
Submitted:
01 October 2024
Posted:
04 October 2024
You are already at the latest version
Abstract
The discovery of antibiotics was one of the greatest achievements in human history, however, antibiotic resistance evolved no later than the introduction of antibiotics. The rapid evolution of antibiotic-resistant pathogens soon became a nightmare and remained a global healthcare threat. There is an urgent need to have new alternatives or new strategies to combat the multi-drug resistant superbugs such as methicillin-resistant Staphylococcus aureus (MRSA), vancomycin-resistant enterococci (VRE), carbapenem-resistant Pseudomonas aeruginosa (CR-PA), extended-spectrum β-lactamases (ESBL) bearing multidrug-resistant Acinetobacter baumannii (MDR-AB), Escherichia coli, and Klebsiella pneumonia. Antimicrobial peptides (AMPs) have been considered promising agents equipped with unique mechanisms of action along with several other benefits to fight the battle against drug-resistant superbugs. Overall, the current review summarizes the mechanisms of drug-resistant development, the mechanism of action adopted by AMPs to combat drug-resistant pathogens, and the immunomodulatory properties of AMPs. Additionally, we have also reviewed the synergistic potential of AMPs with conventional antibiotics and the associated challenges and limitations of AMPs in the way of pharmacological development for therapeutic applications in clinical settings.
Keywords:
Superbugs
; Antibiotics
; Antimicrobial peptide
; Antibiotic-resistance
; Multidrug-resistance
; Infections
1. Introduction
Globally world is facing nearly 5 million deaths due to microbial resistance every year, according to the World Health Organization, making it vital to identify possible alternative drug candidates or therapeutic strategies as soon as possible [1]. Drug-resistant bacteria are increasing worldwide, causing the age of antibiotics to end with the pressing need to have novel alternatives to fight drug-resistant superbugs [2]. Due to AMR’s complexity, multiple approaches are required to deal with the problem. Some of them are: developing new antimicrobials, improving diagnostics, making existing drugs more effective, and developing new drug combinations. It is therefore increasingly necessary to find antibacterial therapeutics that reduce the risk of causing antimicrobial resistance [3]. AMPs (antimicrobial peptides) are emerging as viable alternatives to conventional antibiotics in the battle against rapidly evolving antibiotic resistance. It was unknown back then that MDR (multi-drug resistance) would become such a huge crisis over the years when AMPs were discovered, so translational applications for AMPs were not the priority at the time. Due to the subsequent rise in MDR strains, novel antimicrobial agents are now necessarily required and on the way to development [4].
As self-defense mechanisms, natural AMPs are potent inhibitory agents that are effective against the broadest spectrum of microorganisms, from bacteria to viruses. There are currently over 3300 AMPs in the antimicrobial peptide database (APD3). The composition of AMPs and their length vary widely. AMPs typically consist of short chains of amino acids (somewhere between 10 and 50), have a positive charge (between +2 and +11), are 2-9 kDa in size, and contain a majority of hydrophobic residues [5]. A key function of AMPs is to disrupt bacterial cell membranes, modulate the host’s immune response, and control inflammation [6]. The diverse antimicrobial mechanisms of AMPs make them promising therapeutic candidates, in contrast to conventional small-molecule drugs, which target cellular processes such as protein and cell wall synthesis) [7]. Additionally, AMPs are advantageous because they disrupt membranes directly, and their action times are quick, therefore, it is difficult for microbes to develop resistance against them [8,9,10]. Throughout the past few decades, we have gained a deep understanding of antibiotics and how bacteria resist their inhibitory or killing effects, as well as the significance of context in determining the effects of many resistance mechanisms, for example, a diverse growth condition can dramatically alter the expression of resistance genes. Overall, an understanding of bacterial resistance development mechanisms also provides insights into mechanisms used by AMPs to overcome bacteria resistance (Figure 1).
This article reviews the emergency of having new alternatives to cope drug resistance superbugs along with highlighting the various mechanisms of action used by AMPs to fight against the infections caused by drug-resistant superbugs. In addition, AMPs are discussed in conjunction with advanced new technologies for the treatment of drug-resistant bacterial infections and challenges in the way of AMPs to pharmacological development for therapeutic applications in clinical settings.
2. Antibiotic Resistance Drivers and Mechanisms
As microorganisms become resistant to antibiotics as soon as they are introduced, the new antibiotics remain ineffective for the treatment of common infections. Hospitals and other sectors in both developing and developed countries use antibiotics too frequently and irresponsibly, contributing to frequent mutations and selection pressures that lead to rapid resistance development. Resistance to antibiotics is a grave threat primarily due to poor hygiene and sanitation, as well as globalization in terms of trade, travel, and transmission [11]. Additionally, many factors contribute to drug resistance, including agriculture and animal husbandry. A direct route for transmitting resistant bacteria between humans and animals is the food chain, for example, cattle are often given antibiotics (even when they are healthy) to prevent illness, which leads to overuse of these antibiotics, since they are similar to those used clinically, for instance, poultry has been identified as a source of drug-resistant E. coli in rural villages of Barcelona [12,13]. Further, infections are often treated with broad-spectrum antibiotics in hospitals, which allows higher opportunities for resistance development. Evidence shows that the rate of resistance to antibiotics may increase with increased antibiotic usage, while resistance rates may fall with reduced antibiotic usage. Further, frequent re-administration of antibiotics over time also provides enough chances for resistance development, since antibiotics provide selective pressure on bacteria [13]. Unfortunately due to a lack of proper diagnostic tools, along with unnecessary antibiotic administration, and their low cost, antimicrobial resistance is rising uncontrollably [12,13].
The antibiotic resistance mechanism of currently used conventional antibiotics, especially target cell walls, DNA replication, and translational activity to eliminate pathogenic bacteria. Additionally, naturally resisting antibiotics or mutations in the bacterial chromosomes are also contributing factors for resistance development. The resistant mutant may continue to exist as soon as the antibiotic kills the normal bacterium while transmitting the antibiotic resistance genes by horizontal or vertical transfer.
In summary, bacteria adopted various mechanisms to develop drug-resistant that includes (i) Poor drug influx through porin channels such as, a mutation in the outer membrane of P. aeruginosa may lead to resistance to imipenem, (ii) Increased efflux of drugs from drug-resistant bacterial cells via efflux pumps along with the modification in regulatory proteins, (iii) Modifications to genomes affecting or protecting target areas such as Group C and G Streptococci, have mutations in erm genes that prevent binding to macrolides, lincosamides, and streptogramins, (iv) Covalent bonding is another common method of developing resistance to drugs for example NDM-1 (metallo β-lactamase 1, a carbapenemase-active-lactamase) that resists all antibiotics used to treat severe infections, (v) The R plasmid, a plasmid that carries several resistant genes which plays a crucial role in drug-resistance transmission such as MGE pheromone, that involved with enterococcal pheromone-responsive plasmids and assists resistance development in E. faecalis, (vi) Biofilm development that become resistant to available antibiotics. The biofilm matrix which is made up of exo-polysaccharides slows down the diffusion and reduces the bioavailability of antibiotics by increased viscosity [14]. (vii) Surface remodeling such as lipid A, which is a part of the Gram-negative outer membrane component LPS, that is modified with phosphoethanolamine or 4-amino-4-deoxy-l-arabinose residues in polymyxin B-resistant P. aeruginosa, V. cholerae, A. baumannii, and S. enterica [15,16]. Polymyxin B resistance is conferred by LPS modifications, which are primarily caused by regulatory complexes like PmrAB and PhoPQ, which alter gene expression [17].
3. AMP Mechanisms of Action in Treating Drug-Resistant Bacteria
In the past few years, MDR bacteria have emerged as a result of antibiotic abuse that specifically includes MRSA, MDR-AB, VRE, and CR-PA. As antibiotics have failed to treat bacterial infections, therefore, treatment and prevention have become increasingly important. The use of AMPs is one of the promising new approaches for treating multidrug-resistant bacteria due to their unique antibacterial mechanism by hitting more targets to kill bacteria. AMPs, unlike antibiotics, directly act on bacterial membranes including biofilms. Additionally, AMPs resensitize, and inhibit biofilm growth, via regulating immune responses, and thus regulate intracellular bacteriostatic functions. Overall, these factors and specific mechanisms of AMPs indicate their efficacy as an epistatic alternative to conventional antibiotics.
3.1. AMPs Mediated Membrane Disruption
The mechanism of action of AMPs involves the disruption of membranes which typically relies on a variety of composition of pathogen and host membranes to impart selectivity. As opposed to the mammalian cell surface that has primarily neutrally charged phospholipids such as phosphatidylcholine and sphingolipids, bacteria, on the other hand, have a significant fraction of negatively charged phospholipids like cardiolipin, phosphatidylglycerol (PG), and phosphatidylserine (PS), which is a favorable factor for positively charged AMPs to act upon [18]. Furthermore, Gram-positive bacteria have large amounts of teichoic acid which is negatively charged in their peptidoglycan cell wall surrounding their membrane. In Gram-negative bacteria, the outer leaflets of their outer membranes are predominantly composed of lipopolysaccharide which is also negatively charged [19]. There are negatively charged phospholipids like PS in mammalian cell membranes, but they are broadly distributed in the cytosol leaflet of the bilayer, so AMPs usually don’t target mammalian cells [20]. Overall, the outer surface of bacteria has an overall negative charge that results in electrostatic interactions with AMPs that enable the primary binding and membrane-specific killing by AMPs [21]. Studies have shown that AMP activity is decreased significantly by decreasing AMP positive charge below a certain threshold [22]. Increasing the positive charge of AMPs should generally result in the improved antimicrobial activity of those AMPs, but too high a charge could cause off-target toxic effects. A higher charge density is supposed to enhance the AMP-water interaction, stabilizing the transmembrane pores on the hydrophilic surface which is formed by AMPs after the initial binding to bacterial cell membrane. The electrostatic attraction to a membrane that is negatively charged, however, does not seem feasible when various AMPs are anionic. An example of such a type of peptide is DCD-1L [23]. Although DCD-1L has an overall negative charge of -2, it is assumed that only it’s positively charged N-terminus plays a major role in the initial interaction with the target bacterial cell membrane. Additionally, it is believed that the divalent cations such as Mg 2+, Ca2+, and Zn2+, are also involved in improving the antimicrobial activity of DCD-1L by stabilization of membrane-spanning peptide oligomers, or through salt bridge formation with the anionic phospholipids. Next, interaction with the positive phospholipids or hydrophobicity is another mechanism of action for anionic AMPs against superbugs [24]. For example, microcin J25 is taken up through FhuA, an outer membrane protein that is involved in an anionic AMP transport [25].
Further, based on spectrometric data and molecular-dynamic simulations using artificial membrane bilayers and liposomes, several models have been proposed to explain the AMPs mode of action. These models are categorized as porous or nonporous. The barrel-stave model and the toroidal model are two pore-forming models that support the formation of transmembrane pores on the lipid bilayer [22,25,26]. In the barrel-stave model, AMPs interact with the fatty acyl chains surrounding the pore wall, whereas in the toroidal model, they only interact with the lipid head groups. The 26-residue peptide melittin, which is found in bee venom, is one of the most famous AMPs classified in the pore-forming model. In contrast, several non-pore-forming models have also been proposed for AMP mechanism of action, such as the carpet model and the detergent-like model [21,27]. In the carpet model, AMPs lie on the surface of the membrane and interact only with the lipid head group while the detergent-like model shows how AMPs dissolve and remove lipids from a lipid bilayer in a detergent-like fashion. Interestingly, one AMP may disrupt bacterial cell membranes through more than one mechanism depending on its concentration, conditions, and lipid composition [26]. AMPs have been studied primarily using artificial membranes, while studies of their effects on real membranes have been scarce. An AMP must accumulate to a critical concentration on the membrane surface before it can disrupt the target bacterial membrane. Depending on whether diffusion barriers exist in the outer membrane or the periplasmic space, they may be prevented from partitioning onto the membrane. There are fewer rate-limiting steps for Gram-positive bacteria because AMPs only need to diffuse through nano-sized pores in the peptidoglycan [28]. It is the peptidoglycan layer of the target bacterial membrane that facilitates the accumulation of AMPs on the target bacterial membrane via the availability of negatively charged teichoic acid that helps in providing an overall negative charge to facilitate the interaction with positively charged AMPs [26]. Further, in the case of Gram-negative pathogens, AMPs first need to penetrate the outer envelope and then the cell membrane such as polymyxin B has strong antimicrobial activity against Gram-negative pathogens because it disrupts both the outer and cytoplasmic membranes. It has been shown that the removal of the lipid tail from the polymyxin B, allows it to permeabilize only the outer membrane of the Gram-negative bacteria and lose its antimicrobial properties [27]. This suggested the importance of lipid moiety in the antimicrobial action of polymyxin B and other lipid-containing AMPs.
3.2. AMPs Mediated Intracellular Targeting
AMPs are short polycationic peptides that have a wide range of antimicrobial properties. Membrane-lysis is recognized as the primary antibacterial mechanism, which directly impairs the integrity of the bacterial membranes and cell walls. AMPs also form transmembrane channels when they self-aggregate or polymerize, resulting in cytoplasm leakage and cell death. However, increasing evidence suggests that AMPs are capable of exerting intracellular inhibition as the primary or supporting mechanism for effective killing. Several AMPs inhibit multiple intracellular targets, such as indolicidin, buforin II, DM3, and microcin J25 [29]. In many studies, AMPs have proven to affect various physiological processes in cells, including the synthesis of DNA and proteins, the folding of proteins, enzyme activity, and the synthesis of cell walls [30]. Burofin II (Lys-C-cleaved derivative of buforin I), is a transmembrane active AMP that induces membrane permeation in Escherichia coli due to its proline hinge region [31]. In the gel retardation experiments, buforin II is found to exhibit a high affinity with nucleic acids and it is strongly implied that it inhibits the cellular processes by interfering with RNA and DNA metabolism [32]. Next, indolicidin, an AMP of cathelicidin family is reported to inhibit DNA biosynthesis exclusively via continuous entry into the bacterial cells without any observed cell lysis [33,34,35,36]. Further, detailed studies confirmed the strong binding of indolicidin to the duplex DNA [CG], [AG], and [AT], while weak biding to [GT] [37]. It has also been reported that indolicidin inhibits DNA replication and transcription by stabilizing the central PWWP motif on the duplex DNA [38]. Microcin J25 is reported to interfere with the transcription by binding to bacterial RNA polymerase [39]. Another example is lactoferricin B, which killed the target bacterial by interfering with the phosphorylation of the bacterial two-component system response regulators BasR and CreB along with the accumulation of pyruvate in bacterial cells [40,41].
