
Submitted:
10 October 2024
Posted:
11 October 2024
You are already at the latest version
Abstract
Diabetic retinopathy is the most common retinal vascular disease, affecting the retina's blood vessels and causing chronic inflammation, oxidative stress, and ultimately, vision loss. Diabetes-induced elevated glucose levels increase glycolysis, the main pathway for methylglyoxal (MGO) formation. MGO is a highly reactive dicarbonyl and the most rapid glycation compound to form endogenous advanced glycation end products (AGEs). MGO can act both intra- and extracellularly by glycating molecules and activating the RAGE pathway. This review summarizes the sources of MGO formation and its actions on various cell pathways in retinal cells such as oxidative stress, glycation, autophagy, ER stress, and mitochondrial dysfunction. Finally, the detoxification of MGO by glyoxalases is discussed.
Keywords:
Methylglyoxal
; diabetic retinopathy
; oxidative stress
; inflammation
; autophagy
; ER stress
; glyoxalase
1. Sources of Methylglyoxal and Its Association with Diabetic Retinopathy
Diabetes mellitus (DM) is one of the most widespread diseases today. According to the International Diabetes Federation, 1 in 10 people suffers from this condition [1]. Elevated blood glucose levels cause dysfunction in various tissues, reducing people’s quality of life.
The accumulation of advanced glycation end products (AGEs) during DM progression is associated with biochemical tissue dysfunctions, such as nephropathy, retinopathy, and peripheral neuropathy [2,3,4]. Diabetic retinopathy (DR) results from chronic DM and is the most common retinal vascular disease. Statistics show that three out of four people who have lived with DM for more than 15 years develop DR [5]. Currently, DR is the fifth leading cause of vision impairment. Elevated levels of glucose, blood pressure, and glycated proteins are significantly associated with the DR progression [6]. Common methods to study DR progression in animals include streptozotocin- and alloxan-induced models, high-fat diets (HFD), and genetic approaches to diabetes progression [7,8]. Another potential way to study DR is through the application of AGEs and methylglyoxal (MGO) [9].
MGO is a highly reactive dicarbonyl compound that plays a significant role in the pathogenesis of DM, its metabolic complication, and other age-related chronic inflammatory diseases such as cardiovascular diseases and cancer [10,11,12,13,14]. MGO is formed as a byproduct of glycolysis and is the primary and fastest cause that is known to nonenzymatically modify protein and DNA by glycation and lead to AGEs formation [11,15].
Until now, several pathways of MGO formation have been identified. The major source coming from glucose is via the polyol pathway, where aldose reductase causes glucose reduction to sorbitol, and sorbitol dehydrogenase converts sorbitol to fructose [16]. Aldolase then converts fructose to dihydroxyacetone phosphate. Both aldose reductase and sorbitol dehydrogenase activities are increased in DM conditions [17]. MGO synthase has been found to convert dihydroxyacetone phosphate into MGO irreversibly. Under DM conditions, DHAP levels are significantly elevated to increase MGO formation [18].
MGO can also be produced from the metabolism of proteins and fatty acids although only a small amount [19]. The increased formation of ketones, such as acetone, found in DM can activate CYP2E1 (an acetone monoxidase) in the liver to form MGO. Formation of MGO from proteins is catalyzed by semicarbazide-sensitive amine oxidase (SSAO) which converts aminoacetone to acetone, acetol, and finally to MGO [20]. Figure 1 shows a schematic representation of the major sources of MGO production.
In addition to the generation of MGO from the oxidation of carbohydrates, lipids, and proteins as mentioned above in various tissues, vitamin A-derived lipofuscin pigments bisretinoids are also a unique and novel source of MGO in the eye [21,22]. Upon irradiation with short wavelength visible light, the bisretinoid fluorophores such as A2E and all-trans-retinal dimer are formed as by-products of vitamin A cycling in the retina, and accumulate in retinal pigment epithelial (RPE) cells. Further photooxidation and photodegradation of both bisretinoids lead to the formation of MGO. The elevation of photocleavage products of bisretinoids that especially occurs in aging PRE cells is highly implicated in age-related macular degeneration (AMD) [21,23,24]. MGO production is significantly increased in chronic retinal diseases, such as DR and AMD [12]. In this review, we focus on the pathogenetic role of MGO in DR, and the molecular mechanisms of its actions in retinal cells, majorly in RPE, Müller, vascular, and microglial cells.
2. Modes of Action of MGO in DR
MGO has been shown to affect numerous cellular targets, signaling pathways, and stress responses in retinal cells, leading to impaired retinal function. Two major action modes of MGO, i.e., direct non-enzymatic glycation and RAGE pathway activation, are integrated and contribute to its pathogenic actions.
2.1. MGO-Induced Retina Proteins Glycation and Oxidative Stress
Reactive dicarbonyls play a crucial role in protein glycation due to their high reactivity. Glycation adducts, such as hydroimidazolone AGEs, are produced from MGO and are the major AGEs formed in the retina, nerves, glomeruli, and plasma proteins [3]. The high levels of protein glycation via the AGEs/MGO pathway, especially Nε-carboxymethyl lysine adducts in serum, vitreous, and retina are correlated with the progression and various secondary complications of DM [25,26,27]. These include neuropathy of the retina [2,28,29,30,31], nephropathy [32,33], impaired wound healing [34], cardiovascular complications [12,26,35,36], salivary protein glycation [37], and neurodegeneration [29]. MGO is also suggested to be involved in insulin resistance and beta-cell dysfunction, contributing to DM development and creating a feedback loop between glycation and hyperglycemia [26,32]. Protein glycation in the retina contributes to vision loss during DR progression. The accumulation of glycated proteins in the lens causes blurred vision in the early stages of DR and may lead to complete vision loss in advanced stages [38].
Besides the well-known glycated hemoglobin (HbA1c) which is a disease progression marker of DM, there are several identified glycated proteins of AGEs that contribute to abnormalities in retinal cell pathways, ROS formation, and cytotoxicity [39]. These include crystallins, albumin, low-density lipoprotein (LDL), and extracellular matrix. Water-soluble lens proteins, such as crystallins, are crucial for maintaining retinal transparency and participating in metabolic and regulatory functions [40]. Abdullah et al. found that MGO-induced glycation of camel lens ζ-crystallin alters its secondary structure, and reduces its solubility. They also found that MGO can glycate proteins more rapidly than high glucose levels [41]. Moreover, α, β-crystallin in RPE cells exhibits anti-apoptotic activity against MGO treatment. The interaction between α, β -crystallin and caspase subtypes -2L, -2S, -3, -4, -7, -8, -9, and -12 can be disrupted by MGO treatment, causing caspase release and cellular apoptosis [42]. Albumin glycation that forms under hyperglycemic conditions in the retinal blood vessels of diabetic patients is involved in DR. Glycated albumin can induce cell death in retinal pericytes via ROS production [43], retinal RPE dysfunction [44], vascular injury [45], blood-retinal barrier permeabilization [46], and inflammation in retinal microglia [47]. Glycated LDL may mediate capillary injury in DR [48]. Moreover, glycated extracellular matrix proteins, such as fibronectin and laminin, are detected in diabetic animals [49], and in endothelial and Müller cells in the retinas of patients with DR [50].
Oxidative stress is the most important key contributor to the pathogenesis of DR and AGE-induced retinal injury [51]. Accumulating lines of evidence have demonstrated that AGEs act as pro-oxidant metabolites, which leads elevation of intracellular ROS and lipoxidation [52]. All these studies suggest that antioxidants and inhibitors of advanced glycation such as Nrf2 activators and pigment epithelium-derived factor (PEDF) are therapeutic strategies to ameliorate DR [46,53,54].
2.2. MGO-Induced RAGE Activation
MGO’s mode of action is not limited to protein glycation but also involves the activation of the receptor for AGEs (RAGE). The AGEs-RAGE axis can trigger a range of signaling events that are associated with diabetes [55,56]. The RAGE activation in RPE and Müller cells induces the NF-κB, PI3K/AKT/GSK3B, Ras/MEK/ERK, p38, and JNK pathways and increases NADPH oxidase activity [57, 58
[57,58]. Moreover, RAGE activation by AGEs also mediates cellular dysfunction and apoptosis in RPE cells. [13,59]. Not only in the retina, the involvement of the MGO-RAGE axis in multiple pathological consequences related to neurodegeneration has been demonstrated in the brain, where RAGE activation mediates disruption of the blood-brain barrier, neuroinflammation, remodeling of extracellular matrix, dysregulation of polyol pathway and antioxidant enzymes [60].
Studies further show the positive feedback regulation of the AGEs-RAGE pathway and the interplay between RAGE and TLR4 in the pathogenesis of DR. In a diabetic mouse model, it was found that diabetes progression and MGO accumulation can activate and upregulate RAGE expression in the retina [61]. This event might result from the ROS-NF-κB axis [62,63]. Ramya et al. found that MGO and its induced AGEs promote inflammation and TLR4 expression in endothelial cells via the RAGE pathway [64]. Moreover, high mobility group box 1 (HMGB1) is an inflammatory alarmin to initiate the host defense system. Studies indicate the role of HMGB1 in the pathogenesis of type 2 DM depends on the activation of the RAGE and TLR4 molecules that contribute to the production of pro-inflammatory cytokines [65,66]. Moreover, AGEs can also increase HMGB1 secretion in retinal ganglion cells, which in turn activates RAGE for VEGF-A production [67]. With these findings, manipulation of RAGE activity by either reducing RAGE expression [68] or antagonizing RAGE [69,70] becomes a promising strategy to prevent AGEs-induced retinal cell damage and the progression of diabetic complications.
