
Submitted:
01 October 2024
Posted:
02 October 2024
You are already at the latest version
Abstract
This review investigates the impact of bioactive molecules produced by probiotics on child health, focusing on their roles in modulating gut microbiota, enhancing immune function, and supporting overall development. Key metabolites, including short-chain fatty acids (SCFAs), bacteriocins, exopolysaccharides (EPS), vitamins, and gamma-aminobutyric acid (GABA), are highlighted for their ability to maintain gut health, regulate inflammation, and support neurodevelopment. Clinical studies demonstrate that specific probiotics and their metabolites can reduce gastrointestinal disorders, enhance immune responses, and decrease the incidence of allergies and respiratory infections in pediatric populations. Additionally, postbiotics—bioactive compounds from probiotic fermentation—offer promising benefits, such as improved gut barrier function, reduced inflammation, and enhanced nutrient absorption, while presenting fewer safety concerns compared to live probiotics. By examining the mechanisms of action and clinical evidence, this review underscores the potential of integrating probiotics and postbiotics into pediatric healthcare strategies to promote optimal growth, development, and overall health in children.
Keywords:
probiotics
; short chain fatty acids
; exopolissaccharide
; vitamins
; peptides
; postbiotics
1. Introduction
Childhood is a critical period characterized by rapid growth, development, and maturation of various physiological systems, including the gastrointestinal tract and the immune system. During this stage, maintaining a balanced gut microbiota is essential, as it plays a vital role in nutrient absorption, metabolism, immune development, and protection against pathogens [1,2]. Disruptions in the gut microbiome, such as those caused by antibiotics, poor nutrition, or infections, can lead to various health issues in children, including gastrointestinal disorders, allergies, and impaired immune responses [3]. Recent research has highlighted the potential of probiotics—live microorganisms that confer health benefits to the host—to modulate the gut microbiota and improve pediatric health outcomes [4,5].
A key aspect of probiotics’ benefits lies in the bioactive metabolites they produce. These metabolites, including short-chain fatty acids (SCFAs), bacteriocins, exopolysaccharides (EPS), vitamins, and neuroactive compounds like gamma-aminobutyric acid (GABA), are crucial mediators of the beneficial effects associated with probiotics [6,7]. For instance, SCFAs, such as acetate, propionate, and butyrate, play significant roles in maintaining gut barrier integrity, modulating immune function, and providing energy to colonocytes, which are particularly important in infants and young children whose immune and digestive systems are still developing [8,9]
Probiotic metabolites also contribute to the regulation of inflammatory responses, which is crucial in preventing conditions such as necrotizing enterocolitis (NEC), a severe gastrointestinal disorder that predominantly affects preterm infants [10,11]. Metabolites like bacteriocins, antimicrobial peptides produced by probiotics, help in protecting against pathogenic bacteria by disrupting their cell membranes, thereby preventing infections such as antibiotic-associated diarrhea. In addition, exopolysaccharides (EPS) produced by probiotic strains support biofilm formation, promoting long-term colonization of the gut and protection against pathogens [12,13,14].
Furthermore, these metabolites have a direct impact on nutrient absorption, a vital function during childhood, a period marked by increased nutritional demands for growth and development. For example, certain metabolites enhance the bioavailability of essential vitamins and minerals, which are critical for bone growth, cognitive development, and overall health [15]. Additionally, bioactive molecules like GABA, produced by certain probiotic strains, have been implicated in supporting neurodevelopment and emotional regulation through their interaction with the gut-brain axis [16,17].
Given the diverse and critical roles of these bioactive metabolites in promoting child health, there is growing interest in understanding their specific mechanisms of action and potential applications in pediatric care. This review aims to provide a comprehensive overview of the various metabolites produced by probiotics and their impact on child health. We will discuss the specific types of metabolites, such as SCFAs, bacteriocins, EPS, vitamins, and neuroactive compounds, and explore how these molecules contribute to gastrointestinal health, immune modulation, nutrient absorption, and neurodevelopment in children. By examining the evidence, this review seeks to highlight the potential of incorporating probiotic-derived metabolites into pediatric healthcare strategies to enhance the health and well-being of children.
2. Adaptive Role of Metabolites Produced by Probiotic Bacteria
From an evolutionary perspective, the ability of probiotic bacteria to produce metabolites enables them to adapt effectively to their host [18,19]. Bacteria that generate beneficial metabolites gain a survival advantage by outcompeting harmful microbes and fostering symbiotic relationships with their hosts. This evolutionary trait allows beneficial bacteria to thrive and proliferate within the gut environment, supporting host health and contributing to the stability of the microbial community [20,21].
For example, probiotic bacteria produce various metabolites and molecules as part of their complex survival strategies within the gut environment, ensuring their survival and competitiveness while significantly contributing to the host’s health [22]. They compete for space and nutrients by producing metabolites such as SCFAs and bacteriocins, which inhibit the growth of pathogenic bacteria and other microbes [23]. SCFAs, like acetate, propionate, and butyrate lower the gut pH, creating a less hospitable for harmful pathogens and favoring the growth of beneficial bacteria, thus maintaining their niche within the gut microbiota [24]. Additionally, bacteriocins, which are ribosomally synthesized antimicrobial peptides, target and kill competing bacterial strains by permeabilizing their cell membranes, ensuring the dominance of probiotic strains in their ecological niche [25].
Furthermore, EPS, high-molecular-weight polymers secreted by probiotic bacteria, play a crucial role in biofilm formation. Biofilms are structured communities of bacteria encased in a self-produced matrix that adheres to surfaces that protect the bacteria from environmental stresses, including desiccation, antibiotics, and the host immune system [14]. This stable environment enhances bacterial survival and promotes long-term colonization in the gut [26].
In addition, the production of metabolites like vitamins and SCFAs fosters a symbiotic relationship with the host [27] For example, vitamins such as B12 and folate, synthesized by bacteria like Lactobacillus reuteri, are essential for the host’s metabolic processes, including DNA synthesis and energy metabolism [28,29]. By providing these essential nutrients, probiotics contribute to the host’s nutritional status and overall health, creating a mutually beneficial relationship [30].
Moreover, SCFAs produced by probiotic bacteria serve as an energy source for colonocytes and play a role in maintaining the integrity of the gut barrier, which is crucial for preventing the translocation of pathogens and toxins from the gut into the bloodstream, thereby protecting the host from infections and inflammation [31]. Additionally, SCFAs have been shown to modulate the host immune system, promoting anti-inflammatory responses and enhancing immune tolerance [32]. A summary of the metabolites and their effects are illustrated in Figure 1.
In conclusion, probiotic bacteria produce these metabolites not out of altruism but as part of their survival strategies within the gut environment. The range of survival strategies employed by these bacteria is as diverse as the array of molecules they produce, which, while primarily serving their own persistence and competitiveness, ultimately have significant and beneficial effects on the host’s health.
3. Overview of Probiotics Benefits in Child Health
Probiotics are increasingly recognized for their beneficial effects on pediatric health, particularly in managing gastrointestinal disorders and enhancing immune function [4]. Common probiotic strains found in pediatric populations, such as Lactobacillus and Bifidobacterium species, are often incorporated into new probiotic foods due to their health-promoting effects, including immunomodulation, control of diarrhea, and potential benefits in managing inflammatory bowel diseases like Crohn’s disease [33,34]. Although there is a widespread belief that these effects are strain-specific, evidence from randomized controlled trials (RCTs) indicates that the clinical benefits of probiotics in children are more likely to be genus- or species-specific, with few significant differences observed between strains of the same species [4].
Probiotics have demonstrated efficacy in treating various gastrointestinal disorders in children, including necrotizing enterocolitis (NEC), antibiotic-associated diarrhea, and Helicobacter pylori infections. Both single-organism and multi-strain probiotic formulations have shown positive outcomes in these conditions [35,36,37]. As ongoing research continues to support their integration into pediatric healthcare, probiotics hold promise for reducing the incidence and severity of gastrointestinal disorders and improving overall health outcomes in children.
Beyond gastrointestinal health, probiotic strains and their metabolites contribute to children’s well-being in multiple ways. The bioactive molecules produced by probiotics can modulate the immune response, strengthening the body’s defense against infections and illnesses [38]. This immune modulation can potentially decrease the frequency and severity of common childhood ailments, such as colds and respiratory infections [39].
Probiotics also play a vital role in enhancing the absorption and utilization of essential nutrients, ensuring that children receive adequate vitamins, minerals, and other nutrients necessary for optimal growth and development [40]. This is particularly important during childhood, a critical period for growth and maturation. Additionally, some metabolites produced by probiotic strains possess anti-inflammatory properties that help reduce inflammation in the gut and throughout the body [41]. By promoting a balanced inflammatory response, probiotics may lower the risk of inflammation-related conditions in children, contributing to their overall health and well-being [41,42]. Furthermore, probiotics have shown potential in managing allergies in children by modulating immune responses, reducing inflammation, and promoting regulatory T cell activity [43]. For example, strains like Lactobacillus rhamnosus GG have been demonstrated to alleviate allergic symptoms, such as atopic dermatitis and food allergies, by balancing the immune system and enhancing anti-inflammatory pathways.
A comprehensive understanding of the metabolites produced by probiotics and their mechanisms of action are essential for their effective integration into pediatric healthcare. This knowledge, combined with ongoing research, offers significant potential for reducing the incidence and severity of gastrointestinal disorders and enhancing overall health outcomes in children.
4. Specific Probiotic Strains for Children’s Health
When it comes to enhancing children’s health, selecting specific probiotic strains plays a crucial role in providing targeted benefits. Research has highlighted the efficacy of particular probiotic strains in promoting gut health, bolstering the immune system, and alleviating various health issues in children [44,45]. The gut microbiota of children is composed of numerous microorganisms, each potentially contributing to child health and development. However, this section will focus on those microorganisms with the strongest scientific evidence supporting their positive effects in children, as well as those with the most detailed descriptions of the metabolites they produce.
4.1. Lactobacillus rhamnosus
L. rhamnosus is a facultative anaerobic bacterium known for its extensive genomic diversity and metabolic flexibility. Its genome is highly adaptable, featuring a broad array of genes encoding transporters and enzymes that facilitate the utilization of various carbohydrates, including lactose, glucose, galactose, and fucose. This adaptability enables L. rhamnosus to thrive in different niches within the gastrointestinal tract [46,47].
The bacterium’s ability to metabolize a wide range of carbohydrates is complemented by its production of key metabolites. In children, L. rhamnosus produces several important metabolites, including lactic acid, acetic acid, and short-chain fatty acids (SCFAs) such as butyrate. These metabolites are crucial for maintaining a low pH in the gut, which inhibits the growth of pathogenic bacteria and fosters a healthy gut environment [48].
Additionally, L. rhamnosus metabolizes fucose through unique pathways, resulting in the production of lactic acid, 1,2-propanediol, acetic acid, formic acid, and carbon dioxide. These metabolites are vital for energy production and biomass formation, which distinguish L. rhamnosus from other lactic acid bacteria [48].
L. rhamnosus also possesses several genes responsible for synthesizing antimicrobial peptides, including bacteriocins, which inhibit the growth of pathogenic bacteria [49]. Furthermore, it produces EPS that are crucial for biofilm formation. These EPS enhance the bacterium’s ability to adhere to intestinal surfaces and resist environmental stressors, such as bile salts and low pH [50].
The presence of genes encoding antioxidant enzymes, such as superoxide dismutase (SOD) and glutathione reductase, further contributes to its protective capabilities. These enzymes help protect both the bacterium and host cells from oxidative stress [50,51].
L. rhamnosus has also been shown to modulate host immune responses by interacting with dendritic cells and promoting the production of anti-inflammatory cytokines, such as IL-10. This immunomodulatory effect underscores its value in maintaining gut health [46,47].
Moreover, L. rhamnosus produces inosine, a metabolite with antioxidant, anti-inflammatory, anti-infective, and neuroprotective properties. The production of inosine is notably upregulated in L. rhamnosus compared to other Lactobacillus species, suggesting a unique metabolic profile that enhances its probiotic and postbiotic activities [52].
4.2. Bifidobacterium infantis
B. longum subsp. infantis is particularly adapted to the infant gut environment, largely due to its unique genomic configuration that enables the metabolism of human milk oligosaccharides (HMOs). The genomics of B. infantis reveal specialized adaptations for thriving in the infant gut, including unique metabolic pathways, genetic stability, and competitive advantages through bacteriocin production [53].
The genome of B. infantis contains a vast array of genes encoding glycosyl hydrolases and transport proteins that specifically target and degrade HMOs, providing a competitive advantage in the infant gut [54,55].
Additionally, B. infantis produces key metabolites, such as acetate and lactate, which have been shown to strengthen the intestinal barrier by enhancing tight junction integrity [56]. The bacterium’s genome also contains genes for producing various short-chain fatty acids (SCFAs), which serve as energy sources for colonocytes and play a role in maintaining gut homeostasis [57]. Although the genome of B. infantis does not directly contain genes for producing indole-3-lactic acid (ILA), its production of ILA results from its ability to metabolize tryptophan present in the growth medium, especially when grown on HMOs [58].
Furthermore, B. infantis stands out among Bifidobacterium species for its numerous bacteriocin gene clusters, including lanthipeptides and thiopeptides. These bacteriocins provide a competitive edge in colonizing the infant gut by inhibiting harmful microorganisms [59].
In addition to its gastrointestinal benefits, B. infantis produces inosine, a metabolite of B. infantis, has been shown to exert cardioprotective effects. It mitigates cardiac inflammation and cell death during ischemia/reperfusion injury by activating the adenosine A2A receptor, which reduces pro-inflammatory cytokines and supports ATP generation through the purine salvage pathway [60].
4.3. Streptococcus thermophilus
S. thermophilus, a widely recognized probiotic bacterium found in fermented dairy products, plays a pivotal role in influencing gut health and host metabolism through its diverse metabolic activities. One of its primary metabolites is lactate, produced via the glycolysis pathway [61]. Lactate is crucial for lowering the pH in the gut, which not only facilitates milk coagulation during fermentation but also modulates the colon epithelium. This modulation affects the expression of various transporters and proteins involved in cell cycle regulation, thereby positively impacting gut health and function [62].
In addition to lactate, S. thermophilus produces a range of amino acids, including leucine, isoleucine, proline, aspartic acid, and tryptophan. These amino acids are essential for the bacterium’s growth and significantly contribute to the sensory properties and quality of fermented milk products. The strain also generates various fatty acids and other metabolites, such as 2-hydroxybutyric acid, D-glycerol-D-galactose-heptanol, and hydra starch, which influence the flavor profile and overall characteristics of the fermentation process [63].
Metabolic pathways in S. thermophilus involve the breakdown and utilization of several amino acids, including cysteine, methionine, glutamate, glutamine, arginine, aspartate, asparagine, and alanine. These amino acids are integral to the synthesis of glutathione, a potent antioxidant that helps the bacterium combat oxidative stress and environmental challenges [64]. Moreover, S. thermophilus affects gut metabolism by altering tryptophan metabolism, leading to reduced levels of indole derivatives and increased production of serotonin. This metabolic shift has broader implications for mood regulation and gut-brain interactions [65].
Furthermore, S. thermophilus produces a range of antibiotic-like compounds and bactericidal proteins, such as bacteriocins, which are instrumental in reducing uremia and inhibiting the growth of pathogenic microbes. These antimicrobial properties enhance the probiotic benefits of S. thermophilus, making it a valuable contributor to gut health and a potent preventive measure against infections caused by harmful bacteria [66]. Overall, the metabolic versatility and probiotic attributes of S. thermophilus underscore its significance in both industrial applications and health-related functions.
4.4. Lactobacillus acidophilus
Lactobacillus acidophilus is a probiotic microorganism known for producing a range of metabolites that contribute to gut health, including conjugated linoleic acid (CLA), exopolysaccharides (EPSs), and bacteriocins (BACs). The production of these metabolites is influenced by several factors, such as initial pH, temperature, incubation time, yeast extract concentration, and the availability of free linoleic acid [67].
Bacteriocins, which are antimicrobial peptides produced by L. acidophilus, play a key role in inhibiting the growth of pathogenic bacteria [68]. Unlike conventional antibiotics, these bacteriocins have a relatively narrow spectrum of activity and can be degraded by proteases in the gastrointestinal tract. This may limit their direct antimicrobial efficacy but also reduces their potential to disrupt beneficial microbiota [69]. Certain bacteriocin-producing strains, such as L. acidophilus JCM1132, have been shown to alter gut microbiota composition in healthy mice, reducing inflammatory responses and potentially preventing metabolic diseases. This finding underscores the probiotic potential of bacteriocin-producing strains in modulating gut health [70].
Another important metabolite produced by L. acidophilus is valeric acid, a short-chain fatty acid (SCFA) that has been demonstrated to suppress the development of non-alcoholic fatty liver disease-associated hepatocellular carcinoma (NAFLD-HCC) by inhibiting specific cellular pathways [71].
L. acidophilus also produces several antigenic proteins, such as GroEL (HSP60), enolase, and transcription factors EF-Ts and EF-Tu. These proteins are recognized by serum IgG antibodies in children, particularly those with autoimmune conditions like type 1 diabetes and celiac disease, suggesting a complex interaction between the immune system and commensal bacteria [72].
The strain L. acidophilus 5e2 synthesizes exopolysaccharides composed of glucose, galactose, and glucosamine, which may promote gut health by fostering beneficial microbial communities. Additionally, Lactobacillus species produce biosurfactants such as surlactin, which have the capacity to reduce surface tension and inhibit pathogen adhesion, thereby maintaining a balanced microbiota and providing protection against infections [73,74].
4.5. Saccharomyces boulardii
S. boulardii, a probiotic yeast, is well-known for its therapeutic benefits, particularly in gastrointestinal health. Recent research has expanded our understanding of the metabolites produced by S. boulardii, highlighting its potential applications in gastrointestinal disorders, cancer treatment, and as a microbial cell factory.
Saccharomyces cerevisiae var. boulardii synthesizes a diverse range of bioactive metabolites, including polyphenolic compounds such as vanillic acid, cinnamic acid, and phenyl ethyl alcohol, as well as essential nutrients like vitamin B6. These compounds contribute to its antioxidant capacity and provide a foundation for its anti-carcinogenic, antibacterial, antiviral, and general health-promoting properties [75,76]. Additionally, S. boulardii produces molecules like erythromycin and amphetamine, further enhancing its therapeutic profile.
One of the notable metabolites produced by S. boulardii, particularly by the strain S. boulardii-B508, is the Saccharomyces anti-inflammatory factor (SAIF). This factor has been shown to reduce the burden of Mycobacterium intracellulare in human macrophages by inducing apoptosis in infected cells and inhibiting IL-8 expression through the suppression of NF-κB activation, a key regulator of the human inflammatory response [77]. S. boulardii also synthesizes a phosphatase capable of dephosphorylating endotoxins such as the lipopolysaccharide (LPS) of Escherichia coli, thereby reducing their cytotoxic effects. Furthermore, it produces a 54-kDa serine protease that decreases intestinal permeability and inhibits the secretion of water and electrolytes, enhancing its ability to protect against bacterial toxins and exert anti-inflammatory effects in the gastrointestinal tract [78,79].
Moreover, S. boulardii generates high levels of acetic acid at 37 °C, a characteristic linked to unique mutations in the SDH1 and WHI2 genes, which are not found in S. cerevisiae. These genetic traits enable the yeast to thrive in acidic environments, providing resistance to gastric conditions and supporting its efficacy as a probiotic [80,81]. Additionally, S. boulardii produces a serine protease that cleaves Clostridioides difficile toxin A, stimulates the production of antibodies against this toxin, and modulates inflammatory responses by promoting anti-inflammatory molecules like peroxisome proliferator-activated receptor-gamma (PPAR-γ) [82].
In summary, the array of bioactive compounds produced by S. boulardii, including organic acids, enzymes, polyphenols, and proteases, underlines its diverse probiotic activities and therapeutic potential, making it a valuable tool in managing various health conditions.
5. Bioactive Metabolites Produced by Probiotics: Mechanisms of Action and Their Role in Enhancing Pediatric Health
5.1. Immunomodulatory Metabolites
Probiotic bacteria, particularly strains of Bifidobacterium and Lactobacillus, produce a variety of immunomodulatory metabolites that play crucial roles in maintaining gut homeostasis and modulating the host immune system. These metabolites, which include short-chain fatty acids (SCFAs), bacteriocins, indole derivatives, and vitamins, have been shown to suppress inflammation, enhance microbial diversity, and improve intestinal barrier function by altering intestinal permeability and strengthening intercellular junctions [83]. Additionally, Lactobacillus and Bifidobacterium species are well-characterized for their production of SCFAs and vitamins, which enhance intestinal barrier integrity and promote anti-inflammatory responses [84]. Moreover, gut microbial-derived metabolites such as polyamines, choline-derived compounds, and secondary bile acids have been identified as immunoregulatory molecules that specifically affect adaptive immune responses, particularly T helper 17 and regulatory T cells, thereby influencing health and disease outcomes [85].
These strains produce immunomodulatory metabolites that positively impact the immune system in children by enhancing innate immunity, regulating pro-inflammatory cytokine expression, and preventing tissue damage from excessive inflammatory responses [86,87]. Specifically, Lactobacillus rhamnosus GG has been shown to induce beneficial Th1 immunomodulatory effects, helping manage conditions like cow’s milk allergy and atopic dermatitis by promoting IL-10 production [88].
Additionally, probiotics modulate the infant microbiota, induce immune mediator production, and influence cytokine production by intestinal cells, showcasing their ability to shape the immune response [86]. These effects are strain-specific and impact the immune system through various pathways, such as suppressing inflammation via the NF-κB pathway and enhancing phagocytic activity [89]. Furthermore, probiotics have been associated with reducing allergic reactions by downregulating Th2-related responses, inhibiting pro-inflammatory cytokine production, and modulating immune system components, ultimately promoting anti-inflammatory and immunomodulatory effects [90]. For example, lactic acid bacteria (LAB) are particularly effective in preventing allergic diseases like atopic eczema in infants by enhancing the body’s capacity to produce immune-enhancing cytokines such as interferon-gamma (IFN-γ) [91]. Specific probiotic strains also significantly increase the production of intestinal immunoglobulin A (IgA), which enhances mucosal immunity and protects against gastrointestinal infections [92]. Moreover, probiotics can influence systemic immunity by promoting the activity of natural killer (NK) cells and the differentiation of T-helper cells, both crucial for fighting infections and maintaining immune balance [93].
Lactobacillus gasseri TCI515, a probiotic strain, exemplifies this role by enhancing the expression of innate immunity-regulating genes and inhibiting pro-inflammatory cytokine gene expression, thereby improving innate immunity and preventing tissue damage from excessive inflammatory responses [87] Similarly, LAB fermentation of herbal medicines generates metabolites like exopolysaccharides, SCFAs, and bacteriocins, which have immunomodulatory properties and interact with the immune system, potentially boosting the innate immune response in children [94].
Overall, probiotic bacteria play a crucial role in producing metabolites that interact with and modulate the immune system, offering significant benefits for children’s health and well-being.
