Submitted:
30 September 2024
Posted:
01 October 2024
You are already at the latest version
Abstract
The use of probiotics has been gaining popularity in terms of inclusion into human diets over recent years. Based on properties exerted by these living organisms, several benefits have been elucidated and conferred to the host. Bacteria has been more commonly used in probiotic preparations, in comparison to yeast candidates, however, yeast exhibit several beneficial properties such as the prevention and treatment of diarrhoea, production of antimicrobial agents, prevention of pathogen adherence to intestinal sites, maintain microbial balance, modulation of the immune system, resistant to some antibiotics, amongst others. This review details the use of yeast organisms as biotherapeutics and has a special focus on production considerations and their formulation into different delivery formats.
Keywords:
Probiotics
; yeast
; biotherapeutics
; formulation
1. Introduction
Probiotics and their use for human health implications have been studied extensively, and in more recent years, their acceptance for use by the global population has seen a positive trajectory. According to the most widely accepted definition, a probiotic is known to be “live microorganisms which when administered in adequate amounts confer a health benefit on the host” [1].
Generally, only bacterial and yeast organisms have been classified as probiotics, with the most common being Lactobacilli, Bifidobacteria, Enterococci, Faecalibacterium, Clostridia and more recently Propionibacteria [2]. Upon application, these organisms have been found to prevent and treat various clinical diseases, improve the intestinal micro-environment, prevent physiological stress and the proliferation of pathogens, improve health of the intestinal epithelium, modulate immunological homeostasis amongst others [3]. Historically, negative perceptions plagued yeast cultures, as these organisms were generally referred to as pathogenic, disease-causing microbes. However, several studies centred on the use of non-pathogenic yeasts, that possess probiotic properties have emerged, which highlights their innate ability to influence physiology, metabolism, and immune homeostasis in the colon [4]. Yeasts have been studied and have proven to be effective starter cultures, and significant interest has been noted in their use in various biotechnological applications [5].
Yeasts make up <0.1% of the human microbiome Most yeast isolates that have been isolated from the human microbiome include Candida albicans, Torulopsis glabratra, Candida tropicalis, Malassezia spp., and Saccharomyces spp [6], [7]. Other probiotic yeast candidates include Cryptococcus spp. Candida famata [9], C. tropicalis [10] , Debaryomyces hansenii [11], Issatchenkia orientalis [10], Kluyveromyces lactis [12], Kluyveromyces marxianus [12], [13], [14], Metschnikowia gruessii [13], Pichia jadinii (Buerth et al. 2016), Pichia kluyveri [10], Pichia kudriavzevii [10], Pichia pastoris [15], Pichia guilliermondii [16], Wickeramomyces anomalus [16]. These organisms, due to their ability to resist low pHs, to produce digestive enzymes, bile salts, organic acids, make them ideal candidates to serve as probiotics [7].
Yeasts that have been classified as generally regarded as safe (GRAS), have been shown to have several health implications on the human host. These influences may include but are not limited to being effective on gut microbiota dysbiosis, possess anti-inflammatory, anti-proliferative, anti-cancer and anti-allergenic properties [17,18].
Saccharomyces boulardii, Saccharomyces cerevisiae, Candida spp, are the most common yeasts used as probiotics, as is used most for the treatment of Clostridium difficile diarrhoea [19]. S. boulardii, was first isolated from litchis in Indochina, and is not autochthonous in the microbiome [7]. However, this non-pathogenic yeast is known to have the following characteristics that advocates its use as a probiotic (Figure 1).
According to Arevalo-Villena et al. [20], when developing a yeast probiotic product, the following characteristics ought to be sought, in a candidate organism (Table 1). Other factors that may be considered include the assimilation of cholesterol assimilation, the deconjugation of bile salts, demonstration of antioxidant, haemolytic, cytotoxicity, activity, as well as ability to produce cytokines and phytase.
2. Status of the Biomanufacturing of Yeast Probiotics
Yeast biomass production is a common practice across the globe [28]. During probiotic production, for the interest of this review, yeast probiotics; the primary focus of process optimisation is to maximise volumetric organism productivity [29].
2.1. Key Bioprocess Considerations
The main contributing factors to this key process indicator are composition and components present in the growth medium, operational parameters used in the process, such as temperature, pH, aeration, as well as mode of cultivations, either batch, fed batch or continuous.
Despite there being several commercial probiotics available, the focus of producers is to ensure that these products can be produced economically to increase market share [29]. At present, major producers of probiotics have to date, developed highly efficient, refined and vertically integrated microbial production systems [29]. Upon biomanufacturing of probiotics, the following characteristics are of vital importance, to promote the uptake of the technology (Figure 2).
2.2. Advances in Yeast Probiotic Manufacture
Conventional yeast production requires a cultivation medium that typically contains carbon, nitrogen, vitamins and trace metals [30]. As the growth medium for probiotic production is a major consideration in process development, other non-conventional fermentation feedstocks are being considered to produce probiotics to minimise production costs (Table 2). Agro-industrial residues such as molasses has been used for the cultivation of yeast probiotics [17]. Molasses is a viscous, sugar rich nutrient source, that contains ~34% sucrose, glucose, fructose and other minerals. In sugar producing countries, such as Brazil, and South Africa, ~10 million tons of sugar cane molasses is discharged [17], hence making it a suitable nutrient feedstock for the large-scale production and manufacture of probiotics.
Additionally, with the rise in food production, food waste volumes are also on the increase. Hence, the food waste that is accumulated, is rich in proteins carbohydrates and lipids, and therefore can be used as a suitable substrate to cultivate microorganisms (Sharma 2021). This circular economy or in some instances referred to as bioeconomy initiatives, are intended to reduce the economic, societal and environmental costs, and to drive waste to wealth activities (Sharma 2021). Probiotics are not usually the intended product produced using waste valorisation initiatives, however, there has been some successful attempts in demonstrating the concept in studies conducted by Patil et al., (2022) and [31]. These studies demonstrated the production of Kluyveromyces, Torula, Candida and Saccharomyces spp as well as Saccharomyces boulardii CCT 4308, using coffee pulp and sugar cane molasses, respectively. It is envisaged that more instances of successful waste valorisation are expected in this area of R&D in the coming years.
2.3. Challenges Associated with Yeast Probiotic Manufacturing
Less established entities that are interested in the use, manufacture and/or supply of probiotics may not have access to the skills, expertise and infrastructure required to produce these bio-based products. These entities include but are not limited to small and medium enterprise farmers, start-up biotech-based companies and the research and development community. Although these institutions and stakeholders may have a high degree of interest in probiotic development and manufacture, they may not necessarily have the high levels of competencies in probiotic development, manufacture and supply in comparison to the established players [29].
In these instances, niche probiotics may be developed, or novel candidate products may be identified through collaborative R&D partnerships, however the developmental pathway for these concepts to commercial scale are in some instances not clear and seemed to be filled with challenges and high risk. The disadvantage of these unexploited R&D initiatives, is that the value of research investments may not be realised, or the needs identified upon specific product development remain unmet [29].
Success in these instances has been attained with the creation of non-classical R&D pathways to commercialisation. Feedback loops initiated within product and process development, agile manufacturing, market and user testing, coupled to the intellectual property management and regulatory frameworks (if applicable) are necessary to rapidly develop and deliver products through the value chain [29]. This also entails the specification of production performance targets, final product adherence to end use specifications, market and financial data as well as production capacity to fulfil market demand. These factors are essential in de-risking the success if the probiotic product and its adoption for use.
2.4. Manufacturing Considerations to Produce Yeast Probiotics
During product and process development, it is a key development area to determine base line performance of the production process. Thereafter ancillary development steps may be taken to further retrofit production process, especially that of cost-sensitive unit operations. Simulations and process modelling offers a useful tool, termed – in-silico, to enable the optimisation of key process steps in the production process. With the use of this strategy, it is envisaged that significant time and expenses are saved by removing the need to conduct several actual cultivations at laboratory and pilot scales, which ultimately achieves economic impact.
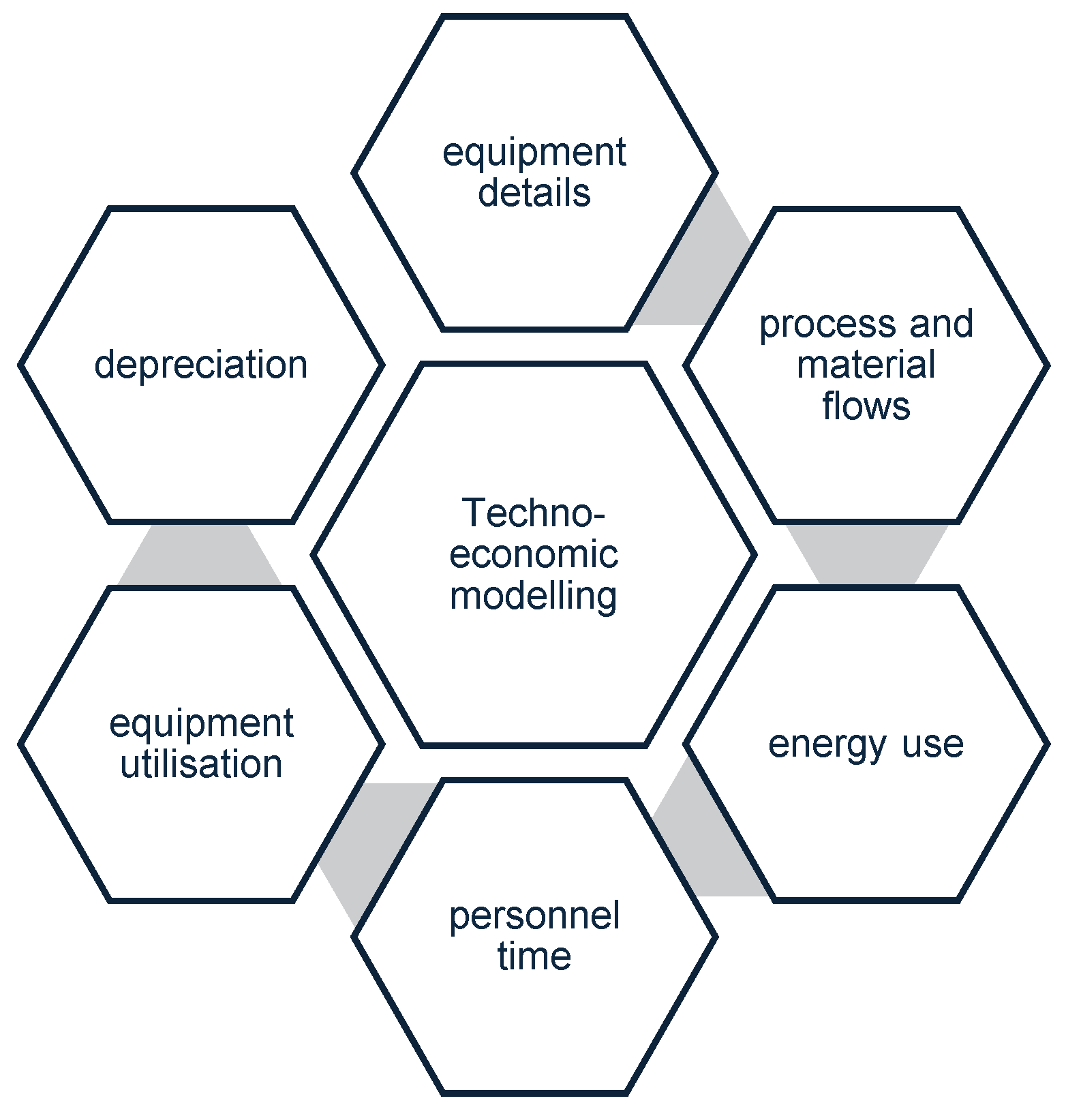
Another important consideration for probiotic production is the techno-economic modelling and assessment. This exercise details the technical and economic details of the process and includes the following components:
Figure 3.
Aspects that contribute to the techno-economic feasibility of the production process.
2.5. Location of Known Producers and Global Manufacturers of Yeast Probiotics
A key component of successfully commercialising a probiotic, is the access to suitable manufacturing expertise, particularly in developing countries. Infrastructure requirements are capital intense and are limited in availability. Probiotic technologies require a production scale pipeline that facilitates technology development from small scale to manufacturing scale.
Large enterprises that have shown efficient production competencies have skills and infrastructure that is currently producing a wide variety of products. These production facilities are fully utilised, using tight production scheduling strategies that give them the edge of new market entrants, or smaller entities that lack vertical process integration.
Biomanufacturing entities on the African continent are limited, and scarcely available. The global probiotic market consists of various entities and are categorised into three tiers (BCC Market Research, 2022). These companies and their respective tiering are listed in Table 2. As can be seen, tier 1 companies that occupy ~40 to 45 % of the global market, are found in the United States of America, Europe and Japan, with most tier 2 companies presiding in these regions.

Table 2.
Major global producers (tier 1) of probiotics (BCC Market Research 2022).
| Name | Country |
|---|---|
| ADM, | USA |
| Abbott, | USA |
| Asahi Group Holdings Ltd., | Japan |
| Chobani LLC, | USA |
| Chr. Hansen Inc., | Denmark |
| DSM | Netherlands |
| Danone Inc., IFF, | France |
| Kerry, | Ireland |
| Estee Lauder Inc., | USA |
| Morinaga Milk Industry Co. Ltd., | Japan |
| NESTLÉ, | Switzerland |
| Yakult Honsha Co. Ltd. | Japan |
3. The Use of Genetically Modified Organisms (GMO) as Probiotics.
Conventionally, wild type organisms have been applied as probiotics, however, the advances in genome editing and associated tools have unlocked the possibility of being able to engineer probiotics to deliver customised therapeutics [32]. Ma et al. [33] has provided an extensive review on the theoretical basis for probiotic gene editing technology, as well as the use of these engineered probiotics for the treatment of diseases. These diseases range from inflammatory bowel disease, cancer, obesity and diabetes amongst others, and have been tested both in human and animal models. Interestingly, only 8% of the genetically modified organisms listed in the review were yeast probiotics, with the remaining belonging to their bacterial counterparts. It is envisaged that the advent of genome editing may impart a variety of benefits to human health especially in the treatment of specific diseases. To date, there has been significant hesitation and consumer resistance for use of genetically modified organisms as probiotics, however, this aversion towards its use may be reduced with the progression of several clinical studies that are currently ongoing [34].