3. MGO Alters Various Cellular Pathways
3.1. Inhibition of Autophagy in Retinal Cells
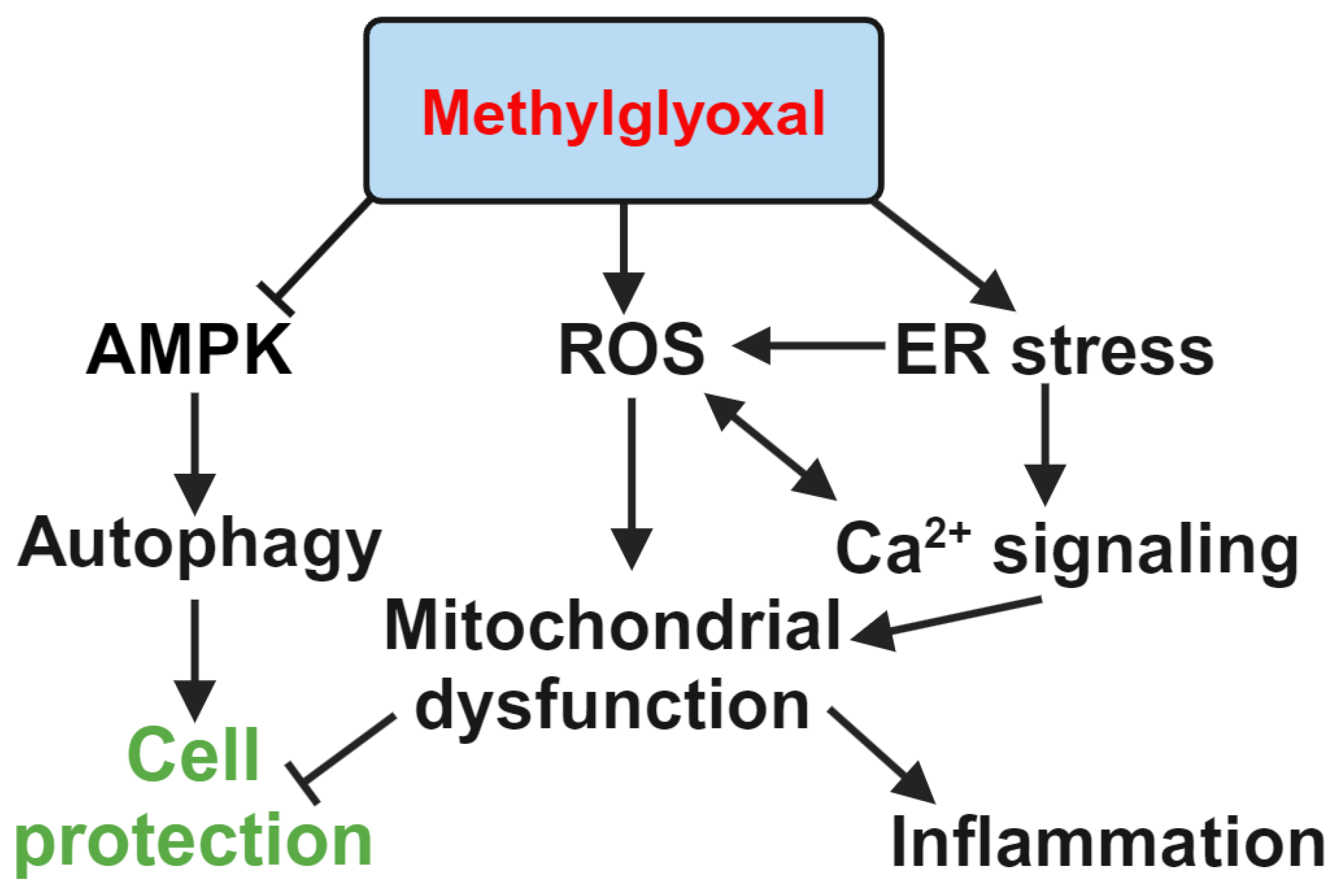
Autophagy is a catabolic process that degrades and recycles damaged organelles, cellular components, and cytoplasmic proteins to maintain cellular homeostasis. The primary function of RPE cells is to maintain healthy photoreceptors [71]; however, their functions also include light absorption, phagocytosis, barrier function, and participation in the retinoid cycle. Disturbed autophagy accelerates the progression of age-related macular degeneration [72]. RPE cells dysfunction can be caused by oxidative stress [73,74], senescence [75,76], type 2 DM [77,78], and MGO [2,9,79,80]. Impaired RPE function promotes photoreceptor damage and other cellular injuries, increases chronic inflammation, and ultimately leads to vision loss [81]. RPE cells exhibit high levels of autophagy proteins to maintain their phagocytic functions [82,83]. Normal autophagy levels prevent RPE cell dysfunction, as demonstrated in the studies of MGO-, UVA-, and NaIO3-induced damage [79,84,85]. Abnormal levels of autophagy, either decreased [85] or increased [86], promote cellular damage.
In models of high glucose and diabetes, decreased phagocytosis and autophagy levels are linked to the cellular signaling interplay between AMPK, AKT, and mTOR pathways. High glucose treatment in ARPE-19 cells reduces AMPK but increases AKT activities, downregulating mTOR-related autophagy gene expression [87,88]. Feng et al. demonstrated the protective role of autophagy in DR. RPE cells exposed to high glucose shows decreased autophagy levels and lysosomal membrane permeabilization, which is abolished by HMGB1 silencing-dependent restoration of the degradative capacity of autophagy [89]. In our previous work, we found that MGO treatment causes significant apoptosis in ARPE-19 cells [79]. MGO treatment affects autophagy proteins similarly to high glucose, downregulating the LC3II/LC3I ratio via AMPK suppression, suggesting that MGO-induced autophagy downregulation in RPE cells promotes cell death.
3.2. Induction of Oxidative Stress-Associated Inflammation in Retinal Cells
Inflammation induced by various stress factors, along with related oxidative stress, leads to cellular dysfunction. High glucose levels have been shown to promote oxidative stress and inflammation in RPE cells [90,91], and both ROS and/or reactive nitrogen species (RNS) production can be induced by high glucose in RPE cells [92,93], retinal ganglion cells [94], photoreceptors cells [95], and retinal microvascular endothelial cells [96]. Likewise, MGO can increase the production of ROS and RNS, damaging cellular components, leading to mitochondrial dysfunction, cell death, and inflammation in various cell types including in PRE [79,97], retinal pericytes [98], endothelial cells [99], eosinophils [100], macrophages [101], and hepatocytes [102].
MGO has also been found to induce inflammation responses in the retina of humans and rats with DM. Ma et al. demonstrated that MGO treatment elevates the expression of cyclooxygenase (COX)-2, chemokine receptor CXCR4, IL-6, IL-8, monocyte chemoattractant protein-1 (MCP-1), and ICAM-1 genes [103]. MGO-modified fibronectin was shown to upregulate CD40, intercellular adhesion molecule 1 (ICAM-1), and CCL2 in endothelial and Müller cells, leading to enhanced CD40-dependent pro-inflammatory responses. Moreover, increased CD40 expression in endothelial and Müller cells from patients with DR was observed by confocal microscopy, which was associated with increased carboxymethyl lysine expression in fibronectin and laminin [50]. In human retinal endothelial cells, MGO also can upregulate lysyl oxidase expression via RAGE, leading to proinflammatory and matrix stiffening [104]. Recently Wang et al. reported that MGO can induce pyroptosis in endothelial cells via NLRP3 inflammasome activation and oxidative stress [99]. Moreover, MGO displays the ability to reduce the immunosuppressive activity of retinal pericytes to inhibit activated T cell proliferation via expressing PD-L1. This finding reveals that the protective function of pericytes against inflammation-mediated apoptosis in the retina is impaired by hyperglycemic conditions [105]. As mentioned above, MGO-induced inflammation might result from the direct activation of RAGE and the consequential activation of TLR [64,100].
3.3. Inflammation and Microglia Activation
Müller cells are the principal macroglial cells of the retina. Their main functions include providing structural support (by maintaining the structural integrity of retinal layers), regulating the retinal environment (by balancing ions, removing excess neurotransmitters like glutamate, and maintaining K⁺ concentrations in retinal tissue), light conduction, nutrient transport, and neuroprotection (by secreting neurotrophic factors) [106]. Microglia are the resident immune cells of the CNS, including the retina, and play a critical role in responding to injury, infection, and other pathological changes while maintaining retinal homeostasis [107]. Studies indicate that Müller cells and microglia work together to maintain retinal homeostasis, with Müller cells playing a role in regulating microglial activation and function [108]. This cell-cell interaction is crucial for maintaining a balanced immune response in the retina [109]. However, in DR, microglia become over-activated and contribute to disease progression. The inflammatory response induced by activated microglia can damage the BRB, leading to increased vascular permeability [110], which contributes to retinal edema and hemorrhage, the hallmark features of DR [111]. Persistent activation of microglia in the diabetic retina leads to a chronic inflammatory state, exacerbating retinal damage and accelerating the progression of DR from the non-proliferative to the proliferative stage.
Reber et al. demonstrated that glyoxal causes significant morphological changes in the E1A-NR3 retinal neuron cell line, including cell membrane blebbing, aggregation of intracellular organelles, time-dependent acidification (to pH 7.2), loss of mitochondrial membrane potential, and apoptosis [111]. MGO and high glucose conditions activate Müller cells CD40 receptor and produce pro-inflammatory cytokines (TNF-α, IL-1β, CCL2) via upregulation of the CD40/MAPK/NF-κB pathway [50,112]. Portillo et al. further highlight the role of stress-induced Müller cells-microglia activation (Figure 2). CD40 activation not only promotes cytokine synthesis but also stimulates the TRAF2/Src/PLCγ1 pathway, leading to ATP release from Müller cells, activation of microglial P2X7 receptors, and their chronic activation [113,114,115].
Vascular endothelial growth factor (VEGF), an angiogenic factor associated with abnormal blood vessel formation, is mostly produced by activated Müller cells in the retina [116]. In a zebrafish embryo model of MGO-induced retinal angiogenesis, MGO was found to promote angiogenesis through VEGF production [117]. These MGO-induced changes are reversed by the VEGF inhibitor PTK787, proving the important role of VEGF synthesis under MGO treatment. Bautista-Pérez et al. further confirm this hypothesis in a streptozotocin-induced diabetic rat model [27]. They show that high glucose levels in diabetic conditions lead to a pro-inflammatory state in retinal cells via the RAGE/NF-κB pathway, which is also linked to the production of VEGF and the progression of proliferative DR.
3.4. Endoplasmic Reticulum (ER)-Stress And Calcium Signaling
The ER is an intracellular Ca²⁺ storage organelle that plays a crucial role in signal transduction and protein folding. ER stress primarily triggers three major signaling pathways: IRE1α/XBP1, PERK/eIF2α/ATF4, and ATF6. It can also lead to apoptosis and cell growth arrest via the CHOP protein [118,119]. ER stress plays a significant role in the progression of DR [103]. In our previous work, we found that MGO induces RPE cell death through ER stress-related ROS production and mitochondrial dysfunction [80]. MGO triggers both necrotic and apoptotic cell death modes. MGO increases the expression of proteins such as GRP78, CHOP, ATF6, and ATF4, along with phosphorylation of eIF2α and PERK, and induces spliced XBP1 and ATF6 formation in a time-dependent manner. In the first 2 hours following MGO treatment, cellular and mitochondrial ROS levels increase. N-acetylcysteine can protect RPE cells from MGO-induced cell death. Additionally, 4-PBA, salubrinal (ER stress inhibitors), and BAPTA/AM (calcium chelator) can also prevent cell death, suggesting the involvement of ER stress and calcium signaling [80]. Furthermore, we demonstrated that ER stress inhibitors can reduce ROS production, prevent intracellular calcium level increases, and protect against MMP loss. Additionally, store-operated calcium entry (SOCE) inhibitors, MRS1845 and YM-58483, but not the IP3 receptor inhibitor xestospongin C, were able to block MGO-induced ROS production, MMP loss, and the sustained increase in intracellular calcium in RPE cells. These findings suggest that MGO is capable of inducing an ER stress response, leading to calcium signaling changes, mitochondrial dysfunction, and cell death [80].
