
Submitted:
30 September 2024
Posted:
03 October 2024
You are already at the latest version
Abstract
Fused in sarcoma (FUS) is involved in the formation of nuclear biomolecular condensates associated with poly(ADP-ribose) [PAR] synthesis catalyzed by a DNA damage sensor such as PARP1. Here, we studied FUS microphases’ separation induced by poly(ADP-ribosyl)ated PARP1WT [PAR-PARP1WT] or its mutants PARP1Y986S and PARP1Y986H respectively synthesizing (short PAR)-PARP1Y986S or (short hyperbranched PAR)-PARP1Y986H using dynamic light scattering, fluorescence microscopy, turbidity assays, and atomic force microscopy. We observed that biologically relevant cations such as Mg2+, Ca2+, or Mn2+ or polyamines (spermine4+ or spermidine3+) were essential for the assembly of FUS with PAR-PARP1WT and FUS with PAR-PARP1Y986S in vitro. We estimated the range of the FUS-to-PAR-PARP1 molar ratio and the cation concentration that are favorable for the stability of the protein’s microphase-separated state. We also found that FUS microphase separation induced by PAR-PARP1Y986H (i.e., a PARP1 mutant attaching short hyperbranched PAR to itself) can occur in the absence of cations. The dependence of PAR-PARP1–induced FUS microphase separation on cations and on branching of PAR structure points to a potential role of the latter in the regulation of formation of FUS-related biological condensates and requires further investigation.
Keywords:
FUS
; poly(ADP-ribosyl)ated PARP1
; mutant PARP1(Y986H)
; mutant PARP1(Y986S)
; microphase separation
1. Introduction
Fused in sarcoma (FUS, also known as translocated in liposarcoma [TLS]), along with Ewing’s sarcoma (EWS) and TATA-binding-protein–associated factor 15 (TAF15), is a member of the highly conserved FET family of RNA-binding proteins that regulate mRNA metabolism and may be involved in DNA repair and participate in the organization of biomolecular condensates in vivo [1,2,3]. FUS belongs to the class of intrinsically disordered proteins and contains an N-terminal low-complexity domain (LCD, also called the prion-like domain), three intrinsically disordered arginine/glycine/glycine (RGG)-rich regions, a conserved RNA recognition motif (RRM), a zinc finger (ZnF) motif, and a proline-tyrosine nuclear localization signal (PY-NLS) at the C terminus [4,5]. The LCD domain and RGG-rich regions have been shown to promote FUS phase separation that can be achieved through FUS self-assembly and/or binding to other proteins and/or nucleic acids [6,7]. For a long time, FUS has been considered a regulator of RNA metabolism [8]; nevertheless, more and more studies are revealing direct involvement of FUS in DNA repair [2,9,10]. In this regard, DNA damage sensors such as nuclear PARPs (PARP1 and -2: members of the ADP-ribosyltransferase diphtheria toxin-like [ARTD] family) can mediate FUS participation in DNA repair events [11,12,13,14,15]. Nuclear PARP1 and PARP2 recognize damaged DNA—in particular DNA strand breaks caused by genotoxic agents (free radicals, ionizing radiation, or alkylating agents) or by activities of repair enzymes—and catalyze the synthesis of poly(ADP-ribose) [PAR] covalently attached to proteins, mainly PARPs themselves, using NAD+ as a substrate [11,16]. Thus, PAR can be considered a nucleic-acid-like polymer that shares several features with single-stranded RNA or DNA, although PAR has mostly branched-chain structure and “star” shape [17,18]. PARP1-dependent PAR synthesis has been shown to promote the recruitment of repair enzymes and other factors to DNA lesions, thereby accelerating the repair process [19,20,21,22]. It has been suggested that PAR serves as a “seed” for promotion of FUS condensation near DNA damage [7,14,15,23,24,25].
In vitro, the recruitment of FUS to damaged DNA via binding to poly(ADP-ribosyl)ated [PARylated] PARP1 leads to the formation of large supramolecular assemblies containing damaged DNA, PARylated PARP1 (hereafter: PAR-PARP1), and FUS [15]. Moreover, FUS has been reported to form liquid-like condensates with short homogeneous and long heterogeneous protein-free PAR [7,26,27,28].
Although the FUS assembly and liquid-like phase separation (LLPS) in the presence of PAR have been actively studied in vitro [14,26,27,28], the features of FUS phase separation in the presence of PARylated PARPs are still not well understood due to the limited knowledge about the effect of cations. It has been found that both the assembly of multimolecular compartments containing FUS, PAR-PARP1, and damaged DNA and FUS-dependent stabilization of PARylated-PARP1–DNA condensates take place in the presence of Mg2+ [15,29]. Currently, physiologically relevant metal cations such as Mg2+ and Ca2+ are regarded as regulators of LLPS of both proteins and nucleic acids [30,31]; in particular, divalent cations (Mg2+ or Ca2+) cause condensation–compaction and microphase separation of protein-free PAR in vitro [26,28,32,33]. Moreover, we have previously observed that such cations as Mg2+, Ca2+, Mn2+, spermidine3+ (Spd3+), or spermine4+ (Spn4+) induce liquid-like self-assembly of PAR-PARP1 into multimolecular associates [34,35,36].
In the present work, we attempted to elucidate how the ability of FUS to form higher-order assemblies can be implemented in the presence of PAR-PARP1. Effects of biologically relevant polyamines, of divalent cations, and of PAR structure were investigated. By dynamic light scattering (DLS), fluorescence microscopy, atomic force microscopy (AFM), and turbidity assays, we characterized the FUS microphase separation induced by wild-type PARylated PARP1 (PAR-PARP1WT) or its mutants PARP1Y986S and PARP1Y986H respectively synthesizing (short PAR)-PARP1Y986S and (short hyperbranched PAR)-PARP1Y986H. We then compared their properties as molecular “seeds” for the promotion of higher-order assembly of FUS. For this purpose, we conducted experiments under the conditions where FUS was primarily monomeric and its higher-order assembly was initiated by addition of PAR-PARP1. We found that PAR-PARP1WT and PAR-PARP1Y986S are not able to induce FUS microphase separation. PARP1’s cofactors such as Mg2+, Ca2+, or Mn2+ or polyamines (Spd3+ or Spn4+) were required for the assembly of FUS with PAR-PARP1WT and FUS with PAR-PARP1Y986S in vitro. The range of the FUS-to-PAR-PARP1 molar ratio and cation concentration were estimated that are favorable for the stability of the protein phase-separated state. We also found that the FUS microphase separation induced by PAR-PARP1Y986H (a PARP1 mutant synthesizing short hyperbranched PAR) can proceed in the absence of cations. The dependence of PAR-PARP1–induced FUS microphase separation on cations and branched PAR structure points to a potential role of the latter in the regulation of formation of FUS-related biological condensates and requires further research.
2. Results
2.1. Cations Promote the Assembly of FUS into Higher-Order Structures in the Presence of PAR-PARP1
Although the mechanism that drives PAR-dependent FUS condensation is well-studied [7,14,26,27,28], little is known regarding the effects of PAR-PARP1 on phase behavior of FUS. Earlier, by AFM, FUS has been shown to interact with PAR-PARP1, thereby leading to the formation of supramolecular complexes or multimolecular compartments containing FUS, PAR-PARP1, and damaged DNA [15]. A recent study has revealed that FUS stabilizes PARP1–DNA condensates, which otherwise dissociate after PARP1 PARylation [29]. In the cell, PAR mainly exists as a polymer covalently attached to proteins, mainly to PARP1 [37], and protein-free PAR can arise transiently via hydrolysis of PAR attached to proteins [38]. Therefore, to assess biological relevance of this phenomenon, here we tested the propensity of FUS for liquid-like assembly under the conditions where negatively charged poly(ADP-ribose) is completely attached to PARP1, i.e., using PAR-PARP1. For this purpose, we employed DLS to monitor changes in hydrodynamic size of FUS in the presence of various concentrations of PAR-PARP1. For these experiments, we used FUS concentrations of 5–10 μM and PARP1 at ~2 μM, which are close to their physiological concentrations [7,39], and try to found the PAR-PARP1 concentration that promotes FUS higher-order assembly (Figure 1). We observed that PAR-PARP1 was not able to induce the assembly of FUS into higher-order structures or protein-rich microphases even at a high FUS-to-PARP1 molar ratio, i.e., when the FUS concentration was one to two orders of magnitude higher than the PAR-PARP1 concentration (Figure 1).
Only small particles with a hydrodynamic radius of 12–19 nm were detectable under such conditions (Figure 1). The size of these particles was close to the size of PAR-PARP1 (Figure 1).
In our previous work, we noticed that aggregates containing FUS, PAR-PARP1, and damaged DNA arise in the presence of Mg2+ [15]; moreover, FUS is reported to stabilize PAR-PARP1–DNA condensates also in the presence of Mg2+ [29]. Therefore, we hypothesized that PAR-PARP1 could seed FUS liquid-like assembly in the presence of Mg2+. Given that the cations at low concentration do not trigger the self-assembly of PAR-PARP1 [36], FUS titration with increasing concentration of PAR-PARP1 was performed at submillimolar concentration of Mg2+ or other biologically relevant cations such as Ca2+, Mn2+, Spd3+, or Spn4+ (Figure 2a–e). Indeed, the automodified PARP1 promoted liquid-like assembly of FUS in the presence of the cations, as evidenced by the emergence of large particles with a radius of 540–834 nm (Figure 2 and Figure S1a–e).
The titration of FUS indicated that FUS higher-order assemblies arose at FUS/PAR-PARP1 molar ratios in the range of 200:1 to 50:1 (Figure 2a–e). Thus, only substoichiometric levels of PAR-PARP1 (at concentration 200- or 50-fold below that of FUS) promoted the protein assembly, because after the increase of PAR-PARP1 concentration, the assembly disappeared (Figure 2 and Figure S1a–e). Similar re-entrant phase behavior of FUS with protein-free PAR, but not with PAR-PARP1, has been reported elsewhere [26,27]. Although FUS–PAR condensates are stable within a certain concentration range of PAR, they dissolve at a high PAR concentration [26,27]. Accordingly, the formation of relatively stable PAR-PARP1-FUS supramolecular assemblies was detectable at a high FUS-to-PAR-PARP1 molar ratio and after the addition of cations.
Both FUS and PARP1 are abundant cellular proteins, and their intracellular concentration has been estimated as 2–8 μM for FUS and 2.0 μM for PARP1 in HeLa cells [7,39]. In our system, we also tested whether the presence of divalent cations was sufficient to induce FUS assembly when PARP1 was added at a micromolar concentration similar to that of FUS, which was within the concentrations found in the cells. By DLS measurements, we analyzed the phase behavior of the proteins within a low FUS-to-PAR-PARP1 molar range (3:1 to 1:1) and after increasing concentrations of Mg2+, Ca2+, Mn2+, Spn4+, or Spd3+ (Figure 3 and Figure S2). The DLS data suggested that the cations at millimolar concentration promoted the FUS assembly even when PAR-PARP1 concentration was close to that of FUS (Figure 3). Addition of EDTA, which chelates divalent cations Mg2+, Ca2+, and Mn2+ [40], to the FUS–PAR-PARP1 mixture resulted in assembly dissolution.
These data strongly support the idea that intermolecular interactions of FUS and PAR-PARP1 are stabilized by cations, even at a low FUS-to-PAR-PARP1 molar ratio that promotes disruption of FUS assemblies (Figure 2 and Figure 3). Consequently, we supposed that neutralization of negative charges on the PAR phosphate backbone by cations is key to FUS phase separation in the presence of PAR-PARP1.
Because these cations may promote the self-assembly of PAR-PARP1 even in the absence of FUS [36], two types of assembly may ensue: PAR-PARP1 or PAR-PARP1–FUS. Therefore, we tested by fluorescence microscopy whether FUS coassembles with PAR-PARP1. To this end, we utilized Alexa Fluor 488 (AF488)-labeled FUS and Cyanine 3 (Cy3)-labeled PARP1 and analyzed the formation of condensates at a low FUS-to-PARP1 molar ratio (4:1) to visualize both the labeled PARP1 and FUS (Figure 4). An optimal concentration of Mg2+ was selected to promote the phase separation (15 mM). Starting with the mixture of FUS and PARP1 at micromolar concentration (Figure 3a), we added NAD+, which resulted in the formation of small droplets after 30 min incubation of the protein mixture (Figure 3b). Subsequent incubation of the mixture gave rise to larger assemblies, as evidenced by fluorescence microcopy (Figure 3c). For instance, the microscopy of the PARylated Cy3-PARP1 and AF488-FUS showed overlapping green (AF488) and red (Cy3) fluorescence signals, indicating simultaneous presence of the two proteins in the assemblies, i.e., FUS–PAR-PARP1 coassembly in vitro (Figure 3b,c).
