
Submitted:
03 October 2024
Posted:
04 October 2024
You are already at the latest version
Abstract
The number of environmental surveillance reports of the ocean ecosystem through systematic studies on marine mammal health and disease is increasing. Understanding host-microbe interactions in cetaceans may contribute to the identification of compromised populations, microbial markers of disease, and ultimately to scientifically base population management decisions. This study is the first assessment of the oral microbiome of three pygmy sperm whales (Kogia breviceps), found stranded in Atlantic Iberian waters, by 16S rDNA-amplicon sequencing. A constrained canonical analysis approach showed that the oral microbiomes were shaped by the host species when compared to the oral microbiomes of three Odontoceti species obtained in a previous study (Delphinus delphis, Stenella coeruleoalba, and Phocoena phocoena). A total of 9 ASVs, belonging to the genera Campylobacter (2 ASVs), Phocoenobacter (4 ASVs), Psychrobacter (1 ASV), Tenacibaculum (1 ASV), and Terasakiispira (1ASV), contributed exclusively for the pigmy sperm whales clustering. Evaluating these potential microbial signatures from the K. breviceps specimens, we were able to highlight features evidencing high homology with known marine pathogens, such as Actinobacillus delphinicola, Pasteurella skyensis, Tenacibaculum gallaicum, and Te-nacibaculum soleae, thus suggesting a disease association. These should be further explored as mi-crobiota fingerprints towards cetacean population control and biomonitoring.
Keywords:
oral cavity
; oral microbiome
; metabarcoding
; pygmy sperm whale
; cetaceans
1. Introduction
Fisheries and other human-driven activities, together with climate change and habitat degradation, interfere with the sustainability of aquatic ecosystems and the conservation of marine species [1,2]. Owing to their sensitivity to environmental stressors, marine mammals are often considered ‘ecosystem sentinels’. Being primary or secondary consumers, cetaceans are of particular interest as their condition and well-being reflect those of lower trophic levels in the food chain [3]. Several cetacean species inhabit the Atlantic Iberian waters, whose populations may represent a potentially important tool to obtain information on the bioaccumulation of contaminants in the Iberian marine ecosystem [4,5].
Cetacean strandings in the continental coast of Portugal are frequent. Between the year 2000 and 2016, the national marine animal strandings network reported an average of 236 stranded cetaceans per year, where disease was the attributed cause of death in 3.3% of the analyzed individuals [6], possibly caused by viral or bacterial infections [7,8,9,10].
Within the Kogiidae family, the genus Kogia is currently comprised of only two species, Kogia sima (dwarf sperm whale), and Kogia breviceps (pygmy sperm whale) and represents one of the least known groups of marine cetaceans [11]. Pygmy sperm whales have been observed as solitary animals or in small groups (3-6 individuals). They are slow swimmers, preferring deep waters in tropical to warm temperate zones [12]. Previous studies have identified the feeding habits of these cetaceans, primarily comprising cephalopods, although fish and crustaceans are also part of their diet [13]. In Portugal, the number of observations of live Kogia breviceps specimens is low and the population is categorized as Data Deficient [14]. Between the year 2000 and 2016, four live stranded K. breviceps were assisted by the rehabilitation team in the Portuguese coast [6].
Owing to sampling constraints, studying the microbiome by resorting to non-invasive sampling is crucial to cetacean research. Research currently aims to identify biomarkers to quickly identify unhealthy individuals by assessing disease-associated microbiota alterations [15]. Significantly, pathogenic microbes are often implicated in large stranding events in cetaceans [16,17,18,19,20].
Given the limited data on the pygmy sperm whale and the possible implications of using the microbiome profiles as a biomonitoring tool, a broader knowledge of its microbiome is needed. The few microbiome studies of K. breviceps currently available focus on the gastrointestinal tract [21,22]. Due to its non-invasive nature and possibility of systematic use in either dead or live animals, we assessed the oral cavity of three specimens of K. breviceps stranded in the Atlantic Iberian coast. These specimens were found in a critical clinical state and died before reaching the rehabilitation center. Therefore, our analyses also intend to contextualize the microbial diversity regarding the observation of potentially pathogenic microorganisms, that may have impacted their health status.
2. Materials and Methods
2.1. Sample Collection
Three Kogia breviceps specimens were initially assisted along the western Atlantic Iberian coast, by the Northern Regional Strandings Network. This Regional Strandings network is integrated in the Portuguese Marine Animal Strandings Network, which is coordinated by the Instituto para a Conservação da Natureza e Florestas from Portugal (ICNF) (http://www.icnf.pt/portal/icnf). In the case of the animals found initially alive, refloating was not a possibility due to their critical clinical state, and despite the efforts of the rescuing teams, the animals eventually died before reaching the rehabilitation center.
Following the procedure previously established in Godoy-Vitorino et al. [8], samples were collected from the oral cavity by swabbing the gingival sulcus of the lower and upper jaws with sterile nylon fiber swabs (FLOQSwabTM, Copan). When possible, information regarding the cause of death of the specimens was registered during necropsy procedures [23], and post-mortem analysis for the occurrence of gross pathologies was performed according to standard procedures [24]. All samples were archived in the Marine Animal Tissue Bank (13PT0124/S), recognised by the ICNF, with CITES permit code PT009 to maintain samples. The analysis carried out in the present study focus on the oral microbiome of three pygmy sperm whales (K. breviceps), further including the comparison to the microbial composition of the oral cavity of a previous pool of other Odontoceti cetaceans [9], particularly, 18 common dolphins (Delphinus delphis), 10 striped dolphins (Stenella coeruleoalba) and 10 harbour porpoises (Phocoena phocoena) (Supplementary Table S1).
2.2. Genomic DNA Extraction, PCR Amplifications and Sequencing
The genomic DNA acquired by the oral cavity swabs was extracted using the PureLinkTM Genomic DNA Mini Kit. PCR amplification of the 16S rRNA gene targeting the hypervariable region V4 (forward primer: 5’ GTGCCAGCMGCCGCGGTAA 3’; reverse primer: 5’ GGACTACHVGGGTWTCTAATCC 3’). The generated amplicon covered a region of 251 bp, and the DNA was processed according to Illumina instructions to generate Nextera XT paired-end libraries (2x250 bp).
2.3. Sequencing Raw Data Processing and Data Analysis
Read pairs were trimmed with Sickle [25], to remove adapter and primers sequences, as well as nucleotides corresponding to low-quality base calls (minimum Phred score of 20 and a sliding window of 10% of the read length). The BayesHammer module in SPAdes [26] was used for error correction of the paired-end reads. Read pairs were merged with VSEARCH [27] and prepared with strict quality and size filtering (minimum length of overlap between reads = 20 bp; minimum length of the merged sequence = 200 bp; maximum expected error of 0.5; maximum number of different bases in the overlap = 2) into uniform error-free sequences, standardized to a fixed length of 251 nucleotides. The Quantitative Insights Into Microbial Ecology (QIIME2) software package [28], combined with its Deblur plugin [29], was used for sequence denoising (quality score threshold = 20) and chimera removal, generating the final set of amplicon sequence variants (ASVs). Previous data was reanalyzed to standardize the identifications. To format, manage and manipulate the SILVA database (v138, non-redundant, 99%; [30]) into dereplicated reference sequences matching the locus amplified with the 515f/806r primer pair, the RESCRIPt [31] and feature-classifier plugins of QIIME2 were used, for further usage of this dereplicated database for taxonomic classification of the ASVs, clustered at 99% nucleotide identity. Before the analysis of the microbial profiles, the ASV table was filtered out of unclassified sequences and low-frequency counts (singletons, considered as potential artifacts).
The analysis of the community structure between samples was carried out after performing a total sum scaling normalization, transforming abundances into relative frequency, and rarefaction of the ASV tables to an even sampling depth of 24.030 sequences per sample for ordination analyses, thus eliminating any bias due to differing sampling depth during the sequencing process [32].
To determine if there were characteristic microbial community profiles for each species, profiles from all samples were compared by performing a cluster analysis based on the normalized abundance of each ASV within the samples. Hierarchical cluster analysis was performed using the R [33], via the cluster R package [34]. A Bray–Curtis dissimilarity matrix was created from the proportioned data, and this matrix was used to perform the cluster analysis using the UPGMA method.
The alpha-diversity metrics (richness – the number of ASVs and Shannon diversity index) were estimated with the amp_alphadiv function available from the ampvis2 R package [35], and the differences observed between groups of samples were tested with the Kruskal-Wallis chi-squared test, followed by pairwise Wilcoxon test between groups (kruskal.test and pairwise.wilcox.test functions).
The constrained canonical correspondence analysis (CCA) was performed according to each variable of the metadata collected, by using the cca and anova.cca functions from the vegan R package [36], with 999 permutation testing. The comparisons of the microbial communities by CCA were performed using the ASV table with a square root transformation of the relative abundance (Hellinger transformation performed with the decostand function), to reduce the range of the data and to make it suitable for analysis by linear methods, as described before [9]. Representation of the CCA plot was performed with the amp_ordinate function from the ampvis2 R package [35].
