
Submitted:
01 October 2024
Posted:
02 October 2024
You are already at the latest version
Abstract
The membrane dipole potential that arises from the interfacial water and constitutive dipolar groups of lipid molecules modulates the interaction of amphiphiles and proteins with membranes. Consequently, its determination for lipid mixtures resembling the existing diversity in biological membranes is very relevant. In this work the dipole potential of monolayers, formed at the air-water interface, from pure or mixed lipids (1-palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine (POPC), 1-palmitoyl-2-oleoyl-sn-glycero-3-phosphoethanolamine (POPE), 1-palmitoyl-2-ol0eoyl-sn-glycero-3-phosphatidyserine (POPS), Sphingomyelin (SpM) and cholesterol) were measured and correlated with the mean area per lipid. The results showed that, as previously observed, cholesterol increases the dipole potential in correspondence with the decrease in the average area per lipid and that the replacement of phosphocholine lipids by sphingomyelin leads to a decrease in the dipole potential. We have also observed that 20 mol% of POPE induces a monolayer condensation which is not affecting the measured dipole potential, except when cholesterol is added to the mixture. Additionally, at the small mole fractions encountered in biomembranes, the presence of the negatively charged lipid POPS increases the dipole potential of monolayers even though an enhancement in the monolayer packing is not evident. This increase in the dipole potential due to POPS and cholesterol and the smaller dipole potential of monolayers containing large amounts of SpM generates a non-zero transbilayer dipole potential in the asymmetric plasma membrane of eukaryotic cells.
Keywords:
Dipole Potential
; Cholesterol
; POPC
; POPE
; POPS
; Surface Pressure
; Monolayers
; Area per Lipid
1. Introduction
The total electric profile of a membrane may be defined by three distinct types of electrostatic potential: the transmembrane potential, the surface charge potential and the dipole potential [1,2,3,4]. The first is a consequence of the charge displacement from one side to the other of a membrane, due to a difference in ion concentrations between both aqueous compartments. The surface charge potential, which is described by the Gouy-Chapman theory, results from the presence of charged groups in the lipids and ion distribution in the electrical double layer on the membrane surface. Third, but not less important, is originated from the preferred alignment of water molecules and certain constitutive dipoles of lipids. The dipole potential was first described by Liberman and Topaly in 1969, through membrane conductivity changes upon addition of large hydrophobic anions and cations (TPPB-, TPPB+). They observed that the membrane conductivity was much larger for the hydrophobic anions than for the cations. This difference was attributed to their distinct partition into the center of the membrane and they assumed that the presence of a more positive potential within the membrane was the main reason for the permeability differences [5]. The main sources contributing to the membrane internal dipole potential are: (i) the lipid carbonyl groups, (ii) the dipole resulting from the phospholipid headgroup (choline-phosphate dipole in the case of phosphatidylcholines), (iii) the terminal methyl group in the lipid alkyl chain and (iv) interfacial water hydrating the phospholipid headgroups [1,2,3,4,6,7,8,9,10]. Studies in monolayers with phosphatidylcholines revealed a dipole potential of approximately 400 mV. This potential generates a considerably higher membrane electric field than that of the surface charge or the transmembrane potentials, suggesting that it has an important biological role. In fact, the binding affinity or the orientation of a membrane peptide was shown to be affected by the dipole potential [11,12]. We have recently demonstrated that the dipole potential exerts an important influence on the association of amphiphiles with ordered lipid membranes, changing their membrane-affinity and transversal location, with effects on amphiphile aggregation and ionization [13]. The effect of the dipole potential on the permeability of charged molecules was at the basis of its identification, and has been observed by several authors [14,15]. Molecular dynamics simulations have shown that this effect is relevant even for uncharged polar molecules, with the dipole moment of the permeating molecule re-orienting as it moves across the bilayer [16]. The dipole potential has also been recognized as a modulator on the activity of membrane embedded peptides [17] and on the kinetics of charge transfer reactions in reaction centers [18]. It was shown by Monte Carlo simulations that the dipole potential is also an important parameter in phase miscibility and topology [19].
The dipole potential of lipid bilayers, between the membrane/water interface and the bilayer center, cannot be measured directly and this has hindered its characterization. Its effect on the relative permeability of hydrophobic cations and anions has been the classical methodology followed, but it is tedious and not free from artifacts. More recently, several fluorescent probes which are sensitive to the local electrical field have been developed [20], and cryoelectron microscopy (cryoEM) has been used to characterize the dipole potential profile in rapidly frozen liposomes [21]. However, the use of fluorescent probes to quantitatively characterize the dipole potential relies on calibration with other methodologies and the interpretation of cryoEM rests on several assumptions (further details may be found in references [22,23]).
The simplest and commonly used methodology to quantitatively characterize the dipole potential is the use lipid monolayers. Briefly, it consists in measuring the potential difference across an air/water interface, first in the absence and then in the presence of a formed lipid monolayer [6,24].
Results from monolayer studies and from the relative permeability of hydrophobic ions have shown that the dipole potential can be changed through the addition of certain dipolar molecules such as phloretin [25] , cholesterol and its analogue 6-ketocholestanol [14,26,27,28]. Sterols are present in large quantities in the biological membranes of eukaryotic cells and the effect of cholesterol in the membrane dipole moment was first addressed by Szabo in 1974 [14]. Using hydrophobic ions as molecular probes it was observed that upon the addition of cholesterol to a neutral membrane there was a 30-fold increase in anion permeability, compared to a 100-fold decrease in cation permeability. This distinct behavior for anion and cation was compatible with an increase in dipole potential of cholesterol-containing membranes. Later this observation was confirmed through measurements of the dipole potential of cholesterol-containing Egg Phosphatadylcholine (EggPC) monolayers [28]. The main reason why cholesterol increases the membrane potential is by enhancing the lipid packing in the membrane, the so-called cholesterol condensation effect [29], although changes in the alignment of interfacial water are also involved [30].
Biological membranes are composed of several types of lipids whose relative proportions vary significantly among distinct cells and within cell membranes. Additionally, the distribution of the lipids in both bilayer leaflets is not homogeneous, depending on the chemical environment that is in direct contact with each monolayer [31] and on the asymmetric synthesis and active translocation by membrane lipids and proteins [32]. Typically, the erythrocyte membrane contains 30-50% cholesterol (Chol), 15-20% sphingomyelin (SpM) and 40-50% glycerophospholipids; being 20-25% phosphatidylcholines (PC), 10-25% phosphatidylethanolamines (PE) and 5-15% of phosphatidylserine (PS) [32,33,34,35,36]. While SpM, PC and Chol are the major components of the outer exoplasmic leaflet of the plasma membranes in eukaryotic cells, the composition of the inner leaflet is mainly PC, PE and PS [32,33,37].
The dipole potential of phosphatidylcholines, sphingomyelin and their mixtures with cholesterol has been a subject widely discussed in literature, to get insights on the sources contributing to the dipole potential [38,39,40,41,42,43]. The quantification of the membrane dipole potential with different lipid compositions is also important to understand its role in the partition and permeation of amphiphilic molecules, especially those having a preferential dipole moment orientation. In the last years, our group has characterized the interaction of different amphiphiles and drugs with membranes of distinct lipid composition [44,45,46,47,48,49], and the magnitude of the dipole potential stands up as a membrane property that may play a crucial role [13,50]. Moreover, changes in the membrane dipole potential have been proposed to explain some unspecific drug effects [51,52,53,54,55,56,57]. In this respect, the different dipole potential of distinct biomembranes influence the drug-membrane affinity and localization, and the presence of the drug affects membrane’s dipole potential with impact on the structure and function of membrane-associated proteins [58,59].
In spite of its importance, the dipole potential of biologically relevant lipid mixtures has not been systematically addressed in literature. In this work we fulfill this gap with the characterization of the effects of the head groups PC, PE and PS, at biologically relevant molar fractions, as well as their mixtures with cholesterol. The systems used are monolayers formed at the air-water interface, allowing the direct characterization of the dipole potential.
2. Results
2.1. Area per Lipid in Monolayers
The area per lipid allows to quantitatively evaluate differences in lipid packing within a monolayer and to correlate it with the membrane dipole potential produced by mixing different lipids. Moreover, together with the results of dipole potential, it allows the rationalization of the effect (condensation/expansion/reorientation) of different lipids in more complex lipid mixtures. In this work we have calculated the area per lipid from the changes in the surface pressure as a function of the amount of lipid added to the air/water interface (Figure 1). The chosen lipid compositions mimic the lipid bilayer of distinct biological membranes of eukaryotic cells (which data is not available in the literature) as well as simpler mixtures required for the interpretation of the final results. As a reference we have also measured the area per lipid in the well characterized monolayer of pure POPC.
The mean area per lipid values most commonly reported in literature are those at a surface pressure of 30 mN/m which is assumed to be equivalent to a bilayer model [60]. In order to analyze our results comparing with previously reported data, a polynomial equation was fitted to the experimental π-area isotherms, for areas smaller than 100 Å2 (liquid expanded/condensed state) (Figure 2), and the area per lipid at a surface pressure of 30 mN/m was calculated, Table 1. The π-area isotherm profile and the area per lipid obtained for pure POPC at πsat and 30 mN/m are similar to values obtained experimentally [24,61,62] and by molecular dynamics simulations [63]. The addition of 20 mol% POPE changes the π-area isotherm to smaller areas, decreasing the area per lipid, both at 30 mN/m and at the saturation pressure, in agreement with the smaller area observed for pure POPE monolayers [24]. In the presence of 10% of POPS, the π-area isotherm is shifted both to larger areas and pressures, leading to a significantly larger area per lipid at 30 mN/m. It should however be noted that at the saturation pressure the area per lipid is similar to that of pure POPC, indicating that the electrostatic repulsion between the negatively charged serine groups is somehow compensated by lipid-lipid interactions at pressures near saturation. The replacement of 30 mol% POPC by cholesterol in the POPC:POPE (8:2) monolayer leads to a significant decrease in the area at 30 mN/m but little effect at saturation. This is in contrast with the effect observed in pure POPC where the monolayer is condensed at all pressures due to the presence of cholesterol. Finally, the quaternary mixture, that mimics the inner leaflet of plasma membrane, showed an area per lipid equal to that of pure POPC monolayers at saturation and slightly smaller at 30 mN/m. The comparison of the π-area isotherm profiles of POPC:Chol:POPE and POPC:Chol:POPE:POPS shows that the inclusion of the charged lipid shifts the curve to larger areas and pressures. This suggests that the condensation effect induced by cholesterol and POPE is compensated by the electrostatic repulsion between the charged POPS.