Overall, these experiments suggested that FUS coassembly with PAR-PARP1 can be influenced by divalent cations or polyamines. Therefore, we were able to vary phase separation of FUS either by decreasing PAR-PARP1 concentration or by adding cations that stabilize the condensates (Figure 2 and Figure 3). On the basis of these results, we next studied the impact of PAR structure on the assembly formation in the PAR-PARP1–FUS system.
2.2. Frequency of Branching of PAR Influences the Assembly of FUS into Higher-Order Structures in the Presence of PAR-PARP1
PAR produced by PARP1 is a polymer with branching structure and may contain up to 200 ADP-ribose units with branch points (~1%) occurring approximately every 20–50 units [17,41,42]. Taking into account that protein-free PAR’s length influences FUS condensation [26], we hypothesized that the changing of PAR structure, namely, formation of short, long, and/or hyperbranched polymers—when the polymer is covalently attached to PARP1—may also modulate FUS phase separation. To clarify the influence of PAR structure on FUS’s phase behavior, we used point mutants of PARP1 that are a less active than the wild-type enzyme and synthesize short (PARP1Y986S) or short hyperbranched (PARP1Y986H) PAR that causes alterations in the morphology of PAR-PARP1 molecules [42,43,44]. Under our experimental conditions, the PARP1Y986S mutant manifested 15% residual activity relative to PARP1WT, whereas the PARP1Y986H mutant showed 50% activity (Figure S3).
On the one hand, it has been found that protein-free PAR contributes to phase separation of FUS by acting as a molecular “seed” that promotes protein assembly [7,14,26,27]. On the other hand, PAR readily disrupts FUS assemblies when present in a large molar excess over FUS [26,27]. Accordingly, a FUS–PAR system involves phase transition of FUS from one phase to two phases and back to one phase in response to an increase of PAR concentration. This observation suggested that the less effective PAR synthesis by the PARP1 mutants may contribute to the stability of higher-order assembly of FUS in the presence of PAR-PARP1. Therefore, we tested FUS assembly in the presence of PAR- PARP1Y986H or PAR-PARP1Y986S. As in the case of PARP1WT (Figure 1), PARylation of either PARP1Y986H or PARP1Y986S did not trigger their self-assembly (Figure S4). Then, we again employed DLS to monitor changes in the hydrodynamic size of FUS at a high FUS-to-PAR-PARP1Y986H (or FUS-to-PAR-PARP1Y986S) molar ratio and submillimolar concentration of Mg2+ (Figure 5a,b and Figure S5a,b).
In the case of PARPY986S (producing short PAR), higher-order assemblies were observed at a high FUS-to-PAR-PARP1Y986S molar ratio (145:1 to 75:1) (Figure 5a and Figure S5a). Thus, the FUS–PARP1Y986S assembly was stable within a molar ratio that was close to the ratio observed with PARP1WT (Figure 2a and Figure 5a). The results indicated that the less extensive PARP1 autoPARylation and biosynthesis of shorter PAR only slightly affect the FUS-to-PARP1 molar ratio that favors higher-order assembly of FUS with PAR-PARP1.
At the same time, FUS possesses a strong ability to form higher-order assemblies (Rh > 200 nm) with PAR-PARP1Y986H, which produces a hyperbranched polymer (Figure 5b and Figure S5b). In fact, FUS retained the capacity for microphase separation in a broad range of protein-to-PAR-PARP1Y986H molar ratios (200:1 to 10:1), whereas in the cases of PAR-PARP1WT and PAR-PARP1Y986S, large particles were detected in a narrower range of the FUS-to-PARP1 molar ratio (200:1.0 to 50:1.00) (Figure 2b and Figure 5b). Our results also revealed that the PAR-PARP1Y986H mutant (synthesizing hyperbranched PAR) is able to induce FUS assembly even in the absence of cations (Figure 5c and Figure S5c).
Similar to PARP1WT, these mutants could also promote FUS higher-order assembly at a low FUS-to-PARP1 molar ratio in the presence of Mg2+ when large particles with Rh (~267 nm) and Rh (~103 nm) were detectable (Figure 5d and Figure S5d). Consequently, FUS’s microphase-separated state strongly depends on the FUS-to-PAR-PARP1 molar ratio, which differed between the PARP1 mutants producing PAR of different lengths and branching modes. Nevertheless, cations (Mg2+) were still needed to induce the assembly at a low FUS-to-PAR-PARP1Y986H molar ratio (Figure 5d and Figure S5d).
To confirm that the PAR-PARP1–FUS assembly is affected by the structure of PAR, we tested whether the cation concentration (in particular Mg2+) needed to induce FUS microphase separation differed between PARPWT, PARP1Y986S, and PARP1Y986H (Figure 6, Figures S6 and S7). For this purpose, we compared the assembly of FUS with PAR-PARP1WT or its mutant in the presence of different concentrations of Mg2+ by measuring turbidity, which correlates with protein phase separation [45] (Figure 6, Figures S6 and S7). It was observed that at a low FUS/PAR-PARP1 molar ratio (~1:1), already 9 mM Mg2+ can promote microphase separation after mixing of FUS and (highly branched PAR)-PARP1Y986H (Figure 6). In contrast to PARP1Y986H, Mg2+ concentration higher than 11.3 mM was required for inducing FUS microphase separation in the presence of either PAR-PARPWT or PAR-PARP1Y986S (Figure 6). Thus, the biosynthesis of highly branching PAR during PARP1 automodification required lower Mg2+ concentrations to promote FUS phase separation. We also tested the effect of 1,6-hexanediol: a compound widely used to disrupt weak protein–protein hydrophobic interactions underlying FUS LLPS [46].
A significant decrease in the turbidity was observed after the addition of 1,6-hexanediol to a PAR-PARP1–FUS solution, suggesting that nonionic interactions play an important part in this Mg2+-dependent assembly of FUS and PAR-PARP1 molecules. For a FUS–PARP1Y986H mixture, addition of 2.8% of 1,6-hexanediol diminished the turbidity twofold, suggesting that 1,6-hexanediol is efficient in disrupting the microphases (Figure 6). On the other hand, addition of 1,6-hexanediol up to 15% to a FUS-and-PAR-PARP1WT or FUS-and-PAR-PARP1Y986S solution respectively gave only a 17% and 13% reduction in turbidity (Figure 6).
Therefore, in contrast to FUS–PAR-PARP1WT or FUS–PAR-PARP1Y986S, the FUS–PAR-PARP1Y896H assembly has higher susceptibility to dissolution by 1,6-hexanediol, indicating that this microphase-separated state depends to a lesser extent on the presence of a cation decreasing the involvement of electrostatic interactions. Accordingly, hydrophobic interactions notably contribute to FUS–PAR-PARP1Y896H microphase separation. Although a noticeable reduction in turbidity by 13–50% for PAR-PARP1WT and its mutants was registered at concentrations of 2.8% to 15% 1,6-hexanediol, the reagent was not able to completely disrupt FUS–PAR-PARP1 assemblies coming into being in the presence of Mg2+.
Only the addition of EDTA readily disrupted the assemblies, and the turbidity of the solution returned to a value similar to that of the solution with 4 mM Mg2+ (Figure 6).
These results indicated that the assembly of FUS with PAR-PARP1 is indeed a mainly cation-regulated process; however, hydrophobic interactions sensitive to 1,6-hexanediol treatment also help to stabilize the condensates.
As reported elsewhere, mutations Y986S or Y986H in PARP1 alter both overall protein activity and PAR chain structure [42,43,44]. Consequently, these mutations could influence the size of PAR-PARP1–FUS assemblies. To clarify this issue, we visualized a mixture of either PAR-PARP1WT or PARP1Y986H with FUS by AFM (Figure 7). To evaluate the size of PAR-PARP1–FUS assemblies in the presence of Mg2+, first, AFM imaging of PARP1WT and PARP1Y986H was undertaken after incubation with a gapped pBR322 (pBR) plasmid in the presence of NAD+ (Figure 7).
As for PARP1WT, the PARylated molecules had a size (diameter) up to 52 nm, whereas in the case of PARP1Y986H, the size was only ≤40 nm (Figure 7b). Consistently with our previous AFM data [44], modified molecules of PARP1Y986H synthesizing hyperbranched PAR were found to carry highly packed polymer chains in contrast to PARP1WT (Figure 7). The activation of PARP1WT and its mutants as detected by AFM enabled us to measure the average size of the assemblies engendered by PARylated proteins after the addition of FUS (Figure 7). We noticed that a decrease in the size of PAR-PARP1Y986H was accompanied by a diminishing size of the assemblies that were formed by FUS in the presence of PAR-PARP1Y986H (Figure 7).
Consequently, the structure of PAR, mainly its branching frequency, is an important factor for FUS higher-order assembly with PAR-PARP1 and influences the size of assemblies and the cation concentration required for the stabilization of the FUS microphase separation process.
3. Discussion
During the past decade, functions of RNA-binding proteins in the DNA damage response have been revised due to the discovery of biomolecular condensates generated via protein liquid–liquid phase separation [47,48]. From this point of view, the accumulation of repair proteins at DNA damage sites and the formation of DNA repair foci may be associated with condensation [7,13,14,15]. PAR and FET family proteins (FUS, EWS, and TAF15) seem to play a specific role in the formation of DNA damage–induced condensates because PAR contributes to the recruitment of FET proteins to a DNA damage site and also promotes their LLPS [7,12,13,14,15]. Since the discovery of the PAR-dependent involvement of FUS in DNA repair in 2013 [12], there has been a substantial increase in the number of articles revealing the direct interactions between FUS and PARP1 and PAR [7,13,14,15,49,50]. Indeed, the emergence of FUS-rich assemblies in DNA damage regions after laser microirradiation is observed depending on PARP1 activation [7,13,14]. Moreover, FUS has been shown to interact with PAR-PARP1 [49,50] and to regulate PARP1-dependent PAR synthesis in HeLa cells after genotoxic stress [50] and possibly directly participates in the formation of repairosome compartments [14,15,29]. In vitro, FUS has been found to interact with protein-free PAR, and FUS’s binding to PAR promotes LLPS of FUS and possibly its aggregation [7,14,17,26,27,28].