2.4. Identification of Discriminatory Bacterial Fingerprints According to Host Species
The potential signature bacterial genera and ASVs contributing to the CCA clustering profiles were assessed by the linear discriminant analysis (LDA) effect size algorithm (LEfSe). The LEfSe from the MicrobiomeAnalyst web-tool [37] was carried out with the original ASV table, transformed with total sum scaling, to identify variations in the abundance of taxa showing a significant p-value after correction by false discovery rate (FDR-corrected p-value herein, < 0.05). This approach involves the non-parametric factorial Kruskal-Wallis sum-rank test to identify taxa with significant differential abundances, according to the grouping variables of interest, followed by LDA to estimate the effect size of each differentially abundant taxa [38]. The threshold of the LDA score to identify the major bacterial taxa driving the clustering of the samples was set to 2, as previously described [9].
Resorting to pathogen databases [39,40,41] and previous studies on marine mammals [15,16,18,19,42,43,44], genera and ASVs associated with known or possible pathogens were highlighted, aiming to associate specific hits with the confirmed disease status of the cetaceans. This custom database included putative pathogens of any marine mammal body site identified from published studies, while also including any human and animal bacterial pathogens, to account for bacteria not yet identified as disease-causing in marine mammals. BLASTn analysis of the representative sequences of the ASVs was performed against the NCBI 16S rRNA database, to identify the closest homologs of relevant taxa [45].
3. Results
3.1. Structure of the Oral Community of Three Kogia Breviceps Specimens
This study is the first report of the microbial diversity observed in the oral cavity of Kogia breviceps. Despite including only three K. breviceps specimens, whose intrinsic heterogeneity underlies a considerable degree of variability of the assessed microbial profiles, our analysis provided valuable and unprecedented insights on the oral microbiome of pygmy sperm whales. A total of 372,655 MiSeq paired reads were filtered and merged into 283,390 high quality-filtered sequences comprising the V4 region of the 16S rRNA gene, resulting into a total of 433 ASVs (Table 1, Supplementary Figure S1A, Supplementary Table S2).
Sequencing reads from the three sampled pigmy sperm whales were taxonomically classified as 15 phyla, 28 classes, 46 orders, 77 families and 88 genera. From these 433 ASVs, 60 were shared by all pygmy sperm whale specimens, distributed by 8 phyla, 14 classes, 22 orders, 35 families and 34 genera.
The oral microbiome of K. breviceps was dominated by members belonging to the phyla Proteobacteria, Firmicutes, and Bacteroidetes. The overall most abundant classes were those of Clostridia, Gammaproteobacteria, Bacteroidia, Deltaproteobacteria, and Spirochaetia.
3.2. The Variable Composition of the Oral Microbiota Allows Discrimination of the Analyzed Cetacean Species
In 2019, Soares-Castro and collaborators [9] published the microbiome composition from the oral cavity of three other species of Odontoceti cetaceans. Therefore, aiming to highlight the singularities of the microbial community of the sampled pygmy sperm whales, we integrated that data with the oral microbiome of the stranded K. breviceps (Supplementary Table S1). The Bray-Curtis distances between the sampled microbial communities of the cetacean species resulted in the clear clustering of the pigmy sperm whales (Supplementary Figure S1B), hinting at a possible host-specificity exhibited by their oral microbiota.
The sampled bacterial communities were subjected to a Canonical Correspondence Analysis (CCA) to discriminate the biological and ecological factors underlying their variability (CCA is sensitive to the less abundant and unique species in the samples). When considering the samples from K. breviceps, the ordination trend observed was according to the animal phylogeny (Figure 1A), grouping the samples of the same species. From this clustering profile, microbial communities of the oral cavity were different between K. breviceps samples vs. the P. phocoena and the D. delphis-S. coeruleoalba group. The occurrence of mixed groups of cetaceans has been reported, such as the association of specimens from D. delphis and S. coeruleoalba [46,47], which ultimately could lead to a more similar microbiota between animals, as we observed. On the other hand, the pygmy sperm whale does not typically form associations with other cetaceans, thus this microbiota divergence is not surprising and is in agreement with the distant phylogenetic relationship of the family Kogiidae [48].
Furthermore, the oral microbiome associated with K. breviceps was the smallest of the cetacean species here compared, comprised of a total of 433 ASVs (Figure 1B). From a total of 1795 classified ASVs, the four Odontoceti species only shared 117 (around 6.5%). Evidencing the divergence between their communities, the other species shared 290 ASVs (around 16%), which could be influenced by the phylogeny, habitat and food preferences of the pygmy sperm whales.
3.3. Potential Microbial Signatures within the Oral Cavity of Kogia breviceps
From the 433 ASVs detected within the oral cavities of K. breviceps, 60 ASVs were present in all three samples (Figure 2, Supplementary Table S3). Some of these taxa have been previously identified as an abundant bacterial component of the microbiome of marine mammals [9,49,50,51,52,53]. The bacterial genus Psychrobacter (ASV113, with a 95% nucleotide identity corresponding to the deep-sea species Psychrobacter pacificensis) was shared with P. phocoena and S. coeruleoalba, the genus Porphyromonas (ASV881, with a 93% nucleotide identity corresponding to the potential human pathogen Porphyromonas pasteri; [54]) with P. phocoena, the genus Sphingomonas (ASV1644, with 100% nucleotide identity corresponding to the type strain of Sphingomonas aquatica) with D. delphis and S. coeruleoalba, and the family Muribaculaceae (ASV1646, with a lower nucleotide homology – 88%, corresponding to Duncaniella freteri, a disease-associated bacteria found in mice; [55]) shared with D. delphis.
This LEfSe analysis identified a total of 28 ASVs as sole contributors for the host species clustering (p-value < 0.05, detailed in Supplementary Table S4; Table 2). Out of these 28 ASVs, 5 were shared by all pygmy sperm whales (Figure 2), belonging to the genera Phocoenobacter (ASV156), Defluviitaleaceae UCG-011 (ASV251), and Tenacibaculum (ASV1034), and two features with no classification at the genus level, from the Eubacteriales Family XIII. Incertae Sedis (ASV1094 and ASV1665). Additionally, 10 bacterial genera and 57 ASVs were identified as mainly contributing to this cluster, with statistical significance (Supplementary Table S4). Highlighting the taxa that were also shared by all K. breviceps, 28 ASVs were found to be mainly contributing to the pigmy sperm whales cluster.
The taxa identified by the LEfSe analysis showing statistical significance at the genus and ASV level (p-value < 0.05) could be considered as potential microbial signatures of the associated groups, based on the host species variable. This study highlighted 9 ASVs (from which 2 were shared by all K. breviceps specimens) solely contributing to the pigmy sperm whales clustering meeting these criteria belonging to the genera Campylobacter (ASV519, ASV753), Phocoenobacter (ASV156, ASV725, ASV1104, ASV1155), Psychrobacter (ASV888), Tenacibaculum (ASV1034) and Terasakiispira (ASV189). Additionally, 14 ASVs (from which 9 were shared by all K. breviceps specimens) mainly contributing to this cluster could also be considered potential bacterial fingerprints, belonging to the genera Campylobacter (ASV99, ASV635), Desulfoplanes (ASV1255), Desulfovibrio (ASV1007), Oceanispirochaeta (ASV1121), Phocoenobacter (ASV1624), Sediminispirochaeta (ASV849, ASV909), Spirochaeta (ASV148, ASV544, ASV1283), Tenacibaculum (ASV765) and Terasakiispira (ASV205).
3.4. Hints of Disease-Association from Potential Pathogenic Features
The LEfSe analysis also identified potential discriminatory bacterial fingerprints according to the cause of death, pinpointing 4 bacterial genera and 9 ASVs as significant contributors to the Disease cluster, from which, 6 ASVs were not highlighted in the host species analysis (Supplementary Table S4). The ASVs 558 and 946 from the Bacillus genus were identified as potential microbial signatures of the diseased animals, evidencing a nucleotide identity of 100% with, respectively, the coral-associated Bacillus algicola [57], and the virulent-able Bacillus subtilis [58]. Moreover, the genera Bradyrhizobium (ASV239) and Porphyromonas (ASVs 881, 1444, and 1661), specifically P. pasteri, previously associated with an increased decline in human lung function [54], were also considered as potential indicators of disease.
BLASTn was used to assess the homology between the 9 ASV sequences associated with the Disease cluster from the current study and the representative sequences of the OTUs available from a previous study [9]. This comparison allowed us to identify seven common representative sequences, with 100% nucleotide identity observed within the 251 bp comprising the V4 region of the 16S rRNA gene, targeted in the amplicon sequencing approach of the present study, from which four were also previously associated with the health status of cetaceans, specifically contributing to the Disease cluster (Supplementary Table S5).
4. Discussion
There has been an effort to characterize the cetacean microbiota, however, most focus on the most prominent species and readily sampled and/or the gut microbiome. Considering its non-invasiveness and the possibility to apply in dead or live specimens, we assess the oral cavity of three specimens of K. breviceps stranded in the Atlantic Iberian coast. This data was complemented with a previous study in other Odontoceti cetaceans [9], and a clustering analysis of the microbial communities evidenced clear discrimination according to animal phylogeny. The occurrence of mixed groups of cetaceans has been reported, such as the association of specimens from D. delphis and S. coeruleoalba [46,47], which ultimately could lead to a more similar microbiota between animals, as we observed. On the other hand, the pygmy sperm whale is considered to be a solitary cetacean, thus this microbiota divergence is not surprising and is in agreement with the distant phylogenetic relationship of the family Kogiidae.
