
Submitted:
11 October 2024
Posted:
11 October 2024
You are already at the latest version
Abstract
Lidocaine, a commonly used local anesthetic, has demonstrated modulatory effects in various cells. This study investigates the impact of lidocaine on cytokine production in peripheral blood mononuclear cells (PBMCs) from healthy donors and tumor-infiltrating immune cells (TIICs) from gastric cancer patients. Flow cytometry and cytokine assays revealed that lidocaine inhibited IFN-γ and IL-12 production in CD8+ and CD14+ PBMCs, respectively, while enhancing anti-inflammatory cytokines IL-10, TGF-β, and IL-35 in CD4+CD25+ and CD14+ PBMCs. In contrast, lidocaine promoted the secretion of pro-inflammatory IFN-γ and IL-12 in CD8+ and CD14+ TIICs, while reducing IL-10, TGF-β, and IL-35 levels, thus shifting CD14+ macrophages toward an M1-like phenotype. Additionally, lidocaine decreased PD-1 expression and increased IFN-γ production in CD8+ TIICs via NF-κB activation, enhancing their cytotoxicity against primary gastric cancer cells (PGCCs). Importantly, lidocaine did not affect the viability of normal PBMCs, TIICs, or PGCCs at concentrations up to 1.5 mM. These findings suggest that lidocaine reprograms the tumor immune microenvironment, supporting its potential as an adjunct therapy in gastric cancer by enhancing anti-tumor immunity.
Keywords:
lidocaine
; gastric cancer
; immune regulation
; tumor-infiltrating immune cells (TIICs)
; cytokine secretion
; peripheral blood mononuclear cells (PBMCs)
; primary gastric cancer cells (PGCCs)
; regulatory T(Treg)
; tumor microenvironment (TME)
; cytotoxic T cells (CTLs)
1. Introduction
Lidocaine, a commonly used local anesthetic, also has anti-inflammatory activity in various diseases. Lidocaine also regulates the immune system [1]. Although lidocaine inhibits natural killer (NK) cell function at high concentrations, it stimulates the killing activity of these cells at therapeutic plasma concentrations [2]. Ramirez and colleagues demonstrated that lidocaine enhances NK cell killing activity against three different leukemia cell lines [3]. In the past, there were many reports on the regulation of immune cells by lidocaine, the production of inflammatory cytokines, and the regulation of nuclear factor-kappaB (NF-κB) [4,5,6]. It was found that lidocaine downregulates NF-κB signaling and inhibits cytokine production and T cell proliferation [6,7]. Uncontrolled immune responses are associated with almost all kinds of cancer. Immune cells play a crucial role in anti-cancer responses and cytokine production. Therefore, inhibition of the activation of regulatory T (Treg) cells and their related cytokine production is considered an important strategy to deal with cancer [8]. Treg cells are a subset of CD4+CD25+ T cells that potently suppress many immune responses [9].
IFN-γ, a cytokine essential for both innate and adaptive immune responses, is produced principally by CD4+ and CD8+ T cells. It is critical for successful clearance of intracellular pathogens and also in host defense against malignant transformation [10]. IFN-γ production should therefore be subject to intense positive and negative regulation in cells of the immune system [11]. The specific immunity is controlled by specific cytokines [12]. Most of the IL-12-induced effects are mediated by IFN-γ [13]. IL-12 induces immune responses against tumors through their direct effects on tumors via angiogenesis and lymphocytes [13]. CD14+ macrophages commonly exist in two distinct subsets: classically activated (M1) macrophages, which are pro-inflammatory and associated with T-helper (Th)1 cytokines such as IFN-γ and IL-12, and alternatively activated (M2) macrophages, characterized by markers such as TGF-β and IL-10 [14]. M2 macrophages secrete high amounts of IL-10 and TGF-β to suppress the inflammation [14]. IL-10 is an important anti-inflammatory cytokine produced under different conditions of immune activation by a variety of cell types, including T cells and macrophages [15]. It seems to be a double-edged sword in the host defense.
IL-35 is secreted by forkhead box P3 (Foxp3)+ cluster of differentiation CD4+CD25+ Tregs or a Foxp3-Treg population and has previously been proposed as a novel immune-suppressing cytokine and a key effector molecule of Tregs function [16]. It shares the IL-12 p35 and Epstein-Barr virus-induced gene 3 subunits with IL-12 and IL-27, respectively [17]. IL-35 suppresses the activity of Th1, Th2 and Th17 cells, and expands CD4+CD25+Foxp3+Tregs [16]. It is also required for maximal Treg activity and it alone is sufficient to suppress T-cell proliferation [18]. Moreover, IL-35 stimulation increased the inhibitory function of CD4+CD25+Foxp3+ Tregs and enhances IL-35/IL-10 productions [18]. However, the precise underlying mechanism behind the involvement of IL-35 in lidocaine effects has yet to be elucidated in normal human PBMCs and gastric TIICs.
Accumulating evidence indicates that CD4+CD25+Treg cells have a powerful ability to suppress the host immune response [8]. However, recent studies have shown that tumor cells can recruit these Treg cells to suppress antitumor immunity in the tumor microenvironment (TME), thereby limiting the efficiency of cancer immunotherapy [9]. Our strategy is to use lidocaine to overcome tumor-associated immunosuppression for successful gastric cancer immunotherapy. It was reported that PD-1 play a vital role in inhibiting immune responses [19,20]. Monoclonal antibodies target PD-1 can boost the immune response against cancer cells [19,20]. PD-1 blockades have also been associated with the development of cytotoxic CD8+ T cells and an imbalance of the immune system [20,21]. This is the first report to reveal that lidocaine blockade of PD-1 and increase IFN-γ through NF-κB signaling. Lidocaine-treated gastric CD8+ TIICs augmented the anti-tumor response, killing primary gastric cancer cells (PGCCs).
2. Materials and Methods
2.1. Isolation and Culture of PBMCs from Healthy Adult Volunteers
Blood samples were obtained from healthy adult volunteers (participants without known medical conditions) (n=23) at Taichung Veterans General Hospital, following institutional review board (IRB) approval and consent. The protocol was approved by the Ethical and Scientific Committee of Taichung Veterans General Hospital (IRB no. SF22141B#1). Peripheral blood was collected in EDTA tubes, and peripheral blood mononuclear cells (PBMCs) were isolated using Histopaque 1.077 g/ml (Sigma Chemicals, Darmstadt, Germany), as previously described [22]. The cell pellet (PBMCs) was resuspended in RPMI medium supplemented with 10% fetal bovine serum (FBS), and 100 U/ml penicillin and streptomycin.
2.2. Reagents and Antibodies
Reagents and sources were as follows: FITC-conjugated IgG1 anti-human CD14 (BD Bioscience, San Diego, USA); FITC-conjugated IgG1 anti-human CD8 (eBioscience); FITC-conjugated IgG1 anti-human CD25 (Elabscience); IgG1-FITC isotype control (Caltag Laboratories, Inc., Burlingame, USA); PE-conjugated IgG1 anti-human CD4 (Elabscience, San Diego, USA); PE-conjugated IgG1 anti-human IFN-γ (Elabscience, San Diego, USA); IgG1-PE isotype control (Caltag Laboratories, Inc., Burlingame, USA). NF-κB-specific inhibitor BAY11-7082 was purchased from Merck (San Diego, USA).
2.3. Isolation of TIICs from Gastric Cancer Patients
Ex vivo malignant gastric tissues were obtained from patients undergoing routine planned cancer-related surgery (n=21). Written informed consent was obtained from each patient in accordance with local institutional ethics review and approved by the Ethical and Scientific Committee of Taichung Veterans General Hospital (TCVGH-IRB no. SF22141B#1). The generation of TIIC cultures from tumors has been detailed elsewhere [23]. In brief, tumor specimens removed from cancer patients were placed on a plate with 5% FBS in Hank’s balanced salt solution buffer (Gibco, New York, USA) on ice and disintegrated using scissors. The homogenate was collected and treated with 1 mg/ml type IV collagenase (Sigma, St. Louis, USA) and 0.05 mg/ml DNase (Promega, Madison, USA) for 30 minutes at 37°C with gentle agitation. The digested extract was screened using a 100-mesh filter, and the filtrate was washed with 5% FBS in Hank’s balanced salt solution buffer and centrifuged at 600xg for 7 minutes at 4°C. The cell pellet obtained was treated with ACK erythrocyte lysis buffer (155 mM NH4Cl, 10 mM KHCO3, and 1 mM Na2EDTA, pH 7.3) for 5 min at room temperature. Finally, TIICs were resuspended in RPMI 1640 medium with 10% FBS. Cells were harvested by 7 to 14 days of culture. Each initial well was considered to be an independent TIICs culture and was maintained separately from the others.
2.4. Primary Gastric Cancer Cells (PGCCs) Culture from Fresh Surgical Malignant Gastric Tissues
Specimens were collected in Dulbecco’s modified Eagle’s medium (Biochrom co, Berlin, Germany) containing 1% Penicillin/Streptomycin for transport to our laboratory. Primary PGCCs were purified and maintained in Dulbecco’s modified Eagle’s medium supplemented with 10% FBS as previously described [24,25]. On the next day, the cell culture was rinsed with PBS twice to remove non-adherent cells. The medium was changed every 3-7 days, depending on the density of cell growth. The colonies increased in size and spread out, resulting in some cells separating at the periphery of the colonies after 2 weeks of culture. PGCCs were identified using granulin (GRN) markers [24]. These primary cells were maintained in culture for up to 4-8 weeks.
2.5. Sorting of CD8+T cells, CD4+CD25+ Treg, CD14+Macrophages, and GRN+ PGCCs
PBMCs were obtained from healthy donors, TIICs and PGCCs were obtained from gastric cancer patients after written informed consent and approval by the Ethical and Scientific Committee of Taichung Veterans General Hospital (IRB no. SF22141B#1). Cells were stained with the CD8 Ab for cytotoxic T cells (CTLs), CD4 Ab and CD25 Ab for Tregs, CD14 Ab for macrophage cells. PGCCs were identified using GRN markers [24]. Sample acquisition and cell sorting were managed on the BD FACSMelody™ cell sorter (BD Biosciences, San Jose, USA) and analyzed using BD Chorus software.
2.6. Human CD8+, CD4+CD25+, CD14+PBMCs and CD8+, CD4+CD25+, CD14+ TIICs Viability
Human immune cells were assessed by flow cytometric analysis using propidium iodide-stained cells. Initially, 104 cells were incubated in 96-well plates in the absence or presence of lidocaine at concentrations ranging from 0.25 mM to 1.5 mM. After 72h treatment, cells were washed with PBS and fixed with 70% ethanol for 1 h on ice. Pelleted cells were then incubated with RNase-A (0.1 mg/mL) and propidium iodide (40 μg/mL) for 1 h with shaking and protected from light. The percentage of the sub-G1 population was determined by flow cytometry.
2.7. CD8+ Primary T Cells and CD14+ Primary Macrophage Activation
CD8+PBMCs and CD14+PBMCs were triggered by phorbol myristate acetate (PHA) 2.5 mg/ml and phytohaemagglutinin (PMA) 50 ng/ml in the absence or presence of lidocaine concentrations ranging from 0.25 mM to 1.5 mM, or cultured in media alone as a negative control.
2.8. Analysis of Cytokines by ELISA
Human IFN-γ, IL-12, IL-10, TGF-β, and IL-35 protein levels were quantified using a sandwich ELISA. The production of IFN-γ, IL-12, IL-10 and TGF-β were analyzed by human IL-10 ELISA kit (Arigo Biolaboratories, Taipei, Taiwan), human IL-12, TGF-β ELISA kit (Elabscience, Houston, USA), and human IFN-γ ELISA KIT (Gen-Probe, San Diego, USA) respectively. Initially, 100 μL of supernatant was added to the ELISA plates with pre-coated monoclonal antibody at 37 °C for 2 h. After washing with PBS three times, the plates were blocked with complete RPMI medium containing 10% fetal bovine serum (FBS) for 2 h at 37 °C. Finally, 100 μL of streptavidin-HRP was added to each well for 20 min at room temperature. After washing 3 times, 100 μL of 3, 3′,5′,5′-tetramethylbenzidine (TMB) coloring agent was added to each well. Finally, color development was initiated by adding 100 μL of TMB buffer (100 μL/well) and terminated by adding 100 μL of H2SO4. The OD450 nm value was measured with an ELISA reader [22]. IL-35 concentration was assayed using a human IL-35 ELISA kit (#88-7357; eBioscience Inc., San Diego, USA) according to the manufacturer’s protocol. The optical density was measured at 450 nm. The reported concentration of IL-35 was determined by subtracting the concentration of IL-35 in PBS alone and represented by relative units compared to standard samples provided in the kit. All assays were performed in triplicate.
2.9. Foxp3 Staining and Intracellular Cytokine Staining
CD4+CD25+ Tregs showed Foxp3 expression as determined by FACS intra-cytoplasmic staining with APC-conjugated IgG1 anti-Foxp3Ab (eBioscience, San Diego, USA). For IL-10, TGF-β, and IL-35 detection, PE-conjugated IgG1 fluorescent antibodies were utilized (all from BD Pharmingen, San Diego, USA). We used the Intracellular Cytokine Staining Kit (BD biosciences) consisting of fixation/permeabilization buffer. In brief, the cells were fixed using fixation/permeabilization buffer, followed by washing the cells with PBS and then staining the cells using anti–Foxp3 and anti–IL-10, TGF-β, and IL-35 antibodies. The cells were finally suspended in 0.5 ml staining buffer and analyzed as previously described [26].