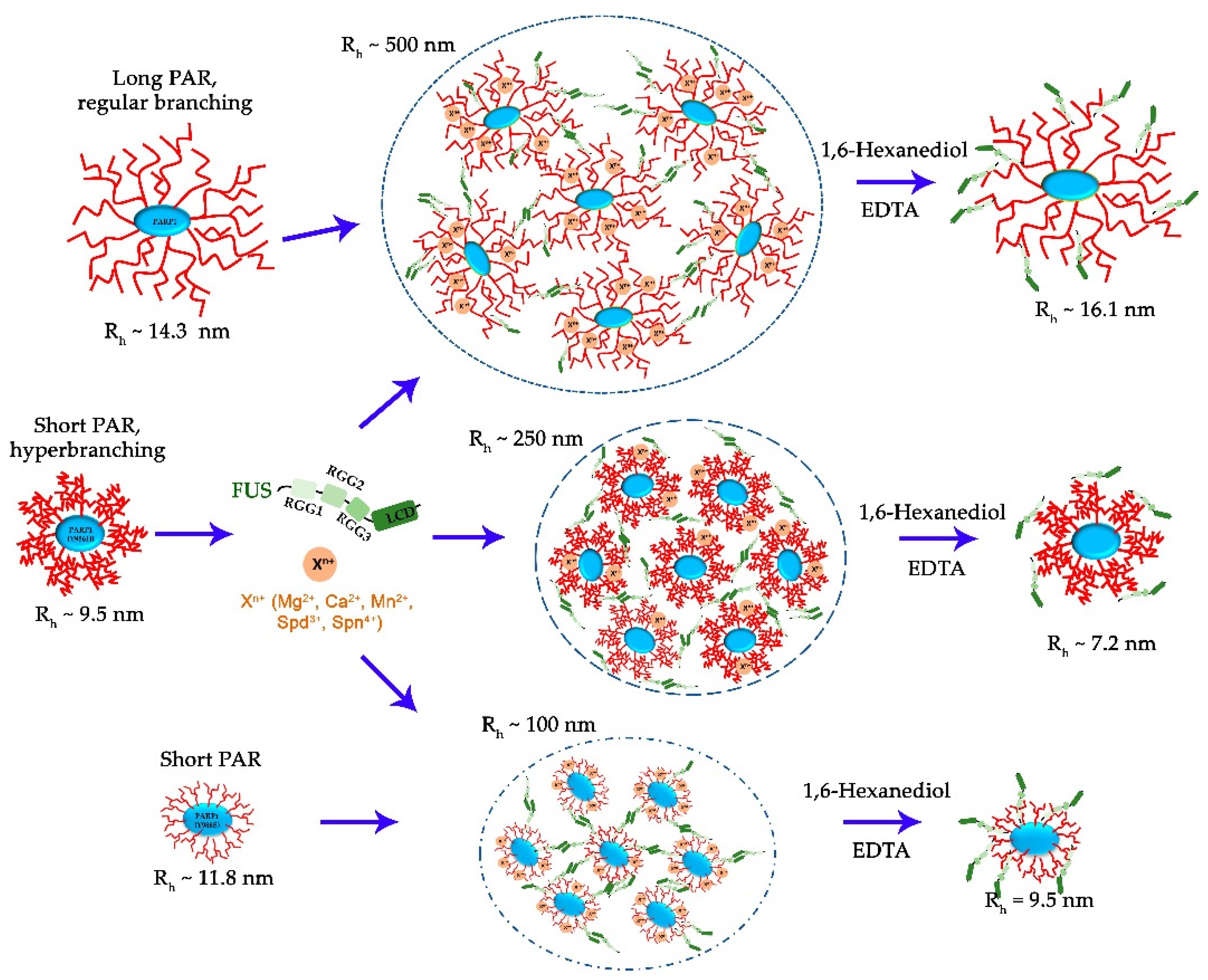
In the current study, we employed a simple in vitro model of FUS phase separation that allows us to identify functional features of the interaction between FUS and PAR-PARP1 that lead to FUS microphase separation. Both free and protein-associated PAR can have similar effects on the seeding of FUS phase separation: a low PAR concentration promotes LLPS of FUS, but when the PAR concentration exceeds a certain threshold, the condensate will begin to disintegrate [26,27]. In contrast to protein-free PAR, we failed to detect FUS assembly with PAR-PARP1 in the absence of cations; however, such bivalent cations as Mg2+, Ca2+, or Mn2+ or natural polyamines (Spd3+ or Spn4+) promoted the phase separation of FUS both at a high and a low FUS–to-PARP1 molar ratio (Figure 2 and Figure 3). In the presence of a submillimolar level of the cations (0.1–0.5 mM), PAR-PARP1 induced the assembly of FUS into higher-order structures in a narrow range of the FUS-to-PARP1 molar ratio (~200:1 to 50:1; Figure 2); whereas a millimolar cation concentration (2 mM Mn2+, 12.5 mM Mg2+, 7 mM Ca2+, 12 mM Spd3+, or 2.3 mM Spn4+) stabilized FUS–PAR-PARP1 assembly at a low FUS-to-PARP1 ratio (~3:1 to 1:1; Figure 3). Some authors have estimated that the physiological concentrations of FUS and PARP1 within HeLa cells are approximately 8.0–1.4 μM and 2.0 μM, respectively [7,39]; accordingly, under physiological conditions, PAR-PARP1 induces FUS phase separation only at its high local concentration. Nevertheless, such cations as Mg2+, Ca2+, Spd3+, or Spn4+ promoted FUS microphase separation with PAR-PARP1 even at a FUS-to-PARP1 molar ratio close to 1:1 (Figure 3 and Figure 6). Indeed, these cations are abundant in eukaryotic cells, which generally contain 1 mM polyamines, 17–20 mM Mg2+, and 0.1 µM to 1 mM Ca2+ [51,52,53]. Such cations as Mg2+, Ca2+, Mn2+, Spd3+, and Spn4+ play an important part in the regulation of many RNA- and DNA-dependent processes and can serve as cofactors of PARP1 [32,54,55,56,57,58]. Moreover, some of these cations (Mg2+, Ca2+, and Spn4+) are reported to regulate the formation of membraneless organelles, to modulate phase behavior of both intrinsically disordered proteins and RNA, DNA, and PAR, which are a key components of cellular protein condensates and aggregates [30,31,59]. Moreover, we have shown recently that cations are able to promote liquid-like self-assembly of PARylated PARP1 in the absence of FUS [36].
Here we demonstrated that the near-physiological concentration of the cations tested in our experiments has a strong impact on phase behavior of FUS in the presence of PAR-PARP1 in vitro. The present work also addresses effects of PAR structure on FUS phase separation. As mentioned above, protein-free PAR induces FUS LLPS at a low concentration but dissolution at a high concentration [26,27]. Furthermore, it was expected here that PAR-PARP1—similarly to protein-free PAR—would reversibly modulate microphase separation of FUS, but modified PARP1 was unable to trigger FUS assembly without cations (Figure 1). Given that the noncovalent FUS–PAR interaction and FUS assembly are tightly linked within the phase separation process [7,14,15,26,27,28], PAR structure, namely, length and branching frequency may be regarded as a key factor for the regulation of FUS phase separation [26,33]. To explore possible relations between FUS assembly and the PAR structure produced by PARP1, we also investigated the properties of PARP1 mutants in the “seeding” of higher-order assembly of FUS (Figure 5, Figure 6 and Figure 7). We found that the PARP1 mutations—that cause both a reduction in the magnitude of PARP1 autoPARylation and changes in the structure of PAR—strongly influence the FUS assembly; this effect was especially evident in the case of PARP1Y986H, which produces short hyperbranched PAR (Figure 5 and Figure 6). In the experiments with FUS and the PARP1 mutants, we noted that not the PAR length but the level of PAR branching is the main factor that influences the assembly of FUS into higher-order structures in the presence of PAR-PARP1 (Figure 5, Figure 6 and Figure 7). This finding implies that promotion of FUS microphase separation substantially depends on the structure of PAR because hyperbranched PAR is more effective than long or short PAR having regular or slightly enhanced branching, as in the case of PARP1WT and its PARP1Y986S mutant (Figure 2 and Figure 5). In the presence of Mg2+, (hyperbranched short PAR)-PARP1Y986H and (short PAR)-PARP1Y986S (or PAR-PARP1WT) can induce FUS microphase separation at a FUS-to-PARP1 molar ratio differing by a factor of five (Figure 2a and Figure 5a,b). Moreover, hyperbranched PAR-PARP1Y986H seeded FUS microphase separation in the absence of cations (Figure 5c). Thus, we believe that the PAR-induced assembly and disassembly of FUS higher-order structures are affected by PAR’s structural characteristics such as branching and length.
It has been demonstrated that both PAR branching frequency and chain length can affect noncovalent protein binding and PAR-dependent protein localization and undergo changes in nuclear structures within the cell [1,21,60,61]. Furthermore, PAR branching frequency is reported to vary during different phases of the DNA damage–induced PARylation reaction, implying biological relevance of PAR structure [1]. Taking into account that (hyperbranched PAR)-PARP1Y986H has an obvious preference for the seeding of FUS phase separation (Figure 5b), PAR structure may perform a pivotal function in the modulation of formation efficiency of FUS-dependent condensates associated with PARP1 activation at DNA damage sites.
This relation also opens up new opportunities for a better understanding of the mechanisms by which PAR-PARP1 and PAR structures can influence FUS phase separation and should elucidate the potential function of cations in the regulation of FUS-dependent condensate formation (Figure 8). Moreover, we recently showed that PAR-PARP1 undergoes self-assembly into supramolecular complexes in the presence of cations; besides, we observed that the self-assembly of PARP1 regulates the level of its automodifications and stimulates activities of poly(ADP-ribose)glycohydrolase to hydrolyze PAR and DNA polymerase β, a key DNA polymerase of base excision repair and DNA single-strand repair pathways [Sukhanova 2024]. At the same time, there are data about PAR-dependent organization of biomolecular condensates with the participation of FUS both in vitro and ex vivo [7,14,15,26,27]. Additional research will clarify whether FUS condensation and FUS’s interaction with PAR-PARP1 have functional consequences such as regulation of PARP1-dependent DNA repair pathways or activity of PAR-degrading enzymes.
4. Materials and Methods
4.1. Plasmid Construction, Proteins, and Reagents
Plasmid pET32a-hPARP1-His encoding cDNA of wild-type recombinant human PARP1 with a His tag is a kind gift from Dr. M. Satoh (Université Laval, Québec, Canada). pET32a-hPARP1-His was used to generate mutation Y986S or Y986H within the PARP1 coding sequence [44].
The recombinant wild-type PARP1 (PARP1WT) and mutants PARP1Y986S and PARP1Y986H were overexpressed in Escherichia coli (E. coli) Rosetta (DE3)pLysS (Novagen) and purified by means of Ni-NTA agarose (GE Healthcare, United States) affinity chromatography, HiTrap Heparin High Performance (GE Healthcare, United States) affinity chromatography, and deoxyribonucleic-acid−cellulose (single-stranded calf thymus DNA) (Sigma-Aldrich, United States) affinity chromatography as described before [44]. Recombinant FUS was expressed in E. coli strain BL21(DE3) and purified as described previously [15].
Sulfo-Cy3 NHS ester and AF488 NHS ester were purchased from Lumiprobe RUS Ltd. (Russia). Radioactive [α-32P]ATP was prepared in the Laboratory of Biotechnology at the ICBFM (SB RAS, Novosibirsk, Russia). Oligodeoxynucleotides were synthesized by the Laboratory of Biomedical Chemistry (ICBFM SB RAS, Novosibirsk, Russia). NAD+ and β-nicotinamide mononucleotide were purchased from Sigma-Aldrich (United States), and the pBR plasmid from New England BioLabs (United States). Reagents for buffers and electrophoresis components were bought from Sigma-Aldrich (United States), PanReacAppliChem (Germany) (acrylamide/bis-acrylamide and urea), Molecular Group (dithiothreitol [DTT]), and Merk (NaCl), whereas protein molecular weight markers from ThermoScientific (Lithuania). Olaparib (AZD2281, Ku-0059436) was purchased from Apexbio Technology (United States).
4.2. Preparation of the DNA Duplex and Damaged Plasmid
A DNA duplex (30-mer with a one-nucleotide gap) (DNA-gap) was obtained via hybridization of an oligonucleotide (3′-cccaaccaaacgc g taagtgtcaagaggcg-5′) with complementary oligonucleotides (5′-OH-gggttggtttgcg-3′ and 5′-phosphate-attcacagttctccgc-3′) in a 1.0:1.5 ratio. The oligonucleotide mixture was incubated for 3 min at 95 °C and then slowly cooled to room temperature.
The pBR plasmid containing single-strand DNA breaks was prepared using heat and acid treatment to create abasic sites followed by apurinic/apyrimidinic-site cleavage by means of apurinic/apyrimidinic endonuclease 1 activity as described previously [15].
4.3. Preparation of PAR-PARP1
PAR-PARP1 was prepared in a reaction mixture (20 µL) consisting of 2.5 µM PARP1WT (or its mutant PARP1Y986S or PARP1Y986H), 2.0 µM DNA duplex, 200 mM NaCl, 300 mM urea, 25 mM HEPES-NaOH pH 7.5, and 1 mM DTT. Samples were equilibrated for 1 min, and then the PARP1 activation was initiated by the addition of NAD+ to a final concentration of 1 mM. The reaction mixtures were incubated at 30 °C for 40 min. The reactions were stopped by the addition of olaparib to a final concentration of 200 µM.