5.2. Anti-Inflammatory Metabolites
Probiotics have been studied for their potential to produce anti-inflammatory metabolites, such as short-chain fatty acids (SCFAs) like butyrate and acetate, which may modulate immune responses and reduce inflammation in children [95].However, the evidence is mixed and often varies depending on the specific strains used, the health status of the children, and the biomarkers measured. For example, a recent study involving a combination of Lactobacillus acidophilus and Bifidobacterium lactis demonstrated a significant reduction in inflammatory markers, such as MPIF-1 and MIP-3α, in children, suggesting an anti-inflammatory effect through immune modulation [96]. Similarly, another study found that synbiotic supplementation in overweight children led to decreased levels of inflammatory markers like tumor necrosis factor-α and interleukin-6, although these effects were associated with weight reduction rather than a direct anti-inflammatory action [97,98].
n children with various diseases, probiotics have shown potential in reducing systemic inflammation, particularly in conditions such as allergies, autoimmune diseases, and severe illnesses [98]. However, a study investigating the long-term effects of Lactobacillus paracasei supplementation during weaning found no significant impact on metabolic and inflammatory profiles at school age, suggesting that early probiotic intervention may not have lasting anti-inflammatory benefits [99]. Thus, the effectiveness of these biotics appears to be influenced by the health status of the children, with greater benefits observed in those with specific conditions, indicating that they may be more effective for certain groups.
Given that the molecular mechanisms underlying the anti-inflammatory effects of specific probiotic molecules are not well understood, a recent in vitro study demonstrated that secreted metabolites from Bifidobacterium infantis and Lactobacillus acidophilus exert anti-inflammatory effects in immature human enterocytes by modulating genes involved in immune response, cell survival, and NF-κB signaling. These metabolites reduce IL-6 and IL-8 production, suggesting their potential to mitigate inflammation in conditions such as necrotizing enterocolitis in children [100].
Additionally, another in vitro study evaluating the anti-inflammatory effects of biomolecules in probiotics commonly used in children showed that intestinal bacterial metabolites produced by Bifidobacterium animalis subsp. lactis LKM512 can suppress TNF-alpha production in J774.1 cells stimulated by lipopolysaccharide (LPS), suggesting that yogurt consumption containing this strain may help reduce inflammatory cytokines produced by macrophages [101].
In conclusion, while probiotics and their metabolites show promising potential as anti-inflammatory agents in children, further investigation is needed to fully elucidate the molecular mechanisms underlying these effects and to determine optimal conditions for their application. Such insights could guide the development of targeted probiotic therapies, particularly for conditions like necrotizing enterocolitis, allergies, and autoimmune diseases, ultimately improving therapeutic outcomes for pediatric populations. A summary of the role of probiotic metabolites in children’s health is shown in Figure 2.
5.3. Nutrient Absorption-Enhancing Metabolites
Probiotics have been shown to enhance nutrient absorption in children by producing beneficial metabolites that improve gut health and nutrient uptake. These metabolites include vitamins, minerals, and short-chain fatty acids (SCFAs), which play crucial roles in maintaining a healthy gut microbiome and improving overall health outcomes in children [4].
A recent study demonstrated that probiotics significantly increased the blood levels of vitamins and minerals such as Vitamin D, Vitamin A, calcium, zinc, and iron in children over a 10-week period. This finding suggests that probiotics can enhance the absorption of these essential nutrients, potentially improving the nutritional status and immunity of children [102]. Probiotics have also been shown to regulate lipid metabolism, which is crucial for nutrient absorption. In overweight or obese children, probiotics helped reduce levels of LDL cholesterol and leptin while increasing HDL cholesterol and adiponectin, indicating improved lipid profiles and metabolic health[103,104].
The administration of prebiotic-enhanced lipid-based nutrient supplements (LNSp) in undernourished infants led to a significant increase in the production of SCFAs such as acetate, butyrate, and propionate. These SCFAs are known to enhance gut health and nutrient absorption by promoting a beneficial gut microbiota composition. Similarly, the use of multi-strain probiotics in obese children increased the abundance of beneficial bacteria like Lactobacillus spp. and Bifidobacterium animalis, which are associated with improved SCFA production and lipid metabolism [105].
A recent study assessed the effectiveness of probiotics in improving health outcomes for children with severe acute malnutrition (SAM) in the Democratic Republic of Congo. The findings suggested that probiotics contributed to better health metrics, including weight gain, shorter recovery times, and overall nutritional improvement. The study concluded that incorporating probiotics into rehabilitation protocols could enhance the recovery of malnourished children [106].
In vitro approaches have shown that a mixture of Saccharomyces boulardii, Lactobacillus acidophilus, Lactobacillus rhamnosus, and Bifidobacterium breve, in combination with the enzyme amylase, disrupts pathogenic gastrointestinal biofilms, improving nutrient absorption by enhancing permeability and increasing the penetration of proteins and vitamins through intestinal cell monolayers [107].
While the evidence supports the role of probiotics in enhancing nutrient absorption through the production of beneficial metabolites, the effects can vary based on the probiotic strains used, the duration of supplementation, and the health status of the children. Further studies are needed to optimize probiotic interventions for different pediatric populations and to fully understand the mechanisms behind these benefits.
5.4. Gut Microbiota Balancing and Barrier-Enhancing Metabolites
A healthy microbiota involves maintaining an appropriate diversity and abundance of beneficial microorganisms that outnumber and outcompete potentially pathogenic or harmful ones [108]. When this delicate equilibrium is altered, the microbiota becomes imbalanced, a condition known as dysbiosis [109]. In children, dysbiosis is associated with alterations of gut function such as diarrhea [110], infant colic [111] or even autoimmune and atopic diseases such as asthma or rhinitis [112].
Balancing the gut microbiota is essential for optimal digestive health in children, and probiotic metabolites contribute significantly to this balance [113,114]. The bioactive molecules produced by probiotics help maintain a healthy gut environment by improving the balance of the gut microbiota [4]. By supporting a diverse and beneficial microbial community in the gut, these metabolites promote digestive health, prevent conditions such as diarrhea and colic, and contribute to overall well-being in children.
Short-chain fatty acids (SCFAs) are primary metabolites produced during the fermentation of dietary fibers by probiotic bacteria in the colon. These SCFAs, particularly butyrate, play a critical role in modulating the composition and activity of the gut microbiota [115]. Butyrate, for instance, is known for its ability to promote the growth of beneficial bacteria such as Faecalibacterium prausnitzii, a bacterium associated with anti-inflammatory properties [116]. At the same time, SCFAs help lower the pH of the gut environment, creating conditions unfavorable for pathogenic bacteria [117], thereby reducing the risk of gastrointestinal infections like diarrhea and colic in children.
In addition to promoting a healthy microbial balance, SCFAs have been found to enhance the production of mucin, a key component of the gut mucus layer that provides a protective barrier against pathogens [118]. By increasing mucin production, SCFAs help fortify the gut lining, reducing the likelihood of pathogens adhering to and invading the epithelial cells of the intestine. Furthermore, butyrate serves as a primary energy source for colonocytes [119,120], the cells lining the colon, thereby maintaining the integrity and function of the intestinal barrier [121].
Tight junctions are critical components that regulate the permeability of the gut barrier. Probiotic-derived metabolites like butyrate have been demonstrated to upregulate the expression of tight junction proteins, including claudin and occluding [122], which are essential for maintaining the integrity of the epithelial barrier. By enhancing these tight junctions, probiotics help prevent “leaky gut,” a condition characterized by increased intestinal permeability that allows toxins, microbes, and other harmful substances to enter the bloodstream, potentially triggering systemic inflammation and immune responses.
Additionally, certain metabolites produced by probiotics, such as lactate and hydrogen peroxide, have antimicrobial properties that contribute to maintaining a healthy gut barrier. Lactate, for example, can inhibit the growth of pathogenic bacteria by lowering the pH of the gut environment [123]. Meanwhile, hydrogen peroxide, produced by various microbial strains, has direct bactericidal effects against pathogens further protecting the integrity of the gut barrier [124].
6. Types and Mechanisms of Action of Probiotic Metabolites in Children
Probiotic metabolites, the bioactive compounds produced by beneficial microorganisms, play a crucial role in promoting and maintaining health in children. These metabolites, which include short-chain fatty acids (SCFAs), bacteriocins, exopolysaccharides (EPS), and vitamins, among others, exert a variety of beneficial effects on the host. Understanding the types and mechanisms of action of these probiotic metabolites is essential for leveraging their full therapeutic potential. This section delves into the various types of probiotic metabolites and explores the specific mechanisms through which they exert their health-promoting effects in children. A summary of the main metabolites produced by probiotic bacteria in children is shown in Figure 3.
6.1. Vitamins
Probiotic bacteria play a vital role in synthesizing essential vitamins that significantly impact the overall health and development of children. These beneficial bacteria, particularly certain strains of lactic acid bacteria (LAB) and bifidobacteria, can produce various vitamins necessary for multiple physiological processes, such as growth, development, and immune function [125]. In this section, we explore the types of vitamins produced by probiotic bacteria, their impact on children’s health, and the mechanisms through which these vitamins exert their beneficial effects.
Vitamin B Complex
Probiotic bacteria are known to synthesize several B vitamins, including B1 (thiamine), B2 (riboflavin), B3 (niacin), B5 (pantothenic acid), B6 (pyridoxine), B7 (biotin), B9 (folate), and B12 (cobalamin). These vitamins are critical for energy metabolism, DNA synthesis, red blood cell formation, and the proper functioning of the nervous system. Ensuring adequate levels of these vitamins is crucial for children’s growth and development, as they support numerous metabolic pathways and physiological processes [15,125].
Research has shown that specific probiotic strains, such as those within the genera Lactobacillus and Bifidobacterium, are highly effective at producing B vitamins. For example, Lactobacillus fermentum has been identified as a robust producer of folate and vitamin B12, achieving production levels of up to 801.79 μg/mL for folate [126].These strains can enhance the nutritional profile of various foods, such as fermented dairy products, making them an excellent source of B vitamins.
Oligosaccharides, which are prebiotic fibers that serve as a food source for probiotic bacteria, have been found to enhance the vitamin-producing capabilities of certain probiotic strains. Studies indicate that the presence of oligosaccharides can increase the synthesis of B vitamins by enhancing the bacteria’s properties, such as hydrophobicity, auto-aggregation, and biofilm formation [127].
Several probiotic strains, including Lactobacillus reuteri, Lactobacillus acidophilus, Streptococcus thermophilus, and Lactobacillus rhamnosus GG, are recognized for their ability to produce essential B-group vitamins, such as vitamin B12, riboflavin, folate, and thiamine, which are vital for numerous metabolic and physiological processes [128]. For example, L. reuteri strains produce corrinoids related to vitamin B12, while L. acidophilus strains increase riboflavin levels during fermentation, enhancing the nutritional value of foods like soymilk [129,130]. Additionally, S. thermophilus can boost the production of bioactive folate forms [131].
Leveraging these probiotic strains’ vitamin-producing capabilities could help develop fortified, vitamin-rich functional foods, especially beneficial for promoting children’s growth and development.
Vitamin K
Vitamin K, particularly in its K2 form (menaquinone), is another vital nutrient synthesized by probiotic bacteria [132]. This vitamin plays a crucial role in blood clotting, bone health, and cardiovascular function. The synthesis of vitamin K2 by gut bacteria is especially significant in children, who may have limited dietary intake of this nutrient due to selective eating habits or inadequate nutrition [133,134].
The probiotic bacterium Bacillus clausii has been demonstrated to produce vitamin K2, effectively correcting coagulation disorders in infants following antibiotic treatment by normalizing prothrombin levels [135]. Similarly, Lactococcus lactis is a known producer of vitamin K2, particularly in the context of fermented foods, where different strains produce varying amounts of the vitamin. Under specific cultivation conditions, these bacteria can enhance the delivery of vitamin K2 through extracellular vesicles [136].
Additionally, certain neonatal gut bacteria, such as Enterobacter agglomerans, Serratia marcescens, and Enterococcus faecium, have been identified as producers of menaquinones, a form of vitamin K2. This suggests that even in early life, the gut microbiota can contribute significantly to vitamin K production, which is essential for proper blood clotting processes [137]. However, antibiotic treatments in infants can disrupt the gut microbiota, leading to decreased vitamin K production. Studies have shown that administering probiotics like B. clausii can help restore normal prothrombin levels, highlighting the role of gut bacteria in vitamin K synthesis and coagulation [135].
There are far fewer bacteria known to produce vitamin K than those that synthesize B vitamins; however, as the gut microbiota of infants and children continues to be decoded, more microorganisms capable of synthesizing vitamin K are likely to be discovered, given its importance in pediatric health.
It is important to highlight that probiotics not only produce vitamins but also enhance their absorption and bioavailability. For instance, a study involving children aged 8–13 showed that probiotic supplementation significantly increased blood levels of vitamins D and A compared to a placebo group over a 10-week period, suggesting that probiotics may improve the absorption and serum concentrations of essential vitamins [102]. Another study reported improvements in gut health markers, such as increased bifidobacteria and reduced inflammatory markers, in children receiving prebiotics and vitamin supplements, demonstrating the potential of probiotics to enhance both gut health and vitamin status [138].
The ability of probiotics to synthesize and enhance the absorption of vitamins has broad implications for child health. By ensuring the production of essential nutrients, such as B vitamins and vitamin K, probiotics support various physiological processes, including energy metabolism, immune response, and bone mineralization. Furthermore, vitamins produced by commensal bacteria may influence immune responses, suggesting roles beyond basic nutrition, such as regulating gene expression and enhancing nutrient absorption [15,31]. A summary of the probiotic bacteria involved in the synthesis of vitamins is described in Table 1.
6.2. Short-Chain Fatty Acids (SCFAs)
Short-chain fatty acids (SCFAs) such as acetate, propionate, and butyrate are key metabolites that play a vital role in supporting gut health in children. SCFAs are essential for maintaining a healthy digestive system [140]. One of their primary functions is to nourish the cells lining the intestines, known as colonocytes [119], which are crucial for forming a robust gut barrier that prevents harmful substances from entering the bloodstream [119]. Additionally, SCFAs are critical for the development, regulation, and maturation of the immune system [141,142]. Notably, butyrate, a specific SCFA, has demonstrated anti-inflammatory properties [140], helping to protect against conditions such as colic, gastrointestinal infections, and inflammatory bowel disease [143].
SCFAs also contribute to maintaining an optimal gut environment by lowering the pH [144,145], which supports the growth of beneficial bacteria while inhibiting gut pathogens [146]. Furthermore, these molecules enhance the gut barrier by promoting the production of mucins, substances that protect the gut lining and prevent the adherence and invasion of pathogens [147].
Certain probiotic strains, such as Lactobacillus and Bifidobacterium, are known to produce SCFAs in the gut [31,148,149,150]. These beneficial bacteria are commonly found in fermented foods like yogurt or in dietary supplements specifically formulated for children. When children consume foods rich in probiotics, these bacteria proliferate in the gut and produce SCFAs by fermenting dietary fibers present in fruits, vegetables, and whole grains [151,152].
7. Antimicrobial Peptides
Antimicrobial peptides (AMPs) are small molecules that serve as natural antibiotics produced by the body and certain probiotic bacteria [153,154]. AMPs represent a promising avenue for enhancing child health by modulating the gut microbiota and providing a natural defense against pathogens [154]. These bioactive molecules, which include bacteriocins and other peptide-based antimicrobials, have been shown to selectively inhibit harmful bacteria while promoting the growth of beneficial microbes [155]. Furthermore, unlike conventional antibiotics, AMPs have a lower propensity for resistance development, making them a safer alternative for managing pediatric infections and promoting overall gut health [156,157,158].
Although there is currently limited direct evidence for the use of probiotic-derived antimicrobial peptides (AMPs) specifically in pediatric health, several lines of research suggest their potential as novel therapeutic agents for children. Extensive studies have demonstrated the broader benefits of probiotic-derived AMPs for human health, particularly their antimicrobial, immunomodulatory, and microbiota-regulating properties [155,159,160,161]. Additionally, AMPs from other sources have proven effective in managing pediatric infections, and clinical trials have highlighted cationic antimicrobial peptides as promising alternatives for treating infections in neonates and children, especially in cases involving antibiotic-resistant pathogens [162]. Further research into the use of probiotic-derived AMPs could open new pathways for developing innovative, safe, and effective treatments to manage infections and enhance health outcomes in children.
8. Enzymes
The enzymes produced by probiotics hold significant potential in improving children’s health by supporting essential digestive functions and enhancing nutrient absorption [163]. These beneficial microorganisms produce enzymes that break down complex nutrients, making them more accessible for a child’s developing digestive system. As children’s gut microbiota continues to evolve, the enzymatic activity provided by probiotics plays a pivotal role in promoting a balanced gut environment, which is crucial for immune function, growth, and overall well-being [164].
One of these enzymes derived from probiotics is β-glucosidase, an enzyme important for breaking down complex carbohydrates into simpler sugars, which can enhance the probiotic’s effectiveness in the gut [165]. Bifidobacterium species, early colonizers of the infant gut, produce β-galactosidase to metabolize milk-based diets. This enzyme cleaves the glycosidic bond in lactose through hydrolysis, producing the monosaccharides glucose and galactose, which are essential for energy production and growth in infants [166]. In addition to hydrolysis, β-galactosidase can transfer galactosyl units to other sugar molecules through transgalactosylation. This activity forms galactooligosaccharides (GOSs), which are beneficial prebiotics that promote the growth of healthy gut microbiota [166,167,168]. In infants with nutritional disorders such as celiac disease and cystic fibrosis, β-galactosidase activity can be affected. For example, lactase activity is significantly reduced in celiac disease, while hetero-β-galactosidase activity remains relatively stable [169].
Similarly, lacto-N-biosidase (LNBase) plays a crucial role in the digestion of human milk oligosaccharides (HMOs). This enzyme, primarily found in Bifidobacterium spp., facilitates the breakdown of complex sugars into simpler forms that can be utilized by the infant’s gut microbiota [164]. For instance, LNBase from Bifidobacterium bifidum (LnbB) is essential for the degradation of HMOs, specifically lacto-N-tetraose, into lacto-N-biose I and lactose, which is vital for the early-life microbiota in infants [170].
LNBase operates via a substrate-assisted catalytic mechanism, with a unique metabolic pathway specific to lacto-N-biose I, a major core structure in HMOs [171]. Its activity is modulated by specific amino acids, such as His263, which plays a critical role in the catalytic process by altering the pKa of the acid/base residue [170]. The stability of LNBase during digestion and its ability to modulate gut microbiota composition, increasing the abundance of beneficial bacteria like B. bifidum, underscores its potential in alleviating infant food allergies and promoting overall gut health [164].
There are other enzymes from probiotics that contribute significantly to the nutrition and health of infants. For example, proteases are important for breaking down proteins into peptides and amino acids, facilitating better protein absorption and utilization in the body. This is particularly beneficial for infants and children, who require adequate protein for growth and development. Some proteases, particularly those from the genus Lactobacillus, have also shown antimicrobial properties, protecting infants against infections by targeting the cell walls of pathogenic bacteria [172]. Bile salt hydrolase (BSH), also known as choloylglycine hydrolase, is another enzyme commonly found in probiotic bacteria like Bifidobacterium or Lactobacillus species [173]. This enzyme plays a crucial role in the deconjugation of bile salts [174,175]. Nuñez et al. (2022) provided evidence of the presence of BSH genes in the early infant microbiota, suggesting that this metabolic activity may occur as early as one day after birth [174,175].
The enzymatic activities of probiotics contribute significantly to infant and child health by enhancing digestion, nutrient absorption, and gut microbiota composition. These findings highlight the importance of continued research into probiotic-derived enzymes as valuable tools for improving pediatric health outcomes.
9. Exopolysaccharides (EPSs)
EPSs are complex carbohydrate polymers secreted by probiotic bacteria such as Lactobacillus, Bifidobacterium, Streptococcus, Weissella, during metabolic processes [176]. Structurally, exopolysaccharides consist of repeating units of glucose, galactose, mannose and rhamnose, which can form homo or heteropolysaccharides, often linked by glycosidic bonds [177]. These polysaccharides can either be covalently anchored to the cell surface, forming capsular polysaccharides [178], or be secreted into the extracellular environment, leading to the formation of a mucilaginous layer. This latter form plays a critical role in the development of bacterial biofilms [178,179]. These types of polysaccharides can vary widely in composition, branching, and molecular weight, contributing to their diverse functional properties and activities [180].
One of the most notable is their bifidogenic activity, which refers to their ability to selectively stimulate the growth of specific members of the infant gut microbiota, specifically of Bifidobacterium species [181]. In infants, Bifidobacterium typically accounts for about 90% of intestinal bacteria [182].
The bifidogenic effects of EPSs arise from their complex carbohydrate structures, which are resistant to digestion in the upper gastrointestinal tract. Upon reaching the colon, these EPSs serve as prebiotics. Lv et al. (2024) isolated and purified EPSs from Bifidobacterium animalis subsp. Lactis SF (SF-EPS) from the feces sample of a healthy infant), and their probiotic potential was evaluated in vitro. SF-EPS regulated the gut microbiota by increasing the relative abundances of Faecalibacterium, Anaerostipes, and Bifidobacterium, while reducing the abundance of Enterobacter and Klebsiella. Furthermore, SF-EPS enhanced the production of SCFAs by intestinal microorganisms. These findings suggest that SF-EPS may serve as a potential prebiotic for use in functional foods [14].
Exopolysaccharides also exhibit potent biological activities, including antioxidant properties, free radical scavenging, and the reduction of oxidative stress [14,183]. Tarique et al. (2024) showed the antioxidant potential of EPSs from Enterococcus faecium and S. thermophilus. This potential could be due to the different sugars and their arrangements which can affect the ability of EPSs to interact with and neutralize free radicals [26].
EPS also have immunoregulatory properties. EPS from Bifidobacterium longum subsp. infantis E4 demonstrated significant immunomodulatory and anti-inflammatory effects in vitro. These EPS enhanced macrophage activity and reduced inflammatory markers, indicating potential benefits for immune health in infants [184]. Additionally, some studies have found that EPSs from probiotics have other properties such as: antitumor activity [185,186,187], antibacterial activity [188] antiviral protection [189], and lipid regulation potential [190,191]. EPSs also enhance adherence and subsequent colonization of microflora on host cells [14].
In summary, the combination of different biological activities makes EPSs promising candidates for functional foods and therapeutic agents aimed at improving children’s health.
10. Neurotransmitters
10.1. Gamma-Aminobutyric Acid (GABA)
GABA is a crucial neurotransmitter for children, playing a key role in brain development, emotional regulation, and cognitive functions like learning and memory. It helps maintain a balance between excitatory and inhibitory signals in the brain, which is important for reducing anxiety, promoting restful sleep, and managing stress [192,193,194]. Adequate GABA levels support healthy neural circuit formation, contribute to emotional stability, and help children with behavioral control, making it essential for overall mental and physical well-being [195,196]. Interestingly, these levels have been found to increase with age [197], and this difference in GABA levels seems to be related to how fast children can learn. Experimental results indicate children show more flexible GABA-related inhibitory processing compared to adults, allowing for quicker adaptation to stabilize learning [194].
While Lactobacillaceae species are recognized as primary producers of GABA, Bifidobacterium species have been identified as the most efficient GABA-producers [195]. Notably, the presence of gad genes responsible for GABA synthesis are also found in other probiotic strains [198,199]. The production of GABA by these microorganisms has suggested a link between the gut microbiota and neurological health [199]. GABA is produced by the enzyme glutamate decarboxylase (GAD), which requires pyridoxal-5′-phosphate (PLP) and works through the irreversible α-decarboxylation of l-glutamate, consuming one cytoplasmic proton [200]. In bacteria, the import of glutamate and the export of GABA typically occur simultaneously through a specific glutamate/gamma-aminobutyrate antiporter, with the gadB and gadC genes encoding these enzymes [201].
