [50] Ivermectin is seen as a potential therapeutic inhibitor against RNA viruses because it blocks the nuclear transport pathway of viral proteins, which is essential to the host's antiviral response. [216,217] Ivermectin has recently been shown to have the ability to reduce viral RNA to 5000 times after 48 hours of SARS-CoV-2 infection. [202] Research to establish the ideal dosage utilising the well-established safety profile for anti-parasitic therapy is part of demonstrating the efficacy of ivermectin in COVID-19 treatment. [45,218] Further investigation is necessary to determine its efficacy against COVID-19. Additionally, ivermectin has demonstrated a broad spectrum of antiviral activity. By preventing NS3 helicase activity, it directly prevents the yellow fever virus from replicating. [219] Moreover, it stops importin α/β/1, which facilitates the passage of proteins between the nucleus and cytoplasm, which is necessary for HIV-1 replication and dengue viruses. [46,215,219] Recently, a case-controlled retrospective analysis suggested that ivermectin medication at a dose of 150 mcg/Kg might reduce the hospital admissions length and the mortality risk. Before ivermectin's efficacy can be verified in patients with SARS-CoV-2 infection, randomised controlled clinical trials are required. [50] For the treatment or prophylaxis of COVID-19, hydroxychloroquine and ivermectin combination medicine was recommended. Because of its dual effects on viral replication and viral assembly, this combination may have a synergistic effect. [220] Although the pharmacokinetic evaluation revealed that greater dosages were required to achieve antiviral activity, administering the prescribed inhibitory concentration in humans is likewise highly difficult. [221]
6.3.3. Interferon α and β
Interferons (IFNs), a broad-spectrum antiviral medication, are cytokines that activate the innate immune system in response to the viral infection. The antiviral activities and immunomodulatory of IFN can simulate protein production. IFN may also enhance the immune cells of the host cell's particular cytotoxic action. [221] Furthermore, the interferon (IFN) response is the first crucial one of protection against viruses. Type I and type III IFN responses aganist viral infections are stimulated by recognition of the innate immune sensing. [222] Many disorders, such as multiple sclerosis (MS) and viral hepatitis, have already been treated with pegylated and recombinant IFN α/β. As a result, the INF proposal against COVID-19 has demonstrated widespread concern. [210,223] Therefore, it is essential to thoroughly understand the biology of coronavirus infections to include rational therapeutic strategies and assess their clinical efficacy in COVID-19. [224] Although the cytokine development or robust chemokine or clinical investigations have revealed that the IFN reaction in patients is not mediated by significant IFN-I development. [216] When inflammatory chemokines and cytokines type I were analysed in the serum of COVID-19 patients, no substantial amounts were discovered, but pro-inflammatory chemokines and cytokines were found to be present in high levels. By tracking the transcriptome of SARS-CoV infected cells over time, other investigations shown that the IFN action on the virus can be delayed rather than completely eliminated. It was also shown that IFNs can inhibit the release of pro-inflammatory cytokines. [225] A systematic evaluation of 8 types of study, about 116 patients, found that using interferon in combination with ribavirin induced adverse side effects, including one patient who had evident hemolysis and two patients who had an increase in pancreatic enzymes. [226]
6.3.4. Teicoplanin
Teicoplanin , is an antibiotic glycopeptide frequently used to treat bacterial infections. Additionally, it is being utilised in treating SARS-CoV and is on prescription drugs lists for COVID-19. Teicoplanin is frequently used to treat viruses such as influenza, HIV, flavivirus, hepatitis C, ebola, and coronavirus; SARS-CoV and MERS-CoV. It can also cure gram-positive bacterial infections, particularly streptococcal and staphylococcal infections. [227] Teicoplanin inhibits the release of the viral genome and the viral replication cycle by cleaving the spike protein at low pH with cathepsin L; which enters the cell and targets the S protein in case of COVID-19, at the late endosomes during the early stages of viral replication. [228] The cathepsin L cleavage site has been preserved in the SARS-CoV and COVID-19 S proteins. The IC50 for teicoplanin in vitro is 1.66 M, which is much lower than the amount of 8.78 M for a 400 mg daily dose in human blood. [206]
6.3.5. Emetine
Emetine, (Figure 9) a protein synthesis inhibitor, is used to treat amebiasis as an anti-protozoan; it also works to prevent malaria by interacting with the ribosomal E site on Plasmodium falciparum. Its therapeutic utility has recently been constrained due to potential cardiotoxicity. Many RNA and DNA viruses, such as Zika virus, Cytomegalovirus, Ebola virus, Buffalo poxvirus, HIV-1, Plague of the Tiny Ruminants virus, Echovirus-1, Newcastle virus, Herpesvirus Bovine 1, Herpes Simplex Virus-2, Metapneumovirus, Rift River Fever virus, and influenza viruses were tested for its antiviral effects. [229] Additionally, emetine was noted in vitro to suppress MHV-A59, SARS-CoV, and MERS-CoV. At 0.5 M, it was discovered to block the replication of SARS-CoV-2 effectively. In vitro, SARS-CoV-2 therapeutic plasma levels can approach 0.075 g/mL below EC50. The plasma has a toxic concentration of 0.5 g/mL. Remdesivir with 6.25 M combined with emetine with 0.195 μm may decrease the viral generation by 64.9%; more in vivo studies are warranted. [202]
6.3.6. Chlorpromazine
The phenothiazine derivative chlorpromazine (CPZ), (Figure 9) which was chosen for Largactil, as the French brand name, has a wide range of properties, including antiviral, antifungal ones, anxiolytic, antiemetic, as well as immunomodulatory effects, the ability to modulate blood-brain barrier function, the ability to inhibit clathrin-mediated endocytosis, and others. It works via chlorpromazine-HCl, preventing the modulator assembly on cell surfaces and endosomes, which stops the virus from entering host cells. In addition, chlorpromazine is used to treat schizophrenia and other psychotic illnesses, tetanus symptoms, nausea, acute intermittent porphyria, chronic hiccups, and anxiety. Recently, in vitro investigations found that the CPZ had anti-MERS-CoV and anti-SARS-CoV-1 action. Without an intensive care unit (ICU), it is thought that CPZ may reduce COVID-19 infection in patients who need respiratory support. [202,230]
6.3.7. Aplidin
It was declared that aplidin had antiviral action, on March 2020. It is described that Multiple Myeloma is treated with aplidin on a large scale. The key to the virus's proliferation and spread is elongation factor 1 alpha (EF1A), which has been proven to be affected by aplidin in vitro tests. The antiviral activity of aplidin was initially investigated in human hepatoma cell line and HCoV-229E-GFP-related viruses. The early results are generally positive for SARS-CoV-2. [231]
6.3.8. Rapamycin
Rapamycin was initially employed as an antifungal and later as an immunosuppressive medication for patients undergoing organ transplants. [232] It causes the signal transduction pathway to be interrupted downstream, which leads to the phosphorylation of mTOR. In some viral infections, such as H1N1 pneumonia, Andes virus, MERS-CoV, and HCV, mTOR inhibition prevents viral duplication and improves clinical outcomes. The well-known mechanism of action of the immunosuppressive drug rapamycin involves blocking mTOR kinase. A crucial part of viral replication is played by mTOR, and more especially by the protein complex mTORC1 that mTOR forms. It was demonstrated that rapamycin affected the PI3K/AKT/mTOR pathway, which prevented MERS infection. According to earlier research, rapamycin can regulate the production of virus particles, cytokine storms, and aid in the treatment of the illness. Consequently, it appears that rapamycin is an appealing choice for drug repurposing. Furthermore, compared to widely used antivirals, it might be a better option for COVID-19 therapy. Furthermore, the rapid mutation rate of viral RNA is unlikely to lessen its efficiency. [233,234,235,236]
6.3.9. Lianhuaqingwen Capsule
In particular, lianhuaqingwen (LH) is utilised to treat influenza. [237] It reduces many symptoms such as muscle ache, headache, hyperpyrexia or fever, running nose, cough, aversion to colds, and nasal obstruction. The recommended dose is four capsules, three times a day. [129] LH was utilised by Chinese physicians to treat both mild and severe COVID-19 cases. As a potential treatment for SARS-CoV-2, LH has demonstrated effective anti-inflammatory and antiviral properties in vitro against coronaviruses. [238] Therefore, a different randomised controlled experiment (RCT) has shown that antiviral activity with LH can, in most cases, significantly relieve the COVID-19 symptoms, such as fever, fatigue, and cough. It may also significantly shorten and ease pneumonia symptoms without showing any noticeable negative effects. [239] Additionally, the antiviral potency of LH and other conventional Chinese medicines during a pandemic also effectively contributed to the treatment of SARs-CoV-2. [129]
6.3.10. Convalescent Plasma
Convalescent plasma for COVID-19, also called "survivor's plasma," is blood plasma obtained from COVID-19-recovered patients. Recently, the FDA approved its use in hospitalised COVID-19 patients by issuing an Emergency Use Authorization. This demonstrated that COVID-19 plasma or clean monoclonal antibodies might be extracted from patients who have totally recovered and subsequently administered to another new patient for treatment. [240] Through Feb. 2020, about five COVID-19 patients in Shenzhen, China, received convalescent plasma treatment. Patients in this trial developed a specific anti-SARS-CoV-2 antibody 10 to 22 days after receiving convalescent plasma therapy. [241] Reduced viral loads and improved results in sequential organ failure were observed in four of the five patients. Their virus test was still negative after the transfusion for 12 days. After two weeks of admission, the four patients were taken off artificial ventilation. However, three individuals were discharged from the hospital after over 50 days. [242] Convalescent plasma therapy has positive results, despite the trial's extremely small sample size and then in the US, this approach of care is advised. [240]
6.3.11. Metformin
The hyperinflammatory state is a hallmark of COVID-19. Monocytes and macrophages are essential immune cells that metabolically reprogrammed when stimulated with different stimuli including SARS-CoV-2 spike protein. [243] Thus drugs which regulate immunometabolism could inhibit this inflammatory response. Pre-treatment of monocytes with metformin strongly suppressed spike protein-mediated metabolic reprogramming that also suppresses inflammatory responses to SARS-CoV-2. This has potential implications for the treatment of hyper-inflammation during COVID-19. [243]
7. Neutralizing Antibodies for SARS-CoV-2
Despite the fact that it has been effective in certain patients, convalescent plasma's potential is still debatable. In fact, allergic reactions, transmitted infections due to transfusion (ex. HCV, HBV, HIV), lung injury were observed in certain convalescent plasma trials. Furthermore, only a portion of plasma antibodies will be neutralizing; meanwhile the non-neutralizing antibodies will attach to non-spike protein antigens, compromising antibody responses and causing additional tissue damage. Moreover, convalescent plasma antibody titer is low and blood resources are limited. All of these drawbacks resulted in limiting the use of convalescent plasma treatment. Conversely, anti-SARS-CoV-2 monoclonal antibodies overcomes all the drawbacks of convalescent plasma by being able to specifically target the neutralizing sites and be manufactured in large quantities with ease of scalability. [244] Bamlanivimab, also known as LY-CoV555, was the first monoclonal antibody discovered to be effective against COVID-19 infection. LY-CoV555 exhibited potent binding and neutralizing action to ACE2 (Figure 10). Even at low doses, it could decrease the viral amount in respiratory tract samples. [245]
Another monoclonal antibody called etesevimab has always been utilized alongside with bamlanivimab. This combination has shown more efficiency than bamlanivimab monotherapy in reducing the viral load in outpatients with mild-to-moderate symptoms in addition to reducing the risk of hospitalization and death linked to COVID-19. [247,248] On February 9, 2021, they were approved for emergency use jointly due to their significant effectiveness in treating individuals with mild-to-moderate COVID-19. Yet, due to the Omicron variant's high frequency, the FDA has withdrawn the use of these monoclonal antibodies for COVID-19 treatment due to its ineffectiveness compared to Omicron variance. [248,249] However, the FDA and the National Institutes of Health have approved bebtelovimab, as the only effective neutralizing monoclonal antibody for the treatment of high-risk COVID-19 patients. [250] Additionally, Iketani et al. verified that, with the exception of bebtelovimab, three Omicron sub lineages demonstrated resistance to seventeen different neutralizing antibodies. [251,252]
8. Some Recently Synthesized Compounds and Approved for COVID-19 Treatment
8.1. Molnupiravir (MK-4482, EIDD-2801) (Ridgeback Biotherapeutics/MSD)
Molnupiravir, a prodrug antiviral medicine, Figure 11, was used to orally treat influenza A and B viruses, was just licenced in the United Kingdom and available in November 2021. It is a synthetic nucleoside N4-hydroxycytidine (NHC) derivative that inhibits specific RNA viruses by causing copying errors during RNA replication. Compared to its precursor NHC, this medication has a higher oral bioavailability in non-human primates and ferrets. It is also effectively digested in vivo after absorption, releasing the active compound into the plasma. Using in vitro studies, [253] NHC can prevent SARS-CoV-2 and other related coronaviruses. [254,255] The nucleoside analogue introduced by the viral RdRp during viral RNA synthesis leads to error catastrophe and suppression of RNA synthesis. [256] This property makes it a viable option for treating COVID-19. [257,258,259] Molnupiravir has demonstrated effectiveness in reducing viral loads and lung pathology in Syrian hamsters and human lung-only mice, whether administered before or after SARS-CoV-2 infection. [254,260] In ferrets, post-infection treatment with molnupiravir lowered virus levels in nasal lavages and impeded transmission to untreated contact animals. Moreover, hamsters infected with the B.1.1.7 (alpha variant) and B.1.351 (beta variant) of COVID-19 showed resistance to SARS-CoV-2 when treated with molnupiravir. A randomized, double-blind, placebo-controlled phase 1 trial with healthy volunteers revealed that oral doses ranging from 50 to 1600 mg of molnupiravir were well-tolerated, with only a few mild side effects reported. [261] Molnupiravir is being examined in phase 3 clinical trials for COVID-19 outpatient therapy (NCT04575584), postexposure prophylaxis (NCT04939428), and inpatient therapy (NCT04575597). According to MSD release and Ridgeback Biotherapeutics, [262] molnupiravir reduced the risk of hospitalisation or death in patients with mild-to-moderate COVID-19 disease around 50% compared to placebo and approved by the U.K.'s Medicines and Healthcare Products Regulatory Agency. [263]
8.2. Paxlovid (Pf-07321332)
Pfizer Inc. produced PF-07321332, an oral antiviral medication. It acts as an active 3CLpro protease inhibitor. The drug combination of PF-07321332/ritonavir for the COVID-19 treatment underwent phase III research and was marketed under the name Paxlovid. [167,168] As a combination, [264] ritonavir delays the cytochrome enzymes' metabolism of the PF-07321332, keeping larger levels of the primary medication in the blood. When taken within three days of the onset symptom, Pfizer's phase 2/3 results showed an 89% reduction in hospitals, it has been released in November 2021. [169,170] Ritonavir is coadministered in small doses to slow down PF-07321332 metabolism. [265]
9. COVID-19 and Cancer
It is commonly known that viral infections increase the likelihood of developing tumors. According to estimates, viruses are the primary cause of carcinogenic illnesses, which account for 15.4% of the cases of cancer. Numerous RNA viruses have been linked to increased cancer risk, and many can result in chronic infections. [266,267]
The majority of viruses use oncogenic processes that entail the continuous production of particular gene products that interact with cellular gene products to control proliferative or anti-apoptotic activity. For up to six months following a negative SARS-CoV-2 test, remnant SARS-CoV-2 nucleocapsid proteins were found in a number of extrapulmonary tissue samples, including those from the ileum, appendix, colon, lymph nodes and liver in individuals who had recovered with COVID-19. [268,269]
Additionally; it has been noted that acute COVID-19 infection patients have changed microbiota. This change was typified by an increase in opportunistic pathogens and a decrease in commensals, or beneficial bacteria, in the gut. [270]
Moreover, previous research has found that the tumor-suppressor proteins and SARS-CoV-2 spike overlap and that autoimmune cross-reactivity may be a possible mechanism behind future cancer recurrence after exposure to SARS-CoV-2.
In order to regulate p53, which poses a threat to SARS-CoV-2, it has evolved tactics similar to those of other viruses (such as the Epstein-Barr virus). Since the apoptotic signaling system depends heavily on the onco-suppressive protein p53, it has been suggested that long-term SARS-CoV-2 p53 suppression may have carcinogenic consequences. [267,271]
SARS-CoV-2-caused COVID-19 infection is deemed fatal since it has a wide-ranging impact on various organs, primarily the respiratory system. It damages the neurological, cardiovascular, and pulmonary systems, among other organs, leading to organ failure. Investigations should focus on how inflammation brought on by SARS-CoV-2 affects cancer cells and the environment around tumours. [272,273] The microenvironment tumour may change due to COVID-19, encouraging cancer cell growth and reawakening the dormant cancer cell (DCC). [274,275,276] When SARS-CoV-2 infection occurs, DCCs can reawaken and populate the pre-metastatic in the lungs and other organs, which can result in the spread of tumours. The most severe clinical effects of COVID-19 are DCC reawakening and subsequent neutrophil and monocyte/macrophage activation with an unregulated cascade of proinflammatory cytokines. The role of COVID-19 in inflammation, tumour growth, and tumour cell metastasis demand further investigation; the findings of these investigations will contribute to creating new targeted medicines for the treatment of COVID-19-positive patients as well as for the prevention of cancer. [277,278,279]
10. Conclusions and Public Health Perspectives
The WHO declared a pandemic on March 11, 2020, in response to the new coronavirus in the human, which sparked a global threat. It is undoubtedly one of the worst public health disasters two years later. Neither the SARS-CoV-2 development nor the severe and widespread effects of COVID-19 infections were anticipated. However, the quick reaction to the COVID-19 pandemic, and the responses taken by the WHO, governments, business, international researchers, and health authorities, has strengthened public health resilience and assisted in reducing adverse effects on society. These initiatives openly disclosed data on infection rates and fatalities in terms of clinical trials. The severity of SARSCoV-2 was lessened by open research, such as the early disclosure of the viral genome, patient trial validation of vaccine candidates, industry involvement in the development, and governments' speedy licensing of new diagnostic tests and vaccinations. Researchers and experts are determined to develop innovative therapeutic tactics quickly and plan to combat the terrible COVID-19 epidemic. Creating new vaccines and employing some FDA-approved medications that might be tested against COVID-19 and are viewed as repurposed drugs are two therapeutic approaches. The effectiveness of many classes of currently licensed and candidate vaccines and repurposed medications (such as interferons, non-antivirals and antivirals, and antiparasitic medicines), against COVID-19 infections have been reported. The effectiveness of several drugs has been presented and categorised according to their mechanisms of action against SARS-CoV-2. In-depth discussion was given in this study of antiviral medications, along with protease inhibitors, fusion inhibitors, M2 ion-channel protein blockers, neuraminidase inhibitors, neutralizing antibodies, and other non-antiviral drugs that may have effects against SARS-CoV-2. The recently FDA-approved drugs molnupiravir and PF-07321332 shed insight on their mode of action and eligibility as cutting-edge oral medications that fight SARS-CoV-2 by reducing hospitalizations for COVID-19 patients. Regarding the effectiveness of the vaccinations, it has been noted that Pfizer, Moderna, Fosun Pharma, BioNTech, and NIAID vaccines may be found to be the most effective to combat COVID-19. These vaccines are now being used, although they have significant drawbacks, including viral and host issues. As a result, choosing to get immunised with any of the licenced vaccinations must be done so under under medical supervision and after taking the results of current clinical studies into account. Several lessons have been learned from the pandemic, including the urgency of large-scale vaccine production and distribution, the need for point-of-care diagnostic tests and, the importance of addressing the trade in wild animals and ecosystem destruction as significant contributors to the spread of infectious diseases. The COVID-19 pandemic's lessons such as the necessity of point-of-care diagnostic testing, the urgency of producing and distributing vaccines on a broad scale, and the significance of managing ecosystem devastation and trading in wild animals as major factors to the propagation of infectious illnesses; will be crucial for addressing future dangers to the public's health, particularly those brought on by new viruses or diseases.