2.10. Detection of PD-1+ Analysis of TIICs by Flow Cytometry
After washing three times with PBS buffer; 104 cells of TIICs were divided into 1.5 cc centrifuge tubes. Fluorescent-conjugated anti-PD-1 antibodies; along with their corresponding fluorochrome-conjugated mouse immunoglobulin G isotype controls; were all obtained from PharMingen (San Diego; USA). Antibodies were added; and the reaction was carried out in the dark at 4°C for 30 min. After washing once; the cells were resuspended in PBS and analyzed by flow cytometry using BD FACSCalibur (Becton Dickinson; USA); as previously described [22]. Representative results are shown in histograms based on 104 gated cells in all conditions, and cell viability was >95%, as assessed by propidium iodide exclusion. Similar results were observed using at least 3 different TIICs donors.
2.11. Apoptosis Assays
Apoptosis was assessed by flow cytometric analysis of cells stained with annexin V-fluorescein isothiocyanate (FITC) and 7-AAD according to manufacturer’s instruction (Annexin V-FITC/7-AAD Apoptosis Kit, Elabscience, Houston, USA). TIICs and PGGCs co-culture cells were seeded at 106 cells/mL in 6-well plates followed by treated with lidocaine (1.5mM) or medium only for 72h. Briefly, cells were washed and resuspended in 500 μL PBS followed incubation with 10 μL 7-AAD and 10 μL annexin V-FITC at room temperature in the dark for 15 min and analyzed by flow cytometry.
2.12. Detection of Cytotoxicity of CD8+TIICs and PGCCs
The cytotoxicity of CD8+TIICs and PGCCs were estimated by quantification of LDH activity in the culture medium by using the QuantiChromTM LDH Cytotoxicity Assay Kit (BioAssay Systems, Hayward, USA) [27,28]. Briefly, cytotoxicity assays were carried out in 96-well plates with a final sample volume of 100 μl/well. Target cells (PGCCs) in 50 μl/well were co-cultured with effector cells (CD8+TIICs) at various effector to target ratios (5:1) for 4h [29].
2.13. Lidocaine Treated-CD8+TIICs Mediated Cytotoxicity Assay Using Time-Resolved Fluorometry
Lidocaine (1.5 mM) treated CD8+TIICs mediated cytotoxicity was determined using the DELFIA® EuTDA Cytotoxicity Reagents (PerkinElmer Life Sciences, Waltham, MA, USA), as described previously [30,31,32]. Briefly, target cells (PGGCs) were incubated with freshly prepared 10 μM BATDA (a fluorescence enhancing ligand) in 2 ml of culture medium for 30 min at 37°C, and washed. Next, 100 μl of BATDA-labeled target cells (PGCCs) were transferred into a round bottom sterile plate and co-cultured with lidocaine (1.5 mM) treated CD8+TIICs for 2 hours at effector/target ratios was 5:1. After incubation, 20 μl of supernatant from each well was transferred to the wells of flat-bottom 96 well plates. 180 μl of europium (Eu) solution was then added to form highly fluorescent and stable chelates (EuTDA), and the fluorescence of these chelates were measured by time resolved fluorometry (Enspire 2300-0000, PerkinElmer). The percent of specific release was calculated using (experimental release – spontaneous release)/ (maximum release – spontaneous release) X 100(%). All experiments were performed in triplicate.
2.14. Statistical Analysis
Figures were generated using GraphPad Prism 8.0 software (GraphPad Software, Inc., La Jolla, USA). Statistical analyses were also performed using this software. Differences between means were evaluated using the student’s t-test and were deemed significant at *p ≤ 0.05 **p ≤ 0.01.
3. Results
3.1. Lidocaine Inhibits IFN-γ Production by CD8+PBMCs and IL-12 Production by CD14+PBMCs
CD8 is a marker of cytotoxic T lymphocytes (CTLs), with CD8+ cells being preferred immune cells for targeting cancer [10]. CD14 is a key marker of macrophages [33]. To evaluate the effects of lidocaine on the secretion of IFN-γ from CD8+ and IL-12 from CD14+ PBMCs, cells were stimulated with the potent activators PMA and PHA, then cultured in the absence or presence of graded concentrations of lidocaine. The supernatant was collected at 72 hours and analyzed for the IFN-γ or IL-12 level. We found that the addition of lidocaine significantly inhibited the amount of IFN-γ secreted into the supernatant in a dose-dependent manner from CD8+ (Figure 1A) and IL-12 secreted into the supernatant in a dose-dependent manner from CD14+ PBMCs (Figure 1B).
3.2. Lidocaine Increases the Production of Anti-Inflammatory Cytokine IL-10 by CD4+CD25+PBMCs and CD14+PBMCs
CD4+CD25+T cells, also known as Tregs, strengthen immune tolerance [34]. To evaluate the effects of lidocaine on the secretion of IL-10 from CD4+CD25+ and CD14+ PBMCs, recent studies have disclosed variable effects of IL-10 on human M2 macrophage lineage cells [35]. We hypothesized that the secretion of IL-10 by CD14+ PBMCs might be stimulated by lidocaine. Cells were cultured in the absence or presence of graded concentrations of lidocaine. The supernatant was collected at 72 hours and analyzed for IL-10 content. IL-10 analysis showed a significant increase with graded concentrations of lidocaine in both CD4+CD25+PBMCs (Figure 2A) and CD14+ PBMCs (Figure 2B).
3.3. Lidocaine Increases the CD4+CD25+Tregs-Related Cytokine TGF-β and M2 CD14+Macrophage-Related Cytokine TGF-β
Recent studies have revealed the secretion effects of TGF-β on human M2-type macrophages and Tregs [36]. Cells were stimulated in the absence or presence of graded concentrations of lidocaine. The supernatant was collected at 72 hours and analyzed for the content of TGF-β. The current studies therefore examined in detail the effects of lidocaine on CD4+CD25+ and CD14+ PBMCs, as well as their production of TGF-β in response to lidocaine. TGF-β analysis showed a significant increase with graded concentrations of lidocaine in both CD4+CD25+ (Figure 2C) and CD14+ PBMCs (Figure 2D).
3.4. Lidocaine Increases a Novel Immunomodulator Cytokine IL-35 by CD4+CD25+PBMCs and CD14+PBMCs
To evaluate the effects of lidocaine on the secretion of IL-35 from CD4+CD25+ and CD14+ PBMCs, cells were stimulated in the absence or presence of graded concentrations of lidocaine. The supernatant was collected at 72 hours and analyzed for the content of IL-35. This study examined in detail the regulation of lidocaine on CD14+ PBMCs and CD4+CD25+ PBMCs, and their production of IL-35 in response to lidocaine. IL-35 analysis showed a significant increase with graded concentrations of lidocaine in both CD4+CD25+ (Figure 2E) and CD14+ PBMCs (Figure 2F). This is the first study to disclose variable secretion of IL-35 by lidocaine on human M2 CD14+ lineage cells. The study suggests that IL-35 is involved in the lidocaine-induced switch of M1 macrophages of CD14+ PBMCs to an M2-like phenotype macrophage of CD14+ PBMCs.
3.5. Lidocaine Does Not Affect the Viability of CD4+CD25+, CD8+, and CD14+ PBMCs.
Having shown that an effect of lidocaine on cytokine secretion could potentially result from reduced cellular function or cytotoxicity, we next aimed to determine the effect of lidocaine on the viability of CD4+CD25+, CD8+, and CD14+ PBMCs. Cell death was determined by a sub-G1 assay after 72 hours of culture with the designated concentrations of lidocaine. We found that increasing concentrations of lidocaine had only a slight and nonsignificant effect on the viability of CD4+CD25+, CD8+, and CD14+ PBMCs (Figure S1). This suggests that the mechanism for cytokine production under lidocaine treatment is mediated through non-cytotoxic effects.
3.6. Lidocaine Increases the Anti-Cancer-Related Cytokines IFN-γ by CD8+ TIICs and IL-12 by CD14+ TIICs
CD14+ tumor-infiltrating macrophages represent one of the main tumor-infiltrating immune cell types and are generally categorized into either of two functionally contrasting subtypes, namely classical activated M1 macrophages and alternatively activated M2 macrophages [37]. To evaluate the effects of lidocaine on the secretion of IFN-γ from CD8+ and IL-12 from CD14+ TIICs, cells were stimulated with graded concentrations of lidocaine. The supernatant was collected at 72 hours and analyzed for the content of IFN-γ or IL-12. Regulation and production of the anti-cancer-related cytokines IFN-γ in CD8+ TIICs and IL-12 in CD14+ TIICs by lidocaine were analyzed. IFN-γ showed a significant increase with graded concentrations of lidocaine in CD8+TIICs (A) and IL-12 showed a significant increase with graded concentrations of lidocaine in CD14+TIICs (B).
3.7. Lidocaine Inhibits IL-10, TGF-β, and IL-35 Production by CD4+CD25+ and CD14+TIICs
To evaluate the effects of lidocaine on the secretion of IL-10, TGF-β, and IL-35 from CD4+CD25+ and CD14+ TIICs, cells were stimulated with graded concentrations of lidocaine. The supernatant was collected at 72 hours and analyzed for the levels of IL-10, TGF-β, and IL-35. We found that lidocaine significantly inhibits the amount of IL-10, TGF-β, and IL-35 secreted by CD4+CD25+ (Figure 4A, C, E) and CD14+ TIICs (Figure 4B, D, F) in a dose-dependent manner. This study suggests that IL-35 is involved in the lidocaine-induced switch of M2 macrophages in CD14+ TIICs to an M1-like phenotype macrophage. M1-type macrophages have anti-tumor effects, enabling them to distinguish tumor cells from normal cells [37]. Lidocaine modulates the immunosuppression of normal PBMCs and enhances the anti-tumor effects of gastric TIICs (Figure 5).
3.8. Lidocaine Does Not Affect the Viability of CD4+CD25+, CD8+, and CD14+ TIICs.
To gain further insight into the responsible mechanism for the observed effect, we next aimed to determine the effect of lidocaine on the viability of CD4+CD25+, CD8+, and CD14+ TIICs. Cell death was determined by a sub-G1 assay after 72 hours of culture with the designated concentrations of lidocaine. We found that increasing concentrations of lidocaine have only a slight effect on the viability of CD4+CD25+, CD8+, and CD14+ TIICs (Figure S2). This suggests that the mechanism for cytokine production and inhibition under lidocaine treatment is mediated through non-cytotoxic effects.
3.9. Lidocaine Inhibits IL-10, TGF-β and IL-35 Production by the CD4+CD25+ Foxp3+ TIICs
While CD4 and CD25 are markers commonly associated with regulatory T cells (Tregs), these markers alone do not specifically identify Tregs. A more accurate identification requires the inclusion of additional markers such as FoxP3, which is a transcription factor crucial for Treg development and function. Currently, cell sorting can only use two colors at most, so we used PE and FITC to sort the traditional CD4 and CD25 markers. In the subsequent experiment shown in Figure 6, we used the FoxP3 marker additionally to repeat the experiment. Novel immunomodulator cytokine IL-35, IL-10, and TGF-β expression were also determined by intracellular cytokine staining. CD4+CD25+ TIICs were stimulated by lidocaine (1.5 mM) for 72 hours. We found that lidocaine decreases IL-35 production by CD4+CD25+Foxp3+ TIICs (A, B). Lidocaine also inhibits IL-10 production (Figure 6C, D) and TGF-β (Figure 6E, F) by CD4+CD25+Foxp3+ TIICs. This is the first study to reveal variable secretion of IL-35 by lidocaine from CD4+CD25+Foxp3+ TIICs.
3.10. A significant Decrease in PD-1 and Significant Increase IFN-γ Expression Was Observed in Lidocaine-Treated CD8+ TIICs through the NF-κB Signaling Pathway
IFN-γ has direct cytotoxic effects on tumor cells, thus potential cytotoxic effects on anti-tumor immune cells [38]. A significant increase in IFN-γ level was observed in lidocaine-treated CD8+ TIICs (Figure 7A, B). Previous studies have revealed that PD-1 inhibitory pathways optimize anti-tumor CD8+ T cell responses [39]. Reduced PD-1 expression enhances the antitumor effect and decreases immune escape [40]. PD-1 play vital roles in inhibiting immune responses [19]. To evaluate the effects of lidocaine on the expression of PD-1 in CD8+ TIICs, flow cytometry was used to assess PD-1 expression levels in lidocaine (1.5 mM)-treated CD8+ TIICs. A significant decrease in PD-1 expression was observed in lidocaine-treated CD8+ TIICs (Figure 7C, D). Lidocaine triggers anti-tumor immunity by restricting PD-1 expression and increase IFN-γ expression in CD8+ TIICs within the local gastric tumor microenvironment (TME). Previous study had indicated that PD-1 and IFN-γ downstream activation of NF-κB [41,42,43]. As shown in Figure 7, treatment with the NF-κB inhibitor resulted in reduced expression of IFN-γ and increased expression of PD-1 in lidocaine treated-CD8+TIICs, suggesting that lidocaine induced CD8+TIICs secretion of IFN-γ and decreased PD-1 expression through the NF-κB signaling pathway. Lidocaine treated led to CD8+ T cells increased IFN-γ secretion and decreased PD-1 expression through NF-kB. Inhibited expression of NF-kb decreased expression of IFN-γ and increased PD-1 expression. In conclusion, inhibiting NF-kb expression decreased the CD8+ T cell function caused by lidocaine. The current data highlight the importance of lidocaine in gastric cancer by increase CD8+T cell function.
