Submitted:
10 October 2024
Posted:
10 October 2024
You are already at the latest version
Abstract
Nanotechnology is being used in agriculture to improve plant nutrition while maintaining soil texture and safeguarding it against microbial illnesses. Examples of such applications include the utilization of nano-fertilizers, nano-pesticides, and nano-herbicides. Nanotechnology plays a crucial role in maintaining soil health, hence promoting the overall well-being of plants. Nanoparticles have been shown to enhance agricultural productivity and output, mitigate the opposing effects of chemical runoff and nutrient loss. Several factors, including concentrations, physiochemical properties, and plant species, influence the impact of nanoparticles on plants. Several nanoparticles have been shown to impact plant physiology, leading to enhanced biomass output and germination rate. Nanoparticles have the ability to alter molecular pathways in plants via their impact on gene expression. The presence of quantum dots, carbon nanotubes, ZnO, Ag, Fe2O3, Se, Au, TiO2, Al2O3, SiO2, and non-metal oxides of nanoparticles is of significant importance in the promotion of plant development and growth. Extensive research has been conducted on the role of NPs in the reduction of growth, inhibition of chlorophyll, and enhancement of photosynthetic efficiency. The main purpose of this study was to offer a comprehensive overview of studies that have examined the impacts, translocation, and interactions of nanoparticles with plants.
Keywords:
Nanotechnology
; microbial illness
; pathway
; productivity
; translocation
; interactions
1. Introduction
In 2050, the world’s population will reach 9.8 billion, which is a one third increase over the current population, by a report of Food and Agriculture Organization (FAO) (van Dijk et al. 2021). To fulfil the nutritional requirements of this population, agricultural output must be increased and food security must be enhanced. Pollution, among other environmental factors, and biotic and abiotic stressors experienced by certain soils are preventing agricultural output from keeping pace with population growth (Godoy et al. 2021). There is a lot of pressure to find new ways to make food production better, safer, more efficient, and less harmful to the environment because to the enormous demand for high-quality agricultural goods (van Dijk et al. 2021).
Nanotechnology, which has found several uses in areas including materials science, medicine, physics, and chemistry, has also been effectively used in farming in the last few decades (Omanović-Mikličanin and Maksimović 2016). The utilization of nanoparticles in various applications has hastened the transformation of traditional farming and food production due to the fast expansion of nanotechnology (Figure 1) (Chaud et al. 2021). These particles can serve as nanosensors, antimicrobial, nanofungicides, nanoherbicides, and nanofertilizing agents (Fatima et al. 2021), and as stimulants for plant growth, enhancing resistance to adverse conditions like abiotic stress due to contamination of heavy metal in the soil (Rodríguez-Seijo et al. 2022). To keep agricultural systems viable in the long run, it is important to minimize the negative impacts that might come from employing nanoparticles (Ali et al. 2021).
This manuscript specifically examines the use of inorganic nanoparticles in crop germination, with a focus on both the potential negative and positive effects (Ashraf et al. 2021). Understanding the process of germination is crucial for the survival and conservation of plant species. It helps in improving growth and yield of crops, which is essential for modern agriculture. Therefore, any factor that impacts germination will inevitably impact the subsequent plant development (Khan et al. 2022).
Understanding germination is crucial for restoring degraded soils and promoting vegetation growth. It also plays a dynamic role in food production, as it directly impacts the productivity of crops (Arnott et al. 2021). Germination is typically assessed using two parameters. The first is the germination percentage (GP), which calculates the proportion of seeds that have successfully germinated out of the total number of seeds. The second parameter is the seedling vigour index (SVI), which considers factors such as root and leaf weight or length. Various methods can be used to calculate the SVI (González-Feijoo et al. 2023).
Every year, a multitude of new NPs are introduced, each with enhanced capabilities and a wide range of applications. Biological responses to NPs are influenced by their physicochemical characteristics, like size, zeta potential concentration (Acharya et al. 2019). Biologists have discovered that NPs have an extensive range of potential that can greatly enhance plant productivity. These include acting as germination enhancers, creating nanofertilizers, delivering herbicides, detecting pests through nanosensors, and using nanoporous zeolites for controlled water and fertilizer release. Nevertheless, certain NPs can have negative effects on plants, inhibiting seed germination or being toxic to young seedlings (Hayes et al. 2020).
Various processes such as leaching, hydrolysis, photolysis, and decomposition can render certain fertilizers unavailable to plants, even though they are crucial for plant development and growth (Kah et al. 2019). There have been numerous innovative solutions developed in recent years to address the issue of food waste and improve crop yields. Nanopesticides and nanofertilizers are just a couple of examples of these novel solutions (Omara et al. 2019).
Biologists have discovered that nanofertilizers and nano encapsulated nutrients have the ability to regulate the release of chemical fertilizers, which in turn boost the activity of the desired plants (Iqbal et al. 2019). Various NPs are currently under investigation to determine their effectiveness in shielding plants from various environmental pressures and promoting plant growth (Rajput et al. 2021). Plant biotechnology offers exciting opportunities to manipulate gene expression and cellular properties, expanding our understanding of plant biology. Aside from their significant contributions to agriculture and environmental remediation, NPs also have a diverse array of applications in biosensors (Khan et al. 2019).
Plant uptake and translocation are influenced by various factors, such as the size, types, concentration, reactivity, toxicity, pore sizes, surface charge, and other properties of nanoparticles (Hu et al. 2020). When nanoparticles enter treated surfaces, they have the ability to modify their characteristics, reactivity, and bioavailability to living organisms (Singh et al. 2022). As a biologist, our aim is to present a comprehensive analysis of the advantages and disadvantages associated with the utilization of nanoscale materials in agriculture (Figure 2).
Micronutrients such as Zn, Mn, Mo, Fe, Cu, B, etc. are widely recognized as crucial for the process of growth and development. The implementation of the green revolution and innovative agricultural methods has led to a substantial rise in crop productivity, but at the expense of soil micronutrients such as Zn, Mo, and Fe (Dhaliwal et al. 2021). The use of nanotechnology (NT) has the potential to enhance the accessibility of micronutrients for plants. The enhancement of soil health and vitality may be achieved by the use of nano-formulations of micronutrients, which can be administered to plants through spraying or incorporated into the soil for root absorption (Peteu et al. 2010).
Zinc is crucial for plant metabolism when present at levels below the threshold, but it becomes harmful when present in quantities that exceed the optimal range (Natasha et al. 2022). Zn deficiency is a disorder that has been extensively studied and documented. Zn insufficiency is often seen in semi-arid/arid regions due to reduced solubility and heightened fixation (Younas et al. 2023). Hence, the amount of Zn available in dry soils varies from 0.1 to 2.0 mg/kg, which is inadequate to satisfy the plant’s minimum micronutrient requirement of 60 mg/kg. Therefore, additional Zn treatments are necessary to optimize plant productivity. As to the World Health Organization (WHO), around 50% of the global population is affected by this deficiency, which makes Zn one of the micronutrients with the lowest recommended consumption (Beal et al. 2017).
2. Plants Growth
Physiological reactions are the first notable modifications that transpire upon plant exposure to diverse environmental factors. The physiological reactions of plants to nanoparticles may exhibit both positive and negative effects, which are contingent upon the quantities of NPs and the specific plant species (Gupta et al. 2018). Research has consistently shown that the use of nanoparticles at lower concentrations promotes plant growth and facilitates the production of secondary metabolites (Karimi et al. 2018).
Zn (Zn) plays a vital role as a microelement in several biochemical processes inside plants, such as development and reproduction (Veena & Puthur, 2022). Multiple studies (Choukri et al. 2022) have provided evidence of the advantageous impacts of ZnO NPs on plant development.
To mitigate the ingress of extraneous substances, such as nanoparticles, the cellular walls of plants include a diverse array of functional groups, such as carboxylate, hydroxyl, phosphate, and many others. Biomolecules, such as proteins, polysaccharides, and cellulose, are formed by the combination of these groups (Vinopal et al. 2007). The key factor influencing absorption and transport of nanoparticles is the plant species. Therefore, a comprehensive mechanism that includes the entire system—roots, stems, and leaves—introduces nanoparticles into plants through interactions with soil, water, and several other environmental factors. Furthermore, the presence of nanoparticles in soil might lead to interactions with the root system, ultimately resulting in cellular absorption (Tripathi et al. 2017). Nanoparticles that possess a diameter comparable to that of the cell wall are capable of traversing its sieving mechanisms and ultimately reaching the plasma membrane. The diameter of the cell wall varies between 5 and 20 nm. Refer to Table 1.
Studies examined the physiological alterations that led to the growth-promoting effects of ZnO nanoparticles. These changes included the modulation of plant pigments associated with biomass accumulation, such as chlorophyll and carotenoids (Faizan et al. 2018), an enhancement in nutrient uptake efficiency (Chanu and Upadhyaya 2019), and an elevation in antioxidant metabolism (Venkatachalam et al. 2017).
