Submitted:
29 September 2024
Posted:
01 October 2024
You are already at the latest version
Abstract
Through the integration of genomic information, transcriptome sequencing data, and bioinformatics methods, we conducted a comprehensive identification of the ALDH gene family in melon. We explored the impact of this gene family on melon growth, development, and their expression patterns in various tissues and under different stress conditions. Our study discovered a total of 17 ALDH genes spread across chromosomes 1, 2, 3, 4, 5, 7, 8, 11, and 12 in the melon genome. Through a phylogenetic analysis, these genes were classified into 10 distinct subfamilies. Notably, genes within the same subfamily exhibited consistent gene structures and conserved motifs. Our study discovered a pair of fragment repeats within the melon ALDH gene. Furthermore, there was a noticeable collinearity relationship between the melon's ALDH gene and that of Arabidopsis (12 times), and rice (3 times). Transcriptome data reanalysis revealed that some ALDH genes consistently expressed highly across all tissues and developmental stages, while others were tissue or stage-specific. We analysed the ALDH gene's expression patterns under six stress types, namely salt, chilling, waterlogged, powdery mildew, fusarium wilt, and gummy stem blight. The results showed differential expression of MELO3C024328 and MELO3C004430 under all stress conditions, signifying their crucial roles in melon growth and stress response. RT-qPCR analysis further corroborated these findings. This study paves the way for future genetic improvements in melon molecular breeding.
Keywords:
ALDH gene family
; melon
; abiotic and biotic stress
; RNA-seq
1. Introduction
Throughout their lifecycle, plants are subjected to various forms of abiotic and biological stresses. These stresses promptly induce the synthesis of reactive oxygen species (ROS) within the plants, subsequently leading to an accumulation of aldehydes [1]. Aldehydes, which are metabolic byproducts of carbohydrates, lipids, and amino acids, can, in appropriate quantities, positively influence the physiological metabolism of plants. However, excessive concentrations of aldehydes can cause cellular damage, disrupt metabolic processes, and lead to the formation of toxic substances detrimental to plant health, compromising their normal growth [2]. Hence, the selective removal and regulation of aldehyde concentrations within plants hold significant implications for their growth and development, mandating strict regulation [3].
Aldehyde dehydrogenase (ALDH), often referred to as the "aldehyde scavenger," is effective in eliminating active aldehyde molecules in plants [4]. ALDH is a category of enzymes that encode NAD(P)+-dependent reactions, showcasing a diversity of amino acid sequences and incorporating different motifs such as the cysteine active site (PS00070), glutamic acid active site (PS00687), and the Rossmann fold [5]. To mitigate the damage inflicted by excessive aldehyde accumulation during plant growth, the ALDH gene is induced to express differently under stress, subsequently leading to a substantial increase in aldehyde dehydrogenase proteins [6]. Consequently, the toxic aldehydes generated due to stress are irreversibly oxidized into their non-toxic carboxylic acid counterparts, thus enhancing the plant's stress tolerance and reducing stress-induced damage. This indicates a critical detoxification mechanism in plants [7]. Furthermore, ALDH also plays a pivotal role in plant metabolism, such as glycolysis, amino acid metabolism, and carnitine biosynthesis [8]. Under osmotic stress, ALDH helps maintain cellular osmotic equilibrium in plants through the synthesis of osmoregulators [9].
Aldehyde dehydrogenase (ALDH) is predominantly found in both eukaryotes and prokaryotes. As the genome sequencing information for an ever-increasing number of species becomes available, the presence of ALDH genes in more species is being discovered. The ALDH Gene Naming Committee (ANGC) defines a classification system based on protein sequence similarity between a new gene and an identified gene. If this similarity is less than 40%, the gene is considered as part of a new family; if the similarity ranges between 40% and 60%, it is classified within a family; if the similarity exceeds 60%, it is allocated to a subfamily [10]. Currently, the identified ALDH genes are categorized into 24 families, namely ALDH1-ALDH24, with 14 families found in plants. These 14 include ALDH2, ALDH3, ALDH5, ALDH6, ALDH7, ALDH10, ALDH11, ALDH12, ALDH18, ALDH19, ALDH21, ALDH22, ALDH23, and ALDH24, with the last seven exclusive to plants [11,12]. Over recent years, we have seen an increasing number of ALDH genes being identified in various plants, each with varying quantities, such as 14, 20, 28, 29, 53, 19, 27, 27, 23, 22, and 39 in Arabidopsis [5], rice [13], maize [14], tomato [15], soybean [16], sorghum [17], Chinese cabbage [18], grape [19], potato [20], and apple [21] respectively. These genes play a significant role in the regulation of stress response, growth, and development in plants, hence critical for normal plant growth [22,23]. For instance, AtALDH3I1 and AtALDH7B4 in Arabidopsis are seen to reduce MDA (malondialdehyde) and lipid peroxidation when their expression is up-regulated, improving the plant's tolerance to drought and salt stress [24]. Overexpression of BrALDH7B2 in tobacco can enhance photosynthesis, reduce ROS production, thereby improving salt tolerance [25]. When wheat is subjected to drought and salt stress, TraeALDH7B1-5A is highly expressed in various parts of the plant, and overexpression of this gene in transgenic Arabidopsis significantly improves plant tolerance to drought stress [26]. Similarly, rice exhibited elevated expression levels of OsALDH2-4, OsALDH3-4, OsALDH7, OsALDH18-2, and OsALDH12 when subjected to drought stress, surpassing control levels by twofold [13]. Virus-Induced Gene Silencing (VIGS) experiment performed on the cotton genes Gohir.A11G040800 and Gohir.D06G046200 demonstrated increased sensitivity of the silenced plants to salt stress as compared to the control, indicating the potential involvement of these genes in cotton's salt stress response [27]. In soybeans, acetaldehyde dehydrogenase can reduce the content of toxic malondialdehyde (MDA), and overexpression of ALDH3H1 increases soybean resistance to salt stress [28]. ZmALDH9, ZmALDH13, and ZmALDH17 have been reported to participate in the maize response to drought stress and are involved in plant acid tolerance and pathogen infection response [14].
Cucumis melo L., a member of the genus Cucumis in the Cucurbitaceae family, is an annual horticultural crop of high economic worth, primarily due to its flavorful fruits and substantial nutritional value [29-31]. In 2012, the genome of Muskmelon DHL92 was sequenced by the Spanish Agricultural Genomics Research Center, paving the way for bioinformatics research related to muskmelon [32]. As bioinformatics technology continued to advance, Ruggieri carried out an upgrade of the DHL92 genome to version V3.6.1 in 2018, resulting in an enhanced sequence and annotation information [33]. Later in 2019, Castanera further improved the V3.6.1 version of DHL92 to version V4.0 [34]. Leveraging the available melon genomic information, researchers worldwide have conducted numerous gene family studies, focusing on gene families such as SUAR[35], ARF[36], LBD[37], SUN[38], and JMJ-C[39]. However, no research has yet been reported on the ALDH gene family in melon. In this study, we identified 17 ALDH genes in the melon genome using the available genomic data and conducted a thorough biogenic analysis, examining aspects such as chromosome localization, phylogeny, gene structure, and collinearity. To explore the expression patterns of the ALDH gene in different melon tissues under various stresses, we utilized published transcriptome data to analyze ALDH gene expression levels. The findings of this study establish a basis for future research on the ALDH gene family in melon and provide potential target candidate genes for resistance breeding in melon.
2. Results
2.1. Identification and Physicochemical Characteristics of the ALDH Family Member in Melon
This research identified 17 ALDH genes in the melon genome (DHL92V4) through the use of the hidden Markov model PF00171, and discovered conserved ALDH domains in Pfam, SMART, and NCBI conserved domain databases. An analysis of the physical and chemical properties of the ALDH family members in melon revealed that the smallest CDS gene was MELO3C004383 (1149 bp), while the largest was MELO3C007705 (3132 bp). The encoded amino acid length ranged from 382aa (MELO3C004383) to 1043aa (MELO3C007705); molecular weights fell between 41812.14 kD (MELO3C004383) and 114376.9 kD (MELO3C007705); and theoretical isoelectric point values varied from 5.32 (MELO3C017125) to 8.89 (MELO3C010493). Our instability coefficient analysis indicated that only one ALDH gene in melon, MELO3C007705, yielded a stable protein (instability coefficient >40), while the rest were classified as unstable proteins (instability coefficient <40). The fat coefficients for the 17 ALDH genes spanned a range from 77.52 (MELO3C007705) to 105.54 (MELO3C008245). An evaluation of their average hydrophilic values revealed that 12 ALDH genes had discernible hydrophilic values (mean hydrophilic value <0), with the remaining 5 classified as hydrophobic proteins. The mitochondria housed the most ALDH genes, with a total of six. This was followed by the chloroplast with five genes, the cytoplasm with four, and the nuclear and plasma membrane regions each containing one.

Table 1.
Information and physicochemical properties of 17 CmALDH members.
| Gene ID | CDS size (bp) | Number of Amino Acid(aa) |
Molecular Weight (kDa) |
Theoretical pI |
Instability Index | Aliphatic Index | Grand Average of Hydropathicity | Prediction of Subcellular Location |
|---|---|---|---|---|---|---|---|---|
| MELO3C018583 | 1614 | 537 | 57554.02 | 6.47 | 34.29 | 90.63 | 0.041 | Mitochondrial |
| MELO3C024345 | 1365 | 454 | 48695.21 | 6.19 | 37.34 | 96.17 | 0.085 | Chloroplast |
| MELO3C025328 | 1701 | 566 | 61733.01 | 8.81 | 31.75 | 87.05 | -0.069 | Mitochondrial |
| MELO3C017125 | 1512 | 503 | 54555.85 | 5.32 | 25.17 | 92.94 | -0.001 | Cytoplasmic |
| MELO3C017100 | 1665 | 554 | 60067.44 | 7.23 | 33.22 | 89.10 | -0.084 | Chloroplast |
| MELO3C008245 | 2154 | 717 | 77669.64 | 6.20 | 32.92 | 105.54 | -0.096 | Mitochondrial |
| MELO3C009229 | 1941 | 646 | 69861.68 | 5.45 | 34.89 | 104.78 | -0.032 | Cytoplasmic |
| MELO3C014601 | 1446 | 481 | 54034.52 | 8.55 | 39.02 | 95.90 | -0.043 | Plasma Membrane |
| MELO3C004383 | 1149 | 382 | 41812.14 | 8.30 | 26.07 | 93.19 | -0.022 | Chloroplast |
| MELO3C004430 | 1620 | 539 | 58481.63 | 7.60 | 37.92 | 91.37 | -0.002 | Chloroplast |
| MELO3C010494 | 1296 | 431 | 47243.77 | 8.11 | 34.49 | 104.29 | 0.046 | Cytoplasmic |
| MELO3C010493 | 1560 | 519 | 57189.43 | 8.89 | 35.71 | 99.79 | 0.02 | Mitochondrial |
| MELO3C017542 | 1395 | 464 | 51874.30 | 8.76 | 37.35 | 97.93 | -0.06 | Cytoplasmic |
| MELO3C007705 | 3132 | 1043 | 114376.90 | 8.10 | 48.04 | 77.52 | -0.38 | Nuclear |
| MELO3C019622 | 1626 | 541 | 57939.60 | 6.98 | 38.09 | 95.21 | 0.039 | Chloroplast |
| MELO3C021272 | 1815 | 604 | 66876.10 | 7.20 | 36.64 | 92.33 | -0.032 | Mitochondrial |
| MELO3C002203 | 1620 | 539 | 60367.76 | 6.33 | 39.65 | 95.45 | -0.09 | Mitochondrial |
2.2. Chromosome Distribution of Melon ALDH Genes
Utilizing the location information of ALDH gene family members on melon chromosomes, a distribution map of the ALDH gene in melon was constructed using TBtools software (Figure 1). The findings revealed that ALDH genes were present on all chromosomes except for 6, 9, and 10, which lacked these genes, and a certain degree of variation in distribution quantity. For instance, the highest number of distributed genes were found on chromosomes 2 and 7, with three on each. Conversely, chromosomes 3, 4, 8, and 12 harbored the fewest genes, each with only one. This suggests that chromosomes 2 and 7 may play a significant role in the evolution of ALDH in melon. By observing gene positioning on the chromosomes, it was observed that ALDH genes in melon predominantly resided on the termini of chromosomes, with some located centrally. Furthermore, one pair of tandem repeat genes and fragment repeat genes, namely MELO3C010494/MELO3C010493 and MELO3C008245/MELO3C009229 respectively, were identified within the 17 ALDH genes of melon.