3.5. AMP-Activated Protein Kinase and Mitochondrial Stability
AMPK is one of the key proteins involved in metabolism and energy homeostasis [120]. Impaired glucose metabolism in diabetic conditions leads to reduced glucose uptake and an altered ATP/ADP ratio, which downregulates AMPK activity [121]. It is also known that AMPK activation induces antioxidant responses through the PGC-1α, FOXO, and Nrf2 pathways. The interplay between SIRT1 and AMPK promotes antioxidant responses in diabetic conditions [122]. AMPK also plays a role in lipid metabolism. Reduced AMPK activity is implicated in lipid dysregulation, including dysregulated de novo lipogenesis, elevated acetyl-CoA carboxylase signaling, and increased fatty acid synthase activity, all of which contribute to lipotoxicity [123].
MGO-modified proteins cause mitochondrial dysfunction and oxidative stress and inhibit autophagy in AMD. Impaired autophagy accelerates disease progression by promoting retinal cell death and inflammation via the NLRP3 inflammasome pathway. Metformin, an AMPK activator that promotes autophagy can inhibit the senescent cell phenotype during AMD progression through PGC-1α, SIRT1, and AMPK pathways [72].
Zou et al. found that Wnt inhibitory factor 1 (WIF1) can suppress VEGF expression, mitochondrial autophagy-related proteins, and mitochondrial dysfunction in high glucose-treated ARPE-19 cells. An in vivo study showed that WIF1 reduces DR by downregulating the AMPK/mTOR axis [124]. In our previous study on MGO-induced ARPE-19 cell death and retinopathy, the critical role of AMPK signaling is also observed [97]. MGO inhibits AMPK activity and autophagy (indicated by LC3II accumulation) while decreasing the expression of mitochondrial biogenesis and dynamics markers (MFN1, PGC-1α, and TFAM). These changes lead to mitochondrial fission and reduce mitochondrial biogenesis. Both AMPK activators (metformin and A769662) reduce these effects of MGO, which is confirmed using AMPK inhibitor (compound C) and siAMPK transfection. Additionally, AMPK activity is also involved in the upregulation of the Glo1 protein. Animal study confirms our findings, showing that intravitreal injection of MGO causes cotton wool spots and macular edema. However, metformin and A769662 treatments in animal groups provide protective effects against MGO. Functional, histological, and optical coherence tomography analyses support the protective actions of AMPK activators against MGO-induced retinal damage. Song et al. demonstrate AMPK’s protective effect in AGEs-induced apoptosis in photoreceptor 661W cells and streptozotocin-induced photoreceptor cell degeneration [125]. MGO and AGEs treatments in 661W cells induce apoptosis by promoting Bax but reducing Bcl-2 protein levels, as well as disrupting autophagy. AGEs treatment also causes mitochondrial dysfunction, abnormal mitochondrial morphology, downregulation of mitochondrial biogenesis-associated proteins such as TFAM, Nrf1, and PGC-1α, and antioxidant protein expression. These changes in 661W cells are prevented by metformin pretreatment, suggesting AMPK’s protective role. The same effect is confirmed in a diabetes-induced DR model in mice, which is also prevented by metformin administration [126].
4. Role of Glyoxlases in MGO Detoxification
There are several detoxification systems for MGO in the body, including kidney clearance, aldehyde dehydrogenase, aldo-keto reductase, and the glyoxalase pathways. The glyoxalase system is considered the most important for MGO detoxification in the retina. The first step of MGO detoxification is its reaction with glutathione for the hemithioacetal formation. Glyoxalase 1 (Glo1) is a Zn²⁺-dependent enzyme that converts hemithioacetal to S-D-lactoylglutathione. It is regulated by proteins such as E2F4, NF-κB, Nrf2, and PKA [127]. Nrf2 acts as an inducer of Glo1, promoting its expression. PKA can phosphorylate Glo1 at Thr107, which decreases its activity and leads to caspase-dependent cell death [128].
Glyoxalase 2 (Glo2) exists in both cytosolic and mitochondrial fractions, where it hydrolyzes S-D-lactoylglutathione to produce D-lactate and regenerates glutathione [129]. Glo2’s catalytic domain contains Fe²⁺ and Zn²⁺, but only Zn²⁺ can regulate its activity. Glo2 also has a p53-responsive region that is activated by p63 and p73, increasing its expression [130]. Lastly, the glyoxalase 3 (Glo3) protein, which converts MGO to D-lactate without the need for glutathione, is present in bacteria.
Figure 4.
Role of glyoxalase system in MGO detoxification.
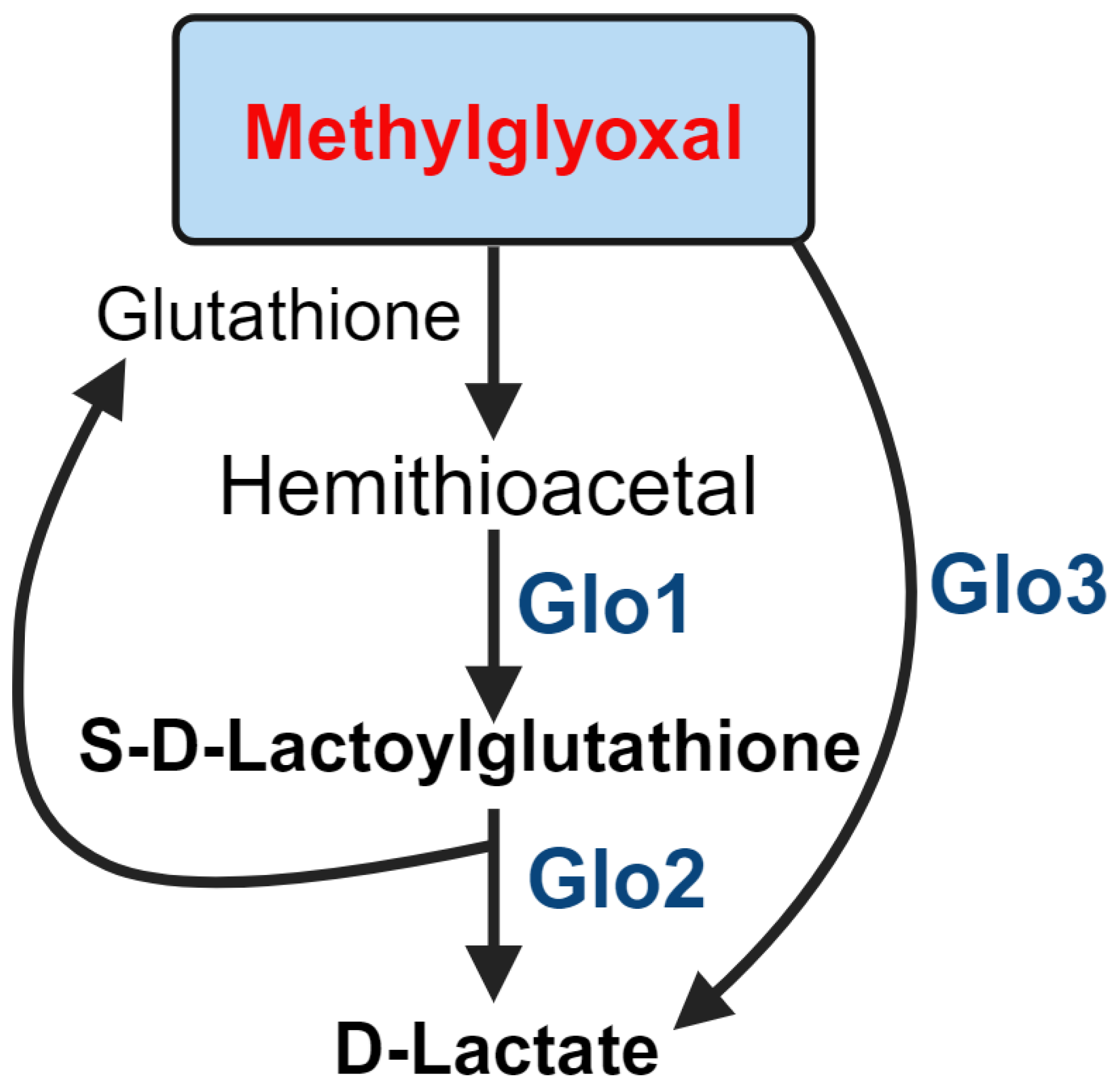
The MGO detoxification glyoxalase system is closely linked to diabetes complications, including nephropathy, retinopathy, neuropathy, and cardiovascular disease [131]. Increased MGO formation in hyperglycemia is associated with the downregulation of Glo1 protein due to inflammatory signaling [132]. In DR, Glo1 expression and activity are downregulated in diabetic patients [133]. It has been found that knocking out transient receptor potential cation channels (TRPC) 1/4/5/6 isoforms in mice increases Glo1 activity and expression in diabetic conditions, protecting the retina from DR-related changes and reducing MGO formation [134].
Several studies on manipulating Glo1 levels in the cell and animal models have demonstrated its crucial role against diabetes. In zebrafish, CRISPR-Cas9 knockout of Glo1 potentiates HFD-induced MGO formation, elevates fasting glucose levels, impairs glucose tolerance, and promotes new retinal blood vessel formation [135]. Bernel et al. showed that Glo1 overexpression in streptozotocin-treated rats prevents DR-related retinal damage by reducing AGEs formation, decreasing GFAP levels, increasing Kir4.1 protein expression in Müller cells, and reducing the formation of new blood vessels in the retina [136]. In Drosophila melanogaster, Glo1 knockout accelerates diabetes progression, increases MGO concentration, promotes lipid accumulation, elevates blood glucose levels, and decreases insulin sensitivity [137]. The Glo1 inducer tRES-HESP (a combination of trans-resveratrol and hesperetin) has been shown to reduce the expression of RAGE and cell adhesion molecules, as well as decrease inflammation in human aortic endothelial cells [138]. In fibroblasts and HepG2 cells, tRES-HESP can reduce basal levels of RAGE and MMP3 proteins and increase glutathione levels [139].
