Submitted:
02 October 2024
Posted:
03 October 2024
You are already at the latest version
Abstract
To quantify the ecological impacts of organic sediments and environmental dredging, benthic mollusks were chosen to serve as bioindicators of environmental change, measured as sediment organic content and associated parameters. Data on species richness, biodiversity, and abundances were collected alongside sediment and near-bottom water quality data before, during, and after environmental dredging. Organic sediment content was found to have an inverse logarithmic relationship with benthic mollusk biodiversity, species richness, and abundance. Sediments with lower organic content had higher biodiversity (up to 1.337), while sediments with higher organic content had low biodiversity (0-0.6667). Post-hoc analyses found that percent dissolved oxygen, which correlates with sediment organic content, was responsible for 29.31-34.12% of the benthic mollusk community variation.
Keywords:
mollusk
; muck
; eutrophication
; abundance
; sediment
; pollution
; biodiversity
1. Introduction
Nutrient loading is a major anthropogenic stressor afflicting global estuaries. “Eutrophication is regarded as one of the greatest threats to coastal ecosystem health” [1]. While some level of nutrients are in estuaries naturally due to geological weathering and upwelling, extreme nutrient loading is likely due to a combination of anthropogenic influences. Issues related to eutrophication are an increase in harmful algal blooms (HABs) and turbidity. These issues can lead to hypoxic conditions, fish kills, decline in submerged aquatic vegetation, a shift to pelagic-dominated productivity, and loss of biodiversity. Each of these ecosystem effects can have significant economic impacts (e.g., tourism, fisheries, recreation, and real estate) [1]. An estuary facing these issues is the Indian River Lagoon (IRL, Florida), a shallow polluted subtropical estuary with a low flushing rate [2]. Bloom events and anthropogenic influences in this estuary have led to the accumulation of fine-grained, organic-rich sediments on the bottom of the estuary, colloquially known as muck.
Muck is a primary repository for toxic metals, organic substances, and anthropogenic nutrients [7]. IRL muck is comprised of 10-30% organic matter, 60% silt and clay, and a high water content (about 75% by weight) [7]. It covers around 10% of the IRL, ranging from a few centimeters to several meters in thickness [8]. Muck aggravates the IRL’s habitat degradation by contributing additional nutrients for HABs, blocking sunlight, depleting oxygen, and smothering benthic organisms [7,9,10]. Turbidity, HABs, and hypoxia can lead to loss of seagrass meadows and other important benthic habitats [11]. Habitat loss decreases biodiversity, species richness, and benthic population abundances. Hypoxic conditions alter community composition and may selectively eliminate sensitive species, thereby promoting the growth and recruitment of those with a higher tolerance for less ideal conditions [12].
Restoration projects for polluted and impaired estuaries may include flow redirection, point-source pollution reduction, or dredging to remove legacy loads of pollutants [6]. Environmental dredging is a remedial process attempting to remove contaminated sediments, improve water quality, and restore benthic habitats [13]. In the complex Indian River Lagoon System, environmental dredging for restoration has been carried out in several locations including Crane Creek, the Eau Gallie River, Sykes Creek, and the Mims Boat Ramp canal.
Mollusks are diverse in estuaries and coastal habitats, second only to arthropods in metazoan taxonomic diversity [14,15,16], and arguably more reliably identified to lower taxonomic levels than other benthic groups [12]. Many benthic mollusks are relatively sessile, and even errant species tend to stay within a localized region. They have longer life cycles and are sensitive to environmental changes [16]. Due to these characteristics, they may serve as effective bioindicators of environmental change, sediment health, and water quality [17,18,19,20,21]. In IRL benthic habitats, bivalves and gastropods are especially abundant [14]. Some common species of gastropods include Acteocina canaliculata, Astyris lunata, Haminoea succinea, and Phrontis vibex [22]. Some common species of bivalves include Amygdalum papyrium, Mulinia lateralis, Parastarte triquetra, and Mercenaria mercenaria [22].
Many mollusks have lower metabolic and oxygen consumption rates relative to other benthic invertebrates [23,24,25,26], helping them persist in polluted organic sediments. Bayne and Newell [23] compared mollusk oxygen consumption rates based on trophic roles with those of other taxa and found them to be lower. This included grazers like the periwinkle snail Littorina littorea (oxygen consumption rate 2.052 ml h-1), the suspension-feeding oyster Crassostrea virginica (oxygen consumption rate 0.496 ml h-1), and the predatory snail Nassarius reticulatus (oxygen consumption rate 1.63 ml h-1). In contrast, Fox & Simmonds [24] found arthropod consumption rates were much higher, including the amphipod Gammarus marinus (oxygen consumption rate 191-367 ml g-1 hr-1), the amphipod Gammarus locusta (oxygen consumption rate 188-207 ml g-1 hr-1), and the isopod Idotea neglecta (oxygen consumption rate 125-321 ml g-1 hr-1). Salvato et al. [26] compared the effects of several environmental conditions on oxygen consumption by the decapod crustaceans Palaemon serratus (mean oxygen consumption rate 0.3519 ml O2 g-1 wet tissue hr-1) and Panaeus monodon (mean oxygen consumption rate 0.5287 ml O2 g-1 wet tissue hr-1), and the prosobranch gastropods Trunculariopsis trunculus (mean oxygen consumption rate 0.0394ml O2 g-1 wet tissue hr-1) and Nassarius mutabilis (mean oxygen consumption rate 0.0687 ml O2 g-1 wet tissue hr-1). Pamatmat [25] also compared the aerobic and anaerobic metabolic rates of bivalves to benthic polychaetes and found that bivalves were lower. Lower mollusk oxygen consumption rates help them survive in more polluted conditions relative to other benthic taxa. Some mollusks are also able to withstand physiological stress caused by toxic H2S, often associated with muck sediments [17,27]. Theede et al. [28] found that Mytilus edulis can survive 25 days in water containing H2S, and 35 days in water with 0.15ml O2 l-1. H2S tolerance is important for benthic organisms in order to survive in hypoxic and anoxic environments [27,28].
Despite being seemingly well-suited to polluted estuarine conditions [12,17,21], fine gills and other structures may be clogged by silt and clay particles, potentially suffocating benthic mollusks [29,30]. Pearson [30] found that benthic communities exposed to anthropogenically polluted effluents typically exhibit low diversity, are poorly organized, and are composed of small r-selected species. Both Pearson and Rhoads & Young [30,31] suggest that, in some systems, this could be due to trophic amensalism, where deposit feeders exclude filter-feeding neighbors by disturbing and resuspending sediments, clogging the filter-feeding mechanisms of less tolerant competitors. Since muck is easily resuspended [29], smothering or suffocation is a significant threat to benthic mollusks in degraded habitats. Some mollusks in the IRL persist in polluted conditions, yet may remain stressed in organic sediments [29]. Moraitis et al. [21] stated that effective bioindicators, like the presence of benthic estuarine mollusks, are easily monitored and display the effects of environmental change.
This study addresses the following hypotheses:
(1) There is an inverse relationship in the biodiversity, species richness, and abundances of mollusks with sediment percent organic content.
(2) Removal of sediments with high concentrations of fine-grained organic matter by dredging will increase the biodiversity, species richness, and abundances of benthic mollusks.
2. Materials and Methods
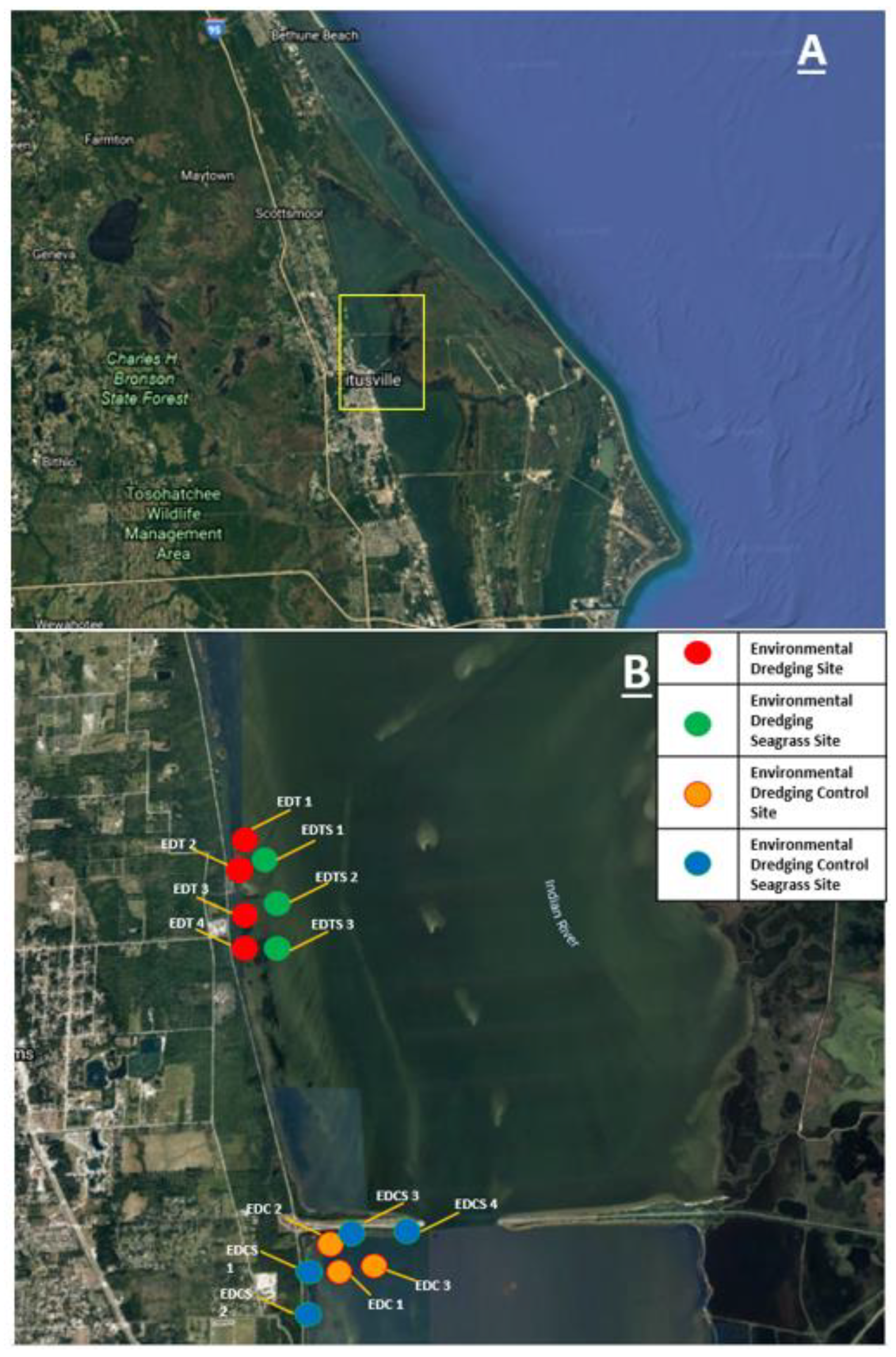
To determine the impacts of organic sediments and dredging restoration on benthic mollusk communities, benthic grabs, sediment samples, and water quality data were collected before, during, and after dredging at muck sites-to-be-dredged (“Environmental Dredging Treatment” or EDT), as well as undredged control muck sites (“Environmental Dredging Control” or EDC). For comparison, seagrass sites adjacent to muck dredging and muck control sites were also sampled. Seagrass sites adjacent to EDT (dredged) sites are identified as “Environmental Dredging Treatment – Seagrass” (EDTS). Seagrass sites adjacent to EDC (undredged) muck sites are identified as “Environmental Dredging Control – Seagrass” (EDCS) (Figure 1). Data were collected quarterly from March 2017 until October 2020 from 14 stations in Mims, Fl (Figure 1). All 14 stations are located within the Indian River Lagoon (IRL) proper (Figure 1 and Appendix Table A2). All stations were sampled every quarter (season) from 2017-2020 with the exception of the control stations in winter of 2017 and summer 2019, and all stations in winter of 2020.
Figure 1.
A) The region in the northern Indian River Lagoon (IRL) in which this study was conducted, including treatment and control sites. B) Stations were environmentally dredged muck stations (EDT, red), seagrass stations near dredging (EDTS, green), environmental dredging control (undredged) muck stations (EDC, orange), and seagrasses near undredged (control) muck stations (EDCS, blue).
Figure 1.
A) The region in the northern Indian River Lagoon (IRL) in which this study was conducted, including treatment and control sites. B) Stations were environmentally dredged muck stations (EDT, red), seagrass stations near dredging (EDTS, green), environmental dredging control (undredged) muck stations (EDC, orange), and seagrasses near undredged (control) muck stations (EDCS, blue).
Water quality was recorded at each sampling station and included temperature, salinity, percent saturation of dissolved oxygen (DO), mg L-1 DO, and turbidity. The Secchi depth (0.5-1 m deep) and total water column depth (0.5-5 m deep) were also determined for each sampling station. A Yellow Springs Instrument (YSI) Multimeter was calibrated prior to sampling against factory standards and used to measure water quality at the Secchi depth and near the bottom.
On a given date, four sediment grabs were collected at each station (Figure 1B) with a Petite Ponar grab (benthic sampling area of 225 cm2), with one grab for sediment analysis and three grabs for the faunal survey. Total sediment volumes were recorded for calculations of organism densities and grab penetration depth. Benthic fauna grabs were sieved through a 0.5mm sieve and placed in labeled plastic bags. One unsieved grab from each station was bagged for sediment characterization (% water content, % organic content, and % silt/clay content). Samples were then transported to the laboratory in a cooler. Fauna samples were frozen pending microscope sorting, and sediment samples were refrigerated while they awaited processing.
Fauna samples were thawed and sorted via light stereomicroscopy (8 – 35x magnification). Higher volume samples were split 1-4 times to allow for processing fauna in a timely manner. Fauna were identified to the lowest possible taxonomic level. Only whole organisms, apparently alive at the time of collection, were counted. After processing, each sample was preserved in a 4% formaldehyde solution for long-term storage. Counts were extrapolated to whole grabs and field densities.
In order to determine the silt/clay content of the sediments with visibly higher muck content (Folk 1974), ten grams of sample were used. For sandier samples, more sediment was required for accuracy, and 30 grams of sediment were used to determine silt/clay content. Samples were then sieved through 63-micron mesh to separate silt/clay from larger sediment particles. Both smaller sample components were dried by baking at 105 degrees Celsius for 24 hours, then re-weighed, with the relative proportions yielding the % silt/clay content [32] In a similar process, the % water content of the sediments was determined by weighing un-sieved sediments prior to and after baking them at 105 degrees Celsius for 24 hours [33] Sediment organic content was determined with the mass loss-on-ignition (LOI) method, where sediment is dried, ground, and weighed before and after baking at 550° C for 4 hours [34,35].
Biodiversity was calculated using the Shannon-Weiner index. Abundances, species richness, and biodiversity are being used to evaluate mollusk community responses to dredging. Generalized linear models were utilized to compare how dredged conditions, seasons, time since dredging, sediment organic content, silt/clay content, and water content affect mollusk species richness, biodiversity, and abundance. 2-Way ANOVA tests were used to examine the influence of the categorical factors, time and season, on the biodiversity, species richness, and abundance of benthic mollusks. ANOSIM tests were conducted to assess the impact of percent sediment organic, silt/clay, percent water content, percent dissolved oxygen, salinity, water temperature, and, where applicable, seagrass percent cover on the biodiversity, species richness, and abundance of benthic mollusks. The ANOSIMs were paired with non-metric multi-dimensional scaling (nMDS) to represent community similarities and dissimilarities in multivariate space. All statistical tests, nMDS plots, and figures were carried out and created in R and Microsoft Excel.
3. Results
3.1. Presence and Absence of Species
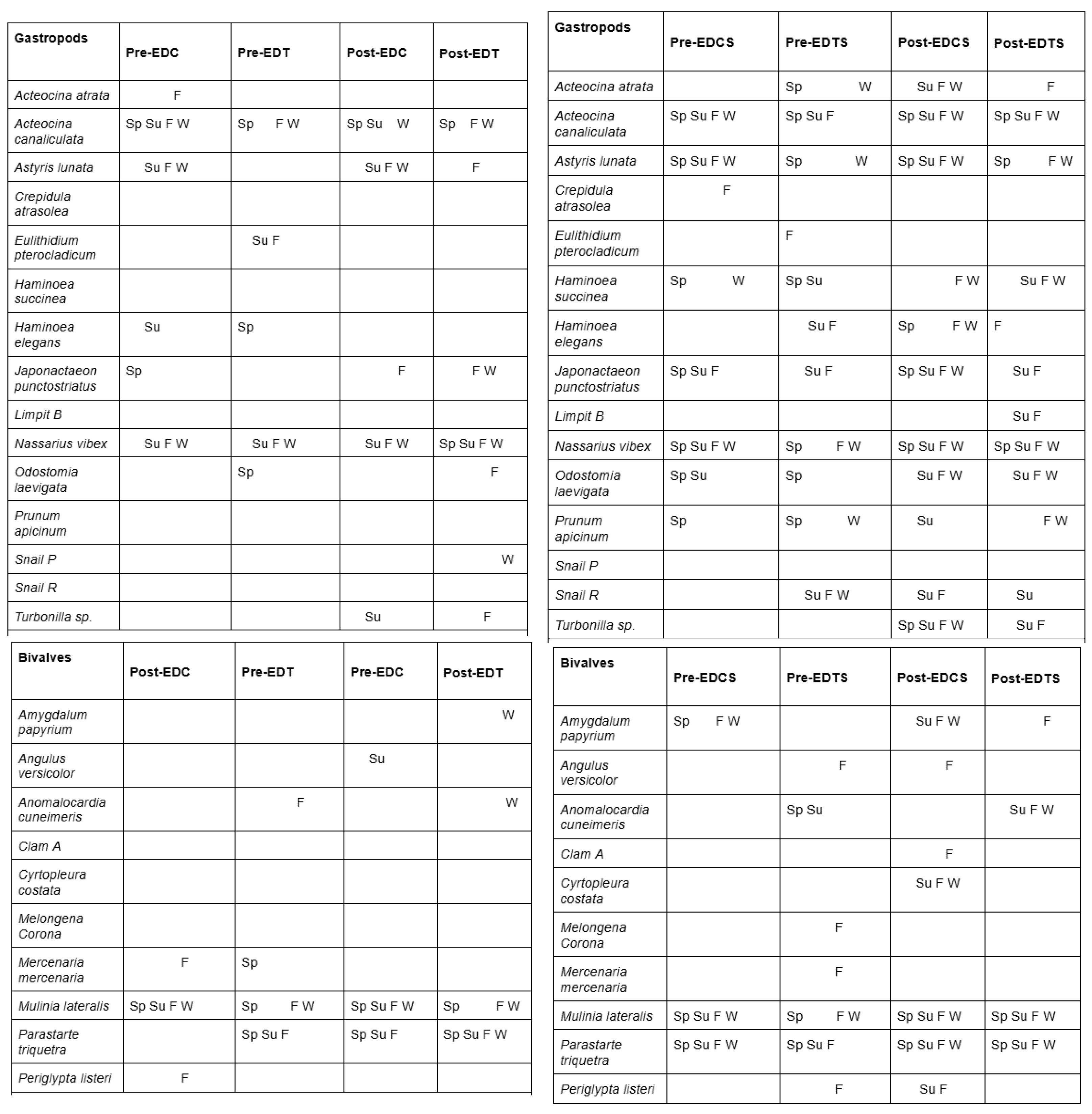
A total of twenty-seven mollusk species were identified at both the treatment and control sites over the course of this study. These represented two classes in Phylum Mollusca: Gastropoda and Bivalvia.

Table 1.
(Left) Presence/absence table of Gastropods and Bivalves at the Environmental Dredging Treatment (EDT) Environmental Dredging Control (EDC) stations by season. (Right) Presence/absence table of Gastropods and Bivalves at the adjacent undredged Seagrass stations (“Environmental Dredging Treatment – Seagrass” or EDTS, and “Environmental Dredging Control – Seagrass” or EDCS). Presense is annotated according to the season the species was observed: Spring (Sp), Summer (Su), Fall (F), Winter (W).
Table 1.
(Left) Presence/absence table of Gastropods and Bivalves at the Environmental Dredging Treatment (EDT) Environmental Dredging Control (EDC) stations by season. (Right) Presence/absence table of Gastropods and Bivalves at the adjacent undredged Seagrass stations (“Environmental Dredging Treatment – Seagrass” or EDTS, and “Environmental Dredging Control – Seagrass” or EDCS). Presense is annotated according to the season the species was observed: Spring (Sp), Summer (Su), Fall (F), Winter (W).
Environmental Dredged Treatment stations (EDT) had an average biodiversity of 0.103 m-2, an average species richness of 0.55 m-2, and an average abundance of 211.11 m-2 before dredging. After dredging, these values were 0.141 m-2, 0.6363 m-2, and 182.27 m-2. Control stations (EDC) had an average biodiversity of 0.21 m-2, an average species richness of 1.056 m-2, and an average abundance of 530.86 organisms m-2 before dredging. After dredging, these values were 0.042 m-2, 0.642 m-2, and 767.08 m-2. Environmental Dredged Seagrass sites (EDTS), adjacent to EDT stations, had an average biodiversity of 0.6 m-2, an average species richness of 2.133 m-2, and an average abundance of 758.52 m-2 before dredging. After dredging, these values were 0.52 m-2, 2.72 m-2, and 4481.55 m-2. Control Seagrass stations, adjacent to EDC stations, had an average biodiversity of 0.65 m-2, an average species richness of 2.23 m-2, and an average abundance of 919.44 m-2 before dredging. After dredging, these values were 0.484 m-2, 3.202 m-2, and 24609.2 m-2.
While only 30% of the identified species were bivalves, the two species with the highest observed abundances were the clams Parastarte triquetra and Mulinia lateralis. These species were particularly abundant in sandier stations, such as EDTS and EDCS (Figure 1). The highest observed abundances of P. triquetra and M. lateralis were 1.7x105 and 3.7x105 individuals m-2 at EDCS in 2020 and 2019, respectively (Figure 1). The adjacent seagrass stations (EDTS and EDCS) were also found to be the stations with the overall highest species richness (the peak richness of 6.33 was found at EDCS in the winter of 2019). All 27 species were found to average 5445 organisms m-2 or greater with an average species richness of 1.69 over the course of this study.
2-Way ANOVAS were conducted on EDT stations, EDC stations, and the adjacent seagrass stations for each of the 27 individual species identified over the course of this study, as well as on a categorical breakdown of gastropods and bivalves at each of the EDT and EDC stations. The results of these ANOVAs have been summarized in table 4.
Individual ANOVA results can be found in Appendix B, and p-values can be found in Appendix A.
Table 4.
A) Significant 2-Way ANOVAs conducted on the biodiversity, species richness, and abundance of benthic mollusks in the different seasons of the study. Xs indicate statistically significant distinctness of the season for that parameter (α=0.05) B) 2-Way ANOVAs for each species’ abundances, conducted season by season. Xs indicate statistically significant distinctness of the season for that species’.bundance (α=0.05).
Table 4.
A) Significant 2-Way ANOVAs conducted on the biodiversity, species richness, and abundance of benthic mollusks in the different seasons of the study. Xs indicate statistically significant distinctness of the season for that parameter (α=0.05) B) 2-Way ANOVAs for each species’ abundances, conducted season by season. Xs indicate statistically significant distinctness of the season for that species’.bundance (α=0.05).
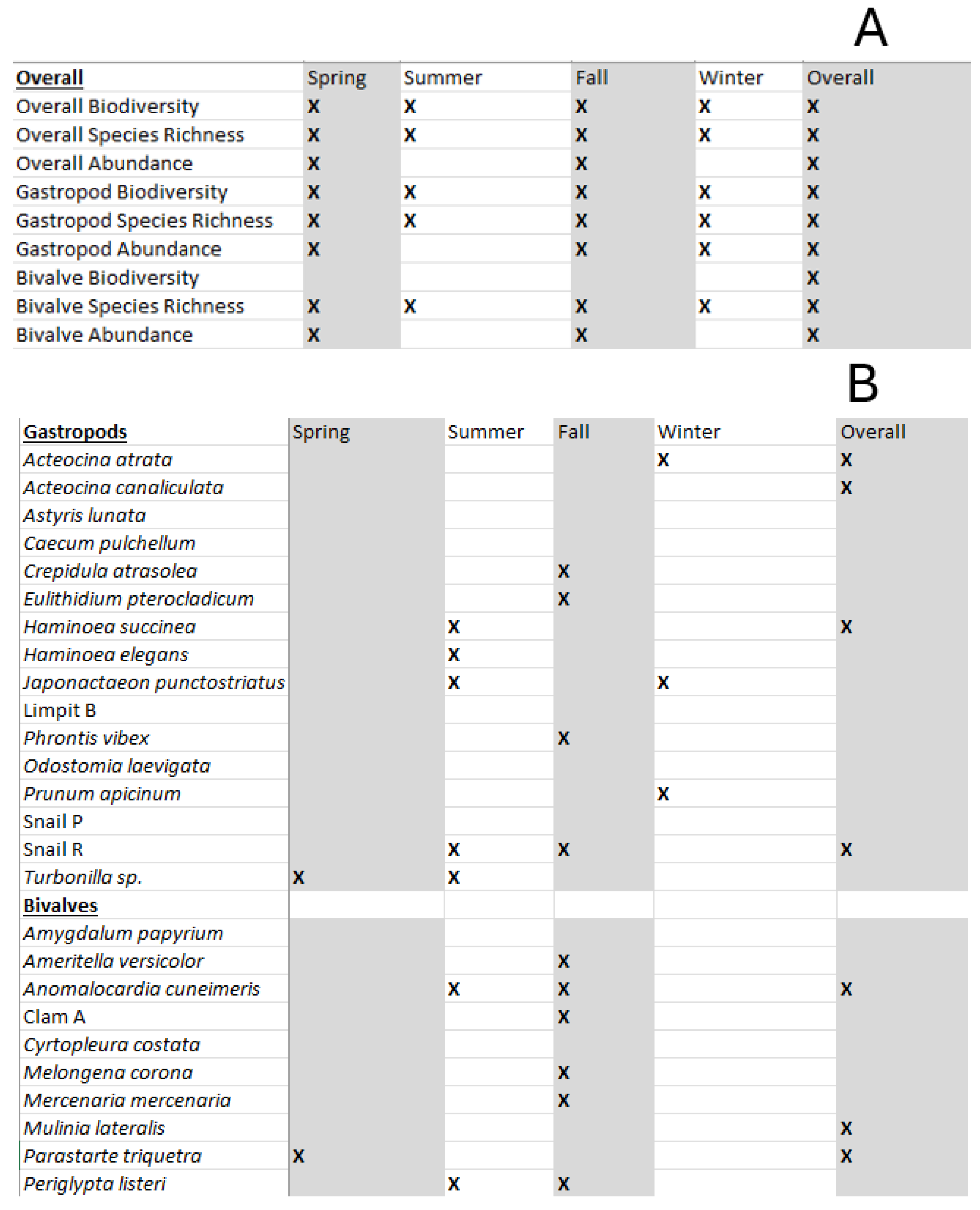
Figure 2.
A) Mollusk abundance m-2 vs. sediment % organic content at muck stations (EDT=orange; EDC=blue). R2 was 0.1 and 0.01 for EDT and EDC, respectively, and significant for EDT (p-0.02). B) Mollusk abundance m-2 vs sediment % organic content at seagrass sites near muck (EDTS=orange; EDCS=blue). R2 was 0.12 and 0.001 for EDTS and EDCS, respectively, and significant for EDTS (p=0.028). C) Benthic mollusk abundance m-2 vs sediment % silt-clay content at muck station (EDT=orange; EDC=blue). R2 was 0.0625 and 0.56 for EDT and EDC, respectively, and significant in both cases (p= 0.048 and 0.0001, respectively).
Figure 2.
A) Mollusk abundance m-2 vs. sediment % organic content at muck stations (EDT=orange; EDC=blue). R2 was 0.1 and 0.01 for EDT and EDC, respectively, and significant for EDT (p-0.02). B) Mollusk abundance m-2 vs sediment % organic content at seagrass sites near muck (EDTS=orange; EDCS=blue). R2 was 0.12 and 0.001 for EDTS and EDCS, respectively, and significant for EDTS (p=0.028). C) Benthic mollusk abundance m-2 vs sediment % silt-clay content at muck station (EDT=orange; EDC=blue). R2 was 0.0625 and 0.56 for EDT and EDC, respectively, and significant in both cases (p= 0.048 and 0.0001, respectively).
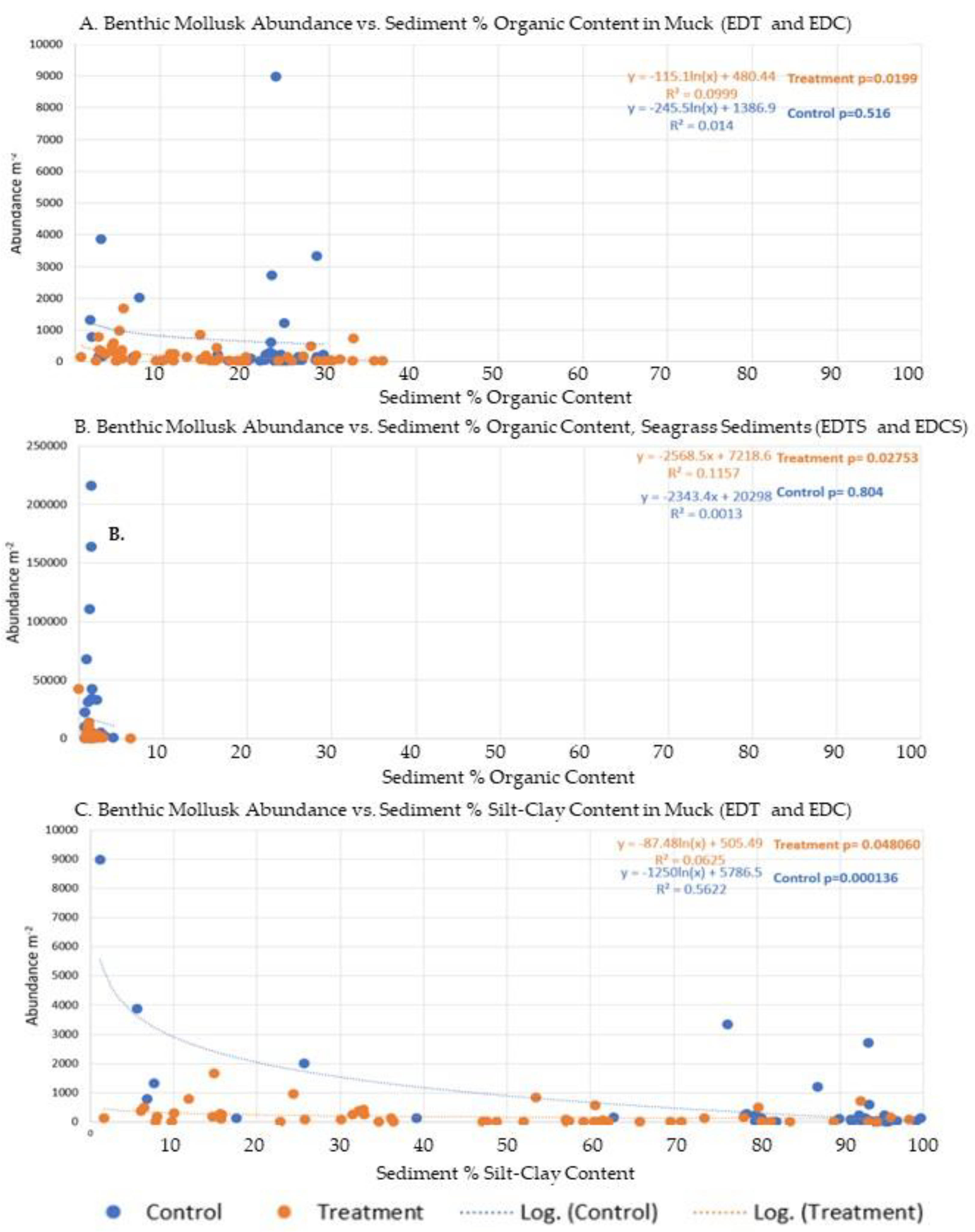
Figure 3.
A) Benthic mollusk collective biodiversity (m-2) vs sediment percent organic content at dredging treatment (orange) and control stations (blue). Regression relationships were weak (R2=0.15 and 0.2, respectively), but significant in both cases (p=0.004 and 0.009 respectively))> B) Benthic mollusk collective biodiversity (m-2) vs sediment percent silt clay content at dredging treatment (orange) and control stations (blue). Regression relationships were moderate to weak (R2 = 0.06 and 0.24, respectively), but significant in the control (p-0.00074).
Figure 3.
A) Benthic mollusk collective biodiversity (m-2) vs sediment percent organic content at dredging treatment (orange) and control stations (blue). Regression relationships were weak (R2=0.15 and 0.2, respectively), but significant in both cases (p=0.004 and 0.009 respectively))> B) Benthic mollusk collective biodiversity (m-2) vs sediment percent silt clay content at dredging treatment (orange) and control stations (blue). Regression relationships were moderate to weak (R2 = 0.06 and 0.24, respectively), but significant in the control (p-0.00074).
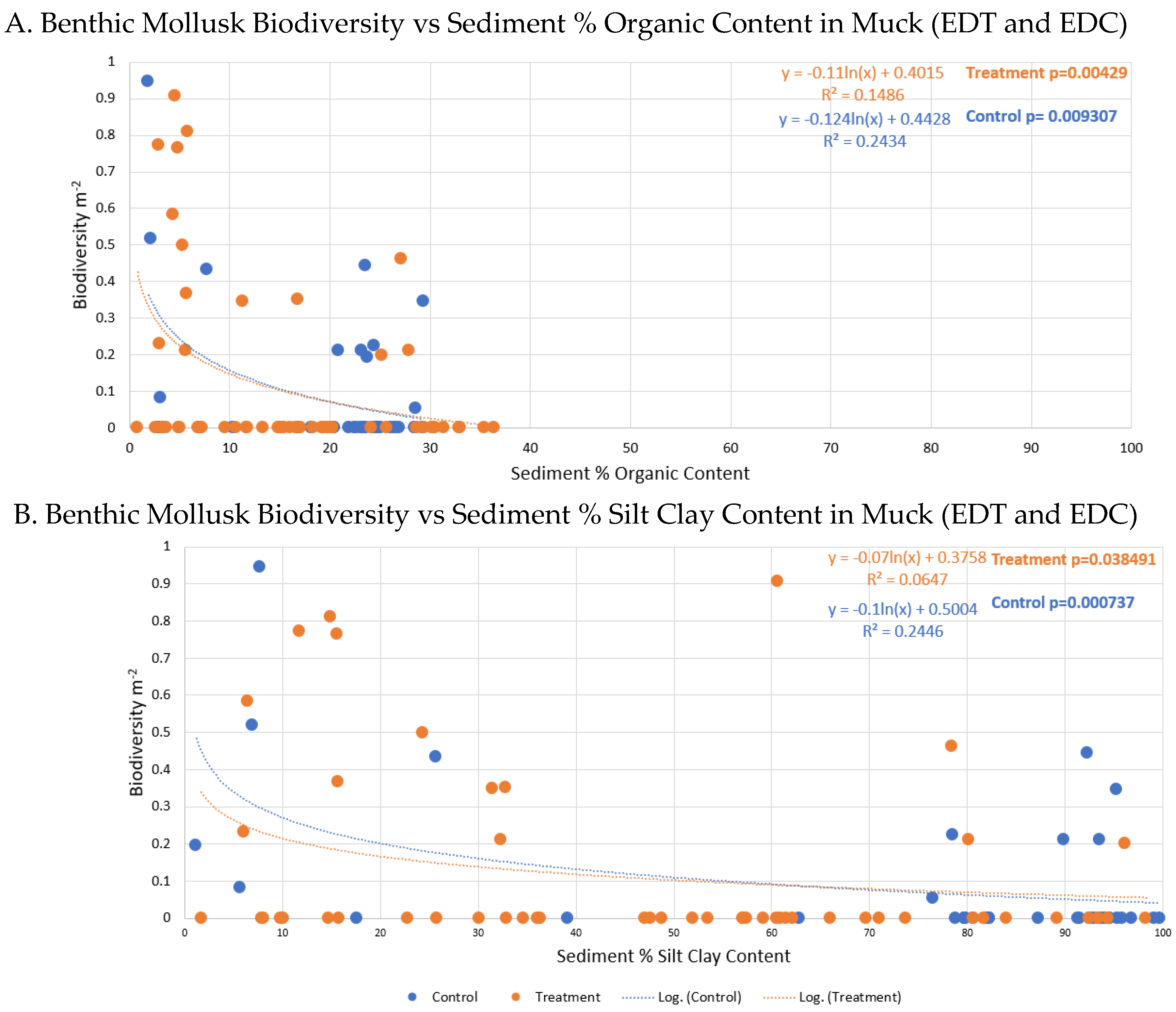
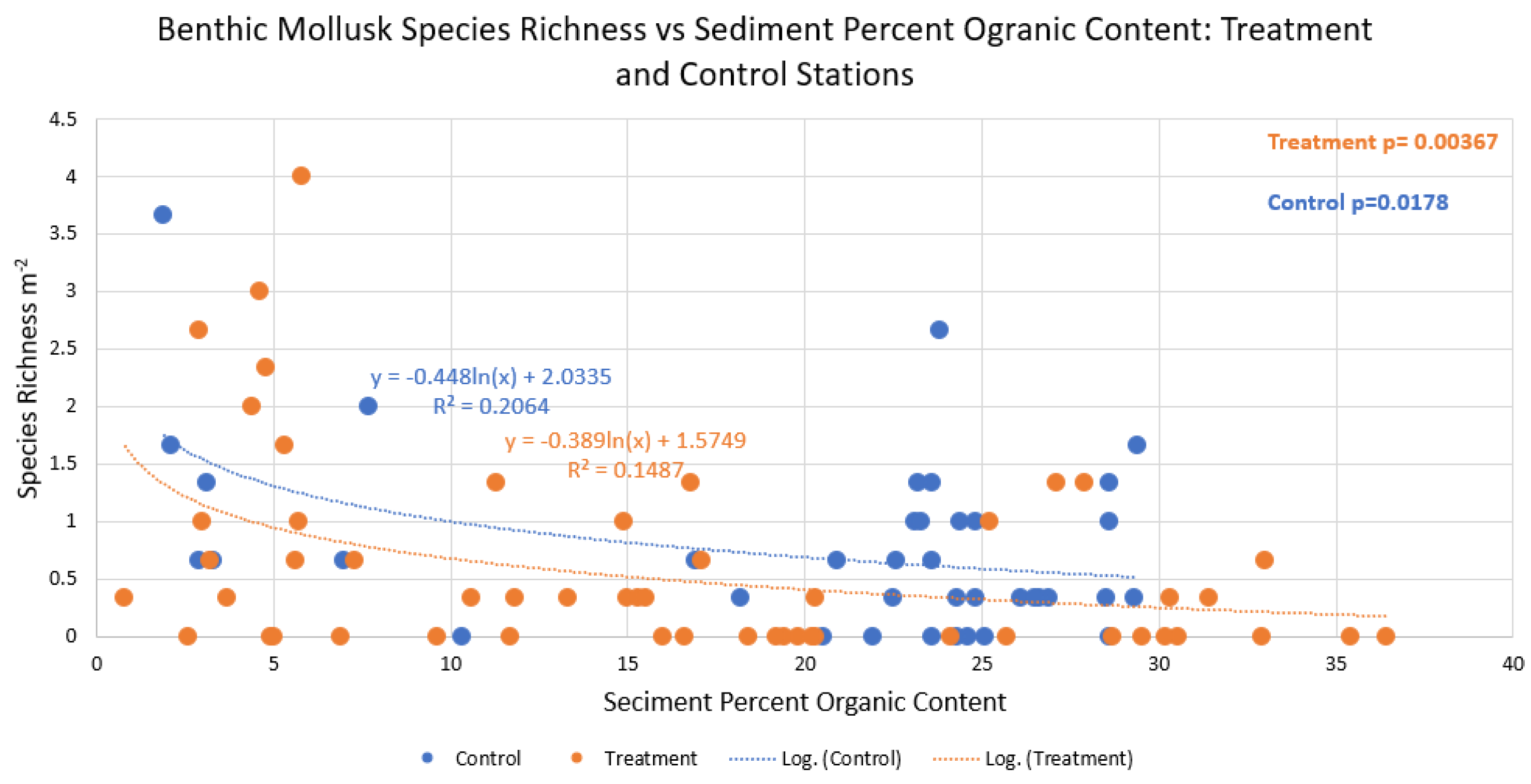
Figure 4.