4.4. Hydrodynamic Size Measurement by DLS
DLS measurements were carried out in a low-volume quartz batch cuvette (ZEN 2112) using a Zetasizer Nano ZS instrument (Malvern Instruments Ltd., Malvern, UK) at 25 °C. All stock solutions of DNA and proteins were filtered through a polyethersulfone membrane (0.2 μm pore size). The measurements and data processing were performed as described elsewhere [34,36].
FUS, PARP1WT, PARP1Y986H, and PARP1Y986S hydrodynamic size assays were performed in reaction mixtures consisting of DLS buffer (25 mM HEPES-NaOH pH 7.5, 200 mM NaCl, 300 mM urea, and 1 mM DTT), 10 μM FUS, 2.5 µM PARP1WT, 2.5 µM PARP1Y986H or 2.5 µM PARP1Y986S as specified in figure legends. The samples were equilibrated at 30 ° for 1 min, and then Rh measurement was performed.
For analysis of PAR-PARP1WT, PAR-PARP1Y986H, or PAR-PARP1Y986S hydrodynamic size, 2.5 µM PARP1WT, PARP1Y986H, or PARP1Y986S was incubated with 2.5 µM DNA-gap in DLS buffer. Samples were equilibrated for 1 min, after which PARP1 activation was initiated by the addition of NAD+ to a final concentration of 1 mM. The reaction mixtures were incubated at 30 °C for 40 min, and next Rh measurement was performed.
For analysis of hydrodynamic size at a high FUS-to-PARP1 molar ratio, 10 µM FUS was mixed with 50–650 nM PAR-PARP1WT, PARP1Y986S, or PARP1Y986H in DLS buffer (25 mM HEPES-NaOH pH 7.5, 200 mM NaCl, 300 mM urea, and 1 mM DTT) in the absence or presence of 0.1 mM Mn2+, 0.1 mM Spn4+, 0.4 mM Spd3+, 0.5 mM Ca2+, or 0.5 mM Mg2+ as indicated in figure legends. The samples were equilibrated at 30 °C for 1 min, after which Rh was measured.
For the analysis of hydrodynamic size at a low FUS-to-PARP1 molar ratio, 1.0–1.5 µM PARPARP1 was mixed with 5–4 µM FUS in DLS buffer, the samples were equilibrated for 1 min, and Rh was measured. After that, the reactions were supplemented with Mn2+ up to 2 mM, Mg2+ up to 12.5 mM, Ca2+ up to 7 mM, Spd3+ up to 12 mM, or Spn4+ up to 2.3 mM. The samples were equilibrated at 30 °C for 1 min, and Rh was measured again.
4.5. Turbidity Measurements
For assessment of the turbidity of FUS solutions in the presence of PAR-PARP1, 1.8 µM FUS was incubated with 2 µM PAR-PARP1WT, PAR-PARP1Y986S, or PAR-PARP1Y986H in a buffer consisting of 100 mM NaCl, 20 mM HEPES-NaOH pH 7.5, and 300 mM urea at 30 °C for 5 min. After that, the reactions were supplemented with 4–16 mM Mg2+ as indicated in a figure legend, and an absorption spectrum of the solutions was recorded. Then, either 1,6-hexanediol (2.8–15.0%) or EDTA (30 mM) was introduced as indicated in a figure legend, and the absorption spectrum of the solutions was recorded again.
4.6. Fluorescent Labeling of FUS and PARP1 and Fluorescence Microscopy
For labeling of FUS, purified FUS (19 nmol, 4.6 mg/mL) was incubated with AF488 (45 nmol) for 1 h at 25 °C and then at 4 °C overnight in a buffer consisting of 6 mM HEPES-NaOH pH 7.5, 50 mM NaCl, 2.0 M urea, and 100 mM NaHCO3. The unreacted AF488 dye was removed via dialysis against a buffer composed of 20 mM HEPES-NaOH pH 7.5, 200 mM NaCl, 6 M urea, and 1 mM DTT followed by concentration of the AF488-labeled FUS (AF-FUS) in ultrafiltration spin columns. For labeling of PARP1, purified PARP1 (1.2 nmol, 3.0 mg/mL) was incubated with Cy3 (13 nmol) for 1 h at 25 °C and then at 4 °C overnight in a buffer consisting of 50 mM HEPES-NaOH pH 7.5, 400 mM NaCl, and 2 mM tris(2-carboxyethyl)phosphine) (TCEP). The unreacted Cy3 dye was removed via dialysis against a buffer composed of 50 mM HEPES-NaOH pH 7.5, 200 mM NaCl, and 1 mM TCEP followed by concentration of the Cy3-labeled PARP1 (Cy3-PARP1) in ultrafiltration spin columns.
Concentrations of the conjugates AF488-FUS and Cy3-PARP1 and the degree of labeling (DOL) were determined using the following extinction coefficients: ϵ280 = 70,390 M−1cm−1 for FUS, ϵ280 = 120,000 M−1cm−1 for PARP1, ϵ555 = 150,000 M−1cm−1 and a correction factor (CF280) of 0.09 for Cy3, and ϵ495 = 71,800 M−1cm−1 and CF280 = 0.1 for AF488. The DOL was calculated via the formula
where ɛprot and ɛmax are the molar extinction coefficients of the protein and dye, respectively; Amax is at the absorption maximum of the dye; and A280 is absorbance of the protein at 280 nm.
DOL = Amax × ɛprot/((A280 − Amax × CF280) × ɛ280),
The DOL was estimated at 12% for FUS and 90% for PARP1. The extinction coefficients (ɛprot) of FUS and PARP1 were based on Expasy Protparam data, and the extinction coefficients (ɛmax) for AF488 and Cy3 were taken from Lumiprobe protocols.
4.7. Preparation of Samples for AFM Experiments and Image Analysis
Complexes of PAR-PARP1 and FUS for AFM were generated in reaction mixtures (20 μL) composed of 12.5 mM HEPES-NaOH pH 8.0, 12.5 mM NaCl, 1 mM DTT, 5 mM MgCl2, 100 mM urea, either 30 nM PARP1WT or PARP1Y986H, and 12.5 nM damaged pBR. The samples were equilibrated for 1 min, and then the PARP1 activation was initiated by the addition of NAD+ to a final concentration of 0.3 mM, with incubation at 37 °C for 5 min, after which FUS was introduced at a final concentration of 400 nM followed by incubation at 37 °C for 1 min. Next, the samples were diluted 10-fold with a buffer (12.5 mM HEPES-NaOH pH 8.0, 12.5 mM NaCl, and 1 mM DTT). To adsorb the molecules on mica, putrescine was added to the solution to a final concentration of 1 mM, after which a 10 μL droplet was deposited on the surface of freshly cleaved mica and processed and imaged as described before [15,18]. AFM images were captured in air using a Nanoscope V Multimode 8 (Bruker, Santa Barbara, CA) in PeakForce Tapping mode using Scanasyst-Air probes (Bruker). Continuous force–distance curves were thus recorded with an amplitude of 100–300 nm at low frequency (1–2 kHz). PeakForce Tapping mode decreases lateral and shear forces. Images were captured at 2048 × 2048 pixels at a line rate of 1.5 Hz.
The minimum diameter (nm) of the circle in which PARylated proteins or complexes could be enclosed was measured in AFM images with the help of the “section” tool in the Nanoscope Analysis software.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Typical volume-weighted size distributions for the mixture of FUS with PAR-PARP1WT in the presence of submillimolar concentration of cations; Figure S2: Typical volume-weighted size distributions for the mixture of FUS with PAR-PARP1WT in the presence of millimolar concentration of cations; Figure S3: PARylation of PARP1WT, PARP1Y986H and PARP1Y986S according to SDS-PAGE and phosphorimaging; Figure S4: Typical volume-weighted size distributions for PARP1Y986H, PAR-PARP1Y986H, PARP1Y986S and PAR-PARP1Y986S; Figure S5: Typical volume-weighted size distributions for FUS – PAR-PARP1Y986S and FUS – PAR-PARP1Y98H mixtures; Figure S6: Absorbance spectra of the mixtures consisting of FUS and PARylated PARP1WT, FUS and PARylated PARP1Y986H, FUS and PARylated PARP1Y986S in the presence of different concentration of Mg2. Figure S7: Absorbance spectra of the mixtures consisting of FUS and PARylated PARP1WT, FUS and PARylated PARP1Y986H, FUS and PARylated PARP1Y986S and FUS and PARP1WT in the presence of EDTA and different concentration of 1,6-hexanediol.
Author Contributions
Conceptualization, O.I.L. and M.V.S.; methodology, M.V.S., A.S.S., K.N.N., L.H. and R.O.A.; validation, R.O.A.., A.S.S., L.H. and M.V.S.; formal analysis, R.O.A., A.S.S., and M.V.S.; investigation, R.O.A., A.S.S., K.N.N., L.H. and M.V.S.; resources, O.I.L., D.P., and M.V.S.; data curation, R.O.A., A.S.S., and M.V.S.; writing—original draft preparation, M.V.S. and R.O.A.; writing—review and editing, O.I.L., D.P., and M.V.S.; visualization, R.O.A., A.S.S., and M.V.S.; supervision, O.I.L., D.P., and M.V.S.; project administration, O.I.L., D.P., and M.V.S.; funding acquisition, O.I.L., D.P., and M.V.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Russian Science Foundation (grant number 20-14-00086) and by the Russian Ministry of Science and Higher Education (grant number 121031300041-4; expression and purification of recombinant proteins).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in the study are included in the article and supplementary material; further inquiries can be directed to the corresponding author.
Acknowledgments
The authors gratefully acknowledge UEVE Université Paris-Saclay and Genopole EVRY for the constant support of the Laboratoire Structure-Activité des Biomolécules Normales et Pathologiques (INSERM U1204, University of Evry/Paris-Saclay), as well as the INSERM PRI grant [RaPiD]. The authors also thank Nikolai A. Shevchuk for proofreading and comments.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of the data; in the writing of the manuscript; or in the decision to publish the results.