2.3. Phylogenetic Tree Analysis of ALDH Family Genes
To elucidate the phylogenetic relationship of ALDH genes in melon, we procured the protein sequences of acknowledged ALDH genes from Arabidopsis and rice genome database. These sequences were aligned with the 17 ALDH protein sequences in melon to facilitate phylogenetic analysis and construct a phylogenetic tree (Figure 2). Subsequently, melon's ALDH genes were partitioned into 10 subfamilies based on Arabidopsis and rice groupings. Family 3 was identified as the most abundant, containing four melon ALDH genes, followed by Family 2 with three. Aside from Family 6 and 18, which each comprised of two melon ALDH genes, the remaining six subfamilies each held a single ALDH gene.
We discovered seven pairs of direct homologous genes between the ALDH family of muskmelon and Arabidopsis, namely MELO3C004430/AT2G24270, MELO3C017125/AT1G74920, MELO3C025328/AT3G24503, MELO3C017100/AT3G48000, MELO3C002203/AT5G62530, MELO3C007705/AT2G14170, and MELO3C017542/AT4G36250. Additionally, we found one family of orthologous genes between melon and rice ALDH, specifically MELO3C024345/Os09g0440300. In the melon ALDH gene family, only one pair of paralogs genes, MELO3C010493/MELO3C010494, was identified. Evolutionarily related ALDH genes, which exhibit similar biological functions and gene structures, were noted. Future studies can predict the biological function of the melon ALDH gene based on previously reported ALDH genes in Arabidopsis and rice.
2.4. Gene Structure and Conserved Motif Analysis of Melon ALDH Genes
We conducted a phylogenetic tree construction using the protein sequences from 17 melon ALDH genes, underpinned by gene structure and conserved motif analysis in correlation with melon genome files (Figure 3). The findings demonstrated that melon's ALDH gene family could be segregated into 10 distinct subfamilies. ALDH genes originating from the same subfamily exhibited a tendency to cluster, which essentially aligns with the ALDH gene clustering outcomes from melon, Arabidopsis, and rice (Figure 2).
In the context of multi-gene families, the structural disparities in exon-introns within the same family serve as pivotal factors in the evolutionary process. As per the gene structure of ALDH genes in the melon family, the exon count fluctuates between 7 to 20. The genes MELO3C019622 and MELO3C008245 are characterized by the highest number of exons, tallying at 20, succeeded by MELO3C018583 and MELO3C007705 from the same subfamily, each having 19 exons. The lowest count is attributed to MELO3C004383, with a mere 7 exons. Genes within the same subfamily generally exhibit similarities in their exon-intron structure. The resemblance in these gene structures, coupled with high sequence homology, suggests that gene duplication events transpired in the evolution of the melon's ALDH gene family.
Conserved motif analysis revealed that the motif sequences of Family 3, 22, 11, 5, 10, and 2 were nearly identical, in the sequence of 3, 1, and 2. The motifs in Family 6 and Family 7 displayed the same sequence, 3, 1. Similarly, the motifs in Family 12 and Family 18 followed the same sequence, incorporating only motif 3. This suggests that ALDH genes within these similar subfamilies could potentially share similar biological functions.
2.5. Synteny Analysis of ALDH Genes among Melon, Arabidopsis, and Rice
To deepen our understanding of the evolutionary details of ALDH genes in melon, we conducted a synteny analysis of these genes (Figure 4). Our findings revealed that, out of the 17 ALDH genes in melon, there was only one pair of repeated gene fragments, specifically MELO3C008245/MELO3C009229. Furthermore, we analyzed ALDH orthologous genes between melon, Arabidopsis, and rice.Among the melon's ALDH family, there were nine genes (MELO3C018583, MELO3C024345, MELO3C025328, MELO3C017100, MELO3C008245, MELO3C010494, MELO3C0117542, MELO3C019622, MELO3C021272) and twelve in Arabidopsis (AT1G23800, AT1G44170, AT1G54100, AT1G74920, AT1G79440, AT2G14170, AT2G39800, AT3G66658, AT3G24503, AT3G48170, AT3G55610, AT4G36250). We identified 12 collinearity relationships among these. We also discovered three collinearity relationships between three melon ALDH genes (MELO3C024345, MELO3C017125, MELO3C008245) and three rice ALDH genes (Os01g0848200, Os04g0464200, Os09g0440300). However, the remaining six ALDH genes in melon (MELO3C014601, MELO3C004383, MELO3C004430, MELO3C010493, MELO3C007705, MELO3C002203) exhibited no collinearity with those in Arabidopsis and rice. Indicated that these genes demonstrate a certain degree of conservation within the ALDH family.
2.6. Analysis of the Cis-Acting Elements in Melon ALDH Genes
Based on the cis-acting element analysis, we identified 12 distinct cis-acting elements in the promoter sequences of the melon ALDH genes (Figure 5A). Light responsiveness elements constituted the majority, accounting for 45.6% of the total and were uniformly distributed among all 17 ALDH genes in melon. The second most prevalent were the hormone-related cis-acting elements (abscisic acid responsiveness, MeJA-responsiveness, and salicylic acid responsiveness), contributing to 28.6% of the total, following photoresponsive elements (Figure 5B). Additionally, we examined elements related to stress response and endosperm expression. Our findings suggest that the ALDH gene could respond to a range of hormones and stress conditions. Thus, the identified response elements could significantly impact the stress response capability of the ALDH gene under stress conditions.
2.7. Tissue-Specific Expression Analysis of Melon ALDH Genes
Utilizing previously published transcriptome data from an array of melon tissues and fruit at different developmental stages, we were able to analyze the tissue-specific expression profiles of the melon's ALDH gene family members during various stages of development (Figure 6). The histospecificity analysis of the ALDH gene family revealed high expression levels of MELO3C018583 and MELO3C017125 across all organs. Notably, MELO3C010493 is expressed specifically in male flowers ang showed nearly an 8-fold greater expression level compared to other organs. Similar to MELO3C025328 and MELO3C009229, which presented specific expression only in the roots and male flowers. Conversely, MELO3C008245 and MELO3C021272 exhibited low expression in male flowers, but high expression in other organs. MELO3C024345 and MELO3C004383 were highly expressed in all organs except the ovary, where expression levels were significantly lower. The expression of MELO3C019622 and MELO3C002203 were more evenly distributed across all organs. Interestingly, MELO3C004430 displayed significantly higher expression levels in both male and female flowers than other organs, which suggests it may play a role in the formation of melon flower organs and fruits. Among the 17 ALDH genes, only MELO3C007705 showed no specific organ expression and maintained low expression levels, suggesting this gene is not involved in the growth and development of melon organs.
The expression profiles of ALDH family genes during different stages of fruit development showed that MELO3C018583 and MELO3C017125 were highly expressed at all four stages (growing, ripening, climacteric, and post-climacteric). This pattern, combined with their organ-specific expression, suggests these genes participate in melon organ development and have a role in fruit ripening and development. MELO3C024345, MELO3C010494, MELO3C019622, and MELO3C002203 also displayed high expression levels in all four stages, second only to MELO3C018583 and MELO3C017125. However, MELO3C0225328, MELO3C009229, MELO3C014601, and MELO3C017542 exhibited very low or no expression throughout the entire fruit development stage. MELO3C010493 and MELO3C004383 showed stage-specific expression during the ripening and growing stages, respectively, while MELO3C017100 was significantly more expressed during the growing stage than at other stages. No expression was observed during the climacteric and post-climacteric stages, suggesting this gene may be involved only in the growth and ripening of muskmelon fruits.
2.8. Expression Patterns Analysis of Melon ALDH Genes under Abiotic Stresses
To investigate the expression patterns of the ALDH gene in melon under varying abiotic stress conditions, we used published transcriptome sequencing data from melon studies conducted under salt, chilling, and waterlogged stress. This allowed us to analyze ALDH gene expression levels under these stresses (Figure 7).
Under salt stress (Figure 7A), only five ALDH genes – MELO3C021272, MELO3C007705, MELO3C014601, MELO3C017542, and MELO3C010493 – were not differentially expressed compared to the control. Conversely, MELO3C017100, MELO3C025328, and MELO3C004430 showed significant down-regulation in both salt-sensitive and salt-resistant materials. Other genes, particularly MELO3C004383 and MELO3C024345, were significantly up-regulated in these same materials.
In response to chillingstress (Figure 7B), only MELO3C004430 was significantly up-regulated in both chilling sensitive and resistant materials when compared with the control. MELO3C004383 showed a down-regulation in sensitive materials and an up-regulation in resistant materials, whereas MELO3C025328 exhibited an opposed pattern – up-regulation in sensitive and down-regulation in resistant materials. Other genes – MELO3C017100, MELO3C007705, MELO3C021272, MELO3C014601, and MELO3C017542 – were significantly down-regulated in both types of materials. However, down-regulated expressions of MELO3C018583, MELO3C002203, MELO3C019622, and MELO3C024345 were only observed in sensitive materials.
When under waterlogging stress (Figure 7C), MELO3C025328 was significantly down-regulated after 24 hours of treatment, but showed a significant up-regulation after 72 hours. Both MELO3C017542 and MELO3C004430 showed significant down-regulation in all four treatments. MELO3C017100, MELO3C014601, and MELO3C010493 did not present differential expression at the 6-hour mark but demonstrated down-regulation at 24, 48, and 72 hours.
2.9. Expression Patterns Analysis of Melon ALDH Genes under Biotic Stresses
To comprehend the expression patterns of muskmelon under various biological stresses, we reanalyzed the published transcriptomic data of muskmelon under powdery mildew, fusarium wilt, and gummy stem blight stress. This reanalysis was performed in conjunction with the melon genome data (DHL92 V4) using TBtools software to generate expression heat maps (Figure 8).
During powdery mildew stress (Figure 8A), MELO3C004430 and MELO3C025328 exhibited identical differential expression patterns. Their expression levels significantly decreased and were downregulated after 24 hours of treatment in the sensitive material, then increased significantly and were upregulated following 168 hours of treatment. MELO3C014601 showed downregulation in sensitive materials but upregulation in resistant materials. MELO3C004383 demonstrated a conflux of both upregulated and downregulated expressions in resistant materials. Both MELO3C021272 and MELO3C017100 were significantly downregulated in both sensitive and resistant materials, while MELO3C008245 showed significant downregulation only in sensitive materials. Conversely, MELO3C018583, MELO3C024345, and MELO3C017125 were significantly downregulated in resistant materials.
Under fusarium wilt stress (Figure 8B), compared with the control materials, MELO3C008245, MELO3C024345, and MELO3C017125 showed the highest expression and significantly upregulated in both sensitive and resistant materials. Similarly, MELO3C009229 was significantly upregulated in both sensitive and resistant materials. In contrast, MELO3C025328 and MELO3C007705 demonstrated significant downregulation in both types of materials. MELO3C017125, MELO3C002203, and MELO3C010494 were significantly upregulated in sensitive materials, while MELO3C004430 and MELO3C017542 were significantly downregulated. Notably, MELO3C019622 was only significantly upregulated in resistant materials.
Under gummy stem blight stress (Figure 8C), five ALDH genes – MELO3C024345, MELO3C010494, MELO3C018583, MELO3C004383, and MELO3C002203 – were significantly upregulated in both sensitive and resistant materials compared with control materials. Excluding MELO3C010494, the expression levels of the other four ALDH genes showed significant changes in sensitive materials. MELO3C017100 was downregulated in sensitive materials yet significantly upregulated in resistant ones. Conversely, MELO3C004430 and MELO3C008245 were significantly downregulated in sensitive materials. MELO3C014601, MELO3C017542, and MELO3C025328 were significantly upregulated in resistant materials. Lastly, MELO3C019622 demonstrated a significant downregulation in resistant materials.
2.10. Regulation Patterns of Melon ALDH Genes under Stresses
The expression profiles of the melon ALDH gene family under various stresses were utilized to summarize and label the stress response of the ALDH gene family, and a heat map of ALDH genes under differing stresses was created using TBtools software (Figure 9). As indicated in the figure, 17 ALDH genes in melon participate in the plant's stress response to varying extents. Notably, the ALDH genes MELO3C025328 and MELO3C004430 exhibited significant differential expression in both abiotic and biological stress scenarios. This indicates that these two genes are capable of initiating a positive response under stress conditions. Hence, they may serve as key candidate genes for future studies examining melon's stress responses. Interestingly, while MELO3C017100 did not exhibit different expression under fusarium wilt stress, it was significantly downregulated under all other five forms of stress. Some ALDH genes in melon, such as MELO3C010493, only showed downregulation under waterlogging stress conditions. In the context of abiotic stress, most of the 17 ALDH genes were upregulated under salt stress, but downregulated under chilling and waterlogging stresses. Under biological stress, the majority of these ALDH genes were downregulated in response to powdery mildew stress but upregulated in response to fusarium wilt and gummy stem blight stress.These findings highlight the differential expression of melon's ALDH gene family under abiotic and biological stresses, providing a favorable starting point for further research into the molecular biological functions of the ALDH gene family in melons.
