
Submitted:
01 October 2024
Posted:
01 October 2024
You are already at the latest version
Abstract
This review explores the intricate relationship between the microbiome and cancer metabolism, focusing on the Warburg effect—a shift where cancer cells rely on glycolysis for energy even in oxygen-rich environments. This metabolic reprogramming fuels tumor growth and alters the tumor microenvironment. Recent studies highlight the microbiome's influence on cancer metabolism, suggesting that microbial imbalances can either promote or hinder the Warburg effect. Microbiome alterations impact metabolic pathways, immune responses, and gene expression, which can accelerate or mitigate cancer progression. We examine how dysbiosis affects the Warburg effect and its implications for tumor growth, metastasis, and treatment resistance. Additionally, we discuss the potential of microbiome-targeted therapies, such as probiotics and fecal microbiota transplants, to modulate cancer metabolism. These interventions offer the possibility of reversing or controlling the metabolic shifts in cancer cells, enhancing the efficacy of traditional treatments like chemotherapy and immunotherapy. Despite promising developments, challenges remain in identifying key microbial species and pathways, as well as in validating microbiome-targeted therapies through large-scale clinical trials. Nonetheless, the intersection of microbiome research and cancer metabolism presents an exciting frontier for innovative therapies. This review offers a fresh perspective on cancer metabolism by integrating microbiome insights, highlighting the potential for interdisciplinary research to enhance our understanding of cancer progression and treatment strategies.
Keywords:
Microbiome
; Dysbiosis
; Cancer Metabolism
; Metabolic reprogramming
; Microbiome-targeted therapies
1. Overview of Gut Microbiome & Influence on Metabolism
The microbiome refers to the microorganisms symbiotically living in the different body locations of the host, which includes the oral cavity, respiratory tract, skin, gut, and vagina[1]. Among these, the gut microbiome, well-known for its complexity, consists of multiple organisms from different kingdoms, including bacteria, archaea, fungi, and viruses, which reside on the mucosa of the host’s gastrointestinal (GI) tract[1]. Currently, bacteria are the most studied microorganisms in the gut microbiome, consisting of phyla Firmicutes, Bacteroidetes, Proteobacteria, and Actinobacteria[2]. These microorganisms convert the substances in the host GI tract and into a variety of microbiome metabolites, such as short-chain fatty acids (SCFAs), metabolites of amino acids, vitamins, conjugated lipids, and secondary bail acids, to interact with host tissues and cells to affect host physiology[3]. The gut microbiome shifts when the host suffers from diseases, such as autoimmune disease, type 2 diabetes, brain disease, inflammatory bowel disease, liver disease, etc[4]. Thus, it is critical to study the gut microbiome’s role in the host's physiological and pathophysiological state.
One of the most important aspects that is affected by the gut microbiome is the host metabolism. The study associated the gut microbiome and metabolism, indicating that individuals who take antibiotics have reduced weight compared to those who do not[5]. Later studies using germ-free mice or antibiotic-treated mice also showed reduced body weight[5]. Fecal transplantation in germ-free using fecal material from healthy individuals alleviates weight loss, which establishes the connection of the gut microbiome that can affect host body weight. This link was further supported by the difference in the gut microbiome between obese and lean mice/people[5,6].
Later, studies take this further by investigating how the gut microbiome cross-talks with host tissue and cells to affect host energy intake and expenditure. Currently, the gut microbiome regulates energy intake majorly in two ways. First, metabolites from the gut microbiome directly or indirectly participate in the caloric extraction from the food. One example is that the SCFAs, the fermented products from indigestible fiber, are involved in the different tissue energy metabolism, allowing the host to access the inaccessible calories without the gut microbiome. Butyrate is used as the energy source for the colonocytes[7]. Propionate is the substrate for gluconeogenesis in the intestine and liver[7,8]. Astrocytes use acetate as an alternative energy source other than glucose[9]. SCFAs also modulate incretin hormones to regulate appetite. Previous publications have shown that SCFAs regulate GLP-1 and PYY secretion from the enteroendocrine cells via an FFAR2-mediated pathway, indirectly involving energy intake by controlling intestine motility and appetite[10,11]. Acetate, the most abundant SCFA, elevates parasympathetic output, leading to increased food intake[12]. Second, metabolites from the gut microbiome modulate energy expenditure through various mechanisms. Butyrate has been shown to increase energy expenditure in muscle, brown adipose tissue (BAT), and beige adipose tissue. Regarding increased BAT thermogenesis, butyrate has been shown to increase sympathetic output to the BAT and BAT UCP1 expression, the key gene for thermogenesis[13]. Butyrate enhances energy expenditure in beige adipose tissue and muscle by elevating fatty acid oxidation. These studies further reveal the gut microbiome's role in the host energy balance[13].
The gut microbiome also plays important roles in carbohydrates, amino acids, and lipid metabolism. Studies have shown that the gut microbiome regulates the insulin signaling pathway, enteroendocrine cell functions, and bile acid signaling via imidazole propionate, SCFAs, and FXR/TGR5 to maintain whole-body glucose metabolism homeostasis[5,14]. Amino acids and their derivatives produced by the gut microbiome, especially from tryptophan and its metabolites, significantly impact intestine permeability and immunity[15]. The gut microbiome also impacts lipid metabolism by further processing the lipid. One example is that one bacterium can metabolize cholesterol into a sterol, which is hardly absorbed by the intestine[16]. Also, the gut microbiome can convert the primary bile acids into secondary bile acids to regulate lipid metabolism[17]. This evidence further implies the complicated role of the gut microbiome in host metabolism. With cancer being a disease that presents with a significantly changed metabolism, the gut microbiome can serve as a therapeutic target.
2. Correlation between microbiome, population-based cancer epidemiology and epigenetics
According to World Health Organization data, in 2024, there were 19.9 million new cases of cancer and almost 10,000,000 deaths from cancer. The increase in cancer burden over the next two decades is expected to be approximately 60%, requiring further research on the correlation between microbiome, cancer, and preventative strategies. Convincing evidence has supported that diet influences and reduces the risk of different types of cancers in healthy populations. A recent meta-analysis has shown that a Mediterranean diet high in fruits, vegetables, and whole grains reduces the risk of colorectal cancer[18,19]. On the other hand, malnutrition leads to alteration in body composition, which is frequently seen in cancer patients[20,21]. Many prospective cohort studies have analyzed the benefits and harms of different foods and diets in cancer patients pre- and post-diagnosis[22,23,24,25,26,27].
Many studies investigating a periodontal pathogen called Fusobacterium nucleatum in colorectal cancer (CRC) patients have shown that their number increases as the disease progresses from an adenoma to cancer, and the higher tumor burden of F. nucleatum is correlated to worse outcomes in terms of cancer-specific survival[28,29,30,31,32]. These outcomes might be linked to the innate ability of the bacteria to activate autophagy-related pathways in the tumor environment, promoting resistance to chemotherapeutic agents like 5 Fluorouracil and Oxaliplatin[32]. Additional findings prove a unique interaction between the metabolome (i.e., polyamines) and the gut microbiota, which is significantly disrupted in CRC[33]. It is also associated with preexisting inflammation or colitis in about 2% of cases where adherent-invasive Escherichia coli strain NC101 and enterotoxigenic Bacteroides fragilis toxin are involved in cancer development[34,35]. Fusobacterium was also linked to non-colitis-associated CRC in patients[36,37]. Furthermore, a few Clostridium species and E. coli strains produce deoxycholic acid (DCA) and colibactin, respectively, which are examples of genotoxic metabolites produced by gut bacteria that contribute to cancer development[38].
Flemer et al. investigated different bacteria’s Co abundance Groups (CAG). The outcome of this study showed that increased CAGs of Prevotella and Bacteroidetes were associated with improved survival. Other bacteria, such as Faecalibacterium prausnitzii, also showed better survival outcomes[30,39]. Changes in tumor microenvironment have also been seen at phylum, family, and genus levels. Najafi et al. conducted a meta-analysis and found a significant difference in actinobacteria species between case and control groups at the phylum level. At the family level, Corynebacteriaceae and Halomonadaceae levels were significantly decreased in the tumor tissue. Finally, at the genus level, decreased levels of Lachnoanaerobaculum, Corynebacterium, and Halomonas were seen in the tumor[40]. Subsequently, studies done on H. Pylori-infected patients have shown that eradicating the infection resulted in a significant decrease in gastric cancer occurrence compared to the patients who did not receive treatment for eradication[41]. Epstein-Barr virus (EBV) infection has also been shown to increase the risk of gastric cancer by 18fold. Even though the prevalence of EBV is higher in men than women, the prevalence of gastric cancer related to EBV has been higher in women[42].
Of the many types of human papillomavirus (HPV), more than 30 infect the genital tract. The association between certain oncogenic (high-risk) strains of HPV and cervical cancer is well established[43]. HPV 16 and 18 are the main causative organisms for cervical cancer, which is one of the leading causes of death for women worldwide. The HPV early region (E) oncoproteins are linked to etiopathogenesis and aid in cancer development. Li et al., in their meta-analysis, found that HPV infection was linked to a higher incidence of esophageal SCC[44]. Since the prevalence of high-risk HPV infection is higher in regions with an elevated risk for esophageal cancer compared to low-moderate risk regions, it is possible that HPV infection plays a role in esophageal carcinogenesis. High-risk HPV subtypes can be particularly taken into consideration among risk factors; however, more research would be needed to establish the link between HPV and esophageal cancer[45].
3. Impact of Microbiome Dysbiosis on cancer metabolism
Host gut microbiome alterations influence cancer progression and metabolism through various multifaceted mechanisms. These mechanisms can foster a tumor microenvironment, enhance or inhibit treatment efficacy, create unique tumor-specific microbial profiles, and contain carcinogenic-metabolite-producing bacteria. Changes in the diversity and composition of gut microbiota are associated with cancer. Certain microbial species or communities may either promote cancer or protect against it. For instance, high microbial diversity and alterations in the gut microbiota are linked to CRC patients. They exacerbate cancer development through chronic gut inflammation from dysbiosis, or microbial imbalance, and further contribute to the tumorigenic microenvironment. The effects of gut microbial changes can produce carcinogenic metabolites as a byproduct, which further contribute to cancer development and risk. Imbalances of these metabolites include alterations in genotoxic metabolites such as short-chain fatty acids (SCFAs) and bile acids, which directly damage DNA, contributing to cancer progression[46]. Microbial pathogens also influence carcinogenesis by inducing or exacerbating inflammatory processes. For example, Lactobacillus iners, a bacterium commonly found in tumors, has been associated with a more hostile tumor environment through changes in metabolism and an increased resistance to chemoradiation therapy. The bacterium produces high lactate levels in a tumor microenvironment that causes metabolic changes within tumor cells[47]. The changes involve altering energy production pathways and enhancing their ability to survive and resist treatment. Further research is needed to investigate the mechanistic alterations within these pathways.