Benthic mollusk collective species richness (m-2) vs sediment percent organic content at dredging treatment (orange) and control stations (blue). Regression relationships were weak (R2 = 0.15 and 0.21, respectively), but significant in both the treatment and control (p=0.004 and 0.01 respectively).
Figure 4.
Benthic mollusk collective species richness (m-2) vs sediment percent organic content at dredging treatment (orange) and control stations (blue). Regression relationships were weak (R2 = 0.15 and 0.21, respectively), but significant in both the treatment and control (p=0.004 and 0.01 respectively).
The organic content of the sediments ranged from 0.7% to 36.4%. The higher organic content sediments (0.8% to 36.4%) were found in the Environmental Dredged Treatment (EDT) and Control (EDC) stations (Figure 1: EDC 1, 2, and 3, and EDT 1,2,3, and 4). Accordingly, these organic-rich stations consistently had the lowest overall biodiversity (0-0.46), species richness (0-2.67), and abundances (0-3318.52 organisms m-2). Sediments lower in percent organic content (0.7% to 6.1%) were located near the adjacent seagrass beds. Seagrass-adjacent sediments with higher organic content generally had low biodiversity (0-0.6667). Seagrass-adjacent sediments with lower organic content generally had higher biodiversity (0-1.337). Generalized linear models found that sediment percent organic content had an inverse logarithmic correlation with overall mollusk biodiversity, species richness, and abundance at the EDT and EDC stations. Sediment percent organic content also had an inverse relationship with benthic mollusk abundance at the adjacent seagrass stations.
When data were separated by taxonomic family, gastropod biodiversity was found to range between 0-0.54 with higher sediment organic content, and 0-1.28 with lower sediment organic content. Bivalve biodiversity was found to range between 0-0.63 with higher sediment organic content, and 0-0.67 with lower sediment organic content.
The water content of the sediments ranged from 21.3% to 86.9%. Sediments generally higher in water content, ranging from 26.3% to 86.9%, were found in the treatment and control muck (EDT and EDC) stations, whereas sediments generally lower in water content, ranging 21.3% to 49.1% water content, were located near the adjacent seagrass stations.
The sediment % silt-clay content of the sediments ranged from 0.2% to 99.7%. Sediments generally higher in silt clay content, ranging from 1.2% to 99.7%, were found in the treatment and control muck (EDT and EDC) stations, whereas sediments generally lower in silt clay content, ranging from 0.2% to 73.7% were found in the adjacent seagrass stations. Generalized linear models found an inverse relationship between sediment percent silt clay content and benthic mollusk biodiversity and abundance.
The percent dissolved oxygen of the water ranged from 0.01-141.4%. The treatment and control muck (EDT and EDC) stations typically had the lowest percent dissolved oxygen (0.01-114%). The adjacent seagrass stations typically had the highest percent dissolved oxygen (18.6-141.4%).
ANOSIM was performed on these data, which found that treatment, year, sediment percent organic content, and sediment percent water content were statistically significant factors (p<0.05). When data was separated by family at the adjacent seagrass stations (Figure 1), it was found that year and treatment were statistically significant (p<0.05) for both gastropods and bivalves. Salinity and water temperature were found to be statistically significant for bivalves at the treatment (EDT) stations.

An even distribution between groups is determined by the R-value closer to 0 than 1. High and low ranks are evenly distributed for comparisons within and between groups (Table 5). Higher R-values indicate dissimilarity of groups. The ANOSIMs were paired with non-metric multi-dimensional scaling (nMDS) models to provide a visual representation of similarities and dissimilarities between groups. Overall, there was a compelling trend in grouping between stations. At all of the treatment and control muck (EDT and EDC) stations, there was a large amount of overlap with all groups, with some separation of the Treatment-Before-Dredging (Pre-EDT) group. At adjacent seagrass stations, there was also considerable overlap, with some separation of the Treatment Post Dredging (Post-EDT) group. All individual nMDS results can be found in Appendix C. Examples of the aforementioned separation pattern are included below.
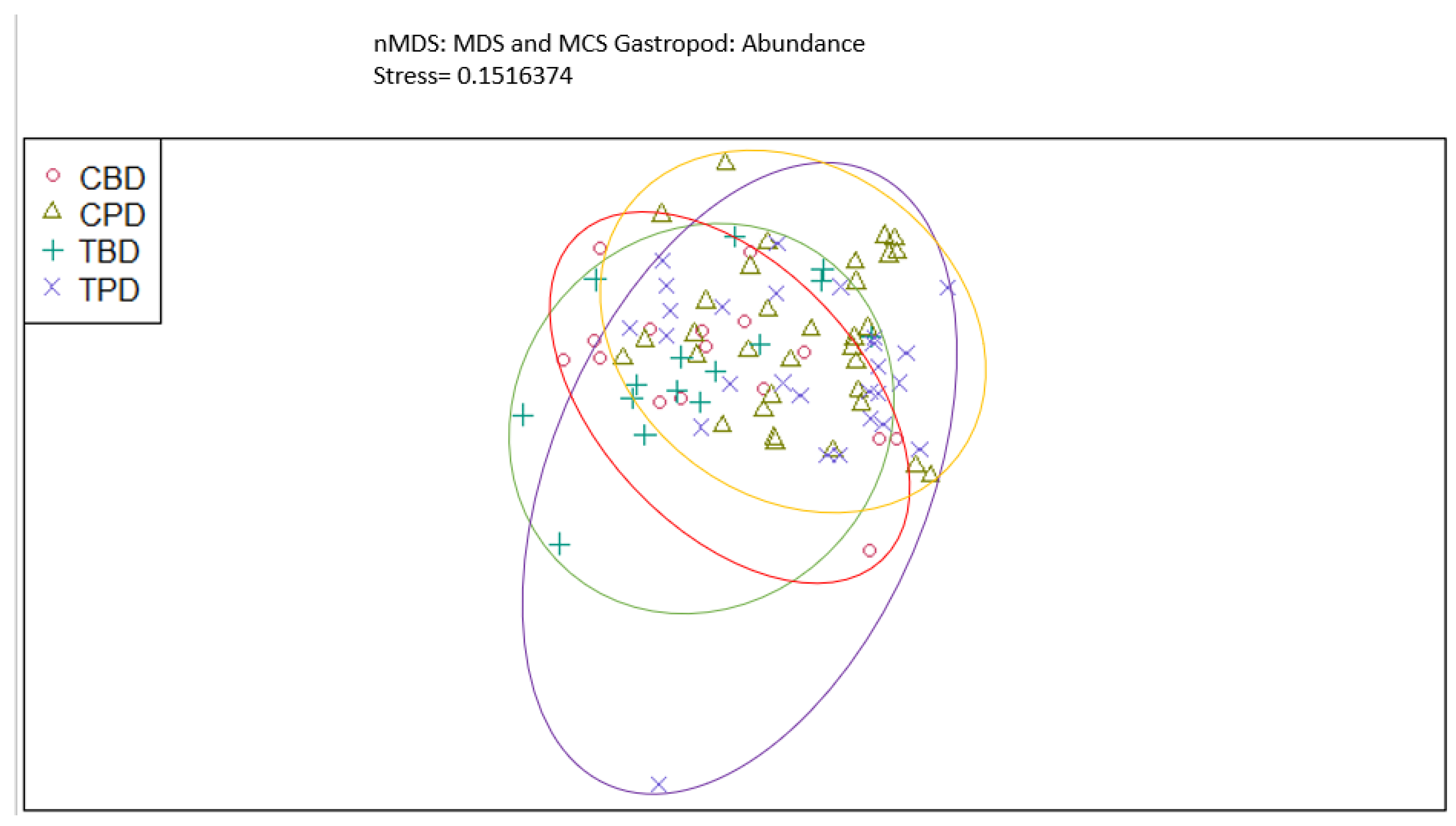
Figure 8.
Non-Metric Multidimensional Scaling reveals pooled mollusk biodiversity at the treatment and control muck stations similarities and dissimilarities in multivariate space. CBD=Control Before Dredging; CPD=Control Post Dredging; TBD=Treatment Before Dredging; TPD=Treatment Post Dredging. Non-overlapping portions of ellipses represent the proportion of group distinctiveness or dissimilarity.
Figure 8.
Non-Metric Multidimensional Scaling reveals pooled mollusk biodiversity at the treatment and control muck stations similarities and dissimilarities in multivariate space. CBD=Control Before Dredging; CPD=Control Post Dredging; TBD=Treatment Before Dredging; TPD=Treatment Post Dredging. Non-overlapping portions of ellipses represent the proportion of group distinctiveness or dissimilarity.
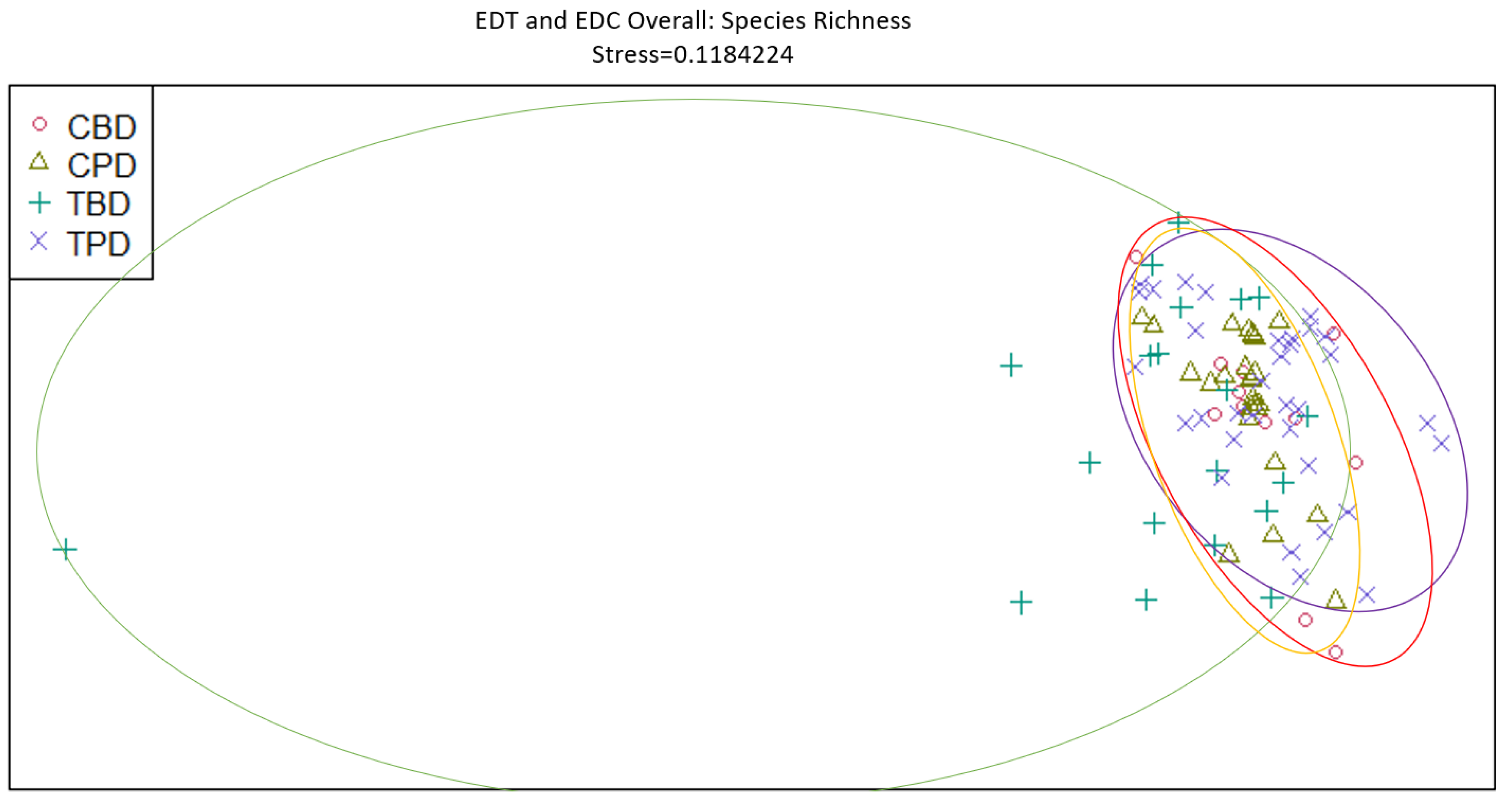
Figure 9.
Non-Metric Multidimensional Scaling reveals pooled mollusk species richness at the treatment and control muck stations similarities and dissimilarities in multivariate space. CBD=Control Before Dredging; CPD=Control Post Dredging; TBD=Treatment Before Dredging; TPD=Treatment Post Dredging. Non-overlapping portions of ellipses represent the proportion of group distinctiveness or dissimilarity.
Figure 9.
Non-Metric Multidimensional Scaling reveals pooled mollusk species richness at the treatment and control muck stations similarities and dissimilarities in multivariate space. CBD=Control Before Dredging; CPD=Control Post Dredging; TBD=Treatment Before Dredging; TPD=Treatment Post Dredging. Non-overlapping portions of ellipses represent the proportion of group distinctiveness or dissimilarity.
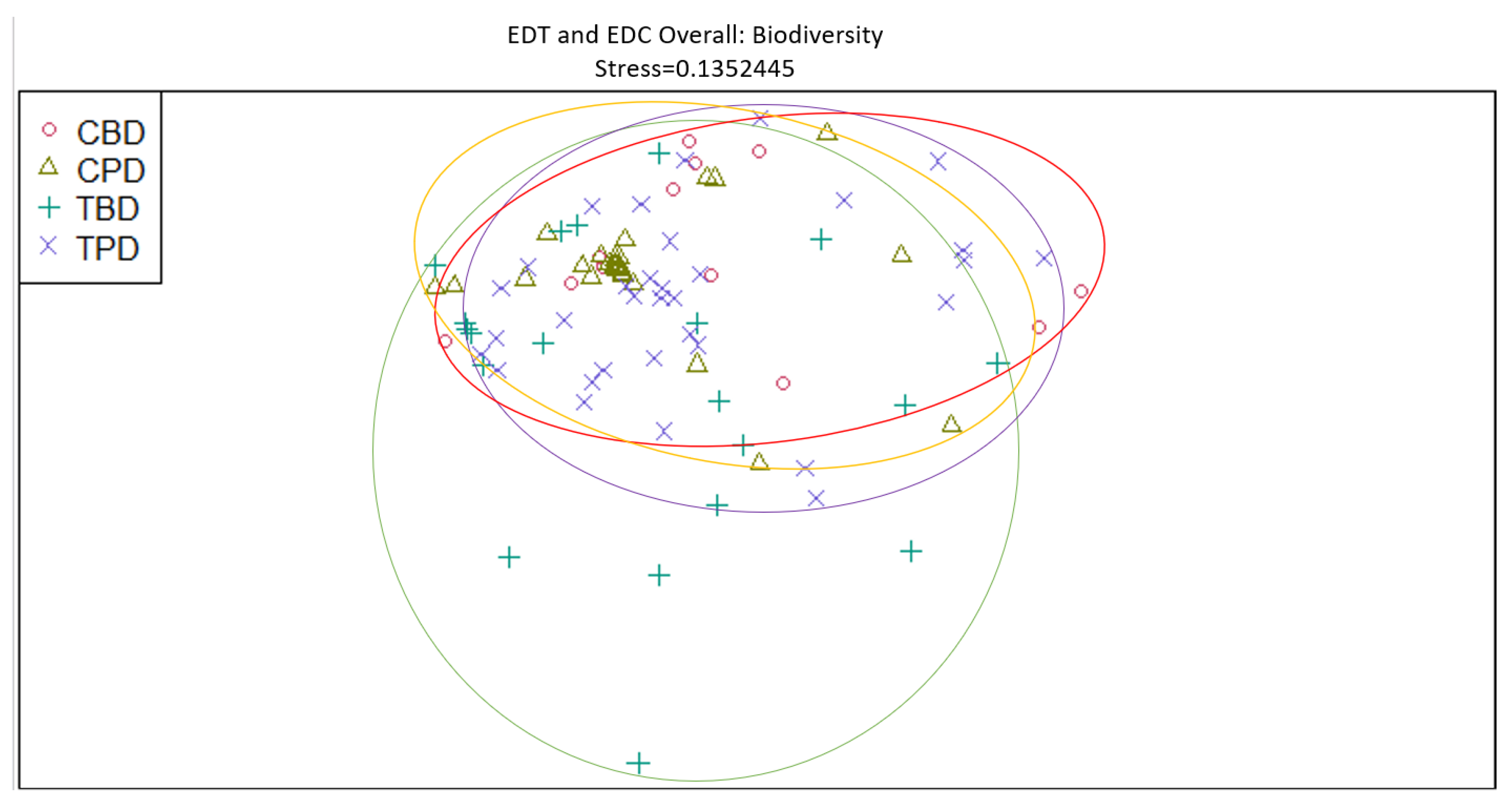
Figure 10.
Non-Metric Multidimensional Scaling reveals pooled mollusk abundance at the treatment and control muck stations similarities and dissimilarities in multivariate space. CBD=Control Before Dredging; CPD=Control Post Dredging; TBD=Treatment Before Dredging; TPD=Treatment Post Dredging. Non-overlapping portions of ellipses represent the proportion of group distinctiveness or dissimilarity.
Figure 10.
Non-Metric Multidimensional Scaling reveals pooled mollusk abundance at the treatment and control muck stations similarities and dissimilarities in multivariate space. CBD=Control Before Dredging; CPD=Control Post Dredging; TBD=Treatment Before Dredging; TPD=Treatment Post Dredging. Non-overlapping portions of ellipses represent the proportion of group distinctiveness or dissimilarity.
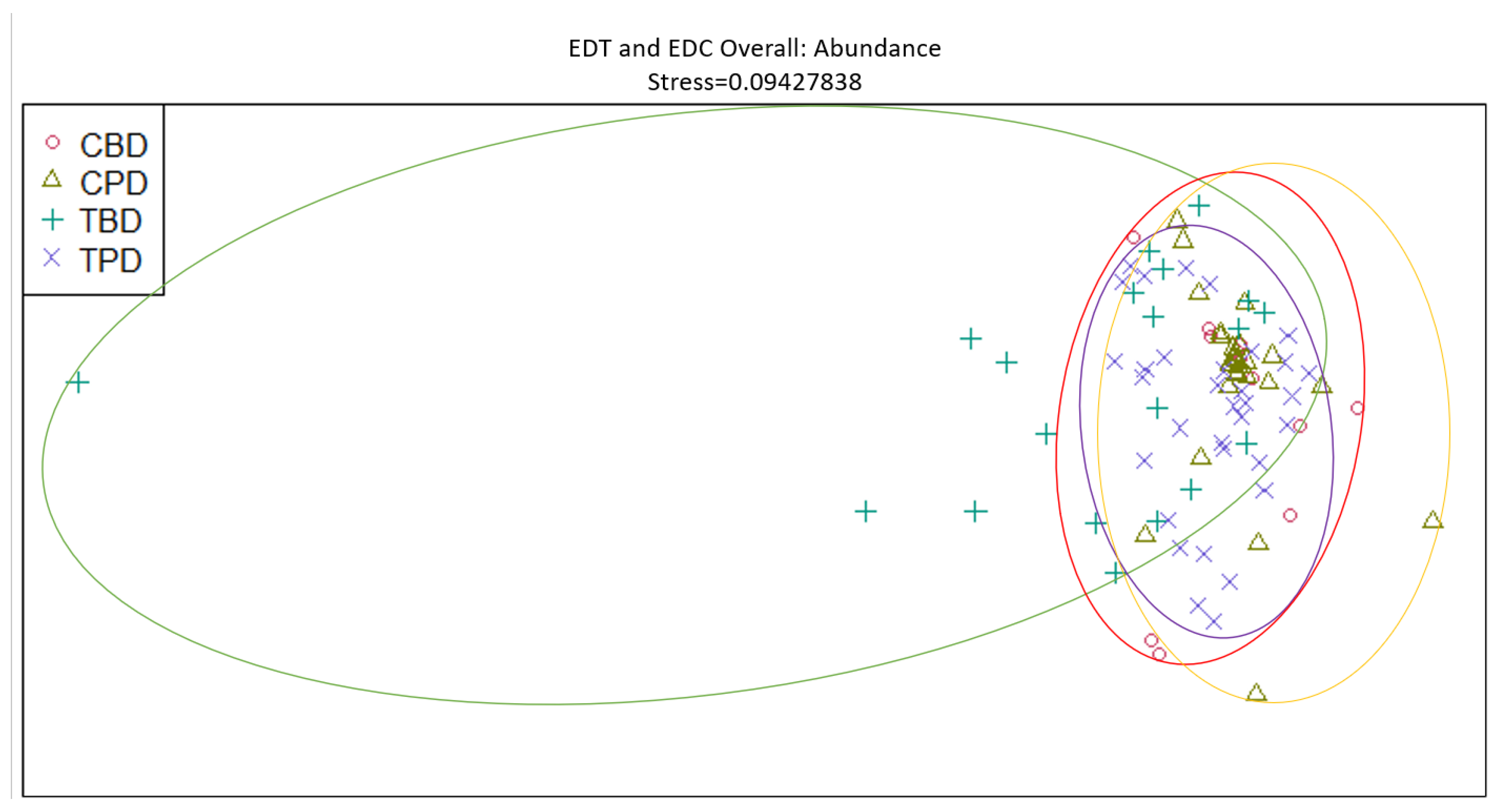
Figure 11.
Non-Metric Multidimensional Scaling reveals pooled mollusk biodiversity at the adjacent seagrass stations similarities and dissimilarities in multivariate space. CBD=Control Before Dredging; CPD=Control Post Dredging; TBD=Treatment Before Dredging; TPD=Treatment Post Dredging. Non-overlapping portions of ellipses represent the proportion of group distinctiveness or dissimilarity.
Figure 11.
Non-Metric Multidimensional Scaling reveals pooled mollusk biodiversity at the adjacent seagrass stations similarities and dissimilarities in multivariate space. CBD=Control Before Dredging; CPD=Control Post Dredging; TBD=Treatment Before Dredging; TPD=Treatment Post Dredging. Non-overlapping portions of ellipses represent the proportion of group distinctiveness or dissimilarity.
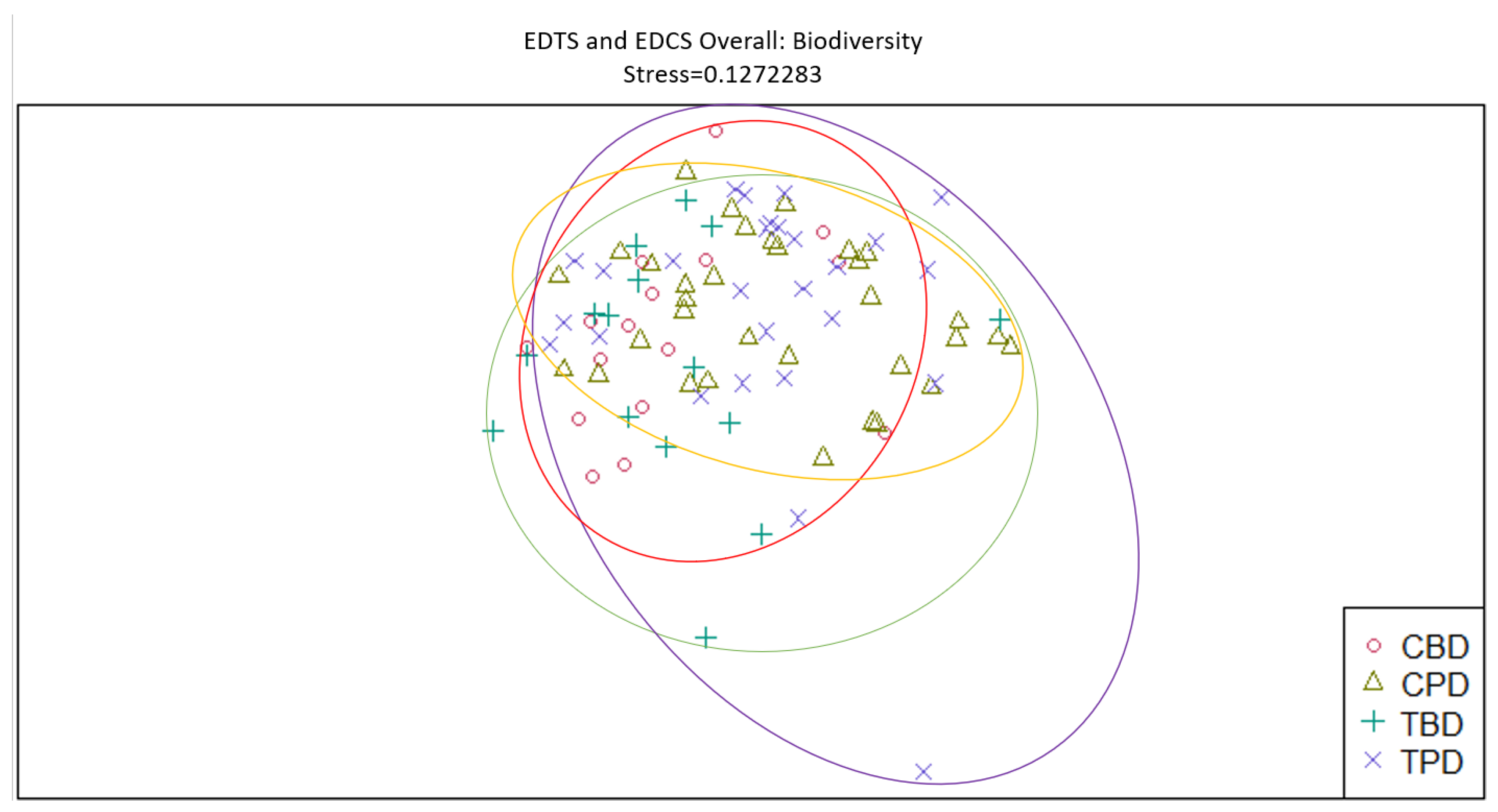
Figure 12.
Non-Metric Multidimensional Scaling reveals pooled mollusk species richness at the adjacent seagrass stations similarities and dissimilarities in multivariate space. CBD=Control Before Dredging; CPD=Control Post Dredging; TBD=Treatment Before Dredging; TPD=Treatment Post Dredging. Non-overlapping portions of ellipses represent the proportion of group distinctiveness or dissimilarity.
Figure 12.
Non-Metric Multidimensional Scaling reveals pooled mollusk species richness at the adjacent seagrass stations similarities and dissimilarities in multivariate space. CBD=Control Before Dredging; CPD=Control Post Dredging; TBD=Treatment Before Dredging; TPD=Treatment Post Dredging. Non-overlapping portions of ellipses represent the proportion of group distinctiveness or dissimilarity.
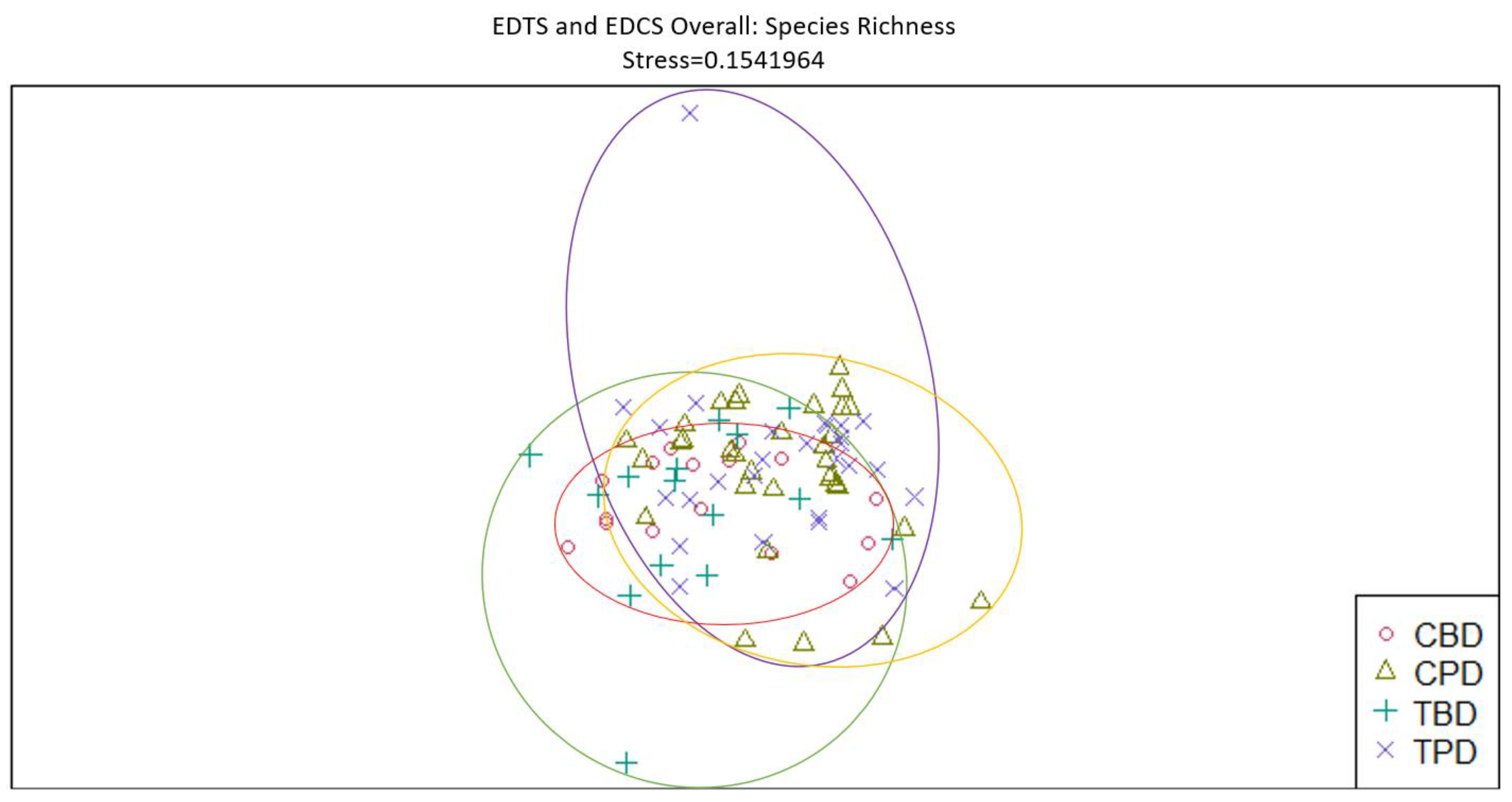
Figure 13.
Non-Metric Multidimensional Scaling reveals the pooled mollusk species richness at the adjacent seagrass stations similarities and dissimilarities in multivariate space. CBD=Control Before Dredging; CPD=Control Post Dredging; TBD=Treatment Before Dredging; TPD=Treatment Post Dredging. Non-overlapping portions of ellipses represent the proportion of group distinctiveness or dissimilarity.
Figure 13.
Non-Metric Multidimensional Scaling reveals the pooled mollusk species richness at the adjacent seagrass stations similarities and dissimilarities in multivariate space. CBD=Control Before Dredging; CPD=Control Post Dredging; TBD=Treatment Before Dredging; TPD=Treatment Post Dredging. Non-overlapping portions of ellipses represent the proportion of group distinctiveness or dissimilarity.
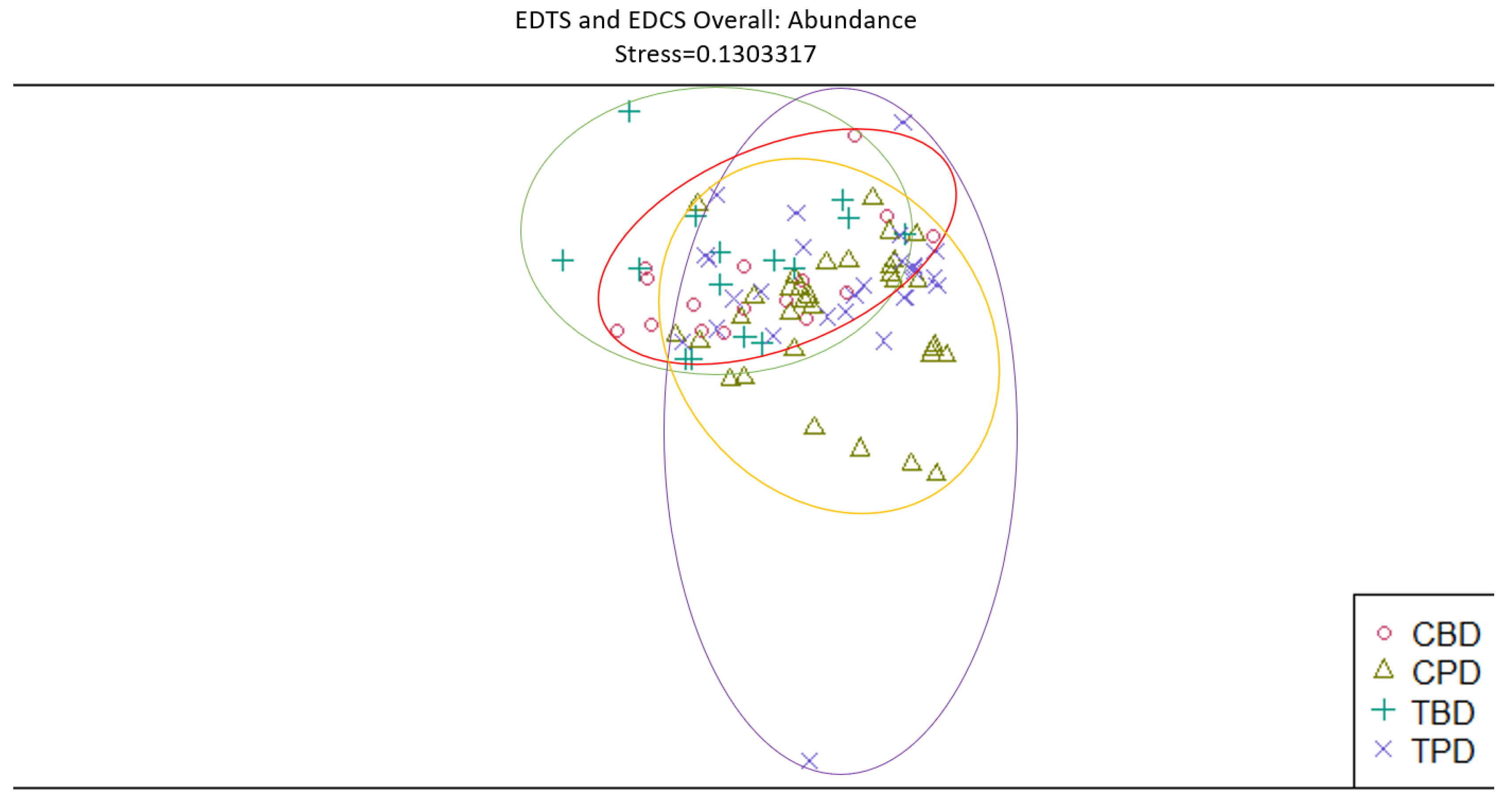
Figure 14.