2.2. Dipole Potential in Lipid Monolayers
The magnitude of the dipole potential depends on the orientation and density of dipoles present in lipid monolayers and this parameter is important for understanding the lipid organization in bilayers [67], as well as the interactions established and the function of membrane-associated molecules [13,53,57]. The comprehension of the dipole potential variation in biologically relevant lipid mixtures requires a primary analysis of less complex mixtures and pure lipids. Consequently, the dipole potential of pure POPC, POPE, SpM and POPS in monolayers formed at the air-water interface was obtained first (Figure 3). In agreement with other results with negatively charged lipids [66,68], the measured potential for POPS is lower than the dipole potential due to the surface potential (equation 2) and for those monolayers we have estimated the surface dipole potential using the Gouy-Chapman theory (equation 3). The contribution of the surface potential to the measured potential in pure POPS monolayers is 135 mV, considering an area similar to POPC (49.4 Å2) and a formal charge of -1 per lipid which generates a surface charge density (σ) of -3.1 C/m2 [69].
The results show, in agreement with other monolayer studies [24] and molecular dynamics simulations [70], that ethanolamine and serine headgroups promote an increase in the dipole potential when compared to choline. As observed previously, the dipole potential of SpM monolayers is significantly smaller than that of POPC despite the same phosphate-choline head group [71].
The increase in dipole potential in POPE and POPS monolayers is in parallel with an increase in the lipid packing. In contrast, a much smaller dipole potential is obtained for SpM despite a higher lipid packing as observed by other authors [72,73]. SpM alkyl chain motional constraints and its ability to establish a network of hydrogen bonds between headgroups increases the lipid packing in these membranes [74,75,76]. However, this increase in packing does not lead to an increase in dipole potential. Thus, the dipole potential is being influenced by structural differences due to the presence of an OH in SpM instead of the carbonyl groups in POPC and/or due to the interfacial hydration water layer.
The addition of increasing percentages of cholesterol to POPC increases the dipole potential from 417 mV to ~490 mV, at 50% cholesterol, which has been attributed to the increase in membrane lipid packing in those membranes [65,72] (Figure 4). This dipole potential variation (17 %) is in agreement with the observed difference for EggPC:Cholesterol monolayer at equimolar concentrations [28]. The addition of 40 mol% cholesterol to SpM leads to a 20 % increase in the dipole potential, in agreement with the 26 % increase observed in brain SpM at 50 mol% cholesterol [71]. However, the larger increase observed in the dipolar potential of SpM:Chol monolayers is not sufficient to approach the dipole potential of POPC:Chol monolayers (at 30 or 50 mol% cholesterol). It is noteworthy that despite the distinct physical state of pure POPC and SpM monolayers (liquid disordered and gel, respectively), and the corresponding much larger area per lipid of the former, the area per lipid in POPC and SpM monolayers saturated with cholesterol is similar. The dipole potential of the cholesterol saturated monolayers is however much smaller in SpM:Chol.
The addition of biologically relevant amounts of POPS (10%) and POPE (20%) to POPC leads to a small increase in the dipole potential. Surprisingly, in spite of the much larger dipole potential of pure POPE and the opposite variations in the mean area per lipid, the increase due to the addition of 20 mol% POPE is smaller than that generated by the addition of 10 mol% POPS. This can only be understood in terms of a larger dipole moment of POPS when present at small molar fractions in POPC mixed monolayers as compared with pure POPS monolayers, and is in agreement with an increase in the ionization constant of POPS at high surface charge densities [77,78].
Upon the addition of POPE (20%) and in the presence of cholesterol there is a dipole potential increase of approximately 4% from 463 mV in POPC:CHOL (7:3) to 482 mV in POPC:CHOL:POPE (5:3:2). The replacement of 10 mol% POPC by POPS in this ternary mixture, representing the inner leaflet of plasma membrane, leads to a small decrease in the dipole potential, from 482 to 474 mV. This is surprising in face of the increase in dipole potential observed at 10 mol% POPS in POPC, and raises questions regarding the lateral homogeneity of this lipid composition.
The dipole potentials shown in Table 1 and discussed above correspond to monolayers at the saturation pressure. Although this is the condition at which the lipids in the monolayer are in equilibrium with bilayers in the aqueous phase, it is generally assumed that the properties of the bilayers are better described by monolayers at smaller lateral pressures. This is supported by comparisons of the mean area per lipid, which are smaller in monolayers at the saturation pressure than in bilayers, and by studies of partition into monolayers and bilayers that show that partition into densely packed monolayers is hindered [79]. Comparisons of the main phase transition temperature in bilayers and monolayers at different lateral pressures have shown, using the method of monolayer compression, that the lateral pressure of monolayers at which they behave as bilayers is dependent on the lipid composition but usually smaller than the saturation pressure [80]. It has however been shown that a good correspondence between the temperature of the main phase transition in bilayers and in monolayers at the saturation pressure is obtained when the constant area method is followed [67], as in this work. Some controversy notwithstanding, it is usually considered that monolayers at a lateral pressure of 30 mN/m [60,81] are good models for bilayers [80].
Given that at the saturation pressure and at 30mN/m the lipid monolayer is in the same phase, the orientation of the lipids should be maintained and only the density of the dipoles contributes to the observed changes in the dipole potential. According to this assumption, the dipole potential at 30 mN/m () was calculated using the Helmholtz equation for a parallel-plate capacitor [24], equation 1,
where ∆V is the dipole potential variation through the hydrated lipid monolayer (mV), A is the mean area per lipid (Å2) and μ⊥ is the component of the molecular dipole moment perpendicular to the lipid-water interface (mD).
The surface potential at a lateral pressure of 30 mN/m calculated from equation 1, , is represented in Figure 5 as a function of the surface potential measured at the saturation lateral pressure, .
A positive relation between the dipole potential at the two lateral pressures is observed, but the correlation is poor (R2=0.54). This results from the distinct area variations (at vs 30 mN/m) for the different lipid compositions. For the monolayers that present a small area variation at lateral pressures above 30 mN/m, the calculated is similar to the measured . This is the case for monolayers with large compressibility modulus such as SpM- and cholesterol-enriched membranes [65]. Lipid monolayers in a more expanded state are compressed more easily and show a large area variation at 30 mN/m and above. The value of is therefore much smaller than .
3. Discussion
The comparison between the areas obtained for the pure lipid monolayers with those obtained for the mixtures may give insights regarding the lipid-lipid interactions established and the ideality of the mixture. If the interactions between distinct lipids in the monolayer are the same, the overall area is expected to be a weighted average of those of the pure components. On the other hand, if the distinct components are phase separated the overall area is also expected to be the sum of the contributions from each phase. It is therefore not possible to conclude regarding phase homogeneity in the mixture based only on the area comparison. One may nevertheless obtain important information, and this will be analyzed below for the areas obtained at 30 mN/m.
The area calculated for an ideal SpM:Chol mixture (6:4) is 44 Å2, assuming an area for cholesterol equal to 38 Å2 [72], compared to the experimentally obtained value of 39 Å2. The addition of cholesterol to POPC monolayers leads to average calculated areas equal to 57 and 51 Å2 at 30 and 50 mol%, while the measured values are 48 and 44 Å2, respectively. The calculated areas in cholesterol-enriched monolayers are therefore always much larger than the measured values, in agreement with the well-established cholesterol condensing effect [29,72].
When the area of the POPC:POPS (9:1) mixture is calculated from the area of pure POPC and POPS the value obtained per lipid (64 Å2) is significantly smaller than the one measured experimentally (70 Å2). This indicates that at 10 mol% of POPS the attractive interactions between the lipids are smaller and/or the repulsions are larger, this being compatible with a homogeneous lipid distribution where the hydrogen bonds between POPS molecules are not established, and the electrostatic repulsion leads to a larger separation between them.
The area calculated for POPC:POPE (8:2) is slightly higher than the measured one (64 vs 60 Å2) and the same behavior is obtained for the POPC:Chol:POPE membranes (56 vs 54 Å2). This may be interpreted as phase separation between POPC and POPE, especially in the presence of cholesterol. In fact, it has been shown that cholesterol induces an ordering effect on the acyl chains that is more pronounced in POPC compared to POPE [82] and that the hydrogen and electrostatic interactions between POPE headgroups are stronger than the interactions between POPE and Cholesterol [83,84]. This would favor coexistence of two liquid phases being one more enriched in PC and cholesterol than the other. The formation of cholesterol-rich domains in POPC:Chol:POPE monolayers has been observed by Atomic Force Microscopy [84]. When POPS is added to this ternary mixture, the calculated area (54 Å2) is smaller than the one observed (62 Å2). This variation is essentially the same as obtained in the POPC:POPS (9:1) mixture and may be interpreted as electrostatic repulsion between POPS.
The dipole potential is strongly affected by the lipid area and, as expected, the addition of cholesterol to POPC and SpM monolayers leads to an increase in the dipole potential obtained.
The addition of POPS to POPC monolayers leads to an increase in the area per lipid, however the dipole potential is significantly higher. This apparent contradiction is due to the much larger dipole potential of POPS which overcompensates the decrease in dipole density. The dipole potential of pure POPE is even larger than that of POPS and a small condensation is actually observed in the POPC:POPE (8:2) mixture. A larger increase in the dipole potential would therefore be expected for this mixture. This is not observed and the dipole potential measured for this mixture is almost the same as that of POPC. When cholesterol is added to this mixture, the dipole potential is significantly increased (481 vs 424 mV), although no condensing effect is observed. This behavior points towards significant structural rearrangements in POPE-containing monolayers either at the lipids or hydrating water layer, and may be a result of phase separation as discussed above. The addition of 10 mol% POPS leads to a small area expansion that is accompanied by the expected decrease in the dipole potential.