Furthermore, researchers have looked at the liver to provide potential therapeutic targets against host-microbiome tumorigenesis as it is largely involved in bile acid metabolism. Feng et al. highlighted the bidirectional relationship between the liver and the gut microbiome in response to changes in bile acid production and metabolism, contributing to the progression of cancer cachexia, a complex syndrome in cancer patients, including weight loss and malnutrition[48]. Galeano et al. used imaging and molecular techniques to analyze various cancer tumor samples to create a map of the spatial distribution of microbial species to understand the tumor microenvironment. Different microbial species were localized in distinct regions of the tumor. The presence of certain microorganisms influenced growth patterns, immune cell interactions, and resistance to treatment[49]. High microbial diversity is associated with greater cellular and spatial heterogeneity that causes changes in tumor cell characteristics[49]. The microbial communities within tumors impact how a tumor develops and responds to treatment, thus serving as a potential target against microbiota-resistant treatment. Additionally, Yachida et al. examined gut microbiota changes at different stages of CRC using metagenomic and metabolomic analyses to understand the relationship between microbial communities and cancer progression. Their results showed alterations of metabolic activity by distinct microbial communities in the early and late stages of CRC, suggesting cancer progression may result from stage-specific microbial metabolic products. Identifying microbiota profiles at different stages could be used as biomarkers and targeted therapeutics for CRC[50].
4. Factors Influencing Cancer Progression
Genetic and environmental factors increase the risk of cancer progression due to microbial changes. The microbiome can influence the host’s genetic and epigenetic landscape, creating microbial metabolites and interactions that can lead to genetic mutations or epigenetic modifications, further increasing cancer susceptibility. High-fat and cholesterol diets contribute to the development of fatty liver diseases. The gut microbiota plays an important role in the progression of fatty liver disease to hepatocellular carcinoma (HCC). Zhang et al. showed alterations in the gut microbiota due to dietary cholesterol influencing metabolite production during metabolism and further exacerbating cancer development and inflammation[50].
Concerning immune system modulators, specific microbial profiles regulate the immune responses by interacting with the T cells and macrophages of the host immune system, thereby influencing tumor development. These. Interestingly, researchers have also found that microbial-immune modulation can inhibit or enhance the efficacy of immunotherapy. For example, some bacteria might boost the efficacy of checkpoint inhibitors by promoting a more robust anti-tumor immune response. At the same time, some weaken the efficacy and metabolism of chemotherapeutic agents and targeted therapies by microbial enzymes, which metabolize these drugs into more or less active forms. This is how host metabolites communicate with host cellular pathways involved in drug metabolism and cancer development, such as signaling pathways that cause tumor growth. Microbial alterations can disrupt the integrity of the gut barrier, leading to greater systemic inflammation and a reduction in treatment responsiveness and the body’s overall immune function [50].
Host-microbiome interactions play a large role in cancer progression, especially lung and intestinal cancer. Dong et al. investigated the diverse microbial communities in the lung, gut, and oral cavity of lung cancer patients to examine new strategies for cancer prevention. Alterations in the microbiota can influence immune responses within the lung, thus advancing tumor development and resistance to immunotherapy. Personalized medicine approaches to enhance treatment efficacy are important against harmful microbial communities harboring the tumor microenvironment[51]. These mechanisms illustrate the complex interplay between gut microbiome and cancer, illustrating how microbial composition, metabolism, immune modulation, and inflammation exacerbate cancer progression. Identifying specific microbial profiles and restoring a healthy microbiome balance can help prevent or treat certain cancers.
5. Microbiota dysbiosis in Cancer & Mechanism of Microbial Oncogenesis
Microbiota dysbiosis is defined as an imbalance in the microbial communities within the body, which mainly includes bacteria, viruses, fungi, and other microorganisms[52]. The gut microbiome is crucial in regulating the immune system. Dysbiosis has gained widespread recognition as a significant factor in the development of various types of cancer by causing chronic inflammation and immune dysregulation[53]. Some of the microbes have a direct correlation with specific cancers. Helicobacter pylori, Human Papillomavirus (HPV), and Fusobacterium nucleatum are associated with gastric adenocarcinoma, cervical cancer, and colorectal cancer, respectively[54,55,56]. In addition, microbiota-related metabolites have also been associated with cancer development. The production of short-chain fatty acids (SCFA) is protective due to their anti-inflammatory and anti-carcinogenic properties; however, with dysbiosis, there is a decrease in the level, which subsequently leads to an increase in harmful metabolites such as secondary bile acids[57]. These secondary bile acids cause DNA damage, promote inflammation, and generate reactive oxygen species, thereby contributing to carcinogenesis[58]. In addition, they also activate various signaling pathways such as nuclear factor kappa B (NF-KB) and WNT, both of which are involved in cellular proliferation and survival, further promoting tumorigenesis[59]. This has been implicated in the development of liver and colorectal cancers.
5.1. Contact-dependent mechanism
The contact-dependent mechanism of microbial oncogenesis involving the gut microbiome is an interplay between host factors and the gut microbiome. It mainly includes interbacterial signaling, cellular adhesion, and invasion. The initial step of gut colonization occurs because of the interaction between the pathogen and the host components, such as cell surfaces and extracellular matrices [60]. This is followed by using proteins that deliver toxins to neighboring bacteria, leading to growth inhibition and changes in genetic expression [61]. Recent studies have discussed the role of bacterial protein and molecular trafficking in modifying the gut microbiome, ultimately influencing cancer development. Garcia-Bayona et al. stated the contact-dependent mechanism for some bacteria, like Caulobacter crescentus [62]. In addition, fungal infections caused by Candida albicans also involve kinase activation through physical contact, which leads to invasive growth and biofilm development [63]. Liang et al. 2014 described the development of colorectal cancer in a mouse model with colitis, thereby highlighting the role of inflammation in gut microbial composition [64].
5.2. Contact-independent mechanism
Recent studies have highlighted contact-independent mechanisms of microbial carcinogenesis that do not require direct physical interaction between microbes and host cells. These mechanisms include modulation of inflammation, DNA damage, chronic inflammation, and harmful bacterial metabolites. Additionally, recent studies have mentioned the influence of gut microbiomes on gut epithelia with a prior predisposition of mutant p53 activity [65]. An important consideration for colonization resistance in gut microbiota is the interplay between the contact-dependent and contact-independent mechanisms facilitated by molecular trafficking [66]. This has been implicated in promoting tumorigeneses in various cancers, including hepatocellular carcinoma and colorectal cancer [67].
5.3. Immunological mechanisms
Immunological factors also play a key role in microbial carcinogenesis in the gut microbiome. Recent studies have highlighted the impact of gut microbiome on the host immune regulation, thereby influencing disease outcomes and leading to cancer development [68]. An important factor that has led to cancer development is the disruption in the immune surveillance mechanism. Understanding the complex interaction between microbial species and innate and acquired immunity is important. Tekle et al. talk about the various immune mechanisms involved in carcinogenesis[69]. Dendritic cells and molecular processes like mitophagy and apoptosis have been postulated to play a key role in microbial carcinogenesis[70]. After interacting with particular bacteria, the dendritic cells from the gut-associated lymphoid tissue and spleen activate the type 1 interferon (IFN) mediated signaling, leading to tumorigenesis[69]. Regarding molecular processes like mitophagy and apoptosis, Roberti et al. found that gut microbiota can enhance these processes, contributing to great carcinogenesis[71].
6. Biomarkers – Current microbiota markers in clinical practice
The growing clinical use of microbiota markers is important due to their ability to help diagnose and treat conditions. Microbiota markers have become a frontier in research when it comes to carcinoma. They can be used to diagnose and treat a wide range of diseases, potentially improving public health outcomes in the future. Certain bacteria seem to interact with the oral epithelial cells to cause cancer, including P. gingivalis and F. nucleatum, as they stimulate cell growth [72]. The role of the microbiome in cancer remains unclear overall; however, we do see an elevation or reduction in the level of bacteria when carcinoma is present[72]. A known mechanism that makes the likelihood of cancer higher is the administration of antibiotics. When they are administered, antibiotics replace the previous microbiota, rearranging the gut and acting as a potential risk factor for the development of certain forms of cancer. In liver cancer, Akkermansia mucinphila, and Facalibacterium are elevated. In lymphoma, for example, Ruminococcus is elevated. These bacteria are known as responders-enriched bacteria. Probiotics, too, are involved in the development of cancer. In the modern day, selective antibiotics are used to negate the risk of the destruction of beneficial bacteria in the gut [73].
On the other hand, some bacteria have an overall benefit when dealing with cancer. These include Bifidobacteria longum, Lactobacillus acidophilus, and Saccharomyces boulardii. They contain SCFA, Vitamins, antioxidants, and glucosinolates. These limit pathogen growth, reduce DNA damage and inflammation, and hinder tumor growth. On the flip side, in our diet, we find that there may be E. coli, Salmonella enterica, and Fusobacterium nucleatum, which, on the other hand, lead to increased tumor growth, DNA alkylation, DNA damage, and inflammation [74]. In addition to the specific bacterial correlation, a link exists between the microbiome's diversity and cancer development. People migrating to the West have witnessed a noticeable decline in gut microbiome diversity. Furthermore, the overexpression of certain bacteria is a key indicator to understand the cancer stage. In some other cancers like adrenocortical carcinoma, studies have shown a correlation between Bacteroides, Streptomyces, and the chance of survival. The microbiome plays a crucial role in not only indicating cancer and its potential stage but also in its role of survival and treatment [72].
7. Treatment:
Multiple treatment modalities have been described in the literature with a prime focus on the gut microbiota—these range from pharmacological agents like antibiotics to dietary interventions like probiotics and prebiotics. Certain biotherapeutic modalities, such as fecal microbiota transplantation, have also gained widespread attention and need special mention.