Non-Metric Multidimensional Scaling reveals the spread of 27 identified mollusk species abundances at all stations similarities and dissimilarities in multivariate space. MCM=Mims Control Muck; MCS=Mims Control Seagrass; MDM=Mims Dredged Muck; MDS=Mims Dredged Seagrass. Non-overlapping portions of ellipses represent the proportion of group distinctiveness or dissimilarity.
Figure 14.
Non-Metric Multidimensional Scaling reveals the spread of 27 identified mollusk species abundances at all stations similarities and dissimilarities in multivariate space. MCM=Mims Control Muck; MCS=Mims Control Seagrass; MDM=Mims Dredged Muck; MDS=Mims Dredged Seagrass. Non-overlapping portions of ellipses represent the proportion of group distinctiveness or dissimilarity.
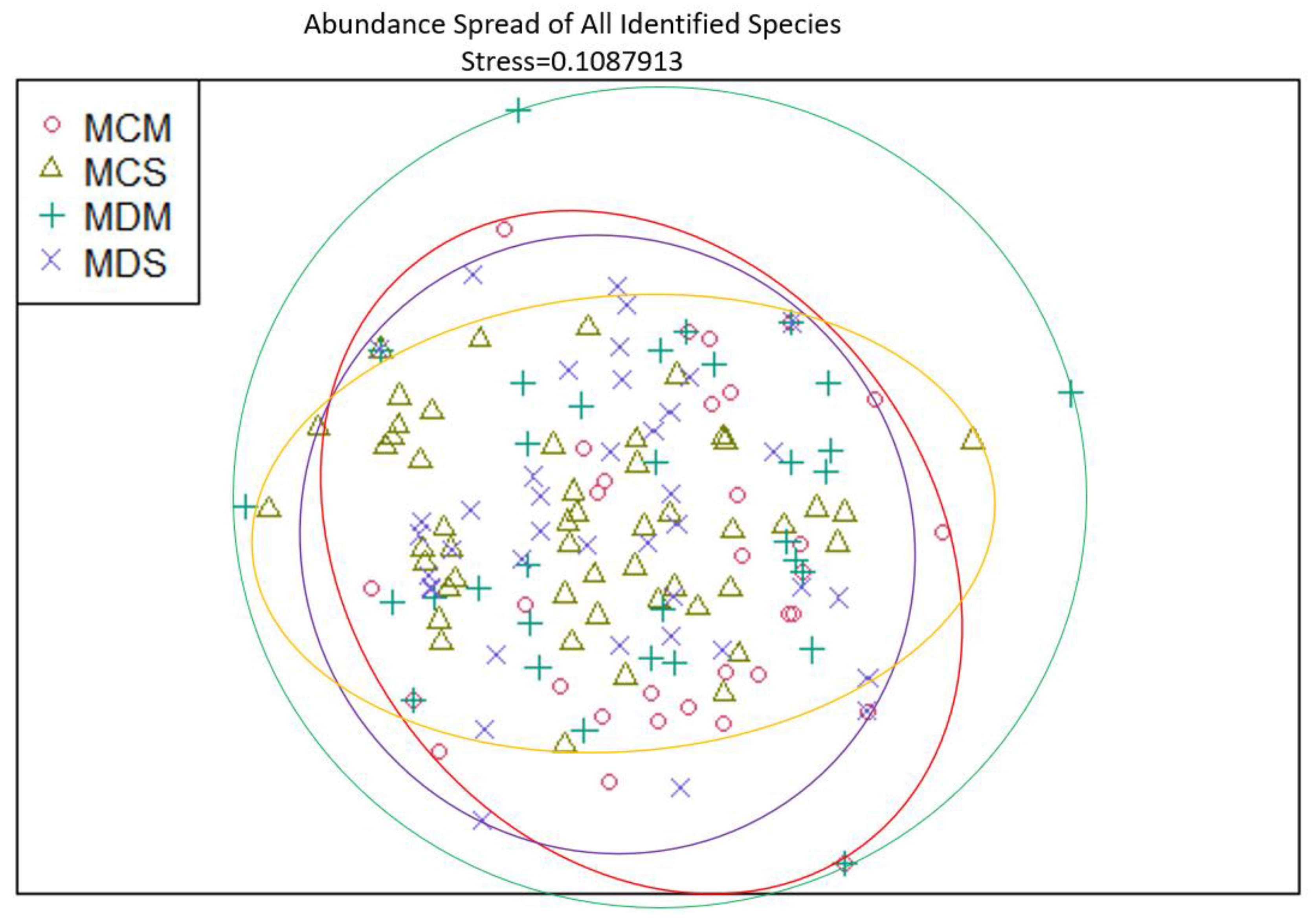
The abundance spread of each of the 27 identified species were also analyzed as a community. These results also reflect the aforementioned overlap pattern.
4. Discussion
The purpose of this study was to determine the effects of environmental dredging on benthic mollusk communities in a shallow and diverse estuary. We also examined taxonomic differences in dredging responses. Results show an inverse logarithmic relationship between benthic mollusk community composition, biodiversity, species richness, and abundance and sediment % organic content.
Sediment percent organic content and silt-clay content have inverse relationships with benthic mollusk biodiversity, species richness, and abundances (Figure 2, Figure 3, Figure 4, Figure 5, Figure 6 and Figure 7), supporting the first hypothesis of this study, that biological parameters will have an inverse relationship with sediment organic content. This inverse relationship, with population measurements plummeting as muck organic content increases, arises from increased stress stemming from the many impacts of living in polluted sediments.
In this study, treatment and control muck (EDT and EDC) stations had higher sediment % organic content (Figure 1). Those muck stations also had lower percent dissolved oxygen in the boundary bottom water (47.55% ± 29.99%) and, accordingly, the lowest overall biodiversity (0.11 ± 0.227 m-2), species richness (0.67 ± 0.84 m-2), and abundances (405.636 ± 1118.44 organisms m-2). Sediment organic content holds the potential to represent benthic community health because it has a substantial and direct influence on benthic biochemistry [29]. As fine-grained benthic organic matter (a.k.a. muck) accumulates, it binds to silt and clay particulates [36,37]. Benthic bacteria then break down and digest organic matter, depleting dissolved oxygen and increasing H2S [38]. This recycling process is a natural phenomenon in aquatic and marine ecosystems. However, with excessive concentrations of organic matter, this process exposes benthic life to hypoxia and toxic hydrogen sulfide. These conditions can alter community composition by excluding sensitive species and restricting sediment bioturbation [39,40,41]. Lack of bioturbation can perpetuate hypoxic conditions in sediments. This creates a positive feedback loop, further increasing inhospitable conditions [42].
Negative impacts of degraded sediments on benthic mollusk communities have been observed and modeled in other systems. The Pearson and Rosenberg model [43] showcases a pattern of faunal richness and abundance increasing with lower levels of sediment organic content and then decreasing in response to higher concentrations. Other studies have also found that species richness and biodiversity increase with organic matter concentration, up to about 3% in some cases [44], but then rapidly plummet as organic content continues to increase [29,45,46]. These corroborating studies all show sediment relationships similar to those found in this study. However, the Pearson and Rosenberg model may need to be modified for coastal environments due to system energy disparities, since systems with less wave energy have more sedimentation and lower dissolved oxygen [47]. Abundant oxygen and low toxicity generally foster healthier, more diverse, and abundant benthic communities. As adverse conditions increase, some taxa are better adapted than others for withstanding the impacts.
There was no discernable difference in benthic mollusk community composition between muck that was dredged vs. undredged. Removal of sediments with high concentrations of fine-grained organic matter by dredging did not increase the biodiversity, species richness, or overall community abundance of benthic mollusks during the time period of the study. Since the decay of organic matter creates stressful hypoxic or anoxic conditions, low dissolved oxygen is a likely limiting factor for some benthic infauna [29,44,49]. However, this did not translate to substantial distinctiveness between communities in this study (Figure 8, Figure 9, Figure 10, Figure 11, Figure 12, Figure 13 and Figure 14). This could be due to the lower metabolic and oxygen consumption rates of mollusks, which allow them to persist in polluted sediments where oxygen is more limiting [23,24,25,26]. When oxygen is not a limiting factor, more tolerant species will gain an advantage in polluted/organic-rich sediments over the more sensitive competitors [29,42,43]. This can be seen with the high abundances of Parastarte triquetra and Ameritella versicolor in this study. P. triquetra was the most abundant mollusk in this study, reaching counts of 8.6 x 103 individuals m-2 at the control muck stations, and up to 2.6 x 105 individuals m-2 at the control seagrass stations. A. versicolor was recorded at only 3 station sampling events in this study, reaching counts of 15 individuals m-2 at the control muck stations, and 356 individuals m-2 at the control seagrass stations. It may be that lower metabolic rates in mollusks prevent oxygen levels from rapidly shaping benthic communities. However, variability was high and no significant pre- vs. post-dredging differences were observed in the community (Figure 8, Figure 9, Figure 10, Figure 11, Figure 12, Figure 13 and Figure 14). One possible explanation for this is that dredging does not remove all of the muck overlaying the prime sediments, and organic sediment conditions persist after environmental dredging is complete.
Anoxia resulting from the mass decomposition of organic matter fosters the occurrence of sulfur-reducing bacteria [49,50]. These bacteria release H2S that can saturate the benthic boundary layer and interstitial water, rendering hypoxic habitats still more hostile to fauna and flora [38]. Fauna with H2S resistance tend to survive better in hypoxic environments [27,28]. Gray et al. [48] observed that crustaceans and echinoderms are most sensitive to polluted fine-grained organic-rich sediments. Anelids are more tolerant than these two groups and mollusks are the most tolerant of organic sediments. This is consistent with their documented ability to withstand the physiological stressors of hypoxia and H2S [27].
Competition is a major factor in the shaping of community structures in many ecosystems [51]. However, Thistle [52] found that colonizing species’ patterns of succession may be driven by the nature of the resource base, rather than competition. Additionally, Virnstein [53] observed that, in the absence of predation, Mulinia lateralis persisted at high densities and could exclude other species. In this study, M. lateralis was one of the most abundant species identified, reaching densities of 4267 individuals m-2 at the control seagrass stations and 3793 individuals m-2 at the control muck stations. However, Virnstein [53] postulated that the potential for M. lateralis to reach these densities and exclude other species could have been due to predation on settling larvae or another amensalistic interaction, as some species are poor “predation avoiders”. Other corroborating predator exclusion studies have also reported an absence of competitive exclusion, even where benthic fauna occurred at higher densities [54,55]. Peterson [56] postulated that this absence of competitive exclusion in soft sediments could be due to reduced opportunities for interference competition, and, rather, increased opportunities for amensalistic interactions. Since the introduction of the trophic amensalism hypothesis [31], it has been used to explain the distribution of deposit feeders and suspension feeders in numerous polluted and other soft-sediment benthic environments [51,56,57,58,59]. Rhoades and Young’s trophic amensalism hypothesis proposes that deposit feeders exclude filter-feeding neighbors by disturbing and resuspending fine-grained sediments that can clog their fine gills and feeding structures [31]. However, the results of this study did not see a discrepancy in the abundances of suspension-feeding vs. deposit-feeding mollusks before or after dredging, despite the polluted sediment conditions.
Many benthic macroinvertebrates are used as bioindicators for environmental change because they are easy to monitor, have limited escape mechanisms to avoid disturbances and provide a good indication of change over time [60,61]. Numerous studies have shown that the benthos responds relatively rapidly to stressors, both natural and anthropogenic [62]. Benthic mollusks are typically very abundant, relatively sessile, and tend to have longer life cycles [17,62]. Some studies on macrofaunal succession found that mollusks dominate less polluted habitats and that degraded habitats replace mollusks with unshelled organisms (e.g., polychaetes) [61]. However, our study found that mollusks can tolerate very degraded conditions, reaching densities as high as 9.0 x 103 individuals m-2 and 1.7 x 103 individuals m-2 at control and treatment (EDC and EDT) sites, respectively. In one study conducted by Pelletiera et al., it was found that the snail Acteocina sp. is a “pollution-sensitive species”, making its absence a potential bioindicator of polluted sediments. Two species of Acteocina were observed in this study, Acteocina atrata and Acteocina canaliculata. A. atrata was only recorded at 13 station sampling events over the course of this study, 12 of which were at seagrass stations. A. atrata reached counts of 830 individuals m-2 at the control seagrass-adjacent stations and 59 individuals m-2 at the one control muck station. A. canaliculata was occurred more frequently and was far more dense than A. atrata. A. canaliculata was recorded at 79 station sampling events over the course of this study, reaching counts of 1.8 x 103 individuals m-2 at the control seagrass stations and 2.7 x 102 individuals m-2 at the control muck stations. Two-Way ANOVAs were conducted on all 27 species identified over the course of this study. The ANOVAs conducted on both A. atrata and A. canaliculata found statistically significant differences in the abundances of both species between the seagrass stations and the muck stations. These findings, along with the documented sensitivity to pollution, could make A. atrata and A. canaliculata potential indicator species for future studies on benthic restoration. An index of ecological integrity could also be adopted into future studies, as they are able to quantify the quality/health of an environment in order to aid in forming management decisions regarding environmental conditions by integrating physical habitat structure with benthic communities [62,63]. Borjaa et al. found that if the same ecological basis was used, different indices could produce a level of agreement. This means that different indices produce comparable results when used in the same environment.
Figure 15.
Diagram of environmental dredging and its residuals [63]. Dredging machinery is used to remove polluted or harmful sediments. It is often not possible, however, to remove 100% of contaminated sediments and adjacent areas may retain thick organic-rich sediments that may slide into the dredge hole after dredging is completed, re-contaminating the sediments.
Figure 15.
Diagram of environmental dredging and its residuals [63]. Dredging machinery is used to remove polluted or harmful sediments. It is often not possible, however, to remove 100% of contaminated sediments and adjacent areas may retain thick organic-rich sediments that may slide into the dredge hole after dredging is completed, re-contaminating the sediments.
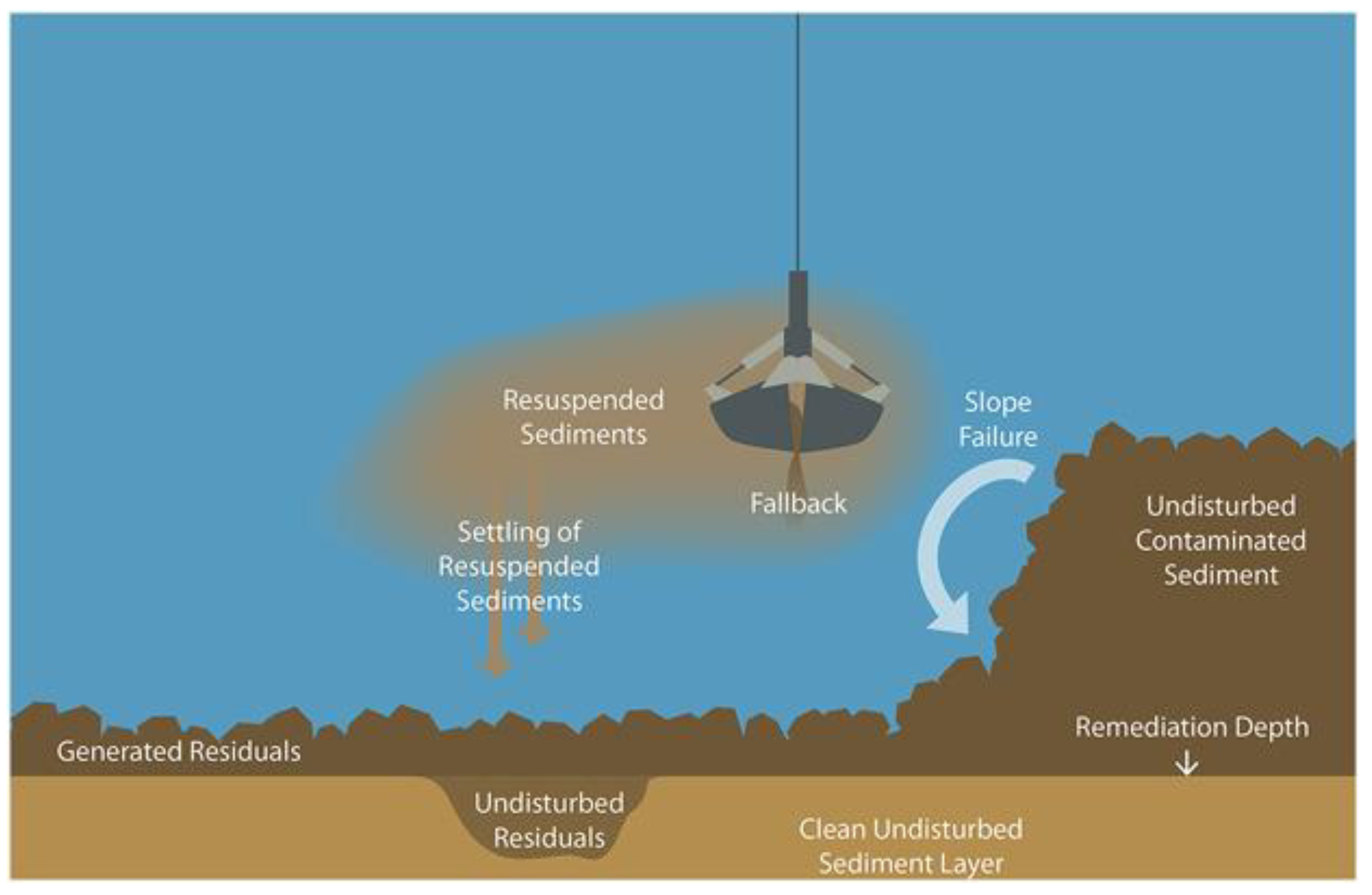
There was no shift in the benthic mollusk community found due to dredging, which could be due to mollusks tolerance of degraded sediment conditions [23,24,25,26]. This study focused on the benthic mollusk communities at the bottom of the dredge line. There could be significant changes found in adjacent communities, or if muck was removed more thoroughly by environmental dredging. There is still much to be learned about the causal relationships relating dredging processes to potential impacts [65]. One study highlighted the “4 Rs of Environmental Dredging”: Resuspension, Release, Residuals, and Risk [66]. Resuspension of polluted sediments can cause them to spread and contaminate neighboring areas. The removal of surface sediments, and the resuspension of sediments (Figure 15) can release deeply sequestered muck contaminants into the water column [65]. Currently, one of the top objectives of environmental dredging is to dredge with enough accuracy that deeper, clean sediments are not impacted or removed [64,67]. However, this approach inevitably leaves behind a residual layer of contaminated/polluted sediment. Additionally, muck from the top of the dredge slope may slide down and join the residual layer (Figure 15), potentially re-establishing high organic content and preventing significant shifts in benthic faunal communities. The aforementioned sloughing of the organic-rich sediments to the bottom of the dredge pit could create a scenario in which the downslope environment experiences little to no change following dredging. dredging deeper and completely removing the residual layer, on the other hand, could result in cleaner, less organic sediments that remain such for a longer period of time following dredging. This could potentially support a more abundant and diverse benthic community, including sensitive filter- and deposit-feeders. The latter would increase sediment bioturbation, potentially breaking up the aforementioned positive feedback loop of anoxic, organic-rich, sediment accumulation.
5. Conclusions
Benthic mollusk abundance, biodiversity, and species richness have inverse logarithmic relationships with sediment organic content. This could be due to hypoxic and/or anoxic conditions in boundary bottom water [29]. However, the removal of some sediments with high concentrations of fine-grained organic matter by dredging did not substantially increase the biodiversity, species richness, and abundance of benthic mollusks. It may be that the high tolerance of many mollusks to stressful environmental conditions allows mollusks to persist under both types of sediment conditions. Alternatively, it may take benthic communities a longer period of time to adjust to new sediment conditions.
Author Contributions
Conceptualization, R.H.S. and K.B.J.; methodology, R.H.S and K.B.J.; validation, K.B.J.; formal analysis, R.H.S.; investigation, R.H.S.; resources, K.B.J.; data curation, R.H.S.; writing—original draft preparation, R.H.S.; writing—review and editing, R.H.S and K.B.J.; visualization, R.H.S.; supervision, K.B.J.; project administration, K.B.J.; funding acquisition, K.B.J. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Brevard County Natural Resources Department and the Citizens of Brevard County through the Save our Indian River Lagoon Project Fund.
Data Availability Statement
All data can be accessed at this link.
Acknowledgments
We would like to thank the citizens of Brevard County for funding this project with the Save our Indian River Lagoon Project Fund, the Brevard County Natural Resources Department, and the members of the Johnson Lab at the Florida Institute of Technology for their assistance.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Appendix A
Table A1.
2-way ANOVA P-Values for each individual species and overall for each season and location.
Table A1.
2-way ANOVA P-Values for each individual species and overall for each season and location.
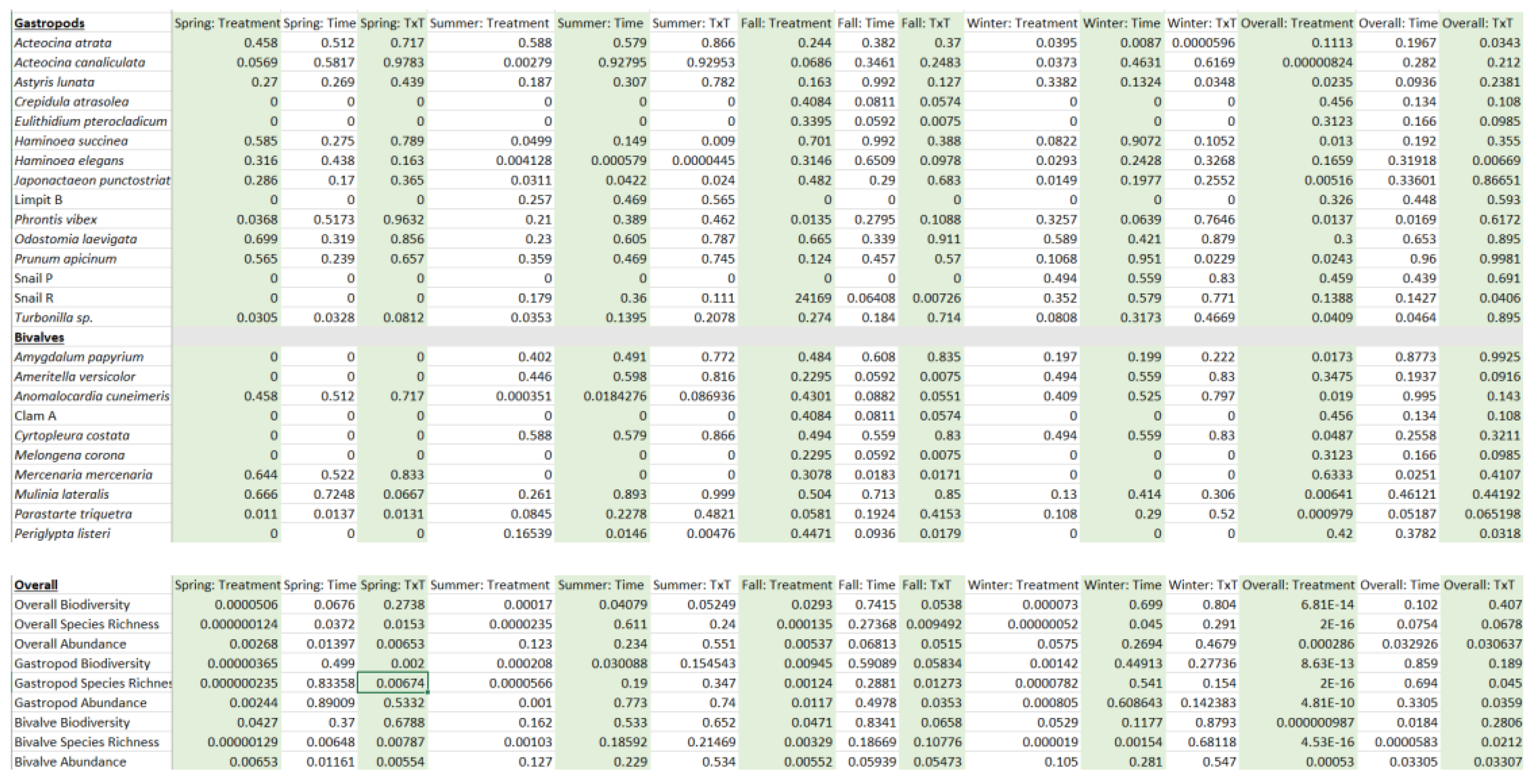
Table A2.
GPS Coordinates for control and treatment sampling stations (see Figure 1).
Table A2.
GPS Coordinates for control and treatment sampling stations (see Figure 1).
Appendix B
Figure A1.
2-way ANOVA figure for Acteocina atrata Abundance vs Treatment in the winter.
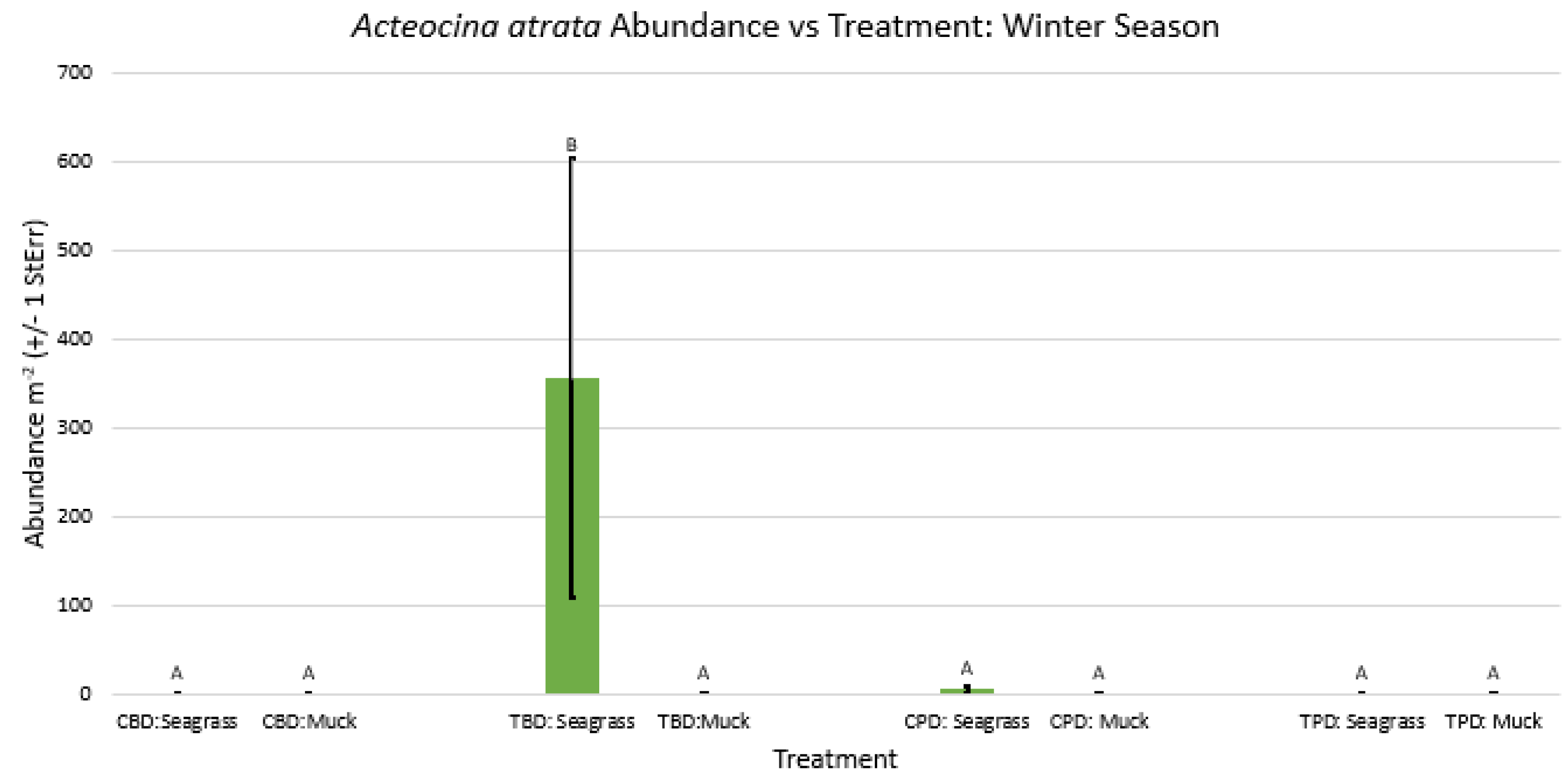
Figure A2.
2-way ANOVA figure for Acteocina atrata Abundance vs Treatment overall.
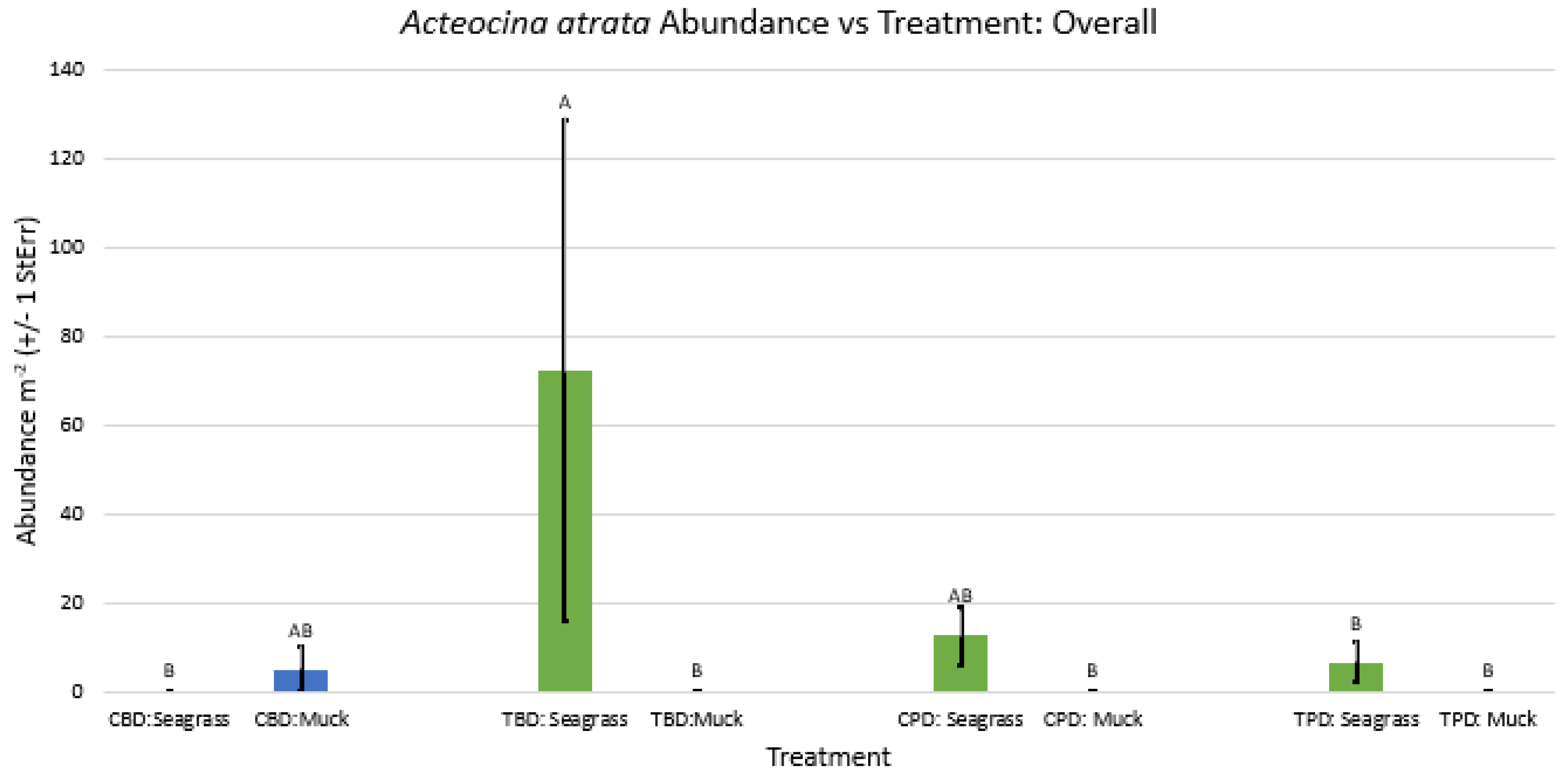
Figure A3.
2-way ANOVA figure for Ameritella versicolor Abundance vs Treatment in the fall.
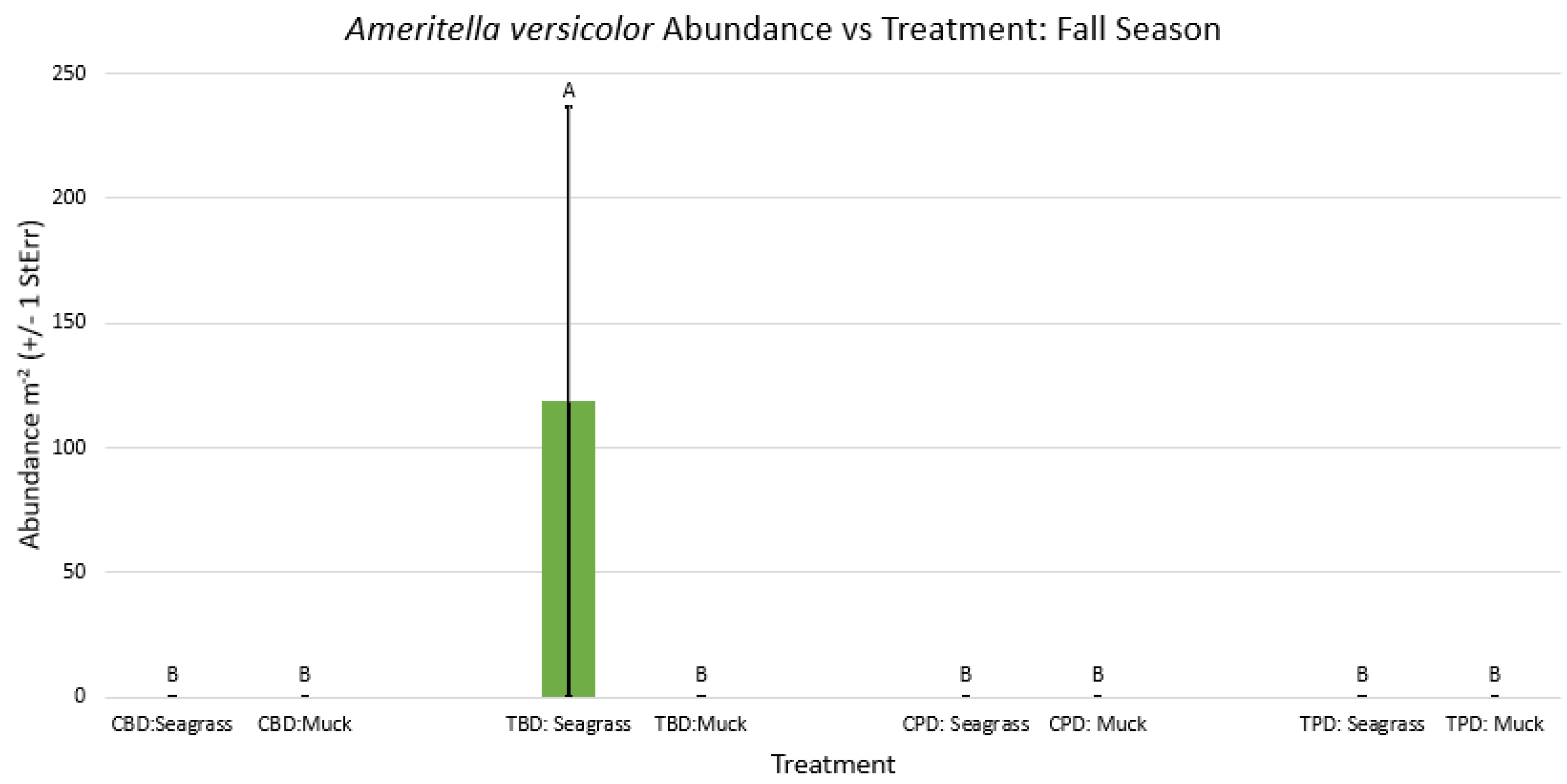
Figure A4.