Focusing on the LEfSe analysis for the host species clustering, 5 ASVs were shared by all K. breviceps specimens and were not detected in the other Odontoceti: the ASV1665, with 94% nucleotide homology corresponding to proteolytic bacteria Anoxynatronum buryatiense, the bacteria Pasteurella skyensis (ASV156; nucleotide identity of 98%), which can cause pasteurellosis, an extremely serious disease in fish [59], and the ASV1034 showing 98% of homology with the fish pathogen Tenacibaculum soleae [56]. Suggesting a possible association with fecal pollution, the ASV251 evidenced a 91% nucleotide identity with Anaerostipes hadrus, a dominant species within the human colonic microbiota [60], and ASV1094 with 93% nucleotide identity corresponding to Ihubacter massiliensis, also isolated from the human gut [61].
Examining the homology results for features contributing, but not exclusively, to the clustering of the pigmy sperm whales, the two species of Arcobacter correlate with high levels of fecal pollution in waters [62], with ASV99 being associated with Arcobacter canalis (nucleotide identity of 90%), previously isolated from a water canal contaminated with urban sewage [63], and with a potential association with animal illness, as ASV1160 evidenced 98% nucleotide homology with Arcobacter venerupis, from which virulence genes have been identified [44]. The ASV861 was associated with Aminipila butyrica (96% of nucleotide identity), a bacterium previously isolated from waste samples.
The genus Fusibacter was also potentially represented, with ASV171 being associated with the Fusibacter paucivorans (88% identity), isolated from, ASV356 with Fusibacter tunisiensis (93% identity), and ASV1128 with Fusibacter fontis (93% identity). These species were, respectively, previously linked to an oil-producing well, the treatment of olive-mill wastewater, and a mesothermal spring. Interestingly, ASV527 could also be linked to crude oil, as it evidenced 96% identity with the sulfate-reducing bacterium Desulfotignum toluenicum.
Other ASVs evidenced higher sequence identity to species previously isolated from marine sediments, such as Anaerotignum propionicum (ASV81, 92%; ASV1776, 90%), Anaerovorax odorimutans (ASV1774, 96%), Cellulosilyticum lentocellum (ASV1388, 87%), Marinifilum fragile (ASV141, 94%), Marinospirillum alkaliphilum (ASV205, 89%), Oceanispirochaeta sediminicola (ASVs 148, 1121 and 1283, 92-94%), Petrocella atlantisensis (ASV757, 92%) and Pseudodesulfovibrio indicus (ASV1007, 98%). Moreover, similarity to species isolated from coral reefs, as the ASV1098 showing 89% identity with Aureibacter tunicatorum, or from marine mollusks, as the ASV603 with 90% similarity to Oceaniserpentilla haliotis, was also observed.
Given the disease-association of all the assayed K. breviceps specimens (Table 1), our custom database was surveyed and some of the features highlighted in the pigmy sperm whales specimens are of potential pathogens in marine environments, as the aforementioned ASV156, and the ASVs 725, 1104, 1155 and 1624, evidencing a nucleotide identity from 96 to 99% with the fish pathogen Pasteurella skyensis, and ASV909 with 92% homology with Porphyromonas pasteri. Another example was ASV383, exclusively found in pigmy sperm whales in this study, thus a noteworthy feature potentially associated with disease in these cetaceans, as its closest homolog was a reported pathogen of marine mammals, with 98% nucleotide homology with Actinobacillus delphinicola [16].
Two ASVs, solely contributing to the pigmy sperm whale cluster, whose closest homologs belong to the genera Arcobacter and Campylobacter (ASVs 519 and 753), belonging to the Campylobacteraceae family, have been linked to human and animal disease [64,65] but have also been found in marine sediments [66] and the dental plaque of healthy captive delphinids [67]. Previous research has suggested that Arcobacter may be better adapted than Campylobacter to survive in aquatic environments, and, as previously discussed, is associated with fecal pollution [62].
Bacterial taxa, for which available characterization studies are mainly in a human infection context, may predispose interpretations regarding the health status of cetaceans. For example, Comamonas kerstersii, associated with severe diseases in humans, like abdominal infection and bacteremia [68], evidenced a 99% identity with ASV1298. The feature ASV635 showed the highest identity with Helicobacter bilis, an opportunistic pathogen in humans, triggering multiple diseases [69]. The presence of the ASVs 777 and 1264, with 96% similarity with an animal [70] pathogen, Streptobacillus moniliformis, is also noteworthy, along with ASVs 111 and 458, with 95-98% nucleotide similarity with Sneathia sanguinegens, and ASV948, with 92% identity with Treponema medium, other examples of human pathogens.
The ASV1252 evidenced an identity of 97% with Mycoplasma mucosicanis, and the ASV1410 had an identity of 98% with Mycoplasma opalescens. Mycoplama spp. were associated with respiratory disease in fish and marine mammals, particularly in pinnipeds [16,71]. The ASV883, classified as Mucispirillum schaedleri (with a nucleotide identity of 100%), is a suggested pathobiont, a commensal that plays a role in the disease of the several mammals described to inhabit [72]. Also in cetaceans, the opportunistic pathogen Pseudomonas aeruginosa (ASV486; nucleotide identity of 100%) is associated with pneumonia, one of the most common causes of morbidity [73].
Moreover, several members of the genus Tenacibaculum, including the closest homologs here reported for ASVs 765 (nucleotide identity of 99% with Tenacibaculum gallaicum) and 1034 (nucleotide identity of 96% with Tenacibaculum soleae), are linked to the presence of disease (tenacibaculosis) and high mortality rate in multiple fish [41,56,74] and few bivalves [75,76]. As suggested in other studies, shrimp and shellfish could be an important reservoir and source of infection of these bacteria [38,77], and one could postulate that the feeding habits of K. breviceps played a role in their death. However, care must be taken when associating these and other features with potential microbial signatures of the compromised health status of the pigmy sperm whales, as no healthy counterpart was assayed and the pathogenic character may be due to a compromised immune system of the assayed specimens. As all K. breviceps described in this preliminary study correspond to disease strandings and death before reaching the rehabilitation center, future comparison of the oral cavity microbiota of pygmy sperm whale healthy individuals would provide valuable insights for the identification of disease markers in this species. The potential role of the here emphasized ASVs in the disease of K. breviceps should, nevertheless, be further explored as potential fingerprints to develop (i) specific diagnostic assays for cetacean population conservation and (ii) bio-monitoring approaches to assess the health of marine ecosystems from the Iberian Atlantic basin, using cetaceans as bioindicators aiming to facilitate population control and monitoring towards conservation.
5. Conclusions
This is the first study of the oral microbiota of Kogia breviceps specimens. Despite possessing the smallest oral microbiome of the studied cetacean species, samples from K. breviceps showed a less variable average number of ASVs and a microbiome sharing more features. Even though this may be biased by the low number of sampled specimens, it was possible to highlight the variation in the microbial community according to host phylogeny, host biogeography, feeding, deep diving and social habits, thereby complementing and agreeing with previous studies [9,68,69,70]. From the total 1795 ASVs detected in this study, the four Odontoceti species shared 117 (around 6.5%), whereas the species D. delphis, S. coeruleoalba, and P. phocoena shared 517 ASVs (around 29%), evidencing the divergence between their communities, which could be owed to the distant phylogenetic relationship of the Kogia species [48], but also potentially related to the solitary social habits of the pygmy sperm whale. As only the bacterial taxa showing significant differential abundance at the genus and ASV levels were considered as potential microbial signatures for the respective grouping variable, this preliminary study identified 22 ASVs belonging to the genera Campylobacter (4 ASVs), Desulfoplanes (1 ASV), Desulfovibrio (1ASV), Oceanispirochaeta (1 ASV), Phocoenobacter (5 ASVs), Psychrobacter (1 ASV), Sediminispirochaeta (2 ASVs), Spirochaeta (3 ASVs), Tenacibaculum (2 ASVs) and Terasakiispira (2 ASVs) as potential fingerprints of the Kogia breviceps specimens. A comparison regarding the cause of death of the cetaceans allowed us to identify 9 ASVs associated with the Disease cluster, belonging to the genera Bacillus (2 ASVs), Bradyrhizobium (1 ASV), Porphyromonas (4 ASVs) and Oceanivirga (2 ASVs). Furthermore, features highlighted from our analysis for the Kogia breviceps specimens were surveyed for potential marine pathogens, aiming to associate them with the health status of these cetaceans.
The knowledge regarding the role of the species detected in the oral cavity of cetaceans (and other mammals) is scarce, highlighting our insufficient familiarity when it comes to marine mammal-hosted microbial communities and the need for continued study of the relationships between these animals and their microbiota. These are pioneering findings that could serve as the basis for comparisons in non-invasive biomonitoring studies to assess health changes or deviations.
Supplementary Materials
The essential data generated and analyzed during this study is included in the Supplementary Material with doi: https://doi.org/10.34622/datarepositorium/VEVBVI.
Author Contributions
CE: PMS designed the study, CE, MF, MS were involved in animal rescue and swab collection. CSM, PS-C, PMS analyzed the sequencing data. CSM, PS-C, CE, PMS drafted the manuscript. All authors read, revised, and approved the manuscript.