Bac7, is a large 60 amino acid-long AMP isolated from bovine neutrophils, however, its N-terminal 35-residue (Bac7 1-35) is reported to have a role in antimicrobial activity via ribosome binding and inhibition of protein translation [42,43]. Next, a hybrid AMP, DM3, is demonstrated to exhibit broad-spectrum bactericidal activity in a rapid time interval while A genome-wide transcriptomic analysis revealed significant changes in several important intracellular pathways including those associated with DNA replication and transcription, RpoD, and RNA polymerase sigma factors down-regulation, and amino acid biosynthesis pathways [44,45].
3.3. Immune Activity Regulation by AMPs
In several studies, it has been shown that AMPs can suppress or enhance innate and adaptive immune system responses, demonstrating their immunomodulatory properties. A variety of factors influence this capacity, including environmental factors, the cells involved, receptor interactions, signaling pathways involved, and transcription factors that bind to them [46]. Several AMPs have been shown to act in a pleiotropic manner depending on the cell they are acting on, while their concentration plays a crucial role in the mechanism of action [47]. AMPs are found in NK cells and neutrophils as well as in innate immunity, including α defensins and cathelicidins. Extracellular release of AMPs can also act as a chemokine because they recruit diverse types of cells to the infection site, including neutrophils, eosinophils, mast cells, monocytes, and lymphocytes [30,48]. Human Neutrophil Peptide (HNP-1) increases pathogen elimination by stimulating macrophages for phagocytosis of apoptotic peptides and neutrophils [49]. Cathelicidin LL-37 has also been observed stimulating neutrophil phagocytosis. There is a fundamental mechanism that involves the chemokine CXCL8 synthesis which is regulated by p38 (MAPK) and ERK (extracellular signal-regulated kinase) signaling pathways [50]. For initiating adaptive antimicrobial immune responses, the recruitment of immune cells to infection sites includes dendritic cells, macrophages, antigen-presenting cells (APCs), B lymphocytes, and macrophages [51]. Based on their interactions particularly with CCR6, the HNP1, HNP-2, HNP-3, and hBD1-3 are the selective chemoattractants for immature dendritic cells [52,53,54]. Apart from recruitment, AMPs could also promote and regulate dendritic cell maturation because mouse β-defensin 2 upregulates the expression of costimulatory receptors, such as CD40, CD80, and MHC II, while the similar phenomenon reported in the case of hBD1, hBD3, and HNP-1. Through immune regulation, AMPs show promising effects in the area of sepsis because AMPs can confer bactericidal and immunomodulatory effects, which leads to the development of combination therapies with bactericidal and anti-inflammatory AMPs [55,56]. The initial focus of AMP research was on classical natural AMPs, but they could also improve immunomodulatory activities for the applications in various therapeutic applications.
3.4. AMPs as Role Players in Eradicating Biofilm-Mediated Drug Resistance
Biofilms may be as most adaptable microbial nature’s feature. As a consequence, when pathogenic microbes aggregate into the biofilms, become an important virulence factor. Furthermore, microbial biofilms reduce the effectiveness of antimicrobial compounds and modulate the immune response, thereby causing antimicrobial resistance that enables persistent infection establishment. AMR is extremely complex, in which biofilms play a significant role in driving the resistance. Bacteria residing in biofilms exhibit a 100 to 1000-fold enhanced AMR than those living in a planktonic state. One of the studies examined that 100% of S. epidermidis strains tested were susceptible to antibiotic vancomycin in the planktonic state while nearly 75% of the biofilms were found fully resistant [57]. Similarly in the case of K. pneumoniae, which was found to be susceptible when tested in a planktonic state but becomes resistant in biofilms to certain antibiotics [58]. Biofilm develops a surface resistance and it is hard for antibiotics to penetrate this slimy and sticky surface. Multilayered, localized, and heterogeneous communities of bacteria make up a bacterial biofilm, which is surrounded by a matrix of extracellular polymeric substances (EPSs) [59]. Biomolecules in EPSs include polysaccharides, proteins, extracellular DNA, and lipids, which makes it difficult for the antibiotics to penetrate and reach their target bacteria. Additionally, because of the slow diffusion within the biofilms, antibiotics could be deactivated near the level of the surface rather than reaching the depth of the cell. Microenvironmental resistance inside biofilms is another aspect that makes it challenging for antibiotics at the deeper levels where waste, metabolic by-products, and nutrients, are also accumulated. Furthermore, an anaerobic environment could also be possible within the biofilms that drastically reduced oxygen levels. When these factors are combined, antibiotics vary in their effects depending on their structure and action as ciprofloxacin and tobramycin are found to have less bactericidal effect at low oxygen levels, whereas changes in pH could impact negatively aminoglycoside action [60]. Additionally, bacterial persister cell resistance is another phenomenon for the bacteria residing deeper in biofilm layers, where they become more adept at evading antibiotic therapy. Moreover, as a method of survival, a subpopulation of bacteria develops spore-like characteristics (persister cells) which make them resistant to antibiotics or chemical treatments. In the presence of antibiotics, these persister cells remain in a dormant state and don’t divide. Additionally, persister cells do not genetically modify to resist and survive antibiotic treatment, and they will return to their pre-persister susceptibility profile once released from biofilms and start dividing again [61]. According to an estimation, up to 500–50,000 times the antimicrobial resistance of microorganisms can be enhanced by biofilm formation [62]. It is generally known that AMPs are highly stable at a wide range of pH and temperature, properties that are beneficial in scaling up production and incorporating them into deliverable products. Even well-established biofilms have been eliminated by low concentrations of AMPs with inhibitory and disruptive properties. As well as acting at different stages of biofilm formation, AMPs can inhibit adhesion, inhibit biofilm formation, and kill pre-formed biofilms, all of which are potential applications of AMPs [63].
For example, natural AMP LL-37 from the cathelicidin family and synthetic AMP NA-CATH:ATRA1-ATRA1 are known to inhibit the growth of biofilms by S. aureus below a concentration of 3 μg/mL [64]. It has been reported that LL-37 stimulates twitching motility, down-regulates the Las and Rhl QS system, and decreases P. aeruginosa cell attachment to medical devices and tissues while in the case of S. epidermidis it inhibits both initial attachment and biofilm formation [65,66]. Several anti-biofilm agents, such as lactoferrin, conjugated lactoferricin, melimine, and citropin 1.1, are effective in medical device infections caused by S. aureus and P. aeruginosa, especially when administered in combination with conventional antibiotics, such as rifampicin and minocycline [67]. In a recent study, four chimeric AMPs are shown to inhibit multidrug-resistant biofilms of Acinetobacter baumannii in synergy with conventional antibiotics, while demonstrating low cytotoxicity against human skin cells [68]. Overall, AMPs displayed antibiofilm activities by direct disruption or degradation of biofilm-embedded bacterial cell membrane potential, disrupting bacterial cell signaling systems, degradation of polysaccharide and biofilm matrix, inhibition of the alarm system to avoid bacterial stringent responses, and by reduction of the expression of genes responsible for biofilm formation and protein transport [69].
4. AMPs Approved for Clinical Use or under Clinical Trials
The development of AMP-based therapies could eventually replace conventional antibiotics with broad-spectrum AMPs. To date, only a few AMPs have been approved for use in clinical settings (Table 1). Vancomycin is one of the most efficient AMP (tricyclic glycopeptide), originally produced and isolated from Streptococcus orientalis that has been approved for the treatment of serious Gram-positive bacterial infections [11]. It is efficiently active against Gram-positive bacilli as it enables to inhibit formation of the cell wall of Gram-positive bacteria. As a first-line antibiotic, vancomycin is also used to treat multiple drug-resistant infections such as endocarditis, bacteremia, pneumonia, osteomyelitis, and cellulitis [70]. Gramicidin (15 amino acids) is a natural AMP produced by Bacillus brevis. Gramicidin is the first peptide antibiotic discovered in 1939 and later got FDA approval for topical application against Gram-positive bacteria [71]. Bacitracin (12 amino acids) is another FDA-approved natural AMP produced by Bacillus licheniformis M1. It shows potent activity against Gram-positive bacteria including MRSA [72]. Next, colisitin (10 amino acids) is produced by Paenibacillus polymyxa and is FDA-approved. It is considered a last resort treatment for Gram-negative bacterial infections including pneumonia and also shows potent antibiofilm activities [73]. Daptomycin is a cyclic lipopeptide produced by Streptomyces roseosporus. It has been approved to treat serious skin infections caused by Gram-positive bacteria which include methicillin-resistant Staphylococcus aureus (MRSA) and vancomycin-resistant Enterococci (VRE) [74]. Next, polymyxin B is a cyclic cationic AMP produced by Bacillus polymyxa. It has potent activity mainly against Gram-negative bacteria including K. pneumoniae, P. aeruginosa, and A. baumannii [75,76]. Also, many potential AMPs are already under clinical trials to be used against various infections (Table 2). In addition to this, there are many AMPs reported with potential activities against various drug-resistant bacteria and biofilms (Table 3). All the experimental evidences suggested that AMPs could be an efficient alternative to combat superbug infections, especially when there are great opportunity to have enormous new AMPs. Interestingly, the bacterial communities such as human gut microbiome which is constantly evolving under the pressure of other human metabolites and various interacting factors. Additionally, bacterial communities from marine ecosystem have been shown to have huge possibilities of potentially novel AMPs to fight against drug-resistant super bugs [77].
5. Synergistic Action of AMPs with Conventional Antibiotics
It has been found that many of the conventional antibiotics become more efficient when used along with AMPs. This suggested the synergistic action potential of AMPs with antibiotics or other antimicrobials which is mainly due to their membrane-specific mechanisms of action that facilitate the entry of antibiotics in the bacterial cell for intracellular targeting (Table 4). Combining AMPs with other antibacterial agents provides a new set of drugs to achieve synergistic action, thus overcoming the limitations of using a single drug to treat a particular infection or disease. The synergistic effect of AMPs could allow the reuse of standard antibiotics and result in better expected clinical outcomes including increased efficacy, reduced toxicity, and delayed development of resistance. It has been demonstrated that a defensin-like bacteriocin, laterosporulin10 produced by Brevibacillus laterosporus SKDU10, and HNP-1 exhibits synergistic activity with rifampicin (a frontline anti-Mtb drug) against pathogenic stain Mycobacterium tuberculosis H37Rv, and significantly reduces the MIC values of rifampicin [78,79]. This is interesting, especially against drug-resistant Mtb, when there is no efficient antibiotic available for 100% eradication. Nisin and its variants displayed synergism with various antibiotics (Ramoplanin, Polymyxin E, Clarithromycin, Amoxicillin, Penicillin, Streptomycin, Ceftiofur, Tetracycline, Ampicillin, Chloramphenicol, Kanamycin, Lincomycin, Rifampicin, and Vancomycin) against several pathogens including, MRSA, P. aeruginosa, and S. suis [80,81,82,83]. In one of the studies, the synergistic action of colistin, pediocin, or nisin Z is confirmed with ampicillin, rifampicin, and penicillin, which results in reduced MIC values against antibiotic resistance P. fluorescens [84]. In another study, melamine with ciprofloxacin was confirmed to have efficient synergistic action that eliminates fluoroquinolone-resistant P. aeruginosa [80]. Next, variants of indolicidin are confirmed to have a synergistic mechanism of action with tobramycin, polymyxin B, amikacin, and gentamicin to combat drug-resistant P. aeruginosa [85]. Further, arenicin-1, a marine AMP synergistically kills S. dermis, S. aureus, E. coli, and P. aeruginosa when used in combination with conventional antibiotics including erythromycin, ampicillin, and chloramphenicol. In addition to assisting in antibiotic absorption, arenicin-1 produces hydroxyl radicals in the cells thereby blocking bacterial growth [86]. Whilst there are various reports showing AMPs act synergistically with authorized antibiotics, further studies are required to determine the specific mechanisms of action involved (Table 4). The use of AMPs in combination with antibiotics provides a huge arsenal of new antimicrobials to fight against drug-resistant superbugs.
6. Challenges with AMPs for Therapeutic Applications against Superbugs
For future development of AMPs as therapeutics, limitations must be addressed despite their promising features. Although AMPs showed promising bioactivities against infections caused by superbugs, many limitations or disadvantages have to be overcome (Table 5). Stability is the biological system is one of the major challenges for AMPs when they are exposed to biological fluids such as serum and saliva. Unfortunately, AMPs showed significantly reduced antimicrobial potency in biological fluids when compared to the non-physiological conditions. Especially, AMPs get inactivated by host and bacterial proteases during infection due to high salt concentrations, anionic proteins, and polysaccharides in biological fluids [87]. Further, several pros and cons are mentioned in Table 5 that have a great impact on the way AMPs are developed for therapeutic applications. Additionally, the production of AMPs at the commercial level is expensive when compared to conventional antibiotics. Also, the identification, extraction, isolation, and purification of AMPs is very challenging. Especially, in the case of bacterial AMPs which are considered an enormous source of potentially novel AMPs due to huge unexplored diversity, however, several posttranslational modifications and the presence of other secondary metabolites make their purification and characterization more challenging. Further, the size of AMPs is one of the major obstacles in the way of their development in pharmacological applications. Non-specific cytotoxicity and limited routs of delivery are the major challenges due to the large size of AMPs when compared to conventional antibiotics. Overall, AMPs are providing an excellent and huge arsenal to combat drug-resistant superbugs, however, there is a long way to go before they can be used in therapeutic settings while considering the cons associated.