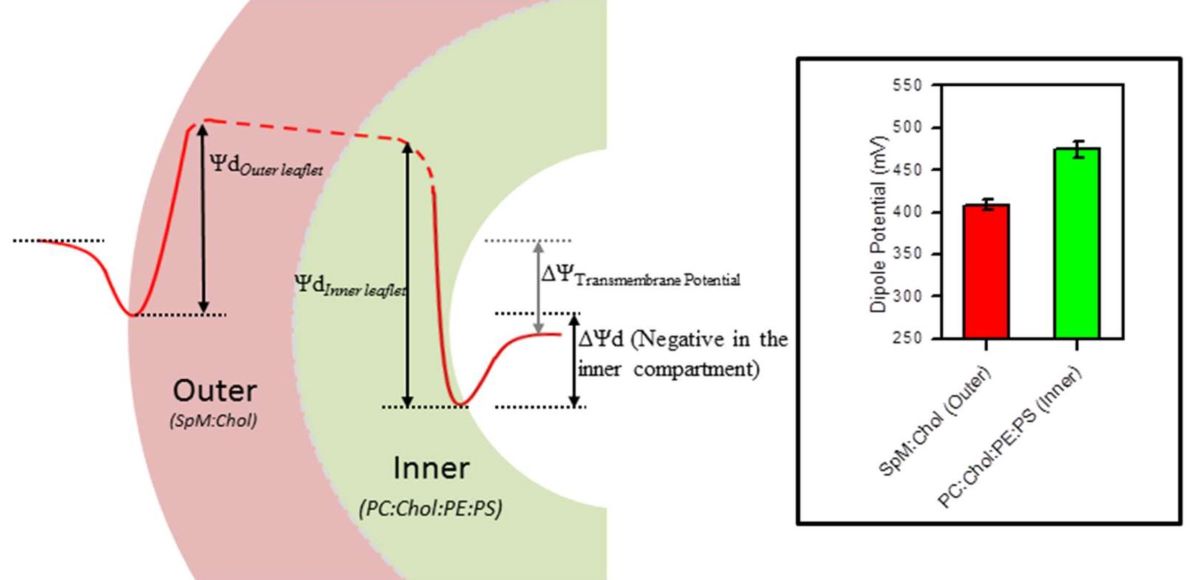
Biological membranes are often asymmetric, with distinct lipid compositions in the two leaflets, as a result of the asymmetric synthesis of lipids and proteins, their catalyzed translocation and the intrinsically slow passive diffusion through the membrane. From the dipole potentials obtained in air/water monolayers, one can calculate the transbilayer dipole potential. A POPC and POPE asymmetric bilayer leads to a non-zero transbilayer dipole potential (~77 mV) in good agreement with experimental results in bacterial PE and 1,3-diolein [85] and further observed by molecular dynamics simulations [86].
The plasma membrane of eukaryotic cells also presents an asymmetric lipid transmembrane distribution, being enriched in SpM and cholesterol in the outer leaflet and with significant amounts of POPE and POPS in the inner leaflet. The negatively charged POPS contributes in two ways to the transmembrane potential. It imposes a negative surface potential in the inner leaflet, and the larger dipole potential associated with this leaflet (negative in the aqueous membrane interface) increases the difference in the electrostatic potential between the two leaflets. The transmembrane dipole potential of this membrane may be calculated considering SpM:Chol (6:4) and POPC:POPE:Chol:POPS (4:3:2:1) for the outer and inner leaflet respectively. This leads to 66 mV, negative in the inner leaflet with respect to the outer one, reinforcing the measured transmembrane potential in eukaryotic cells [87].
Moreover, the depletion of POPS from the inner leaflet leads to a smaller dipole potential, and this may have important biological implications. POPS is known as a recognition molecule in apoptotic cells and once exposed to the outer leaflet it imposes a negative surface potential and serves as a signal for phagocytosis [88]. The decrease observed in the transmembrane potential due to the movement of POPS from the inner to the outer leaflet will also influence the normal function of membrane proteins and may be another important key signal in programmed cell death.
4. Materials and Methods
4.1. Materials
1-palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine (POPC), 1-palmitoyl-2-oleoyl-sn-glycero-3-phosphoethanolamine (POPE), 1-palmitoyl-2-oleoyl-sn-glycero-3-phospho-L-serine sodium salt (POPS), cholesterol (Chol), and egg-sphingomyelin (SpM) were from Avanti Polar Lipids, Inc. (Alabaster, Alabama, USA). All other reagents and solvents were of analytical grade, or higher purity, from Sigma-Aldrich Química S.A. (Sintra, Portugal). High quality deionized water, with a conductivity of 0.09 μS/cm, was used to perform the experiments.
4.2. Monolayer Surface Pressure Experiments
The formation of monolayers at an aqueous surface was monitored through changes in the surface pressure, using Kibron μTrough S equipment, with the addition of lipid at a constant area and temperature. Small aliquots of a chloroform solution containing the lipids of interest (0.5 mM) were carefully spread on a clean surface of an aqueous buffer (NaCl 150 mM, sodium azide 0.02%, EDTA 1 mM and HEPES 10 mM at pH 7.4) and left to equilibrate until the surface pressure reaches a constant value. When the addition of further lipid results in no visible changes in surface pressure (saturation) the excess lipid form aggregates in the aqueous subphase and the thermodynamic and interfacial properties are comparable to one leaflet of the lipid bilayer. This procedure allows for a spontaneous lipid stabilization in the aqueous-air interface without applying any lateral pressure [67,89]. The areas per lipid at the saturation pressure (πsat) were obtained from the variation of the surface pressure versus the amount of lipid through the interception of two lines, one describing the high pressure region (small slope) and the other describing the region with the steepest slope (Figure 6).
4.3. Measurement of the Potential Difference across Lipid Monolayers
The potential was measured in lipid monolayers formed at the air-aqueous interface. The system consists of a high impedance circuit which connects an ionizing electrode of polonium in the air and a reference (Ag/AgCl) electrode immersed in the aqueous subphase. The monolayers are formed, at the air-aqueous solution interface (NaCl 150 mM, sodium azide 0.02%, EDTA 1 mM and HEPES 10 mM at pH 7.4), by gently adding aliquots of lipids (dissolved in chloroform) with a micro syringe until the saturation point after which further addition of lipid does not induce any change in the measured dipole potential (Vlipid). The difference between the potential of the air-aqueous solution interface without any lipid (Vsolution) and the potential arising after the spreading of the lipids (Vlipid) allows the evaluation of the monolayer dipole potential [89]. For the case of zwitterionic lipids, the difference between the measured potentials results from the dipoles of the hydrated lipid monolayer and is directly the dipole potential. For charged lipid molecules the measured potential comprehends a contribution of the water and lipid dipole potentials as well as from the surface charged potential (Ψ0) (equation 2). Therefore, in order to obtain the dipole potential in monolayers containing charged lipids we must first determine Ψ0, which may be done using the Gouy-Chapman theory and the electric charge density (σ) at the experimentally determined area per lipid in the saturation pressure (πsat) (equation 3):
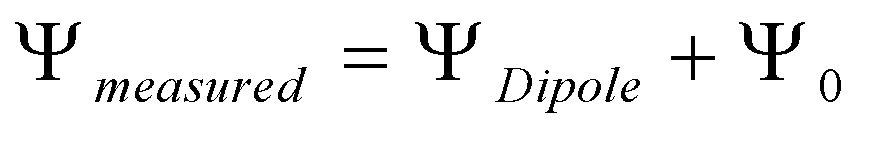 (2)
(2)
Author Contributions
Conceptualization, R.M.S.C. and M.J.M.; methodology, R.M.S.C. and E.A.D.; formal analysis, R.M.S.C. and M.J.M.; investigation, R.M.S.C. and F.L.; resources, E.A.D. and M.J.M.; writing—original draft preparation, R.M.S.C.; writing—review and editing, L.M.S.L. and M.J.M.; supervision, E.A.D. and M.J.M.; funding acquisition, L.M.S.L. and M.J.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Fundação para a Ciência e Tecnologia, Portugal, projects UIDB/00313/2020 (https://doi.org/10.54499/UIDB/00313/2020) and UIDP/00313/2020 (https://doi.org/10.54499/UIDP/00313/2020).
Data Availability Statement
The data presented in this study will be sent to interested researchers upon request to the corresponding authors.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- McLaughlin, S. Electrostatic Potentials at Membrane-Solution Interfaces. In; Bronner, F., Kleinzeller, A., Eds.; Current Topics in Membranes and Transport; Academic Press, 1977; Vol. 9, pp. 71–144.