3.11. Lidocaine Does Not Affect the Viability of Primary Gastric Cancer Cells
A recent study suggested that lidocaine (5 mM and 10 mM) significantly induces apoptosis in human gastric cancer cell lines [44]. Cell death of normal PBMCs and gastric TIICs was determined by a sub-G1 assay after 72 hours of culture with lidocaine at concentrations of 5 mM and 10 mM. We found that both 5 mM and 10 mM concentrations of lidocaine have a significant effect on the viability of gastric TIICs (Figure 8 A, B) and normal PBMCs (Figure 8 C, D), suggesting a cytotoxic effect. Primary gastric cancer cell death was determined by a sub-G1 assay after 72 hours of culture with lidocaine at concentrations of 0.5 mM and 1.5 mM. We found that increasing concentrations (0-1.5 mM) of lidocaine has only a slight and nonsignificant effect on primary gastric cancer cell viability (Figure 8 E, F).
3.12. Lidocaine-Treated CD8+TIICs Induced Primary Gastric Cancer Cell Death
To investigate whether lidocaine-treated CD8+ TIICs induce PGCCs death in TME, we examined responsiveness of PGCCs co-cultured with lidocaine-treated CD8+ TIICs. Lidocaine-treated CD8+TIICs and lidocaine-treated PGCCs only induced minimal cell death, whereas lidocaine-treated CD8+TIICs co-culture with PGCCs induced strong cell death (Figure 8 G, H). In vitro co-cultures revealed that cell death of PGCCs were enhanced by lidocaine-treated CD8+TIICs.
3.13. Lidocaine Enhances the Killing Activity of CD8+ TIICs against Gastric Cancer
To investigate whether lidocaine-treated CD8+ TIICs can kill PGCCs, we examined the responsiveness of PGCCs co-cultured with lidocaine-treated CD8+ TIICs for 48 hours and 72 hours. Lidocaine-untreated CD8+ TIICs showed minimal cell death of PGCCs, whereas lidocaine-treated CD8+TIICs induced strong cell death. The cytotoxic effect on PGCCs was assessed by an LDH release assay. As shown in Figure S3, lidocaine-treated CD8+TIICs displayed strong cell killing activity on PGCCs.
The cytotoxic effects of T cells on tumor cells in vitro are usually difficult to observe, and it is suggested to supplement related experiments. Using LDH release to observe is not accurate enough, as it cannot be determined whether this LDH comes from CD8+T cells or tumor cells. Therefore, CD8+TIICs cytotoxicity was assessed using DELFIA EuTDA cell cytotoxicity assays. Tumor cells (PGGCs) were labeled with BATDA (a fluorescence enhancing ligand). Subsequently, lidocaine-treated CD8+TIICs were added to the BATDA labeled PGCCs. Fluorescence only comes from lysis tumor cells (PGCCs) was measured. Lidocaine-untreated CD8+TIICs showed minimal cytotoxicity of PGCCs, whereas lidocaine-treated CD8+TIICs induced strong cytotoxic effect of PGCCs. As shown in Figure 9, lidocaine-treated CD8+ TIICs displayed strong cell killing activity on PGCCs.
Lidocaine blockade of PD-1 and increase IFN-γ triggers cytotoxic CD8+ T cell activation through NF-kb pathway (Figure 7), resulting in the killing of PGCCs through immunogenic cell death (Figure 9). Taken together, our findings reveal that lidocaine enhances anti-gastric cancer immunity of TIICs, indicating a potential therapeutic value for gastric cancer (Figure 10).
4. Discussion
Current animal models of gastric cancer have certain limitations and cannot fully mimic the complexity of human gastric cancer [45]. This has led us to use gastric cancer specimens obtained from patients to simulate in vivo conditions, reflecting the pathophysiology of human diseases more accurately. We found that lidocaine has potential applications in gastric cancer research, particularly due to its anti-tumor and anti-inflammatory effects. Given the limitations of gastric cancer animal models, using gastric cancer specimens from patients along with advanced culture techniques can provide a more accurate assessment of the efficacy and mechanisms of lidocaine.
This study investigates the role of lidocaine in modulating the immune response and its anti-tumor effect in gastric primary tumor-infiltrating immune cells (TIICs) within the tumor microenvironment (TME). The findings suggest that lidocaine regulates the suppressive activity of human Treg cells or alternative (M2) macrophages, providing new opportunities to improve the outcome of cancer immunotherapy. Given that long-term treatment effects of multiple immunotherapy approaches have been unsatisfactory in solid tumors, especially in gastric cancer, these findings hold promise for advancing treatment strategies [46]. Gastric cancer may partly result from the immunosuppressive status in the TME mediated by negative immune cells such as Tregs or M2 macrophages, and immune inhibitory cytokines such as IL-10 and TGF-β [46,47]. However, little is known about cytokine expression in lidocaine-treated gastric TIICs. Thus, the present study investigates cytokine expression levels in lidocaine-treated gastric tumor-infiltrating immune cells (TIICs) to explore the potential role of lidocaine in anti-cancer activity within the gastric TME.
IL-10 is a well-known anti-inflammatory cytokine that limits the immune response during infections and is produced by nearly every type of cells in the immune system [48]. It inhibits inflammatory cytokines [49]. This study is the first to show that lidocaine causes an increase in IL-10 levels in lidocaine-treated CD4+CD25+ and CD14+ PBMCs, while a decrease in IL-10 levels is observed in lidocaine-treated CD4+CD25+ and CD14+ TIICs. CD14+ tumor-infiltrating macrophages expressing IL-10 have been found and enriched in gastric cancer patients to facilitate immune evasion [50]. Lidocaine suppresses IL-10 secretion in CD14+ tumor-infiltrating macrophages and CD4+CD25+ tumor-infiltrating Tregs. Taken together, it is likely that IL-10 is involved in lidocaine-modulated mechanisms against gastric cancer. PD-1 have been shown to have inhibitory functions in T cells [51]. Interleukin-10 receptor signaling promotes the maintenance of PD-1+CD8+ T cell population [52]. The blockade of PD-1, combined with IL-10 neutralization, augmented the anti-tumor response [53]. Our results demonstrate a decrease in PD-1 expression in lidocaine-treated CD8+TIICs through NF-kb pathway, suggesting potential for therapy in gastric cancer.
Distinct from other members of the IL-12 family, IL-35 is a novel inhibitory cytokine that suppresses T cell proliferation [52]. Recently, IL-35 has been proposed as a novel immunosuppressive cytokine [54]. It has been demonstrated to promote tumor angiogenesis and inhibit the antitumor cytotoxic CD8+ T cell response [55]. In addition, IL-35 expression is considered to be associated with colorectal cancer progression and prognosis [56]. We first found that lidocaine, in a dose-dependent manner, significantly reduces levels of IL-35 in CD4+CD25+ or CD14+ TIICs. The present report analyzes the expression of IL-35 in lidocaine-treated PBMCs and TIICs. To the best of our knowledge, this is the first study to explore the function of the novel cytokine IL-35 in lidocaine-treated PBMCs and TIICs.
The present study suggests that in normal immune cells, lidocaine exerts its functions by inhibiting the production of IFN-γ and IL-12, while increasing the production of anti-inflammatory cytokines IL-10 and TGF-β, as well as a novel immunomodulator cytokine IL-35 from normal PBMCs. Our findings provide new insights into the anti-inflammatory mechanisms of lidocaine and a novel molecular target. In the TME, Tregs and tumor-infiltrating macrophages are considered to be sources of IL-35 [57]. It was reported that tumor-derived IL-35 increases tumorigenesis with a pro-tumor effect, and IL-35 production in the TME increases suppressor cells [58]. Like TGF-β and IL-10, IL-35 can also induce the development of CD4+CD25+Foxp3+ Treg population [16]. CD4+CD25+Foxp3+ Tregs are recruited and expanded in tumors and constitute an important mechanism utilized by tumor cells to evade protective immunity and support metastatic growth [59]. Lidocaine exerts its functions on TME by inhibiting the production of IL-10 and TGF-β as well as a novel immunomodulator cytokine IL-35 from CD4+CD25+Foxp3+TIICs. The differential anti-inflammatory and pro-inflammatory responses to lidocaine observed between TIICs and normal PBMCs may be attributed to the distinct microenvironments and functional states of these cell types. TIICs are directly exposed to the gastric tumor microenvironment, which can influence their response to lidocaine differently compared to PBMCs, which are typically isolated from normal peripheral blood and not directly exposed to tumor-associated factors.
Our findings reveal that lidocaine modulates the antitumor effects by decreasing IL-10, TGF-β, IL-35 level in CD4+CD25+Foxp3+TIICs, decreasing PD-1 and increase IFN-g expression in cytotoxic CD8+TIICs through NF-kb pathway. Lidocaine is a common local anesthetic; however, recent studies suggest that it induces apoptosis in gastric cancer cell lines in vitro [44], although the use of 5 and 10 mM lidocaine has been found to be cytotoxic. We have found that lidocaine (0-1.5 mM) has no cytotoxicity in either TIICs or PGGCs. Lidocaine-untreated CD8+TIICs showed minimal cell death of PGCCs, whereas lidocaine (1.5 mM)-treated CD8+TIICs induced strong cell death. Lidocaine (1.5 mM)-treated CD8+ TIICs displayed strong cell killing activity on PGCCs. The lidocaine (1.5 mM) blockade of PD-1 triggers cytotoxic CD8+ T cell activation, resulting in the killing of PGCCs through immunogenic cell death. Administered local anesthetics such as lidocaine have favorable effects on overall gastric cancer patients’ TIICs. Lidocaine anesthesia may thus influence the TME of gastric cancer. This study further demonstrates that lidocaine may be involved in the anti-cancer effect of gastric cancer.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Lidocaine does not affect CD4+CD25+PBMCs, CD8+PBMCs and CD14+ PBMCs; Figure S2: Lidocaine does not affect CD4+CD25+TIICs, CD8+TIICs and CD14+TIICs viability; Figure S3: Lidocaine-sensitized CD8+TIICs dependent immunogenic cell death of PGCCs.
Author Contributions
Y.W.-M.C. conceived this project. I.C.-F.W., T.L. and H.W. designed this project. Y.W., M.C., I.C., F.W., T.L. and H.W. performed this research. B.N. and H.L. analyzed and interpreted the data. Y.W. and H.L. wrote this manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Ditmanson Medical Foundation of Chia-Yi Christian Hospital under grant number R111-03 and Ministry of Science and Technology of Taiwan (112-2313-B-005-050-MY3), The iEGG and Animal Biotechnology Center from The Feature Areas Research Center Program within the framework of the Higher Education Sprout Project by the Ministry of Education (MOE) in Taiwan (112S0023A and 113S0023A), Taichung Veterans General Hospital (TCVGH-NCHU 1127605 and TCVGH-NCHU-1137607).
Institutional Review Board Statement
Written informed consent was obtained from each patient in accordance with local institutional ethics review and has been approved by the Ethical and Scientific Committee of Taichung Veterans General Hospital (TCVGH-IRB no. SF22141B#1).
Informed Consent Statement
Informed consent was obtained from all patients involved in the study.
Data Availability Statement
The data that support the findings of our study are available on request from the corresponding author.