Bifidobacterium species such as Bifidobacterium adolescentis, Bifidobacterium dentium, and Bifidobacterium longum have demonstrated the capacity to synthesize GABA through the decarboxylation of glutamate by GAD enzymes. This process not only helps in maintaining the gut’s acid-base balance but also contributes to the pool of bioactive GABA within the host [198,202,203,204,205].
GABA-producing probiotic strains significantly impact the gut-brain axis, a communication network between the gastrointestinal tract and the central nervous system, involving the gut microbiota, immune system, enteric nervous system (ENS), and central nervous system [206]. GABA can modulate ENS activity, affecting gut motility, secretion, and blood flow [207], and may influence the central nervous system via the vagus nerve, impacting neuropsychiatric conditions [208,209]. Increased GABA levels in the gut have been linked to reduced stress and anxiety-like behaviors [193], with specific bacteria like Bacteroides and GABA-producing Bifidobacterium strains playing roles in mental health and reducing systemic inflammation associated with mood disorders and neurodegenerative diseases [199,209].
Research has shown that the maternal gut microbiome can promote fetal brain development, affect brain connectivity, and influence cognitive and behavioral development in offspring, suggesting that a healthy maternal microbiome rich in GABA-producing bacteria can positively influence the infant’s neurological outcomes [210].
Previous studies demonstrated that the gut microbiota diversity in children with Autism Spectrum Disorder (ASD) underwent significant changes, with alterations in Bifidobacterium being linked to the severity of ASD [211]. Infants at higher risk for ASD have a decreased abundance of Bifidobacterium and an increased abundance of Clostridium and Klebsiella compared to those at lower risk. Also, fecal GABA levels were lower in infants with a higher likelihood of ASD, with GABA levels showing a positive correlation with Bifidobacterium [212].
Analyses on children with attention deficit hyperactivity disorder (ADHD) have found lower levels of GABA and reduced presence of lactic acid bacteria. These bacteria are known to be involved in the production of GABA, suggesting that the reduction of lactic acid bacteria in the gut of infants with ADHD could be associated with lower GABA levels [213,214].
Furthermore, clinical trials have suggested that probiotics containing GABA-producing Bifidobacterium or Lactobacillaceae species can be effective in treating gastrointestinal issues and enhancing overall mental health in children. For example, trials involving the administration of GABA-producing species to very low-birth-weight preterm infants demonstrated improved gut health and reduced incidences of necrotizing enterocolitis, a severe intestinal condition [215,216,217].
10.2. Other Neurotransmitters Produced by Probiotics
In addition to GABA, probiotic microorganisms are gaining recognition for their capacity to produce various neurotransmitters, which can significantly influence host health via the gut-brain axis. A well-documented example is serotonin (5-HT), a neurotransmitter crucial for regulating mood, appetite, and sleep. Certain probiotic strains, such as Enterococcus and Streptococcus, have been shown to synthesize serotonin, suggesting their potential impact on emotional and psychological well-being [218].
Dopamine, another essential neurotransmitter associated with reward and motivation, is produced by probiotic strains like Bacillus and Lactobacillus species [219]. The production of dopamine by these probiotics could affect neurological functions and behaviors, highlighting a possible route for influencing the host’s nervous system. Similarly, Lactobacillus plantarum is known to produce acetylcholine, which plays a critical role in learning, memory, and muscle activation [220]. The presence of acetylcholine in the gut underscores the complex communication pathways of the gut-brain axis.
Probiotics like Escherichia coli and Bacillus subtilis have also demonstrated the ability to synthesize norepinephrine, a neurotransmitter involved in alertness and the body’s “fight or flight” response [221]. This ability suggests that these microorganisms could help modulate stress and mood responses. Additionally, Lactobacillus reuteri is known to produce histamine, which is critical for immune responses and gut motility [222]. The production of glutamate, an excitatory neurotransmitter important for synaptic plasticity, has been observed in strains of Lactobacillus and Bifidobacterium [223].
Overall, the capacity of probiotics to synthesize these neurotransmitters supports their potential role in managing neuropsychiatric and gastrointestinal disorders by modulating the gut-brain axis, providing a promising avenue for therapeutic intervention.
11. Bioactive Postbiotic Fractions
Postbiotics refer to bioactive compounds produced by microorganisms during fermentation, encompassing microbial cells, cell constituents, and metabolites. According to the International Scientific Association for Probiotics and Prebiotics (ISAPP), postbiotics are defined as “a preparation of inanimate microorganisms and/or their components that confers a health benefit on the host” [224]. Postbiotics are key mediators of microbiota-host interactions. Unlike probiotics, which are live microorganisms, postbiotics do not contain live bacteria but consist of beneficial byproducts released during the microorganisms’ life cycle [225,226]. Postbiotics can be classified based on their composition, including lipids (e.g., butyrate, propionate), proteins (e.g., lactocepin), carbohydrates (e.g., polysaccharides, teichoic acids), vitamins, organic acids (e.g., lactic acid), and complex molecules (e.g., lipoteichoic acids) [225,226,227]. Additionally, they can be categorized by their physiological functions, such as immunomodulatory, anti-inflammatory, hypocholesterolemic, anti-obesity, antihypertensive, anti-proliferative, and antioxidant effects (Figure 4) [228,229]. These compounds are widely present in foods such as yogurt, kefir, and pickled vegetables, and can also be intentionally applied in functional foods such as infant formulas [225,226,230].
The significance of postbiotics lies in their potential health benefits and greater stability compared to probiotics. Because postbiotics lack live bacteria, they are more stable and have a longer shelf life, making them easier to store and use in therapeutic and nutritional applications [231]. The unique structure of postbiotics can exert a range of beneficial effects on the host through diverse cellular and molecular mechanisms. Postbiotics play a crucial role in children’s health by supporting gut balance, boosting immunity, and promoting digestive health. They help reduce inflammation, enhance nutrient absorption, and protect against infections, contributing to overall well-being in growing children [225,232]. Additionally, the use of postbiotics is associated with fewer safety concerns than probiotics, as there is no risk of infection from live bacteria. This makes them particularly suitable for vulnerable populations, including infants, the elderly, and immunocompromised individuals [225,226,233].
Postbiotics exhibit significant immunomodulatory and anti-inflammatory effects, playing a critical role in enhancing both innate and adaptive immune responses. These non-living microbial products and metabolic byproducts influence immune pathways, modulating cytokine production and regulating immune cell activity. One of the key mechanisms by which postbiotics exert immunomodulatory effects is through the activation of immune receptors such as Toll-like receptors (TLRs). For instance, heat-inactivated Lactobacillus casei Zhang has been shown to enhance macrophage-mediated innate immunity by increasing the transcription of TLRs (TLR2, TLR3, TLR4, and TLR9) and stimulating pro-inflammatory cytokines, which strengthen the body’s defense against infections [234]. Moreover, postbiotics derived from Lactobacillus gasseri TMC0356 have demonstrated a more potent effect on immune activity than their probiotic counterparts, inducing higher levels of interleukin-12 (IL-12) in macrophages [235]. Similarly, heat-inactivated Lactobacillus acidophilus A2 and Lactobacillus gasseri A5 have been found to enhance Th1-mediated immunity by promoting the production of IL-10 and IL-12 p70 and increasing interferon-gamma (IFN-γ) in splenocytes, further demonstrating the capacity of postbiotics to modulate immune responses [236].
The anti-inflammatory properties of postbiotics are equally noteworthy. By regulating cytokine production, postbiotics can help reduce inflammatory responses. For example, supernatants of Faecalibacterium prausnitzii have been shown to alleviate colitis in mice by increasing the production of the anti-inflammatory cytokine IL-10 while reducing the pro-inflammatory cytokine IL-12, likely through the inhibition of NF-κB activation [237]. Additionally, Lactobacillus paracasei B21060 postbiotics have demonstrated protective effects against inflammation caused by Salmonella in human colon tissues [238]. Another example of the immunomodulatory action of postbiotics involves Lactobacillus reuteri 17938, which promotes the production of the anti-inflammatory cytokine IL-10 in dendritic cells, leading to an enhanced regulatory T-cell response [239]. Additionally, culture supernatants from Lactobacillus casei DG were shown to mitigate inflammation in ileal and colonic cultures of post-infectious bowel syndrome patients, suggesting postbiotics can target localized inflammation in the gut [240].
Recent research has increasingly focused on the role of fermented infant formulas and their postbiotic components in enhancing infant health, particularly in reducing the severity of gastrointestinal and allergic conditions. A study by Béghin et al. (2021) explored the effects of a fermented infant formula (FF) with Bifidobacterium breve C50 and Streptococcus thermophilus O65 combined with prebiotic oligosaccharides on gut microbiota composition and immune function in healthy term infants. The findings indicated that this combination led to a gut microbiota composition and metabolic activity more similar to that of breastfed infants, with a significant increase in secretory IgA (SIgA) levels, highlighting its potential to enhance early immune defense [241]. In contrast, a study by Thibault et al. (2004) focused on the impact of an FF with the same probiotic bacteria on the incidence and severity of acute diarrhea in healthy infants aged 4 to 6 months. While the formula did not significantly alter the incidence or duration of diarrhea, it did reduce the severity of episodes, likely due to the bifidogenic and immunomodulatory properties of the fermentation products [242].
Additionally, Morisset et al. (2011) found that a non-hydrolyzed, fermented infant formula containing heat-killed Bifidobacterium breve C50 and Streptococcus thermophilus O65 significantly reduced the incidence of digestive and respiratory events in infants at high risk of allergy. Although it did not decrease the overall incidence of cow’s milk allergy (CMA), it lowered the proportion of positive skin prick tests for cow’s milk and the occurrence of potentially allergic adverse events (PAAEs) related to digestion and respiration. These results suggest that the postbiotic components of the formula may play a role in improving oral tolerance and reducing allergy-like symptoms in high-risk infants [243]. Similarly, Nocerino et al. (2017) and Corsello et al. (2017) investigated the effects of postbiotics, specifically L. paracasei fermented cow’s milk, in preventing common infectious diseases (CIDs) in children. Both trials showed that dietary supplementation with postbiotics significantly reduced the incidence of acute gastroenteritis and upper respiratory tract infections compared to a placebo. Children consuming the fermented milk had fewer episodes of illness, reduced medication use, and missed fewer days of school or daycare [244,245].
In summary, postbiotics offer a range of immunomodulatory and anti-inflammatory benefits, making them promising therapeutic agents. Their ability to regulate immune responses, reduce inflammation, and support overall immune health highlights their potential in managing conditions such as inflammatory bowel disease and other immune-related disorders. By leveraging postbiotics, health professionals can modulate immune responses without relying on live probiotics, offering an alternative approach for those who may be sensitive to live bacterial strains [225,246,247].
12. Clinical Applications and Health Implications
Since a comprehensive analysis published by Dronkers et al. in 2020 on 1341 studies retrieved from the ClinicalTrials.gov database using the search term “probiotics”, showed that 56% were conducted in the USA or Europe, and with around 100 studies registered annually since 2010 with a tendency to increase in numbers over the most recent years [248]. The vast majority were interventional (95.6%) but almost half in healthy participants (43.8%), and only 31.8% of those studies were in children (birth-17 years old). Of 852 study registrations which could be analyzed, Lactobacillus rhamnosus GG (LGG) was the probiotic strain most frequently registered (146 studies) followed by Bifidobacterium animalis ssp. lactis BB12 with 55 studies, while VSL#3, a consortium of 3 different Bifidobacteria, 4 Lactobacillus, and 1 Streptococcus thermophilus strains, was the most registered multispecies preparation in 74 studies.
Over the last five years, ClinicalTrials.gov has listed 730 studies, of which 167 are in children and 157 are interventional studies in children. Two studies were withdrawn (one in colic and other in peanut allergy); five were prematurely terminated (two in respiratory infections, one in atopic dermatitis, one in surgical procedures and one in healthy children), and 48 are completed addressing a range of conditions including allergic diseases, respiratory infections, cystic fibrosis, obesity, ADHD, gastrointestinal diseases, and on Autism Spectrum Disorder (ASD). This indicates that 84 studies (53.5%) are registered but still ongoing. Despite this focus, the volume of published clinical trials in children has been decreasing according to the Pubmed.gov database, from 69 publications in 2020 and 2021, to 55 articles in 2022 and just 46 in the most recent year.
In recent years, the role of probiotics in managing ASD has been studied, as potential benefits of probiotics were suggested by exploring the gut-brain axis, offering hope for improved management strategies [249]. However, a meta-analysis published in the Journal of Medical Microbiology in 2022, has shown no significant benefit of probiotics for ASD treatment [250]. This study highlighted that the lack of standardization in trials—such as variations in strains, dosages, and protocols—complicates the ability to conduct comprehensive meta-analyses with robust findings.
Current Practice Guidelines
The clinical application of probiotics is still under discrepancies between the existing guidelines, particularly for acute gastroenteritis in children, and is summarize in Table 2:
American Academy of Pediatrics (AAP): Probiotics such as L. rhamnosus GG and S. boulardii are recommended for reducing the duration of acute gastroenteritis in children. However, routine use in healthy children is not broadly advised. Probiotics may help prevent antibiotic-associated diarrhea and necrotizing enterocolitis (NEC) in preterm infants, though caution is advised in immunocompromised patients [251].
European Society for Pediatric Gastroenterology, Hepatology and Nutrition (ESPGHAN): Supports probiotics for conditions like acute gastroenteritis and prevention of NEC in preterm infants but stresses strain-specific efficacy and proper quality control [252].
British Society of Pediatric Gastroenterology, Hepatology and Nutrition (BSPGHAN): Aligns with ESPGHAN, emphasizing selective use of probiotics for gastrointestinal issues and the importance of strain-specific evidence in children [251].
For instance, the American Gastroenterological Association (AGA) is against the use of probiotics [251,253].These inconsistencies arise from several factors, including variability in literature review processes—such as incomplete searches or differing inclusion criteria—which can lead to conflicting recommendations. Additionally, some guidelines may place more weight on single randomized controlled trials (RCTs), while others incorporate a broader evidence base [254].
Differences in study populations, including socioeconomic levels, medical issues, and geographical features, also influence these recommendations. Potential conflicts of interest (COI) bias from industry sponsorship may further impact guideline development. Lastly, the evolving nature of research means that guidelines may not always reflect the most current evidence. These factors highlight the need for more standardized approaches and regular updates to clinical guidelines to support better decision-making and foster ongoing research [254].

Table 2.
Current Guidelines on Probiotics for most common conditions (modified from [255]).
Table 2.
Current Guidelines on Probiotics for most common conditions (modified from [255]).
| Disorder | Probiotic Strain | Recommended Dose | Evidence Level ** |
|---|---|---|---|
| Acute gastroenteritis [Reduced the risk of diarrhea lasting ≥ 48 h; reduced the mean duration of diarrhea [256]] | Probiotics as a general group | N/A | 1 |
| L. rhamnosus GG [257] | ≥1010 cfu/day, for 5–7 days | 1 | |
| S. boulardii * [258] | 250–750 mg/day, for 5–7 days | 1 | |
| L. reuteri DSM 17,938 [259] | 1 × 108 to 4 × 108 cfu/day, for 5 days | 1 | |
|
L. rhamnosus 19070-2 & L. reuteri DSM 12,246 [260] |
2 × 1010 cfu for each strain/day, for 5 days | 1 | |
| B. lactis B94 [261] | 5 × 1010 cfu once daily, for 5 days | 3 | |
| L. paracasei B21060 [262] | 2.5 × 109 cfu, twice daily, for 5 days | 3 | |
| L. rhamnosus strains 573L/1; 573L/2; 573L/3 [263] | 1.2 × 1010 cfu, twice daily, for 5 days | 3 | |
| L. delbrueckii var. bulgaricus, L. acidophilus, S thermophilus, B. bifidum (LMG- P17550, LMG-P 17549, LMG-P 17503, LMG-P 17500) [264]. | 109 cfu, 109 cfu, 109 cfu, 5 × 108 cfu/dose, for 5 days | 3 | |
| B. lactis Bi-07, L. rhamnosus HN001, and L. acidophilus NCFM [265] | Then 1 × 1010 cfu once a day, for the duration of diarrhea plus 7 days | 3 | |
| Prevention of AAD (Reduced risk of AAD [266]) | Probiotics as a general group | N/A | 1 |
| S. boulardii * [267] | ≥5 billion cfu per day, for the duration of antibiotic treatment | 1 | |
| L. rhamnosus GG [268] | ≥5 billion cfu per day, for the duration of antibiotic treatment | 1 | |
| Multispecies probiotic (Bifidobacterium bifidum W23, B. lactis W51, Lactobacillus acidophilus W37, Lactobacillus acidophilus W55, Lacticaseibacillus paracasei W20, Lactoplantibacillus plantarum W62, Lacticaseibacillus rhamnosus W71, and Ligilactobacillus salivarius W24] [269] | 10 billion cfu per day, for the duration of antibiotic treatment and for 7 days after | 3 | |
| L. rhamnosus (strains E/N, Oxy, and Pen) [270] | 2 × 1010 cfu, twice daily, for the duration of antibiotic treatment | 3 | |
| Prevention of C. difficile diarrhea | S. boulardii * [267] | 250–500 mg | 1 |
| Prevention of nosocomial diarrhea | L. rhamnosus GG [271,272] | At least 109 cfu/day, for the duration of the hospital stay | 1 |
| Prevention of necrotizing enterocolitis [273,274,275] | Systematic reviews and meta-analyses (> 10,000 neonates) of RCTs | 1 | |
| L. rhamnosus GG [276] | From 1 × 109 to 6 × 109 cfu | 1 | |
| B. infantis BB-02, B. lactis BB-12, and S. thermophilus TH-4 [276] | 3.0 to 3.5 × 108 cfu (of each strain) | 1 | |
| B. animalis subsp. lactis Bb-12 or B94 [276] | 5 × 109 cfu | 3 | |
|
L. reuteri ATCC 55730* or DSM 17938 *this strain is no longer available. [276,277] |
1 × 108 cfu (various regimens) | 1 | |
| B. longum subsp. infantis ATCC 15,697 + L. acidophilus ATCC 4356 [277,278] | 125 mg/kg/dose twice daily with breast milk until discharge | 3 | |
| B. longum subsp. longum 35,624 + L. rhamnosus GG [278] | 5 × 108 cfu and 5 × 108 cfu, respectively | 3 | |
|
Helicobacter pylori infection [279,280,281,282,283] |
Probiotics as a general group | 1 | |
| S. boulardii* [284,285] | 500 mg | 1 | |
| Infantile colic [286,287,288,289,290,291] | Probiotics as a general group | N/A | 1 |
| L. reuteri DSM 17,938 [292,293] | 108 cfu/day for at least 21 days | 1 | |
| B. lactis Bb12 [294,295] | 1 × 109 cfu/day, for 21–28 days | 2 | |
| L. rhamnosus 19070-2 and L. reuteri 12,246 [296] | 250 × 10⁶ cfu, respectively, for 28 days | 3 | |
| L. paracasei DSM 24733, L. plantarum DSM 24730, L. acidophilus DSM 24735, L. delbrueckii subsp. bulgaricus DSM 24734), B. longum DSM 24736, B. breve DSM 24732, and B. infantis DSM 24737, and S. thermophilus DSM 24,731 [297] | 5 billion cfu, for 21 days | 3 | |
| Infantile colic—prevention | L. reuteri DSM 17,938 [298] | 108 cfu/day, to newborns each day for 90 days | 1 |
| Functional abdominal pain/IBS | L. reuteri DSM 17,938 [299,300] | 108 cfu to 2 × 108 cfu/day | 1 |
| L. rhamnosus GG [299,301] | 109 cfu to 3 × 109 cfu twice daily | 1 | |
| Ulcerative colitis [302] | Probiotics as a group | N/A | 1 |
| A mixture of 8 strains (L. paracasei DSM 24733, L. plantarum DSM 24730, L. acidophilus DSM 24735, L. delbrueckii subsp. bulgaricus DSM 24734, B. longum DSM 24736, B. infantis DSM 24737, B. breve DSM 24732, and S. thermophilus DSM 247), as adjuvant therapy or in those intolerant to 5-ASA [303] | Daily dosages: 4–6 y (17–23 kg): 450 billion; 7–9 y (24–33 kg): 900 billion; 11–14 y (34–53 kg): 1350 billion; 15–17 y (54–66 kg): 1800 billion. |
3 | |
| Pouchitis | A mixture of 8 strains (L. paracasei DSM 24733, L. plantarum DSM 24730, L. acidophilus DSM 24735, L. delbrueckii subsp. bulgaricus DSM 24734, B. longum DSM 24736, B. infantis DSM 24737, B. breve DSM 24732, and S. thermophilus DSM 247) [304,305] | Daily dosages: 4–6 y (17–23 kg): 450 billion; 7–9 y (24–33 kg): 900 billion; 11–14 y (34–53 kg): 1350 billion; 15–17 y (54–66 kg): 1800 billion. |
3 |
* Most studies with the strain S. boulardii CNCM I-745; ** (1) Systematic review or meta-analysis of randomized control trials; (2) Randomized control. trials; (3) Quasi-experimental studies; (4) Non-experimental studies.
13. Challenges of Using Bioactive Molecules from Probiotics for Pediatric Diseases
As has been explained in other sections, the use of bioactive molecules derived from probiotics in pediatric diseases offer considerable potential to enhance children’s health and well-being. However, there are several challenges that still need to be addressed. Overcoming these challenges is essential to fully unlock the therapeutic benefits of probiotics.
The use of probiotics in children, particularly in the most vulnerable such as preterm infants, immunocompromised, or those with underlying health conditions, is still limited by safety concerns. Although many studies confirm the safety of probiotics, there have been isolated cases of sepsis or gastrointestinal mucormycosis linked to contaminated probiotic products, highlighting the need for stringent quality control measures during manufacturing [306]. Furthermore, the reported risk of cross-colonization with probiotic strains in neonatal intensive care units (NICUs) suggested that despite probiotics offered potential benefits, they must be administered cautiously, supported by comprehensive safety assessments [306]. The Agency for Healthcare Research and Quality reviewed 622 studies on probiotic safety and found that while there is no evidence of increased risk from randomized controlled trials, the literature lacks systematic reporting on adverse events, making it difficult to assess rare risks with confidence [307]. Therefore, it is crucial to implement standardized protocols for safety outcomes in clinical trials, ensuring comprehensive and transparent data collection. Enhanced guidelines would enable a more accurate assessment of the benefits/risks associated with probiotics use and support informed decision-making in both research and clinical practice.
Translating promising findings from preclinical and clinical studies into clinical guidelines is an imminent challenge. Research conducted in diverse geographic regions may not be generalizable due to variations in genetics, diet, sanitation, and endemic enteropathogens [308]. Systematic reviews, have failed to recommend yet specific strains due to inconsistency in results and lack of standardized protocols regarding species, dosage, and administration duration [248].Variations in study designs, including differences in probiotic strains, dosages, and treatment durations, have led to inconsistent results and have hindered the development of clear guidelines. Moreover, the absence of standardized protocols and regulatory frameworks complicates the approval and adoption of probiotics for pediatric use. To overcome these obstacles, more well-designed randomized controlled trials (RCTs) and meta-analyses are needed to establish evidence-based guidelines for probiotic use in children.
