
Submitted:
14 October 2024
Posted:
15 October 2024
You are already at the latest version
Abstract
Bacterial keratitis caused by Pseudomonas aeruginosa is indeed a serious concern due to its potential to cause blindness and its resistance to antibiotics, partly attributed to biofilm formation and cytotoxicity to the cornea. The present study uses meta-analysis of transcriptomics dataset to identify important genes and pathways in biofilm formation of P. aeruginosa induced keratitis. By combining data from several studies, meta-analysis can enhance statistical power and robustness, enabling the identification of 83 differentially expressed candidate genes including fis that could serve as therapeutic targets. The approach of combining meta-analysis with virtual screening and in vitro methods provides a comprehensive strategy for identifying potential target genes and pathway crucial for bacterial biofilm formation and development anti-biofilm medications against P. aeruginosa infections. The study identified 83 candidate genes that exhibited differential expression in the biofilm state, with fis proposed as an ideal target for therapy for P. aeruginosa biofilm formation. These techniques, meta-analysis, virtual screening, and in-vitro methods were used in combination to diagnostically identify these genes which play a significant role in biofilms. This finding has highlighted a hallmark target list for P. aeruginosa anti-biofilm potential treatments.
Keywords:
Bacterial keratitis
; Biofilm
; fis
; genes
; meta-transcriptome analysis
; Pseudomonas aeruginosa
1. Introduction
A dangerous and potentially blinding corneal infection, the transparent anterior layer of the eye that encompasses the iris and pupil, is called bacterial keratitis. Bacteria enter the cornea and cause infection, which results in inflammation and damage. The adaptable opportunistic pathogen Pseudomonas aeruginosa may result in numerous problems, including corneal infections. There are numerous virulence factors in P. aeruginosa that aid in its establishment and infliction of harm in the host tissue when it comes to corneal infections. The production of biofilms and cytotoxicity to cornea have been directly connected to P. aeruginosa's enhanced resistance to antibiotics [1].
P. aeruginosa strain PAO1 is a well-known model organism for studying biofilm development and related activities. Because PAO1 is well-known for its capacity to build durable biofilms, it is an invaluable tool for comprehending the physiological and molecular components of biofilm formation [2]. Attachment and early colonization are two important characteristics of PAO1 biofilms [3]. To begin the process of forming a biofilm, PAO1 uses pili and specific adhesins to adhere to surfaces. Such factors as flagella and type IV pili mediate initial adhesion [4]. Extracellular Polymeric Substances (EPS) components, such as pellicle polysaccharide (Pel), alginate, and polysaccharide synthesis locus (Psl) are produced by PAO1 and are essential for the stability and structure of biofilms. Antimicrobial drugs and environmental pressures are protected from by EPS [5]. PAO1 controls gene epression linked to biofilm development and coordinates the formation of biofilms by means of Quorum Sensing (QS) systems, namely the Las, PQS, and Rhl systems. In addition, QS regulates the synthesis of EPS components and virulence factors [6]. PAO1 biofilms have a complex architecture that includes microcolonies, water channels, and blank spaces [7]. PAO1 biofilm formation is controlled by sophisticated regulatory networks involving various transcriptional regulators (e.g., FleQ and AlgU), cyclic-di-GMP signaling, and two-component systems (e.g., GacS/GacA) [8].
Biofilm formation and gene expression are influenced by variables such as nutrient supply, oxygen levels, and surface characteristics. During biofilm formation, metabolic changes take place that result in changed gene expression and the creation of EPS [9]. The rate of metabolism of PAO1 biofilms is lesser than that of planktonic cells. Because of resistant cells and decreased antimicrobial agent penetration, PAO1 biofilms exhibit enhanced tolerance and resistance to antibiotics [10]. In clinical settings, biofilm-associated antibiotic resistance poses a serious problem. PAO1 biofilms have been linked to persistent infections linked to wounds, medical device-related illnesses, and cystic fibrosis [11,12]. The discovery of innovative treatments targeting biofilm-specific mechanisms such as EPS production and QS is aided by an understanding of PAO1 biofilm biology. In order to increase the effectiveness of antibiotics, research is concentrated on breaking down biofilms using antimicrobial peptides and nanoparticles [13,14,15]. P. aeruginosa PAO1 biofilms have a complex architecture, produce EPS, and use quorum sensing and metabolic adjustments to regulate themselves. Studying PAO1 biofilms provides vital insights into biofilm biology and antibiotic resistance mechanisms. This has implications for creating effective tactics to combat diseases linked to biofilms [16].