7.1. Antibiotics
Bacterial association with systemic lymphoid tissue has sparked research on microbial alteration as a potent immunotherapeutic tactic. Early research shows that intratumoral microbiota is prevalent and immunologically active in most patients, underscoring the need to consider the microbial environment and its interactions[75,76]. For instance, in pancreatic cancer, changes in gut microbiota may impact the intratumoral microbiome's makeup through pancreatic duct communication [77]. On the other hand, antibiotics have a variety of consequences. When used to clear the pancreatic intratumoral microbiome, they paradoxically increase the efficacy of immunotherapy by upregulating PD-1 expression, even though they may compromise the immune response by disturbing the gut microbiome. Antimicrobial therapy for cancer involves using medications such as antibiotics, antivirals, and vaccines against hepatitis B and human papillomavirus. The focus for such treatment regimens is microbiological carcinogens, like the Hepatitis C virus and H. pylori, which can cause stomach lymphomas [78,79]. According to studies done on pancreatic, lung, and colon malignancies, eliminating the intratumoral microbiota may alter the tumor microenvironment (TME) from immunogenic to tolerogenic by reducing inflammation that impedes tumor growth and slows down cellular division [80,81,82]. Clinical evidence indicates systemic antibiotics may lower survival rates and undermine immune checkpoint inhibitors' efficacy. Preclinical evidence suggests that antibiotics or gut bacterial translocation may trigger leukemic development in genetically vulnerable hosts with hematologic malignancies [83].
7.2. Nutrition, postbiotics, and probiotics
A few practical strategies for modifying the microbiota are using postbiotics, prebiotics, and modifying diet. Despite the prevalence of epidemiological correlations, the exact mechanisms linking certain foods to cancer remain unclear due to challenges in collecting dietary data. Prebiotics with promising results in preclinical models, like resistant starch, inulin, and mucin, are currently being studied in humans to see their impact. In melanoma and colon cancer, they have enhanced the antitumor immunity and the therapeutic response (e.g., NCT03870607, NCT03950635). Chemicals derived from microbes may have advantages in composition and repeatability despite postbiotics receiving less scientific attention. Because of their potential modes of action and associated safety concerns, probiotics, described as live microorganisms offering health advantages when taken in moderation, have garnered a lot of attention in the setting of CRC. Probiotics were first thought to alter gut flora [84,85]. However, in addition to changing the microbiota, their role now includes altering it to suit the host's physiology and metabolism. They can produce lactic and acetic acid, or bacteriocins, which limit bacterial development and lower the pH. In addition, they also interact directly with lethal bacteria to restrict their growth [86]. Probiotics have an immunomodulatory effect in the gut and can either improve immunosurveillance or alter colonic inflammation, depending on the specific activity of each strain. They have been shown by Klaenhammer et al. to alter macrophage subsets and decrease colonic inflammation[87]. By binding with toll-like receptors (TLRs), specific probiotic strains of Bifidobacterium infantis and Bifidobacterium breve stimulate intestinal dendritic cells (DCs), eliciting the expression of Foxp3+ regulatory T cells (Treg), type 1 regulatory T cells (Tr1) and stimulating the release of IL-10. They also trigger the metabolism of retinoid acid. Other probiotic bacteria, such as Lactobacillus acidophilus and Lactobacillus rhamnosus, can block STAT3 and NF-κB signaling to limit Th17 cell production, release IL23 and IL17, and promote a shift in macrophage phenotype from pro-inflammatory M1 to immunosuppressive M2. Probiotics, such as Lactobacillus rhamnosus, Lactobacillus plantarum, and Escherichia coli, improve the integrity of the gut barrier by upregulating the synthesis of mucin and the expression of tight junction proteins [88,89,90,91,92]. Several studies have shown that probiotics reduce the colonization of pathogens such as Staphylococcus aureus and Clostridium difficile, which supports their utility in preventing intestinal infections [93,94]. The usefulness of probiotics in treating and preventing colorectal cancer has been the subject of numerous studies. A meta-analysis of data from multiple trials revealed a significant association between probiotic consumption and a decreased risk of colorectal cancer (CRC) [86].
7.3. Transplantation of Fecal Microbiota and Identified Microbial Consortiums
Fecal microbiota transplantation (FMT), specified microbial consortia, and commercial probiotics are three ways to modulate the gut microbiota in cancer patients. FMT, a recently developed biotherapeutic modality, restores microbial equilibrium in patients with microbial dysbiosis by introducing a disease-free, healthy microbial population from healthy donors. It appears to have some potential in treating colitis linked to immunotherapy and is useful in treating Clostridium difficile-induced colitis. However, its stability and long-term effectiveness need to be studied extensively [95,96]. According to clinical trials, FMT from donors who respond to immunotherapy may improve anticancer immune responses (e.g., NCT03353402) [97]. Microbial consortia, ranging from complex consortia to monoclonal bacterial strains, are also being evaluated in trials. Several microbial metabolites, including cadaverine, indoles, short-chain fatty acids (SCFAs), and nisin, have been shown to have anticancer properties in breast cancer cases [98].
Probiotics may increase the risk of bacterial translocation and systemic invasion in patients with weakened immune systems. There have also been reports of CMV infection following FMT, norovirus gastroenteritis, and Escherichia coli bacteremia [99,100]. Unknown components transferred during FMT provide a risk of changing the recipient's microbiota and perhaps causing chronic illnesses. Research has demonstrated that pro-atherogenic trimethylamine-N-oxide (TMAO) synthesis and metabolic phenotypes associated with obesity are transferred from donors to recipients [86,101]. Due to the paucity of clinical data, long-term follow-ups are required to establish causality. Despite these concerns, FMT is better than transient therapies like probiotics and prebiotics because it has advantages over other modulation tactics, including higher microbial diversity, less disruption of gut ecology than antibiotics, and the possibility for long-term engraftment[86].
7.4. Utilizing Exogenous Microbiota in Cancer Therapy
The creation of exogenous bacteria and viruses for cancer treatment has advanced significantly. Talimogene laherparepvec (T-VEC) for metastatic melanoma and Mycobacterium bovis (BCG vaccination) for non-muscle invasive bladder cancer are two notable FDA-approved treatments. Oncolytic viruses are covered in depth elsewhere, but it is essential to understand that synthetic biological methods that reduce systemic toxicities and boost local antitumor immunity and bacterial cancer treatments (BCTs) are receiving widespread attention. Clinical trials (e.g., NCT04167137) are investigating the potential of BCT drugs despite regulatory obstacles; however, commercial feasibility is still a barrier[102]. Because engineered microorganisms naturally target tumor tissues, they hold considerable promise as cancer therapy. When given intratumorally or intravenously, this tumor tropism causes an approximately 10,000-fold concentration of microorganisms in tumors compared to the liver, spleen, and lungs [103]. Using this, scientists have created a unique therapeutic strategy in which these tumor-homing bacteria deliver deadly chemicals[57]. Using several delivery methods, genetically engineered strains of Salmonella, Bifidobacterium, Listeria, Clostridium, Shigella, Lactococcus, Vibrio, and Escherichia have shown anticancer effects in preclinical mice[103]. While some treatment modalities use bacteria as "intratumoral bioreactors" to continually generate and release therapeutic compounds extracellularly during colonization, others involve the intracellular delivery of medications via phagocytic absorption. One noteworthy technique is engineered bacterial lysis, which minimizes colony size and lowers systemic toxicity by releasing therapeutic proteins only when the bacterial population reaches a particular density [104]. To release chemokines, hemolysins, or pro-apoptotic proteins cyclically within the tumor microenvironment (TME), for example, Din et al. modified non-pathogenic E. coli and Salmonella to lyse at a threshold population density [104].
7.5. Intratumoral Microbiota's Impact in Boosting Antitumor Immunity
Through several processes, the intratumoral microbiota is essential for boosting antitumor immunity and the effectiveness of immunotherapy. These processes demonstrate how important the intratumoral microbiota is for boosting anticancer immunity and enhancing immunotherapy's effectiveness [105].
- a)
- Intratumoral Bifidobacterium can activate dendritic cells (DCs) through the STING signaling pathway. Furthermore, A. muciniphila can create STING agonists, which cause intratumoral monocytes to secrete IFN-I. This procedure increases communication between natural killer (NK) cells and DCs further encouraging macrophage conversion [106].
- b)
- T and NK Cell Activation: By encouraging the recruitment and activation of CD8+ T cells, several intratumoral microbiota, including Saccharopolyspora, Lachnoclostridium, EBV, and HBV, improve antitumor immunity. Patients survive longer because of this activation, mediated by chemokines produced from the intratumoral microbiome such as CXCL9, CXCL10, and CCL5[106]. Trimethylamine N-oxide (TMAO) generated from Clostridiales can cause PERK-mediated endoplasmic reticulum (ER) stress, which can result in more CD8+ T cell-mediated antitumor immunity and tumor cell pyroptosis. Furthermore, NK cell activity is improved, and tumor regression is induced by increased levels of Bifidobacterium in intratumoral areas, which is caused by an elevated by-product in the diet called Hippurate[106,107].
- c)
- Production of Tertiary Lymphoid Structures (TLS): The development of TLS is caused by intratumoral H. hepaticus, which stimulates anticancer immune responses dependent on Tfh- and B-cells [108].
- d)
7.6. Human Microbiota's Therapeutic Effect in Skin Cancer
To control the tumor microenvironment and affect the growth, spread, and response to treatment, the human microbiome is essential. Several promising treatment approaches are developing, most notably the regulation of skin and gut dysbiosis by prebiotics and probiotics [109]. In this context, prebiotics—which are substrates that host bacteria specifically use for advantageous effects—and probiotics—which are live microorganisms good for the skin or intestinal flora—offer great promise [109]. According to clinical data, oral probiotics are effective in treating various gastrointestinal (GI) conditions, including GI malignancies [110]. These probiotics reduce symptoms related to GI malignancies and have antiproliferative or pro-apoptotic actions[88,110,111]. For instance, oral administration of Lactobacillus rhamnosus GG dramatically decreased the incidence of GI cancer in high-risk people, according to a clinical trial conducted by Smith et al. Combining Lactobacillus and Bifidobacterium probiotics relieved GI cancer-related symptoms and shrunk tumor size, according to another study by Johnson et al. Furthermore, repairing the microbiota in the stomach can indirectly help dermatological problems like psoriasis, acne, and atopic dermatitis[108]. Clinical trials have demonstrated oral probiotics to considerably lessen the severity of atopic dermatitis symptoms in young individuals [112]. Additionally, research by Lee et al. showed that supplementing with probiotics significantly improved adult patients' acne severity levels.