Authors Contribution
R.F.B., A.V.S. and D.G. contributed equally to the conception and design of the study, as well as the analysis and interpretation of the data. They both played a significant role in drafting and revising the manuscript critically for important intellectual content. V.T. participated in the study's design, data analysis, and interpretation. She was also involved in drafting and revising the manuscript, providing important intellectual input. A.M.A. contributed to the acquisition of data, data analysis, and interpretation. He also played a role in drafting and revising the manuscript. K.A., M.M.E. was involved in the conception of the study and the acquisition of data. They provided critical input during the drafting and revision of the manuscript. All authors read and approved the manuscript.
Funding
Not applicable.
Availability of data and materials
The datasets and materials used in this research are available upon request.
Acknowledgments
Authors are gratefully acknowledging the National Research Center, Cairo, Egypt for their support.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Ethical Approval
Not applicable.
References
- Weiss, S.R.; Leibowitz, J.L. Chapter 4 - Coronavirus Pathogenesis. In: Maramorosch K, Shatkin AJ, Murphy FA, eds. Vol 81. Advances in Virus Research. Academic Press 2011, 81, 85–164. [Google Scholar] [CrossRef]
- Su, S.; Wong, G.; Shi, W.; Liu, J.; Lai, A.C.K.; Zhou, J.; Liu, W.; Bi, Y.; Gao, G.F. Epidemiology, Genetic Recombination, and Pathogenesis of Coronaviruses. Trends Microbiol. 2016, 24, 490–502. [Google Scholar] [CrossRef]
- Tripathy, S.; Dassarma, B.; Roy, S.; Chabalala, H.; Matsabisa, M.G. A review on possible modes of action of chloroquine/hydroxychloroquine: repurposing against SAR-CoV-2 (COVID-19) pandemic. Int. J. Antimicrob. Agents 2020, 56, 106028. [Google Scholar] [CrossRef]
- Tufan, A.; GÜLER, A.A.; Matucci-Cerinic, M. COVID-19, immune system response, hyperinflammation and repurposing antirheumatic drugs. Turk. J. Med Sci. 2020, 50, 620–632. [Google Scholar] [CrossRef]
- Hong, S.I.; Ryu, B.-H.; Chong, Y.P.; Lee, S.; Kim, S.; Kim, H.C.; Hong, K.-W.; Bae, I.-G.; Cho, O.-H. Five severe COVID-19 pneumonia patients treated with triple combination therapy with lopinavir/ritonavir, hydroxychloroquine, and interferon β-1b. Int. J. Antimicrob. Agents 2020, 56, 106052. [Google Scholar] [CrossRef]
- Saha, B.K.; Bonnier, A.; Chong, W. Antimalarials as Antivirals for COVID-19: Believe it or Not! Am. J. Med. Sci. 2020, 360, 618–630. [Google Scholar] [CrossRef]
- El-Masry, R.M.; Al-Karmalawy, A.A.; Alnajjar, R.; Mahmoud, S.H.; Mostafa, A.; Kadry, H.H.; Abou-Seri, S.M.; Taher, A.T. Newly synthesized series of oxoindole–oxadiazole conjugates as potential anti-SARS-CoV-2 agents:in silicoandin vitrostudies. New J. Chem. 2022, 46, 5078–5090. [Google Scholar] [CrossRef]
- Roshdy WH, khalifa MK, San JE, et al. SARS-CoV-2 Genetic diversity and lineage dynamics of in Egypt. medRxiv. Published online , 2022:2022.01.05.22268646. 5 January. [CrossRef]
- Hosseini, F.S.; Amanlou, M. Anti-HCV and anti-malaria agent, potential candidates to repurpose for coronavirus infection: Virtual screening, molecular docking, and molecular dynamics simulation study. Life Sci. 2020, 258, 118205–118205. [Google Scholar] [CrossRef]
- Kandeel, M.; Al-Nazawi, M. Virtual screening and repurposing of FDA approved drugs against COVID-19 main protease. Life Sci. 2020, 251, 117627–117627. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wang, X.-J. Potential inhibitors against 2019-nCoV coronavirus M protease from clinically approved medicines. J. Genet. Genom. 2020, 47, 119–121. [Google Scholar] [CrossRef] [PubMed]
- Elfiky, A.A. Anti-HCV, nucleotide inhibitors, repurposing against COVID-19. Life Sci. 2020, 248, 117477–117477. [Google Scholar] [CrossRef]
- Li F, Michelson AP, Foraker R, Zhan M, Payne PRO. Repurposing drugs for COVID-19 based on transcriptional response of host cells to SARS-CoV-2. arXiv. Published online , 2020. Accessed March 23, 2023. https://arxiv.org/abs/2006. 1 June 0122.
- Samaee, H.; Mohsenzadegan, M.; Ala, S.; Maroufi, S.S.; Moradimajd, P. Tocilizumab for treatment patients with COVID-19: Recommended medication for novel disease. Int. Immunopharmacol. 2020, 89, 107018. [Google Scholar] [CrossRef]
- El Kantar, S.; Nehmeh, B.; Saad, P.; Mitri, G.; Estephan, C.; Mroueh, M.; Akoury, E.; Taleb, R.I. Derivatization and combination therapy of current COVID-19 therapeutic agents: a review of mechanistic pathways, adverse effects, and binding sites. Drug Discov. Today 2020, 25, 1822–1838. [Google Scholar] [CrossRef]
- Peretto, G.; Sala, S.; Caforio, A.L.P. Acute myocardial injury, MINOCA, or myocarditis? Improving characterization of coronavirus-associated myocardial involvement. Eur. Hear. J. 2020, 41, 2124–2125. [Google Scholar] [CrossRef]
- Zhang, R.; Li, Y.; Zhang, A.L.; Wang, Y.; Molina, M.J. Identifying airborne transmission as the dominant route for the spread of COVID-19. Proc. Natl. Acad. Sci. 2020, 117, 14857–14863. [Google Scholar] [CrossRef]
- Falahi, S.; Kenarkoohi, A. Transmission routes for SARS-CoV-2 infection: review of evidence. New Microbes New Infect. 2020, 38, 100778–100778. [Google Scholar] [CrossRef]
- Marquès, M.; Domingo, J.L. Contamination of inert surfaces by SARS-CoV-2: Persistence, stability and infectivity. A review. Environ. Res. 2020, 193, 110559–110559. [Google Scholar] [CrossRef]
- Farne, H.; Kumar, K.; Ritchie, A.I.; Finney, L.J.; Johnston, S.L.; Singanayagam, A. Repurposing Existing Drugs for the Treatment of COVID-19. Ann. Am. Thorac. Soc. 2020, 17, 1186–1194. [Google Scholar] [CrossRef]
- The RECOVERY Collaborative Group. Dexamethasone in Hospitalized Patients with Covid-19. N. Engl. J. Med. 2021, 384, 693–704. [Google Scholar] [CrossRef]
- Mahmoud, D.B.; Bakr, M.M.; Al-Karmalawy, A.A.; Moatasim, Y.; El Taweel, A.; Mostafa, A. Scrutinizing the Feasibility of Nonionic Surfactants to Form Isotropic Bicelles of Curcumin: a Potential Antiviral Candidate Against COVID-19. Aaps Pharmscitech 2021, 23, 1–12. [Google Scholar] [CrossRef]
- C. R., V.; Sharma, R.; Jayashree, M.; Nallasamy, K.; Bansal, A.; Angurana, S.K.; Mathew, J.L.; Sankhyan, N.; Dutta, S.; Verma, S.; et al. Epidemiology, Clinical Profile, Intensive Care Needs and Outcome in Children with SARS-CoV-2 Infection Admitted to a Tertiary Hospital During the First and Second Waves of the COVID-19 Pandemic in India. Indian J. Pediatr. 2022, 90, 131–138. [Google Scholar] [CrossRef]
- Shehata, M.M.; Mahmoud, S.H.; Tarek, M.; Al-Karmalawy, A.A.; Mahmoud, A.; Mostafa, A.; Elhefnawi, M.M.; Ali, M.A. In Silico and In Vivo Evaluation of SARS-CoV-2 Predicted Epitopes-Based Candidate Vaccine. Molecules 2021, 26, 6182. [Google Scholar] [CrossRef] [PubMed]
- Harvey, W.T.; Carabelli, A.M.; Jackson, B.; Gupta, R.K.; Thomson, E.C.; Harrison, E.M.; Ludden, C.; Reeve, R.; Rambaut, A.; Consortium, C.-G.U.; et al. SARS-CoV-2 variants, spike mutations and immune escape. Nat. Rev. Microbiol. 2021, 19, 409–424. [Google Scholar] [CrossRef]
- He, X.M.; Hong, W.Q.; Pan, X.Y.; Lu, G.W.; Wei, X.W. SARS-CoV-2 Omicron variant: Characteristics and prevention. MedComm 2021, 2, 838–845. [Google Scholar] [CrossRef]
- Kaur, U.; Acharya, K.; Mondal, R.; Singh, A.; Saso, L.; Chakrabarti, S.S. Should ACE2 be given a chance in COVID-19 therapeutics: A semi-systematic review of strategies enhancing ACE2. Eur. J. Pharmacol. 2020, 887, 173545–173545. [Google Scholar] [CrossRef]
- De Luca V, Angeli A, Nocentini A, et al. Leveraging SARS-CoV-2 Main Protease (Mpro) for COVID-19 Mitigation with Selenium-Based Inhibitors. Int J Mol Sci. 2024;25(2):971. [CrossRef]
- Barghash, R.F.; Fawzy, I.M.; Chandrasekar, V.; Singh, A.V.; Katha, U.; Mandour, A.A. In Silico Modeling as a Perspective in Developing Potential Vaccine Candidates and Therapeutics for COVID-19. Coatings 2021, 11, 1273. [Google Scholar] [CrossRef]
- Awad, A.M.; Hansen, K.; Del Rio, D.; Flores, D.; Barghash, R.F.; Kakkola, L.; Julkunen, I.; Awad, K. Insights into COVID-19: Perspectives on Drug Remedies and Host Cell Responses. Biomolecules 2023, 13, 1452. [Google Scholar] [CrossRef]
- Gemmati D, Longo G, Gallo I, et al. Host genetics impact on SARS-CoV-2 vaccine-induced immunoglobulin levels and dynamics: The role of TP53, ABO, APOE, ACE2, HLA-A, and CRP genes. Front Genet. 2022, 13, 3145. [CrossRef]
- Roy, A.V.; Chan, M.; Banadyga, L.; He, S.; Zhu, W.; Chrétien, M.; Mbikay, M. Quercetin inhibits SARS-CoV-2 infection and prevents syncytium formation by cells co-expressing the viral spike protein and human ACE2. Virol. J. 2024, 21, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Tebha SS, Tameezuddin A, Bajpai S, Zaidi AK. SARS-CoV-2—Virus structure and life cycle. Prog Mol Biol Transl Sci. 2024;202:1-23. [CrossRef]
- Sun, Z.; Ostrikov, K.K. Future antiviral surfaces: Lessons from COVID-19 pandemic. Sustain. Mater. Technol. 2020, 25, e00203. [Google Scholar] [CrossRef]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.-H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Liu, Y.; Yang, Y.; Zhang, P.; Zhong, W.; Wang, Y.; Wang, Q.; Xu, Y.; Li, M.; Li, X.; et al. Analysis of therapeutic targets for SARS-CoV-2 and discovery of potential drugs by computational methods. Acta Pharm. Sin. B 2020, 10, 766–788. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.K.; Brindisi, M.; Shahabi, D.; Chapman, M.E.; Mesecar, A.D. Drug Development and Medicinal Chemistry Efforts toward SARS-Coronavirus and Covid-19 Therapeutics. ChemMedChem 2020, 15, 907–932. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.V.; Katz, A.; Maharjan, R.S.; Gadicherla, A.K.; Richter, M.H.; Heyda, J.; del Pino, P.; Laux, P.; Luch, A. Coronavirus-mimicking nanoparticles (CorNPs) in artificial saliva droplets and nanoaerosols: Influence of shape and environmental factors on particokinetics/particle aerodynamics. Sci. Total. Environ. 2022, 860, 160503–160503. [Google Scholar] [CrossRef]
- Yan, R.; Zhang, Y.; Li, Y.; Xia, L.; Guo, Y.; Zhou, Q. Structural basis for the recognition of SARS-CoV-2 by full-length human ACE2. Science 2020, 367, 1444–1448. [Google Scholar] [CrossRef]
- Shang, J.; Ye, G.; Shi, K.; Wan, Y.; Luo, C.; Aihara, H.; Geng, Q.; Auerbach, A.; Li, F. Structural basis of receptor recognition by SARS-CoV-2. Nature 2020, 581, 221–224. [Google Scholar] [CrossRef]
- Devaux, C.A.; Rolain, J.-M.; Colson, P.; Raoult, D. New insights on the antiviral effects of chloroquine against coronavirus: what to expect for COVID-19? Int. J. Antimicrob. Agents 2020, 55, 105938–105938. [Google Scholar] [CrossRef]
- Zhang, T.; Wu, Q.; Zhang, Z. Probable Pangolin Origin of SARS-CoV-2 Associated with the COVID-19 Outbreak. Curr. Biol. 2020, 30, 1346–1351. [Google Scholar] [CrossRef]
- Hilgenfeld, R. From SARS to MERS: crystallographic studies on coronaviral proteases enable antiviral drug design. FEBS J. 2014, 281, 4085–4096. [Google Scholar] [CrossRef]
- Wang, H.; Xue, S.; Yang, H.; Chen, C. Recent progress in the discovery of inhibitors targeting coronavirus proteases. Virol. Sin. 2016, 31, 24–30. [Google Scholar] [CrossRef]
- Shannon, A.; Le, N.T.-T.; Selisko, B.; Eydoux, C.; Alvarez, K.; Guillemot, J.-C.; Decroly, E.; Peersen, O.; Ferron, F.; Canard, B. Remdesivir and SARS-CoV-2: Structural requirements at both nsp12 RdRp and nsp14 Exonuclease active-sites. Antivir. Res. 2020, 178, 104793–104793. [Google Scholar] [CrossRef] [PubMed]
- Ferron, F.; Subissi, L.; De Morais, A.T.S.; Le, N.T.T.; Sevajol, M.; Gluais, L.; Decroly, E.; Vonrhein, C.; Bricogne, G.; Canard, B.; et al. Structural and molecular basis of mismatch correction and ribavirin excision from coronavirus RNA. Proc. Natl. Acad. Sci. USA 2018, 115, E162–E171. [Google Scholar] [CrossRef] [PubMed]
- Nagy, P.D.; Pogany, J. The dependence of viral RNA replication on co-opted host factors. Nat. Rev. Microbiol. 2011, 10, 137–149. [Google Scholar] [CrossRef]
- Nal, B.; Chan, C.; Kien, F.; Siu, L.; Tse, J.; Chu, K.; Kam, J.; Staropoli, I.; Crescenzo-Chaigne, B.; Escriou, N.; et al. Differential maturation and subcellular localization of severe acute respiratory syndrome coronavirus surface proteins S, M and E. J. Gen. Virol. 2005, 86, 1423–1434. [Google Scholar] [CrossRef]
- Klumperman, J.; Locker, J.K.; Meijer, A.; Horzinek, M.C.; Geuze, H.J.; Rottier, P.J. Coronavirus M proteins accumulate in the Golgi complex beyond the site of virion budding. J. Virol. 1994, 68, 6523–34. [Google Scholar] [CrossRef]
- Frediansyah, A.; Tiwari, R.; Sharun, K.; Dhama, K.; Harapan, H. Antivirals for COVID-19: A critical review. Clin. Epidemiology Glob. Heal. 2020, 9, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Vaccines – COVID19 Vaccine Tracker. Accessed , 2023. https://covid19.trackvaccines. 17 April.