Distinctions between planktonic and biofilm bacteria is the main justification for the creation of anti-infective methods against biofilms [17]. Numerous approaches, from molecular ones like Northern blotting and PCR [18,19,20] to high-throughput ones like proteomics and transcriptomics, are used in research to find key biofilm genes [21,22,23]. However, because of the variations in the methods employed and the strain dependence of the molecular components, the collection of discovered genes found in every study varies and exhibits minimal overlap between one another [24]. Many biological issues, including the pathogenicity of bacteria like P. aeruginosa, can be studied by analyzing gene expression changes and examining the underlying mechanisms using datasets from transcriptomic technologies like RNA-seq and microarray [25]. Better, more trustworthy results are obtained from extensive expression profile analyses that include greater quantity of samples, yet organizing such an experiment is not always possible. A meta-analysis approach is used in these situations, which includes the datasets and findings from numerous earlier investigations. This method removes inconsistencies, lifts the sample size limit, and finds genes that are consistently changed in different research, all while increasing statistical power [24].
A meta-analysis of transcriptome data related to P. aeruginosa biofilms entails collecting and analyzing various research to uncover similar trends and key genes/pathways involved in biofilm formation. utilizing me-ta-analysis and feature selection, the current study aims to identify a signature collection of putative genes that distinguish significantly in P. aeruginosa biofilms utilizing transcriptome data obtained from various investigations. To discover differential expression, random-effects modelling was applied to public gene expression datasets of P. aeruginosa in planktonic and biofilm settings. Additional feature selection techniques were used to identify potential genes involved in biofilm development, which were confirmed using incredibly accurate classifiers. In addition, based on the data from meta-analysis, virtual screening and invitro approaches were applied to identify a P. aeruginosa biofilm inhibitor.
2. Materials and Methods
2.1. Chemicals, Microbial Strains and Culture Conditions
The Pseudomonas aeruginosa (MTCC1688) was acquired from the Institute of Microbial Technology in Chandigarh, India. It was cultivated on Nutrient Agar Medium (NA; cat. no. MM012; Himedia, India) and kept at 4 °C. Brain Heart Infusion (BHI) broth was inoculated with a single colony of P. aeruginosa. (Cat No. M210 Himedia, India) and then the suspensions were incubated for an entire night in a shaker at 200 rpm and 37 °C. Himedia, India supplied the dexamethasone (at No. C80687; 99% purity). All chemicals and media utilised in the current investigation, unless otherwise noted, were purchased from Himedia (India) and Merck (India).
2.2. Dataset Collection
Using keywords, the National Centre for Biotechnology Information's public expression profile database Gene Expression Omnibus (GEO) was searched for expression records of P. aeruginosa planktonic and matching biofilm forms. Platform variations and sample preparations are mostly responsible for significant variations observed in comparable experiments that distinguish between two conditions. As a result, only samples from the same studies and platform that met both test and control conditions were retrieved. Four data sets, derived from two distinct methodologies (microarray and RNA sequencing), were recognised (Table 1).
2.3. Data Processing and DEG Screening
In order to perform the meta-analysis and identify shared DEGs within the expression profiles, Network Analyst, an online tool for integrative meta-analysis, was used [25]. The ComBat approach was used to remove the batch effects after the normalised expression profiles in the required format were uploaded. The random effect size in the meta-analysis was selected in accordance with the outcomes of the Cochran's Q-test, which estimates statistical heterogeneity [26,27]. The DESeq2 tool in the R programming was used to perform differential expression analysis of the datasets; metrics such as p-values, P-Value adjusted, and Log2 Fold Change (Log2FC) were analysed for each gene.