7. Conclusions and Future Directions
Since the discovery of nisin, the first bacterially produced AMP, much debate has surrounded the possibility of AMPs being considered a new anti-infective. It is been more than 35 years since nisin was FDA-approved as a safe food preservative and consumed along with food while no side effects reported so far. Also, nisin has been tested in multiple studies against many drug-resistant bacteria, however still far from therapeutic applications. There are many AMP derivatives and mimetics under clinical trial that suggest the potential of AMPs against drug-resistant superbugs. Overall, a multidisciplinary environment is essential to the development of new AMPs for therapeutic or pharmacological applications, involving several research areas, such as microbiology, medicinal chemistry, synthetic chemistry, and preclinical studies. By facilitating such collaborative and multidisciplinary work, it is possible to develop AMP-based antimicrobials that are effective against drug-resistant superbugs. Even though the current methods and technologies could be improved, we have also discussed the synergistic action of AMPs with conventional antibiotics that might provide a completely new and efficient set of antimicrobial formulations to fight against superbug infections. Overall, AMPs could be a promising, efficient, and enormous arsenal for alternatives to conventional antibiotics to combat drug-resistant superbug infections, however, further, detailed studies are needed to address the ongoing issues.
Author Contributions
SK and PB wrote the original draft. DR, SMM, and PB revised and edited the manuscript. PB proofread and edited the final draft. PB conceived the original idea and led the manuscript.
Acknowledgments
PB is thankful to the Animals Sciences Research Center, University of Missouri, Columbia for providing the necessary space and facilities to complete this work.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Salam, M.A.; Al-Amin, M.Y.; Salam, M.T.; Pawar, J.S.; Akhter, N.; Rabaan, A.A.; Alqumber, M.A.A. Antimicrobial Resistance: A Growing Serious Threat for Global Public Health. Healthc. 2023, 11, 1–20. [Google Scholar] [CrossRef]
- Lee Ventola, M.C. The Antibiotic Resistance Crisis Part 1: Causes and Threats. Compr. Biochem. 1963, 11, 181–224. [Google Scholar] [CrossRef]
- Matthyssen, T.; Li, W.; Holden, J.A.; Lenzo, J.C.; Hadjigol, S.; O’Brien-Simpson, N.M. The Potential of Modified and Multimeric Antimicrobial Peptide Materials as Superbug Killers. Front. Chem. 2022, 9, 1–23. [Google Scholar] [CrossRef]
- Terreni, M.; Taccani, M.; Pregnolato, M. New antibiotics for multidrug-resistant bacterial strains: Latest research developments and future perspectives. Molecules 2021, 26, 2671. [Google Scholar] [CrossRef]
- Mahlapuu, M.; Björn, C.; Ekblom, J. Antimicrobial peptides as therapeutic agents: Opportunities and challenges. Crit. Rev. Biotechnol. 2020, 40, 978–992. [Google Scholar] [CrossRef]
- Lei, J.; Sun, L.C.; Huang, S.; Zhu, C.; Li, P.; He, J.; Mackey, V.; Coy, D.H.; He, Q.Y. The antimicrobial peptides and their potential clinical applications. Am. J. Transl. Res. 2019, 11, 3919–3931. [Google Scholar]
- Walsh, C. Where will new antibiotics come from? Pharm. Eng. 2017, 37, 72. [Google Scholar] [CrossRef]
- Nagarajan, D.; Roy, N.; Kulkarni, O.; Nanajkar, N.; Datey, A.; Ravichandran, S.; Thakur, C.; Sandeep, T.; Aprameya, I.V.; Sarma, S.P.; et al. W76: A designed antimicrobial peptide to combat carbapenem- And tigecycline-resistant Acinetobacter baumannii. Sci. Adv. 2019, 5. [Google Scholar] [CrossRef]
- Chen, S.P.; Chen, E.H.L.; Yang, S.Y.; Kuo, P.S.; Jan, H.M.; Yang, T.C.; Hsieh, M.Y.; Lee, K.T.; Lin, C.H.; Chen, R.P.Y. A Systematic Study of the Stability, Safety, and Efficacy of the de novo Designed Antimicrobial Peptide PepD2 and Its Modified Derivatives Against Acinetobacter baumannii. Front. Microbiol. 2021, 12, 1–12. [Google Scholar] [CrossRef]
- Mohan, N.M.; Zorgani, A.; Jalowicki, G.; Kerr, A.; Khaldi, N.; Martins, M. Unlocking NuriPep 1653 From Common Pea Protein: A Potent Antimicrobial Peptide to Tackle a Pan-Drug Resistant Acinetobacter baumannii. Front. Microbiol. 2019, 10, 1–16. [Google Scholar] [CrossRef]
- Laxminarayan, R.; Duse, A.; Wattal, C.; Zaidi, A.K.M.; Wertheim, H.F.L.; Sumpradit, N.; Vlieghe, E.; Hara, G.L.; Gould, I.M.; Goossens, H.; et al. Antibiotic resistance-the need for global solutions. Lancet Infect. Dis. 2013, 13, 1057–1098. [Google Scholar] [CrossRef]
- Dadgostar, P.; Ng, W.J.; Hing, C.L.; Loo, C.B.; Hoh, E.K.; Loke, I.L.; Ee, K.Y. Ginger-Enriched Honey Attenuates Antibiotic Resistant Pseudomonas aeruginosa Quorum Sensing Virulence Factors and Biofilm Formation. Antibiotics 2023, 12, 1123. [Google Scholar] [CrossRef]
- Zaman, S.B.; Hussain, M.A.; Nye, R.; Mehta, V.; Mamun, K.T.; Hossain, N. A Review on Antibiotic Resistance: Alarm Bells are Ringing. Cureus 2017, 9. [Google Scholar] [CrossRef]
- Ruddaraju, L.K.; Pammi, S.V.N.; Guntuku, G.; Padavala, V.S.; Kolapalli, V.R.M. A review on anti-bacterials to combat resistance: From ancient era of plants and metals to present and future perspectives of green nano technological combinations. Asian J. Pharm. Sci. 2020, 15, 42–59. [Google Scholar] [CrossRef]
- Han, M.; Zhu, Y.; Creek, D.J.; Lin, Y.; Anderson, D.; Shen, H.; Tsuji, B.; Gutu, A.D.; Moskowitz, S.M.; Velkov, T.; et al. Alterations of Metabolic and Lipid Profiles in Polymyxin-. Antimicrob. Agents Chemother. 2018, 62, 1–14. [Google Scholar] [CrossRef]
- Hankins, J.V.; Madsen, J.A.; Giles, D.K.; Brodbelt, J.S.; Trent, M.S. Amino acid addition to Vibrio cholerae LPS establishes a link between surface remodeling in Gram-positive and Gram-negative bacteria. Proc. Natl. Acad. Sci. USA 2012, 109, 8722–8727. [Google Scholar] [CrossRef]
- Gan, B.H.; Gaynord, J.; Rowe, S.M.; Deingruber, T.; Spring, D.R. The multifaceted nature of antimicrobial peptides: Current synthetic chemistry approaches and future directions. Chem. Soc. Rev. 2021, 50, 7820–7880. [Google Scholar] [CrossRef]
- Zhang, Y.M.; Rock, C.O. Membrane lipid homeostasis in bacteria. Nat. Rev. Microbiol. 2008, 6, 222–233. [Google Scholar] [CrossRef]
- Matsuzaki, K. Control of cell selectivity of antimicrobial peptides. Biochim. Biophys. Acta-Biomembr. 2009, 1788, 1687–1692. [Google Scholar] [CrossRef]
- Verkleij, A.J.; Zwaal, R.F.; Roelofsen, B.; Comfurius, P.; Kastelijn, D.; Van Deenen, L.L. The asymmetric distribution of phospholipids in the human red cell membrane. Bba 1973, 21, 1154–1157. [Google Scholar]
- Yount, M.R.Y. and N.Y. Mechanisms of antimicrobial peptide action and resistance. Pharmacol. Rev. 2003, 55, 453–499. [Google Scholar] [CrossRef]
- Jiang, Z.; Vasil, A.I.; Hale, J.D.; Hancock, R.E.W.; Vasil, M.L.; Hodges, R.S. Effects of net charge and the number of positively charged residues on the biological activity of amphipathic α-helical cationic antimicrobial peptides. Biopolym.-Pept. Sci. Sect. 2008, 90, 369–383. [Google Scholar] [CrossRef]
- Paulmann, M.; Arnold, T.; Linke, D.; Özdirekcan, S.; Kopp, A.; Gutsmann, T.; Kalbacher, H.; Wanke, I.; Schuenemann, V.J.; Habeck, M.; et al. Structure-activity analysis of the dermcidin-derived peptide DCD-1L, an anionic antimicrobial peptide present in human sweat. J. Biol. Chem. 2012, 287, 8434–8443. [Google Scholar] [CrossRef]
- Cytryńska, M.; Zdybicka-Barabas, A. Defense peptides: Recent developments. Biomol. Concepts 2015, 6, 237–251. [Google Scholar] [CrossRef]
- Bellomio, A.; Vincent, P.A.; De Arcuri, B.F.; Farias, R.N.; Morero, R.D. Microcin J25 has dual and independent mechanisms of action in Escherichia coli: RNA polymerase inhibition and increased superoxide production. J. Bacteriol. 2007, 189, 4180–4186. [Google Scholar] [CrossRef]
- Malanovic, N.; Lohner, K. Gram-positive bacterial cell envelopes: The impact on the activity of antimicrobial peptides. Biochim. Biophys. Acta-Biomembr. 2016, 1858, 936–946. [Google Scholar] [CrossRef]
- Ofek, I.; Cohen, S.; Rahmani, R.; Kabha, K.; Tamarkin, D.; Herzig, Y.; Rubinstein, E. Antibacterial synergism of polymyxin B nonapeptide and hydrophobic antibiotics in experimental gram-negative infections in mice. Antimicrob. Agents Chemother. 1994, 38, 374–377. [Google Scholar] [CrossRef]
- Vollmer, W.; Blanot, D.; De Pedro, M.A. Peptidoglycan structure and architecture. FEMS Microbiol. Rev. 2008, 32, 149–167. [Google Scholar] [CrossRef]
- Le, C.F. Intracellular Targeting Mechanisms by Antimicrobial Peptides. Antimicrob. Agents Chemother. 2017, 61, 1–16. [Google Scholar] [CrossRef]
- Nicolas, P. Multifunctional host defense peptides: Intracellular-targeting antimicrobial peptides. FEBS J. 2009. [CrossRef]
- Park, C.B.; Yi, K.S.; Matsuzaki, K.; Kim, M.S.; Kim, S.C. Structure-activity analysis of buforin II, a histone H2A-derived antimicrobial peptide: The proline hinge is responsible for the cell-penetrating ability of buforin II. Proc. Natl. Acad. Sci. USA 2000, 97, 8245–8250. [Google Scholar] [CrossRef]
- Park, C.B.; Kim, H.S.; Kim, S.C. Mechanism of action of the antimicrobial peptide buforin II: Buforin II kills microorganisms by penetrating the cell membrane and inhibiting cellular functions. Biochem. Biophys. Res. Commun. 1998, 244, 253–257. [Google Scholar] [CrossRef]
- Selsted, M.E.; Novotny, M.J.; Morris, W.L.; Tang, Y.Q.; Smith, W.; Cullor, J.S. Indolicidin, a novel bactericidal tridecapeptide amide from neutrophils. J. Biol. Chem. 1992, 267, 4292–4295. [Google Scholar] [CrossRef]
- Falla, T.J.; Nedra Karunaratne, D.; Hancock, R.E.W. Mode of action of the antimicrobial peptide indolicidin. J. Biol. Chem. 1996, 271, 19298–19303. [Google Scholar] [CrossRef]
- Subbalakshmi, C.; Krishnakumari, V.; Nagaraj, R.; Sitaram, N. Requirements for antibacterial and hemolytic activities in the bovine neutrophil derived 13-residue peptide indolicidin. FEBS Lett. 1996, 395, 48–52. [Google Scholar] [CrossRef]
- Subbalakshmi, C.; Sitaram, N. Mechanism of antimicrobial action of indolicidin. FEMS Microbiol. Lett. 1998, 160, 91–96. [Google Scholar] [CrossRef]
- Hsu, C.H.; Chen, C.; Jou, M.L.; Lee, A.Y.L.; Lin, Y.C.; Yu, Y.P.; Huang, W.T.; Wu, S.H. Structural and DNA-binding studies on the bovine antimicrobial peptide, indolicidin: Evidence for multiple conformations involved in binding to membranes and DNA. Nucleic Acids Res. 2005, 33, 4053–4064. [Google Scholar] [CrossRef]