4. General Routes of Administration of Yeast Probiotics
The practise of using probiotics has become widely accepted as a natural means to stimulate health for humans. Today, probiotics are used as health supplements in food, as pharmaceuticals or chemical supplements [35]. If a probiotic is classified as pharmaceutical or drug for treatment of a disease or disorder, stricter requirements are necessary to substantiate the claims stated by the manufacturer. It must be proven safe and effective for its intended use through clinical trials and be approved by the Food and Drug Administration (FDA) before it can be sold. Depending on the intended use of a probiotic, if to be used as a drug or a dietary supplement or a nutraceutical, regulatory requirements differ [36]. According to the definition provided by the FDA; a drug is an article intended for use in the diagnosis, cure, mitigation, treatment, or prevention of dis-ease [37].
With regards to nutraceuticals, these are known as pharmaceutical alternatives that exert physiological benefits, used to promote health and support the structure or function of the human body [38]. Nutraceuticals are regarded as safe and less likely to have side effects, as such, probiotics are generally classified as nutraceuticals. The safety of probiotics is apparently due to the absence of toxicity in their populations. Additionally, most probiotics form part of the natural microbiome of the human host and therefore are applied back to a known environment (WHO, 2002).
In most instances, the optimal concentration of active probiotic cells, required to confer a positive benefit to the host is not known. What is known, is that the probiotic needs to be in high enough concentration to survive some of the physiological barriers upon transit to the target site. Another important consideration is the method selected for use to deliver the probiotic to the intended site.
5. Conventional Pharmaceutical Methods Used to Administer Probiotics
5.1. Oral Delivery Systems
The most common delivery of probiotics is oral administration. This infers that the final step in the probiotic production process is the formulation and packaging of a probiotic into a delivery system that will be able maintain functionality of the probiotic through the harsh gastric environment. Conventionally used delivery systems include tablets, capsules, hydrogels, granules and others as described in Figure 4.
Oral administration of the probiotic product is a widely accepted channel for the delivery of drugs and probiotic microorganisms in several disease and treatment applications. Consequently, it presents the biggest challenges in administering probiotics, as the live cell preparations found in final formulated products, need to ultimately survive the gastric environment in the stomach which can reach a pH below 3 [39]. Most authors that have deduced upon application of probiotics, that when conferred in adequate amounts, these organisms are able to confer health benefits to the host. This infers that for every step in the probiotic production process, viability needs to be maintained, and cell losses need to be kept to a minimum, since it is envisaged that a certain component of the probiotic population will die upon exposure to the gastric environment.
Table 3.
Some of the benefits that are offered from oral administrations of probiotics and addition in food products.
Table 3.
Some of the benefits that are offered from oral administrations of probiotics and addition in food products.
| Benefits | Organisms of interest | Reference |
|---|---|---|
| Inhibition of Cd absorption | L. Plantarum | [40] |
| Protection of the intestinal barrier – by alleviation of Cd-induced oxidative stress | ||
| Enhancement of antimicrobial activity | L. paracasei and L. casei | [41] |
| Reduction of hypertension effects | S. cerevisiae | [42] |
| Modification of the fecal resistome during Helocobacter pylori treatment – reduction of antibiotic resistance | S. boulardii | [43] |
| Potential in removal of toxins | S. cerevisiae W13 and S. boulardii ATCC MYA-796 | [44] |
| Improvement of glycaemic indices in type II diabetic patients. | S. cerevisiae | [45] |
| Inhibition and reduction of Gardnerella vaginalis biofilms in mice | S. cerevisiae CNCM I-3856 and L. rhamnosus ATCC 53103 | [46] |
| Cholesterol reduction |
Pichia fermentans BY5 Pichia kudriavzevii BY10 Pichia kudriavzevii BY15 Yarrowia lipolytica HY4 |
[47] |
| Better sensory properties with lower ethanol content | Meyerozyma caribbica 9D | [48] |
| Production of alcohol-free and low-alcohol products |
S. boulardii | [49] |
5.2. Transdermal Delivery Systems
According to Chen et al. [50]; methods that are simple and effective for delivery of probiotics into the dermis for the regulation are lacking. Yeast organisms, such as S. boulardii has been used for the treatment of skin ailments, including acne, due to its anti-inflammatory properties. However, in this instance, the probiotic product was administered dermally, as one would expect, but rather as an oral supplement. Upon ingestion, the anti-inflammatory effects of the organisms are exerted towards the skin. S. boulardii demonstrated the ability to produce acetic acid which is known to exert antifungal and antimicrobial effects. This preparation, when used topically, was shown to reduce the bacterial load responsible for causing infections or skin conditions such as acne, rosacea, seborrheic dermatitis (scalp eczema), and eczema. In this instance, yeast by-products and not the organism itself was used as a topical treatment. All the S. boulardii strains tested in this study, secreted an antimicrobial agent that demonstrated an inhibitory effect on E. coli and bioactivity against Candida albicans hyphae [50].
6. Other Probiotic Delivery Systems
It is known that the most common and effective delivery route for probiotics is orally, as the intended destination for the product is the GI tract of the host [51]. However, during processing of pharmaceutical products in general, such as tablets, high processing temperatures may be used to obtain aesthetically pleasing final products, which may kill the probiotic microorganisms of interest. This occurrence may be applicable to many formulations that contain viable organisms and proves to be the biggest challenge that needs to be overcome in terms of final product processing.
In order to circumvent this significant challenge, fermented also known as functional foods are used as a delivery system, and do not require high temperature processing. Instead, indigenous microorganisms already present in the ingredient mix, are activated and thereby replicate to higher concentrations. This is termed as non-conventional delivery methods, despite it being consumed by several populations dating back to early human civilisation.
Fermented foods derived from animals and plants are vital in human diets as they contain beneficial microorganisms and compounds including organic acids, ethanol or antimicrobial compounds [52]. These fermented foods are termed functional foods, which are foods that offer benefits that are more than their nutritional value and are divided into dairy and non-dairy options.
6.1. Functional Foods as a Source of Probiotics
6.1.1. Dairy - Based Probiotics
In the early 1990s, Nobel Laureate, Elie Metchnikoff (1845–1916) whilst working in Bulgaria noticed how a certain population in the country had lived a longer life span than others. Upon further investigation, the researcher discovered that this population consumed a fermented drinking yoghurt daily [53]. This preliminary research laid a foundation for the study of probiotics detailing the use of functional foods as a dietary supplement. These foods contained beneficial microorganisms; either bacterial or yeast cultures or a combination of both. S. boulardii a known yeast probiotic, has been isolated from dairy products including milk, yoghurt, cream, cheese and kefir.
Yeasts have shown the ability to produce enzymes that synthesise milk proteins. However, this property is only activated once lactic acid bacteria (LAB), breakdown the lactose present in dairy-based foods, into glucose and galactose. Once this conversion is done, both the yeasts and LAB cultures, use the available sugar as a carbon source to grow and replicate [54]. [55] added that yeasts such as S. boulardii should be included in products solely as a probiotic as it offers no benefits to some dairy products such as yoghurts, however in cheese making, yeasts have a more important contributions to the process of cheese maturation. These yeasts contribute to the development of the flavour of the cheese, as well as texture due to proteolysis, lipolysis and utilisation of lactic acid [55], [56], [57].
Dairy products have been the most utilized carrier/ formulation of bacterial probiotics with limited applications for yeasts strains [58]. Upon assessing the literature landscape, it was found that the Saccharomyces, Pichia, Candida, Meyerozyma, Debaryomyces and Kleuveromyces genera found in different types of fermented vegetables, cheese, and kefir [59], [60], [61], [62]. Additionally, Wickerhamomyces, Torulaspora, Yarrowia and Metschnikowia are the other yeast genera present in fermented fish, legumes and meat products [58], [63]. The use of dairy-based food as a means for probiotic delivery is largely affected by poor shelf-life. As a result, non-dairy based probiotic formulations have been evaluated for their potential as an alternative solution.
6.1.2. Non-Dairy Based Probiotics
It has been found that in most instances, yeasts such as S. boulardii does not naturally occur in food and usually added as a supplement. S. boulardii is commonly added into cereals and legumes to stabilise nutrients using its enzymes [53]. One of the nutrients that is present and broken down by the organisms is phytates or phytic acid which is the primary storage compound for phosphorus in seeds. This compound binds to metals rendering them insoluble and thus inaccessible as nutritional components. S. boulardii is added to synthesise the phytates using phytases which in turn enhance the bioavailability and absorption or important essential minerals such as iron, zinc, magnesium and phosphorus [53], [64], [65].
Lazo-Velez et al. [66] proposed that S. boulardii be supplemented with cereal based or low water activity foods to be used as vehicles to administer this probiotic yeast. Additionally, the application of S. boulardii has been successfully added to fermented drinks which include beers, grain drinks, malts and fruit/veggie juices [67], [68]
7. Advancements in Probiotic Delivery Systems
The administering probiotics has evolved into various methods based on the need. Typically, probiotic administration has been done through oral ingestion, however, new advances include the application of probiotics into the nostrils using nasal sprays, applications through the vagina or as topical application, termed transdermal applications.
An additional mechanism of oral supplementation includes the sublingual routes, whereby the probiotic is applied under the tongue where it is absorbed rapidly. This route of administration is currently being researched, along with the advent of rectal Suppositories or probiotic based enemas. Probiotic enemas contain a solution of probiotics, which is administered through the rectum and colon, and has been gaining popularity as an alternative remedy for gut health, immune system support, and some diseases of the digestive tract.
Other avenues being explored for probiotic administration indicate that meat and meat products are emerging as potential routes when supplemented with probiotics. Members of the Lactobacillus and Bifidobacterium genus, are the most commonly used probiotics in meat and yeasts such as S. boulardii may be explored in future research. The application of probiotics in meat products have been predominantly used as bioprotective cultures against harmful and pathogenic bacteria. This probiotic effect is mainly centred on the production of bacteriocins which aid the host [69].
8. Formulation Techniques Used for Yeast Probiotics
Probiotics administered orally transit through the mouth, stomach, small intestine, and the colon where they are subjected to saliva, acidic conditions, pancreatic juices, bile acids and digestive enzymes as well as competition with host microbiota for nutrients and adhesion sites [70]. As a result, these probiotics are shown to lose viability upon transit. This is attributed to their high sensitivity to gastrointestinal conditions, extrinsic factors such as processing techniques and storage conditions as well as intrinsic factors which include water activity, antimicrobial components and redox potential in the product matrix [71]. As stated above, a probiotic must maintain its viable activity to confer a therapeutic effect to the host. Typically, the intended viable population is targeted to be within the range of ~1.0 × 1006 as a minimum, up to a maximum of ~1.0 × 1009 CFU.ml-1 viable cells to be considered effective [72].
The challenges associated with poor survival of probiotics during processing and passage to the GIT have been studied extensively [71], [73], [74], [75], [76], [77], [78]. According to Sehrawat et al. [79], before the introduction of new technologies, there was even more challenges in the use of probiotics as starter cultures, as these preparations were used in liquid form, which was associated with low shelf-life, high risk of bacteriophage infections as well as high production and transportation costs [71], [80]. Therefore, the increased demand for use of probiotics in food and pharmaceutical industries based on their demonstrated efficacy on health and nutritional benefits has prompted the intensive research being conducted on solving these production and viability hurdles.
8.1. Immobilization
Immobilization has shown to be an effective method for the preservation of yeasts. It is typically used as a method of entrapment of bioactive materials in protective matrices, and several reports have indicated its suitability to enhance viability of many probiotic bacterial [81] and yeast cultures [82]. However, in some instances, yeast probiotics cells were found to be not entirely protected, as a small percentage of the immobilized material is still exposed to the external environment at the surface of the carrier and may as a consequence, be deemed inefficient [81].
8.2. Encapsulation
Encapsulation is one of the most utilized methods for protection of probiotics from harsh conditions and is defined as a process that involves packaging of live probiotic cells in a food-grade material such as polymers, proteins, and fats [47], [83]. In this process, encapsulated cells are contained within the coating material, which is formed continuously around an inner core matrix [81]. Additionally, these techniques improve the bioavailability of encapsulated probiotics by facilitating controlled release at the target site, the large intestine [84], [85]. Encapsulation is categorized into two classes based on particle size such as microencapsulation (3-800 µm) and nanoencapsulation (10-1000 nm). Since microbes are the size of a micron, microencapsulation is the only possible technique for encapsulating all probiotics including yeasts. Microencapsulation techniques such as extrusion, emulsion, spray drying, spray chilling, fluidized bed, freeze drying, spray-freeze drying, coacervation, and electrospraying are currently utilized to formulate probiotics [72], [85], [86].
In general, encapsulation is carried out in three steps. The initial step involves incorporation of the microbial cells into a solid or liquid matrix. The second step includes spraying and dispersion of the solid and liquid matrix, respectively. In the third phase, stabilization of the system is carried out either through physical (evaporation, solidification as well as coalescence) or chemical (polymerization) and gelation processes.
Encapsulation was suitably demonstrated for yeast probiotic applications in a study conducted by Patarroyo et al. [87] whereby Kluyveromyces lactis was encapsulated in cross-linked gelatin hydrogels, which a commercially available and relatively inexpensive material that will easily allow for industrial scale-up. The encapsulation enhanced rigidity of the final probiotic product as cell viability levels were enhanced by 50% under simulated GIT conditions [87].