References
- Kovar, H.Dr. Jekyll and Mr. Hyde: The two faces of the FUS/EWS/TAF15 protein family. Sarcoma. 2011, 2011, 837474. [CrossRef]
- Wang, H.; Hegde, M.L. New mechanisms of DNA repair defects in fused in sarcoma–associated neurodegeneration: Stage set for DNA repair-based therapeutics? J. Exp. Neurosci. 2019, 13, 1179069519856358. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Choi, J.M.; Holehouse, A.S.; Lee, H.O.; Zhang, X.; Jahnel, M.; Maharana, S.; Régis Lemaitre, R.; Pozniakovsky, A.; Drechsel, D.; Ina Poser, I.; Pappu, R.V.; Alberti, S.; Hyman, A. A. A molecular grammar governing the driving forces for phase separation of prion-like RNA binding proteins. Cell. [CrossRef]
- Iko, Y.; Kodama, T.S.; Kasai, N.; Oyama, T.; Morita, E.H.; Muto, T.; Okumura, M.; Fujii, R.; Takumi, T.; Tate, S.-I.; Morikawa, K. Domain architectures and characterization of an RNA-binding protein, TLS. J. Biol. Chem. 2004, 279, 44834–44840. [Google Scholar] [CrossRef]
- Deng, H.; Gao, K.; Jankovic, J. The role of FUS gene variants in neurodegenerative diseases. Nat. Rev. Neurol. 2014, 10, 337–348. [Google Scholar] [CrossRef]
- Kato, M.; Han, T.W.; Xie, S.; Shi, K.; Du, X.; Wu, L.C.; Mirzaei, H.; Goldsmith, E.J.; Longgood, J.; Pei, J.; Grishin, N.V.; Frantz, D.E.; Schneider, J.W.; Chen, S.; Li, L.; Sawaya, M.R.; Eisenberg, D.; Tycko, R.; McKnight, S.L. Cell-free formation of RNA granules: Low complexity sequence domains form dynamic fibers within hydrogels. Cell 2012, 149, 753–767. [Google Scholar] [CrossRef]
- Patel, A.; Lee, H.O.; Pozniakovski, A.; Jawerth, L.; Poser, I.; Maghelli, N.; Royer, L.A.; Martin Weigert, M.; Myers, E.W.; Grill, S.; Drechsel, D.; Hyman, A.A.; Alberti, S. A liquid-to-solid phase transition of the ALS protein FUS accelerated by disease mutation. Cell 2015, 162, 1066–1077. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Warraich, S.T.; Nicholson, G.A.; Blair, I.P. Fused in sarcoma/translocated in liposarcoma: A multifunctional DNA/RNA binding protein. Int. J. Biochem. Cell Biol. 2010, 42, 1408–1411. [Google Scholar] [CrossRef]
- Sama, R.R.K.; Ward, C.L.; Bosco, D.A. Functions of FUS/TLS from DNA repair to stress response: Implications for ALS. ASN Neuro 2014, 6, 1759091414544472. [Google Scholar] [CrossRef] [PubMed]
- Sukhanova, M.V., Singatulina, A.S., Pastré, D., & Lavrik, O. I. Fused in sarcoma (FUS) in DNA repair: tango with poly (ADP-ribose) polymerase 1 and compartmentalisation of damaged DNA. Int. J. Mol. Sci. 2020, 21, 7020. [CrossRef]
- Lüscher, B.; Ahel, I.; Altmeyer, M.; Ashworth, A.; Bai, P.; Chang, P.; Cohen, M.; Corda, D.; Dantzer, F.; Daugherty, M.; et al. ADP-ribosyltransferases, an update on function and nomenclature. FEBS J. 2022, 289, 7399–7410. [Google Scholar] [CrossRef]
- Mastrocola, A.S.; Kim, S.H.; Trinh, A.T.; Rodenkirch, L.A.; Tibbetts, R.S. The RNA binding protein fused in sarcoma (FUS) functions downstream of PARP in response to DNA damage. J. Biol. Chem. 2013, 288, 24731–24741. [Google Scholar] [CrossRef]
- Rulten, S.L.; Rotheray, A.; Green, R.L.; Grundy, G.J.; Moore, D.A.; Gomez-Herreros, F.; Hafezparast, M.; Caldecott, K.W. PARP-1 dependent recruitment of the amyotrophic lateral sclerosis-associated protein FUS/TLS to sites of oxidative DNA damage. Nucleic Acids Res. 2014, 42, 307–314. [Google Scholar] [CrossRef]
- Altmeyer, M.; Neelsen, K.J.; Teloni, F.; Pozdnyakova, I.; Pellegrino, S.; Grofte, M.; Rask, M.B.; Streicher, W.; Jungmichel, S.; Nielsen, M.L.; Lukas, J. Liquid demixing of intrinsically disordered proteins is seeded by poly (ADP-ribose). Nat. Commun. 2015, 6, 8088. [Google Scholar] [CrossRef]
- Singatulina, A.S.; Hamon, L.; Sukhanova, M.V.; Desforges, B.; Joshi, V.; Bouhss, A.; Lavrik, O.I.; Pastré, D. PARP-1 activation directs FUS to DNA damage sites to form PARG-reversible compartments enriched in damaged DNA. Cell Rep. 2019, 27, 809–1821. [Google Scholar] [CrossRef]
- Ame, J.C.; Spenlehauer, C.; de Murcia, G. The PARP superfamily. Bioessays 2004, 26, 882–893. [Google Scholar] [CrossRef]
- Althaus, F.R.; Richter, C.; Althaus, F.R.; Richter, C. Poly (ADP-ribose): Structure, properties and quantification. In ADP-Ribosylation of Proteins: Enzymology and Biological Significance. Publisher: Springer Berlin, Heidelberg, Germany, 1987; 37, pp. 3-11.
- Sukhanova, M. V., Abrakhi; Lavrik, O. I. Single molecule detection of PARP1 and PARP2 interaction with DNA strand breaks and their poly (ADP-ribosyl) ation using high-resolution AFM imaging. Nucleic Acids Res. 2016, 44, e60. [Google Scholar] [CrossRef]
- Teloni, F.; Altmeyer, M. Readers of poly (ADP-ribose): designed to be fit for purpose. Nucleic Acids Res. 2015, 44, 993–1006. [Google Scholar] [CrossRef]
- Koczor, C.A.; Saville, K.M.; Andrews, J.F.; Clark, J.; Fang, Q.; Li, J.; Al-Rahahleh, R.Q.; Ibrahim, Md.; McClellan, S.; Makarov, M.V.; Migaud, M.E.; Sobol, R.W. Temporal dynamics of base excision/single-strand break repair protein complex assembly/disassembly are modulated by the PARP/NAD+/SIRT6 axis. Cell Rep. 2021, 37, 109917–10. [Google Scholar] [CrossRef]
- Reber, J. M.; Mangerich, A. Why structure and chain length matter: on the biological significance underlying the structural heterogeneity of poly (ADP-ribose). Nucleic Acids Res. 2021, 49, 8432–8448. [Google Scholar] [CrossRef]
- Smith, R.; Zentout, S.; Rother, M.; Bigot, N.; Chapuis, C.; Mihuț, A.; Zobel, F.F.; Ahel, I.; van Attikum, H.; Timinszky, G.; Huet, S. HPF1-dependent histone ADP-ribosylation triggers chromatin relaxation to promote the recruitment of repair factors at sites of DNA damage. Nat. Struct. Mol. Biol. 2023, 30, 678–691. [Google Scholar] [CrossRef]
- Leung, A. K. Poly (ADP-ribose): a dynamic trigger for biomolecular condensate formation. Trends Cell Biol. 2020, 30, 370–383. [Google Scholar] [CrossRef]
- Alemasova, E.E.; Lavrik, O.I. A sePARate phase? Poly (ADP-ribose) versus RNA in the organization of biomolecular condensates. Nucleic Acids Res. 2022, 50, 10817–10838. [Google Scholar] [CrossRef]
- Rhine, K.; Odeh, H.M.; Shorter, J.; Myong, S. Regulation of biomolecular condensates by Poly(ADP-ribose). Chemical Rev. 2023, 123, 9065–9093. [Google Scholar] [CrossRef]
- Rhine, K.; Dasovich, M.; Yoniles, J.; Badiee, M.; Skanchy, S.; Ganser, L.R.; Ge, Y.; Fare, C.M.; Shorter, J.; Leung, A.K.L.; Myong, S. Poly(ADP-ribose) drives condensation of FUS via a transient interaction. Mol. Cell 2022, 82, 969–985. [Google Scholar] [CrossRef]
- Sukhanova, M.V.; Anarbaev, R.O.; Maltseva, E.A.; Pastré, D.; Lavrik, O.I. FUS Microphase Separation: Regulation by Nucleic Acid Polymers and DNA Repair Proteins. Int. J. Mol. Sci. 2022, 23, 13200. [Google Scholar] [CrossRef]
- Badiee, M.; Kenet, A.L.; Ganser, L.R.; Paul, T.; Myong, S.; Leung, A.K. Switch-like compaction of poly(ADP-ribose) upon cation binding. Proc. Natl. Acad. Sci. U.S.A. 2023, 120, e2215068120. [Google Scholar] [CrossRef]
- Chappidi, N.; Quail, T.; Doll, S.; Vogel, L.T.; Aleksandrov, R.; Felekyan, S.; Kühnemuth, R.; Stoynov, S.; Seidel, C.A.M.; Brugués, J.; Jahnel, M.; Franzmann, T.M.; Alberti, S. PARP1-DNA co-condensation drives DNA repair site assembly to prevent disjunction of broken DNA ends. Cell, 2024; 187, 945–961.e18. [Google Scholar] [CrossRef]
- Wang, W. J.; Tan, C. P.; Mao, Z. W. Metals and inorganic molecules in regulating protein and nucleic acid phase separation. Curr. Opin. Chem. Biol. 2023, 74, 102308. [Google Scholar] [CrossRef]
- Sołtys, K.; Tarczewska, A.; Bystranowska, D. Modulation of biomolecular phase behavior by metal ions. Biochim. Biophys. Acta, Mol. Cell Res. 2023, 1870, 119567. [Google Scholar] [CrossRef]
- Müller, K.H.; Hayward, R.; Rajan, R.; Whitehead, M.; Cobb, A.M.; Ahmad, S.; Sun, M.; Goldberga, I.; Li, R.; Bashtanova, U.; Puszkarska, A.M.; Reid, D.G.; Brooks, R.A.; Skepper, J.N.; Bordoloi, J.; Chow, W.Y.; Oschkinat, H.; Groombridge, A.; Scherman, O.A.; Harrison, J.A.; Verhulst, A.; D’Haese, P.C.; Neven, E.; Needham, L.-M.; Lee, S.F.; Shanahan, C.M.; Duer, M.J. Poly(ADP-Ribose) links the DNA damage response and biomineralization. Cell Rep. 2019, 27, 3124–3138. [Google Scholar] [CrossRef]
- Wang, T.; Coshic, K.; Badiee, M.; Aksimentiev, A.; Pollack, L.; Leung, A.K.L. Cation-induced intramolecular coil-to-globule transition in poly (ADP-ribose). Nat. Commun. 2024, 15, 7901. [Google Scholar] [CrossRef]
- Vasil’eva, I.A.; Anarbaev, R.O.; Moor, N.A.; Lavrik, O.I. Dynamic light scattering study of base excision DNA repair proteins and their complexes. Biochim. Biophys. Acta. 2019, 1867, 297–305. [Google Scholar] [CrossRef]