- Honig, B.H.; Hubbell, W.L.; Flewelling, R.F. Electrostatic Interactions In Membranes And Proteins. Annu Rev Biophys 1986, 15, 163–193. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin, S. The Electrostatic Properties of Membranes. Annu Rev Biophys 1989, 18, 113–136. [Google Scholar] [CrossRef] [PubMed]
- Cevc, G. Membrane Electrostatics. Biochimica et Biophysica Acta (BBA) - Reviews on Biomembranes 1990, 1031, 311–382. [Google Scholar] [CrossRef]
- Liberman, Y.A.; Topaly, V.P. Permeability Of Bimolecular Phospholipid Membranes For Fat-Soluble Ions. BIOPHYSICS-USSR 1969, 14, 477. [Google Scholar]
- Lairion, F.; Disalvo, E.A. Effect of Dipole Potential Variations on the Surface Charge Potential of Lipid Membranes. J Phys Chem B 2009, 113, 1607–1614. [Google Scholar] [CrossRef] [PubMed]
- Gawrisch, K.; Ruston, D.; Zimmerberg, J.; Parsegian, V.A.; Rand, R.P.; Fuller, N. Membrane Dipole Potentials, Hydration Forces, and the Ordering of Water at Membrane Surfaces. Biophys J 1992, 61, 1213–1223. [Google Scholar] [CrossRef]
- Frias, M.A.; Cejas, J.P.; Rosa, A.S.; Disalvo, E.A. Relevance of Water in Biological Membranes. Chem Phys 2023, 566, 111784. [Google Scholar] [CrossRef]
- Li, S.; Wu, L.; Zhang, X.; Jiang, X. The Structure of Water Bonded to Phosphate Groups at the Electrified Zwitterionic Phospholipid Membranes/Aqueous Interface. Angewandte Chemie International Edition 2020, 59, 6627–6630. [Google Scholar] [CrossRef]
- Mai, T.L.; Derreumaux, P.; Nguyen, P.H. Structure and Elasticity of Mitochondrial Membranes: A Molecular Dynamics Simulation Study. J Phys Chem B 2023, 127, 10778–10791. [Google Scholar] [CrossRef]
- Zhan, H.; Lazaridis, T. Influence of the Membrane Dipole Potential on Peptide Binding to Lipid Bilayers. Biophys Chem 2012, 161, 1–7. [Google Scholar] [CrossRef]
- Cladera, J.; O’Shea, P. Intramembrane Molecular Dipoles Affect the Membrane Insertion and Folding of a Model Amphiphilic Peptide. Biophys J 1998, 74, 2434–2442. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, R.M.S.; Martins, P.A.T.; Ramos, C. V; Cordeiro, M.M.; Leote, R.J.B.; Razi Naqvi, K.; Vaz, W.L.C.; Moreno, M.J. Effect of Dipole Moment on Amphiphile Solubility and Partition into Liquid Ordered and Liquid Disordered Phases in Lipid Bilayers. Biochimica et Biophysica Acta (BBA) - Biomembranes 2020, 1862, 183157. [Google Scholar] [CrossRef] [PubMed]
- Szabo, G. Dual Mechanism for the Action of Cholesterol on Membrane Permeability. Nature 1974, 252, 47–49. [Google Scholar] [CrossRef] [PubMed]
- Flewelling, R.F.; Hubbell, W.L. Hydrophobic Ion Interactions with Membranes. Thermodynamic Analysis of Tetraphenylphosphonium Binding to Vesicles. Biophys J 1986, 49, 531–540. [Google Scholar] [CrossRef]
- Filipe, H.A.L.; Loura, L.M.S.; Moreno, M.J. Permeation of a Homologous Series of NBD-Labeled Fatty Amines through Lipid Bilayers: A Molecular Dynamics Study. Membranes (Basel) 2023, 13. [Google Scholar] [CrossRef]
- Luchian, T.; Mereuta, L. Phlorizin- and 6-Ketocholestanol-Mediated Antagonistic Modulation of Alamethicin Activity in Phospholipid Planar Membranes. Langmuir 2006, 22, 8452–8457. [Google Scholar] [CrossRef]
- Pilotelle-Bunner, A.; Beaunier, P.; Tandori, J.; Maroti, P.; Clarke, R.J.; Sebban, P. The Local Electric Field within Phospholipid Membranes Modulates the Charge Transfer Reactions in Reaction Centres. Biochimica et Biophysica Acta (BBA) - Bioenergetics 2009, 1787, 1039–1049. [Google Scholar] [CrossRef]
- Amazon, J.J.; Feigenson, G.W. Lattice Simulations of Phase Morphology on Lipid Bilayers: Renormalization, Membrane Shape, and Electrostatic Dipole Interactions. Phys Rev E 2014, 89, 22702. [Google Scholar] [CrossRef]
- Clarke, R.J. Effect of Lipid Structure on the Dipole Potential of Phosphatidylcholine Bilayers. Biochimica et Biophysica Acta (BBA) - Biomembranes 1997, 1327, 269–278. [Google Scholar] [CrossRef]
- Wang, L.; Bose, P.S.; Sigworth, F.J. Using Cryo-EM to Measure the Dipole Potential of a Lipid Membrane. Proceedings of the National Academy of Sciences 2006, 103, 18528–18533. [Google Scholar] [CrossRef]
- Almgren, M.; Gimel, J.C.; Wang, K.; Karlsson, G.; Edwards, K.; Brown, W.; Mortensen, K. SDS Micelles at High Ionic Strength. A Light Scattering, Neutron Scattering, Fluorescence Quenching, and CryoTEM Investigation. J Colloid Interface Sci 1998, 202, 222–231. [Google Scholar] [CrossRef]
- Sarkar, P.; Chattopadhyay, A. Membrane Dipole Potential: An Emerging Approach to Explore Membrane Organization and Function. J Phys Chem B 2022, 126, 4415–4430. [Google Scholar] [CrossRef] [PubMed]
- Brockman, H. Dipole Potential of Lipid Membranes. Chem Phys Lipids 1994, 73, 57–79. [Google Scholar] [CrossRef]
- Andersen, O.S.; Finkelstein, A.; Katz, I.; Cass, A. Effect of Phloretin on the Permeability of Thin Lipid Membranes. Journal of General Physiology 1976, 67, 749–771. [Google Scholar] [CrossRef]
- Franklin, J.C.; Cafiso, D.S. Internal Electrostatic Potentials in Bilayers: Measuring and Controlling Dipole Potentials in Lipid Vesicles. Biophys J 1993, 65, 289–299. [Google Scholar] [CrossRef] [PubMed]
- Simon, S.A.; McIntosh, T.J.; Magid, A.D.; Needham, D. Modulation of the Interbilayer Hydration Pressure by the Addition of Dipoles at the Hydrocarbon/Water Interface. Biophys J 1992, 61, 786–799. [Google Scholar] [CrossRef]
- McIntosh, T.J.; Magid, A.D.; Simon, S.A. Cholesterol Modifies the Short-Range Repulsive Interactions between Phosphatidylcholine Membranes. Biochemistry 1989, 28, 17–25. [Google Scholar] [CrossRef]
- Smaby, J.M.; Brockman, H.L.; Brown, R.E. Cholesterol’s Interfacial Interactions with Sphingomyelins and-Phosphatidylcholines: Hydrocarbon Chain Structure Determines the Magnitude of Condensation. Biochemistry 1994, 33, 9135–9142. [Google Scholar] [CrossRef]
- Orlikowska-Rzeznik, H.; Versluis, J.; Bakker, H.J.; Piatkowski, L. Cholesterol Changes Interfacial Water Alignment in Model Cell Membranes. J Am Chem Soc 2024, 146, 13151–13162. [Google Scholar] [CrossRef]
- den Kamp, J.A.F. Lipid Asymmetry in Membranes. Annu Rev Biochem 1979, 48, 47–71. [Google Scholar] [CrossRef]
- Devaux, P.F. Protein Involvement in Transmembrane Lipid. Annu Rev Biophys 1992, 21, 417–439. [Google Scholar] [CrossRef]
- Rothman, J.E.; Lenard, J. Membrane Asymmetry. Science (1979) 1977, 195, 743–753. [Google Scholar] [CrossRef] [PubMed]
- Leidl, K.; Liebisch, G.; Richter, D.; Schmitz, G. Mass Spectrometric Analysis of Lipid Species of Human Circulating Blood Cells. Biochimica et Biophysica Acta (BBA) - Molecular and Cell Biology of Lipids 2008, 1781, 655–664. [Google Scholar] [CrossRef] [PubMed]
- Dodge, J.T.; Phillips, G.B. Composition of Phospholipids and of Phospholipid Fatty Acids and Aldehydes in Human Red Cells. J Lipid Res 1967, 8, 667–675. [Google Scholar] [CrossRef]
- Verkleij, A.J.; Zwaal, R.F.A.; Roelofsen, B.; Comfurius, P.; Kastelijn, D.; van Deenen, L.L.M. The Asymmetric Distribution of Phospholipids in the Human Red Cell Membrane. A Combined Study Using Phospholipases and Freeze-Etch Electron Microscopy. Biochimica et Biophysica Acta (BBA) - Biomembranes 1973, 323, 178–193. [Google Scholar] [CrossRef]
- Kiessling, V.; Wan, C.; Tamm, L.K. Domain Coupling in Asymmetric Lipid Bilayers. Biochimica et Biophysica Acta (BBA) - Biomembranes 2009, 1788, 64–71. [Google Scholar] [CrossRef]
- Starke-Peterkovic, T.; Turner, N.; Vitha, M.F.; Waller, M.P.; Hibbs, D.E.; Clarke, R.J. Cholesterol Effect on the Dipole Potential of Lipid Membranes. Biophys J 2006, 90, 4060–4070. [Google Scholar] [CrossRef]
- Falkovich, S.G.; Martinez-Seara, H.; Nesterenko, A.M.; Vattulainen, I.; Gurtovenko, A.A. What Can We Learn about Cholesterol’s Transmembrane Distribution Based on Cholesterol-Induced Changes in Membrane Dipole Potential? J Phys Chem Lett 2016, 7, 4585–4590. [Google Scholar] [CrossRef]
- Shen, H.; Deng, M.; Wu, Z.; Zhang, J.; Zhang, Y.; Gao, C.; Cen, C. Effect of Cholesterol on Membrane Dipole Potential: Atomistic and Coarse-Grained Molecular Dynamics Simulations. J Chem Theory Comput 2018, 14, 3780–3795. [Google Scholar] [CrossRef]
- Oakes, V.; Domene, C. Stereospecific Interactions of Cholesterol in a Model Cell Membrane: Implications for the Membrane Dipole Potential. J Membr Biol 2018, 251, 507–519. [Google Scholar] [CrossRef]
- Clarke, R.J. Effect of Cholesterol on the Dipole Potential of Lipid Membranes. In Cholesterol Modulation of Protein Function: Sterol Specificity and Indirect Mechanisms; Rosenhouse-Dantsker, A., Bukiya, A.N., Eds.; Springer International Publishing: Cham, 2019; ISBN 978-3-030-04278-3. [Google Scholar]
- Zakany, F.; Kovacs, T.; Panyi, G.; Varga, Z. Direct and Indirect Cholesterol Effects on Membrane Proteins with Special Focus on Potassium Channels. Biochimica et Biophysica Acta (BBA) - Molecular and Cell Biology of Lipids 2020, 1865, 158706. [Google Scholar] [CrossRef] [PubMed]
- Martins, P.T.; Velazquez-Campoy, A.; Vaz, W.L.C.; Cardoso, R.M.S.; Valério, J.; Moreno, M.J. Kinetics and Thermodynamics of Chlorpromazine Interaction with Lipid Bilayers: Effect of Charge and Cholesterol. J Am Chem Soc 2012, 134, 4184–4195. [Google Scholar] [CrossRef] [PubMed]