2.11. RT-qPCR Analysis of the Melon ALDH Gene Family
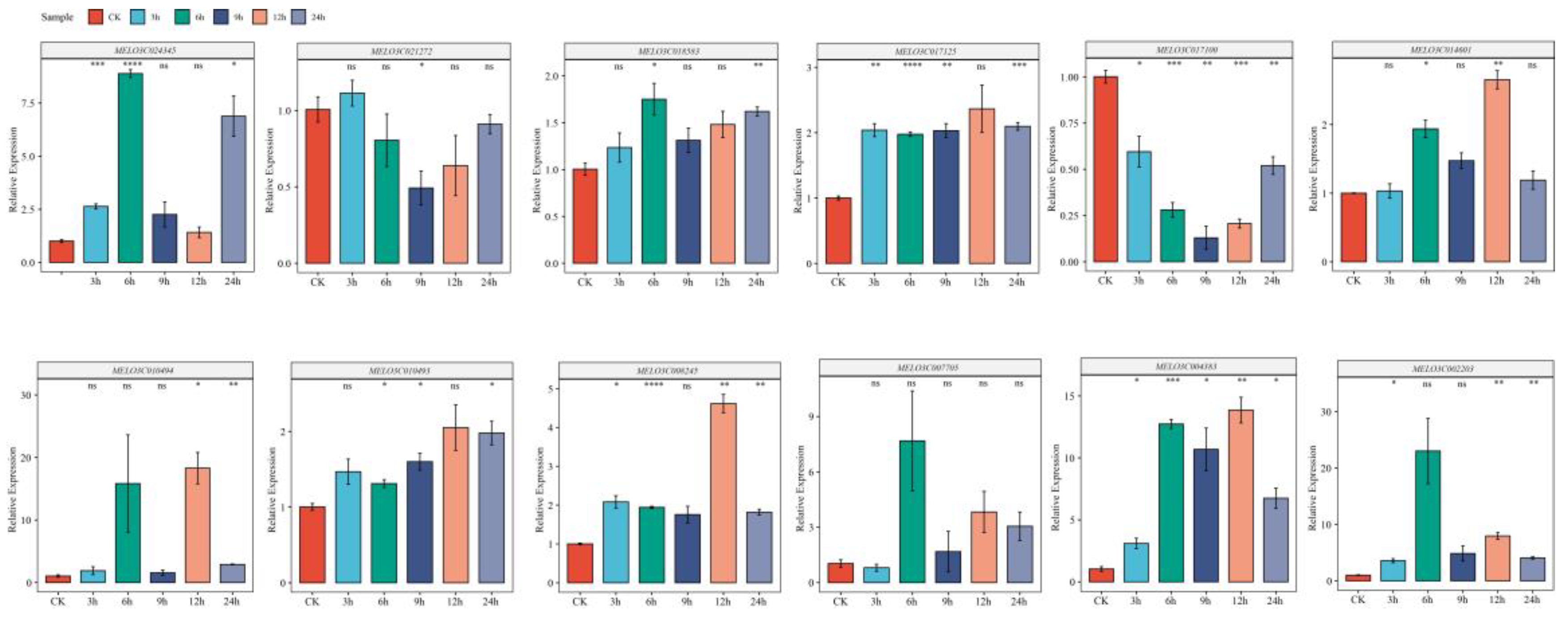
To evaluate the precision of our transcriptomic data analysis, we selected 10 genes from the CmALDH gene family. We assessed the impact of salt stress treatment (300mM) at varying time intervals (0h, 3h, 6h, 12h, and 24h) on their expression levels using the RT-qPCR method (Figure 10). In this investigation, the transcriptomic data was salt stress treatment after 24 hours. According to our RT-qPCR test results, the expression patterns were generally consistent for the two sets of data we focused on. Such as MELO3C024345, MELO3C018583, MELO3C007705, and MELO3C002203, peaked in expression after 6 hours of NaCl treatment (with CK serving as the control), showing significant differences in comparison to the control. Conversely, other genes, including MELO3C017125, MELO3C014601, MELO3C010494, MELO3C010493, MELO3C008245, and MELO3C004383, exhibited their highest expression levels and significant differences at the 12-hour mark. Of these, the gene MELO3C017100 displayed a substantial decrease in expression following stress treatment, reaching its lowest point after 9 hours, before demonstrating an increasing trend. The expression level of MELO3C004383 showed a rising pattern post-stress, exhibiting significant differences compared to the control. This increase was particularly notable at the 12-hour point, after which it started to decrease.
3. Discussion
Research into plant responses to both abiotic and biological stresses has consistently attracted the attention of scientists. Stress has the potential to not only diminish the yield and quality of food crops [59] but also, in severe instances, result in plant death, thereby incurring substantial losses in agricultural production [60]. The ALDH enzyme, aside from its protective and detoxifying influence, also plays a crucial role in plant stress response [61]. The most prominently studied gene within the ALDH gene family is the betaine aldehyde dehydrogenase gene. This gene, belonging to the ALDH10 family, is a well-established osmoregulator and has demonstrated an effective response to adversity. It becomes upregulated in response to both drought and salt stress, enhancing plant resilience to these stresses [62]. As scientific and technological progress has allowed for the publication of an increasing amount of genome and transcriptome sequencing information, it's become particularly valuable to utilize this data to identify stress-related genes. This approach not only increases the usefulness of the existing data but also holds significant implications for related research [63]. Melon, the second cucurbit crop to have its whole genome sequenced (as early as 2012) [32], has yet to have any studies published on its ALDH gene family. This lack of information significantly hinders research into melon's biological functions linked to ALDH genes. Therefore, this study aims to fill this gap by conducting a whole genome identification and expression pattern analysis of the ALDH gene family in melons. The results will provide a theoretical framework for future ALDH gene family studies in melons and contribute to the identification of advantageous genes for resistance breeding in melons.
In this study, we identified 17 ALDH genes in the melon genome using the latest genomic information and advanced bioinformatics analysis methods. These findings differ from the number of ALDH genes identified in other species, which may be due to variations in the number of ALDH gene family members across different species and distinct stages of evolution [64]. It could also be linked to the specific characteristics of the melon's chromosomes and genome [65]. In our phylogenetic analysis, these 17 melon ALDH genes were classified into 10 subfamilies, each with a largely consistent gene structure and motif arrangement. Comparative analysis with the model plants Arabidopsis and rice revealed that there are 7 pairs of directly homologous genes with Arabidopsis, and only one pair with rice's ALDH gene family. This suggests potential similarities in biological functions, allowing for future melon studies to refer to the homologous genes in Arabidopsis and rice. Our gene duplication analysis within the melon's ALDH gene family revealed a pair of tandem and fragment repeats, suggesting that the amplification of the melon's ALDH gene was solely due to these repeats, a finding that aligns with reports on gene families in other plants [66].
Transcriptomics forms a critical component of genomics research. Bolstered by the ongoing advancements in high-throughput sequencing technology, transcriptome sequencing is becoming increasingly prevalent in plant-related studies. This has led to deepened insights into gene expression regulatory mechanisms and cellular functional diversity in plants [67,68]. Numerous transcriptome sequencing projects have used melons as their subject, with the resulting data undergoing validation via qRT-PCR and rigorous peer review before publication [69,70]. This process has led to an extensive database of melon transcriptomes. This study leverages this wealth of published transcriptome sequencing data to analyze the expression patterns of 17 ALDH genes in melons across different tissues, developmental stages, and stress conditions.
ALDH's role in plant organ development has been widely reported. For instance, the ALDH2B2 (Rf2a) gene is linked to cytoplasmic male sterility in maize and plays a key role in anther development [71]. ALDH is also highly expressed in rice anthers during early meiosis to microspore [72]. The ALDH3F1 mutant triggers early flowering in plants, while its overexpression causes delayed flowering [73]. In grapes, the expression levels of three genes (ALDH2B8, ALDH3H5, and ALDH18B1) noticeably increase during fruit development and maturation, while the levels of ALDH2B4 and ALDH5F1 significantly decrease [11]. In this study, we found that all 17 ALDH genes in melons were expressed to varying degrees in different melon tissues, with some genes exhibiting tissue-specific expression. This specific expression pattern across various melon tissues contributes to the synergistic regulation of melon growth and development. Only four ALDH genes were not expressed at different developmental stages of melon fruit. Notably, the MELO3C017100 gene exhibited the highest expression level during the fruit growth stage and in male and female flowers and the ovary. However, its expression level dropped dramatically as the fruit matured, indicating that this gene primarily contributes to melon fruit growth.
The ALDH gene family's response to stress in plants has become a key focus of recent studies, suggesting that this gene family is crucial for plant stress resistance. In European sweet cherry, fluorescence quantitative experiments confirmed that PaALDH17's expression under salt stress was significantly higher than that of other genes. PaALDH17 was subsequently cloned and transferred to Arabidopsis, where transgenic PaALDH17 plants exhibited stronger stress resistance and fewer changes in various physiological indices under salt stress, compared to wild-type plants [74]. In grapes, VvALDH2B4 and VvALDH2B8 expression levels significantly increased under drought and salt stress [11]. Moreover, overexpressing VvALDH2B4 in wild Chinese grapes led to a decrease in MDA levels, significantly improving resistance to pathogenic bacteria [75]. In bamboo, ALDH gene expression significantly increased under drought stress compared to the control, with PeALDH2B2 responding to drought stress by interacting with PeGPB1[76]. Wang et al. [16] discovered that the wheat ALDH gene TraeALDH7B1-5A exhibited significant differences under both drought and salt stress. Its soybean homologous gene, GmALDH7B1, was significantly upregulated in both roots and leaves under 20% PEG treatment. To understand the melon ALDH gene family's role in abiotic stress, this study analyzed the ALDH family genes' expression patterns under salt, chilling, and waterlogging stress using published melon transcriptome sequencing data. It was revealed that over half of the melon ALDH genes displayed significantly upregulated expression under salt stress, in line with previous reports on plant ALDH genes' upregulated expression under salt and drought stress [20]. In contrast, most melon ALDH genes' expression was significantly downregulated under chilling and waterlogging stress. This suggests that these ALDH genes are negatively regulated when transcribed into mRNA under stress, leading to reduced expression [77]. Interestingly, some melon ALDH genes were differentially expressed under all three abiotic stresses, while others were only differentially expressed under one stress. For instance, MELO3C017125, MELO3C009229, MELO3C010494, and MELO3C008245 were differentially expressed solely under salt stress, while their expression levels remained unchanged under chilling and waterlogged stress. MELO3C021272 and MELO3C007705 were differentially expressed only under chilling stress, and MELO3C010493 was differentially expressed only under waterlogging stress. These results suggest that while some ALDH genes can be induced under various abiotic stresses, others are only induced by a specific abiotic stress. In addition to abiotic stress analysis, this study also examined the ALDH gene family's expression patterns in melons under biological stress. It was observed that the 17 melon ALDH genes were primarily downregulated under powdery mildew stress and primarily upregulated under fusarium wilt stress, with a higher number of differentially expressed genes than under powdery mildew stress. These ALDH genes were positively responsive to these stresses and played a role in plant disease resistance [78]. Among these, MELO3C021272 was significantly downregulated under powdery mildew stress but not expressed under fusarium wilt stress. Similarly, MELO3C007705 and MELO3C009229 were differentially expressed under fusarium wilt stress. However, MELO3C010493 was not differentially expressed under any of these three biological stresses, suggesting that this gene did not contribute to melon's response to these biological stresses. The functional characteristics of melon ALDH genes showed that 17 ALDH genes were differentially expressed under both abiotic and biological stresses, with MELO3C024328 and MELO3C004430 being differentially expressed under all stresses (salt, chilling, waterlogging, powdery mildew, fusarium wilt, and gummy stem blight). Interestingly, MELO3C024328 was upregulated and downregulated in chilling, waterlogged, and powdery mildew stress, suggesting that these two melon ALDH genes play a role in plants facing these stresses.