Acknowledgments
The authors thank all staff members who assisted with the study.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Loftus, J.P.; Williams, J.M.; Belknap, J.K.; Black, S.J. In vivo priming and ex vivo activation of equine neutrophils in black walnut extract-induced equine laminitis is not attenuated by systemic lidocaine administration. Vet Immunol Immunopathol 2010, 138, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Renzi, P.M.; Ginns, L.C. Effect of lidocaine on natural killer activity: Rapid inhibition of lysis. Immunopharmacol Immunotoxicol 1990, 12, 417–437. [Google Scholar] [CrossRef] [PubMed]
- Elizagaray, M.L.; Mazitelli, I.; Pontoriero, A.; Baumeister, E.; Docena, G.; Raimondi, C.; Correger, E.; Rumbo, M. Lidocaine reinforces the anti-inflammatory action of dexamethasone on myeloid and epithelial cells activated by inflammatory cytokines or SARS-CoV-2 infection. Biomedical journal 2022. [Google Scholar] [CrossRef] [PubMed]
- Feng, G.; Liu, S.; Wang, G.L.; Liu, G.J. Lidocaine attenuates lipopolysaccharide-induced acute lung injury through inhibiting NF-kappaB activation. Pharmacology 2008, 81, 32–40. [Google Scholar] [CrossRef]
- Sun, H.; Sun, Y. Lidocaine inhibits proliferation and metastasis of lung cancer cell via regulation of miR-539/EGFR axis. Artificial cells, nanomedicine, and biotechnology 2019, 47, 2866–2874. [Google Scholar] [CrossRef]
- Chen, L.J.; Ding, Y.B.; Ma, P.L.; Jiang, S.H.; Li, K.Z.; Li, A.Z.; Li, M.C.; Shi, C.X.; Du, J.; Zhou, H.D. The protective effect of lidocaine on lipopolysaccharide-induced acute lung injury in rats through NF-κB and p38 MAPK signaling pathway and excessive inflammatory responses. European review for medical and pharmacological sciences 2018, 22, 2099–2108. [Google Scholar]
- Lahat, A.; Ben-Horin, S.; Lang, A.; Fudim, E.; Picard, O.; Chowers, Y. Lidocaine down-regulates nuclear factor-kappaB signalling and inhibits cytokine production and T cell proliferation. Clinical and experimental immunology 2008, 152, 320–327. [Google Scholar] [CrossRef] [PubMed]
- Johnson BD, Jing W, Orentas RJ. CD25+ regulatory T cell inhibition enhances vaccine-induced immunity to neuroblastoma. J Immunother 2007, 30, 203–214. [Google Scholar] [CrossRef]
- Hesse, M.; Piccirillo, C.A.; Belkaid, Y.; Prufer, J.; Mentink-Kane, M.; Leusink, M.; Cheever, A.W.; Shevach, E.M.; Wynn, T.A. The pathogenesis of schistosomiasis is controlled by cooperating IL-10-producing innate effector and regulatory T cells. J Immunol 2004, 172, 3157–3166. [Google Scholar] [CrossRef]
- Farhood B, Najafi M, Mortezaee K. CD8(+) cytotoxic T lymphocytes in cancer immunotherapy: A review. J Cell Physiol 2019, 234, 8509–8521. [Google Scholar] [CrossRef]
- Dinarello, C.A. Anti-cytokine therapeutics and infections. Vaccine 2003, 21 Suppl. S2, S24–S34. [Google Scholar] [CrossRef] [PubMed]
- Macedo, N.; Miller, D.M.; Haq, R.; Kaufman, H.L. Clinical landscape of oncolytic virus research in 2020. Journal for immunotherapy of cancer 2020, 8. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Cao, S.; Mitsuhashi, M.; Xiang, Z.; Ma, X. Genome-wide analysis of molecular changes in IL-12-induced control of mammary carcinoma via IFN-gamma-independent mechanisms. Journal of immunology (Baltimore, Md: 1950) 2004, 172, 4111–4122. [Google Scholar] [CrossRef] [PubMed]
- Hung, C.H.; Hsu, H.Y.; Chiou, H.C.; Tsai, M.L.; You, H.L.; Lin, Y.C.; Liao, W.T.; Lin, Y.C. Arsenic Induces M2 Macrophage Polarization and Shifts M1/M2 Cytokine Production via Mitophagy. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef] [PubMed]
- Kang MJ, Jang AR, Park JY, Ahn JH, Lee TS, Kim DY, Lee MS, Hwang S, Jeong YJ, Park JH. IL-10 Protects Mice From the Lung Infection of Acinetobacter baumannii and Contributes to Bacterial Clearance by Regulating STAT3-Mediated MARCO Expression in Macrophages. Frontiers in immunology 2020, 11, 270.
- Collison, L.W.; Workman, C.J.; Kuo, T.T.; Boyd, K.; Wang, Y.; Vignali, K.M.; Cross, R.; Sehy, D.; Blumberg, R.S.; Vignali, D.A. The inhibitory cytokine IL-35 contributes to regulatory T-cell function. Nature 2007, 450, 566–569. [Google Scholar] [CrossRef]
- Feng, J.; Wu, Y. Interleukin-35 ameliorates cardiovascular disease by suppressing inflammatory responses and regulating immune homeostasis. International immunopharmacology 2022, 110, 108938. [Google Scholar] [CrossRef]
- He, W.; Hao, S.; Dong, X.; Zhang, D.; Jia, Z. Circulating cytokine profile and modulation of regulatory T cells in chronic hepatitis B patients with type 2 diabetes mellitus. Bosnian journal of basic medical sciences 2022. [Google Scholar] [CrossRef]
- Han Y, Liu D, Li L. PD-1/PD-L1 pathway: Current researches in cancer. Am J Cancer Res 2020, 10, 727–742. [Google Scholar]
- Wei SC, Levine JH, Cogdill AP, Zhao Y, Anang NAS, Andrews MC, Sharma P, Wang J, Wargo JA, Pe’er D et al. : Distinct Cellular Mechanisms Underlie Anti-CTLA-4 and Anti-PD-1 Checkpoint Blockade. Cell 2017, 170, 1120–1133. [Google Scholar] [CrossRef]
- Huang Q, Wu X, Wang Z, Chen X, Wang L, Lu Y, Xiong D, Liu Q, Tian Y, Lin H et al. : The primordial differentiation of tumor-specific memory CD8(+) T cells as bona fide responders to PD-1/PD-L1 blockade in draining lymph nodes. Cell 2022, 185, 4049–4066. [Google Scholar] [CrossRef] [PubMed]
- Chen YY, Yang WC, Chang YK, Wang CY, Huang WR, Li JY, Chuang KP, Wu HY, Chang CD, Nielsen BL et al. : Construction of polycistronic baculovirus surface display vectors to express the PCV2 Cap(d41) protein and analysis of its immunogenicity in mice and swine. Vet Res 2020, 51, 112. [Google Scholar] [CrossRef]
- Dudley, M.E.; Wunderlich, J.R.; Shelton, T.E.; Even, J.; Rosenberg, S.A. Generation of tumor-infiltrating lymphocyte cultures for use in adoptive transfer therapy for melanoma patients. J Immunother 2003, 26, 332–342. [Google Scholar] [CrossRef]
- Aziz, F.; Yang, X.; Wen, Q.; Yan, Q. A method for establishing human primary gastric epithelial cell culture from fresh surgical gastric tissues. Mol Med Rep 2015, 12, 2939–2944. [Google Scholar] [CrossRef]
- Smoot, D.T.; Sewchand, J.; Young, K.; Desbordes, B.C.; Allen, C.R.; Naab, T. A method for establishing primary cultures of human gastric epithelial cells. Methods Cell Sci 2000, 22, 133–136. [Google Scholar] [CrossRef]
- Sultan, M.; Alghetaa, H.; Mohammed, A.; Abdulla, O.A.; Wisniewski, P.J.; Singh, N.; Nagarkatti, P.; Nagarkatti, M. The Endocannabinoid Anandamide Attenuates Acute Respiratory Distress Syndrome by Downregulating miRNA that Target Inflammatory Pathways. Front Pharmacol 2021, 12, 644281. [Google Scholar] [CrossRef] [PubMed]
- Vergis, J.; Malik, S.V.S.; Pathak, R.; Kumar, M.; Kurkure, N.V.; Barbuddhe, S.B.; Rawool, D.B. Exploring Galleria mellonella larval model to evaluate antibacterial efficacy of Cecropin A (1-7)-Melittin against multi-drug resistant enteroaggregative Escherichia coli. Pathog Dis 2021, 79. [Google Scholar] [CrossRef] [PubMed]
- Ilkow CS, Marguerie M, Batenchuk C, Mayer J, Ben Neriah D, Cousineau S, Falls T, Jennings VA, Boileau M, Bellamy D et al. : Reciprocal cellular cross-talk within the tumor microenvironment promotes oncolytic virus activity. Nat Med 2015, 21, 530–536. [Google Scholar] [CrossRef]
- Zhou Y, Chen CL, Jiang SW, Feng Y, Yuan L, Chen P, Zhang L, Huang S, Li J, Xia JC et al. : Retrospective analysis of the efficacy of adjuvant CIK cell therapy in epithelial ovarian cancer patients who received postoperative chemotherapy. Oncoimmunology 2019, 8, e1528411. [Google Scholar] [CrossRef]
- Snyder KM, Dixon KJ, Davis Z, Hosking M, Hart G, Khaw M, Matson A, Bjordahl R, Hancock B, Shirinbak S et al.: iPSC-derived natural killer cells expressing the FcγR fusion CD64/16A can be armed with antibodies for multitumor antigen targeting. J Immunother Cancer 2023, 11.
- Yao L, Hou J, Wu X, Lu Y, Jin Z, Yu Z, Yu B, Li J, Yang Z, Li C et al.: Cancer-associated fibroblasts impair the cytotoxic function of NK cells in gastric cancer by inducing ferroptosis via iron regulation. Redox Biol 2023, 67, 102923.
- Sun, L.; Jiang, G.; Ng, Y.Y.; Xiao, L.; Du, Z.; Wang, S.; Zhu, J. T cells with split CARs specific for NKG2D ligands and PD-L1 exhibit improved selectivity towards monocyte-derived cells while effective in eliminating acute myeloid leukaemia in vivo. J Cancer Res Clin Oncol 2023, 149, 10189–10201. [Google Scholar] [CrossRef] [PubMed]
- Jimenez-Duran G, Luque-Martin R, Patel M, Koppe E, Bernard S, Sharp C, Buchan N, Rea C, de Winther MPJ, Turan N et al.: Pharmacological validation of targets regulating CD14 during macrophage differentiation. EBioMedicine 2020, 61, 103039.