5. Conclusions
DR is one of the most common retinal vascular diseases promoting new blood vessel formation, chronic inflammation, and oxidative stress. Increased glucose levels and impaired lipids synthesis promote reactive dicarbonyl MGO formation. MGO is the most rapid glycation compound, which acts both intra- and extracellularly by non-enzymatically glycating proteins and activating the RAGE pathway, respectively. Its action causes several cellular responses that lead to retinal cell dysfunction and cell death. These include autophagy reduction, oxidative and ER stress, mitochondrial dysfunction, cellular calcium overload, AMPK inhibition, RAGE activation, and inflammation. In addition to the supplementation of antioxidants to inhibit AGEs formation and oxidative stress-mediated cell dysfunction, towards newer molecular targets on glyoxlase, glycation, RAGE, and aldose reductase for chronic DR is a new research field.
Author Contributions
Conceptualization, VK, CMC, and WWL; investigation, VK; writing—original draft preparation, VK; writing—review and editing, VK, WWL, CMC; visualization, VK; supervision, WWL and CMC; funding acquisition, CMC. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Science and Technology Council, grant number NSTC113-2314-B-567-001.
Data Availability Statement
Data used for this review is available online on PubMed.
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- International Diabetes Federation. IDF Diabetes Atlas, 10th edn. Brussels, Belgium: International Diabetes Federation, 2021.
- Milne, R.; Brownstein, S. Advanced glycation end products and diabetic retinopathy. Amino Acids 2013, 44, 1397–1407. [Google Scholar] [CrossRef] [PubMed]
- Karachalias, N.; Babaei-Jadidi, R.; Ahmed, N.; Thornalley, P.J. Accumulation of fructosyl-lysine and advanced glycation end products in the kidney, retina and peripheral nerve of streptozotocin-induced diabetic rats. Biochem. Soc. Trans. 2003, 31 Pt 6, 1423–1425. [Google Scholar] [CrossRef] [PubMed]
- Zong, H.; Ward, M.; Stitt, A.W. AGEs, RAGE, and diabetic retinopathy. Curr. Diab. Rep. 2011, 11, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Fung, T.H.; Patel, B.; Wilmot, E.G.; Amoaku, W.M. Diabetic retinopathy for the non-ophthalmologist. Clin. Med. (Lond.) 2022, 22, 112–116. [Google Scholar] [CrossRef] [PubMed]
- Hainsworth, D.P.; Bebu, I.; Aiello, L.P.; Sivitz, W.; Gubitosi-Klug, R.; Malone, J.; White, N.H.; et al. Risk factors for retinopathy in type 1 diabetes: the DCCT/EDIC study. Diabetes Care 2019, 42, 875–882. [Google Scholar] [CrossRef]
- Olivares, A.M.; Althoff, K.; Chen, G.F.; Wu, S.; Morrisson, M.A.; DeAngelis, M.M.; Haider, N. Animal Models of Diabetic Retinopathy. Curr. Diab. Rep. 2017, 17, 93. [Google Scholar] [CrossRef]
- Sadikan, M.Z.; Abdul Nasir, N.A.; Lambuk, L.; Mohamud, R.; Reshidan, N.H.; Low, E.; Singar, S.A.; et al. Diabetic retinopathy: a comprehensive update on in vivo, in vitro and ex vivo experimental models. BMC Ophthalmol. 2023, 23, 421. [Google Scholar] [CrossRef]
- Schlotterer, A.; Kolibabka, M.; Lin, J.; Acunman, K.; Dietrich, N.; Sticht, C.; Fleming, T.; et al. Methylglyoxal induces retinopathy-type lesions in the absence of hyperglycemia: studies in a rat model. FASEB J. 2019, 33, 4141–4153. [Google Scholar] [CrossRef]
- Bellier, J.; Nokin, M.J.; Larde, E.; Karoyan, P.; Peulen, O.; Castronovo, V.; Bellahcene, A. Methylglyoxal, a potent inducer of AGEs, connects between diabetes and cancer. Diabetes Res. Clin. Pract. 2019, 148, 200–211. [Google Scholar] [CrossRef]
- Schalkwijk, C.G.; Stehouwer, C.D.A. Methylglyoxal, a Highly Reactive Dicarbonyl Compound, in Diabetes, Its Vascular Complications, and Other Age-Related Diseases. Physiol. Rev. 2020, 100, 407–461. [Google Scholar] [CrossRef]
- Mukohda, M.; Okada, M.; Hara, Yukio. ; Yamawaki, H. Exploring Mechanisms of Diabetes-Related Macrovascular Complications: Role of Methylglyoxal, a Metabolite of Glucose on Regulation of Vascular Contractility. J. of Pharmacol. Sci. 2012, 118, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Fleming, T.H.; Humpert, P.M.; Nawroth, P.P.; Bierhaus, A. Reactive metabolites and AGE/RAGE-mediated cellular dysfunction affect the aging process: a mini-review. Gerontology 2011, 57, 435–443. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.I.; Ashfaq, F.; Alsayegh, A.A.; Hamouda, A.; Khatoon, F.; Altamimi, T.N.; Alhodieb, F.S.; Beg, M.M.A. Advanced glycation end product signaling and metabolic complications: Dietary approach. World J. Diabetes 2023, 14, 995–1012. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, A.L.; de Oliveira, M.G.; Mónica, F.Z.; Antunes, E. Methylglyoxal and Advanced Glycation End Products (AGEs): Targets for the Prevention and Treatment of Diabetes-Associated Bladder Dysfunction? Biomedicines 2024, 12, 939. [Google Scholar] [CrossRef]
- Yan, L.J. Redox imbalance stress in diabetes mellitus: Role of the polyol pathway. Animal Model Exp. Med. 2018, 1, 7–13. [Google Scholar] [CrossRef]
- Tang, W.H.; Martin, K.A.; Hwa, J. Aldose reductase, oxidative stress, and diabetic mellitus. Front. Pharmacol. 2012, 3, 87. [Google Scholar] [CrossRef]
- Kalapos, M.P. Methylglyoxal and glucose metabolism: a historical perspective and future avenues for research. Drug Metabol. Drug. Interact. 2008, 23(1-2), 69-91. [CrossRef]
- Yu, P. H.; Wright, S.; Fan, E. H.; Lun, Z. R.; Gubisne-Harberle, D. Physiological and pathological implications of semicarbazide-sensitive amine oxidase. Biochim. Biophys. Acta 2003, 1647, 193–199. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information (2024). PubChem Pathway Summary for Pathway PWY-5451, acetone degradation I (to methylglyoxal), Source: BioCyc. Retrieved October 3, 2024 from https://pubchem.ncbi.nlm.nih. 3 October 5451.
- Yoon, K.D.; Yamamoto, K.; Ueda, K.; Zhou, J.; Sparrow, J.R. A novel source of methylglyoxal and glyoxal in retina: implications for age-related macular degeneration. PLoS One 2012, 7, e41309. [Google Scholar] [CrossRef]
- Ueda, K.; Zhao, J.; Kim, H.J.; Sparrow, J.R. Photodegradation of retinal bisretinoids in mouse models and implications for macular degeneration. Proc. Natl. Acad. Sci. USA 2016, 113, 6904–6909. [Google Scholar] [CrossRef]
- Wu, Y.; Yanase, E.; Feng, X.; Siegel, M.M.; Sparrow, J.R. Structural characterization of bisretinoid A2E photocleavage pro Alouffi ducts and implications for age-related macular degeneration. Proc. Natl. Acad. Sci. USA 2010, 107, 7275–7280. [Google Scholar] [CrossRef]
- Zhou, J.; Ueda, K.; Zhao, J.; Sparrow, J.R. Correlations between Photodegradation of Bisretinoid Constituents of Retina and Dicarbonyl Adduct Deposition. J. Biol. Chem. 2015, 290, 27215–27227. [Google Scholar] [CrossRef] [PubMed]
- Choudhuri, S.; Dutta, D.; Sen, A.; Chowdhury, I.H.; Mitra, B.; Mondal, L.K.; Saha, A.; et al. Role of N-ε- carboxy methyl lysine, advanced glycation end products and reactive oxygen species for the development of nonproliferative and proliferative retinopathy in type 2 diabetes mellitus. Mol. Vis. 2013, 19, 100–113. [Google Scholar] [PubMed]
- Rabbani, N.; Xue, M.; Thornalley, P.J. Hexokinase-2-Linked Glycolytic Overload and Unscheduled Glycolysis-Driver of Insulin Resistance and Development of Vascular Complications of Diabetes. Int. J. Mol. Sci. 2022, 23, 2165. [Google Scholar] [CrossRef] [PubMed]
- Bautista-Perez, R.; Cano-Martinez, A.; Gutierrez-Velazquez, E.; Martinez-Rosas, M.; Perez-Gutierrez, R.M.; Jimenez-Gomez, F.; Flores-Estrada, J. Spinach Methanolic Extract Attenuates the Retinal Degeneration in Diabetic Rats. Antioxidants (Basel) 2021, 10, 717. [Google Scholar] [CrossRef]
- Alouffi, S.; Khan, M.W.A. Dicarbonyls Generation, Toxicities, Detoxifications and Potential Roles in Diabetes Complications. Curr. Protein Pept. Sci. 2020, 21, 890–898. [Google Scholar] [CrossRef]
- Muronetz, V.I. , Melnikova, A.K.; Seferbekova, Z.N.; Barinova, K.V.; Schmalhausen, E.V. Glycation, Glycolysis, and neurodegenerative diseases: Is There Any Connection? Biochemistry (Mosc.) 2017, 82, 874–886. [Google Scholar] [CrossRef]
- Oshitari, T. Advanced Glycation End-Products and Diabetic Neuropathy of the Retina. Int. J. Mol. Sci. 2023, 24, 2927. [Google Scholar] [CrossRef]
- Hammes, H.P.; Alt, A.; Niwa, T.; Clausen, J.T.; Bretzel, R.G.; Brownlee, M.; Schleicher, E.D. Differential accumulation of advanced glycation end products in the course of diabetic retinopathy. Diabetologia 1999, 42, 728–736. [Google Scholar] [CrossRef]
- Matafome, P.; Rodrigues, T.; Sena, C.; Seiça, R. Methylglyoxal in Metabolic Disorders: Facts, Myths, and Promises. Med. Res. Rev. 2017, 37, 368–403. [Google Scholar] [CrossRef]
