
Submitted:
01 October 2024
Posted:
01 October 2024
You are already at the latest version
Abstract
Recent clinical trials using synthetic incretin hormones, glucagon-like peptide 1 (GLP-1) and glucose-dependent insulinotropic polypeptide (GIP) receptor agonists have demonstrated that these treatments ameliorated many complications related to obesity, emphasizing the significant impact of body weight on overall health.
Incretins are enteroendocrine hormones secreted by gut endothelial cells triggered by nutrient ingestion. The phenomenon that oral ingestion of glucose elicited much higher insulin secretion than intra-venous injection of equimolar glucose is known as incretin effect. This also alludes to the thesis that food intake is the root cause of insulin resistance.
Amylin is co-expressed with insulin from the pancreas β-cells but does not have insulinotropic function. Amylin suppresses glucagon secretion, slowing gastric emptying, and suppressing the central nervous system (CNS) reward center leading to weight loss. However, amylin can self-aggregate and cause serious cytotoxicity and may cause β-cell apoptosis.
Glucagon is secreted by the pancreatic α-cells and participates in glucose homeostasis in glucose-dependent manner. In hypoglycemia, glucagon increases blood glucose level by glycogenolysis and gluconeogenesis and inhibits glycogenesis in the liver. Some triple agonists in combination with glucagon and dual incretins are already being developed.
These advances bring to the question “Are the benefits of these anti-obesity treatments sustainable?” Chronic agonism may decrease the number of receptors. Also, long-term stimulation may cause β-cell exhaustion and failure. Additionally, instead of endogenous control of the appetite, exogenous control of satiety and food intake may hinder the long-term sustainability of these treatments. We will discuss the incretins’ mechanism of action, challenges, and future directions.
Keywords:
Glucagon-like peptide 1
; Glucose-dependent insulinotropic polypeptide
; Satiety
; Proopiomelanocortin (POMC)
; Neuropeptide Y (NPY)
; Agouti-related peptide (AgRP)
; GABAergic neurons
Introduction
By the definition of La Barre[1], an incretin should satisfy two criteria: (a) it must be released by nutrients, especially carbohydrates and; (b) it must stimulate insulin secretion in the presence of elevated blood glucose levels. Among the potential incretin candidates such as gastrin, secretin, cholecystokinin (CCK), vasoactive intestinal peptide (VIP), peptide histidine isoleucine (PHI), gastrin-releasing peptide (GRP), entero-glucagon, glucagonlike peptides (GLPs), and glucose-dependent insulinotropic polypeptides (GIP), GLP-1 and GIP are the most important and most researched incretins.
GIP is secreted from K cells in the small intestine by proteolysis of pre-pro-GIP,[2] while GLP-1 is expressed in the intestinal L cells and derived from pro-glucagon through differential proteolytic cleavage.[3] Both GIP and GLP-1 act on G protein-coupled receptors on islet β cells to stimulate insulin secretion. [4] Although both stimulate insulin secretion, there are subtle differences between these two molecules. GLP-1 suppresses appetite via neuronal inputs, while GIP’s role in appetite is conflicting. Although animal studies report GIP’s involvement in appetite suppression.[5,6,7] and neurons expressing the GIP receptors are identified in the brain of both mice and humans, human studies including randomized controlled trials, reported almost no effects of GIP on appetite suppression. [8,9,10] We believe that appetite control involves numerous hormones and neurons, so focusing on just one neuron or molecule as was done in most animal studies might not provide a complete picture. Moreover, each molecule can be stimulated or inhibited. Thus, the end results may have many possible outcomes.
The clinical successes of the incretin trials utilizing GLP-1 receptor agonists (GLP-1ras) and GIP receptor agonists (GIPras) have been nothing short of “astonishing” in weight reduction,[11,12] and glucoregulation in type 2 diabetes (T2D)[13] and prediabetes.[14] Further, incretin efficacies are reported in the prevention of cardiovascular complications,[15] and delaying the complication of diabetes-related kidney dysfunction.[16] The most recent trials utilizing dual (combination of GLP-1 and GIP) or triple receptor agonists demonstrated more robust weight reduction and glycemic control than GLP-1 receptor agonists (GLP-ras) alone.[17,18,19,20,21]
We have summarized some key randomized trial results showing these benefits in Table 1. These are not a complete collection of references but only essential trials that demonstrated the superiority of incretin agonism compared to traditional diabetes treatments or other GLP-1 ras.

Table 1.
Key randomized trial results on incretin agonism and health outcomes.
| First author, trial name/ID, year, phase | Sample size, duration | Target population, methods | Objectives, Results (s), & comments |
|---|---|---|---|
| Le Roux, [NCT01272219], 2017, Phase 3[14] |
N=2254, 68 weeks or 160 weeks |
Prediabetic obese and overweight with co-morbidities cohort Drug: Lira 3.0mg or placebo SC injection once daily |
Objectives: Weight reduction /maintenance, T2D onset delay. B Results: y 160 weeks, DM Dx given to (2%) of 1472 in Lira vs. (6%) of 738 in the placebo. Time to DM diagnosis was 99 wks in Lira vs. 87 wks in placebo. |
| Husain M, PIONEER 6 [NCT02692716], 2019, phase 3[15] | N=3183, Median 62 weeks |
T2D with high-cardiovascular risk cohort Drug: Sema or placebo Oral administration once daily |
Objectives: Cardiovascular safety of oral Sema, QD in T2D patients. Primary outcome: incidence of MACE Results: MACE occurred in 3.8% in Sema vs. 4.8% in placebo including 15 CVD mort. In Sema Vs. 30 in placebo. |
| Frías JP, AWARD-11 [NCT03495102], 2021, phase 3[13] | N=1842, 52 weeks (36 weeks primary endpoint) |
T2D Patients inadequately controlled with metformin Drug: Dula 1.5mg, 3.0mg or 4.5mg SC injection once weekly |
Objectives: Change in HbA1c by week 36 from baseline. Results: At 36wks, Dula 4.5 mg superior to 1.5mg with [ETD] -0.24% but Tx estimand of 3.0 mg was not significant (P = 0.096). However, vomiting nearly doubled in 4.5 mg level. (5.6% vs. 9.3%) |
| Rubino D, STEP-4 [NCT03548987], 2021, phase 3a[11] | N=803, 68 weeks |
Obese or overweight cohort without T2D Drug: Sema 2.4mg or placebo SC injection once weekly Primary outcome: Change in body weight (%) |
Objectives: Comparison of SC Sema continued or switch to placebo both with lifestyle intervention. wt. change week 20- 68: Sema -7.9% vs placebo +6.9%. G-I adverse events: Sema 49.1% vs placebo 26.1% (1.88 times more in Sema group.) |
| Frías JP, SURPASS-2 [NCT03987919], 2021, phase 3[17] | N=1879, 40 weeks |
Metformin-treated T2D cohort Drug: Sema 1mg or Tirzep 5mg, 10mg, 15mg SC injection once weekly |
Objectives: Compare effect of Sema and Tirzep on blood sugar levels. Outcome=Change in HbA1c by week 40. The diff. btw groups Tirzep 5-mg, 10-mg, and 15-mg and Sema were -0.15%, −0.39% & −0.45%, respectively. SAE: 5-7% in Tirzep vs. 3% in Sema. |
| Ludvik B, SURPASS-3 [NCT03882970], 2021, phase 3[22] | N=1444, 52 weeks |
Metformin-treated or metformin with SGLT2i- treated T2D cohort Drug: Tirzep 5mg, 10mg, 15mg or insulin degludec 100 U/mL (titrated) SC injection once weekly (Tirzep), SC injection once daily (insulin degludec) |
Objectives: Assess safety and efficacy of Tirzep versus insulin degludec on blood sugar levels Results: Non-inferiority of Tirzep to insulin. HbA1c change in Tirzep 5, 10, 15 mg at wk 52 were -1·93%, -2·20%, -2·37%, respectively and -1.34% in insulin. G-I adverse events: 7% in Tirzep vs. 1% in insulin group. Hypoglycemia: 4% in Tirzep vs. 7% in insulin gr. |
| Del Prato S, SURPASS-4 [NCT03730662], 2021, phase 3[23] | N=2002, 52 weeks (treatment continued until maximum 104 weeks) |
Metformin-treated, sulfonylurea-treated, SGLT2i-treated T2D cohort Drug: Tirzep 5mg, 10mg, 15mg or glargine 100 U/mL (titrated) SC once weekly (Tirzep), SC once daily (glargine) |
Objective: Assess efficacy and safety of Tirzep versus insulin glargine in adults with high CVD risk and T2D. Primary outcome: Non-inferiority of Tirzep 10mg or/and 15mg versus glargine. Mean HbA1c change at week 52: -2·43% and -2·58%, with Tirzep 10, 15 mg respectively vs.-1·44% with glargine. |
| Rubino DM, STEP-8 [NCT04074161], 2022, phase 3b[12] | N=338, 68 weeks |
Obese or overweight cohort without T2D Drug: Sema 2.4mg or Lira 3.0mg or placebo (matching for both conditions) SC injection once weekly (Sema), SC injection once daily (Lira) |
Objectives: Assess the efficacy of once-weekly Sema vs. once-daily Lira on weight loss. Change in body weight (%) by week 68. Mean Wt. change from baseline: -15.8% with Sema, -6.4% with Lira, -1.9% with placebo. G-I adverse events: 84.1% with Sema, 82.7% with Lira. |
| Wilding J, STEP 1- extension [NCT03548935], 2022[24] | N=327, 1 year after withdrawal from STEP-1 |
Extension analysis Previous drug: Sema 2.4mg or placebo |
Objectives: body weight changes and cardio-metabolic factors following Sema withdrawal. Primary outcome: One year after withdrawal of weekly Sema 2.4 mg + lifestyle intervention, participants regained two-thirds of their prior weight loss. |