Alginate, starch, k-carrageenan, chitosan, xanthan gum and cellulose acetate phthalate, gelatin, and milk proteins are some of the known polymers used in encapsulation of probiotics to date [88], [89], [90]. The encapsulating materials are selected based on their ability to stabilize the final product, non-toxicity, protective effect to the cells and possess a satisfactory control in the release of the bioactive material in the intestinal tract [85], [86]. Extrusion, spray drying, coacervation, liposomes, and emulsions are encapsulation techniques that are conventionally used in the food industry.
8.3. Extrusion
Although extrusion is largely employed in encapsulation of bacterial cells, it is a low-cost, easy technique that is carried out under mild conditions and results in high viability of encapsulated probiotics. As described by Rodrigues et al. Rodrigues et al. (2020), extrusion involves the use of hydrocolloid solutions containing microbial cultures. The mixture is then extruded through a nozzle in crosslinking solution which provides instant transition of the hydrocolloid solution to gel which results in the formation of beads [86]. These beads are stable at low pH levels and deform under alkaline conditions. In a study by Graff et al. Graff et al. (2008), S. boulardii was encapsulated with alginate microspheres coated with chitosan by extrusion. This report revealed that after 120 min at pH 1.1, encapsulated yeast cells remained entrapped in the microspheres whereas 99% of the non-encapsulated probiotic survived was lost. The authors further stated that exposure to pH 6.8 resulted in the release of viable yeast cells, demonstrating the effectiveness of this technique [91], [92], [93], [94].
8.4. Spray Drying
In spray drying, hot gas is used to atomize a liquid product into powder, instantly. It is a cost-effective and rapid microencapsulation method which results in high productivity. Spray-drying is the most common process in the food industry [85], [95]. However, the harsh conditions such as high temperature, dehydration, osmotic stress, and pressure applied during the process also pose detrimental effects to the probiotics being processed. These conditions result in alteration of cell membrane components such as fatty acids, proteins, and lipids which eventual cause cell death [96]. Improvement of cell viability during spray drying has been achieved through optimization of the process conditions and the use of lower temperature has proved to be effective as a result of reduced heat damage [86], [97].
8.5. Spray Chilling
Spray chilling is similar to spray drying as small droplets are also produced in this technique. The matrix (formed by lipids) and the encapsulated agent are dispersed by atomization in a cold air chamber which enables solidification of the particles [86]. Although it is less exploited, this process is an excellent alternative for encapsulation of probiotic due to its cost-effectiveness and applicability at industrial scale [98], [99]. In a study by Arslan-Tontul & Erbas [100], encapsulation of S. boulardii by spray drying and spray chilling using gum Arabic and b-cyclodextrin as an encapsulation material resulted in enhanced heat and survivability in the gut system [100].
8.6. Emulsions
During the preparation of emulsions, two immiscible liquids are dispersed in the presence of a stabilizing agent. An additional solidifying agent is used to separate the dispersed droplets. The emulsion is referred to as a water-in-oil (W/O) if the dispersed phase is aqueous whereas the opposite is named oil-in-water (O/W) or reverse phase. Simple emulsions are formed by two phases and addition of another phase results in double emulsions such as water-in-oil-in-water (W/O/W) or oil-in-water-in-oil (O/W/O) [85]. This technique improves solubility, activity as well as stability of immiscible compounds and it is widely employed in the food and pharmaceutical industries. This system, particularly, the dispersed aqueous phase, is mostly used in encapsulation of probiotics due to hydrophilic properties of microbial cells. In a study by Suvarna et al. [92], effects on encapsulation using emulsification were reported on four probiotic yeasts such as Pichia barkeri VIT-SJSN01, Yarrowia lipolytica VIT-ASN04, Wickerhamomyces anomalus VIT-ASN01 and Saccharomyces cerevisiae VIT-ASN03. This resulted in enhanced survival during storage and in simulated GIT conditions [92].
8.7. Fluidized Bed Drying
The fluidized bed drying technology is carried out through atomization of a coating over solid particles in suspensions. It is mainly used for coating, granulation and drying. Fluidized bed drying is a rapid, low-cost process that has high productivity [83]. This process is attractive as it allows for use of various encapsulating materials such as lipid, proteins and polysaccharides. The particles to be encapsulated are kept in constant motion due to air flow in a heated chamber. The coating particle size is reduced, forming a solid homogenous layer [86]. The ability of this technique to protect yeast cells along with the use of Hongqu rice peptides as a microencapsulation during thermal processing was investigated by [101]. it was found that the drying rate and yeast viability was significantly in comparison to free cells [101]. Another potential probiotic yeast, Meyerozyma guilliemondii Lv196 and stable granulated prototypes were reported with 0.2% loss of viability over 15 months of storage at room temperature [102], [103], [104], [105].
8.8. Supercritical Technology
Other microencapsulation techniques include supercritical technology and freeze-drying. Supercritical technology employs the use of supercritical fluids. These are solvents which describe the state of a material above its critical point at which its vapour/liquid phase equilibrium can exist. Above these conditions, the liquid-gas phase transition disappears and the properties, such as diffusion coefficient and density, continuously change with variation in pressure or temperature. Supercritical processes result in micro- or even nanoparticles with narrow size distribution and can also be used to achieve microencapsulation and surface coating of probiotics [96]. Supercritical carbon dioxide (scCO2) is one of the most used supercritical fluid due to its environmental compatibility and low reactivity and low critical parameters. in supercritical technique, the probiotic cells are first immobilized during the process of interpolymer complex formation in scCO2 and then the probiotic microcapsule is obtained by gasifying the scCO2 through depressurizing [106], [107], [108]. This technique has mainly been applied in encapsulation of probiotic bacteria.
8.9. Freeze Drying
Freeze drying is one of the well-established processes in probiotic processing. The technology involves freezing of microbial cells at extremely low temperature and drying though sublimation under high vacuum [96]. In comparison to spray drying, the operating conditions are less harsh and usually results in high survival rates. However, this process formation of extracellular crystals which results in high osmotic pressure cause cell damage. Therefore, the use of cryoprotective agents is generally applied to protect the cells. These cryoprotectants can be low molecular weight sugars such as glucose, lactose, mannose, trehalose and sorbitol or high molecular weight polysaccharides and proteins [96], [109]. This technique has been successfully applied in encapsulation of the commercially available yeast probiotic, S. boulardii. In a study by Thomas et al. [110], S. boulardii was encapsulated using layers of chitosan and dextran sulphate., whereby, the coated cells were subsequently frozen in liquid nitrogen before freeze-drying. This resulted in enhanced viability and the permeability of the encapsulated cells [110]. Illustration of the commonly used microencapsulation techniques described above as well as other methods such as refractive window drying, electro-spraying and electrospinning are outlined in Figure 5 [76].
8.10. New Advances in Probiotic Formulations
Co-encapsulation of probiotics with prebiotics and use of duocaps are emerging technologies applied to further enhance survivability and probiotic efficacy. Co-encapsulation improves the oral delivery of viable cells towards the target site. As reviewed by Rashidinejad et al. [75], various studies have been reported and several polymers and polysaccharides such as inulin, fructo-oligosaccharides and lactulose have been used. As illustrated in Figure 6, co-encapsulation provides a protective layer to probiotic cells while enhancing viability through the coexistence with a prebiotic enhances its self-proliferation [75]. Co-encapsulated particles are referred to as synbiotics. However, to the best of our knowledge, there has not been any reports on co-encapsulation of yeast probiotics.
9. Application of probiotics for preventative health benefits
9.1. Gut Microbiome Initiatives
Since the discovery of microorganisms in the 17th century; technologies and knowledge in this field have advanced rapidly, consequently resulting in microbiome mapping initiatives becoming a reality in the 21st century. Arnold et al. [111] stated that , microbiome research is an intrinsically multidisciplinary field, that has been able to reap the benefits of technological advancements in systems and synthetic biology, biomaterials engineering, and traditional microbiology. Prior to microbiome mapping, DNA technology and improvements thereof; paved the way for the advancements in whole genome sequencing and microbial population study shifts in the human body. Further advancements have resulted in the knowledge on how specific microbial compounds and activities result in health benefits, which has been developing area of research and developed [112].
The human body hosts complex microbial communities, wherein the combined membership of these organisms outnumbers our own cells by at least a factor of 10. The total number of microorganisms in the human body can reach ~100 trillion. The cells are responsible for awarding us with crucial traits which include our reliance on them to aid in nutrition, resist pathogens, and educate our immune system [113]. In comparison to other parts of the body, the human gut has the largest number of microbes, as both the gut and skin are immensely immersed with microbiota. It is estimated that the skin has about 1012 cells while the gut accounts for 1014 cells [114], [115].
The subsequent sections will focus on the gut and skin microbiome, which interestingly, share astoundingly similar characteristics as they are highly analogous to each other, both in terms of purpose and functionality [116]. According to O’Neill et al. [117], both organs are highly innervated and vascularised. Both these organs are essential for immune and neuroendocrine function. Furthermore, the inner surface of the gut and the outer surface of the skin are both covered by epithelial cells (ECs) which have direct contact with the exogenous environment [118].
According to Thursby and Juge [119], the human gastrointestinal (GI) tract represents one of the largest interfaces (250–400 m2) between the host, environmental factors and antigens in the human body. In an average lifespan, around 60 tonnes of food pass through the human GI tract. This includes an abundance of microorganisms from the environment which poses a major threat to gut integrity. The digestive process starts after the ingestion of food in the mouth where the food is grinded by teeth into smaller particles which are then emptied into the mouth. Due to the harsh environment in the stomach the microbial community that exists is at a low concentration of ~102 cells. Once food is passed from the stomach, the contents called chyme are emptied slowly into the small intestine. In the small intestine, the duodenum, jejunum, and ileum; food is mixed with digestive juices from the pancreas, liver, and intestine, and push the mixture forward for further digestion. In the small intestine the microbial community can reach between 104 to 106 cells (Figure 1). Thereafter, all non- absorbed nutrients and waste matter that was not absorbed or used; is passed into the colon where there are ~between 1012 to 1014 cells (Figure 7) [120].
The microorganisms, bacteria, archaea and eukarya, that colonise the GI tract may exert countless benefits through a wide array of physiological functions. These may include but are not limited to improving gut integrity or shaping the intestinal epithelium, protecting against pathogens, harvesting energy, and regulating host immunity. However, there is potential for these mechanisms to be disrupted because of an altered microbial composition, known as dysbiosis [121,122,123,124,125].
Thursby and Juge, (2017) also added the role of gut microbiota in human health has gained increasing attention. Recent studies has shown that diverse groups of bacteria species colonise the gut, and the composition is strongly linked to every person’s individual GI health. Additionally, there is growing evidence that indicates that by administering probiotics, the microbial ecosystem is modified, thereby exerting a variety of health benefits including a prevention and/or treatment of diseases (Gareau et al. 2010).
Microbial colonization of the GI tract mucosal tissue starts from infancy, these early life events have a long-standing consequence to the development of the human body and how it responds to diseases. During development from infancy, the developing microbiome is responsible for facilitating tolerance to environmental exposures or contributing to the development of disease, including inflammatory bowel disease, allergy, and asthma. Recent studies conducted by Gensollen et al.[126] stated that there is a critical period during early development wherein the disruption of optimal host-commensal interactions can lead to persistent and in some cases irreversible defects in the development and training of specific immune subsets.
The role of the microorganisms that form the microbiome is to facilitate metabolism, such as breaking down indigestible complex polysaccharides into essential nutrients such as vitamin K and B12, butyrate, and propionate [127]. The latter has been found to have a positive effect on the epithelial barrier integrity, which plays a crucial role in protecting microbiota from pathogenic microorganisms and avoiding inflammation in the gut [128]. Researchers that focus on the composition of the human microbiome have found that the most abundant genera of fungi in descending order of abundance in the human gut are Saccharomyces, Malassezia, and Candida [129]; with eight out of 15 genera comprising ascomycetes and approximately 5–65% comprising of Saccharomyces.
In recent microbiome studies, Saccharomyces strains have been observed in up to 96.8% of samples [130,131,132]. Since fungi are harboured in the gut environment, it follows that some resident species might provide a symbiotic benefit to the human host. The role of microbiome in the GI tract and its influence on human health has unlocked a significant area of interest, and further investigation, particularly the profiling of the African microbiome, is vital for further discovery in the development of niche treatment technologies for the global population.
9.2. Skin Microbiome
The skin is the largest and most external barrier of the body with the outer environment; therefore, it is considered the external interface between the body and the environment [133]. The skin is richly perfused with immune cells and heavily colonized by microbial cells, which in turn, trains immune cells and determines the well-being of the host. Also, it is worth noting that despite the skin covering many areas of the human body, the population and microbial concentration differs per area. It has also been found that a shift in population can also shift depending on the external environment, disease and diet [134].
The skin epidermis, including sweat and sebaceous glands, provide a total skin surface of about 25 m2 and forms one of the largest epithelial surfaces for interaction with microbes [116]. Epithelial cells cover the surfaces of the body such as skin, airways, or the intestinal tract and provide an important link between the outside environment and the body interior [135].
Like those in our gut, skin microorganisms have important functions in the protection against invading pathogens, the teaching of our immune system and the breakdown of natural products [136,137,138,139]. According to Byrd et al. [140], several skin microbiome assessment surveys have to date, used amplicon sequencing, however in recent years; major technical breakthroughs have occurred, which uses shotgun metagenomic sequencing. The advantage of using the latter approach is that it does not sequence specific target regions. This technique simultaneously captures all genetic material in a sample, including human, bacterial, fungal, archaeal and viral microorganisms, providing vital information on the microbial composition.
9.3. Case Studies Assessing the Use of Yeast Probiotics and Its Impact on the Host Microbiome
The microbiome of a healthy individual consists of balanced populations of both beneficial and harmful microorganisms [141]. These play a major role in providing the host with physiological, metabolic, and immune functions useful in warding off pathogens and any imbalance results in increased levels of harmful microbes. There is a mutual relationship between the gut and human flora. The colon harbors the highest population, however, only < 0.1% of these are fungi and Saccharomyces and Candida are the dominant genera [142,143,144]. As the most commercialized probiotic yeast, S. boulardii is widely used in treatment of gut-related diseases such as Traveler’s diarrhea, AIDS-associated syndrome, irritable bowel syndrome and Crohn’s disease. Oral administration of S. boulardii alone or in combination with other probiotics has proven to induce changes in the gut microbial combinations in various clinical reports [143,145,146,147]. S. boulardii influences the host microbiome by direct inhibition of pathogenic intestinal microbes and normalizing the pH of the gastrointestinal tract, this is achieved by reducing the pathogenicity of toxic microorganisms [144].