Skin and/or gut microbiota disturbances are associated with the development of skin cancer, particularly when UV radiation is present. Recent research has demonstrated the ability of oral probiotics to counteract UV-induced immunosuppression and slow the growth of skin tumors [110]. For example, Wang et al.'s randomized controlled experiment discovered that probiotics taken orally significantly decreased the incidence of UV-induced skin tumors in mice. According to another clinical trial by Kim et al., probiotic supplementation boosted human subjects' immune responses against UV-induced skin damage. Topical probiotics have also shown promise in treating various skin conditions by directly affecting the skin microbiome. For example, topical application of specific probiotics improved conditions like atopic dermatitis and acne and decreased pathogen colonization on the skin [107]. Although clinical studies on the direct effects of topical probiotics on skin cancer are lacking, their potential mechanisms include modulating the skin and intratumoral microenvironment, enhancing immune surveillance, and suppressing chronic inflammation [107].
In conclusion, the therapeutic impact of human microbiota, particularly through probiotics, shows promising potential in managing skin cancer and related dermatological conditions. Further clinical trials are warranted to validate these findings and explore the optimal probiotic formulations and dosages for maximal therapeutic benefits.
8. Environmental risk factors influencing microorganisms-cancer interactions
Environmental factors highly influence the interplay between gut microbiome and cancer metabolism. They play a key role in shaping the microbiome's composition and function, thereby affecting the carcinogenesis process [111]. An important environmental risk factor is diet which has a direct effect on the diversity of gut microbiome [112]. Diets rich in high fat and low fiber have been linked to microbial dysbiosis, leading to inflammation and carcinogenesis. Dietary fiber gets fermented by gut bacteria into butyrate, which reduces the risk of colorectal cancer [113]. This alteration impacts the metabolic pathways involved in the Warburg effect, where cancer cells use glycolysis to generate energy over oxidative phosphorylation, even in the presence of oxygen [112,114]. Diet-induced alteration in the gut microbiome modulates tumor etiology, progression, and response to cancer treatment [115].
Another crucial environmental factor to note is the presence of environmental pollutants and toxins. These include heavy metals and persistent organic pollutants that act through complex mechanisms to cause tumorigenesis [116,117]. An important pollutant to consider is polycyclic aromatic hydrocarbons (PAH), which promote a pro-inflammatory environment [118]. Consequently, it is essential to understand the crucial role of aryl hydrocarbon receptor (AhR) in metabolizing PAHs into carcinogenic metabolites [119]. These metabolites cause carcinogenesis through mechanisms that are still under investigation. Subsequently, an important consideration is a synergistic interaction between different pollutants, even at low concentrations, that cause carcinogenesis by producing reactive oxygen species and activating AhR [120]. Clause et al. 2016 highlighted the relationship between gut microbiome and environmental pollutants which have been found to contribute to their toxicity [121].
Factors such as physical activity and stress also play a crucial role in the link between gut microbiome and carcinogenesis [122]. Regular physical exercise is known to have a positive impact on the gut microbiota, reducing the level of inflammation and risk of cancer [123,124]. Studies have found that chronic psychological stress leads to alteration in the microbiome, increases intestinal permeability, promotes systemic inflammation, and metabolic derangements leading to cancer progression [125,126]. Furthermore, these changes alter the signaling molecules that drive the Warburg effect, leading to carcinogenesis [127]. Understanding these environmental risk factors that modulate the gut microbiome is quintessential. Addressing these factors through lifestyle modifications may possibly counteract the Warburg effect.
9. Limitations of this review and future directions:
Much research has been done on the role of gut microbiome in cancer metabolism and the Warburg effect. However, some limitations need special mention. Many microorganisms have been identified that influence cancer metabolism, but the scientific community has still not been able to pinpoint specific bacteria or microbial metabolites that impact cancer metabolism[128,129,130]. From an individual standpoint, there is significant variability in diet, genetics, lifestyle, and environment, complicating the ability to generalize findings and postulate a universal understanding of all types of cancers[131,132]. We still have not reached a stage where we can decipher the combination of factors that could lead to cancer development in a specific individual. Several studies have explained the correlation between gut microbiome alterations and cancer development. Still, we cannot identify molecular and cellular mechanisms, particularly regarding cancer treatment[131]. This necessitates the need for more in-depth studies at the molecular level to elucidate the exact mechanism. Most of the studies have been done on animal models, particularly mice, which may not fully replicate human physiology, and their relevance to humans is questionable. This is due to the inherent difference between human and murine gut microbiome. The translation from animal studies to human clinical studies demands careful validation to ensure strict adherence to ethical standards[133].
It is important to realize the importance of future research using advanced metagenomic techniques to understand the correlation between gut microbiome and cancer development[134,135]. This will aid in identifying specific microbes and their metabolites that are involved in causing cancer. In addition, it is important to understand the temporal relationship between gut microbiota and cancer development, for which large-scale, longitudinal cohort studies would be required[136]. In vitro and in vivo experiments focusing on cancer cell metabolic pathways must be conducted[137]to understand the mechanism better. We need to reach a stage where we can develop a personalized microbiome-based treatment regimen tailored to the individual’s microbiome and cancer type, possibly by using custom-designed probiotics or specific microbiota transplantation[50,138]. Further progress in cancer treatment also demands well-designed clinical trials to evaluate the efficacy and safety of microbiome-targeted cancer treatment. Such a holistic approach will help the scientific community progress in combating cancer and improving overall patient outcomes.
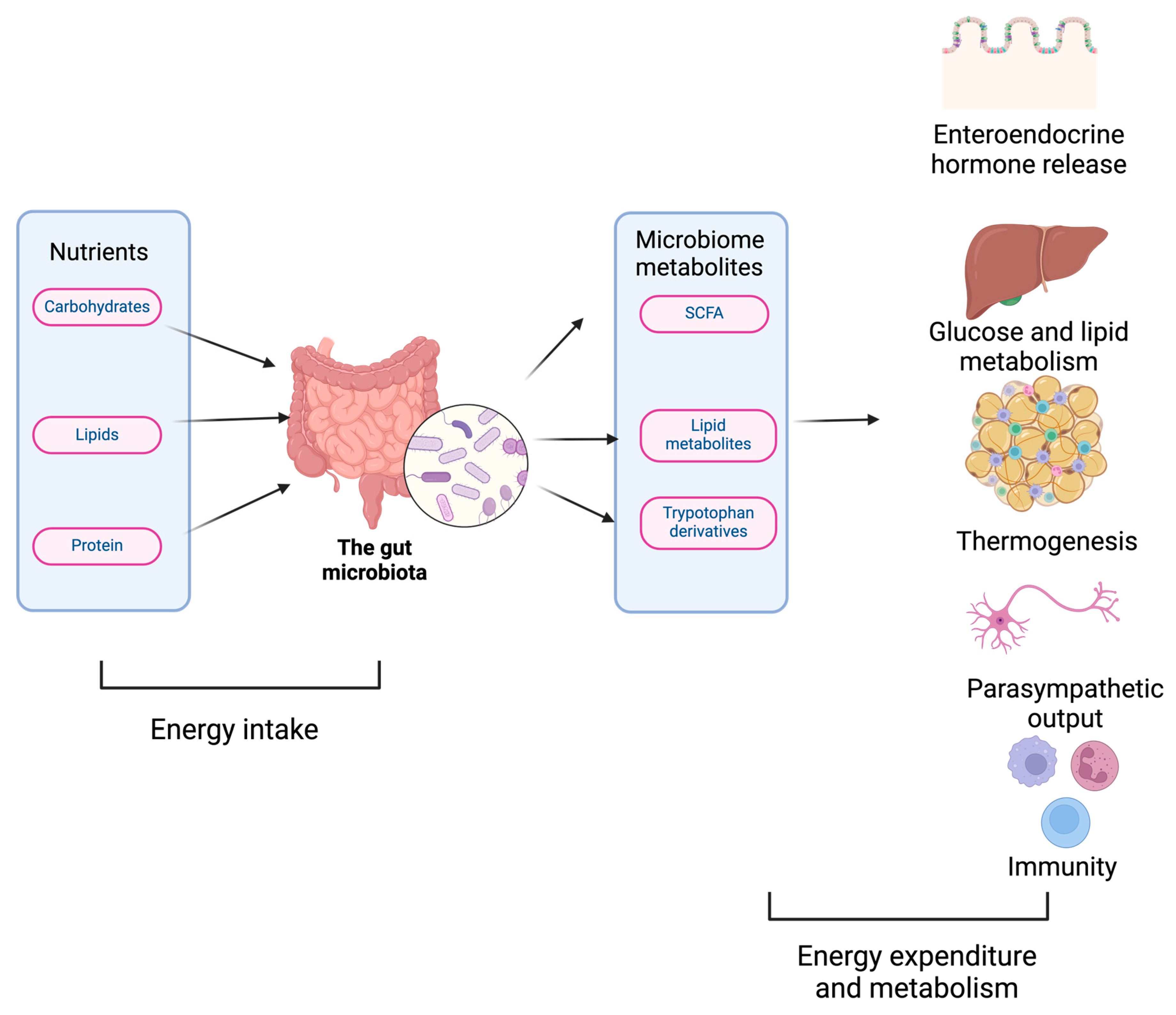
Figure 1.
The gut microbiome affects body physiology: The gut microbiome regulates physiology by modulating energy intake (nutrient digestion), energy expenditure (thermogenesis), glucose, amino acid, and lipid metabolism.
Figure 1.
The gut microbiome affects body physiology: The gut microbiome regulates physiology by modulating energy intake (nutrient digestion), energy expenditure (thermogenesis), glucose, amino acid, and lipid metabolism.