- Vasil’eva, I.; Moor, N.; Anarbaev, R.; Kutuzov, M.; Lavrik, O. Functional roles of PARP2 in assembling protein–protein complexes involved in base excision DNA repair. Int. J. Mol. Sci. 2021, 22, 4679. [Google Scholar] [CrossRef]
- Sukhanova, M.V.; Anarbaev, R.O.; Maltseva, E.A.; Kutuzov, M.M.; Lavrik, O.I. Divalent and multivalent cations control liquid-like assembly of poly (ADP-ribosyl) ated PARP1 into multimolecular associates in vitro. Commun. Biol. 2024, 7, 1148. [Google Scholar] [CrossRef]
- Hassa, P.O. The molecular “Jekyll and Hyde” duality of PARP1 in cell death and cell survival. Front. Biosci. 2009, 14, 72–111. [Google Scholar] [CrossRef]
- O’Sullivan, J., Ferreira, M.T., Gagné, J.P., Sharma, A.K., Hendzel, M.J., Masson, J.Y., Poirier, G.G. Emerging roles of eraser enzymes in the dynamic control of protein ADP-ribosylation. Nat. Commun. 2019, 10, 1182. [CrossRef]
- Hein, M.Y.; Hubner, N.C.; Poser, I.; Cox, J.; Nagaraj, N.; Toyoda, Y.; Gak, I.A.; Weisswange, I.; Mansfeld, J.; Buchholz, F.; Hyman, A.A.; Mann, M. A human interactome in three quantitative dimensions organized by stoichiometries and abundances. Cell 2015, 163, 712–723. [Google Scholar] [CrossRef]
- Chaberek, Jr.S.; Martell, A.E. (1955). Interaction of divalent metal ions with N-hydroxyethylethylenediaminetriacetic acid. J. Am. Chem. Soc. 1955, 77, 1477–1480. [Google Scholar] [CrossRef]
- Miwa, M.; Saikawa, N.; Yamaizumi, Z.; Nishimura, S.; Sugimura, T. Structure of poly(adenosine diphosphate ribose): identification of 2’-(1’‘-ribosyl-2’‘-(or 3’‘-)(1’‘‘-ribosyl))adenosine-5’,5’‘,5’‘‘-tris(phosphate) as a branch linkage. Proc. Nat. Acad. Sci. U.S.A. 1979, 76, 595–599. [Google Scholar] [CrossRef]
- Aberle, L.; Krüger, A.; Reber, J.M.; Lippmann, M.; Hufnagel, M.; Schmalz, M.; Trussina, I.R.E.A.; Schlesiger, S.; Zubel, T.; Schütz, K.; Marx, A.; Hartwig, A.; Ferrando-May, E.; Bürkle, A.; Mangerich, A. PARP1 Catalytic Variants Reveal Branching and Chain Length-specific Functions of poly(ADP-Ribose) in Cellular Physiology and Stress Response. Nucleic Acids Res. 2020, 48, 10015–10033. [Google Scholar] [CrossRef]
- Rolli, V.; O’Farrell, M.; Ménissier-de Murcia, J.; de Murcia, G. Random Mutagenesis of the Poly(ADP-Ribose) Polymerase Catalytic Domain Reveals Amino Acids Involved in Polymer Branching. Biochemistry 1997, 36, 12147–12154. [Google Scholar] [CrossRef]
- Naumenko, K.N.; Sukhanova, M.V.; Hamon, L.; Kurgina, T.A.; Anarbaev, R.O.; Mangerich, A.; Pastre, D.; Lavrik, O. I. The C-terminal domain of Y-box binding protein 1 exhibits structure-specific binding to poly (ADP-ribose), which regulates PARP1 activity. Fron. Cell Dev. Biol. 2022, 10, 831741. [Google Scholar] [CrossRef]
- Babinchak, W.M.; Surewicz, W. K. Studying protein aggregation in the context of liquid-liquid phase separation using fluorescence and atomic force microscopy, fluorescence and turbidity assays, and FRAP. Bio Protoc. 2020, 10, e3489. [Google Scholar] [CrossRef]
- Berkeley, R.F.; Kashefi, M.; Debelouchina, G.T. Real-time observation of structure and dynamics during the liquid-to-solid transition of FUS LC. Biophys. J. 2021, 120, 1276–1287. [Google Scholar] [CrossRef]
- Miné-Hattab, J.; Liu, S.; Taddei, A. Repair Foci as Liquid Phase Separation: Evidence and Limitations. Genes 2022, 13, 1846. [Google Scholar] [CrossRef]
- Dall’Agnese, G.; Dall’Agnese, A.; Banani, S.F.; Codrich, M.; Malfatti, M.C.; Antoniali, G.; Tell, G. Role of condensates in modulating DNA repair pathways and its implication for chemoresistance. J. Biol. Chem. 2023, 299, 104800. [Google Scholar] [CrossRef]
- Wang, H.; Guo, W.; Mitra, J.; Hegde, P.M.; Vandoorne, T.; Eckelmann, B.J.; Mitra, S.; Tomkinson, A.E.; Van Den Bosch, L.; Hegde, M.L. Mutant FUS causes DNA ligation defects to inhibit oxidative damage repair in Amyotrophic Lateral Sclerosis. Nat. Commun. 2018, 9, 3683. [Google Scholar] [CrossRef]
- Mamontova, E.M.; Clément, M.J.; Sukhanova, M.V.; Joshi, V.; Bouhss, A.; Rengifo-Gonzalez, J.C.; Desforges, B.; Hamon, L.; Lavrik, O.I.; Pastré, D. FUS RRM regulates poly (ADP-ribose) levels after transcriptional arrest and PARP-1 activation on DNA damage. Cell Rep. 2023, 42, 113199. [Google Scholar] [CrossRef]
- Igarashi, K.; Kashiwagi, K. Modulation of cellular function by polyamines. Int. J. Biochem. Cell Biol. 2010, 42, 39–51. [Google Scholar] [CrossRef]
- Romani, A.M. Cellular magnesium homeostasis. Arch. Biochem. Biophys. 2011, 512, 1–23. [Google Scholar] [CrossRef]
- Bagur, R.; Hajnóczky, G. Intracellular Ca2+ sensing: its role in calcium homeostasis and signaling. Mol. Cell 2017, 66, 780–788. [Google Scholar] [CrossRef]
- Lee, C.Y.; Su, G.C.; Huang, W.Y.; Ko, M.Y.; Yeh, H.Y.; Chang, G.D.; Lin, S.J.; Chi, P. Promotion of homology-directed DNA repair by polyamines. Nat. Commun. 2019, 10, 65. [Google Scholar] [CrossRef]
- Haberland, V.; M., Magin, S.; Iliakis, G.; Hartwig, A. Impact of Manganese and Chromate on Specific DNA Double-Strand Break Repair Pathways. Int. J. Mol. Sci. 2023, 24, 10392. [CrossRef]
- Gafter, U.; Malachi, T.; Ori, Y.; Breitbart, H. The role of calcium in human lymphocyte DNA repair ability. J. Lab. Clin. Med. 1997, 130, 33–41. [Google Scholar] [CrossRef]
- Kun, E.; Kirsten, E.; Mendeleyev, J.; Ordahl, C.P. Regulation of the Enzymatic Catalysis of Poly(ADP-ribose) Polymerase by dsDNA, Polyamines, Mg2+, Ca2+, Histones H1and H3, and ATP. Biochemistry 2004, 43, 210–216. [Google Scholar] [CrossRef]
- Naumenko, K.N., Sukhanova, M.V., Hamon, L., Kurgina, T.A., Alemasova, E.E., Kutuzov, M.M., Pastre, D.; Lavrik, O.I. Regulation of poly (ADP-Ribose) polymerase 1 activity by Y-Box-Binding protein 1. Biomolecules 2020, 10, 1325. [CrossRef]
- Percival, M.; Pantoja, C.F.; Cima-Omori, M.S.; Becker, S.; Zweckstetter, M. (2024). Polyamines promote disordered protein phase separation. bioRxiv 2024, 2024–05. [Google Scholar] [CrossRef]
- Fahrer, J.; Kranaster, R.; Altmeyer, M.; Marx, A.; Bürkle, A. Quantitative Analysis of the Binding Affinity of poly(ADP-Ribose) to Specific Binding Proteins as a Function of Chain Length. Nucleic Acids Res. 2007, 35, e143. [Google Scholar] [CrossRef]
- Pleschke, J.M.; Kleczkowska, H.E.; Strohm, M.; Althaus, F.R. Poly (ADP-ribose) binds to specific domains in DNA damage checkpoint proteins. J. Biol. Chem. 2000, 275, 40974–40980. [Google Scholar] [CrossRef]
Figure 1.
Typical volume-weighted size distributions for FUS, PARP1WT, PAR-PARP1WT, and a FUS–PAR-PARP1WT mixture. The profiles were obtained by means of experimental autocorrelation functions in the Zetasizer Nano ZS software. Average hydrodynamic radii (Rh) computed from the distributions are presented as well. Rh is the average value estimated from at least three DLS experiments. Size measurement was performed on FUS, PARP1WT, PAR-PARP1, and the mixture of FUS with PAR-PARP1WT in reaction mixtures consisting of either 2.5 µM PARP1, 10 μM FUS, and 2.5 µM PAR-PARP1WT or 10 μM FUS and 60–540 nM PAR-PARP1. The Rh values were measured directly after 3-min incubation of FUS with PAR-PARP1WT.
Figure 1.
Typical volume-weighted size distributions for FUS, PARP1WT, PAR-PARP1WT, and a FUS–PAR-PARP1WT mixture. The profiles were obtained by means of experimental autocorrelation functions in the Zetasizer Nano ZS software. Average hydrodynamic radii (Rh) computed from the distributions are presented as well. Rh is the average value estimated from at least three DLS experiments. Size measurement was performed on FUS, PARP1WT, PAR-PARP1, and the mixture of FUS with PAR-PARP1WT in reaction mixtures consisting of either 2.5 µM PARP1, 10 μM FUS, and 2.5 µM PAR-PARP1WT or 10 μM FUS and 60–540 nM PAR-PARP1. The Rh values were measured directly after 3-min incubation of FUS with PAR-PARP1WT.
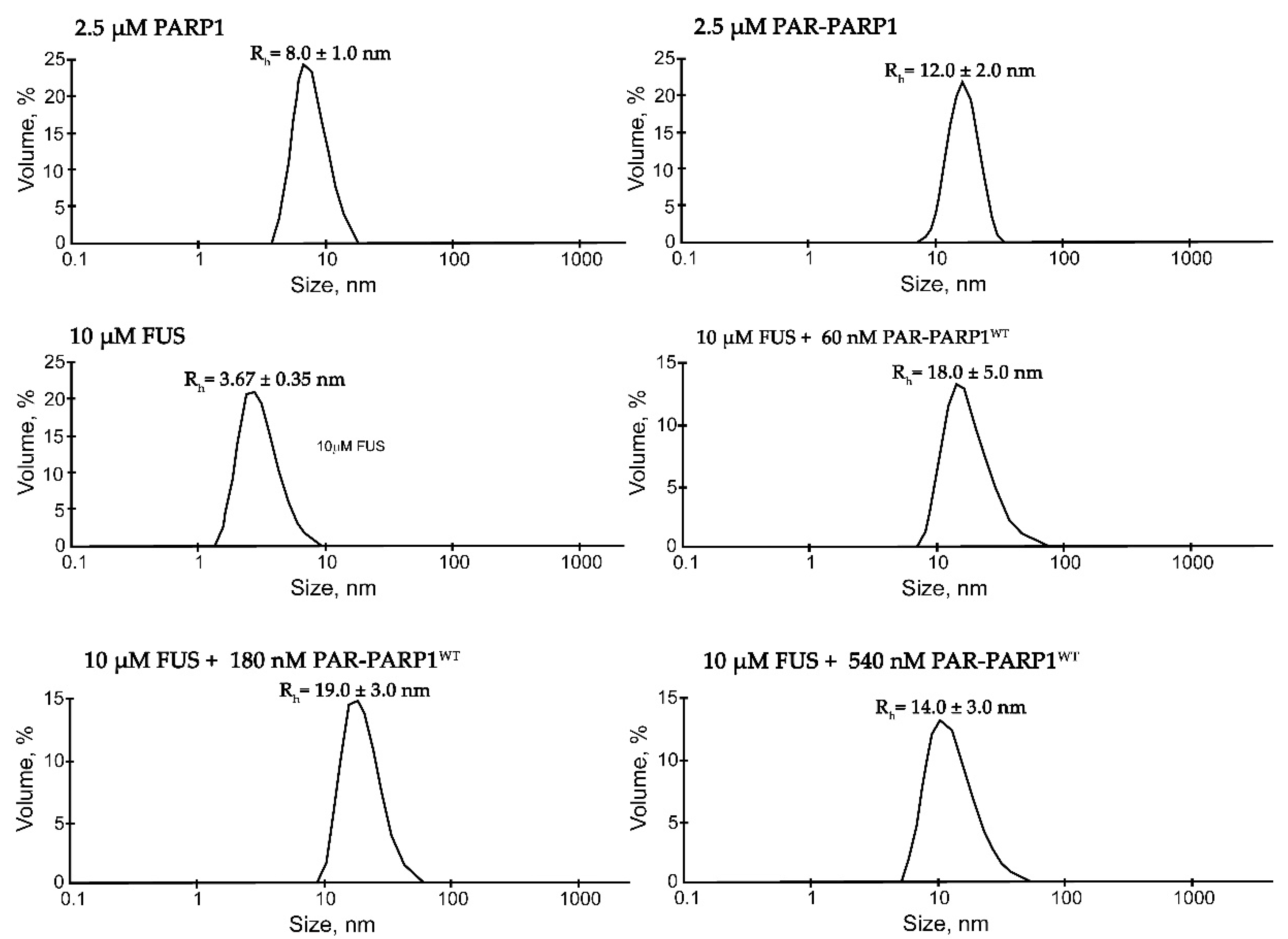
Figure 2.
Submillimolar concentration of a cation promotes FUS microphase separation at a high FUS-to-PAR-PARP1 molar ratio. Rh of FUS–PAR-PARP1 mixtures is presented as a function of the FUS-to-PAR-PARP1 molar ratio. Rh is the average value estimated from at least three DLS experiments. Analyses of FUS higher-order structure in the presence of PAR-PARP1WT were performed in reaction mixtures consisting of 10 μM FUS and 50–540 nM PAR-PARP1 as well as 0.5 mM Mg2+ (a), 0.1 mM Mn2+ (b), 0.5 mM Ca2+ (c), 0.4 mM Spd3+ (d), or 0.1 mM Spn4+ (e), as indicated in the figure. The underlining shows the FUS-to-PAR-PARP1 molar ratio at which we observed bimodal particle size distributions (Figure S1).
Figure 2.