3. Nanoparticle Absorption by Plants
NPs breach the root cell membrane, enter the plant’s vascular system, and reach the leaves via a complex chain reaction (Tripathi et al. 2017). Certain nanoparticles may diffuse across lipid bilayers and enter cells via endocytosis by creating pores, binding to ion channels and aquaporins, etc. (Schmidt, 2015).
NPs may enter plant cells via the apoplastic and symplastic transport mechanisms. Smaller NPs penetrate the cell wall more readily, while larger ones pass via stomata, hydathodes, and the flower stigma (Hossain et al. 2016). Stomata can open and shut, despite their abundance. Nanoparticles bigger than 40 nm may pass the plant’s stomata and hydathodes to reach the leaf’s spongy and palisade parenchyma (Tripathi et al. 2017). NPs may enter the seed coat via parenchymatous intercellular spaces (Banerjee et al. 2019). However, aquaporins reassemble the AQP-1 and Galphai-3 regulatory complex to limit seed coat NP entry (Abu-Hamdah et al. 2004).
3.1. Foliar Uptake and Translocation of Nanoparticles
Foliar NPs penetrate plants via leaf epidermis or stomata. After absorption, they enter vascular tissue via apoplastic or symplastic pahways. NPs are largely deposited in vacuoles and cell walls, although vascular tissue (xylem and phloem) transports them upward and downward (Su et al., 2019). Plant physiological traits, atmospheric abiotic variables (humidity, light, and temperature), and NP attributes (size, shape, and charge) all influence NP penetration, transfer, and accumulation. Cucumis sativus exhibits foliar uptake of 8 ± 1 nm CeO2 NPs (Hong et al. 2014) and Lactuca sativa uptake of 50-100 nm Cu(OH)2 NPs (Zhao et al. 2016). Figure 3 shows that foliar spraying NPs is more effective and environmentally favourable than root or soil exposure (Fan et al. 2020).
3.2. Transformation and Uptake of Nanoparticles by Root
Roots have the ability to directly absorb nanoparticles from the soil and then transfer them to plant parts. For example, the roots of Arabidopsis thaliana can absorb Au NPs ranging in size from 7 to 108 nm (Taylor et al. 2014). Similarly, the roots of Arabidopsis thaliana can take up SiO2 NPs with sizes up to 200 nm (Slomberg and Schoenfisch, 2012). Wheat root has the ability to absorb TiO2 nanoparticles ranging in size from 36 to 140 nm (Larue et al. 2012). According to Larue et al. (2012), nanoparticles with a size smaller than 36 nm were able to enter the roots of plants and subsequently spread to other parts of the plant without undergoing any transformation. However, NPs ranging in size from 36 to 140 nm did not migrate to the shoots and instead accumulated in the root parenchyma. Furthermore, NPs larger than 140 nm did not accumulate in the root. Figure 4 illustrates the transportation of nanoparticles in plants via the apoplastic and symplastic pathways (Avellan et al. 2021).
The apoplastic route involves NPs passing via epidermis and cortex to endodermis, where casparian strip is formed in radial and transverse walls to inhibit NP and macromolecule access into the vascular system. The casparian strip is detached at the lateral root junction or underdeveloped at the root tip, so NPs may bypass it and enter the vascular system (Schymura et al. 2017). In the symplastic route, NPs reach the cytoplasm or plasmodesmata of neighboring cells via plasma membrane (PM). Transmission Electron Microscope (TEM) showed that Arabidopsis thaliana root tips subjected to 20, 40, and 80 nm Ag NPs accumulated Ag NPs in root cell plasmodesmata and middle lamella. Au NPs were found in poplar root cell cytoplasm, plasmodesmata, mitochondria, plastids, and cell walls using TEM subjected to 15, 25, and 50 nm Au NPs (Zhai et al. 2014).
4. Transformation of Nanoparticles
The conversion of nanoparticles takes place at several plant sites, including the root, leaf surface, and internal tissues (Mia et al. 2021). This study provides a concise overview of the sites where nanoparticles undergo transformation, the variables that influence NPs transformation, and the processes of absorption and translocation of changed NPs in plants.
4.1. Transformation in Soil
The presence of inorganic and organic components, as well as soil texture, can have an impact on the transformation of NPs in soil. Clay soil, with its limited pore space for air, creates a reducing environment that can result in a decrease in NPs. This decrease can occur through various mechanisms, including:
- Leaching: NPs being washed away by water, potentially entering groundwater or nearby water sources.
- Dissolution: NPs breaking down into smaller species or ions, losing their nanoparticle properties.
- Adsorption: NPs adhering to surfaces, such as soil particles or plant cell walls, reducing their availability.
- Degradation: NPs being broken down by chemical or biological processes, such as oxidation or enzymatic activity.
- Uptake and internalization: NPs being taken up by plants or microorganisms, reducing their external concentration.
-
Sedimentation: NPs settling out of solution due to gravity, reducing their concentration in the surrounding medium.
- ➢
- When dealing with sandy soil, it’s worth noting that NPs tend to undergo oxidation due to the higher oxygen content. For instance, the presence of an Ag2O layer around Ag NPs can lead to the dissolution and subsequent release of Ag+ (Li et al. 2017).
- ➢
- The weathering process can have a significant impact on the availability of copper nanoparticles in soil, as well as its uptake and movement within lettuce plants (Servin et al. 2017).
4.2. Plant-Mediated Transformation of NPs
- ➢
- A complicated process produces differences in organic matter, mineral components, soil pH, and microbial community that alter the transformations of NPs during their dissolution and transformation in the rhizosphere. Many chemical reactions, including dissolution, accompany the change of metal-based NPs, which may also include sulfidation, phosphorylation, chelation, or reduction (Zhang et al. 2020).
- ➢
- The chemical modification of NPs at the Phyllosphere via interactions with the epiphytes (bacteria, fungus, and yeast) on the surface of the leaf might alter the aggregation state (Zhang et al. 2020).
- ➢
- Depending on the species of plant, the kind and degree of NP transformation may differ. As an example, the root exudates of cucumbers (Cucumis sativus) are firmly bound to by CuO NPs (∼40 nm), which causes the transformation of CuO NPs to Cu(I) and Cu (II) and decreases the absorption and buildup of Cu (Huang et al. 2017).
5. Physiological Effects of Nanoparticles in Plants
Figure 5 shows the positive and negative impacts of NPs on morphophysiological traits, plant growth, and agricultural crop output. Factors influencing NPs’ effects include plant species, developmental stage, application technique, dosage, and supplementing level. In order to increase crop yields and plant development, many agricultural products nowadays are generated from nanotechnology (Awasthi et al. 2020). These goods are superior to traditional farming methods and pesticides because to their many desirable qualities, including their compact size, low toxicity, ease of handling, extended storage life, and high efficacy (Hong et al. 2021).
6. Impression of Different NPs on the Physiological Processes for Plant Development, Growth, and Maturation
6.1. Copper Nanoparticles
CuO NP exposure had a substantial influence on the germination, biomass, shoot development, and other processes of many plant species (Rajput et al. 2018). The toxicity of CuO nanoparticles did not have any impact on seed germination in maize plants. The nanoparticles were transported to the shoots via the xylem and then transported back to the roots via the phloem. Brassica napus seedlings were cultivated in MS medium supplemented with CuO NPs (0, 10, 100, and 1000 mg L-1) for a duration of 10 days (Wang et al. 2012). The administration of a maximum dose of 10 mg L-1 led to the initiation of growth, whereas higher concentrations (100 and 1000 mg L-1) caused a decrease in root dry weight and an increase in shoot elongation (Rahmani et al. 2016).
The growth of Lemna minor was impeded by the presence of Cu2+ in the culture medium at lower concentrations, in contrast to greater concentrations of CuO (Song et al. 2016). Applying a colloidal solution of CuNP (0.5 mg L-1) and CoNP (0.8 mg L-1) along with MS media to the Mentha longifoila plant resulted in a significant increase in height and growth, as well as a 29.4-33.9 % increase in internodes, a 55.6-26.2 % increase in shoots, and a 30-40 % increase in reproduction coefficient (TalankovaSereda et al. 2016). Copper nanoparticles, produced from tea extract, had beneficial effects on the growth of seedlings and the signalling of nitric oxide when exposed to Lactuca sativa at a concentration of 20 μg mL-1 or lower (Pelegrino et al., 2020).