References
- Hou, K.; Wu, Z.X.; Chen, X.Y.; Wang, J.Q.; Zhang, D.; Xiao, C.; Zhu, D.; Koya, J.B.; Wei, L.; Li, J.; et al. Microbiota in health and diseases. Signal Transduct Target Ther 2022, 7, 135. [Google Scholar] [CrossRef] [PubMed]
- Fujisaka, S.; Watanabe, Y.; Tobe, K. The gut microbiome: a core regulator of metabolism. J Endocrinol 2023, 256. [Google Scholar] [CrossRef] [PubMed]
- Montenegro, J.; Armet, A.M.; Willing, B.P.; Deehan, E.C.; Fassini, P.G.; Mota, J.F.; Walter, J.; Prado, C.M. Exploring the Influence of Gut Microbiome on Energy Metabolism in Humans. Adv Nutr 2023, 14, 840–857. [Google Scholar] [CrossRef] [PubMed]
- Ruan, W.; Engevik, M.A.; Spinler, J.K.; Versalovic, J. Healthy Human Gastrointestinal Microbiome: Composition and Function After a Decade of Exploration. Dig Dis Sci 2020, 65, 695–705. [Google Scholar] [CrossRef]
- Cox, T.O.; Lundgren, P.; Nath, K.; Thaiss, C.A. Metabolic control by the microbiome. Genome Med 2022, 14, 80. [Google Scholar] [CrossRef]
- Backhed, F.; Ding, H.; Wang, T.; Hooper, L.V.; Koh, G.Y.; Nagy, A.; Semenkovich, C.F.; Gordon, J.I. The gut microbiota as an environmental factor that regulates fat storage. Proc Natl Acad Sci U S A 2004, 101, 15718–15723. [Google Scholar] [CrossRef]
- Zhu, L.B.; Zhang, Y.C.; Huang, H.H.; Lin, J. Prospects for clinical applications of butyrate-producing bacteria. World J Clin Pediatr 2021, 10, 84–92. [Google Scholar] [CrossRef]
- De Vadder, F.; Kovatcheva-Datchary, P.; Goncalves, D.; Vinera, J.; Zitoun, C.; Duchampt, A.; Backhed, F.; Mithieux, G. Microbiota-generated metabolites promote metabolic benefits via gut-brain neural circuits. Cell 2014, 156, 84–96. [Google Scholar] [CrossRef]
- Nogal, A.; Valdes, A.M.; Menni, C. The role of short-chain fatty acids in the interplay between gut microbiota and diet in cardio-metabolic health. Gut Microbes 2021, 13, 1–24. [Google Scholar] [CrossRef]
- Caengprasath, N.; Gonzalez-Abuin, N.; Shchepinova, M.; Ma, Y.; Inoue, A.; Tate, E.W.; Frost, G.; Hanyaloglu, A.C. Internalization-Dependent Free Fatty Acid Receptor 2 Signaling Is Essential for Propionate-Induced Anorectic Gut Hormone Release. iScience 2020, 23, 101449. [Google Scholar] [CrossRef]
- Forbes, S.; Stafford, S.; Coope, G.; Heffron, H.; Real, K.; Newman, R.; Davenport, R.; Barnes, M.; Grosse, J.; Cox, H. Selective FFA2 Agonism Appears to Act via Intestinal PYY to Reduce Transit and Food Intake but Does Not Improve Glucose Tolerance in Mouse Models. Diabetes 2015, 64, 3763–3771. [Google Scholar] [CrossRef] [PubMed]
- Perry, R.J.; Peng, L.; Barry, N.A.; Cline, G.W.; Zhang, D.; Cardone, R.L.; Petersen, K.F.; Kibbey, R.G.; Goodman, A.L.; Shulman, G.I. Acetate mediates a microbiome-brain-beta-cell axis to promote metabolic syndrome. Nature 2016, 534, 213–217. [Google Scholar] [CrossRef] [PubMed]
- Peng, K.; Dong, W.; Luo, T.; Tang, H.; Zhu, W.; Huang, Y.; Yang, X. Butyrate and obesity: Current research status and future prospect. Front Endocrinol (Lausanne) 2023, 14, 1098881. [Google Scholar] [CrossRef]
- Ma, H.; Patti, M.E. Bile acids, obesity, and the metabolic syndrome. Best Pract Res Clin Gastroenterol 2014, 28, 573–583. [Google Scholar] [CrossRef] [PubMed]
- Roager, H.M.; Licht, T.R. Microbial tryptophan catabolites in health and disease. Nat Commun 2018, 9, 3294. [Google Scholar] [CrossRef] [PubMed]
- Kenny, D.J.; Plichta, D.R.; Shungin, D.; Koppel, N.; Hall, A.B.; Fu, B.; Vasan, R.S.; Shaw, S.Y.; Vlamakis, H.; Balskus, E.P.; et al. Cholesterol Metabolism by Uncultured Human Gut Bacteria Influences Host Cholesterol Level. Cell Host Microbe 2020, 28, 245–257.e246. [Google Scholar] [CrossRef]
- Jones, B.V.; Begley, M.; Hill, C.; Gahan, C.G.; Marchesi, J.R. Functional and comparative metagenomic analysis of bile salt hydrolase activity in the human gut microbiome. Proc Natl Acad Sci U S A 2008, 105, 13580–13585. [Google Scholar] [CrossRef]
- Schwingshackl, L.; Schwedhelm, C.; Galbete, C.; Hoffmann, G. Adherence to Mediterranean Diet and Risk of Cancer: An Updated Systematic Review and Meta-Analysis. Nutrients 2017, 9. [Google Scholar] [CrossRef]
- Schwingshackl, L.; Hoffmann, G. Adherence to Mediterranean diet and risk of cancer: an updated systematic review and meta-analysis of observational studies. Cancer Med 2015, 4, 1933–1947. [Google Scholar] [CrossRef]
- Cederholm, T.; Barazzoni, R.; Austin, P.; Ballmer, P.; Biolo, G.; Bischoff, S.C.; Compher, C.; Correia, I.; Higashiguchi, T.; Holst, M.; et al. ESPEN guidelines on definitions and terminology of clinical nutrition. Clin Nutr 2017, 36, 49–64. [Google Scholar] [CrossRef]
- Muscaritoli, M.; Lucia, S.; Farcomeni, A.; Lorusso, V.; Saracino, V.; Barone, C.; Plastino, F.; Gori, S.; Magarotto, R.; Carteni, G.; et al. Prevalence of malnutrition in patients at first medical oncology visit: the PreMiO study. Oncotarget 2017, 8, 79884–79896. [Google Scholar] [CrossRef] [PubMed]
- Holmes, M.D.; Wang, J.; Hankinson, S.E.; Tamimi, R.M.; Chen, W.Y. Protein Intake and Breast Cancer Survival in the Nurses' Health Study. J Clin Oncol 2017, 35, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Meyerhardt, J.A.; Niedzwiecki, D.; Hollis, D.; Saltz, L.B.; Hu, F.B.; Mayer, R.J.; Nelson, H.; Whittom, R.; Hantel, A.; Thomas, J.; et al. Association of dietary patterns with cancer recurrence and survival in patients with stage III colon cancer. JAMA 2007, 298, 754–764. [Google Scholar] [CrossRef] [PubMed]
- Fadelu, T.; Zhang, S.; Niedzwiecki, D.; Ye, X.; Saltz, L.B.; Mayer, R.J.; Mowat, R.B.; Whittom, R.; Hantel, A.; Benson, A.B.; et al. Nut Consumption and Survival in Patients With Stage III Colon Cancer: Results From CALGB 89803 (Alliance). J Clin Oncol 2018, 36, 1112–1120. [Google Scholar] [CrossRef]
- Richman, E.L.; Stampfer, M.J.; Paciorek, A.; Broering, J.M.; Carroll, P.R.; Chan, J.M. Intakes of meat, fish, poultry, and eggs and risk of prostate cancer progression. Am J Clin Nutr 2010, 91, 712–721. [Google Scholar] [CrossRef]
- Ratjen, I.; Schafmayer, C.; di Giuseppe, R.; Waniek, S.; Plachta-Danielzik, S.; Koch, M.; Nothlings, U.; Hampe, J.; Schlesinger, S.; Lieb, W. Postdiagnostic Mediterranean and Healthy Nordic Dietary Patterns Are Inversely Associated with All-Cause Mortality in Long-Term Colorectal Cancer Survivors. J Nutr 2017, 147, 636–644. [Google Scholar] [CrossRef]
- Jochems, S.H.J.; van Osch, F.H.M.; Reulen, R.C.; van Hensbergen, M.; Nekeman, D.; Pirrie, S.; Wesselius, A.; van Schooten, F.J.; James, N.D.; Wallace, D.M.A.; et al. Fruit and vegetable intake and the risk of recurrence in patients with non-muscle invasive bladder cancer: a prospective cohort study. Cancer Causes Control 2018, 29, 573–579. [Google Scholar] [CrossRef]
- Flanagan, L.; Schmid, J.; Ebert, M.; Soucek, P.; Kunicka, T.; Liska, V.; Bruha, J.; Neary, P.; Dezeeuw, N.; Tommasino, M.; et al. Fusobacterium nucleatum associates with stages of colorectal neoplasia development, colorectal cancer and disease outcome. Eur J Clin Microbiol Infect Dis 2014, 33, 1381–1390. [Google Scholar] [CrossRef]
- Mima, K.; Nishihara, R.; Qian, Z.R.; Cao, Y.; Sukawa, Y.; Nowak, J.A.; Yang, J.; Dou, R.; Masugi, Y.; Song, M.; et al. Fusobacterium nucleatum in colorectal carcinoma tissue and patient prognosis. Gut 2016, 65, 1973–1980. [Google Scholar] [CrossRef]
- Wei, Z.; Cao, S.; Liu, S.; Yao, Z.; Sun, T.; Li, Y.; Li, J.; Zhang, D.; Zhou, Y. Could gut microbiota serve as prognostic biomarker associated with colorectal cancer patients' survival? A pilot study on relevant mechanism. Oncotarget 2016, 7, 46158–46172. [Google Scholar] [CrossRef]
- Yan, X.; Liu, L.; Li, H.; Qin, H.; Sun, Z. Clinical significance of Fusobacterium nucleatum, epithelial-mesenchymal transition, and cancer stem cell markers in stage III/IV colorectal cancer patients. Onco Targets Ther 2017, 10, 5031–5046. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.; Guo, F.; Yu, Y.; Sun, T.; Ma, D.; Han, J.; Qian, Y.; Kryczek, I.; Sun, D.; Nagarsheth, N.; et al. Fusobacterium nucleatum Promotes Chemoresistance to Colorectal Cancer by Modulating Autophagy. Cell 2017, 170, 548–563.e516. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Misra, B.B.; Liang, L.; Bi, D.; Weng, W.; Wu, W.; Cai, S.; Qin, H.; Goel, A.; Li, X.; et al. Integrated microbiome and metabolome analysis reveals a novel interplay between commensal bacteria and metabolites in colorectal cancer. Theranostics 2019, 9, 4101–4114. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Rhee, K.J.; Albesiano, E.; Rabizadeh, S.; Wu, X.; Yen, H.R.; Huso, D.L.; Brancati, F.L.; Wick, E.; McAllister, F.; et al. A human colonic commensal promotes colon tumorigenesis via activation of T helper type 17 T cell responses. Nat Med 2009, 15, 1016–1022. [Google Scholar] [CrossRef]