- Yu Q, Xu M, Yu F, Jin Y. CD4(+)CD25(+) regulatory T cells as a therapeutic target in rheumatoid arthritis. Cent Eur J Immunol 2014, 39, 100–103. [Google Scholar]
- Liu Q, Yang C, Wang S, Shi D, Wei C, Song J, Lin X, Dou R, Bai J, Xiang Z et al. : Wnt5a-induced M2 polarization of tumor-associated macrophages via IL-10 promotes colorectal cancer progression. Cell Commun Signal 2020, 18, 51. [Google Scholar] [CrossRef] [PubMed]
- Ma X, Gao Y, Chen Y, Liu J, Yang C, Bao C, Wang Y, Feng Y, Song X, Qiao S. M2-Type Macrophages Induce Tregs Generation by Activating the TGF-β/Smad Signalling Pathway to Promote Colorectal Cancer Development. Onco Targets Ther 2021, 14, 5391–5402. [CrossRef]
- Pan, Y.; Yu, Y.; Wang, X.; Zhang, T. Tumor-Associated Macrophages in Tumor Immunity. Front Immunol 2020, 11, 583084. [Google Scholar] [CrossRef]
- Gocher, A.M.; Workman, C.J.; Vignali, D.A.A. Interferon-γ: Teammate or opponent in the tumour microenvironment? Nat Rev Immunol 2022, 22, 158–172. [Google Scholar] [CrossRef]
- Banta KL, Xu X, Chitre AS, Au-Yeung A, Takahashi C, O’Gorman WE, Wu TD, Mittman S, Cubas R, Comps-Agrar L et al. : Mechanistic convergence of the TIGIT and PD-1 inhibitory pathways necessitates co-blockade to optimize anti-tumor CD8(+) T cell responses. Immunity 2022, 55, 512–526. [Google Scholar] [CrossRef]
- Yang, R.; Pei, T.; Huang, R.; Xiao, Y.; Yan, J.; Zhu, J.; Zheng, C.; Xiao, W.; Huang, C. Platycodon grandiflorum Triggers Antitumor Immunity by Restricting PD-1 Expression of CD8(+) T Cells in Local Tumor Microenvironment. Front Pharmacol 2022, 13, 774440. [Google Scholar] [CrossRef]
- Garg, S.K.; Welsh, E.A.; Fang, B.; Hernandez, Y.I.; Rose, T.; Gray, J.; Koomen, J.M.; Berglund, A.; Mulé, J.J.; Markowitz, J. Multi-Omics and Informatics Analysis of FFPE Tissues Derived from Melanoma Patients with Long/Short Responses to Anti-PD1 Therapy Reveals Pathways of Response. Cancers (Basel) 2020, 12. [Google Scholar] [CrossRef] [PubMed]
- Bally AP, Lu P, Tang Y, Austin JW, Scharer CD, Ahmed R, Boss JM. NF-κB regulates PD-1 expression in macrophages. J Immunol 2015, 194, 4545–4554. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Sun, Z.; Wang, Y.; Miao, C. Long-term use of indomethacin leads to poor prognoses through promoting the expression of PD-1 and PD-L2 via TRIF/NF-κB pathway and JAK/STAT3 pathway to inhibit TNF-α and IFN-γ in hepatocellular carcinoma. Exp Cell Res 2015, 337, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Ye, L.; Zhang, Y.; Chen, Y.J.; Liu, Q. Anti-tumor effects of lidocaine on human gastric cancer cells in vitro. Bratisl Lek Listy 2019, 120, 212–217. [Google Scholar] [CrossRef] [PubMed]
- Hayakawa, Y.; Fox, J.G.; Gonda, T.; Worthley, D.L.; Muthupalani, S.; Wang, T.C. Mouse models of gastric cancer. Cancers (Basel) 2013, 5, 92–130. [Google Scholar] [CrossRef]
- Chow, A.; Perica, K.; Klebanoff, C.A.; Wolchok, J.D. Clinical implications of T cell exhaustion for cancer immunotherapy. Nat Rev Clin Oncol 2022, 19, 775–790. [Google Scholar] [CrossRef]
- Wen, Z.; Sun, H.; Zhang, Z.; Zheng, Y.; Zheng, S.; Bin, J.; Liao, Y.; Shi, M.; Zhou, R.; Liao, W. High baseline tumor burden-associated macrophages promote an immunosuppressive microenvironment and reduce the efficacy of immune checkpoint inhibitors through the IGFBP2-STAT3-PD-L1 pathway. Cancer Commun (Lond) 2023, 43, 562–581. [Google Scholar] [CrossRef]
- Steen, E.H.; Wang, X.; Balaji, S.; Butte, M.J.; Bollyky, P.L.; Keswani, S.G. The Role of the Anti-Inflammatory Cytokine Interleukin-10 in Tissue Fibrosis. Adv Wound Care (New Rochelle) 2020, 9, 184–198. [Google Scholar] [CrossRef]
- Degboé, Y.; Rauwel, B.; Baron, M.; Boyer, J.F.; Ruyssen-Witrand, A.; Constantin, A.; Davignon, J.L. Polarization of Rheumatoid Macrophages by TNF Targeting Through an IL-10/STAT3 Mechanism. Frontiers in immunology 2019, 10, 3. [Google Scholar] [CrossRef]
- Zhang H, Li R, Cao Y, Gu Y, Lin C, Liu X, Lv K, He X, Fang H, Jin K et al. : Poor Clinical Outcomes and Immunoevasive Contexture in Intratumoral IL-10-Producing Macrophages Enriched Gastric Cancer Patients. Ann Surg 2022, 275, e626–e635. [Google Scholar] [CrossRef]
- Ahn, E.; Araki, K.; Hashimoto, M.; Li, W.; Riley, J.L.; Cheung, J.; Sharpe, A.H.; Freeman, G.J.; Irving, B.A.; Ahmed, R. Role of PD-1 during effector CD8 T cell differentiation. Proc Natl Acad Sci U S A 2018, 115, 4749–4754. [Google Scholar] [CrossRef] [PubMed]
- Cai, Z.; Zhang, S.; Wu, P.; Ren, Q.; Wei, P.; Hong, M.; Feng, Y.; Wong, C.K.; Tang, H.; Zeng, H. A novel potential target of IL-35-regulated JAK/STAT signaling pathway in lupus nephritis. Clinical and translational medicine 2021, 11, e309. [Google Scholar] [CrossRef]
- Moeini, S.; Saeidi, M.; Fotouhi, F.; Mondanizadeh, M.; Shirian, S.; Mohebi, A.; Gorji, A.; Ghaemi, A. Synergistic effect of programmed cell death protein 1 blockade and secondary lymphoid tissue chemokine in the induction of anti-tumor immunity by a therapeutic cancer vaccine. Arch Virol 2017, 162, 333–346. [Google Scholar] [CrossRef] [PubMed]
- Wu W, Jiang H, Li Y, Yan MX. IL-35 expression is increased in laryngeal squamous cell carcinoma and in the peripheral blood of patients. Oncol Lett 2017, 13, 3303–3308. [Google Scholar] [CrossRef] [PubMed]
- Jiang H, Zhang T, Yan MX, Wu W. IL-35 inhibits CD8(+) T cells activity by suppressing expression of costimulatory molecule CD28 and Th1 cytokine production. Transl Cancer Res 2019, 8, 1319–1325. [Google Scholar] [CrossRef]
- Zeng JC, Zhang Z, Li TY, Liang YF, Wang HM, Bao JJ, Zhang JA, Wang WD, Xiang WY, Kong B et al. : Assessing the role of IL-35 in colorectal cancer progression and prognosis. Int J Clin Exp Pathol 2013, 6, 1806–1816. [Google Scholar]
- Huang Y, Hu H, Liu L, Ye J, Wang Z, Que B, Liu W, Shi Y, Zeng T, Shi L et al.: Interleukin-12p35 Deficiency Reverses the Th1/Th2 Imbalance, Aggravates the Th17/Treg Imbalance, and Ameliorates Atherosclerosis in ApoE-/- Mice. Mediators Inflamm 2019, 2019, 3152040.
- Olson, B.M.; Jankowska-Gan, E.; Becker, J.T.; Vignali, D.A.; Burlingham, W.J.; McNeel, D.G. Human prostate tumor antigen-specific CD8+ regulatory T cells are inhibited by CTLA-4 or IL-35 blockade. J Immunol 2012, 189, 5590–5601. [Google Scholar] [CrossRef]
- Vinay DS, Ryan EP, Pawelec G, Talib WH, Stagg J, Elkord E, Lichtor T, Decker WK, Whelan RL, Kumara H et al.: Immune evasion in cancer: Mechanistic basis and therapeutic strategies. Semin Cancer Biol 2015, 35 Suppl, S185–s198.
Figure 1.
Lidocaine reduces the secretion levels of IFN-γ by CD8+ PBMCs and IL-12 by CD14+ PBMCs. The effects of lidocaine (0.25 mM to 1.5 mM) on PMA and PHA-stimulated IFN-γ secretion from CD8+ PBMCs (A) were investigated. The IFN-γ level in the supernatant was determined at 72 hours by ELISA. Additionally, CD14+ PBMCs (B) were stimulated by PMA and PHA, then cultured in the absence or presence of graded concentrations of lidocaine. The IL-12 level in the supernatant was also determined at 72 hours by ELISA. Data are from distinct samples and are presented as the mean± SEM from three different experiments, each performed in duplicate. *P < 0.05, **P < 0.01. NC: negative control (non-stimulated cells).
Figure 1.
Lidocaine reduces the secretion levels of IFN-γ by CD8+ PBMCs and IL-12 by CD14+ PBMCs. The effects of lidocaine (0.25 mM to 1.5 mM) on PMA and PHA-stimulated IFN-γ secretion from CD8+ PBMCs (A) were investigated. The IFN-γ level in the supernatant was determined at 72 hours by ELISA. Additionally, CD14+ PBMCs (B) were stimulated by PMA and PHA, then cultured in the absence or presence of graded concentrations of lidocaine. The IL-12 level in the supernatant was also determined at 72 hours by ELISA. Data are from distinct samples and are presented as the mean± SEM from three different experiments, each performed in duplicate. *P < 0.05, **P < 0.01. NC: negative control (non-stimulated cells).
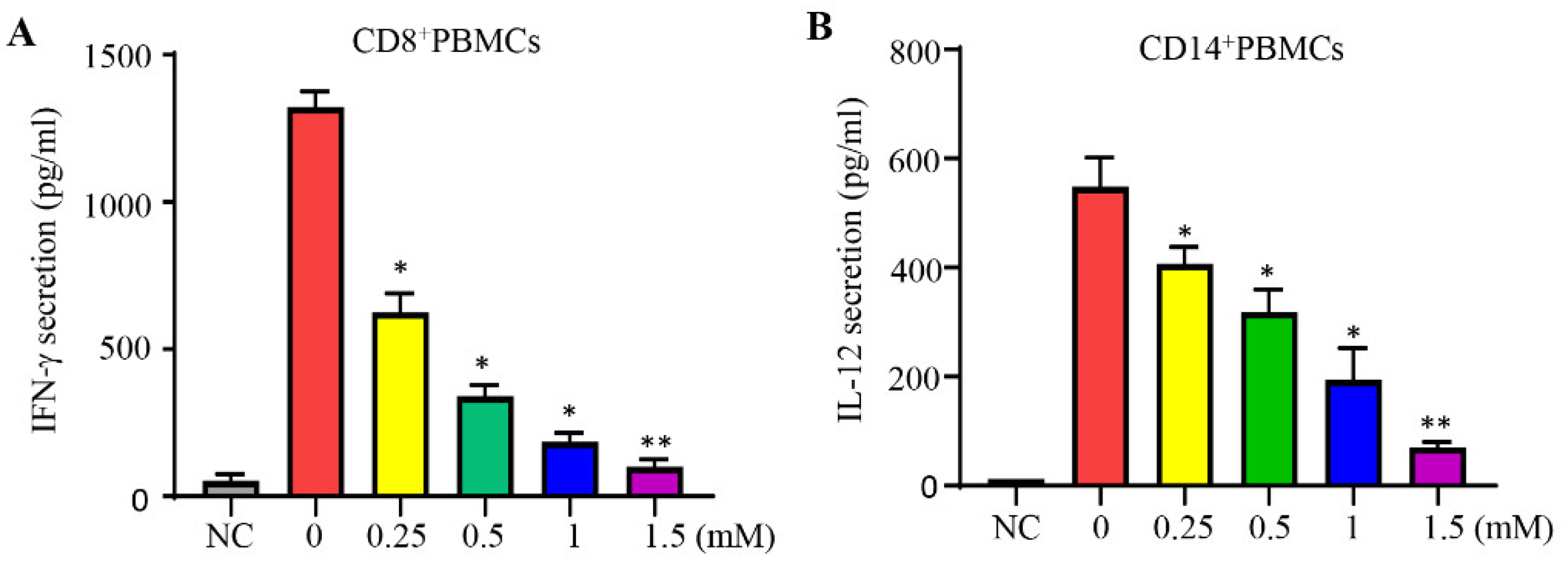
Figure 2.
Lidocaine induces the secretion of IL-10, TGF-β, and IL-35 by CD4+C25+ and CD14+ PBMCs. The effect of lidocaine on the secretion of IL-10 (A, B), TGF-β (C, D), and IL-35 (E, F) from CD4+CD25+ and CD14+ PBMCs was investigated. CD4+CD25+ and CD14+ PBMCs were cultured in the absence or presence of graded concentrations of lidocaine (0.25 mM to 1.5 mM). The levels of IL-10, TGF-β, and IL-35 in the supernatant were determined at 72 hours by ELISA. Data are from distinct samples and are presented as the mean± SEM from three different experiments, each performed in duplicate. *P < 0.05, **P < 0.01.
Figure 2.
Lidocaine induces the secretion of IL-10, TGF-β, and IL-35 by CD4+C25+ and CD14+ PBMCs. The effect of lidocaine on the secretion of IL-10 (A, B), TGF-β (C, D), and IL-35 (E, F) from CD4+CD25+ and CD14+ PBMCs was investigated. CD4+CD25+ and CD14+ PBMCs were cultured in the absence or presence of graded concentrations of lidocaine (0.25 mM to 1.5 mM). The levels of IL-10, TGF-β, and IL-35 in the supernatant were determined at 72 hours by ELISA. Data are from distinct samples and are presented as the mean± SEM from three different experiments, each performed in duplicate. *P < 0.05, **P < 0.01.
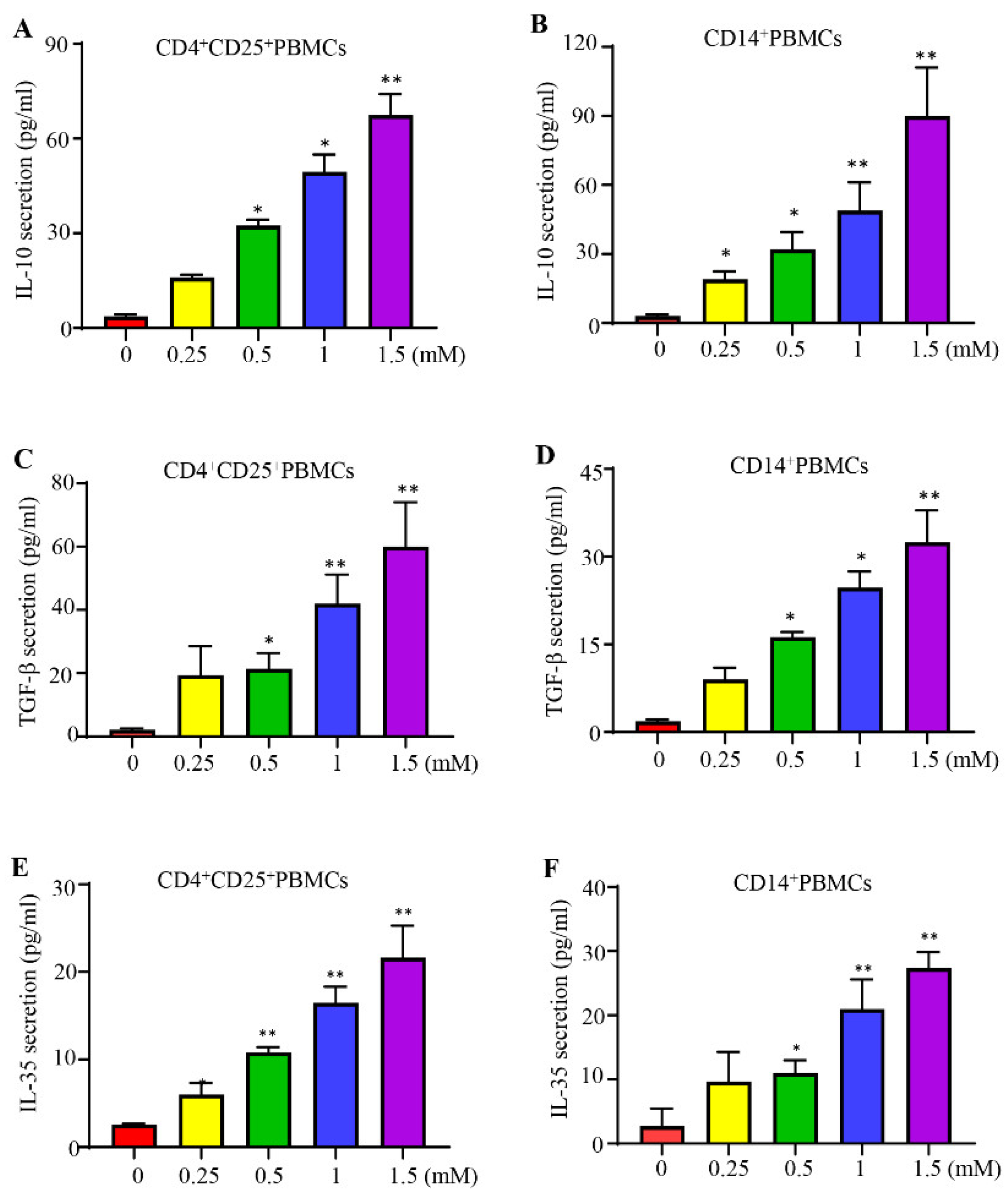
Figure 3.