| Heerspink H, SURPASS-4 Post Hoc Analysis, 2022[16] | N=2002, Median 85 weeks (104 weeks max) |
Metformin-treated, sulfonylurea-treated, SGLT2i-treated T2D cohort Drug: Tirzep 5mg, 10mg, 15mg or glargine 100U/mL (titrated) SC injection once weekly (Tirzep), SC injection once daily (glargine) |
Objectives: Compare the effects of Tirzep and insulin glargine on kidney. Primary outcome: tirzepatide slowed the eGFR decline (1.4 vs 3.6 mL/min ) and UACR increased with insulin while with Tirzep decreased by -6.8% compared with insulin glargine. |
| Dahl D, SURPASS-5 [NCT04039503], 2022, phase 3[25] | N= 475, 40 weeks |
T2D with titrated insulin glargine on glycemic control cohort Drug: Tirzep 5mg, 10mg, 15mg or placebo SC injection once weekly |
Objectives: Assess efficacy and safety of Tirzep in T2D patients receiving inadequate glycemic control. Primary outcome: Mean changew in HbA1c were −2.40% , −2.34%, and −0.86% with 10mg, 15-mg Tirzep and placebo, respectively. |
| Lincoff AM, SELECT [NCT03574597], 2023, phase 3[26] | N=17604, Mean 137 weeks (Mean follow up 160 weeks) |
Obese or overweight cohort with CVD and without T2D Drug: Sema 2.4mg or placebo SC injection once weekly |
Objectives: Assess reduction in risk of having cardiovascular events. Primary outcome= MACE (CVD mortality+ nonfatal MI+ nonfatal stroke). 6.5% MACE in Sema 8.0% in placebo (Risk Diff.=1.5%) SAE leading to permanent discontinuation was doubled in Sema. (16.6% in Sema, 8.2% in placebo). |
| Jastreboff AM, [NCT04881760], 2023, phase 2[21] | N=338, 48 weeks |
Obese or overweight with weight-related comorbidities cohort without T2D Drug: Reta 1mg, 4mg, 8mg, 12mg or placebo SC injection once weekly Retatrutide=multireceptor agonist of (GLP-1+ GIP+glucagon) |
Objectives: Assess efficacy of Reta on body weight loss. Primary outcome: Change in body weight (%) by week 24. Results: Wt. change at 24 weeks -7.2% (1-mg), -12.9% (4-mg), -17.3% (8-mg), & 17.5% in the 12-mg retatrutide groups, -1.6% placebo. HR peaked at 24 weeks and declined thereafter. NB: comparator should have been Tirzep, not placebo to show adding glucagon would be safe. |
| Aronne L, SURMOUNT-4 [NCT04660643], 2024, phase 3[27] | N=670, 88 weeks (36 weeks onward placebo could be administered) |
Cohort: Obese or overweight without T2D Drug: Tirzep 10mg, 15mg Mean wt. loss 20.9%. At wk 36 randomized to continue Sema or placebo. |
Objectives: Assess Tirzep effect on maintenance of body weight reduction. Primary outcome: Mean change in weight from week 36 until week 88 (%). Results: Switched to placebo group regained 14% wt. (67%) continuing tirzepatide lost additional 5.5%. |
| Loomba R, [NCT04166773], 2024, phase 2[28] | N=190, 52 weeks |
Cohort: Confirmed-MASH with liver fibrosis Drug: Tirzep 5mg, 10mg, 15mg or placebo SC injection once weekly |
Objectives: Assess safety and efficacy of Tirzep as a MASH treatment. Primary outcome: Resolution of MASH without worsening of fibrosis by week 52. Results: Risk diff. 34%, 46%, & 53% at Tirzep 5-mg, 10-mg, 15-mg respectively. |
| Sanyal AJ, [NCT04771273], 2024, phase 2[29] | N=293, 48 weeks (24 weeks rapid-dose phase, 24 weeks maintenance phase) |
Confirmed-MASH with fibrosis cohort Drug: Survo 2.4mg, 4.8mg, 6.0mg or placebo SC injection once weekly |
Objectives: Assess safety, tolerability and efficacy of Survo (Dual agonist of Glucagon and GLP-1 ra)as a MASH treatment. Primary outcome: Reduction in MASH with no worsening of fibrosis by week 48. Results: risk diff. of liver fat decrease were 49%, 53%, & 43% in the 2.4mg, 4.8mg, 6.0mg Survo groups respectively. more nausea in Survo (66% vs. 23%), diarrhea (49% vs. 23%), and vomiting (41% vs. 4%); SAE were similar. |
| Sanyal AJ, [NCT04881760], 2024, phase 2a[30] | N=98, 48 weeks |
Obese or overweight with weight-related comorbidities cohort without T2D Drug: Reta 1mg, 4mg, 8mg, 12mg or placebo SC injection once weekly |
Objectives: Assess safety, tolerability and efficacy of Reta for body weight loss, assess liver fat at 24 weeks. Results: At 24 weeks, normal LF was achieved by 27%, 52%, 79%, & 86% with 1 mg, 4 mg, 8 mg, 12 mg of Reta and 0% (placebo). LF reductions were related to changes in Wt., abdominal fat and metabolic measures of insulin sensitivity. |
Nota Bene: In real-world data, age at the time of GLP-1 initiation, and GLP-1 ra cessation increased the risk of major adverse cardiac events (MACE).[31] These facts strongly suggest that age and other factors can be important confounders.
The two incretins have shared functions as well as divergent functions. They share the insulinotropic function but when combined, they seem to have additive action. GIP has more powerful insulinotropic action and is responsible for 44% of the total insulin responses and GLP-1 contributes 22%. [32] We will review the role of incretins and other molecules involved in glucose homeostasis and weight management in the next section.
- a.
- GLP-1 and GIP Mechanism of Action
When GLP-1 and GIP bind to their cognate receptors, they stimulate insulin secretion from the pancreatic β-cell through the incretin effects.[33] Both incretin-related receptors (GIPr) and GLP-1r belong to the class B family of 7-transmembrane G protein-coupled receptors (GPCR). GIP and GLP-1 share the insulinotropic actions, but other functions maybe divergent. For example, GIP stimulates glucagon secretion from pancreatic α-cells in hypoglycemia in healthy persons, but in T2D, the glucagonotropic function of GIP is dysregulated.[34] While GLP-1 is known to inhibit glucagon secretion possibly via somatostatin,[35] delay gastric emptying, and suppress food intake, GIP does not appear to slow gastric emptying.[36] Reports regarding GIP’s role in appetite suppression are also conflicting: Animal studies and in vitro studies reported GIP’s role in appetite suppression, [5,6] while human studies did not. [8,10] Another reason for combining GLP-1 and GIP agonists is that GIP has an anti-emetic function and counteracts nausea and vomiting evoked by GLP-1 ra.[37]
- b.
- Amylin Mechanism of Action
Amylin, also known as islet amyloid polypeptide, is co-expressed with insulin from the pancreatic β-cells but has no insulinotropic function. For this reason, amylin is not considered an incretin. [38] Amylin reduces endogenous glucose production by suppressing glucagon secretion, slows gastric emptying, and suppresses CNS reward centers leading to weight loss. However, amylin as the name suggests, ‘islet amyloid polypeptide’ can self-aggregate and cause endoplasmic reticulum stress, serious cytotoxicity, and may cause β-cell death. [39] Therefore, the clinical utility of amylin is very limited.
- c.
- Glucagon Mechanism of Action
Glucagon is a peptide with 29 amino acids and is secreted by α-cells of the pancreas. It is not an incretin but intimately participates in glucoregulation and body weight management. Glucagon receptors (GCGR) are expressed in many organs including liver, kidney and heart among other organs. The main biological function of glucagon is counter-balancing insulin in hypoglycemia. It is also involved in hepatic lipids and amino acids metabolism. Additionally, glucagon is known to enhance satiety and suppress food intake, and it has become an attractive molecule for body weight management.[40] Glucagon also promotes lipogenesis and ketone body formation from non-carbohydrate energy. In times of high energy demand, glucagon converts fatty acids to acetyl-coenzyme A via β-oxidation in the liver.[41] Additionally, glucagon activates the signaling pathway to inhibit hepatic de novo lipogenesis and prevent the onset of hepatic steatosis.[42] Although activation of GCGR is involved in the body weight-lowering action of oxyntomodulin (OXM), OXM’s involvement in weight loss appears to be redundant with GLP-1 agonism. In T2D, however, glucagon regulation is abnormal.[43] The newly developed triple agonist, retatrutide, containes of GLP-1, GIP and GCGR agonist. However, adding 3 powerful molecules together may beget a hidden danger in a long-term usage. Let us recapitulate the benefits of incretins.
Increasing insulin secretion and improving glycemic control; Some studies also reported insulin sensitivity. However, insulin sensitivity was reported to be associated with inflammation arising from obesity. [44,45]
Suppress food intake by slowing gastric emptying and increasing satiety which lead to weight loss. [10] Interestingly, semaglutide did not delay gastric emptying assessed using paracetamol absorption in a recent trial.[46] In a murine model, GLP-1 did not slow the gastric emptying.[47] As we have stated earlier, manipulating the neurohormonal axis by incretins may be the cause of weight loss. The same pathway also reduces craving for alcohol intake. Unfortunately, when incretin’s manipulation of neuronal pathways is terminated, approximately 2/3 of the lost weight was regained. [24,48]
May potentiate functional β-cell regeneration in animal or in vitro studies. [49] However, clear evidence of functional β-cell regeneration or proliferation in humans is lacking. [35]
May prevent bone fractures.[50] However, not all studies reported beneficial effects of incretin agonism on bone mineral density (BMD). Two randomized trials reported that exercise might be a better option for BMD than GLP-1 agonism.[51,52] Further well-conducted studies are needed.