Submillimolar concentration of a cation promotes FUS microphase separation at a high FUS-to-PAR-PARP1 molar ratio. Rh of FUS–PAR-PARP1 mixtures is presented as a function of the FUS-to-PAR-PARP1 molar ratio. Rh is the average value estimated from at least three DLS experiments. Analyses of FUS higher-order structure in the presence of PAR-PARP1WT were performed in reaction mixtures consisting of 10 μM FUS and 50–540 nM PAR-PARP1 as well as 0.5 mM Mg2+ (a), 0.1 mM Mn2+ (b), 0.5 mM Ca2+ (c), 0.4 mM Spd3+ (d), or 0.1 mM Spn4+ (e), as indicated in the figure. The underlining shows the FUS-to-PAR-PARP1 molar ratio at which we observed bimodal particle size distributions (Figure S1).
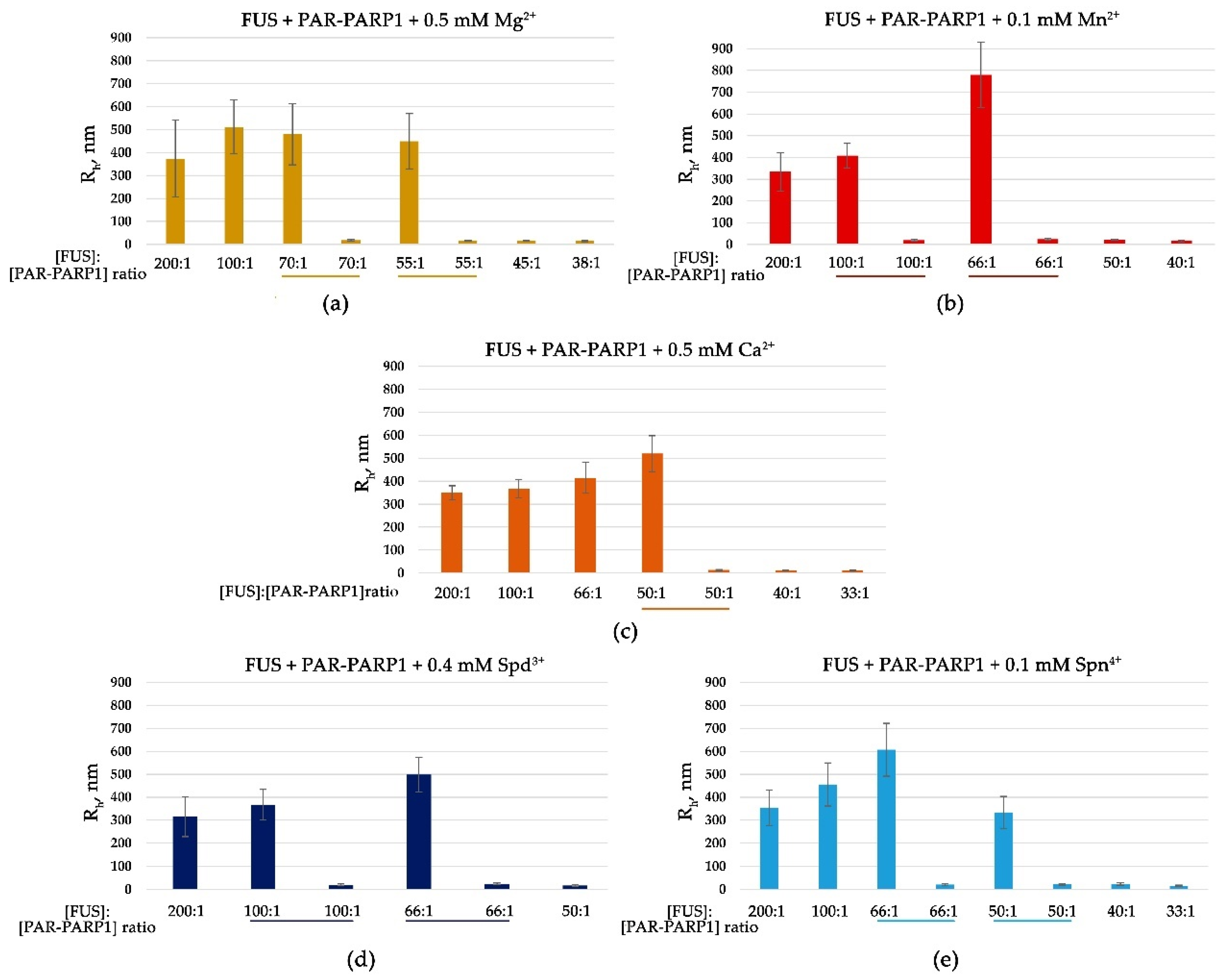
Figure 3.
Millimolar concentration of a cation promotes FUS microphase separation at a low FUS-to-PAR-PARP1 molar ratio. FUS higher-order structure analyses in the presence of PAR-PARP1WT were performed in reaction mixtures consisting of 4–5 μM FUS and 1.5–1.8 µM PAR-PARP1. PAR-PARP1 was mixed with FUS, the samples were equilibrated for 1 min, and Rh was measured next (Figure S2). After that, the reactions were supplemented with 2 mM Mn2+, 12.5 mM Mg2+, 7 mM Ca2+, 12 mM Spd3+, or 2.3 mM Spn4+, and Rh was measured again (Figure S2). To disrupt FUS–PAR-PARP1 assemblies stabilized by a cation, EDTA (5–30 mM) was added as indicated in the figure, and Rh was measured in the EDTA-treated samples (Figure S2).
Figure 3.
Millimolar concentration of a cation promotes FUS microphase separation at a low FUS-to-PAR-PARP1 molar ratio. FUS higher-order structure analyses in the presence of PAR-PARP1WT were performed in reaction mixtures consisting of 4–5 μM FUS and 1.5–1.8 µM PAR-PARP1. PAR-PARP1 was mixed with FUS, the samples were equilibrated for 1 min, and Rh was measured next (Figure S2). After that, the reactions were supplemented with 2 mM Mn2+, 12.5 mM Mg2+, 7 mM Ca2+, 12 mM Spd3+, or 2.3 mM Spn4+, and Rh was measured again (Figure S2). To disrupt FUS–PAR-PARP1 assemblies stabilized by a cation, EDTA (5–30 mM) was added as indicated in the figure, and Rh was measured in the EDTA-treated samples (Figure S2).
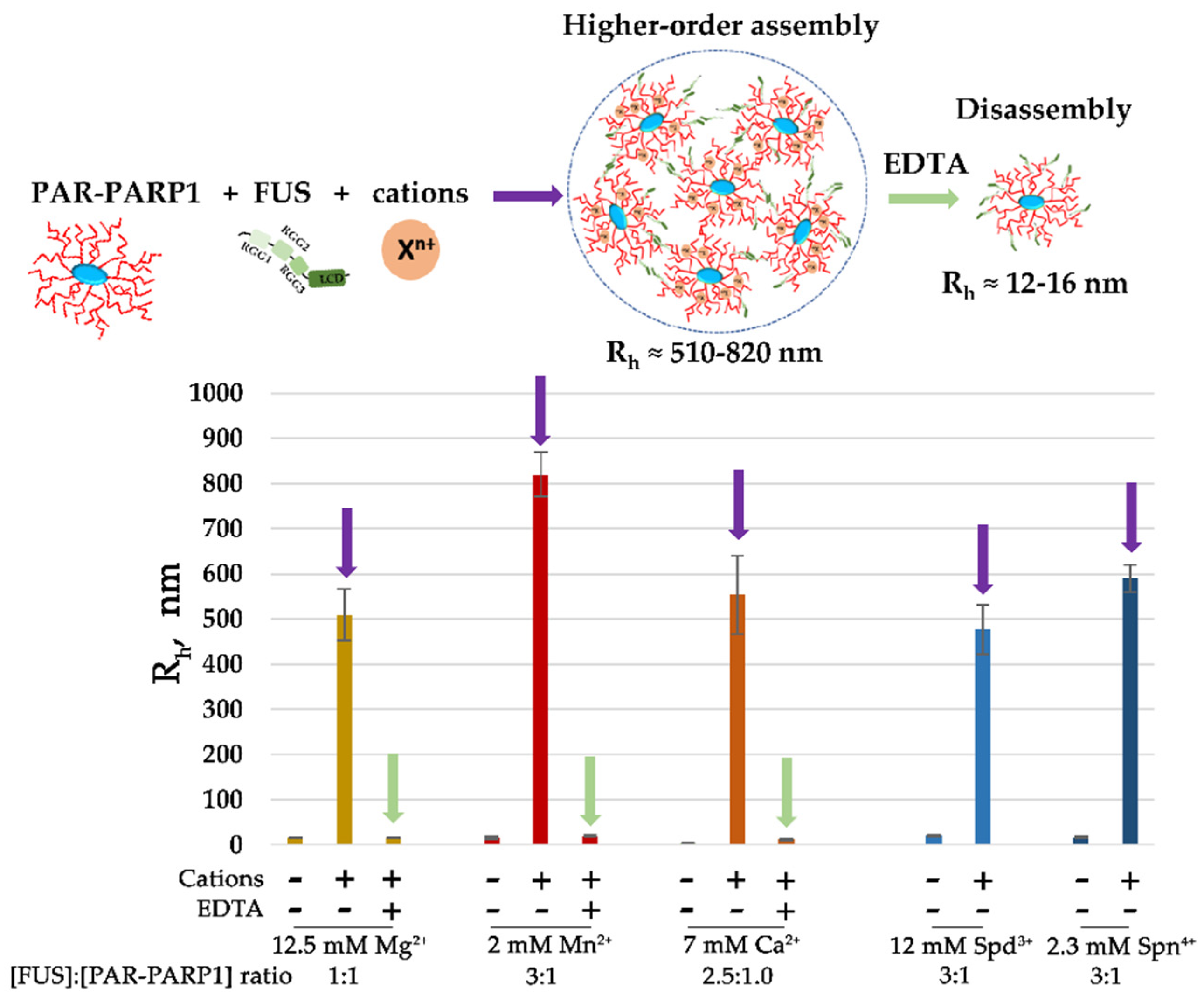
Figure 4.
Coassembly of FUS and PAR-PARP1 in the presence of Mg2+. Fluorescence images of 10 µM AF488-FUS and 2.5 µM PARP1 in the presence of 15 mM Mg2+ (a) or 10 µM AF488-FUS and 2.5 µM PARP1 in the presence of 15 mM Mg2+ and 1 mM NAD+ (b,c). The fluorescence photos were captured before and after the addition of NAD+ to FUS–PARP1 mixtures and 30–60 min incubation at room temperature. The reaction mixtures (20 µL) contained a buffer (25 mM HEPES-NaOH pH 7.5, 200 mM NaCl, 300 nM urea, and 2 mM dithiothreitol [DTT]), 10 µM AF488-FUS, 2.5 µM Cy3-PARP1, 15 mM Mg2+, and 1 mM NAD+ as indicated.
Figure 4.
Coassembly of FUS and PAR-PARP1 in the presence of Mg2+. Fluorescence images of 10 µM AF488-FUS and 2.5 µM PARP1 in the presence of 15 mM Mg2+ (a) or 10 µM AF488-FUS and 2.5 µM PARP1 in the presence of 15 mM Mg2+ and 1 mM NAD+ (b,c). The fluorescence photos were captured before and after the addition of NAD+ to FUS–PARP1 mixtures and 30–60 min incubation at room temperature. The reaction mixtures (20 µL) contained a buffer (25 mM HEPES-NaOH pH 7.5, 200 mM NaCl, 300 nM urea, and 2 mM dithiothreitol [DTT]), 10 µM AF488-FUS, 2.5 µM Cy3-PARP1, 15 mM Mg2+, and 1 mM NAD+ as indicated.

Figure 5.
The PAR-PARP1 mutant producing hyperbranched PAR is more effective in the seeding of higher-order assembly of FUS. Rh is presented as a function of the FUS-to-PAR-PARP1 molar ratio. Rh is the average value estimated from at least three DLS experiments (Figure S5). (a,b) Analyses of FUS higher-order structure in the presence of either PAR-PARP1Y986S or PAR-PARP1Y986H were performed in reaction mixtures consisting of 10 μM FUS and either 60–285 nM PAR-PARP1Y986S (a) or 50–1000 nM PAR-PARP1Y986H (b) in the presence of 0.5 mM Mg2+ as indicated in the figure. (c) Analyses of FUS higher-order structure in the presence of PAR-PARP1Y986H were performed in reaction mixtures consisting of 10 μM FUS and 60–250 nM PAR-PARP1Y986H. (d) FUS higher-order structure analyses in the presence of either PAR-PARP1Y986S or PAR-PARP1Y986H were performed in reaction mixtures consisting of 1.8 μM FUS and 2 µM PAR-PARP1. PAR-PARP1 was mixed with FUS, the samples were equilibrated for 1 min, and Rh was measured. After that, the reactions were supplemented with 12.5 mM Mg2+, and Rh was measured again. To disrupt FUS–PAR-PARP1 assemblies stabilized by a cation, EDTA (30 mM) was introduced as indicated in the figure, and Rh was measured in the EDTA-treated samples.