The effect of lidocaine on IFN-γ secretion by CD8+ TIICs and IL-12 secretion by CD14+ TIICs. The effects of lidocaine on IFN-γ secretion from CD8+ TIICs (A) and IL-12 secretion from CD14+ TIICs (B) were investigated. CD8+ and CD14+ TIICs were stimulated in the absence or presence of graded concentrations of lidocaine (0.25 mM to 1.5 mM). The levels of IFN-γ and IL-12 in the supernatant were determined at 72 hours by ELISA. Results obtained from three different donors are shown. Data are from distinct samples and are presented as the mean± SEM. *P < 0.05, **P < 0.01.
Figure 3.
The effect of lidocaine on IFN-γ secretion by CD8+ TIICs and IL-12 secretion by CD14+ TIICs. The effects of lidocaine on IFN-γ secretion from CD8+ TIICs (A) and IL-12 secretion from CD14+ TIICs (B) were investigated. CD8+ and CD14+ TIICs were stimulated in the absence or presence of graded concentrations of lidocaine (0.25 mM to 1.5 mM). The levels of IFN-γ and IL-12 in the supernatant were determined at 72 hours by ELISA. Results obtained from three different donors are shown. Data are from distinct samples and are presented as the mean± SEM. *P < 0.05, **P < 0.01.
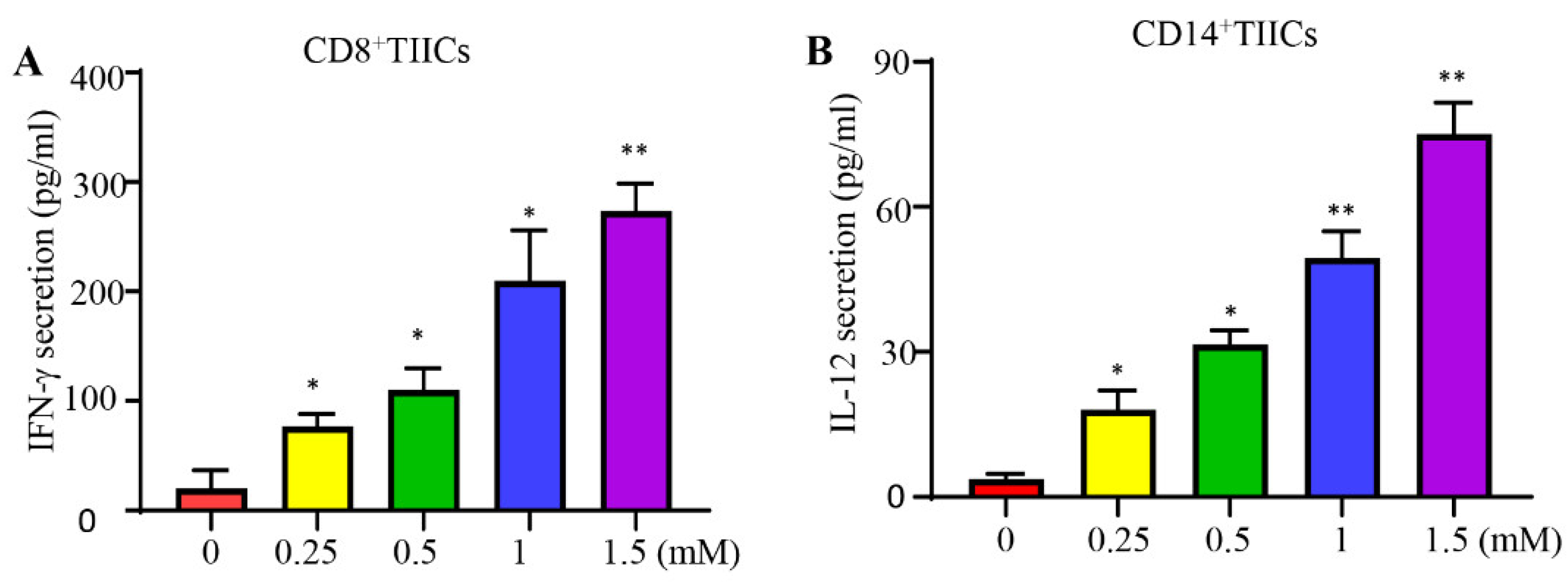
Figure 4.
The effect of lidocaine on IL-10, TGF-β, and IL-35 induced by CD4+CD25+ and CD14+ TIICs. The effect of lidocaine on IL-10 (A, B), TGF-β (C, D), and IL-35 (E, F) secretion from CD4+CD25+ and CD14+ TIICs were investigated. CD4+CD25+ and CD14+ TIICs were cultured in the absence or presence of graded concentrations of lidocaine (0.25 mM to 1.5 mM). The levels of IL-10, TGF-β, and IL-35 in the supernatant were determined at 72 hours by ELISA. Results obtained from three different donors are shown. Data are from distinct samples and are presented as the mean± SEM from three different experiments, each performed in duplicate. *P < 0.05, **P < 0.01.
Figure 4.
The effect of lidocaine on IL-10, TGF-β, and IL-35 induced by CD4+CD25+ and CD14+ TIICs. The effect of lidocaine on IL-10 (A, B), TGF-β (C, D), and IL-35 (E, F) secretion from CD4+CD25+ and CD14+ TIICs were investigated. CD4+CD25+ and CD14+ TIICs were cultured in the absence or presence of graded concentrations of lidocaine (0.25 mM to 1.5 mM). The levels of IL-10, TGF-β, and IL-35 in the supernatant were determined at 72 hours by ELISA. Results obtained from three different donors are shown. Data are from distinct samples and are presented as the mean± SEM from three different experiments, each performed in duplicate. *P < 0.05, **P < 0.01.
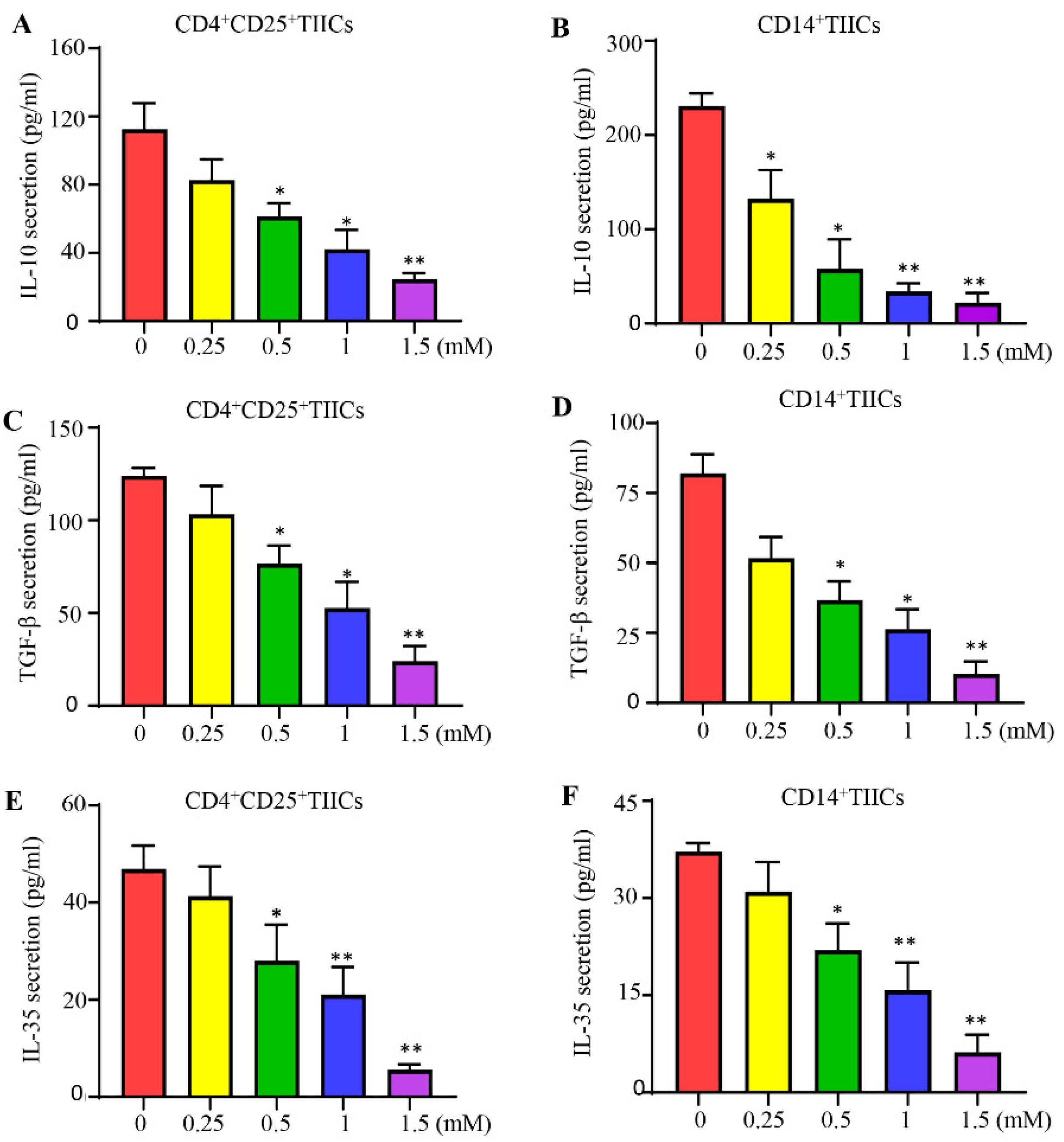
Figure 5.
Schematic diagram: lidocaine-modulated immunosuppression of normal PBMCs and anti-tumor effect of gastric TIICs. This diagram illustrates the dual effects of lidocaine on immune cells: 1. PBMCs: Demonstrates how lidocaine modulates immunosuppression in normal PBMCs, highlighting changes in cytokine secretion. 2. TIICs: Illustrates the anti-tumor effect of lidocaine on gastric TIICs, focusing on its impact on cytokine secretion.
Figure 5.
Schematic diagram: lidocaine-modulated immunosuppression of normal PBMCs and anti-tumor effect of gastric TIICs. This diagram illustrates the dual effects of lidocaine on immune cells: 1. PBMCs: Demonstrates how lidocaine modulates immunosuppression in normal PBMCs, highlighting changes in cytokine secretion. 2. TIICs: Illustrates the anti-tumor effect of lidocaine on gastric TIICs, focusing on its impact on cytokine secretion.