Clinical trials in pediatric populations present additional ethical challenges and special regulatory compliance. Moreover, differences in diet, genetics, and environmental factors across different populations limits generalization of the research findings. Regulatory agencies should develop frameworks that promote innovation while ensuring safety and efficacy, facilitating global standardization in probiotic research and applications.
14. Future Perspectives and Opportunities
Successful integration of probiotic-derived bioactive molecules in the pediatric population is also an opportunity to enhance treatment options and potentially improve outcomes. By addressing the previously discussed challenges there are substantial strides toward safer, more effective, and widely accepted probiotic-based therapies for pediatric populations.
14.1. Fostering of Clinical Research
Current situation of clinical trials with probiotics in the pediatric population reveals a growing interest in the field, while the decrease in the number of publications might indicate stronger review criteria on the available data. Future allocation of resources, multidisciplinary teams, and active collaboration are essential to strengthen clinical research oriented to answer relevant and important questions.
14.2. Evidence-Based Clinical Guidelines
The current data available and its variability offers an opportunity to foster broader and global collaborations among health care providers, their institutions, research-oriented organizations, academia, and industry. Therefore, creating stronger evidence for the use of probiotics in children should be a must. Development of updated guidelines, based on evidence, that consider regional differences in diet, genetics, and environmental factors will follow the idea of personalized treatments.
14.3. Strengthening Ethical Frameworks for Pediatric Research
Pediatric research presents unique opportunities, although their ethical challenges, which can be addressed by developing innovative frameworks balancing patient protection with medical progress. Regulatory agencies can set international ethical standards for probiotic trials, including informed consent and independent ethical reviews. Transparent data sharing, and international collaboration can build trust and accelerate the transition from research to clinical practice.
14.4. Enhancing Professional Education and Interdisciplinary Collaboration
Promoting literacy in basic sciences, e.g., biochemistry and microbiology, among healthcare professionals through interdisciplinary workshops, seminars, and updated curricula fosters innovation and informed decision-making. Integrating probiotic science into medical education and offering continuous professional development can lead to the discovery of new probiotic strains or bioactive compounds with unique benefits for children, enhancing clinical practice and research.
Author Contributions
Conceptualization, L.P.G. and C.B.-O., writing—original draft preparation, L.P.G., C.B.-O., S.E.C.-P., J.Z.-M., E.T. and C.E.; writing—review and editing, L.P.G., C.B.-O., S.E.C.-P., J.Z.-M., E.T. and C.E.; supervision, L.P.G. and C.B.-O.; project administration, L.P.G. and C.B.-O. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Universidad San Francisco de Quito, through funding ID: 17884 “Development of a simple and efficient system for prototyping metabolic pathways in the probiotic yeast S. boulardii.”.
Informed Consent Statement
Not applicable.
Acknowledgments
NA.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Buonocore, G. Microbiota and gut immunity in infants and young children. Global Pediatrics 2024, 9, 100202. [CrossRef]
- Samara State Medical University; Sskvortsova, O.V.; Migacheva, N.B.; Samara State Medical University; Kaganova, T.I.; Samara State Medical University; Lyamin, A.V.; Samara State Medical University; Antipov, V.A.; Samara State Medical University Role of gut microbiota and selected metabolic products in the development of childhood obesity. bccm 2024, 17, 81–87. [CrossRef]
- Hoskinson, C.; Medeleanu, M.V.; Reyna, M.E.; Dai, D.L.Y.; Chowdhury, B.; Moraes, T.J.; Mandhane, P.J.; Simons, E.; Kozyrskyj, A.L.; Azad, M.B.; et al. Antibiotics taken within the first year of life are linked to infant gut microbiome disruption and elevated atopic dermatitis risk. J. Allergy Clin. Immunol. 2024, 154, 131–142. [CrossRef]
- Sharma, S.; Singh, N. Probiotics for Children. In Probiotics: A comprehensive guide to enhance health and mitigate disease; CRC Press: Boca Raton, 2024; pp. 95–128 ISBN 9781003452249.
- Bompalli, L.K. The Next-Gen Innovative Therapeutic Potential of Probiotics: Insights into Gut Microbiota Modulation and Health Promotion. Unpublished 2024. [CrossRef]
- Eastwood, J.; van Hemert, S.; Poveda, C.; Elmore, S.; Williams, C.; Lamport, D.; Walton, G. The effect of probiotic bacteria on composition and metabolite production of faecal microbiota using in vitro batch cultures. Nutrients 2023, 15. [CrossRef]
- Jang, H.J.; Lee, N.-K.; Paik, H.-D. A narrative review on the advance of probiotics to metabiotics. J. Microbiol. Biotechnol. 2024, 34, 487–494. [CrossRef]
- Alsharairi, N.A. Therapeutic Potential of Gut Microbiota and Its Metabolite Short-Chain Fatty Acids in Neonatal Necrotizing Enterocolitis. Life (Basel) 2023, 13. [CrossRef]
- Golpour, F.; Abbasi-Alaei, M.; Babaei, F.; Mirzababaei, M.; Parvardeh, S.; Mohammadi, G.; Nassiri-Asl, M. Short chain fatty acids, a possible treatment option for autoimmune diseases. Biomed. Pharmacother. 2023, 163, 114763. [CrossRef]
- Calvo, L.N.; Greenberg, R.G.; Gray, K.D. Safety and Effectiveness of Probiotics in Preterm Infants with Necrotizing Enterocolitis. Neoreviews 2024, 25, e193–e206. [CrossRef]
- Sharif, S.; Meader, N.; Oddie, S.J.; Rojas-Reyes, M.X.; McGuire, W. Probiotics to prevent necrotising enterocolitis in very preterm or very low birth weight infants. Cochrane Database Syst. Rev. 2023, 7, CD005496. [CrossRef]
- Bartram, E.; Asai, M.; Gabant, P.; Wigneshweraraj, S. Enhancing the antibacterial function of probiotic Escherichia coli Nissle: when less is more. Appl. Environ. Microbiol. 2023, 89, e0097523. [CrossRef]
- Thuy, T.T.D.; Lu, H.-F.; Bregente, C.J.B.; Huang, F.-C.A.; Tu, P.-C.; Kao, C.-Y. Characterization of the broad-spectrum antibacterial activity of bacteriocin-like inhibitory substance-producing probiotics isolated from fermented foods. BMC Microbiol. 2024, 24, 85. [CrossRef]
- Lv, H.; Teng, Q.; Chen, J.; Peng, L.; Ren, Z.; Ma, L.; Yang, W.; Yu, B.; Wu, Z.; Wan, C. Probiotic potential of a novel exopolysaccharide produced by Bifidobacterium animalis subsp. Lactis SF. LWT 2024, 193, 115764. [CrossRef]
- LeBlanc, J.G.; Laiño, J.E.; del Valle, M.J.; de Giori, G.S.; Sesma, F.; Taranto, M.P. B-Group Vitamins Production by Probiotic Lactic Acid Bacteria. In Biotechnology of lactic acid bacteria: novel applications; Mozzi, F., Raya, R.R., Vignolo, G.M., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 2015; pp. 279–296 ISBN 9781118868386.
- Chowdhury, S.R. Psychobiotics and gut–brain axis—a new paradigm for improving mental health. In Probiotics: A comprehensive guide to enhance health and mitigate disease; CRC Press: Boca Raton, 2024; pp. 146–158 ISBN 9781003452249.
- Lozano, J.; Fabius, S.; Fernández-Ciganda, S.; Urbanavicius, J.; Piccini, C.; Scorza, C.; Zunino, P. Beneficial effect of GABA-producing Lactiplantibacillus strain LPB145 isolated from cheese starters evaluated in anxiety- and depression-like behaviours in rats. Benef. Microbes 2024, 15, 465–479. [CrossRef]
- Ambat, J.; Antonio, J.; Bala, A.; Suntornsaratoon, P.; Flores, J.; Li, W.V.; Jones, A. Metabolomic and transcriptomic correlative analyses in germ free mice link probiotic-associated metabolites to host intestinal fatty acid metabolism and b-oxidation. Physiology (Bethesda) 2024, 39. [CrossRef]
- Indira, M.; Venkateswarulu, T.C.; Abraham Peele, K.; Nazneen Bobby, M.; Krupanidhi, S. Bioactive molecules of probiotic bacteria and their mechanism of action: a review. 3 Biotech 2019, 9, 306. [CrossRef]
- Shealy, N.G.; Yoo, W.; Byndloss, M.X. Colonization resistance: metabolic warfare as a strategy against pathogenic Enterobacteriaceae. Curr. Opin. Microbiol. 2021, 64, 82–90. [CrossRef]
- Roy D’Souza, S.; Singh, S.; Ravi, L. Secondary metabolites produced from symbiotic microbes. In Microbial Symbionts; Elsevier, 2023; pp. 803–830 ISBN 9780323993340.
- Kumar, M.K.; Morya, S. Probiotics: Role of bioactives in foods and dietary supplements. Pharma Innovation 2022, 11, 2592–2597. [CrossRef]
- Sharma, D.; Rashid, G.; Sharma, L. Probiotics. In Probiotics: A comprehensive guide to enhance health and mitigate disease; CRC Press: Boca Raton, 2024; pp. 325–343 ISBN 9781003452249.
- Baldassare, M.A.; Bhattacharjee, D.; Coles, J.D.; Nelson, S.; McCollum, C.A.; Seekatz, A.M. Butyrate enhances Clostridioides difficile sporulation in vitro. J. Bacteriol. 2023, 205, e0013823. [CrossRef]
- Arbulu, S.; Kjos, M. Revisiting the multifaceted roles of bacteriocins : the multifaceted roles of bacteriocins. Microb. Ecol. 2024, 87, 41. [CrossRef]
- Tarique, M.; Ali, A.H.; Kizhakkayil, J.; Liu, S.-Q.; Oz, F.; Dertli, E.; Kamal-Eldin, A.; Ayyash, M. Exopolysaccharides from Enterococcus faecium and Streptococcus thermophilus: Bioactivities, gut microbiome effects, and fermented milk rheology. Food Chemistry: X 2024, 21, 101073. [CrossRef]
- Shenderov, B.A.; Sinitsa, A.V.; Zakharchenko, M.M.; Lang, C. Metabolic relationship between the host and its gut microbiota. In Metabiotics: present state, challenges and perspectives; Springer International Publishing: Cham, 2020; pp. 15–21 ISBN 978-3-030-34166-4.
- Oleskin, A.V.; Shenderov, B.A. Neuromodulatory effects and targets of the SCFAs and gasotransmitters produced by the human symbiotic microbiota. Microb. Ecol. Health Dis. 2016, 27, 30971. [CrossRef]
- Engevik, M.A.; Morra, C.N.; Röth, D.; Engevik, K.; Spinler, J.K.; Devaraj, S.; Crawford, S.E.; Estes, M.K.; Kalkum, M.; Versalovic, J. Microbial metabolic capacity for intestinal folate production and modulation of host folate receptors. Front. Microbiol. 2019, 10, 2305. [CrossRef]
- Chávarri, M.; Diez-Gutiérrez, L.; Marañón, I.; del Carmen Villarán, M.; Barrón, L.J.R. The role of probiotics in nutritional health: probiotics as nutribiotics. In Probiotics in the prevention and management of human diseases; Elsevier, 2022; pp. 397–415 ISBN 9780128237335.
- LeBlanc, J.G.; Chain, F.; Martín, R.; Bermúdez-Humarán, L.G.; Courau, S.; Langella, P. Beneficial effects on host energy metabolism of short-chain fatty acids and vitamins produced by commensal and probiotic bacteria. Microb. Cell Fact. 2017, 16, 79. [CrossRef]
- Duan, H.; Wang, L.; Huangfu, M.; Li, H. The impact of microbiota-derived short-chain fatty acids on macrophage activities in disease: Mechanisms and therapeutic potentials. Biomed. Pharmacother. 2023, 165, 115276. [CrossRef]
- Siddiqui, A.; Haider, R.; Aaqil, S.I.; Vohra, L.I.; Qamar, K.; Jawed, A.; Fatima, N.; Adnan, A.; Parikh, V.; Ochani, S.; et al. Probiotic formulations and gastro-intestinal diseases in the paediatric population: a narrative review. Ann Med Surg 2024, 86, 2836–2847. [CrossRef]
- Schneider, R.; Sant’Anna, A. Using probiotics in paediatric populations. Paediatr. Child Health 2022, 27, 482–491. [CrossRef]
- Thomas, J.P.; Raine, T.; Reddy, S.; Belteki, G. Probiotics for the prevention of necrotising enterocolitis in very low-birth-weight infants: a meta-analysis and systematic review. Acta Paediatr. 2017, 106, 1729–1741. [CrossRef]
- Zhang, Y.; Xu, Q.; Zhang, F.; Sun, C. Probiotics for Preventing Necrotizing Enterocolitis: A Meta-Analysis with Trial Sequential Analysis. J. Clin. Pharm. Ther. 2023, 2023, 1–15. [CrossRef]
- Isaeva, G.; Isaeva, R. Probiotics in the treatment of Helicobacter pylori infection: reality and perspective. Minerva Gastroenterol (Torino) 2022, 68, 277–288. [CrossRef]
- Jain, M.; Agarwal, A.; Singh, K.; Jarouliya, U. Probiotics and immune response in disease prevention and treatment. In Immune-Boosting Nutraceuticals for Better Human Health: Novel Applications; Apple Academic Press: New York, 2023; pp. 99–129 ISBN 9781003371069.
- Thoda, C.; Touraki, M. Immunomodulatory properties of probiotics and their derived bioactive compounds. Appl. Sci. 2023, 13, 4726. [CrossRef]
- Zambelli, R.A.; Mendonça, L.G. de Application of probiotics in food industry. In Probiotics: A Comprehensive Guide to Enhance Health and Mitigate Disease; CRC Press: Boca Raton, 2024; pp. 282–304 ISBN 9781003452249.
- Kumar, H.; Dhalaria, R.; Guleria, S.; Cimler, R.; Choudhary, R.; Dhanjal, D.S.; Singh, R.; Kimta, N.; Dulta, K.; Pathera, A.K.; et al. To exploring the role of probiotics, plant-based fermented products, and paraprobiotics as anti-inflammatory agents in promoting human health. Journal of Agriculture and Food Research 2023, 14, 100896. [CrossRef]
- Kadia, B.M.; Allen, S.J. Effect of Pre-, Pro-, and Synbiotics on Biomarkers of Systemic Inflammation in Children: A Scoping Review. Nutrients 2024, 16. [CrossRef]
- Di Costanzo, M.; Vella, A.; Infantino, C.; Morini, R.; Bruni, S.; Esposito, S.; Biasucci, G. Probiotics in infancy and childhood for food allergy prevention and treatment. Nutrients 2024, 16. [CrossRef]
- Abavisani, M.; Ebadpour, N.; Khoshrou, A.; Sahebkar, A. Boosting vaccine effectiveness: The groundbreaking role of probiotics. Journal of Agriculture and Food Research 2024, 16, 101189. [CrossRef]
- Bradford, G.; Asgari, B.; Smit, B.; Hatje, E.; Kuballa, A.; Katouli, M. The Efficacy of Selected Probiotic Strains and Their Combination to Inhibit the Interaction of Adherent-Invasive Escherichia coli (AIEC) with a Co-Culture of Caco-2:HT29-MTX Cells. Microorganisms 2024, 12. [CrossRef]
- You, L.; Lv, R.; Jin, H.; Ma, T.; Zhao, Z.; Kwok, L.-Y.; Sun, Z. A large-scale comparative genomics study reveals niche-driven and within-sample intra-species functional diversification in Lacticaseibacillus rhamnosus. Food Res. Int 2023, 173, 113446. [CrossRef]
- Ceapa, C.; Davids, M.; Ritari, J.; Lambert, J.; Wels, M.; Douillard, F.P.; Smokvina, T.; de Vos, W.M.; Knol, J.; Kleerebezem, M. The Variable Regions of Lactobacillus rhamnosus Genomes Reveal the Dynamic Evolution of Metabolic and Host-Adaptation Repertoires. Genome Biol. Evol. 2016, 8, 1889–1905. [CrossRef]
- Cheong, Y.E.; Kim, J.; Jin, Y.-S.; Kim, K.H. Elucidation of the fucose metabolism of probiotic Lactobacillus rhamnosus GG by metabolomic and flux balance analyses. J. Biotechnol. 2022, 360, 110–116. [CrossRef]
- Zhang, J.; Li, K.; Bu, X.; Cheng, S.; Duan, Z. Characterization of the anti-pathogenic, genomic and phenotypic properties of a Lacticaseibacillus rhamnosus VHProbi M14 isolate. PLoS ONE 2023, 18, e0285480. [CrossRef]
- Talbi, C.; Elmarrakchy, S.; Youssfi, M.; Bouzroud, S.; Belfquih, M.; Sifou, A.; Bouhaddou, N.; Badaoui, B.; Balahbib, A.; Bouyahya, A.; et al. Bacterial exopolysaccharides: from production to functional features. Prog Micobes Mol Biol 2023, 6. [CrossRef]
- Zheng, J.; Ahmad, A.A.; Yang, Y.; Liang, Z.; Shen, W.; Feng, M.; Shen, J.; Lan, X.; Ding, X. Lactobacillus rhamnosus CY12 Enhances Intestinal Barrier Function by Regulating Tight Junction Protein Expression, Oxidative Stress, and Inflammation Response in Lipopolysaccharide-Induced Caco-2 Cells. Int. J. Mol. Sci. 2022, 23. [CrossRef]
- Spaggiari, L.; Pedretti, N.; Ricchi, F.; Pinetti, D.; Campisciano, G.; De Seta, F.; Comar, M.; Kenno, S.; Ardizzoni, A.; Pericolini, E. An Untargeted Metabolomic Analysis of Lactobacillus (L.) rhamnosus, L. acidophilus, L. plantarum and L. reuteri Reveals an Upregulated Production of Inosine from L. rhamnosus. 2024. [CrossRef]
- Tso, L.; Bonham, K.S.; Fishbein, A.; Rowland, S.; Klepac-Ceraj, V. Targeted High-Resolution Taxonomic Identification of Bifidobacterium longum subsp. infantis Using Human Milk Oligosaccharide Metabolizing Genes. Nutrients 2021, 13. [CrossRef]
- Román, L.; Melis-Arcos, F.; Pröschle, T.; Saa, P.A.; Garrido, D. Genome-scale metabolic modeling of the human milk oligosaccharide utilization by Bifidobacterium longum subsp. infantis. mSystems 2024, 9, e0071523. [CrossRef]
- Wong, C.B.; Huang, H.; Ning, Y.; Xiao, J. Probiotics in the New Era of Human Milk Oligosaccharides (HMOs): HMO Utilization and Beneficial Effects of Bifidobacterium longum subsp. infantis M-63 on Infant Health. Microorganisms 2024, 12. [CrossRef]
- Reens, A.L.; Cosetta, C.M.; Saur, R.; Trofimuk, O.; Brooker, S.L.; Lee, M.L.; Sun, A.K.; McKenzie, G.J.; Button, J.E. Tunable control of B. infantis abundance and gut metabolites by co-administration of human milk oligosaccharides. Gut Microbes 2024, 16, 2304160. [CrossRef]
- Sela, D.A.; Chapman, J.; Adeuya, A.; Kim, J.H.; Chen, F.; Whitehead, T.R.; Lapidus, A.; Rokhsar, D.S.; Lebrilla, C.B.; German, J.B.; et al. The genome sequence of Bifidobacterium longum subsp. infantis reveals adaptations for milk utilization within the infant microbiome. Proc Natl Acad Sci USA 2008, 105, 18964–18969. [CrossRef]
- Ehrlich, A.M.; Pacheco, A.R.; Henrick, B.M.; Taft, D.; Xu, G.; Huda, M.N.; Mishchuk, D.; Goodson, M.L.; Slupsky, C.; Barile, D.; et al. Indole-3-lactic acid associated with Bifidobacterium-dominated microbiota significantly decreases inflammation in intestinal epithelial cells. BMC Microbiol. 2020, 20, 357. [CrossRef]
- Yu, D.; Pei, Z.; Chen, Y.; Wang, H.; Xiao, Y.; Zhang, H.; Chen, W.; Lu, W. Bifidobacterium longum subsp. infantis as widespread bacteriocin gene clusters carrier stands out among the Bifidobacterium. Appl. Environ. Microbiol. 2023, 89, e0097923. [CrossRef]
- Zhang, H.; Wang, J.; Shen, J.; Chen, S.; Yuan, H.; Zhang, X.; Liu, X.; Yu, Y.; Li, X.; Gao, Z.; et al. Prophylactic supplementation with Bifidobacterium infantis or its metabolite inosine attenuates cardiac ischemia/reperfusion injury. iMeta 2024, 3, e220. [CrossRef]
- Kapse, N.; Pisu, V.; Dhakephalkar, T.; Margale, P.; Shetty, D.; Wagh, S.; Dagar, S.; Dhakephalkar, P.K. Unveiling the Probiotic Potential of Streptococcus thermophilus MCC0200: Insights from In Vitro Studies Corroborated with Genome Analysis. Microorganisms 2024, 12. [CrossRef]
- Rau, M.H.; Gaspar, P.; Jensen, M.L.; Geppel, A.; Neves, A.R.; Zeidan, A.A. Genome-Scale Metabolic Modeling Combined with Transcriptome Profiling Provides Mechanistic Understanding of Streptococcus thermophilus CH8 Metabolism. Appl. Environ. Microbiol. 2022, 88, e0078022. [CrossRef]
- Li, Y.; Wang, Y.; Li, B.; Hou, B.; Hung, W.; He, J.; Jiang, Y.; Zhang, Y.; Man, C. Streptococcus thermophilus JM905-Strain Carbon Source Utilization and Its Fermented Milk Metabolic Profile at Different Fermentation Stages. Foods 2023, 12. [CrossRef]
- Qiao, Y.; Liu, G.; Leng, C.; Zhang, Y.; Lv, X.; Chen, H.; Sun, J.; Feng, Z. Metabolic profiles of cysteine, methionine, glutamate, glutamine, arginine, aspartate, asparagine, alanine and glutathione in Streptococcus thermophilus during pH-controlled batch fermentations. Sci. Rep. 2018, 8, 12441. [CrossRef]
- Yu, P.; Jiang, Y.; Pan, Y.; Guo, M.; Yang, B.; Liu, X.; Zhao, J.; Zhang, H.; Chen, W. Altered microbiome and metabolome features provide clues in understanding strain- specific regulation of Streptococcus thermophilus in the host. Res. Sq. 2022. [CrossRef]
- Vitetta, L.; Llewellyn, H.; Oldfield, D. Gut Dysbiosis and the Intestinal Microbiome: Streptococcus thermophilus a Key Probiotic for Reducing Uremia. Microorganisms 2019, 7. [CrossRef]
- Amiri, S. Co-production of parabiotic metabolites by Lactobacillus acidophilus LA5 and Bifidobacterium animalis subsp. lactis BB12 in dairy effluents. College & Research Libraries 2021.
- Rani, P.; Tiwari, S.K. Health benefits of bacteriocins produced by probiotic lactic acid bacteria. In Microbial Biomolecules; Elsevier, 2023; pp. 97–111 ISBN 9780323994767.