Several reports suggest that relative hyperglucagonemia contributes to fasting and postprandial hyperglycemia in T2D, and glycemic control may be achieved by blocking glucagon action. [53] Moreover, numerous reports suggested the deleterious effects of GLP-1 ra as well as glucagon on heart rate and other cardiac functions. [54,55,56,57] Please refer to the “Challenges” section for further reading.
2. Role of Incretins in the Neurohormonal Axis of Appetite Control
Homeostatic feeding is a mechanism where energy for basic metabolic processes and survival will be obtained, while hedonic feeding is driven by sensory perception or pleasure. [58] Homeostatic feeding is tightly controlled by many molecules, hormones, and neuronal elements. These include sensing nutrients in the central nervous system (CNS), integrating afferent stimuli, reflecting the energy balance, and adjusting subsequent food intake. [59] The incretin effect is largely mediated by neuroendocrine actions and is correlated with the size of the meal. [38]
In the neuroendocrine control of food intake, the brainstem and hypothalamus are the core CNS areas because they receive, convey, and integrate peripheral signals. The area postrema (AP) and nucleus tractus solitarius (NTS) in the brainstem convey the peripheral signals, consisting of nutrients, hormones, and vagal afferent inputs, to the arcuate nucleus (ARC) of the hypothalamus. The ARC contains both orexigenic and anorexigenic neurons. The former expresses neuropeptide Y (NPY) and agouti-related peptides (AgRP) and the latter expresses pro-opiomelanocortin (POMC) and cocaine / amphetamine-related transcript (CART). They collectively process the received information and regulate eating and attain energy homeostasis. [60,61]
POMC cells activate melanocortin 4 receptor (MC4R) expressing neurons in the paraventricular nucleus of the hypothalamus (PVH) and other brain regions, thereby inhibiting food intake and increasing energy expenditure. The MC4R gene is involved in the brain's regulation of appetite and weight. Conversely, NPY/ AgRP neurons antagonize these effects. [62]
Also, leptin and serotonin are involved in regulating energy balance, appetite, and bone mass. Although both leptin and serotonin depolarize POMC neurons [63,64], there is a distinct selectivity in the responsive neurons. Namely, serotonin-responsive POMC neurons are not activated by leptin. Also, these two groups of neurons are anatomically segregated: leptin-activated POMC cells are located more laterally in the ARC than the serotonin-responsive cells. [64] Serotonin modulates the endogenous release of both agonists and antagonists of the melanocortin receptors, which are a core component of the CNS circuitry controlling body weight homeostasis. It should be noted that non-homeostatic or hedonic feeding can override this homeostatic pathway and result in overeating and obesity. Therefore, preventing hedonic feeding by food choices may be beneficial for weight homeostasis.
A sophisticated murine study reported the presence of GABAergic neurons in the dorsal vagal complex as a new player in the governing feeding behavior. [7] However, it was reported that GABAergic neurons, do not appear to express AgRP and reduce inhibitory tone to postsynaptic POMC neurons.[65] Vong’s study actually challenges the recent sophisticated studies in mice and insinuates that these studies might be flawed. The authors of murine models have shown that GABAergic neurons inhibit NPY [7] or food intake. [6] Moreover, the effects of GABAergic signals are not limited to NPY/ AgRP inhibition. GABAergic signals also inhibit (POMC) neurons during fasting.[61]
Both anorexigenic neurons as well as orexigenic neurons could be inhibited, or stimulated, depending on the energy status of the host.[61] Therefore, a multitude of feeding possibilities exists even without accounting for the brain region involved. The probability of non-exclusive event involving multiple molecules with 2 possible functions (inhibited or stimulated) is impossible to estimate in animal studies. The key point is that GLP-1 and GIP receptor agonists pharmacologically manipulate the neural signals and consequently, obese individuals lose appetite as well as body weight.
Peripheral injection of fluorescently labeled liraglutide revealed the presence of the drug in the circum-ventricular organs. [66] In this study, murine brain slices showed that GLP-1 directly stimulates POMC/CART neurons and indirectly inhibits neurons expressing neuropeptide Y (NPY) and agouti-related peptide (AgRP) via GABA-dependent signaling. [66] The labeled liraglutide was internalized in the neurons expressing proopiomelanocortin (POMC) and cocaine- and amphetamine-regulated transcript (CART).[66]
Beck also reported that the physiological activation of neuropeptides is dependent on energy availability of the host.[61,67] When energy is deprived or restricted, NPY is activated and when energy availability returns to normal, NPY synthesis returns to baseline level. Also, NPY metabolism is regulated by diet type, especially carbohydrate and fat content.[67] More recent findings support the view that the hypophagia was mediated by GLP-1 receptors in the brain.[68]
Toda summarized these facts as “arcuate melanocortin neurons consist of two distinct neuronal populations: (POMC)-expressing neurons and NPY/AgRP)-expressing neurons.”[62] However, it should be noted that POMC cells are activated by energy surfeits (excess) signals and inhibited by energy deficits.[61] In other words, the energy excess or deficits are the drivers of POMC cells. So, if the person’s energy state is the driver of POMC cell activation, then the energy state of humans should be controlled to activate or inhibit the POMC cells. We need to pause here to remind ourselves, that in a string of biologic actions, the earliest event is the cause. Therefore, food intake and resultant energy state of the host are the cause of POMC cell activation. Our view is supported by Beck and Rau who stated that “POMC cell stimulation or inhibition is the consequence of food intake and the resultant energy state.”[61,67]
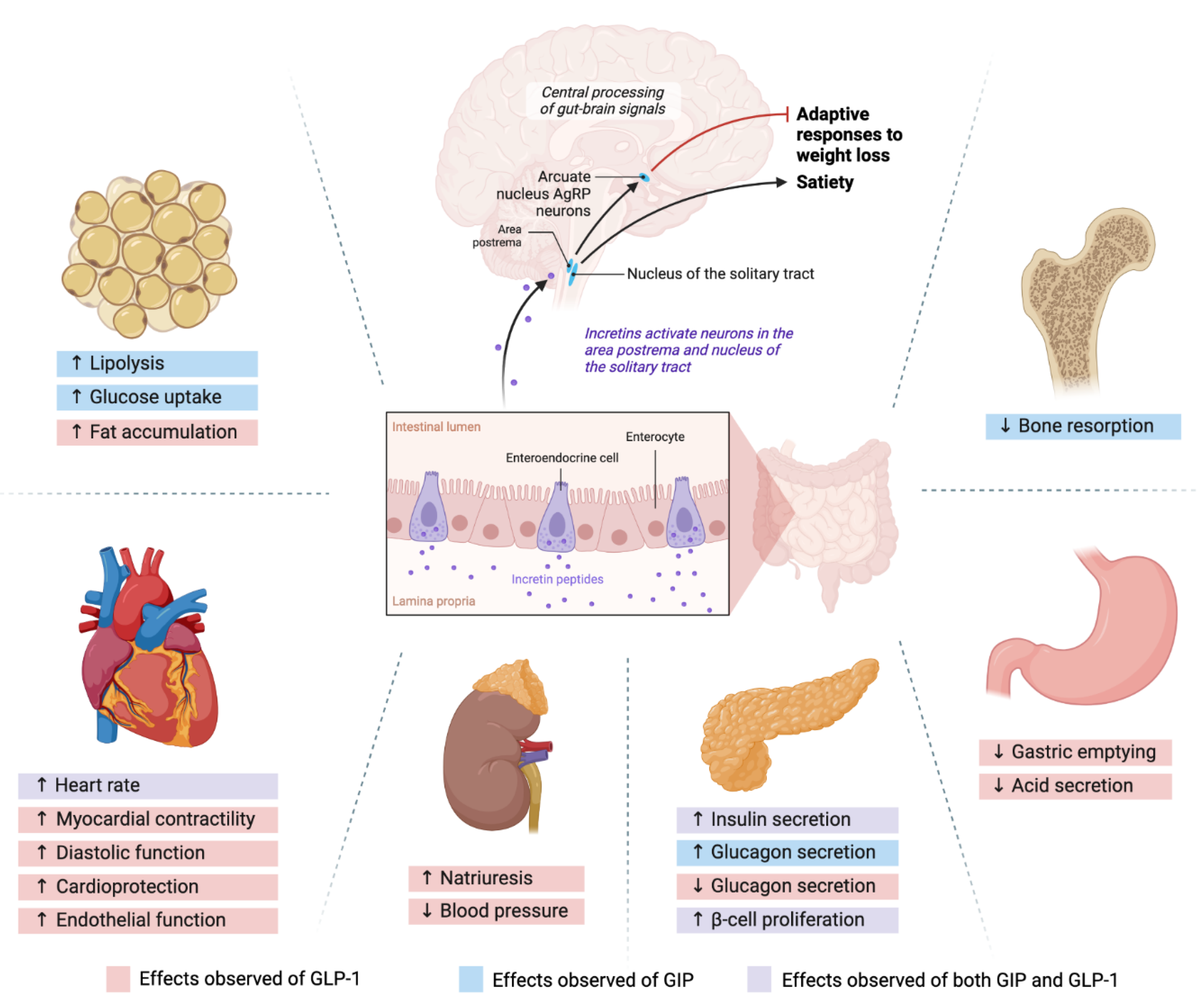
The pharmacological incretin agonism manipulates these neural axes, and control the person’s appetite. However, once these neural inhibitions are lifted, weight regain can occur.[24,48] After discontinuation of tirzepatide, the study participants regained almost 2/3 of their lost weight within 4 weeks.[48] We will suggest a strategy to mitigate the weight regain after discontinuation of incretin agonists in the FUTURE DIRECTIONS section. These mechanisms and neurohormonal pathways are illustrated in Figure 1.