Funding
This work was supported by the project CetSenti RECI/AAGGLO/0470/2012, and by the GenomePT project (POCI-01-0145-FEDER-022184), supported by COMPETE 2020 - Operational Programme for Competitiveness and Internationalisation (POCI), Lisboa Portugal Regional Operational Programme (Lisboa2020), Algarve Portugal Regional Operational Programme (CRESC Algarve2020), under the PORTUGAL 2020 Partnership Agreement, funded by national funds through the Fundação para a Ciência e a Tecnologia (FCT) I.P. and the European Regional Development Fund (ERDF), through the COMPETE2020-Programa Operacional Competitividade e Internacionalização (POCI). Thanks are due to Fundação para a Ciência e a Tecnologia (FCT) I.P. for financial support to CBMA (UIDB/04050/2020), Associate Laboratory ARNET (LA/P/0069/2020), and to CESAM (UIDP/50017/2020, UIDB/50017/2020 and LA/P/0094/2020): https://doi.org/10.54499/UIDB/04050/2020, https://doi.org/10.54499/LA/P/0069/2020, https://doi.org/10.54499/UIDP/50017/2020, https://doi.org/10.54499/UIDB/50017/2020, https://doi.org/10.54499/LA/P/0094/2020
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The sequences datasets used in this study have been deposited in NCBI submission portal with the BioProject database ID PRJNA1096153. The datasets of a previous study in other three cetacean species [9] are available in the BioProject database ID PRJNA494623, associated with the SRA accession numbers ranging from SRR7963801 to SRR7963838.
Acknowledgments
The authors would like to acknowledge to Jörg Becker and João Sobral for the sequencing services provided at the Instituto Gulbenkian de Ciência.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Fleming, L.E.; Broad, K.; Clement, A.; Dewailly, E.; Elmir, S.; Knap, A.; Pomponi, S.A.; Smith, S.; Solo Gabriele, H.; Walsh, P. Oceans and Human Health: Emerging Public Health Risks in the Marine Environment. Mar Pollut Bull 2006, 53, 545–560. [CrossRef]
- Halpern, B.S.; Walbridge, S.; Selkoe, K.A.; Kappel, C. V.; Micheli, F.; D’Agrosa, C.; Bruno, J.F.; Casey, K.S.; Ebert, C.; Fox, H.E.; et al. A Global Map of Human Impact on Marine Ecosystems. Science (1979) 2008, 319, 948–952. [CrossRef]
- Bossart, G.D. Marine Mammals as Sentinel Species for Oceans and Human Health. Vet Pathol 2011, 48, 676–690. [CrossRef]
- Ferreira, M.; Monteiro, S.S.; Torres, J.; Oliveira, I.; Sequeira, M.; López, A.; Vingada, J.; Eira, C. Biological Variables and Health Status Affecting Inorganic Element Concentrations in Harbour Porpoises (Phocoena Phocoena) from Portugal (Western Iberian Peninsula). Environmental Pollution 2016, 210, 293–302. [CrossRef]
- Monteiro, S.S.; Torres, J.; Ferreira, M.; Marçalo, A.; Nicolau, L.; Vingada, J. V.; Eira, C. Ecological Variables Influencing Trace Element Concentrations in Bottlenose Dolphins (Tursiops Truncatus, Montagu 1821) Stranded in Continental Portugal. Science of The Total Environment 2016, 544, 837–844. [CrossRef]
- Vingada, J.; Eira, C. Conservation of Cetaceans and Seabirds in Continental Portugal. The LIFE+ MarPro Project. Final Project Report NAT/PT/00038; 2018;
- Bento, M.C.; Canha, R.; Eira, C.; Vingada, J.; Nicolau, L.; Ferreira, M.; Domingo, M.; Tavares, L.; Duarte, A. Herpesvirus Infection in Marine Mammals: A Retrospective Molecular Survey of Stranded Cetaceans in the Portuguese Coastline. Infection, Genetics and Evolution 2019, 67, 222–233. [CrossRef]
- Bento, M.C.R. de M.; Eira, C.I.C.S.; Vingada, J.V.; Marçalo, A.L.; Ferreira, M.C.T.; Fernandez, A.L.; Tavares, L.M.M.; Duarte, A.I.S.P. New Insight into Dolphin Morbillivirus Phylogeny and Epidemiology in the Northeast Atlantic: Opportunistic Study in Cetaceans Stranded along the Portuguese and Galician Coasts. BMC Vet Res 2016, 12, 1–12. [CrossRef]
- Soares-Castro, P.; Araújo-Rodrigues, H.; Godoy-Vitorino, F.; Ferreira, M.; Covelo, P.; López, A.; Vingada, J.; Eira, C.; Santos, P.M. Microbiota Fingerprints within the Oral Cavity of Cetaceans as Indicators for Population Biomonitoring. Scientific Reports 2019 9:1 2019, 9, 1–15. [CrossRef]
- Godoy-Vitorino, F.; Rodriguez-Hilario, A.; Alves, A.L.; Gonçalves, F.; Cabrera-Colon, B.; Mesquita, C.S.; Soares-Castro, P.; Ferreira, M.; Marçalo, A.; Vingada, J.; et al. The Microbiome of a Striped Dolphin (Stenella Coeruleoalba) Stranded in Portugal. Res Microbiol 2017, 168, 85–93. [CrossRef]
- Chivers, S.J.; Leduc, R.G.; Robertson, K.M.; Barros, N.B.; Dizon, A.E. Genetic Variation of Kogia Spp. with Preliminary Evidence for Two Species of Kogia Sima. Mar Mamm Sci 2005, 21, 619–634. [CrossRef]
- Hodge, L.E.W.; Baumann-Pickering, S.; Hildebrand, J.A.; Bell, J.T.; Cummings, E.W.; Foley, H.J.; McAlarney, R.J.; McLellan, W.A.; Pabst, D.A.; Swaim, Z.T.; et al. Heard but Not Seen: Occurrence of Kogia Spp. along the Western North Atlantic Shelf Break. Mar Mamm Sci 2018, 34, 1141–1153. [CrossRef]
- Moura, J.F.; Acevedo-Trejos, E.; Tavares, D.C.; Meirelles, A.C.O.; Silva, C.P.N.; Oliveira, L.R.; Santos, R.A.; Wickert, J.C.; Machado, R.; Siciliano, S.; et al. Stranding Events of Kogia Whales along the Brazilian Coast. PLoS One 2016, 11, e0146108. [CrossRef]
- Ferreira, M.; Eira, C.; López, A.; Sequeira, M. Kogia Breviceps Cachalote-Pigmeu. Livro Vermelho Dos Mamíferos de Portugal Continental. In; Mathias, M.D.L., Fonseca, C., Rodrigues, L., Grilo, C., Lopes-Fernandes, M., M. Palmeirim, J., Santos-Reis, M., Alves, P.C., Cabral, J.A., Ferreira, M., Mira, A., Eira, C., Negrões, N., Paupério, J., Pita, R., Rainho, A., Rosalino, L.M., Tapisso, J.T., Vingada, J., Eds.; FCiências.ID / ICNF, 2023; pp. 1–371 ISBN 978-989-53724-1-6.
- Sehnal, L.; Brammer-Robbins, E.; Wormington, A.M.; Blaha, L.; Bisesi, J.; Larkin, I.; Martyniuk, C.J.; Simonin, M.; Adamovsky, O. Microbiome Composition and Function in Aquatic Vertebrates: Small Organisms Making Big Impacts on Aquatic Animal Health. Front Microbiol 2021, 12, 567408. [CrossRef]
- Waltzek, T.B.; Cortés-Hinojosa, G.; Wellehan, J.F.X.; Gray, G.C. Marine Mammal Zoonoses: A Review of Disease Manifestations. Zoonoses Public Health 2012, 59, 521–535. [CrossRef]
- Li, C.; Tan, X.; Bai, J.; Xu, Q.; Liu, S.; Guo, W.; Yu, C.; Fan, G.; Lu, Y.; Zhang, H.; et al. A Survey of the Sperm Whale (Physeter Catodon) Commensal Microbiome. PeerJ 2019, 2019, e7257. [CrossRef]
- Marón, C.F.; Kohl, K.D.; Chirife, A.; Di Martino, M.; Fons, M.P.; Navarro, M.A.; Beingesser, J.; McAloose, D.; Uzal, F.A.; Dearing, M.D.; et al. Symbiotic Microbes and Potential Pathogens in the Intestine of Dead Southern Right Whale (Eubalaena Australis) Calves. Anaerobe 2019, 57, 107–114. [CrossRef]
- Terracciano, G.; Fichi, G.; Comentale, A.; Ricci, E.; Mancusi, C.; Perrucci, S. Dolphins Stranded along the Tuscan Coastline (Central Italy) of the “Pelagos Sanctuary”: A Parasitological Investigation. Pathogens 2020, Vol. 9, Page 612 2020, 9, 612. [CrossRef]
- Marangi, M.; Airoldi, S.; Beneduce, L.; Zaccone, C. Wild Whale Faecal Samples as a Proxy of Anthropogenic Impact. Scientific Reports 2021 11:1 2021, 11, 1–11. [CrossRef]
- Erwin, P.M.; Rhodes, R.G.; Kiser, K.B.; Keenan-Bateman, T.F.; McLellan, W.A.; Pabst, D.A. High Diversity and Unique Composition of Gut Microbiomes in Pygmy (Kogia breviceps) and Dwarf (K. sima) Sperm Whales. Scientific Reports 2017 7:1 2017, 7, 1–11. [CrossRef]
- Denison, E.R.; Rhodes, R.G.; McLellan, W.A.; Pabst, D.A.; Erwin, P.M. Host Phylogeny and Life History Stage Shape the Gut Microbiome in Dwarf (Kogia Sima) and Pygmy (Kogia Breviceps) Sperm Whales. Scientific Reports 2020 10:1 2020, 10, 1–13. [CrossRef]
- Kuiken, T.; García-Hartmann, M. Dissection Techniques and Tissue Sampling. Proceedings of the Proceedings of the first ECS workshop on Cetacean pathology, 1991.