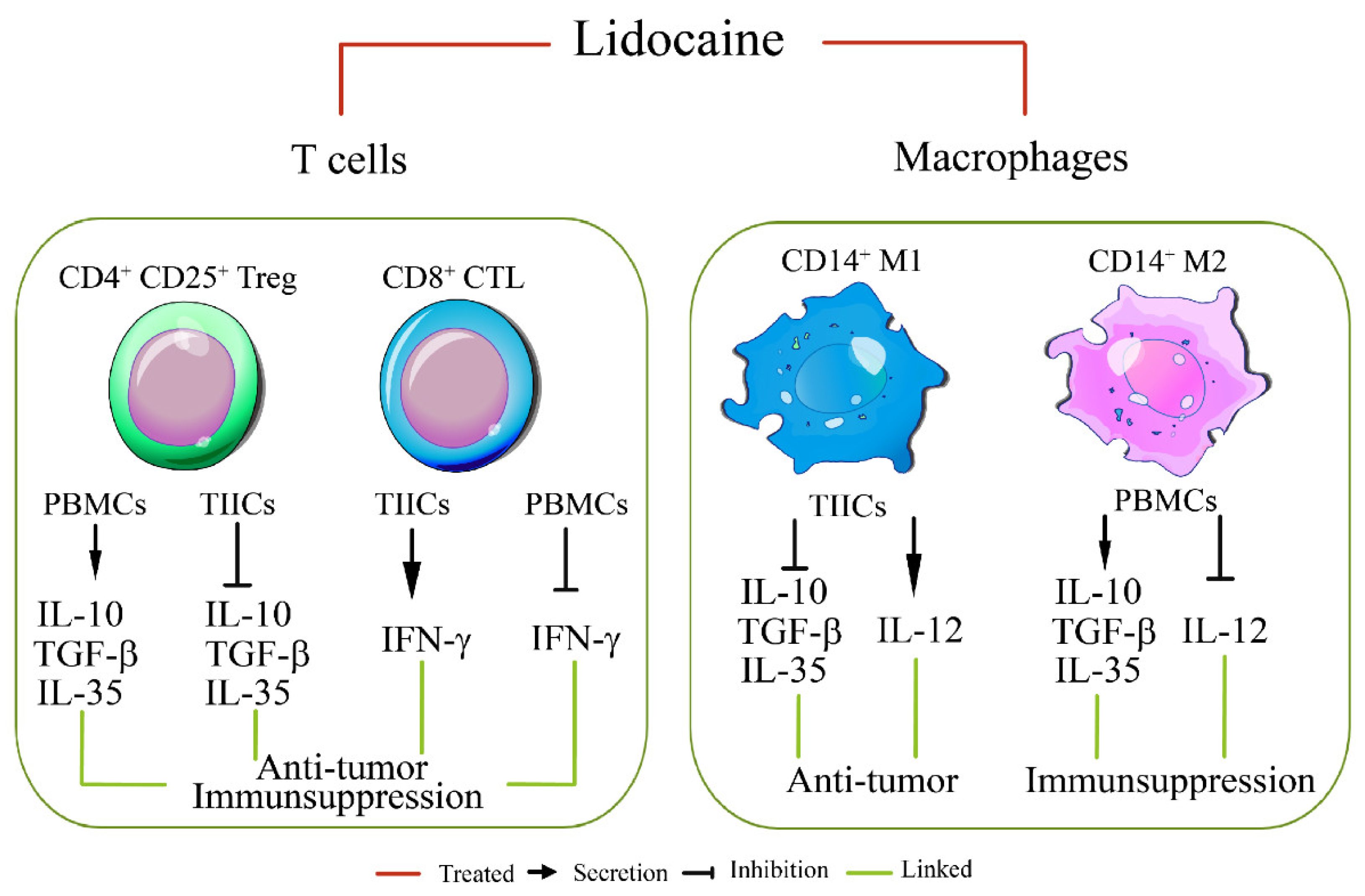
Figure 6.
Lidocaine inhibits IL-35, IL-10, and TGF-β production by CD4+CD25+Foxp3+ TIICs.Sorted CD4+CD25+TIICs were treated with 1.5 mM lidocaine for 72 hours followed by staining with anti-IL-35, anti-IL-10, anti-TGF-β, and anti-Foxp3 antibodies for flow cytometry analysis. Isotype controls were used to distinguish between positive and negative cells for IL-35, IL-10, TGF-β, and Foxp3. Typical flow cytometry dot plot analysis revealed the percentage of (A, B) CD35+Foxp3+CD4+CD25+ TIICs, (C, D) IL-10+Foxp3+CD4+CD25+ TIICs, and (E, F) TGF-β+Foxp3+CD4+CD25+ TIICs treated with lidocaine (1.5 mM). Data are from distinct samples and presented as the mean± SEM in three different experiments, each performed in duplicate. *P < 0.05, **P < 0.01.
Figure 6.
Lidocaine inhibits IL-35, IL-10, and TGF-β production by CD4+CD25+Foxp3+ TIICs.Sorted CD4+CD25+TIICs were treated with 1.5 mM lidocaine for 72 hours followed by staining with anti-IL-35, anti-IL-10, anti-TGF-β, and anti-Foxp3 antibodies for flow cytometry analysis. Isotype controls were used to distinguish between positive and negative cells for IL-35, IL-10, TGF-β, and Foxp3. Typical flow cytometry dot plot analysis revealed the percentage of (A, B) CD35+Foxp3+CD4+CD25+ TIICs, (C, D) IL-10+Foxp3+CD4+CD25+ TIICs, and (E, F) TGF-β+Foxp3+CD4+CD25+ TIICs treated with lidocaine (1.5 mM). Data are from distinct samples and presented as the mean± SEM in three different experiments, each performed in duplicate. *P < 0.05, **P < 0.01.
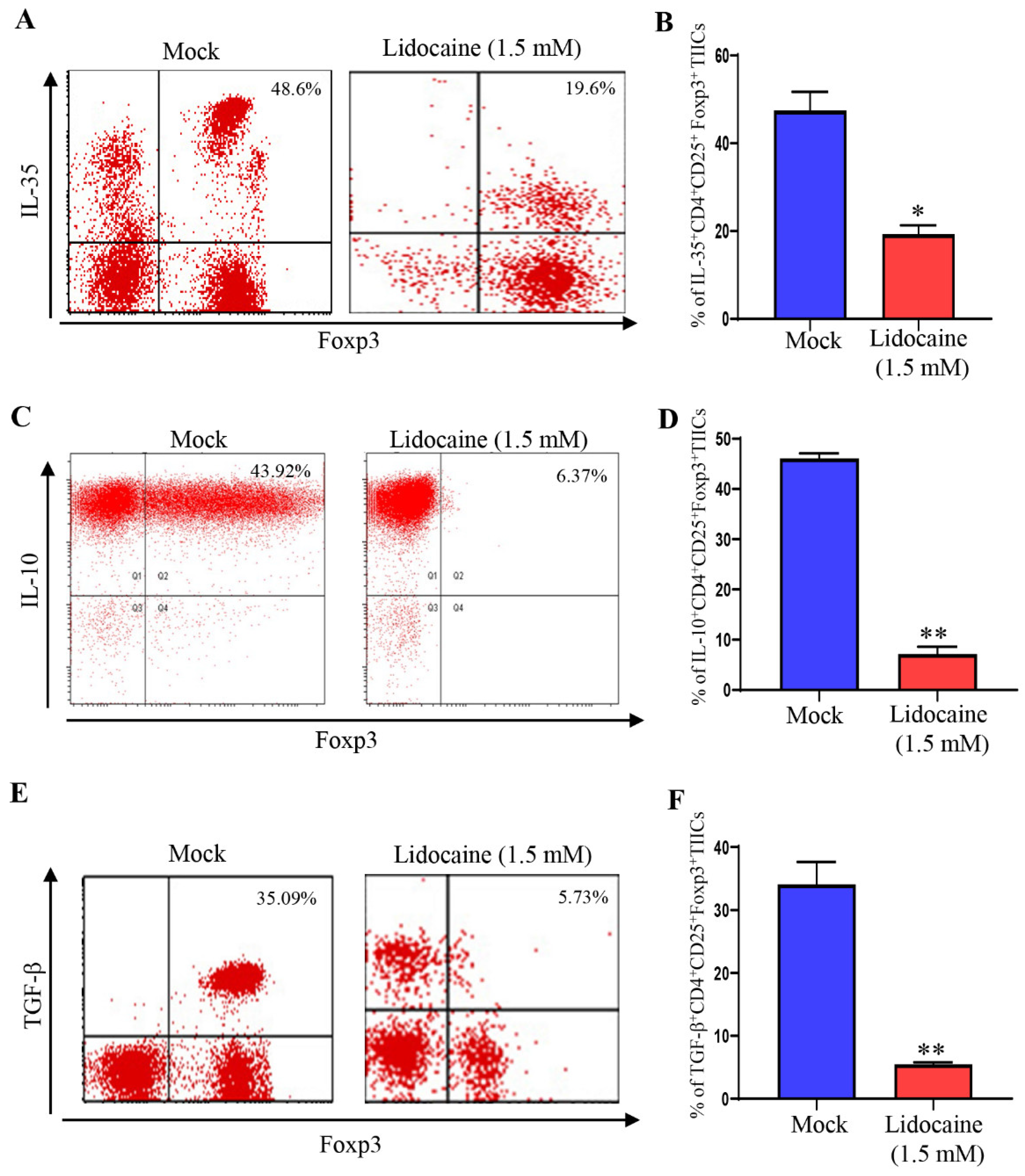
Figure 7.
Lidocaine enhances antitumor immunity by reducing PD-1 and increase IFN-γ expression on CD8+ TIICs through NF-κB signaling pathway. Gastric CD8+ TIICs treated with lidocaine (1.5 mM) were analyzed by flow cytometry. Single-cell suspensions obtained from sorted CD8+ TIICs were stained to detect IFN-γ (A, B) and PD-1 (C, D). Analysis of IFN-γ production and PD-1 expression by lidocaine treated-CD8+TIICs with NF-κB inhibitor. CD8+TIICs were incubated for 1 h with or without 10 μM BAY11-7082 and then treated with lidocaine for 72 h. IFN-γ and PD-1 was measured by flow cytometry. All flow cytometry analyses were gated on total live cells. Data are from distinct samples and presented as the mean± SEM. *p < 0.05, **p < 0.01; n ≥ 3.
Figure 7.
Lidocaine enhances antitumor immunity by reducing PD-1 and increase IFN-γ expression on CD8+ TIICs through NF-κB signaling pathway. Gastric CD8+ TIICs treated with lidocaine (1.5 mM) were analyzed by flow cytometry. Single-cell suspensions obtained from sorted CD8+ TIICs were stained to detect IFN-γ (A, B) and PD-1 (C, D). Analysis of IFN-γ production and PD-1 expression by lidocaine treated-CD8+TIICs with NF-κB inhibitor. CD8+TIICs were incubated for 1 h with or without 10 μM BAY11-7082 and then treated with lidocaine for 72 h. IFN-γ and PD-1 was measured by flow cytometry. All flow cytometry analyses were gated on total live cells. Data are from distinct samples and presented as the mean± SEM. *p < 0.05, **p < 0.01; n ≥ 3.
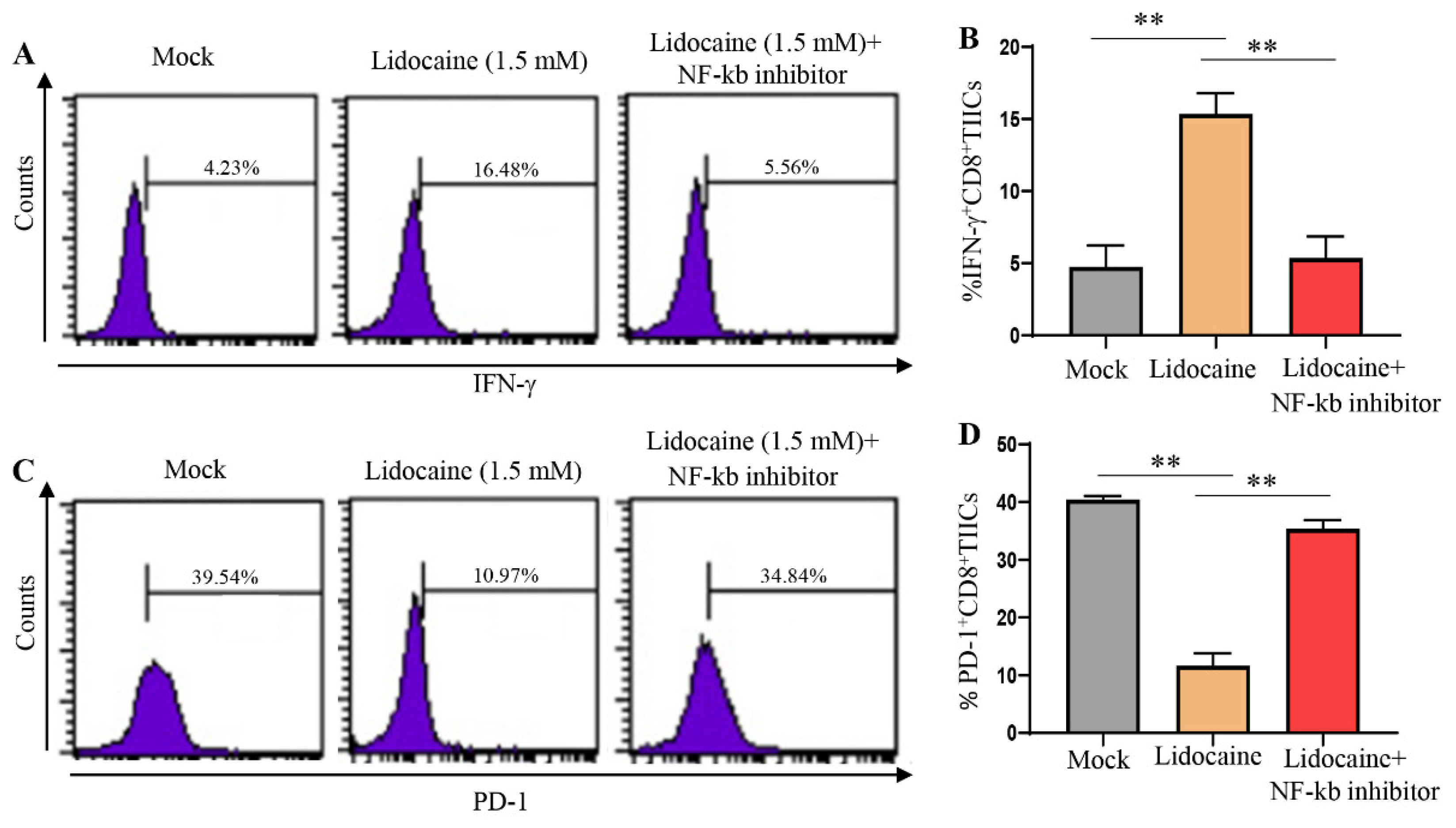
Figure 8.