3. Challenges in Incretin Agonism
- a.
- Involvement of β-Arrestins
As stated in section1-a, incretin and glucagon receptors (GCGR) belong to the class B GPCR and their efficacy is limited by β-arrestins. [69] β-arrestin-1 (β1arrs) and β-arrestin-2 (Β2arrs) are best known for their ability to mediate the desensitization and internalization of GPCRs. [70] The effect of B2arrs on insulin secretion was dominated by GLP-1R- and no clear GCGR-dependent effects are observed.[69]
Β2arrs are ubiquitously expressed and function as negative regulators of GPCR signaling by inhibiting GPCR and G protein coupling, via uncoupling cyclic AMP (cAMP)/protein kinase A (PKA) signaling at physiological levels of GLP-1.[71] This is a process called desensitization which dampens insulin secretion. Next, β2-arres scaffold enzymes, phosphodiesterase and diacylglycerol kinase, degrade second messengers generated by G protein activity. This will solidify the desensitization process.[72]
At pharmacological doses, the activation of extracellular signal-related kinase (ERK)/cAMP-responsive element-binding protein (CREB) requires Β2arrs. Also, GIP-dependent insulin secretion needs Β2arrs in human islets.[71]
β2arres involvement is a potential drawback of incretin agonism because it will cause plateau in the drug efficacies. Avoiding Β2arrs-dependent GPCR desensitization may alleviate the problem of tachyphylaxis of GLP-1 and GIP agonism.[73] The recent development of Tirzepatide is based on the previous study that revealed islets β-arrestin1 limits the insulin response of GLP-1, but not GIP, thus may not affect insulin secretion by GIP. [74]
- b.
- β-Cell Exhaustion and Failure
Continuing stimulation of pancreatic β-cells may cause receptor downregulation and desensitization. A good example of this phenomenon is the insulin resistance in T2D patients.[75] Forcing the β-cells to secrete insulin when they are overwhelmed by insulin demand may accelerate β-cell failure as shown with sulfonylurea administration.[76] Simply put, chronic agonism eventually reduces the number of receptors leading to a condition similar to failure of function.[77] [78]
The mechanism of β-cell failure is presumed that overstimulation of β-cell increases cytoplasmic Ca++ levels, and persistent elevation of cytoplasmic Ca++ may trigger apoptosis of β-cells.[79] Under chronic agonism, some β-cells fail to differentiate properly and lose their identity.[80]
While these examples describe overstimulation of β-cells by hyperglycemia in T2D, chronic agonism by incretins may exert similar effects. [76,81,82] Indeed, animal studies reported increased pyknotic nuclei (a sign of apoptosis) of β-cells and acinar inflammation in rats given a high dose of exenatide.[83]
4. Potential Adverse Effects of Incretin Agonism
Although newer incretins appear to be much safer, increased risk of biliary disease,[12] pancreatitis,[84] bowel obstruction,[85] and gastroparesis have been noted. [86] Also an increased risk of pancreatic or thyroid cancers has been reported. [87,88] However, conflicting non-significant results on cancer risk were also observed. [89] But this study has very short follow-up (3.9 years) and compared GLP-1ra to dipeptidyl peptidase 4 inhibitors (DPP4i) which is on the causal pathway of GLP-1ra and thus, is an inappropriate comparator.
It is plausible that insulin secretion modulated by incretin agonism may affect pancreatitis and pancreatic cancer, because of their shared pathway. Indeed increased risks for multiple pathologies were reported.[90] Sodhi et al. [90] using a large health claims data set reported that the use of GLP-1ra compared with bupropion/naltrexone was associated with a 9-fold increased risk of pancreatitis, a 4.2-fold increase of bowel obstruction, and a 3.7- fold increase in gastroparesis. [90]
The cause of pancreatitis associated with GLP-1ra is hypothesized as incretin agonism may promote pancreatic hyperplasia, leading to increased pancreatic weight and exocrine duct occlusion. [91] However, the incidence of pancreatitis resulting from the incretin agonism is relatively rare and only large data sets such as pharmacovigilance data or insurance claims data will show significant results.
Using large historical population data, Dankner et al. reported no increase in GLP-1ra-associated pancreatic cancer incidence compared to insulin and metformin.[92] However, this study is fraught with selection bias.[92] Insulin is a well-known oncogene and this will generate selection bias. Choosing a worse comparator generate an aura of erroneously making GLP-1ras apper innocuous. [92] Further well-conducted studies are needed.
Other adverse effects such as retinopathy,[93] gastroparesis and bowel obstruction[85] were also reported. Lu and colleagues [94] explained the mechanism of intestinal obstruction as “long-term application of GLP-1ras may also elevate the release of endogenous GLP-2. GLP-2 is a cell-specific growth hormone regulating the growth of the small intestine, colonic villi and crypts, increasing villi height, and reducing antral motility.”. [94] This study provides the scientific basis for the intestinal blockage and raises concerns for the long-term use of incretin agonism. [94]
Recently several cases of pancreatitis and one fatality associated with Tirzepatide [95] have been reported. As more people use this class of medications, more serious adverse events may emerge. Especially, for tirzepatide with added GIP which masks the nausea and vomiting,[37] and allows the patients to tolerate the medication better and to take it for a longer duration. This may increase more serious adverse events.
Additionally, a case of leukocytoclastic vasculitis (LCV) induced by once-weekly subcutaneous semaglutide has also been reported. [96] Although 50% of LCV can be idiopathic, immune dysregulation is presumed cause. Especially, complete resolution of the lesions shortly after the discontinuation of semaglutide proves the causal role of semaglutide on LCV. [96]
Also, GLP-1R and GCGR agonism may increase heart rate (HR) [54,57,97,98] and sympathetic tone.[99,100] Both glucagon[101] and GLP-1[102] are known to increase sympathetic tone.[103,104] Although some studies reported no increase in HR , animal studies suggest that GLP-1 acts directly on the sinus node.[55,56]
The mechanism that glucagon receptor (GCGR) activation leads to increased HR is via adenylyl cyclase activation and increases in 3',5'-c- AMP production in the myocardium. [54] For this reason, glucagon is used as an antidote for β-blocker overdose.[101] It is conceivable that glucagon is associated with sympathetic tone because hypoglycemia activates stress response and the hypothalamic-pituitary-adrenal axis. [105,106,107,108]
The mechanism that GLP-1R contributes to increased HR is reasoned that the preproglucagon neurons that mediate GLP-1-associated anorexia are situated in the brainstem to receive signals of stress. This suggests a potential link with the sympathetic nervous system. [102,103] However, another study reported that the initial increase in sympathetic tone was reversed by GLP-1R activation via by acetylcholine and nitric oxide.[109] Making it more confusing, in a human trial, the addition of liraglutide to exercise appeared to blunt the beneficial effects of exercise on left ventricular diastolic function. [52] Many systematic reviews do suggest cardiac protective effects of incretins. [110,111] However, the absolute benefit size was only a 1.5% decrease. [112] Also, a meta-analysis of the CVD benefits of dulaglutide showed that all individual studies include relative risk of 1 (no effect). This means that the results are not statistically significant. [113]
5. Future Directions
How do we harness the observed beneficial effects of incretin agonism on glucose levels and weight loss minimizing potential harm? These are the wishes of experts and patients alike. They all raised the concern for potential adverse events from a long-term use of incretin agonism, sustainability of their efficacy, and the high cost. [114,115,116] We concur with the experts and suggest that the remarkable weight loss and glycemic control by incretin agonism should be juxtaposed to the high cost, it’s unsustainability of efficacy, and potential adverse effects. As we stated in the neuro-hormonal control of appetite section, manipulating neuronal axis will fail when the manipulation terminates. Thus, tapering GLP-1 agonism and enhancing lifestyle modification may be the key to healthy weight maintenance. It is pivotal to teach the patients so that they can make healthier food choices that maintain satiety [117], encouraging physical activities. A group in Denmark had done this by slowly tapering semaglutide and applying intense lifestyle modification in a trial.[118] More longer-term studies are needed emphasizing the lifestyle modifications in addition to tapering GLP-1ras.
References
- BARRE, L. Sur les possibilities d'un traitement du diabete par. L'incretine. Bull Acad R Med Belg. 1932;12:620-34.
- Gupta K, Raja A. Physiology, Gastric Inhibitory Peptide. 2024.