- Deliorman Orhan, D. Bacteriocins produced by probiotic microorganisms. In Advances in Probiotics; Elsevier, 2021; pp. 277–291 ISBN 9780128229095.
- Wang, G.; Yu, Y.; Garcia-Gutierrez, E.; Jin, X.; He, Y.; Wang, L.; Tian, P.; Liu, Z.; Zhao, J.; Zhang, H.; et al. Lactobacillus acidophilus JCM 1132 Strain and Its Mutant with Different Bacteriocin-Producing Behaviour Have Various in Situ Effects on the Gut Microbiota of Healthy Mice. Microorganisms 2019, 8. [CrossRef]
- Lau, H.C.-H.; Zhang, X.; Ji, F.; Lin, Y.; Liang, W.; Li, Q.; Chen, D.; Fong, W.; Kang, X.; Liu, W.; et al. Lactobacillus acidophilus suppresses non-alcoholic fatty liver disease-associated hepatocellular carcinoma through producing valeric acid. EBioMedicine 2024, 100, 104952. [CrossRef]
- Prangli, A.-L.; Utt, M.; Talja, I.; Sepp, E.; Mikelsaar, M.; Rajasalu, T.; Uibo, O.; Tillmann, V.; Uibo, R. Antigenic proteins of Lactobacillus acidophilus that are recognised by serum IgG antibodies in children with type 1 diabetes and coeliac disease. Pediatr. Allergy Immunol. 2010, 21, e772-9. [CrossRef]
- Brzozowski, B.; Bednarski, W.; Dziuba, B. Functional properties of Lactobacillus acidophilus metabolites. J. Sci. Food Agric. 2009, 89, 2467–2476. [CrossRef]
- Reid, G.; Heinemann, C.; Velraeds, M.; van der Mei, H.C.; Busscher, H.J. [31] Biosurfactants produced by Lactobacillus. In Biofilms; Methods in Enzymology; Elsevier, 1999; Vol. 310, pp. 426–433 ISBN 9780121822118.
- Datta, S.; Timson, D.J.; Annapure, U.S. Antioxidant properties and global metabolite screening of the probiotic yeast Saccharomyces cerevisiae var. boulardii. J. Sci. Food Agric. 2017, 97, 3039–3049. [CrossRef]
- Ansari, F.; Alian Samakkhah, S.; Bahadori, A.; Jafari, S.M.; Ziaee, M.; Khodayari, M.T.; Pourjafar, H. Health-promoting properties of Saccharomyces cerevisiae var. boulardii as a probiotic; characteristics, isolation, and applications in dairy products. Crit. Rev. Food Sci. Nutr. 2023, 63, 457–485. [CrossRef]
- Bai, A.; Weaver, M.; Bao, F.; Chan, E.D.; Bai, X. Saccharomyces boulardii Produces a Factor That Inhibits Mycobacterium intracellulare Burden in Human Macrophages. AIM 2016, 06, 965–974. [CrossRef]
- Buts, J.-P.; Dekeyser, N.; Stilmant, C.; Delem, E.; Smets, F.; Sokal, E. Saccharomyces boulardii produces in rat small intestine a novel protein phosphatase that inhibits Escherichia coli endotoxin by dephosphorylation. Pediatr. Res. 2006, 60, 24–29. [CrossRef]
- Vandenplas, Y.; Brunser, O.; Szajewska, H. Saccharomyces boulardii in childhood. Eur. J. Pediatr. 2009, 168, 253–265. [CrossRef]
- Pais, P.; Almeida, V.; Yılmaz, M.; Teixeira, M.C. Saccharomyces boulardii: What Makes It Tick as Successful Probiotic? J Fungi (Basel) 2020, 6. [CrossRef]
- Offei, B.; Vandecruys, P.; De Graeve, S.; Foulquié-Moreno, M.R.; Thevelein, J.M. Unique genetic basis of the distinct antibiotic potency of high acetic acid production in the probiotic yeast Saccharomyces cerevisiae var. boulardii. Genome Res. 2019, 29, 1478–1494. [CrossRef]
- Im, E.; Pothoulakis, C. [Recent advances in Saccharomyces boulardii research]. Gastroenterol. Clin. Biol. 2010, 34 Suppl 1, S62-70. [CrossRef]
- dos Santos Freitas, A.; da Silva Fernandes, L.J.; Coelho-Rocha, N.D.; de Jesus, L.C.L.; de Rezende Rodovalho, V.; da Silva, T.F.; de Oliveira Carvalho, R.D.; Azevedo, V. Immunomodulatory and antiinflammatory mechanisms of probiotics. In Probiotics; Elsevier, 2022; pp. 321–341 ISBN 9780323851701.
- Tao, T.; Zhang, L.; Yu, T.; Ma, J.; Lu, S.; Ren, J.; Li, X.; Guo, X. Exopolysaccharide production by Lactobacillus plantarum T10 is responsible for the probiotic activity in enhancing intestinal barrier function in vitro and in vivo. Food Funct. 2024, 15, 3583–3599. [CrossRef]
- Chia, T.-Y.; Zolp, A.; Miska, J. Polyamine immunometabolism: central regulators of inflammation, cancer and autoimmunity. Cells 2022, 11. [CrossRef]
- Filidou, E.; Kandilogiannakis, L.; Shrewsbury, A.; Kolios, G.; Kotzampassi, K. Probiotics: Shaping the gut immunological responses. World J. Gastroenterol. 2024, 30, 2096–2108. [CrossRef]
- Verhoeven, D. Immunometabolism and innate immunity in the context of immunological maturation and respiratory pathogens in young children. J. Leukoc. Biol. 2019, 106, 301–308. [CrossRef]
- Fong, F.L.Y.; Shah, N.P.; Kirjavainen, P.; El-Nezami, H. Mechanism of Action of Probiotic Bacteria on Intestinal and Systemic Immunities and Antigen-Presenting Cells. Int. Rev. Immunol. 2016, 35, 179–188. [CrossRef]
- Yousefi, B.; Eslami, M.; Ghasemian, A.; Kokhaei, P.; Salek Farrokhi, A.; Darabi, N. Probiotics importance and their immunomodulatory properties. J. Cell. Physiol. 2019, 234, 8008–8018. [CrossRef]
- Kleniewska, P.; Pawliczak, R. Can probiotics be used in the prevention and treatment of bronchial asthma? Pharmacol. Rep. 2024. [CrossRef]
- Nova, E.; Wärnberg, J.; Gómez-Martínez, S.; Díaz, L.E.; Romeo, J.; Marcos, A. Immunomodulatory effects of probiotics in different stages of life. Br. J. Nutr. 2007, 98 Suppl 1, S90-5. [CrossRef]
- Fukushima, Y.; Kawata, Y.; Hara, H.; Terada, A.; Mitsuoka, T. Effect of a probiotic formula on intestinal immunoglobulin A production in healthy children. Int. J. Food Microbiol. 1998, 42, 39–44. [CrossRef]
- Fong, F.L.Y.; El-Nezami, H.; Mykkänen, O.; Kirjavainen, P.V. The effects of single strains and mixtures of probiotic bacteria on immune profile in liver, spleen, and peripheral blood. Front. Nutr. 2022, 9, 773298. [CrossRef]
- Zhu, H.; Guo, L.; Yu, D.; Du, X. New insights into immunomodulatory properties of lactic acid bacteria fermented herbal medicines. Front. Microbiol. 2022, 13, 1073922. [CrossRef]
- Licciardi, P.V.; Wong, S.-S.; Tang, M.L.; Karagiannis, T.C. Epigenome targeting by probiotic metabolites. Gut Pathog. 2010, 2, 24. [CrossRef]
- DeMuri, G.P.; Lehtoranta, L.M.; Eickhoff, J.C.; Lehtinen, M.J.; Wald, E.R. Ex vivo peripheral blood mononuclear cell response to R848 in children after supplementation with the probiotic Lactobacillus acidophilus NCFM/Bifidobacterium lactis Bi-07. Benef. Microbes 2021, 12, 85–93. [CrossRef]
- Duan, Y.; Wang, L.; Ma, Y.; Ning, L.; Zhang, X. A meta-analysis of the therapeutic effect of probiotic intervention in obese or overweight adolescents. Front Endocrinol (Lausanne) 2024, 15, 1335810. [CrossRef]
- Atazadegan, M.A.; Heidari-Beni, M.; Entezari, M.H.; Sharifianjazi, F.; Kelishadi, R. Effects of synbiotic supplementation on anthropometric indices and body composition in overweight or obese children and adolescents: a randomized, double-blind, placebo-controlled clinical trial. World J. Pediatr. 2023, 19, 356–365. [CrossRef]
- Karlsson Videhult, F.; Andersson, Y.; Öhlund, I.; Stenlund, H.; Hernell, O.; West, C.E. Impact of probiotics during weaning on the metabolic and inflammatory profile: follow-up at school age. Int. J. Food Sci. Nutr. 2015, 66, 686–691. [CrossRef]
- Guo, S.; Guo, Y.; Ergun, A.; Lu, L.; Walker, W.A.; Ganguli, K. Secreted Metabolites of Bifidobacterium infantis and Lactobacillus acidophilus Protect Immature Human Enterocytes from IL-1β-Induced Inflammation: A Transcription Profiling Analysis. PLoS ONE 2015, 10, e0124549. [CrossRef]
- Matsumoto, M.; Benno, Y. Anti-inflammatory metabolite production in the gut from the consumption of probiotic yogurt containing Bifidobacterium animalis subsp. lactis LKM512. Biosci. Biotechnol. Biochem. 2006, 70, 1287–1292. [CrossRef]
- Ballini, A.; Gnoni, A.; De Vito, D.; Dipalma, G.; Cantore, S.; Gargiulo Isacco, C.; Saini, R.; Santacroce, L.; Topi, S.; Scarano, A.; et al. Effect of probiotics on the occurrence of nutrition absorption capacities in healthy children: a randomized double-blinded placebo-controlled pilot study. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 8645–8657. [CrossRef]
- Chen, A.-C.; Fang, T.-J.; Ho, H.-H.; Chen, J.-F.; Kuo, Y.-W.; Huang, Y.-Y.; Tsai, S.-Y.; Wu, S.-F.; Lin, H.-C.; Yeh, Y.-T. A multi-strain probiotic blend reshaped obesity-related gut dysbiosis and improved lipid metabolism in obese children. Front. Nutr. 2022, 9. [CrossRef]
- Li, Y.; Liu, T.; Qin, L.; Wu, L. Effects of probiotic administration on overweight or obese children: a meta-analysis and systematic review. J. Transl. Med. 2023, 21, 525. [CrossRef]
- Toe, L.C.; Kerckhof, F.-M.; De Bodt, J.; Morel, F.B.; Ouedraogo, J.-B.; Kolsteren, P.; Van de Wiele, T. A prebiotic-enhanced lipid-based nutrient supplement (LNSp) increases Bifidobacterium relative abundance and enhances short-chain fatty acid production in simulated colonic microbiota from undernourished infants. FEMS Microbiol. Ecol. 2020, 96. [CrossRef]
- Kambale, R.M.; Ntagazibwa, J.N.; Kasengi, J.B.; Zigashane, A.B.; Francisca, I.N.; Mashukano, B.N.; Amani Ngaboyeka, G.; Bahizire, E.; Zech, F.; Bindels, L.B.; et al. Probiotics for children with uncomplicated severe acute malnutrition (PruSAM study): A randomized controlled trial in the Democratic Republic of Congo. Am. J. Clin. Nutr. 2023, 117, 976–984. [CrossRef]
- M., G.; A., G.; C, H.; M, R.; N, I.; Ts, M. The probiotic BIOHM improves nutrient absorption by disrupting gastrointestinal biofilms. J. Prob. Health 2019, 07. [CrossRef]
- Fan, Y.; Pedersen, O. Gut microbiota in human metabolic health and disease. Nat. Rev. Microbiol. 2021, 19, 55–71. [CrossRef]
- Lozupone, C.A.; Stombaugh, J.I.; Gordon, J.I.; Jansson, J.K.; Knight, R. Diversity, stability and resilience of the human gut microbiota. Nature 2012, 489, 220–230. [CrossRef]
- Bandsma, R.H.J.; Sadiq, K.; Bhutta, Z.A. Persistent diarrhoea: current knowledge and novel concepts. Paediatr. Int. Child Health 2019, 39, 41–47. [CrossRef]
- Campbell, D.I.; McPhail, G.; Lunn, P.G.; Elia, M.; Jeffries, D.J. Intestinal inflammation measured by fecal neopterin in Gambian children with enteropathy: association with growth failure, Giardia lamblia, and intestinal permeability. J. Pediatr. Gastroenterol. Nutr. 2004, 39, 153–157. [CrossRef]
- Sjögren, Y.M.; Jenmalm, M.C.; Böttcher, M.F.; Björkstén, B.; Sverremark-Ekström, E. Altered early infant gut microbiota in children developing allergy up to 5 years of age. Clin. Exp. Allergy 2009, 39, 518–526. [CrossRef]
- Bhattacharjee, R.; Pathak, K.; Das, J.; Bordoloi, S.; Pathak, M.P.; Barbhuiya, P.A.; Saikia, R. Therapeutic potential of gut microbiota in child health. AIA 2023, 21. [CrossRef]
- Navarro-Tapia, E.; Sebastiani, G.; Sailer, S.; Toledano, L.A.; Serra-Delgado, M.; García-Algar, Ó.; Andreu-Fernández, V. Probiotic supplementation during the perinatal and infant period: effects on gut dysbiosis and disease. Nutrients 2020, 12. [CrossRef]
- Maiuolo, J.; Bulotta, R.M.; Ruga, S.; Nucera, S.; Macrì, R.; Scarano, F.; Oppedisano, F.; Carresi, C.; Gliozzi, M.; Musolino, V.; et al. The Postbiotic Properties of Butyrate in the Modulation of the Gut Microbiota: The Potential of Its Combination with Polyphenols and Dietary Fibers. Int. J. Mol. Sci. 2024, 25. [CrossRef]
- Dogra, S.K.; Dardinier, A.; Mainardi, F.; Siegwald, L.; Bartova, S.; Roy, C. le; Chou, C.J. Application of computational data modeling to a large-scale population cohort assists the discovery of specific nutrients that influence beneficial human gut bacteria Faecalibacterium prausnitzii. BioRxiv 2022. [CrossRef]
- Kadry, A.A.; El-Antrawy, M.A.; El-Ganiny, A.M. Impact of short chain fatty acids (SCFAs) on antimicrobial activity of new β-lactam/β-lactamase inhibitor combinations and on virulence of Escherichia coli isolates. J. Antibiot. 2023, 76, 225–235. [CrossRef]
- Liang, L.; Liu, L.; Zhou, W.; Yang, C.; Mai, G.; Li, H.; Chen, Y. Gut microbiota-derived butyrate regulates gut mucus barrier repair by activating the macrophage/WNT/ERK signaling pathway. Clin. Sci. 2022, 136, 291–307. [CrossRef]
- Ornelas, A.; Welch, N.; Countess, J.A.; Zhou, L.; Wang, R.X.; Dowdell, A.S.; Colgan, S.P. Mimicry of microbially-derived butyrate reveals templates for potent intestinal epithelial HIF stabilizers. Gut Microbes 2023, 15, 2267706. [CrossRef]
- Recharla, N.; Geesala, R.; Shi, X.-Z. Gut microbial metabolite butyrate and its therapeutic role in inflammatory bowel disease: A literature review. Nutrients 2023, 15. [CrossRef]
- Conder, E.; Shay, H.C.; Vekaria, H.; Erinkitola, I.; Bhogoju, S.; Goretsky, T.; Sullivan, P.; Barrett, T.; Kapur, N. Butyrate-induced mitochondrial function improves barrier function in inflammatory bowel disease (ibd). Gastroenterology 2023, 164, S91. [CrossRef]
- Novella Nicese, M.; van den Berg, B.; Sol, W.; van Raalte, D.; Nieuwdorp, M.; Rotmans, J. MO618: influence of butyrate and acetate on behavior and metabolism of human glomerular microvascular endothelial cells. Nephrol. Dial. Transplant 2022, 37. [CrossRef]
- Wang, S.P.; Rubio, L.A.; Duncan, S.H.; Donachie, G.E.; Holtrop, G.; Lo, G.; Farquharson, F.M.; Wagner, J.; Parkhill, J.; Louis, P.; et al. Pivotal Roles for pH, Lactate, and Lactate-Utilizing Bacteria in the Stability of a Human Colonic Microbial Ecosystem. mSystems 2020, 5. [CrossRef]
- Zhang, C.; Zhang, S.; Liu, W.; Guo, T.; Gu, R.; Kong, J. Potential Application and Bactericidal Mechanism of Lactic Acid-Hydrogen Peroxide Consortium. Appl. Biochem. Biotechnol. 2019, 189, 822–833. [CrossRef]
- Gu, Q.; Li, P. Biosynthesis of vitamins by probiotic bacteria. In Probiotics and prebiotics in human nutrition and health; Rao, V., Rao, L.G., Eds.; InTech, 2016 ISBN 978-953-51-2475-7.
- Odumosu, B.T.; Bamidele, T.A.; Ofem, D.W.; Agbozo, F.; Olasehinde, G.I. Screening, isolation and biotechnological potentials of foodborne Lactobacillus fermentum strains MT903311 and MT903312. Heliyon 2023, 9, e14959. [CrossRef]
- Yin, J.; Peng, X.; Yang, A.; Lin, M.; Ji, K.; Dai, X.; Huang, J.; Li, L.; Feng, L. Impact of oligosaccharides on probiotic properties and B vitamins production: a comprehensive assessment of probiotic strains. Int. J. Food Sci. Technol. 2024, 59, 6044–6064. [CrossRef]
- Uddin, M.; Uddin, S.; Deb, B.; Hussain, T.; Rafi, S.; Islam, A. The role of lactobacillus reuteri probiotic for preventing functional gastrointestinal disorders in toddlerhood. IJG 2024, 8, 14–21. [CrossRef]
- Torres, A.C.; Vannini, V.; Font, G.; Saavedra, L.; Taranto, M.P. Novel pathway for corrinoid compounds production in lactobacillus. Front. Microbiol. 2018, 9, 2256. [CrossRef]
- Langa, S.; Peirotén, Á.; Rodríguez, S.; Calzada, J.; Prieto-Paredes, R.; Curiel, J.A.; Landete, J.M. Riboflavin bio-enrichment of soy beverage by selected roseoflavin-resistant and engineered lactic acid bacteria. Int. J. Food Microbiol. 2024, 411, 110547. [CrossRef]
- Meucci, A.; Rossetti, L.; Zago, M.; Monti, L.; Giraffa, G.; Carminati, D.; Tidona, F. Folates biosynthesis by Streptococcus thermophilus during growth in milk. Food Microbiol. 2018, 69, 116–122. [CrossRef]
- Smajdor, J.; Jedlińska, K.; Porada, R.; Górska-Ratusznik, A.; Policht, A.; Śróttek, M.; Więcek, G.; Baś, B.; Strus, M. The impact of gut bacteria producing long chain homologs of vitamin K2 on colorectal carcinogenesis. Cancer Cell Int. 2023, 23, 268. [CrossRef]
- Parama Iswara, R.; Yunus, K.; Pati, N. Vitamin K Deficiency. In Causes and management of nutritional deficiency disorders; Nayak, A., Misra, S., Eds.; Advances in medical diagnosis, treatment, and care; IGI Global, 2024; pp. 214–218 ISBN 9798369329474.
- A. Aljafary, M.; Alshwyeh, H.; Alahmadi, N.; Shehzad, A.; Tombuloglu, H.; Gaymalov, Z.; Homieda, A.; Al-Suhaimi, E. Physiological and cellular functions of vitamin K on cardiovascular function. In Vitamin K—Recent Topics on the Biology and Chemistry; Kagechika, H., Shirakawa, H., Eds.; Biochemistry; IntechOpen, 2022; Vol. 27 ISBN 978-1-83969-391-5.
- Ivanko, O.G.; Solianyk, O.V. Antibiotic-associated disorders of prothrombin synthesis and their probiotic correction with B. Clausii in breastfeed infants. ZMJ 2018, 0. [CrossRef]
- Liu, Y. Good things come in small packages : delivery of vitamin K2 to human cells by extracellular vesicles from Lactococcus lactis. Doctoral dissertation, Wageningen University., 2022.