- Pugliares, K.R.; Bogomolni, A.L.; Touhey, K.M.; Herzig, S.M.; Harry, C.T.; Moore, M.J. Marine Mammal Necropsy : An Introductory Guide for Stranding Responders and Field Biologists. 2007. [CrossRef]
- Joshi, N.; Fass, J. Sickle: A Windowed Adaptive Trimming for Fastq Files Using Quality Available online: https://github.com/najoshi/sickle (accessed on 13 May 2024).
- Nikolenko, S.I.; Korobeynikov, A.I.; Alekseyev, M.A. BayesHammer: Bayesian Clustering for Error Correction in Single-Cell Sequencing. BMC Genomics 2013, 14, 1–11. [CrossRef]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A Versatile Open Source Tool for Metagenomics. PeerJ 2016, 2016, e2584. [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, Interactive, Scalable and Extensible Microbiome Data Science Using QIIME 2. Nature Biotechnology 2019 37:8 2019, 37, 852–857. [CrossRef]
- Amir, A.; McDonald, D.; Navas-Molina, J.A.; Kopylova, E.; Morton, J.T.; Xu, Z.Z.; Kightley, E.P.; Thompson, L.R.; Hyde, E.R.; Gonzalez, A.; et al. Deblur Rapidly Resolves Single-Nucleotide Community Sequence Patterns. mSystems 2017, 2. [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res 2013, 41, D590–D596. [CrossRef]
- Robeson, M.S.; O’Rourke, D.R.; Kaehler, B.D.; Ziemski, M.; Dillon, M.R.; Foster, J.T.; Bokulich, N.A. RESCRIPt: Reproducible Sequence Taxonomy Reference Database Management. PLoS Comput Biol 2021, 17, e1009581. [CrossRef]
- Chao, A.; Jost, L. Coverage-Based Rarefaction and Extrapolation: Standardizing Samples by Completeness Rather than Size. Ecology 2012, 93, 2533–2547. [CrossRef]
- .
- Maechler, M.; Rousseeuw, P.; Struyf, A.; Hubert, M.; Hornik, K. Cluster: Cluster Analysis Basics and Extensions. 2023.
- Andersen, K.S.; Kirkegaard, R.H.; Karst, S.M.; Albertsen, M. Ampvis2: An R Package to Analyse and Visualise 16S RRNA Amplicon Data. bioRxiv 2018, 299537. [CrossRef]
- Dixon, P. VEGAN, a Package of R Functions for Community Ecology. Journal of Vegetation Science 2003, 14, 927–930. [CrossRef]
- Dhariwal, A.; Chong, J.; Habib, S.; King, I.L.; Agellon, L.B.; Xia, J. MicrobiomeAnalyst: A Web-Based Tool for Comprehensive Statistical, Visual and Meta-Analysis of Microbiome Data. Nucleic Acids Res 2017, 45, W180–W188. [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic Biomarker Discovery and Explanation. Genome Biol 2011, 12, 1–18. [CrossRef]
- Amos, B.; Aurrecoechea, C.; Barba, M.; Barreto, A.; Basenko, E.Y.; Bażant, W.; Belnap, R.; Blevins, A.S.; Böhme, U.; Brestelli, J.; et al. VEuPathDB: The Eukaryotic Pathogen, Vector and Host Bioinformatics Resource Center. Nucleic Acids Res 2022, 50, D898–D911. [CrossRef]
- Venn-Watson, S.; Smith, C.R.; Jensen, E.D. Primary Bacterial Pathogens in Bottlenose Dolphins Tursiops Truncatus: Needles in Haystacks of Commensal and Environmental Microbes. Dis Aquat Organ 2008, 79, 87–93. [CrossRef]
- Martins, P.; Cleary, D.F.R.; Pires, A.C.C.; Rodrigues, A.M.; Quintino, V.; Calado, R.; Gomes, N.C.M. Molecular Analysis of Bacterial Communities and Detection of Potential Pathogens in a Recirculating Aquaculture System for Scophthalmus Maximus and Solea Senegalensis. PLoS One 2013, 8, e80847. [CrossRef]
- Apprill, A.; Miller, C.A.; Moore, M.J.; Durban, J.W.; Fearnbach, H.; Barrett-Lennard, L.G. Extensive Core Microbiome in Drone-Captured Whale Blow Supports a Framework for Health Monitoring. mSystems 2017, 2. [CrossRef]
- Raverty, S.A.; Rhodes, L.D.; Zabek, E.; Eshghi, A.; Cameron, C.E.; Hanson, M.B.; Schroeder, J.P. Respiratory Microbiome of Endangered Southern Resident Killer Whales and Microbiota of Surrounding Sea Surface Microlayer in the Eastern North Pacific. Scientific Reports 2017 7:1 2017, 7, 1–12. [CrossRef]
- Jurelevicius, D.; Cotta, S.R.; Montezzi, L.F.; Dias, A.C.F.; Mason, O.U.; Picão, R.C.; Jansson, J.K.; Seldin, L. Enrichment of Potential Pathogens in Marine Microbiomes with Different Degrees of Anthropogenic Activity. Environmental Pollution 2021, 268, 115757. [CrossRef]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and Applications. BMC Bioinformatics 2009, 10, 1–9. [CrossRef]
- Frantzis, A.; Herzing, D.L. Mixed-Species Associations of Striped Dolphins (Stenella Coeruleoalba), Short-Beaked Common Dolphins (Delphinus Delphis), and Risso’s Dolphins (Grampus Griseus) in the Gulf of Corinth (Greece, Mediterranean Sea). Aquat Mamm 2002, 28, 188–197.
- Quérouil, S.; Silva, M.A.; Cascão, I.; Magalhães, S.; Seabra, M.I.; Machete, M.A.; Santos, R.S. Why Do Dolphins Form Mixed-Species Associations in the Azores? Ethology 2008, 114, 1183–1194. [CrossRef]
- McGowen, M.R.; Spaulding, M.; Gatesy, J. Divergence Date Estimation and a Comprehensive Molecular Tree of Extant Cetaceans. Mol Phylogenet Evol 2009, 53, 891–906. [CrossRef]
- Apprill, A.; Robbins, J.; Eren, A.M.; Pack, A.A.; Reveillaud, J.; Mattila, D.; Moore, M.; Niemeyer, M.; Moore, K.M.T.; Mincer, T.J. Humpback Whale Populations Share a Core Skin Bacterial Community: Towards a Health Index for Marine Mammals? PLoS One 2014, 9, e90785. [CrossRef]
- Chiarello, M.; Villéger, S.; Bouvier, C.; Auguet, J.C.; Bouvier, T. Captive Bottlenose Dolphins and Killer Whales Harbor a Species-Specific Skin Microbiota That Varies among Individuals. Scientific Reports 2017 7:1 2017, 7, 1–12. [CrossRef]
- Robles-Malagamba, M.J.; Walsh, M.T.; Ahasan, M.S.; Thompson, P.; Wells, R.S.; Jobin, C.; Fodor, A.A.; Winglee, K.; Waltzek, T.B. Characterization of the Bacterial Microbiome among Free-Ranging Bottlenose Dolphins (Tursiops Truncatus). Heliyon 2020, 6, e03944. [CrossRef]
- Toro-Valdivieso, C.; Toro, F.; Stubbs, S.; Castro-Nallar, E.; Blacklaws, B. Patterns of the Fecal Microbiota in the Juan Fernández Fur Seal (Arctocephalus Philippii). Microbiologyopen 2021, 10, e1215. [CrossRef]
- Glaeser, S.P.; Silva, L.M.R.; Prieto, R.; Silva, M.A.; Franco, A.; Kämpfer, P.; Hermosilla, C.; Taubert, A.; Eisenberg, T. A Preliminary Comparison on Faecal Microbiomes of Free-Ranging Large Baleen (Balaenoptera Musculus, B. Physalus, B. Borealis) and Toothed (Physeter Macrocephalus) Whales. Microb Ecol 2022, 83, 18–33. [CrossRef]
- Webb, K.; Zain, N.M.M.; Stewart, I.; Fogarty, A.; Nash, E.F.; Whitehouse, J.L.; Smyth, A.R.; Lilley, A.K.; Knox, A.; Williams, P.; et al. Porphyromonas Pasteri and Prevotella Nanceiensis in the Sputum Microbiota Are Associated with Increased Decline in Lung Function in Individuals with Cystic Fibrosis. J Med Microbiol 2022, 71, 001481. [CrossRef]
- Forster, S.C.; Clare, S.; Beresford-Jones, B.S.; Harcourt, K.; Notley, G.; Stares, M.D.; Kumar, N.; Soderholm, A.T.; Adoum, A.; Wong, H.; et al. Identification of Gut Microbial Species Linked with Disease Variability in a Widely Used Mouse Model of Colitis. Nature Microbiology 2022 7:4 2022, 7, 590–599. [CrossRef]
- López, J.R.; Piñeiro-Vidal, M.; García-Lamas, N.; de la Herran, R.; Navas, J.I.; Hachero-Cruzado, I.; Santos, Y. First Isolation of Tenacibaculum Soleae from Diseased Cultured Wedge Sole, Dicologoglossa Cuneata (Moreau), and Brill, Scophthalmus Rhombus (L.). J Fish Dis 2010, 33, 273–278. [CrossRef]
- Sabdono, A.; Radjasa, O.K. Anti-Bacterial Property of a Coral-Associated Bacterium Bacillus Sp. against Coral Pathogenic Bbd (Black Band Disease). J. Coastal Dev 2006, 9, 175–182.