- Holst, J.J. The Physiology of Glucagon-like Peptide 1. Physiol. Rev. 2007, 87, 1409–1439. [Google Scholar] [CrossRef] [PubMed]
- Svendsen, B.; Pais, R.; Engelstoft, M.S.; Milev, N.B.; Richards, P.; Christiansen, C.B.; Egerod, K.L.; Jensen, S.M.; Habib, A.M.; Gribble, F.M.; et al. GLP1- and GIP-producing cells rarely overlap and differ by bombesin receptor-2 expression and responsiveness. J. Endocrinol. 2015, 228, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Adriaenssens, A.E.; Biggs, E.K.; Darwish, T.; Tadross, J.; Sukthankar, T.; Girish, M.; Polex-Wolf, J.; Lam, B.Y.; Zvetkova, I.; Pan, W.; et al. Glucose-Dependent Insulinotropic Polypeptide Receptor-Expressing Cells in the Hypothalamus Regulate Food Intake. Cell Metab. 2019, 30, 987–996. [Google Scholar] [CrossRef] [PubMed]
- Liskiewicz, A.; Khalil, A.; Liskiewicz, D.; Novikoff, A.; Grandl, G.; Maity-Kumar, G.; Gutgesell, R.M.; Bakhti, M.; Bastidas-Ponce, A.; Czarnecki, O.; et al. Glucose-dependent insulinotropic polypeptide regulates body weight and food intake via GABAergic neurons in mice. Nat. Metab. 2023, 5, 2075–2085. [Google Scholar] [CrossRef]
- Martinez de Morentin PB, Gonzalez JA, Dowsett GKC, Martynova Y, Yeo GSH, Sylantyev S, et al. A brainstem to hypothalamic arcuate nucleus GABAergic circuit drives feeding. Curr Biol. 2024;34(8):1646-56.e4. [CrossRef]
- Asmar, M.; Tangaa, W.; Madsbad, S.; Hare, K.; Astrup, A.; Flint, A.; Bülow, J.; Holst, J.J. On the role of glucose-dependent insulintropic polypeptide in postprandial metabolism in humans. Am. J. Physiol. Metab. 2010, 298, E614–E621. [Google Scholar] [CrossRef]
- Meier, J.J.; Goetze, O.; Anstipp, J.; Hagemann, D.; Holst, J.J.; Schmidt, W.E.; Gallwitz, B.; Nauck, M.A. Gastric inhibitory polypeptide does not inhibit gastric emptying in humans. Am. J. Physiol. Metab. 2004, 286, E621–E625. [Google Scholar] [CrossRef]
- Bergmann, N.C.; Lund, A.; Gasbjerg, L.S.; Meessen, E.C.E.; Andersen, M.M.; Bergmann, S.; Hartmann, B.; Holst, J.J.; Jessen, L.; Christensen, M.B.; et al. Effects of combined GIP and GLP-1 infusion on energy intake, appetite and energy expenditure in overweight/obese individuals: a randomised, crossover study. Diabetologia 2019, 62, 665–675. [Google Scholar] [CrossRef]
- Rubino D, Abrahamsson N, Davies M, Hesse D, Greenway FL, Jensen C, et al. Effect of Continued Weekly Subcutaneous Semaglutide vs Placebo on Weight Loss Maintenance in Adults With Overweight or Obesity: The STEP 4 Randomized Clinical Trial. JAMA. 2021;325(14):1414-25. [CrossRef]
- Rubino, D.M.; Greenway, F.L.; Khalid, U.; O’neil, P.M.; Rosenstock, J.; Sørrig, R.; Wadden, T.A.; Wizert, A.; Garvey, W.T.; STEP 8 Investigators; et al. Effect of Weekly Subcutaneous Semaglutide vs Daily Liraglutide on Body Weight in Adults With Overweight or Obesity Without Diabetes. JAMA 2022, 327, 138–150. [Google Scholar] [CrossRef]
- Frias, J.P.; Bonora, E.; Ruiz, L.N.; Li, Y.G.; Yu, Z.; Milicevic, Z.; Malik, R.; Bethel, M.A.; Cox, D.A. Efficacy and Safety of Dulaglutide 3.0 mg and 4.5 mg Versus Dulaglutide 1.5 mg in Metformin-Treated Patients With Type 2 Diabetes in a Randomized Controlled Trial (AWARD-11). Diabetes Care 2021, 44, 765–773. [Google Scholar] [CrossRef]
- Fujioka, K.; Lau, D.C.W.; Van Gaal, L.; Wilding, J.P.H.; Skjøth, T.V.; Manning, L.S.; Pi-Sunyer, X.; Hamann, A.; Barakat, A.; Blüher, M.; et al. 3 years of liraglutide versus placebo for type 2 diabetes risk reduction and weight management in individuals with prediabetes: a randomised, double-blind trial. Lancet 2017, 389, 1399–1409. [Google Scholar] [CrossRef]
- Husain, M.; Birkenfeld, A.L.; Donsmark, M.; Dungan, K.; Eliaschewitz, F.G.; Franco, D.R.; Jeppesen, O.K.; Lingvay, I.; Mosenzon, O.; Pedersen, S.D.; et al. Oral Semaglutide and Cardiovascular Outcomes in Patients with Type 2 Diabetes. N. Engl. J. Med. 2019, 381, 841–851. [Google Scholar] [CrossRef] [PubMed]
- Heerspink, H.J.L.; Sattar, N.; Pavo, I.; Haupt, A.; Duffin, K.L.; Yang, Z.; Wiese, R.J.; Tuttle, K.R.; I Cherney, D.Z. Effects of tirzepatide versus insulin glargine on kidney outcomes in type 2 diabetes in the SURPASS-4 trial: post-hoc analysis of an open-label, randomised, phase 3 trial. Lancet Diabetes Endocrinol. 2022, 10, 774–785. [Google Scholar] [CrossRef] [PubMed]
- Frías, J.P.; Davies, M.J.; Rosenstock, J.; Pérez Manghi, F.C.; Fernández Landó, L.; Bergman, B.K.; Liu, B.; Cui, X.; Brown, K. Tirzepatide versus Semaglutide Once Weekly in Patients with Type 2 Diabetes. N. Engl. J. Med. 2021, 385, 503–515. [Google Scholar] [CrossRef] [PubMed]
- Rosenstock, J.; Frias, J.; Jastreboff, A.M.; Du, Y.; Lou, J.; Gurbuz, S.; Thomas, M.K.; Hartman, M.L.; Haupt, A.; Milicevic, Z.; et al. Retatrutide, a GIP, GLP-1 and glucagon receptor agonist, for people with type 2 diabetes: a randomised, double-blind, placebo and active-controlled, parallel-group, phase 2 trial conducted in the USA. Lancet 2023, 402, 529–544. [Google Scholar] [CrossRef]
- Rosenstock, J.; Wysham, C.; Frías, J.P.; Kaneko, S.; Lee, C.J.; Landó, L.F.; Mao, H.; Cui, X.; A Karanikas, C.A.; Thieu, V.T. Efficacy and safety of a novel dual GIP and GLP-1 receptor agonist tirzepatide in patients with type 2 diabetes (SURPASS-1): a double-blind, randomised, phase 3 trial. Lancet 2021, 398, 143–155. [Google Scholar] [CrossRef]
- Jastreboff, A.M.; Jastreboff, A.M.; Aronne, L.J.; Aronne, L.J.; Ahmad, N.N.; Ahmad, N.N.; Wharton, S.; Wharton, S.; Connery, L.; Connery, L.; et al. Tirzepatide Once Weekly for the Treatment of Obesity. New Engl. J. Med. 2022, 387, 205–216. [Google Scholar] [CrossRef]
- Jastreboff, A.M.; Kaplan, L.M.; Frías, J.P.; Wu, Q.; Du, Y.; Gurbuz, S.; Coskun, T.; Haupt, A.; Milicevic, Z.; Hartman, M.L. Triple–Hormone-Receptor Agonist Retatrutide for Obesity — A Phase 2 Trial. New Engl. J. Med. 2023, 389, 514–526. [Google Scholar] [CrossRef]
- Ludvik, B.; Giorgino, F.; Jódar, E.; Frias, J.P.; Landó, L.F.; Brown, K.; Bray, R.; Rodríguez. Once-weekly tirzepatide versus once-daily insulin degludec as add-on to metformin with or without SGLT2 inhibitors in patients with type 2 diabetes (SURPASS-3): a randomised, open-label, parallel-group, phase 3 trial. Lancet 2021, 398, 583–598. [Google Scholar] [CrossRef]
- Del Prato S, Kahn SE, Pavo I, Weerakkody GJ, Yang Z, Doupis J, et al. Tirzepatide versus insulin glargine in type 2 diabetes and increased cardiovascular risk (SURPASS-4): a randomised, open-label, parallel-group, multicentre, phase 3 trial. Lancet. 2021;398(10313):1811-24. [CrossRef]
- Wilding, J.P.; Batterham, R.L.; Davies, M.; Van Gaal, L.F.; Kandler, K.; Konakli, K.; Lingvay, I.; McGowan, B.M.; Oral, T.K.; Rosenstock, J.; et al. Author response for "Weight regain and cardiometabolic effects after withdrawal of semaglutide: the STEP 1 trial extension". 2022. [Google Scholar] [CrossRef]
- Dahl, D.; Onishi, Y.; Norwood, P.; Huh, R.; Bray, R.; Patel, H.; Rodríguez. Effect of Subcutaneous Tirzepatide vs Placebo Added to Titrated Insulin Glargine on Glycemic Control in Patients With Type 2 Diabetes. JAMA 2022, 327, 534–545. [Google Scholar] [CrossRef]
- Lincoff, A.M.; Brown-Frandsen, K.; Colhoun, H.M.; Deanfield, J.; Emerson, S.S.; Esbjerg, S.; Hardt-Lindberg, S.; Hovingh, G.K.; Kahn, S.E.; Kushner, R.F.; et al. Semaglutide and Cardiovascular Outcomes in Obesity without Diabetes. New Engl. J. Med. 2023, 389, 2221–2232. [Google Scholar] [CrossRef]
- Aronne, L.J.; Sattar, N.; Horn, D.B.; Bays, H.E.; Wharton, S.; Lin, W.-Y.; Ahmad, N.N.; Zhang, S.; Liao, R.; Bunck, M.C.; et al. Continued Treatment With Tirzepatide for Maintenance of Weight Reduction in Adults With Obesity. JAMA 2024, 331, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Loomba, R.; Hartman, M.L.; Lawitz, E.J.; Vuppalanchi, R.; Boursier, J.; Bugianesi, E.; Yoneda, M.; Behling, C.; Cummings, O.W.; Tang, Y.; et al. Tirzepatide for Metabolic Dysfunction–Associated Steatohepatitis with Liver Fibrosis. New Engl. J. Med. 2024, 391, 299–310. [Google Scholar] [CrossRef] [PubMed]