- Ghosh, A.; Kar, R.K.; Jana, J.; Saha, A.; Jana, B.; Krishnamoorthy, J.; Kumar, D.; Ghosh, S.; Chatterjee, S.; Bhunia, A. Indolicidin targets duplex DNA: Structural and mechanistic insight through a combination of spectroscopy and microscopy. ChemMedChem 2014, 9, 2052–2058. [Google Scholar] [CrossRef]
- Adelman, K.; Yuzenkova, J.; La Porta, A.; Zenkin, N.; Lee, J.; Lis, J.T.; Borukhov, S.; Wang, M.D.; Severinov, K. Molecular mechanism of transcription inhibition by peptide antibiotic Microcin J25. Mol. Cell 2004, 14, 753–762. [Google Scholar] [CrossRef]
- Ho, Y.H.; Sung, T.C.; Chen, C.S. Lactoferricin B inhibits the phosphorylation of the two-component system response regulators BasR and CreB. Mol. Cell. Proteomics 2012, 11, 1–10. [Google Scholar] [CrossRef]
- Tu, Y.H.; Ho, Y.H.; Chuang, Y.C.; Chen, P.C.; Chen, C.S. Identification of lactoferricin B intracellular targets using an escherichia coli proteome chip. PLoS ONE 2011, 6, e0028197. [Google Scholar] [CrossRef]
- Schnapp, D.; Kemp, G.D.; Smith, V.J. Purification and characterization of a proline-rich antibacterial peptide, pdf. 1996, 539, 532–539. [Google Scholar]
- Mardirossian, M.; Grzela, R.; Giglione, C.; Meinnel, T.; Gennaro, R.; Mergaert, P.; Scocchi, M. The host antimicrobial peptide Bac71-35 binds to bacterial ribosomal proteins and inhibits protein synthesis. Chem. Biol. 2014, 21, 1639–1647. [Google Scholar] [CrossRef]
- Le, C.F.; Yusof, M.Y.M.; Hassan, H.; Sekaran, S.D. In vitro properties of designed antimicrobial peptides that exhibit potent antipneumococcal activity and produces synergism in combination with penicillin. Sci. Rep. 2015, 5, 1–8. [Google Scholar] [CrossRef]
- Le, C.F.; Gudimella, R.; Razali, R.; Manikam, R.; Sekaran, S.D. Transcriptome analysis of Streptococcus pneumoniae treated with the designed antimicrobial peptides, DM3. Sci. Rep. 2016, 6, 1–9. [Google Scholar] [CrossRef]
- Hancock, R.E.W.; Haney, E.F.; Gill, E.E. The immunology of host defence peptides: Beyond antimicrobial activity. Nat. Rev. Immunol. 2016, 16, 321–334. [Google Scholar] [CrossRef]
- Duarte-Mata, D.I.; Salinas-Carmona, M.C. Antimicrobial peptides’ immune modulation role in intracellular bacterial infection. Front. Immunol. 2023, 14, 1–14. [Google Scholar] [CrossRef]
- Hiemstra, P.S. Defensins and cathelicidins in inflammatory lung disease: Beyond antimicrobial activity. Biochem. Soc. Trans. 2006, 34, 276–278. [Google Scholar] [CrossRef]
- Tan, B.H.; Meinken, C.; Bastian, M.; Bruns, H.; Legaspi, A.; Ochoa, M.T.; Krutzik, S.R.; Bloom, B.R.; Ganz, T.; Modlin, R.L.; et al. Macrophages Acquire Neutrophil Granules for Antimicrobial Activity against Intracellular Pathogens. J. Immunol. 2006. [Google Scholar] [CrossRef]
- Zheng, Y.; Niyonsaba, F.; Ushio, H.; Nagaoka, I.; Ikeda, S.; Okumura, K.; Ogawa, H. Cathelicidin LL-37 induces the generation of reactive oxygen species and release of human α-defensins from neutrophils. Br. J. Dermatol. 2007. [Google Scholar] [CrossRef]
- Suarez-Carmona, M.; Hubert, P.; Delvenne, P.; Herfs, M. Defensins: “Simple” antimicrobial peptides or broad-spectrum molecules? Cytokine Growth Factor Rev. 2015, 26, 361–370. [Google Scholar] [CrossRef]
- Yang, D.; Chen, Q.; Chertov, O.; Oppenheim, J.J. Human neutrophil defensins selectively chemoattract naive T and immature dendritic cells. J. Leukoc. Biol. 2000, 68, 9–14. [Google Scholar] [CrossRef]
- García, J.R.C.; Jaumann, F.; Schulz, S.; Krause, A.; Rodríguez-Jiménez, J.; Forssmann, U.; Adermann, K.; Klüver, E.; Vogelmeier, C.; Becker, D.; et al. Identification of a novel, multifunctional β-defensin (human β-defensin 3) with specific antimicrobial activity: Its interaction with plasma membranes of Xenopus oocytes and the induction of macrophage chemoattraction. Cell Tissue Res. 2001, 306, 257–264. [Google Scholar] [CrossRef]
- Röhrl, J.; Yang, D.; Oppenheim, J.J.; Hehlgans, T. Human β-Defensin 2 and 3 and Their Mouse Orthologs Induce Chemotaxis through Interaction with CCR2. J. Immunol. 2010, 184, 6688–6694. [Google Scholar] [CrossRef]
- Lim, H.K.; O’Neill, H.C. Identification of Stromal Cells in Spleen Which Support Myelopoiesis. Front. Cell Dev. Biol. 2019, 7, 1–13. [Google Scholar] [CrossRef]
- Engelmayer, J.; Blezinger, P.; Varadhachary, A. Talactoferrin Stimulates Wound Healing With Modulation of Inflammation. J. Surg. Res. 2008, 149, 278–286. [Google Scholar] [CrossRef]
- Gaio, V.; Cerca, N. Cells released from S. epidermidis biofilms present increased antibiotic tolerance to multiple antibiotics. PeerJ 2019, 2019. [Google Scholar] [CrossRef]
- Vuotto, C.; Longo, F.; Balice, M.P.; Donelli, G.; Varaldo, P.E. Antibiotic resistance related to biofilm formation in Klebsiella pneumoniae. Pathogens 2014, 3, 743–758. [Google Scholar] [CrossRef]
- Amankwah, S.; Abdella, K.; Kassa, T. Bacterial biofilm destruction: A focused review on the recent use of phage-based strategies with other antibiofilm agents. Nanotechnol. Sci. Appl. 2021, 14, 161–177. [Google Scholar] [CrossRef]
- Shariati, A.; Arshadi, M.; Khosrojerdi, M.A.; Abedinzadeh, M.; Ganjalishahi, M.; Maleki, A.; Heidary, M.; Khoshnood, S. The resistance mechanisms of bacteria against ciprofloxacin and new approaches for enhancing the efficacy of this antibiotic. Front. Public Heal. 2022, 10, 1025633. [Google Scholar] [CrossRef]
- Wood, T.K.; Knabel, S.J.; Kwan, B.W. Bacterial persister cell formation and dormancy. Appl. Environ. Microbiol. 2013, 79, 7116–7121. [Google Scholar] [CrossRef]
- Khatoon, Z.; McTiernan, C.D.; Suuronen, E.J.; Mah, T.F.; Alarcon, E.I. Bacterial biofilm formation on implantable devices and approaches to its treatment and prevention. Heliyon 2018, 4, e01067. [Google Scholar] [CrossRef]
- Chung, P.Y.; Khanum, R. Antimicrobial peptides as potential anti-biofilm agents against multidrug-resistant bacteria. J. Microbiol. Immunol. Infect. 2017, 50, 405–410. [Google Scholar] [CrossRef]
- Dean, S.N.; Bishop, B.M.; van Hoek, M.L. Natural and synthetic cathelicidin peptides with anti-microbial and anti-biofilm activity against Staphylococcus aureus. BMC Microbiol. 2011, 11, 114. [Google Scholar] [CrossRef]
- Overhage, J.; Campisano, A.; Bains, M.; Torfs, E.C.W.; Rehm, B.H.A.; Hancock, R.E.W. Human host defense peptide LL-37 prevents bacterial biofilm formation. Infect. Immun. 2008, 76, 4176–4182. [Google Scholar] [CrossRef]
- Hell, É.; Giske, C.G.; Nelson, A.; Römling, U.; Marchini, G. Human cathelicidin peptide LL37 inhibits both attachment capability and biofilm formation of Staphylococcus epidermidis. Lett. Appl. Microbiol. 2010, 50, 211–215. [Google Scholar] [CrossRef]
- Yoshinari, M.; Kato, T.; Matsuzaka, K.; Hayakawa, T.; Shiba, K. Prevention of biofilm formation on titanium surfaces modified with conjugated molecules comprised of antimicrobial and titanium-binding peptides. Biofouling 2010, 26, 103–110. [Google Scholar] [CrossRef]
- Gopal, R.; Kim, Y.G.; Lee, J.H.; Lee, S.K.; Chae, J.D.; Son, B.K.; Seo, C.H.; Park, Y. Synergistic effects and antibiofilm properties of chimeric peptides against multidrug-resistant acinetobacter baumannii strains. Antimicrob. Agents Chemother. 2014, 58, 1622–1629. [Google Scholar] [CrossRef]
- Yasir, M.; Willcox, M.D.P.; Dutta, D. Action of antimicrobial peptides against bacterial biofilms. Materials 2018, 11, 2468. [Google Scholar] [CrossRef]
- Mermer, S.; Turhan, T.; Bolat, E.; Aydemir, S.; Yamazhan, T.; Pullukcu, H.; Arda, B.; Sipahi, H.; Ulusoy, S.; Sipahi, O.R. Ceftaroline versus vancomycin in the treatment of methicillin-resistant Staphylococcus aureus (MRSA) in an experimental MRSA meningitis model. J. Glob. Antimicrob. Resist. 2020, 22, 147–151. [Google Scholar] [CrossRef]
- Gause, G.F.; Brazhnikova, M.G. Gramicidin S and its use in the treatment of infected wounds. Nature 1944, 154, 703. [Google Scholar] [CrossRef]
- UMBREIT, W.W. Mechanisms of antibacterial action. Pharmacol. Rev. 1953, 5, 275–284. [Google Scholar] [CrossRef]
- Sahoo, S.; Mohanty, J.N.; Routray, S.P.; Khandia, R.; Das, J.; Shah, S.; Swarnkar, T. Colistin the last resort drug in 21st century antibiotics to combat Multidrug resistance superbugs. J. Exp. Biol. Agric. Sci. 2023, 11, 919–929. [Google Scholar] [CrossRef]
- Ye, Y.; Xia, Z.; Zhang, D.; Sheng, Z.; Zhang, P.; Zhu, H.; Xu, N.; Liang, S. Multifunctional pharmaceutical effects of the antibiotic daptomycin. Biomed Res. Int. 2019, 2019, 8609218. [Google Scholar] [CrossRef]
- Landman, D.; Georgescu, C.; Martin, D.A.; Quale, J. Polymyxins revisited. Clin. Microbiol. Rev. 2008, 21, 449–465. [Google Scholar] [CrossRef]
- Xuan, J.; Feng, W.; Wang, J.; Wang, R.; Zhang, B.; Bo, L.; Chen, Z.S.; Yang, H.; Sun, L. Antimicrobial peptides for combating drug-resistant bacterial infections. Drug Resist. Updat. 2023, 68. [Google Scholar] [CrossRef]
- Baindara, P.; Dinata, R.; Mandal, S.M. Marine Bacteriocins: An Evolutionary Gold Mine to Payoff Antibiotic Resistance. Mar. Drugs 2024, 22. [Google Scholar] [CrossRef]
- Kalita, A.; Verma, I.; Khuller, G.K. Role of human neutrophil peptide-1 as a possible adjunct to antituberculosis chemotherapy. J. Infect. Dis. 2004, 190, 1476–1480. [Google Scholar] [CrossRef]
- Baindara, P.; Singh, N.; Ranjan, M.; Nallabelli, N.; Chaudhry, V.; Pathania, G.L.; Sharma, N.; Kumar, A.; Patil, P.B.; Korpole, S. Laterosporulin10: A novel defensin like class iid bacteriocin from brevibacillus sp. strain SKDU10 with inhibitory activity against microbial pathogens. Microbiol. (United Kingdom) 2016, 162, 1286–1299. [Google Scholar] [CrossRef]
- Naghmouchi, K.; Le Lay, C.; Baah, J.; Drider, D. Antibiotic and antimicrobial peptide combinations: Synergistic inhibition of Pseudomonas fluorescens and antibiotic-resistant variants. Res. Microbiol. 2012, 163, 101–108. [Google Scholar] [CrossRef]
- Brumfitt, W.; Salton, M.R.J.; Hamilton-Miller, J.M.T. Nisin, alone and combined with peptidoglycan-modulating antibiotics: Activity against methicillin-resistant Staphylococcus aureus and vancomycin-resistant enterococci. J. Antimicrob. Chemother. 2002, 50, 731–734. [Google Scholar] [CrossRef]
- Giacometti, A.; Cirioni, O.; Barchiesi, F.; Fortuna, M.; Scalise, G. In-vitro activity of cationic peptides alone and in combination with clinically used antimicrobial agents against Pseudomonas aeruginosa. J. Antimicrob. Chemother. 1999, 44, 641–645. [Google Scholar] [CrossRef]
- Lebel, G.; Piché, F.; Frenette, M.; Gottschalk, M.; Grenier, D. Antimicrobial activity of nisin against the swine pathogen Streptococcus suis and its synergistic interaction with antibiotics. Peptides 2013, 50, 19–23. [Google Scholar] [CrossRef]