- Gu, H.J.; Sun, Q.L.; Luo, J.C.; Zhang, J.; Sun, L. A First Study of the Virulence Potential of a Bacillus Subtilis Isolate From Deep-Sea Hydrothermal Vent. Front Cell Infect Microbiol 2019, 9. [CrossRef]
- Birkbeck, T.H.; Laidler, L.A.; Grant, A.N.; Cox, D.I. Pasteurella Skyensis Sp. Nov., Isolated from Atlantic Salmon (Salmo Salar L.). Int J Syst Evol Microbiol 2002, 52, 699–704. [CrossRef]
- Allen-Vercoe, E.; Daigneault, M.; White, A.; Panaccione, R.; Duncan, S.H.; Flint, H.J.; O’Neal, L.; Lawson, P.A. Anaerostipes Hadrus Comb. Nov., a Dominant Species within the Human Colonic Microbiota; Reclassification of Eubacterium Hadrum Moore et al. 1976. Anaerobe 2012, 18, 523–529. [CrossRef]
- Ndongo, S.; Lagier, J.C.; Fournier, P.E.; Raoult, D.; Khelaifia, S. “Ihubacter Massiliensis”: A New Bacterium Isolated from the Human Gut. New Microbes New Infect 2016, 13, 104–105. [CrossRef]
- Collado, L.; Inza, I.; Guarro, J.; Figueras, M.J. Presence of Arcobacter Spp. in Environmental Waters Correlates with High Levels of Fecal Pollution. Environ Microbiol 2008, 10, 1635–1640. [CrossRef]
- Pérez-Cataluña, A.; Salas-Massó, N.; Figueras, M.J. Arcobacter Canalis Sp. Nov., Isolated from a Water Canal Contaminated with Urban Sewage. Int J Syst Evol Microbiol 2018, 68, 1258–1264. [CrossRef]
- Lee, C.; Agidi, S.; Marion, J.W.; Lee, J. Arcobacter in Lake Erie Beach Waters: An Emerging Gastrointestinal Pathogen Linked with Human-Associated Fecal Contamination. Appl Environ Microbiol 2012, 78, 5511–5519. [CrossRef]
- Stoddard, R.A.; Atwill, E.R.; Gulland, F.M.D.; Miller, M.A.; Dabritz, H.A.; Paradies, D.M.; Worcester, K.R.; Jang, S.; Lawrence, J.; Byrne, B.A.; et al. Risk Factors for Infection with Pathogenic and Antimicrobial-Resistant Fecal Bacteria in Northern Elephant Seals in California. Public Health Rep 2008, 123, 360–370. [CrossRef]
- Fera, M.T.; Maugeri, T.L.; Gugliandolo, C.; Beninati, C.; Giannone, M.; La Camera, E.; Carbone, M. Detection of Arcobacter Spp. in the Coastal Environment of the Mediterranean Sea. Appl Environ Microbiol 2004, 70, 1271–1276. [CrossRef]
- Goldman, C.G.; Matteo, M.J.; Loureiro, J.D.; Almuzara, M.; Barberis, C.; Vay, C.; Catalano, M.; Heredia, S.R.; Mantero, P.; Boccio, J.R.; et al. Novel Gastric Helicobacters and Oral Campylobacters Are Present in Captive and Wild Cetaceans. Vet Microbiol 2011, 152, 138–145. [CrossRef]
- Almuzara, M.N.; Cittadini, R.; Ocampo, C.V.; Bakai, R.; Traglia, G.; Ramirez, M.S.; Del Castillo, M.; Vay, C.A. Intra-Abdominal Infections Due to Comamonas Kerstersii. J Clin Microbiol 2013, 51, 1998–2000. [CrossRef]
- Peng, W.; Zhao, X.; Li, X. Helicobacter Bilis Contributes to the Occurrence of Inflammatory Bowel Disease by Inducing Host Immune Disorders. Biomed Res Int 2022, 2022, 1837850. [CrossRef]
- Simpson, V.R.; Davison, N.J.; Dagleish, M.P. Causes of Mortality and Lesions Observed Post Mortem in European Moles (Talpa Europaea) in Cornwall, South-West England: Disease in Wildlife or Exotic Species. J Comp Pathol 2019, 167, 18–25. [CrossRef]
- Sarzosa, M.S.; Duignan, P.; Derango, E.J.; Field, C.; Ríos, C.; Sanchez, S.; Espinoza, E.; Loyola, A.; Rueda, D.; Páez-Rosas, D. Occurrence of Mycoplasmas in Galapagos Sea Lions (Zalophus Wollebaeki) and Their Association with Other Respiratory Pathogens. J Wildl Dis 2021, 57, 623–627. [CrossRef]
- Herp, S.; Durai Raj, A.C.; Salvado Silva, M.; Woelfel, S.; Stecher, B. The Human Symbiont Mucispirillum Schaedleri: Causality in Health and Disease. Med Microbiol Immunol 2021, 210, 173–179. [CrossRef]
- Venn-Watson, S.; Daniels, R.; Smith, C. Thirty Year Retrospective Evaluation of Pneumonia in a Bottlenose Dolphin Tursiops Truncatus Population. Dis Aquat Organ 2012, 99, 237–242. [CrossRef]
- Fernández-Álvarez, C.; Santos, Y. Identification and Typing of Fish Pathogenic Species of the Genus Tenacibaculum. Appl Microbiol Biotechnol 2018, 102, 9973–9989. [CrossRef]
- Nowlan, J.P.; Lumsden, J.S.; Russell, S. Advancements in Characterizing Tenacibaculum Infections in Canada. Pathogens 2020, Vol. 9, Page 1029 2020, 9, 1029. [CrossRef]
- Burioli, E.A.V.; Varello, K.; Trancart, S.; Bozzetta, E.; Gorla, A.; Prearo, M.; Houssin, M. First Description of a Mortality Event in Adult Pacific Oysters in Italy Associated with Infection by a Tenacibaculum Soleae Strain. J Fish Dis 2018, 41, 215–221. [CrossRef]
- Rungrassamee, W.; Klanchui, A.; Maibunkaew, S.; Chaiyapechara, S.; Jiravanichpaisal, P.; Karoonuthaisiri, N. Characterization of Intestinal Bacteria in Wild and Domesticated Adult Black Tiger Shrimp (Penaeus Monodon). PLoS One 2014, 9, e91853. [CrossRef]
- Nelson, T.M.; Rogers, T.L.; Brown, M. V. The Gut Bacterial Community of Mammals from Marine and Terrestrial Habitats. PLoS One 2013, 8. [CrossRef]
- Archie, E.A.; Tung, J. Social Behavior and the Microbiome. Curr Opin Behav Sci 2015, 6, 28–34. [CrossRef]
- Delport, T.C.; Power, M.L.; Harcourt, R.G.; Webster, K.N.; Tetu, S.G. Colony Location and Captivity Influence the Gut Microbial Community Composition of the Australian Sea Lion (Neophoca Cinerea). Appl Environ Microbiol 2016, 82, 3440–3449. [CrossRef]
Figure 1.
Comparison of the microbial communities of the oral cavity of cetaceans according to the species of the sampled animal. A. The Canonical Correspondence Analysis (CCA, p-value = 0.001) was performed after subsampling the ASV table to even sequencing depth and at the bacterial species level, with Hellinger transformation of the abundances. Similar results were obtained when grouping the ASVs by bacterial genera. The color frames group the samples according to the constrained variable. The major contributions of the host species variable are shown as % in the first and second components of the CCA plot (CCA1 and CCA2, respectively). B. Representation of the number of total ASVs in each cetacean group and the number of ASVs shared between groups. Kogia breviceps (n=3), Delphinus delphis (n = 18), Phocoena phocoena (n = 10), and Stenella coeruleoalba (n = 10).
Figure 1.
Comparison of the microbial communities of the oral cavity of cetaceans according to the species of the sampled animal. A. The Canonical Correspondence Analysis (CCA, p-value = 0.001) was performed after subsampling the ASV table to even sequencing depth and at the bacterial species level, with Hellinger transformation of the abundances. Similar results were obtained when grouping the ASVs by bacterial genera. The color frames group the samples according to the constrained variable. The major contributions of the host species variable are shown as % in the first and second components of the CCA plot (CCA1 and CCA2, respectively). B. Representation of the number of total ASVs in each cetacean group and the number of ASVs shared between groups. Kogia breviceps (n=3), Delphinus delphis (n = 18), Phocoena phocoena (n = 10), and Stenella coeruleoalba (n = 10).