4. Materials and Methods
4.1. Identification and Chromosomal Distribution of Melon ALDH Gene Family Members
We retrieved the melon DHL92 protein group sequence file from the Cucurbitaceae genome database (http://cucurbitgenomics.org/v2/, accessed 5 May 2024), and constructed a local database [40]. Subsequently, the HMM model file for the ALDH gene family (PF00171) was downloaded from the InterPro database (https://www.ebi.ac.uk/interpro/, accessed 5 May 2024) [41]. We utilized the hmmsearch program [42] to screen the ALDH gene ID (E<1×10^5) in the melon protein database, and a Perl script was employed to extract the protein sequence data of the candidate genes. These sequences were then uploaded to the online platforms InterPro [43], SMART [44], and NCBI [45] for domain verification of identified ALDH gene candidates in melon. Sequences that did not contain the ALDH domain were eliminated, aiding in the identification of ALDH gene family members in melon. The ExPASy online tool [46] (https://web.expasy.org/protparam/, accessed 5 May 2024) was used to analyze the physical and chemical properties of the melon ALDH family genetic information. Lastly, the chromosome distribution map of the ALDH family genes in melon was constructed using the TBtools software [47].
4.2. Phylogenetic Analysis of ALDH Family Genes From Melon, Arabidopsis, and Rice
The sequence data for the ALDH gene family members in melon were uploaded to the online platform, GSDS (http://gsds.cbi.pku.edu.cn/, accessed 8 May 2024) [48], to execute a gene structure analysis. For the examination of conserved motifs within the ALDH gene family, the same sequence information was submitted to the online resource, MEME (http://meme-suite.org/tools/meme, accessed 8 May 2024) [49], with all parameters set to their default values. In order to construct a phylogenetic tree, we combined ALDH protein sequences of Arabidopsis and rice with those of melon. This collective dataset was then imported into the MEGA11 software [50]. The construction process employed the maximum likelihood method, with all parameters set to their default values.
4.3. Genetic Characterization and Phylogenetic Analysis of ALDH Family In Melon
The MCScanX software [51] was employed to examine tandem and fragment duplications within the ALDH gene family in melon. Furthermore, it was used to investigate the collinearity relationship among the ALDH gene families in Arabidopsis, rice, and melon. The findings were then visualized using the TBtools software.
4.4. Tissue-Specific Expression of Melon ALDH Family Genes
We reanalyzed publicly available melon transcriptome data from different tissues (PRJNA803327) [30] and developmental stages (PRJNA543288) [52] in conjunction with melon genome information. Subsequently, TBtools was utilized to generate a heatmap representation of ALDH gene family expression in various melon tissues and at different stages.
4.5. Analysis of Expression Patterns of Melon ALDH Gene Family Under Various Stresses
In order to investigate the effect of different stresses, we evaluated published melon transcriptome data under conditions including salt stress (PRJNA296827) [53], chilling stress (PRJNA553119) [54], waterlogging stress (PRJNA726294) [55], powdery mildew (PRJNA358655) [56], fusarium wilt (PRJNA842515) [57], and gummy stem blight (PRJNA681992) [58]. This data was reanalyzed in combination with melon genomic data, enabling us to identify differentially expressed genes. A heatmap of their expression under different stress conditions in melon was again visualized using TBtools.
4.6. RT-qPCR Analysis of Melon Under Salt Stress
We used "TQ1", a highly inbred muskmelon strain, provided by the Institute of Vegetables, Anhui Academy of Agricultural Sciences, as the experimental subject. Five leaves from a melon seedling at the single stage were subjected to salt stress (300mM NaCl) for 0, 3, 6, 9, 12, and 24 hours, respectively. The leaves were then flash-frozen in liquid nitrogen for RNA extraction and reverse transcription, followed by RT-qPCR. The reaction system consisted of: ddH2O (7µL), 2×Mix (10µL), cDNA (1µL), and both positive and negative primers (1µL each). The results obtained from three biological replicates were analyzed using the -△△ct method and processed via Excel 2021.
5. Conclusions
In this research, we undertook the first-ever identification of the ALDH gene family within the melon genome. We identified a total of 17 ALDH gene members, which were scattered across chromosomes 1, 2, 3, 4, 5, 7, 8, 11, and 12. Phylogenetic analysis grouped these genes into 10 distinct subfamilies, each exhibiting a largely similar gene structure. Collinearity analysis revealed a pair of fragment repeats among the 17 melon ALDH genes. Additionally, there were 12 collinear relationships between melon and Arabidopsis and three collinear relationships between melon and rice. Analysis of ALDH gene expression patterns during different developmental stages and in various melon tissues suggested that these genes contribute to melon growth, development, and fruit ripening. The examination of ALDH gene expression patterns under diverse stress conditions showed that these genes were differentially expressed at varying degrees in response to different stresses in melons. Particularly, the distinct expressions of MELO3C024328 and MELO3C004430 signpost these genes' crucial role in melon growth and development. RT-qPCR analysis results showed that most genes were highly expressed, signifying their participation in salt stress responses. Together, these findings pave the way for further exploration of the ALDH gene family in melons and offer promising genes for enhancing melon resistance breeding.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: Primers used for RT-qPCR.
Author Contributions
C.Y. and X.L. conceived the research and designed the experiments. D.Y. performed the research, analyzed the data, and wrote the manuscript. H.C., Y.Z. and Q.D. participated in downloading the transcriptome sequencing data and helped with the bioinformatics analysis. Y.W., M.W. and Y.Z. reviewed and edited the manuscript. Q.Z. and G.X. constructive suggestions were provided. All authors have read and agreed to the published version of the manuscript.
Funding
Please add: This research was funded by Anhui Province Key Research and Development Plan Basic Field Project (2023z04020019); the Youth Development Fund from Anhui Academy of Agricultural Sciences (QNYC-202110); the National Modern Agricultural Industrial Technology System Project (CARS-25); and the Anhui Province Vegetable industry Technology System.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bartels, D. Targeting detoxification pathways: an efficient approach to obtain plants with multiple stress tolerance? Trends Plant Sci 2001, 6, 284–286. [Google Scholar] [CrossRef] [PubMed]
- Lindahl, R. Aldehyde dehydrogenases and their role in carcinogenesis. Crit Rev Biochem Mol 1992, 27, 283–335. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Brocker, C.; Koppaka, V.; etc. Aldehyde dehydrogenases in cellular responses to oxidative/electrophilic stress. Free Radical Bio Med 2013, 56, 89–101. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, A.; Rzhetsky, A.; Hsu, L.C.; etc. Human aldehyde dehydrogenase gene family. Eur J Biochem 1998, 251, 549–557. [Google Scholar] [CrossRef]
- Kirch, H.H.; Bartels, D.; Wei, Y.; etc. The ALDH gene superfamily of Arabidopsis. Trends Plant Sci 2004, 9, 371–377. [Google Scholar] [CrossRef]
- Perozich, J.; Nicholas, H.; Wang, B.C.; etc. Relationships within the aldehyde dehydrogenase extended family. Protein Sci 1999, 8, 137–146. [Google Scholar] [CrossRef]
- Kotchoni, S.O.; Kuhns, C.; Ditzer, A.; etc. Over-expression of different aldehyde dehydrogenase genes in Arabidopsis thaliana confers tolerance to abiotic stress and protects plants against lipid peroxidation and oxidative stress. Plant Cell Environ 2006, 29, 1033–1048. [Google Scholar] [CrossRef]
- Islam, M.S.; Ghosh, A. Evolution, family expansion, and functional diversification of plant aldehyde dehydrogenases. Gene 2022, 829, 146522. [Google Scholar] [CrossRef]
- Brocker, C.; Lassen, N.; Estey, T.; etc. Aldehyde dehydrogenase 7A1 (ALDH7A1) is a novel enzyme involved in cellular defense against hyperosmotic stress. J Biol Chem 2010, 285, 18452–18463. [Google Scholar] [CrossRef]
- Vasiliou, V.; Bairoch, A.; Tipton, K.F.; etc. Eukaryotic aldehyde dehydrogenase (ALDH) genes: human polymorphisms, and recommended nomenclature based on divergent evolution and chromosomal mapping. Pharmacogenetics 1999, 9, 421–434. [Google Scholar]
- Zhang, Y.; Mao, L.; Wang, H.; etc. Genome-wide identification and analysis of grape aldehyde dehydrogenase (ALDH) gene superfamily. Plos One 2012, 7, e32153. [Google Scholar] [CrossRef] [PubMed]
- Hou, Q.; Bartels, D. Comparative study of the aldehyde dehydrogenase (ALDH) gene superfamily in the glycophyte Arabidopsis thaliana and Eutrema halophytes. Ann Bot-London 2015, 115, 465–479. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Han, B. Evolutionary and expression study of the aldehyde dehydrogenase (ALDH) gene superfamily in rice (Oryza sativa). Gene 2009, 431, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.L.; Zhang, Q.; Zhou, M.; etc. Aldehyde dehydrogenase protein superfamily in maize. Funct Integr Genomic 2012, 12, 683–691. [Google Scholar] [CrossRef]
- Jimenez-Lopez, J.C.; Lopez-Valverde, F.J.; Robles-Bolivar, P.; etc. Genome-Wide Identification and Functional Classification of Tomato (Solanum lycopersicum) Aldehyde Dehydrogenase (ALDH) Gene Superfamily. Plos One 2016, 11, e0164798. [Google Scholar] [CrossRef]
- Wang, W.; Jiang, W.; Liu, J.; etc. Genome-wide characterization of the aldehyde dehydrogenase gene superfamily in soybean and its potential role in drought stress response. Bmc Genomics 2017, 18, 518. [Google Scholar] [CrossRef]
- Islam, M.S.; Mohtasim, M.; Islam, T.; etc. Aldehyde dehydrogenase superfamily in sorghum: genome-wide identification, evolution, and transcript profiling during development stages and stress conditions. Bmc Plant Biol 2022, 22, 316. [Google Scholar] [CrossRef]
- Jiang, X.; Ren, J.; Ye, X.; etc. Genome-wide identification and analysis of the aldehyde dehydrogenase gene superfamily in Chinese cabbage (Brassica rapa subsp. pekinensis). Can J Plant Sci 2019, 99, 420–436. [Google Scholar] [CrossRef]
- Zhang, Y.; Mao, L.; Wang, H.; etc. Genome-wide identification and analysis of grape aldehyde dehydrogenase (ALDH) gene superfamily. Plos One 2012, 7, e32153. [Google Scholar] [CrossRef]
- Islam, M.S.; Hasan, M.S.; Hasan, M.N.; etc. Genome-wide identification, evolution, and transcript profiling of Aldehyde dehydrogenase superfamily in potato during development stages and stress conditions. Sci Rep-Uk 2021, 11, 18217–18284. [Google Scholar] [CrossRef]
- Li, X.; Guo, R.; Li, J.; etc. Genome-wide identification and analysis of the aldehyde dehydrogenase (ALDH) gene superfamily in apple (Malus x domestica Borkh.). Plant Physiol Bioch 2013, 71, 268–282. [Google Scholar] [CrossRef] [PubMed]
- Tola, A.J.; Jaballi, A.; Germain, H.; etc. Recent Development on Plant Aldehyde Dehydrogenase Enzymes and Their Functions in Plant Development and Stress Signaling. Genes-Basel 2020, 12, 51. [Google Scholar] [CrossRef] [PubMed]
- Brocker, C.; Vasiliou, M.; Carpenter, S.; etc. Aldehyde dehydrogenase (ALDH) superfamily in plants: gene nomenclature and comparative genomics. Planta 2013, 237, 189–210. [Google Scholar] [CrossRef] [PubMed]