2.4. Gene Ontology and Pathway Analysis
The annotation, visualization, and integrated discovery (DAVID) online database was utilized to analyse the resulting meta-DEGs for gene ontology (GO), which covers biological process, cellular component, and molecular function as well as pathway enrichment [28,29]. DAVID analysis yielded the functional annotation clustering and Kyoto encyclopaedia of genes and genomes (KEGG) pathways. Biological process-related clusters were combined when the enrichment score (ES) threshold was set at 0.5. The enriched GO, pathway were visualization using Hiplot (ORG) [30].
2.5. Virtual Screening for Identification of Drug Candidate
The target proteins' (3-D) experimentally determined structures (fis; PDB id: 6m10) were obtained from the Protein Data Bank (https://www.rcsb.pdb.org/) [31]. UCSF Chimaera v1.16's conjugate gradient and steepest descent algorithms were employed to minimise the energies of fis protein following the addition of polar hydrogen atoms and other missing atoms, the removal of heteroatoms and co-crystallized water, and the assignment of partial charge [32]. The produced receptor was uploaded to the DrugRep webserver to perform receptor-based virtual screening [33]. Pocket 1 was chosen to be docked (centre coordinates: -17.0, -8.2, and 9.4, size values: 23, 19, and 16). The library chosen for virtual screening was Approved Drug Library. The selected, ligand (dexamethasone) was reanalysed for its interaction with fis using Autodock tool. To elucidate the pattern of interactions between the fis and dexamethasone, Discovery Studio Visualizer 2022 was used27. The fis in complex with dexamethasone was examined in terms of their binding affinity, conformation, and interactions (docking score). Using the CABS-Flex 2.0 system, which relies on coarse-grained protein movements, Molecular dynamics (MD) simulations of complex and the native protein (fis) were carried out [34]. There were other distance constraints, such as a global weight of 1.0, in addition to more than 50 cycles and 50 trajectory frames in 10 ns each. The complexes' mobility was represented in terms of root-mean-square fluctuations (RMSF).
2.6. Antibacterial Activity of Dexamethasone against P. aeruginosa
Using a well-diffusion approach, the antimicrobial effect of dexamethasone against P. aeruginosa was analysed [35,36]. Nutrient Agar (Himedia, India) was used to cultivate P. aeruginosa for a duration of 24 hours at 37 °C. A 0.85% NaCl (w/v) sterile saline solution was used to provide inoculations of bacteria equivalent to 0.5 McFarland (1 X 108 CFU/mL). The aforementioned suspension was distributed on Mueller-Hinton agar (MHA) plates. A 6 mm diameter sterile dicsc was placed on the agar medium, and loaded with dexamethasone (0.5, 1, 2, and 4 mg/disc). As positive control, disc containing 20 µg ciprofloxacin was placed on the plates and incubated at 37°C for 24 hours. The zone of inhibition (mm in diameter) was used to measure the activity against P. aeruginosa.
2.7. Minimal Inhibitory Concentration (MIC)
Planktonic suspended cells were used to estimate the least inhibitory concentrations (MICs) of P. aeruginosa using broth micro-dilution on microtitre plates [37]. The plates were made using aseptic methods. After pipetting 100 µL of each chemical, dexamethasone (100 mg/ml) in DMSO into each well, the two substances were serially diluted to their lowest concentration 0.625 mg/ml and 1.25 µg/mL, respectively. Following that, 10 µL of bacterial suspension (1 X 106 CFU/mL) was added to the wells, and the mixture was kept at 37 °C for 24 hours. Once incubated, 30 µL of resazurin (0.015%) was added to each well, and the wells were incubated for a further two to four hours to observe any colour changes. After measuring plate absorbance at 570 nm, the quantity of dexamethasone/ciprofloxacin that suppressed cell growth was named as the minimum inhibitory concentration (MIC).