6.2. Iron Oxide Nanoparticles
Applying iron oxide (Fe2O3) nanoparticles to plants greatly enhances their growth, ability to withstand stress, and nutritional condition. According to Yasmeen et al. (2015), the germination percentage of wheat (Triticum aestivum) was enhanced by immersing it in distilled water and thereafter subjecting it to incubation in a solution containing iron nanoparticles. Nevertheless, the absence of NPs in the distilled water resulted in a decrease in root development, while the presence of NPs in the distilled water led to an increase in root growth. Both the positively and negatively charged ions of iron oxide at concentrations of 3 and 25 mg L-1 influenced the physiology of A. thaliana. According to Bombin et al. (2015), the seedling and root length remained unchanged when exposed to a dosage of 3 mg L-1. However, these lengths significantly reduced when exposed to a dose of 25 mg L-1.
Researchers have shown that applying iron oxide and chelated iron EDTA to Acinetobacter hypogaea enhances the biomass, germination, and development of peanut plants by increasing enzyme antioxidant activities and phytohormone levels. According to Rui et al. (2016), the application of Fe2O3 to plants resulted in enhanced iron availability, making it a viable option for fertilization purposes. Liu et al. (2016) revealed that the exposure of L. sativa seedlings to Fe2O3 NP (5-20 ppm) resulted in a 12-26 % improvement in root elongation. According to research done by Tombuloglu et al. (2019), the accumulation of Fe3O4 in Hordeum vulgare resulted in enhanced plant growth and higher photosynthetic efficiency.
6.3. Silver Nanoparticles
Because of its antimicrobial characteristics, silver (Ag) has found extensive use in industry and medicine, exposing it to more plants and humans than ever before. The usage of Ag NPs in agriculture has shown promising results due to their many favorable impacts on plant growth and development (Almutairi 2016). When silver nanoparticles were used excessively, they increased the synthesis and activity of antioxidants such carotenoids and proline, as well as peroxidases and catalases. Additionally, at higher dosages, it enhanced seed germination and development in Lolium multiflorum and Eruca sativa, while it reduced root length in V. radiata and Sorghum bicolor (Aqeel et al. 2022). The effects of heat stress in T. aestivum were mitigated by Ag NPs (Iqbal et al. 2019). Numerous biochemical indicators, such as leaf area, root and shoot length, carbohydrate and protein contents, and antioxidant enzyme activity, were enhanced in plants that were exposed to Ag NPs. These plants included B. juncea, common bean, and maize (Salama 2012).
6.4. Carbon Nanotubes
Carbon nanotubes (CNTs) are gaining attention in the diagnostic, biomedical, and agricultural fields because of their varied physiochemical properties (Patel et al. 2020). Thanks to their one-of-a-kind physicochemical properties, CNTs are great for regulating plant development, absorbing water, and supplementing nutrients (Achari et al. 2020). Nanoparticles containing carbon, such as C60, SWCNTs, and MWCNTs, have many uses in the scientific community. By influencing gene expression, SWCNTs improve rice seedlings’ water intake and speed up the germination process (Zhang et al. 2017).
Tomato seedlings (Solanum lycopersicum) were shown to germinate and grow more rapidly when exposed to CNTs as compared to a control group. Additionally, the CNTs facilitated water absorption by penetrating the plants’ outer layer (epidermis) (Fincheira et al. 2020). According to Rahmani et al. (2020), a lower concentration of oxidized multi-walled carbon nanotubes (∼30 nm) might be an effective treatment for B. juncea effect are:
- Enhanced growth and biomass production (e.g., TiO2 NPs)
- Improved photosynthesis and light absorption (e.g., ZnO NPs)
- Increased water uses efficiency and drought tolerance (e.g., SiO2 NPs)
- Enhanced nutrient uptake and transport (e.g., Fe3O4 NPs)
- Altered hormone regulation and signaling (e.g., Au NPs)
- Increased stress tolerance and antioxidant activity (e.g., CeO2 NPs)
- Modified cell wall composition and structure (e.g., Ag NPs)
- Changed gene expression and regulation (e.g., CuO NPs)
The use of MWCNTs has been shown to hasten seed germination in G. max and H. vulgare plants, with no detrimental effects on the plants’ future growth (Samadi et al. 2020). Water channel protein-encoding genes were more abundant in treated seeds compared to control seeds. Using a lower dose of MWCNTs improved water delivery in Z. mays plants (Tiwari et al. 2014).
The same results have been shown for B. napus and C. arietinum plants. Also, via influencing changes in root plasma membrane lipid content, stiffness, and permeability, MWCNTs enhance aquaporin transduction in NaCl-challenged conditions (Martinez-Ballesta et al. 2020).
7. Nanoparticles Influence the Structure and Function of Plants’ Photosynthetic Systems
Plants and algae both use photosynthesis to transform the energy from the sun into chemical energy. The amount of energy that plants are able to shift over their life cycle is a meagre 2-4% (Kirschbaum 2011). Photosynthesis, mineral and nutrient translocation, and the oxygen cycle are all processes in which plants play an essential role. The nutrients that plants take in may be either essential or non-essential; nonetheless, there is a concentration at which they start to become toxic (Mitra 2017). By using gene editing and nanotechnology, scientists can improve the photosynthetic system and efficiency of plants. The interaction between plants and NPs will inevitably have an effect on translocation as well as the acceleration of plant biotechnology.
Toxic NPs like CuO and Ag disrupt the structure and function of the photosynthetic apparatus.
Inhibitory Effects:
- Ag NPs (silver nanoparticles)
- CuO NPs (copper oxide nanoparticles)
- ZnO NPs (zinc oxide nanoparticles)
can inhibit photosynthesis by:
- Damaging chloroplasts and disrupting electron transport chains
- Reducing light absorption and pigment content
- Altering stomatal aperture and gas exchange
- Inducing oxidative stress and antioxidant defenses
On the other hand, NPs like:
- TiO2 NPs (titanium dioxide nanoparticles)
- SiO2 NPs (silicon dioxide nanoparticles)
- Fe3O4 NPs (iron oxide nanoparticles)
can enhance photosynthesis by:
- Increasing light absorption and scattering
- Improving electron transport and ATP production
- Enhancing stomatal conductance and CO2 uptake
- Reducing oxidative stress and promoting antioxidant activity
NPs cause chloroplast anomalies such as reduced photosynthetic pigment concentration (particularly chlorophyll) and grana disruption. Nones are inefficient in photosynthesis and photosystem II. A number of undesirable effects persisted even after the introduction of CeO2 and TiO2 NPs improved electron transport between PS II and I and Rubisco activity (Tighe-Neira et al. 2018). The use of SWCNTs resulted in a tripling of chloroplast electron transport rate and photosynthetic activity. Nano TiO2-induced carboxylation by Rubisco activation enhances photosynthetic carbon absorption (Ali et al. 2021). SiO2 NPs enhanced photosynthesis by influencing carbonic anhydrase and photosynthetic pigment activity (Poddar 2020).
8. Nanoparticles and Antioxidant Capacity
Exposure to NPs has the potential to activate antioxidant defence mechanisms, cause oxidative damage, and generate reactive oxygen species (ROS) (Rico et al. 2015). An integral aspect of the antioxidant defence system are enzymatic antioxidants such as glutathione reductase (GR), glutathione, as well as chloramphenicol acetyltransferase (CAT), superoxide dismutase (SOD), glutathione peroxidase (GPOX), and glutathione reductase (GR) (Kaur et al. 2019). As per Rico et al. (2015), SOD is responsible for converting superoxide ions into hydrogen peroxide, whereas CAT and GPOX are responsible for stifling peroxy radicals and reactive oxygen species, respectively. The generation of reactive oxygen species (ROS) by NPs via APOX involves the direct reduction of hydrogen peroxide into water (Mittler 2016). The anti-oxidant enzyme activities of several NPs. A few examples are the following: nFe2O4, nCeO2, and nCo3O4 improve catalase activity; nFe3O4, nCeO2, nMnO2, nCuO, and nAu enhance GPOX activity; and nCeO2 and fullerene generate superoxide dismutase (Tripathi et al. 2017).
There is a lack of evidence connecting the enzyme activity disruptions seen in plants exposed to NPs to the chemical features of NPs or to the enzyme interactions with these NPs as the origin of these changes, despite the fact that several nanophytotoxicity studies have proven this. It turned out that NPs affected enzyme activity in different ways, for sure. While nTiO2 enhanced the GPOX, SOD, and CAT activities in Lemna minor and spinach, respectively, it decreased the GR and APOX activities in Vicia faba (Lei et al. 2008). This makes it difficult to isolate the NPs that affect certain enzymes.