- Mataraci, E.; Dosler, S. In vitro activities of antibiotics and antimicrobial cationic peptides alone and in combination against methicillin-resistant Staphylococcus aureus biofilms. Antimicrob. Agents Chemother. 2012, 56, 6366–6371. [Google Scholar] [CrossRef]
- De Gier, M.G.; Bauke Albada, H.; Josten, M.; Willems, R.; Leavis, H.; Van Mansveld, R.; Paganelli, F.L.; Dekker, B.; Lammers, J.W.J.; Sahl, H.G.; et al. Synergistic activity of a short lipidated antimicrobial peptide (lipoAMP) and colistin or tobramycin against Pseudomonas aeruginosa from cystic fibrosis patients. Medchemcomm 2016, 7, 148–156. [Google Scholar] [CrossRef]
- Duong, L.; Gross, S.P.; Siryaporn, A. Developing Antimicrobial Synergy With AMPs. Front. Med. Technol. 2021, 3, 640981. [Google Scholar] [CrossRef]
- Bowdish, D.; Davidson, D.; Hancock, R. A Re-evaluation of the Role of Host Defence Peptides in Mammalian Immunity. Curr. Protein Pept. Sci. 2005, 6, 35–51. [Google Scholar] [CrossRef]
- Arsene, M.M.J.; Jorelle, A.B.J.; Sarra, S.; Viktorovna, P.I.; Davares, A.K.L.; Ingrid, N.K.C.; Steve, A.A.F.; Andreevna, S.L.; Vyacheslavovna, Y.N.; Carime, B.Z. Short review on the potential alternatives to antibiotics in the era of antibiotic resistance. J. Appl. Pharm. Sci. 2022, 12, 029–040. [Google Scholar] [CrossRef]
- Miller, W.R.; Bayer, A.S.; Arias, C.A. Mechanism of action and resistance to daptomycin in Staphylococcus aureus and enterococci. Cold Spring Harb. Perspect. Med. 2016, 6, a026997. [Google Scholar] [CrossRef]
- Trimble, M.J.; Mlynárčik, P.; Kolář, M.; Hancock, R.E.W. Polymyxin: Alternative mechanisms of action and resistance. Cold Spring Harb. Perspect. Med. 2016, 6, a025288. [Google Scholar] [CrossRef]
- Prince, A.; Sandhu, P.; Kumar, P.; Dash, E.; Sharma, S.; Arakha, M.; Jha, S.; Akhter, Y.; Saleem, M. Lipid-II Independent Antimicrobial Mechanism of Nisin Depends on Its Crowding and Degree of Oligomerization. Sci. Rep. 2016, 6, 37908. [Google Scholar] [CrossRef]
- Srinivas, N.; Jetter, P.; Ueberbacher, B.J.; Werneburg, M.; Zerbe, K.; Steinmann, J.; Van Der Meijden, B.; Bernardini, F.; Lederer, A.; Dias, R.L.A.; et al. Peptidomimetic antibiotics target outer-membrane biogenesis in pseudomonas aeruginosa. Science (80-. ). 2010, 327, 1010–1013. [Google Scholar] [CrossRef]
- Landa, A.; Jiménez, L.; Willms, K.; Jiménez-García, L.F.; Lara-Martínez, R.; Robert, L.; Cirioni, O.; Barańska-Rybak, W.; Kamysz, W. Antimicrobial peptides (Temporin A and Iseganan IB-367): Effect on the cysticerci of Taenia crassiceps. Mol. Biochem. Parasitol. 2009, 164, 126–130. [Google Scholar] [CrossRef]
- Alam, M.Z.; Wu, X.; Mascio, C.; Chesnel, L.; Hurdle, J.G. Mode of action and bactericidal properties of surotomycin against growing and nongrowing clostridium difficile. Antimicrob. Agents Chemother. 2015, 59, 5165–5170. [Google Scholar] [CrossRef]
- Rubinchik, E.; Dugourd, D.; Algara, T.; Pasetka, C.; Friedland, H.D. Antimicrobial and antifungal activities of a novel cationic antimicrobial peptide, omiganan, in experimental skin colonisation models. Int. J. Antimicrob. Agents 2009, 34, 457–461. [Google Scholar] [CrossRef]
- Crowther, G.S.; Baines, S.D.; Todhunter, S.L.; Freeman, J.; Chilton, C.H.; Wilcox, M.H. Evaluation of NVB302 versus vancomycin activity in an in vitro human gut model of Clostridium difficile infection. J. Antimicrob. Chemother. 2013, 68, 168–176. [Google Scholar] [CrossRef]
- Malanovic, N.; Leber, R.; Schmuck, M.; Kriechbaum, M.; Cordfunke, R.A.; Drijfhout, J.W.; De Breij, A.; Nibbering, P.H.; Kolb, D.; Lohner, K. Phospholipid-driven differences determine the action of the synthetic antimicrobial peptide OP-145 on Gram-positive bacterial and mammalian membrane model systems. Biochim. Biophys. Acta-Biomembr. 2015, 1848, 2437–2447. [Google Scholar] [CrossRef]
- Sivertsen, A.; Isaksson, J.; Leiros, H.K.S.; Svenson, J.; Svendsen, J.S.; Brandsdal, B.O. Synthetic cationic antimicrobial peptides bind with their hydrophobic parts to drug site II of human serum albumin. BMC Struct. Biol. 2014, 14, 1–14. [Google Scholar] [CrossRef]
- Van Groenendael, R.; Kox, M.; Van Eijk, L.T.; Pickkers, P. Immunomodulatory and kidney-protective effects of the human chorionic gonadotropin derivate EA-230. Nephron 2018, 140, 148–151. [Google Scholar] [CrossRef]
- PJM Brouwer, C. Structure-Activity Relationship Study of Synthetic Variants Derived from the Highly Potent Human Antimicrobial Peptide hLF(1-11). Cohesive J. Microbiol. Infect. Dis. 2018, 1, 1–19. [Google Scholar] [CrossRef]
- Guo, L.; McLean, J.S.; Yang, Y.; Eckert, R.; Kaplan, C.W.; Kyme, P.; Sheikh, O.; Varnum, B.; Lux, R.; Shi, W.; et al. Precision-guided antimicrobial peptide as a targeted modulator of human microbial ecology. Proc. Natl. Acad. Sci. USA 2015, 112, 7569–7574. [Google Scholar] [CrossRef]
- Fulco, P.; Wenzel, R.P. Ramoplanin: A topical lipoglycodepsipeptide antibacterial agent. Expert Rev. Anti. Infect. Ther. 2006, 4, 939–945. [Google Scholar] [CrossRef]
- Bulger, E.M.; Maier, R.V.; Sperry, J.; Joshi, M.; Henry, S.; Moore, F.A.; Moldawer, L.L.; Demetriades, D.; Talving, P.; Schreiber, M.; et al. A novel drug for treatment of necrotizing soft-tissue infections: A randomized clinical trial. JAMA Surg. 2014, 149, 528–536. [Google Scholar] [CrossRef]
- Muchintala, D.; Suresh, V.; Raju, D.; Sashidhar, R.B. Synthesis and characterization of cecropin peptide-based silver nanocomposites: Its antibacterial activity and mode of action. Mater. Sci. Eng. C 2020, 110, 110712. [Google Scholar] [CrossRef]
- Leeds, J.A.; Sachdeva, M.; Mullin, S.; Dzink-Fox, J.; LaMarche, M.J. Mechanism of action of and mechanism of reduced susceptibility to the novel anti-Clostridium difficile compound LFF571. Antimicrob. Agents Chemother. 2012, 56, 4463–4465. [Google Scholar] [CrossRef]
- Peyrusson, F.; Butler, D.; Tulkens, P.M.; Van Bambeke, F. Cellular pharmacokinetics and intracellular activity of the novel peptide deformylase inhibitor GSK1322322 against Staphylococcus aureus laboratory and clinical strains with various resistance phenotypes: Studies with human THP-1 monocytes and J774 murine. Antimicrob. Agents Chemother. 2015, 59, 5747–5760. [Google Scholar] [CrossRef]
- Mensa, B.; Howell, G.L.; Scott, R.; DeGrado, W.F. Comparative mechanistic studies of brilacidin, daptomycin, and the antimicrobial peptide LL16. Antimicrob. Agents Chemother. 2014, 58, 5136–5145. [Google Scholar] [CrossRef]
- Ooi, N.; Miller, K.; Hobbs, J.; Rhys-Williams, W.; Love, W.; Chopra, I. XF-73, a novel antistaphylococcal membrane-active agent with rapid bactericidal activity. J. Antimicrob. Chemother. 2009, 64, 735–740. [Google Scholar] [CrossRef]
- Itoh, H.; Tokumoto, K.; Kaji, T.; Paudel, A.; Panthee, S.; Hamamoto, H.; Sekimizu, K.; Inoue, M. Total Synthesis and Biological Mode of Action of WAP-8294A2: A Menaquinone-Targeting Antibiotic. J. Org. Chem. 2018, 83, 6924–6935. [Google Scholar] [CrossRef]
- Miyake, O.; Ochiai, A.; Hashimoto, W.; Murata, K. Origin and Diversity of Alginate Lyases of Families PL-5 and -7 in Sphingomonas sp. Strain A1. J. Bacteriol. 2004, 186, 2891–2896. [Google Scholar] [CrossRef]
- Yu, H.B.; Kielczewska, A.; Rozek, A.; Takenaka, S.; Li, Y.; Thorson, L.; Hancock, R.E.W.; Guarna, M.M.; North, J.R.; Foster, L.J.; et al. Sequestosome-1/p62 is the key intracellular target of innate defense regulator peptide. J. Biol. Chem. 2009, 284, 36007–36011. [Google Scholar] [CrossRef]
- Kruszewska, D.; Sahl, H.G.; Bierbaum, G.; Pag, U.; Hynes, S.O.; Ljungh, Å. Mersacidin eradicates methicillin-resistant Staphylococcus aureus (MRSA) in a mouse rhinitis model. J. Antimicrob. Chemother. 2004, 54, 648–653. [Google Scholar] [CrossRef]
- Ekkelenkamp, M.B.; Hanssen, M.; Hsu, S.T.D.; De Jong, A.; Milatovic, D.; Verhoef, J.; Van Nuland, N.A.J. Isolation and structural characterization of epilancin 15X, a novel lantibiotic from a clinical strain of Staphylococcus epidermidis. FEBS Lett. 2005, 579, 1917–1922. [Google Scholar] [CrossRef]
- Mota-Meira, M.; Morency, H.; Lavoie, M.C. In vivo activity of mutacin B-Ny266. J. Antimicrob. Chemother. 2005, 56, 869–871. [Google Scholar] [CrossRef]
- Halliwell, S.; Warn, P.; Sattar, A.; Derrick, J.P.; Upton, M. A single dose of epidermicin NI01 is sufficient to eradicate MRSA from the nares of cotton rats. J. Antimicrob. Chemother. 2017, 72, 778–781. [Google Scholar] [CrossRef]
- Kokai-Kun, J.F.; Walsh, S.M.; Chanturiya, T.; Mond, J.J. Lysostaphin cream eradicates Staphylococcus aureus nasal colonization in a cotton rat model. Antimicrob. Agents Chemother. 2003, 47, 1589–1597. [Google Scholar] [CrossRef]
- Kers, J.A.; Sharp, R.E.; Defusco, A.W.; Park, J.H.; Xu, J.; Pulse, M.E.; Weiss, W.J.; Handfield, M. Mutacin 1140 lantibiotic variants are efficacious against Clostridium difficile infection. Front. Microbiol. 2018, 9, 415. [Google Scholar] [CrossRef]
- Kers, J.A.; DeFusco, A.W.; Park, J.H.; Xu, J.; Pulse, M.E.; Weiss, W.J.; Handfield, M. OG716: Designing a fit-for-purpose lantibiotic for the treatment of Clostridium difficile infections. PLoS ONE 2018, 13, e0197467. [Google Scholar] [CrossRef]
- Pulse, M.E.; Weiss, W.J.; Kers, J.A.; DeFusco, A.W.; Park, J.H.; Handfield, M. Pharmacological, toxicological, and dose range assessment of OG716, a novel lantibiotic for the treatment of clostridium difficile-associated infection. Antimicrob. Agents Chemother. 2019, 63, 10–1128. [Google Scholar] [CrossRef]
- Boakes, S.; Ayala, T.; Herman, M.; Appleyard, A.N.; Dawson, M.J.; Cortés, J. Generation of an actagardine A variant library through saturation mutagenesis. Appl. Microbiol. Biotechnol. 2012, 95, 1509–1517. [Google Scholar] [CrossRef]
- Netz, D.J.A.; Pohl, R.; Beck-Sickinger, A.G.; Selmer, T.; Pierik, A.J.; Bastos, M.D.C.D.F.; Sahl, H.G. Biochemical characterisation and genetic analysis of aureocin A53, a new, atypical bacteriocin from Staphylococcus aureus. J. Mol. Biol. 2002, 319, 745–756. [Google Scholar] [CrossRef]
- Vassiliadis, G.; Destoumieux-Garzón, D.; Lombard, C.; Rebuffat, S.; Peduzzi, J. Isolation and characterization of two members of the siderophore-microcin family, microcins M and H47. Antimicrob. Agents Chemother. 2010, 54, 288–297. [Google Scholar] [CrossRef]
- Gavrish, E.; Sit, C.S.; Cao, S.; Kandror, O.; Spoering, A.; Peoples, A.; Ling, L.; Fetterman, A.; Hughes, D.; Bissell, A.; et al. Lassomycin, a ribosomally synthesized cyclic peptide, kills mycobacterium tuberculosis by targeting the ATP-dependent protease ClpC1P1P2. Chem. Biol. 2014, 21, 509–518. [Google Scholar] [CrossRef]