- Kotchoni, S.O.; Kuhns, C.; Ditzer, A.; etc. Over-expression of different aldehyde dehydrogenase genes in Arabidopsis thaliana confers tolerance to abiotic stress and protects plants against lipid peroxidation and oxidative stress. Plant Cell Environ 2006, 29, 1033–1048. [Google Scholar] [CrossRef]
- Gautam, R.; Meena, R.K.; Woch, N.; etc. Ectopic expression of BrALDH7B2 gene encoding an antiquitin from Brassica rapa confers tolerance to abiotic stresses and improves photosynthetic performance under salt stress in tobacco. Environ Exp Bot 2020, 180, 104223. [Google Scholar] [CrossRef]
- Chen, J.; Wei, B.; Li, G.; etc. TraeALDH7B1-5A, encoding aldehyde dehydrogenase 7 in wheat, confers improved drought tolerance in Arabidopsis. Planta 2015, 242, 137–151. [Google Scholar] [CrossRef]
- Gu, H.; Pan, Z.; Jia, M.; etc. Genome-wide identification and analysis of the cotton ALDH gene family. Bmc Genomics 2024, 25, 513. [Google Scholar] [CrossRef]
- Cao, Y.; Wang, J.; Zhao, S.; etc. Overexpression of the aldehyde dehydrogenase AhALDH3H1 from Arachis hypogaea in soybean increases saline-alkali stress tolerance. Front Plant Sci 2023, 14, 1165384. [Google Scholar] [CrossRef]
- Rolim, P.M.; Fidelis, G.P.; Padilha, C.; etc. Phenolic profile and antioxidant activity from peels and seeds of melon (Cucumis melo L. var. reticulatus) and their antiproliferative effect in cancer cells. Braz J Med Biol Res 2018, 51, e6069. [Google Scholar] [CrossRef]
- Tian, Z.; Han, J.; Che, G.; etc. Genome-wide characterization and expression analysis of SAUR gene family in Melon (Cucumis melo L.). Planta 2022, 255, 123. [Google Scholar] [CrossRef]
- Ma, M.; Liu, S.; Wang, Z.; etc. Genome-Wide Identification of the SUN Gene Family in Melon (Cucumis melo) and Functional Characterization of Two CmSUN Genes in Regulating Fruit Shape Variation. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Mas, J.; Benjak, A.; Sanseverino, W.; etc. The genome of melon (Cucumis melo L.). P Natl Acad Sci Usa 2012, 109, 11872–11877. [Google Scholar] [CrossRef] [PubMed]
- Ruggieri, V.; Alexiou, K.G.; Morata, J.; etc. An improved assembly and annotation of the melon (Cucumis melo L.) reference genome. Sci Rep-Uk 2018, 8, 8088. [Google Scholar] [CrossRef]
- Castanera, R.; Ruggieri, V.; Pujol, M.; etc. An Improved Melon Reference Genome With Single-Molecule Sequencing Uncovers a Recent Burst of Transposable Elements With Potential Impact on Genes. Front Plant Sci 2019, 10, 1815. [Google Scholar] [CrossRef]
- Tian, Z.; Han, J.; Che, G.; etc. Genome-wide characterization and expression analysis of SAUR gene family in Melon (Cucumis melo L.). Planta 2022, 255, 123. [Google Scholar] [CrossRef]
- Wu, B.; Wang, L.; Pan, G.; etc. Genome-wide characterization and expression analysis of the auxin response factor (ARF) gene family during melon (Cucumis melo L.) fruit development. Protoplasma 2020, 257, 979–992. [Google Scholar] [CrossRef]
- Zheng, L.; Chao, Y.; Wang, Y.; etc. Genome-Wide Analysis of the LBD Gene Family in Melon and Expression Analysis in Response to Wilt Disease Infection. Genes-Basel 2024, 15. [Google Scholar] [CrossRef]
- Ma, M.; Liu, S.; Wang, Z.; etc. Genome-Wide Identification of the SUN Gene Family in Melon (Cucumis melo) and Functional Characterization of Two CmSUN Genes in Regulating Fruit Shape Variation. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- Jin, W.; Yan, W.; Ma, M.; etc. Genome-wide identification and expression analysis of the JMJ-C gene family in melon (Cucumis melo L.) reveals their potential role in fruit development. Bmc Genomics 2023, 24, 771. [Google Scholar] [CrossRef]
- Yu, J.; Wu, S.; Sun, H.; etc. CuGenDBv2: an updated database for cucurbit genomics. Nucleic Acids Res 2023, 51, D1457–D1464. [Google Scholar] [CrossRef]
- Paysan-Lafosse, T.; Blum, M.; Chuguransky, S.; etc. InterPro in 2022. Nucleic Acids Res 2023, 51, D418–D427. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Clements, J.; Eddy, S.R. HMMER web server: interactive sequence similarity searching. Nucleic Acids Res 2011, 39, W29–W37. [Google Scholar] [CrossRef]
- Paysan-Lafosse, T.; Blum, M.; Chuguransky, S.; etc. InterPro in 2022. Nucleic Acids Res 2023, 51, D418–D427. [Google Scholar] [CrossRef]
- Letunic, I.; Khedkar, S.; Bork, P. SMART: recent updates, new developments and status in 2020. Nucleic Acids Res 2021, 49, D458–D460. [Google Scholar] [CrossRef]
- Marchler-Bauer, A.; Bryant, S.H. CD-Search: protein domain annotations on the fly. Nucleic Acids Res 2004, 32, W327–W331. [Google Scholar] [CrossRef]
- Duvaud, S.; Gabella, C.; Lisacek, F.; etc. Expasy, the Swiss Bioinformatics Resource Portal, as designed by its users. Nucleic Acids Res 2021, 49, W216–W227. [Google Scholar] [CrossRef]
- Chen, C.; Wu, Y.; Li, J.; etc. TBtools-II: A "one for all, all for one" bioinformatics platform for biological big-data mining. Mol Plant 2023, 16, 1733–1742. [Google Scholar] [CrossRef]
- Hu, B.; Jin, J.; Guo, A.Y.; etc. GSDS 2.0: an upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; etc. The MEME Suite. Nucleic Acids Res 2015, 43, W39–W49. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol Biol Evol 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, H.; Wang, X.; etc. Detection of colinear blocks and synteny and evolutionary analyses based on utilization of MCScanX. Nat Protoc 2024, 19, 2206–2229. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Bai, S.; Dang, Z.; etc. Genome-wide identification and characterization of long non-coding RNAs involved in fruit ripening and the climacteric in Cucumis melo. Bmc Plant Biol 2019, 19, 369. [Google Scholar] [CrossRef] [PubMed]
- Tian, Z.; Han, J.; Che, G.; etc. Genome-wide characterization and expression analysis of SAUR gene family in Melon (Cucumis melo L.). Planta 2022, 255, 123. [Google Scholar] [CrossRef] [PubMed]
- DIAO, Q.; CAO, Y.; FAN, H.; etc. Transcriptome analysis deciphers the mechanisms of exogenous nitric oxide action on the response of melon leaves to chilling stress. Biol Plantarum 2020, 64, 465–472. [Google Scholar] [CrossRef]
- Zhang, H.; Li, G.; Yan, C.; etc. Depicting the molecular responses of adventitious rooting to waterlogging in melon hypocotyls by transcriptome profiling. 3 Biotech 2021, 11, 351. [Google Scholar] [CrossRef]
- Zhu, Q.; Gao, P.; Wan, Y.; etc. Comparative transcriptome profiling of genes and pathways related to resistance against powdery mildew in two contrasting melon genotypes. Sci Hortic-Amsterdam 2018, 227, 169–180. [Google Scholar] [CrossRef]
- Yang, T.; Liu, J.; Li, X.; etc. Transcriptomic Analysis of Fusarium oxysporum Stress-Induced Pathosystem and Screening of Fom-2 Interaction Factors in Contrasted Melon Plants. Front Plant Sci 2022, 13, 961586. [Google Scholar] [CrossRef]
- Intana, W.; Wonglom, P.; Suwannarach, N.; etc. Trichoderma asperelloides PSU-P1 Induced Expression of Pathogenesis-Related Protein Genes against Gummy Stem Blight of Muskmelon (Cucumis melo) in Field Evaluation. J Fungi 2022, 8. [Google Scholar] [CrossRef]
- Strzemski, M.; Dresler, S. Impact of Biotic/Abiotic Stress Factors on Plant Specialized Metabolites. Int J Mol Sci 2024, 25. [Google Scholar] [CrossRef]
- Fang, Y.; Jiang, J.; Ding, H.; etc. Phospholipase C: Diverse functions in plant biotic stress resistance and fungal pathogenicity. Mol Plant Pathol 2023, 24, 1192–1202. [Google Scholar] [CrossRef]
- Jacques, F.; Zhao, Y.; Kopecna, M.; etc. Roles for ALDH10 enzymes in gamma-butyrobetaine synthesis, seed development, germination, and salt tolerance in Arabidopsis. J Exp Bot 2020, 71, 7088–7102. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Sanchez, A.G.; Gonzalez-Segura, L.; Mujica-Jimenez, C.; etc. Amino acid residues critical for the specificity for betaine aldehyde of the plant ALDH10 isoenzyme involved in the synthesis of glycine betaine. Plant Physiol 2012, 158, 1570–1582. [Google Scholar] [CrossRef] [PubMed]
- Dmitriev, A.A.; Pushkova, E.N.; Melnikova, N.V. [Plant Genome Sequencing: Modern Technologies and Novel Opportunities for Breeding]. Mol Biol (Mosk) 2022, 56, 531–545. [Google Scholar] [CrossRef] [PubMed]
- Jain, M.; Tyagi, A.K.; Khurana, J.P. Genome-wide analysis, evolutionary expansion, and expression of early auxin-responsive SAUR gene family in rice (Oryza sativa). Genomics 2006, 88, 360–371. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Koo, D.H.; Li, Y.; etc. Chromosome rearrangements during domestication of cucumber as revealed by high-density genetic mapping and draft genome assembly. Plant J 2012, 71, 895–906. [Google Scholar] [CrossRef]
- Zhang, S.; Chen, C.; Li, L.; etc. Evolutionary expansion, gene structure, and expression of the rice wall-associated kinase gene family. Plant Physiol 2005, 139, 1107–1124. [Google Scholar] [CrossRef]
- Wang, L.; Li, P.; Brutnell, T.P. Exploring plant transcriptomes using ultra high-throughput sequencing. Brief Funct Genomics 2010, 9, 118–128. [Google Scholar] [CrossRef]
- Shulse, C.N.; Cole, B.J.; Ciobanu, D.; etc. High-Throughput Single-Cell Transcriptome Profiling of Plant Cell Types. Cell Rep 2019, 27, 2241–2247. [Google Scholar] [CrossRef]
- Yang, D.; Li, Y.; Zhu, M.; etc. Genome-Wide Identification and Expression Analysis of the Cucumber FKBP Gene Family in Response to Abiotic and Biotic Stresses. Genes-Basel 2023, 14. [Google Scholar] [CrossRef]
- Zhang, K.; He, S.; Sui, Y.; etc. Genome-Wide Characterization of HSP90 Gene Family in Cucumber and Their Potential Roles in Response to Abiotic and Biotic Stresses. Front Genet 2021, 12, 584886. [Google Scholar] [CrossRef]
- Liu, F.; Cui, X.; Horner, H.T.; etc. Mitochondrial aldehyde dehydrogenase activity is required for male fertility in maize. Plant Cell 2001, 13, 1063–1078. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Zhang, Z.; Zhao, Z.; etc. The mitochondrial aldehyde dehydrogenase OsALDH2b negatively regulates tapetum degeneration in rice. J Exp Bot 2020, 71, 2551–2560. [Google Scholar] [CrossRef] [PubMed]
- Gan, Q.; Luan, M.; Hu, M.; etc. Functional study of CYP90A1 and ALDH3F1 gene obtained by transcriptome sequencing analysis of Brassica napus seedlings treated with brassinolide. Front Plant Sci 2022, 13, 1040511. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Wang, X.; Wang, W.; etc. Genome-wide identification and expression analysis of the ALDH gene family and functional analysis of PaALDH17 in Prunus avium. Physiol Mol Biol Pla 2024, 30, 633–645. [Google Scholar] [CrossRef]
- Wen, Y.; Wang, X.; Xiao, S.; etc. Ectopic expression of VpALDH2B4, a novel aldehyde dehydrogenase gene from Chinese wild grapevine (Vitis pseudoreticulata), enhances resistance to mildew pathogens and salt stress in Arabidopsis. Planta 2012, 236, 525–539. [Google Scholar] [CrossRef]
- Xu, J.; Liu, L.; Huang, H.; etc. Genome-wide characterization and gene expression analyses of ALDH gene family in response to drought stress in moso bamboo (Phyllostachys edulis). Plant Physiol Bioch 2023, 202, 107954. [Google Scholar] [CrossRef]
- Wang, L.; Dai, W.; Shi, Y.; etc. Cloning and activity analysis of the highly expressed gene VviABCG20 promoter in seed and its activity is negatively regulated by the transcription factor VviDof14. Plant Sci 2022, 315, 111152. [Google Scholar] [CrossRef]
- Arick, M.N.; Hsu, C.Y. Differential Gene Expression Analysis of Plants. Methods Mol Biol 2018, 1783, 279–298. [Google Scholar] [CrossRef]
Figure 1.