A recent study representing the effect of the use of S, boulardii on the gut microbiota was reported by Yang et al. Yang et al. (2022). The potential of this probiotic yeast in treatment of non-alcoholic steatohepatitis (NASH) in mice through gut-liver axis was demonstrated. NASH is a non-alcoholic fatty liver disease associated with inflammation, damage, and presence of excess fat in the liver. Yang et al. [149], fed NASH-inducing diet [Methionine-choline-deficient (MCD)] to all test mice and the control group was given normal chow diet (NCD). Florastor®, a commercial product containing lyophilized S. boulardii CNCM I-745 as a main component was also administered by gavage to random mice (both on MCD diet and the control group) five days a week. After 8 weeks, the mouse fecal genomic DNA was extracted, sequenced, and analyzed. The positive effect of administering S. boulardii to MCD diet-fed mice was evident (summarized in Table 4) by the microbial composition presented at family level [148]. It was concluded that this probiotic played a role in restoring the gut microbiome diversity that was reduced by the diet. Additionally, the MCD diet resulted in changes in the mycobiome, dominated by Pichia and Trichosporon. This was an indication of the robustness of these fungal genera under severe conditions in the gut [148]. Furthermore, the positive impacts of the gut mycobiota on regulating functions of other human organs such as brain, pancreas, liver, and kidney as well as overall host immunity towards intestinal and extraintestinal diseases has been comprehensively reviewed by [150].
Co-supplementation of multi-strain probiotic has also shown to have even outstanding benefits. The World Health Organisation stated that “mental health is critically important for everyone, everywhere” (WHO, 2002), the positive impacts of probiotics in cognitive performance were reported by Bloemendaal et al. [152]. This was determined by the increase in population of plant fibre degrading bacteria that produce short-chain fatty acids which are known for their beneficial effect on gut and brain health [152].
In another study, benefits of co-supplementation of bacterial (Lacticasebacillus rhamnosus) and fungal (S. boulardii) probiotics protected the gut microbiome post antibiotic administration in vitro [153]. Here, the human intestinal ecosystem was simulated using SHIME model. Three regions of the gastrointestinal tract were represented, upper part, proximal and distal colon. Mucin-covered mucosms were included in the proximal colon to simulate luminal mucus-associated microbiota and the parameters in the reactors were stabilized for two weeks. The study involved two healthy human adults who consented to give feacal samples. After inoculation, baseline conditions were established and then a 5-day antibiotic (amoxicillin and clavulanic acid) treatment was initiated. The study was conducted in parallel where one set was dosed with probiotics (L. rhamnosus and S. boulardii). Composition of the gut microbiota was then profiled. Although, the overall population was donor-dependent, there was a clear protective impact of the yeast probiotic towards L. rhamnosus against antibiotics. Furthermore, the presence of each or both probiotics significantly enhanced abundance of other Lactobacillaceae, Bifidobacteriaceae and Lachnospiraceae. This demonstrated the ability of probiotics to restore, stimulate and strengthen the composition as well as functionality of the microbial community negatively impacted by the use of antibiotics [153].
Functionality of yeasts as probiotics are not only limited to their use as whole cells therapeutics. A review conducted by Saber et al. [153], indicated that their metabolic by-products, such as folic acid and β-glucan may have an effect on cancerous cells, by being able to affect pathogenic bacteria, inactivate carcinogenic compounds particularly those derived from food, being able to improve intestinal barrier function, modulate host immune responses, antitoxic functions, apoptosis and anti-proliferative effects [4].
9.4. The Use of Yeast Probiotics in Skin Applications
Further to the limited instances of the use of yeasts as probiotics, employed as food supplements and/or additives, there are lesser studies that focus on the use of these organisms for skin applications. This scantity could be due to the fact that, S. boulardii the most commonly studied yeast probiotic, is mostly active in the colon and can grow at low pH levels (2.0-3.0); whereas the skin pH is 5.5. Other yeast genera such as Candida, Cryptococcus, Epidermophyton, Hortaea, Malassezia, Microsporum and Trichophyton are well-known for causing vaginal yeast infections, athlete’s foot, jock itch, ringworm or thrush owing to their ability to ability to penetrate tissues [154], and limited report of beneficial yeasts are limited in contrast.
10. Conclusionary Remarks and Future Prospects
The use of probiotics has gained significant momentum in terms of advocation for use amongst the global population. Advent of genetic engineered probiotics may be more effective, cheaper production costs, higher stability and specificity for the treatment of a plethora of human ailments and disorders [3]. Yet, the application of these GMOs faces significant hurdles, particularly in terms of biosafety considerations upon ingestion by the host. Several clinical trials have been conducted to date; however, their effect cannot be guaranteed to achieve their intended effect, and therefore prevents effective deployment. With more in depth understanding into the human microbiome and its relation to disease mechanisms, the safety and endorsement of engineered probiotics, both bacterial and yeast, may gain acceptance for use, particularly, when conventional health strategies prove ineffective [3]. Additionally, in terms of advances in probiotic production and formulation, significant strides have been made to deliver highly efficacious probiotic treatments for the treat of several human metabolic disorders.
References
- C. Hill et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed]
- C. Altieri, ‘Dairy propionibacteria as probiotics: recent evidences. World J Microbiol Biotechnol 2016, 32. [Google Scholar] [CrossRef]
- J. Ma et al. Engineered probiotics’, Microb Cell Fact 2022, 21, 72. [CrossRef]
- Saber, B. Saber, B. Alipour, Z. Faghfoori, and A. Yari Khosroushahi, ‘Cellular and molecular effects of yeast probiotics on cancer. 2016, 43, 96–115. [Google Scholar] [CrossRef] [PubMed]
- M. Arevalo-Villena, A. Briones-Perez, M. R. Corbo, M. Sinigaglia, and A. Bevilacqua, ‘Biotechnological application of yeasts in food science: Starter cultures, probiotics and enzyme production’. J Appl Microbiol 2017, 123, 1360–1372. [CrossRef]
- S. Sen and T. J. Mansell, ‘Yeasts as probiotics: Mechanisms, outcomes, and future potential’, Fungal Genetics and Biology 2020, 137. [CrossRef]
- D. Czerucka, T. Piche, and P. Rampal, ‘Review article: yeast as probiotics –Saccharomyces boulardii’. Aliment Pharmacol Ther 2007, 26, 767–778. [CrossRef]
- H. S. ßanlidere, A. Gl U, E. Demir, € Oz, and Z. € Ub E Y D E € On, ‘Assimilation of cholesterol and probiotic characterisation of yeast strains isolated from raw milk and fermented foods. Wiley Online Library 2015, 69, 63–70. [CrossRef]
- Al-Seraih, C. Flahaut, F. Krier, B. Cudennec, and D. Drider, ‘Characterization of Candida famata Isolated from Poultry Feces for Possible Probiotic Applications’. Probiotics Antimicrob Proteins 2015, 7, 233–241. [CrossRef]
- R. Ogunremi, A. I. Sanni, and R. Agrawal, ‘Probiotic potentials of yeasts isolated from some cereal-based Nigerian traditional fermented food products. J Appl Microbiol 2015, 119, 797–808. [CrossRef]
- H. S. Ochangco, A. H. S. Ochangco, A. Gamero, I. M. Smith, J. E. Christensen, L. Jespersen, and N. Arneborg, ‘In vitro investigation of Debaryomyces hansenii strains for potential probiotic properties’, World J Microbiol Biotechnol 2016, 32. [CrossRef]
- Binetti, M. Carrasco, J. Reinheimer, and V. Suárez, ‘Yeasts from autochthonal cheese starters: technological and functional properties’. J Appl Microbiol 2013, 115, 434–444. [CrossRef]
- M. Smith, A. Baker, N. Arneborg, and L. Jespersen, ‘Non-Saccharomyces yeasts protect against epithelial cell barrier disruption induced by Salmonella enterica subsp. enterica serovar Typhimurium. Lett Appl Microbiol 2015, 61, 491–497. [CrossRef] [PubMed]
- G. Diosma, D. E. Romanin, M. F. Rey-Burusco, A. Londero, and G. L. Garrote, ‘Yeasts from kefir grains: Isolation, identification, and probiotic characterization. World J Microbiol Biotechnol 2014, 30, 43–53. [CrossRef]
- R. C. França et al. Pichia pastoris X-33 has probiotic properties with remarkable antibacterial activity against Salmonella Typhimurium. Appl Microbiol Biotechnol 2015, 99, 7953–7961. [CrossRef]
- S. Bonatsou, A. Benítez, F. Rodríguez-Gómez, E. Z. Panagou, and F. N. Arroyo-López, ‘Selection of yeasts with multifunctional features for application as starters in natural black table olive processing’. Food Microbiol 2015, 46, 66–73. [CrossRef] [PubMed]
- D. H. Kitamura, L. P. de Souza Vandenberghe, C. Rodrigues, D. N. X. Salmon, G. V. de Melo Pereira, and C. R. Soccol, ‘Selenium-Enriched Probiotic Saccharomyces boulardii CCT 4308 Biomass Production Using Low-Cost Sugarcane Molasses Medium’. Brazilian Archives of Biology and Technology 2021, 64, 1–14. [CrossRef]
- Saber, B. Alipour, Z. Faghfoori, and A. Yari Khosroushahi, ‘Cellular and molecular effects of yeast probiotics on cancer’. Crit Rev Microbiol 2017, 43, 96–115. [Google Scholar] [CrossRef] [PubMed]
- S. Sen and T. J. Mansell, ‘Yeasts as probiotics: Mechanisms, outcomes, and future potential’. Fungal Genetics and Biology 2020, 137, 103333. [CrossRef]
- M. Arevalo-Villena, A. Briones-Perez, M. R. Corbo, M. Sinigaglia, and A. Bevilacqua, ‘Biotechnological application of yeasts in food science: Starter cultures, probiotics and enzyme production’. J Appl Microbiol 2017, 123, 1360–1372. [CrossRef]
- Papadimitriou, *!!! REPLACE !!!*; et al. Discovering probiotic microorganisms: in vitro, in vivo, genetic and omics approaches’, Front Microbiol 2015, 6. [CrossRef]
- T. Trunk, H. S. Khalil, and J. C. Leo, ‘Bacterial autoaggregation. AIMS Microbiol 2018, 4, 140–164. [CrossRef]
- Javed; et al. In Silico and In Vitro Analysis of Helicobacter pullorum Type Six Secretory Protein Hcp and Its Role in Bacterial Invasion and Pathogenesis’, Curr Microbiol 2022, 79. [CrossRef]
- Pillinger, B. Weber, B. Standen, M. C. Schmid, and J. C. Kesselring, ‘Multi-strain probiotics show increased protection of intestinal epithelial cells against pathogens in rainbow trout (Oncorhynchus mykiss)’. Aquaculture 2022, 560, 738487. [CrossRef]
- H. Ziar and A. Riazi, ‘Polysorbate 80 improves the adhesion and survival of yogurt starters with cholesterol uptake abilities’. Saudi J Biol Sci 2022, 29, 103367. [CrossRef]
- G. Barzoki, S. S. Malekshahi, and M. Shayestehpour, ‘In vitro evaluation of antiviral activity of Shouchella clausii probiotic strain and bacterial supernatant against herpes simplex virus type 1’. Arch Microbiol 2022, 204, 522. [CrossRef]
- Fakruddin, M. N. Hossain, and M. M. Ahmed, ‘Antimicrobial and antioxidant activities of Saccharomyces cerevisiae IFST062013, a potential probiotic’, BMC Complement Altern Med, 2017; 17, 1–11. [Google Scholar]
- Halász and, R. Lásztity, ‘Use of yeast biomass in food production’, Use of Yeast Biomass in Food Production, 1–312, 2017.
- R. E. Speight, L. Navone, L. K. Gebbie, J.-A. L. Blinco, and W. L. Bryden, ‘Platforms to accelerate biomanufacturing of enzyme and probiotic animal feed supplements: discovery considerations and manufacturing implications’. Anim Prod Sci 2022, 62, 1113–1128. [CrossRef]
- B. Hahn-Hägerdal, K. Karhumaa, C. U. Larsson, M. Gorwa-Grauslund, J. Görgens, and W. H. van Zyl, ‘Role of cultivation media in the development of yeast strains for large scale industrial use’, Microb Cell Fact 2005, 4. [CrossRef]
- D. H. Kitamura, L. P. de S. Vandenberghe, C. Rodrigues, D. N. X. Salmon, G. V. de M. Pereira, and C. R. Soccol, ‘Selenium-Enriched Probiotic Saccharomyces boulardii CCT 4308 Biomass Production Using Low-Cost Sugarcane Molasses Medium’, Brazilian Archives of Biology and Technology 2021, 64. [CrossRef]
- O’Toole, J. Marchesi, C. H.-N. microbiology, and undefined 2017, ‘Next-generation probiotics: the spectrum from probiotics to live biotherapeutics’, nature.com, 2017. [CrossRef]
- J. Ma et al. Engineered probiotics. Microb Cell Fact 2022, 21, 72. [CrossRef]
- Anadón, M. Rosa Martínez-Larrañaga, and M. Aranzazu Martínez, ‘Probiotics for animal nutrition in the European Union. Regulation and safety assessment’. Regulatory Toxicology and Pharmacology 2006, 45, 91–95. [CrossRef]
- W. O’Toole, J. R. Marchesi, and C. Hill, Next-generation probiotics: the spectrum from probiotics to live biotherapeutics. Nat Microbiol 2017, 2, 17057. [CrossRef]
- de Simone, ‘The Unregulated Probiotic Market’. Clinical Gastroenterology and Hepatology 2019, 17, 809–817. [CrossRef] [PubMed]
- V. Venugopalan, K. A. Shriner, and A. Wong-Beringer, ‘Regulatory Oversight and Safety of Probiotic Use’. Emerg Infect Dis 2010, 16, 1661–1665. [CrossRef]
- H. Nasri, A. Baradaran, H. Shirzad, and M. Rafieian-Kopaei, ‘New Concepts in Nutraceuticals as Alternative for Pharmaceuticals’. Int J Prev Med 2014, 5, 1487–1499.