- Fiolet, T.; Kherabi, Y.; MacDonald, C.-J.; Ghosn, J.; Peiffer-Smadja, N. Comparing COVID-19 vaccines for their characteristics, efficacy and effectiveness against SARS-CoV-2 and variants of concern: a narrative review. Clin. Microbiol. Infect. 2021, 28, 202–221. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhao, X.; Zhang, R.; Ye, B.; Yuan, X.; Zhang, D.; Li, L.; Tian, J.; Guo, Y.; Gao, G.F.; et al. Heterologous BBIBP-CorV/ZF2001 vaccination augments neutralization against SARS-CoV-2 variants: A preliminary observation. Lancet Reg. Heal. - West. Pac. 2022, 21, 100440. [Google Scholar] [CrossRef]
- Enjuanes L, Zuñiga S, Castaño-Rodriguez C, Gutierrez-Alvarez J, Canton J, Sola I. Molecular Basis of Coronavirus Virulence and Vaccine Development. Adv Virus Res. 2016;96:245-286. [CrossRef]
- Heinz, F.X.; Stiasny, K. Distinguishing features of current COVID-19 vaccines: knowns and unknowns of antigen presentation and modes of action. npj Vaccines 2021, 6, 1–13. [Google Scholar] [CrossRef]
- Asthana, A.; Gaughan, C.; Dong, B.; Weiss, S.R.; Silverman, R.H. Specificity and Mechanism of Coronavirus, Rotavirus, and Mammalian Two-Histidine Phosphoesterases That Antagonize Antiviral Innate Immunity. mBio 2021, 12, e0178121. [Google Scholar] [CrossRef]
- Kyriakidis, N.C.; López-Cortés, A.; González, E.V.; Grimaldos, A.B.; Prado, E.O. SARS-CoV-2 vaccines strategies: a comprehensive review of phase 3 candidates. npj Vaccines 2021, 6, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Granados-Riveron, J.T.; Aquino-Jarquin, G. Engineering of the current nucleoside-modified mRNA-LNP vaccines against SARS-CoV-2. Biomed. Pharmacother. 2021, 142, 111953–111953. [Google Scholar] [CrossRef]
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Pérez Marc, G.; Moreira, E.D.; Zerbini, C.; et al. Safety and efficacy of the BNT162b2 mRNA COVID-19 vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef] [PubMed]
- Dagan, N.; Barda, N.; Kepten, E.; Miron, O.; Perchik, S.; Katz, M.A.; Hernán, M.A.; Lipsitch, M.; Reis, B.; Balicer, R.D. BNT162b2 mRNA Covid-19 Vaccine in a Nationwide Mass Vaccination Setting. N. Engl. J. Med. 2021, 384, 1412–1423. [Google Scholar] [CrossRef]
- Baden, L.R.; El Sahly, H.M.; Essink, B.; Kotloff, K.; Frey, S.; Novak, R.; Diemert, D.; Spector, S.A.; Rouphael, N.; Creech, C.B.; et al. Efficacy and Safety of the mRNA-1273 SARS-CoV-2 Vaccine. N. Engl. J. Med. 2021, 384, 403–416. [Google Scholar] [CrossRef]
- Feikin, D.R.; Feikin, D.R.; Higdon, M.M.; Higdon, M.M.; Abu-Raddad, L.J.; Abu-Raddad, L.J.; Andrews, N.; Andrews, N.; Araos, R.; Araos, R.; et al. Duration of effectiveness of vaccines against SARS-CoV-2 infection and COVID-19 disease: results of a systematic review and meta-regression. Lancet 2022, 399, 924–944. [Google Scholar] [CrossRef] [PubMed]
- Sievers, B.L.; Cheng, M.T.K.; Csiba, K.; Meng, B.; Gupta, R.K. SARS-CoV-2 and innate immunity: the good, the bad, and the “goldilocks”. Cell. Mol. Immunol. 2023, 21, 171–183. [Google Scholar] [CrossRef]
- Ogando, N.S.; Ferron, F.; Decroly, E.; Canard, B.; Posthuma, C.C.; Snijder, E.J. The Curious Case of the Nidovirus Exoribonuclease: Its Role in RNA Synthesis and Replication Fidelity. Front. Microbiol. 2019, 10, 1813. [Google Scholar] [CrossRef]
- Ogando, N.S.; Zevenhoven-Dobbe, J.C.; van der Meer, Y.; Bredenbeek, P.J.; Posthuma, C.C.; Snijder, E.J. The Enzymatic Activity of the nsp14 Exoribonuclease Is Critical for Replication of MERS-CoV and SARS-CoV-2. J. Virol. 2020, 94. [Google Scholar] [CrossRef]
- Flores-Alanis, A.; Cruz-Rangel, A.; Rodríguez-Gómez, F.; González, J.; Torres-Guerrero, C.A.; Delgado, G.; Cravioto, A.; Morales-Espinosa, R. Molecular Epidemiology Surveillance of SARS-CoV-2: Mutations and Genetic Diversity One Year after Emerging. Pathogens 2021, 10, 184. [Google Scholar] [CrossRef]
- Lauring, A.S.; Hodcroft, E.B. Genetic Variants of SARS-CoV-2—What Do They Mean? JAMA 2021, 325, 529–531. [Google Scholar] [CrossRef] [PubMed]
- Korber, B.; Fischer, W.M.; Gnanakaran, S.; Yoon, H.; Theiler, J.; Abfalterer, W.; Hengartner, N.; Giorgi, E.E.; Bhattacharya, T.; Foley, B.; et al. Tracking Changes in SARS-CoV-2 Spike: Evidence that D614G Increases Infectivity of the COVID-19 Virus. Cell 2020, 182, 812–827. [Google Scholar] [CrossRef] [PubMed]
- Plante, J.A.; Liu, Y.; Liu, J.; Xia, H.; Johnson, B.A.; Lokugamage, K.G.; Zhang, X.; Muruato, A.E.; Zou, J.; Fontes-Garfias, C.R.; et al. Spike mutation D614G alters SARS-CoV-2 fitness. Nature 2021, 592, 116–121. [Google Scholar] [CrossRef] [PubMed]
- da Silva, S.J.R.; de Lima, S.C.; da Silva, R.C.; Kohl, A.; Pena, L. Viral Load in COVID-19 Patients: Implications for Prognosis and Vaccine Efficacy in the Context of Emerging SARS-CoV-2 Variants. Front. Med. 2022, 8, 836826. [Google Scholar] [CrossRef]
- da Silva, S.J.R.; Pena, L. Collapse of the public health system and the emergence of new variants during the second wave of the COVID-19 pandemic in Brazil. One Heal. 2021, 13, 100287–100287. [Google Scholar] [CrossRef]
- Coronavirus disease (COVID-19): Variants of SARS-COV-2. Accessed , 2023. https://www.who.int/emergencies/diseases/novel-coronavirus-2019/question-and-answers-hub/q-a-detail/coronavirus-disease-%28covid-19%29-variants-of-sars-cov-2? 5 April.
- WHO. COVID-19 Variants. 2023. https://www.who.int/activities/tracking-SARS-CoV-2-variants (2023-08-23).
- Hemo, M.K.M.; Islam, A. JN. 1 as a new variant of COVID-19 – editorial. Ann. Med. Surg. 2024, 86, 1833–1835. [Google Scholar] [CrossRef]
- Rahimi, F.; Abadi, A.T.B. Implications of the Emergence of a New Variant of SARS-CoV-2, VUI-202012/01. Arch. Med Res. 2021, 52, 569–571. [Google Scholar] [CrossRef]
- Utsumi, T.; Lusida, M.I.; Dinana, Z.; Wahyuni, R.M.; Yamani, L.N.; Juniastuti; Soetjipto; Matsui, C. ; Deng, L.; Abe, T.; et al. Occurrence of norovirus infection in an asymptomatic population in Indonesia. Infect. Genet. Evol. 2017, 55, 1–7. [Google Scholar] [CrossRef]
- Davies, N.G.; Abbott, S.; Barnard, R.C.; Jarvis, C.I.; Kucharski, A.J.; Munday, J.D.; Pearson, C.A.B.; Russell, T.W.; Tully, D.C.; Washburne, A.D.; et al. Estimated transmissibility and impact of SARS-CoV-2 lineage B.1.1.7 in England. Science 2021, 372, eabg3055. [Google Scholar] [CrossRef]
- Davies, N.G.; Jarvis, C.I.; Edmunds, W.J. Increased hazard of death in community-tested cases of SARS-CoV-2 Variant of Concern 202012/01. medRxiv 2021. [Google Scholar] [CrossRef]
- Calistri, P.; Amato, L.; Puglia, I.; Cito, F.; Di Giuseppe, A.; Danzetta, M.L.; Morelli, D.; Di Domenico, M.; Caporale, M.; Scialabba, S.; et al. Infection sustained by lineage B.1.1.7 of SARS-CoV-2 is characterised by longer persistence and higher viral RNA loads in nasopharyngeal swabs. Int. J. Infect. Dis. 2021, 105, 753–755. [Google Scholar] [CrossRef]
- Frampton, D.; Rampling, T.; Cross, A.; Bailey, H.; Heaney, J.; Byott, M.; Sconza, R.; Price, J.; Margaritis, M.; Bergstrom, M.; et al. Genomic characteristics and clinical effect of the emergent SARS-CoV-2 B.1.1.7 lineage in London, UK: a whole-genome sequencing and hospital-based cohort study. Lancet Infect. Dis. 2021, 21, 1246–1256. [Google Scholar] [CrossRef]
- Bal A, Destras G, Gaymard A, et al. Two-step strategy for the identification of SARS-CoV-2 variant of concern 202012/01 and other variants with spike deletion H69–V70, France, August to 20. Eurosurveillance. 2021;26(3):2100008. 20 December. [CrossRef]
- Funk T, Pharris A, Spiteri G, et al. Characteristics of SARS-CoV-2 variants of concern B.1.1.7, B.1.351 or P.1: data from seven EU/EEA countries, weeks 38/2020 to 10/2021. Eurosurveillance. 2021;26(16):2100348. [CrossRef]
- Challen, R.; Brooks-Pollock, E.; Read, J.M.; Dyson, L.; Tsaneva-Atanasova, K.; Danon, L. Risk of mortality in patients infected with SARS-CoV-2 variant of concern 202012/1: matched cohort study. BMJ 2021, 372, n579. [Google Scholar] [CrossRef] [PubMed]
- Rees-Spear, C.; Muir, L.; Griffith, S.A.; Heaney, J.; Aldon, Y.; Snitselaar, J.L.; Thomas, P.; Graham, C.; Seow, J.; Lee, N.; et al. The effect of spike mutations on SARS-CoV-2 neutralization. Cell Rep. 2021, 34, 108890. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Nair, M.S.; Liu, L.; Iketani, S.; Luo, Y.; Guo, Y.; Wang, M.; Yu, J.; Zhang, B.; Kwong, P.D.; et al. Antibody resistance of SARS-CoV-2 variants B.1.351 and B.1.1.7. Nature 2021, 593, 130–135. [Google Scholar] [CrossRef]
- Planas, D.; Bruel, T.; Grzelak, L.; Guivel-Benhassine, F.; Staropoli, I.; Porrot, F.; Planchais, C.; Buchrieser, J.; Rajah, M.M.; Bishop, E.; et al. Sensitivity of infectious SARS-CoV-2 B.1.1.7 and B.1.351 variants to neutralizing antibodies. Nat. Med. 2021, 27, 917–924. [Google Scholar] [CrossRef] [PubMed]
- Rondinone, V.; Pace, L.; Fasanella, A.; Manzulli, V.; Parisi, A.; Capobianchi, M.R.; Ostuni, A.; Chironna, M.; Caprioli, E.; Labonia, M.; et al. VOC 202012/01 Variant Is Effectively Neutralized by Antibodies Produced by Patients Infected before Its Diffusion in Italy. Viruses 2021, 13, 276. [Google Scholar] [CrossRef] [PubMed]
- Abu-Raddad, L.J.; Chemaitelly, H.; Butt, A.A. Effectiveness of the BNT162b2 Covid-19 Vaccine against the B.1.1.7 and B.1.351 Variants. N. Engl. J. Med. 2021, 385, 187–189. [Google Scholar] [CrossRef]
- Muik, A.; Wallisch, A.-K.; Sänger, B.; Swanson, K.A.; Mühl, J.; Chen, W.; Cai, H.; Maurus, D.; Sarkar, R.; Türeci, Ö.; et al. Neutralization of SARS-CoV-2 lineage B.1.1.7 pseudovirus by BNT162b2 vaccine–elicited human sera. Science 2021, 371, 1152–1153. [Google Scholar] [CrossRef]
- Sapkal, G.N.; Yadav, P.D.; Ella, R.; Deshpande, G.R.; Sahay, R.R.; Gupta, N.; Vadrevu, K.M.; Abraham, P.; Panda, S.; Bhargava, B. Inactivated COVID-19 vaccine BBV152/COVAXIN effectively neutralizes recently emerged B.1.1.7 variant of SARS-CoV-2. J. Travel Med. 2021, 28. [Google Scholar] [CrossRef]
- Wang GL, Wang ZY, Duan LJ, et al. Susceptibility of Circulating SARS-CoV-2 Variants to Neutralization. N Engl J Med. 2021;384(24):2354-2356. [CrossRef]
- Emary, K.R.W.; Golubchik, T.; Aley, P.K.; Ariani, C.V.; Angus, B.; Bibi, S.; Blane, B.; Bonsall, D.; Cicconi, P.; Charlton, S.; et al. Efficacy of ChAdOx1 nCoV-19 (AZD1222) vaccine against SARS-CoV-2 variant of concern 202012/01 (B.1.1.7): an exploratory analysis of a randomised controlled trial. Lancet 2021, 397, 1351–1362. [Google Scholar] [CrossRef] [PubMed]
- Ikegame, S.; Siddiquey, M.N.A.; Hung, C.-T.; Haas, G.; Brambilla, L.; Oguntuyo, K.Y.; Kowdle, S.; Chiu, H.-P.; Stevens, C.S.; Vilardo, A.E.; et al. Neutralizing activity of Sputnik V vaccine sera against SARS-CoV-2 variants. Nat. Commun. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Tegally, H.; Wilkinson, E.; Giovanetti, M.; Iranzadeh, A.; Fonseca, V.; Giandhari, J.; Doolabh, D.; Pillay, S.; San, E.J.; Msomi, N.; et al. Detection of a SARS-CoV-2 variant of concern in South Africa. Nature 2021, 592, 438–443. [Google Scholar] [CrossRef] [PubMed]
- Salehi-Vaziri, M.; Fazlalipour, M.; Khorrami, S.M.S.; Azadmanesh, K.; Pouriayevali, M.H.; Jalali, T.; Shoja, Z.; Maleki, A. The ins and outs of SARS-CoV-2 variants of concern (VOCs). Arch. Virol. 2022, 167, 327–344. [Google Scholar] [CrossRef]
- Edara VV, Norwood C, Floyd K, et al. Infection- and vaccine-induced antibody binding and neutralization of the B.1.351 SARS-CoV-2 variant. Cell Host Microbe. 2021; 29(4): 516-521.e3. Accessed April 5, 2023. [Google Scholar]
- Garcia-Beltran, W.F.; Lam, E.C.; St Denis, K.; Nitido, A.D.; Garcia, Z.H.; Hauser, B.M.; Feldman, J.; Pavlovic, M.N.; Gregory, D.J.; Poznansky, M.C.; et al. Multiple SARS-CoV-2 variants escape neutralization by vaccine-induced humoral immunity. Cell 2021, 184, 2372–2383. [Google Scholar] [CrossRef]
- Kuzmina, A.; Khalaila, Y.; Voloshin, O.; Keren-Naus, A.; Boehm-Cohen, L.; Raviv, Y.; Shemer-Avni, Y.; Rosenberg, E.; Taube, R. SARS-CoV-2 spike variants exhibit differential infectivity and neutralization resistance to convalescent or post-vaccination sera. Cell Host Microbe 2021, 29, 522–528. [Google Scholar] [CrossRef]
- Zhou, D.; Dejnirattisai, W.; Supasa, P.; Liu, C.; Mentzer, A.J.; Ginn, H.M.; Zhao, Y.; Duyvesteyn, H.M.; Tuekprakhon, A.; Nutalai, R.; et al. Evidence of escape of SARS-CoV-2 variant B.1.351 from natural and vaccine-induced sera. Cell 2021, 184, 2348–2361. [Google Scholar] [CrossRef] [PubMed]
- Madhi, S.A.; Baillie, V.; Cutland, C.L.; Voysey, M.; Koen, A.L.; Fairlie, L.; Padayachee, S.D.; Dheda, K.; Barnabas, S.L.; Bhorat, Q.E.; et al. Efficacy of the ChAdOx1 nCoV-19 Covid-19 Vaccine against the B.1.351 Variant. N. Engl. J. Med. 2021, 384, 1885–1898. [Google Scholar] [CrossRef]
- Iturriza-Gomara, M.; O’brien, S.J. Foodborne viral infections. Curr. Opin. Infect. Dis. 2016, 29, 495–501. [Google Scholar] [CrossRef]
- Faria, N.R.; Mellan, T.A.; Whittaker, C.; Claro, I.M.; Candido, D.D.S.; Mishra, S.; Crispim, M.A.E.; Sales, F.C.S.; Hawryluk, I.; McCrone, J.T.; et al. Genomics and epidemiology of the P.1 SARS-CoV-2 lineage in Manaus, Brazil. Science 2021, 372, 815–821. [Google Scholar] [CrossRef]
- Naveca, F.G.; Nascimento, V.; de Souza, V.C.; Corado, A.d.L.; Nascimento, F.; Silva, G.; Costa. ; Duarte, D.; Pessoa, K.; Mejía, M.; et al. COVID-19 in Amazonas, Brazil, was driven by the persistence of endemic lineages and P.1 emergence. Nat. Med. 2021, 27, 1230–1238. [Google Scholar] [CrossRef] [PubMed]
- Romano, C.M.; Felix, A.C.; de Paula, A.V.; de Jesus, J.G.; Andrade, P.S.; Cândido, D.; de Oliveira, F.M.; Ribeiro, A.C.; da Silva, F.C.; Inemami, M.; et al. SARS-CoV-2 reinfection caused by the P.1 lineage in Araraquara city, Sao Paulo State, Brazil. Rev. do Inst. de Med. Trop. de Sao Paulo 2021, 63, e36. [Google Scholar] [CrossRef]
- Levidiotou, S.; Gartzonika, C.; Papaventsis, D.; Christaki, C.; Priavali, E.; Zotos, N.; Kapsali, E.; Vrioni, G. Viral agents of acute gastroenteritis in hospitalized children in Greece. Clin. Microbiol. Infect. 2009, 15, 596–598. [Google Scholar] [CrossRef] [PubMed]
- Sabino, E.C.; Buss, L.F.; Carvalho, M.P.S.; Prete, C.A., Jr.; Crispim, M.A.E.; Fraiji, N.A.; Pereira, R.H.M.; Parag, K.V.; da Silva Peixoto, P.; Kraemer, M.U.G.; et al. Resurgence of COVID-19 in Manaus, Brazil, despite high seroprevalence. Lancet 2021, 397, 452–455. [Google Scholar] [CrossRef]
- Chen, R.E.; Zhang, X.; Case, J.B.; Winkler, E.S.; Liu, Y.; VanBlargan, L.A.; Liu, J.; Errico, J.M.; Xie, X.; Suryadevara, N.; et al. Resistance of SARS-CoV-2 variants to neutralization by monoclonal and serum-derived polyclonal antibodies. Nat. Med. 2021, 27, 717–726. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Casner, R.G.; Nair, M.S.; Wang, M.; Yu, J.; Cerutti, G.; Liu, L.; Kwong, P.D.; Huang, Y.; Shapiro, L.; et al. Increased resistance of SARS-CoV-2 variant P.1 to antibody neutralization. Cell Host Microbe 2021, 29, 747–751. [Google Scholar] [CrossRef]
- Dejnirattisai, W.; Zhou, D.; Supasa, P.; Liu, C.; Mentzer, A.J.; Ginn, H.M.; Zhao, Y.; Duyvesteyn, H.M.; Tuekprakhon, A.; Nutalai, R.; et al. Antibody evasion by the P.1 strain of SARS-CoV-2. Cell 2021, 184, 2939–2954. [Google Scholar] [CrossRef]
- Fabiani, M.; Margiotti, K.; Viola, A.; Mesoraca, A.; Giorlandino, C. Mild Symptomatic SARS-CoV-2 P.1 (B.1.1.28) Infection in a Fully Vaccinated 83-Year-Old Man. Pathogens 2021, 10, 614. [Google Scholar] [CrossRef]
- Ahmad, A.; Fawaz, M.A.M.; Aisha, A. A comparative overview of SARS-CoV-2 and its variants of concern. 2022, 30, 328–343. [CrossRef]
- Campbell, F.; Archer, B.; Laurenson-Schafer, H.; Jinnai, Y.; Konings, F.; Batra, N.; Pavlin, B.; Vandemaele, K.; Van Kerkhove, M.D.; Jombart, T.; et al. Increased transmissibility and global spread of SARS-CoV-2 variants of concern as at June 2021. Eurosurveillance 2021, 26, 2100509. [Google Scholar] [CrossRef]
- Ong, S.W.X.; Chiew, C.J.; Ang, L.W.; Mak, T.M.; Cui, L.; Toh, M.P.H.S.; Lim, Y.D.; Lee, P.H.; Lee, T.H.; Chia, P.Y.; et al. Clinical and Virological Features of Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Variants of Concern: A Retrospective Cohort Study Comparing B.1.1.7 (Alpha), B.1.351 (Beta), and B.1.617.2 (Delta). Clin. Infect. Dis. 2021, 75, e1128–e1136. [Google Scholar] [CrossRef]
- Williams, G.H.; Llewelyn, A.; Brandao, R.; Chowdhary, K.; Hardisty, K.-M.; Loddo, M. SARS-CoV-2 testing and sequencing for international arrivals reveals significant cross border transmission of high risk variants into the United Kingdom. eClinicalMedicine 2021, 38, 101021. [Google Scholar] [CrossRef] [PubMed]
- Infections caused by the Delta variant (B.1.617.2) of SARS-CoV-2 are associated with increased viral loads compared to infections with the Alpha variant (B.1.1.7) or non-Variants of Concern. Published online , 2021. 4 August. [CrossRef]