2.8. Inhibition Assay in Biofilm Formation
The ¼ MIC of dexamethasone was treated with suspensions of bacteria in BHI broth enriched with 1% (w/v) sucrose (1 × 107 CFU/mL) for 48 hours at 37°C in a 96-well cell culture plate with a flat bottom. The addition of sucrose aided in the development of bacterial biofilms [38]. In addition to the dexamethasone treatment, appropriate culture and media controls were included. Any residual cells that weren't adherent to the biofilm were then eliminated by rinsing with sterile PBS after the medium and the supernatant carrying dispersed planktonic cells were removed. The potential of dexamethasone to suppress biomass and metabolism on P. aeruginosa biofilm was ascertained through the use of the XTT assay and crystal violet staining. For the XTT experiment, 100 µL of fresh, sterile medium was added to each well. Subsequently, 50 µL of a detection solution containing XTT (2,3-Bis-(2-Methoxy-4-Nitro-5-Sulfophenyl)-2H-Tetrazolium-5-Carboxanilide) and phenazine methosulfate (PMS) was introduced into each well, and it was incubated for four hours at 37°C. The absorbance at 450 nm was then measured after the plates were rapidly agitated for 10 seconds to mix the dye with the solution [39]. In order to evaluate the dexamethasone's capacity to inhibit P. aeruginosa biofilm biomass, 100 µL of 0.1% w/v crystal violet solution was introduced into each well. After that, the wells were allowed to incubate for ten minutes. Each well was washed using sterile distilled water after the unattached crystal violet solution was removed. Next, to measure the absorbance of colour intensity at 595 nm, Each well received 95% ethanol, and the plate was shaken gently for 30 minutes at room temperature [40]. The following formula was used to determine the percentage of biofilm inhibition throughout the triplicate experiment:
Biofilm inhibition (%) = 100 − ((OD570 in sample treatment - OD570 in untreated bacteria)/(OD570 in untreated sample - OD570 in untreated bacteria) × 100)
2.9. Fluorescent Microscopic Analysis of Biofilm
The fluorescence microscopic studies of dexamethasone impacts on P. aeruginosa biofilms were performed out as previously described, with a few minor variations [41,42]. In short, P. aeruginosa biofilms were formed on 12 well culture plates for 72 hours at 37 °C using varied dexamethasone concentrations (range from 0.25 to 4 mg/ml) in BHI medium enriched with 2% (w/v) glucose. The wells were delicately washed with antiseptic phosphate buffered saline (PBS) before staining with acridine orange for 30 minutes. A fluorescent microscope (Optika, Germany) was utilised to record five arbitrary images, with the intensity of light, the background, and contrast remaining constant during the experiment.
2.10. Statistical Analysis
Each experiment was run three times in triplicate. Graphpad Prism 10.2 software was used to estimate the intergroup differences using one way ANOVA and T-test. The mean and standard error mean (SEM) of the data is displayed. A p-value was considered statistically significant if it was less than 0.05.
3. Results
3.1. Dataset Selection and Quality Control
Initially, 4 expression profiling by high throughput RNA sequencing and 2 expression profiling by microarray, from biofilm and planktonic samples were retrieved from GEO database. However, based on the initial Principal component analysis (PCA) 2 high throughput RNA sequencing datasets were removed from further analysis. It was determined that the four profiles with untreated planktonic and biofilm were appropriate for additional pre-analysis stages. The GEO accessions GSE30021, GSE120760, GSE136111, and GSE223663 corresponded to the chosen expression profiles. Table 1 contains comprehensive details about the chosen datasets. For the meta-analysis, 12 biofilm and 12 planktonic samples were chosen using PCA, outlier removal, normalisation, and batch effect removal procedures (Figure 1 A and B).
3.2. Meta-Analysis and DEG Identification
It was successful to apply meta-analysis (random effect size) to the expression profiles' pre-processed matrix files. Based on the findings of the Cochran's Q-test and the literature review, the random effect size was chosen as the meta-analysis approach. Meta-analysis using random effect size revealed 530 genes were commonly expressed in all 4 expression profiles. Using an adjusted p-value of 0.05, 83 genes that includes 45 upregulated and 38 downregulated genes were determined to be meta-DEGs (Table 2). Supplementary file 1 (S1) has the entire list of the DEGs that have been identified. Heatmap of top 50 meta-DEGs are shown Figure 2A and the volcano plot of DEGs were shown in figure 2B. The top 50 meta-DEGs sorted by their effect sizes (fold changes) are listed in Table 3 and Table 4.