- Rabbani, N.; Thornalley, P.J. Advanced glycation end products in the pathogenesis of chronic kidney disease. Kidney Int. 2018, 93, 803–813. [Google Scholar] [CrossRef]
- Alqahtani, A.S.; Li, K.M.; Razmovski-Naumovski, V.; Kam, A.; Alam, P.; Li, G.Q. Attenuation of methylglyoxal-induced glycation and cellular dysfunction in wound healing by Centella cordifolia. Saudi. J. Biol. Sci. 2021, 28, 813–824. [Google Scholar] [CrossRef] [PubMed]
- Pinto, R.S.; Machado, U.F.; Passarelli, M. Advanced glycation end products as biomarkers for cardiovascular disease: browning clarifying atherogenesis. Biomark. Med. 2020, 14, 611–614. [Google Scholar] [CrossRef] [PubMed]
- Delligatti, C.E.; Kirk, J.A. Glycation in the cardiomyocyte. Vitam. Horm. 2024, 125, 47–88. [Google Scholar] [CrossRef] [PubMed]
- Muraoka, M.Y.; Justino, A.B.; Caixeta, D.C.; Queiroz, J.S.; Sabino-Silva, R.; Salmen Espindola, F. Fructose and methylglyoxal-induced glycation alters structural and functional properties of salivary proteins, albumin and lysozyme. PLoS One 2022, 17, e0262369. [Google Scholar] [CrossRef]
- Bejarano, E.; Taylor, A. Too sweet: Problems of protein glycation in the eye. Exp. Eye Res. 2019, 178, 255–262. [Google Scholar] [CrossRef]
- Kang, Q.; Yang, C. Oxidative stress and diabetic retinopathy: Molecular mechanisms, pathogenetic role and therapeutic implications. Redox Biol. 2020, 37, 101799. [Google Scholar] [CrossRef]
- Bhat, S.P. Crystallins, genes and cataract. Progress in Drug Res. 2003, 60. [Google Scholar] [CrossRef]
- Abdullah, E.M.; Haq, S.H.; Ahmed, M.A.; Khan, J.M.; Alamery, S.F.; Malik, A. Structural stability and solubility of glycated camel lens ζ-crystallin. Int. J. Biol. Macromol. 2020, 158, 384–393. [Google Scholar] [CrossRef]
- Jeong, W.J.; Rho, J.H.; Yoon, Y.G.; Yoo, S.H.; Jeong, N.Y.; Ryu, W.Y.; Ahn, H.B.; et al. Cytoplasmic and nuclear anti-apoptotic roles of αB-crystallin in retinal pigment epithelial cells. PLoS One 2012, 7, e45754. [Google Scholar] [CrossRef]
- Kim, J.; Kim, K.S.; Shinn, J.W.; Oh, Y.S.; Kim, H.T.; Jo, I.; Shinn, S.H. The effect of antioxidants on glycated albumin-induced cytotoxicity in bovine retinal pericytes. Biochem. Biophys. Res. Commun. 2002, 292, 1010–1016. [Google Scholar] [CrossRef]
- Dahrouj, M.; Desjardins, D.M.; Liu, Y.; Crosson, C.E.; Ablonczy, Z. Receptor mediated disruption of retinal pigment epithelium function in acute glycated-albumin exposure. Exp. Eye Res. 2015, 137, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.S.; Kim, J.; Jo, K.; Lee, Y.M.; Sohn, E.; Yoo, N.H.; Kim, J.S. OSSC1E-K19, a novel phytochemical component of Osteomeles schwerinae, prevents glycated albumin-induced retinal vascular injury in rats. Mol. Med. Rep. 2015, 12, 7279–7284. [Google Scholar] [CrossRef] [PubMed]
- Nakagami, Y.; Masuda, K.; Hatano, E.; Inoue, T.; Matsuyama, T.; Iizuka, M.; Ono, Y.; et al. Novel Nrf2 activators from microbial transformation products inhibit blood-retinal barrier permeability in rabbits. Br. J. Pharmacol. 2015, 172, 1237–1249. [Google Scholar] [CrossRef] [PubMed]
- Dong, N.; Xu, B.; Shi, H. Long noncoding RNA MALAT1 acts as a competing endogenous RNA to regulate Amadori-glycated albumin-induced MCP-1 expression in retinal microglia by a microRNA-124-dependent mechanism. Inflamm. Res. 2018, 67, 913–925. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.X.; Wang, J.J.; Dashti, A.; Wilson, K.; Zou, M.H.; Szweda, L.; Ma, J.X.; Lyons, T.J. Pigment epithelium-derived factor mitigates inflammation and oxidative stress in retinal pericytes exposed to oxidized low-density lipoprotein. J. Mol. Endocrinol. 2008, 41, 135–143. [Google Scholar] [CrossRef]
- Duran-Jimenez, B.; Dobler, D.; Moffatt, S.; Rabbani, N.; Streuli, C.H.; Thornalley, P.J.; Tomlinson, D.R.; Gardiner, N.J. Advanced glycation end products in extracellular matrix proteins contribute to the failure of sensory nerve regeneration in diabetes. Diabetes 2009, 58, 2893–2903. [Google Scholar] [CrossRef]
- Portillo, J.C.; Pfaff, A.; Vos, S.; Weng, M.; Nagaraj, R.H.; Subauste, C.S. Advanced Glycation End Products Upregulate CD40 in Human Retinal Endothelial and Müller Cells: Relevance to Diabetic Retinopathy. Cells 2024, 13, 429. [Google Scholar] [CrossRef]
- Ighodaro, O.M. Molecular pathways associated with oxidative stress in diabetes mellitus. Biomed. Pharmacother. 2018, 108, 656–662. [Google Scholar] [CrossRef]
- Augustine, J.; Troendle, E.P.; Barabas, P.; McAleese, C.A.; Friedel, T.; Stitt, A.W.; Curtis, T.M. The Role of Lipoxidation in the Pathogenesis of Diabetic Retinopathy. Front. Endocrinol. (Lausanne) 2021, 11, 621938. [Google Scholar] [CrossRef]
- Yatoh, S.; Mizutani, M.; Yokoo, T.; Kozawa, T.; Sone, H.; Toyoshima, H.; Suzuki, S.; et al. Antioxidants and an inhibitor of advanced glycation ameliorate death of retinal microvascular cells in diabetic retinopathy. Diabetes Metab. Res. Rev. 2006, 22, 38–45. [Google Scholar] [CrossRef]
- Sheikpranbabu, S.; Haribalaganesh, R.; Gurunathan, S. Pigment epithelium-derived factor inhibits advanced glycation end-products-induced cytotoxicity in retinal pericytes. Diabetes Metab. 2011, 37, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.; Zhang, Y.; Huang, Y.; Deng, H. Pathophysiology of RAGE in inflammatory diseases. Front Immunol. 2022, 13, 931473. [Google Scholar] [CrossRef] [PubMed]
- Barile, G.R.; Pachydaki, S.I.; Tari, S.R.; Lee, S.E.; Donmoyer, C.M.; Ma, W.; Rong, L.L.; et al. The RAGE axis in early diabetic retinopathy. Invest. Ophthalmol. Vis. Sci. 2005, 46, 2916–2924. [Google Scholar] [CrossRef] [PubMed]
- Zong, H.; Ward, M.; Madden, A.; Yong, P.H.; Limb, G.A.; Curtis, T.M.; Stitt, A.W. Hyperglycaemia-induced pro-inflammatory responses by retinal Müller glia are regulated by the receptor for advanced glycation end-products (RAGE). Diabetologia 2010, 53, 2656–2666. [Google Scholar] [CrossRef] [PubMed]
- Taguchi, K.; Fukami, K. RAGE signaling regulates the progression of diabetic complications. Front. Pharmacol. 2023, 14, 1128872. [Google Scholar] [CrossRef]
- Wang, X.L.; Yu, T.; Yan, Q.C.; Wang, W.; Meng, N. : Li, X.J.; Luo, Y.H. AGEs Promote Oxidative Stress and Induce Apoptosis in Retinal Pigmented Epithelium Cells RAGE-dependently. J. Mol. Neurosci. [CrossRef]
- Bhattacharya, R.; Alam, M.R.; Kamal, M.A.; Seo, K.J.; Singh, L.R. AGE-RAGE axis culminates into multiple pathogenic processes: a central road to neurodegeneration. Front. Mol. Neurosci. 2023, 16, 1155175. [Google Scholar] [CrossRef]
- McVicar, C.M.; Ward, M.; Colhoun, L.M.; Guduric-Fuchs, J.; Bierhaus, A.; Fleming, T.; Schlotterer, A.; et al. Role of the receptor for advanced glycation endproducts (RAGE) in retinal vasodegenerative pathology during diabetes in mice. Diabetologia 2015, 58, 1129–1137. [Google Scholar] [CrossRef]
- Yao, D.; Brownlee, M. Hyperglycemia-induced reactive oxygen species increase expression of the receptor for advanced glycation end products (RAGE) and RAGE ligands. Diabetes 2010, 59, 249–255. [Google Scholar] [CrossRef]
- Hu, Z.; Fang, W.; Liu, Y.; Liang, H.; Chen, W.; Wang, H. Acute glucose fluctuation promotes RAGE expression via reactive oxygen species-mediated NF-κB activation in rat podocytes. Mol. Med. Rep. 2021, 23, 330. [Google Scholar] [CrossRef]
- Ramya, R.; Coral, K.; Bharathidevi, S.R. RAGE silencing deters CML-AGE induced inflammation and TLR4 expression in endothelial cells. Exp. Eye Res. 2021, 206, 108519. [Google Scholar] [CrossRef]
- Wang, Y.; Zhong, J.; Zhang, X.; Liu, Z.; Yang, Y.; Gong, Q.; Ren, B. The Role of HMGB1 in the Pathogenesis of Type 2 Diabetes. J. Diabetes Res. 2016, 2016, 2543268. [Google Scholar] [CrossRef] [PubMed]
- Behl, T.; Sharma, E.; Sehgal, A.; Kaur, I.; Kumar, A.; Arora, R.; Pal, G.; et al. Expatiating the molecular approaches of HMGB1 in diabetes mellitus: Highlighting signalling pathways via RAGE and TLRs. Mol. Biol. Rep. 2021, 48, 1869–1881. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.J.; Hsiao, C.C.; Yang, I.H.; Chou, M.H.; Wu, C.L.; Wei, Y.C.; Chen, C.H.; Chuang, J.H. High-mobility group box 1 protein is implicated in advanced glycation end products-induced vascular endothelial growth factor A production in the rat retinal ganglion cell line RGC-5. Mol. Vis. 2012, 18, 838–850. [Google Scholar] [PubMed]
- Maeda, S.; Matsui, T.; Ojima, A.; Takeuchi, M.; Yamagishi, S. Sulforaphane inhibits advanced glycation end product-induced pericyte damage by reducing expression of receptor for advanced glycation end products. Nutr. Res. 2014, 34, 807–813. [Google Scholar] [CrossRef]
- Li, G.; Tang, J.; Du, Y.; Lee, C.A.; Kern, T.S. Beneficial effects of a novel RAGE inhibitor on early diabetic retinopathy and tactile allodynia. Mol. Vis. 2011, 17, 3156–3165. [Google Scholar]