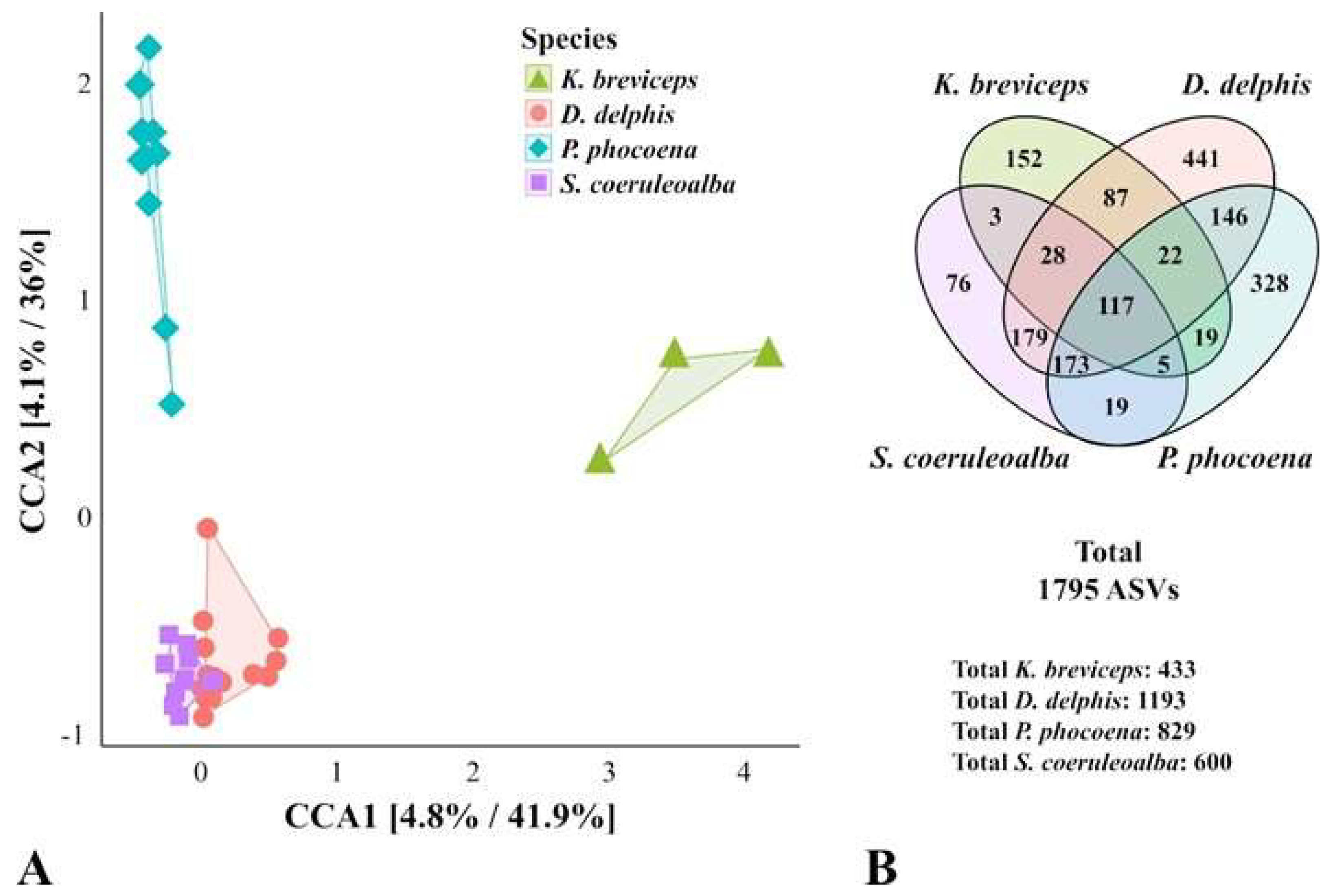
Figure 2.
Composition of the microbial communities within the oral cavities of K. breviceps. A. Representation of the number of total ASVs in each sampled K. breviceps and the number of ASVs shared between the three sampled specimens, highlighting the ones present in all K. breviceps samples and how they distribute in the other assayed Odontoceti species. B. Relative abundance between the assayed Odontoceti species of the 60 ASVs shared by all K. breviceps specimens. The significance of potential microbial signatures (highlighted with “*”, p-value < 0.05 at the genus and ASV level) was identified by the linear discriminant analysis (LDA) effect size algorithm (LEfSe), detailed in Supplementary Table S4. Taxonomic validation of the representative ASVs was performed with BLASTn analysis. Features possibly associated with disease, given the closest homolog identified, are represented with a dagger (“†”). Kbr Kogia breviceps; Dde, Delphinus delphis; Pph, Phocoena phocoena; Sco, Stenella coeruleoalba.
Figure 2.
Composition of the microbial communities within the oral cavities of K. breviceps. A. Representation of the number of total ASVs in each sampled K. breviceps and the number of ASVs shared between the three sampled specimens, highlighting the ones present in all K. breviceps samples and how they distribute in the other assayed Odontoceti species. B. Relative abundance between the assayed Odontoceti species of the 60 ASVs shared by all K. breviceps specimens. The significance of potential microbial signatures (highlighted with “*”, p-value < 0.05 at the genus and ASV level) was identified by the linear discriminant analysis (LDA) effect size algorithm (LEfSe), detailed in Supplementary Table S4. Taxonomic validation of the representative ASVs was performed with BLASTn analysis. Features possibly associated with disease, given the closest homolog identified, are represented with a dagger (“†”). Kbr Kogia breviceps; Dde, Delphinus delphis; Pph, Phocoena phocoena; Sco, Stenella coeruleoalba.
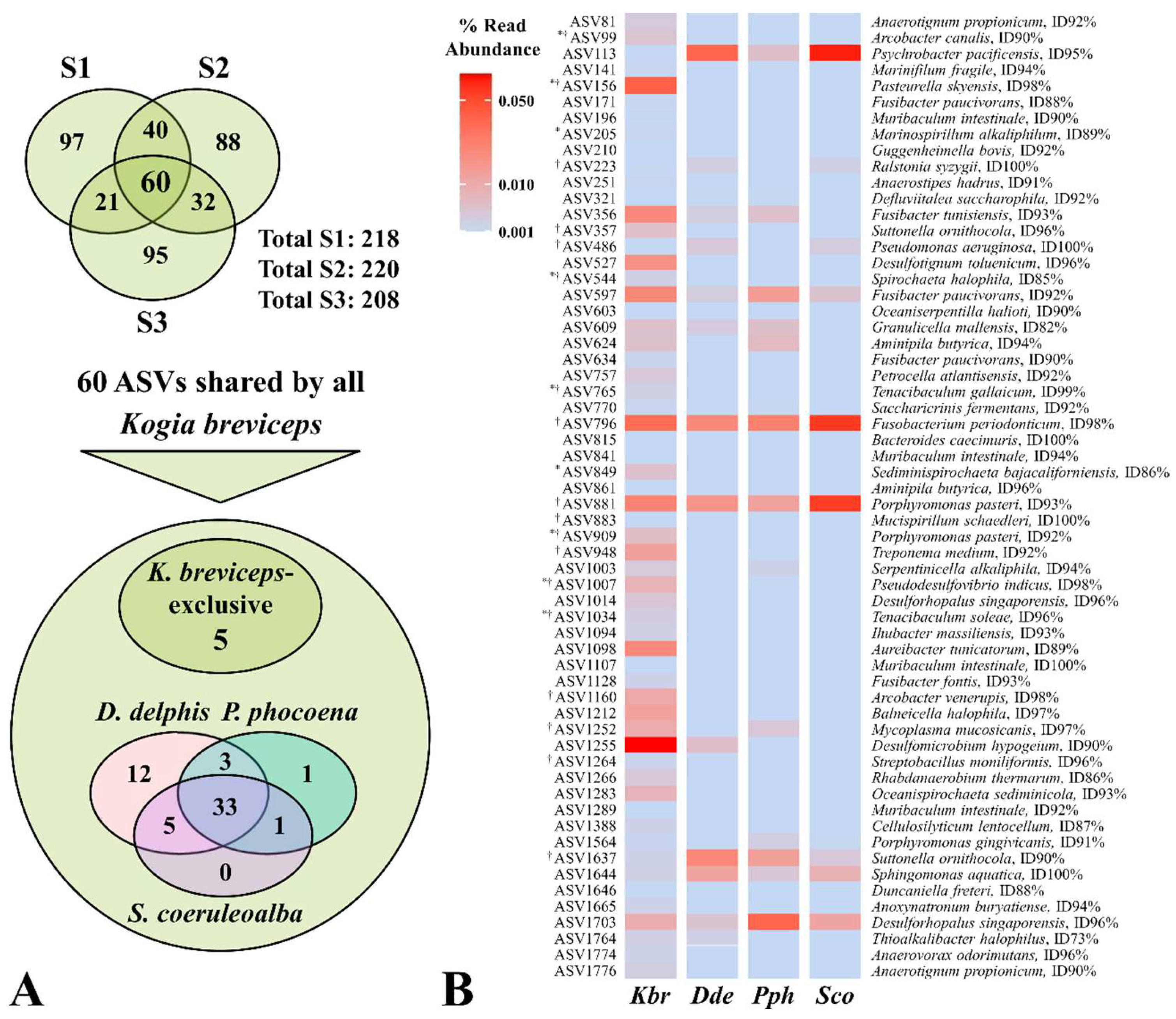

Table 1.
Metadata and number of classified sequences and ASVs estimated in the samples of the oral cavity of the three specimens of Kogia breviceps. The number of unique ASVs per sample, considering all assayed Odontoceti species is also presented.
Table 1.