- Cooke, G.; Behan, J.; Costello, M. Newly identified vitamin K-producing bacteria isolated from the neonatal faecal flora. Microb. Ecol. Health Dis. 2006, 18, 133–138. [CrossRef]
- Kaushal, D.; Kalsi, G. Dietary intervention of prebiotics and vitamins on gut health of children. Nutrition & Food Science 2022, 52, 1045–1054. [CrossRef]
- Kim, J.-Y.; Choi, E.-J.; Lee, J.-H.; Yoo, M.-S.; Heo, K.; Shim, J.-J.; Lee, J.-L. Probiotic Potential of a Novel Vitamin B2-Overproducing Lactobacillus plantarum Strain, HY7715, Isolated from Kimchi. Appl. Sci. 2021, 11, 5765. [CrossRef]
- Salvi, P.S.; Cowles, R.A. Butyrate and the intestinal epithelium: modulation of proliferation and inflammation in homeostasis and disease. Cells 2021, 10. [CrossRef]
- Round, J.L.; Mazmanian, S.K. Inducible Foxp3+ regulatory T-cell development by a commensal bacterium of the intestinal microbiota. Proc Natl Acad Sci USA 2010, 107, 12204–12209. [CrossRef]
- Nilsen, M.; Nygaard, U.C.; Brodin, P.; Carlsen, K.C.L.; Fredheim, C.; Haugen, G.; Hedlin, G.; Jonassen, C.M.; Jonsmoen, U.L.A.; Lakshmikanth, T.; et al. Gut bacteria at 6 months of age are associated with immune cell status in 1-year-old children. Scand. J. Immunol. 2024, 99, e13346. [CrossRef]
- Treem, W.R.; Ahsan, N.; Shoup, M.; Hyams, J.S. Fecal short-chain fatty acids in children with inflammatory bowel disease. J. Pediatr. Gastroenterol. Nutr. 1994, 18, 159–164. [CrossRef]
- Lopetuso, L.R.; Scaldaferri, F.; Petito, V.; Gasbarrini, A. Commensal Clostridia: leading players in the maintenance of gut homeostasis. Gut Pathog. 2013, 5, 23. [CrossRef]
- Hossain, M.S.; Das, S.; Gazi, M.A.; Alam, M.A.; Haque, N.M.S.; Mahfuz, M.; Ahmed, T.; Damman, C.J. Association of faecal pH with childhood stunting: Results from a cross-sectional study. bmjpo 2019, 3, e000549. [CrossRef]
- van Limpt, C.; Crienen, A.; Vriesema, A.; Knol, J. 134 effect of colonic short chain fatty acids, lactate and PH on the growth of common gut pathogens. Pediatr. Res. 2004, 56, 487–487. [CrossRef]
- Hatayama, H.; Iwashita, J.; Kuwajima, A.; Abe, T. The short chain fatty acid, butyrate, stimulates MUC2 mucin production in the human colon cancer cell line, LS174T. Biochem. Biophys. Res. Commun. 2007, 356, 599–603. [CrossRef]
- Chen, Y. The SCFAs Production of Syntrophic Culture of L. johnsonii SZ-YL and A. Muciniphila in Different Macrobutrients. HSET 2023, 30, 24–33. [CrossRef]
- Ferrari, S.; Galla, R.; Mulè, S.; Rosso, G.; Brovero, A.; Macchi, V.; Ruga, S.; Uberti, F. The Role of Bifidobacterium bifidum novaBBF7, Bifidobacterium longum novaBLG2 and Lactobacillus paracasei TJB8 to Improve Mechanisms Linked to Neuronal Cells Protection against Oxidative Condition in a Gut-Brain Axis Model. Int. J. Mol. Sci. 2023, 24. [CrossRef]
- Cheng, Y.; Liu, J.; Ling, Z. Short-chain fatty acids-producing probiotics: A novel source of psychobiotics. Crit. Rev. Food Sci. Nutr. 2022, 62, 7929–7959. [CrossRef]
- Vital, M.; Karch, A.; Pieper, D.H. Colonic Butyrate-Producing Communities in Humans: an Overview Using Omics Data. mSystems 2017, 2. [CrossRef]
- Hojsak, I.; Benninga, M.A.; Hauser, B.; Kansu, A.; Kelly, V.B.; Stephen, A.M.; Morais Lopez, A.; Slavin, J.; Tuohy, K. Benefits of dietary fibre for children in health and disease. Arch. Dis. Child. 2022, 107, 973–979. [CrossRef]
- Cardoso, M.H.; Meneguetti, B.T.; Oliveira-Júnior, N.G.; Macedo, M.L.R.; Franco, O.L. Antimicrobial peptide production in response to gut microbiota imbalance. Peptides 2022, 157, 170865. [CrossRef]
- Hassan, M.; Kjos, M.; Nes, I.F.; Diep, D.B.; Lotfipour, F. Natural antimicrobial peptides from bacteria: characteristics and potential applications to fight against antibiotic resistance. J. Appl. Microbiol. 2012, 113, 723–736. [CrossRef]
- Huang, F.; Teng, K.; Liu, Y.; Cao, Y.; Wang, T.; Ma, C.; Zhang, J.; Zhong, J. Bacteriocins: potential for human health. Oxid. Med. Cell. Longev. 2021, 2021, 5518825. [CrossRef]
- Spohn, R.; Daruka, L.; Lázár, V.; Martins, A.; Vidovics, F.; Grézal, G.; Méhi, O.; Kintses, B.; Számel, M.; Jangir, P.K.; et al. Integrated evolutionary analysis reveals antimicrobial peptides with limited resistance. Nat. Commun. 2019, 10, 4538. [CrossRef]
- Andersson, D.I.; Hughes, D.; Kubicek-Sutherland, J.Z. Mechanisms and consequences of bacterial resistance to antimicrobial peptides. Drug Resist. Updat. 2016, 26, 43–57. [CrossRef]
- Yeaman, M.R.; Yount, N.Y. Mechanisms of antimicrobial peptide action and resistance. Pharmacol. Rev. 2003, 55, 27–55. [CrossRef]
- Mandal, S.M.; Pati, B.R.; Chakraborty, R.; Franco, O.L. New insights into the bioactivity of peptides from probiotics. Front Biosci (Elite Ed) 2016, 8, 450–459. [CrossRef]
- Heilbronner, S.; Krismer, B.; Brötz-Oesterhelt, H.; Peschel, A. The microbiome-shaping roles of bacteriocins. Nat. Rev. Microbiol. 2021, 19, 726–739. [CrossRef]
- Shin, J.M.; Gwak, J.W.; Kamarajan, P.; Fenno, J.C.; Rickard, A.H.; Kapila, Y.L. Biomedical applications of nisin. J. Appl. Microbiol. 2016, 120, 1449–1465. [CrossRef]
- Ashby, M.; Petkova, A.; Hilpert, K. Cationic antimicrobial peptides as potential new therapeutic agents in neonates and children: a review. Curr. Opin. Infect. Dis. 2014, 27, 258–267. [CrossRef]
- Arun, K.B.; Madhavan, A.; Emmanual, S.; Sindhu, R.; Binod, P.; Pandey, A. Enzymes in probiotics and genetically modified foods. In Value-Addition in Food Products and Processing Through Enzyme Technology; Elsevier, 2022; pp. 13–23 ISBN 9780323899291.
- Xie, W.; Li, W.; Gao, F.; Li, T. In vitro digestion and fermentation of lacto- N -biose, a core building block of human milk oligosaccharides. Int. J. Food Sci. Technol. 2023, 58, 4511–4518. [CrossRef]
- Florindo, R.N.; Souza, V.P.; Manzine, L.R.; Camilo, C.M.; Marana, S.R.; Polikarpov, I.; Nascimento, A.S. Structural and biochemical characterization of a GH3 β-glucosidase from the probiotic bacteria Bifidobacterium adolescentis. Biochimie 2018, 148, 107–115. [CrossRef]
- Ambrogi, V.; Bottacini, F.; O’Callaghan, J.; Casey, E.; van Breen, J.; Schoemaker, B.; Cao, L.; Kuipers, B.; O’Connell Motherway, M.; Schoterman, M.; et al. Infant-Associated Bifidobacterial β-Galactosidases and Their Ability to Synthesize Galacto-Oligosaccharides. Front. Microbiol. 2021, 12, 662959. [CrossRef]
- Xin, Y.; Guo, T.; Zhang, Y.; Wu, J.; Kong, J. A new β-galactosidase extracted from the infant feces with high hydrolytic and transgalactosylation activity. Appl. Microbiol. Biotechnol. 2019, 103, 8439–8448. [CrossRef]
- Enzymatic Production of Galactooligosaccharides by β-Galactosidase from Bifidobacterium longum BCRC 15708. Available online: https://pubs.acs.org/doi/10.1021/jf063126%2B (accessed on 5 September 2024).
- Antonowicz, I.; Shwachman, H.; Sotoo, I. Beta-galactosidase and beta-glucuronidase activities in intestinal mucosa of infants and children. Pediatrics 1971, 47, 737–744. [CrossRef]
- Cuxart, I.; Coines, J.; Esquivias, O.; Faijes, M.; Planas, A.; Biarnés, X.; Rovira, C. Enzymatic Hydrolysis of Human Milk Oligosaccharides. The Molecular Mechanism of Bifidobacterium Bifidum Lacto-N-biosidase. ACS Catal. 2022, 12, 4737–4743. [CrossRef]
- Ito, T.; Katayama, T.; Hattie, M.; Sakurama, H.; Wada, J.; Suzuki, R.; Ashida, H.; Wakagi, T.; Yamamoto, K.; Stubbs, K.A.; et al. Crystal structures of a glycoside hydrolase family 20 lacto-N-biosidase from Bifidobacterium bifidum. J. Biol. Chem. 2013, 288, 11795–11806. [CrossRef]
- El-Sayed, A.I.M.; El-Borai, A.M.; Akl, S.H.; El-Aassar, S.A.; Abdel-Latif, M.S. Identification of Lactobacillus strains from human mother milk and cottage cheese revealed potential probiotic properties with enzymatic activity. Sci. Rep. 2022, 12, 22522. [CrossRef]
- Li, C.; Ji, Q.; He, T.; Liu, Y.; Ma, Y. Characterization of a recombinant bile salt hydrolase (BSH) from Bifidobacterium bifidum for its glycine-conjugated bile salts specificity. Biocatal. Biotransformation 2021, 39, 61–70. [CrossRef]
- Öztürk, M.; Kılıçsaymaz, Z.; Önal, C. Site-Directed Mutagenesis of Bile Salt Hydrolase (BSH) from Lactobacillus plantarum B14 Confirms the Importance of the V58 and Y65 Amino Acids for Activity and Substrate Specificity. Food Biotechnol. 2023, 37, 74–88. [CrossRef]
- Núñez-Sánchez, M.A.; Herisson, F.M.; Keane, J.M.; García-González, N.; Rossini, V.; Pinhiero, J.; Daly, J.; Bustamante-Garrido, M.; Hueston, C.M.; Patel, S.; et al. Microbial bile salt hydrolase activity influences gene expression profiles and gastrointestinal maturation in infant mice. Gut Microbes 2022, 14, 2149023. [CrossRef]
- Ma, T.; Shen, X.; Shi, X.; Sakandar, H.A.; Quan, K.; Li, Y.; Jin, H.; Kwok, L.-Y.; Zhang, H.; Sun, Z. Targeting gut microbiota and metabolism as the major probiotic mechanism—An evidence-based review. Trends Food Sci. Technol. 2023, 138, 178–198. [CrossRef]
- Kaur, N.; Dey, P. Bacterial exopolysaccharides as emerging bioactive macromolecules: from fundamentals to applications. Res. Microbiol. 2023, 174, 104024. [CrossRef]
- Alessandri, G.; van Sinderen, D.; Ventura, M. The genus bifidobacterium: From genomics to functionality of an important component of the mammalian gut microbiota running title: Bifidobacterial adaptation to and interaction with the host. Comput. Struct. Biotechnol. J. 2021, 19, 1472–1487. [CrossRef]
- Xie, Y.; Ye, Z.; Wan, X.; Deng, H.; Sun, W.; He, X.; Chen, K. Screening of exopolysaccharide-producing Enterobacter aerogenes NJ1023 and its cadaverine biosynthesis promotion. Front. Microbiol. 2023, 14, 1200123. [CrossRef]
- Salimi, F.; Farrokh, P. Recent advances in the biological activities of microbial exopolysaccharides. World J. Microbiol. Biotechnol. 2023, 39, 213. [CrossRef]
- Milani, C.; Duranti, S.; Bottacini, F.; Casey, E.; Turroni, F.; Mahony, J.; Belzer, C.; Delgado Palacio, S.; Arboleya Montes, S.; Mancabelli, L.; et al. The first microbial colonizers of the human gut: composition, activities, and health implications of the infant gut microbiota. Microbiol. Mol. Biol. Rev. 2017, 81. [CrossRef]
- Sadeghi, M.; Haghshenas, B.; Nami, Y. Bifidobacterium exopolysaccharides: new insights into engineering strategies, physicochemical functions, and immunomodulatory effects on host health. Front. Microbiol. 2024, 15. [CrossRef]
- Caggianiello, G.; Kleerebezem, M.; Spano, G. Exopolysaccharides produced by lactic acid bacteria: from health-promoting benefits to stress tolerance mechanisms. Appl. Microbiol. Biotechnol. 2016, 100, 3877–3886. [CrossRef]
- Yue, Y.; Wang, Y.; Yang, R.; Liu, D.; Cheng, Y.; Li, S.; Sun, L.; Li, B.; Huo, G. Exopolysaccharides of Bifidobacterium longum subsp. infantis E4 on the immune and anti-inflammatory effects in vitro. J. Funct. Foods 2023, 107, 105699. [CrossRef]
- Wu, J.; Zhang, Y.; Ye, L.; Wang, C. The anti-cancer effects and mechanisms of lactic acid bacteria exopolysaccharides in vitro: A review. Carbohydr. Polym. 2021, 253, 117308. [CrossRef]
- Ren, Z.; Chen, S.; Lv, H.; Peng, L.; Yang, W.; Chen, J.; Wu, Z.; Wan, C. Effect of Bifidobacterium animalis subsp. lactis SF on enhancing the tumor suppression of irinotecan by regulating the intestinal flora. Pharmacol. Res. 2022, 184, 106406. [CrossRef]
- Wang, L.; Wang, Y.; Li, Q.; Tian, K.; Xu, L.; Liu, G.; Guo, C. Exopolysaccharide, Isolated From a Novel Strain Bifidobacterium breve lw01 Possess an Anticancer Effect on Head and Neck Cancer—Genetic and Biochemical Evidences. Front. Microbiol. 2019, 10, 1044. [CrossRef]
- Angelin, J.; Kavitha, M. Exopolysaccharides from probiotic bacteria and their health potential. Int. J. Biol. Macromol. 2020, 162, 853–865. [CrossRef]
- Kim, K.; Lee, G.; Thanh, H.D.; Kim, J.-H.; Konkit, M.; Yoon, S.; Park, M.; Yang, S.; Park, E.; Kim, W. Exopolysaccharide from Lactobacillus plantarum LRCC5310 offers protection against rotavirus-induced diarrhea and regulates inflammatory response. J. Dairy Sci. 2018, 101, 5702–5712. [CrossRef]
- Bhat, B.; Bajaj, B.K. Hypocholesterolemic potential and bioactivity spectrum of an exopolysaccharide from a probiotic isolate Lactobacillus paracasei M7. Bioactive Carbohydrates and Dietary Fibre 2019, 19, 100191. [CrossRef]
- Zhang, Z.; Zhou, Z.; Li, Y.; Zhou, L.; Ding, Q.; Xu, L. Isolated exopolysaccharides from Lactobacillus rhamnosus GG alleviated adipogenesis mediated by TLR2 in mice. Sci. Rep. 2016, 6, 36083. [CrossRef]
- Porges, E.C.; Woods, A.J.; Edden, R.A.E.; Puts, N.A.J.; Harris, A.D.; Chen, H.; Garcia, A.M.; Seider, T.R.; Lamb, D.G.; Williamson, J.B.; et al. Frontal Gamma-Aminobutyric Acid Concentrations Are Associated With Cognitive Performance in Older Adults. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2017, 2, 38–44. [CrossRef]
- Bravo, J.A.; Forsythe, P.; Chew, M.V.; Escaravage, E.; Savignac, H.M.; Dinan, T.G.; Bienenstock, J.; Cryan, J.F. Ingestion of Lactobacillus strain regulates emotional behavior and central GABA receptor expression in a mouse via the vagus nerve. Proc Natl Acad Sci USA 2011, 108, 16050–16055. [CrossRef]
- Frank, S.M.; Becker, M.; Qi, A.; Geiger, P.; Frank, U.I.; Rosedahl, L.A.; Malloni, W.M.; Sasaki, Y.; Greenlee, M.W.; Watanabe, T. Efficient learning in children with rapid GABA boosting during and after training. Curr. Biol. 2022, 32, 5022-5030.e7. [CrossRef]
- Yunes, R.A.; Poluektova, E.U.; Dyachkova, M.S.; Klimina, K.M.; Kovtun, A.S.; Averina, O.V.; Orlova, V.S.; Danilenko, V.N. GABA production and structure of gadB/gadC genes in Lactobacillus and Bifidobacterium strains from human microbiota. Anaerobe 2016, 42, 197–204. [CrossRef]
- Allen, M.J.; Sabir, S.; Sharma, S. GABA Receptor. In StatPearls; StatPearls Publishing: Treasure Island (FL), 2024.
- Saleh, M.G.; Papantoni, A.; Mikkelsen, M.; Hui, S.C.N.; Oeltzschner, G.; Puts, N.A.; Edden, R.A.E.; Carnell, S. Effect of age on GABA+ and glutathione in a pediatric sample. AJNR Am J Neuroradiol 2020, 41, 1099–1104. [CrossRef]
- Monteagudo-Mera, A.; Fanti, V.; Rodriguez-Sobstel, C.; Gibson, G.; Wijeyesekera, A.; Karatzas, K.-A.; Chakrabarti, B. Gamma aminobutyric acid production by commercially available probiotic strains. J. Appl. Microbiol. 2023, 134. [CrossRef]
- Braga, J.D.; Thongngam, M.; Kumrungsee, T. Gamma-aminobutyric acid as a potential postbiotic mediator in the gut-brain axis. npj Sci. Food 2024, 8, 16. [CrossRef]
- Ueno, H. Enzymatic and structural aspects on glutamate decarboxylase. Journal of Molecular Catalysis B: Enzymatic 2000, 10, 67–79. [CrossRef]
- Ma, D.; Lu, P.; Yan, C.; Fan, C.; Yin, P.; Wang, J.; Shi, Y. Structure and mechanism of a glutamate-GABA antiporter. Nature 2012, 483, 632–636. [CrossRef]
- Altaib, H.; Kozakai, T.; Badr, Y.; Nakao, H.; El-Nouby, M.A.M.; Yanase, E.; Nomura, I.; Suzuki, T. Cell factory for γ-aminobutyric acid (GABA) production using Bifidobacterium adolescentis. Microb. Cell Fact. 2022, 21, 33. [CrossRef]
- Tamés, H.; Sabater, C.; Margolles, A.; Ruiz, L.; Ruas-Madiedo, P. Production of GABA in milk fermented by Bifidobacterium adolescentis strains selected on the bases of their technological and gastrointestinal performance. Food Res. Int 2023, 171, 113009. [CrossRef]
- Pokusaeva, K.; Johnson, C.; Luk, B.; Uribe, G.; Fu, Y.; Oezguen, N.; Matsunami, R.K.; Lugo, M.; Major, A.; Mori-Akiyama, Y.; et al. GABA-producing Bifidobacterium dentium modulates visceral sensitivity in the intestine. Neurogastroenterol. Motil. 2017, 29. [CrossRef]
- Park, K.-B.; Ji, G.-E.; Park, M.-S.; Oh, S.-H. Expression of rice glutamate decarboxylase in Bifidobacterium longum enhances gamma-aminobutyric acid production. Biotechnol. Lett. 2005, 27, 1681–1684. [CrossRef]
- Cocean, A.-M.; Vodnar, D.C. Exploring the gut-brain Axis: Potential therapeutic impact of Psychobiotics on mental health. Prog Neuropsychopharmacol Biol Psychiatry 2024, 134, 111073. [CrossRef]
- Annahazi, A.; Schemann, M. The enteric nervous system: “A little brain in the gut.” Neuroforum 2020, 26, 31–42. [CrossRef]
- Bonaz, B.; Sinniger, V.; Pellissier, S. The Vagus Nerve in the Neuro-Immune Axis: Implications in the Pathology of the Gastrointestinal Tract. Front. Immunol. 2017, 8, 1452. [CrossRef]
- Barrio, C.; Arias-Sánchez, S.; Martín-Monzón, I. The gut microbiota-brain axis, psychobiotics and its influence on brain and behaviour: A systematic review. Psychoneuroendocrinology 2022, 137, 105640. [CrossRef]
- Sajdel-Sulkowska, E.M. The Impact of Maternal Gut Microbiota during Pregnancy on Fetal Gut-Brain Axis Development and Life-Long Health Outcomes. Microorganisms 2023, 11. [CrossRef]
- Ding, X.; Xu, Y.; Zhang, X.; Zhang, L.; Duan, G.; Song, C.; Li, Z.; Yang, Y.; Wang, Y.; Wang, X.; et al. Gut microbiota changes in patients with autism spectrum disorders. J. Psychiatr. Res. 2020, 129, 149–159. [CrossRef]
- Zuffa, S.; Schimmel, P.; Gonzalez-Santana, A.; Belzer, C.; Knol, J.; Bölte, S.; Falck-Ytter, T.; Forssberg, H.; Swann, J.; Diaz Heijtz, R. Early-life differences in the gut microbiota composition and functionality of infants at elevated likelihood of developing autism spectrum disorder. Transl. Psychiatry 2023, 13, 257. [CrossRef]
- Cassidy-Bushrow, A.E.; Sitarik, A.R.; Johnson, C.C.; Johnson-Hooper, T.M.; Kassem, Z.; Levin, A.M.; Lynch, S.V.; Ownby, D.R.; Phillips, J.M.; Yong, G.J.M.; et al. Early-life gut microbiota and attention deficit hyperactivity disorder in preadolescents. Pediatr. Res. 2023, 93, 2051–2060. [CrossRef]
- Pärtty, A.; Kalliomäki, M.; Wacklin, P.; Salminen, S.; Isolauri, E. A possible link between early probiotic intervention and the risk of neuropsychiatric disorders later in childhood: a randomized trial. Pediatr. Res. 2015, 77, 823–828. [CrossRef]
- Braga, T.D.; da Silva, G.A.P.; de Lira, P.I.C.; de Carvalho Lima, M. Efficacy of Bifidobacterium breve and Lactobacillus casei oral supplementation on necrotizing enterocolitis in very-low-birth-weight preterm infants: a double-blind, randomized, controlled trial. Am. J. Clin. Nutr. 2011, 93, 81–86. [CrossRef]
- Dilli, D.; Aydin, B.; Fettah, N.D.; Özyazıcı, E.; Beken, S.; Zenciroğlu, A.; Okumuş, N.; Özyurt, B.M.; İpek, M.Ş.; Akdağ, A.; Turan, Ö.; Bozdağ, Ş. The propre-save study: effects of probiotics and prebiotics alone or combined on necrotizing enterocolitis in very low birth weight infants. J. Pediatr. 2015, 166, 545–51.e1. [CrossRef]
- Murphy, K.; Ross, R.P.; Ryan, C.A.; Dempsey, E.M.; Stanton, C. Probiotics, prebiotics, and synbiotics for the prevention of necrotizing enterocolitis. Front. Nutr. 2021, 8, 667188. [CrossRef]
- O’Mahony, S.M.; Clarke, G.; Borre, Y.E.; Dinan, T.G.; Cryan, J.F. Serotonin, tryptophan metabolism and the brain-gut-microbiome axis. Behav. Brain Res. 2015, 277, 32–48. [CrossRef]
- Wall, R.; Cryan, J.F.; Ross, R.P.; Fitzgerald, G.F.; Dinan, T.G.; Stanton, C. Bacterial neuroactive compounds produced by psychobiotics. Adv. Exp. Med. Biol. 2014, 817, 221–239. [CrossRef]
- Yano, J.M.; Yu, K.; Donaldson, G.P.; Shastri, G.G.; Ann, P.; Ma, L.; Nagler, C.R.; Ismagilov, R.F.; Mazmanian, S.K.; Hsiao, E.Y. Indigenous bacteria from the gut microbiota regulate host serotonin biosynthesis. Cell 2015, 161, 264–276. [CrossRef]
- Asano, Y.; Hiramoto, T.; Nishino, R.; Aiba, Y.; Kimura, T.; Yoshihara, K.; Koga, Y.; Sudo, N. Critical role of gut microbiota in the production of biologically active, free catecholamines in the gut lumen of mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 303, G1288-95. [CrossRef]
- Thomas, C.M.; Versalovic, J. Probiotics-host communication: Modulation of signaling pathways in the intestine. Gut Microbes 2010, 1, 148–163. [CrossRef]
- Barrett, E.; Ross, R.P.; O’Toole, P.W.; Fitzgerald, G.F.; Stanton, C. γ-Aminobutyric acid production by culturable bacteria from the human intestine. J. Appl. Microbiol. 2012, 113, 411–417. [CrossRef]
- Salminen, S.; Collado, M.C.; Endo, A.; Hill, C.; Lebeer, S.; Quigley, E.M.M.; Sanders, M.E.; Shamir, R.; Swann, J.R.; Szajewska, H.; et al. The International Scientific Association of Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of postbiotics. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 649–667. [CrossRef]
- Ozma, M.A.; Abbasi, A.; Akrami, S.; Lahouty, M.; Shahbazi, N.; Ganbarov, K.; Pagliano, P.; Sabahi, S.; Köse, Ş.; Yousefi, M.; Dao, S.; et al. Postbiotics as the key mediators of the gut microbiota-host interactions. Infez. Med. 2022, 30, 180–193. [CrossRef]
- Aguilar-Toalá, J.E.; Garcia-Varela, R.; Garcia, H.S.; Mata-Haro, V.; González-Córdova, A.F.; Vallejo-Cordoba, B.; Hernández-Mendoza, A. Postbiotics: An evolving term within the functional foods field. Trends Food Sci. Technol. 2018, 75, 105–114. [CrossRef]
- Konstantinov, S.R.; Kuipers, E.J.; Peppelenbosch, M.P. Functional genomic analyses of the gut microbiota for CRC screening. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 741–745. [CrossRef]
- Sawada, H.; Furushiro, M.; Hirai, K.; Motoike, M.; Watanabe, T.; Yokokura, T. Purification and characterization of an antihypertensive compound from Lactobacillus casei. Agric. Biol. Chem. 1990, 54, 3211–3219.