- Wu, Q.; Liu, H.; Zhou, M. Fangchinoline Ameliorates Diabetic Retinopathy by Inhibiting Receptor for Advanced Glycation End-Products (RAGE)-Nuclear Factor Kappa-Light-Chain-Enhancer of Activated B Cells (NF-κB) Pathway in Streptozotocin (STZ)-Induced Diabetic Rats. Med. Sci. Monit. 2019, 25, 1113–1121. [Google Scholar] [CrossRef]
- Simon, M.V. , Vera, M.S.; Tenconi, P.E.; Soto, T.; Prado Spalm, F.H.; Torlaschi, C.; Mateos, M.V., Rotstein, N.P. Sphingosine-1-phosphate and ceramide-1-phosphate promote migration, pro-inflammatory and pro-fibrotic responses in retinal pigment epithelium cells. Exp. Eye Res. 2022, 224, 109222. [Google Scholar] [CrossRef]
- Nita, M.; Grzybowski, A. Interplay between reactive oxygen species and autophagy in the course of age-related macular degeneration. EXCLI J. 2020, 19, 1353–1371. [Google Scholar] [CrossRef]
- Zhang, S.M.; Fan, B.; Li, Y.L.; Zuo, Z.Y.; Li, G.Y. Oxidative Stress-Involved Mitophagy of Retinal Pigment Epithelium and Retinal Degenerative Diseases. Cell Mol. Neurobiol. 2023, 43, 3265–3276. [Google Scholar] [CrossRef]
- Datta, S.; Cano, M.; Ebrahimi, K.; Wang, L.; Handa, J.T. The impact of oxidative stress and inflammation on RPE degeneration in non-neovascular AMD. Prog. Retin. Eye Res. 2017, 60, 201–218. [Google Scholar] [CrossRef]
- Hashikawa, K.I.; Tsuruta, A.; Yamakawa, W.; Yasukochi, S.; Koyanagi, S.; Ohdo, S. Senescence-induced alteration of circadian phagocytic activity of retinal pigment epithelium cell line ARPE-19. Biochem. Biophys. Res. Commun. 2023, 658, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Cai, B.; Li, Y.; Su, W.; Zhao, X.; Gong, B.; Li, Z.; Zhang, X.; Wu, Y.; et al. HMGB1 and Caveolin-1 related to RPE cell senescence in age-related macular degeneration. Aging (Albany NY) 2019, 11(13), 4323- 4337. [CrossRef]
- Qi, H.; Liu, T.; Liu, J.; Teng, Q.; Ma, Z.; Wang, S.; Wen, S.; et al. Thioredoxin1 is a target to attenuate diabetes-induced RPE cell dysfunction in human ARPE19 cells by alleviating oxidative stress. Mol. Med. Rep. 2023, 28, 134. [Google Scholar] [CrossRef] [PubMed]
- Clarkson-Townsend, D.A.; Douglass, A.J.; Singh, A.; Allen, R.S.; Uwaifo, I.N.; Pardue, M.T. Impacts of high fat diet on ocular outcomes in rodent models of visual disease. Exp. Eye Res. 2021, 204, 108440. [Google Scholar] [CrossRef] [PubMed]
- Sekar, P.; Hsiao, G.; Hsu, S.H.; Huang, D.Y.; Lin, W.W.; Chan, C.M. Metformin inhibits methylglyoxal-induced retinal pigment epithelial cell death and retinopathy via AMPK-dependent mechanisms: Reversing mitochondrial dysfunction and upregulating glyoxalase 1. Redox Biol. 2023, 64, 102786. [Google Scholar] [CrossRef]
- Chan, C.M.; Huang, D.Y.; Huang, Y.P.; Hsu, S.H.; Kang, L.Y.; Shen, C.M.; Lin, W.W. Methylglyoxal induces cell death through endoplasmic reticulum stress-associated ROS production and mitochondrial dysfunction. J. Cell. Mol. Med. 2016, 20, 1749–1760. [Google Scholar] [CrossRef]
- Antonetti, D.A.; Silva, P.S.; Stitt, A.W. Current understanding of the molecular and cellular pathology of diabetic retinopathy. Nat. Rev. Endocrinol. 2021, 17, 195–206. [Google Scholar] [CrossRef]
- Intartaglia, D.; Giamundo, G.; Conte, I. Autophagy in the retinal pigment epithelium: a new vision and future challenges. FEBS J. 2022, 289, 7199–7212. [Google Scholar] [CrossRef]
- Si, Z.; Zheng, Y.; Zhao, J. The Role of Retinal Pigment Epithelial Cells in Age-Related Macular Degeneration: Phagocytosis and Autophagy. Biomolecules 2023, 13, 901. [Google Scholar] [CrossRef]
- Wu, A.Y.; Sekar, P.; Huang, D.Y.; Hsu, S.H.; Chan, C.M.; Lin, W.W. Spatiotemporal roles of AMPK in PARP-1- and autophagy-dependent retinal pigment epithelial cell death caused by UVA. J. Biomed. Sci. 2023, 30, 91. [Google Scholar] [CrossRef]
- Chan, C.M.; Huang, D.Y.; Sekar, P.; Hsu, S.H.; Lin, W.W. Reactive oxygen species-dependent mitochondrial dynamics and autophagy confer protective effects in retinal pigment epithelial cells against sodium iodate-induced cell death. J. Biomed. Sci. 2019, 26, 40. [Google Scholar] [CrossRef]
- Lakkaraju, A.; Umapathy, A.; Tan, L.X.; Daniele, L.; Philp, N.J.; Boesze-Battaglia, K.; Williams, D.S. The cell biology of the retinal pigment epithelium. Prog. Retin. Eye Res. 2020, 100846. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.Y.; Peng, J.; He, F.; Tursun, X.; Li, S.P.; Xin, X.L.; Aisa, H.A. Shabyar Ameliorates High Glucose Induced Retinal Pigment Epithelium Injury Through Suppressing Aldose Reductase and AMPK/mTOR/ULK1 Autophagy Pathway. Front. Pharmacol. 2022, 13, 852945. [Google Scholar] [CrossRef] [PubMed]
- Zou, J.; Tan, W.; Liu, K.; Chen, B.; Duan, T.; Xu, H. Wnt inhibitory factor 1 ameliorated diabetic retinopathy through the AMPK/mTOR pathway-mediated mitochondrial function. FASEB J. 2022, 36, e22531. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Liang, L.; Zhang, S.; Yang, J.; Yue, Y.; Zhang, X. HMGB1 downregulation in retinal pigment epithelial cells protects against diabetic retinopathy through the autophagy-lysosome pathway. Autophagy 2022, 18, 320–339. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, J.; Zhang, C.; Zhang, J.; Gu, L.; Luo, D.; Qiu, Q. Diabetic Macular Edema: Current Understanding, Molecular Mechanisms and Therapeutic Implications. Cells 2022, 11, 3362. [Google Scholar] [CrossRef]
- Zhang, C.; Gu, L.; Xie, H.; Liu, Y.; Huang, P.; Zhang, J.; Luo, D.; Zhang, J. Glucose transport, transporters and metabolism in diabetic retinopathy. Biochim. Biophys. Acta Mol. Basis Dis. 2024, 1870, 166995. [Google Scholar] [CrossRef]
- Singh, L.P. Thioredoxin Interacting Protein (TXNIP) and Pathogenesis of Diabetic Retinopathy. J Clin. Exp. Ophthalmol. 2013, 4. [Google Scholar] [CrossRef]
- Bang, E.; Park, C.; Hwangbo, H.; Shim, J.H.; Leem, S.H.; Hyun, J.W.; Kim, G.Y.; Choi, Y.H. Spermidine Attenuates High Glucose-Induced Oxidative Damage in Retinal Pigment Epithelial Cells by Inhibiting Production of ROS and NF-κB/NLRP3 Inflammasome Pathway. Int. J. Mol. Sci. 2023, 24, 10550. [Google Scholar] [CrossRef]
- Li, J.Q.; Shi, Y.H. ; Min-Xu; Shi, C.X., Teng-Wang, Wang, T.H., Zuo, Z.F., Eds.; Liu, X.Z. Discovery of astragaloside IV against high glucose-induced apoptosis in retinal ganglion cells: Bioinformatics and in vitro studies. Gene 2024, 905, 148219. [Google Scholar] [CrossRef]
- Gao, S.; Gao, S.; Wang, Y.; Li, N.; Yang, Z.; Yao, H.; Chen, Y.; et al. Inhibition of Ferroptosis Ameliorates Photoreceptor Degeneration in Experimental Diabetic Mice. Int. J. Mol. Sci. 2023, 24, 16946. [Google Scholar] [CrossRef]
- Liu, K.; Gao, X.; Hu, C.; Gui, Y.; Gui, S.; Ni, Q.; Tao, L.; Jiang, Z. Capsaicin ameliorates diabetic retinopathy by inhibiting poldip2-induced oxidative stress. Redox Biol. 2022, 56, 102460. [Google Scholar] [CrossRef]
- Chang, Y.C.; Hsieh, M.C.; Wu, H.J.; Wu, W.C.; Kao, Y.H. Methylglyoxal, a reactive glucose metabolite, enhances autophagy flux and suppresses proliferation of human retinal pigment epithelial ARPE-19 cells. Toxicol. in Vitro 2015, 29, 1358–1368. [Google Scholar] [CrossRef] [PubMed]
- Kim, O.S.; Kim, J.; Kim, C.S.; Kim, N.H.; Kim, J.S. KIOM-79 prevents methyglyoxal-induced retinal pericyte apoptosis in vitro and in vivo. J. Ethnopharmacol. 2010, 129, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, J.; Zheng, Y.; Jiang, J.; Wang, L.; Wu, J.; Zhang, C.; Luo, M. Glucose metabolite methylglyoxal induces vascular endothelial cell pyroptosis via NLRP3 inflammasome activation and oxidative stress in vitro and in vivo. Cell Mol. Life Sci. 2024, 81, 401. [Google Scholar] [CrossRef] [PubMed]
- Medeiros, M.L.; Oliveira, A.L.; Mello, G.C.; Antunes, E. Metformin Counteracts the Deleterious Effects of Methylglyoxal on Ovalbumin-Induced Airway Eosinophilic Inflammation and Remodeling. Int. J. Mol. Sci. 2023, 24, 9549. [Google Scholar] [CrossRef]
- Spagnuolo, L.; Della Posta, S.; Fanali, C.; Dugo, L.; De Gara, L. Chemical Composition of Hazelnut Skin Food Waste and Protective Role against Advanced Glycation End-Products (AGEs) Damage in THP-1-Derived Macrophages. Molecules 2023, 28, 2680. [Google Scholar] [CrossRef]
- Kang, J.; Jeong, Y.J.; Ha, S.K.; Lee, H.H.; Lee, K.W. Glyoxal-derived advanced glycation end-products, Nε-carboxymethyl-lysine, and glyoxal-derived lysine dimer induce apoptosis-related gene expression in hepatocytes. Mol. Biol. Rep. 2023, 50, 2511–2520. [Google Scholar] [CrossRef]
- Ma, J.H.; Wang, J.J.; Zhang, S.X. The unfolded protein response and diabetic retinopathy. J. Diabetes Res. 2014, 2014, 160140. [Google Scholar] [CrossRef]