9. Conclusion
Nanotechnology, a novel approach, has several scientific uses. Nanoparticles may lead to food security and innovative farming. NPs in farming improve the global economy in various ways. Due to a paucity of data, NPs’ toxic effects are unknown, however NP-plant interaction is sensitive to NP size and may be useful or harmful. They affect plants differently according on their growth stage, exposure duration, the absorption rate, and physiochemical traits. NPs outperform traditional resources in effectiveness and agronomy. Interactions between plants and NPs may identify illnesses on-site, promising sustainable agriculture. Nano-based herbicides, insecticides, fertilizers, fungicides, and sensors have been studied for plant management and controlled release to protect the environment. However, the expanding use of NPs in agriculture and related sectors raises concerns about environmental pollution, thus proactive measures should be taken to avoid their buildup. Due of population growth, agricultural experts worry about food security. Nano-revolution will improve food security, the ecology, and farm sustainability. Molecular science research on plant-NP interactions is critically needed to reduce phytotoxicity and boost agricultural yield for human welfare.
Author Contribution
All the authors contributed in writing review. In addition, all the authors have read and agreed to publish the current version of the manuscript.
Data availability statement
Not applicable
Conflict of interest
We declare that current article is original and has not been submitted for publication, in part or in whole, to any other national or international journal.
References
- Abu-Hamdah R, Cho WJ, Cho SJ, Jeremic A, Kelly M, Ilie, AE, Jena, BP (2004) Regulation of the water channel aquaporin-1: isolation and reconstitution of the regulatory complex. Cell biology international, 28(1), 7-17. [CrossRef]
- Achari GA, Zakane RN, Kowshik, M (2020) Eco-friendly Nanomaterials in Agriculture: Biofortification, Plant Growth Promotion, and Phytopathogen Control. Handbook of Nanomaterials and Nanocomposites for Energy and Environmental Applications, 1-22.
- Acharya P, Jayaprakasha GK, Crosby KM, Jifon JL, Patil BS (2019) Green-synthesized nanoparticles enhanced seedling growth, yield, and quality of onion (Allium cepa L.). ACS Sustainable Chemistry & Engineering, 7(17), 14580-14590. [CrossRef]
- Ali SS, Al-Tohamy R, Koutra E, Moawad MS, Kornaros M, Mustafa AM, Sun, J (2021) Nanobiotechnological advancements in agriculture and food industry: Applications, nanotoxicity, and future perspectives. Science of the Total Environment, 792, 148359. [CrossRef]
- Ali S, Mehmood, A, Khan N (2021) Uptake, translocation, and consequences of nanomaterials on plant growth and stress adaptation. Journal of Nanomaterials, 2021, 1-17. [CrossRef]
- Almutairi ZM (2016) Effect of nano-silicon application on the expression of salt tolerance genes in germinating tomato (‘Solanum lycopersicum’L.) seedlings under salt stress. Plant Omics, 9(1), 106-114.
- Aqeel U, Aftab T, Khan MMA, Naeem M, Khan MN (2022) A comprehensive review of impacts of diverse nanoparticles on growth, development and physiological adjustments in plants under changing environment. Chemosphere, 291, 132672. [CrossRef]
- Arnott A, Galagedara L, Thomas R, Cheema M, Sobze JM (2021) The potential of rock dust nanoparticles to improve seed germination and seedling vigor of native species: A review. Science of The Total Environment, 775, 145139. [CrossRef]
- Ashraf SA, Siddiqui AJ, Abd Elmoneim OE, Khan MI, Patel M, Alreshidi M, Adnan M (2021) Innovations in nanoscience for the sustainable development of food and agriculture with implications on health and environment. Science of the Total Environment, 768, 144990. [CrossRef]
- Avellan A, Yun J, Morais BP, Clement ET, Rodrigues SM, Lowry GV (2021) Critical review: Role of inorganic nanoparticle properties on their foliar uptake and in planta translocation. Environmental science & technology, 55(20), 13417-13431. [CrossRef]
- Awasthi G, Singh T, Tiwari Y, Awasthi A, Tripathi RD, Shrivastava S, Awasthi KK (2020) A review on nanotechnological interventions for plant growth and production. Materials Today: Proceedings, 31, 685-693. [CrossRef]
- Banerjee K, Pramanik P, Maity A, Joshi DC, Wani SH, Krishnan P (2019) Methods of using nanomaterials to plant systems and their delivery to plants (mode of entry, uptake, translocation, accumulation, biotransformation and barriers). In Advances in Phyto nanotechnology (pp. 123-152). Academic Press. [CrossRef]
- Beal T, Massiot E, Arsenault JE, Smith MR, Hijmans RJ (2017) Global trends in dietary micronutrient supplies and estimated prevalence of inadequate intakes. PloS one, 12(4), e0175554. [CrossRef]
- Bombin S, LeFebvre M, Sherwood J, Xu Y, Bao Y, Ramonell KM (2015) Developmental and reproductive effects of iron oxide nanoparticles in Arabidopsis thaliana. International journal of molecular sciences, 16(10), 24174-24193. [CrossRef]
- Chanu TT, Upadhyaya H (2019). Zn oxide nanoparticle-induced responses on plants: a physiological perspective. In Nanomaterials in plants, algae and microorganisms (pp. 43-64). Academic Press. [CrossRef]
- Chaud M, Souto EB, Zielinska A, Severino P, Batain F, Oliveira-Junior J, Alves T. (2021) Nanopesticides in agriculture: Benefits and challenge in agricultural productivity, toxicological risks to human health and environment. Toxics, 9(6), 131. [CrossRef]
- Choukri M, Abouabdillah A, Bouabid R, Abd-Elkader OH, Pacioglu O, Boufahja F, Bourioug M (2022) Zn application through seed priming improves productivity and grain nutritional quality of silage corn. Saudi Journal of Biological Sciences, 29(12), 103456. [CrossRef]
- Dhaliwal SS, Sharma V, Mandal A, Naresh RK, Verma G (2021) Improving soil micronutrient availability under organic farming. In Advances in organic farming (pp. 93-114). Woodhead Publishing. [CrossRef]
- Elsherif DE, Abd-ElShafy E, Khalifa AM (2023) Impacts of ZnO as a nanofertilizer on fenugreek: some biochemical parameters and SCoT analysis. Journal of Genetic Engineering and Biotechnology, 21(1), 52. [CrossRef]
- Fan X, Cao X, Zhou H, Hao L, Dong W, He C, Zheng Y (2020) Carbon dioxide fertilization effect on plant growth under soil water stress associates with changes in stomatal traits, leaf photosynthesis, and foliar nitrogen of bell pepper (Capsicum annuum L.). Environmental and Experimental Botany, 179, 104203. [CrossRef]
- Fatima F, Hashim A, Anees S (2021) Efficacy of nanoparticles as nanofertilizer production: a review. Environmental Science and Pollution Research, 28(2), 1292-1303.
- Fincheira P, Tortella G, Duran N, Seabra AB, Rubilar O (2020) Current applications of nanotechnology to develop plant growth inducer agents as an innovation strategy. Critical reviews in biotechnology, 40(1), 15-30. [CrossRef]
- Geisler-Lee J, Wang Q, Yao Y, Zhang W, Geisler M, Li K, Ma X (2013) Phytotoxicity, accumulation and transport of silver nanoparticles by Arabidopsis thaliana. Nanotoxicology, 7(3), 323-337. [CrossRef]
- Godoy F, Olivos-Hernández K, Stange C, Handford M (2021) Abiotic stress in crop species: improving tolerance by applying plant metabolites. Plants, 10(2), 186. [CrossRef]
- González-Feijoo R, Rodríguez-Seijo A, Fernández-Calviño D, Arias-Estévez M, Arenas-Lago D (2023) Use of three different nanoparticles to reduce Cd availability in soils: effects on germination and early growth of Sinapis alba L. Plants, 12(4), 801. [CrossRef]
- Gupta SD, Agarwal A, Pradhan S (2018) Phytostimulatory effect of silver nanoparticles (AgNPs) on rice seedling growth: An insight from antioxidative enzyme activities and gene expression patterns. Ecotoxicology and environmental safety, 161, 624-633. [CrossRef]
- Hong J, Wang C, Wagner DC, Gardea-Torresdey JL, He F, Rico CM (2021) Foliar application of nanoparticles: mechanisms of absorption, transfer, and multiple impacts. Environmental Science: Nano, 8(5), 1196-1210. [CrossRef]
- Hossain Z, Mustafa G, Sakata K, Komatsu S (2016) Insights into the proteomic response of soybean towards Al2O3, ZnO, and Ag nanoparticles stress. Journal of hazardous materials, 304, 291-305. [CrossRef]
- Hu P, An J, Faulkner MM, Wu H, Li Z, Tian X, Giraldo JP (2020) Nanoparticle charge and size control foliar delivery efficiency to plant cells and organelles. ACS nano, 14(7), 7970-7986. [CrossRef]
- Huang Y, Zhao L, Keller AA (2017) Interactions, transformations, and bioavailability of nano-copper exposed to root exudates. Environmental science & technology, 51(17), 9774-9783. [CrossRef]
- Iqbal M, Raja NI, Mashwani ZUR, Hussain M, Ejaz M, Yasmeen F (2019) Effect of silver nanoparticles on growth of wheat under heat stress. Iranian Journal of Science and Technology, Transactions A: Science, 43, 387-395.