3.3. Functional Classification of DEGs Using Gene Ontology and KEGG Pathways Analysis
DAVID tools version 6.8 was used to perform functional annotation clustering in order to assess the genes that are expressed differently in P. aeruginosa's biofilm and planktonic forms. Gene ontology (GO) analysis divides the three categories of differentially expressed genes into biological processes, molecular function, and cellular components.
Based on the enrichment score (ES), functional annotation clustering created a total of 4 clusters for up-regulated and 2 clusters for down-regulated genes from the 83 DEGs of Biofilm. In case of up-regulated genes, the most enriched clusters consisted of genes associated with translation (ES=3.89) and secondary metabolite biosynthetic process (ES=2.40). Further functional enrichment analysis of DEGs for up-regulated in biofilm samples revealed enrichment of molecular function (ATP binding, structural constituent of ribosome, rRNA and tRNA binding), Biological Process (translation, secondary metabolite biosynthetic process, DNA replication and ATP synthesis coupled proton transport), cellular components (cytosol, large ribosomal subunit and primosome complex) and pathway (ribosome, phenazine biosynthesis, quorum sensing, biofilm formation and oxidative phosphorylation) (Figure 3 A-D). While the functional enrichment analysis of DEGs for down-regulated in biofilm samples revealed a single cluster (ES=1.35) enrichment with heme biosynthetic biological process and porphyrin metabolism pathway (Figure 4).
The study conducted by DAVID yielded pathways from the Kyoto Encyclopaedia of Genes and Genomes (KEGG). The KEGG Pathways ribosome, phenazine biosynthesis, quorum sensing, and biofilm formation were shown to be significant in genes which are up regulated in biofilm (Figure 3D).
Figure 5 displays the genes participate in the ribosomal protein synthesis and translation that includes 5 subunits of 50S ribosome (L3, L9, L28, L32 and L33) and 3 subunits of 30S ribosomes (S3, S4 and S12) are up-regulated in biofilm. While the pathways phenazine biosynthesis, quorum sensing and biofilm formation shared common genes that included PqsA, PqsB, PqsC, PqsD, PhzA and PhzB (Figure 6).
Since, the pathway associated with fis (PA4853) has not yet reported in P. aeruginosa, adaptation of biofilm pathway from Vibrio cholera revealed the up-regulated fis gene is an important contributor of biofilm formation in P. aeruginosa (Figure 7).
Another group of up-regulated genes in biofilm are involved in biosynthesis pyochelin (pchC, pchE and pchG). Whereas the genes participate in pyoverdin biosynthetic pathway that included pvdA and pvdH were down-regulated in biofilm of P. aeruginosa.
3.4. Structure-Based Virtual Screening
Molecular docking scores of approved drug library shown a high affinity for binding of many approved drugs toward fis protein of P. aeruginosa (Supplementary table). Dexamethasone was one among the best potential approved drug that is able to prevent the enzyme from functioning at its active site (docking score ﹣7.9 kcal/mol). Concerning number of hydrogen bond donors/aceptor, dexamethasone is better than other drug molecules. Further analysis of the fis docked with dexamethasone revealed 3 hydrogen bonds between receptor (Try 101, Gln 100 and Lys97) and the ligand (Figure 8). When fis was simulated using MD both alone and in combination with the optimal ligand (dexamethasone), the dexamethasone-fis complex showed a comparatively high RMSF pattern in comparison to the native enzyme, especially at residues 81–102 (Figure 8). Since there were no discernible changes in the mobility of the docked and native proteins, this may be explained by the complex's great stability.
3.5. Antibacterial Activity of Dexamethasone against P. aeruginosa
Dexamethasone does not appear to have any antibacterial activity against P. aeruginosa at any of the tested doses, including the maximum concentration of 8 mg/well, according to the results of this investigation (Figure 9A, B). Furthermore, dexamethasone lacks antibacterial efficacy against P. aeruginosa, as demonstrated by the minimum inhibitory concentration (MIC) analysis performed with resazurin dye reduction assay at the highest test concentration (20 mg/ml) (data not shown).