Apoptotic effects of lidocaine on TIICs, PBMCs, PGCCs, and CD8+TIICs co-cultured with PGCCs assessed by flow cytometry. 5mM and 10mM lidocaine induce TIICs and PBMCs apoptosis but 0.5 and 1.5 lidocaine does not induce PGCCs apoptosis. TIICs (A), PBMCs (C) and PGCCs (E) were assessed by flow cytometric analysis using propidium iodide-stained cells. Firstly, 104 cells were incubated in 96-well plates in the presence or absence of the indicated concentrations of lidocaine. After 72 h treatment, cells were washed with PBS and fixed with 70% ethanol for 1 h on ice. Pelleted cells were incubated with RNaseA (0.1 mg/mL) and propidium iodide (40 μg/mL) for 1 h with shaking and protected from light. The percentage of subG1 population was determined by flow cytometry. (G) Flow cytometry assessment of cell death for lidocaine (1.5 mM)-treated CD8+TIICs, lidocaine (1.5 mM)-treated PGCCs or lidocaine (1.5 mM)-treated CD8+TIICs co-cultured with PGGCs. Data are representative of three independent experiments; n > = 3. (B)(D)(F)(H) Data are from distinct samples and presented as the mean± SEM in three different experiments, each performed in duplicate. *P < 0.05 **P < 0.01.
Figure 8.
Apoptotic effects of lidocaine on TIICs, PBMCs, PGCCs, and CD8+TIICs co-cultured with PGCCs assessed by flow cytometry. 5mM and 10mM lidocaine induce TIICs and PBMCs apoptosis but 0.5 and 1.5 lidocaine does not induce PGCCs apoptosis. TIICs (A), PBMCs (C) and PGCCs (E) were assessed by flow cytometric analysis using propidium iodide-stained cells. Firstly, 104 cells were incubated in 96-well plates in the presence or absence of the indicated concentrations of lidocaine. After 72 h treatment, cells were washed with PBS and fixed with 70% ethanol for 1 h on ice. Pelleted cells were incubated with RNaseA (0.1 mg/mL) and propidium iodide (40 μg/mL) for 1 h with shaking and protected from light. The percentage of subG1 population was determined by flow cytometry. (G) Flow cytometry assessment of cell death for lidocaine (1.5 mM)-treated CD8+TIICs, lidocaine (1.5 mM)-treated PGCCs or lidocaine (1.5 mM)-treated CD8+TIICs co-cultured with PGGCs. Data are representative of three independent experiments; n > = 3. (B)(D)(F)(H) Data are from distinct samples and presented as the mean± SEM in three different experiments, each performed in duplicate. *P < 0.05 **P < 0.01.
Figure 9.
Lidocaine-treated CD8+TIICs dependent immunogenic cell death of PGCCs.Lidocaine (1.5 mM)-treated CD8+TIICs, lidocaine (1.5 mM)-treated PGCCs or lidocaine (1.5 mM)-treated CD8+TIICs co-cultured with PGGCs at the 5:1 E:T ratios. Target cell cytotoxicity was determined at 2 hours by a DELFIA EuTDA assay. Data represent the mean of triplicate experiments, and experiments were repeated at least three times using different donor with similar results. Data are from distinct samples and presented as the mean± SEM. *p < 0.05, **p < 0.01 compared with mock treated cells; n > = 3.
Figure 9.
Lidocaine-treated CD8+TIICs dependent immunogenic cell death of PGCCs.Lidocaine (1.5 mM)-treated CD8+TIICs, lidocaine (1.5 mM)-treated PGCCs or lidocaine (1.5 mM)-treated CD8+TIICs co-cultured with PGGCs at the 5:1 E:T ratios. Target cell cytotoxicity was determined at 2 hours by a DELFIA EuTDA assay. Data represent the mean of triplicate experiments, and experiments were repeated at least three times using different donor with similar results. Data are from distinct samples and presented as the mean± SEM. *p < 0.05, **p < 0.01 compared with mock treated cells; n > = 3.
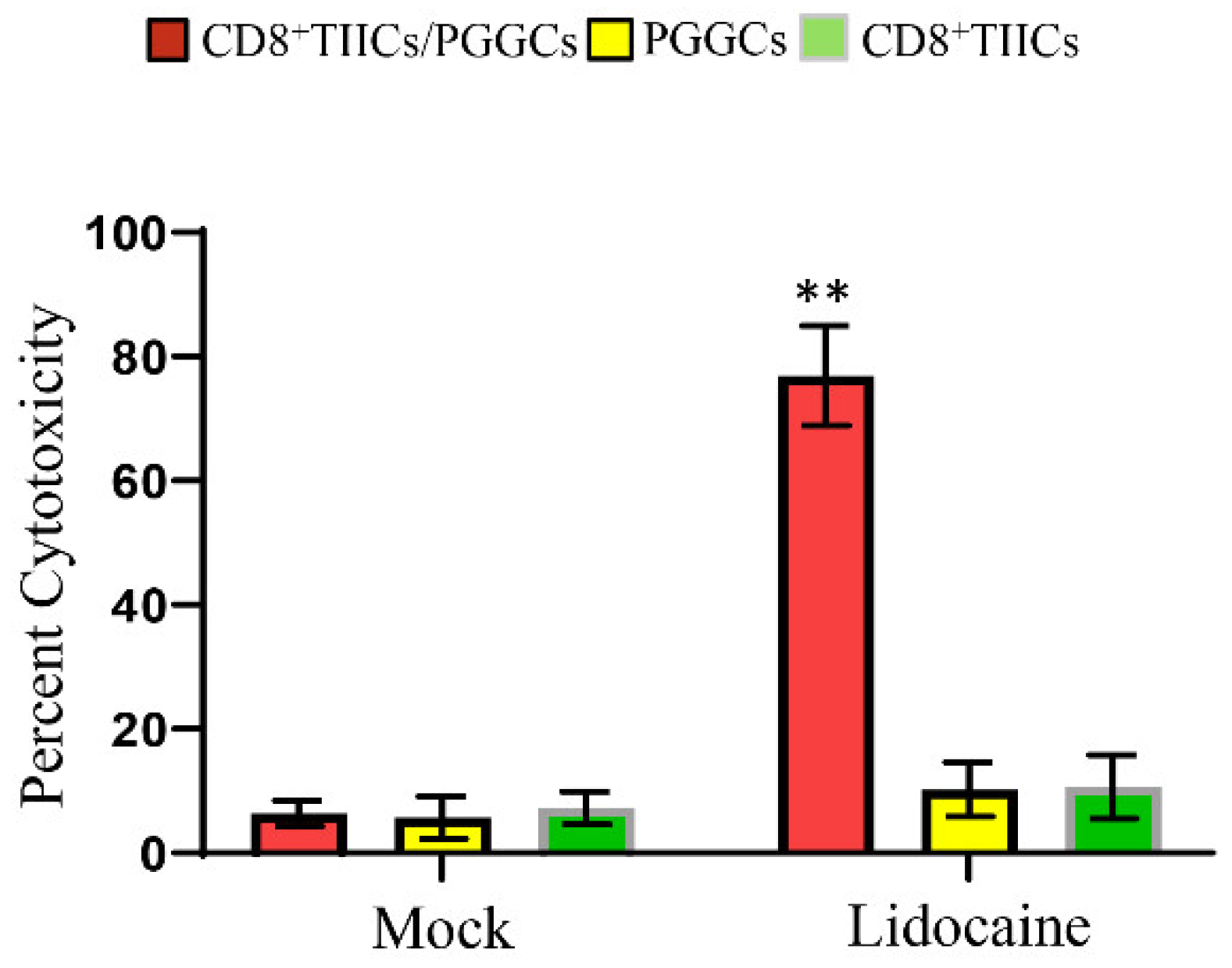
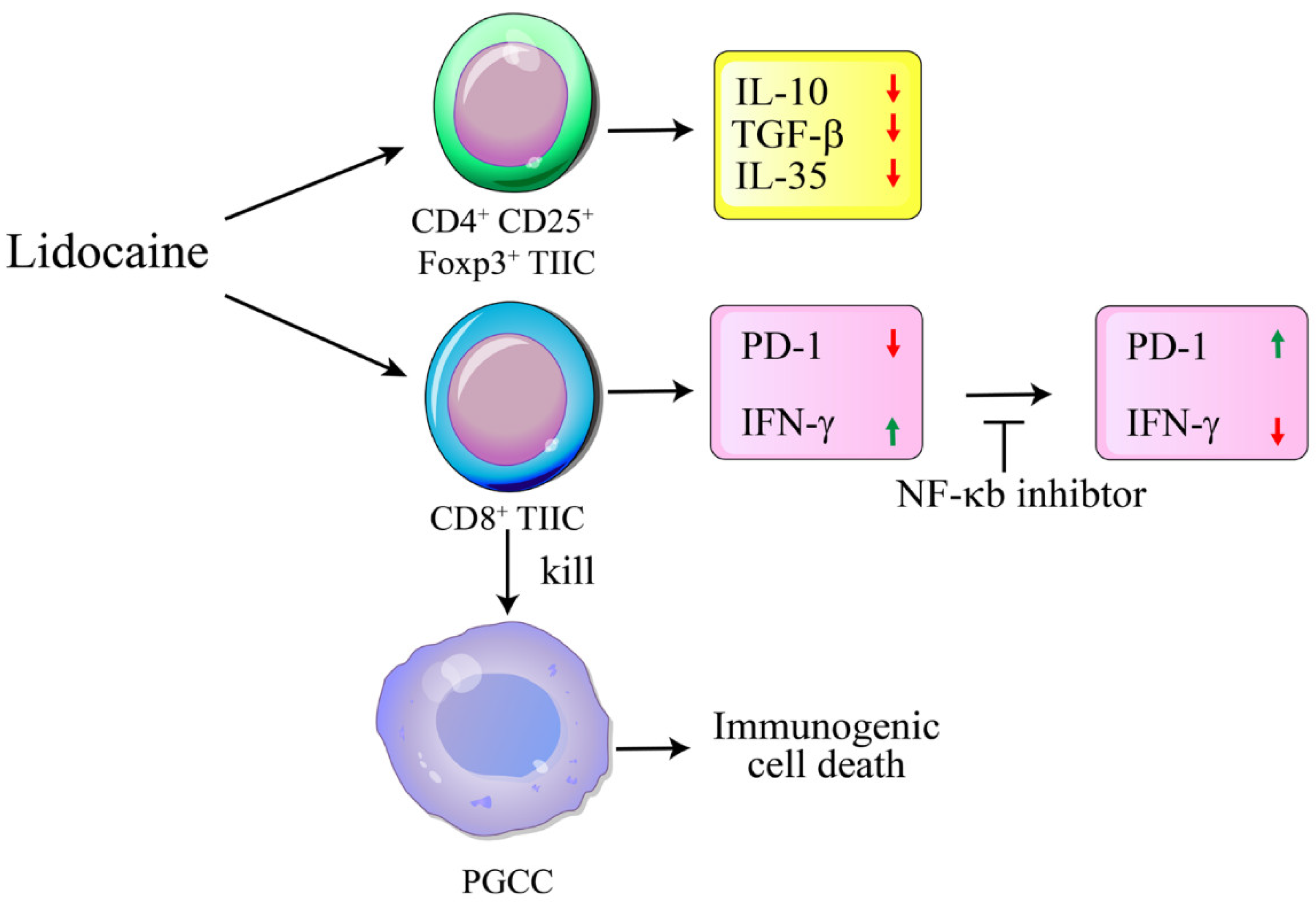
Figure 10.
Schematic diagram illustrating the lidocaine-mediated anti-tumoral mechanism through immunogenic cell death targeting PGCCs. Lidocaine inhibits the production of IL-35, IL-10, and TGF-β by CD4+CD25+Foxp3+ tumor-infiltrating immune cells (TIICs). Additionally, lidocaine enhances antitumor immunity by reducing PD-1 expression and increasing IFN-γ expression on CD8+ TIICs via the NF-κB signaling pathway. The lidocaine-treated CD8+ TIICs subsequently promote immunogenic cell death of PGCCs. Long→: Treated. Short→: Linked. ┬: Inhibition.
Figure 10.
Schematic diagram illustrating the lidocaine-mediated anti-tumoral mechanism through immunogenic cell death targeting PGCCs. Lidocaine inhibits the production of IL-35, IL-10, and TGF-β by CD4+CD25+Foxp3+ tumor-infiltrating immune cells (TIICs). Additionally, lidocaine enhances antitumor immunity by reducing PD-1 expression and increasing IFN-γ expression on CD8+ TIICs via the NF-κB signaling pathway. The lidocaine-treated CD8+ TIICs subsequently promote immunogenic cell death of PGCCs. Long→: Treated. Short→: Linked. ┬: Inhibition.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