- Iqbal M, Umar S, Mahmooduzzafar (2019) Nano-fertilization to enhance nutrient use efficiency and productivity of crop plants. Nanomaterials and plant potential, 473-505.
- Kah M, Tufenkji N, White JC (2019) Nano-enabled strategies to enhance crop nutrition and protection. Nature nanotechnology, 14(6), 532-540.
- Karimi N, Behbahani M, Dini G, Razmjou A (2018) Enhancing the secondary metabolite and anticancer activity of Echinacea purpurea callus extracts by treatment with biosynthesized ZnO nanoparticles. Advances in Natural Sciences: Nanoscience and Nanotechnology, 9(4), 045009. [CrossRef]
- Kaur N, Kaur J, Grewal SK, Singh I (2019) Effect of heat stress on antioxidative defense system and its amelioration by heat acclimation and salicylic acid pre-treatments in three pigeonpea genotypes. Indian Journal of Agricultural Biochemistry, 32(1), 106-110. [CrossRef]
- Khan I, Awan SA, Rizwan M, Hassan ZU, Akram MA, Tariq R, Xie W (2022) Nanoparticle’s uptake and translocation mechanisms in plants via seed priming, foliar treatment, and root exposure: A review. Environmental Science and Pollution Research, 29(60), 89823-89833.
- Khan I, Saeed K, Khan I (2019) Nanoparticles: Properties, applications and toxicities. Arabian journal of chemistry, 12(7), 908-931. [CrossRef]
- Kirschbaum MU (2011) Does enhanced photosynthesis enhance growth? Lessons learned from CO2 enrichment studies. Plant physiology, 155(1), 117-124. [CrossRef]
- Larue C, Veronesi G, Flank AM, Surble S, Herlin-Boime N, Carrière M (2012) Comparative uptake and impact of TiO2 nanoparticles in wheat and rapeseed. Journal of Toxicology and Environmental Health, Part A, 75(13-15), 722-734. [CrossRef]
- Lei Z, Mingyu S, Xiao W, Chao L, Chunxiang Q, Liang C, Fashui H (2008) Antioxidant stress is promoted by nano-anatase in spinach chloroplasts under UV-B radiation. Biological Trace Element Research, 121, 69-79. [CrossRef]
- Li WR, Sun TL, Zhou SL, Ma YK, Shi QS, Xie XB, Huang XM (2017) A comparative analysis of antibacterial activity, dynamics, and effects of silver ions and silver nanoparticles against four bacterial strains. International Biodeterioration & Biodegradation, 123, 304-310. [CrossRef]
- Liu R, Zhang H, Lal R (2016) Effects of stabilized nanoparticles of copper, Zn, manganese, and iron oxides in low concentrations on lettuce (Lactuca sativa) seed germination: nanotoxicants or nanonutrients. Water, Air, & Soil Pollution, 227, 1-14.
- Mazhar Z, Akhtar J, Alhodaib A, Naz T, Zafar MI, Iqbal MM, Naz I (2023) Efficacy of ZnO nanoparticles in Zn fortification and partitioning of wheat and rice grains under salt stress. Scientific reports, 13(1), 2022.
- Mitra G (2017) Essential plant nutrients and recent concepts about their uptake. Essential plant nutrients: Uptake, use efficiency, and management, 3-36.
- Mittler R (2017) ROS is good. Trends in plant science, 22(1), 11-19. [CrossRef]
- Natasha N, Shahid M, Bibi I, Iqbal J, Khalid S, Murtaza B, Arshad M (2022) Zn in soil-plant-human system: A data-analysis review. Science of the Total Environment, 808, 152024. [CrossRef]
- Omanović-Mikličanin M, Maksimović (2016) Nanosensors applications in agriculture and food industry. Glas Hem. Technol. Bosne Herceg., 47, pp. 59-70.
- Omara AED, Elsakhawy T, Alshaal T, El-Ramady H, Kovács Z, Fári M (2019) Nanoparticles: a novel approach for sustainable agro-productivity. Environment, Biodiversity and Soil Security, 3(2019), 29-62. [CrossRef]
- Patel DK, Kim HB, Dutta SD, Ganguly K, Lim KT (2020) Carbon nanotubes-based nanomaterials and their agricultural and biotechnological applications. Materials, 13(7), 1679. [CrossRef]
- Pelegrino MT, Kohatsu MY, Seabra AB, Monteiro LR, Gomes DG, Oliveira HC, Lange CN (2020) Effects of copper oxide nanoparticles on growth of lettuce (Lactuca sativa L.) seedlings and possible implications of nitric oxide in their antioxidative defense. Environmental Monitoring and Assessment, 192, 1-14.
- Peteu SF, Oancea F, Sicuia OA, Constantinescu F, Dinu S (2010) Responsive polymers for crop protection. Polymers, 2(3), 229-251. [CrossRef]
- Plaksenkova I, Kokina I, Petrova A, Jermaļonoka M, Gerbreders V, Krasovska M (2020) The impact of Zn oxide nanoparticles on cytotoxicity, genotoxicity, and miRNA expression in barley (Hordeum vulgare L.) seedlings. The Scientific World Journal, 2020. [CrossRef]
- Poddar K, Sarkar D, Sarkar A (2020) Nanoparticles on photosynthesis of plants: effects and role. Green Nanoparticles: Synthesis and Biomedical Applications, 273-287.
- Rahmani N, Radjabian T, Soltani BM (2020) Impacts of foliar exposure to multi-walled carbon nanotubes on physiological and molecular traits of Salvia verticillata L., as a medicinal plant. Plant physiology and biochemistry, 150, 27-38. [CrossRef]
- Rajput VD, Minkina T, Kumari A, Harish, Singh VK, Verma KK, Keswani C (2021) Coping with the challenges of abiotic stress in plants: New dimensions in the field application of nanoparticles. Plants, 10(6), 1221. [CrossRef]
- Rajput VD, Minkina T, Suskova S, Mandzhieva S, Tsitsuashvili V, Chapligin V, Fedorenko A (2018) Effects of copper nanoparticles (CuO NPs) on crop plants: a mini review. Bio Nanoscience, 8, 36-42.
- Rico CM, Peralta-Videa JR, Gardea-Torresdey JL (2015) Chemistry, biochemistry of nanoparticles, and their role in antioxidant defense system in plants. Nanotechnology and plant sciences: nanoparticles and their impact on plants, 1-17.
- Rodríguez-Seijo A, Soares C, Ribeiro S, Amil BF, Patinha C, Cachada A, Pereira R (2022) Nano-Fe2O3 as a tool to restore plant growth in contaminated soils–Assessment of potentially toxic elements (bio) availability and redox homeostasis in Hordeum vulgare L. Journal of Hazardous Materials, 425, 127999. [CrossRef]
- Rui M, Ma C, Rui Y, Fan X (2016) Iron oxide nanoparticles as a potential iron fertilizer for peanut (Arachis hypogaea). Frontiers in plant science, 7, 195361. [CrossRef]
- Salama HM (2012) Effects of silver nanoparticles in some crop plants, common bean (Phaseolus vulgaris L.) and corn (Zea mays L.). Int Res J Biotechnol, 3(10), 190-197.
- Samadi S, Saharkhiz MJ, Azizi M, Samiei L, Ghorbanpour M (2020) Multi-walled carbon nanotubes stimulate growth, redox reactions and biosynthesis of antioxidant metabolites in Thymus daenensis celak. in vitro. Chemosphere, 249, 126069. [CrossRef]
- Schmidt J (2015) Nanoparticle-induced membrane pore formation studied with lipid bilayer arrays. Biophysical Journal, 108(2), 344a-345a.
- Schymura S, Fricke T, Hildebrand H, Franke K (2017) Elucidating the role of dissolution in CeO2 nanoparticle plant uptake by smart radiolabeling. Angewandte Chemie International Edition, 56(26), 7411-7414. [CrossRef]
- Servin AD, Pagano L, Castillo-Michel H, De la Torre-Roche R, Hawthorne J, Hernandez-Viezcas JA, White JC (2017) Weathering in soil increases nanoparticle CuO bioaccumulation within a terrestrial food chain. Nanotoxicology, 11(1), 98-111. [CrossRef]
- Singh A, Singh S, Prasad SM, Tripathi DK, Singh VP, Ahmad P, Chauhan DK, Prasad SM (2016) Silicon and Nanotechnology Role in Agriculture and Future Perspective in Silicon in Plants in Advances and Future Prospects, CRC Press, p. 392 . [CrossRef]
- Singh Y, Sodhi RS, Singh PP, Kaushal S (2022) Biosynthesis of NiO nanoparticles using Spirogyra sp. cell-free extract and their potential biological applications. Materials Advances, 3(12), 4991-5000. [CrossRef]
- Slomberg DL, Schoenfisch MH (2012) Silica nanoparticle phytotoxicity to Arabidopsis thaliana. Environmental science & technology, 46(18), 10247-10254. [CrossRef]
- Song GuanLing SG, Hou WenHua HW, Gao Yuan GY, Wang Yan WY, Lin Lin LL, Zhang ZhiWei ZZ, Wang HaiXia WH (2016) Effects of CuO nanoparticles on Lemna minor.