Metadata and number of classified sequences and ASVs estimated in the samples of the oral cavity of the three specimens of Kogia breviceps. The number of unique ASVs per sample, considering all assayed Odontoceti species is also presented.
| Samples | S1 | S2 | S3 | |
|---|---|---|---|---|
| Metadata | Gender | Male | Male | Male |
| Sexual maturity | Mature | Mature | Immature | |
| Location | Praia de Mira | Praia do Navio | Praia da Rainha | |
| Geographic region | Western Atlantic Iberian coast | Western Atlantic Iberian coast | Western Atlantic Iberian coast | |
| Cause of death | Disease | Disease | Disease | |
| Sequencing metrics | No. of raw reads | 94.650 | 135.918 | 142.087 |
| No. of filtered reads | 67.509 | 117.826 | 98.055 | |
| No. of classified sequences | 39.523 | 64.461 | 52.972 | |
| Total no. of ASVs | 218 | 220 | 208 | |
| No. of unique ASVs | 38 | 24 | 62 | |
| No. of phyla | 12 | 10 | 13 | |
| No. of classes | 21 | 19 | 23 | |
| No. of orders | 34 | 29 | 35 | |
| No. of families | 57 | 55 | 56 | |
| No. of genera | 67 | 51 | 55 |
Table 2.
Best BLASTn hit for the bacterial ASVs contributing to the Kogia breviceps grouping observed in the CCA, according to the host species variable (p-value<0.05), determined by LEfSe and detailed in Supplementary Table S4. Presence (+) or absence (-) in the other three Odontoceti species is represented, to highlight core features in this study. Bold represents the ASVs present in all sampled pigmy sperm whales. An “*“ indicates the potential microbial signatures (p-value < 0.05 also at the genus level) and a “†” highlights features potentially associated with disease, given the determined closest homolog. Dde, Delphinus delphis; Pph, Phocoena phocoena; Sco, Stenella coeruleoalba.
Table 2.
Best BLASTn hit for the bacterial ASVs contributing to the Kogia breviceps grouping observed in the CCA, according to the host species variable (p-value<0.05), determined by LEfSe and detailed in Supplementary Table S4. Presence (+) or absence (-) in the other three Odontoceti species is represented, to highlight core features in this study. Bold represents the ASVs present in all sampled pigmy sperm whales. An “*“ indicates the potential microbial signatures (p-value < 0.05 also at the genus level) and a “†” highlights features potentially associated with disease, given the determined closest homolog. Dde, Delphinus delphis; Pph, Phocoena phocoena; Sco, Stenella coeruleoalba.
| ASV | Presence in other Odontoceti species | Best BLASTn hit | % Identity |
||
|---|---|---|---|---|---|
| Dde | Pph | Sco | |||
| ASVs contributing solely to K. breviceps cluster | |||||
| ASV32 | - | - | - | Balneicella halophila | 98 |
| ASV77 | - | - | - | Thioalkalibacter halophilus | 74 |
| ASV156 *† | - | - | - | Pasteurella skyensis | 98 |
| ASV170 | - | - | - | Balneicella halophila | 96 |
| ASV189 | - | - | - | Marinospirillum alkaliphilum | 88 |
| ASV251 | - | - | - | Anaerostipes hadrus | 91 |
| ASV383 † | - | - | - | Actinobacillus delphinicola | 98 |
| ASV519 *† | - | - | - | Arcobacter marinus | 92 |
| ASV545 † | - | - | - | Suttonella indologenes | 91 |
| ASV570 | - | - | - | Defluviitalea saccharophila | 93 |
| ASV658 | - | - | - | Altericista lacusladogae | 76 |
| ASV725 *† | - | - | - | Pasteurella skyensis | 98 |
| ASV753 † | - | - | - | Arcobacter aquimarinus | 89 |
| ASV777 † | - | - | - | Streptobacillus moniliformis | 96 |
| ASV817 | - | - | - | Crocinitomix algicola | 88 |
| ASV888 * | - | - | - | Moraxella lincolnii | 96 |
| ASV915 | - | - | - | Anaerostipes hadrus | 87 |
| ASV1034 *† | - | - | - | Tenacibaculum soleae | 96 |
| ASV1036 | - | - | - | Anoxynatronum buryatiense | 94 |
| ASV1084 | - | - | - | Fructobacillus pseudoficulneus | 75 |
| ASV1094 | - | - | - | Ihubacter massiliensis | 93 |
| ASV1104 *† | - | - | - | Pasteurella skyensis | 99 |
| ASV1148 | - | - | - | Labilibacter aurantiacus | 91 |
| ASV1155 *† | - | - | - | Pasteurella skyensis | 99 |
| ASV1497 | - | - | - | Deinococcus petrolearius | 74 |
| ASV1603 | - | - | - | Lutibacter oceani | 97 |
| ASV1647 | - | - | - | Nodosilinea alaskaensis | 78 |
| ASV1665 | - | - | - | Anoxynatronum buryatiense | 94 |
| ASVs mainly contributing to K. breviceps cluster (not exclusively) | |||||
| ASV81 | + | - | + | Anaerotignum propionicum | 92 |
| ASV99 *† | + | - | + | Arcobacter canalis | 90 |
| ASV111 † | + | - | - | Sneathia sanguinegens | 98 |
| ASV141 | + | - | - | Marinifilum fragile | 94 |
| ASV148 * | - | - | + | Oceanispirochaeta sediminicola | 92 |
| ASV171 | + | - | - | Fusibacter paucivorans | 88 |
| ASV205 * | + | - | - | Marinospirillum alkaliphilum | 89 |
| ASV218 | + | - | + | Ruminiclostridium cellobioparum | 86 |
| ASV301 | + | - | - | Spongiimonas flava | 86 |
| ASV356 | + | + | + | Fusibacter tunisiensis | 93 |
| ASV357 † | + | - | - | Suttonella ornithocola | 96 |
| ASV402 | + | - | - | Faecalicatena contorta | 94 |
| ASV426 | + | - | - | Salinivirga cyanobacteriivorans | 92 |
| ASV458 † | + | - | - | Sneathia sanguinegens | 95 |
| ASV492 | + | - | - | Polymorphobacter multimanifer | 82 |
| ASV527 | + | - | + | Desulfotignum toluenicum | 96 |
| ASV544 *† | + | - | - | Spirochaeta halophila | 85 |
| ASV603 | + | - | - | Oceaniserpentilla haliotis | 90 |
| ASV635 *† | + | - | - | Helicobacter bilis | 90 |
| ASV676 | + | - | - | Acholeplasma modicum | 90 |
| ASV754 | + | + | - | Fusibacter paucivorans | 91 |
| ASV757 | + | - | - | Petrocella atlantisensis | 92 |
| ASV765 *† | - | + | - | Tenacibaculum gallaicum | 99 |
| ASV778 | + | - | - | Hydrogenophaga soli | 98 |
| ASV839 | + | - | - | Methylophaga thalassica | 92 |
| ASV849 * | + | + | + | Sediminispirochaeta bajacaliforniensis | 86 |
| ASV861 | + | + | + | Aminipila butyrica | 96 |
| ASV885 | + | - | + | Treponema zuelzerae | 90 |
| ASV898 | - | + | - | Ignavibacterium album | 88 |
| ASV909 *† | + | + | + | Porphyromonas pasteri | 92 |
| ASV948 † | + | + | + | Treponema medium | 92 |
| ASV1007 *† | + | + | + | Pseudodesulfovibrio indicus | 98 |
| ASV1098 | + | - | - | Aureibacter tunicatorum | 89 |
| ASV1121 * | + | - | - | Oceanispirochaeta sediminicola | 94 |
| ASV1128 | + | - | - | Fusibacter fontis | 93 |
| ASV1160 † | + | + | + | Arcobacter venerupis | 98 |
| ASV1252 † | + | + | - | Mycoplasma mucosicanis | 97 |
| ASV1255 | + | + | + | Desulfomicrobium hypogeium | 90 |
| ASV1264 † | + | - | - | Streptobacillus moniliformis | 96 |
| ASV1266 | + | + | - | Rhabdanaerobium thermarum | 86 |
| ASV1283 | + | - | + | Oceanispirochaeta sediminicola | 93 |
| ASV1298 † | + | - | + | Comamonas kerstersii | 99 |
| ASV1342 | + | - | - | Ruminiclostridium cellobioparum | 90 |
| ASV1388 | + | - | + | Cellulosilyticum lentocellum | 87 |
| ASV1404 † | + | - | - | Fusobacterium russii | 98 |
| ASV1410 † | + | - | - | Mycoplasma opalescens | 98 |
| ASV1486 | + | - | - | Labilibacter sediminis | 88 |
| ASV1519 | + | - | - | Paenalcaligenes hermetiae | 97 |
| ASV1535 | - | + | - | Edaphobacter acidisoli | 75 |
| ASV1546 | + | - | - | Defluviitalea raffinosedens | 92 |
| ASV1624 *† | + | + | - | Pasteurella skyensis | 96 |
| ASV1641 | + | - | + | Acholeplasma morum | 88 |
| ASV1698 | + | - | - | Balneicella halophila | 99 |
| ASV1701 | + | - | - | Lachnotalea glycerini | 90 |
| ASV1774 | + | - | - | Anaerovorax odorimutans | 96 |
| ASV1776 | + | - | - | Anaerotignum propionicum | 90 |
| ASV1777 | + | - | - | Crocinitomix algicola | 88 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