- S. Fujimori, ‘Gastric acid level of humans must decrease in the future’. World J Gastroenterol 2020, 26, 6706–6709. [CrossRef]
- Zhai, F. Tian, J. Zhao, H. Zhang, A. Narbad, and W. Chen, ‘Oral administration of probiotics inhibits absorption of the heavy metal cadmium by protecting the intestinal barrier’, Appl Environ Microbiol, 2016; 82, 4429–4440. [Google Scholar] [CrossRef]
- Lebeer; et al. Oral Administration of Probiotics Increases Paneth Cells and Intestinal Antimicrobial Activity’, 2018. [CrossRef]
- F. Hendijani and V. Akbari, ‘Probiotic supplementation for management of cardiovascular risk factors in adults with type II diabetes: A systematic review and meta-analysis’. Clinical Nutrition 2018, 37, 532–541. [CrossRef] [PubMed]
- G. Cifuentes, M. B. Prado, M. Fornasini, H. Cohen, M. E. Baldeón, and P. A. Cárdenas, ‘Saccharomyces boulardii CNCM I-745 supplementation modifies the fecal resistome during Helicobacter pylori eradication therapy’, Helicobacter 2022, 27. [CrossRef]
- M. R. Corbo, D. Campaniello, B. Speranza, C. Altieri, M. Sinigaglia, and A. Bevilacqua, ‘Neutralisation of toxins by probiotics during the transit into the gut: challenges and perspectives’. Int J Food Sci Technol 2018, 53, 1339–1351. [CrossRef]
- P. Hosseinzadeh et al. Brewer’s Yeast Improves Glycemic Indices in Type 2 Diabetes Mellitus’. Int J Prev Med 2013, 4, 1131–1138.
- Y. Zhu, J. Jiang, Y. Yue, Z. Feng, J. Chen, and X. Ye, ‘Influence of mixed probiotics on the the bioactive composition, antioxidant activity and appearance of fermented red bayberry pomace’. LWT 2020, 133, 110076. [CrossRef]
- Staniszewski and M. Kordowska-Wiater, ‘Probiotic and Potentially Probiotic Yeasts—Characteristics and Food Application’. Foods 2021, 10, 1306. [CrossRef] [PubMed]
- J. C. Amorim, R. H. Piccoli, and W. F. Duarte, ‘Probiotic potential of yeasts isolated from pineapple and their use in the elaboration of potentially functional fermented beverages’. Food Research International 2018, 107, 518–527. [CrossRef]
- B. Senkarcinova, I. A. Graça Dias, J. Nespor, and T. Branyik, ‘Probiotic alcohol-free beer made with Saccharomyces cerevisiae var. boulardii’. LWT 2019, 100, 362–367. [CrossRef]
- H.-J. Chen et al. Transdermal Delivery of Living and Biofunctional Probiotics through Dissolvable Microneedle Patches’. ACS Appl Bio Mater 2018, 1, 374–381. [CrossRef]
- M. Govender, Y. E. Choonara, P. Kumar, L. C. du Toit, S. van Vuuren, and V. Pillay, ‘A Review of the Advancements in Probiotic Delivery: Conventional vs. Non-conventional Formulations for Intestinal Flora Supplementation’. AAPS PharmSciTech 2014, 15, 29–43. [CrossRef]
- Y. B. Soemarie, T. Milanda, and M. I. Barliana, ‘Fermented foods as probiotics: A review’. J Adv Pharm Technol Res 2021, 12, 335–339. [CrossRef]
- M. A. Lazo-Velez, S. O. Sern-Salvidar, M. F. Rosales-Medina, M. Tinoco-Alvaear, and M. Briones-Garcia, ‘Application of Saccharomyces cerevisiae var. boulardii in food processing: a review’. J Appl Microbiol 2017, 125, 943–951.
- Parrella; et al. Original article Antioxidant properties of different milk fermented with lactic acid bacteria and yeast’, Int J Food Sci Technol, 1–10, 2012. [CrossRef]
- G. H. Fleet, ‘Yeasts in foods and beverages : impact on product quality and safety’. Current Opinion in Biotechnology 2007 2007, 18, 170–175. [CrossRef]
- E. Addis, G. H. Fleet, J. M. Cox, D. Kolak, and T. Leung, ‘The growth, properties and interactions of yeasts and bacteria associated with the maturation of Camembert and blue-veined cheeses’, 25–36, 2001.
- R. Zamora-Vega et al. Effect of incorporating prebiotics in coating materials for the microencapsulation of Sacharomyces boulardii’. International Journal ofFood Sciences and Nutrition 2012, 63, 930–935. [CrossRef] [PubMed]
- J. P. Tamang and S. Lama, ‘Probiotic properties of yeasts in traditional fermented foods and beverages’. J Appl Microbiol 2022, 132, 3533–3542. [CrossRef] [PubMed]
- L. A. Simões et al. Probiotic properties of yeasts isolated from Brazilian fermented table olives. J Appl Microbiol 2021, 131, 1983–1997. [CrossRef] [PubMed]
- v Merchán, M. J. Benito, A. I. Galván, and S. Ruiz-Moyano Seco de Herrera, ‘Identification and selection of yeast with functional properties for future application in soft paste cheese’. LWT 2020, 124, 109173. [CrossRef]
- G. T. Menezes, C. L. Ramos, G. Cenzi, D. S. Melo, D. R. Dias, and R. F. Schwan, ‘Probiotic Potential, Antioxidant Activity, and Phytase Production of Indigenous Yeasts Isolated from Indigenous Fermented Foods’. Probiotics Antimicrob Proteins 2020, 12, 280–288. [CrossRef]
- R.-T. Hsiung, W.-T. Fang, B. A. LePage, S.-A. Hsu, C.-H. Hsu, and J.-Y. Chou, ‘In Vitro Properties of Potential Probiotic Indigenous Yeasts Originating from Fermented Food and Beverages in Taiwan’. Probiotics Antimicrob Proteins 2021, 13, 113–124. [CrossRef]
- W. Fu, W. Xue, C. Liu, Y. Tian, K. Zhang, and Z. Zhu, ‘Screening of Lactic Acid Bacteria and Yeasts from Sourdough as Starter Cultures for Reduced Allergenicity Wheat Products’. Foods 2020, 9, 751. [CrossRef]
- L. Bohn, A. S. Meyer, and S. K. Rasmussen, ‘Phytate : impact on environment and human nutrition . A challenge for molecular breeding *’ 2008, 9, 165–191. [CrossRef]
- K. Küçükgöz and M. Trzaskowska, ‘Nondairy Probiotic Products : Functional Foods That Require more attention. Nutrients 2022, 14, 753.
- M. A. Lazo-Vélez, S. O. Serna-Saldívar, M. F. Rosales-Medina, M. Tinoco-Alvear, and M. Briones-García, ‘Application of Saccharomyces cerevisiae var. boulardii in food processing: a review’, J Appl Microbiol 2018, 125, 943–951. [CrossRef]
- P. De Paula, D. William, H. Chávez, and L. Lin, ‘Growth Parameters and Survivability of Saccharomyces boulardii for Probiotic Alcoholic Beverages Development’ 2019, 10, no. September, 1–10. [CrossRef]
- Senkarcinova, I. Alexandra, G. Dias, J. Nespor, and T. Branyik, ‘LWT - Food Science and Technology Probiotic alcohol-free beer made with Saccharomyces cerevisiae var . boulardii’. LWT - Food Science and Technology 2019, 100, 362–367. [CrossRef]
- P. Cavalheiro, C. P. Cavalheiro, C. Ruiz-capillas, A. M. Herrero, C. R. De Menezes, L. Lucy, and M. Fries, ‘Application of probiotic delivery systems in meat products’, Trends Food Sci Technol, 2015. [CrossRef]
- Han; et al. Probiotic Gastrointestinal Transit and Colonization After Oral Administration: A Long Journey’, Front Cell Infect Microbiol 2021, 11. [CrossRef]
- R. Sehrawat, S. Abdullah, P. Khatri, L. Kumar, A. Kumar, and A. S. Mujumdar, ‘Role of drying technology in probiotic encapsulation and impact on food safety’. Drying Technology 2022, 40, 1562–1581. [CrossRef]
- J. Palanivelu, S. Thanigaivel, S. Vickram, N. Dey, D. Mihaylova, and I. Desseva, ‘Probiotics in Functional Foods: Survival Assessment and Approaches for Improved Viability’. Applied Sciences 2022, 12, 455. [CrossRef]
- M. A. S. Santos and M. T. C. Machado, ‘Coated alginate–chitosan particles to improve the stability of probiotic yeast’. Int J Food Sci Technol 2021, 56, 2122–2131. [CrossRef]
- R. Rajam and P. Subramanian, ‘Encapsulation of probiotics: past, present and future’. Beni Suef Univ J Basic Appl Sci 2022, 11, 46. [CrossRef]
- Rashidinejad; et al. Co-encapsulation of probiotics with prebiotics and their application in functional/synbiotic dairy products’, Crit Rev Food Sci Nutr 2022, 62, 2470–2494. 62. [CrossRef]
- K. S. Yoha, S. Nida, S. Dutta, J. A. Moses, and C. Anandharamakrishnan, ‘Targeted Delivery of Probiotics: Perspectives on Research and Commercialization’. Probiotics Antimicrob Proteins 2022, 14, 15–48. [CrossRef]
- J. Cielecka-Piontek, M. Dziedziński, O. Szczepaniak, J. Kobus-Cisowska, A. Telichowska, and D. Szymanowska, ‘Survival of commercial probiotic strains and their effect on dark chocolate synbiotic snack with raspberry content during the storage and after simulated digestion’. Electronic Journal of Biotechnology 2020, 48, 62–71. [CrossRef]
- M. Z. A. Chan, M. Toh, and S.-Q. Liu, ‘Growth, survival, and metabolic activities of probiotics Lactobacillus rhamnosus GG and Saccharomyces cerevisiae var. boulardii CNCM-I745 in fermented coffee brews’. Int J Food Microbiol 2021, 350, 109229. [CrossRef]
- R. Sehrawat, S. Abdullah, P. Khatri, L. Kumar, A. Kumar, and A. S. Mujumdar, ‘Role of drying technology in probiotic encapsulation and impact on food safety’, Drying Technology, 1–20, 2022. [CrossRef]
- G. Frakolaki, V. Giannou, D. Kekos, and C. Tzia, ‘A review of the microencapsulation techniques for the incorporation of probiotic bacteria in functional foods’. Crit Rev Food Sci Nutr 2021, 61, 1515–1536. [CrossRef]
- M. Kvakova, I. Bertkova, J. Stofilova, and T. C. Savidge, ‘Co-Encapsulated Synbiotics and Immobilized Probiotics in Human Health and Gut Microbiota Modulation’. Foods 2021, 10, 1297. [CrossRef]
- Ohlmaier-Delgadillo; et al. Ferulated Pectins and Ferulated Arabinoxylans Mixed Gel for Saccharomyces boulardii Entrapment in Electrosprayed Microbeads’, Molecules 2021, 26, 2478. 26. [CrossRef]
- M. Chavarri, I. M. Chavarri, I. Maranon, and M. Carmen, ‘Encapsulation Technology to Protect Probiotic Bacteria’, in Probiotics, InTech, 2012. [CrossRef]
- Terpou, A. Papadaki, I. Lappa, V. Kachrimanidou, L. Bosnea, and N. Kopsahelis, ‘Probiotics in Food Systems: Significance and Emerging Strategies Towards Improved Viability and Delivery of Enhanced Beneficial Value’. Nutrients 2019, 11, 1591. [Google Scholar] [CrossRef] [PubMed]
- P. M. Reque and A. Brandelli, ‘Encapsulation of probiotics and nutraceuticals: Applications in functional food industry’. Trends Food Sci Technol 2021, 114, 1–10. [CrossRef]
- J. Rodrigues, M. F. Cedran, J. L. Bicas, and H. H. Sato, ‘Encapsulated probiotic cells: Relevant techniques, natural sources as encapsulating materials and food applications – A narrative review’. Food Research International 2020, 137, 109682. [CrossRef]
- J. L. Patarroyo, J. S. Florez-Rojas, D. Pradilla, J. D. Valderrama-Rincón, J. C. Cruz, and L. H. Reyes, ‘Formulation and Characterization of Gelatin-Based Hydrogels for the Encapsulation of Kluyveromyces lactis—Applications in Packed-Bed Reactors and Probiotics Delivery in Humans’. Polymers (Basel) 2020, 12, 1287. [CrossRef]
- de la C. Pech-Canul, D. Ortega, A. García-Triana, N. González-Silva, and R. L. Solis-Oviedo, ‘A Brief Review of Edible Coating Materials for the Microencapsulation of Probiotics’. Coatings 2020, 10, 197. [CrossRef]
- L.-F. Călinoiu, B. Ştefănescu, I. Pop, L. Muntean, and D. Vodnar, ‘Chitosan Coating Applications in Probiotic Microencapsulation’. Coatings 2019, 9, 194. [CrossRef]
- Z. Muhammad, R. Ramzan, R. Zhang, and M. Zhang, ‘Resistant Starch-Based Edible Coating Composites for Spray-Dried Microencapsulation of Lactobacillus acidophilus, Comparative Assessment of Thermal Protection, In Vitro Digestion and Physicochemical Characteristics’. Coatings 2021, 11, 587. [CrossRef]
- S. Graff, S. Hussain, J.-C. Chaumeil, and C. Charrueau, ‘Increased Intestinal Delivery of Viable Saccharomyces boulardii by Encapsulation in Microspheres’. Pharm Res 2008, 25, 1290–1296. [CrossRef]
- S. Suvarna, J. Dsouza, M. L. Ragavan, and N. Das, ‘Potential probiotic characterization and effect of encapsulation of probiotic yeast strains on survival in simulated gastrointestinal tract condition’, Food Sci Biotechnol 2018, 27, 745–753. [CrossRef]
- E. Dadkhodazade, E. Khanniri, N. Khorshidian, S. M. Hosseini, A. M. Mortazavian, and E. Moghaddas Kia, ‘Yeast cells for encapsulation of bioactive compounds in food products: A review’, Biotechnol Prog, 2021. [CrossRef]
- B. N. Pham-Hoang, C. Romero-Guido, H. Phan-Thi, and Y. Waché, ‘Encapsulation in a natural, preformed, multi-component and complex capsule: yeast cells. Appl Microbiol Biotechnol 2013, 97, 6635–6645. [CrossRef] [PubMed]
- D. Poddar, J. Palmer, S. Das, M. Gaare, A. Nag, and H. Singh, ‘Effect of Fluidized Bed Drying, Matrix Constituents and Structure on the Viability of Probiotic Lactobacillus paracasei ATCC 55544 during Storage at 4 °C, 25 °C and 37 °C’. Microorganisms 2021, 10, 74. [CrossRef]
- Liu; et al. Protective approaches and mechanisms of microencapsulation to the survival of probiotic bacteria during processing, storage and gastrointestinal digestion: A review’, Crit Rev Food Sci Nutr 2019, 59, 2863–2878. [CrossRef]
- D.-H. Lim, A. Letona, M. Lee, D. Lim, N.-S. Han, and D. Chung, ‘Fluidized-Bed Granulation of Probiotics-Encapsulated Spray-Dried Skim Milk Powder: Effects of a Fluidizing Aid, Moisture-Activation and Dehydration’. Foods 2021, 10, 1600. [CrossRef]
- S. P. Dhakal and J. He, ‘Microencapsulation of vitamins in food applications to prevent losses in processing and storage: A review’. Food Research International 2020, 137, 109326. [CrossRef] [PubMed]
- D. Arepally, R. S. Reddy, T. K. Goswami, and R. Coorey, ‘A Review on Probiotic Microencapsulation and Recent Advances of their Application in Bakery Products’. Food Bioproc Tech 2022, 15, 1677–1699. [CrossRef]
- S. Arslan-Tontul and M. Erbas, ‘Single and double layered microencapsulation of probiotics by spray drying and spray chilling’. LWT - Food Science and Technology 2017, 81, 160–169. [CrossRef]
- B. Chen, Z. Liang, X. Lin, W. Li, X. Lin, and Z. He, ‘Enhanced survival of fluidized bed-dried microencapsulated Saccharomyces cerevisiae cells in the presence of Hongqu rice distiller’s grain peptides. LWT 2022, 163, 113511. [CrossRef]
- M. L. Chaparro, E. Ce´spedes, M. Cruz, C. R. Castillo-Saldarriaga, and M. I. Go´mez-A´ lvarez, ‘FLUIDIZED BED DRYING OF A GRANULATED PROTOTYPE BASED ON A POTENTIAL PROBIOTIC YEAST Meyerozyma guilliermondii: SELECTION OF PROCESS PARAMETERS AND DRYING PROTECTANT’. Rev Mex Ing Quim 2017, 16, 347–357.