3.6. Metabolic and Biomass Inhibitory Potential of Dexamethasone against P. aeruginosa Biofilm
The results showed that dexamethasone suppressed P. aeruginosa biofilm development in a dose-dependent pattern. By using the crystal violet staining method, it was possible to see that even at a dosage of 1 mg/ml, dexamethasone could stop P. aeruginosa biofilms from growing. Dexamethasone was shown to completely prevent the formation of P. aeruginosa biofilms at a dosage of 4 mg/ml (Figure 10A). The findings from the crystal violet staining were in agreement with the outcomes of the XTT reduction experiment. Dexamethasone-induced biofilm inhibition was shown to follow a similar pattern in the XTT reduction assay (Figure 10B). According to these results, dexamethasone may be able to prevent P. aeruginosa biofilms from growing in a dose-dependent way (Figure 10C). The XTT reduction experiment and the crystal violet staining method both offer proof that dexamethasone inhibits the growth of biofilms.
4. Discussion
The biology of P. aeruginosa's ability to produce biofilms has been studied in great detail, although much remains unknown about this disease. Since biofilm structures are inherently more resistant to antibiotics, conventional treatment methods typically employed for bacterial infections are largely useless against them. Additionally complicating matters, such tactics may lead to the appearance of subpopulations that are resistant to antibiotic agents [43]. The fact that the molecular mechanism behind biofilm formation is so diverse, or the inadequate methodology used in earlier research may be responsible for this knowledge gap. In addition to providing useful methods for understanding the underlying biological mechanisms, systems biology can be used to identify new treatment targets for the management and avoidance of P. aeruginosa biofilm formation. Transcriptomics dataset meta-analysis is an effective technique that can yield more consistent findings in selected criteria. Through the integration of expression data obtained from separate research, meta-analysis has the potential to improve a study's statistical power and robustness [44]. The uniformity of meta-analysis study outcomes has rendered them appropriate for forecasting more dependable therapeutic targets and identifying more precise biofilm-associated pathways. Several transcriptomics investigations have been carried out thus far to distinguish the molecular mechanism connected to P. aeruginosa biofilm development and planktonic culture. Nevertheless, as far as we are aware, no meta-analysis investigation involving P. aeruginosa transcriptomics datasets has been done on planktonic culture or biofilm forms. Integrating different sample types (treatment with antibiotics/other chemicals and gene knockout) could increase heterogeneity and have an impact on the final outcome. Hence, in order to understand the specific transcriptomic changes during biofilm formation, the gene-expression profiles associated with antibiotics/other chemicals treatments and gene knockout datasets were left out in order to produce more uniform data. It is commonly discovered that biomarker studies obtained from a single experiment are less accurate due to their small sample sizes and low statistical power [24,45,46]. Therefore, to address these limitations by integrating the information and findings of several studies, a meta-analysis of related but separate investigations is carried out.
In order to compare the differences between P. aeruginosa's planktonic and biofilm development forms from several public expression profiles, the current work employs meta-analysis. The variations in the organism strains and platforms that were employed were corrected using orthology mapping and random-effects modelling, respectively. A meta-analysis of differential expression revealed 83 genes using a mean effect size with adjusted p-value of 0.05. The mean effect size is regarded as the differential expression calculation's equivalent of the log2 fold-change [24].
A significant number of the potential genes that were discovered were shown to serve well-established roles in the production and growth of biofilms, supporting the study's conclusions. The top ranked up-regulated genes candidates 50S ribosomal protein L28 (PA5316; rpmB), rod shape-determining protein MreC (PA4480; mreC) and translation initiation factor IF-2 (PA4744; infB) that was previously implicated in resistance to tobramycin and biofilm formation in P. aeruginosa [47]. In addition, similar to the present study, a gene encoding 30S ribosomal protein S3 (PA4257; rpsC) was reported to be up-regulated in biofilms of E.coli [48]. In addition, genes encoding ribosomal proteins rpsL, rpmG, rplI and rplC were noticed to be up-regulated in biofilms of P. aeruginosa. The gene rpsL that encodes 30S ribosomal protein S12 was earlier reported to be upregulated in biofilms of Staphylococcus aureus [49]. Genes rpsC (50S ribosomal protein L3) and rplI (50S ribosomal protein L9) were also reported to be up-regulated in biofilms of Haemophilus influenza [50]. Gene expression related to ribosome activity and protein synthesis is connected with the first attachment phase and is involved in the creation of peptidoglycan, surface-associated proteins, and capsular polysaccharide/adhesion [51].