2-way ANOVA figure for Eulithidium pterocladicum Abundance vs Treatment in the fall.

Figure A5.
2-way ANOVA figure for Melongena corona Abundance vs Treatment in the fall.
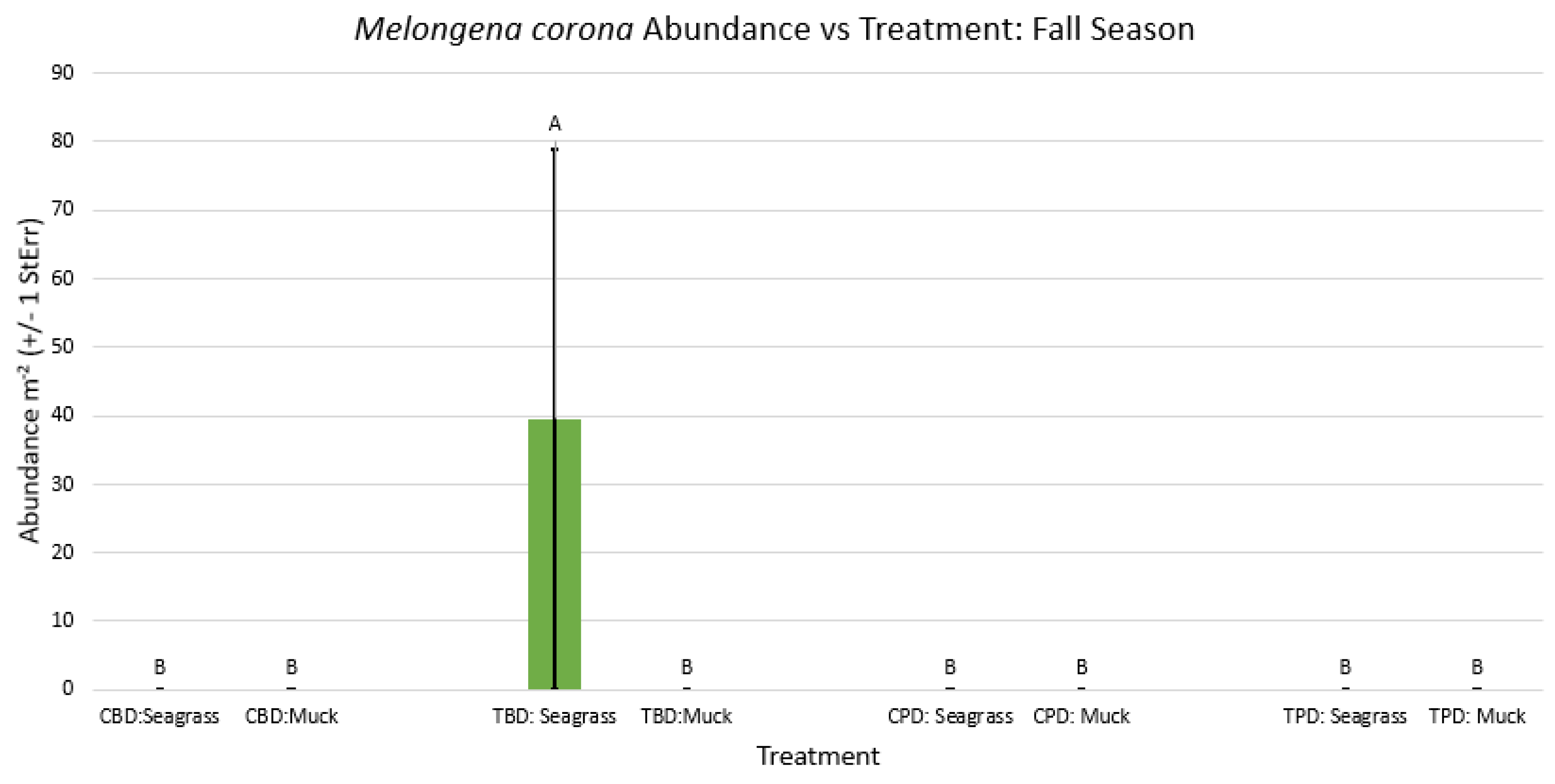
Figure A6.
2-way ANOVA figure for Mercenaria mercenaria Abundance vs Treatment in the fall.
Figure A7.
2-way ANOVA figure for Periglypta listeri Abundance vs Treatment in the summer.
Figure A8.
2-way ANOVA figure for Periglypta listeri Abundance vs Treatment in the fall.
Figure A9.
2-way ANOVA figure for Prunum acicinum Abundance vs Treatment in the winter.
Figure A10.
2-way ANOVA figure for Ameritella versicolor Abundance vs Treatment in the fall.
Figure A11.
2-way ANOVA figure for unidentified Clam A Abundance vs Treatment in the fall.
Figure A12.
2-way ANOVA figure for Crepidula atrasolea Abundance vs Treatment in the fall.
Figure A13.
2-way ANOVA figure for Unidentified Snail R Abundance vs Treatment in the Summer.
Figure A14.
2-way ANOVA figure for Unidentified Snail R Abundance vs Treatment in the fall.
Figure A15.
2-way ANOVA Figure Afor Unidentified Snail R Abundance vs Treatment in all seasons.
Figure A16.
2-way ANOVA Figure Afor Haminoea elegans Abundance vs Treatment in the summer.
Figure A17.
2-way ANOVA Figure Afor Turbonilla sp. Abundance vs Treatment in the spring.
Figure A18.
2-way ANOVA Figure Afor Turbonilla sp. Abundance vs Treatment in the summer.
Figure A19.
2-way ANOVA Figure Afor Anomalacardia cuneimeris Abundance vs Treatment in the summer.
Figure A20.
2-way ANOVA Figure Afor Anomalacardia cuneimeris Abundance vs Treatment in the fall.
Figure A21.
2-way ANOVA Figure Afor Anomalacardia cuneimeris Abundance vs Treatment in all seasons.
Figure A22.
2-way ANOVA Figure Afor Acteocina canaliculata Abundance vs Treatment in all seasons.
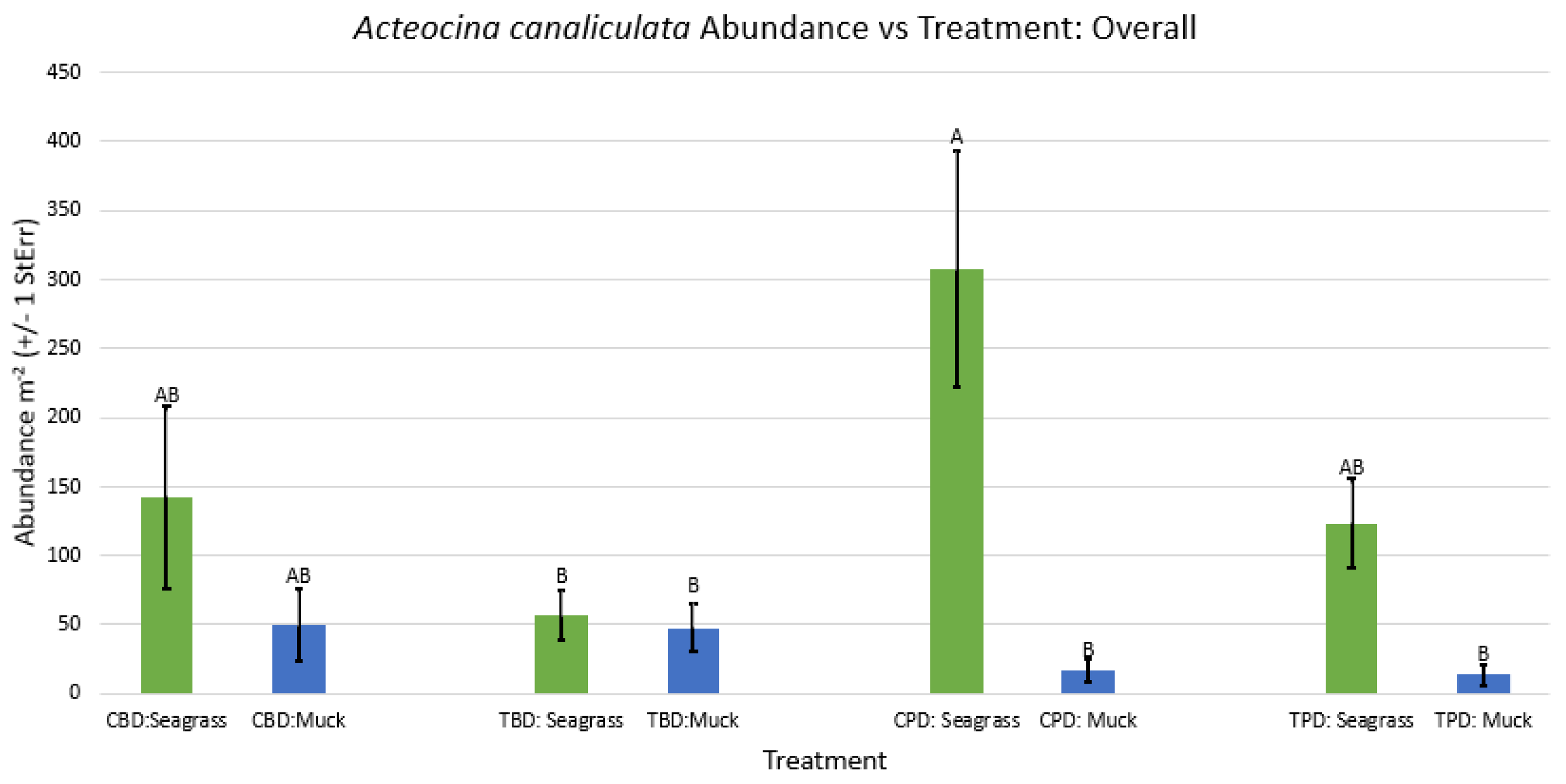
Figure A23.
2-way ANOVA Figure Afor Phrontis vibex Abundance vs Treatment in the fall.
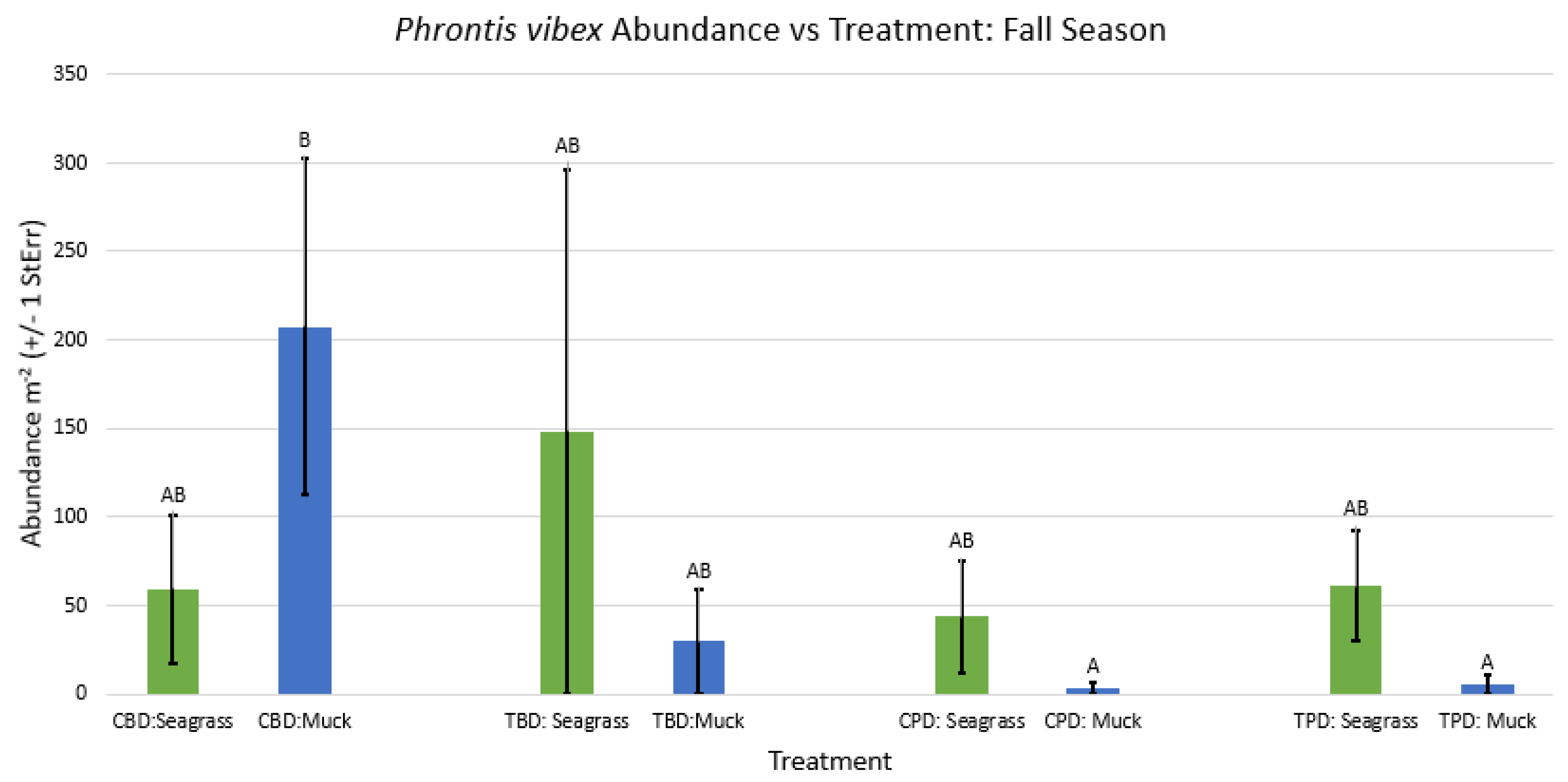
Figure A24.
2-way ANOVA figure for Mulinia lateralis Abundance vs Treatment in all seasons.
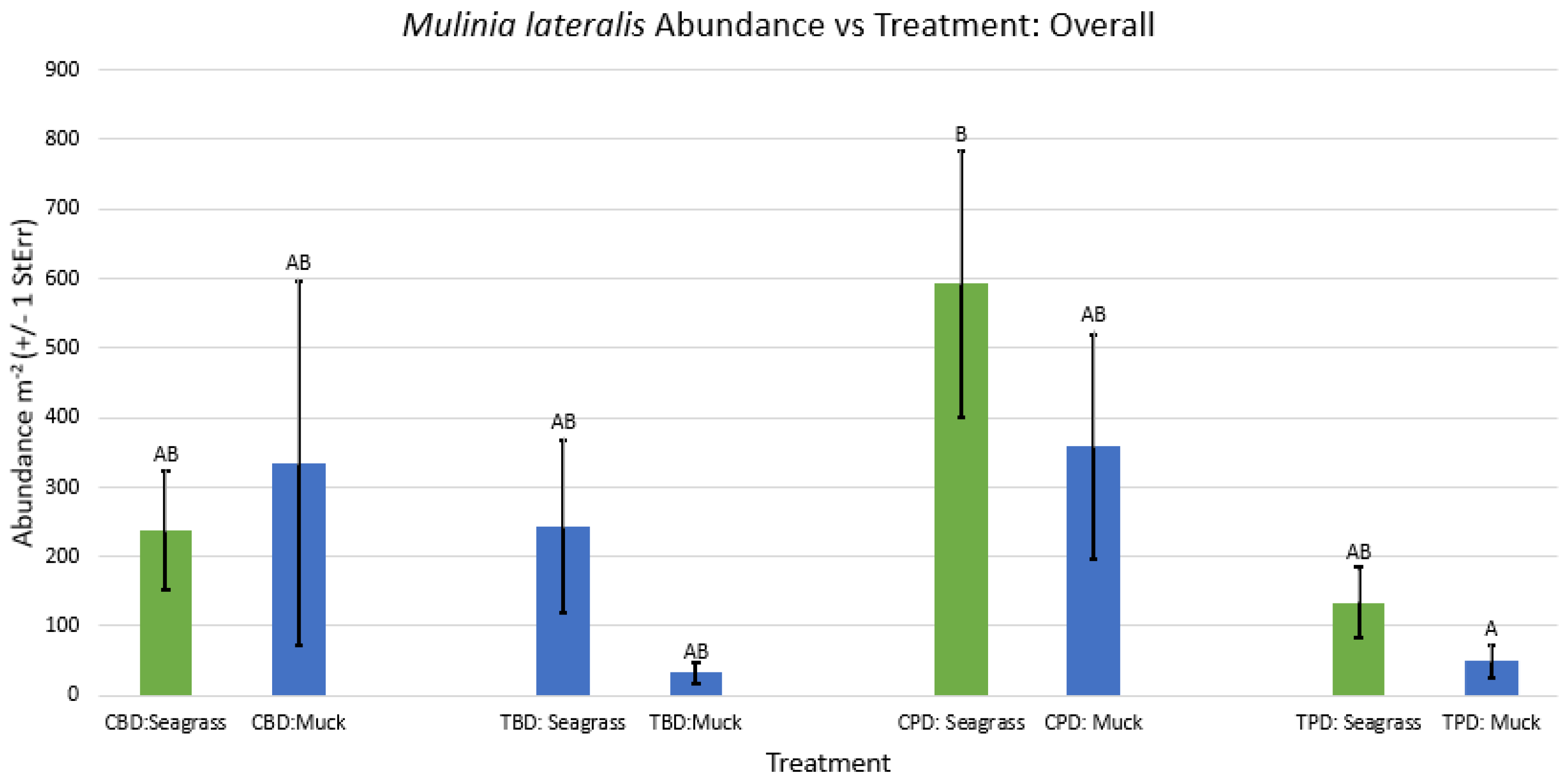
Figure A25.
2-way ANOVA figure for Parastarte triquetra Abundance vs Treatment in the spring.
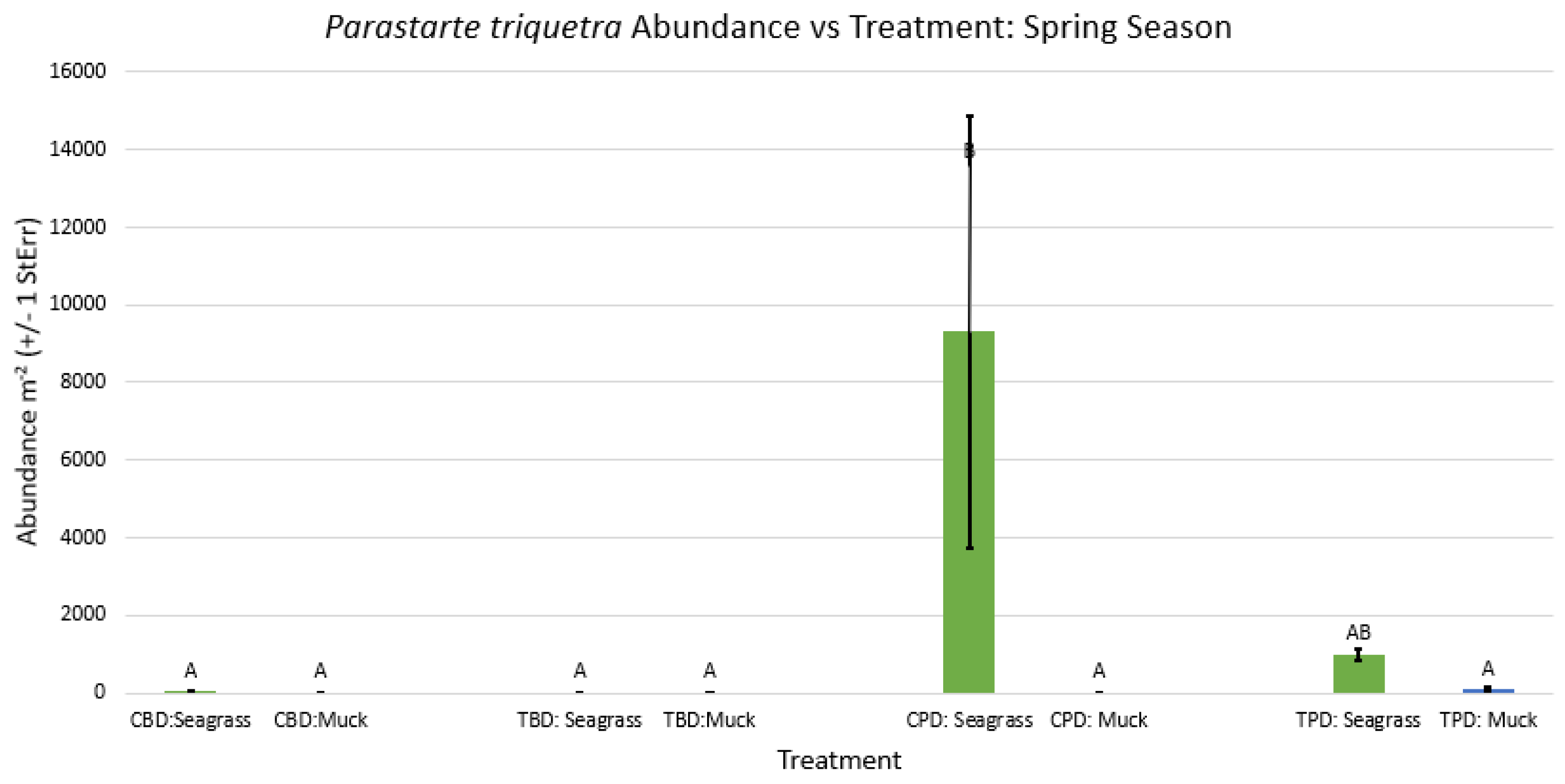
Figure A26.
2-way ANOVA figure for Parastarte triquetra Abundance vs Treatment in all seasons.
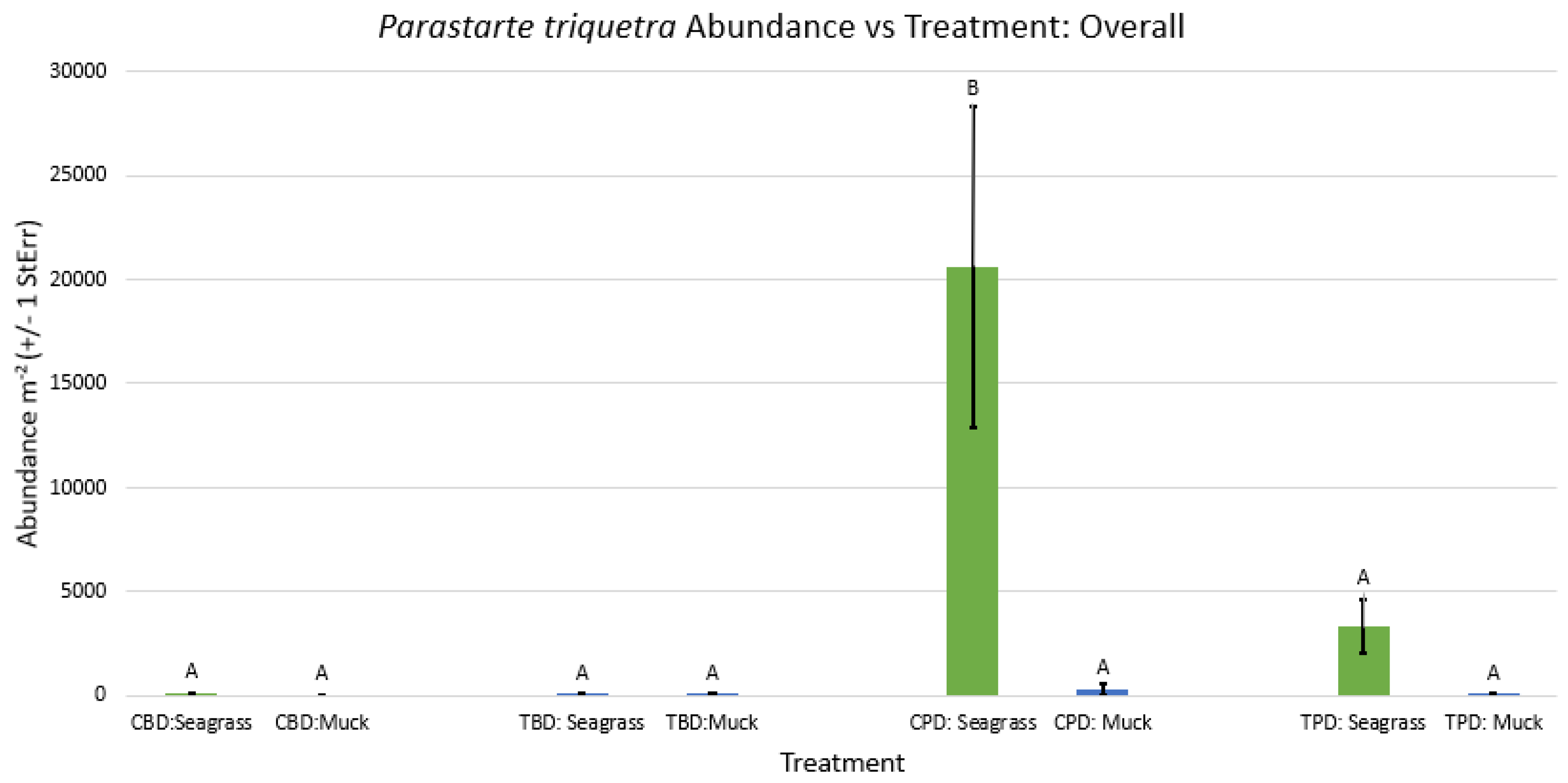
Figure A27.
2-way ANOVA figure for Japonactaeon punctostriatus Abundance vs Treatment in the summer.
Figure A27.
2-way ANOVA figure for Japonactaeon punctostriatus Abundance vs Treatment in the summer.
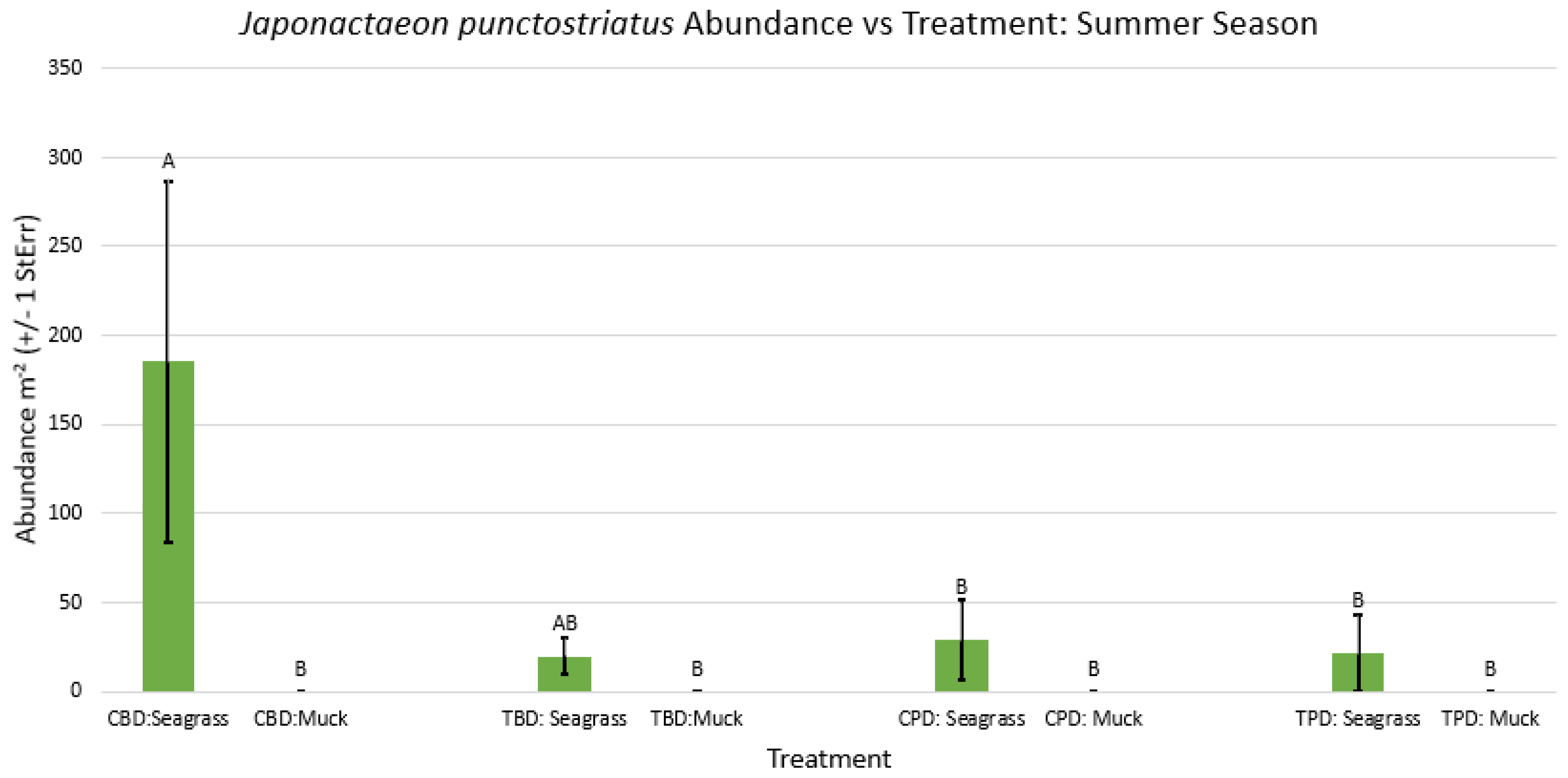
Figure A28.
2-way ANOVA figure for Japonactaeon punctostriatus Abundance vs Treatment in the winter.
Figure A28.
2-way ANOVA figure for Japonactaeon punctostriatus Abundance vs Treatment in the winter.
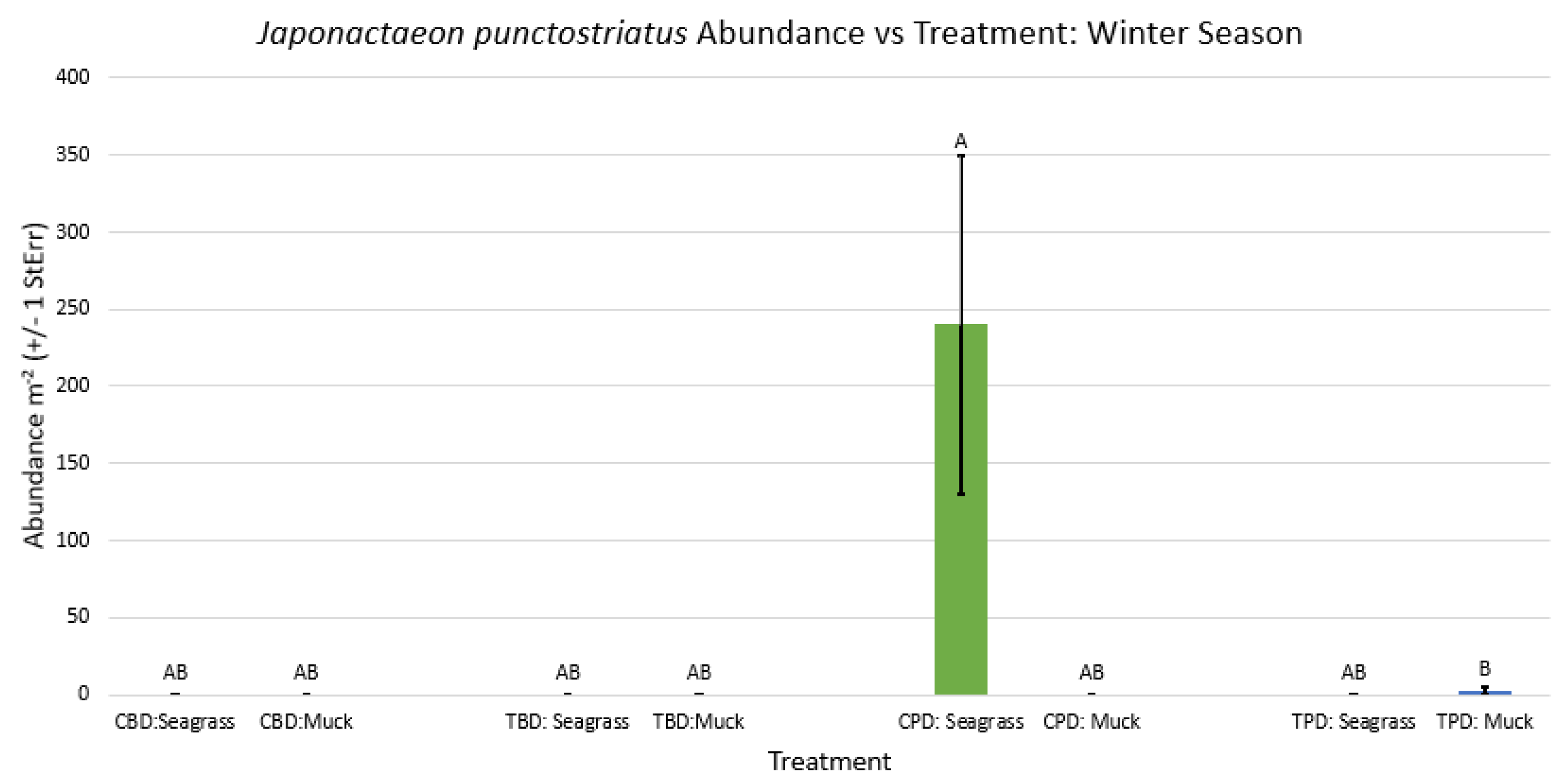
Figure A29.
2-way ANOVA figure for Haminoea succinea Abundance vs Treatment in the summer.
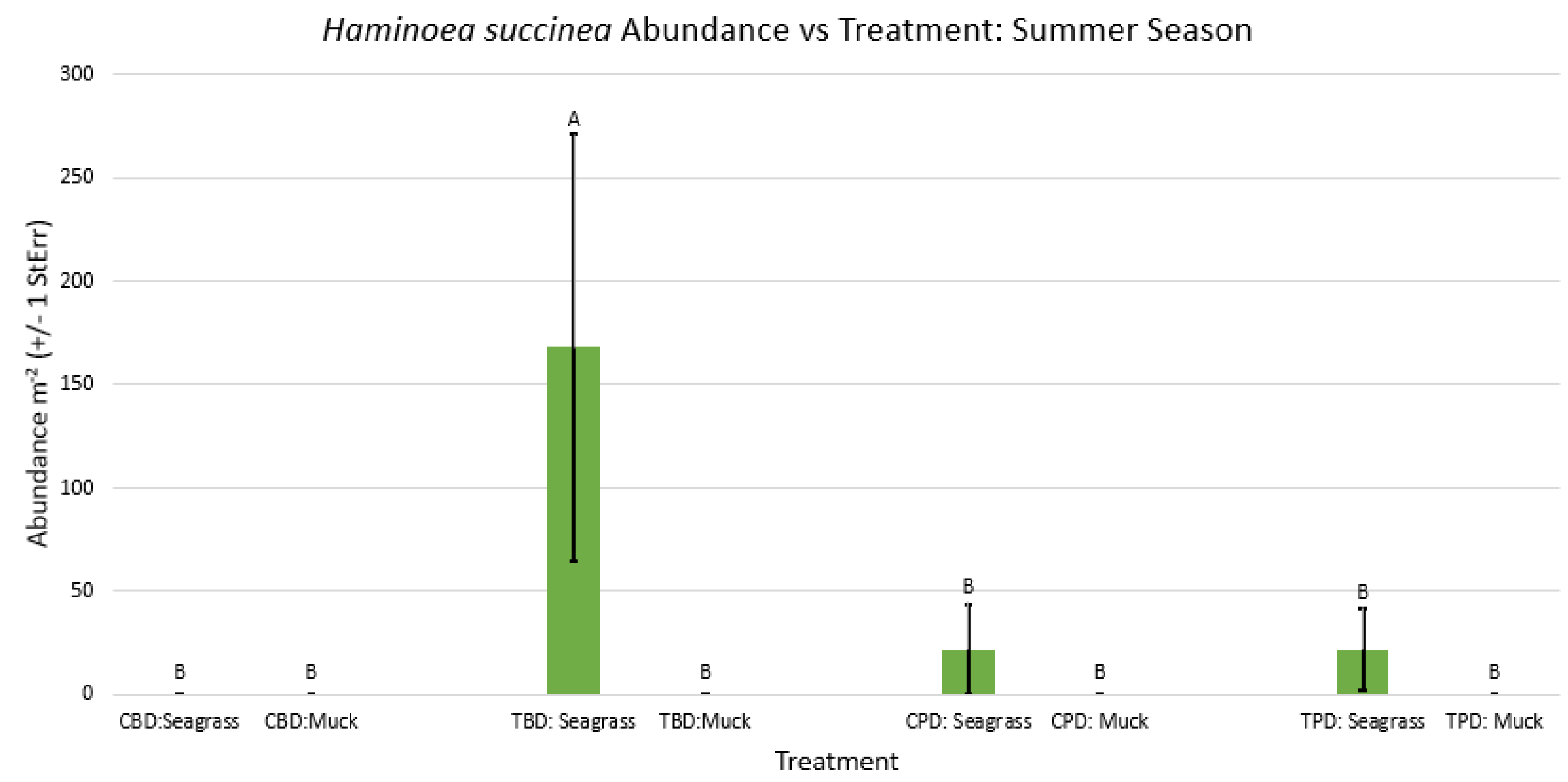
Figure A30.