- Sampaio, J.L.; Moreno, M.J.; Vaz, W.L.C. Kinetics and Thermodynamics of Association of a Fluorescent Lysophospholipid Derivative with Lipid Bilayers in Liquid-Ordered and Liquid-Disordered Phases. Biophys J 2005, 88, 4064–4071. [Google Scholar] [CrossRef]
- Coreta-Gomes, F.M.; Vaz, W.L.C.; Moreno, M.J. Effect of Acyl Chain Length on the Rate of Phospholipid Flip-Flop and Intermembrane Transfer. J Membr Biol 2018, 251, 431–442. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, A.C.; Filipe, H.A.L.; Geraldes, C.F.G.C.; Voth, G.A.; Moreno, M.J.; Loura, L.M.S. Interaction of MRI Contrast Agent [Gd(DOTA)]− with Lipid Membranes: A Molecular Dynamics Study. Inorg Chem 2024, 63, 10897–10914. [Google Scholar] [CrossRef]
- Cordeiro, M.M.; Filipe, H.A.L.; Santos, P. dos; Samelo, J.; Ramalho, J.P.P.; Loura, L.M.S.; Moreno, M.J. Interaction of Hoechst 33342 with POPC Membranes at Different PH Values. Molecules 2023, 28. [Google Scholar] [CrossRef] [PubMed]
- Magalhães, N.; Simões, G.M.; Ramos, C.; Samelo, J.; Oliveira, A.C.; Filipe, H.A.L.; Ramalho, J.P.P.; Moreno, M.J.; Loura, L.M.S. Interactions between Rhodamine Dyes and Model Membrane Systems—Insights from Molecular Dynamics Simulations. Molecules 2022, 27. [Google Scholar] [CrossRef]
- Estronca, L.M.B.B.; João Moreno, M.; Abreu, M.S.C.; Melo, E.; Vaz, W.L.C. Solubility of Amphiphiles in Membranes: Influence of Phase Properties and Amphiphile Head Group. Biochem Biophys Res Commun 2002, 296, 596–603. [Google Scholar] [CrossRef]
- Cejas, J. del P.; Rosa, A.S.; González Paz, A.N.; Disalvo, E.A.; Frías, M. de los A. Impact of Chlorogenic Acid on Surface and Phase Properties of Cholesterol-Enriched Phosphatidylcholine Membranes. Arch Biochem Biophys 2024, 753, 109913. [Google Scholar] [CrossRef]
- Taylor, G.; Nguyen, M.-A.; Koner, S.; Freeman, E.; Collier, C.P.; Sarles, S.A. Electrophysiological Interrogation of Asymmetric Droplet Interface Bilayers Reveals Surface-Bound Alamethicin Induces Lipid Flip-Flop. Biochimica et Biophysica Acta (BBA) - Biomembranes 2019, 1861, 335–343. [Google Scholar] [CrossRef]
- Li, S.; Wu, L.; Zhu, M.; Cheng, X.; Jiang, X. Effect of Dipole Potential on the Orientation of Voltage-Gated Alamethicin Peptides Regulated by Chaotropic Anions. Journal of Electroanalytical Chemistry 2022, 904, 115880. [Google Scholar] [CrossRef]
- de Matos, A.M.; Blázquez-Sánchez, M.T.; Sousa, C.; Oliveira, M.C.; de Almeida, R.F.M.; Rauter, A.P. C-Glucosylation as a Tool for the Prevention of PAINS-Induced Membrane Dipole Potential Alterations. Sci Rep 2021, 11, 4443. [Google Scholar] [CrossRef] [PubMed]
- Pohl, E.E.; Krylov, A. V; Block, M.; Pohl, P. Changes of the Membrane Potential Profile Induced by Verapamil and Propranolol. Biochimica et Biophysica Acta (BBA) - Biomembranes 1998, 1373, 170–178. [Google Scholar] [CrossRef]
- Cafiso, D.S. Dipole Potentials and Spontaneous Curvature: Membrane Properties That Could Mediate Anesthesia. Toxicol Lett 1998, 100–101, 431–439. [Google Scholar] [CrossRef] [PubMed]
- Paiva, T.O.; Bastos, A.E.P.; Marquês, J.T.; Viana, A.S.; Lima, P.A.; de Almeida, R.F.M. M-Cresol Affects the Lipid Bilayer in Membrane Models and Living Neurons. RSC Adv 2016, 6, 105699–105712. [Google Scholar] [CrossRef]
- Clarke, R.J. Dipole-Potential-Mediated Effects on Ion Pump Kinetics. Biophys J 2015, 109, 1513–1520. [Google Scholar] [CrossRef]
- Maggio, B. Modulation of Phospholipase A2 by Electrostatic Fields and Dipole Potential of Glycosphingolipids in Monolayers. J Lipid Res 1999, 40, 930–939. [Google Scholar] [CrossRef]
- Marsh, D. Lateral Pressure in Membranes. Biochimica et Biophysica Acta (BBA) - Reviews on Biomembranes 1996, 1286, 183–223. [Google Scholar] [CrossRef]
- Lantzsch, G.; Binder, H.; Heerklotz, H. Surface Area per Molecule in Lipid/C12Enmembranes as Seen by Fluorescence Resonance Energy Transfer. J Fluoresc 1994, 4, 339–343. [Google Scholar] [CrossRef]
- König, B.; Dietrich, U.; Klose, G. Hydration and Structural Properties of Mixed Lipid/Surfactant Model Membranes. Langmuir 1997, 13, 525–532. [Google Scholar] [CrossRef]
- Mukhopadhyay, P.; Monticelli, L.; Tieleman, D.P. Molecular Dynamics Simulation of a Palmitoyl-Oleoyl Phosphatidylserine Bilayer with Na+ Counterions and NaCl. Biophys J 2004, 86, 1601–1609. [Google Scholar] [CrossRef] [PubMed]
- Rand, R.P.; Parsegian, V.A. Hydration Forces between Phospholipid Bilayers. Biochimica et Biophysica Acta (BBA) - Reviews on Biomembranes 1989, 988, 351–376. [Google Scholar] [CrossRef]
- Smaby, J.M.; Momsen, M.M.; Brockman, H.L.; Brown, R.E. Phosphatidylcholine Acyl Unsaturation Modulates the Decrease in Interfacial Elasticity Induced by Cholesterol. Biophys J 1997, 73, 1492–1505. [Google Scholar] [CrossRef]
- Smaby, J.M.; Brockman, H.L. Surface Dipole Moments of Lipids at the Argon-Water Interface. Similarities among Glycerol-Ester-Based Lipids. Biophys J 1990, 58, 195–204. [Google Scholar] [CrossRef]
- MacDonald, R.C.; Simon, S.A. Lipid Monolayer States and Their Relationships to Bilayers. Proceedings of the National Academy of Sciences 1987, 84, 4089–4093. [Google Scholar] [CrossRef]
- Taylor, D.M.; De Oliveira, O.N.; Morgan, H. Models for Interpreting Surface Potential Measurements and Their Application to Phospholipid Monolayers. J Colloid Interface Sci 1990, 139, 508–518. [Google Scholar] [CrossRef]
- MacDonald, R.C.; Simon, S.A.; Baer, E. Ionic Influences on the Phase Transition of Dipalmitoylphosphatidylserine. Biochemistry 1976, 15, 885–891. [Google Scholar] [CrossRef]
- Ding, W.; Palaiokostas, M.; Wang, W.; Orsi, M. Effects of Lipid Composition on Bilayer Membranes Quantified by All-Atom Molecular Dynamics. J Phys Chem B 2015, 119, 15263–15274. [Google Scholar] [CrossRef]
- McIntosh, T.J.; Simon, S.A.; Needham, D.; Huang, C.H. Interbilayer Interactions between Sphingomyelin and Sphingomyelin/Cholesterol Bilayers. Biochemistry 1992, 31, 2020–2024. [Google Scholar] [CrossRef]
- Lund-Katz, S.; Laboda, H.M.; McLean, L.R.; Phillips, M.C. Influence of Molecular Packing and Phospholipid Type on Rates of Cholesterol Exchange. Biochemistry 1988, 27, 3416–3423. [Google Scholar] [CrossRef]
- Shahane, G.; Ding, W.; Palaiokostas, M.; Orsi, M. Physical Properties of Model Biological Lipid Bilayers: Insights from All-Atom Molecular Dynamics Simulations. J Mol Model 2019, 25, 76. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, T.J.; Simon, S.A.; Needham, D.; Huang, C.H. Structure and Cohesive Properties of Sphingomyelin/Cholesterol Bilayers. Biochemistry 1992, 31, 2012–2020. [Google Scholar] [CrossRef] [PubMed]
- Niemelä, P.; Hyvönen, M.T.; Vattulainen, I. Structure and Dynamics of Sphingomyelin Bilayer: Insight Gained through Systematic Comparison to Phosphatidylcholine. Biophys J 2004, 87, 2976–2989. [Google Scholar] [CrossRef]
- Chiu, S.W.; Vasudevan, S.; Jakobsson, E.; Mashl, R.J.; Scott, H.L. Structure of Sphingomyelin Bilayers: A Simulation Study. Biophys J 2003, 85, 3624–3635. [Google Scholar] [CrossRef]
- Cevc, G.; Watts, A.; Marsh, D. Titration of the Phase Transition of Phosphatidylserine Bilayer Membranes. Effects of PH, Surface Electrostatics, Ion Binding, and Head-Group Hydration. Biochemistry 1981, 20, 4955–4965. [Google Scholar] [CrossRef] [PubMed]
- Tsui, F.C.; Ojcius, D.M.; Hubbell, W.L. The Intrinsic PKa Values for Phosphatidylserine and Phosphatidylethanolamine in Phosphatidylcholine Host Bilayers. Biophys J 1986, 49, 459–468. [Google Scholar] [CrossRef]
- Seelig, A. Local Anesthetics and Pressure: A Comparison of Dibucaine Binding to Lipid Monolayers and Bilayers. Biochimica et Biophysica Acta (BBA) - Biomembranes 1987, 899, 196–204. [Google Scholar] [CrossRef]
- Blume, A. A Comparative Study of the Phase Transitions of Phospholipid Bilayers and Monolayers. Biochimica et Biophysica Acta (BBA) - Biomembranes 1979, 557, 32–44. [Google Scholar] [CrossRef]
- Demel, R.A.; Geurts van Kessel, W.S.M.; Zwaal, R.F.A.; Roelofsen, B.; van Deenen, L.L.M. Relation between Various Phospholipase Actions on Human Red Cell Membranes and the Interfacial Phospholipid Pressure in Monolayers. Biochimica et Biophysica Acta (BBA) - Biomembranes 1975, 406, 97–107. [Google Scholar] [CrossRef]
- Paré, C.; Lafleur, M. Polymorphism of POPE/Cholesterol System: A 2H Nuclear Magnetic Resonance and Infrared Spectroscopic Investigation. Biophys J 1998, 74, 899–909. [Google Scholar] [CrossRef]
- Ohvo-Rekilä, H.; Ramstedt, B.; Leppimäki, P.; Peter Slotte, J. Cholesterol Interactions with Phospholipids in Membranes. Prog Lipid Res 2002, 41, 66–97. [Google Scholar] [CrossRef] [PubMed]
- Sostarecz, A.G.; McQuaw, C.M.; Ewing, A.G.; Winograd, N. Phosphatidylethanolamine-Induced Cholesterol Domains Chemically Identified with Mass Spectrometric Imaging. J Am Chem Soc 2004, 126, 13882–13883. [Google Scholar] [CrossRef] [PubMed]
- Latorre, R.; Hall, J.E. Dipole Potential Measurements in Asymmetric Membranes. Nature 1976, 264, 361–363. [Google Scholar] [CrossRef] [PubMed]
- Gurtovenko, A.A.; Vattulainen, I. Lipid Transmembrane Asymmetry and Intrinsic Membrane Potential: Two Sides of the Same Coin. J Am Chem Soc 2007, 129, 5358–5359. [Google Scholar] [CrossRef] [PubMed]
- Zubay, G. Biochemistry; 3rd ed.; William C Brown: Dubuque, Iowa, 1993.