- Aguilar-Pérez, C.; Gracia, B.; Rodrigues, L.; Vitoria, A.; Cebrián, R.; Deboosère, N.; Song, O.; Brodin, P.; Maqueda, M.; Aínsaa, J.A. Synergy between circular bacteriocin AS-48 and ethambutol against mycobacterium tuberculosis. Antimicrob. Agents Chemother. 2018, 62. [Google Scholar] [CrossRef]
- Hanchi, H.; Hammami, R.; Gingras, H.; Kourda, R.; Bergeron, M.G.; Ben Hamida, J.; Ouellette, M.; Fliss, I. Inhibition of MRSA and of Clostridium difficile by durancin 61A: Synergy with bacteriocins and antibiotics. Future Microbiol. 2017, 12, 205–212. [Google Scholar] [CrossRef]
- Balty, C.; Guillot, A.; Fradale, L.; Brewee, C.; Boulay, M.; Kubiak, X.; Benjdia, A.; Berteau, O. Ruminococcin C, an anti-clostridial sactipeptide produced by a prominent member of the human microbiota Ruminococcus gnavus. J. Biol. Chem. 2019, 294, 14512–14525. [Google Scholar] [CrossRef]
- Gebhart, D.; Lok, S.; Clare, S.; Tomas, M.; Stares, M.; Scholl, D.; Donskey, C.J.; Lawley, T.D.; Govoni, G.R. A modified R-type bacteriocin specifically targeting Clostridium difficile prevents colonization of mice without affecting gut microbiota diversity. MBio 2015, 6. [Google Scholar] [CrossRef]
- Zhu, S.; Gao, B.; Umetsu, Y.; Peigneur, S.; Li, P.; Ohki, S.; Tytgat, J. Adaptively evolved human oral actinomyces-sourced defensins show therapeutic potential. EMBO Mol. Med. 2022, 14. [Google Scholar] [CrossRef]
- Shen, B.; Song, J.; Zhao, Y.; Zhang, Y.; Liu, G.; Li, X.; Guo, X.; Li, W.; Cao, Z.; Wu, Y. Triintsin, a human pathogenic fungus-derived defensin with broad-spectrum antimicrobial activity. Peptides 2018, 107, 61–67. [Google Scholar] [CrossRef]
- Zhang, Z.J.; Wu, C.; Moreira, R.; Dorantes, D.; Pappas, T.; Sundararajan, A.; Lin, H.; Pamer, E.G.; van der Donk, W.A. Activity of Gut-Derived Nisin-like Lantibiotics against Human Gut Pathogens and Commensals. ACS Chem. Biol. 2024, 19, 357–369. [Google Scholar] [CrossRef]
- Baindara, P.; Chaudhry, V.; Mittal, G.; Liao, L.M.; Matos, C.O.; Khatri, N.; Franco, O.L.; Patil, P.B.; Korpole, S. Characterization of the antimicrobial peptide penisin, a class Ia novel lantibiotic from Paenibacillus sp. strain A3. Antimicrob. Agents Chemother. 2016. [Google Scholar] [CrossRef]
- Baindara, P.; Roy, D.; Mandal, S.M. CycP: A Novel Self-Assembled Vesicle-Forming Cyclic Antimicrobial Peptide to Control Drug-Resistant S. aureus. Bioengineering 2024, 11, 855. [Google Scholar] [CrossRef]
- Mygind, P.H.; Fischer, R.L.; Schnorr, K.M.; Hansen, M.T.; Sönksen, C.P.; Ludvigsen, S.; Raventós, D.; Buskov, S.; Christensen, B.; De Maria, L.; et al. Plectasin is a peptide antibiotic with therapeutic potential from a saprophytic fungus. Nature 2005, 437, 975–980. [Google Scholar] [CrossRef]
- Zhu, S.; Gao, B.; Harvey, P.J.; Craik, D.J. Dermatophytic defensin with antiinfective potential. Proc. Natl. Acad. Sci. USA 2012, 109, 8495–8500. [Google Scholar] [CrossRef]
- Dobson, A.; O’Connor, P.M.; Cotter, P.D.; Ross, R.P.; Hill, C. Impact of the broad-spectrum antimicrobial peptide, lacticin 3147, on Streptococcus mutans growing in a biofilm and in human saliva. J. Appl. Microbiol. 2011, 111, 1515–1523. [Google Scholar] [CrossRef]
- Bolocan, A.S.; Pennone, V.; O’Connor, P.M.; Coffey, A.; Nicolau, A.I.; McAuliffe, O.; Jordan, K. Inhibition of Listeria monocytogenes biofilms by bacteriocin-producing bacteria isolated from mushroom substrate. J. Appl. Microbiol. 2017, 122, 279–293. [Google Scholar] [CrossRef]
- Saising, J.; Dube, L.; Ziebandt, A.K.; Voravuthikunchai, S.P.; Nega, M.; Götz, F. Activity of gallidermin on Staphylococcus aureus and Staphylococcus epidermidis biofilms. Antimicrob. Agents Chemother. 2012, 56, 5804–5810. [Google Scholar] [CrossRef]
- Askari, P.; Namaei, M.H.; Ghazvini, K.; Hosseini, M. In vitro and in vivo toxicity and antibacterial efficacy of melittin against clinical extensively drug-resistant bacteria. BMC Pharmacol. Toxicol. 2021, 22. [Google Scholar] [CrossRef]
- Zhu, Y.; Johnson, T.J.; Myers, A.A.; Kanost, M.R. Identification by subtractive suppression hybridization of bacteria-induced genes expressed in Manduca sexta fat body. Insect Biochem. Mol. Biol. 2003, 33, 541–559. [Google Scholar] [CrossRef]
- Jayamani, E.; Rajamuthiah, R.; Larkins-Ford, J.; Fuchs, B.B.; Conery, A.L.; Vilcinskas, A.; Ausubel, F.M.; Mylonakisa, E. Insect-derived cecropins display activity against Acinetobacter baumannii in a whole-animal high-throughput Caenorhabditis elegans model. Antimicrob. Agents Chemother. 2015, 59, 1728–1737. [Google Scholar] [CrossRef]
- Qi, J.; Gao, R.; Liu, C.; Shan, B.; Gao, F.; He, J.; Yuan, M.; Xie, H.; Jin, S.; Ma, Y. Potential role of the antimicrobial peptide tachyplesin III against multidrug-resistant P. aeruginosa and A. baumannii coinfection in an animal model. Infect. Drug Resist. 2019, 12, 2865–2874. [Google Scholar] [CrossRef]
- Ovchinnikova, T.V.; Aleshina, G.M.; Balandin, S.V.; Krasnosdembskaya, A.D.; Markelov, M.L.; Frolova, E.I.; Leonova, Y.F.; Tagaev, A.A.; Krasnodembsky, E.G.; Kokryakov, V.N. Purification and primary structure of two isoforms of arenicin, a novel antimicrobial peptide from marine polychaeta Arenicola marina. FEBS Lett. 2004, 577, 209–214. [Google Scholar] [CrossRef]
- Kim, M.K.; Kang, N.; Ko, S.J.; Park, J.; Park, E.; Shin, D.W.; Kim, S.H.; Lee, S.A.; Lee, J.I.; Lee, S.H.; et al. Antibacterial and antibiofilm activity and mode of action of magainin 2 against drug-resistant acinetobacter baumannii. Int. J. Mol. Sci. 2018, 19, 3041. [Google Scholar] [CrossRef]
- Aleinein, R.A.; Hamoud, R.; Schäfer, H.; Wink, M. Molecular cloning and expression of ranalexin, a bioactive antimicrobial peptide from Rana catesbeiana in Escherichia coli and assessments of its biological activities. Appl. Microbiol. Biotechnol. 2013, 97, 3535–3543. [Google Scholar] [CrossRef]
- Subasinghage, A.P.; Conlon, J.M.; Hewage, C.M. Development of potent anti-infective agents from Silurana tropicalis: Conformational analysis of the amphipathic, alpha-helical antimicrobial peptide XT-7 and its non-haemolytic analogue [G4K]XT-7. Biochim. Biophys. Acta-Proteins Proteomics 2010, 1804, 1020–1028. [Google Scholar] [CrossRef]
- Conlon, J.M.; Al-Ghaferi, N.; Abraham, B.; Sonnevend, A.; Coquet, L.; Leprince, J.; Jouenne, T.; Vaudry, H.; Iwamuro, S. Antimicrobial peptides from the skin of the Tsushima brown frog Rana tsushimensis. Comp. Biochem. Physiol.-C Toxicol. Pharmacol. 2006, 143, 42–49. [Google Scholar] [CrossRef]
- Zhang, L.; Chen, X.; Zhang, Y.; Ma, C.; Xi, X.; Wang, L.; Zhou, M.; Burrows, J.F.; Chen, T. Identification of novel Amurin-2 variants from the skin secretion of Rana amurensis, and the design of cationicity-enhanced analogues. Biochem. Biophys. Res. Commun. 2018, 497, 943–949. [Google Scholar] [CrossRef]
- Mishra, B.; Wang, X.; Lushnikova, T.; Zhang, Y.; Golla, R.M.; Narayana, J.L.; Wang, C.; McGuire, T.R.; Wang, G. Antibacterial, antifungal, anticancer activities and structural bioinformatics analysis of six naturally occurring temporins. Peptides 2018, 106, 9–20. [Google Scholar] [CrossRef]
- Conlon, J.M.; Abraham, B.; Sonnevend, A.; Jouenne, T.; Cosette, P.; Leprince, J.; Vaudry, H.; Bevier, C.R. Purification and characterization of antimicrobial peptides from the skin secretions of the carpenter frog Rana virgatipes (Ranidae, Aquarana). Regul. Pept. 2005, 131, 38–45. [Google Scholar] [CrossRef]
- Abbassi, F.; Lequin, O.; Piesse, C.; Goasdoué, N.; Foulon, T.; Nicolas, P.; Ladram, A. Temporin-SHf, a new type of Phe-rich and hydrophobic ultrashort antimicrobial peptide. J. Biol. Chem. 2010, 285, 16880–16892. [Google Scholar] [CrossRef]
- Conlon, J.M.; Mechkarska, M.; Prajeep, M.; Sonnevend, A.; Coquet, L.; Leprince, J.; Jouenne, T.; Vaudry, H.; King, J.D. Host-defense peptides in skin secretions of the tetraploid frog Silurana epitropicalis with potent activity against methicillin-resistant Staphylococcus aureus (MRSA). Peptides 2012, 37, 113–119. [Google Scholar] [CrossRef]
- Yuan, Y.; Zai, Y.; Xi, X.; Ma, C.; Wang, L.; Zhou, M.; Shaw, C.; Chen, T. A novel membrane-disruptive antimicrobial peptide from frog skin secretion against cystic fibrosis isolates and evaluation of anti-MRSA effect using Galleria mellonella model. Biochim. Biophys. Acta-Gen. Subj. 2019, 1863, 849–856. [Google Scholar] [CrossRef]
- Portelinha, J.; Angeles-Boza, A.M. The Antimicrobial Peptide Gad-1 Clears Pseudomonas aeruginosa Biofilms under Cystic Fibrosis Conditions. ChemBioChem 2021, 22, 1646–1655. [Google Scholar] [CrossRef]
- Menousek, J.; Mishra, B.; Hanke, M.L.; Heim, C.E.; Kielian, T.; Wang, G. Database screening and in vivo efficacy of antimicrobial peptides against methicillin-resistant Staphylococcus aureus USA300. Int. J. Antimicrob. Agents 2012, 39, 402–406. [Google Scholar] [CrossRef]
- Cole, A.M.; Weis, P.; Diamond, G. Isolation and characterization of pleurocidin, an antimicrobial peptide in the skin secretions of winter flounder. J. Biol. Chem. 1997, 272, 12008–12013. [Google Scholar] [CrossRef]
- Zhao, Z.; Ma, Y.; Dai, C.; Zhao, R.; Li, S.R.; Wu, Y.; Cao, Z.; Li, W. Imcroporin, a new cationic antimicrobial peptide from the venom of the scorpion Isometrus maculates. Antimicrob. Agents Chemother. 2009, 53, 3472–3477. [Google Scholar] [CrossRef]
- De Melo, E.T.; Estrela, A.B.; Santos, E.C.G.; Machado, P.R.L.; Farias, K.J.S.; Torres, T.M.; Carvalho, E.; Lima, J.P.M.S.; Silva-Júnior, A.A.; Barbosa, E.G.; et al. Structural characterization of a novel peptide with antimicrobial activity from the venom gland of the scorpion Tityus stigmurus: Stigmurin. Peptides 2015, 68, 3–10. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, L.; Wu, Y.; Wang, L.; Ma, C.; Xi, X.; Bininda-Emonds, O.R.P.; Shaw, C.; Chen, T.; Zhou, M. Evaluation of the bioactivity of a mastoparan peptide from wasp venom and of its analogues designed through targeted engineering. Int. J. Biol. Sci. 2018, 14, 599–607. [Google Scholar] [CrossRef]
- Xiao, Y.; Cai, Y.; Bommineni, Y.R.; Fernando, S.C.; Prakash, O.; Gilliland, S.E.; Zhang, G. Identification and functional characterization of three chicken cathelicidins with potent antimicrobial activity. J. Biol. Chem. 2006, 281, 2858–2867. [Google Scholar] [CrossRef]
- van Dijk, A.; Molhoek, E.M.; Veldhuizen, E.J.A.; van Bokhoven, J.L.M.T.; Wagendorp, E.; Bikker, F.; Haagsman, H.P. Identification of chicken cathelicidin-2 core elements involved in antibacterial and immunomodulatory activities. Mol. Immunol. 2009, 46, 2465–2473. [Google Scholar] [CrossRef]