- Song U, Kim J (2020) Zn oxide nanoparticles: a potential micronutrient fertilizer for horticultural crops with little toxicity. Horticulture, Environment, and Biotechnology, 61(3), 625-631.
- Su, Y., Ashworth, V., Kim, C., Adeleye, A. S., Rolshausen, P., Roper, C., ... & Jassby, D. (2019). Delivery, uptake, fate, and transport of engineered nanoparticles in plants: a critical review and data analysis. Environmental Science: Nano, 6(8), 2311-2331.
- Talankova-Sereda TE, Liapina KV, Shkopinskij EA, Ustinov AI, Kovalyova AV, Dulnev PG, Kucenko NI (2016) The influence of Cu and Co nanoparticles on growth characteristics and biochemical structure of Mentha longifolia in vitro Nanosci. Nanoeng., 4 (2016), pp. 31-39.
- Taylor AF, Rylott EL, Anderson CW, Bruce NC (2014) Investigating the toxicity, uptake, nanoparticle formation and genetic response of plants to gold. PLOS one, 9(4), e93793. [CrossRef]
- Tighe-Neira R, Carmora E, Recio G, Nunes-Nesi A, Reyes-Diaz M, Alberdi M, Inostroza-Blancheteau C (2018) Metallic nanoparticles influence the structure and function of the photosynthetic apparatus in plants Plant Physiology & Biochemistry, 130, pp. 408-417 . [CrossRef]
- Tiwari DK, Dasgupta-Schubert N, Villaseñor Cendejas LM, Villegas J, Carreto Montoya L (2014) Borjas García SE Interfacing carbon nanotubes (CNT) with plants: enhancement of growth, water and ionic nutrient uptake in maize (Zea mays) and implications for nanoagriculture. Appl. Nanosci, 4(5), 577-591.
- Tombuloglu H, Slimani Y, Tombuloglu G, Almessiere M, Baykal A (2019) Uptake and translocation of magnetite (Fe3O4) nanoparticles and its impact on photosynthetic genes in barley (Hordeum vulgare L.). Chemosphere, 226, 110-122. [CrossRef]
- Tripathi A, Liu S, Singh PK, Kumar N, Pandey AC, Tripathi DK, Sahi S (2017) Differential phytotoxic responses of silver nitrate (AgNO3) and silver nanoparticle (AgNps) in Cucumis sativus L. Plant Gene, 11, 255-264. [CrossRef]
- Tripathi DK, Mishra RK, Singh S, Singh S, Vishwakarma K, Sharma S, Chauhan DK (2017) Nitric oxide ameliorates Zn oxide nanoparticles phytotoxicity in wheat seedlings: implication of the ascorbate–glutathione cycle. Frontiers in plant science, 8, 1. [CrossRef]
- Tripathi DK, Singh S, Singh S, Pandey R, Singh VP, Sharma NC, Chauhan DK (2017) An overview on manufactured nanoparticles in plants: uptake, translocation, accumulation and phytotoxicity. Plant physiology and biochemistry, 110, 2-12. [CrossRef]
- Van Dijk M, Morley T, Rau ML, Saghai Y (2021) A meta-analysis of projected global food demand and population at risk of hunger for the period 2010–2050. Nature Food, 2(7), 494-501. [CrossRef]
- Veena M, Puthur JT (2022) Seed nutripriming with Zn is an apt tool to alleviate malnutrition. Environmental Geochemistry and Health, 44(8), 2355-2373.
- Venkatachalam P, Jayaraj M, Manikandan R, Geetha N, Rene ER, Sharma NC, Sahi SV (2017) Zn oxide nanoparticles (ZnONPs) alleviate heavy metal-induced toxicity in Leucaena leucocephala seedlings: a physiochemical analysis. Plant Physiology and Biochemistry, 110, 59-69. [CrossRef]
- Vinopal S, Ruml T, Kotrba P (2007) Biosorption of Cd2+ and Zn2+ by cell surface-engineered Saccharomyces cerevisiae. International Biodeterioration & Biodegradation, 60(2), 96-102. [CrossRef]
- Wang Z, Xie X, Zhao J, Liu X, Feng W, White JC, Xing B (2012) Xylem-and phloem-based transport of CuO nanoparticles in maize (Zea mays L.). Environmental science & technology, 46(8), 4434-4441. [CrossRef]
- Yasmeen F, Razzaq A, Iqbal M, Jhanzab HM (2015) Effect of silver, copper and iron nanoparticles on wheat germination. Int. J. Biosci, 6(4), 112-117. [CrossRef]
- Younas N, Fatima I, Ahmad IA, Ayyaz MK (2023) Alleviation of Zn deficiency in plants and humans through an effective technique; biofortification: A detailed review. Acta Ecologica Sinica, 43(3), 419-425. [CrossRef]
- Zhai G, Walters KS, Peate DW, Alvarez PJ, Schnoor JL (2014). Transport of gold nanoparticles through plasmodesmata and precipitation of gold ions in woody poplar. Environmental science & technology letters, 1(2), 146-151. [CrossRef]
- Zhang H, Yue M, Zheng X, Xie C, Zhou H, Li L (2017) Physiological effects of single-and multi-walled carbon nanotubes on rice seedlings. IEEE transactions on nano bioscience, 16(7), 563-570. [CrossRef]
- Zhang P, Guo Z, Zhang Z, Fu H, White JC, Lynch I (2020) Nanomaterial transformation in the soil–plant system: implications for food safety and application in agriculture. Small, 16(21), 2000705. [CrossRef]
- Zhao L, Ortiz C, Adeleye AS, Hu Q, Zhou H, Huang Y, Keller AA (2016) Metabolomics to detect response of lettuce (Lactuca sativa) to Cu (OH) 2 nanopesticides: oxidative stress response and detoxification mechanisms. Environmental Science & Technology, 50(17), 9697-9707. [CrossRef]
Figure 1.
Effects of spraying essential and non-essential elements.
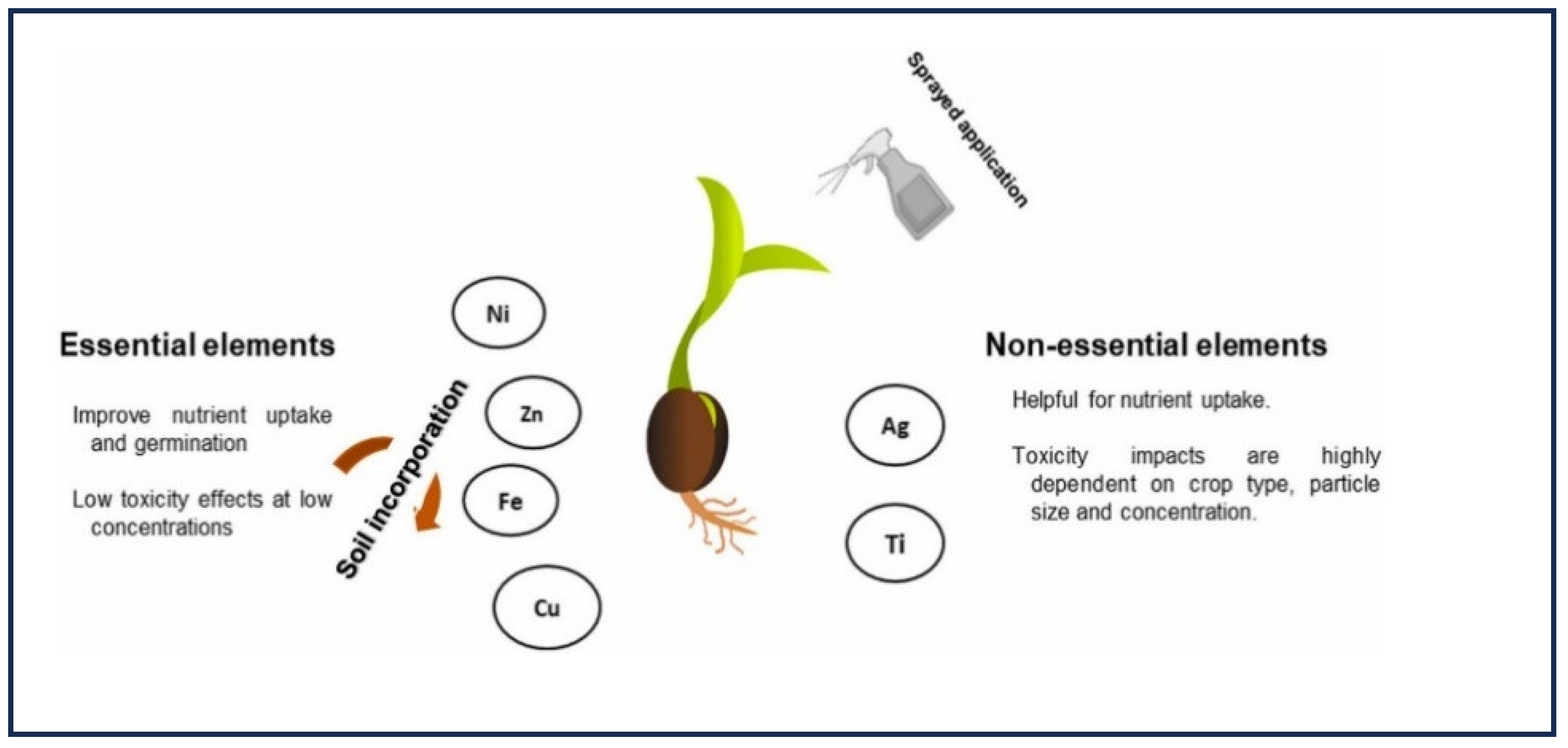
Figure 2.