- Chandrakumar, S.; Santiago Tierno, I.; Agarwal, M.; Matisioudis, N.; Kern, T.S.; Ghosh, K. Subendothelial Matrix Stiffening by Lysyl Oxidase Enhances RAGE-Mediated Retinal Endothelial Activation in Diabetes. Diabetes 2023, 72, 973–985. [Google Scholar] [CrossRef]
- Tu, Z.; Li, Y.; Smith, D.S.; Sheibani, N.; Huang, S.; Kern, T.; Lin, F. Retinal pericytes inhibit activated T cell proliferation. Invest Ophthalmol. Vis. Sci. 2011, 52, 9005–9010. [Google Scholar] [CrossRef]
- Pickel, L.; Kim, S.J.; Hacibekiroglu, S.; Nagy, A.; Lee, J.; Sung, H.K. The Circadian Clock of Müller Glia is Necessary for Retinal Homeostasis and Neuronal Survival. Am. J. Pathol. 0002. [Google Scholar] [CrossRef]
- Rattner, A.; Williams, J.; Nathans, J. Roles of HIFs and VEGF in angiogenesis in the retina and brain. J. Clin. Invest. 2019, 129, 3807–3820. [Google Scholar] [CrossRef]
- Eastlake, K.; Luis, J.; Limb, G.A. Potential of Müller Glia for Retina Neuroprotection. Curr. Eye Res. 2020, 45, 339–348. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Xu, G.T.; Zhang, J.F. Inflammation in diabetic retinopathy: possible roles in pathogenesis and potential implications for therapy. Neural. Regen. Res. 2023, 18, 976–982. [Google Scholar] [CrossRef] [PubMed]
- Sorrentino, F.S.; Allkabes, M.; Salsini, G.; Bonifazzi, C.; Perri, P. The importance of glial cells in the homeostasis of the retinal microenvironment and their pivotal role in the course of diabetic retinopathy. Life Sci. 2016, 162, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Reber, F.; Kasper, M.; Siegner, A.; Kniep, E.; Seigel, G.; Funk, R.H. Alteration of the intracellular pH and apoptosis induction in a retinal cell line by the AGE-inducing agent glyoxal. Graefes. Arch. Clin. Exp. Ophthalmol. 2002, 240, 1022–1032. [Google Scholar] [CrossRef]
- Portillo, J.C.; Yu, J.S.; Vos, S.; Bapputty, R.; Lopez Corcino, Y.; Hubal, A.; Daw, J.; et al. Disruption of retinal inflammation and the development of diabetic retinopathy in mice by a CD40-derived peptide or mutation of CD40 in Müller cells. Diabetologia 2022, 65, 2157–2171. [Google Scholar] [CrossRef]
- Portillo, J.C.; Lopez Corcino, Y.; Miao, Y.; Tang, J.; Sheibani, N.; Kern, T.S.; Dubyak, G.R.; Subauste, C.S. CD40 in Retinal Müller Cells Induces P2X7-Dependent Cytokine Expression in Macrophages/Microglia in Diabetic Mice and Development of Early Experimental Diabetic Retinopathy. Diabetes 2017, 66, 483–493. [Google Scholar] [CrossRef]
- Portillo, J.C.; Corcino, Y.L.; Dubyak, G.R.; Kern, T.S.; Matsuyama, S.; Subauste, C.S. Ligation of CD40 in Human Müller Cells Induces P2X7 Receptor–Dependent Death of Retinal Endothelial Cells. Invest. Ophthalmol. Vis. Sci. 2016, 57, 6278–6286. [Google Scholar] [CrossRef]
- Samuels, I.S.; Portillo, J.C.; Miao, Y.M.; Kern, T.S.; Subaste, C.S. Loss of CD40 attenuates experimental diabetes-induced retinal inflammation but does not protect mice from electroretinogram defects. Vis. Neurosci. 2017, 34, E009. [Google Scholar] [CrossRef]
- Bai, Y.; Ma, J.X.; Guo, J.; Wang, J.; Zhu, M.; Chen, Y.; Le, Y.Z. Müller cell-derived VEGF is a significant contributor to retinal neovascularization. J. Pathol. 2009, 219, 446–454. [Google Scholar] [CrossRef]
- Li, Y.; Zhao, Y.; Sang, S.; Leung, T. Methylglyoxal-Induced Retinal Angiogenesis in Zebrafish Embryo: A Potential Animal Model of Neovascular Retinopathy. J. Ophthalmol. 2019, 2019, 2746735. [Google Scholar] [CrossRef]
- Rozpedek, W.; Pytel, D.; Mucha, B.; Leszczynska, H.; Diehl, J.A.; Majsterek, I. The Role of the PERK/eIF2α/ATF4/CHOP Signaling Pathway in Tumor Progression During Endoplasmic Reticulum Stress. Curr. Mol. Med. 2016, 16, 533–544. [Google Scholar] [CrossRef] [PubMed]
- Sano, R.; Reed, J.C. ER stress-induced cell death mechanisms. Biochim. Biophys. Acta 2013, 1833, 3460–3470. [Google Scholar] [CrossRef] [PubMed]
- Herzig, S.; Shaw, R.J. AMPK: guardian of metabolism and mitochondrial homeostasis. Nat. Rev. Mol. Cell. Biol. 2018, 19, 121–135. [Google Scholar] [CrossRef] [PubMed]
- Entezari, M.; Hashemi, D.; Taheriazam, A.; Zabolian, A.; Mohammadi, S.; Fakhri, F.; Hashemi, M.; Hushmandi, K.; et al. AMPK signaling in diabetes mellitus, insulin resistance and diabetic complications: A pre-clinical and clinical investigation. Biomed. Pharmacother. 2022, 146, 112563. [Google Scholar] [CrossRef]
- Karbasforooshan, H.; Karimi, G. The role of SIRT1 in diabetic retinopathy. Biomed. Pharmacother. 2018, 97, 190–194. [Google Scholar] [CrossRef]
- Zhang, S.; Wei, X.; Bowers, M.; Jessberger, S.; Golczak, M.; Semenkovich, C.F.; Rajagopal, R. Increasing Energetic Demands on Photoreceptors in Diabetes Corrects Retinal Lipid Dysmetabolism and Reduces Subsequent Microvascular Damage. Am. J. Pathol. 2023, 193, 2144–2155. [Google Scholar] [CrossRef]
- Zou, J.; Tan, W.; Liu, K.; Chen, B.; Duan, T.; Xu, H. Wnt inhibitory factor 1 ameliorated diabetic retinopathy through the AMPK/mTOR pathway-mediated mitochondrial function. FASEB J. 2022, 36, e22531. [Google Scholar] [CrossRef]
- Song, S.; Bao, S.; Zhang, C.; Zhang, J.; Li, J.; Li, X.; Chudhary, M.; Ren, X.; Kong, L. Stimulation of AMPK Prevents Diabetes-Induced Photoreceptor Cell Degeneration. Oxid. Med. Cell. Longev. 2021, 2021, 5587340. [Google Scholar] [CrossRef]
- Qian, S.; Qian, Y.; Huo, D.; Wang, S.; Qian, Q. Tanshinone IIa protects retinal endothelial cells against mitochondrial fission induced by methylglyoxal through glyoxalase 1. Europ. J. Pharmacol. 2019, 857, 172419. [Google Scholar] [CrossRef]
- Xue, M.; Rabbani, N.; Momiji, H.; Imbasi, P.; Anwar, M.M.; Kitteringham, N.; Park, B.K.; Souma, T.; et al. Transcriptional control of glyoxalase 1 by Nrf2 provides a stress-responsive defence against dicarbonyl glycation. Biochem. J. 2012, 443, 213–222. [Google Scholar] [CrossRef]
- Van Herreweghe, F.; Mao, J.; Chaplen, F.W.R.; Grooten, J.; Gevaert, K.; Vandekerckhove, J.; Vancompernolle, K. Tumor necrosis factor-induced modulation of glyoxalase I activities through phosphorylation by PKA results in cell death and is accompanied by the formation of a specific methylglyoxal-derived AGE. Proc. Natl. Acad. Sci. USA 2002, 99, 949–954. [Google Scholar] [CrossRef] [PubMed]
- Alhujaily, M. Molecular Assessment of Methylglyoxal-Induced Toxicity and Therapeutic Approaches in Various Diseases: Exploring the Interplay with the Glyoxalase System. Life (Basel) 2024, 14, 263. [Google Scholar] [CrossRef] [PubMed]
- Rabbani, N.; Xue, M.; Thornalley, P.J. Activity, regulation, copy number and function in the glyoxalase system. Biochem. Soc. Trans. 2014, 42, 419–424. [Google Scholar] [CrossRef] [PubMed]
- Maessen, D.E.; Stehouwer, C.D.; Schalkwijk, C.G. The role of methylglyoxal and the glyoxalase system in diabetes and other age-related diseases. Clin. Sci. (Lond) 2015, 128, 839–861. [Google Scholar] [CrossRef]
- Rabbani, N.; Thornalley, P.J. Glyoxalase in diabetes, obesity and related disorders. Semin. Cell. Dev. Biol. 2011, 22, 309–317. [Google Scholar] [CrossRef]
- Rasul, A.; Rashid, A.; Waheed, P.; Khan, S.A. Expression analysis of glyoxalase I gene among patients of diabetic retinopathy. Pak. J. Med. Sci. 2018, 34, 139–143. [Google Scholar] [CrossRef]
- Sachdeva, R.; Schlotterer, A.; Schumacher, D.; Matka, C.; Mathar, I.; Dietrich, N.; Medert, R.; et al. TRPC proteins contribute to development of diabetic retinopathy and regulate glyoxalase 1 activity and methylglyoxal accumulation. Mol. Metab. 2018, 9, 156–167. [Google Scholar] [CrossRef]
- Lodd, E.; Wiggenhauser, L.M.; Morgenstern, J.; Fleming, T.H.; Poschet, G.; Büttner, M.; Tabler, C.T.; et al. The combination of loss of glyoxalase1 and obesity results in hyperglycemia. JCI Insight 2019, 4, e126154. [Google Scholar] [CrossRef]
- Berner, A.K.; Brouwers, O.; Pringle, R.; Klaassen, I.; Colhoun, L.; McVicar, C.; Brockbank, S.; et al. Protection against methylglyoxal-derived AGEs by regulation of glyoxalase 1 prevents retinal neuroglial and vasodegenerative pathology. Diabetologia 2012, 55, 845–854. [Google Scholar] [CrossRef]
- Moraru, A.; Wiederstein, J.; Pfaff, D.; Fleming, T.; Miller, A.K.; Nawroth, P.; Teleman, A.A. Elevated Levels of the Reactive Metabolite Methylglyoxal Recapitulate Progression of Type 2 Diabetes. Cell. Metab, 27. [CrossRef]
- Irshad, Z.; Xue, M.; Ashour, A.; Larkin, J.R.; Thornalley, P.J.; Rabbani, N. Activation of the unfolded protein response in high glucose treated endothelial cells is mediated by methylglyoxal. Sci. Rep. 2019, 2019 9, 7889. [Google Scholar] [CrossRef]
- Xue, M.; Weickert, M.O.; Qureshi, S.; Kandala, N.B.; Anwar, A.; Waldron, M.; Shafie, A.; et al. Improved Glycemic Control and Vascular Function in Overweight and Obese Subjects by Glyoxalase 1 Inducer Formulation. Diabetes 2016, 2016 65, 2282–2294. [Google Scholar] [CrossRef]
Figure 1.