2-way ANOVA figure for Haminoea succinea Abundance vs Treatment in all seasons.
Figure A31.
2-way ANOVA figure for all benthic mollusk species Biodiversity vs Treatment in the spring.
Figure A31.
2-way ANOVA figure for all benthic mollusk species Biodiversity vs Treatment in the spring.
Figure A32.
2-way ANOVA figure for all benthic mollusk species Biodiversity vs Treatment in the summer.
Figure A32.
2-way ANOVA figure for all benthic mollusk species Biodiversity vs Treatment in the summer.
Figure A33.
2-way ANOVA figure for all benthic mollusk species Biodiversity vs Treatment in the fall season at Seagrass and Treatment stations.
Figure A33.
2-way ANOVA figure for all benthic mollusk species Biodiversity vs Treatment in the fall season at Seagrass and Treatment stations.
Figure A34.
2-way ANOVA figure for all benthic mollusk species Biodiversity vs Treatment in the winter.
Figure A34.
2-way ANOVA figure for all benthic mollusk species Biodiversity vs Treatment in the winter.
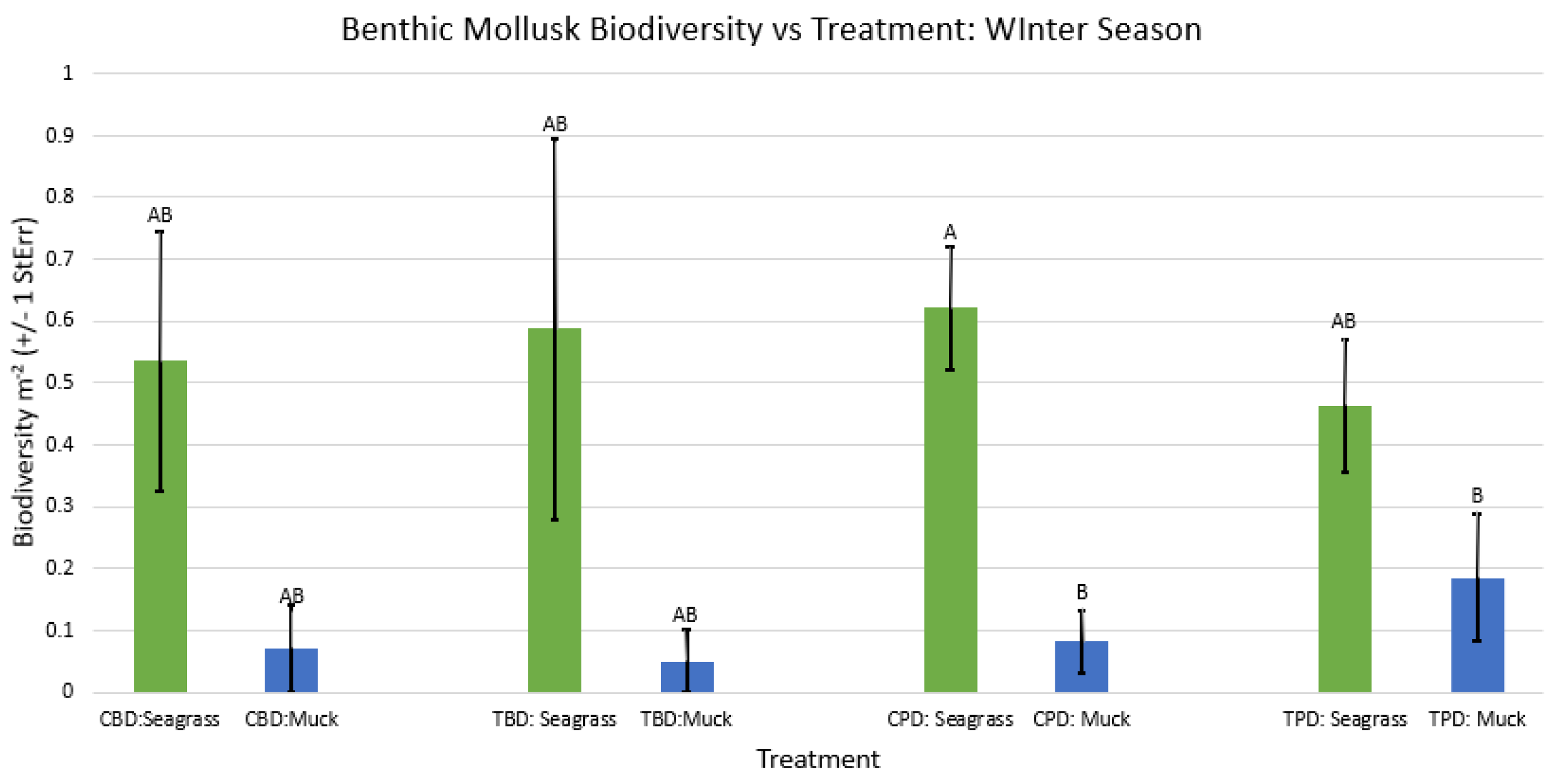
Figure A35.
2-way ANOVA figure for all benthic mollusk species Biodiversity vs Treatment in all seasons.
Figure A35.
2-way ANOVA figure for all benthic mollusk species Biodiversity vs Treatment in all seasons.
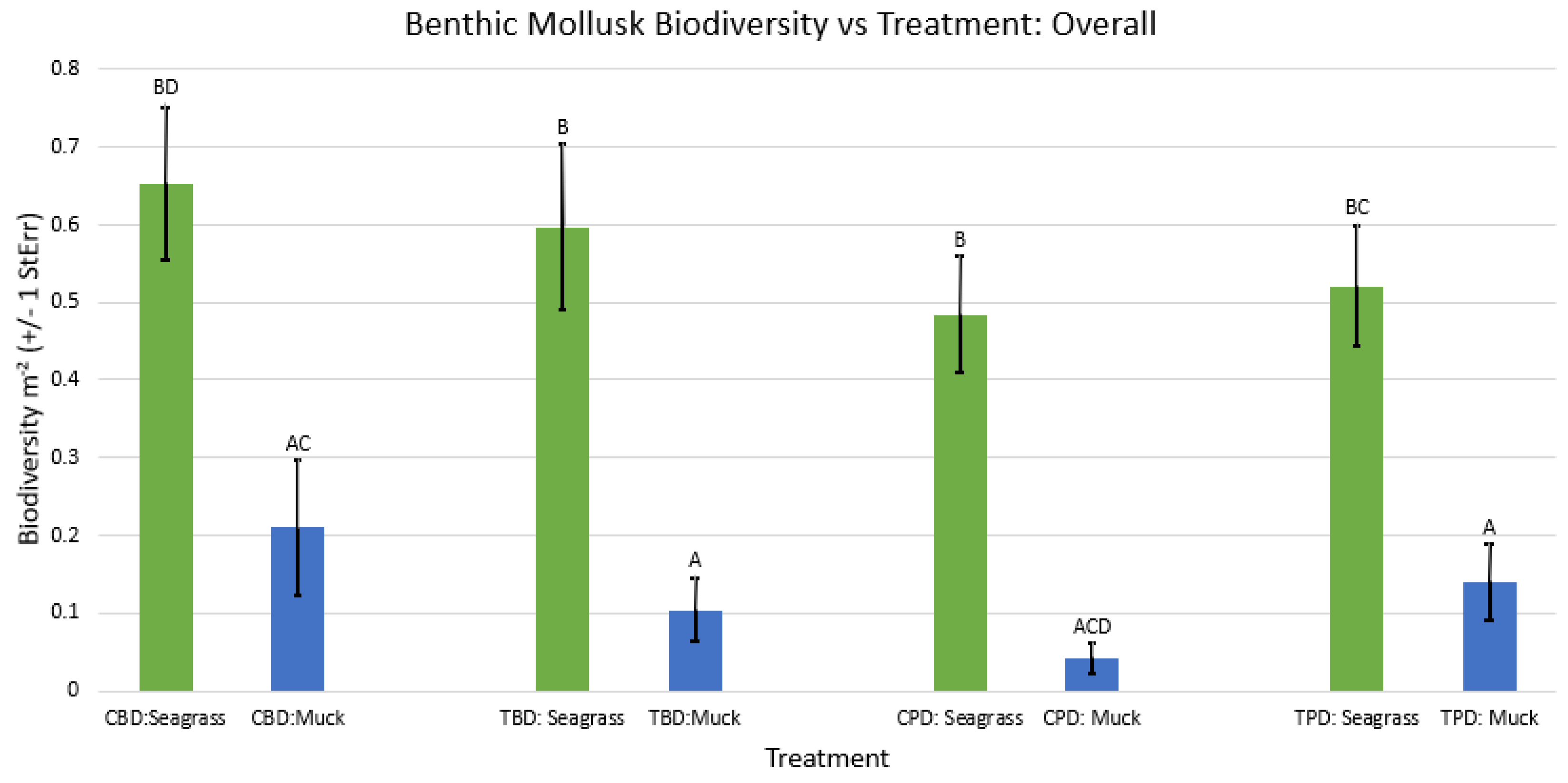
Figure A36.
2-way ANOVA figure for all benthic mollusk species richness vs Treatment in the spring.
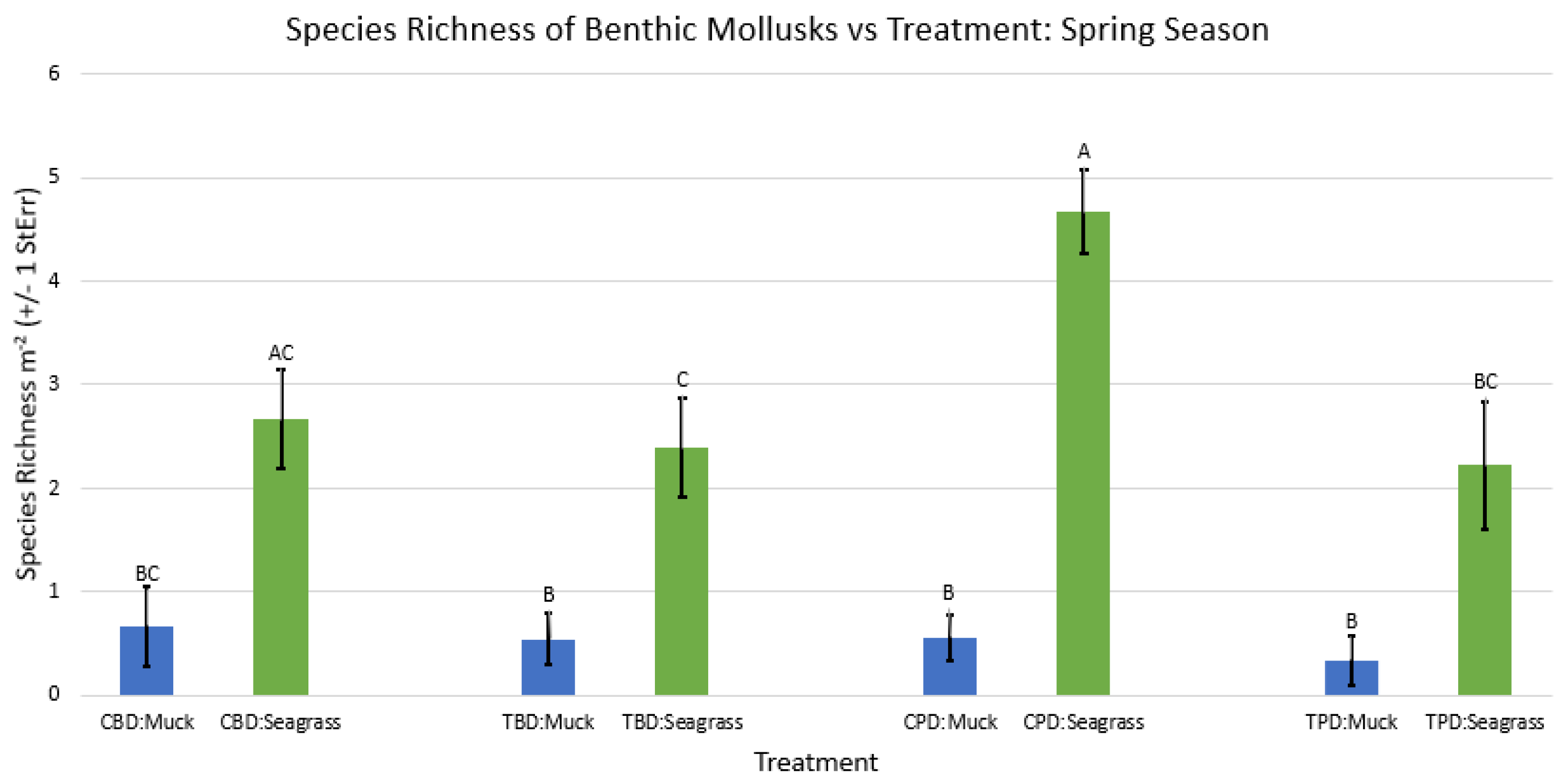
Figure A37.
2-way ANOVA figure for all benthic mollusk species richness vs Treatment in the summer.
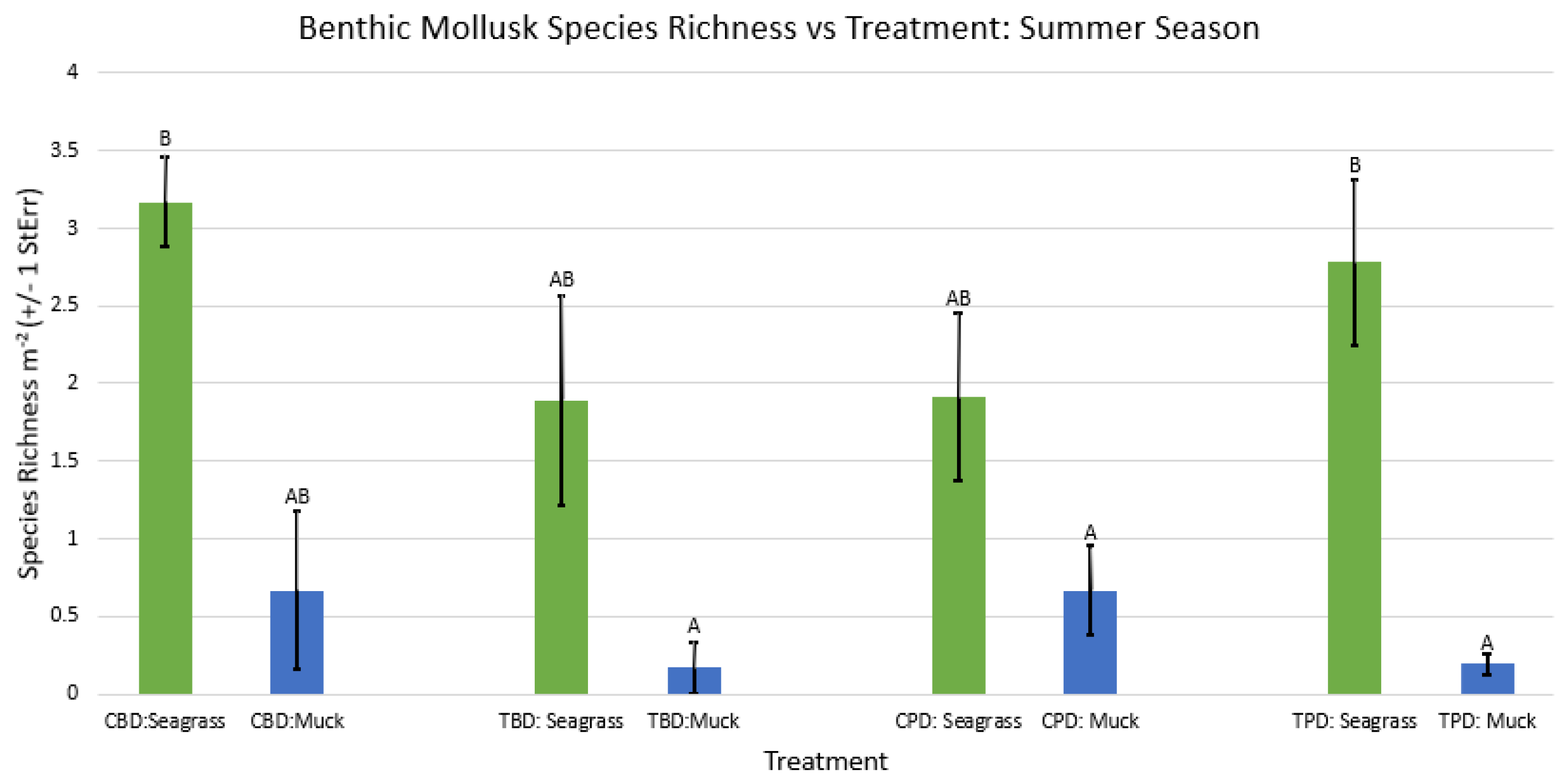
Figure A38.
2-way ANOVA figure for all benthic mollusk species richness vs Treatment in the fall.
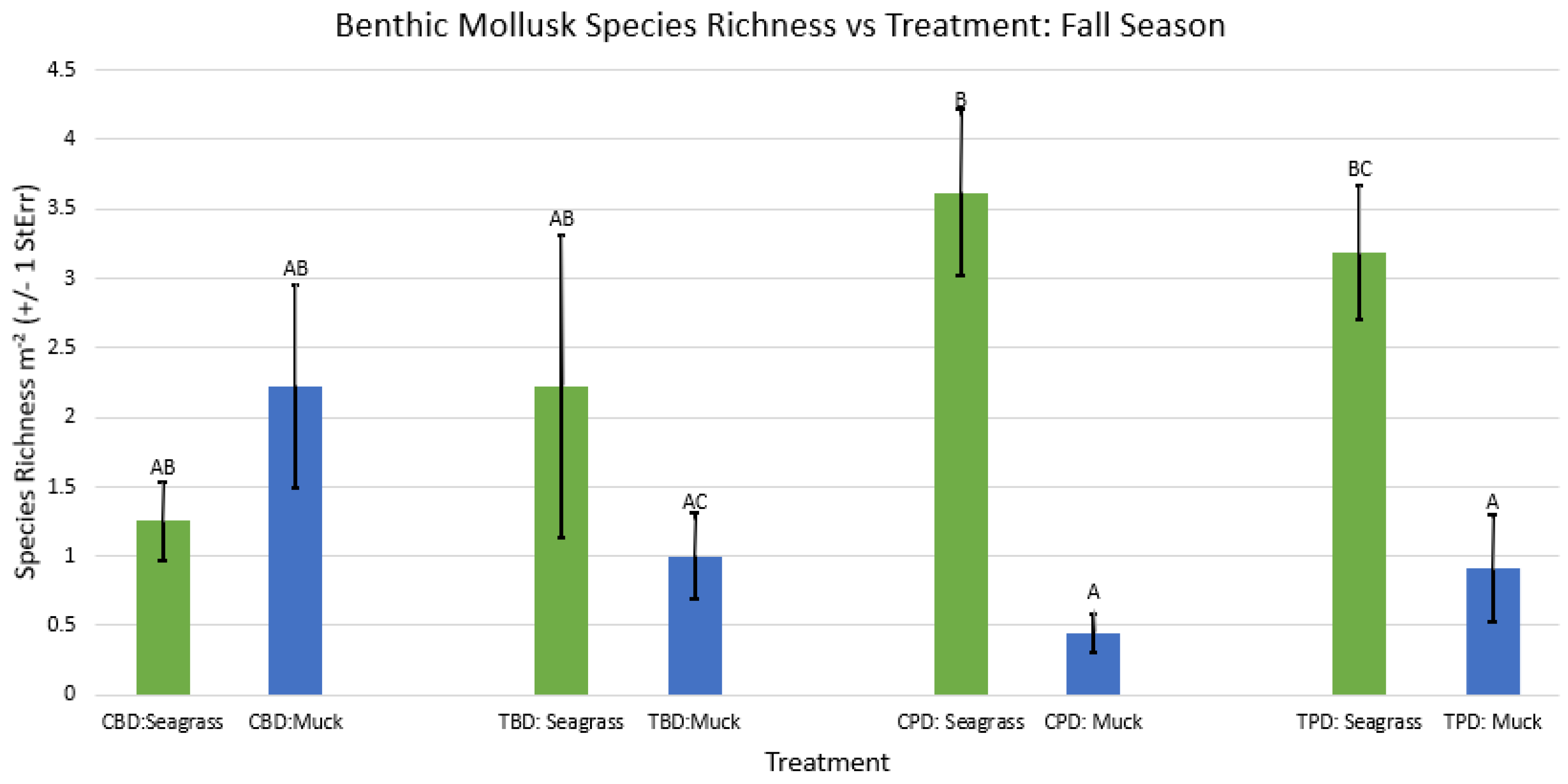
Figure A39.
2-way ANOVA figure for all benthic mollusk species richness vs Treatment in the winter.
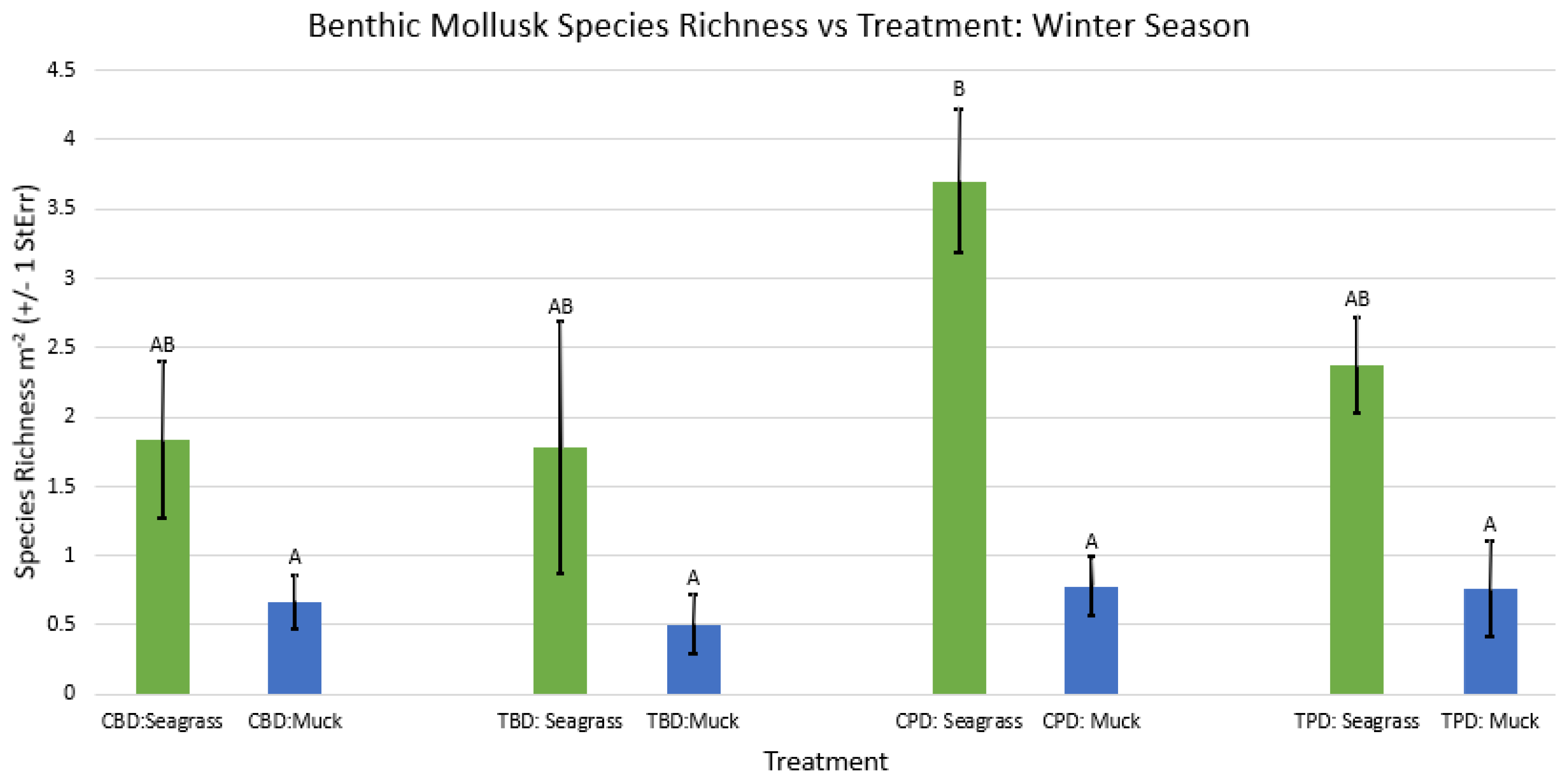
Figure A40.
2-way ANOVA figure for all benthic mollusk species richness vs Treatment in all seasons.
Figure A40.
2-way ANOVA figure for all benthic mollusk species richness vs Treatment in all seasons.
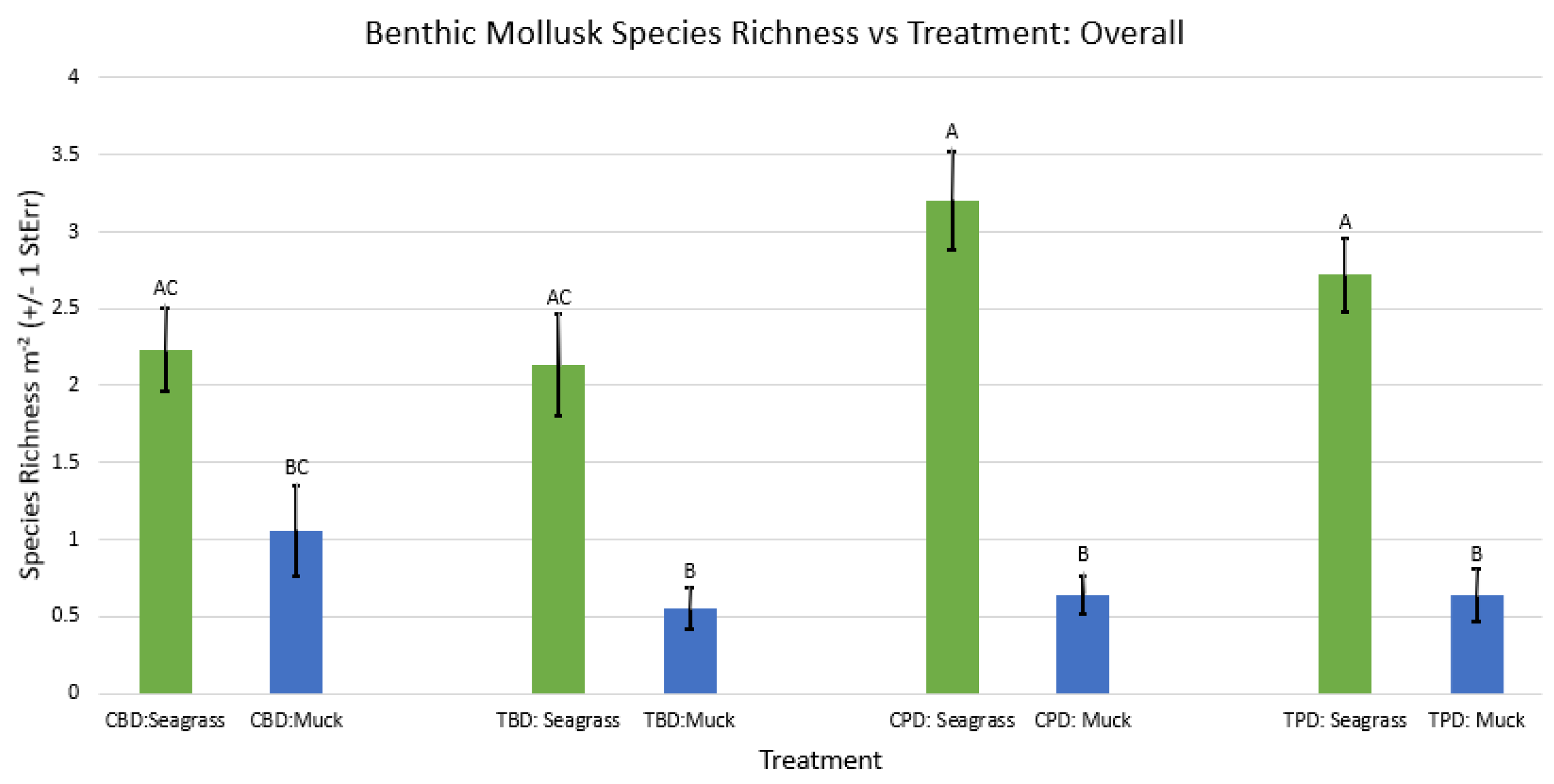
Figure A41.
2-way ANOVA figure for all benthic mollusk abundance vs Treatment in the spring.
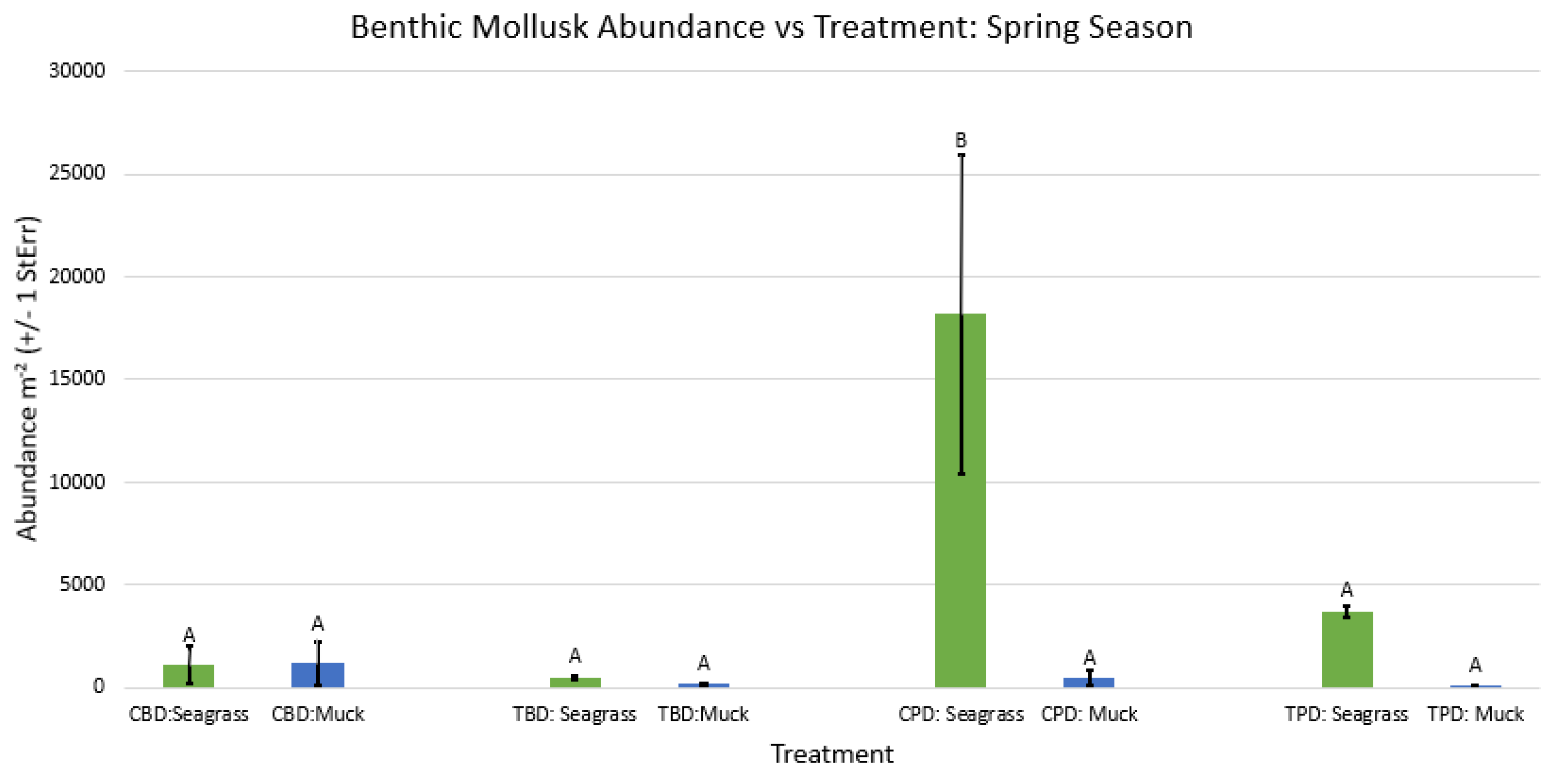
Figure A42.
2-way ANOVA figure for all benthic mollusk abundance vs Treatment in the fall.
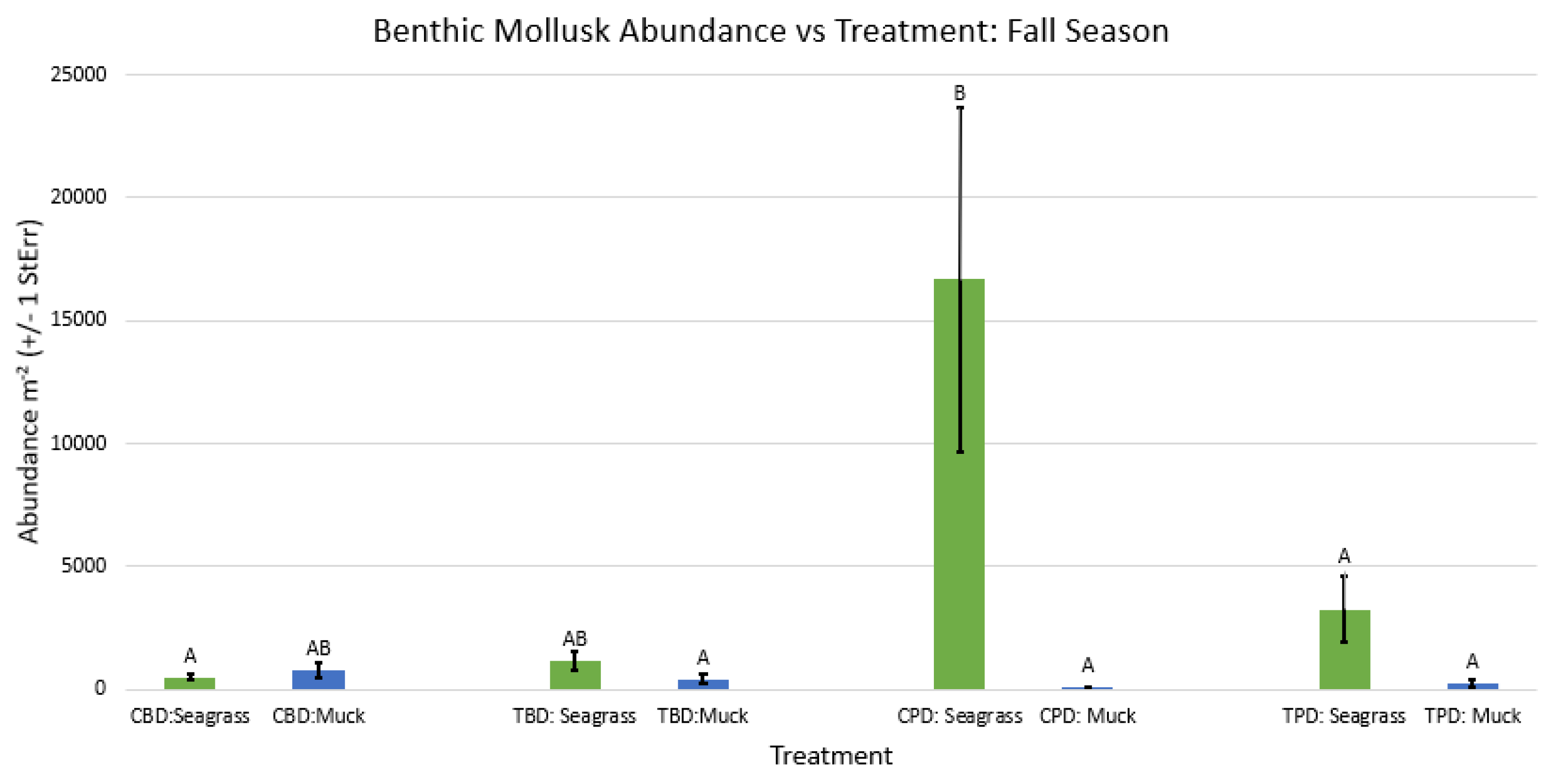
Figure A43.
2-way ANOVA figure for all benthic mollusk abundance vs Treatment in all seasons.