- Sheikh, A.; McMenamin, J.; Taylor, B.; Robertson, C. SARS-CoV-2 Delta VOC in Scotland: demographics, risk of hospital admission, and vaccine effectiveness. Lancet 2021, 397, 2461–2462. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Ginn, H.M.; Dejnirattisai, W.; Supasa, P.; Wang, B.; Tuekprakhon, A.; Nutalai, R.; Zhou, D.; Mentzer, A.J.; Zhao, Y.; et al. Reduced neutralization of SARS-CoV-2 B.1.617 by vaccine and convalescent serum. Cell 2021, 184, 4220–4236. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Liu, Y.; Xia, H.; Zou, J.; Weaver, S.C.; Swanson, K.A.; Cai, H.; Cutler, M.; Cooper, D.; Muik, A.; et al. BNT162b2-elicited neutralization of B.1.617 and other SARS-CoV-2 variants. Nature 2021, 596, 273–275. [Google Scholar] [CrossRef] [PubMed]
- Wall, E.C.; Wu, M.; Harvey, R.; Kelly, G.; Warchal, S.; Sawyer, C.; Daniels, R.; Hobson, P.; Hatipoglu, E.; Ngai, Y.; et al. Neutralising antibody activity against SARS-CoV-2 VOCs B.1.617.2 and B.1.351 by BNT162b2 vaccination. Lancet 2021, 397, 2331–2333. [Google Scholar] [CrossRef]
- Lustig, Y.; Zuckerman, N.; Nemet, I.; Atari, N.; Kliker, L.; Regev-Yochay, G.; Sapir, E.; Mor, O.; Alroy-Preis, S.; Mendelson, E.; et al. Neutralising capacity against Delta (B.1.617.2) and other variants of concern following Comirnaty (BNT162b2, BioNTech/Pfizer) vaccination in health care workers, Israel. Eurosurveillance 2021, 26, 2100557. [Google Scholar] [CrossRef]
- Nasreen, S.; Chung, H.; He, S.; Brown, K.A.; Gubbay, J.B.; Buchan, S.A.; Fell, D.B.; Austin, P.C.; Schwartz, K.L.; Sundaram, M.E.; et al. Effectiveness of COVID-19 vaccines against symptomatic SARS-CoV-2 infection and severe outcomes with variants of concern in Ontario. Nat. Microbiol. 2022, 7, 379–385. [Google Scholar] [CrossRef]
- Chia, P.Y.; Ong, S.W.X.; Chiew, C.J.; Ang, L.W.; Chavatte, J.-M.; Mak, T.-M.; Cui, L.; Kalimuddin, S.; Ni Chia, W.; Tan, C.W.; et al. Virological and serological kinetics of SARS-CoV-2 Delta variant vaccine breakthrough infections: a multicentre cohort study. Clin. Microbiol. Infect. 2021, 28, 612–e1. [Google Scholar] [CrossRef]
- Kang, M.; Yi, Y.; Li, Y.; Sun, L.; Deng, A.; Hu, T.; Zhang, J.; Liu, J.; Cheng, M.; Xie, S.; et al. Effectiveness of Inactivated COVID-19 Vaccines Against Illness Caused by the B.1.617.2 (Delta) Variant During an Outbreak in Guangdong, China. Ann. Intern. Med. 2022, 175, 533–540. [Google Scholar] [CrossRef]
- Yadav, P.D.; Sahay, R.R.; Sapkal, G.; Nyayanit, D.; Shete, A.M.; Deshpande, G.; Patil, D.Y.; Gupta, N.; Kumar, S.; Abraham, P.; et al. Comparable neutralization of SARS-CoV-2 Delta AY.1 and Delta with individuals sera vaccinated with BBV152. J. Travel Med. 2021, 28. [Google Scholar] [CrossRef]
- Gushchin, V.A.; Dolzhikova, I.V.; Shchetinin, A.M.; Odintsova, A.S.; Siniavin, A.E.; Nikiforova, M.A.; Pochtovyi, A.A.; Shidlovskaya, E.V.; Kuznetsova, N.A.; Burgasova, O.A.; et al. Neutralizing Activity of Sera from Sputnik V-Vaccinated People against Variants of Concern (VOC: B.1.1.7, B.1.351, P.1, B.1.617.2, B.1.617.3) and Moscow Endemic SARS-CoV-2 Variants. Vaccines 2021, 9, 779. [Google Scholar] [CrossRef] [PubMed]
- Jongeneelen M, Kaszas K, Veldman D, et al. Ad26.COV2.S elicited neutralizing activity against Delta and other SARS-CoV-2 variants of concern. bioRxiv. Published online , 2021:2021.07.01.450707. 1 July. [CrossRef]
- Belik, M.; Jalkanen, P.; Lundberg, R.; Reinholm, A.; Laine, L.; Väisänen, E.; Skön, M.; Tähtinen, P.A.; Ivaska, L.; Pakkanen, S.H.; et al. Comparative analysis of COVID-19 vaccine responses and third booster dose-induced neutralizing antibodies against Delta and Omicron variants. Nat. Commun. 2022, 13, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Colosi, E.; Bassignana, G.; Barrat, A.; Lina, B.; Vanhems, P.; Bielicki, J.; Colizza, V. Minimising school disruption under high incidence conditions due to the Omicron variant in France, Switzerland, Italy, in January 2022. Eurosurveillance 2023, 28, 2200192. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Li, Z.; Liu, Y. An overview of the safety, clinical application and antiviral research of the COVID-19 therapeutics. J. Infect. Public Heal. 2020, 13, 1405–1414. [Google Scholar] [CrossRef]
- Kadam, R.U.; Wilson, I.A. Structural basis of influenza virus fusion inhibition by the antiviral drug Arbidol. Proc. Natl. Acad. Sci. USA 2017, 114, 206–214. [Google Scholar] [CrossRef]
- Ahmad, G.; Sohail, M.; Bilal, M.; Rasool, N.; Qamar, M.U.; Ciurea, C.; Marceanu, L.G.; Misarca, C. N-Heterocycles as Promising Antiviral Agents: A Comprehensive Overview. Molecules 2024, 29, 2232. [Google Scholar] [CrossRef]
- Agrawal, U.; Raju, R.; Udwadia, Z.F. Favipiravir: A new and emerging antiviral option in COVID-19. Med J. Armed Forces India 2020, 76, 370–376. [Google Scholar] [CrossRef]
- Zhu, Z.; Lu, Z.; Xu, T.; Chen, C.; Yang, G.; Zha, T.; Lu, J.; Xue, Y. Arbidol monotherapy is superior to lopinavir/ritonavir in treating COVID-19. J. Infect. 2020, 81, e21–e23. [Google Scholar] [CrossRef]
- Saul, S.; Einav, S. Old Drugs for a New Virus: Repurposed Approaches for Combating COVID-19. ACS Infect. Dis. 2020, 6, 2304–2318. [Google Scholar] [CrossRef]
- Vanderlinden, E.; Vrancken, B.; Van Houdt, J.; Rajwanshi, V.K.; Gillemot, S.; Andrei, G.; Lemey, P.; Naesens, L. Distinct Effects of T-705 (Favipiravir) and Ribavirin on Influenza Virus Replication and Viral RNA Synthesis. Antimicrob. Agents Chemother. 2016, 60, 6679–6691. [Google Scholar] [CrossRef]
- Singh, A.V.; Kayal, A.; Malik, A.; Maharjan, R.S.; Dietrich, P.; Thissen, A.; Siewert, K.; Curato, C.; Pande, K.; Prahlad, D.; et al. Interfacial Water in the SARS Spike Protein: Investigating the Interaction with Human ACE2 Receptor and In Vitro Uptake in A549 Cells. Langmuir 2022, 38, 7976–7988. [Google Scholar] [CrossRef] [PubMed]
- Uno, Y. Camostat mesilate therapy for COVID-19. Intern. Emerg. Med. 2020, 15, 1577–1578. [Google Scholar] [CrossRef]
- Gasmi, A.; Noor, S.; Tippairote, T.; Dadar, M.; Menzel, A.; Bjørklund, G. Individual risk management strategy and potential therapeutic options for the COVID-19 pandemic. Clin. Immunol. 2020, 215, 108409–108409. [Google Scholar] [CrossRef] [PubMed]
- Kawase, M.; Shirato, K.; van der Hoek, L.; Taguchi, F.; Matsuyama, S. Simultaneous Treatment of Human Bronchial Epithelial Cells with Serine and Cysteine Protease Inhibitors Prevents Severe Acute Respiratory Syndrome Coronavirus Entry. J. Virol. 2012, 86, 6537–6545. [Google Scholar] [CrossRef]
- Coronavirus disease (COVID-19). Accessed , 2023. https://www.who.int/emergencies/diseases/novel-coronavirus-2019? 12 April.
- Liu, F.; Xu, A.; Zhang, Y.; Xuan, W.; Yan, T.; Pan, K.; Yu, W.; Zhang, J. Patients of COVID-19 may benefit from sustained Lopinavir-combined regimen and the increase of Eosinophil may predict the outcome of COVID-19 progression. Int. J. Infect. Dis. 2020, 95, 183–191. [Google Scholar] [CrossRef]
- Chu, C.M.; Cheng, V.C.C.; Hung, I.F.N.; Wong, M.M.L.; Chan, K.H.; Chan, K.S.; Kao, R.Y.T.; Poon, L.L.M.; Wong, C.L.P.; Guan, Y.; et al. Role of lopinavir/ritonavir in the treatment of SARS: Initial virological and clinical findings. Thorax 2004, 59, 252–256. [Google Scholar] [CrossRef] [PubMed]
- Cao, B.; Wang, Y.; Wen, D.; Liu, W.; Wang, J.; Fan, G.; Ruan, L.; Song, B.; Cai, Y.; Wei, M.; et al. A Trial of Lopinavir–Ritonavir in Adults Hospitalized with Severe COVID-19. N. Engl. J. Med. 2020, 382, 1787–1799. [Google Scholar] [CrossRef] [PubMed]
- Phadke, M.; Saunik, S. COVID-19 treatment by repurposing drugs until the vaccine is in sight. Drug Dev. Res. 2020, 81, 541–543. [Google Scholar] [CrossRef]
- Bege, M.; Borbás, A. The Design, Synthesis and Mechanism of Action of Paxlovid, a Protease Inhibitor Drug Combination for the Treatment of COVID-19. Pharmaceutics 2024, 16, 217. [Google Scholar] [CrossRef]
- Sheahan, T.P.; Sims, A.C.; Leist, S.R.; Schäfer, A.; Won, J.; Brown, A.J.; Montgomery, S.A.; Hogg, A.; Babusis, D.; Clarke, M.O.; et al. Comparative therapeutic efficacy of remdesivir and combination lopinavir, ritonavir, and interferon beta against MERS-CoV. Nat. Commun. 2020, 11, 222. [Google Scholar] [CrossRef]
- Eastman, R.T.; Roth, J.S.; Brimacombe, K.R.; Simeonov, A.; Shen, M.; Patnaik, S.; Hall, M.D. Remdesivir: A Review of Its Discovery and Development Leading to Emergency Use Authorization for Treatment of COVID-19. ACS Central Sci. 2020, 6, 672–683. [Google Scholar] [CrossRef] [PubMed]
- Seiwert, S.D.; Andrews, S.W.; Jiang, Y.; Serebryany, V.; Tan, H.; Kossen, K.; Rajagopalan, P.T.R.; Misialek, S.; Stevens, S.K.; Stoycheva, A.; et al. Preclinical Characteristics of the Hepatitis C Virus NS3/4A Protease Inhibitor ITMN-191 (R7227). Antimicrob. Agents Chemother. 2008, 52, 4432–4441. [Google Scholar] [CrossRef]
- Altay, O.; Mohammadi, E.; Lam, S.; Turkez, H.; Boren, J.; Nielsen, J.; Uhlen, M.; Mardinoglu, A. Current Status of COVID-19 Therapies and Drug Repositioning Applications. iScience 2020, 23, 101303. [Google Scholar] [CrossRef]
- Wei, L.; Shang, J.; Ma, Y.; Xu, X.; Huang, Y.; Guan, Y.; Duan, Z.; Zhang, W.; Gao, Z.; Zhang, M.; et al. Efficacy and Safety of 12-week Interferon-based Danoprevir Regimen in Patients with Genotype 1 Chronic Hepatitis C. J. Clin. Transl. Hepatol. 2019, 7, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Feng, B.; Guan, Y.; Zheng, S.; Sheng, J.; Yang, X.; Ma, Y.; Huang, Y.; Kang, Y.; Wen, X.; et al. Efficacy and Safety of All-oral, 12-week Ravidasvir Plus Ritonavir-boosted Danoprevir and Ribavirin in Treatment-naïve Noncirrhotic HCV Genotype 1 Patients: Results from a Phase 2/3 Clinical Trial in China. J. Clin. Transl. Hepatol. 2019, 7, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Nicastri, E.; Petrosillo, N.; Ascoli Bartoli, T.; Lepore, L.; Mondi, A.; Palmieri, F.; D’Offizi, G.; Marchioni, L.; Murachelli, S.; Ippolito, G.; et al. National Institute for the Infectious Diseases “L. Spallanzani” IRCCS. Recommendations for COVID-19 Clinical Management. Infect. Dis. Rep. 2020, 12, 8543. [Google Scholar] [CrossRef]
- Fintelman-Rodrigues, N.; Sacramento, C.Q.; Lima, C.R.; da Silva, F.S.; Ferreira, A.C.; Mattos, M.; de Freitas, C.S.; Soares, V.C.; Dias, S.d.S.G.; Temerozo, J.R.; et al. Atazanavir, Alone or in Combination with Ritonavir, Inhibits SARS-CoV-2 Replication and Proinflammatory Cytokine Production. Antimicrob. Agents Chemother. 2020, 64. [Google Scholar] [CrossRef]
- Stanley, T.L.; Joy, T.; Hadigan, C.M.; Liebau, J.G.; Makimura, H.; Chen, C.Y.; Thomas, B.J.; Weise, S.B.; Robbins, G.K.; Grinspoon, S.K. Effects of switching from lopinavir/ritonavir to atazanavir/ritonavir on muscle glucose uptake and visceral fat in HIV-infected patients. AIDS 2009, 23, 1349–1357. [Google Scholar] [CrossRef]
- Vatansever, E.C.; Yang, K.S.; Drelich, A.K.; Kratch, K.C.; Cho, C.-C.; Kempaiah, K.R.; Hsu, J.C.; Mellott, D.M.; Xu, S.; Tseng, C.-T.K.; et al. Bepridil is potent against SARS-CoV-2 in vitro. Proc. Natl. Acad. Sci. 2021, 118. [Google Scholar] [CrossRef]
- Hall, D.C.; Ji, H.-F. A search for medications to treat COVID-19 via in silico molecular docking models of the SARS-CoV-2 spike glycoprotein and 3CL protease. Travel Med. Infect. Dis. 2020, 35, 101646–101646. [Google Scholar] [CrossRef]
- Yamamoto N, Matsuyama S, Hoshino T, Yamamoto N. Nelfinavir inhibits replication of severe acute respiratory syndrome coronavirus 2 in vitro. bioRxiv. Published online , 2020:2020.04.06.026476. 8 April. [CrossRef]
- Ul Qamar, M.T.; Alqahtani, S.M.; Alamri, M.A.; Chen, L.-L. Structural basis of SARS-CoV-2 3CLpro and anti-COVID-19 drug discovery from medicinal plants. J. Pharm. Anal. 2020, 10, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Khan, R.J.; Jha, R.K.; Amera, G.M.; Jain, M.; Singh, E.; Pathak, A.; Singh, R.P.; Muthukumaran, J.; Singh, A.K. Targeting SARS-CoV-2: a systematic drug repurposing approach to identify promising inhibitors against 3C-like proteinase and 2′-O-ribose methyltransferase. J. Biomol. Struct. Dyn. 2020, 39, 2679–2692. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.T.; Pandey, I.; Zamboni, P.; Gemmati, D.; Kanase, A.; Singh, A.V.; Singh, M.P. Traditional Herbal Remedies with a Multifunctional Therapeutic Approach as an Implication in COVID-19 Associated Co-Infections. Coatings 2020, 10, 761. [Google Scholar] [CrossRef]
- Choi, H.S.; Choi, A.Y.; Kopp, J.B.; Winkler, C.A.; Cho, S.K. Review of COVID-19 Therapeutics by Mechanism: From Discovery to Approval. J. Korean Med Sci. 2024, 39, e134. [Google Scholar] [CrossRef]
- Wu, C.-J.; Jan, J.-T.; Chen, C.-M.; Hsieh, H.-P.; Hwang, D.-R.; Liu, H.-W.; Liu, C.-Y.; Huang, H.-W.; Chen, S.-C.; Hong, C.-F.; et al. Inhibition of Severe Acute Respiratory Syndrome Coronavirus Replication by Niclosamide. Antimicrob. Agents Chemother. 2004, 48, 2693–6. [Google Scholar] [CrossRef]
- Anand, K.; Ziebuhr, J.; Wadhwani, P.; Mesters, J.R.; Hilgenfeld, R. Coronavirus Main Proteinase (3CL pro ) Structure: Basis for Design of Anti-SARS Drugs. Science 2003, 300, 1763–1767. [Google Scholar] [CrossRef]
- Yang, H.; Yang, M.; Ding, Y.; Liu, Y.; Lou, Z.; Zhou, Z.; Sun, L.; Mo, L.; Ye, S.; Pang, H.; et al. The crystal structures of severe acute respiratory syndrome virus main protease and its complex with an inhibitor. Proc. Natl. Acad. Sci. 2003, 100, 13190–13195. [Google Scholar] [CrossRef] [PubMed]
- Chou, K.-C.; Wei, D.-Q.; Zhong, W.-Z. Binding mechanism of coronavirus main proteinase with ligands and its implication to drug design against SARS. Biochem. Biophys. Res. Commun. 2003, 308, 148–151. [Google Scholar] [CrossRef]
- Stockman, L.J.; Bellamy, R.; Garner, P. SARS: Systematic Review of Treatment Effects. PLOS Med. 2006, 3, e343. [Google Scholar] [CrossRef]
- A Burns-Naas, L.; Zorbas, M.; Jessen, B.; Evering, W.; Stevens, G.; Ivett, J.L.; E Ryan, T.; Cook, J.C.; Capen, C.C.; Chen, M.; et al. Increase in thyroid follicular cell tumors in nelfinavir-treated rats observed in a 2-year carcinogenicity study is consistent with a rat-specific mechanism of thyroid neoplasia. Hum. Exp. Toxicol. 2005, 24, 643–654. [Google Scholar] [CrossRef]
- Khaliq, Y.; Gallicano, K.; Tisdale, C.; Carignan, G.; Cooper, C.; McCarthy, A. Pharmacokinetic interaction between mefloquine and ritonavir in healthy volunteers. Br. J. Clin. Pharmacol. 2001, 51, 591–600. [Google Scholar] [CrossRef] [PubMed]
- Trapé, M.; Barnosky, S. Nelfinavir in Expanded Postexposure Prophylaxis Causing Acute Hepatitis With Cholestatic Features TWo Case Reports. Infect. Control. Hosp. Epidemiology 2001, 22, 333–334. [Google Scholar] [CrossRef]
- Müller, A.; Cadenas, E.; Graf, P.; Sies, H. A novel biologically active seleno-organic compound—1. Biochem. Pharmacol. 1984, 33, 3235–3239. [Google Scholar] [CrossRef]
- Parnham, M.J.; Sies, H. The early research and development of ebselen. Biochem. Pharmacol. 2013, 86, 1248–1253. [Google Scholar] [CrossRef]
- Sies, H. Ebselen, a selenoorganic compound as glutathione peroxidase mimic. Free. Radic. Biol. Med. 1993, 14, 313–323. [Google Scholar] [CrossRef]
- Azad, G.K.; Tomar, R.S. Ebselen, a promising antioxidant drug: mechanisms of action and targets of biological pathways. Mol. Biol. Rep. 2014, 41, 4865–4879. [Google Scholar] [CrossRef]
- Kil, J.; Lobarinas, E.; Spankovich, C.; Griffiths, S.K.; Antonelli, P.J.; Lynch, E.D.; Le Prell, C.G. Safety and efficacy of ebselen for the prevention of noise-induced hearing loss: a randomised, double-blind, placebo-controlled, phase 2 trial. Lancet 2017, 390, 969–979. [Google Scholar] [CrossRef]
- Wendel, A.; Fausel, M.; Safayhi, H.; Tiegs, G.; Otter, R. A novel biologically active seleno-organic compound—II. Biochem. Pharmacol. 1984, 33, 3241–3245. [Google Scholar] [CrossRef] [PubMed]
- Martinez, M.A. Compounds with Therapeutic Potential against Novel Respiratory 2019 Coronavirus. Antimicrob. Agents Chemother. 2020, 64, e00399–20. [Google Scholar] [CrossRef]
- Elfiky, A.A. Ribavirin, Remdesivir, Sofosbuvir, Galidesivir, and Tenofovir against SARS-CoV-2 RNA dependent RNA polymerase (RdRp): A molecular docking study. Life Sci. 2020, 253, 117592. [Google Scholar] [CrossRef]
- Amirian, E.S.; Levy, J.K. Current knowledge about the antivirals remdesivir (GS-5734) and GS-441524 as therapeutic options for coronaviruses. One Heal. 2020, 9, 100128. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.-C.; Deng, Q.-X.; Dai, S.-X. Remdesivir for severe acute respiratory syndrome coronavirus 2 causing COVID-19: An evaluation of the evidence. Travel Med. Infect. Dis. 2020, 35, 101647–101647. [Google Scholar] [CrossRef] [PubMed]
- Palazzotti, D.; Sguilla, M.; Manfroni, G.; Cecchetti, V.; Astolfi, A.; Barreca, M.L. Small Molecule Drugs Targeting Viral Polymerases. Pharmaceuticals 2024, 17, 661. [Google Scholar] [CrossRef] [PubMed]
- Martinez, M.A. Efficacy of repurposed antiviral drugs: Lessons from COVID-19. Drug Discov. Today 2022, 27, 1954–1960. [Google Scholar] [CrossRef]
- Agostini, M.L.; Andres, E.L.; Sims, A.C.; Graham, R.L.; Sheahan, T.P.; Lu, X.; Smith, E.C.; Case, J.B.; Feng, J.Y.; Jordan, R.; et al. Coronavirus Susceptibility to the Antiviral Remdesivir (GS-5734) Is Mediated by the Viral Polymerase and the Proofreading Exoribonuclease. mBio 2018, 9, e00221–18. [Google Scholar] [CrossRef]
- Medical Product Alert N°4/2021: Falsified remdesivir. Accessed , 2023. https://www.who. 13 April 2021.