- Sanyal, A.J.; Bedossa, P.; Fraessdorf, M.; Neff, G.W.; Lawitz, E.; Bugianesi, E.; Anstee, Q.M.; Hussain, S.A.; Newsome, P.N.; Ratziu, V.; et al. A Phase 2 Randomized Trial of Survodutide in MASH and Fibrosis. New Engl. J. Med. 2024, 391, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Sanyal, A.J.; Kaplan, L.M.; Frias, J.P.; Brouwers, B.; Wu, Q.; Thomas, M.K.; Harris, C.; Schloot, N.C.; Du, Y.; Mather, K.J.; et al. Triple hormone receptor agonist retatrutide for metabolic dysfunction-associated steatotic liver disease: a randomized phase 2a trial. Nat. Med. 2024, 30, 2037–2048. [Google Scholar] [CrossRef]
- Piccini, S.; Favacchio, G.; Panico, C.; Morenghi, E.; Folli, F.; Mazziotti, G.; Lania, A.G.; Mirani, M. Time-dependent effect of GLP-1 receptor agonists on cardiovascular benefits: a real-world study. Cardiovasc. Diabetol. 2023, 22, 1–14. [Google Scholar] [CrossRef]
- Nauck, M.A.; Meier, J.J. GIP and GLP-1: Stepsiblings Rather Than Monozygotic Twins Within the Incretin Family. Diabetes 2019, 68, 897–900. [Google Scholar] [CrossRef]
- A Nauck, M.; Meier, J.J. The incretin effect in healthy individuals and those with type 2 diabetes: physiology, pathophysiology, and response to therapeutic interventions. Lancet Diabetes Endocrinol. 2016, 4, 525–536. [Google Scholar] [CrossRef]
- El, K.; Campbell, J.E. The role of GIP in α-cells and glucagon secretion. Peptides 2019, 125, 170213–170213. [Google Scholar] [CrossRef]
- Drucker, D.J. Incretin Action in the Pancreas: Potential Promise, Possible Perils, and Pathological Pitfalls. Diabetes 2013, 62, 3316–3323. [Google Scholar] [CrossRef]
- Urva S, Coskun T, Loghin C, Cui X, Beebe E, O'Farrell L, et al. The novel dual glucose-dependent insulinotropic polypeptide and glucagon-like peptide-1 (GLP-1) receptor agonist tirzepatide transiently delays gastric emptying similarly to selective long-acting GLP-1 receptor agonists. Diabetes Obes Metab. 2020;22(10):1886-91. [CrossRef]
- Hayes, M.R.; Borner, T.; De Jonghe, B.C. The Role of GIP in the Regulation of GLP-1 Satiety and Nausea. Diabetes 2021, 70, 1956–1961. [Google Scholar] [CrossRef]
- Hayes, M.R.; Mietlicki-Baase, E.G.; Kanoski, S.E.; De Jonghe, B.C. Incretins and Amylin: Neuroendocrine Communication Between the Gut, Pancreas, and Brain in Control of Food Intake and Blood Glucose. Annu. Rev. Nutr. 2014, 34, 237–260. [Google Scholar] [CrossRef] [PubMed]
- Kiriyama, Y.; Nochi, H. Role and Cytotoxicity of Amylin and Protection of Pancreatic Islet β-Cells from Amylin Cytotoxicity. Cells 2018, 7, 95. [Google Scholar] [CrossRef] [PubMed]
- Kosinski JR, Hubert J, Carrington PE, Chicchi GG, Mu J, Miller C, et al. The glucagon receptor is involved in mediating the body weight-lowering effects of oxyntomodulin. Obesity (Silver Spring). 2012;20(8):1566-71. [CrossRef]
- Geisler, C.E.; Renquist, B.J. Hepatic lipid accumulation: cause and consequence of dysregulated glucoregulatory hormones. J. Endocrinol. 2017, 234, R1–R21. [Google Scholar] [CrossRef]
- Wang, H.; Zhao, M.; Sud, N.; Christian, P.; Shen, J.; Song, Y.; Pashaj, A.; Zhang, K.; Carr, T.; Su, Q. Glucagon regulates hepatic lipid metabolism via cAMP and Insig-2 signaling: implication for the pathogenesis of hypertriglyceridemia and hepatic steatosis. Sci. Rep. 2016, 6, srep32246. [Google Scholar] [CrossRef]
- Christensen, M.; Vedtofte, L.; Holst, J.J.; Vilsbøll, T.; Knop, F.K. Glucose-Dependent Insulinotropic Polypeptide: a bifunctional glucose-dependent regulator of glucagon and insulin secretion in humans. Diabetes 2011, 60, 3103–3109. [Google Scholar] [CrossRef]
- Janket SJ, Javaheri H, Ackerson LK, Ayilavarapu S, Meurman JH. Oral Infections, Metabolic Inflammation, Genetics, and Cardiometabolic Diseases. Journal of dental research. 2015;94(9 Suppl):119s-27s.
- Oussaada, S.; Kilicarslan, M.; de Weijer, B.; Gilijamse, P.; Şekercan, A.; Virtue, S.; Janssen, I.; van de Laar, A.; Demirkiran, A.; van Wagensveld, B.; et al. Tissue-specific inflammation and insulin sensitivity in subjects with obesity. Diabetes Res. Clin. Pr. 2024, 211, 111663. [Google Scholar] [CrossRef]
- Friedrichsen, M.; Breitschaft, A.; Tadayon, S.; Wizert, A.; Skovgaard, D. The effect of semaglutide 2.4 mg once weekly on energy intake, appetite, control of eating, and gastric emptying in adults with obesity. Diabetes, Obes. Metab. 2021, 23, 754–762. [Google Scholar] [CrossRef]
- Maida, A.; Lovshin, J.A.; Baggio, L.L.; Drucker, D.J. The Glucagon-Like Peptide-1 Receptor Agonist Oxyntomodulin Enhances β-Cell Function but Does Not Inhibit Gastric Emptying in Mice. Endocrinology 2008, 149, 5670–5678. [Google Scholar] [CrossRef]
- Rosenstock, J.; Lee, C.J.; Landó, L.F.; Liu, M.; Karanikas, C.A.; Thieu, V.T. Impact on glycated haemoglobin and body weight changes after stopping tirzepatide for 4 weeks in the SURPASS-1 monotherapy trial. Diabetes, Obes. Metab. 2023, 26, 396–399. [Google Scholar] [CrossRef]
- Ackeifi, C.; Wang, P.; Karakose, E.; Fox, J.E.M.; González, B.J.; Liu, H.; Wilson, J.; Swartz, E.; Berrouet, C.; Li, Y.; et al. GLP-1 receptor agonists synergize with DYRK1A inhibitors to potentiate functional human β cell regeneration. Sci. Transl. Med. 2020, 12. [Google Scholar] [CrossRef]
- Cheng, L.; Hu, Y.; Li, Y.; Cao, X.; Bai, N.; Lu, T.; Li, G.; Li, N.; Wang, A.; Mao, X. Glucagon-like peptide-1 receptor agonists and risk of bone fracture in patients with type 2 diabetes: A meta-analysis of randomized controlled trials. Diabetes/Metabolism Res. Rev. 2019, 35, e3168. [Google Scholar] [CrossRef] [PubMed]
- Jensen, S.B.K.; Sørensen, V.; Sandsdal, R.M.; Lehmann, E.W.; Lundgren, J.R.; Juhl, C.R.; Janus, C.; Ternhamar, T.; Stallknecht, B.M.; Holst, J.J.; et al. Bone Health After Exercise Alone, GLP-1 Receptor Agonist Treatment, or Combination Treatment. JAMA Netw. Open 2024, 7, e2416775. [Google Scholar] [CrossRef] [PubMed]
- Jørgensen, P.G.; Jensen, M.T.; Mensberg, P.; Storgaard, H.; Nyby, S.; Jensen, J.S.; Knop, F.K.; Vilsbøll, T. Effect of exercise combined with glucagon-like peptide-1 receptor agonist treatment on cardiac function: A randomized double-blind placebo-controlled clinical trial. Diabetes, Obes. Metab. 2017, 19, 1040–1044. [Google Scholar] [CrossRef]
- Dunning, B.E.; Gerich, J.E. The Role of α-Cell Dysregulation in Fasting and Postprandial Hyperglycemia in Type 2 Diabetes and Therapeutic Implications. Endocr. Rev. 2007, 28, 253–283. [Google Scholar] [CrossRef]
- Hernández-Cascales, J. Does glucagon have a positive inotropic effect in the human heart? Cardiovasc. Diabetol. 2018, 17, 1–4. [Google Scholar] [CrossRef]
- Kaur, S.; A Rose, R. New insights into the effects of glucagon-like peptide-1 on heart rate and sinoatrial node function. Cardiovasc. Res. 2024. [Google Scholar] [CrossRef]
- Lubberding, A.F.; Veedfald, S.; Achter, J.S.; Nissen, S.D.; Soattin, L.; Sorrentino, A.; Vega, E.T.; Linz, B.; Eggertsen, C.H.E.; Mulvey, J.; et al. GLP-1 increases heart rate by a direct action on the sinus node. Cardiovasc. Res. 2024. [Google Scholar] [CrossRef]
- Petersen, K.M.; Bøgevig, S.; Riis, T.; Andersson, N.W.; Dalhoff, K.P.; Holst, J.J.; Knop, F.K.; Faber, J.; Petersen, T.S.; Christensen, M.B. High-Dose Glucagon Has Hemodynamic Effects Regardless of Cardiac Beta-Adrenoceptor Blockade: A Randomized Clinical Trial. J. Am. Hear. Assoc. 2020, 9, e016828. [Google Scholar] [CrossRef]
- Rossi, M.A.; Stuber, G.D. Overlapping Brain Circuits for Homeostatic and Hedonic Feeding. Cell Metab. 2017, 27, 42–56. [Google Scholar] [CrossRef]
- van Bloemendaal, L.; Kulve, J.S.T.; E la Fleur, S.; Ijzerman, R.G.; Diamant, M. Effects of glucagon-like peptide 1 on appetite and body weight: focus on the CNS. J. Endocrinol. 2013, 221, T1–T16. [Google Scholar] [CrossRef]