- S. Hirai and T. Kawasumi, ‘Enhanced lactic acid bacteria viability with yeast coincubation under acidic conditions’. Biosci Biotechnol Biochem 2020, 84, 1706–1713. [CrossRef]
- F. Zahoor, C. Sooklim, P. Songdech, O. Duangpakdee, and N. Soontorngun, ‘Selection of Potential Yeast Probiotics and a Cell Factory for Xylitol or Acid Production from Honeybee Samples’. Metabolites 2021, 11, 312. [CrossRef]
- X. Mo et al. Whole genome sequencing and metabolomics analyses reveal the biosynthesis of nerol in a multi-stress-tolerant. Meyerozyma guilliermondii 2021, 20, 1–11. [CrossRef]
- Aro, E. Järvenpää, J. Mäkinen, M. Lauraeus, R. Huopalahti, and V. Hietaniemi, ‘The utilization of oat polar lipids produced by supercritical fluid technologies in the encapsulation of probiotics’. LWT - Food Science and Technology 2013, 53, 540–546. [Google Scholar] [CrossRef]
- M. S. Thantsha, P. W. Labuschagne, and C. I. Mamvura, ‘Supercritical CO2 interpolymer complex encapsulation improves heat stability of probiotic bifidobacteria. World J Microbiol Biotechnol 2014, 30, 479–486. [CrossRef] [PubMed]
- P. W. Labuschagne, B. Naicker, and L. Kalombo, ‘Micronization, characterization and in-vitro dissolution of shellac from PGSS supercritical CO2 technique. Int J Pharm 2016, 499, 205–216. [CrossRef]
- L. Cao, Q. Xu, Y. Xing, X. Guo, W. Li, and Y. Cai, ‘Effect of skimmed milk powder concentrations on the biological characteristics of microencapsulated Saccharomyces cerevisiae by vacuum-spray-freeze-drying. Drying Technology 2020, 38, 476–494. [CrossRef]
- M. Ben Thomas, M. Vaidyanathan, K. Radhakrishnan, and A. M. Raichur, ‘Enhanced viability of probiotic Saccharomyces boulardii encapsulated by layer-by-layer approach in pH responsive chitosan–dextran sulfate polyelectrolytes’. J Food Eng 2014, 136, 1–8. [CrossRef]
- W. Arnold, J. Roach, and M. A. Azcarate-Peril, ‘Emerging Technologies for Gut Microbiome Research’. Trends Microbiol 2016, 24, 887–901. [CrossRef]
- M. L. Marco, ‘Crystal ball Defining how microorganisms benefit human health’. Microb Biotechnol 2020, 141, 35–40. [CrossRef]
- E. K. Costello, C. L. Lauber, M. Hamady, N. Fierer, J. I. Gordon, and R. Knight, ‘Bacterial community variation in human body habitats across space and time’. Science (1979) 2009, 326, 1694–1697. [CrossRef]
- E. T. Hillman, H. Lu, T. Yao, and C. H. Nakatsu, ‘Microbial Ecology along the Gastrointestinal Tract’. Microbes Environ 2017, 32, 300. [CrossRef]
- R. Khan, F. C. Petersen, and S. Shekhar, ‘Commensal bacteria: An emerging player in defense against respiratory pathogens’. Front Immunol 2019, 10, 1203. [CrossRef] [PubMed]
- B. De Pessemier et al. microorganisms Gut-Skin Axis: Current Knowledge of the Interrelationship between Microbial Dysbiosis and Skin Conditions’, 2021. [CrossRef]
- C. A. O’Neill, G. Monteleone, J. T. McLaughlin, and R. Paus, ‘The gut-skin axis in health and disease: A paradigm with therapeutic implications. BioEssays 2016, 38, 1167–1176. [CrossRef] [PubMed]
- R. Shaykhiev and R. Bals, ‘Interactions between epithelial cells and leukocytes in immunity and tissue homeostasis. J Leukoc Biol 2007, 82, 1–15. [CrossRef] [PubMed]
- E. Thursby and N. Juge, ‘Introduction to the human gut microbiota’, Biochemical Journal 2017, 474, 1823–1836. [CrossRef]
- G. A. M. Cresci and K. Izzo, ‘Gut microbiome’, Adult Short Bowel Syndrome: Nutritional, Medical, and Surgical Management, 45–54, 2018. [CrossRef]
- Gensollen, S. S. Iyer, D. L. Kasper, and R. S. Blumberg, ‘How colonization by microbiota in early life shapes the immune system’. Science (1979) 2016, 352, 539–544. [Google Scholar] [CrossRef]
- G. Den Besten, K. Van Eunen, A. K. Groen, K. Venema, D. J. Reijngoud, and B. M. Bakker, ‘The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism’. J Lipid Res 2013, 54, 2325–2340. [CrossRef]
- J. Baümler and V. Sperandio, ‘Interactions between the microbiota and pathogenic bacteria in the gut’. Nature 2016, 535, no. 7610, 85–93. [CrossRef]
- M. M. Natividad and E. F. Verdu, ‘Modulation of intestinal barrier by intestinal microbiota: Pathological and therapeutic implications. Pharmacol Res 2013, 69, 42–51. [CrossRef]
- E. Thursby and N. Juge, ‘Introduction to the human gut microbiota’. Biochemical Journal 2017, 474, 1823–1836. [CrossRef]
- T. Gensollen, S. S. Iyer, D. L. Kasper, and R. S. Blumberg, ‘How colonization by microbiota in early life shapes the immune system. Science (1979) 2016, 352, no. 6285, 539–544. [CrossRef]
- N. İlhan, ‘Gut Microbiota and Metabolism’, International Journal of Medical Biochemistry 2018, 1. [CrossRef]
- Rowland; et al. Gut microbiota functions: metabolism of nutrients and other food components’, Eur J Nutr 2018, 57, 1. [CrossRef]
- S. Musumeci, M. Coen, A. Leidi, and J. Schrenzel, ‘The human gut mycobiome and the specific role of Candida albicans: where do we stand, as clinicians?’. Clinical Microbiology and Infection 2022, 28, 58–63. [CrossRef] [PubMed]
- S. Dollive et al. A tool kit for quantifying eukaryotic rRNA gene sequences from human microbiome samples’. Genome Biol 2012, 13, R60. [CrossRef]
- K. Nash et al. The gut mycobiome of the Human Microbiome Project healthy cohort’, Microbiome 2017, 5, 1–13. [CrossRef]
- S. Sen and T. J. Mansell, ‘Yeasts as probiotics: Mechanisms, outcomes, and future potential’, 2020. [CrossRef]
- M. Boxberger, V. Cenizo, N. Cassir, and B. Scola, ‘Challenges in exploring and manipulating the human skin microbiome. Microbiome 2021, 9, 1–14. [CrossRef]
- E. Y. Chen, M. A. Fischbach, and Y. Belkaid, ‘Skin microbiota–host interactions’. Nature 2018, 553, 427–436. [CrossRef]
- R. Shaykhiev and R. Bals, ‘Interactions between epithelial cells and leukocytes in immunity and tissue homeostasis. J Leukoc Biol 2007, 82, 1–15. [CrossRef] [PubMed]
- E. A. Grice, ‘The intersection of microbiome and host at the skin interface: genomic-and metagenomic-based insights’, 2015. [CrossRef]
- H. Zhou, L. Shi, Y. Ren, X. Tan, W. Liu, and Z. Liu, ‘Applications of Human Skin Microbiota in the Cutaneous Disorders for Ecology-Based Therapy’. Front Cell Infect Microbiol 2020, 10, 1–12. [CrossRef]
- Y. Belkaid and J. A. Segre, ‘Dialogue between skin microbiota and immunity’. Science Mag 2014, 346, 954–959. [CrossRef]
- L. Byrd, Y. L. Byrd, Y. Belkaid, and J. A. Segre, ‘The human skin microbiome’, Nature Publishing Group 2018, 16. [CrossRef]
- L. Byrd, Y. Belkaid, and J. A. Segre, ‘The human skin microbiome’. Nat Rev Microbiol 2018, 16, 143–155. [CrossRef]
- M. K. Kwofie, N. Bukari, and O. Adeboye, ‘Probiotics Potential of Yeast and Lactic Acid Bacteria Fermented Foods and the Impact of Processing: A Review of Indigenous and Continental Food Products’. Adv Microbiol 2020, 10, 492–507. [CrossRef]
- Kaźmierczak-Siedlecka, J. Ruszkowski, M. Fic, M. Folwarski, and W. Makarewicz, ‘Saccharomyces boulardii CNCM I-745: A Non-bacterial Microorganism Used as Probiotic Agent in Supporting Treatment of Selected Diseases. Curr Microbiol 2020, 77, 1987–1996. [CrossRef] [PubMed]
- P. K. Mukherjee, B. Sendid, G. Hoarau, J.-F. Colombel, D. Poulain, and M. A. Ghannoum, ‘Mycobiota in gastrointestinal diseases. Nat Rev Gastroenterol Hepatol 2015, 12, 77–87. [CrossRef]
- R. Abid et al. Probiotic Yeast Saccharomyces: Back to Nature to Improve Human Health. Journal of Fungi 2022, 8, 444. [CrossRef]
- M.-C. Barc et al. Molecular analysis of the digestive microbiota in a gnotobiotic mouse model during antibiotic treatment: Influence of Saccharomyces boulardii’. Anaerobe 2008, 14, 229–233. [CrossRef]
- Collignon, C. Sandré, and M.-C. Barc, ‘Saccharomyces boulardii modulates dendritic cell properties and intestinal microbiota disruption after antibiotic treatment’. Gastroenterol Clin Biol 2010, 34, S71–S78. [Google Scholar] [CrossRef]
- M. I. Moré and A. Swidsinski, ‘Saccharomyces boulardii CNCM I-745 supports regeneration of the intestinal microbiota after diarrheic dysbiosis – a review’, Clin Exp Gastroenterol 2015, 237. [CrossRef]
- A.-M. Yang et al. Saccharomyces Boulardii Ameliorates Non-alcoholic Steatohepatitis in Mice Induced by a Methionine-Choline-Deficient Diet Through Gut-Liver Axis’, Front Microbiol 2022, 13. [CrossRef]
- A.-M. Yang et al. Saccharomyces Boulardii Ameliorates Non-alcoholic Steatohepatitis in Mice Induced by a Methionine-Choline-Deficient Diet Through Gut-Liver Axis’, Front Microbiol 2022, 13. [CrossRef]
- Y. Wu et al. Effects of Multispecies Probiotic on Intestinal Microbiota and Mucosal Barrier Function of Neonatal Calves Infected With E. coli K99’, Front Microbiol 2022, 12. [CrossRef]
- M. Bloemendaal et al. Probiotics-induced changes in gut microbial composition and its effects on cognitive performance after stress: exploratory analyses’. Transl Psychiatry 2021, 11, 300. [CrossRef]
- Duysburgh, P. van den Abbeele, M. Morera, and M. Marzorati, ‘Lacticaseibacillus rhamnosusGG and Saccharomyces cerevisiae boulardii supplementation exert protective effects on human gut microbiome following antibiotic administration in vitro’. Benef Microbes 2021, 12, 365–379. [Google Scholar] [CrossRef]
- Saber, B. Alipour, Z. Faghfoori, and A. Yari Khosroushahi, ‘Cellular and molecular effects of yeast probiotics on cancer’. Crit Rev Microbiol 2017, 43, 96–115. [Google Scholar] [CrossRef]
- Kainz, M. A. Bauer, F. Madeo, and D. Carmona-Gutierrez, ‘Fungal infections in humans: the silent crisis’. Microbial Cell 2020, 7, 143–145. [Google Scholar] [CrossRef]
- Buerth, C., Tielker, D., & Ernst, J. F. (2016). Candida utilis and Cyberlindnera (Pichia) jadinii: yeast relatives with expanding applications. Applied microbiology and biotechnology, 100, 6981-6990.