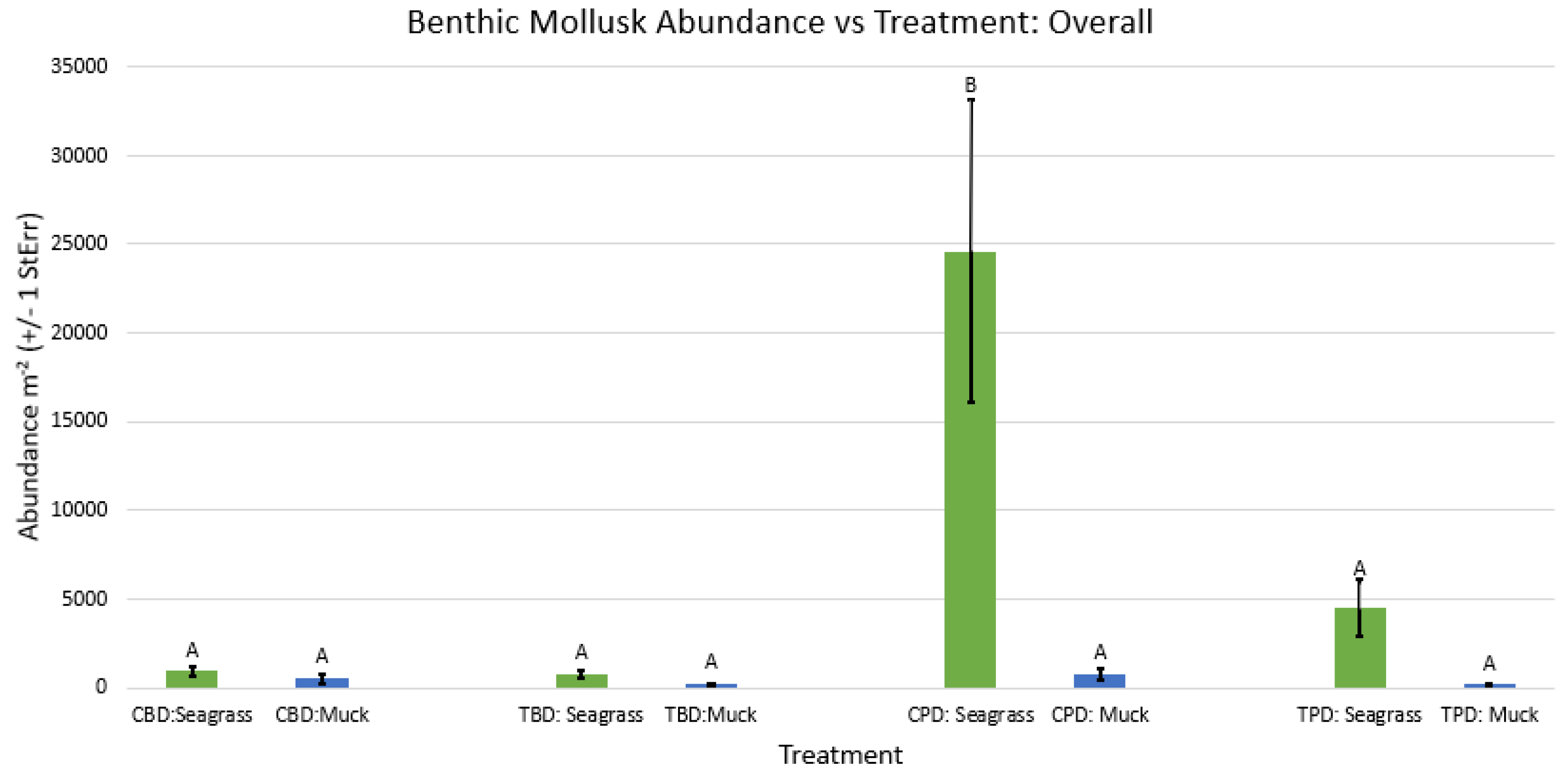
Figure A44.
2-way ANOVA Figure Afor all benthic gastropod biodiversity vs Treatment in the spring.
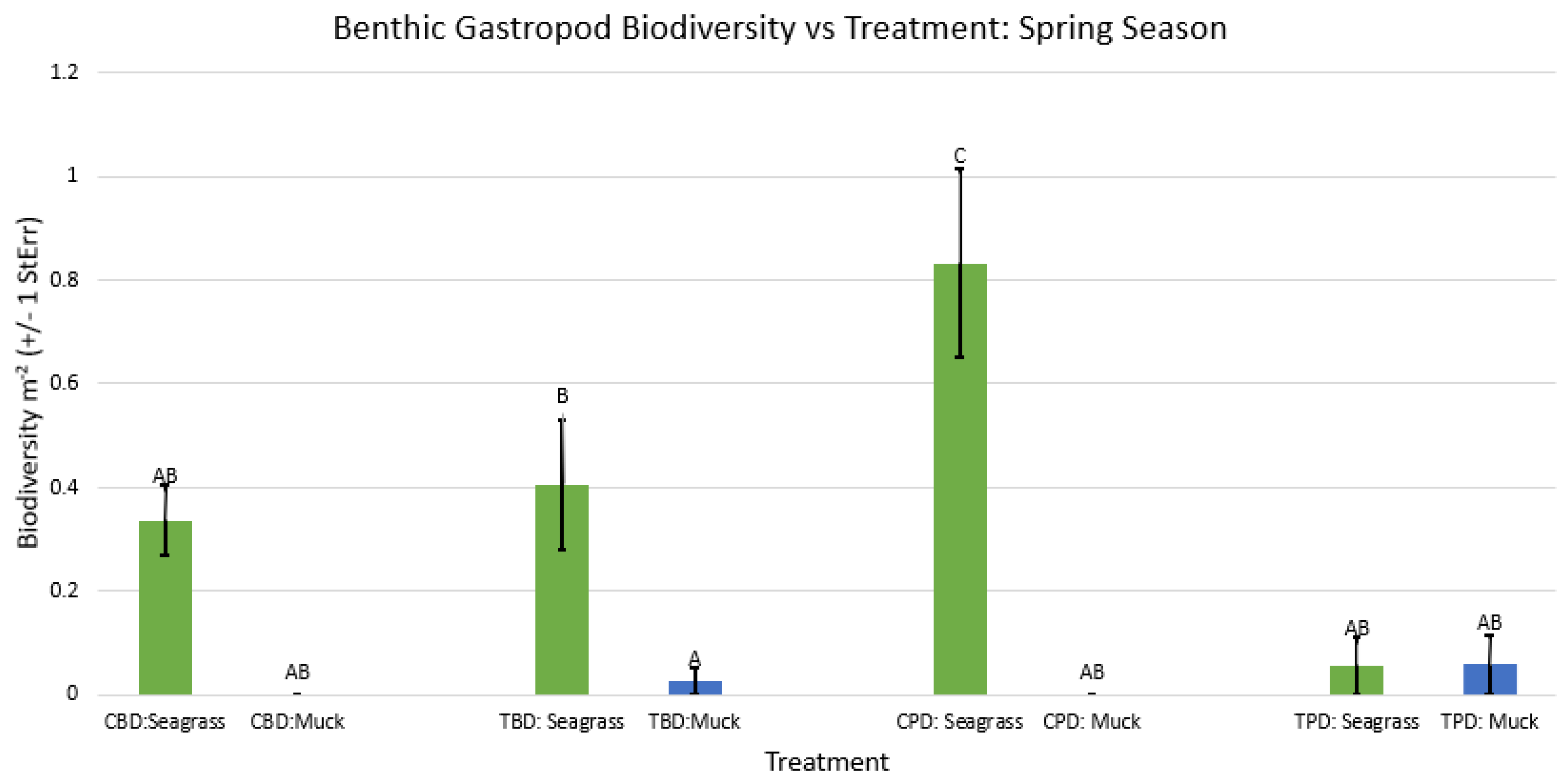
Figure A45.
2-way ANOVA Figure Afor all benthic gastropod biodiversity vs Treatment in the summer.
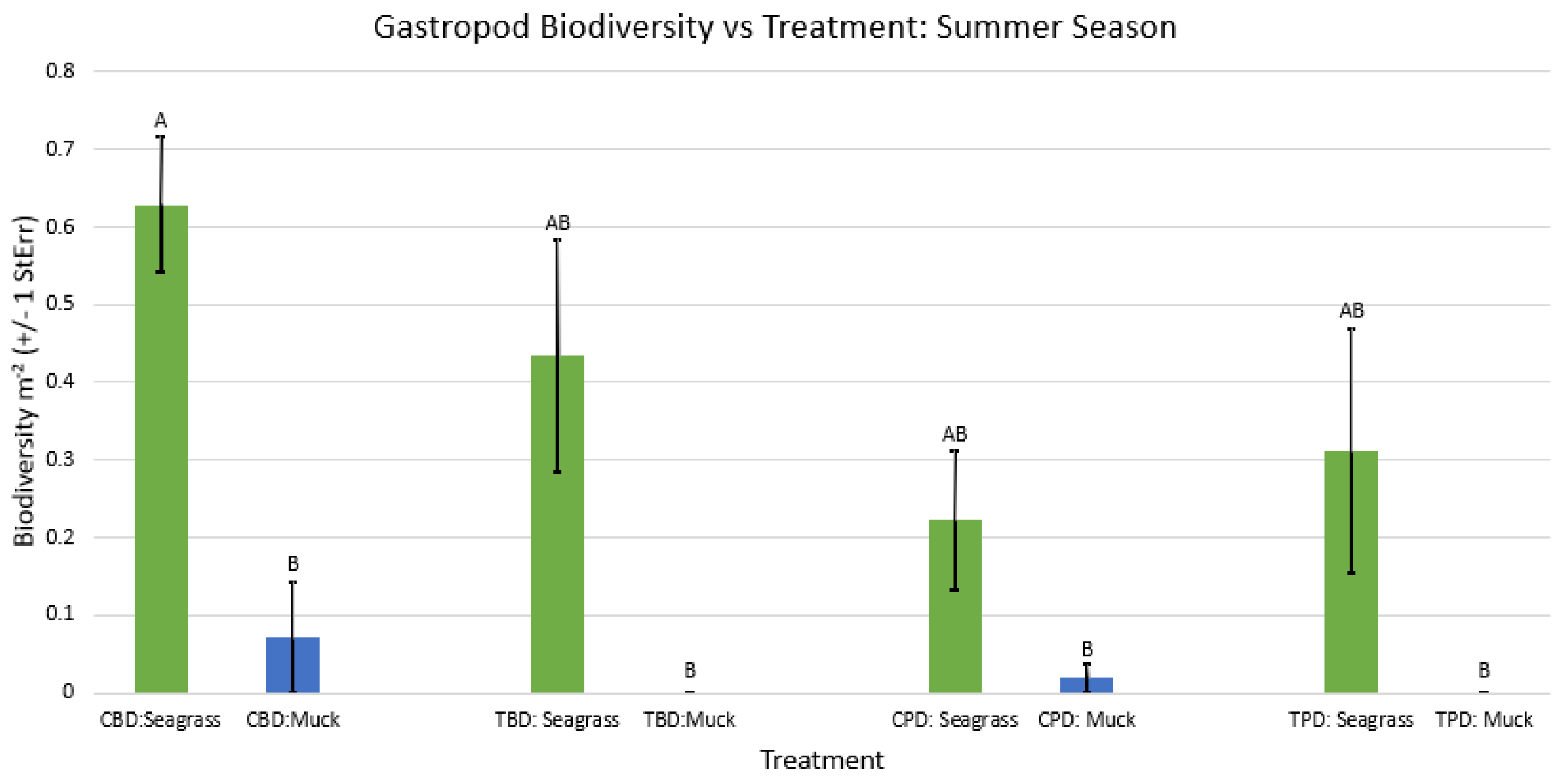
Figure A46.
2-way ANOVA Figure Afor all benthic gastropod biodiversity vs Treatment in the fall.
Figure A47.
2-way ANOVA Figure Afor all benthic gastropod biodiversity vs Treatment in the winter.
Figure A48.
2-way ANOVA Figure Afor all benthic gastropod biodiversity vs Treatment in all seasons.
Figure A49.
2-way ANOVA Figure Afor all benthic gastropod species richness vs Treatment in the spring.
Figure A49.
2-way ANOVA Figure Afor all benthic gastropod species richness vs Treatment in the spring.
Figure A50.
2-way ANOVA Figure Afor all benthic gastropod species richness vs Treatment in the summer.
Figure A50.
2-way ANOVA Figure Afor all benthic gastropod species richness vs Treatment in the summer.
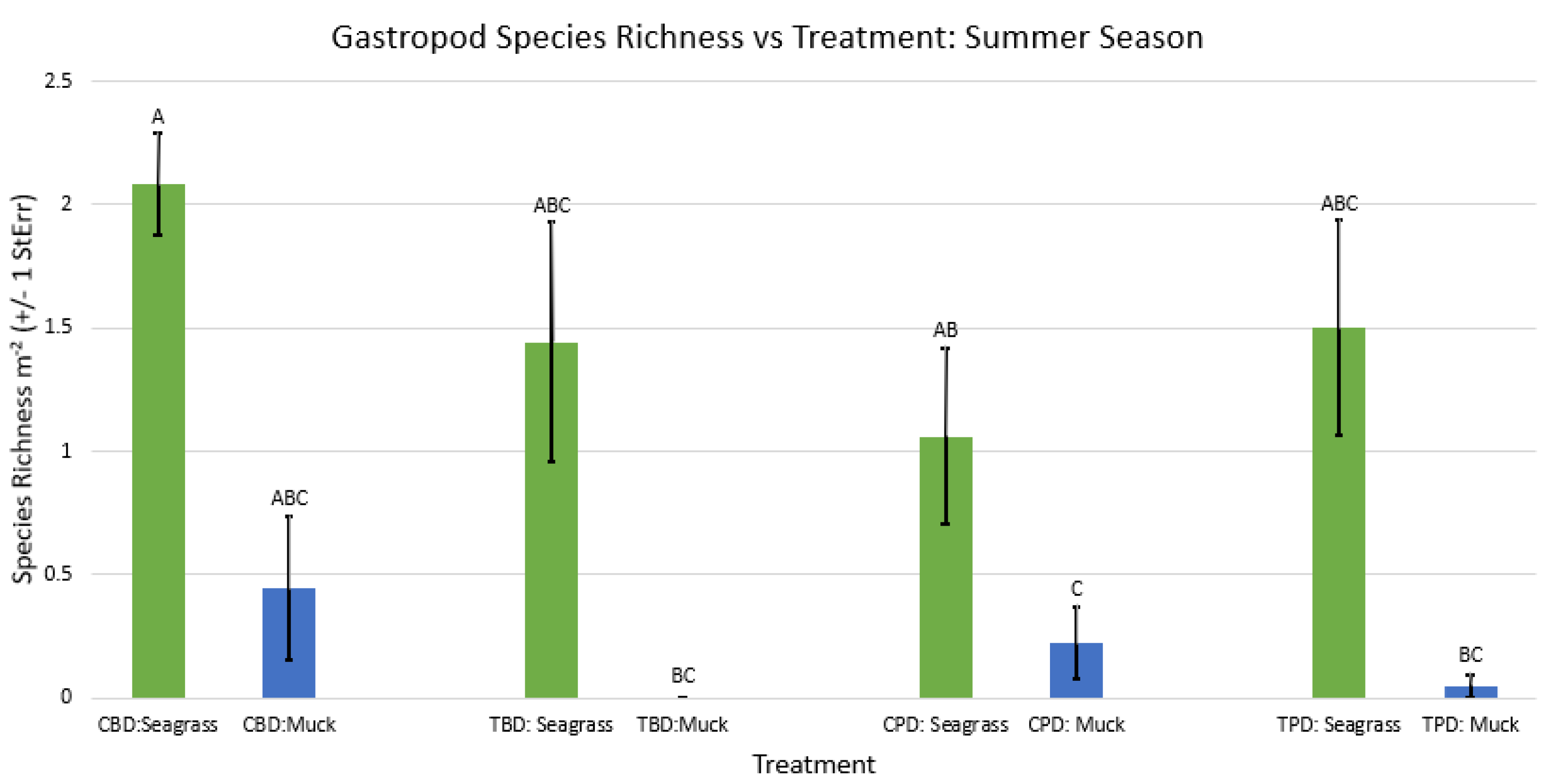
Figure A51.
2-way ANOVA Figure Afor all benthic gastropod species richness vs Treatment in the fall.
Figure A51.
2-way ANOVA Figure Afor all benthic gastropod species richness vs Treatment in the fall.
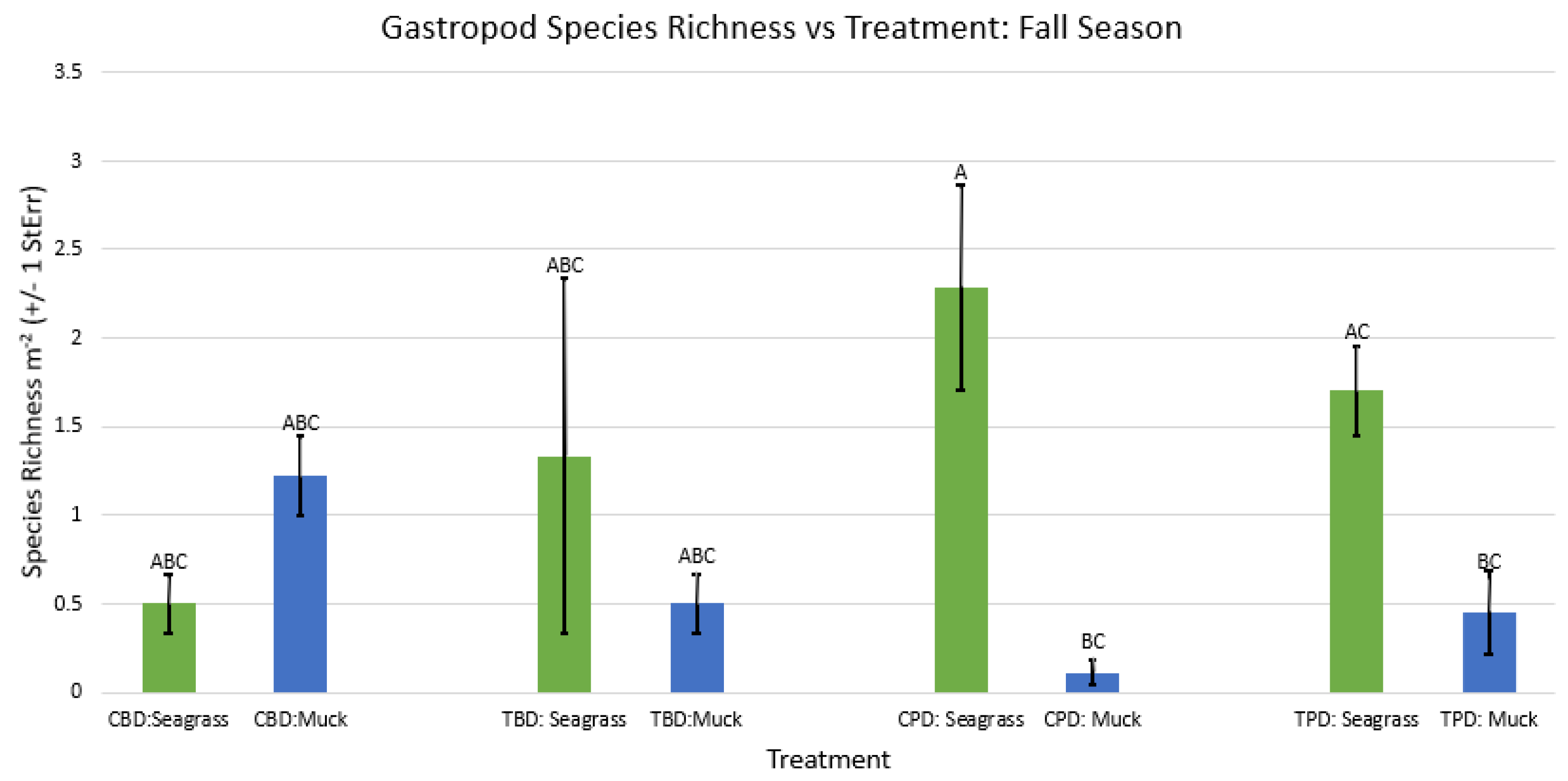
Figure A52.
2-way ANOVA Figure Afor all benthic gastropod species richness vs Treatment in the winter.
Figure A52.
2-way ANOVA Figure Afor all benthic gastropod species richness vs Treatment in the winter.
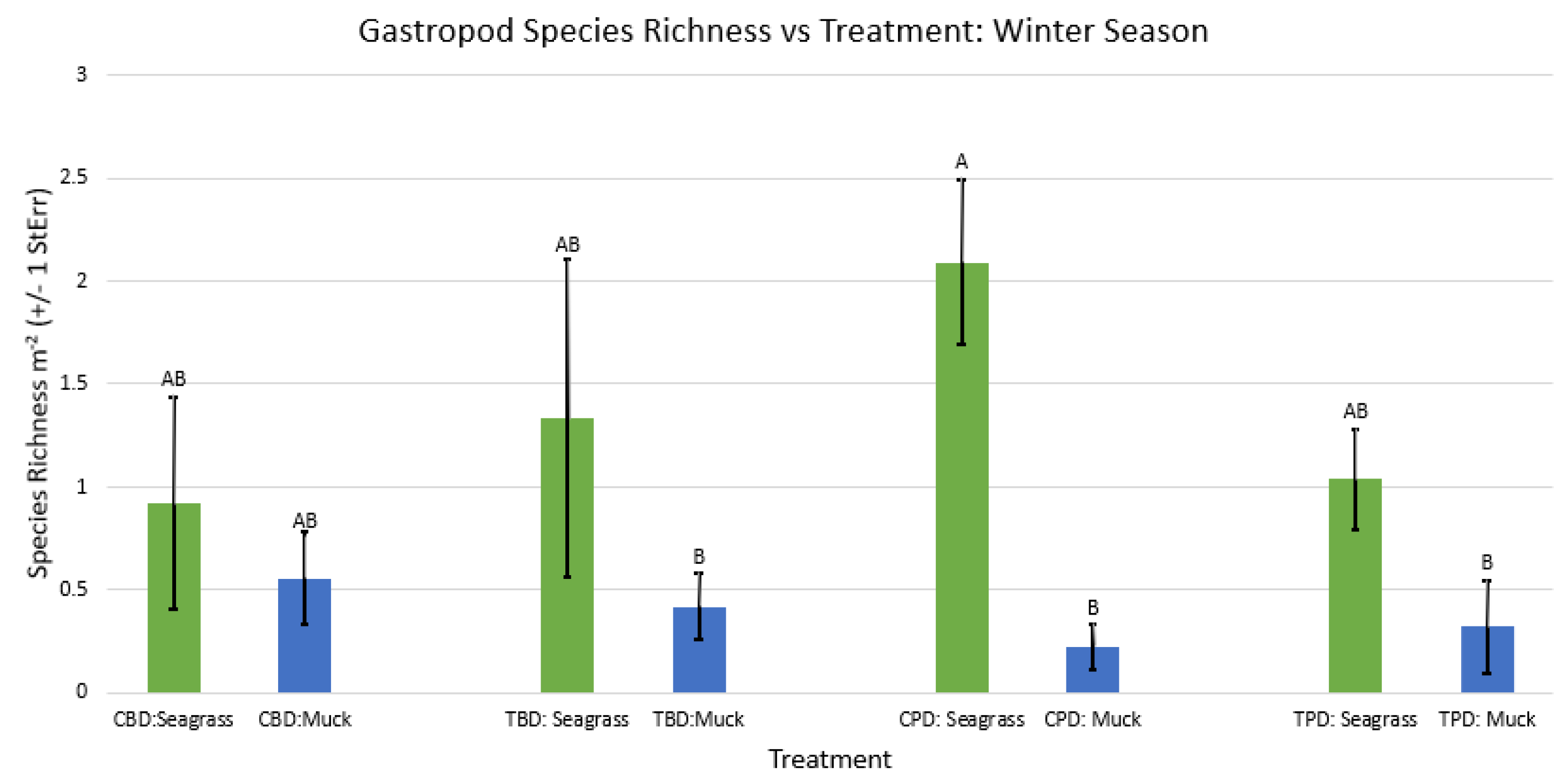
Figure A53.
2-way ANOVA Figure Afor all benthic gastropod species richness vs Treatment in all seasons.
Figure A53.
2-way ANOVA Figure Afor all benthic gastropod species richness vs Treatment in all seasons.
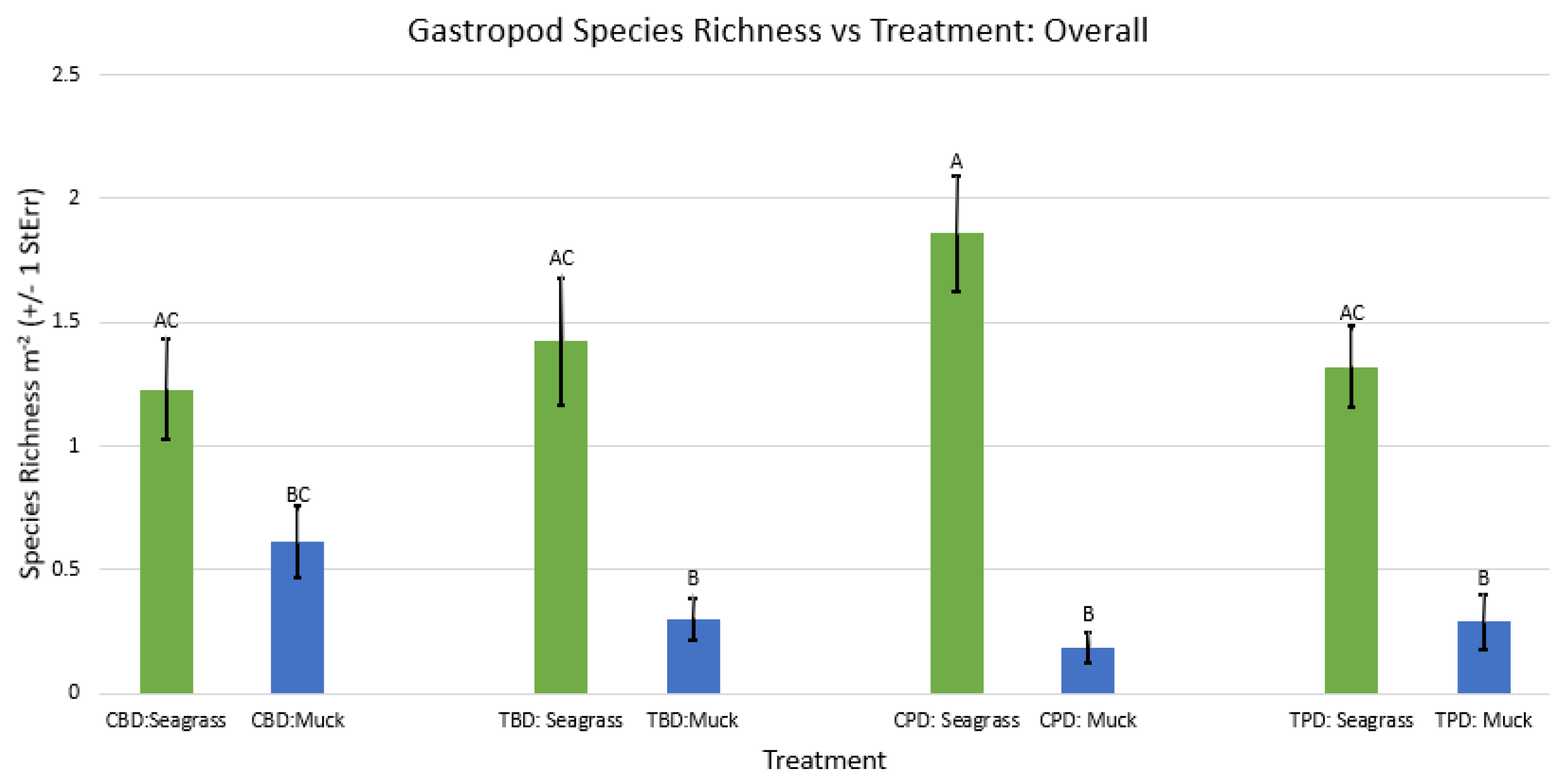
Figure A54.
2-way ANOVA Figure Afor all benthic gastropod abundance vs Treatment in the spring.
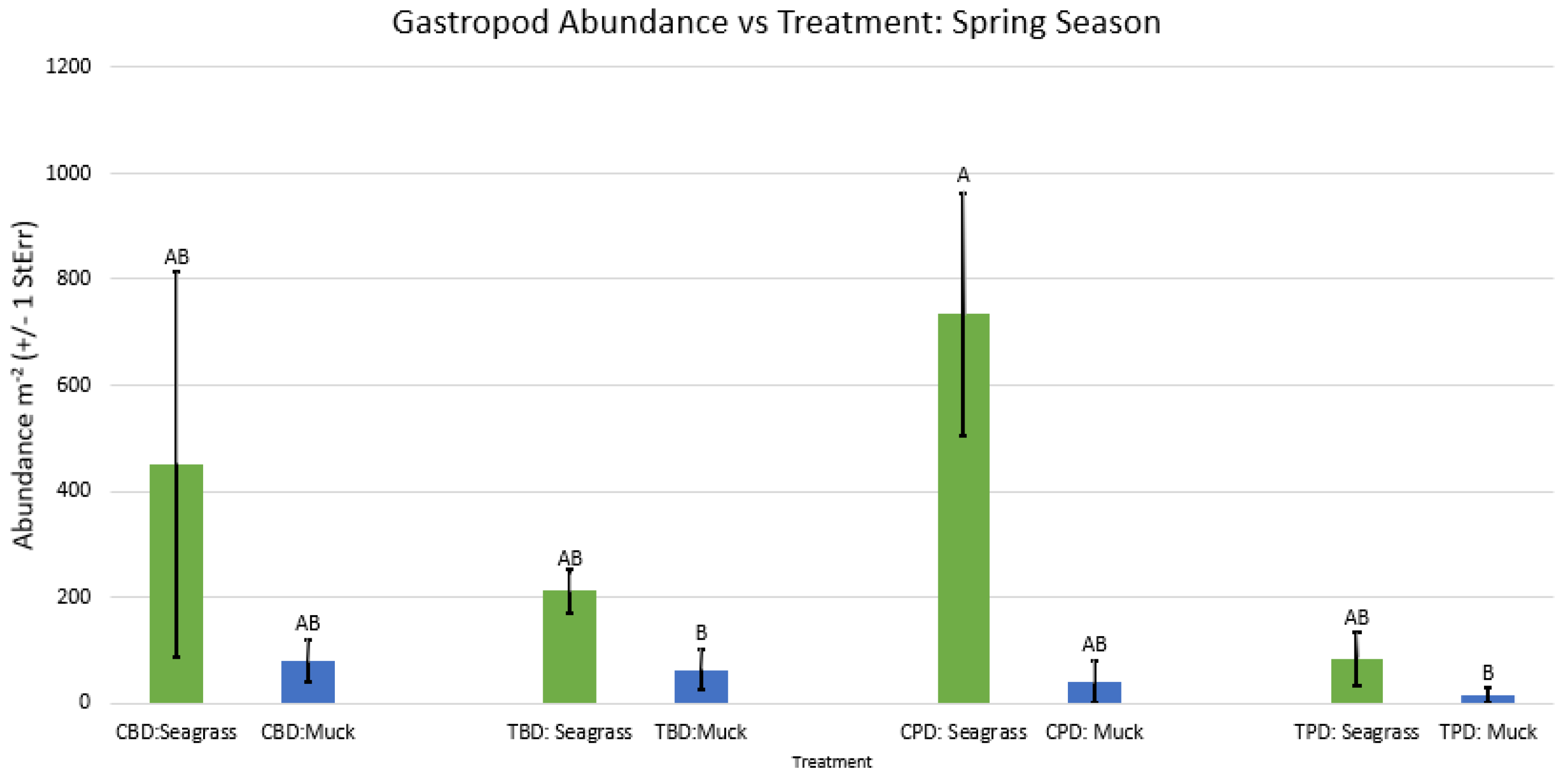
Figure A55.
2-way ANOVA Figure Afor all benthic gastropod abundance vs Treatment in the summer.
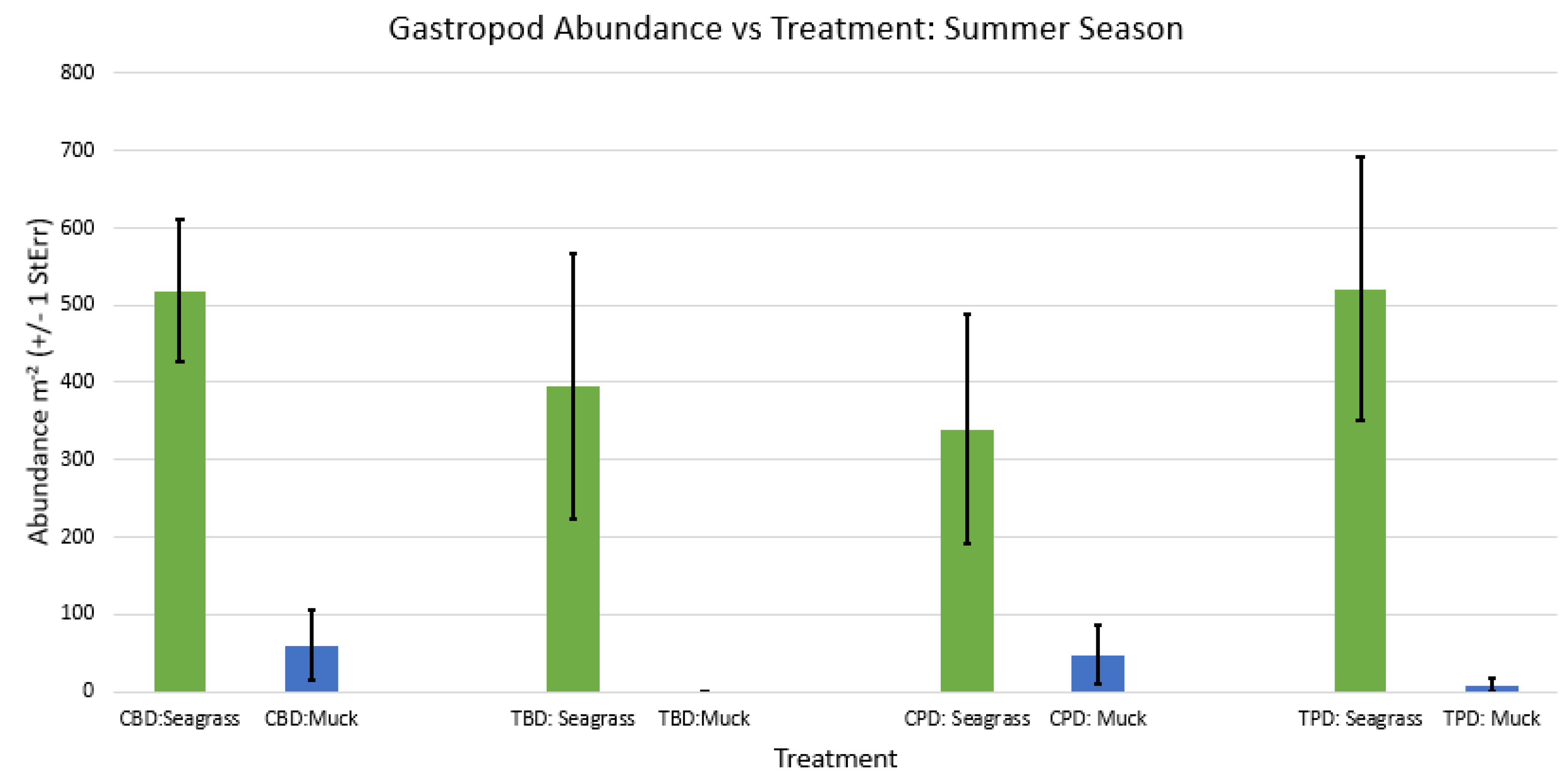
Figure A56.
2-way ANOVA Figure Afor all benthic gastropod abundance vs Treatment in the fall.
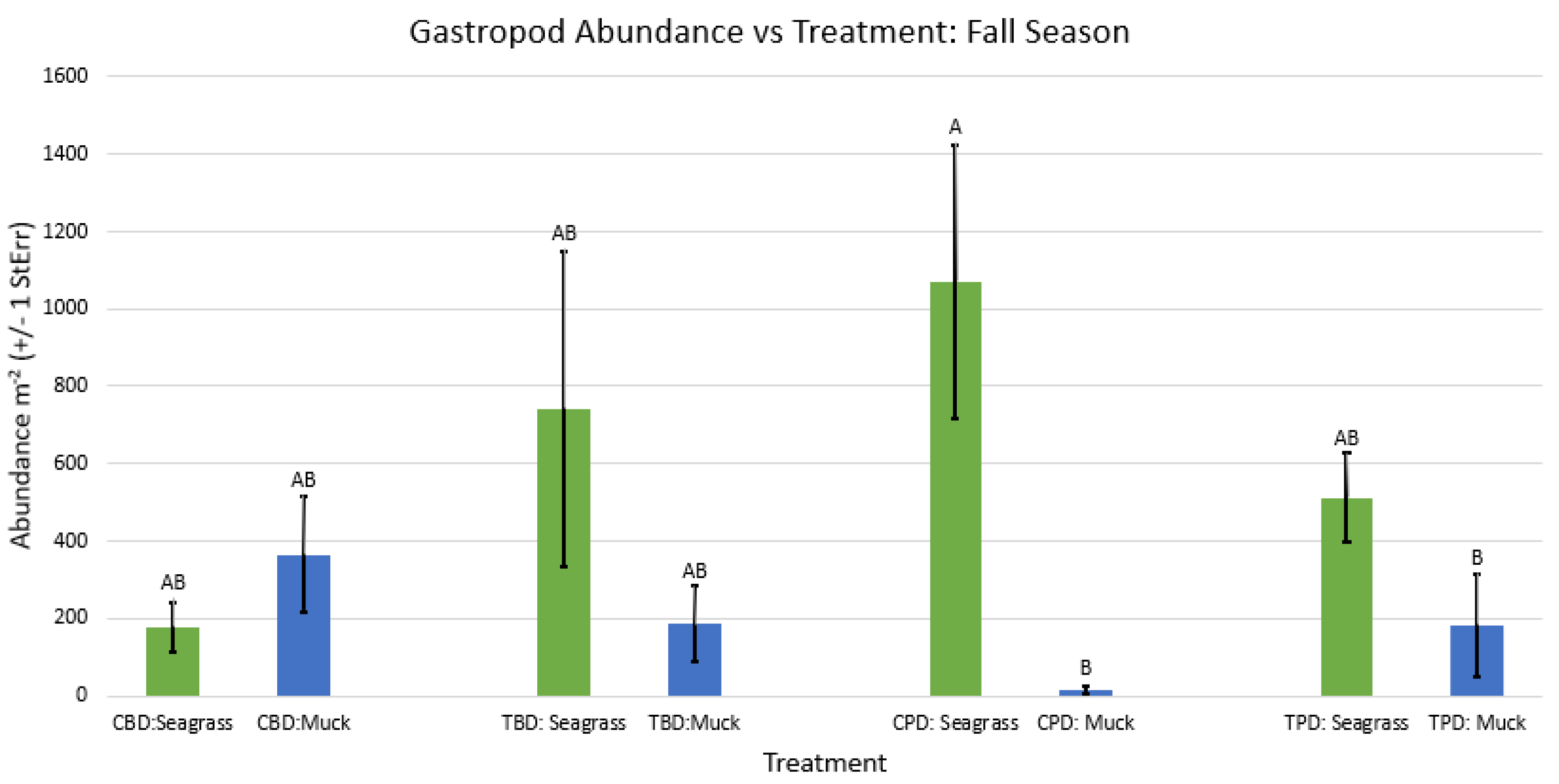
Figure A57.