- Cho, A.; Saunders, O.L.; Butler, T.; Zhang, L.; Xu, J.; Vela, J.E.; Feng, J.Y.; Ray, A.S.; Kim, C.U. Synthesis and antiviral activity of a series of 1′-substituted 4-aza-7,9-dideazaadenosine C-nucleosides. Bioorganic Med. Chem. Lett. 2012, 22, 2705–2707. [Google Scholar] [CrossRef]
- Sheahan, T.P.; Sims, A.C.; Graham, R.L.; Menachery, V.D.; Gralinski, L.E.; Case, J.B.; Leist, S.R.; Pyrc, K.; Feng, J.Y.; Trantcheva, I.; et al. Broad-spectrum antiviral GS-5734 inhibits both epidemic and zoonotic coronaviruses. Sci. Transl. Med. 2017, 9. [Google Scholar] [CrossRef]
- Holshue, M.L.; DeBolt, C.; Lindquist, S.; Lofy, K.H.; Wiesman, J.; Bruce, H.; Spitters, C.; Ericson, K.; Wilkerson, S.; Tural, A.; et al. First Case of 2019 Novel Coronavirus in the United States. N. Engl. J. Med. 2020, 382, 929–936. [Google Scholar] [CrossRef]
- Spinner, C.D.; Gottlieb, R.L.; Criner, G.J.; López, J.R.A.; Cattelan, A.M.; Viladomiu, A.S.; Ogbuagu, O.; Malhotra, P.; Mullane, K.M.; Castagna, A.; et al. Effect of Remdesivir vs Standard Care on Clinical Status at 11 Days in Patients With Moderate COVID-19: A Randomized Clinical Trial. JAMA 2020, 324, 1048–1057. [Google Scholar] [CrossRef]
- Norrie, J.D. Remdesivir for COVID-19: challenges of underpowered studies. Lancet 2020, 395, 1525–1527. [Google Scholar] [CrossRef]
- Malin, J.J.; Suárez, I.; Priesner, V.; Fätkenheuer, G.; Rybniker, J. Remdesivir against COVID-19 and Other Viral Diseases. Clin. Microbiol. Rev. 2020, 34. [Google Scholar] [CrossRef] [PubMed]
- Tzvetkov, N.T.; Peeva, M.I.; Georgieva, M.G.; Deneva, V.; Balacheva, A.A.; Bogdanov, I.P.; Ponticelli, M.; Milella, L.; Kirilov, K.; Matin, M.; et al. Favipiravir vs. Deferiprone: Tautomeric, photophysical, in vitro biological studies, and binding interactions with SARS-Cov-2-MPro/ACE2. Curr. Res. Biotechnol. 2024, 7. [Google Scholar] [CrossRef]
- Furuta, Y.; Gowen, B.B.; Takahashi, K.; Shiraki, K.; Smee, D.F.; Barnard, D.L. Favipiravir (T-705), a novel viral RNA polymerase inhibitor. Antivir. Res. 2013, 100, 446–454. [Google Scholar] [CrossRef] [PubMed]
- Jin, Z.; Smith, L.K.; Rajwanshi, V.K.; Kim, B.; Deval, J. The Ambiguous Base-Pairing and High Substrate Efficiency of T-705 (Favipiravir) Ribofuranosyl 5′-Triphosphate towards Influenza A Virus Polymerase. PLOS ONE 2013, 8, e68347. [Google Scholar] [CrossRef]
- Agrawal, U.; Raju, R.; Udwadia, Z.F. Favipiravir: A new and emerging antiviral option in COVID-19. Med J. Armed Forces India 2020, 76, 370–376. [Google Scholar] [CrossRef]
- Jean, S.-S.; Lee, P.-I.; Hsueh, P.-R. Treatment options for COVID-19: The reality and challenges. J. Microbiol. Immunol. Infect. 2020, 53, 436–443. [Google Scholar] [CrossRef]
- Booth, C.M. Clinical Features and Short-term Outcomes of 144 Patients With SARS in the Greater Toronto Area. JAMA 2003, 289, 2801–2809. [Google Scholar] [CrossRef] [PubMed]
- Sung, J.J.Y.; Wu, A.; Joynt, G.M.; Yuen, K.Y.; Lee, N.; Chan, P.K.S.; Cockram, C.S.; Ahuja, A.T.; Yu, L.M.; Wong, V.W.; et al. Severe acute respiratory syndrome: report of treatment and outcome after a major outbreak. Thorax 2004, 59, 414–420. [Google Scholar] [CrossRef]
- Pandey, A.; Nikam, A.N.; Shreya, A.B.; Mutalik, S.P.; Gopalan, D.; Kulkarni, S.; Padya, B.S.; Fernandes, G.; Mutalik, S.; Prassl, R. Potential therapeutic targets for combating SARS-CoV-2: Drug repurposing, clinical trials and recent advancements. Life Sci. 2020, 256, 117883–117883. [Google Scholar] [CrossRef]
- Cheng, C.-Y.; Lee, Y.-L.; Chen, C.-P.; Lin, Y.-C.; Liu, C.-E.; Liao, C.-H.; Cheng, S.-H. Lopinavir/ritonavir did not shorten the duration of SARS CoV-2 shedding in patients with mild pneumonia in Taiwan. J. Microbiol. Immunol. Infect. 2020, 53, 488–492. [Google Scholar] [CrossRef]
- Poulakou, G.; Barakat, M.; Israel, R.J.; Bacci, M.R. ; Virazole Collaborator Group for COVID-19 Respiratory Distress Ribavirin aerosol in hospitalized adults with respiratory distress and COVID-19: An open-label trial. Clin. Transl. Sci. 2022, 16, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Polo, R.; García-Albéniz, X.; Terán, C.; Morales, M.; Rial-Crestelo, D.; Garcinuño, M.A.; del Toro, M.G.; Hita, C.; Gómez-Sirvent, J.L.; Buzón, L.; et al. Daily tenofovir disoproxil fumarate/emtricitabine and hydroxychloroquine for pre-exposure prophylaxis of COVID-19: a double-blind placebo-controlled randomized trial in healthcare workers. Clin. Microbiol. Infect. 2022, 29, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Hung, I.F.-N.; Lung, K.-C.; Tso, E.Y.-K.; Liu, R.; Chung, T.W.-H.; Chu, M.-Y.; Ng, Y.-Y.; Lo, J.; Chan, J.; Tam, A.R.; et al. Triple combination of interferon beta-1b, lopinavir–ritonavir, and ribavirin in the treatment of patients admitted to hospital with COVID-19: an open-label, randomised, phase 2 trial. Lancet 2020, 395, 1695–1704. [Google Scholar] [CrossRef] [PubMed]
- Novel Coronavirus Information Center. Accessed , 2023. https://www.elsevier. 13 April.
- Warren, T.K.; Wells, J.; Panchal, R.G.; Stuthman, K.S.; Garza, N.L.; Van Tongeren, S.A.; Dong, L.; Retterer, C.J.; Eaton, B.P.; Pegoraro, G.; et al. Protection against filovirus diseases by a novel broad-spectrum nucleoside analogue BCX4430. Nature 2014, 508, 402–405. [Google Scholar] [CrossRef]
- Taylor, R.; Kotian, P.; Warren, T.; Panchal, R.; Bavari, S.; Julander, J.; Dobo, S.; Rose, A.; El-Kattan, Y.; Taubenheim, B.; et al. BCX4430 – A broad-spectrum antiviral adenosine nucleoside analog under development for the treatment of Ebola virus disease. J. Infect. Public Heal. 2016, 9, 220–226. [Google Scholar] [CrossRef]
- Welliver, R.; Monto, A.S.; Carewicz, O.; Schatteman, E.; Hassman, M.; Hedrick, J.; Jackson, H.C.; Huson, L.; Ward, P.; Oxford, J.S. Effectiveness of Oseltamivir in Preventing Influenza in Household ContactsA Randomized Controlled Trial. JAMA 2001, 285, 748–754. [Google Scholar] [CrossRef]
- Yousefi, B.; Valizadeh, S.; Ghaffari, H.; Vahedi, A.; Karbalaei, M.; Eslami, M. A global treatments for coronaviruses including COVID-19. J. Cell. Physiol. 2020, 235, 9133–9142. [Google Scholar] [CrossRef]
- Li, G.; De Clercq, E. Therapeutic Options for the 2019 Novel Coronavirus (2019-nCoV). Nat. Rev. Drug Discov. 2020, 19, 149–150. [Google Scholar] [CrossRef]
- Griffin, S.D.C.; Beales, L.P.; Clarke, D.S.; Worsfold, O.; Evans, S.D.; Jaeger, J.; Harris, M.P.G.; Rowlands, D.J. The p7 protein of hepatitis C virus forms an ion channel that is blocked by the antiviral drug, Amantadine. FEBS Lett. 2003, 535, 34–38. [Google Scholar] [CrossRef]
- Torres, J.; Maheswari, U.; Parthasarathy, K.; Ng, L.; Liu, D.X.; Gong, X. Conductance and amantadine binding of a pore formed by a lysine-flanked transmembrane domain of SARS coronavirus envelope protein. Protein Sci. 2007, 16, 2065–2071. [Google Scholar] [CrossRef]
- Smieszek, S.P.; Przychodzen, B.P.; Polymeropoulos, M.H. Amantadine disrupts lysosomal gene expression: A hypothesis for COVID19 treatment. Int. J. Antimicrob. Agents 2020, 55, 106004–106004. [Google Scholar] [CrossRef] [PubMed]
- Russell, B.; Moss, C.; George, G.; Santaolalla, A.; Cope, A.; Papa, S.; Van Hemelrijck, M. Associations between immune-suppressive and stimulating drugs and novel COVID-19—a systematic review of current evidence. ecancermedicalscience 2020, 14, 1–43. [Google Scholar] [CrossRef] [PubMed]
- Vitiello, A.; Ferrara, F. Perspectives of association Baricitinib/Remdesivir for adults with Covid-19 infection. Mol. Biol. Rep. 2021, 49, 827–831. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekar, V.; Singh, A.V.; Maharjan, R.S.; Dakua, S.P.; Balakrishnan, S.; Dash, S.; Laux, P.; Luch, A.; Singh, S.; Pradhan, M. Perspectives on the Technological Aspects and Biomedical Applications of Virus-Like Particles/Nanoparticles in Reproductive Biology: Insights on the Medicinal and Toxicological Outlook. Adv. NanoBiomed Res. 2022, 2. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, Y.; Qiao, W.; Zhang, J.; Qi, Z. Baricitinib, a drug with potential effect to prevent SARS-COV-2 from entering target cells and control cytokine storm induced by COVID-19. Int. Immunopharmacol. 2020, 86, 106749–106749. [Google Scholar] [CrossRef]
- Wagstaff, K.M.; Sivakumaran, H.; Heaton, S.M.; Harrich, D.; Jans, D.A. Ivermectin is a specific inhibitor of importin α/β-mediated nuclear import able to inhibit replication of HIV-1 and dengue virus. Biochem. J. 2012, 443, 851–856. [Google Scholar] [CrossRef]
- Caly, L.; Wagstaff, K.M.; Jans, D.A. Nuclear trafficking of proteins from RNA viruses: Potential target for antivirals? Antivir. Res. 2012, 95, 202–206. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.N.Y.; Atkinson, S.C.; Wang, C.; Lee, A.; Bogoyevitch, M.A.; Borg, N.A.; Jans, D.A. The broad spectrum antiviral ivermectin targets the host nuclear transport importin α/β1 heterodimer. Antivir. Res. 2020, 177, 104760. [Google Scholar] [CrossRef]
- Tu, Y.-F.; Chien, C.-S.; Yarmishyn, A.A.; Lin, Y.-Y.; Luo, Y.-H.; Lin, Y.-T.; Lai, W.-Y.; Yang, D.-M.; Chou, S.-J.; Yang, Y.-P.; Wang, M.-L.; Chiou, S.-H. A Review of SARS-CoV-2 and the Ongoing Clinical Trials. Int. J. Mol. Sci. 2020, 21, 2657. [Google Scholar] [CrossRef]
- Ali, M.J.; Haider, M.A.; Ahmed, M.U.; Sundas, F.; Hirani, A.; Khan, I.A.; Anis, K.; Karim, A.H. Treatment Options for COVID-19: A Review. Front. Med. 2020, 7, 480. [Google Scholar] [CrossRef]
- Patrì, A.; Fabbrocini, G. Hydroxychloroquine and ivermectin: A synergistic combination for COVID-19 chemoprophylaxis and treatment? J. Am. Acad. Dermatol. 2020, 82, e221–E221. [Google Scholar] [CrossRef] [PubMed]
- Momekov, G.; Momekova, D. Ivermectin as a potential COVID-19 treatment from the pharmacokinetic point of view: antiviral levels are not likely attainable with known dosing regimens. Biotechnol. Biotechnol. Equip. 2020, 34, 469–474. [Google Scholar] [CrossRef]
- Park, A.; Iwasaki, A. Type I and Type III Interferons – Induction, Signaling, Evasion, and Application to Combat COVID-19. Cell Host Microbe 2020, 27, 870–878. [Google Scholar] [CrossRef] [PubMed]
- Scheuplein, V.A.; Seifried, J.; Malczyk, A.H.; Miller, L.; Höcker, L.; Vergara-Alert, J.; Dolnik, O.; Zielecki, F.; Becker, B.; Spreitzer, I.; et al. High Secretion of Interferons by Human Plasmacytoid Dendritic Cells upon Recognition of Middle East Respiratory Syndrome Coronavirus. J. Virol. 2015, 89, 3859–3869. [Google Scholar] [CrossRef]
- Lazear, H.M.; Schoggins, J.W.; Diamond, M.S. Shared and Distinct Functions of Type I and Type III Interferons. Immunity 2019, 50, 907–923. [Google Scholar] [CrossRef]
- Yoshikawa, T.; Hill, T.E.; Yoshikawa, N.; Popov, V.L.; Galindo, C.L.; Garner, H.R.; Peters, C.J.; Tseng, C.-T. (. Dynamic Innate Immune Responses of Human Bronchial Epithelial Cells to Severe Acute Respiratory Syndrome-Associated Coronavirus Infection. PLOS ONE 2010, 5, e8729. [Google Scholar] [CrossRef]
- Uzé, G.; Tavernier, J. High efficiency targeting of IFN-α activity: Possible applications in fighting tumours and infections. Cytokine Growth Factor Rev. 2015, 26, 179–182. [Google Scholar] [CrossRef]
- Raad, I.; Darouiche, R.; Vazquez, J.; Lentnek, A.; Hachem, R.; Hanna, H.; Goldstein, B.; Henkel, T.; Seltzer, E. Efficacy and Safety of Weekly Dalbavancin Therapy for Catheter-Related Bloodstream Infection Caused by Gram-Positive Pathogens. Clin. Infect. Dis. 2005, 40, 374–380. [Google Scholar] [CrossRef]
- Pouya, M.A.; Afshani, S.M.; Maghsoudi, A.S.; Hassani, S.; Mirnia, K. Classification of the present pharmaceutical agents based on the possible effective mechanism on the COVID-19 infection. DARU J. Pharm. Sci. 2020, 28, 745–764. [Google Scholar] [CrossRef]
- Andersen, P.I.; Krpina, K.; Ianevski, A.; Shtaida, N.; Jo, E.; Yang, J.; Koit, S.; Tenson, T.; Hukkanen, V.; Anthonsen, M.W.; et al. Novel Antiviral Activities of Obatoclax, Emetine, Niclosamide, Brequinar, and Homoharringtonine. Viruses 2019, 11, 964. [Google Scholar] [CrossRef]