- Sharma, P., Gaur, V. K., Kim, S. H., & Pandey, A. (2020). Microbial strategies for bio-transforming food waste into resources. Bioresource technology, 299, 122580.
- Patil, S., Pimpley, V., Warudkar, K., & Murthy, P. S. (2022). Valorisation of coffee pulp for development of innovative probiotic beverage using kefir: Physicochemical, antioxidant, sensory analysis and shelf life studies. Waste and Biomass Valorization, 13(2), 905-916.
- BCC Market Research. (2022). Probiotics in Food, Beverages, Dietary Supplements and Animal Feed. FOD035H.
- Group, J.F.W.W. Report on Drafting Guidelines for the Evaluation of Probiotics in Food. London, Ontario, Canada. 2002. Available online: https://www.who.int/foodsafety/fs_management/en/probiotic_guidelines.pdf.
- Gareau, M. G., Sherman, P. M., & Walker, W. A. (2010). Probiotics and the gut microbiota in intestinal health and disease. Nature reviews Gastroenterology & hepatology, 7(9), 503-514.
Figure 1.
Probiotic ability of Saccharomyces boulardii [5].
Figure 1.
Probiotic ability of Saccharomyces boulardii [5].
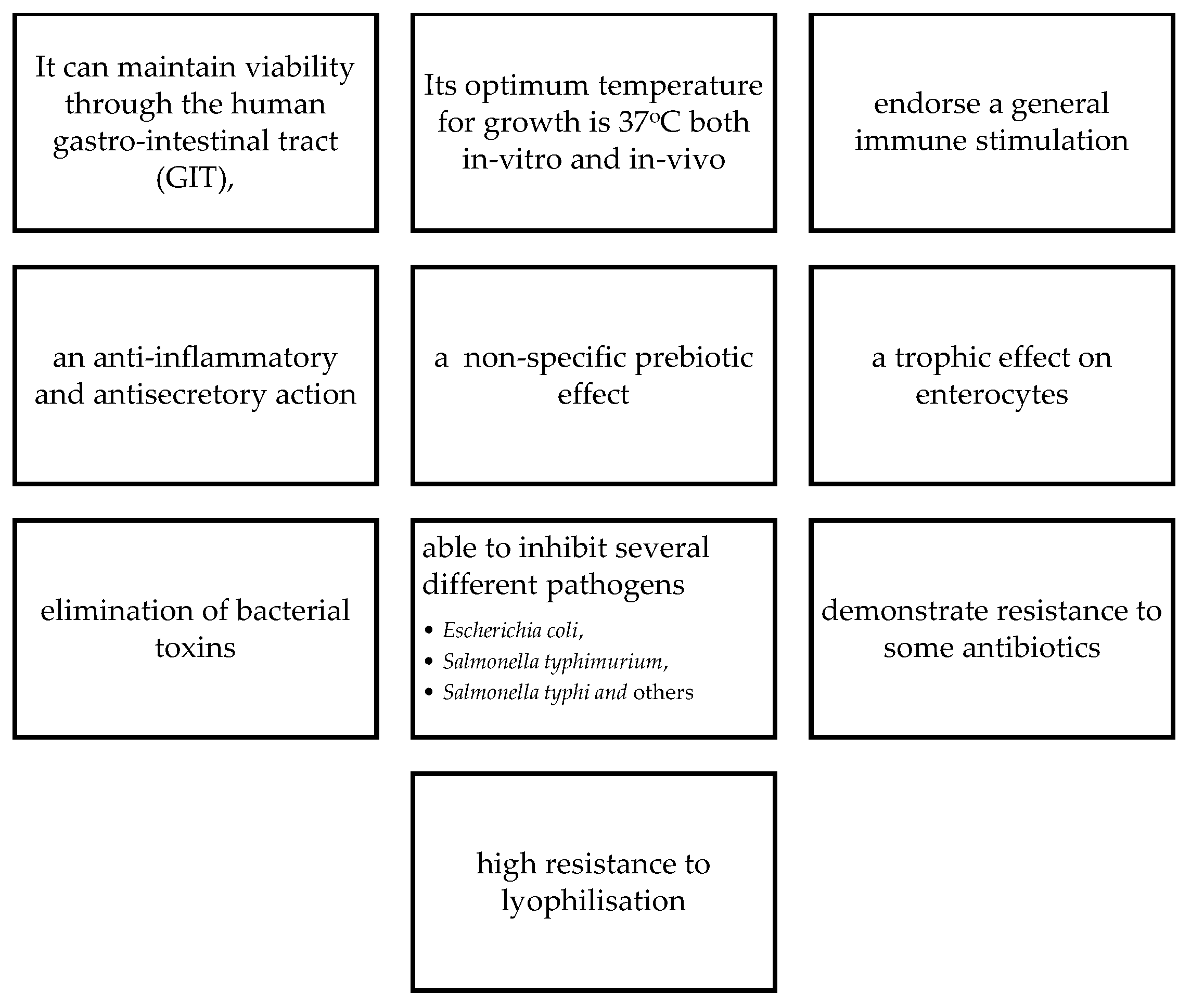
Figure 2.
Factors to consider when developing a yeast probiotic using a classical biotechnological approach.
Figure 2.
Factors to consider when developing a yeast probiotic using a classical biotechnological approach.
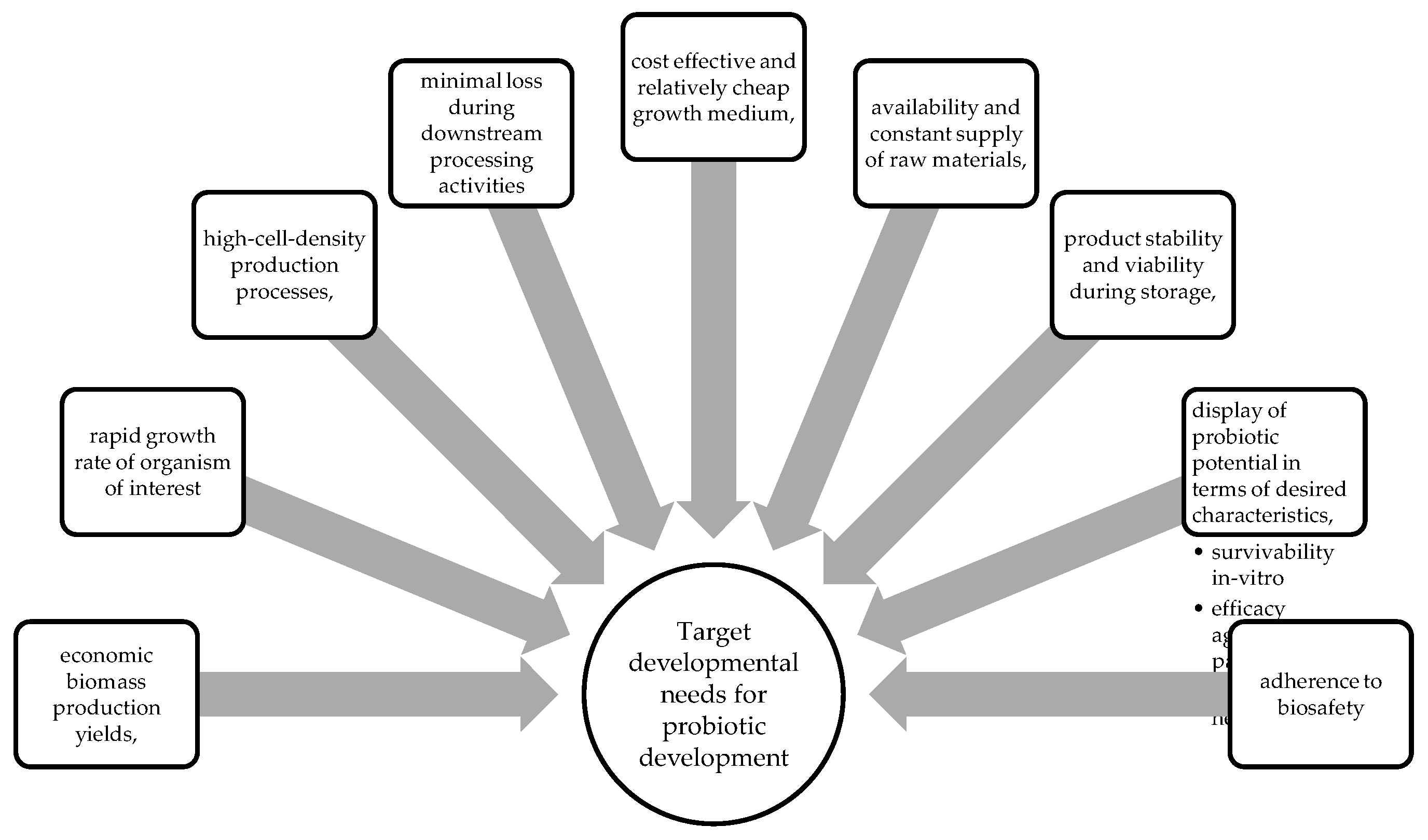
Figure 4.
Different oral delivery systems used to administer probiotics.
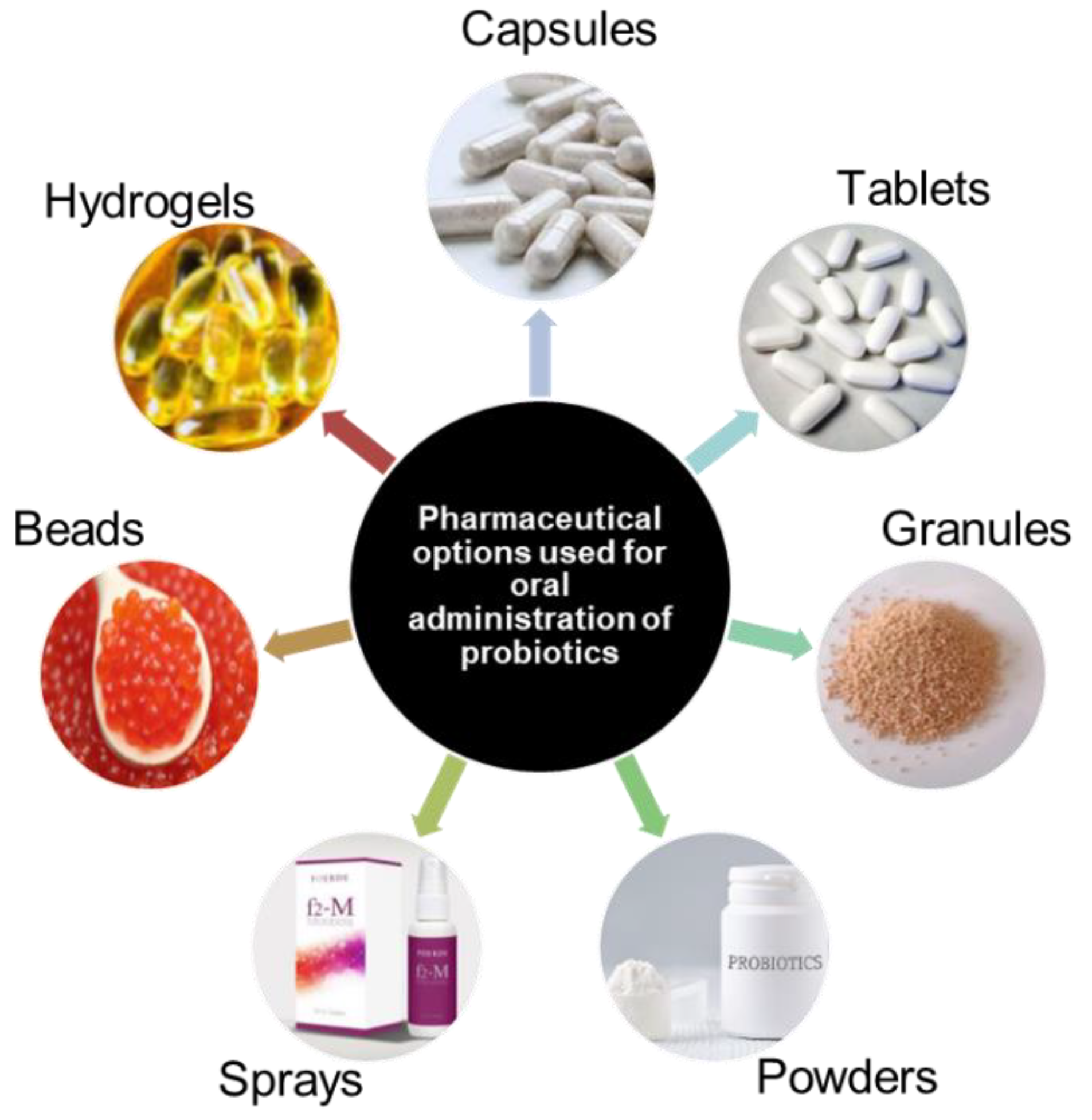
Figure 5.