- Hee Lee, I.N.; Cho, Y.; Lehrer, R.I. Effects of pH and salinity on the antimicrobial properties of clavanins. Infect. Immun. 1997, 65, 2898–2903. [Google Scholar] [CrossRef]
- Skerlavaj, B.; Benincasa, M.; Risso, A.; Zanetti, M.; Gennaro, R. SMAP-29: A potent antibacterial and antifungal peptide from sheep leukocytes. FEBS Lett. 1999, 463, 58–62. [Google Scholar] [CrossRef]
- Skerlavaj, B.; Gennaro, R.; Bagella, L.; Merluzzi, L.; Risso, A.; Zanettit, M. Biological characterization of two novel cathelicidin-derived peptides and identification of structural requirements for their antimicrobial and cell lytic activities. J. Biol. Chem. 1996, 271, 28375–28381. [Google Scholar] [CrossRef]
- Bellamy, W.; Takase, M.; Wakabayashi, H.; Kawase, K.; Tomita, M. Antibacterial spectrum of lactoferricin B, a potent bactericidal peptide derived from the N-terminal region of bovine lactoferrin. J. Appl. Bacteriol. 1992, 73, 472–479. [Google Scholar] [CrossRef]
- Larrick, J.W.; Hirata, M.; Shimomoura, Y.; Yoshida, M.; Zheng, H.; Zhong, J.; Wright, S.C. Antimicrobial activity of rabbit CAP18-derived peptides. Antimicrob. Agents Chemother. 1993, 37, 2534–2539. [Google Scholar] [CrossRef]
- Patil, A.A.; Ouellette, A.J.; Lu, W.; Zhang, G. Rattusin, an intestinal α-defensin-related peptide in rats with a unique cysteine spacing pattern and salt-insensitive antibacterial activities. Antimicrob. Agents Chemother. 2013, 57, 1823–1831. [Google Scholar] [CrossRef]
- Ouellette, A.J.; Hsieh, M.M.; Nosek, M.T.; Cano-Gauci, D.F.; Huttner, K.M.; Buick, R.N.; Selsted, M.E. Mouse Paneth cell defensins: Primary structures and antibacterial activities of numerous cryptdin isoforms. Infect. Immun. 1994, 62, 5040–5047. [Google Scholar] [CrossRef]
- Blower, R.J.; Popov, S.G.; van Hoek, M.L. Cathelicidin peptide rescues G. mellonella infected with B. anthracis. Virulence 2018, 9, 287–293. [Google Scholar] [CrossRef]
- Tang, Y.Q.; Yuan, J.; Ösapay, G.; Ösapay, K.; Tran, D.; Miller, C.J.; Ouellette, A.J.; Selsted, M.E. A cyclic antimicrobial peptide produced in primate leukocytes by the ligation of two truncated α-defensins. Science (80-. ). 1999, 286, 498–502. [Google Scholar] [CrossRef]
- Ciornei, C.D.; Sigurdardóttir, T.; Schmidtchen, A.; Bodelsson, M. Antimicrobial and chemoattractant activity, lipopolysaccharide neutralization, cytotoxicity, and inhibition by serum of analogs of human cathelicidin LL-37. Antimicrob. Agents Chemother. 2005, 49, 2845–2850. [Google Scholar] [CrossRef]
- Giacometti, A.; Cirioni, O.; Greganti, G.; Quarta, M.; Scalise, G. In vitro activities of membrane-active peptides against gram-positive and gram-negative aerobic bacteria. Antimicrob. Agents Chemother. 1998, 42, 3320–3324. [Google Scholar] [CrossRef]
- Peschel, A.; Jack, R.W.; Otto, M.; Collins, L.V.; Staubitz, P.; Nicholson, G.; Kalbacher, H.; Nieuwenhuizen, W.F.; Jung, G.; Tarkowski, A.; et al. Staphylococcus aureus resistance to human defensins and evasion of neutrophil killing via the novel virulence factor MprF is based on modification of membrane lipids with L-lysine. J. Exp. Med. 2001, 193, 1067–1076. [Google Scholar] [CrossRef]
- Alaiwa, M.H.A.; Reznikov, L.R.; Gansemer, N.D.; Sheets, K.A.; Horswill, A.R.; Stoltz, D.A.; Zabner, J.; Welsh, M.J. pH modulates the activity and synergism of the airway surface liquid antimicrobials β-defensin-3 and LL-37. Proc. Natl. Acad. Sci. USA 2014, 111, 18703–18708. [Google Scholar] [CrossRef]
- Ridyard, K.E.; Elsawy, M.; Mattrasingh, D.; Klein, D.; Strehmel, J.; Beaulieu, C.; Wong, A.; Overhage, J. Synergy between Human Peptide LL-37 and Polymyxin B against Planktonic and Biofilm Cells of Escherichia coli and Pseudomonas aeruginosa. Antibiotics 2023, 12, 389. [Google Scholar] [CrossRef]
- Lin, L.; Nonejuie, P.; Munguia, J.; Hollands, A.; Olson, J.; Dam, Q.; Kumaraswamy, M.; Rivera, H.; Corriden, R.; Rohde, M.; et al. Azithromycin Synergizes with Cationic Antimicrobial Peptides to Exert Bactericidal and Therapeutic Activity Against Highly Multidrug-Resistant Gram-Negative Bacterial Pathogens. EBioMedicine 2015, 2, 690–698. [Google Scholar] [CrossRef]
- Desbois, A.P.; Gemmell, C.G.; Coote, P.J. In vivo efficacy of the antimicrobial peptide ranalexin in combination with the endopeptidase lysostaphin against wound and systemic meticillin-resistant Staphylococcus aureus (MRSA) infections. Int. J. Antimicrob. Agents 2010, 35, 559–565. [Google Scholar] [CrossRef]
- Lu, Y.; Tian, H.; Chen, R.; Liu, Q.; Jia, K.; Hu, D.L.; Chen, H.; Ye, C.; Peng, L.; Fang, R. Synergistic Antimicrobial Effect of Antimicrobial Peptides CATH-1, CATH-3, and PMAP-36 With Erythromycin Against Bacterial Pathogens. Front. Microbiol. 2022, 13, 953720. [Google Scholar] [CrossRef]
- Draper, L.A.; Cotter, P.D.; Hill, C.; Ross, R.P. The two peptide lantibiotic lacticin 3147 acts synergistically with polymyxin to inhibit Gram negative bacteria. BMC Microbiol. 2013, 13, 212. [Google Scholar] [CrossRef]
- Mathur, H.; O’Connor, P.M.; Hill, C.; Cotter, P.D.; Ross, R.P. Analysis of anti-clostridium difficile activity of thuricin CD, vancomycin, metronidazole, ramoplanin, and actagardine, both singly and in paired combinations. Antimicrob. Agents Chemother. 2013, 57, 2882–2886. [Google Scholar] [CrossRef]
- Cavera, V.L.; Volski, A.; Chikindas, M.L. The Natural Antimicrobial Subtilosin A Synergizes with Lauramide Arginine Ethyl Ester (LAE), ε-Poly-l-lysine (Polylysine), Clindamycin Phosphate and Metronidazole, Against the Vaginal Pathogen Gardnerella vaginalis. Probiotics Antimicrob. Proteins 2015, 7, 164–171. [Google Scholar] [CrossRef]
- Lobos, O.; Padilla, A.; Padilla, C. In vitro antimicrobial effect of bacteriocin PsVP-10 in combination with chlorhexidine and triclosan against Streptococcus mutans and Streptococcus sobrinus strains. Arch. Oral Biol. 2009, 54, 230–234. [Google Scholar] [CrossRef]
- Herrmann, G.; Yang, L.; Wu, H.; Song, Z.; Wang, H.; Høiby, N.; Ulrich, M.; Molin, S.; Riethmüller, J.; Döring, G. Colistin-tobramycin combinations are superior to monotherapy concerning the killing of biofilm Pseudomonas aeruginosa. J. Infect. Dis. 2010, 202, 1585–1592. [Google Scholar] [CrossRef]
- Rishi, P.; Preet, S.; Bharrhan, S.; Verma, I. In vitro and in vivo synergistic effects of cryptdin 2 and ampicillin against Salmonella. Antimicrob. Agents Chemother. 2011, 55, 4176–4182. [Google Scholar] [CrossRef]
- Jahangiri, A.; Neshani, A.; Mirhosseini, S.A.; Ghazvini, K.; Zare, H.; Sedighian, H. Synergistic effect of two antimicrobial peptides, Nisin and P10 with conventional antibiotics against extensively drug-resistant Acinetobacter baumannii and colistin-resistant Pseudomonas aeruginosa isolates. Microb. Pathog. 2021, 150, 104700. [Google Scholar] [CrossRef]
- Nuding, S.; Frasch, T.; Schaller, M.; Stange, E.F.; Zabel, L.T. Synergistic effects of antimicrobial peptides and antibiotics against clostridium difficile. Antimicrob. Agents Chemother. 2014, 58, 5719–5725. [Google Scholar] [CrossRef]
Figure 1.
Antimicrobial mechanisms of action and antibiotic resistance mechanisms.
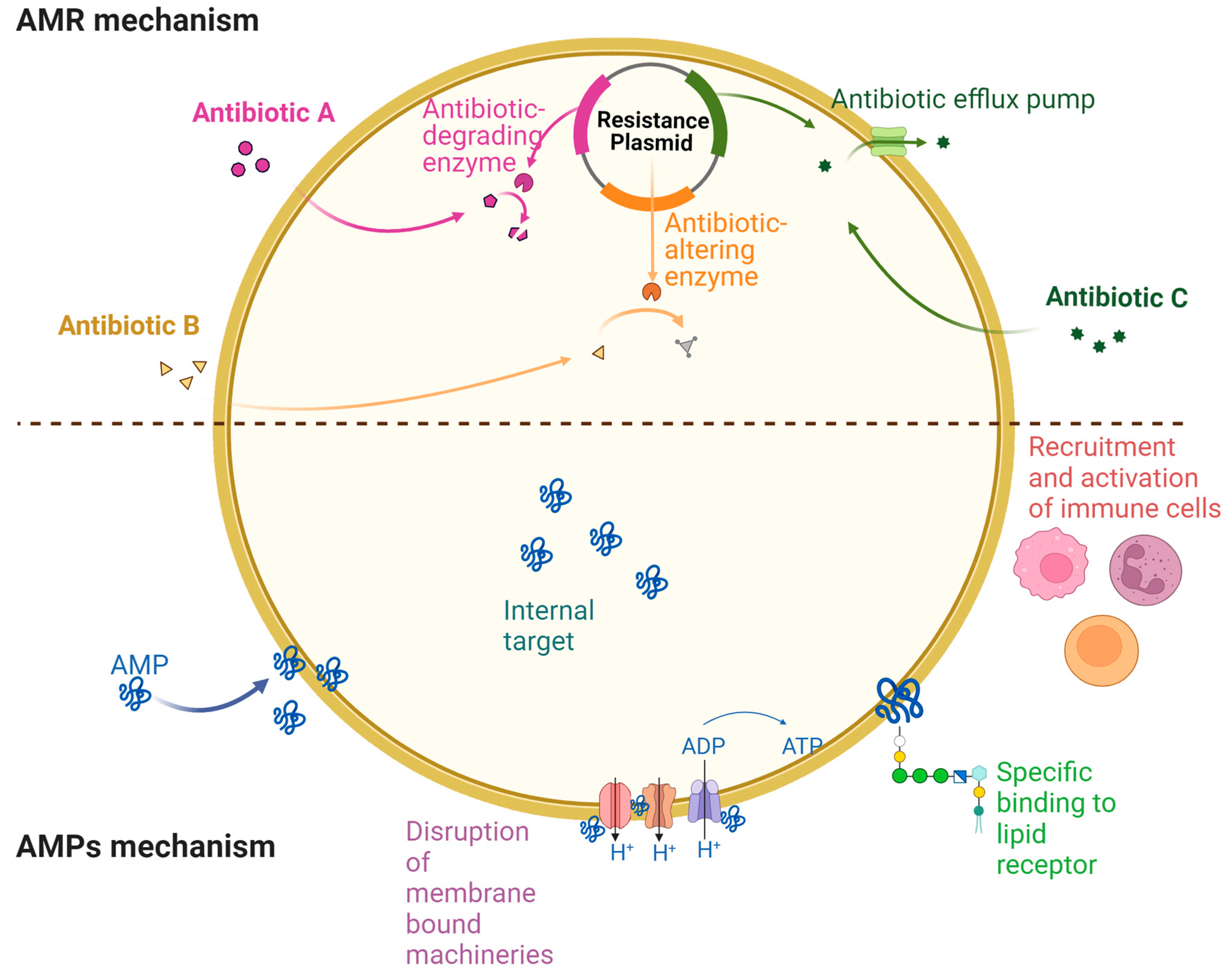

Table 1.
FDA-approved selected AMPs for the treatment of different bacterial infections.
| AMPs | Source/Type | Clinical Use | Mode of Action | Reference |
|---|---|---|---|---|
| Vancomycin | Streptococcus orientalis)/Tricyclic glycopeptide | Complicated infections caused by MRSA, C. difficile, and Gram-positive bacteria | Inhibition of cell wall biosynthesis | [88] |
| Bacitracin | B. licheniformis) | Pneumonia and empyema in infants | Cell wall interference and peptidoglycan synthesis interference | [72] |
| Daptomycin | Streptomyces roseosporus) | Skin bacterial infections | Membrane-lytic peptide, inhibition of DNA, RNA, and protein synthesis | [89] |
| Polymyxin B | Bacteria (Paenibacillus polymyxa) | Bacterial infections caused by Gram-negative bacteria | Targets membrane phospholipids (lipopolysaccharides and lipoproteins) | [90] |
| Gramicidin | Bacteria (B. brevis) | Dermatological and ophthalmological infections | Pore-forming peptide | [71] |
| Colistin | Bacteria (B. polymyxa) | Gram-negative bacterial infections, Pneumonia | Membrane-lytic peptide | [73] |
Table 2.
AMPs under clinical trials to combat biofilms and drug-resistant superbug infections.
| AMPs | Type | Target | Clinical Trial ID | Phase | References |
|---|---|---|---|---|---|
| Nisin | Lantibiotic | Gram-positive bacteria | NCT02928042 NCT02467972 | [91] | |
| Murepavadin (POL7080) | Derivative of protegrin | P. aeruginosa, K. pneumoniae | EUCTR2017-003933-27-EE | II | [92] |
| Iseganan (IB-367) | Derivative of protegrin | Pneumonia/oral mucositis | NCT00118781 NCT00022373 |
III | [93] |
| Surotomycin (CB-315) | Cyclic lipopeptide | C. difficile | NCT01597505 | III | [94] |
| Omiganan (MBI-226) | Derivative of indolicidin | Antisepsis/ catheter infection | NCT00231153 NCT00608959 | III | [95] |
| NVB-302 | Lantibiotic | C. difficile | ISRCTN40071144 | I | [96] |
| OP-145 | Derivative of LL-37 | Chronic middle ear infection | ISRCTN84220089 | II | [97] |
| LTX-109 | Synthetic tripeptide | MRSA, Impetigo | NCT01803035 NCT01158235 | II | [98] |
| EA-230 | Oligopeptide | Sepsis | NCT03145220 | II | [99] |
| hLF1-11 | Human lactoferrin derivative | Bacterial infections | NCT00430469 | II | [100] |
| C16G2 | Synthetic peptide | S. mutans | NCT03004365 | II | [101] |
| Ramoplanin (NTI851) | Glycolipodepsipeptide | C. difficile, VRE | NA | III | [102] |
| p2TA (AB103) | Synthetic peptide | Necrotic tissue infection | NA | III | [103] |
| D2A21 | Synthetic peptide | Burn wound infections | NA | III | [104] |
| LFF571 | Semisynthetic thiopeptide | C. difficile | NCT01232595 | II | [105] |
| GSK132232 (Lanopepde) | Synthetic hydrazide | Bacterial skin infection | NCT01209078 | II | [106] |
| PMX-30063 (Brilacidin) | Defensin mimetic | Acute bacterial skin infection | NCT01211470 NCT02052388 | II | [107] |
| XF-73 (Exeporfinim chloride) | Derivative of porphyrin | Staphylococal infection | NCT03915470 | II | [108] |
| Wap-8294A2 (Lotilibcin) | Naturally produced by Lysobacter species | Gram-positive bacteria | NA | II | [109] |
| PL-5 | Synthetic peptide | Skin infections | NA | I | [110] |
| IDR-1 | Bactenecin | Infection prevention | NA | I | [111] |
| Pexiganan | Analog of magainin isolated from the skin of the African clawed frog | Infections of diabetic foot ulcers, Gram positive and Gram negative bacteria | NCT01590758 | III |
Table 3.