- Plaze, M.; Attali, D.; Prot, M.; Petit, A.-C.; Blatzer, M.; Vinckier, F.; Levillayer, L.; Chiaravalli, J.; Perin-Dureau, F.; Cachia, A.; et al. Inhibition of the replication of SARS-CoV-2 in human cells by the FDA-approved drug chlorpromazine. Int. J. Antimicrob. Agents 2020, 57, 106274. [Google Scholar] [CrossRef] [PubMed]
- Drożdżal, S.; Rosik, J.; Lechowicz, K.; Machaj, F.; Kotfis, K.; Ghavami, S.; Łos, M.J. FDA approved drugs with pharmacotherapeutic potential for SARS-CoV-2 (COVID-19) therapy. Drug Resist. Updat. 2020, 53, 100719–100719. [Google Scholar] [CrossRef] [PubMed]
- Dancey, J. mTOR signaling and drug development in cancer. Nat. Rev. Clin. Oncol. 2010, 7, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Patocka, J.; Kuca, K.; Oleksak, P.; Nepovimova, E.; Valis, M.; Novotny, M.; Klimova, B. Rapamycin: Drug Repurposing in SARS-CoV-2 Infection. Pharmaceuticals 2021, 14, 217. [Google Scholar] [CrossRef]
- Zhou, Y.; Hou, Y.; Shen, J.; Huang, Y.; Martin, W.; Cheng, F. Network-based drug repurposing for novel coronavirus 2019-nCoV/SARS-CoV-2. Cell Discov. 2020, 6, 1–18. [Google Scholar] [CrossRef]
- Husain, A.; Byrareddy, S.N. Rapamycin as a potential repurpose drug candidate for the treatment of COVID-19. Chem. Interactions 2020, 331, 109282–109282. [Google Scholar] [CrossRef]
- Singla, A.; Harun, N.; Dilling, D.F.; Merchant, K.; McMahan, S.; Ingledue, R.; French, A.; Corral, J.A.; Korbee, L.; Kopras, E.J.; et al. Safety and efficacy of sirolimus in hospitalised patients with COVID-19 pneumonia. Respir. Investig. 2024, 62, 216–222. [Google Scholar] [CrossRef]
- Yuan, C.; Guan, Y.M. Efficacy and safety of Lianhua Qingwen as an adjuvant treatment for influenza in Chinese patients: A meta-analysis. Medicine 2024, 103, e36986. [Google Scholar] [CrossRef]
- Runfeng, L.; Yunlong, H.; Jicheng, H.; Weiqi, P.; Qinhai, M.; Yongxia, S.; Chufang, L.; Jin, Z.; Zhenhua, J.; Haiming, J.; et al. Lianhuaqingwen exerts anti-viral and anti-inflammatory activity against novel coronavirus (SARS-CoV-2). Pharmacol. Res. 2020, 156, 104761. [Google Scholar] [CrossRef]
- Hu, K.; Guan, W.-J.; Bi, Y.; Zhang, W.; Li, L.; Zhang, B.; Liu, Q.; Song, Y.; Li, X.; Duan, Z.; et al. Efficacy and safety of Lianhuaqingwen capsules, a repurposed Chinese herb, in patients with coronavirus disease 2019: A multicenter, prospective, randomized controlled trial. Phytomedicine 2020, 85, 153242–153242. [Google Scholar] [CrossRef]
- Kumar, S.; Zhi, K.; Mukherji, A.; Gerth, K. Repurposing Antiviral Protease Inhibitors Using Extracellular Vesicles for Potential Therapy of COVID-19. Viruses 2020, 12, 486. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.; Wang, Z.; Zhao, F.; Yang, Y.; Li, J.; Yuan, J.; Wang, F.; Li, D.; Yang, M.; Xing, L.; et al. Treatment of 5 Critically Ill Patients With COVID-19 With Convalescent Plasma. JAMA 2020, 323, 1582–1589. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Xiong, J.; Bao, L.; Shi, Y. Convalescent plasma as a potential therapy for COVID-19. Lancet Infect. Dis. 2020, 20, 398–400. [Google Scholar] [CrossRef] [PubMed]
- Awad K, Kakkola L, Julkunen I. High Glucose Increases Lactate and Induces the Transforming Growth Factor Beta-Smad 1/5 Atherogenic Pathway in Primary Human Macrophages. Biomedicines. 2024;12(7):1575. [CrossRef]
- Yuan, Y.; Jiao, B.; Qu, L.; Yang, D.; Liu, R. The development of COVID-19 treatment. Front. Immunol. 2023, 14, 1125246. [Google Scholar] [CrossRef]
- Jones BE, Brown-Augsburger PL, Corbett KS, et al. The neutralizing antibody, LY-CoV555, protects against SARS-CoV-2 infection in nonhuman primates. Sci Transl Med. 2021;13(593):1906. [CrossRef]
- Morales-Núñez, J.J.; Muñoz-Valle, J.F.; Torres-Hernández, P.C.; Hernández-Bello, J. Overview of Neutralizing Antibodies and Their Potential in COVID-19. Vaccines 2021, 9, 1376. [Google Scholar] [CrossRef]
- Dougan, M.; Nirula, A.; Azizad, M.; Mocherla, B.; Gottlieb, R.L.; Chen, P.; Hebert, C.; Perry, R.; Boscia, J.; Heller, B.; et al. Bamlanivimab plus Etesevimab in Mild or Moderate Covid-19. New Engl. J. Med. 2021, 385, 1382–1392. [Google Scholar] [CrossRef]
- Li, C.; Phoon, Y.P.; Karlinsey, K.; Tian, Y.F.; Thapaliya, S.; Thongkum, A.; Qu, L.; Matz, A.J.; Cameron, M.; Cameron, C.; et al. A high OXPHOS CD8 T cell subset is predictive of immunotherapy resistance in melanoma patients. J. Exp. Med. 2021, 219. [Google Scholar] [CrossRef]
- VanBlargan, L.A.; Errico, J.M.; Halfmann, P.J.; Zost, S.J.; Crowe, J.E.; Purcell, L.A.; Kawaoka, Y.; Corti, D.; Fremont, D.H.; Diamond, M.S. An infectious SARS-CoV-2 B.1.1.529 Omicron virus escapes neutralization by therapeutic monoclonal antibodies. Nat. Med. 2022, 28, 490–495. [Google Scholar] [CrossRef]
- Liew MNY, Kua KP, Lee SWH, Wong KK. SARS-CoV-2 neutralizing antibody bebtelovimab – a systematic scoping review and meta-analysis. Front Immunol. 2023;14:1100263. [CrossRef]
- Li, C.; Qu, L.; Matz, A.J.; Murphy, P.A.; Liu, Y.; Manichaikul, A.W.; Aguiar, D.; Rich, S.S.; Herrington, D.M.; Vu, D.; et al. AtheroSpectrum Reveals Novel Macrophage Foam Cell Gene Signatures Associated With Atherosclerotic Cardiovascular Disease Risk. Circulation 2021, 145, 206–218. [Google Scholar] [CrossRef]
- Iketani, S.; Iketani, S.; Liu, L.; Liu, L.; Guo, Y.; Guo, Y.; Liu, L.; Liu, L.; Chan, J.F.-W.; Chan, J.F.-W.; et al. Antibody evasion properties of SARS-CoV-2 Omicron sublineages. Nature 2022, 604, 553–556. [Google Scholar] [CrossRef]
- Toots, M.; Yoon, J.-J.; Cox, R.M.; Hart, M.; Sticher, Z.M.; Makhsous, N.; Plesker, R.; Barrena, A.H.; Reddy, P.G.; Mitchell, D.G.; et al. Characterization of orally efficacious influenza drug with high resistance barrier in ferrets and human airway epithelia. Sci. Transl. Med. 2019, 11. [Google Scholar] [CrossRef] [PubMed]
- Rosenke, K.; Hansen, F.; Schwarz, B.; Feldmann, F.; Haddock, E.; Rosenke, R.; Barbian, K.; Meade-White, K.; Okumura, A.; Leventhal, S.; et al. Orally delivered MK-4482 inhibits SARS-CoV-2 replication in the Syrian hamster model. Nat. Commun. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Sheahan, T.P.; Sims, A.C.; Zhou, S.; Graham, R.L.; Pruijssers, A.J.; Agostini, M.L.; Leist, S.R.; Schäfer, A.; Dinnon, K.H., 3rd; Stevens, L.J.; et al. An orally bioavailable broad-spectrum antiviral inhibits SARS-CoV-2 in human airway epithelial cell cultures and multiple coronaviruses in mice. Sci. Transl. Med. 2020, 12, eabb5883. [Google Scholar] [CrossRef] [PubMed]
- Gordon, C.J.; Tchesnokov, E.P.; Schinazi, R.F.; Götte, M. Molnupiravir promotes SARS-CoV-2 mutagenesis via the RNA template. J. Biol. Chem. 2021, 297, 100770. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Chen, Y.; Lu, X.; Zhang, W.; Fang, W.; Yuan, L.; Wang, X. An update on inhibitors targeting RNA-dependent RNA polymerase for COVID-19 treatment: Promises and challenges. Biochem. Pharmacol. 2022, 205, 115279–115279. [Google Scholar] [CrossRef]
- Xu, T.; Zhang, L. Current understanding of nucleoside analogs inhibiting the SARS-CoV-2 RNA-dependent RNA polymerase. Comput. Struct. Biotechnol. J. 2023, 21, 4385–4394. [Google Scholar] [CrossRef]
- Kabinger, F.; Stiller, C.; Schmitzová, J.; Dienemann, C.; Kokic, G.; Hillen, H.S.; Höbartner, C.; Cramer, P. Mechanism of molnupiravir-induced SARS-CoV-2 mutagenesis. Nat. Struct. Mol. Biol. 2021, 28, 740–746. [Google Scholar] [CrossRef]
- Wahl, A.; Gralinski, L.E.; Johnson, C.E.; Yao, W.; Kovarova, M.; Dinnon, K.H.; Liu, H.; Madden, V.J.; Krzystek, H.M.; De, C.; et al. SARS-CoV-2 infection is effectively treated and prevented by EIDD-2801. Nature 2021, 591, 451–457. [Google Scholar] [CrossRef]
- Painter, W.P.; Holman, W.; Bush, J.A.; Almazedi, F.; Malik, H.; Eraut, N.C.J.E.; Morin, M.J.; Szewczyk, L.J.; Painter, G.R. Human Safety, Tolerability, and Pharmacokinetics of Molnupiravir, a Novel Broad-Spectrum Oral Antiviral Agent with Activity against SARS-CoV-2. Antimicrob. Agents Chemother. 2021, 65. [Google Scholar] [CrossRef]
- Syed, Y.Y. Molnupiravir: First Approval. Drugs 2022, 82, 455–460. [Google Scholar] [CrossRef]
- Singh, A.K.; Singh, A.; Singh, R.; Misra, A. Molnupiravir in COVID-19: A systematic review of literature. Diabetes Metab. Syndr. Clin. Res. Rev. 2021, 15, 102329–102329. [Google Scholar] [CrossRef]
- Toussi, S.S.; Hammond, J.L.; Gerstenberger, B.S.; Anderson, A.S. Therapeutics for COVID-19. Nat. Microbiol. 2023, 8, 771–786. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Huang, X.; Ma, Q.; Kuzmič, P.; Zhou, B.; Zhang, S.; Chen, J.; Xu, J.; Liu, B.; Jiang, H.; et al. Preclinical evaluation of the SARS-CoV-2 Mpro inhibitor RAY1216 shows improved pharmacokinetics compared with nirmatrelvir. Nat. Microbiol. 2024, 9, 1075–1088. [Google Scholar] [CrossRef]
- Fujimuro, M. The Interactions between Cells and Viruses. Int. J. Mol. Sci. 2024, 25, 6886. [Google Scholar] [CrossRef] [PubMed]
- Amiama-Roig, A.; Pérez-Martínez, L.; Ledo, P.R.; Verdugo-Sivianes, E.M.; Blanco, J.-R. Should We Expect an Increase in the Number of Cancer Cases in People with Long COVID? Microorganisms 2023, 11, 713. [Google Scholar] [CrossRef] [PubMed]
- Oprinca, G.C.; Mohor, C.-I.; Bereanu, A.-S.; Oprinca-Muja, L.-A.; Bogdan-Duică, I.; Fleacă, S.R.; Hașegan, A.; Diter, A.; Boeraș, I.; Cristian, A.N.; et al. Detection of SARS-CoV-2 Viral Genome and Viral Nucleocapsid in Various Organs and Systems. Int. J. Mol. Sci. 2024, 25, 5755. [Google Scholar] [CrossRef]
- Cheung, C.C.L.; Goh, D.; Lim, X.; Tien, T.Z.; Lim, J.C.T.; Lee, J.N.; Tan, B.; Tay, Z.E.A.; Wan, W.Y.; Chen, E.X.; et al. Residual SARS-CoV-2 viral antigens detected in GI and hepatic tissues from five recovered patients with COVID-19. Gut 2021, 71, 226–229. [Google Scholar] [CrossRef]
- Shimizu, K.; Hirata, H.; Tokuhira, N.; Motooka, D.; Nakamura, S.; Ueda, A.; Tachino, J.; Koide, M.; Uchiyama, A.; Ogura, H.; et al. Dysbiosis of gut microbiota in patients with severe COVID-19. Acute Med. Surg. 2024, 11, e923. [Google Scholar] [CrossRef]
- Kanduc, D. From Anti-Severe Acute Respiratory Syndrome Coronavirus 2 Immune Response to Cancer Onset via Molecular Mimicry and Cross-Reactivity. Glob. Med Genet. 2021, 08, 176–182. [Google Scholar] [CrossRef]
- Lemos, A.E.G.; Silva, G.R.; Gimba, E.R.P.; Matos, A.d.R. Susceptibility of lung cancer patients to COVID-19: A review of the pandemic data from multiple nationalities. Thorac. Cancer 2021, 12, 2637–2647. [Google Scholar] [CrossRef]
- Venkatesulu, B.P.; Chandrasekar, V.T.; Girdhar, P.; Advani, P.; Sharma, A.; Elumalai, T.; Hsieh, C.E.; I Elghazawy, H.; Verma, V.; Krishnan, S. A Systematic Review and Meta-Analysis of Cancer Patients Affected by a Novel Coronavirus. JNCI Cancer Spectr. 2021, 5, pkaa102. [Google Scholar] [CrossRef] [PubMed]
- Peravali, M.; Joshi, I.; Ahn, J.; Kim, C. A Systematic Review and Meta-Analysis of Clinical Characteristics and Outcomes in Patients With Lung Cancer with Coronavirus Disease 2019. JTO Clin. Res. Rep. 2021, 2, 100141–100141. [Google Scholar] [CrossRef] [PubMed]
- Park, S.-Y.; Nam, J.-S. The force awakens: metastatic dormant cancer cells. Exp. Mol. Med. 2020, 52, 569–581. [Google Scholar] [CrossRef]
- Barkan, D.; El Touny, L.H.; Michalowski, A.M.; Smith, J.A.; Chu, I.; Davis, A.S.; Webster, J.D.; Hoover, S.; Simpson, R.M.; Gauldie, J.; et al. Metastatic Growth from Dormant Cells Induced by a Col-I–Enriched Fibrotic Environment. Cancer Res. 2010, 70, 5706–5716. [Google Scholar] [CrossRef] [PubMed]
- Rolfo, C.; Meshulami, N.; Russo, A.; Krammer, F.; García-Sastre, A.; Mack, P.C.; Gomez, J.E.; Bhardwaj, N.; Benyounes, A.; Sirera, R.; et al. Lung Cancer and Severe Acute Respiratory Syndrome Coronavirus 2 Infection: Identifying Important Knowledge Gaps for Investigation. J. Thorac. Oncol. 2021, 17, 214–227. [Google Scholar] [CrossRef]
- Luo, J.; Rizvi, H.; Preeshagul, I.; Egger, J.; Hoyos, D.; Bandlamudi, C.; McCarthy, C.; Falcon, C.; Schoenfeld, A.; Arbour, K.; et al. COVID-19 in patients with lung cancer. Ann. Oncol. 2020, 31, 1386–1396. [Google Scholar] [CrossRef]
- Bora, V.R.; Patel, B.M. The Deadly Duo of COVID-19 and Cancer! Front. Mol. Biosci. 2021, 8, 643004. [Google Scholar] [CrossRef]
Figure 1.
Human Structure, life cycle, mechanism of action of COVID-19. Created with Biorender.