- Schlegel, R.A.; Williamson, P. Phosphatidylserine, a Death Knell. Cell Death Differ 2001, 8, 551–563. [Google Scholar] [CrossRef]
- Luzardo, M. del C.; Peltzer, G.; Disalvo, E.A. Surface Potential of Lipid Interfaces Formed by Mixtures of Phosphatidylcholine of Different Chain Lengths. Langmuir 1998, 14, 5858–5862. [Google Scholar] [CrossRef]
Figure 1.
Formation of the lipid monolayers, at
25±1ºC, followed by the changes in surface pressure as a function of the amount
of lipid added. (a) Pure POPC (□); POPC: POPS (9:1, molar ratio) (O);
POPC:POPE (8:2) (Δ); (b) POPC:CHOL:POPE (5:3:2) (◊); POPC:CHOL:POPE:POPS
(4:3:2:1)( ). The lines are a guide to
the eye.
). The lines are a guide to
the eye.
). The lines are a guide to
the eye.
Figure 1.
Formation of the lipid monolayers, at
25±1ºC, followed by the changes in surface pressure as a function of the amount
of lipid added. (a) Pure POPC (□); POPC: POPS (9:1, molar ratio) (O);
POPC:POPE (8:2) (Δ); (b) POPC:CHOL:POPE (5:3:2) (◊); POPC:CHOL:POPE:POPS
(4:3:2:1)(). The lines are a guide to
the eye.
). The lines are a guide to
the eye.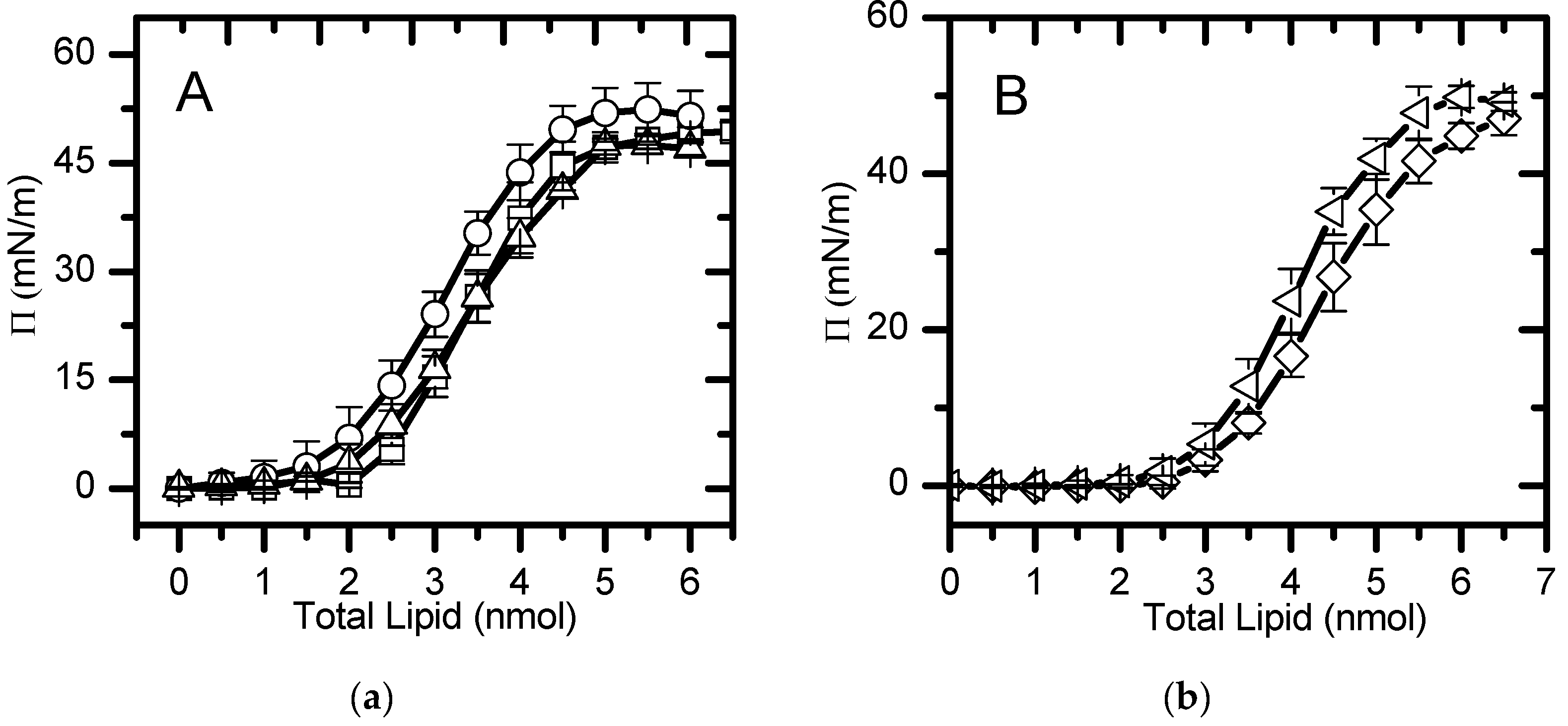
Figure 2.
Mean Area per lipid isotherm for POPC
(□), POPC:POPS (90:10), ○, and POPC:POPE( 80:20) ∆, (a), and
POPC:CHOL:POPE (5:3:2), ◊,
POPC:CHOL:POPE:POPS (4:3:2:1), , (b). The line is
the best fit of a polynomial equation for areas smaller than 100 Å2
in the liquid expanded state.
, (b). The line is
the best fit of a polynomial equation for areas smaller than 100 Å2
in the liquid expanded state.
Figure 2.
Mean Area per lipid isotherm for POPC
(□), POPC:POPS (90:10), ○, and POPC:POPE( 80:20) ∆, (a), and
POPC:CHOL:POPE (5:3:2), ◊,
POPC:CHOL:POPE:POPS (4:3:2:1), , (b). The line is
the best fit of a polynomial equation for areas smaller than 100 Å2
in the liquid expanded state.
, (b). The line is
the best fit of a polynomial equation for areas smaller than 100 Å2
in the liquid expanded state.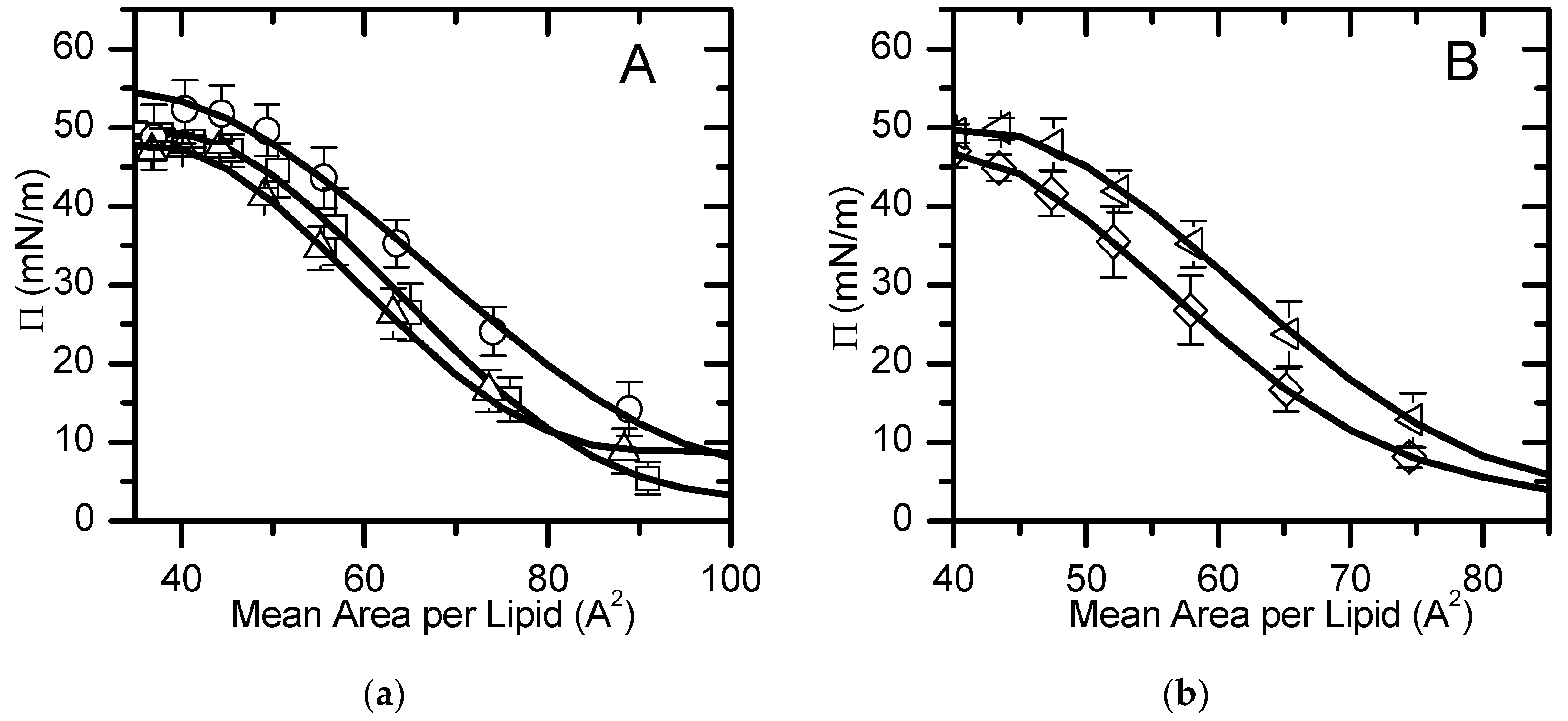
Figure 3.