- Arthur, J.C.; Perez-Chanona, E.; Muhlbauer, M.; Tomkovich, S.; Uronis, J.M.; Fan, T.J.; Campbell, B.J.; Abujamel, T.; Dogan, B.; Rogers, A.B.; et al. Intestinal inflammation targets cancer-inducing activity of the microbiota. Science 2012, 338, 120–123. [Google Scholar] [CrossRef]
- Castellarin, M.; Warren, R.L.; Freeman, J.D.; Dreolini, L.; Krzywinski, M.; Strauss, J.; Barnes, R.; Watson, P.; Allen-Vercoe, E.; Moore, R.A.; et al. Fusobacterium nucleatum infection is prevalent in human colorectal carcinoma. Genome Res 2012, 22, 299–306. [Google Scholar] [CrossRef]
- Kostic, A.D.; Gevers, D.; Pedamallu, C.S.; Michaud, M.; Duke, F.; Earl, A.M.; Ojesina, A.I.; Jung, J.; Bass, A.J.; Tabernero, J.; et al. Genomic analysis identifies association of Fusobacterium with colorectal carcinoma. Genome Res 2012, 22, 292–298. [Google Scholar] [CrossRef]
- Ridlon, J.M.; Devendran, S.; Alves, J.M.; Doden, H.; Wolf, P.G.; Pereira, G.V.; Ly, L.; Volland, A.; Takei, H.; Nittono, H.; et al. The 'in vivo lifestyle' of bile acid 7alpha-dehydroxylating bacteria: comparative genomics, metatranscriptomic, and bile acid metabolomics analysis of a defined microbial community in gnotobiotic mice. Gut Microbes 2020, 11, 381–404. [Google Scholar] [CrossRef]
- Flemer, B.; Herlihy, M.; O'Riordain, M.; Shanahan, F.; O'Toole, P.W. Tumour-associated and non-tumour-associated microbiota: Addendum. Gut Microbes 2018, 9, 369–373. [Google Scholar] [CrossRef]
- Najafi, S.; Abedini, F.; Azimzadeh Jamalkandi, S.; Shariati, P.; Ahmadi, A.; Gholami Fesharaki, M. The composition of lung microbiome in lung cancer: a systematic review and meta-analysis. BMC Microbiol 2021, 21, 315. [Google Scholar] [CrossRef]
- Lee, Y.C.; Chiang, T.H.; Chou, C.K.; Tu, Y.K.; Liao, W.C.; Wu, M.S.; Graham, D.Y. Association Between Helicobacter pylori Eradication and Gastric Cancer Incidence: A Systematic Review and Meta-analysis. Gastroenterology 2016, 150, 1113–1124.e1115. [Google Scholar] [CrossRef] [PubMed]
- Tavakoli, A.; Monavari, S.H.; Solaymani Mohammadi, F.; Kiani, S.J.; Armat, S.; Farahmand, M. Association between Epstein-Barr virus infection and gastric cancer: a systematic review and meta-analysis. BMC Cancer 2020, 20, 493. [Google Scholar] [CrossRef] [PubMed]
- Burd, E.M. Human papillomavirus and cervical cancer. Clin Microbiol Rev 2003, 16, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Gao, C.; Yang, Y.; Zhou, F.; Li, M.; Jin, Q.; Gao, L. Systematic review with meta-analysis: the association between human papillomavirus infection and oesophageal cancer. Aliment Pharmacol Ther 2014, 39, 270–281. [Google Scholar] [CrossRef]
- Turkay, D.O.; Vural, C.; Sayan, M.; Gurbuz, Y. Detection of human papillomavirus in esophageal and gastroesophageal junction tumors: A retrospective study by real-time polymerase chain reaction in an instutional experience from Turkey and review of literature. Pathol Res Pract 2016, 212, 77–82. [Google Scholar] [CrossRef]
- Wong, S.H.; Yu, J. Gut microbiota in colorectal cancer: mechanisms of action and clinical applications. Nat Rev Gastroenterol Hepatol 2019, 16, 690–704. [Google Scholar] [CrossRef]
- Colbert, L.E.; El Alam, M.B.; Wang, R.; Karpinets, T.; Lo, D.; Lynn, E.J.; Harris, T.A.; Elnaggar, J.H.; Yoshida-Court, K.; Tomasic, K.; et al. Tumor-resident Lactobacillus iners confer chemoradiation resistance through lactate-induced metabolic rewiring. Cancer Cell 2023, 41, 1945–1962.e1911. [Google Scholar] [CrossRef]
- Feng, L.; Zhang, W.; Shen, Q.; Miao, C.; Chen, L.; Li, Y.; Gu, X.; Fan, M.; Ma, Y.; Wang, H.; et al. Bile acid metabolism dysregulation associates with cancer cachexia: roles of liver and gut microbiome. J Cachexia Sarcopenia Muscle 2021, 12, 1553–1569. [Google Scholar] [CrossRef]
- Galeano Nino, J.L.; Wu, H.; LaCourse, K.D.; Kempchinsky, A.G.; Baryiames, A.; Barber, B.; Futran, N.; Houlton, J.; Sather, C.; Sicinska, E.; et al. Effect of the intratumoral microbiota on spatial and cellular heterogeneity in cancer. Nature 2022, 611, 810–817. [Google Scholar] [CrossRef]
- Yachida, S.; Mizutani, S.; Shiroma, H.; Shiba, S.; Nakajima, T.; Sakamoto, T.; Watanabe, H.; Masuda, K.; Nishimoto, Y.; Kubo, M.; et al. Metagenomic and metabolomic analyses reveal distinct stage-specific phenotypes of the gut microbiota in colorectal cancer. Nat Med 2019, 25, 968–976. [Google Scholar] [CrossRef]
- Dong, Q.; Chen, E.S.; Zhao, C.; Jin, C. Host-Microbiome Interaction in Lung Cancer. Front Immunol 2021, 12, 679829. [Google Scholar] [CrossRef] [PubMed]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. What is the Healthy Gut Microbiota Composition? A Changing Ecosystem across Age, Environment, Diet, and Diseases. Microorganisms 2019, 7. [Google Scholar] [CrossRef] [PubMed]
- Potrykus, M.; Czaja-Stolc, S.; Stankiewicz, M.; Kaska, L.; Malgorzewicz, S. Intestinal Microbiota as a Contributor to Chronic Inflammation and Its Potential Modifications. Nutrients 2021, 13. [Google Scholar] [CrossRef] [PubMed]
- Ou, S.; Wang, H.; Tao, Y.; Luo, K.; Ye, J.; Ran, S.; Guan, Z.; Wang, Y.; Hu, H.; Huang, R. Fusobacterium nucleatum and colorectal cancer: From phenomenon to mechanism. Front Cell Infect Microbiol 2022, 12, 1020583. [Google Scholar] [CrossRef]
- Wang, X.; Huang, X.; Zhang, Y. Involvement of Human Papillomaviruses in Cervical Cancer. Front Microbiol 2018, 9, 2896. [Google Scholar] [CrossRef]
- Salvatori, S.; Marafini, I.; Laudisi, F.; Monteleone, G.; Stolfi, C. Helicobacter pylori and Gastric Cancer: Pathogenetic Mechanisms. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef]
- Morrison, D.J.; Preston, T. Formation of short chain fatty acids by the gut microbiota and their impact on human metabolism. Gut Microbes 2016, 7, 189–200. [Google Scholar] [CrossRef]
- Yang, R.; Qian, L. Research on Gut Microbiota-Derived Secondary Bile Acids in Cancer Progression. Integr Cancer Ther 2022, 21, 15347354221114100. [Google Scholar] [CrossRef]
- Du, Q.; Geller, D.A. Cross-Regulation Between Wnt and NF-kappaB Signaling Pathways. For Immunopathol Dis Therap 2010, 1, 155–181. [Google Scholar] [CrossRef]
- Finlay, B.B. Cell adhesion and invasion mechanisms in microbial pathogenesis. Curr Opin Cell Biol 1990, 2, 815–820. [Google Scholar] [CrossRef]
- Garcia, E.C.; Perault, A.I.; Marlatt, S.A.; Cotter, P.A. Interbacterial signaling via Burkholderia contact-dependent growth inhibition system proteins. Proc Natl Acad Sci U S A 2016, 113, 8296–8301. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Bayona, L.; Guo, M.S.; Laub, M.T. Contact-dependent killing by Caulobacter crescentus via cell surface-associated, glycine zipper proteins. Elife 2017, 6. [Google Scholar] [CrossRef] [PubMed]
- Kumamoto, C.A. A contact-activated kinase signals Candida albicans invasive growth and biofilm development. Proc Natl Acad Sci U S A 2005, 102, 5576–5581. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Li, H.; Tian, G.; Li, S. Dynamic microbe and molecule networks in a mouse model of colitis-associated colorectal cancer. Sci Rep 2014, 4, 4985. [Google Scholar] [CrossRef] [PubMed]
- White, M.G.; Wargo, J.A. Gut Microbes' Impact on Oncogenic Drivers: Location Matters. Mol Cell 2020, 79, 878–880. [Google Scholar] [CrossRef]
- Boopathi, S.; Liu, D.; Jia, A.Q. Molecular trafficking between bacteria determines the shape of gut microbial community. Gut Microbes 2021, 13, 1959841. [Google Scholar] [CrossRef]
- Huang, C.; Mei, S.; Zhang, X.; Tian, X. Inflammatory Milieu Related to Dysbiotic Gut Microbiota Promotes Tumorigenesis of Hepatocellular Carcinoma. J Clin Gastroenterol 2023, 57, 782–788. [Google Scholar] [CrossRef]
- Maciel-Fiuza, M.F.; Muller, G.C.; Campos, D.M.S.; do Socorro Silva Costa, P.; Peruzzo, J.; Bonamigo, R.R.; Veit, T.; Vianna, F.S.L. Role of gut microbiota in infectious and inflammatory diseases. Front Microbiol 2023, 14, 1098386. [Google Scholar] [CrossRef]
- El Tekle, G.; Andreeva, N.; Garrett, W.S. The Role of the Microbiome in the Etiopathogenesis of Colon Cancer. Annu Rev Physiol 2024, 86, 453–478. [Google Scholar] [CrossRef]