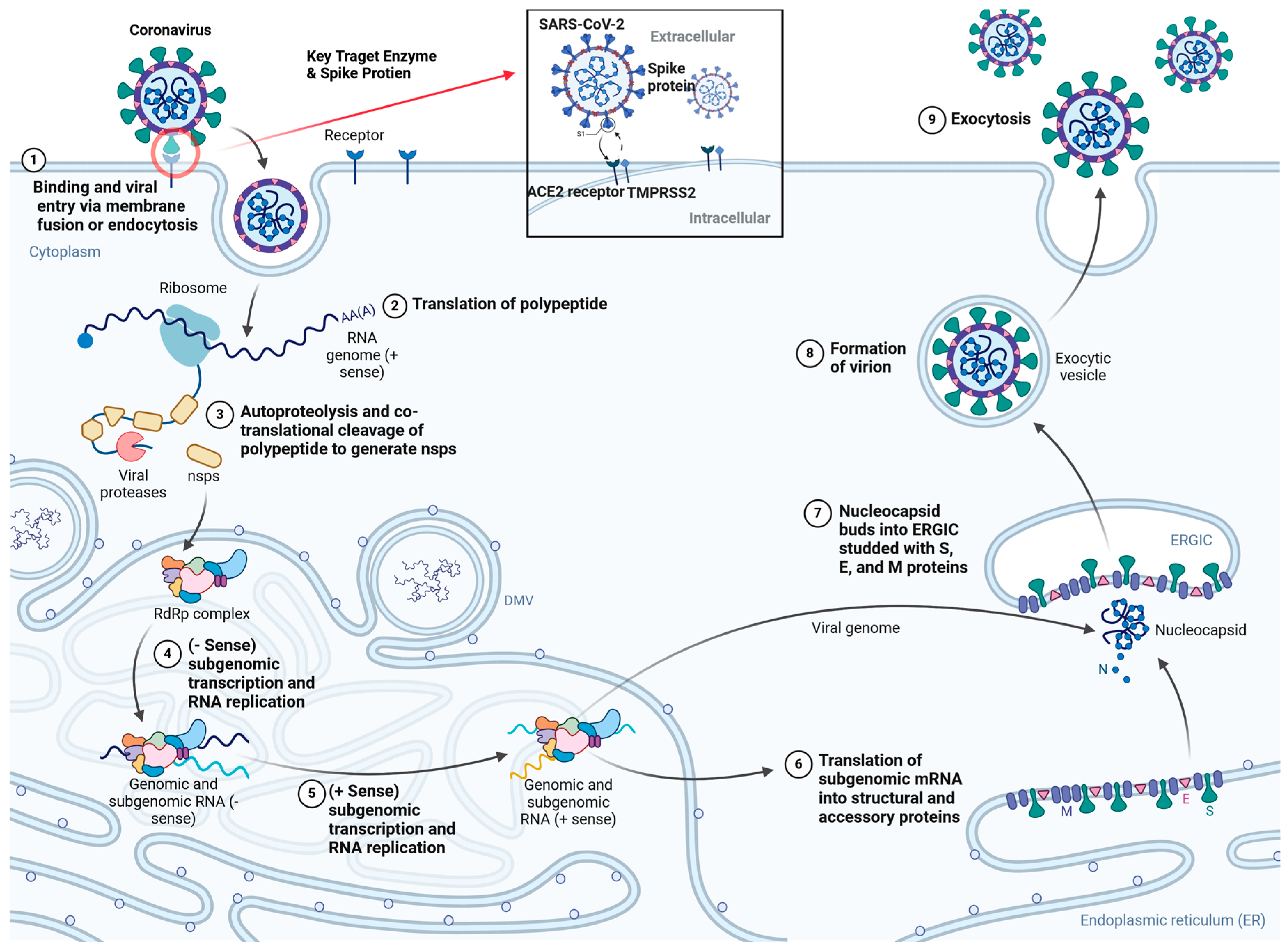
Figure 2.
Schematic Diagram for Vaccines Candidates in human trial. Created with Biorender.
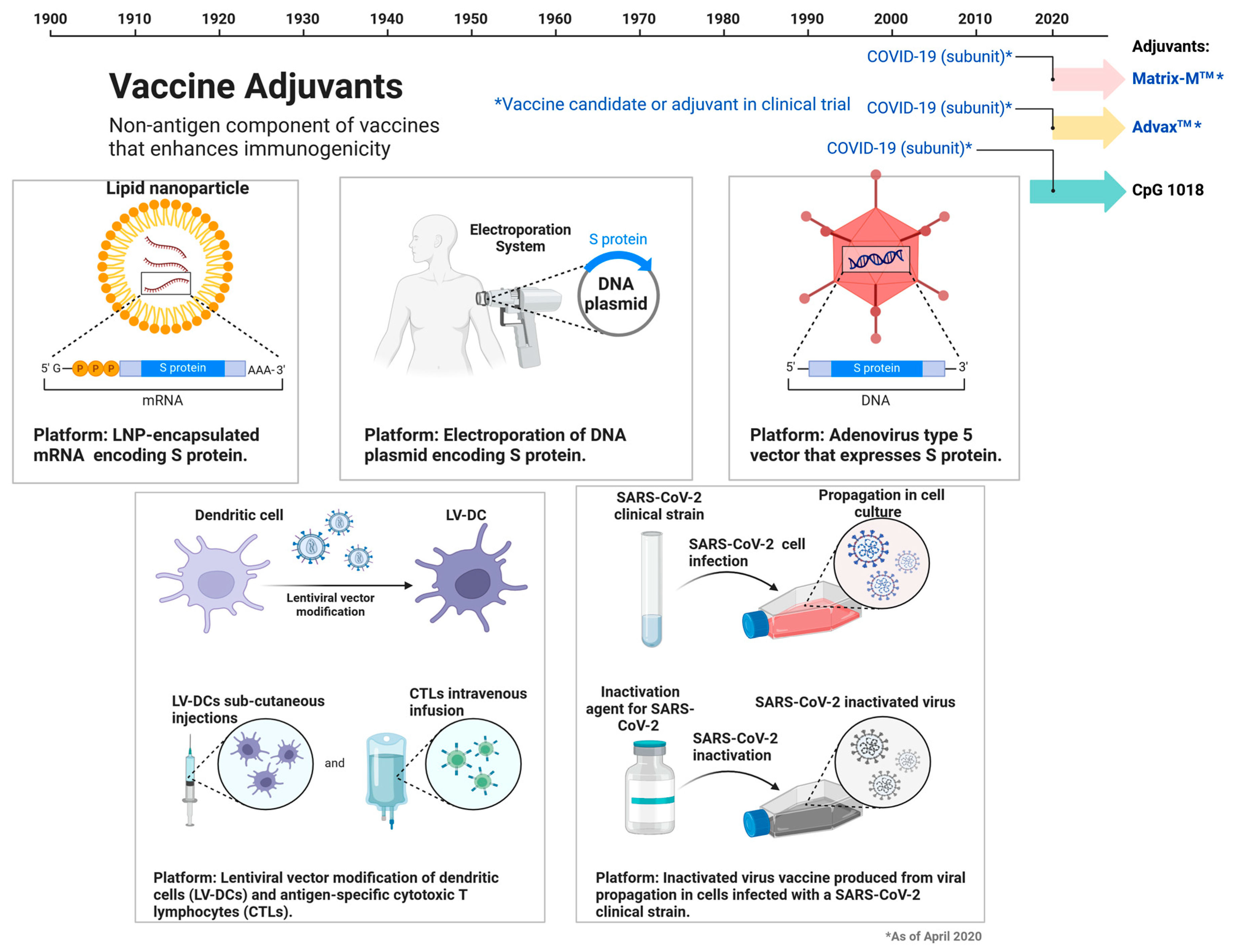
Figure 3.
The SARS-CoV-2 Variants of Concern. Created with Biorender.
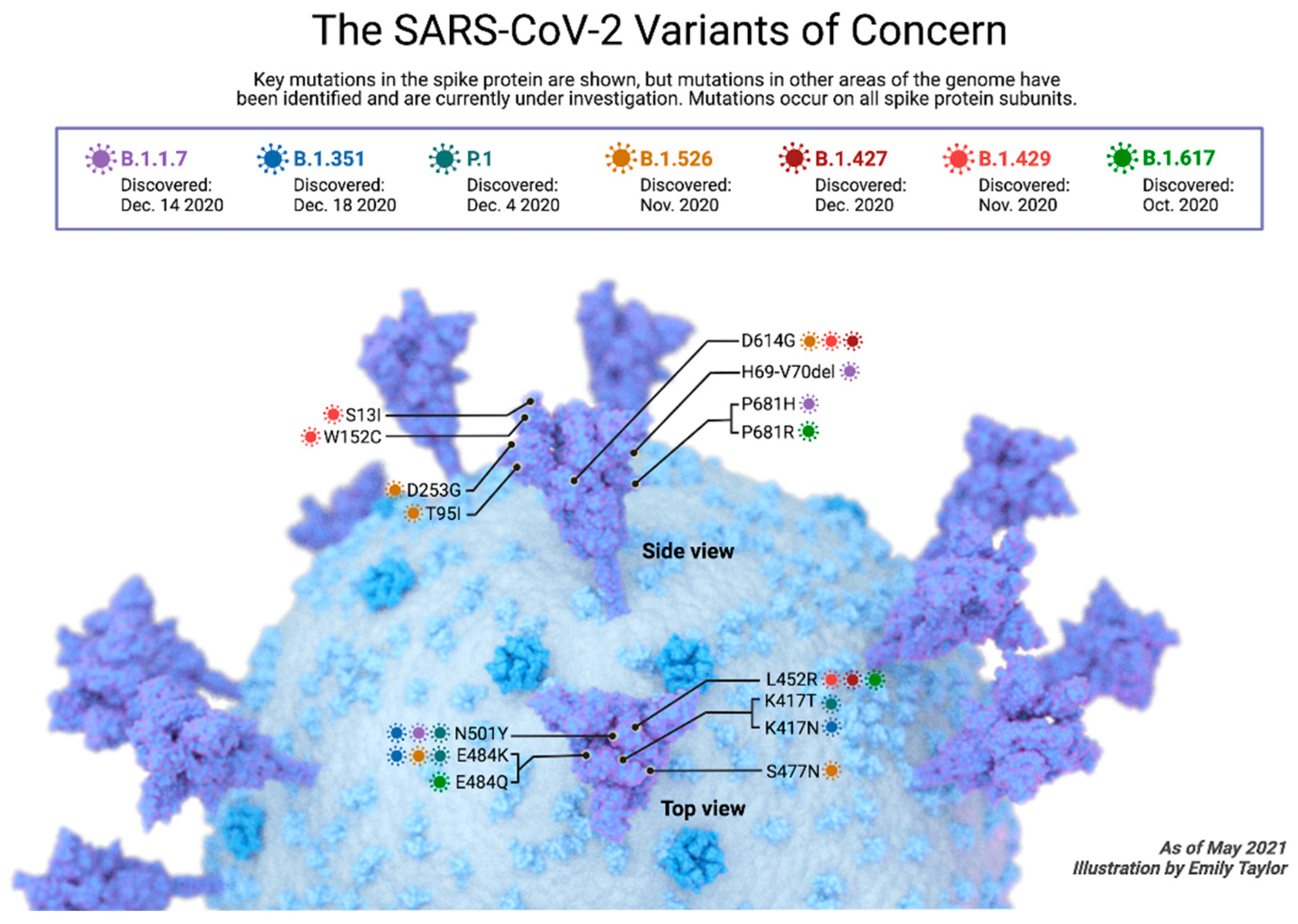
Figure 4.
Chemical structures of Fusion inhibitors that target S spike protein; Umifenovir, Camostat mesylate, and Nafamostat mesylate.
Figure 4.
Chemical structures of Fusion inhibitors that target S spike protein; Umifenovir, Camostat mesylate, and Nafamostat mesylate.
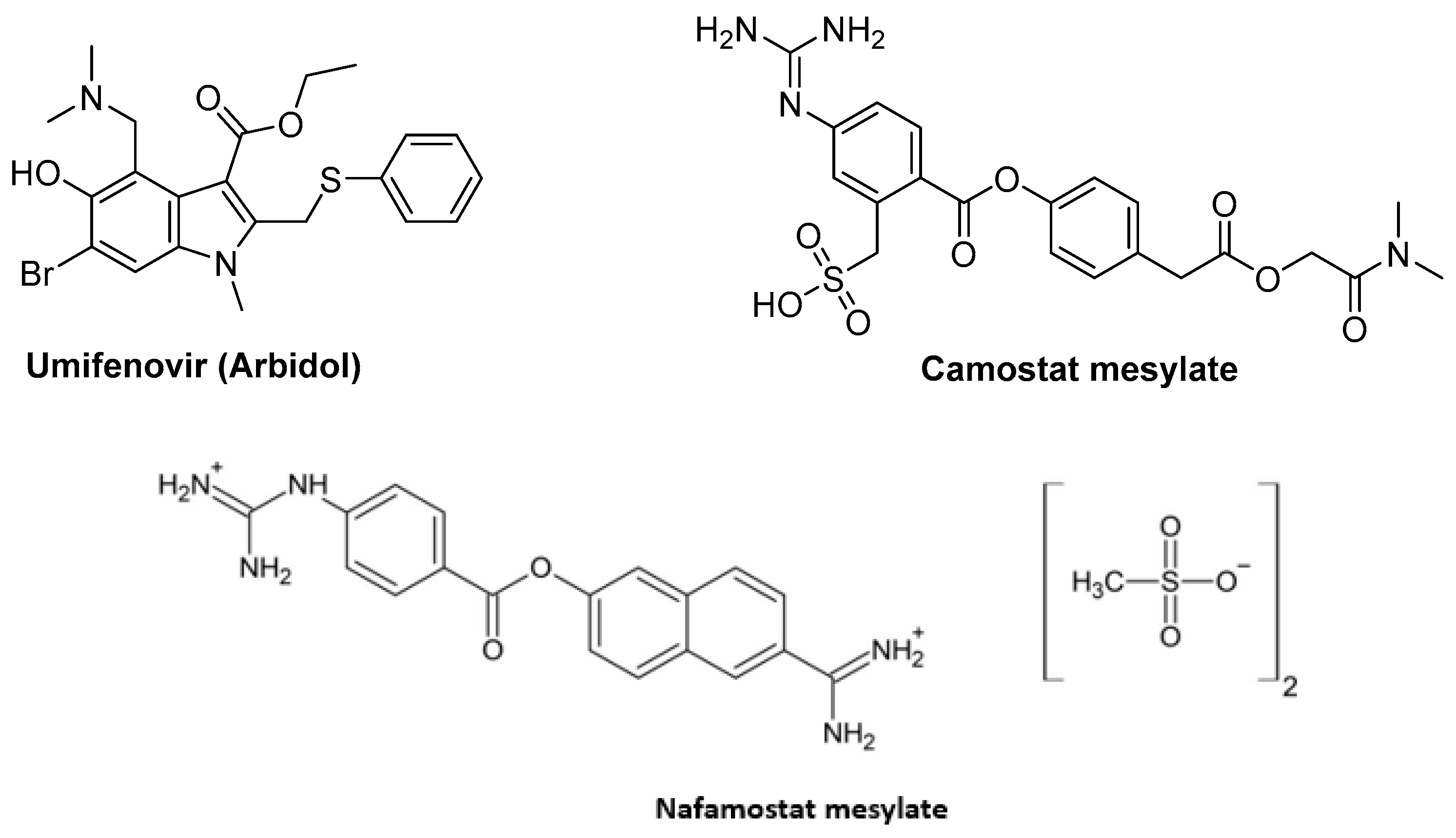
Figure 5.
Chemical structures of Protease inhibitors Lopinavir, Ritonavir, Danoprevir, Saquinavir, Ebselen.
Figure 5.
Chemical structures of Protease inhibitors Lopinavir, Ritonavir, Danoprevir, Saquinavir, Ebselen.
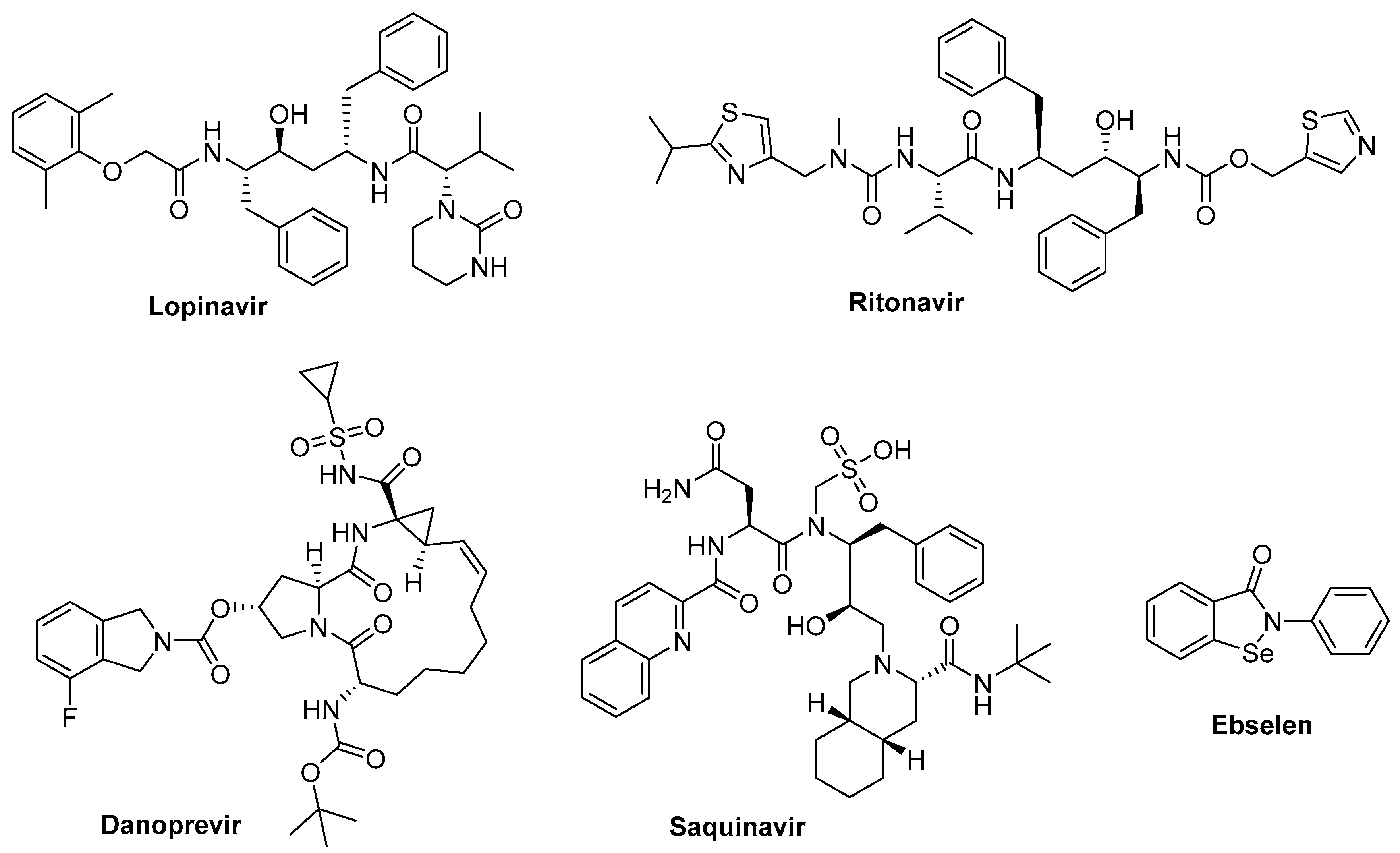
Figure 6.
Chemical structures of RNA-dependent RNA polymerase; Remedsivir, Favipiravir, Ribavirin, Glidesivir.
Figure 6.
Chemical structures of RNA-dependent RNA polymerase; Remedsivir, Favipiravir, Ribavirin, Glidesivir.