Dipole Potential in pure POPC, POPE, POPS and SpM monolayers at πsat with the aqueous buffer as subphase.
Figure 3.
Dipole Potential in pure POPC, POPE, POPS and SpM monolayers at πsat with the aqueous buffer as subphase.
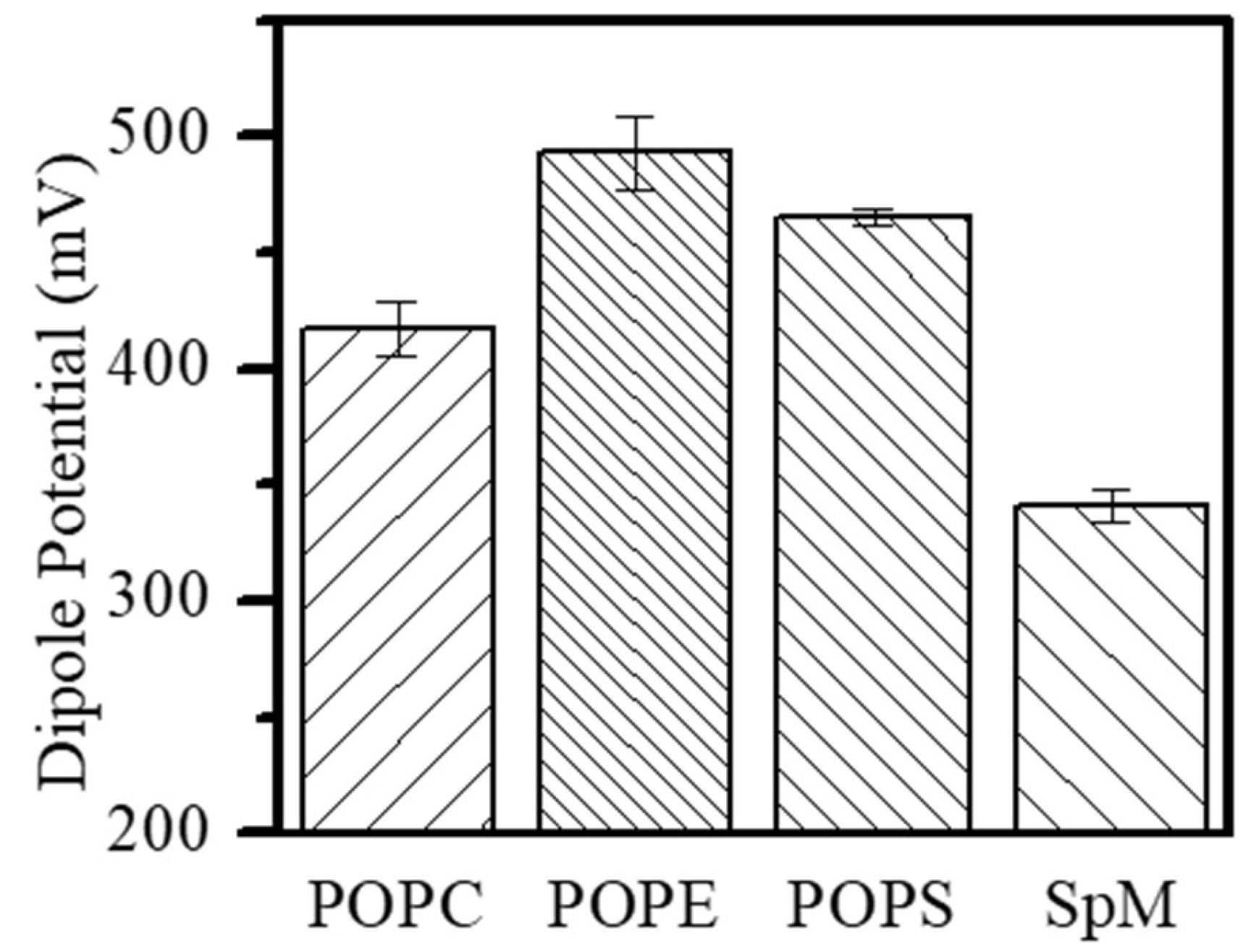
Figure 4.
Dipole Potential in monolayers composed by lipid mixtures, where PC, PS, PE, C and SpM abbreviations refer to POPC, POPS, POPE, Chol and Egg-SpM, respectively. The lipid molar proportion in the mixtures is given in parentheses.
Figure 4.
Dipole Potential in monolayers composed by lipid mixtures, where PC, PS, PE, C and SpM abbreviations refer to POPC, POPS, POPE, Chol and Egg-SpM, respectively. The lipid molar proportion in the mixtures is given in parentheses.
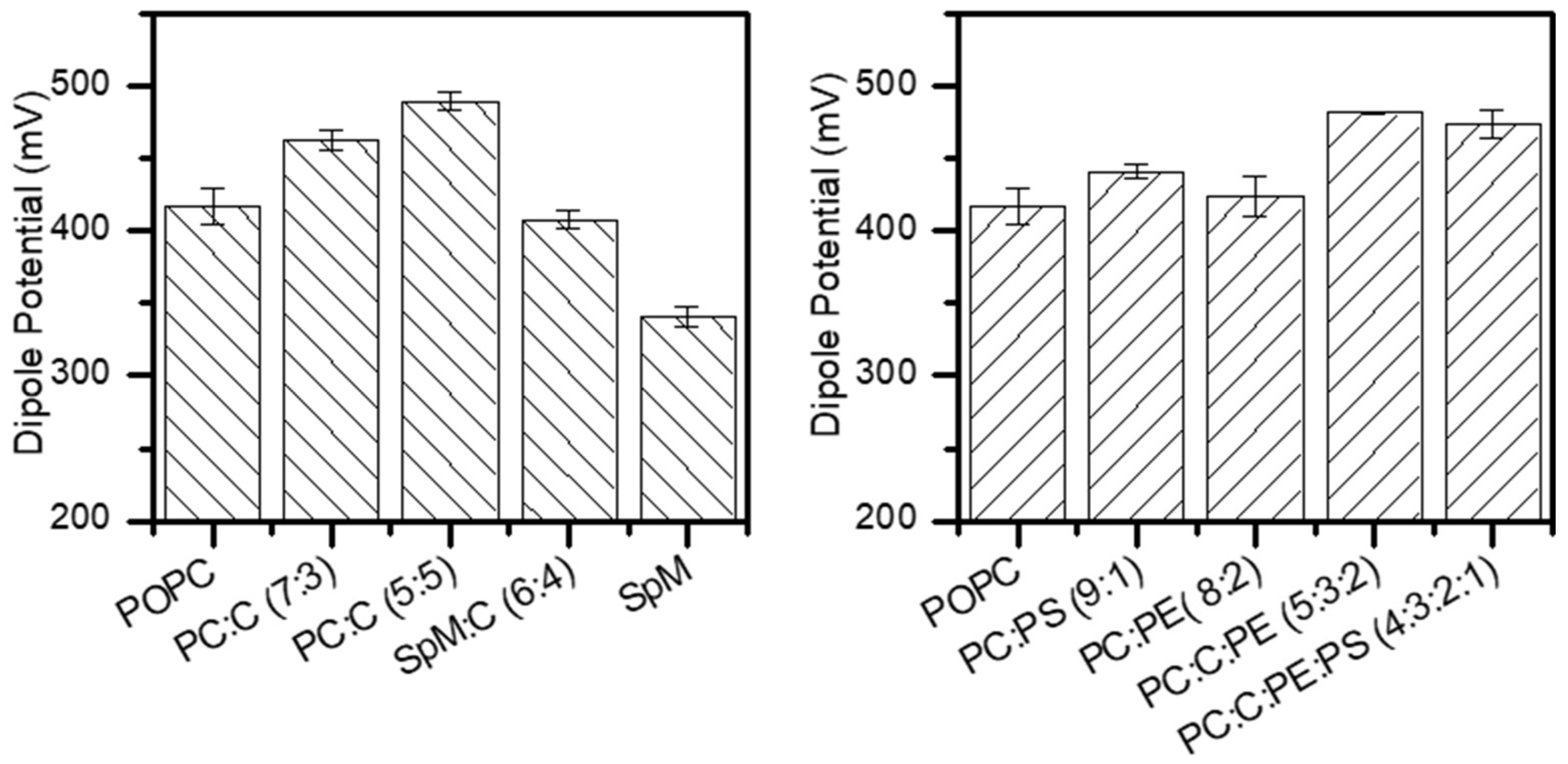
Figure 5.
Dipole potential of monolayers at a lateral pressure of 30 mN/m, calculated from equation 1 (), versus their dipole potential measured at the saturation pressure (). The straight line represents the identity line (=). The lipid monolayers with closer to the identity line are the POPC:Chol mixtures and are represented by (□), pure SpM is also close to the identity line (◊) while monolayers composed of pure POPC, POPE and POPS (◯) and their mixtures (including those with cholesterol) (✩) show the largest deviations.