Distribution of the melon ALDH family genes on the chromosomes. The genes marked in red color were tandem duplication gene pairs, and the genes marked in green color were segmental duplication gene.
Figure 1.
Distribution of the melon ALDH family genes on the chromosomes. The genes marked in red color were tandem duplication gene pairs, and the genes marked in green color were segmental duplication gene.
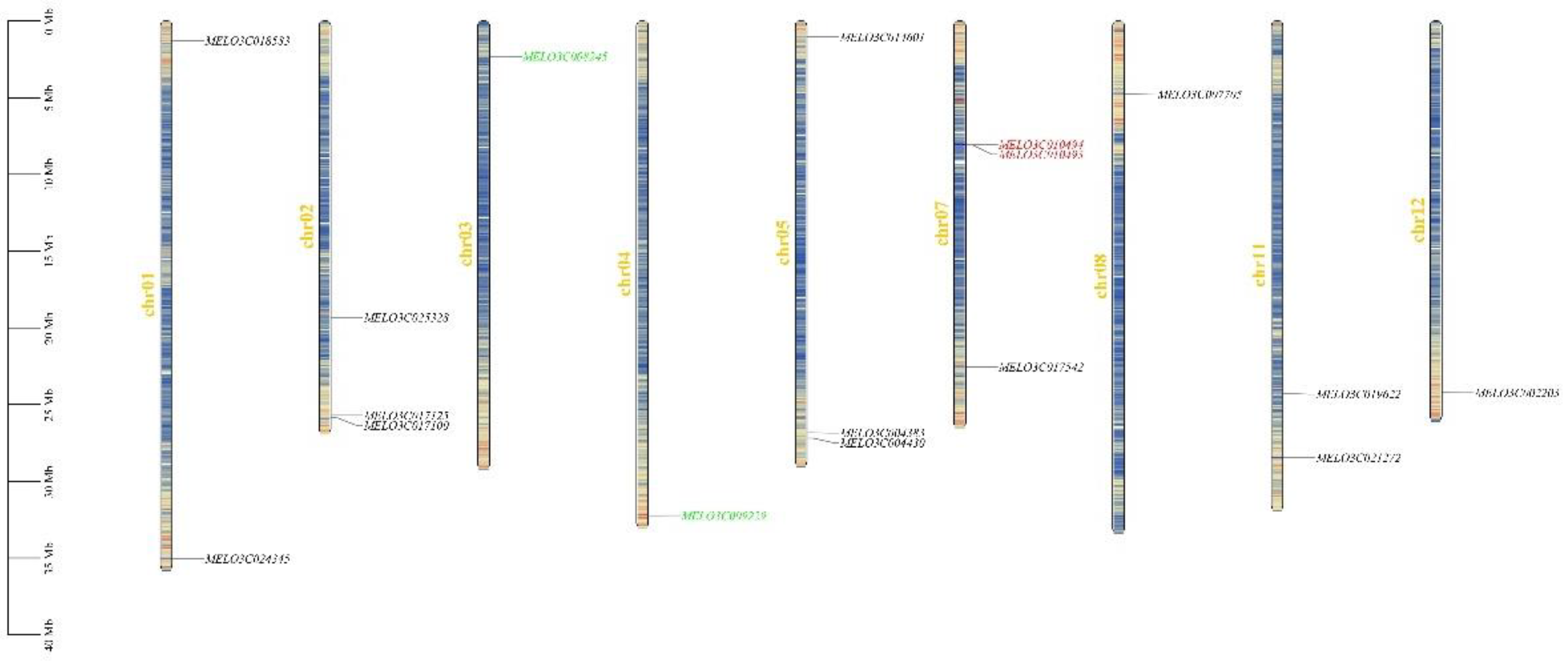
Figure 2.
Phylogenetic relationships of melon, rice, and Arabidopsis ALDH proteins.
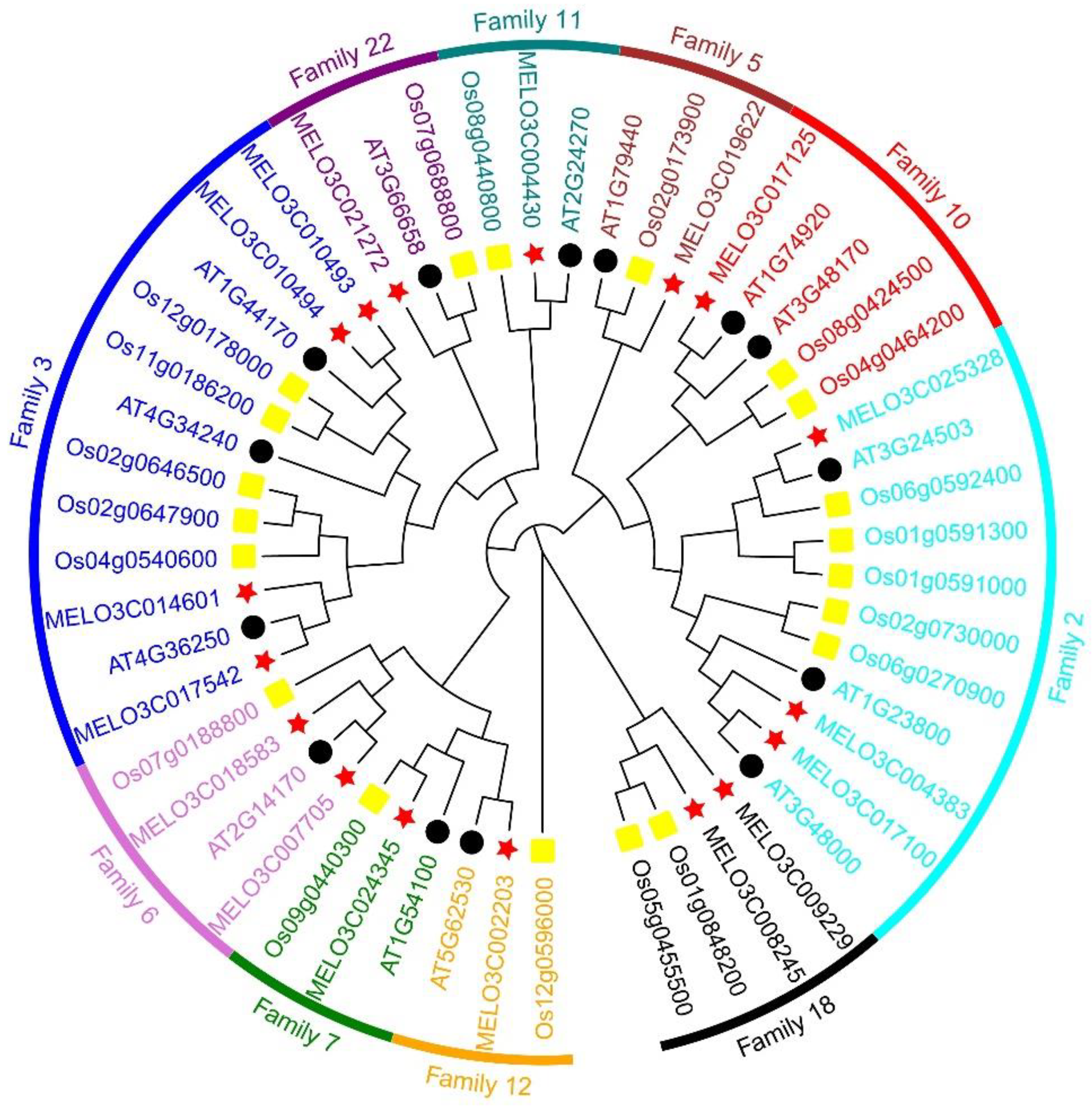
Figure 3.
Gene structures and protein motifs of the melon ALDH gene family.
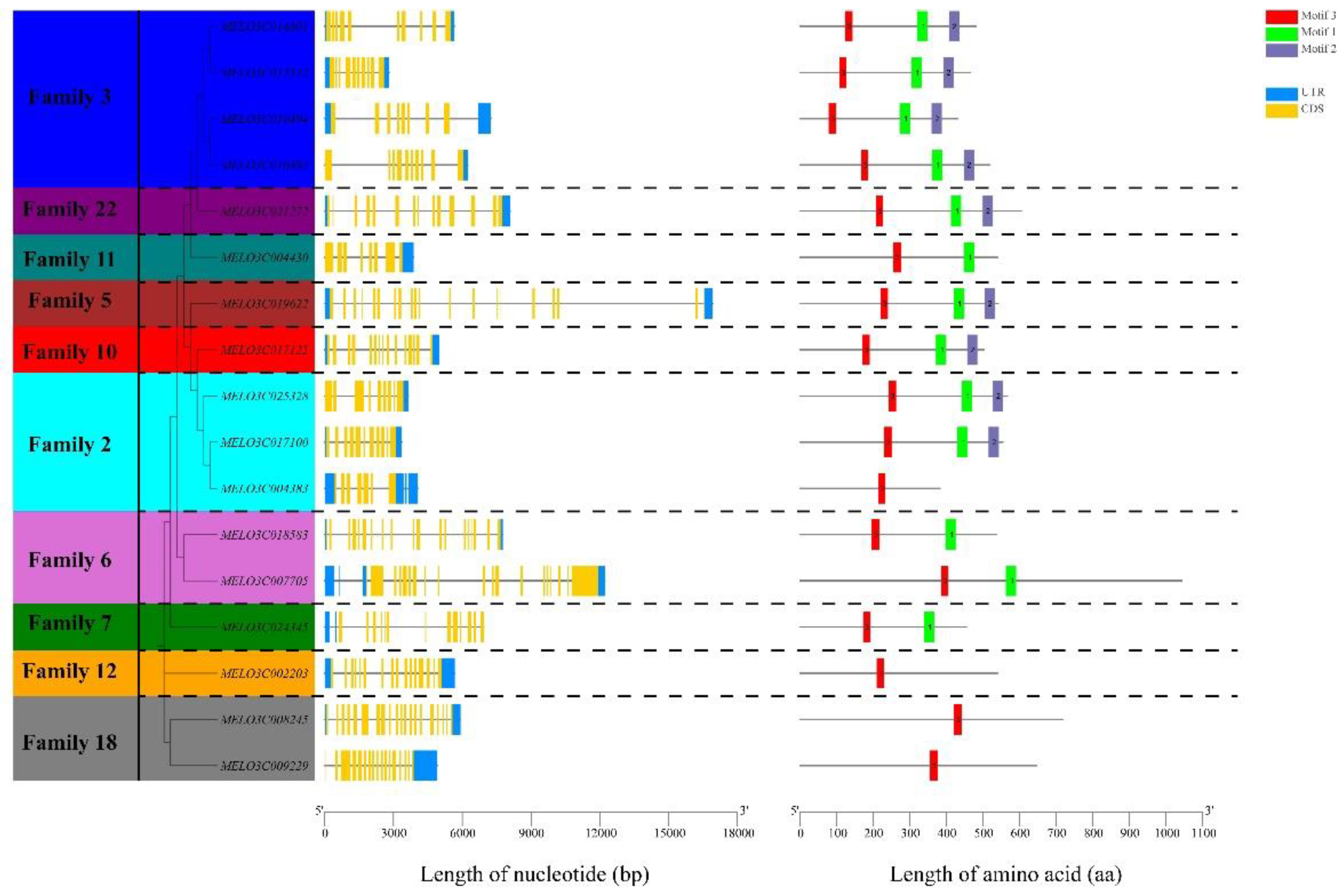
Figure 4.
Assessment of the ALDH gene duplication between melon, Arabidopsis, and rice. The green lines represented the segmentally duplicated ALDH genes in melon. The red lines represented the orthologous relationships of the ALDH genes between melon and Arabidopsis. The blue lines represent the orthologous relationships of the ALDH genes between melon and rice.
Figure 4.
Assessment of the ALDH gene duplication between melon, Arabidopsis, and rice. The green lines represented the segmentally duplicated ALDH genes in melon. The red lines represented the orthologous relationships of the ALDH genes between melon and Arabidopsis. The blue lines represent the orthologous relationships of the ALDH genes between melon and rice.