- Delgado, T.C. Glutamate and GABA in Appetite Regulation. Front. Endocrinol. 2013, 4, 103. [Google Scholar] [CrossRef] [PubMed]
- Rau, A.R.; Hentges, S.T. GABAergic Inputs to POMC Neurons Originating from the Dorsomedial Hypothalamus Are Regulated by Energy State. J. Neurosci. 2019, 39, 6449–6459. [Google Scholar] [CrossRef]
- Toda, C.; Santoro, A.; Kim, J.D.; Diano, S. POMC Neurons: From Birth to Death. Annu. Rev. Physiol. 2017, 79, 209–236. [Google Scholar] [CrossRef] [PubMed]
- Sohn, J.-W.; Xu, Y.; Jones, J.E.; Wickman, K.; Williams, K.W.; Elmquist, J.K. Serotonin 2C Receptor Activates a Distinct Population of Arcuate Pro-opiomelanocortin Neurons via TRPC Channels. Neuron 2011, 71, 488–497. [Google Scholar] [CrossRef] [PubMed]
- Williams, K.W.; Margatho, L.O.; Lee, C.E.; Choi, M.; Lee, S.; Scott, M.M.; Elias, C.F.; Elmquist, J.K. Segregation of Acute Leptin and Insulin Effects in Distinct Populations of Arcuate Proopiomelanocortin Neurons. J. Neurosci. 2010, 30, 2472–2479. [Google Scholar] [CrossRef]
- Vong, L.; Ye, C.; Yang, Z.; Choi, B.; Chua, S., Jr.; Lowell, B.B. Leptin Action on GABAergic Neurons Prevents Obesity and Reduces Inhibitory Tone to POMC Neurons. Neuron 2011, 71, 142–154. [Google Scholar] [CrossRef]
- Secher, A.; Jelsing, J.; Baquero, A.F.; Hecksher-Sørensen, J.; Cowley, M.A.; Dalbøge, L.S.; Hansen, G.; Grove, K.L.; Pyke, C.; Raun, K.; et al. The arcuate nucleus mediates GLP-1 receptor agonist liraglutide-dependent weight loss. J. Clin. Investig. 2014, 124, 4473–4488. [Google Scholar] [CrossRef]
- Beck, B. Neuropeptide Y in normal eating and in genetic and dietary-induced obesity. Philos. Trans. R. Soc. B: Biol. Sci. 2006, 361, 1159–1185. [Google Scholar] [CrossRef]
- Williams, D.L. The diverse effects of brain glucagon-like peptide 1 receptors on ingestive behaviour. Br. J. Pharmacol. 2021, 179, 571–583. [Google Scholar] [CrossRef]
- Jones, B.; McGlone, E.R.; Fang, Z.; Pickford, P.; Corrêa, I.R.; Oishi, A.; Jockers, R.; Inoue, A.; Kumar, S.; Görlitz, F.; et al. Genetic and biased agonist-mediated reductions in β-arrestin recruitment prolong cAMP signaling at glucagon family receptors. J. Biol. Chem. 2021, 296, 100133. [Google Scholar] [CrossRef]
- Bitsi, S.; El Eid, L.; Manchanda, Y.; Oqua, A.I.; Mohamed, N.; Hansen, B.; Suba, K.; Rutter, G.A.; Salem, V.; Jones, B.; et al. Divergent acute versus prolonged pharmacological GLP-1R responses in adult β cell–specific β-arrestin 2 knockout mice. Sci. Adv. 2023, 9, eadf7737. [Google Scholar] [CrossRef] [PubMed]
- Zaïmia, N.; Obeid, J.; Varrault, A.; Sabatier, J.; Broca, C.; Gilon, P.; Costes, S.; Bertrand, G.; Ravier, M.A. GLP-1 and GIP receptors signal through distinct β-arrestin 2-dependent pathways to regulate pancreatic β cell function. Cell Rep. 2023, 42, 113326. [Google Scholar] [CrossRef] [PubMed]
- Jean-Charles, P.-Y.; Kaur, S.; Shenoy, S.K. G Protein–Coupled Receptor Signaling Through β-Arrestin–Dependent Mechanisms. J. Cardiovasc. Pharmacol. 2017, 70, 142–158. [Google Scholar] [CrossRef] [PubMed]
- Peterson, Y.K.; Luttrell, L.M. The Diverse Roles of Arrestin Scaffolds in G Protein–Coupled Receptor Signaling. Pharmacol. Rev. 2017, 69, 256–297. [Google Scholar] [CrossRef]
- Willard, F.S.; Douros, J.D.; Gabe, M.B.; Showalter, A.D.; Wainscott, D.B.; Suter, T.M.; Capozzi, M.E.; van der Velden, W.J.; Stutsman, C.; Cardona, G.R.; et al. Tirzepatide is an imbalanced and biased dual GIP and GLP-1 receptor agonist. J. Clin. Investig. 2020, 5. [Google Scholar] [CrossRef]
- Campbell, J.E.; Newgard, C.B. Mechanisms controlling pancreatic islet cell function in insulin secretion. Nat. Rev. Mol. Cell Biol. 2021, 22, 142–158. [Google Scholar] [CrossRef]
- Aston-Mourney, K.; Proietto, J.; Morahan, G.; Andrikopoulos, S. Too much of a good thing: why it is bad to stimulate the beta cell to secrete insulin. Diabetologia 2008, 51, 540–545. [Google Scholar] [CrossRef]
- Killion, E.A.; Chen, M.; Falsey, J.R.; Sivits, G.; Hager, T.; Atangan, L.; Helmering, J.; Lee, J.; Li, H.; Wu, B.; et al. Chronic glucose-dependent insulinotropic polypeptide receptor (GIPR) agonism desensitizes adipocyte GIPR activity mimicking functional GIPR antagonism. Nat. Commun. 2020, 11, 1–17. [Google Scholar] [CrossRef]
- Campbell, J.E. Targeting the GIPR for obesity: To agonize or antagonize? Potential mechanisms. Mol. Metab. 2020, 46, 101139. [Google Scholar] [CrossRef]
- Grill, V.; Björklund, A. Overstimulation and beta-cell function. Diabetes 2001, 50, S122–4. [Google Scholar] [CrossRef]
- Swisa, A.; Glaser, B.; Dor, Y. Metabolic Stress and Compromised Identity of Pancreatic Beta Cells. Front. Genet. 2017, 08, 21. [Google Scholar] [CrossRef]
- Kolb, H.; Kempf, K.; Röhling, M.; Martin, S. Insulin: too much of a good thing is bad. BMC Med. 2020, 18, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Lebovitz, H.E. Insulin: Potential Negative Consequences of Early Routine Use in Patients With Type 2 Diabetes. Diabetes Care 2011, 34, S225–S230. [Google Scholar] [CrossRef] [PubMed]
- Łabuzek, K.; Kozłowski, M.; Szkudłapski, D.; Sikorska, P.; Kozłowska, M.; Okopień, B. Incretin-based therapies in the treatment of type 2 diabetes — More than meets the eye? Eur. J. Intern. Med. 2013, 24, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Singh S, Chang HY, Richards TM, Weiner JP, Clark JM, Segal JB. Glucagonlike peptide 1-based therapies and risk of hospitalization for acute pancreatitis in type 2 diabetes mellitus: a population-based matched case-control study. JAMA Intern Med. 2013;173(7):534-9. [CrossRef]
- Gudin, B.; Ladhari, C.; Robin, P.; Laroche, M.-L.; Babai, S.; Hillaire-Buys, D.; Faillie, J.-L. Incretin-based drugs and intestinal obstruction: A pharmacovigilance study. Therapies 2020, 75, 641–647. [Google Scholar] [CrossRef]
- Kalas, M.A.; Galura, G.M.; McCallum, R.W. Medication-Induced Gastroparesis: A Case Report. J. Investig. Med. High Impact Case Rep. 2021, 9. [Google Scholar] [CrossRef]
- Monami, M.; Nreu, B.; Scatena, A.; Cresci, B.; Andreozzi, F.; Sesti, G.; Mannucci, E. Safety issues with glucagon-like peptide-1 receptor agonists (pancreatitis, pancreatic cancer and cholelithiasis): Data from randomized controlled trials. Diabetes, Obes. Metab. 2017, 19, 1233–1241. [Google Scholar] [CrossRef]
- Silverii, G.A.; Monami, M.; Gallo, M.; Ragni, A.; Prattichizzo, F.; Renzelli, V.; Ceriello, A.; Mannucci, E. Glucagon-like peptide-1 receptor agonists and risk of thyroid cancer: A systematic review and meta-analysis of randomized controlled trials. Diabetes, Obes. Metab. 2023, 26, 891–900. [Google Scholar] [CrossRef]
- Pasternak, B.; Wintzell, V.; Hviid, A.; Eliasson, B.; Gudbjörnsdottir, S.; Jonasson, C.; Hveem, K.; Svanström, H.; Melbye, M.; Ueda, P. Glucagon-like peptide 1 receptor agonist use and risk of thyroid cancer: Scandinavian cohort study. BMJ 2024, 385, e078225. [Google Scholar] [CrossRef]
- Sodhi, M.; Rezaeianzadeh, R.; Kezouh, A.; Etminan, M. Risk of Gastrointestinal Adverse Events Associated With Glucagon-Like Peptide-1 Receptor Agonists for Weight Loss. JAMA 2023, 330, 1795–1797. [Google Scholar] [CrossRef]
- Eraniyan, K.; Ganti, L.; Sundaramurthi, S.; H, S.; Kumar, N.R.; Dasarathan, S.; D, K. History and Evolution of the Tuning Fork. Cureus 2024, 11. [Google Scholar] [CrossRef] [PubMed]
- Dankner, R.; Murad, H.; Agay, N.; Olmer, L.; Freedman, L.S. Glucagon-Like Peptide-1 Receptor Agonists and Pancreatic Cancer Risk in Patients With Type 2 Diabetes. JAMA Netw. Open 2024, 7, e2350408–e2350408. [Google Scholar] [CrossRef] [PubMed]
- Dungan K, Desantis A. Glucagon-like peptide 1-based therapies for the treatment of type 2 diabetes mellitus. In: Nathan DM, Rubinow K, editors. UpToDate. Waltham, MA: Wolters Kluwer; 2023.