2-way ANOVA Figure Afor all benthic gastropod abundance vs Treatment in the winter.
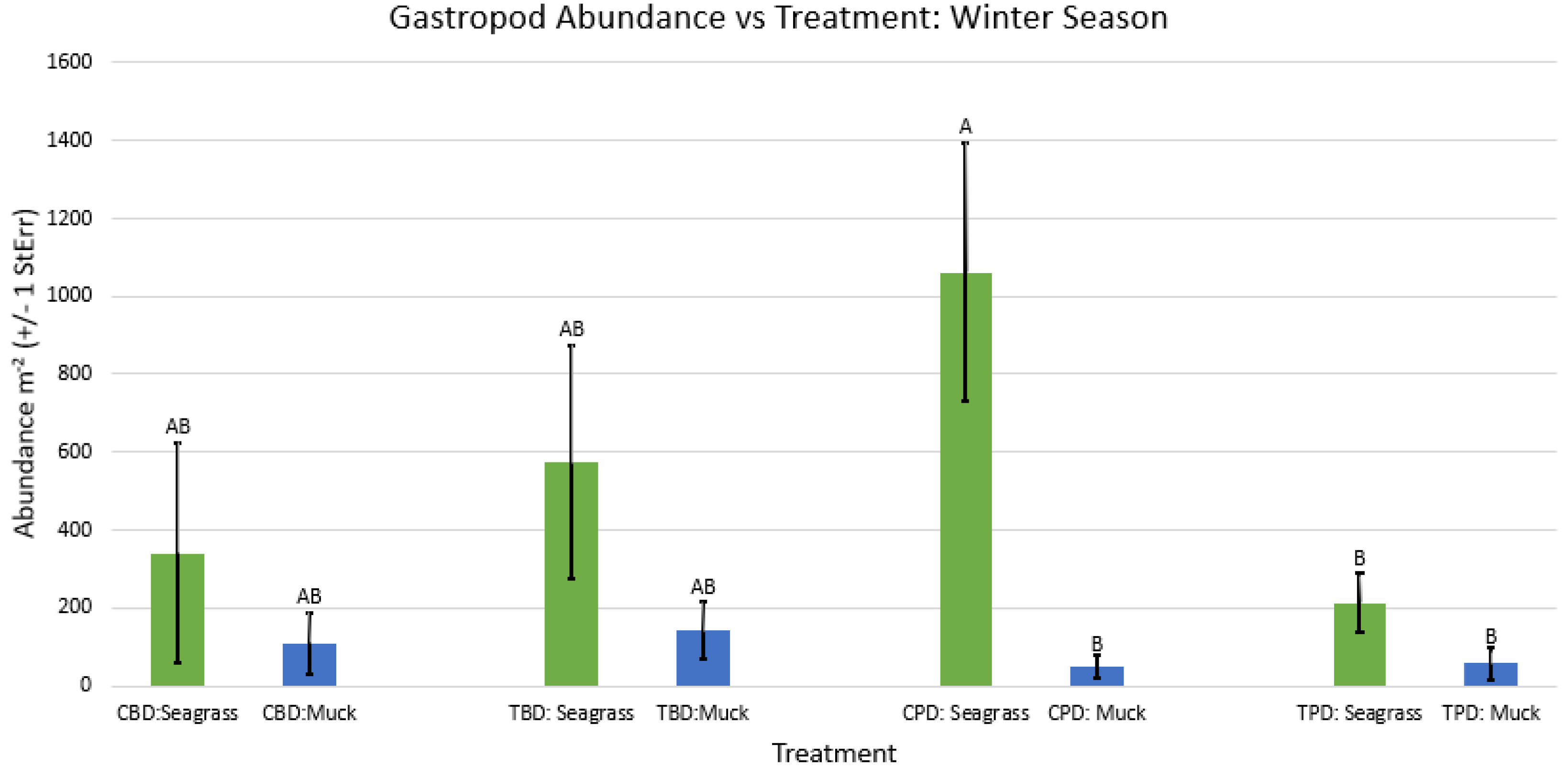
Figure A58.
2-way ANOVA Figure Afor all benthic gastropod abundance vs Treatment in all seasons.
Figure A59.
2-way ANOVA Figure Afor all benthic bivalve abundance vs Treatment in all seasons.
Figure A60.
2-way ANOVA Figure Afor all benthic bivalve species richness vs Treatment in the spring.
Figure A60.
2-way ANOVA Figure Afor all benthic bivalve species richness vs Treatment in the spring.
Figure A61.
2-way ANOVA Figure Afor all benthic bivalve species richness vs Treatment in the summer.
Figure A61.
2-way ANOVA Figure Afor all benthic bivalve species richness vs Treatment in the summer.
Figure A62.
2-way ANOVA Figure Afor all benthic bivalve species richness vs Treatment in the fall.
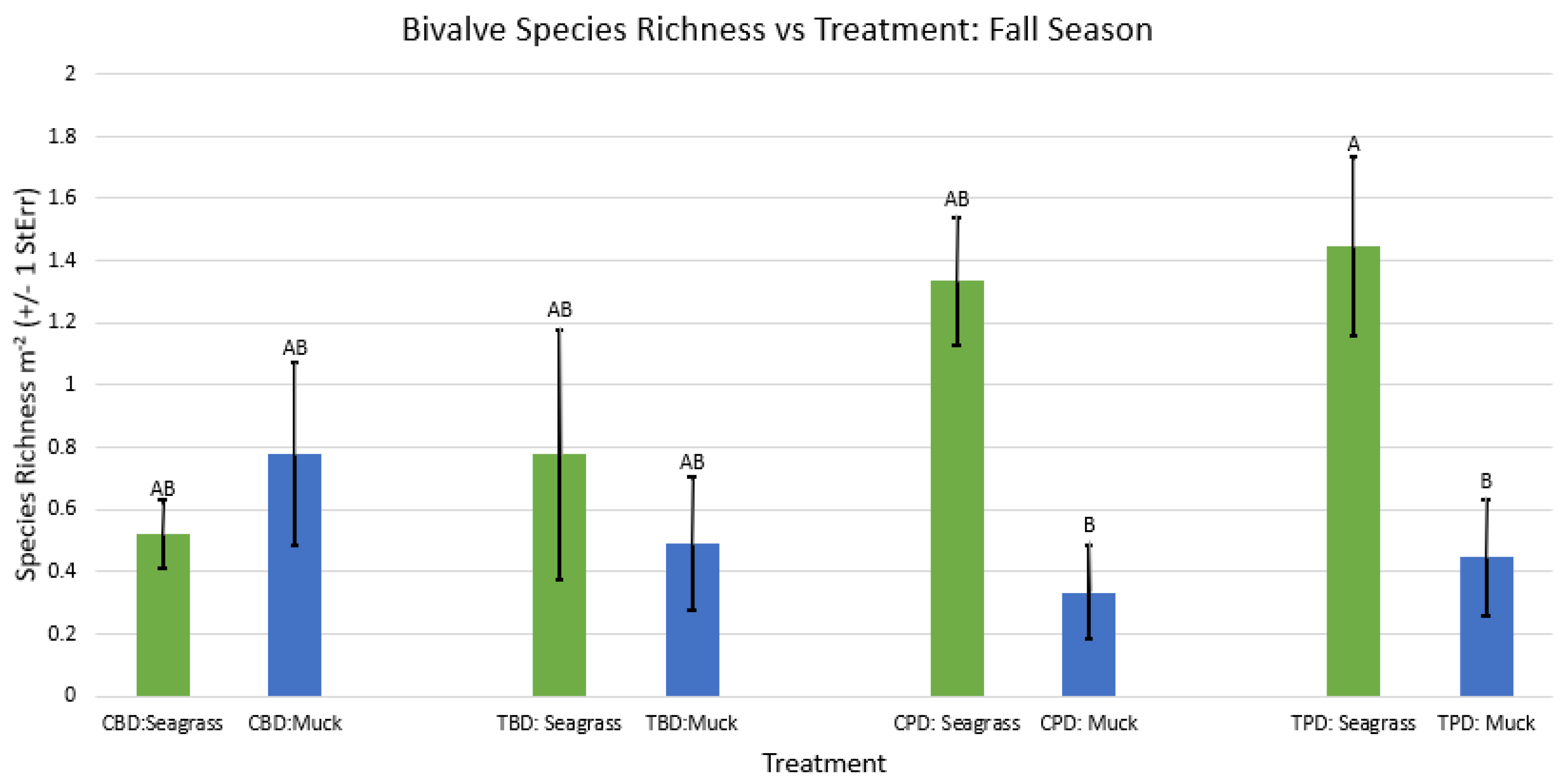
Figure A63.
2-way ANOVA Figure Afor all benthic bivalve species richness vs Treatment in the winter.
Figure A63.
2-way ANOVA Figure Afor all benthic bivalve species richness vs Treatment in the winter.

Figure A64.
2-way ANOVA Figure Afor all benthic bivalve species richness vs Treatment in all seasons.
Figure A64.
2-way ANOVA Figure Afor all benthic bivalve species richness vs Treatment in all seasons.
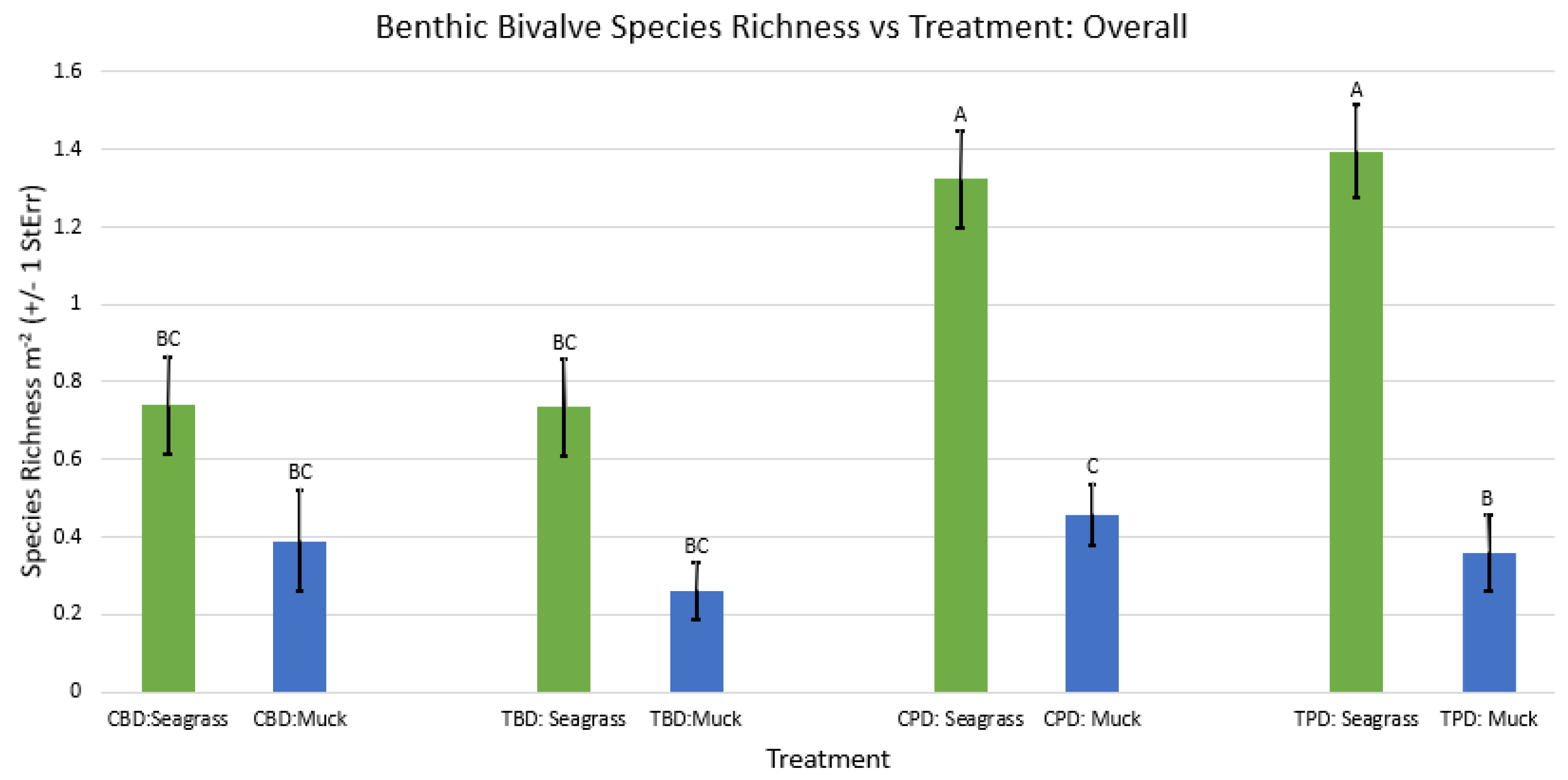
Figure A65.
2-way ANOVA Figure Afor all benthic bivalve abundance vs Treatment in the spring.
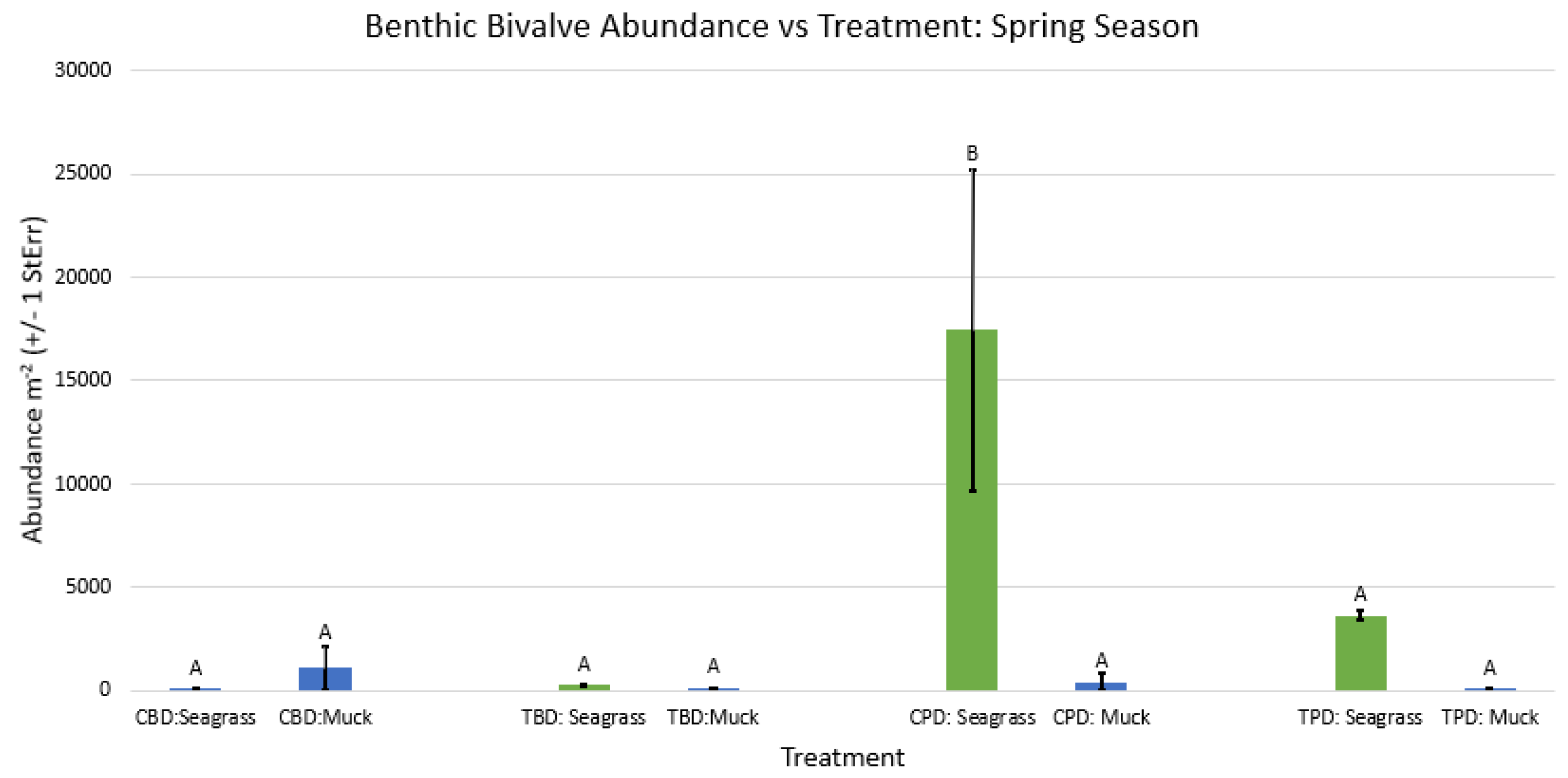
Figure A66.
2-way ANOVA Figure Afor all benthic bivalve abundance vs Treatment in the fall.
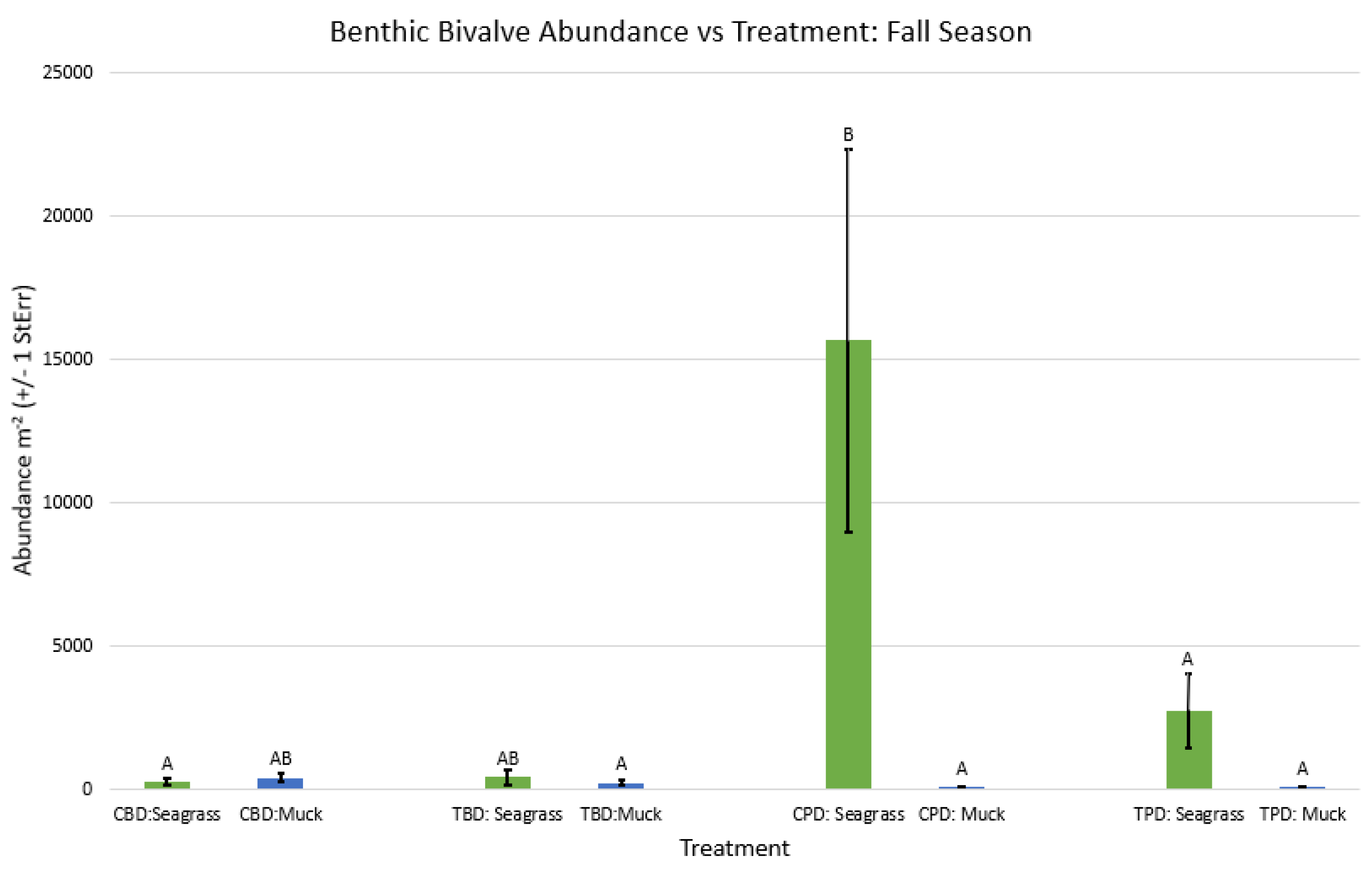
Figure A67.
2-way ANOVA Figure Afor all benthic bivalve abundance vs Treatment in all seasons.
Appendix C: nMDS Figures
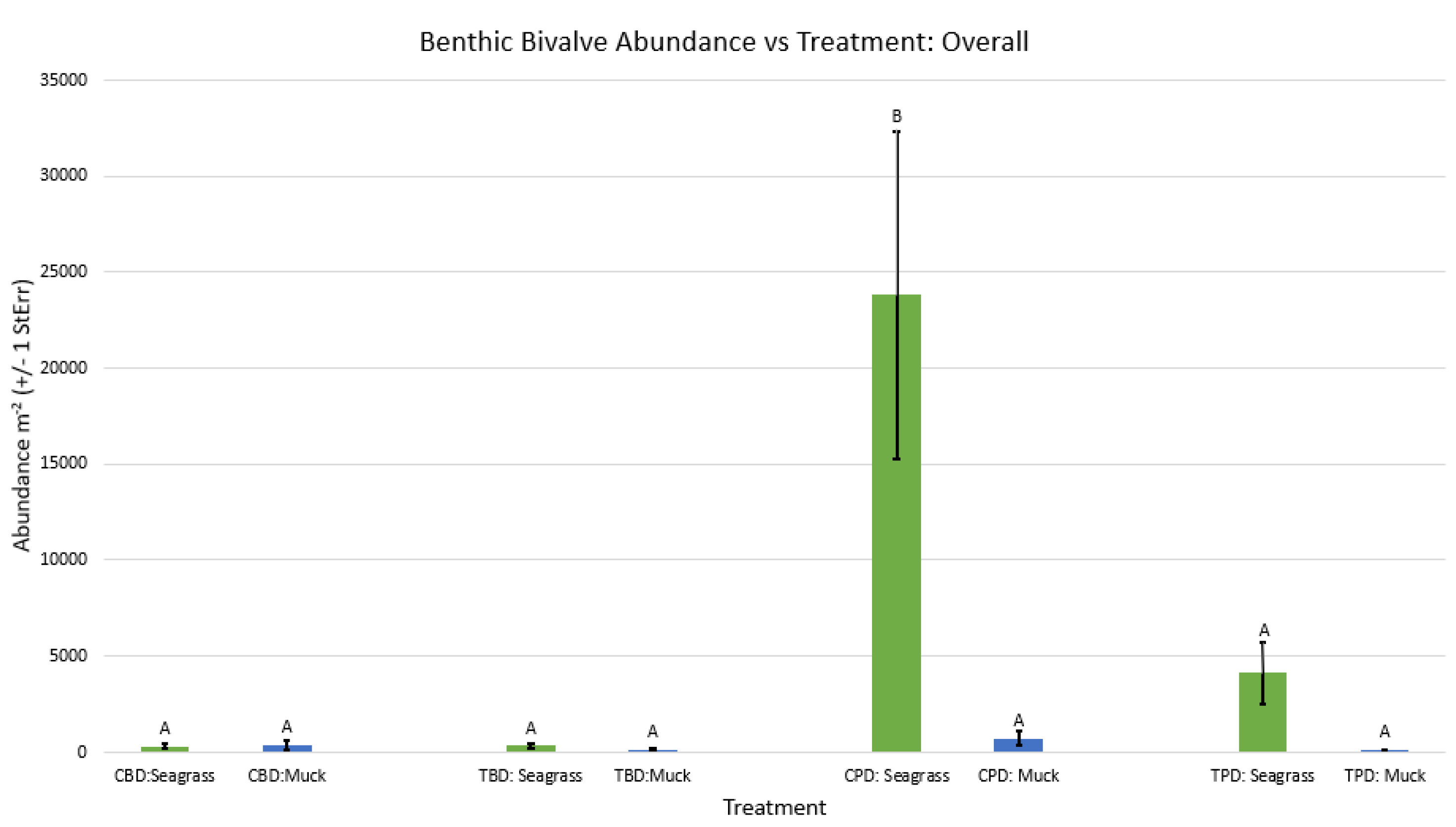
Figure A1.
Non-Metric Multidimensional Scaling showing the pooled benthic bivalve biodiversity at the treatment and control muck stations similarities and dissimilarities in multivariate space. CBD=Control Before Dredging; CPD=Control Post Dredging; TBD=Treatment Before Dredging; TPD=Treatment Post Dredging. Ellipses represent distinct groups based on similarity.
Figure A1.
Non-Metric Multidimensional Scaling showing the pooled benthic bivalve biodiversity at the treatment and control muck stations similarities and dissimilarities in multivariate space. CBD=Control Before Dredging; CPD=Control Post Dredging; TBD=Treatment Before Dredging; TPD=Treatment Post Dredging. Ellipses represent distinct groups based on similarity.

Figure A2.
Non-Metric Multidimensional Scaling showing the pooled benthic bivalve species richness at the treatment and control muck stations similarities and dissimilarities in multivariate space. CBD=Control Before Dredging; CPD=Control Post Dredging; TBD=Treatment Before Dredging; TPD=Treatment Post Dredging. Ellipses represent distinct groups based on similarity.
Figure A2.
Non-Metric Multidimensional Scaling showing the pooled benthic bivalve species richness at the treatment and control muck stations similarities and dissimilarities in multivariate space. CBD=Control Before Dredging; CPD=Control Post Dredging; TBD=Treatment Before Dredging; TPD=Treatment Post Dredging. Ellipses represent distinct groups based on similarity.

Figure A3.
Non-Metric Multidimensional Scaling showing the pooled benthic bivalve abundance at the treatment and control muck stations similarities and dissimilarities in multivariate space. CBD=Control Before Dredging; CPD=Control Post Dredging; TBD=Treatment Before Dredging; TPD=Treatment Post Dredging. Ellipses represent distinct groups based on similarity.
Figure A3.
Non-Metric Multidimensional Scaling showing the pooled benthic bivalve abundance at the treatment and control muck stations similarities and dissimilarities in multivariate space. CBD=Control Before Dredging; CPD=Control Post Dredging; TBD=Treatment Before Dredging; TPD=Treatment Post Dredging. Ellipses represent distinct groups based on similarity.
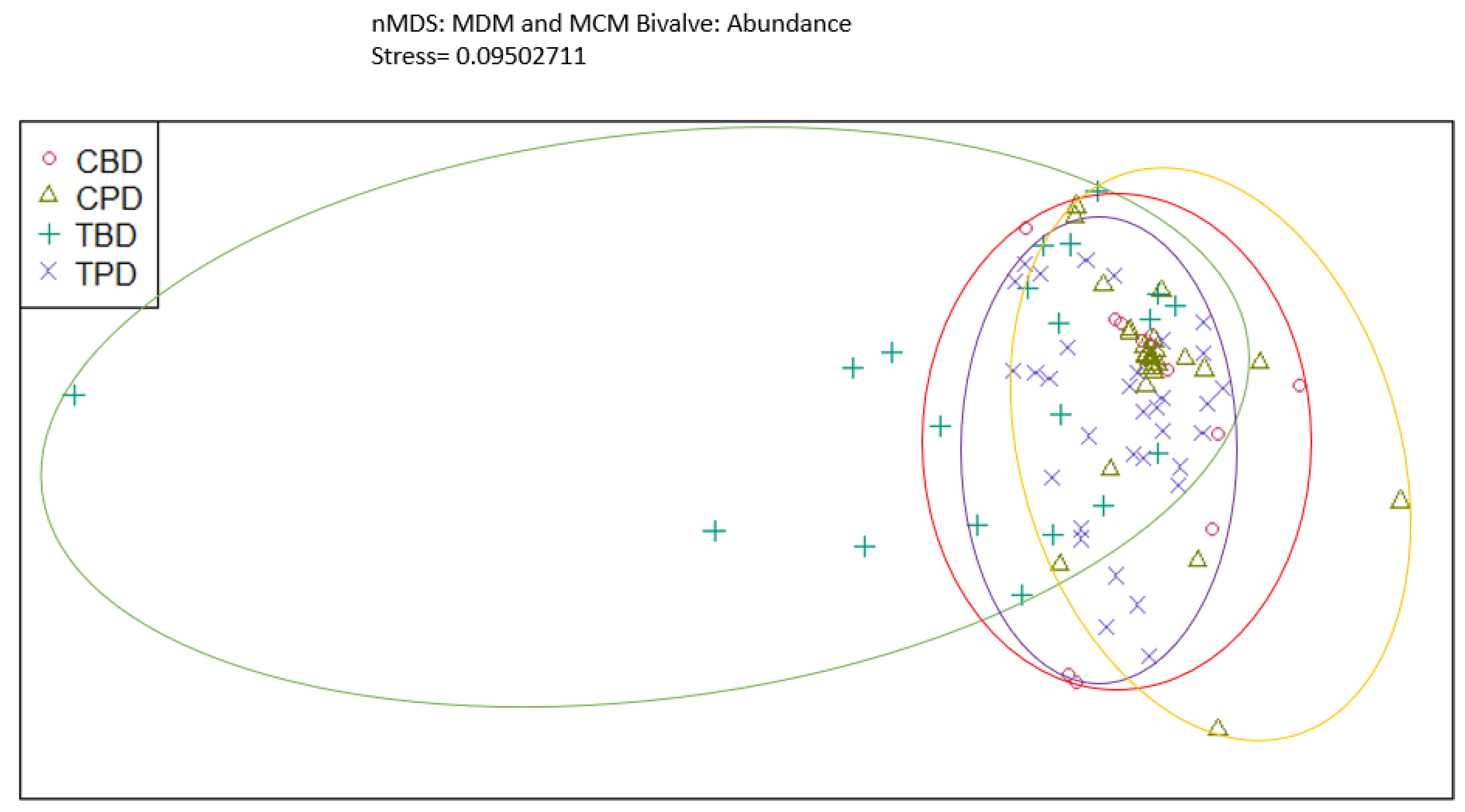
Figure A4.
Non-Metric Multidimensional Scaling showing the pooled benthic bivalve biodiversity at the treatment and control seagrass stations similarities and dissimilarities in multivariate space. CBD=Control Before Dredging; CPD=Control Post Dredging; TBD=Treatment Before Dredging; TPD=Treatment Post Dredging. Ellipses represent distinct groups based on similarity.
Figure A4.
Non-Metric Multidimensional Scaling showing the pooled benthic bivalve biodiversity at the treatment and control seagrass stations similarities and dissimilarities in multivariate space. CBD=Control Before Dredging; CPD=Control Post Dredging; TBD=Treatment Before Dredging; TPD=Treatment Post Dredging. Ellipses represent distinct groups based on similarity.
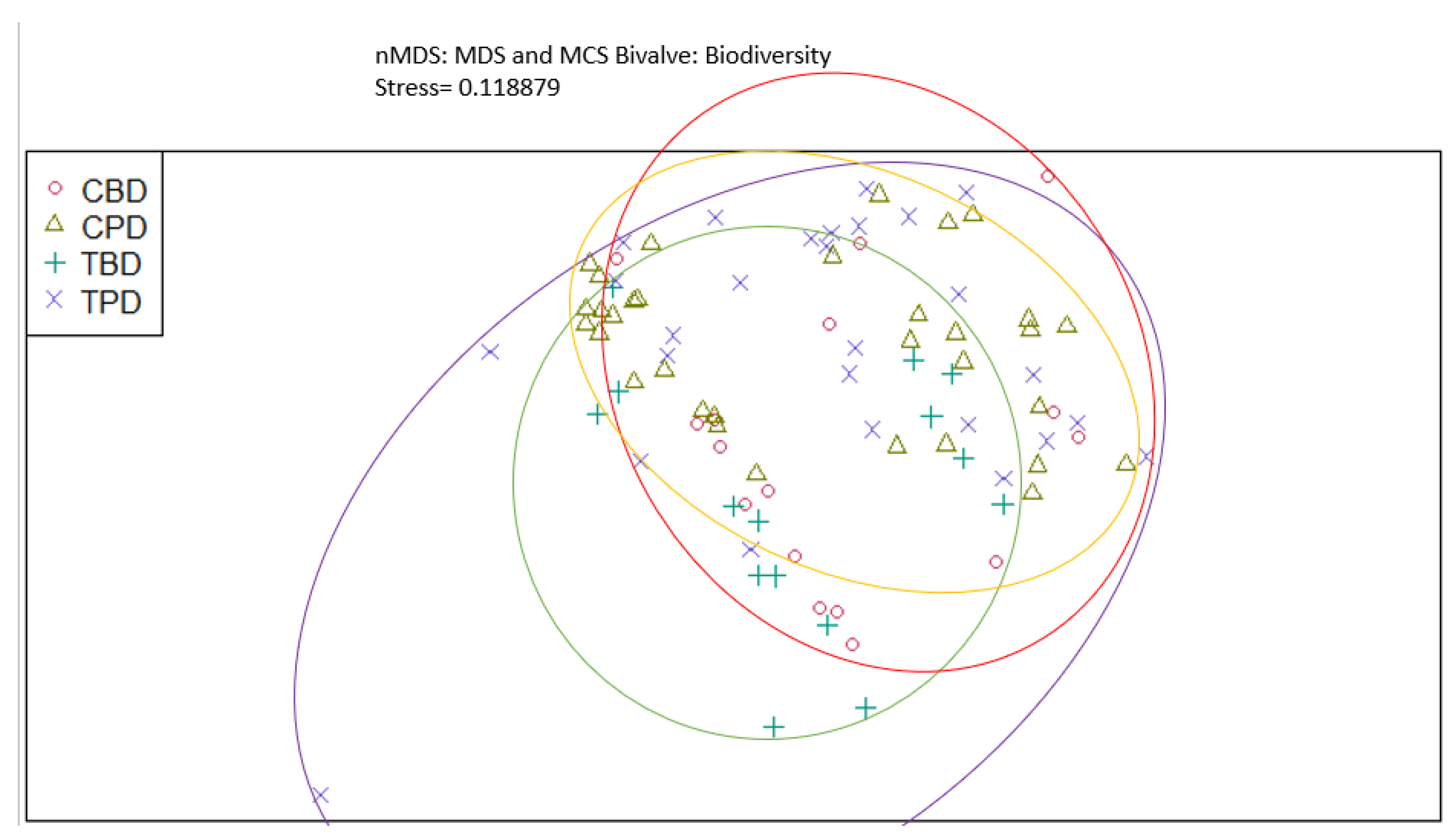
Figure A5.
Non-Metric Multidimensional Scaling showing the pooled benthic bivalve species richness at the treatment and control seagrass stations similarities and dissimilarities in multivariate space. CBD=Control Before Dredging; CPD=Control Post Dredging; TBD=Treatment Before Dredging; TPD=Treatment Post Dredging. Ellipses represent distinct groups based on similarity.
Figure A5.
Non-Metric Multidimensional Scaling showing the pooled benthic bivalve species richness at the treatment and control seagrass stations similarities and dissimilarities in multivariate space. CBD=Control Before Dredging; CPD=Control Post Dredging; TBD=Treatment Before Dredging; TPD=Treatment Post Dredging. Ellipses represent distinct groups based on similarity.