- Takenaka, M.C.; Quintana, F.J. Tolerogenic dendritic cells. Semin Immunopathol 2017, 39, 113–120. [Google Scholar] [CrossRef]
- Roberti, M.P.; Yonekura, S.; Duong, C.P.M.; Picard, M.; Ferrere, G.; Tidjani Alou, M.; Rauber, C.; Iebba, V.; Lehmann, C.H.K.; Amon, L.; et al. Chemotherapy-induced ileal crypt apoptosis and the ileal microbiome shape immunosurveillance and prognosis of proximal colon cancer. Nat Med 2020, 26, 919–931. [Google Scholar] [CrossRef] [PubMed]
- Kandalai, S.; Li, H.; Zhang, N.; Peng, H.; Zheng, Q. The human microbiome and cancer: a diagnostic and therapeutic perspective. Cancer Biol Ther 2023, 24, 2240084. [Google Scholar] [CrossRef]
- Zhang, M.; Liu, J.; Xia, Q. Role of gut microbiome in cancer immunotherapy: from predictive biomarker to therapeutic target. Exp Hematol Oncol 2023, 12, 84. [Google Scholar] [CrossRef] [PubMed]
- Whisner, C.M.; Athena Aktipis, C. The Role of the Microbiome in Cancer Initiation and Progression: How Microbes and Cancer Cells Utilize Excess Energy and Promote One Another's Growth. Curr Nutr Rep 2019, 8, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Poore, G.D.; Kopylova, E.; Zhu, Q.; Carpenter, C.; Fraraccio, S.; Wandro, S.; Kosciolek, T.; Janssen, S.; Metcalf, J.; Song, S.J.; et al. Microbiome analyses of blood and tissues suggest cancer diagnostic approach. Nature 2020, 579, 567–574. [Google Scholar] [CrossRef] [PubMed]
- Nejman, D.; Livyatan, I.; Fuks, G.; Gavert, N.; Zwang, Y.; Geller, L.T.; Rotter-Maskowitz, A.; Weiser, R.; Mallel, G.; Gigi, E.; et al. The human tumor microbiome is composed of tumor type-specific intracellular bacteria. Science 2020, 368, 973–980. [Google Scholar] [CrossRef]
- Aykut, B.; Pushalkar, S.; Chen, R.; Li, Q.; Abengozar, R.; Kim, J.I.; Shadaloey, S.A.; Wu, D.; Preiss, P.; Verma, N.; et al. The fungal mycobiome promotes pancreatic oncogenesis via activation of MBL. Nature 2019, 574, 264–267. [Google Scholar] [CrossRef]
- Stolte, M.; Bayerdorffer, E.; Morgner, A.; Alpen, B.; Wundisch, T.; Thiede, C.; Neubauer, A. Helicobacter and gastric MALT lymphoma. Gut 2002, 50 Suppl 3, III19–24. [Google Scholar] [CrossRef]
- Lowy, D.R.; Schiller, J.T. Preventing Cancer and Other Diseases Caused by Human Papillomavirus Infection: 2017 Lasker-DeBakey Clinical Research Award. JAMA 2017, 318, 901–902. [Google Scholar] [CrossRef]
- Bullman, S.; Pedamallu, C.S.; Sicinska, E.; Clancy, T.E.; Zhang, X.; Cai, D.; Neuberg, D.; Huang, K.; Guevara, F.; Nelson, T.; et al. Analysis of Fusobacterium persistence and antibiotic response in colorectal cancer. Science 2017, 358, 1443–1448. [Google Scholar] [CrossRef]
- Jin, C.; Lagoudas, G.K.; Zhao, C.; Bullman, S.; Bhutkar, A.; Hu, B.; Ameh, S.; Sandel, D.; Liang, X.S.; Mazzilli, S.; et al. Commensal Microbiota Promote Lung Cancer Development via gammadelta T Cells. Cell 2019, 176, 998–1013.e1016. [Google Scholar] [CrossRef] [PubMed]
- Pushalkar, S.; Hundeyin, M.; Daley, D.; Zambirinis, C.P.; Kurz, E.; Mishra, A.; Mohan, N.; Aykut, B.; Usyk, M.; Torres, L.E.; et al. The Pancreatic Cancer Microbiome Promotes Oncogenesis by Induction of Innate and Adaptive Immune Suppression. Cancer Discov 2018, 8, 403–416. [Google Scholar] [CrossRef] [PubMed]
- Le Noci, V.; Guglielmetti, S.; Arioli, S.; Camisaschi, C.; Bianchi, F.; Sommariva, M.; Storti, C.; Triulzi, T.; Castelli, C.; Balsari, A.; et al. Modulation of Pulmonary Microbiota by Antibiotic or Probiotic Aerosol Therapy: A Strategy to Promote Immunosurveillance against Lung Metastases. Cell Rep 2018, 24, 3528–3538. [Google Scholar] [CrossRef]
- Mackowiak, P.A. Recycling metchnikoff: probiotics, the intestinal microbiome and the quest for long life. Front Public Health 2013, 1, 52. [Google Scholar] [CrossRef]
- Fotiadis, C.I.; Stoidis, C.N.; Spyropoulos, B.G.; Zografos, E.D. Role of probiotics, prebiotics and synbiotics in chemoprevention for colorectal cancer. World J Gastroenterol 2008, 14, 6453–6457. [Google Scholar] [CrossRef]
- Fong, W.; Li, Q.; Yu, J. Gut microbiota modulation: a novel strategy for prevention and treatment of colorectal cancer. Oncogene 2020, 39, 4925–4943. [Google Scholar] [CrossRef]
- Klaenhammer, T.R.; Kleerebezem, M.; Kopp, M.V.; Rescigno, M. The impact of probiotics and prebiotics on the immune system. Nat Rev Immunol 2012, 12, 728–734. [Google Scholar] [CrossRef]
- Kumar, M.; Kissoon-Singh, V.; Coria, A.L.; Moreau, F.; Chadee, K. Probiotic mixture VSL#3 reduces colonic inflammation and improves intestinal barrier function in Muc2 mucin-deficient mice. Am J Physiol Gastrointest Liver Physiol 2017, 312, G34–G45. [Google Scholar] [CrossRef]
- Martin, R.; Chamignon, C.; Mhedbi-Hajri, N.; Chain, F.; Derrien, M.; Escribano-Vazquez, U.; Garault, P.; Cotillard, A.; Pham, H.P.; Chervaux, C.; et al. The potential probiotic Lactobacillus rhamnosus CNCM I-3690 strain protects the intestinal barrier by stimulating both mucus production and cytoprotective response. Sci Rep 2019, 9, 5398. [Google Scholar] [CrossRef]
- Wang, L.; Cao, H.; Liu, L.; Wang, B.; Walker, W.A.; Acra, S.A.; Yan, F. Activation of epidermal growth factor receptor mediates mucin production stimulated by p40, a Lactobacillus rhamnosus GG-derived protein. J Biol Chem 2014, 289, 20234–20244. [Google Scholar] [CrossRef]
- Zyrek, A.A.; Cichon, C.; Helms, S.; Enders, C.; Sonnenborn, U.; Schmidt, M.A. Molecular mechanisms underlying the probiotic effects of Escherichia coli Nissle 1917 involve ZO-2 and PKCzeta redistribution resulting in tight junction and epithelial barrier repair. Cell Microbiol 2007, 9, 804–816. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, C.S.; Badia, J.; Bosch, M.; Gimenez, R.; Baldoma, L. Outer Membrane Vesicles and Soluble Factors Released by Probiotic Escherichia coli Nissle 1917 and Commensal ECOR63 Enhance Barrier Function by Regulating Expression of Tight Junction Proteins in Intestinal Epithelial Cells. Front Microbiol 2016, 7, 1981. [Google Scholar] [CrossRef]
- Mills, J.P.; Rao, K.; Young, V.B. Probiotics for prevention of Clostridium difficile infection. Curr Opin Gastroenterol 2018, 34, 3–10. [Google Scholar] [CrossRef]
- Piewngam, P.; Zheng, Y.; Nguyen, T.H.; Dickey, S.W.; Joo, H.S.; Villaruz, A.E.; Glose, K.A.; Fisher, E.L.; Hunt, R.L.; Li, B.; et al. Pathogen elimination by probiotic Bacillus via signalling interference. Nature 2018, 562, 532–537. [Google Scholar] [CrossRef]
- Weingarden, A.R.; Vaughn, B.P. Intestinal microbiota, fecal microbiota transplantation, and inflammatory bowel disease. Gut Microbes 2017, 8, 238–252. [Google Scholar] [CrossRef]
- Smits, L.P.; Bouter, K.E.; de Vos, W.M.; Borody, T.J.; Nieuwdorp, M. Therapeutic potential of fecal microbiota transplantation. Gastroenterology 2013, 145, 946–953. [Google Scholar] [CrossRef]
- Baruch, E.N.; Youngster, I.; Ben-Betzalel, G.; Ortenberg, R.; Lahat, A.; Katz, L.; Adler, K.; Dick-Necula, D.; Raskin, S.; Bloch, N.; et al. Fecal microbiota transplant promotes response in immunotherapy-refractory melanoma patients. Science 2021, 371, 602–609. [Google Scholar] [CrossRef]
- Nandi, D.; Parida, S.; Sharma, D. The gut microbiota in breast cancer development and treatment: The good, the bad, and the useful! Gut Microbes 2023, 15, 2221452. [Google Scholar] [CrossRef]
- Hohmann, E.L.; Ananthakrishnan, A.N.; Deshpande, V. Case Records of the Massachusetts General Hospital. Case 25-2014. A 37-year-old man with ulcerative colitis and bloody diarrhea. N Engl J Med 2014, 371, 668–675. [Google Scholar] [CrossRef]
- Quera, R.; Espinoza, R.; Estay, C.; Rivera, D. Bacteremia as an adverse event of fecal microbiota transplantation in a patient with Crohn's disease and recurrent Clostridium difficile infection. J Crohns Colitis 2014, 8, 252–253. [Google Scholar] [CrossRef]
- Ridaura, V.K.; Faith, J.J.; Rey, F.E.; Cheng, J.; Duncan, A.E.; Kau, A.L.; Griffin, N.W.; Lombard, V.; Henrissat, B.; Bain, J.R.; et al. Gut microbiota from twins discordant for obesity modulate metabolism in mice. Science 2013, 341, 1241214. [Google Scholar] [CrossRef] [PubMed]