- Lu, J.; Liu, H.; Zhou, Q.; Wang, M.-W.; Li, Z. A potentially serious adverse effect of GLP-1 receptor agonists. Acta Pharm. Sin. B 2023, 13, 2291–2293. [Google Scholar] [CrossRef] [PubMed]
- Grennan, K.; Meneley, A.; Shuman, M.; Borg, C.; Janitz, T.; Brahmbhatt, P.; Venegas, C. #1698670 A Case of Fatal Fulminant Necrotizing Pancreatitis in a Patient with Recent Tirzepatide Initiation. Endocr. Pr. 2024, 30, S68. [Google Scholar] [CrossRef]
- Pinheiro, M.M.; de Souza, L.G.; Nunes, G.P.; Martin, I.F.; de Oliveira, Y.U.; Pinheiro, F.M.M.; Costa, L.N.; Caprio, M.; Della-Morte, D.; Infante, M. The first report of leukocytoclastic vasculitis induced by once-weekly subcutaneous semaglutide. Curr. Med Res. Opin. 2024, 40, 1525–1531. [Google Scholar] [CrossRef]
- Lorenz, M.; Lawson, F.; Owens, D.; Raccah, D.; Roy-Duval, C.; Lehmann, A.; Perfetti, R.; Blonde, L. Differential effects of glucagon-like peptide-1 receptor agonists on heart rate. Cardiovasc. Diabetol. 2017, 16, 1–10. [Google Scholar] [CrossRef]
- Nakatani, Y.; Kawabe, A.; Matsumura, M.; Aso, Y.; Yasu, T.; Banba, N.; Nakamoto, T. Effects of GLP-1 Receptor Agonists on Heart Rate and the Autonomic Nervous System Using Holter Electrocardiography and Power Spectrum Analysis of Heart Rate Variability. Diabetes Care 2015, 39, e22–e23. [Google Scholar] [CrossRef]
- Ahrén, B.; Veith, R.C.; Taborsky, G.J. Sympathetic Nerve Stimulation Versus Pancreatic Norepinephrine Infusion in the Dog: 1) Effects on Basal Release of Insulin and Glucagon*. Endocrinology 1987, 121, 323–331. [Google Scholar] [CrossRef]
- Amir, S. Central glucagon-induced hyperglycemia is mediated by combined activation of the adrenal medulla and sympathetic nerve endings. Physiol. Behav. 1986, 37, 563–566. [Google Scholar] [CrossRef]
- Routsolias, J.C.; Berg, S.E.; Paloucek, F.P. Does Glucagon Really Work for Beta Blocker Overdose? J. Med Toxicol. 2022, 19, 7–8. [Google Scholar] [CrossRef]
- Holt, M.K.; Trapp, S. The physiological role of the brain GLP-1 system in stress. Cogent Biol. 2016, 2, 1229086–1229086. [Google Scholar] [CrossRef] [PubMed]
- Llewellyn-Smith, I.; Marina, N.; Manton; Reimann, F. ; Gribble, F.; Trapp, S. Spinally projecting preproglucagon axons preferentially innervate sympathetic preganglionic neurons. Neuroscience 2015, 284, 872–887. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.-Y.; Wang, J.-X.; Chen, J.-L.; Dai, M.; Wang, Y.-M.; Chen, Q.; Li, Y.-H.; Zhu, G.-Q.; Chen, A.-D. GLP-1 in the Hypothalamic Paraventricular Nucleus Promotes Sympathetic Activation and Hypertension. J. Neurosci. 2024, 44. [Google Scholar] [CrossRef]
- Diz-Chaves, Y.; Gil-Lozano, M.; Toba, L.; Fandiño, J.; Ogando, H.; González-Matías, L.C.; Mallo, F. Stressing diabetes? The hidden links between insulinotropic peptides and the HPA axis. J. Endocrinol. 2016, 230, R77–R94. [Google Scholar] [CrossRef] [PubMed]
- Diz-Chaves, Y.; Herrera-Pérez, S.; González-Matías, L.C.; Lamas, J.A.; Mallo, F. Glucagon-Like Peptide-1 (GLP-1) in the Integration of Neural and Endocrine Responses to Stress. Nutrients 2020, 12, 3304. [Google Scholar] [CrossRef]
- Gil-Lozano, M.; Pérez-Tilve, D.; Alvarez-Crespo, M.; Martís, A.; Fernandez, A.M.; Catalina, P.A.F.; Gonzalez-Matias, L.C.; Mallo, F. GLP-1(7-36)-amide and Exendin-4 Stimulate the HPA Axis in Rodents and Humans. Endocrinology 2010, 151, 2629–2640. [Google Scholar] [CrossRef]
- Kuckuck S, van der Valk ES, Scheurink AJW, van der Voorn B, Iyer AM, Visser JA, et al. Glucocorticoids, stress and eating: The mediating role of appetite-regulating hormones. Obes Rev. 2023;24(3):e13539.
- Ang, R.; Mastitskaya, S.; Hosford, P.S.; Basalay, M.; Specterman, M.; Aziz, Q.; Li, Y.; Orini, M.; Taggart, P.; Lambiase, P.D.; et al. Modulation of Cardiac Ventricular Excitability by GLP-1 (Glucagon-Like Peptide-1). Circ. Arrhythmia Electrophysiol. 2018, 11, e006740–e006740. [Google Scholar] [CrossRef]
- Kristensen, S.L.; Rørth, R.; Jhund, P.S.; Docherty, K.F.; Sattar, N.; Preiss, D.; Køber, L.; Petrie, M.C.; McMurray, J.J.V. Cardiovascular, mortality, and kidney outcomes with GLP-1 receptor agonists in patients with type 2 diabetes: a systematic review and meta-analysis of cardiovascular outcome trials. Lancet Diabetes Endocrinol. 2019, 7, 776–785. [Google Scholar] [CrossRef]
- Sattar, N.; Lee, M.M.Y.; Kristensen, S.L.; Branch, K.R.H.; Del Prato, S.; Khurmi, N.S.; Lam, C.S.P.; Lopes, R.D.; McMurray, J.J.V.; Pratley, R.E.; et al. Cardiovascular, mortality, and kidney outcomes with GLP-1 receptor agonists in patients with type 2 diabetes: a systematic review and meta-analysis of randomised trials. Lancet Diabetes Endocrinol. 2021, 9, 653–662. [Google Scholar] [CrossRef]
- Phizackerley, D. Semaglutide reduces the absolute risk of major cardiovascular events by 1.5%. BMJ 2024, 384, q53. [Google Scholar] [CrossRef]
- Heile, M.; Wyne, K.; Billings, L.K.; Cannon, A.; Handelsman, Y.; Shannon, M. Cardiovascular Outcomes with Once-Weekly GLP-1 RAs: Clinical and Economic Implications. J. Manag. Care Spéc. Pharm. 2018, 24, S42–S52. [Google Scholar] [CrossRef] [PubMed]
- Mozaffarian, D. GLP-1 Agonists for Obesity-A New Recipe for Success? JAMA. 1: 2024;331(12), 2024. [Google Scholar] [CrossRef]
- O'Neil, P.M. Long-term maintenance of weight loss. Lancet Public Heal. 2022, 7, e806–e807. [Google Scholar] [CrossRef] [PubMed]
- O'Neil, P. The-upsides-and-downsides-of-blockbuster-weight-loss-drugs. In: Adams H, editor. Medical University of South Carolina: University of South Carolina News; 2023.
- Rebello, C.; Greenway, F.L.; Dhurandhar, N.V. Functional foods to promote weight loss and satiety. Curr. Opin. Clin. Nutr. Metab. Care 2014, 17, 596–604. [Google Scholar] [CrossRef]
- Seier S, Larsen KS, Pedersen J, Biccler J, Gudbergsen H, editors. Tapering semaglutide to the most effective dose: real-world evidence from a digital weight management programme (TAILGATE). 31st European Congress on Obesity; 2024; Venice, Italy: Karger.
Figure 1.
Neuroendocrine axis and incretin actions on multiple organs.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