Selected AMPs showing activity against drug-resistant superbug and biofilms.
| AMPs | Peptide Sequence | Source | Spectrum of Activity | References |
|---|---|---|---|---|
| Mersacidin | CTFTLPGGGGVCTLTSECIC | Bacillus sp. HIL Y-85 54728 | MRSA | [112] |
| Epilancin 15X | SASIVKTTIKASKKLCRGFTLTCGCHFTGKK | S. epidermidis 15X154 | MRSA, Gram-positive | [113] |
| Mutacin B-Ny266 | FKSWSFCTPGCAKTGSFNSYCC | S. mutans Ny266 | MRSA | [114] |
| Epidermicin NI01 | MAAFMKLIQFLATKGQKYVSLAWKHKGTILKWINAGQSFEWIYKQIKKLWA | S. epidermidis 224 | MRSA, Gram-positive | [115] |
| Actagardine A | SSGWVCTLTIECGTVICAC | A. garbadinensis ATCC 31049 | C. difficile, VRE, MRSA | [120] |
| Aureocin A53 | MSWLNFLKYIAKYGKKAVSAAWKYKGKVLEWLNVGPTLEWVWQKLKKIAGL | Staphylococcus aureus A53 | MRSA, Gram positive | [121] |
| LCI | AIKLVQSPNGNFAASFVLDGTKWIFKSKYYDSSKGYWVGIYEVWDRK | Bacillus subtilis strain A014 | MRSA, Gram positive and Gram-negative | |
| Microcin E492 | GETDPNTQLLNDLGNNMAWGAALGAPGGLGSAALGAAGGALQTVGQGLIDHGPVNVFIPVLIGPSWNGSGSGYNSATSSSG | K. pneumoniae RYC492 | K. pneumonia, and other Gram-negative bacteria | [122] |
| Lassomycin | GLRRLFADQLVGRRNI | Lentzea kentuckyensis | M. tuberculosis | [123] |
| Enterocin AS-48 | MAKEFGIPAAVAGTVINVVEAGGWVTTIVSILTAVGSGGLSLLAAAGRESIKAYLKKEIKKKGKRAVIAW | E. faecalis S-48 | MRSA, M. tuberculosis | [124] |
| Ruminococcin C | WGCVCSGSTAVANSHNAGPAYCVGYCGNNGVVTRNANANVAKTA | R. gnavus E1 | C. difficile and other MDR strains | [126] |
| Actinomycesin | GFGCPWNAYECDRHCVSKGYTGGNCRGKIRQTCHCY | Actinomyces sp. | MRSA, Gram-positive | [128] |
| Triintsin | GFGCPLNERECHSHCQSIGRKFGYCGGTLRLTCICGKE | Trichophyton interdigitale | MRSA, Gram-positive, and Gram-negative | [129] |
| Blauticin | ITSKSLCTPGCVTGILMTCPVQTATCGCQITGK | Blautia producta SCSK | MRSA, Gram-positive | [130] |
| Lan-Df | YKSKSVCTPGCPTGILMTCPLKTATCGCHITGK | Dorea formicigenerans | MRSA, Gram-positive | [130] |
| Penisin | NIGLFTSTCFSSQCFSSKCFTDTCFSSNCFTGRHQCGYTHGSC | Paenibacillus sp. A3 | MRSA, Gram-positive, and Gram-negative | [131] |
| Laterosporulin10 | ACVNQCPDAIDRFIVKDKGCHGVEKKYYKQVYVACMNGQHLYCRTEWGGPCQL | B. laterosporus SKDU10 | M. tuberculosis H37Rv, Gram-positive | [79] |
| CycP | CAWLWAPAWLWAC | Synthetic | Drug-resistant S. aureus | [132] |
| Plectasin | GFGCNGPWDEDDMQCHNHCKSIKGYKGGYCAKGGFVCKCY | Pseudoplectania nigrella | MRSA, Gram-positive | [133] |
| Micasin-1 | GFGCPFNENECHAHCLSIGRKFGFCAGPLRATCTCGKQ | Microsporum canis | MRSA, Gram-positive, and Gram-negative | [134] |
| Lacticin 3147 | CSTNTFSLSDYWGNNGAWCTLTHECMAWCK | Lactococcus lactis DPC3147 | S. mutans (Biofilm) | [135] |
| Subtilomycin | TWATIGKTIVQSVKKCRTFTCGCSLGSCSNCN | Bacillus subtilis MMA7 | L. monocytogenes (Biofilm) | [136] |
| Gallidermin | IASKFLCTPGCAKTGSFNSYCC | Staphylococcus gallinarum (F16/P57) | S. aureus, S. epidermidis (Biofilm) | [137] |
| Melittin | GIGAVLKVLTTGLPALISWIKRKRQQ | Apis mellifera | A. baumannii, MRSA, P. aeruginosa, K. pneumonia, and Gram-positive bacteria | [138] |
| MS moricin | GKIPVKAIKQAGKVIGKGLRAINIAGTTHDVVSFFRPKKKKH | Manduca sexta | MRSA, Gram positive and Gram-negative | [139] |
| Cecropin A | RWKVFKKIEKVGRNIRDGVIKAAPAIEVLGQAKAL | Heliothis virescens | A. baumannii, P. aeruginosa, and other Gram-negative, and Gram-positive bacteria | [140] |
| Tachyplesin III | KWCFRVCYRGICYRKCR | Tachypleus gigas | P. aeruginosa, A. baumannii, biofilms, and Gram-positive bacteria | [141] |
| Arenicin-1 | RWCVYAYVRVRGVLVRYRRCW | Arenicola marina | MRSA, Gram-positive, and Gram-negative | [142] |
| Magainin 2 | GIGKFLHSAKKFGKAFVGEIMNS | Xenopus laevis | K. pneumoniae, P. aeruginosa, A. baumannii, Anti-biofilm | [143] |
| Ranalexin | FLGGLIKIVPAMICAVTKKC | Rana catesbeiana (Bullfrog) | MRSA, Gram-positive, and Gram-negative | [144] |
| CPF-ST3 | GLLGPLLKIAAKVGSNLL | Frog | MRSA, Gram-positive, and Gram-negative | [145] |
| Brevinin-1TSa | FLGSIVGALASALPSLISKIRN | Frog | MRSA, Gram-positive, and Gram-negative | [146] |
| Brevinin-1DYa | FLSLALAALPKFLCLVFKKC | Frog | MRSA, Gram-positive, and Gram-negative | [147] |
| Brevinin-1DYb | FLSLALAALPKLFCLIFKKC | Frog | MRSA, Gram-positive, and Gram-negative | [147] |
| Temporin-1Oc | FLPLLASLFSRLF | Frog | MRSA, Gram-positive, | [148] |
| Temporin-1Ga | SILPTIVSFLSKVF | Frog | MRSA, Gram-positive, | [148] |
| Temporin-1Vb | FLSIIAKVLGSLF | Frog | MRSA, Gram-positive | [149] |
| Temporin-SHd | FLPAALAGIGGILGKLF | Frog | MRSA, Gram-positive, and Gram-negative | [150] |
| CPF-SE1 | GFLGPLLKLGLKGVAKVIPHLIPSRQQ | Frog | MRSA, Gram-positive, and Gram-negative | [151] |
| CPF-SE2 | GFLGPLLKLGLKGAAKLLPQLLPSRQQ | Frog | MRSA, Gram-positive, and Gram-negative | [151] |
| Japonicin-2LF | FIVPSIFLLKKAFCIALKKC | Frog | MRSA, Gram-positive, and Gram-negative | [152] |
| Gaduscidin-1 | FIHHIIGWISHGVRAIHRAIH | Gadus morhua (Atlantic cod) | P. aeruginosa biofilms, Gram positive and Gram-negative | [153] |
| Piscidin 1 | FFHHIFRGIVHVGKTIHRLVTG | Fish | MRSA, Gram-positive, and Gram-negative | [154] |
| Pleurocidin | GWGSFFKKAAHVGKHVGKAALTHYL | Pleuronectes americanus | MRSA, Gram-positive, and Gram-negative | [155] |
| Imcroporin | FFSLLPSLIGGLVSAIK | Scorpions | MRSA, Gram-positive, | [156] |
| Stigmurin | FFSLIPSLVGGLISAFK | Scorpions | MRSA, Gram-positive, | [157] |
| MP-C | LNLKALLAVAKKIL | Insects | MRSA, Gram-positive, and Gram-negative | [158] |
| Chicken CATH-1 | RVKRVWPLVIRTVIAGYNLYRAIKKK | Gallus galllus | MRSA, Gram-positive, and Gram-negative | [159] |
| Chicken CATH-2 | RFGRFLRKIRRFRPKVTITIQGSARFG | Chicken | MRSA, Gram-positive, and Gram-negative | [160] |
| Clavanin A | VFQFLGKIIHHVGNFVHGFSHVF | Styela clava | MRSA, Gram-positive, | [161] |
| SMAP-29 | RGLRRLGRKIAHGVKKYGPTVLRIIRIAG | Sheep | MRSA, Gram-positive, and Gram-negative | [162] |
| BMAP-27 | GRFKRFRKKFKKLFKKLSPVIPLLHLG | Cattles | MRSA, Gram-positive, and Gram-negative | [163] |
| BMAP-28 | GGLRSLGRKILRAWKKYGPIIVPIIRIG | Cattles | MRSA, Gram-positive, and Gram-negative | [163] |
| Lactoferricin B | FKCRRWQWRMKKLGAPSITCVRRAF | Bos taurus | MRSA, Gram-positive, and Gram-negative | [164] |
| CAP18 | GLRKRLRKFRNKIKEKLKKIGQKIQGFVPKLAPRTDY | Oryctolagus cuniculus | MRSA, Gram-positive, and Gram-negative | [165] |
| Rattusin | LRVRRTLQCSCRRVCRNTCSCIRLSRSTYAS | Rattus norvegicus | MRSA, Gram-positive, and Gram-negative | [166] |
| Cryptdin-4 | LRGLLCYCRKGHCKRGERVRGTCGIRFLYCCPRR | Mus musculus | MRSA, Gram-positive, and Gram-negative | [167] |
| Protegrin 1 | RGGRLCYCRRRFCVCVGR | Sus scrofa | MRSA, A. baumannii, P. aeruginosa, Anti-biofilm, Gram positive | [168] |
| RTD-1 | GFCRCLCRRGVCRCICTR | Rhesus Macaque (Macaca mulatta) | MRSA, Gram-positive, and Gram-negative | [169] |
| LL-37 | LLGDFFRKSKEKIGKEFKRIVQRIKDFLRNLVPRTES | Human and other mammals | MRSA, P. aeruginosa, A. baumannii, Gram-positive, and Gram-negative | [170] |
| Indolicidin | ILPWKWPWWPWRR | Human and other mammals | MRSA, P. aeruginosa, Gram-positive, and Gram-negative | [171] |
| HNP-1 | ACYCRIPACIAGERRYGTCIYQGRLWAFCC | Homo sapiens | MRSA, C. difficile, Gram-positive, and Gram-negative | [172] |
| hBD-3 | GIINTLQKYYCRVRGGRCAVLSCLPKEEQIGKCSTRGRKCCRRKK | Homo sapiens | MRSA, Anti-biofilm, Gram-positive, and Gram-negative | [173] |
Table 4.
Selected AMPs showing synergistic antimicrobial action with conventional antibiotics.
| AMPs | Antibiotics | Target pathogen | References |
|---|---|---|---|
| Nisin | Ramoplanin, Polymyxin E, Clarithromycin, Amoxicillin, Penicillin, Streptomycin, Ceftiofur, Tetracycline | MRSA, P. aeruginosa, S. suis | [81,82,83] |
| Nisin Z | Ampicillin, Chloramphenicol, Kanamycin, Lincomycin, Penicillin G, Rifampicin, Streptomycin, Tetracycline, Vancomycin | Multi-drug resistant P. fluorescens LRC-R73 | [80] |
| LL-37 | Polymyxin B, Azithromycin | Multi-drug resistant E. coli, P. Aeruginosa, A. baumannii, K. pneumoniae |
[174,175] |
| Ranalexin | Endopeptidase lysostaphin | MRSA | [176] |
| CATH-1, CATH-3, PMAP-36 | Erythromycin | Pathogenic strain of S. aureus, S. enteritidis, and E. coli | [177] |
| Lacticin 3147 | Polymyxin B | S. aureus 5247 | [178] |
| Actagardine | Ramoplanin, Metronidazole, Vancomycin | C. difficile | [179] |
| Thuricin CD | Ramoplanin, Vancomycin | C. difficile | [179] |
| Subtilosin A | Clindamycin phosphate, Metronidazole, Lauramide alginate, Ester poly-lysine | Vaginal pathogen G. vaginalis | [180] |
| PsVP-10 | Chlorhexidine | S. mutans, S. sobrinus | [181] |
| Colistin | Tobramycin, Azithromycin | P. aeruginosa, A. baumannii, K. pneumoniae | [175,182] |
| Cryptdin 2 | Ampicillin | S. typhimurium | [183] |
| Arenicin-1 | Erythromycin, Ampicillin, chloramphenicol | S. dermis, S. aureus, E. coli, and P. aeruginosa | [86] |
| Laterosporulin10 | Rifampicin | M. tuberculosis H37Rv | [79] |
| Gaduscidin-1 | Kanamycin, Ciprofloxacin | P. aeruginosa biofilms | [153] |
| Lactoferricin | Ciprofloxacin, Ceftazidim | P. aeruginosa | [184] |
| Human defensin 5 (HD5) | Meropenem | C. difficile | [185] |
| Human neutrophil peptide-1 (HNP1) | Rifampicin | M. tuberculosis H37Rv | [78] |
| Human β-defensin 3 (HBD3) |
Meropenem, Moxifloxacin, Piperacillin-Tazobactam, Tigecycline | C. difficile | [185] |
Table 5.
AMPs in comparison with conventional antibiotics, concerning pharmacological development for therapeutic applications.
Table 5.
AMPs in comparison with conventional antibiotics, concerning pharmacological development for therapeutic applications.
| Conventional antibiotics | AMPs |
|---|---|
| Not amenable to bioengineering | Highly amenable to bioengineering |
| Non-ribosomal synthesis | Ribosomal synthesis |
| Toxic | A good safety and tolerability profile |
| No immunomodulatory properties | Excellent immunomodulatory properties |
| High serum/ plasma stability (depends on the drug) | Low serum/ plasma stability |
| Usually high (depends on the drug) | Low half-life |
| Not degradable/persistent | Completely metabolized |
| Highly stable | Rapid clearance |
| Less diverse | Highly diverse (endless opportunities, especially in the case of bacterial AMPs) |
| Resistant to biofilms | Strong biofilm activities |
| Easy purification/ high yield | Complex purification/ low yield |
| Narrow spectrum/ Specific target | Broad spectrum/ multiple mechanisms of action |
| Highly prone to resistant development | Less prone to resistant development |
| High solubility | Low solubility |
| Good bioavailability | Depends on size |
| Not affected by digestive enzymes | Prone to digestive enzymes |
| High absorption | Poor absorption |
| Many side effects | Not identified |
| Narrow range for pH and temperature stability | Highly stable at a broad range of pH and temperature |
| Cost-effective | High cost of production |
| Many administration routs | Limited administration routs due to protein degradation issues |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