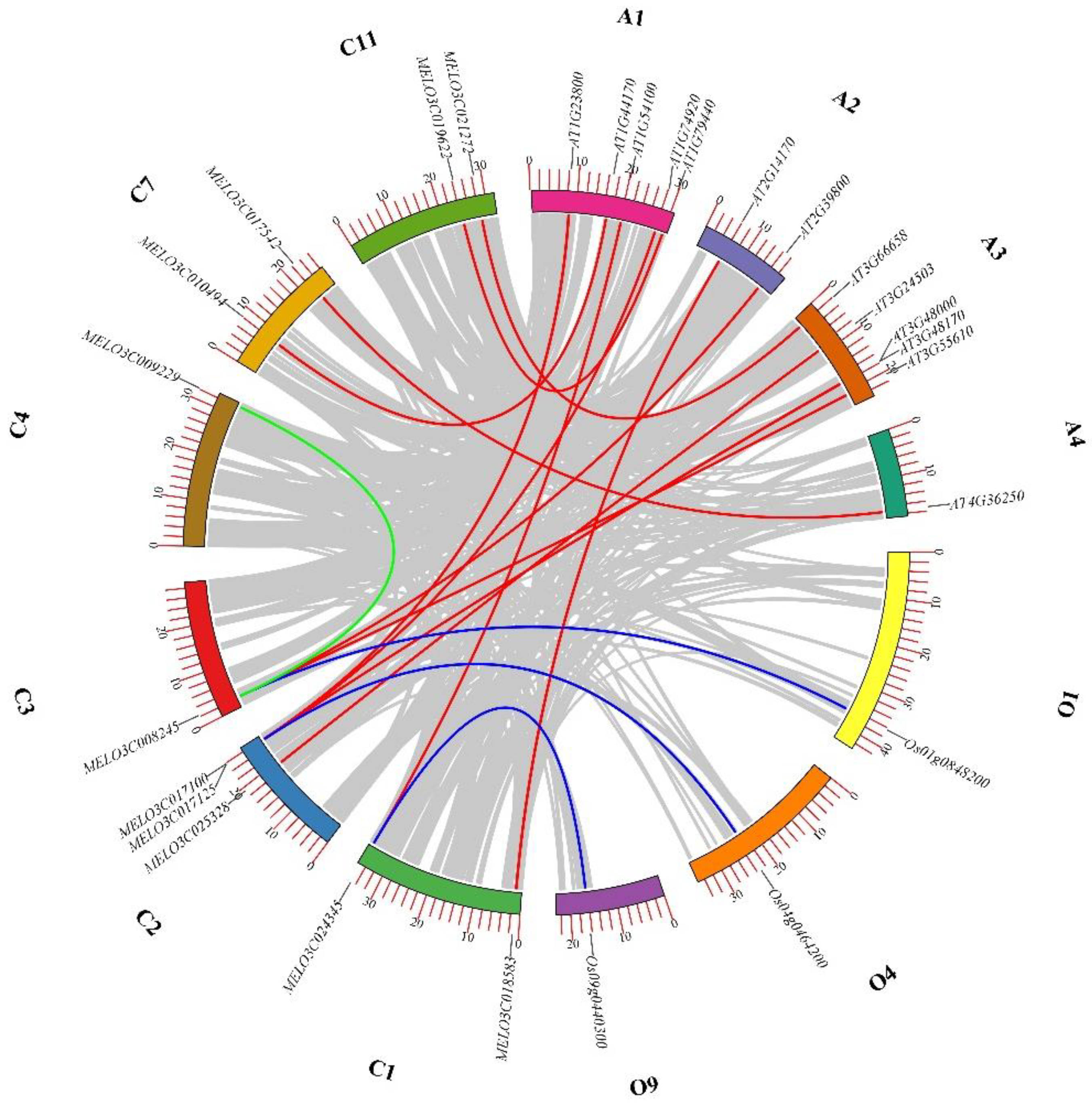
Figure 5.
Cis-regulatory elements in promoter region of melon ALDH genes (A) and their proportions (B).
Figure 5.
Cis-regulatory elements in promoter region of melon ALDH genes (A) and their proportions (B).
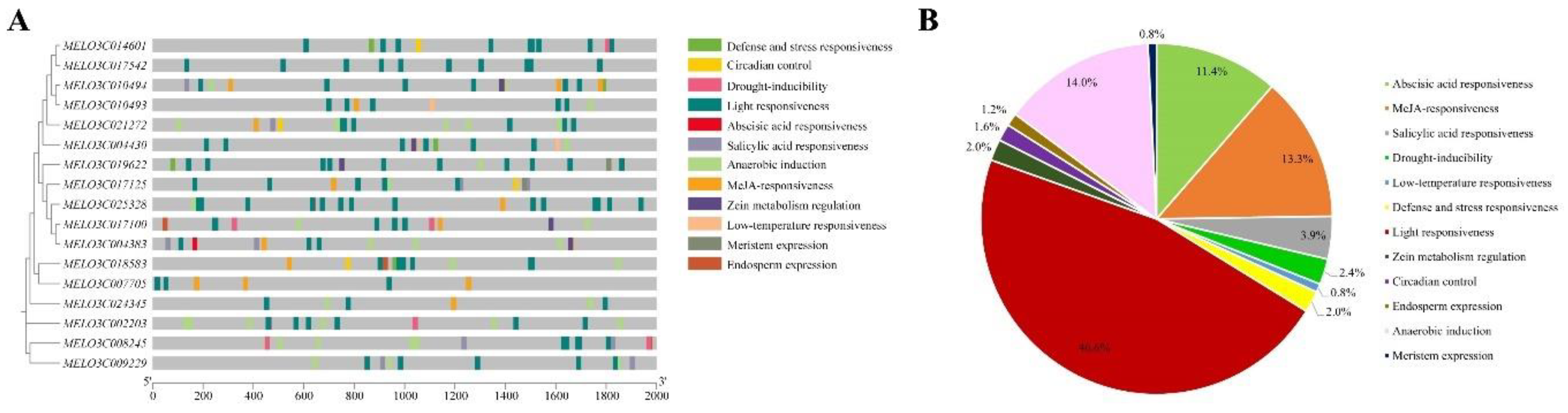
Figure 6.
Expression analysis of melon ALDH genes in different tissue and developmental stages.
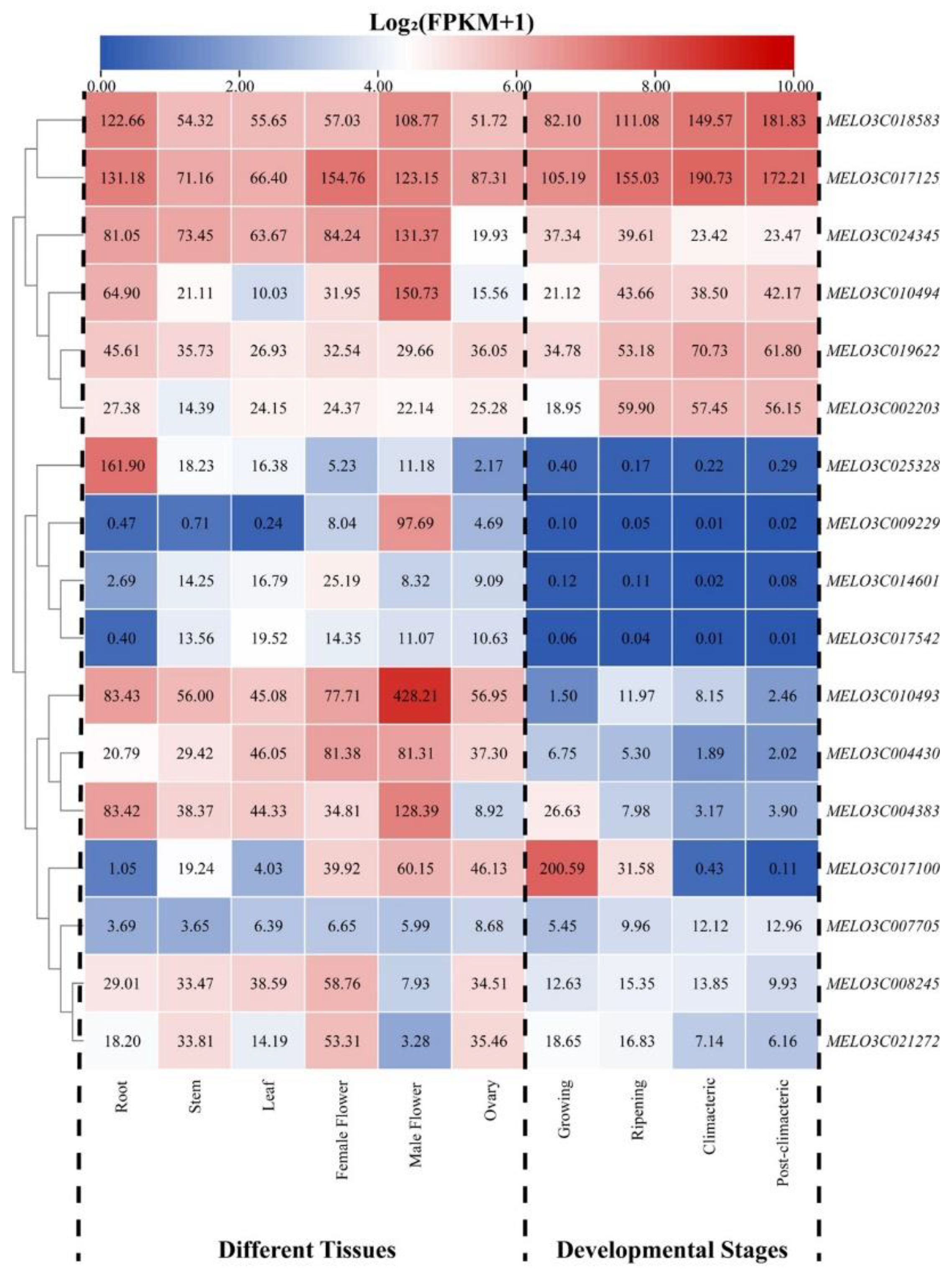
Figure 7.
The expression patterns of melon ALDH genes in response to abiotic stresses. (A)The expression heatmap of melon ALDH genes in response to salt stress. S: susceptible plant; R: resistant plant; CT: control treatment; Salt: salt treatment. (B)The expression heatmap of melon ALDH genes in response to chilling stress. S: susceptible plant; R: resistant plant; 0h, 8h and 72h were treatment for 0h, 8h and 72 hours. (C)The expression heatmap of melon ALDH genes in response to waterlogging stress. 0h, 6h, 24h, 48h and 72h were treatment for 0h, 6h, 24h, 48h and 72h hours. The data in the left expression heatmaps were the original FPKM values; the data in the right boxes were log2 (fold change) values highlighted by red (upregulation) and green (downregulation) colors.
Figure 7.
The expression patterns of melon ALDH genes in response to abiotic stresses. (A)The expression heatmap of melon ALDH genes in response to salt stress. S: susceptible plant; R: resistant plant; CT: control treatment; Salt: salt treatment. (B)The expression heatmap of melon ALDH genes in response to chilling stress. S: susceptible plant; R: resistant plant; 0h, 8h and 72h were treatment for 0h, 8h and 72 hours. (C)The expression heatmap of melon ALDH genes in response to waterlogging stress. 0h, 6h, 24h, 48h and 72h were treatment for 0h, 6h, 24h, 48h and 72h hours. The data in the left expression heatmaps were the original FPKM values; the data in the right boxes were log2 (fold change) values highlighted by red (upregulation) and green (downregulation) colors.