Nanoparticles and its types.
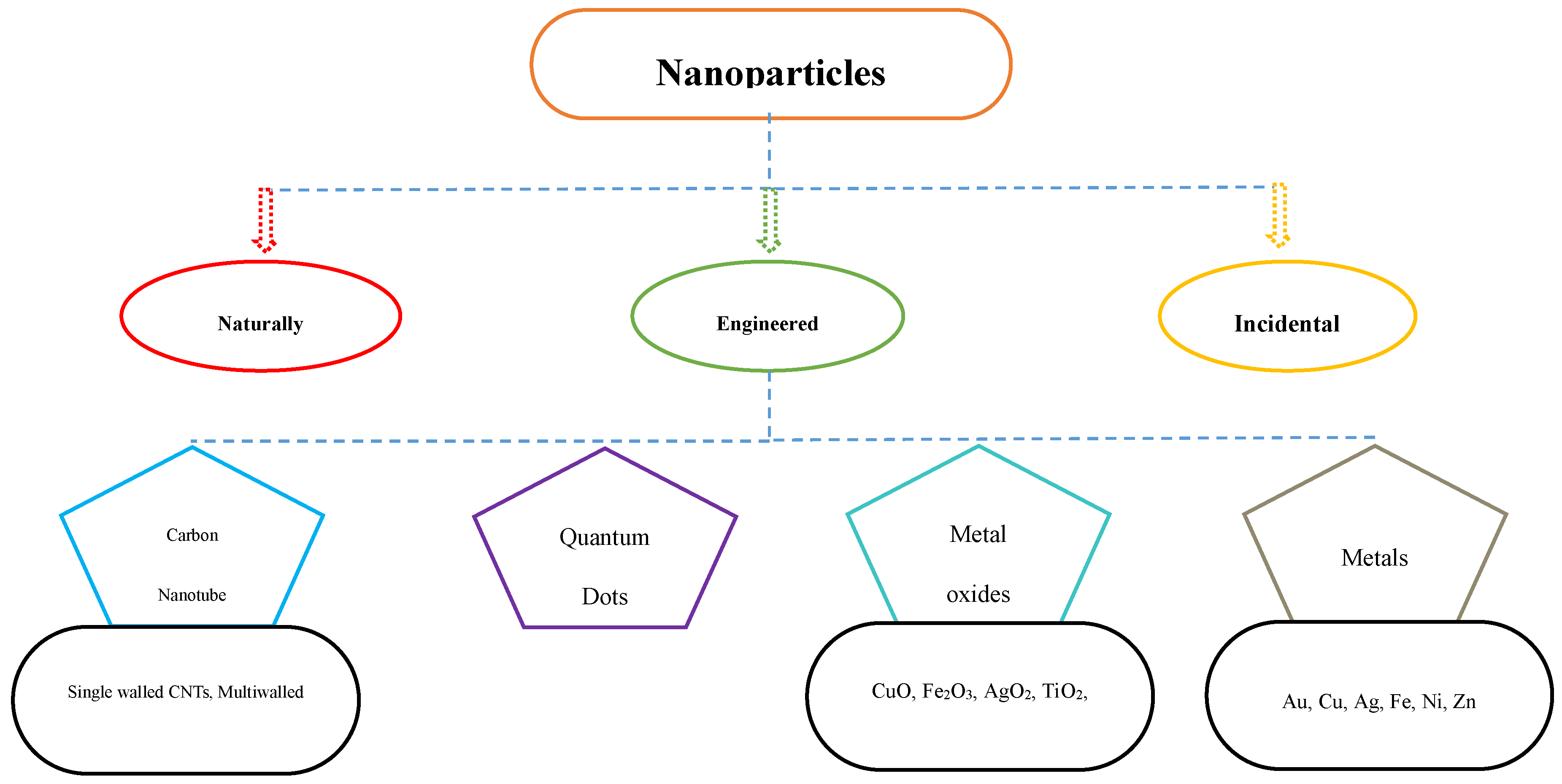
Figure 3.
Foliar applications of NPs and transport via apoplastic and symplastic pathway.
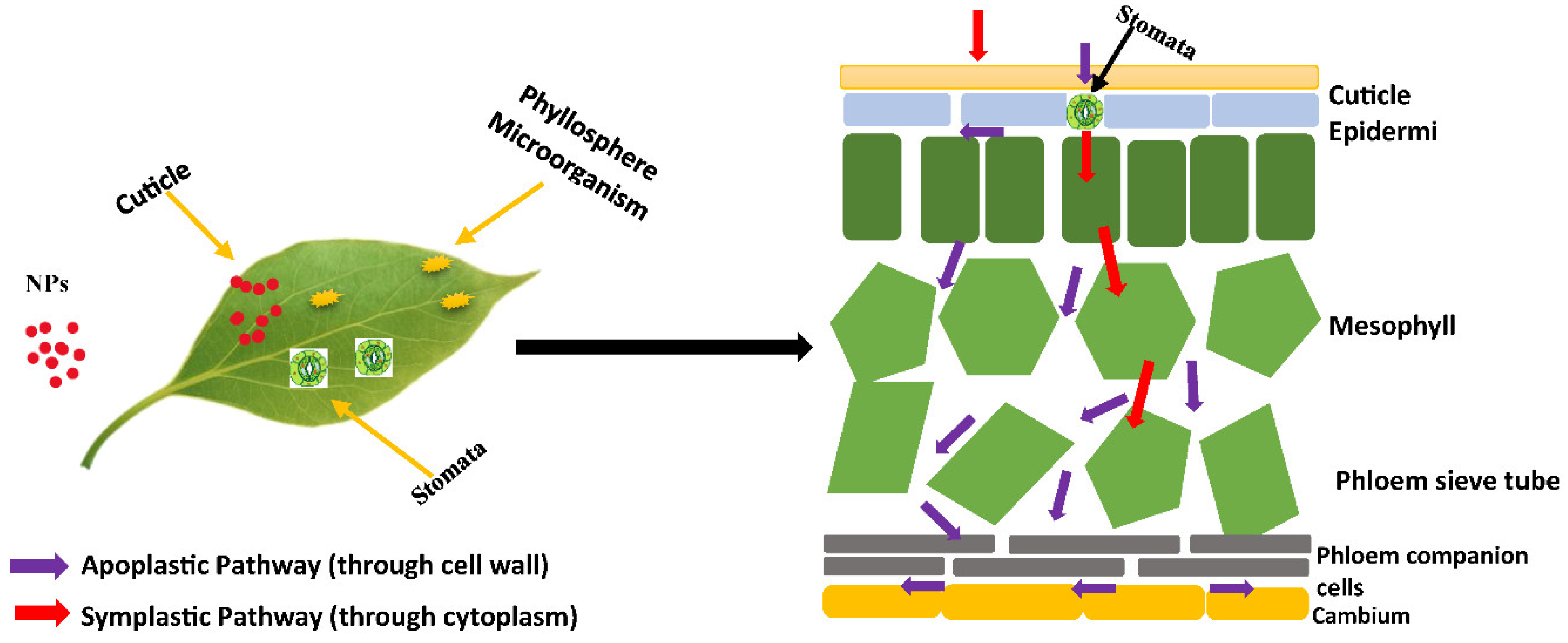
Figure 4.
Root application of NPs and their transport via symplastic and apoplastic pathway.