Diagrammatic representation of the different methods of encapsulation typically used for probiotic applications [76].
Figure 5.
Diagrammatic representation of the different methods of encapsulation typically used for probiotic applications [76].
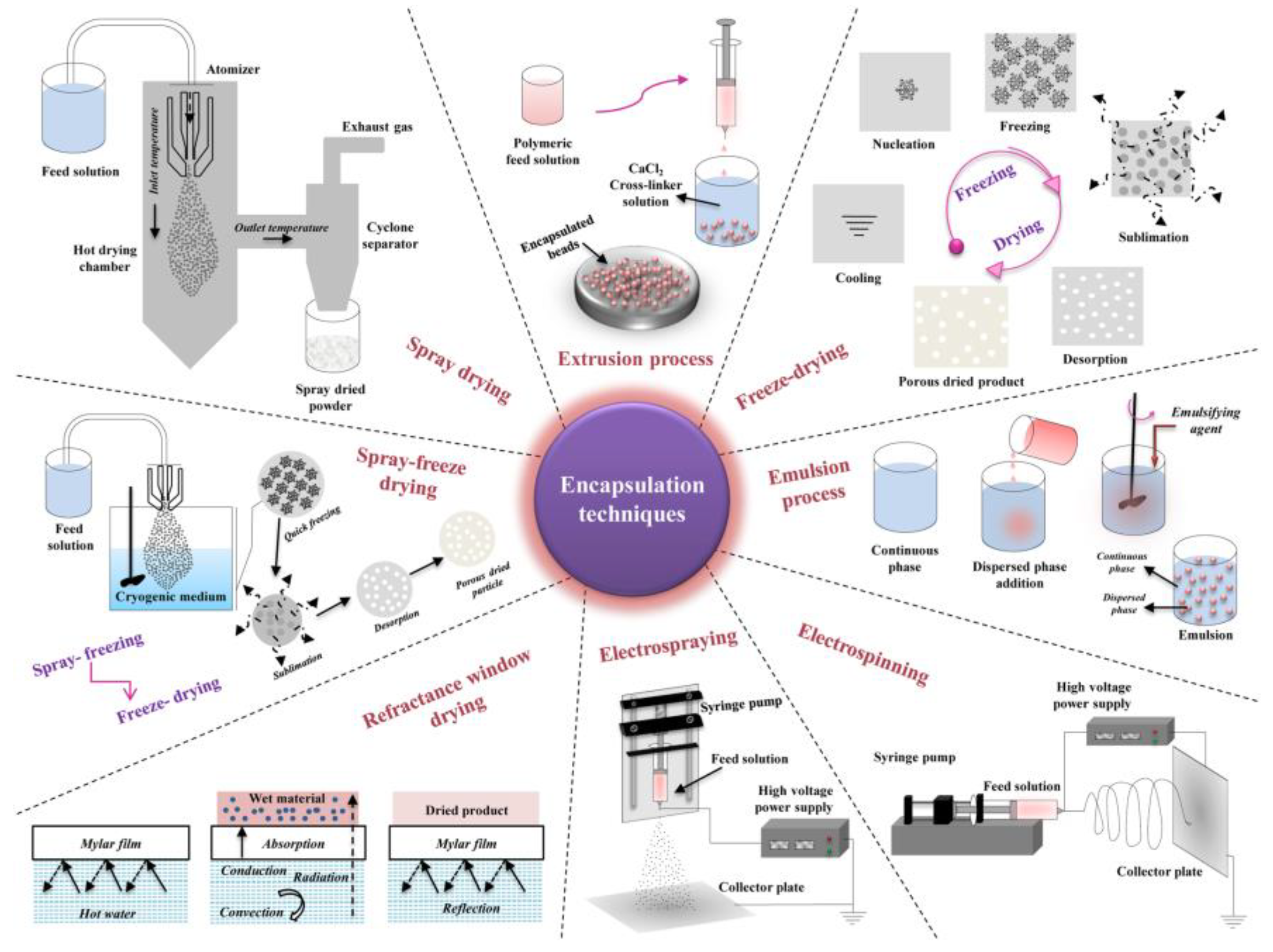
Figure 6.
Illustration of co-encapsulated probiotics, prebiotics in fortified milk product and advantages offered by the technique [75].
Figure 6.
Illustration of co-encapsulated probiotics, prebiotics in fortified milk product and advantages offered by the technique [75].
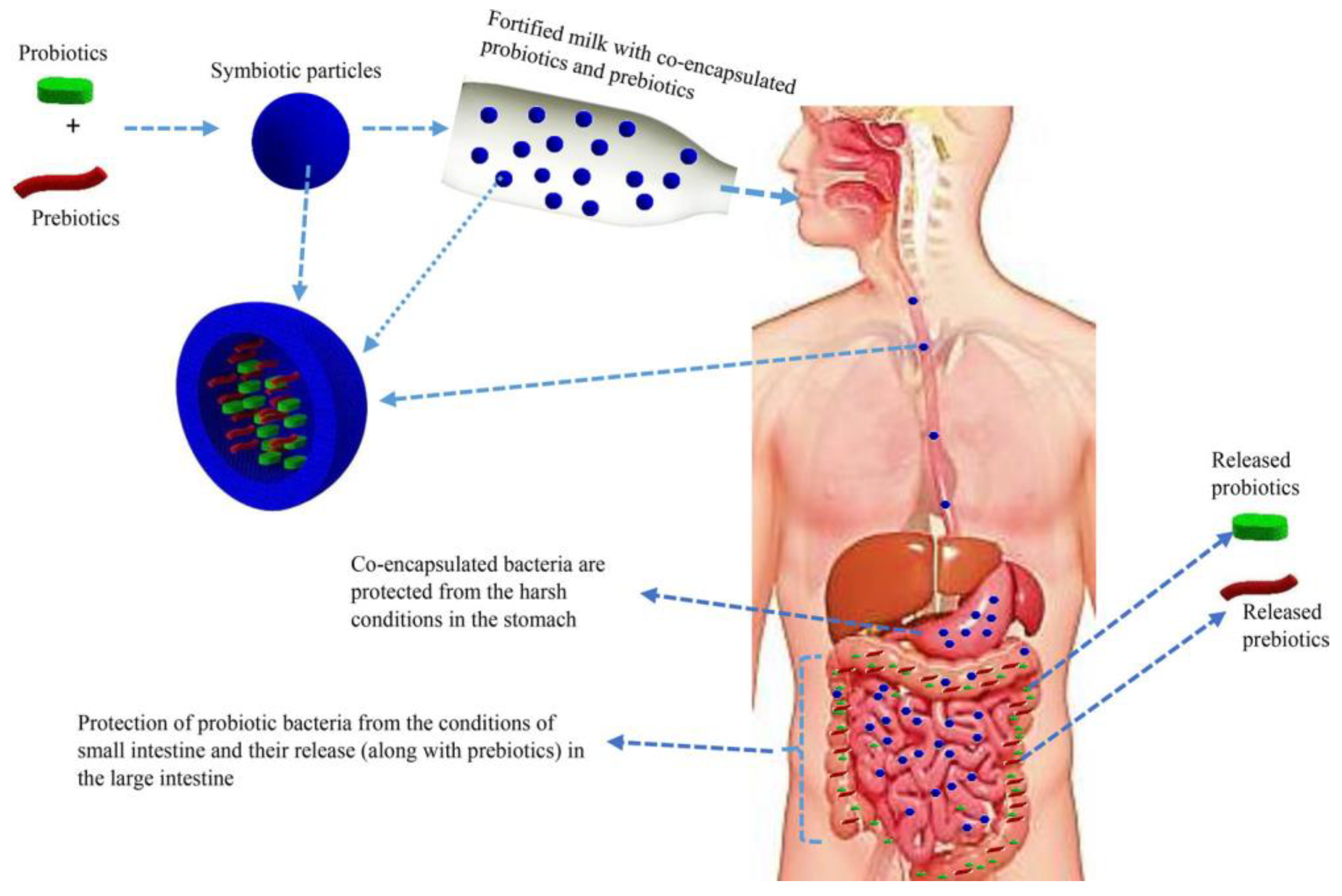
Figure 7.
General composition of the human gut microbiome (Cresci & Izzo, 2018).
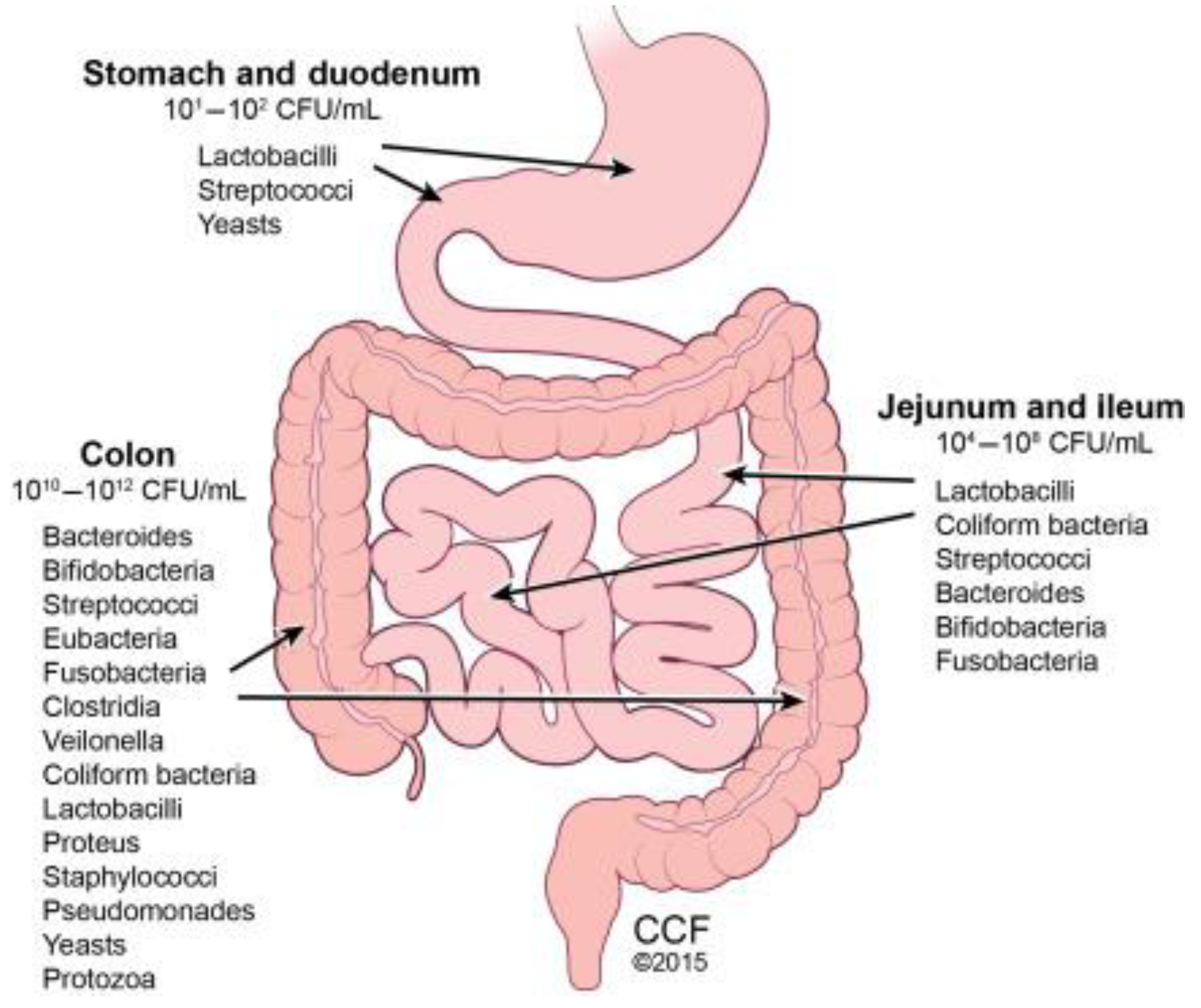
Table 1.
Assessment criteria used for the assessment of yeast probiotic potential.
| Characteristic | Rationale | Reference |
|---|---|---|
| Hydrophobicity | For an organism to show functionality as a probiotic, it needs to display hydrophobicity. The organism of interest needs to demonstrate its ability to adhere/interact with the mucus present within the GIT to confer the probiotic effect. | [21] |
| Auto-aggregation | This is a characteristic wherein cells are able to self-aggregate and adhere to the mucus/mucosal lining in order to form a biofilm. A desirable level of auto-aggregation is ~30 to 60% | [22] |
| Biofilm formation | To show the ability of cells to adhere to each other and the host lining | [23] |
| Adherence ability | To assess the ability of the probiotic cell to adhere to the mucosal lining and confer a probiotic | [24] |
| Survival | To assess the organism’s ability to survive exposure to low pHs (gastric conditions) and the presence of bile salts (0.3%) | [25] |
| Antibiotic resistance | In the instances of yeasts intended for use as a probiotic, antibiotic resistance using a disk assay method will infer information pertaining to the ability of the organism to demonstrate antibiotic resistance. | [26] |
| Antimicrobial activity | To assess the yeast’s ability to demonstrate anti-microbial activity, which is pertinent for the treatment of pathogens | [27] |
Table 4.
Dominating microbial families in mice fed with normal chow diet, MCD only as well as MCD and S. boulardii at family level.
Table 4.
Dominating microbial families in mice fed with normal chow diet, MCD only as well as MCD and S. boulardii at family level.
| Normal chow diet | MCD diet | MCD plus S. boulardii |
|---|---|---|
| Muribaculaceae | Akkermansiaceae | Lachnospiraceae |
| Ruminococcaeceae | Erysipelotrichaceae | Atopobiaceae |
| Lactobacillaceae | Tannerellaceae | Ruminococcaceae |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