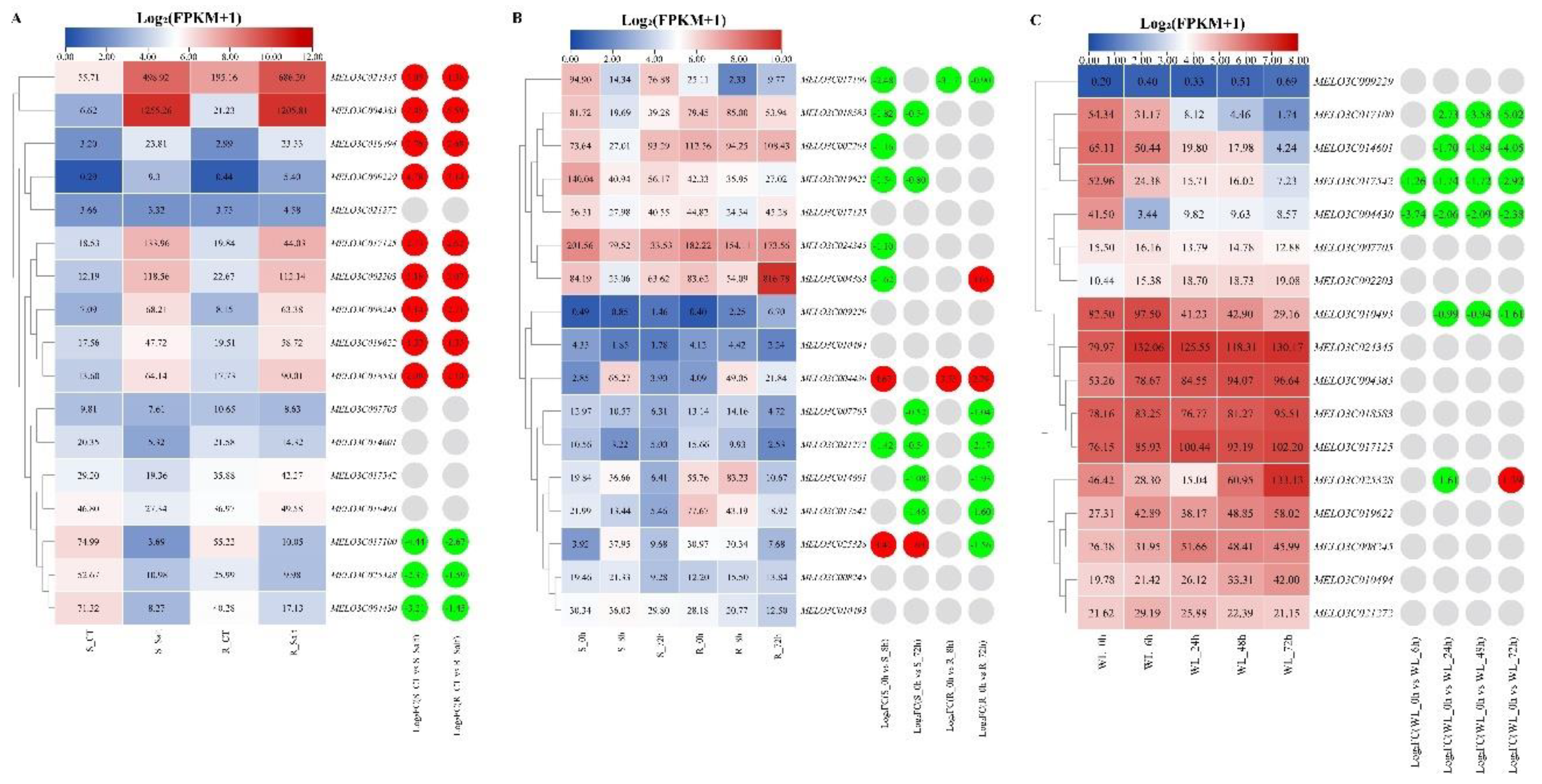
Figure 8.
The expression patterns of melon ALDH genes in response to biotic stresses. (A)The expression heatmap of melon ALDH genes in response to downy mildew stress. S: susceptible plant; R: resistant plant; 0h, 24h, 72h and 168h were 0, 24, 72, and 168 hours post-inoculation, respectively. (B)The expression heatmap of melon ALDH genes in response to Fusarium will stress. S: susceptible plant; R: resistant plant; 0h, 12h, and 72h were 0, 12, and 72 hours post-inoculation, respectively. (C)The expression heatmap of melon ALDH genes in response to gummy stem blight stress. S: susceptible plant; R: resistant plant; 0 day and 2days were 0, and 2 days post-inoculation, respectively. The data in the left expression heatmaps were the original FPKM values; the data in the right boxes were log2 (fold change) values highlighted by red (upregulation) and green (downregulation) colors.
Figure 8.
The expression patterns of melon ALDH genes in response to biotic stresses. (A)The expression heatmap of melon ALDH genes in response to downy mildew stress. S: susceptible plant; R: resistant plant; 0h, 24h, 72h and 168h were 0, 24, 72, and 168 hours post-inoculation, respectively. (B)The expression heatmap of melon ALDH genes in response to Fusarium will stress. S: susceptible plant; R: resistant plant; 0h, 12h, and 72h were 0, 12, and 72 hours post-inoculation, respectively. (C)The expression heatmap of melon ALDH genes in response to gummy stem blight stress. S: susceptible plant; R: resistant plant; 0 day and 2days were 0, and 2 days post-inoculation, respectively. The data in the left expression heatmaps were the original FPKM values; the data in the right boxes were log2 (fold change) values highlighted by red (upregulation) and green (downregulation) colors.
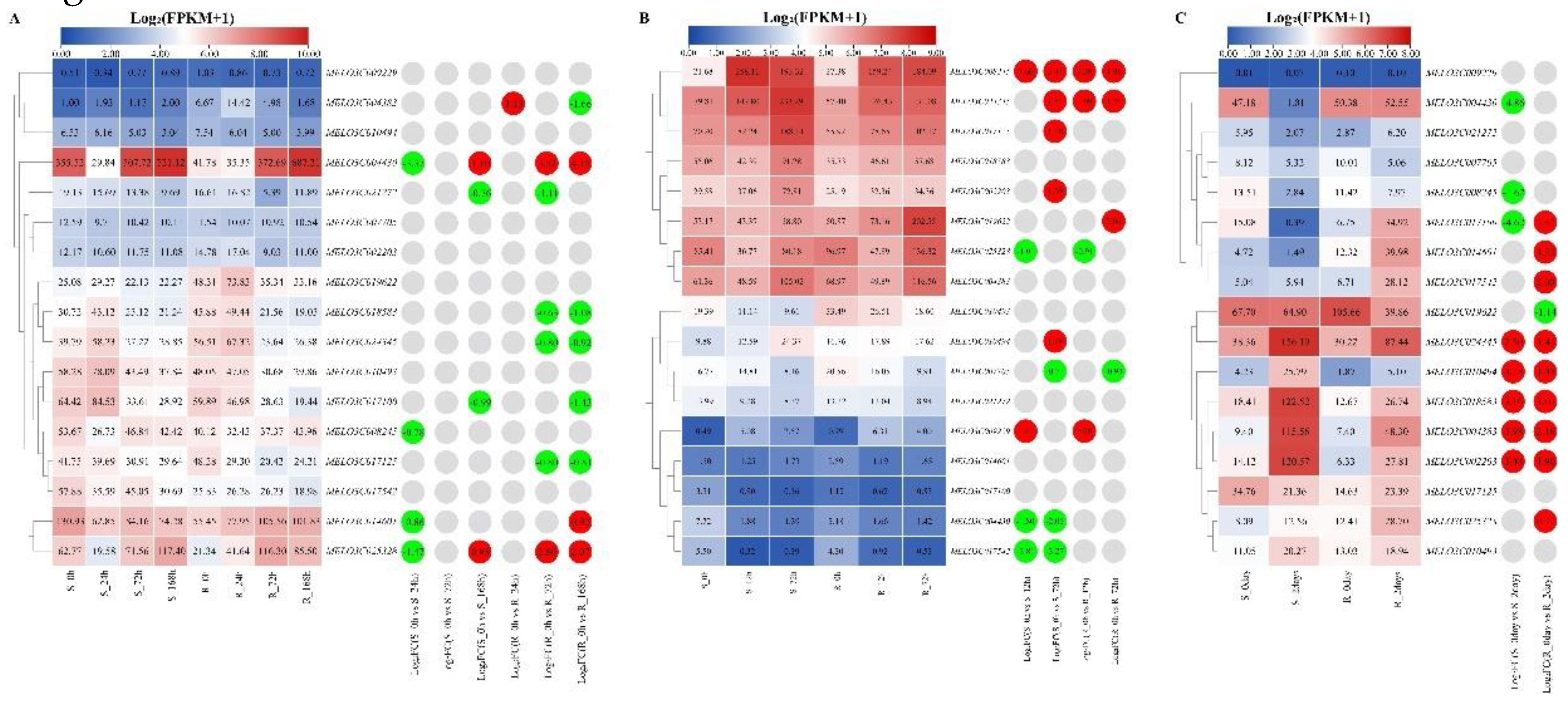
Figure 9.
An expression pattern heatmap of the melon ALDH genes under abiotic and biotic stresses. The gray color represented unchanged expression, red represented upregulated expression, green represented downregulated expression, and blue represented both upregulated and downregulated expression.
Figure 9.
An expression pattern heatmap of the melon ALDH genes under abiotic and biotic stresses. The gray color represented unchanged expression, red represented upregulated expression, green represented downregulated expression, and blue represented both upregulated and downregulated expression.
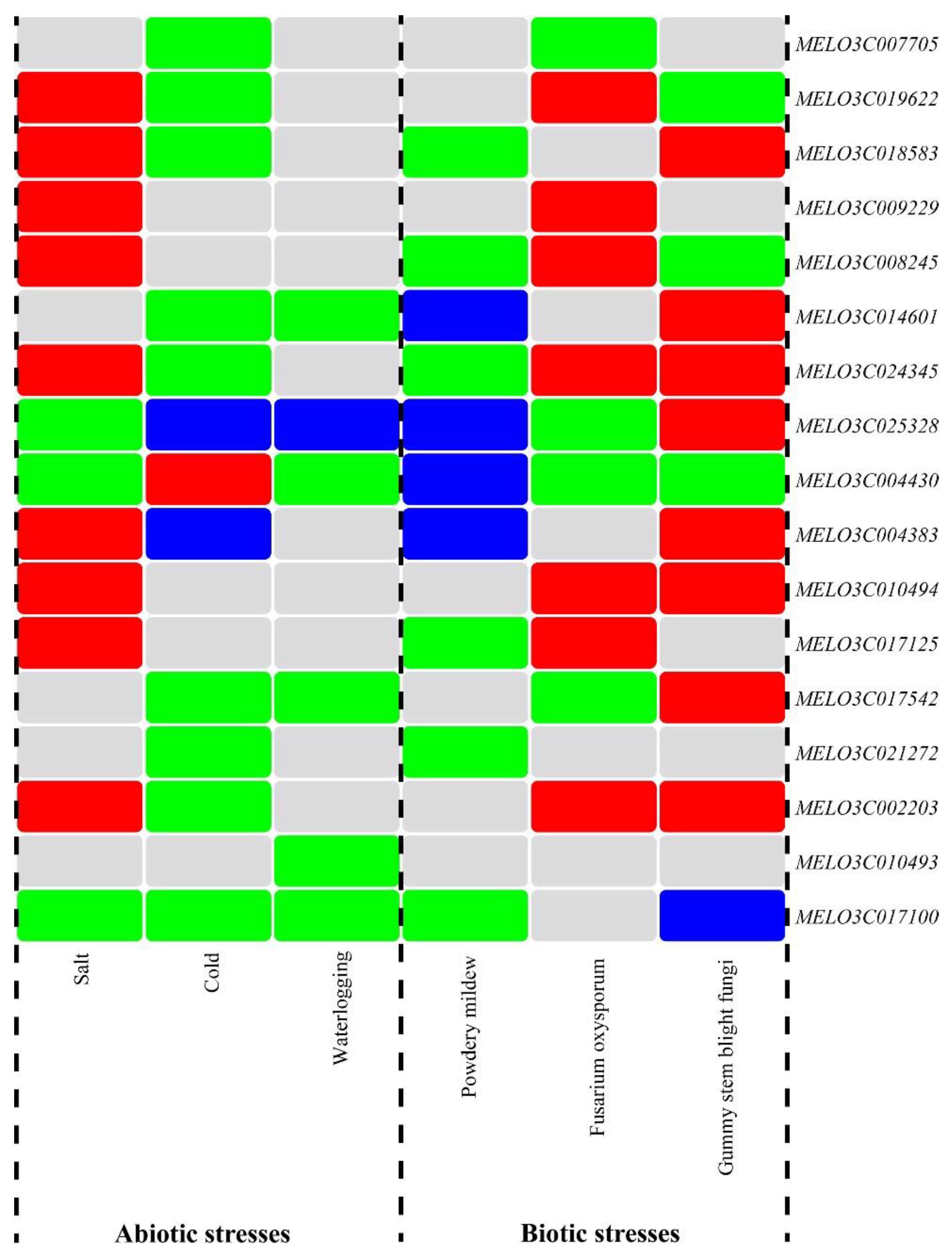
Figure 10.
Relative expression level of 10 CmALDH genes in response to salt treatments. Error bars are standard deviations of three biological replicates. Asterisks were used to indicate the significant degree of the expression level compared to the value of the control (*P < 0.05, **P < 0.01, ***P < 0.001).
Figure 10.
Relative expression level of 10 CmALDH genes in response to salt treatments. Error bars are standard deviations of three biological replicates. Asterisks were used to indicate the significant degree of the expression level compared to the value of the control (*P < 0.05, **P < 0.01, ***P < 0.001).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