Figure 5.
The PAR-PARP1 mutant producing hyperbranched PAR is more effective in the seeding of higher-order assembly of FUS. Rh is presented as a function of the FUS-to-PAR-PARP1 molar ratio. Rh is the average value estimated from at least three DLS experiments (Figure S5). (a,b) Analyses of FUS higher-order structure in the presence of either PAR-PARP1Y986S or PAR-PARP1Y986H were performed in reaction mixtures consisting of 10 μM FUS and either 60–285 nM PAR-PARP1Y986S (a) or 50–1000 nM PAR-PARP1Y986H (b) in the presence of 0.5 mM Mg2+ as indicated in the figure. (c) Analyses of FUS higher-order structure in the presence of PAR-PARP1Y986H were performed in reaction mixtures consisting of 10 μM FUS and 60–250 nM PAR-PARP1Y986H. (d) FUS higher-order structure analyses in the presence of either PAR-PARP1Y986S or PAR-PARP1Y986H were performed in reaction mixtures consisting of 1.8 μM FUS and 2 µM PAR-PARP1. PAR-PARP1 was mixed with FUS, the samples were equilibrated for 1 min, and Rh was measured. After that, the reactions were supplemented with 12.5 mM Mg2+, and Rh was measured again. To disrupt FUS–PAR-PARP1 assemblies stabilized by a cation, EDTA (30 mM) was introduced as indicated in the figure, and Rh was measured in the EDTA-treated samples.
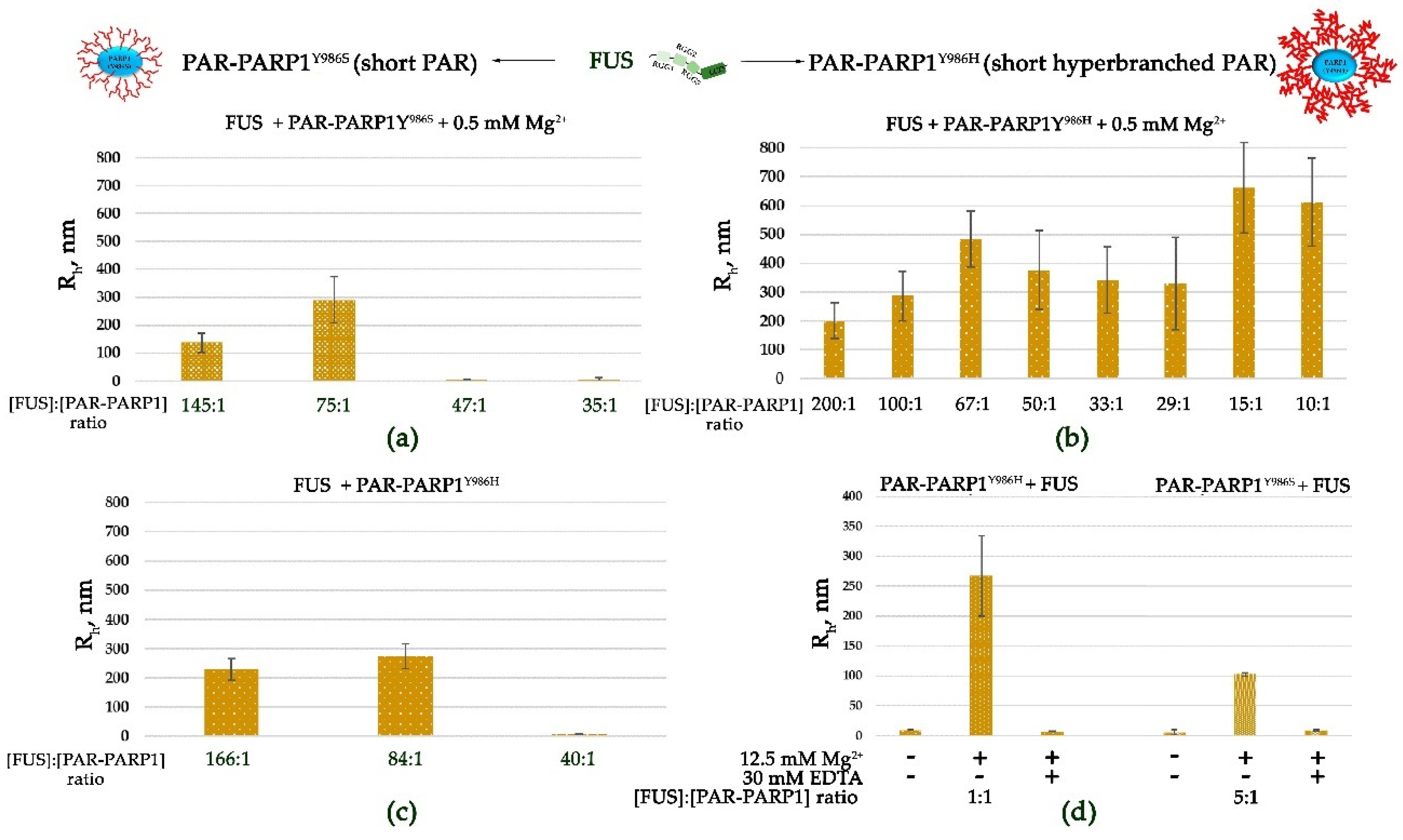
Figure 6.
In contrast to PARP1WT, mutant PAR-PARP1Y986H (producing hyperbranched PAR) seeds higher-order assembly of FUS at a lower concentration of Mg2+ and the assembly has appreciable susceptibility to 1,6-hexanediol (1,6-HD) treatment. Histograms of turbidity of a PAR-PARP1-and-FUS solution (OD at 600 nm) as determined in the presence of different concentrations of Mg2+, 1,6-HD, and EDTA. The mean ± SD of three independent measurements (Figures S6 and S7). ***p < 0.001 and *p < 0.05; t test.
Figure 6.
In contrast to PARP1WT, mutant PAR-PARP1Y986H (producing hyperbranched PAR) seeds higher-order assembly of FUS at a lower concentration of Mg2+ and the assembly has appreciable susceptibility to 1,6-hexanediol (1,6-HD) treatment. Histograms of turbidity of a PAR-PARP1-and-FUS solution (OD at 600 nm) as determined in the presence of different concentrations of Mg2+, 1,6-HD, and EDTA. The mean ± SD of three independent measurements (Figures S6 and S7). ***p < 0.001 and *p < 0.05; t test.
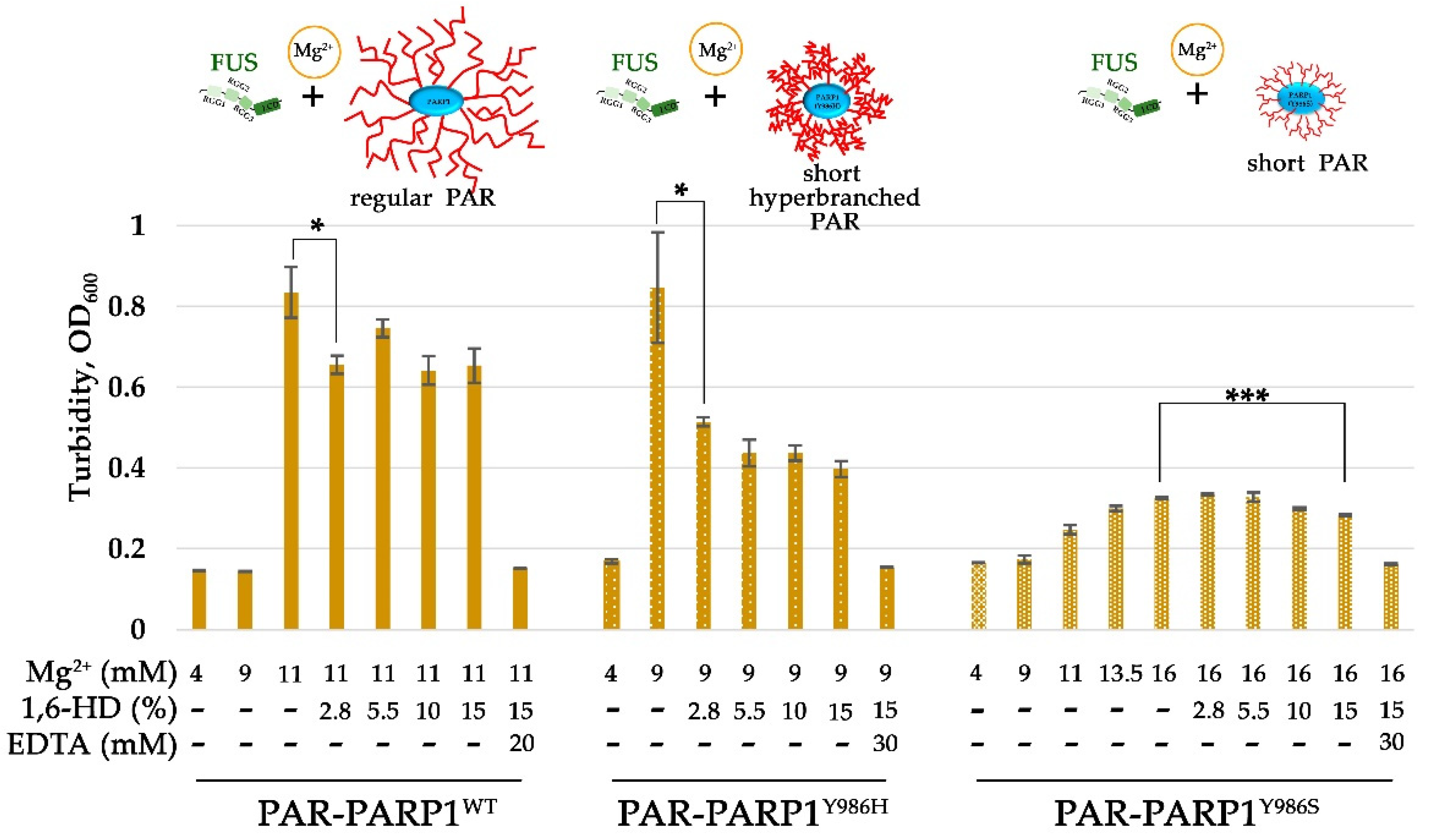
Figure 7.
PAR size and structure affect the size of the higher-order assembly of FUS with PAR-PARP1. (a) AFM visualization of PAR-PARP1WT and PAR-PARP1Y986H and their higher-order assemblies with FUS. Scale bar: 300 nm; Z scale: 3 nm and 6 nm as indicated. (b,c) Measurement of the particle diameter by AFM (horizontal bars denote the mean); n = 100 particles from three independent samples; paired t test, ****P < 0.000001.
Figure 7.
PAR size and structure affect the size of the higher-order assembly of FUS with PAR-PARP1. (a) AFM visualization of PAR-PARP1WT and PAR-PARP1Y986H and their higher-order assemblies with FUS. Scale bar: 300 nm; Z scale: 3 nm and 6 nm as indicated. (b,c) Measurement of the particle diameter by AFM (horizontal bars denote the mean); n = 100 particles from three independent samples; paired t test, ****P < 0.000001.

Figure 8.
The proposed model of cation-dependent assembly of FUS with PAR-PARP1. Cations stabilize FUS association with PAR-PARP1; such assembly is sensitive to chelating agents such as EDTA and to 1,6-hexanediol, which disrupts hydrophobic interactions.
Figure 8.
The proposed model of cation-dependent assembly of FUS with PAR-PARP1. Cations stabilize FUS association with PAR-PARP1; such assembly is sensitive to chelating agents such as EDTA and to 1,6-hexanediol, which disrupts hydrophobic interactions.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