Schematic representation of methylglyoxal sources from carbohydrates, fatty acids, and proteins.
Figure 1.
Schematic representation of methylglyoxal sources from carbohydrates, fatty acids, and proteins.
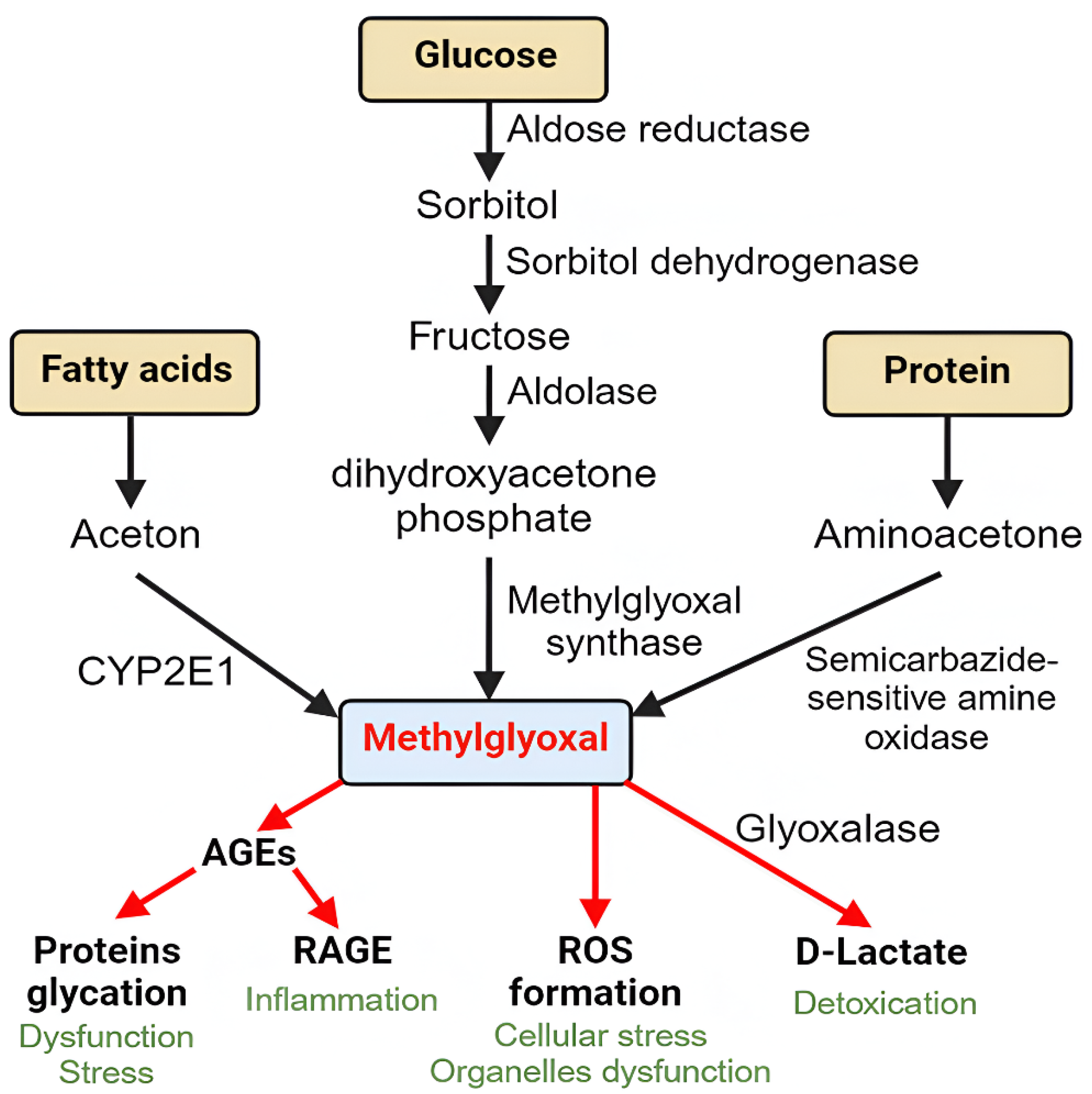
Figure 2.
Roles of CD40 activation and Müller cells-microglial communication in MGO-induced chronic inflammation and promotion of new blood vessel formation in the retina.
Figure 2.
Roles of CD40 activation and Müller cells-microglial communication in MGO-induced chronic inflammation and promotion of new blood vessel formation in the retina.
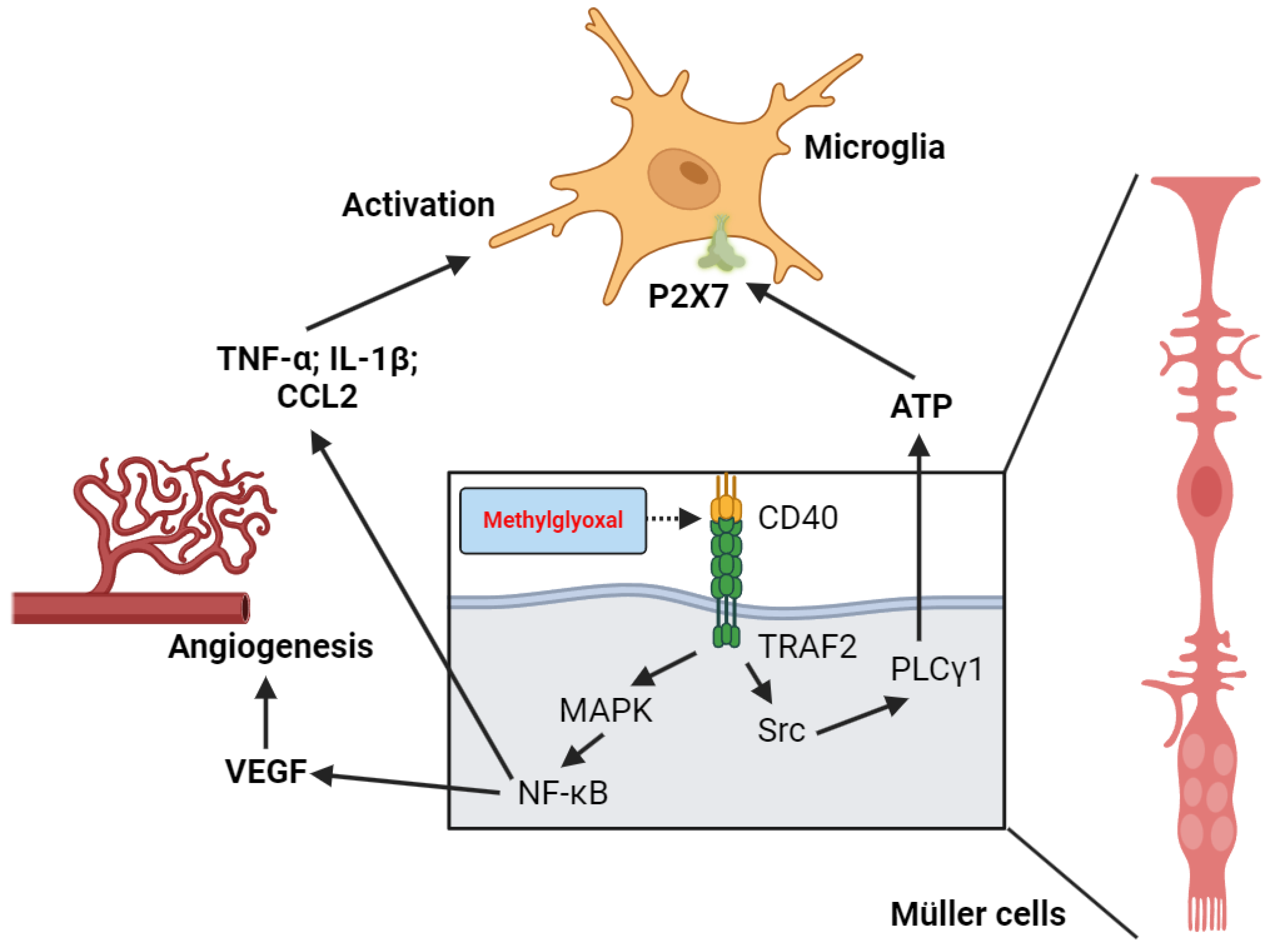
Figure 3.
Representation of MGO-induced pathways activation in RPE cells.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