- Nakamura, F.; Ishida, Y.; Sawada, D.; Ashida, N.; Sugawara, T.; Sakai, M.; Goto, T.; Kawada, T.; Fujiwara, S. Fragmented Lactic Acid Bacterial Cells Activate Peroxisome Proliferator-Activated Receptors and Ameliorate Dyslipidemia in Obese Mice. J. Agric. Food Chem. 2016, 64, 2549–2559. [CrossRef]
- Chaluvadi, S.; Hotchkiss, A.T.; Yam, K.L. Gut Microbiota. In Probiotics, prebiotics, and synbiotics; Elsevier, 2016; pp. 515–523 ISBN 9780128021897.
- Venema, K. Beneficial microbes. Foreword. Benef. Microbes 2012, 3, 1–2. [CrossRef]
- Ozma, M.A.; Khodadadi, E.; Rezaee, M.A.; Asgharzadeh, M.; Aghazadeh, M.; Zeinalzadeh, E.; Ganbarov, K.; Kafil, H.S. Bacterial proteomics and its application in pathogenesis studies. Curr. Pharm. Biotechnol. 2022, 23, 1245–1256. [CrossRef]
- Shigwedha, N.; Sichel, L.; Jia, L.; Zhang, L. Probiotical cell fragments (pcfs) as “novel nutraceutical ingredients.” JBM 2014, 02, 43–55. [CrossRef]
- Wang, Y.; Xie, J.; Wang, N.; Li, Y.; Sun, X.; Zhang, Y.; Zhang, H. Lactobacillus casei Zhang modulate cytokine and toll-like receptor expression and beneficially regulate poly I:C-induced immune responses in RAW264.7 macrophages. Microbiol. Immunol. 2013, 57, 54–62. [CrossRef]
- Miyazawa, K.; He, F.; Kawase, M.; Kubota, A.; Yoda, K.; Hiramatsu, M. Enhancement of immunoregulatory effects of Lactobacillus gasseri TMC0356 by heat treatment and culture medium. Lett. Appl. Microbiol. 2011, 53, 210–216. [CrossRef]
- Chuang, L.; Wu, K.-G.; Pai, C.; Hsieh, P.-S.; Tsai, J.-J.; Yen, J.-H.; Lin, M.-Y. Heat-killed cells of lactobacilli skew the immune response toward T helper 1 polarization in mouse splenocytes and dendritic cell-treated T cells. J. Agric. Food Chem. 2007, 55, 11080–11086. [CrossRef]
- Sokol, H.; Pigneur, B.; Watterlot, L.; Lakhdari, O.; Bermúdez-Humarán, L.G.; Gratadoux, J.-J.; Blugeon, S.; Bridonneau, C.; Furet, J.-P.; Corthier, G.; et al. Faecalibacterium prausnitzii is an anti-inflammatory commensal bacterium identified by gut microbiota analysis of Crohn disease patients. Proc Natl Acad Sci USA 2008, 105, 16731–16736. [CrossRef]
- Tsilingiri, K.; Barbosa, T.; Penna, G.; Caprioli, F.; Sonzogni, A.; Viale, G.; Rescigno, M. Probiotic and postbiotic activity in health and disease: comparison on a novel polarised ex-vivo organ culture model. Gut 2012, 61, 1007–1015. [CrossRef]
- Haileselassie, Y.; Navis, M.; Vu, N.; Qazi, K.R.; Rethi, B.; Sverremark-Ekström, E. Postbiotic Modulation of Retinoic Acid Imprinted Mucosal-like Dendritic Cells by Probiotic Lactobacillus reuteri 17938 In Vitro. Front. Immunol. 2016, 7, 96. [CrossRef]
- Compare, D.; Rocco, A.; Coccoli, P.; Angrisani, D.; Sgamato, C.; Iovine, B.; Salvatore, U.; Nardone, G. Lactobacillus casei DG and its postbiotic reduce the inflammatory mucosal response: an ex-vivo organ culture model of post-infectious irritable bowel syndrome. BMC Gastroenterol. 2017, 17, 53. [CrossRef]
- Béghin, L.; Tims, S.; Roelofs, M.; Rougé, C.; Oozeer, R.; Rakza, T.; Chirico, G.; Roeselers, G.; Knol, J.; Rozé, J.C.; et al. Fermented infant formula (with Bifidobacterium breve C50 and Streptococcus thermophilus O65) with prebiotic oligosaccharides is safe and modulates the gut microbiota towards a microbiota closer to that of breastfed infants. Clin. Nutr. 2021, 40, 778–787. [CrossRef]
- Thibault, H.; Aubert-Jacquin, C.; Goulet, O. Effects of long-term consumption of a fermented infant formula (with Bifidobacterium breve c50 and Streptococcus thermophilus 065) on acute diarrhea in healthy infants. J. Pediatr. Gastroenterol. Nutr. 2004, 39, 147–152. [CrossRef]
- Morisset, M.; Aubert-Jacquin, C.; Soulaines, P.; Moneret-Vautrin, D.A.; Dupont, C. A non-hydrolyzed, fermented milk formula reduces digestive and respiratory events in infants at high risk of allergy. Eur. J. Clin. Nutr. 2011, 65, 175–183. [CrossRef]
- Nocerino, R.; Paparo, L.; Terrin, G.; Pezzella, V.; Amoroso, A.; Cosenza, L.; Cecere, G.; De Marco, G.; Micillo, M.; Albano, F.; et al. Cow’s milk and rice fermented with Lactobacillus paracasei CBA L74 prevent infectious diseases in children: A randomized controlled trial. Clin. Nutr. 2017, 36, 118–125. [CrossRef]
- Corsello, G.; Carta, M.; Marinello, R.; Picca, M.; De Marco, G.; Micillo, M.; Ferrara, D.; Vigneri, P.; Cecere, G.; Ferri, P.; et al. Preventive Effect of Cow’s Milk Fermented with Lactobacillus paracasei CBA L74 on Common Infectious Diseases in Children: A Multicenter Randomized Controlled Trial. Nutrients 2017, 9. [CrossRef]
- Lukovac, S.; Belzer, C.; Pellis, L.; Keijser, B.J.; de Vos, W.M.; Montijn, R.C.; Roeselers, G. Differential modulation by Akkermansia muciniphila and Faecalibacterium prausnitzii of host peripheral lipid metabolism and histone acetylation in mouse gut organoids. MBio 2014, 5. [CrossRef]
- Aguilar-Toalá, J.E.; Santiago-López, L.; Peres, C.M.; Peres, C.; Garcia, H.S.; Vallejo-Cordoba, B.; González-Córdova, A.F.; Hernández-Mendoza, A. Assessment of multifunctional activity of bioactive peptides derived from fermented milk by specific Lactobacillus plantarum strains. J. Dairy Sci. 2017, 100, 65–75. [CrossRef]
- Dronkers, T.M.G.; Ouwehand, A.C.; Rijkers, G.T. Global analysis of clinical trials with probiotics. Heliyon 2020, 6, e04467. [CrossRef]
- Critchfield, J.W.; van Hemert, S.; Ash, M.; Mulder, L.; Ashwood, P. The potential role of probiotics in the management of childhood autism spectrum disorders. Gastroenterol. Res. Pract. 2011, 2011, 161358. [CrossRef]
- Song, W.; Zhang, M.; Teng, L.; Wang, Y.; Zhu, L. Prebiotics and probiotics for autism spectrum disorder: a systematic review and meta-analysis of controlled clinical trials. J. Med. Microbiol. 2022, 71. [CrossRef]
- Depoorter, L.; Vandenplas, Y. Probiotics in pediatrics. A review and practical guide. Nutrients 2021, 13. [CrossRef]
- van den Akker, C.H.P.; van Goudoever, J.B.; Shamir, R.; Domellöf, M.; Embleton, N.D.; Hojsak, I.; Lapillonne, A.; Mihatsch, W.A.; Berni Canani, R.; Bronsky, J.; et al. Probiotics and preterm infants: A position paper by the european society for paediatric gastroenterology hepatology and nutrition committee on nutrition and the european society for paediatric gastroenterology hepatology and nutrition working group for probiotics and prebiotics. J. Pediatr. Gastroenterol. Nutr. 2020, 70, 664–680. [CrossRef]
- Su, G.L.; Ko, C.W.; Bercik, P.; Falck-Ytter, Y.; Sultan, S.; Weizman, A.V.; Morgan, R.L. AGA clinical practice guidelines on the role of probiotics in the management of gastrointestinal disorders. Gastroenterology 2020, 159, 697–705. [CrossRef]
- Weizman, Z.; Vandenplas, Y. Why do clinical guidelines for probiotics differ? Benef. Microbes 2024, 15, 411–415. [CrossRef]
- Guarner, F.; Sanders, M.E.; Szajewska, H.; Cohen, H.; Eliakim, R.; Herrera-deGuise, C.; Karakan, T.; Merenstein, D.; Piscoya, A.; Ramakrishna, B.; et al. World gastroenterology organisation global guidelines: probiotics and prebiotics. J. Clin. Gastroenterol. 2024, 58, 533–553. [CrossRef]
- Collinson, S.; Deans, A.; Padua-Zamora, A.; Gregorio, G.V.; Li, C.; Dans, L.F.; Allen, S.J. Probiotics for treating acute infectious diarrhoea. Cochrane Database Syst. Rev. 2020, 12, CD003048. [CrossRef]
- Szajewska, H.; Berni Canani, R.; Domellöf, M.; Guarino, A.; Hojsak, I.; Indrio, F.; Lo Vecchio, A.; Mihatsch, W.A.; Mosca, A.; Orel, R.; et al. ESPGHAN Special Interest Group on Gut Microbiota and Modifications Probiotics for the management of pediatric gastrointestinal disorders: position paper of the ESPGHAN special interest group on gut microbiota and modifications. J. Pediatr. Gastroenterol. Nutr. 2023, 76, 232–247. [CrossRef]
- Szajewska, H.; Kołodziej, M.; Zalewski, B.M. Systematic review with meta-analysis: Saccharomyces boulardii for treating acute gastroenteritis in children-a 2020 update. Aliment. Pharmacol. Ther. 2020, 51, 678–688. [CrossRef]
- Patro-Gołąb, B.; Szajewska, H. Systematic Review with Meta-Analysis: Lactobacillus reuteri DSM 17938 for Treating Acute Gastroenteritis in Children. An Update. Nutrients 2019, 11. [CrossRef]
- Rosenfeldt, V.; Michaelsen, K.F.; Jakobsen, M.; Larsen, C.N.; Møller, P.L.; Tvede, M.; Weyrehter, H.; Valerius, N.H.; Paerregaard, A. Effect of probiotic Lactobacillus strains on acute diarrhea in a cohort of nonhospitalized children attending day-care centers. Pediatr. Infect. Dis. J. 2002, 21, 417–419. [CrossRef]
- İşlek, A.; Sayar, E.; Yılmaz, A.; Baysan, B.Ö.; Mutlu, D.; Artan, R. The role of Bifidobacterium lactis B94 plus inulin in the treatment of acute infectious diarrhea in children. Turk. J. Gastroenterol. 2014, 25, 628–633. [CrossRef]
- Passariello, A.; Terrin, G.; Cecere, G.; Micillo, M.; De Marco, G.; Di Costanzo, M.; Cosenza, L.; Leone, L.; Nocerino, R.; Canani, R.B. Randomised clinical trial: efficacy of a new synbiotic formulation containing Lactobacillus paracasei B21060 plus arabinogalactan and xilooligosaccharides in children with acute diarrhoea. Aliment. Pharmacol. Ther. 2012, 35, 782–788. [CrossRef]
- Szymański, H.; Pejcz, J.; Jawień, M.; Chmielarczyk, A.; Strus, M.; Heczko, P.B. Treatment of acute infectious diarrhoea in infants and children with a mixture of three Lactobacillus rhamnosus strains--a randomized, double-blind, placebo-controlled trial. Aliment. Pharmacol. Ther. 2006, 23, 247–253. [CrossRef]
- Canani, R.B.; Cirillo, P.; Terrin, G.; Cesarano, L.; Spagnuolo, M.I.; De Vincenzo, A.; Albano, F.; Passariello, A.; De Marco, G.; Manguso, F.; et al. Probiotics for treatment of acute diarrhoea in children: randomised clinical trial of five different preparations. BMJ 2007, 335, 340. [CrossRef]
- Chen, K.; Xin, J.; Zhang, G.; Xie, H.; Luo, L.; Yuan, S.; Bu, Y.; Yang, X.; Ge, Y.; Liu, C. A combination of three probiotic strains for treatment of acute diarrhoea in hospitalised children: an open label, randomised controlled trial. Benef. Microbes 2020, 11, 339–346. [CrossRef]
- Guo, Q.; Goldenberg, J.Z.; Humphrey, C.; El Dib, R.; Johnston, B.C. Probiotics for the prevention of pediatric antibiotic-associated diarrhea. Cochrane Database Syst. Rev. 2019, 4, CD004827. [CrossRef]
- Szajewska, H.; Canani, R.B.; Guarino, A.; Hojsak, I.; Indrio, F.; Kolacek, S.; Orel, R.; Shamir, R.; Vandenplas, Y.; van Goudoever, J.B.; et al. Probiotics for the Prevention of Antibiotic-Associated Diarrhea in Children. J. Pediatr. Gastroenterol. Nutr. 2016, 62, 495–506. [CrossRef]
- Szajewska, H.; Kołodziej, M. Systematic review with meta-analysis: Lactobacillus rhamnosus GG in the prevention of antibiotic-associated diarrhoea in children and adults. Aliment. Pharmacol. Ther. 2015, 42, 1149–1157. [CrossRef]
- Lukasik, J.; Dierikx, T.; Besseling-van der Vaart, I.; de Meij, T.; Szajewska, H.; Multispecies Probiotic in AAD Study Group Multispecies Probiotic for the Prevention of Antibiotic-Associated Diarrhea in Children: A Randomized Clinical Trial. JAMA Pediatr. 2022, 176, 860–866. [CrossRef]
- Ruszczyński, M.; Radzikowski, A.; Szajewska, H. Clinical trial: effectiveness of Lactobacillus rhamnosus (strains E/N, Oxy and Pen) in the prevention of antibiotic-associated diarrhoea in children. Aliment. Pharmacol. Ther. 2008, 28, 154–161. [CrossRef]
- Szajewska, H.; Wanke, M.; Patro, B. Meta-analysis: the effects of Lactobacillus rhamnosus GG supplementation for the prevention of healthcare-associated diarrhoea in children. Aliment. Pharmacol. Ther. 2011, 34, 1079–1087. [CrossRef]
- Hojsak, I.; Szajewska, H.; Canani, R.B.; Guarino, A.; Indrio, F.; Kolacek, S.; Orel, R.; Shamir, R.; Vandenplas, Y.; van Goudoever, J.B.; et al. Probiotics for the prevention of nosocomial diarrhea in children. J. Pediatr. Gastroenterol. Nutr. 2018, 66, 3–9. [CrossRef]
- Beghetti, I.; Panizza, D.; Lenzi, J.; Gori, D.; Martini, S.; Corvaglia, L.; Aceti, A. Probiotics for Preventing Necrotizing Enterocolitis in Preterm Infants: A Network Meta-Analysis. Nutrients 2021, 13. [CrossRef]
- Chi, C.; Li, C.; Buys, N.; Wang, W.; Yin, C.; Sun, J. Effects of Probiotics in Preterm Infants: A Network Meta-analysis. Pediatrics 2021, 147. [CrossRef]
- Gao, X.; Wang, Y.; Shi, L.; Feng, W.; Yi, K. Effect and Safety of Saccharomyces boulardii for Neonatal Necrotizing Enterocolitis in Pre-term Infants: A Systematic Review and Meta-Analysis. J. Trop. Pediatr. 2021, 67. [CrossRef]
- van den Akker, C.H.P.; van Goudoever, J.B.; Szajewska, H.; Embleton, N.D.; Hojsak, I.; Reid, D.; Shamir, R.; ESPGHAN Working Group for Probiotics, Prebiotics & Committee on Nutrition Probiotics for Preterm Infants: A Strain-Specific Systematic Review and Network Meta-analysis. J. Pediatr. Gastroenterol. Nutr. 2018, 67, 103–122. [CrossRef]
- Athalye-Jape, G.; Rao, S.; Patole, S. Lactobacillus reuteri DSM 17938 as a Probiotic for Preterm Neonates: A Strain-Specific Systematic Review. JPEN J Parenter Enteral Nutr 2016, 40, 783–794. [CrossRef]
- Lin, H.-C.; Su, B.-H.; Chen, A.-C.; Lin, T.-W.; Tsai, C.-H.; Yeh, T.-F.; Oh, W. Oral probiotics reduce the incidence and severity of necrotizing enterocolitis in very low birth weight infants. Pediatrics 2005, 115, 1–4. [CrossRef]
- Feng, J.-R.; Wang, F.; Qiu, X.; McFarland, L.V.; Chen, P.-F.; Zhou, R.; Liu, J.; Zhao, Q.; Li, J. Efficacy and safety of probiotic-supplemented triple therapy for eradication of Helicobacter pylori in children: a systematic review and network meta-analysis. Eur. J. Clin. Pharmacol. 2017, 73, 1199–1208. [CrossRef]
- Wen, J.; Peng, P.; Chen, P.; Zeng, L.; Pan, Q.; Wei, W.; He, J. Probiotics in 14-day triple therapy for Asian pediatric patients with Helicobacter pylori infection: a network meta-analysis. Oncotarget 2017, 8, 96409–96418. [CrossRef]
- Zhou, B.-G.; Chen, L.-X.; Li, B.; Wan, L.-Y.; Ai, Y.-W. Saccharomyces boulardii as an adjuvant therapy for Helicobacter pylori eradication: A systematic review and meta-analysis with trial sequential analysis. Helicobacter 2019, 24, e12651. [CrossRef]
- Szajewska, H.; Horvath, A.; Kołodziej, M. Systematic review with meta-analysis: Saccharomyces boulardii supplementation and eradication of Helicobacter pylori infection. Aliment. Pharmacol. Ther. 2015, 41, 1237–1245. [CrossRef]
- Fang, H.-R.; Zhang, G.-Q.; Cheng, J.-Y.; Li, Z.-Y. Efficacy of Lactobacillus-supplemented triple therapy for Helicobacter pylori infection in children: a meta-analysis of randomized controlled trials. Eur. J. Pediatr. 2019, 178, 7–16. [CrossRef]
- Hurduc, V.; Plesca, D.; Dragomir, D.; Sajin, M.; Vandenplas, Y. A randomized, open trial evaluating the effect of Saccharomyces boulardii on the eradication rate of Helicobacter pylori infection in children. Acta Paediatr. 2009, 98, 127–131. [CrossRef]
- Bin, Z.; Ya-Zheng, X.; Zhao-Hui, D.; Bo, C.; Li-Rong, J.; Vandenplas, Y. The Efficacy of Saccharomyces boulardii CNCM I-745 in Addition to Standard Helicobacter pylori Eradication Treatment in Children. Pediatr. Gastroenterol. Hepatol. Nutr. 2015, 18, 17–22. [CrossRef]
- Sung, V.; D’Amico, F.; Cabana, M.D.; Chau, K.; Koren, G.; Savino, F.; Szajewska, H.; Deshpande, G.; Dupont, C.; Indrio, F.; et al. Lactobacillus reuteri to Treat Infant Colic: A Meta-analysis. Pediatrics 2018, 141. [CrossRef]
- Skonieczna-Żydecka, K.; Janda, K.; Kaczmarczyk, M.; Marlicz, W.; Łoniewski, I.; Łoniewska, B. The Effect of Probiotics on Symptoms, Gut Microbiota and Inflammatory Markers in Infantile Colic: A Systematic Review, Meta-Analysis and Meta-Regression of Randomized Controlled Trials. J. Clin. Med. 2020, 9. [CrossRef]
- Ong, T.G.; Gordon, M.; Banks, S.S.; Thomas, M.R.; Akobeng, A.K. Probiotics to prevent infantile colic. Cochrane Database Syst. Rev. 2019, 3, CD012473. [CrossRef]
- Dryl, R.; Szajewska, H. Probiotics for management of infantile colic: a systematic review of randomized controlled trials. Arch. Med. Sci. 2018, 14, 1137–1143. [CrossRef]
- Gutiérrez-Castrellón, P.; Indrio, F.; Bolio-Galvis, A.; Jiménez-Gutiérrez, C.; Jimenez-Escobar, I.; López-Velázquez, G. Efficacy of Lactobacillus reuteri DSM 17938 for infantile colic: Systematic review with network meta-analysis. Medicine (Baltimore) 2017, 96, e9375. [CrossRef]
- Schreck Bird, A.; Gregory, P.J.; Jalloh, M.A.; Risoldi Cochrane, Z.; Hein, D.J. Probiotics for the treatment of infantile colic: A systematic review. J. Pharm. Pract. 2017, 30, 366–374. [CrossRef]
- Harb, T.; Matsuyama, M.; David, M.; Hill, R.J. Infant Colic-What works: A Systematic Review of Interventions for Breast-fed Infants. J. Pediatr. Gastroenterol. Nutr. 2016, 62, 668–686. [CrossRef]
- Szajewska, H.; Gyrczuk, E.; Horvath, A. Lactobacillus reuteri DSM 17938 for the management of infantile colic in breastfed infants: a randomized, double-blind, placebo-controlled trial. J. Pediatr. 2013, 162, 257–262. [CrossRef]
- Nocerino, R.; De Filippis, F.; Cecere, G.; Marino, A.; Micillo, M.; Di Scala, C.; de Caro, C.; Calignano, A.; Bruno, C.; Paparo, L.; et al. The therapeutic efficacy of Bifidobacterium animalis subsp. lactis BB-12® in infant colic: A randomised, double blind, placebo-controlled trial. Aliment. Pharmacol. Ther. 2020, 51, 110–120. [CrossRef]
- Chen, K.; Zhang, G.; Xie, H.; You, L.; Li, H.; Zhang, Y.; Du, C.; Xu, S.; Melsaether, C.; Yuan, S. Efficacy of Bifidobacterium animalis subsp. lactis, BB-12® on infant colic—a randomised, double-blinded, placebo-controlled study. Benef. Microbes 2021, 12, 531–540. [CrossRef]
- Gerasimov, S.; Gantzel, J.; Dementieva, N.; Schevchenko, O.; Tsitsura, O.; Guta, N.; Bobyk, V.; Kaprus, V. Role of Lactobacillus rhamnosus (FloraActiveTM) 19070-2 and Lactobacillus reuteri (FloraActiveTM) 12246 in Infant Colic: A Randomized Dietary Study. Nutrients 2018, 10. [CrossRef]
- Baldassarre, M.E.; Di Mauro, A.; Tafuri, S.; Rizzo, V.; Gallone, M.S.; Mastromarino, P.; Capobianco, D.; Laghi, L.; Zhu, C.; Capozza, M.; et al Effectiveness and Safety of a Probiotic-Mixture for the Treatment of Infantile Colic: A Double-Blind, Randomized, Placebo-Controlled Clinical Trial with Fecal Real-Time PCR and NMR-Based Metabolomics Analysis. Nutrients 2018, 10. [CrossRef]
- Indrio, F.; Di Mauro, A.; Riezzo, G.; Civardi, E.; Intini, C.; Corvaglia, L.; Ballardini, E.; Bisceglia, M.; Cinquetti, M.; Brazzoduro, E.; et al. Prophylactic use of a probiotic in the prevention of colic, regurgitation, and functional constipation: a randomized clinical trial. JAMA Pediatr. 2014, 168, 228–233. [CrossRef]
- Trivić, I.; Niseteo, T.; Jadrešin, O.; Hojsak, I. Use of probiotics in the treatment of functional abdominal pain in children-systematic review and meta-analysis. Eur. J. Pediatr. 2021, 180, 339–351. [CrossRef]
- Weizman, Z.; Abu-Abed, J.; Binsztok, M. Lactobacillus reuteri DSM 17938 for the Management of Functional Abdominal Pain in Childhood: A Randomized, Double-Blind, Placebo-Controlled Trial. J. Pediatr. 2016, 174, 160-164.e1. [CrossRef]
- Gawrońska, A.; Dziechciarz, P.; Horvath, A.; Szajewska, H. A randomized double-blind placebo-controlled trial of Lactobacillus GG for abdominal pain disorders in children. Aliment. Pharmacol. Ther. 2007, 25, 177–184. [CrossRef]
- Kaur, L.; Gordon, M.; Baines, P.A.; Iheozor-Ejiofor, Z.; Sinopoulou, V.; Akobeng, A.K. Probiotics for induction of remission in ulcerative colitis. Cochrane Database Syst. Rev. 2020, 3, CD005573. [CrossRef]
- Miele, E.; Pascarella, F.; Giannetti, E.; Quaglietta, L.; Baldassano, R.N.; Staiano, A. Effect of a probiotic preparation (VSL#3) on induction and maintenance of remission in children with ulcerative colitis. Am. J. Gastroenterol. 2009, 104, 437–443. [CrossRef]
- Mimura, T.; Rizzello, F.; Helwig, U.; Poggioli, G.; Schreiber, S.; Talbot, I.C.; Nicholls, R.J.; Gionchetti, P.; Campieri, M.; Kamm, M.A. Once daily high dose probiotic therapy (VSL#3) for maintaining remission in recurrent or refractory pouchitis. Gut 2004, 53, 108–114.