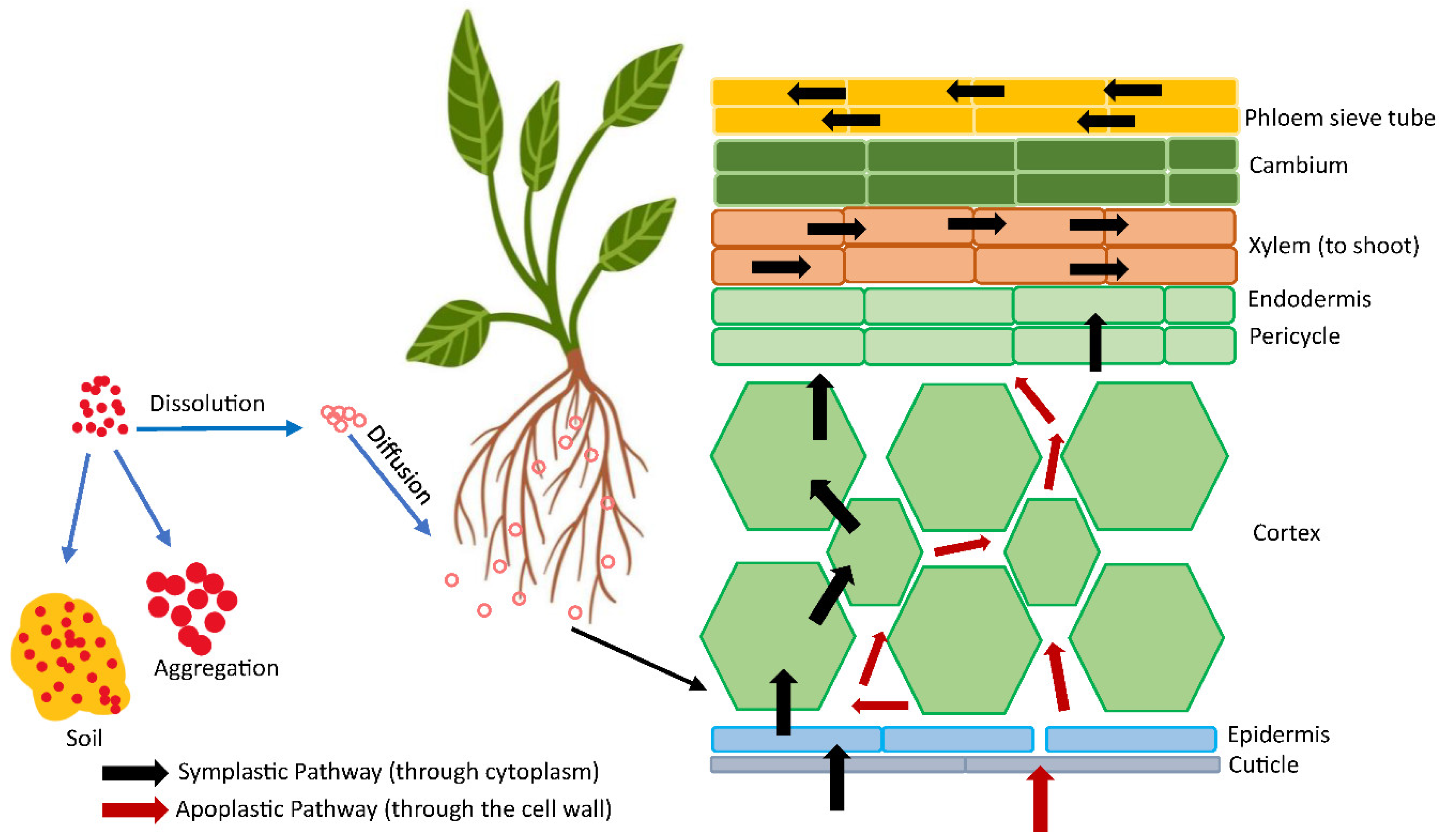
Figure 5.
Effect of foliar and root application of NPs on the growth and yield of plants.
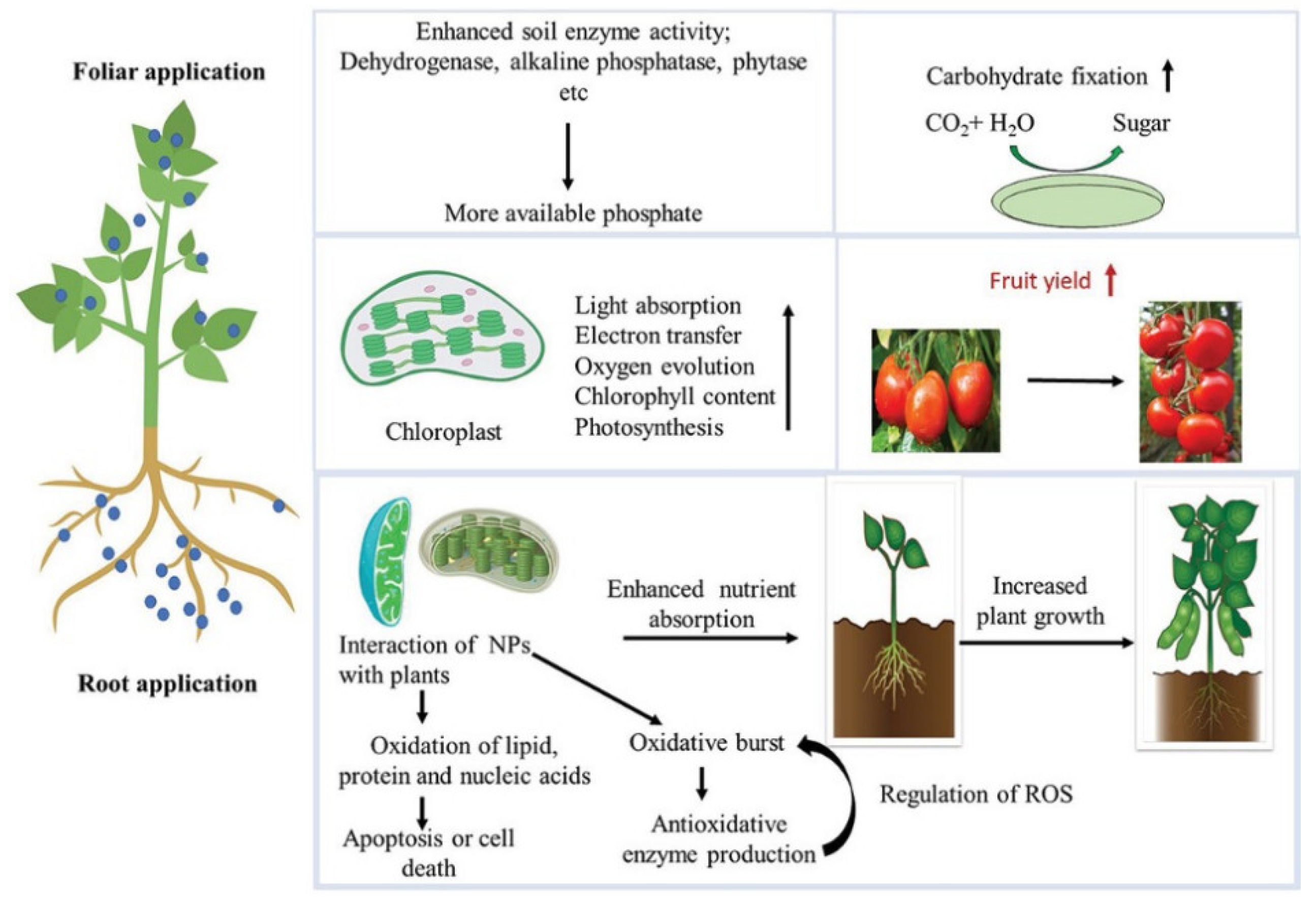

Table 1.
Applications of different NPs on plants to observe the effects in different concentration.
| NPs Application | Size (nm) | Plants | Concentration | Effect | References |
| Soaking | 20 to 45 | Methi | 20 to 30 mg/L | Promoted fresh and dry weights of plant | Elsherif et al. 2023 |
| Adding to soil | 20 to 60 | Wheat | 7mg/kg | Promoted plant height and tillers number | Mazhar et al. 2023 |
| Adding to soil | 20 to 60 | Rice | 10 mg/kg | Enhanced the number of tillers and height of plants | Mazhar et al. 2023 |
| In Petri dishes | 31 | Barley | 1 to 4 mg/L | Improved elongation of root, shoot and germination of seed | Plaksenkova et al. 2021 |
| To soil | More than 100 | Carrot | 1 to 100 mg/kg | Biomass of the plant is increased | Song and Kim, 2020 |
| To soil | More than 100 | Lettuce | 1 to 100 mg/kg | Biomass of the plant is increased | Song and Kim, 2020 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