- Luke, J.J.; Piha-Paul, S.A.; Medina, T.; Verschraegen, C.F.; Varterasian, M.; Brennan, A.M.; Riese, R.J.; Sokolovska, A.; Strauss, J.; Hava, D.L.; et al. Phase I Study of SYNB1891, an Engineered E. coli Nissle Strain Expressing STING Agonist, with and without Atezolizumab in Advanced Malignancies. Clin Cancer Res 2023, 29, 2435–2444. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.H.; Nguyen, V.H.; Jiang, S.N.; Park, S.H.; Tan, W.; Hong, S.H.; Shin, M.G.; Chung, I.J.; Hong, Y.; Bom, H.S.; et al. Two-step enhanced cancer immunotherapy with engineered Salmonella typhimurium secreting heterologous flagellin. Sci Transl Med 2017, 9. [Google Scholar] [CrossRef] [PubMed]
- Din, M.O.; Danino, T.; Prindle, A.; Skalak, M.; Selimkhanov, J.; Allen, K.; Julio, E.; Atolia, E.; Tsimring, L.S.; Bhatia, S.N.; et al. Synchronized cycles of bacterial lysis for in vivo delivery. Nature 2016, 536, 81–85. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Huang, L.; Lin, D.; Lai, X.; Wu, L.; Liao, X.; Liu, J.; Zeng, Y.; Liang, L.; Zhang, G.; et al. GD2-specific chimeric antigen receptor-modified T cells for the treatment of refractory and/or recurrent neuroblastoma in pediatric patients. J Cancer Res Clin Oncol 2022, 148, 2643–2652. [Google Scholar] [CrossRef]
- Yang, L.; Li, A.; Wang, Y.; Zhang, Y. Intratumoral microbiota: roles in cancer initiation, development and therapeutic efficacy. Signal Transduct Target Ther 2023, 8, 35. [Google Scholar] [CrossRef]
- Rizvi, Z.A.; Dalal, R.; Sadhu, S.; Kumar, Y.; Kumar, S.; Gupta, S.K.; Tripathy, M.R.; Rathore, D.K.; Awasthi, A. High-salt diet mediates interplay between NK cells and gut microbiota to induce potent tumor immunity. Sci Adv 2021, 7, eabg5016. [Google Scholar] [CrossRef]
- Overacre-Delgoffe, A.E.; Bumgarner, H.J.; Cillo, A.R.; Burr, A.H.P.; Tometich, J.T.; Bhattacharjee, A.; Bruno, T.C.; Vignali, D.A.A.; Hand, T.W. Microbiota-specific T follicular helper cells drive tertiary lymphoid structures and anti-tumor immunity against colorectal cancer. Immunity 2021, 54, 2812–2824.e2814. [Google Scholar] [CrossRef]
- Woo, Y.R.; Cho, S.H.; Lee, J.D.; Kim, H.S. The Human Microbiota and Skin Cancer. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- Whelan, K. Probiotics and prebiotics in the management of irritable bowel syndrome: a review of recent clinical trials and systematic reviews. Curr Opin Clin Nutr Metab Care 2011, 14, 581–587. [Google Scholar] [CrossRef]
- Szajewska, H.; Guarino, A.; Hojsak, I.; Indrio, F.; Kolacek, S.; Shamir, R.; Vandenplas, Y.; Weizman, Z.; European Society for Pediatric Gastroenterology, H.; Nutrition. Use of probiotics for management of acute gastroenteritis: a position paper by the ESPGHAN Working Group for Probiotics and Prebiotics. J Pediatr Gastroenterol Nutr 2014, 58, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Denipote, F.G.; Trindade, E.B.; Burini, R.C. [Probiotics and prebiotics in primary care for colon cancer]. Arq Gastroenterol 2010, 47, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Iannitti, T.; Palmieri, B. Therapeutical use of probiotic formulations in clinical practice. Clin Nutr 2010, 29, 701–725. [Google Scholar] [CrossRef] [PubMed]
- Rooks, M.G.; Garrett, W.S. Bacteria, food, and cancer. F1000 Biol Rep 2011, 3, 12. [Google Scholar] [CrossRef] [PubMed]
- Bose, S.; Zhang, C.; Le, A. Glucose Metabolism in Cancer: The Warburg Effect and Beyond. Adv Exp Med Biol 2021, 1311, 3–15. [Google Scholar] [CrossRef]
- Liberti, M.V.; Locasale, J.W. The Warburg Effect: How Does it Benefit Cancer Cells? Trends Biochem Sci 2016, 41, 211–218. [Google Scholar] [CrossRef]
- Bultman, S.J. The microbiome and its potential as a cancer preventive intervention. Semin Oncol 2016, 43, 97–106. [Google Scholar] [CrossRef]
- Daschner, P.J.; Ross, S.; Seifried, H.; Kumar, A.; Flores, R. Nutrition and Microbiome Interactions in Human Cancer. J Acad Nutr Diet 2023, 123, 504–514. [Google Scholar] [CrossRef]
- Kim, H.S.; Kim, Y.J.; Seo, Y.R. An Overview of Carcinogenic Heavy Metal: Molecular Toxicity Mechanism and Prevention. J Cancer Prev 2015, 20, 232–240. [Google Scholar] [CrossRef]
- Chen, Q.Y.; DesMarais, T.; Costa, M. Metals and Mechanisms of Carcinogenesis. Annu Rev Pharmacol Toxicol 2019, 59, 537–554. [Google Scholar] [CrossRef]
- Rayan, M.; Sayed, T.S.; Hussein, O.J.; Therachiyil, L.; Maayah, Z.H.; Maccalli, C.; Uddin, S.; Prehn, J.H.M.; Korashy, H.M. Unlocking the secrets: exploring the influence of the aryl hydrocarbon receptor and microbiome on cancer development. Cell Mol Biol Lett 2024, 29, 33. [Google Scholar] [CrossRef] [PubMed]
- Goedtke, L.; Sprenger, H.; Hofmann, U.; Schmidt, F.F.; Hammer, H.S.; Zanger, U.M.; Poetz, O.; Seidel, A.; Braeuning, A.; Hessel-Pras, S. Polycyclic Aromatic Hydrocarbons Activate the Aryl Hydrocarbon Receptor and the Constitutive Androstane Receptor to Regulate Xenobiotic Metabolism in Human Liver Cells. Int J Mol Sci 2020, 22. [Google Scholar] [CrossRef] [PubMed]
- Claus, S.P.; Guillou, H.; Ellero-Simatos, S. The gut microbiota: a major player in the toxicity of environmental pollutants? NPJ Biofilms Microbiomes 2016, 2, 16003. [Google Scholar] [CrossRef]
- Lagunas-Rangel, F.A.; Linnea-Niemi, J.V.; Kudlak, B.; Williams, M.J.; Jonsson, J.; Schioth, H.B. Role of the Synergistic Interactions of Environmental Pollutants in the Development of Cancer. Geohealth 2022, 6, e2021GH000552. [Google Scholar] [CrossRef] [PubMed]
- Wegierska, A.E.; Charitos, I.A.; Topi, S.; Potenza, M.A.; Montagnani, M.; Santacroce, L. The Connection Between Physical Exercise and Gut Microbiota: Implications for Competitive Sports Athletes. Sports Med 2022, 52, 2355–2369. [Google Scholar] [CrossRef] [PubMed]
- Monda, V.; Villano, I.; Messina, A.; Valenzano, A.; Esposito, T.; Moscatelli, F.; Viggiano, A.; Cibelli, G.; Chieffi, S.; Monda, M.; et al. Exercise Modifies the Gut Microbiota with Positive Health Effects. Oxid Med Cell Longev 2017, 2017, 3831972. [Google Scholar] [CrossRef]
- Lou, H.; Liu, X.; Liu, P. Mechanism and implications of pro-nature physical activity in antagonizing psychological stress: the key role of microbial-gut-brain axis. Front Psychol 2023, 14, 1143827. [Google Scholar] [CrossRef]
- Drochioiu, G. Multifactorial Distress, the Warburg Effect, and Respiratory and pH Imbalance in Cancer Development. Stresses 2023, 3, 500–528. [Google Scholar] [CrossRef]
- Madison, A.; Kiecolt-Glaser, J.K. Stress, depression, diet, and the gut microbiota: human-bacteria interactions at the core of psychoneuroimmunology and nutrition. Curr Opin Behav Sci 2019, 28, 105–110. [Google Scholar] [CrossRef]
- Leigh, S.J.; Uhlig, F.; Wilmes, L.; Sanchez-Diaz, P.; Gheorghe, C.E.; Goodson, M.S.; Kelley-Loughnane, N.; Hyland, N.P.; Cryan, J.F.; Clarke, G. The impact of acute and chronic stress on gastrointestinal physiology and function: a microbiota-gut-brain axis perspective. J Physiol 2023, 601, 4491–4538. [Google Scholar] [CrossRef]
- Yu, Y.N.; Fang, J.Y. Gut Microbiota and Colorectal Cancer. Gastrointest Tumors 2015, 2, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Manzoor, S.S.; Doedens, A.; Burns, M.B. The promise and challenge of cancer microbiome research. Genome Biol 2020, 21, 131. [Google Scholar] [CrossRef] [PubMed]
- Wan, Y.Y.; Jena, P.K. Precision dietary supplementation based on personal gut microbiota. Nat Rev Gastroenterol Hepatol 2019, 16, 204–206. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.L.; Vieira-Silva, S.; Liston, A.; Raes, J. How informative is the mouse for human gut microbiota research? Dis Model Mech 2015, 8, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.H. Harness the functions of gut microbiome in tumorigenesis for cancer treatment. Cancer Commun (Lond) 2021, 41, 937–967. [Google Scholar] [CrossRef]
- Chanyi, R.M.; Craven, L.; Harvey, B.; Reid, G.; Silverman, M.J.; Burton, J.P. Faecal microbiota transplantation: Where did it start? What have studies taught us? Where is it going? SAGE Open Med 2017, 5, 2050312117708712. [Google Scholar] [CrossRef]
- Rezasoltani, S.; Ahmadi Bashirzadeh, D.; Nazemalhosseini Mojarad, E.; Asadzadeh Aghdaei, H.; Norouzinia, M.; Shahrokh, S. Signature of Gut Microbiome by Conventional and Advanced Analysis Techniques: Advantages and Disadvantages. Middle East J Dig Dis 2020, 12, 5–11. [Google Scholar] [CrossRef]
- Scott, A.J.; Alexander, J.L.; Merrifield, C.A.; Cunningham, D.; Jobin, C.; Brown, R.; Alverdy, J.; O'Keefe, S.J.; Gaskins, H.R.; Teare, J.; et al. International Cancer Microbiome Consortium consensus statement on the role of the human microbiome in carcinogenesis. Gut 2019, 68, 1624–1632. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