Figure 5.
Dipole potential of monolayers at a lateral pressure of 30 mN/m, calculated from equation 1 (), versus their dipole potential measured at the saturation pressure (). The straight line represents the identity line (=). The lipid monolayers with closer to the identity line are the POPC:Chol mixtures and are represented by (□), pure SpM is also close to the identity line (◊) while monolayers composed of pure POPC, POPE and POPS (◯) and their mixtures (including those with cholesterol) (✩) show the largest deviations.
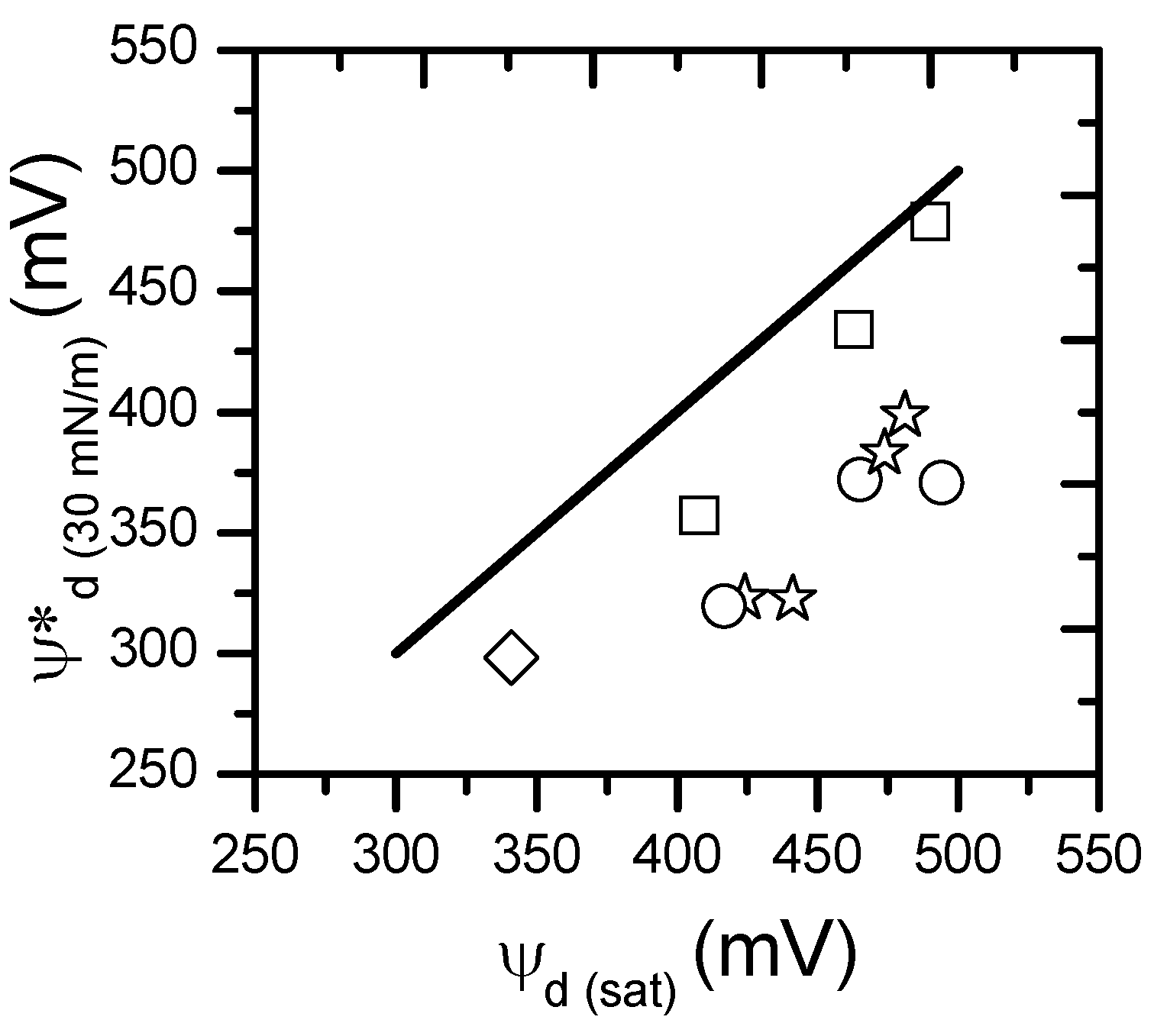
Figure 6.
Typical results for a pressure (π) vs total lipid (nmol) isotherm. The two lines represent the plateau at high pressures and the region with steepest slope, respectively. The saturation point corresponds to the interception of both straight lines.
Figure 6.
Typical results for a pressure (π) vs total lipid (nmol) isotherm. The two lines represent the plateau at high pressures and the region with steepest slope, respectively. The saturation point corresponds to the interception of both straight lines.
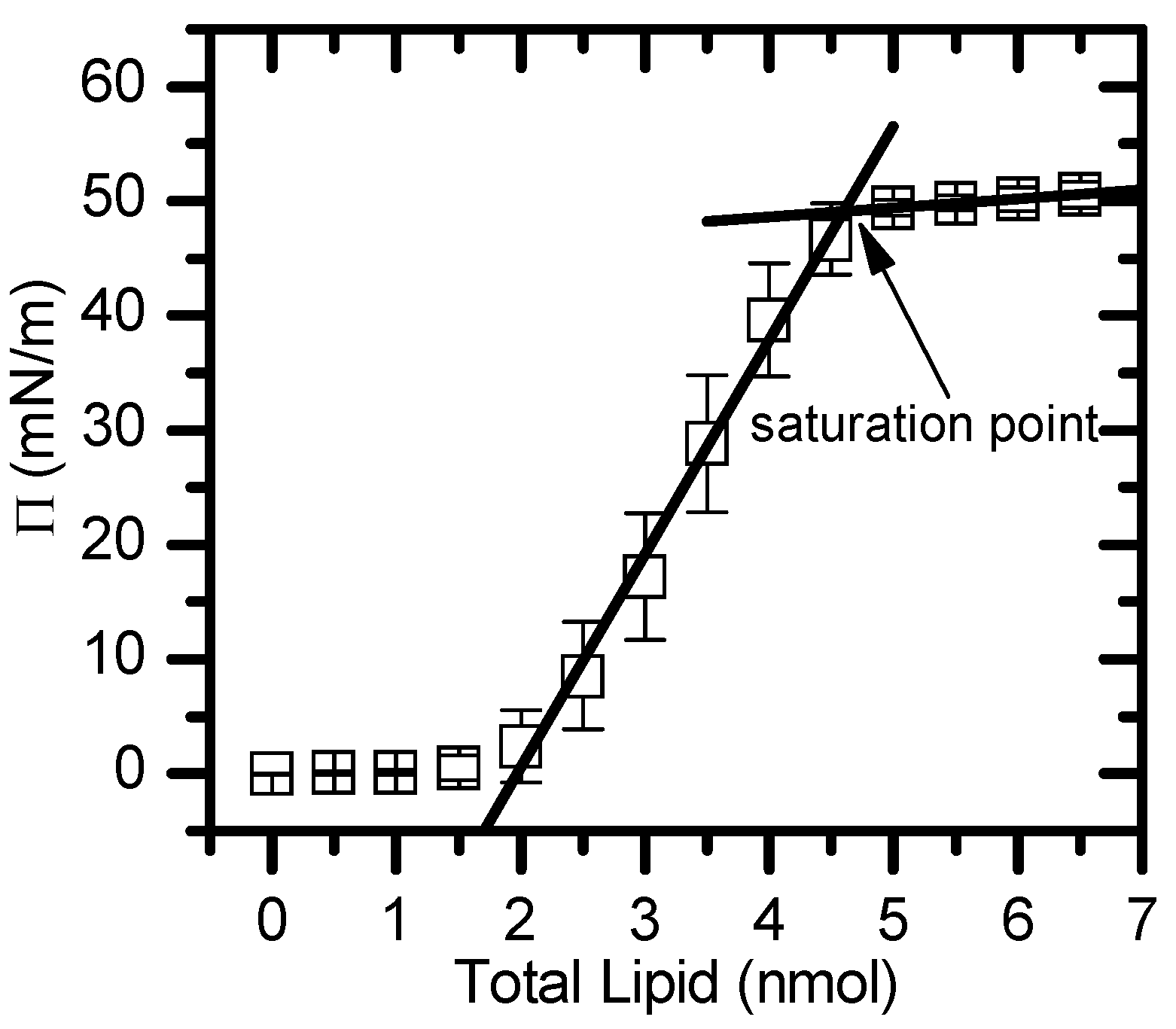

Table 1.
Mean area per lipid and the dipole potential obtained for monolayers with different lipid compositions formed at the air/water interface.
Table 1.
Mean area per lipid and the dipole potential obtained for monolayers with different lipid compositions formed at the air/water interface.
| Area per lipid at π = 30 mN/m (Å2) | Area per lipid at πsat (Å2) | Saturation Pressure (mN/m) | Dipole Potential at πsat (mV) | |
| POPC | 64.5 | 49.4±3.4 | 50 | 417±12 |
| POPS | 55.0a | 44f | - | 465±4.2e |
| POPE | 56.0b | 42f | - | 494±16 |
| SPM | 48.0c | 42c | - | 341±7.1 |
| POPC:POPS (9:1) | 69.8 | 51.0±4.0 | 51 | 441±2.5e |
| POPC:POPE (8:2) | 59.6 | 45.4±3.0 | 47 | 424±15 |
| POPC:CHOL (7:3) | 48.0d | 45d | - | 463±6.9 |
| POPC:CHOL (5:5) | 44.0d | 43d | - | 490±5.7 |
| POPC:CHOL:POPE (5:3:2) | 56.0 | 46.4±2.7 | 46 | 481±1.4 |
| POPC:CHOL:POPE:POPS (4:3:2:1) Inner | 61.5 | 49.7±2.6 | 49 | 474±11e |
| SPM:CHOL (6:4)outer | 39.0 c | - | - | 408±6.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