- Gionchetti, P.; Rizzello, F.; Venturi, A.; Brigidi, P.; Matteuzzi, D.; Bazzocchi, G.; Poggioli, G.; Miglioli, M.; Campieri, M. Oral bacteriotherapy as maintenance treatment in patients with chronic pouchitis: a double-blind, placebo-controlled trial. Gastroenterology 2000, 119, 305–309. [CrossRef]
- Blanchetière, A.; Dolladille, C.; Goyer, I.; Join-Lambert, O.; Fazilleau, L. State of the Art of Probiotic Use in Neonatal Intensive Care Units in French-Speaking European Countries. Children 2023, 10. [CrossRef]
- Hempel, S.; Newberry, S.; Ruelaz, A.; Wang, Z.; Miles, J.N.V.; Suttorp, M.J.; Johnsen, B.; Shanman, R.; Slusser, W.; Fu, N.; et al. Safety of probiotics used to reduce risk and prevent or treat disease. Evid Rep Technol Assess (Full Rep) 2011, 1–645.
- Zhao, F.; Bai, X.; Zhang, J.; Kwok, L.-Y.; Shen, L.; Jin, H.; Sun, T.; Sun, Z.; Zhang, H. Gut Bifidobacterium responses to probiotic Lactobacillus casei Zhang administration vary between subjects from different geographic regions. Appl. Microbiol. Biotechnol. 2022, 106, 2665–2675. [CrossRef]
Figure 1.
Role of metabolites in the adaptation of probiotic bacteria within the gut environment. This figure depicts how probiotic metabolites benefit both the bacteria and the host: 1. Symbiotic Interaction, 2. Self-Protection and 3. Pathogen Inhibition. EPS: Exopolysaccharides. SCFAs: Short chain fatty acids. pH: Potential of hydrogen. ATP: adenosine triphosphate.
Figure 1.
Role of metabolites in the adaptation of probiotic bacteria within the gut environment. This figure depicts how probiotic metabolites benefit both the bacteria and the host: 1. Symbiotic Interaction, 2. Self-Protection and 3. Pathogen Inhibition. EPS: Exopolysaccharides. SCFAs: Short chain fatty acids. pH: Potential of hydrogen. ATP: adenosine triphosphate.
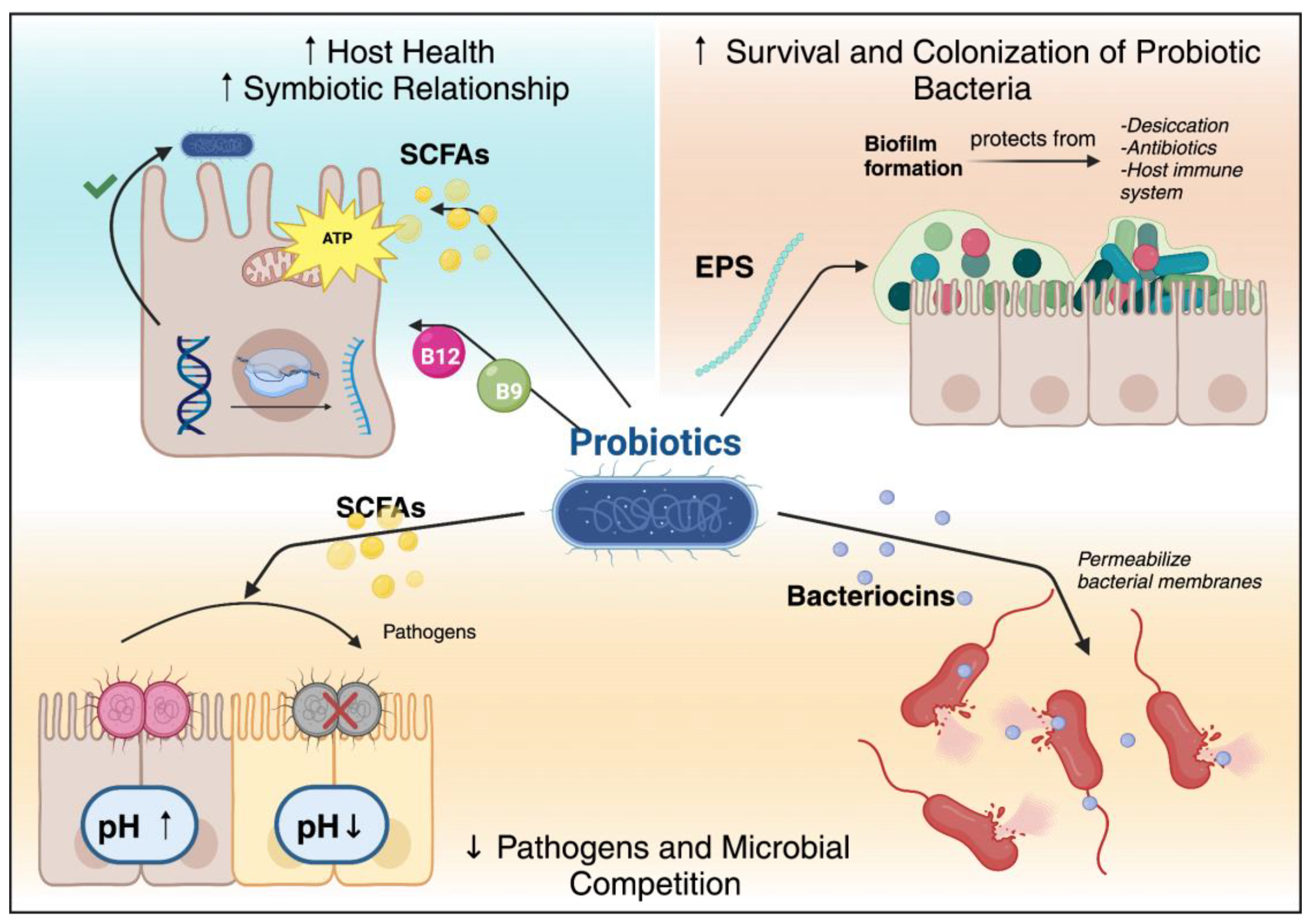
Figure 2.
Role of probiotic metabolites in children’s health. Short-chain fatty acids (SCFAs), exopolysaccharides (EPS), Immunoglobulin A (IgA), interleukin-10 (IL-10), and interferon-gamma (IFN-γ) in immune cells. Nuclear factor kappa B (NF-κB) activation, leading to reduced levels of interleukin-6 (IL-6) and interleukin-8 (IL-8).
Figure 2.
Role of probiotic metabolites in children’s health. Short-chain fatty acids (SCFAs), exopolysaccharides (EPS), Immunoglobulin A (IgA), interleukin-10 (IL-10), and interferon-gamma (IFN-γ) in immune cells. Nuclear factor kappa B (NF-κB) activation, leading to reduced levels of interleukin-6 (IL-6) and interleukin-8 (IL-8).
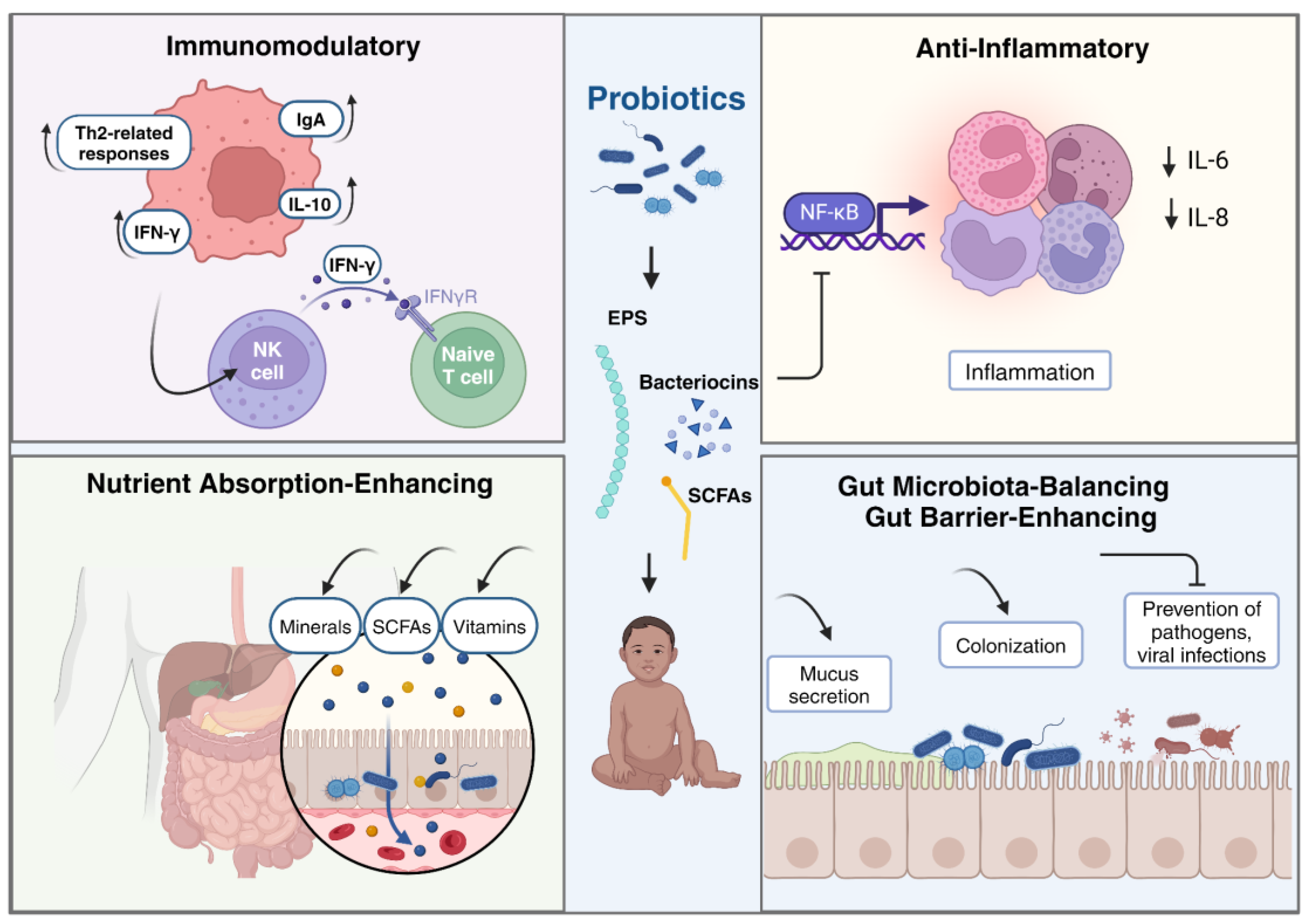
Figure 3.
Major metabolites produced by probiotic bacteria in children.SCFAs: Short-chain Fatty acids. EPS: Exopolysaccharides. GABA: Gamma-Aminobutyric Acid. AMPS: Antimicrobial Peptides.
Figure 3.
Major metabolites produced by probiotic bacteria in children.SCFAs: Short-chain Fatty acids. EPS: Exopolysaccharides. GABA: Gamma-Aminobutyric Acid. AMPS: Antimicrobial Peptides.
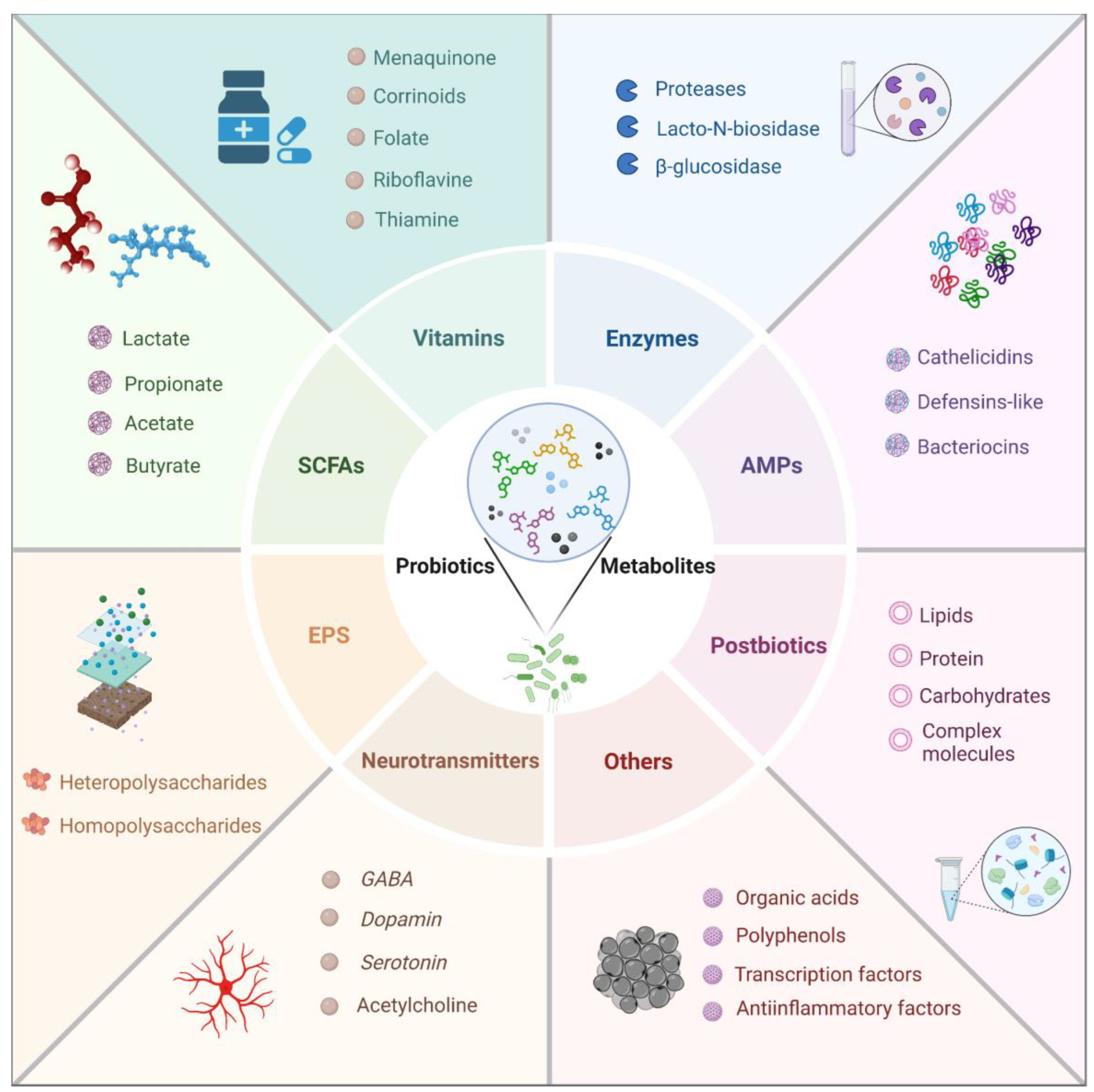
Figure 4.
Postbiotics and their potential positive effects in children.
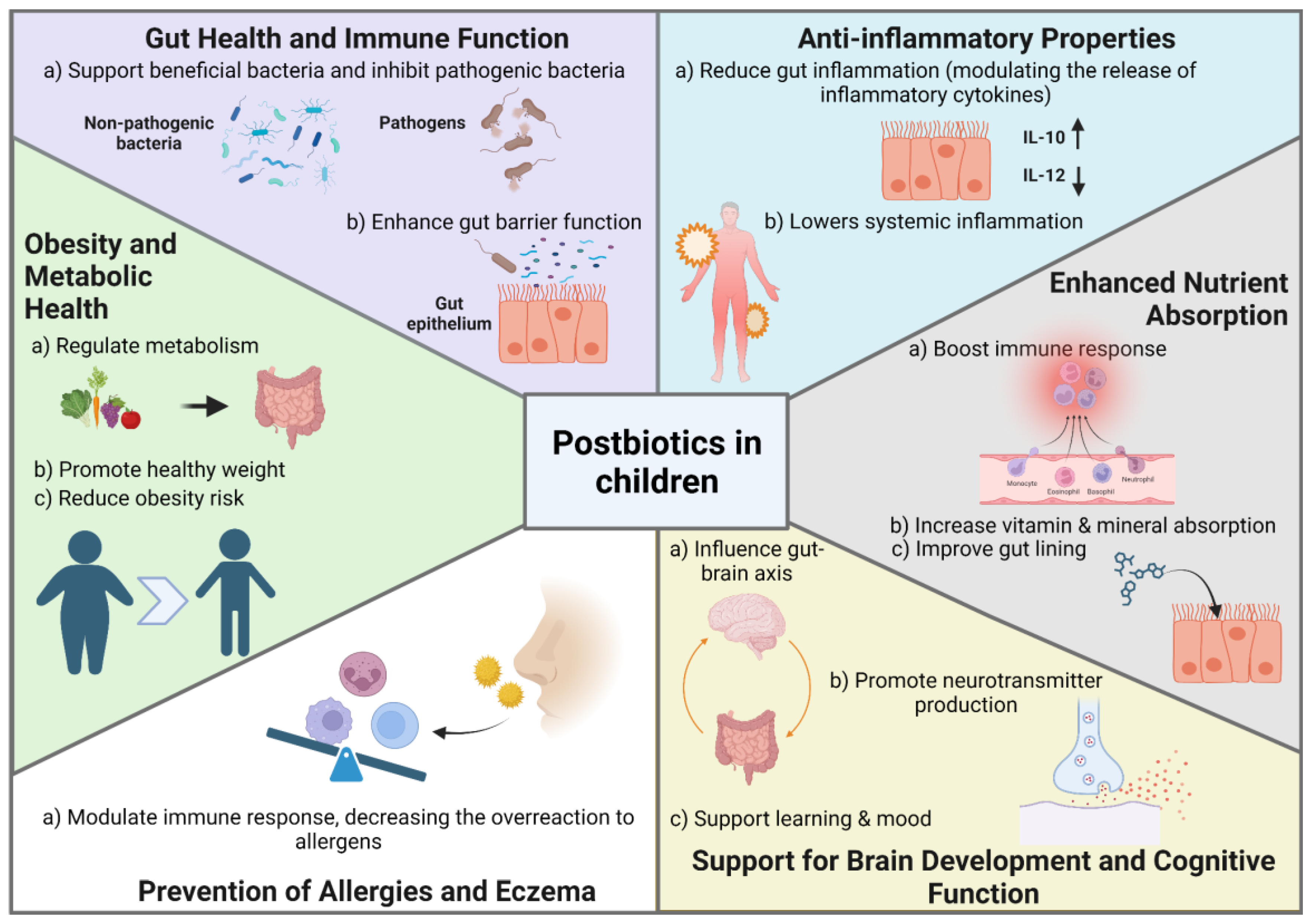
Table 1.
Key vitamins produced by probiotic bacteria.
| Probiotic Strain | Vitamin Produced | Mechanism |
|---|---|---|
| Lactobacillus fermentum [126] | Folate (B9), Vitamin B12 (Cobalamin) | Synthesis of folate and B12 |
| Lactobacillus reuteri (DCM 20016, JCM1112, CRL1324, CRL1327) [31] | Corrinoids (related to Vitamin B12) | Production of corrinoids. |
| Lactobacillus acidophilus (ATCC314, FTDC 8833) [125] | Riboflavin (B2) | Enhance riboflavin production |
| Streptococcus thermophilus (ABM5097) [15] | 5-Methyltetrahydrofolate (5-MTHF) (Folate) | Increase folate production. |
| Lactobacillus rhamnosus GG [31] | Folate (B9), Riboflavin (B2), Thiamine (B1) | Production and releases folate and riboflavin efficiently; low production of intracellular thiamine. |
| Bacillus clausii [135] | Vitamin K2 (Menaquinone) | Production of vitamin K2 |
| Lactococcus lactis [136] | Vitamin K2 (Menaquinone) | Production of vitamin K2 |
| Enterobacter agglomerans, Serratia marcescens, Enterococcus faecium [137] | Menaquinones (Vitamin K2) | Contribute to vitamin K production in the neonatal gut. |
| Various strains (Lactobacillus, Bifidobacterium) [127] | B Vitamins (B1, B2, B3, B5, B6, B7, B9, B12), Vitamin K | Utilize oligosaccharides to enhance hydrophobicity, auto-aggregation, and biofilm formation, improving B vitamin production. |
| Lactobacillus gasseri (FTDC 8131) [139] | Riboflavin (B2) | Interacts with riboflavin; context suggests strain-dependent variability in production or consumption of the vitamin. |
| Bifidobacterium strains (B. longum, B. bifidum) [31] | Thiamine (B1) | Low but significant production of intracellular thiamine without extracellular synthesis; do not produce folates or riboflavin. |
| Children consuming probiotics [102] | Vitamin D, Vitamin A | Probiotics enhance absorption and serum concentrations of vitamins. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