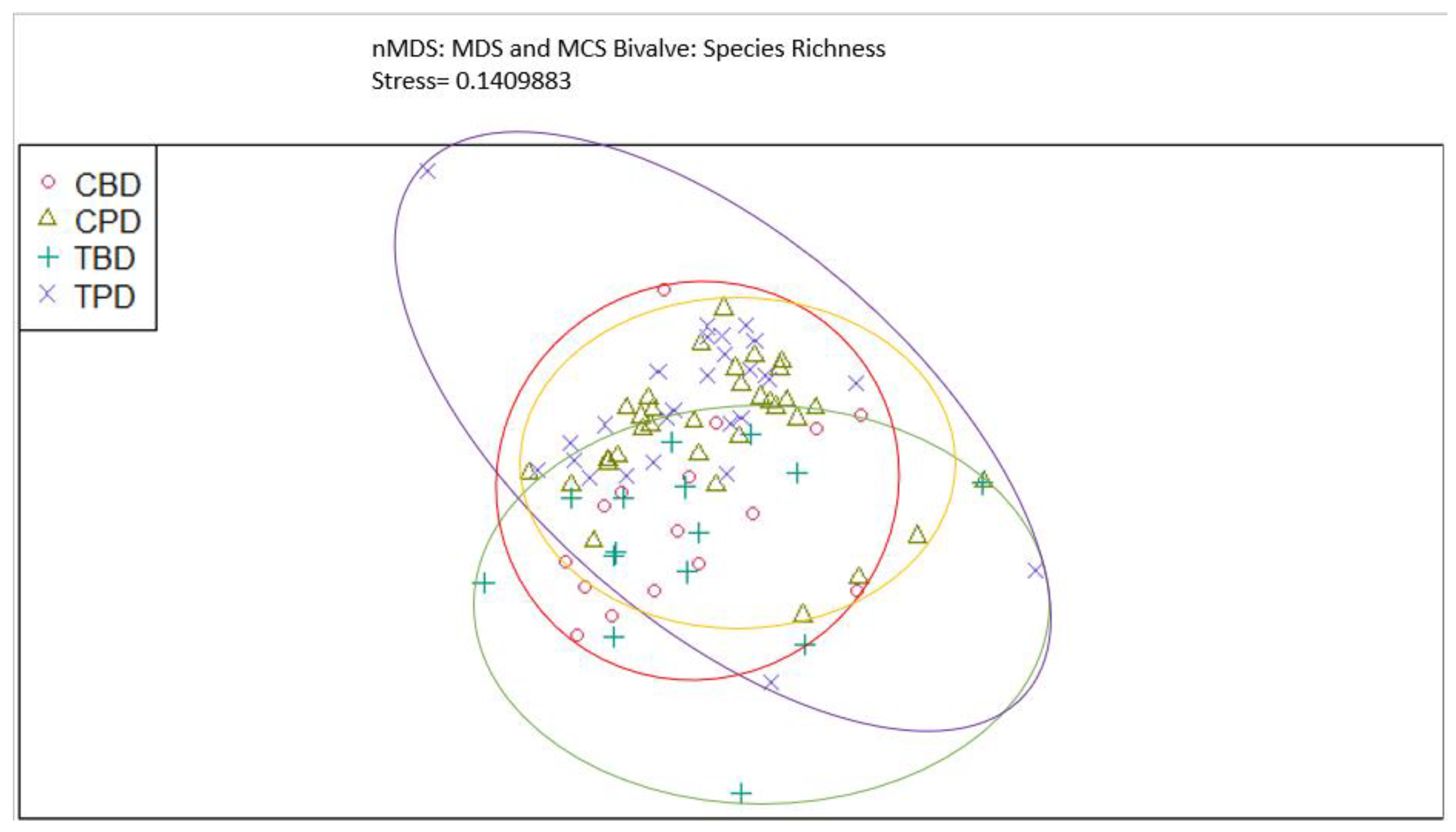
Figure A6.
Non-Metric Multidimensional Scaling showing the pooled benthic bivalve abundances at the treatment and control seagrass stations similarities and dissimilarities in multivariate space. CBD=Control Before Dredging; CPD=Control Post Dredging; TBD=Treatment Before Dredging; TPD=Treatment Post Dredging. Ellipses represent distinct groups based on similarity.
Figure A6.
Non-Metric Multidimensional Scaling showing the pooled benthic bivalve abundances at the treatment and control seagrass stations similarities and dissimilarities in multivariate space. CBD=Control Before Dredging; CPD=Control Post Dredging; TBD=Treatment Before Dredging; TPD=Treatment Post Dredging. Ellipses represent distinct groups based on similarity.
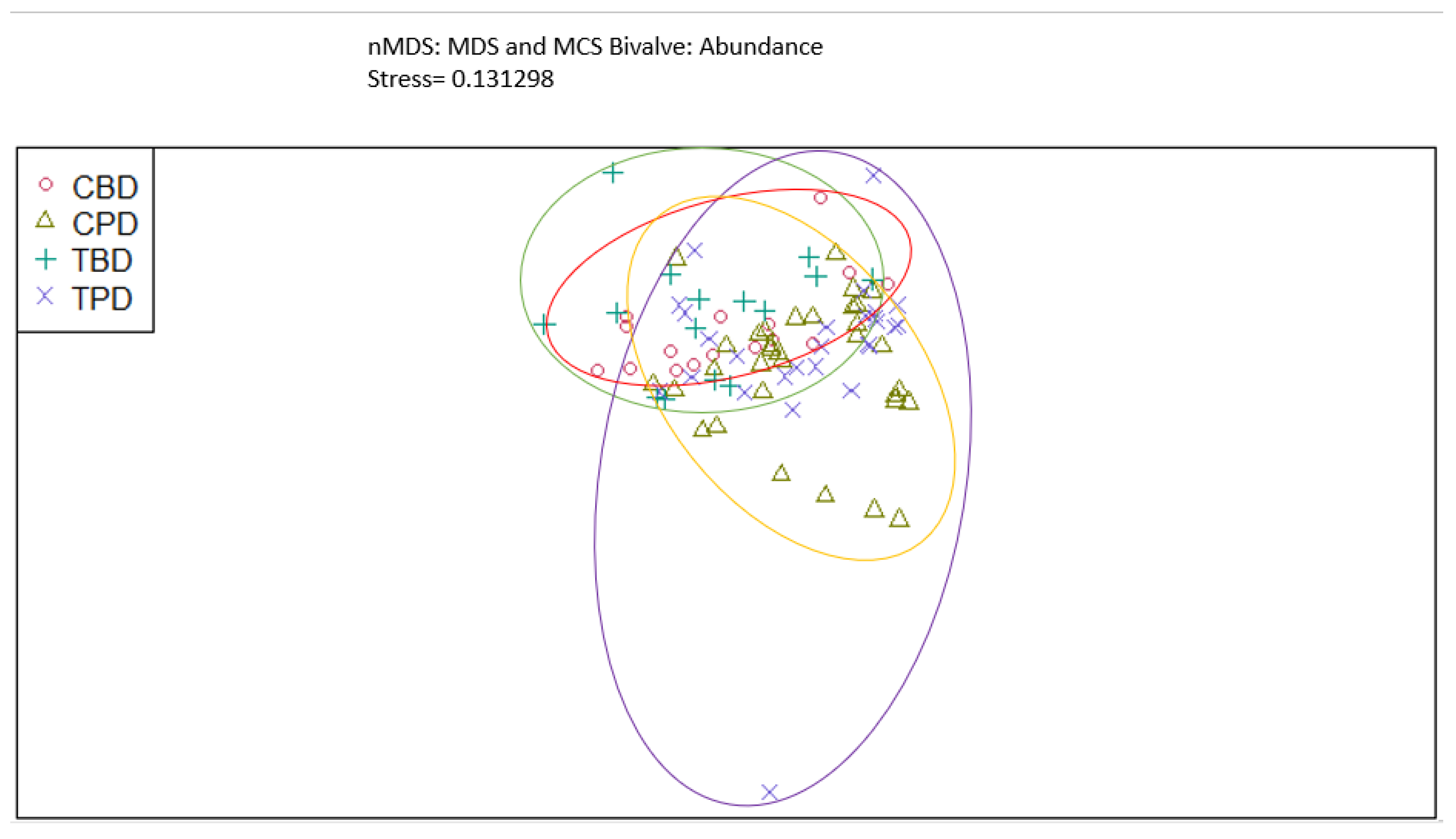
Figure A7.
Non-Metric Multidimensional Scaling showing the pooled benthic gastropod biodiversity at the treatment and control muck stations similarities and dissimilarities in multivariate space. CBD=Control Before Dredging; CPD=Control Post Dredging; TBD=Treatment Before Dredging; TPD=Treatment Post Dredging. Ellipses represent distinct groups based on similarity.
Figure A7.
Non-Metric Multidimensional Scaling showing the pooled benthic gastropod biodiversity at the treatment and control muck stations similarities and dissimilarities in multivariate space. CBD=Control Before Dredging; CPD=Control Post Dredging; TBD=Treatment Before Dredging; TPD=Treatment Post Dredging. Ellipses represent distinct groups based on similarity.

Figure A8.
Non-Metric Multidimensional Scaling showing the pooled benthic gastropod species richness at the treatment and control muck stations similarities and dissimilarities in multivariate space. CBD=Control Before Dredging; CPD=Control Post Dredging; TBD=Treatment Before Dredging; TPD=Treatment Post Dredging. Ellipses represent distinct groups based on similarity.
Figure A8.
Non-Metric Multidimensional Scaling showing the pooled benthic gastropod species richness at the treatment and control muck stations similarities and dissimilarities in multivariate space. CBD=Control Before Dredging; CPD=Control Post Dredging; TBD=Treatment Before Dredging; TPD=Treatment Post Dredging. Ellipses represent distinct groups based on similarity.
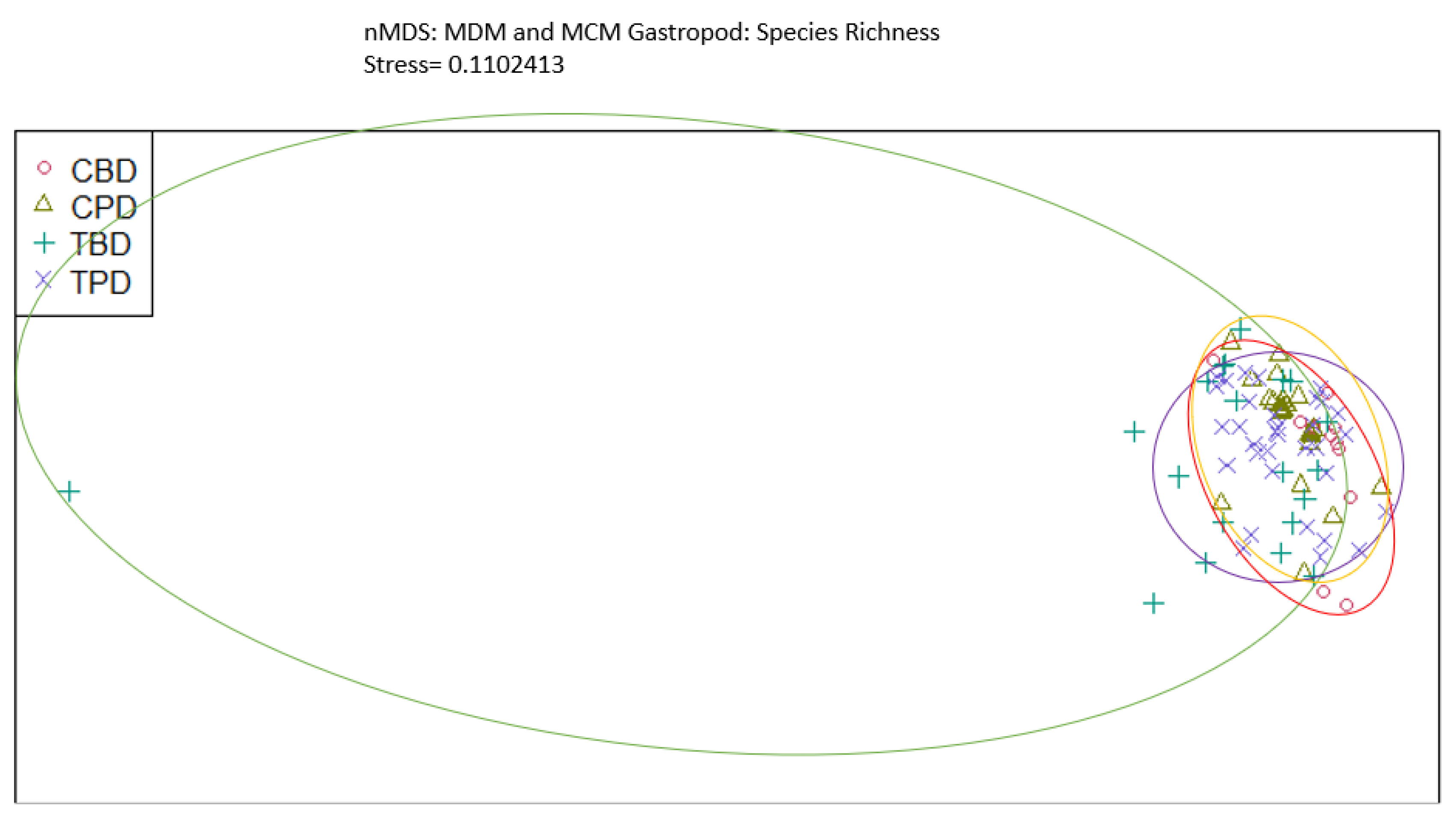
Figure A9.
Non-Metric Multidimensional Scaling showing the pooled benthic gastropod abundances at the treatment and control muck stations similarities and dissimilarities in multivariate space. CBD=Control Before Dredging; CPD=Control Post Dredging; TBD=Treatment Before Dredging; TPD=Treatment Post Dredging. Ellipses represent distinct groups based on similarity.
Figure A9.
Non-Metric Multidimensional Scaling showing the pooled benthic gastropod abundances at the treatment and control muck stations similarities and dissimilarities in multivariate space. CBD=Control Before Dredging; CPD=Control Post Dredging; TBD=Treatment Before Dredging; TPD=Treatment Post Dredging. Ellipses represent distinct groups based on similarity.

Figure A10.
Non-Metric Multidimensional Scaling showing the pooled benthic gastropod biodiversity at the treatment and control seagrass stations similarities and dissimilarities in multivariate space. CBD=Control Before Dredging; CPD=Control Post Dredging; TBD=Treatment Before Dredging; TPD=Treatment Post Dredging. Ellipses represent distinct groups based on similarity.
Figure A10.
Non-Metric Multidimensional Scaling showing the pooled benthic gastropod biodiversity at the treatment and control seagrass stations similarities and dissimilarities in multivariate space. CBD=Control Before Dredging; CPD=Control Post Dredging; TBD=Treatment Before Dredging; TPD=Treatment Post Dredging. Ellipses represent distinct groups based on similarity.
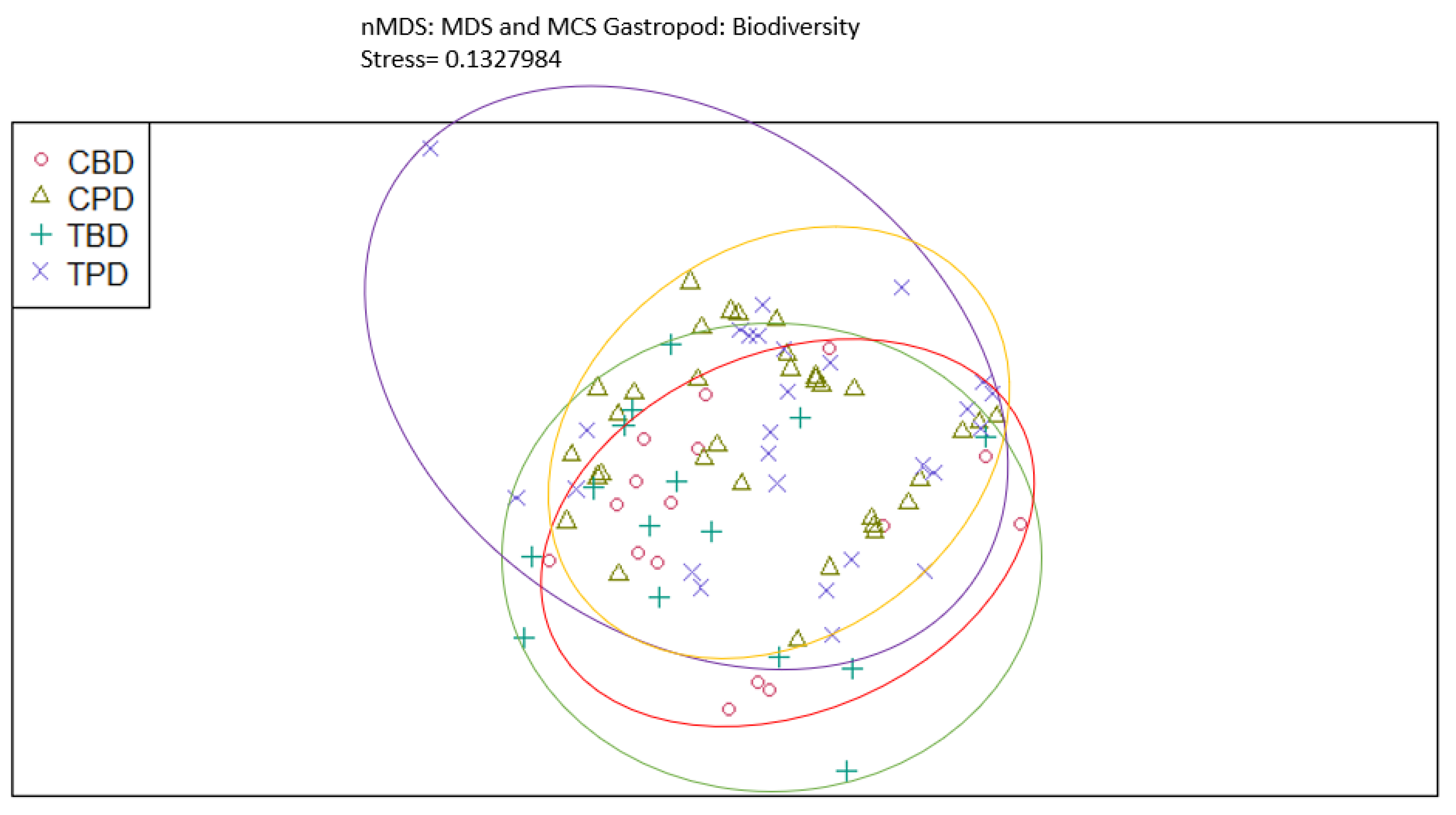
Figure A11.
Non-Metric Multidimensional Scaling showing the pooled benthic gastropod species richness at the treatment and control seagrass stations similarities and dissimilarities in multivariate space. CBD=Control Before Dredging; CPD=Control Post Dredging; TBD=Treatment Before Dredging; TPD=Treatment Post Dredging. Ellipses represent distinct groups based on similarity.
Figure A11.
Non-Metric Multidimensional Scaling showing the pooled benthic gastropod species richness at the treatment and control seagrass stations similarities and dissimilarities in multivariate space. CBD=Control Before Dredging; CPD=Control Post Dredging; TBD=Treatment Before Dredging; TPD=Treatment Post Dredging. Ellipses represent distinct groups based on similarity.
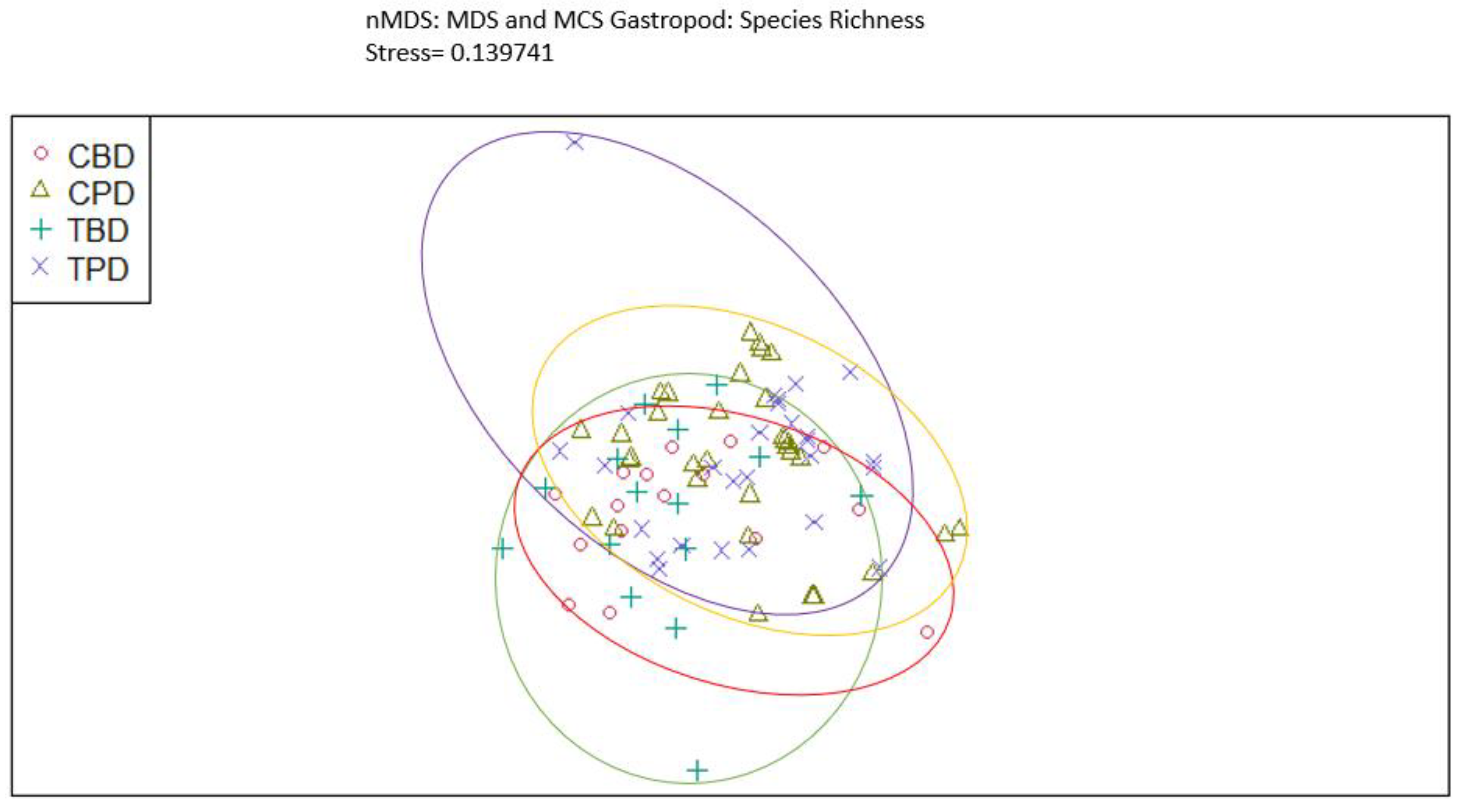
Figure A12.
Non-Metric Multidimensional Scaling showing the pooled benthic gastropod abundances at the treatment and control seagrass stations similarities and dissimilarities in multivariate space. CBD=Control Before Dredging; CPD=Control Post Dredging; TBD=Treatment Before Dredging; TPD=Treatment Post Dredging. Ellipses represent distinct groups based on similarity.
Figure A12.
Non-Metric Multidimensional Scaling showing the pooled benthic gastropod abundances at the treatment and control seagrass stations similarities and dissimilarities in multivariate space. CBD=Control Before Dredging; CPD=Control Post Dredging; TBD=Treatment Before Dredging; TPD=Treatment Post Dredging. Ellipses represent distinct groups based on similarity.
References
- Bricker, S. B., Longstaff, B., Dennison, W., Jones, A., Boicourt, K., Wicks, C., & Woerner, J. (2008). Effects of nutrient enrichment in the nation's estuaries: a decade of change. Harmful Algae, 8(1), 21-32. J. Mar. Sci. Eng. 2023, 11, 444. [CrossRef]
- Tremain, D. M., & Adams, D. H. (1995). Seasonal variations in species diversity, abundance, and composition of fish communities in the northern Indian River Lagoon, Florida. Bulletin of Marine Science, 57(1), 171-192.
- Dawes, C. J., Hanisak, D., & Kenworthy, J. W. (1995). Seagrass biodiversity in the Indian river lagoon. Bulletin of Marine Science, 57(1), 59-66.
- Barile, P. J. (2018). Widespread sewage pollution of the Indian River Lagoon system, Florida (USA) resolved by spatial analyses of macroalgal biogeochemistry. Marine pollution bulletin, 128, 557-574.
- Tetra Tech, Inc and Close Waters LLC. (2018). Save our lagoon project plan for Brevard County, Florida. Report to Brevard County Natural Resources Management Department.
- Fox, A. L., & Trefry, J. H. (2018). Environmental Dredging to Remove Fine Grained, Organic Rich Sediments and Reduce Inputs of Nitrogen and Phosphorus to a Subtropical Estuary. Marine Technology Society Journal, 52(4), 42-57.
- Bradshaw, D. J. , Dickens, N. J., Trefry, J. H., & McCarthy, P. J. (2020). Defining the sediment microbiome of the Indian River Lagoon, FL, USA, an Estuary of National Significance. bioRxiv.
- Erftemeijer, P. L. , & Lewis III, R. R. R. (2006). Environmental impacts of dredging on seagrasses: a review. Marine pollution bulletin, 52(12), 1553-1572.
- Donohue, I., & Garcia Molinos, J. (2009). Impacts of increased sediment loads on the ecology of lakes. Biological Reviews, 84(4), 517-531.
- Trefry, J. H., Trocine, R. P., & Woodall, D. W. (2007). Composition and sources of suspended matter in the Indian River Lagoon, Florida. Florida Scientist, 363-382.
- Nerlović, V., Doğan, A., & Hrs-Brenko, M. (2011). Response to oxygen deficiency (depletion): Bivalve assemblages as an indicator of ecosystem instability in the northern Adriatic Sea. Biologia, 66(6), 1114.
- Cox, A. , Hope, D., Angelica Zamora-Duran, M., & Johnson, K. B. (2018). Environmental Factors Influencing Benthic Polychaete Distributions in a Subtropical Lagoon. Marine Technology Society Journal, 52(4), 58-74.
- Mikkelsen, P. M., Mikkelsen, P. S., & Karlen, D. J. (1995). Molluscan biodiversity in the Indian River lagoon, Florida. Bulletin of Marine Science, 57(1), 94-127.
- Fedosov, A. E., & Puillandre, N. (2012). Phylogeny and taxonomy of the Kermia Pseudodaphnella (Mollusca: Gastropoda: Raphitomidae) genus complex: a remarkable radiation via diversification of larval development. Systematics and Biodiversity, 10(4), 447-477.
- Parkhaev, P. Y. (2017). Origin and the early evolution of the phylum Mollusca. Paleontological Journal, 51(6), 663-686.
- Coelho, J. P., Duarte, A. C., Pardal, M. A., & Pereira, M. E. (2014). Scrobicularia plana (Mollusca, Bivalvia) as a biomonitor for mercury contamination in Portuguese estuaries. Ecological indicators, 46, 447-453.
- Kim, Y. R., Lee, S., Kim, J., Kim, C. J., Choi, K. Y., & Chung, C. S. (2018). Thyasira tokunagai as an ecological indicator for the quality of sediment and benthic communities in the East Sea-Byeong, Korea. Marine pollution bulletin, 135, 873-879.
- Wu, H., Guan, Q., Lu, X., & Batzer, D. P. (2017). Snail (Mollusca: Gastropoda) assemblages as indicators of ecological condition in freshwater wetlands of Northeastern China. Ecological Indicators, 75, 203-209.
- McKeon, C. S., Tunberg, B. G., Johnston, C. A., & Barshis, D. J. (2015). Ecological drivers and habitat associations of estuarine bivalves. PeerJ, 3, e1348.
- Moraitis, M. L., Tsikopoulou, I., Geropoulos, A., Dimitriou, P. D., Papageorgiou, N., Giannoulaki, M., ... & Karakassis, I. (2018). Molluscan indicator species and their potential use in ecological status assessment using species distribution modeling. Marine environmental research, 140, 10-17.
- Johnson, K. B., Shenker, J. M., & Trefry, J. H. (2019, November). Muck Removal Efficiency plus Biological and Chemical Responses/Improvements after Dredging (Subtask 2).
- Bayne, B. L., & Newell, R. C. (1983). Physiological energetics of marine molluscs. In The mollusca (pp. 407-515). Academic Press.
- Fox, H. M., & Simmonds, B. G. (1933). Metabolic rates of aquatic arthropods from different habitats. Journal of Experimental Biology, 10(1), 67-74.
- Pamatmat, M. M. (1983). Measuring aerobic and anaerobic metabolism of benthic.
- infauna under natural conditions. Journal of Experimental Zoology, 228(3), 405-413.
- Salvato, B., Cuomo, V., Di Muro, P., & Beltramini, M. (2001). Effects of environmental parameters on the oxygen consumption of four marine invertebrates: a comparative factorial study. Marine Biology, 138(4), 659-668.
- Bayne, B. L. (Ed.). (1976). Marine mussels: their ecology and physiology (Vol. 10). Cambridge University Press.
- Theede, H., Ponat, A., Hiroki, K., & Schlieper, C. (1969). Studies on the resistance of marine bottom invertebrates to oxygen-deficiency and hydrogen sulphide. Marine Biology, 2(4), 325-337.
- Hope, D. C. (2016). The Tolerance of Benthic Infauna to Fine-Grained Organic Rich Sediments in a Shallow Subtropical Estuary (Doctoral dissertation).
- Pearson, T. H. (1980). Marine pollution effects of pulp and paper industry wastes. Helgoländer Meeresuntersuchungen, 33(1), 340.
- Rhoads, D. C. & Young, D. K., 1970. The influence of deposit feeding organisms on sediment stability and community trophic structure. - J. mar. Res. 28, 150-178.
- Folk, R. L. (1974). Petrology of sedimentary rocks: the University of Texas, Geology 370 K, 383 L, 383 M. Austin, TX: Hemphill.
- Sheridan, G. J., Noske, P. J., Whipp, R. K., & Wijesinghe, N. (2006). The effect of truck traffic and road water content on sediment delivery from unpaved forest roads. Hydrological Processes, 20(8), 1683–1699. [CrossRef]
- Dean Jr, W. E. (1974). Determination of carbonate and organic matter in calcareous sediments and sedimentary rocks by loss on ignition: comparison with other methods. Journal of Sedimentary Research, 44(1).
- Heiri, O. , Lotter, A. F., & Lemcke, G. (2001). Loss on ignition as a method for estimating organic and carbonate content in sediments: reproducibility and comparability of results. Journal of paleolimnology, 25(1), 101-110.
- Milliman, J. D. (1994). Organic matter content in US Atlantic continental slope sediments: decoupling the grain-size factor. Deep Sea Research Part II: Topical Studies in Oceanography, 41(4-6), 797-808.
- Hedges, J. I. , & Keil, R. G. (1995). Sedimentary organic matter preservation: an assessment and speculative synthesis. Marine chemistry, 49(2), 81-115.
- Wang, F. , & Chapman, P. M. (1999). Biological implications of sulfide in sediment—a review focusing on sediment toxicity. Environmental Toxicology and Chemistry, 18(11), 2526-2532.
- Mermillod-Blondin, F. , Rosenberg, R., François-Carcaillet, F., Norling, K., & Mauclaire, L. (2004). Influence of bioturbation by three benthic infaunal species on microbial communities and biogeochemical processes in marine sediment. Aquatic Microbial Ecology, 36(3), 271-284.
- Giblin, A. E. , Hopkinson, C. S., & Tucker, J. (1997). Benthic metabolism and nutrient cycling in Boston Harbor, Massachusetts. Estuaries, 20(2), 346- 364.
- Aller, R. C. (1994). Bioturbation and remineralization of sedimentary organic matter: effects of redox oscillation. Chemical Geology, 114(3), 331-345.
- Gray, J. S. (1982). Effects of pollutants on marine ecosystems. Netherlands Journal of Sea Research, 16, 424-443.
- Pearson, T. H., & Rosenberg, R. (1978). Macrobenthic succession in relation to organic enrichment and pollution of the marine environment. Oceanogr. Mar. Biol. Ann. Rev, 16, 229-311.
- Kodama, K., Lee, J. H., Oyama, M., Shiraishi, H., & Horiguchi, T. (2012). Disturbance of benthic macrofauna in relation to hypoxia and organic enrichment in a eutrophic coastal bay. Marine environmental research, 76, 80-89.
- Hyland, J., Balthis, L., Karakassis, I., Magni, P., Petrov, A., Shine, J., & Warwick, R. (2005). Organic carbon content of sediments as an indicator of stress in the marine benthos. Marine Ecology Progress Series, 295, 91-103.
- Magni, P. , Tagliapietra, D., Lardicci, C., Balthis, L., Castelli, A., Como, S., & Pessa, G. (2009). Animal-sediment relationships: Evaluating the ‘Pearson– Rosenberg paradigm in Mediterranean coastal lagoons. Marine Pollution Bulletin, 58(4), 478-486.
- Puente, A. , & Diaz, R. J. (2015). Response of benthos to ocean outfall discharges: does a general pattern exist?. Marine pollution bulletin, 101(1), 174-181.
- Gray, J. S., Wu, R. S. S., & Or, Y. Y. (2002). Effects of hypoxia and organic enrichment on the coastal marine environment. Marine Ecology Progress Series, 238, 249-279.
- Diaz, R. J. , & Rosenberg, R. (1995). Marine benthic hypoxia: a review of its ecological effects and the behavioural responses of benthic macrofauna. Oceanography and marine biology. An annual review, 33, 245-03.
- Viaroli, P. , Bartoli, M., Giordani, G., Naldi, M., Orfanidis, S., & Zaldivar, J. M. (2008). Community shifts, alternative stable states, biogeochemical controls and feedbacks in eutrophic coastal lagoons: a brief overview. Aquatic Conservation: Marine and Freshwater Ecosystems, 18(S1), S105-S117.
- Glassom, D. (1992). Predation/disturbance effects of greater flamingos (Phoenicopterus ruber) on the benthic communities of two Southern African lagoons (Master's thesis, University of Cape Town).
- Thistle, D. (1981). Natural physical disturbances and communities of marine soft bottoms. Marine Ecology Progress Series, 6(2), 223-228.
- Virnstein, R. W. (1977). The importance of predation by crabs and fishes on benthic infauna in Chesapeake Bay. Ecology, 58(6), 1199-1217.
- Virnstein, R. W. (1978). Predator caging experiments in soft sediments: caution advised. Estuarine interactions, 261-273.
- Quammen, M. L. (1984). Predation by shorebirds, fish, and crabs on invertebrates in intertidal mudflats: an experimental test. Ecology, 65(2), 529-537.
- Peterson, C. H. (1979). Predation, competitive exclusion, and diversity in the soft sediment benthic communities of estuaries and lagoons. In Ecological processes in coastal and marine systems (pp. 233-264). Springer, Boston, MA.
- Probert, P. K. (1984). Disturbance, sediment stability, and trophic structure of soft-bottom communities. Journal of Marine research, 42(4), 893-921.
- Chardy, P., & Clavier, J. (1988). Biomass and trophic structure of the macrobenthos in the south west lagoon of New Caledonia. Marine Biology, 99(2), 195-202.
- Woodin, S. A. (1999). Shallow water benthic ecology: a North American perspective of sedimentary habitats. Australian Journal of Ecology, 24(4), 291-301.
- Pelletiera, M. C., Goldb, A. J., Heltshec, J. F., & Buffumd, H. W. (2010). A method to identify estuarine macroinvertebrate pollution indicator species in the Virginian Biogeographic Province. Ecological Indicators, 10, 1037-1048.
- Engle, V. D., Summers, J. K., & Gaston, G. R. (1994). A benthic index of environmental condition of Gulf of Mexico estuaries. Estuaries, 17(2), 372-384.
- Dauvin, J. C. (2007). Paradox of estuarine quality: Benthic indicators and indices, consensus or debate for the future. Marine Pollution Bulletin, 55, 271-281.
- Borjaa, A. , Dauerb, D., Dıazc, R., Llansód, R. J., Muxikaa, I., Rodrıgueza, J. G., & Schaffnerc, L. (2007). Assessing estuarine benthic quality conditions in Chesapeake Bay: A comparison of three indices.
- Patmont, C., LaRosa, P., Narayanan, R., & Forrest, C. (2018). Environmental dredging residual generation and management. Integrated Environmental Assessment and Management, 14(3), 335-343.
- Bridges, T. S., Gustavson, K. E., Schroeder, P., Ells, S. J., Hayes, D., Nadeau, S. C., ... & Patmont, C. (2010). Dredging processes and remedy effectiveness: Relationship to the 4 Rs of environmental dredging. Integrated Environmental Assessment and Management, 6(4), 619-630.
- Bridges, T. S., Ells, S. J., Hayes, D. F., Mount, D., Nadeau, S. C., Palermo, M. R., ... & Schroeder, P. R. (2008). The four R's of environmental dredging: resuspension, release, residual and risk.
- Palermo, M. R., Schroeder, P. R., Estes, T. J., & Francingues, N. R. (2008). Technical guidelines for environmental dredging of contaminated sediments (No. ERDC/EL TR-08-29). Cold Regions Research and Engineering Laboratory (US).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