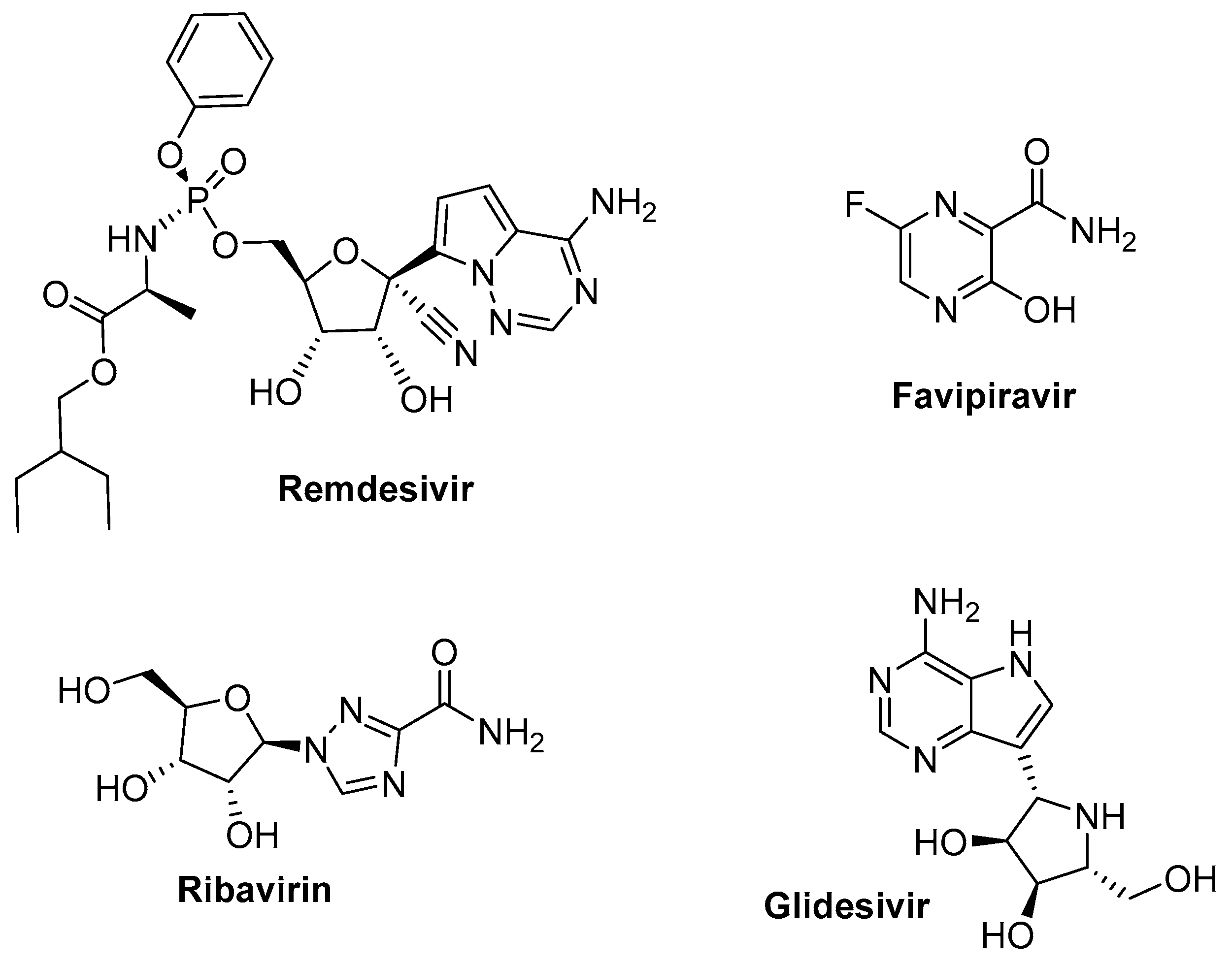
Figure 7.
Chemical structures of Neuraminidase inhibitors, such as Oseltamivir, Zanamivir, and Peramivir.
Figure 7.
Chemical structures of Neuraminidase inhibitors, such as Oseltamivir, Zanamivir, and Peramivir.
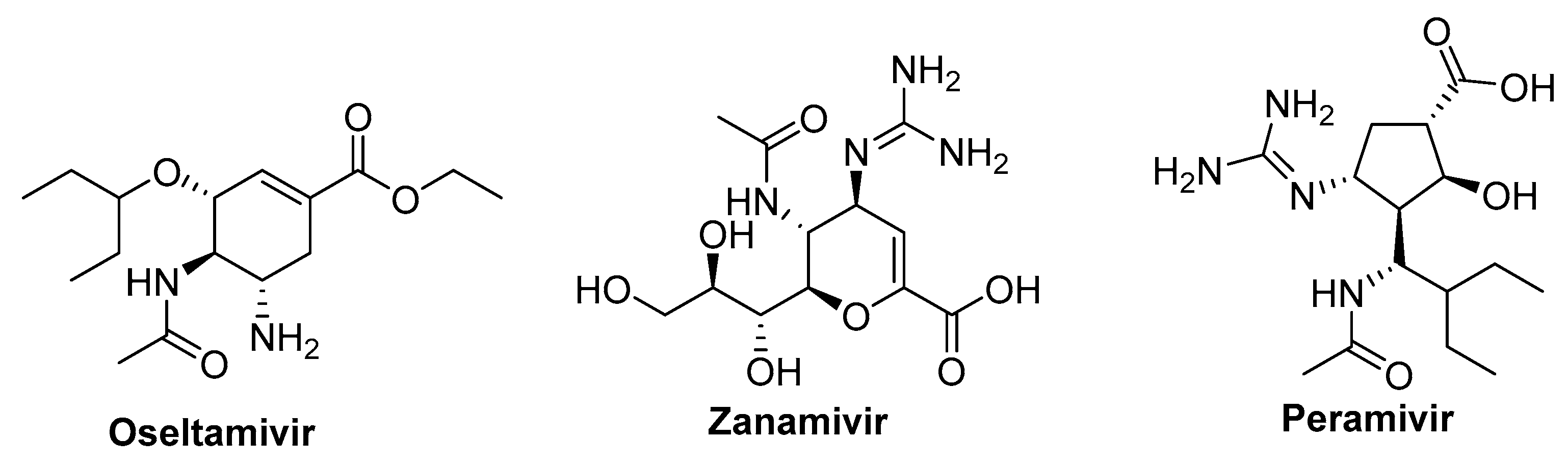
Figure 8.
Chemical structures of M2 channel protein target such as Admantane, Amantadine, and Rimantadine.
Figure 8.
Chemical structures of M2 channel protein target such as Admantane, Amantadine, and Rimantadine.
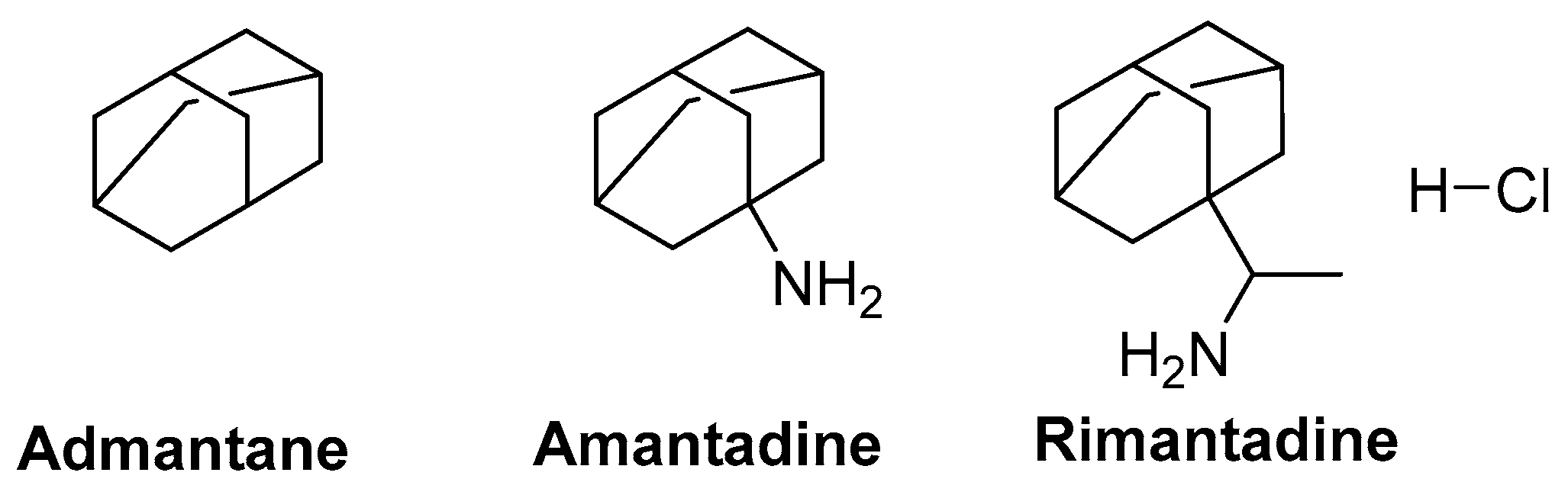
Figure 9.
Chemical structures of some non-antiviral drugs, Chlorpromazine, Emetin, Baricitnib targeting Importin α/β1-mediated nuclear import inhibitors.
Figure 9.
Chemical structures of some non-antiviral drugs, Chlorpromazine, Emetin, Baricitnib targeting Importin α/β1-mediated nuclear import inhibitors.
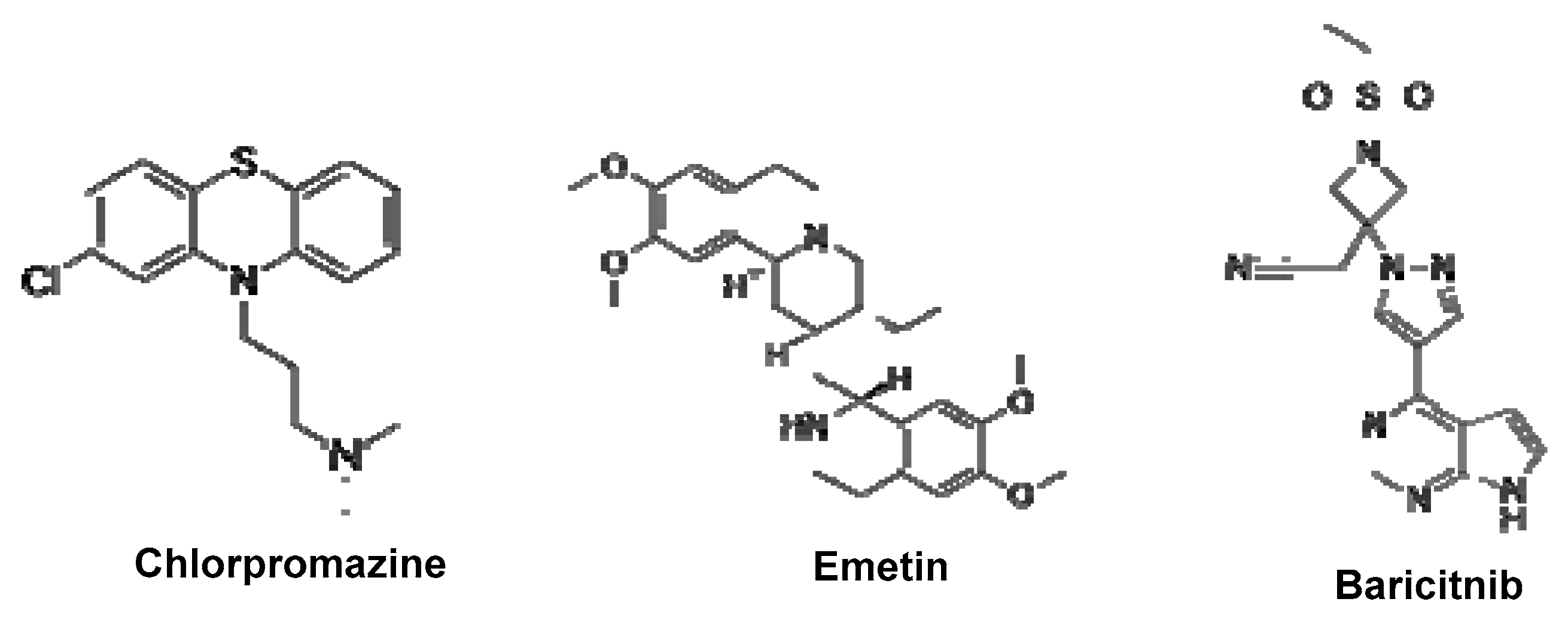
Figure 10.
The mechanism of SARS-CoV-2 neutralizing antibodies. [246].
Figure 10.
The mechanism of SARS-CoV-2 neutralizing antibodies. [246].
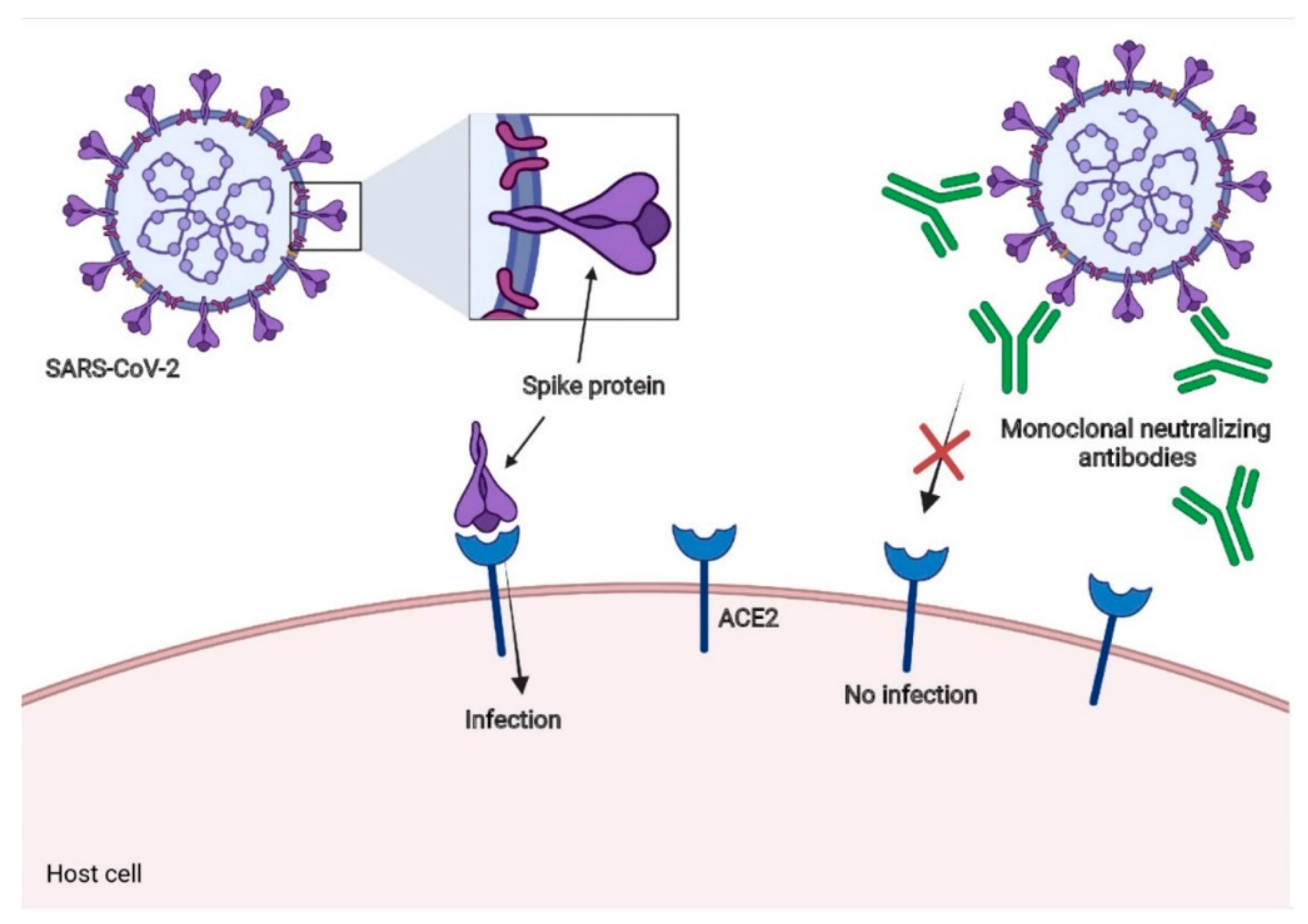
Figure 11.
Chemical structures of Molnupiravir and Paxlovid.
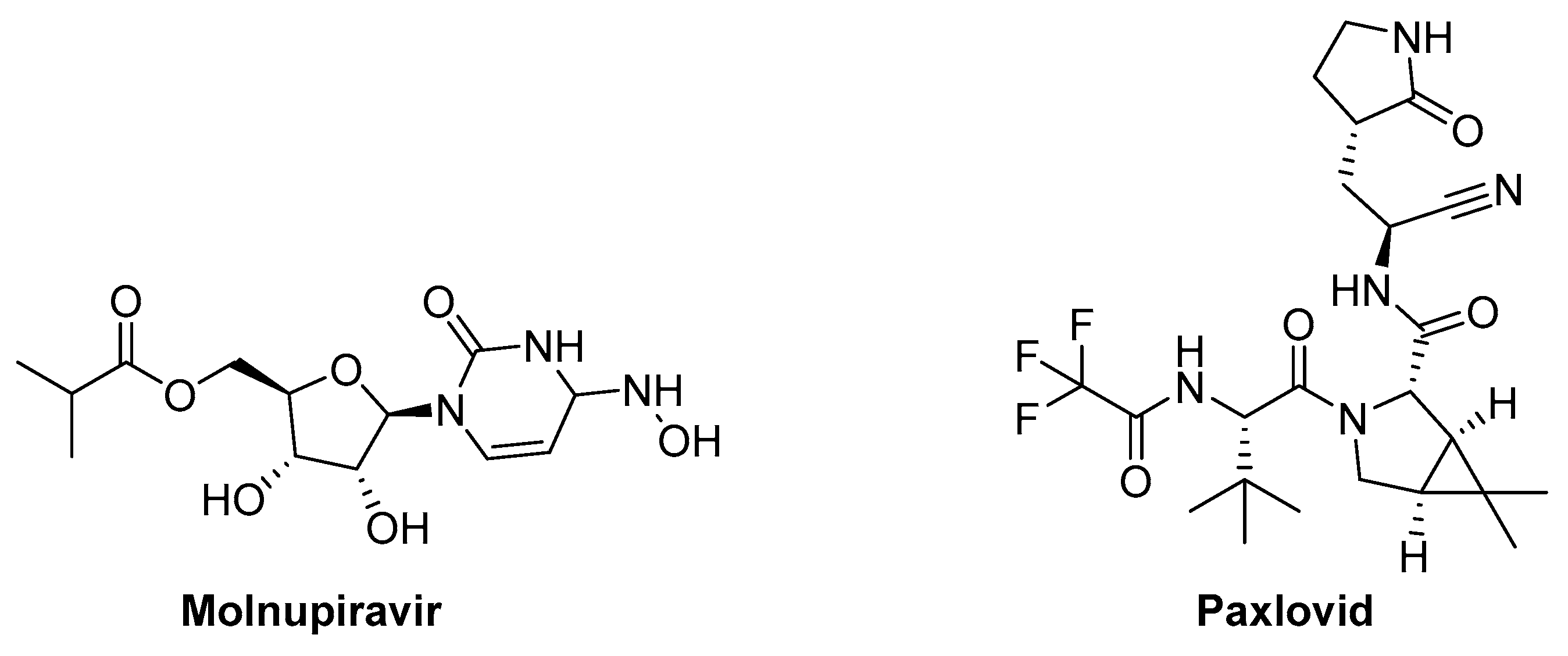
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