In order to defend itself during cyanogenesis, P. aeruginosa expresses a cyanide-insensitive terminal oxidase. Cyanide generation is toxic to nearby species. The cyanide insensitive terminal oxidase (cioB) was Overexpression of cioB prevented cyanide induced dispersal of P. aeruginosa biofilms [52]. It has been exposed that P. aeruginosa biofilms and Rhl, Las, and Pqs genes involved in QS pathways are repressed by inhibition of the uracil biosynthesis pathway [53]. The present meta-analysis of transcriptomic data revealed up-regulation of uridylate kinase encoding gene (pyrH). An earlier investigation on P. aeruginosa AES-1M demonstrated increased expression of genes connected to alginate, biofilm, persistence, and virulence, including dihydroorotase, uridylate kinase, and cardiolipin synthase [54]. Expression of pyrH was reported to up-regulated in Citric Acid insensitive biofilms of TctD-TctE deleted P. aeruginosa [55]. Present study revealed up-regulation of pchE and pchH involved in Pyochelin synthesis in biofilms of P. aeruginosa. Recent investigations conducted in vitro reveal that DNase treatment downregulated the expression of pchE and can limit the production of biofilms by P. aeruginosa and S. aureus [56]. The pchE mutants were also unable to form biofilms and produce phenazines [57]. Peptidyl-tRNA hydrolase's gene was up-regulated as a result of the suhB mutation. In the meanwhile, compared to the wild-type P. aeruginosa strain, the suhB mutant did in fact develop biofilm at higher rates [58,59]. The above studies support the data the was reported in the present study, in which, the expression of gene pth (peptidyl-tRNA hydrolase) was up-regulated in P. aeruginosa biofilm. Moreover, P. aeruginosa mutants containing transposon insertions in the tRNA pseudouridine 55 synthase (truB) gene had poor biofilms made up of tiny aggregates that had a lower biomass [60]. A nucleoid binding protein called factor for inversion stimulation (Fis) attaches to the target gene promoter to influence gene expression widely and its overexpression in P. putida revealed fis as an enhancer of biofilm formation and suppressor of dispersion of biofilm [61]. The results of this study showed that P. aeruginosa biofilms expressed higher levels of Fis. The Fis was reported to regulate the type III secretion system (T3SS) in P. aeruginosa and ciprofloxacin resistance in P. aeruginosa via regulation on pyocin production [62].
Type 1 fimbriae regulation and movability were critical at all stages, whereas matrix synthesis and purine biosynthesis were crucial merely as the biofilm developed. Both mobility and adherence were also critical for the early stages of the biofilm [63]. Up-regulation of purine biosynthesis gene purH in biofilms was found to be vital and essential in the mature biofilm of P. aeruginosa. Disruption of de novo purine biosynthesis by mutation of purH was reported to impair biofilm production in S. aureus and Enterococcus faecalis [64]. The aspartate kinase (AK) gene lysC, which is responsible for aspartic acid phosphorylation, the initial stage of the aspartic amino-acid family's biosynthesis, lysine, methionine, and threonine, was also found to be up-regulated in P. aeruginosa biofilms. Although the role of lysC with respect to biofilm formation is unknown, its mutation was predicted and reported to impair biofilm formation in P. aeruginosa and Vibrio cholera [65,66].
The sodium: solute symporter (PA3234; yjcG) that was top ranked down-regulated genes candidates previously reported to be down-regulated in P. aeruginosa biofilm formation [47]. Kojic acid treatment up-regulated beta-alanine-pyruvate transaminase gene expression and inhibited biofilms formation in Acinetobacter baumannii [67]. Next to yjcG, the gene encoding Beta-alanine: pyruvate transaminase (bauA) was the most down regulated gene in biofilms of P. aeruginosa. The downregulation of the gene dnaB, which codes for a DNA helicase that can unwind long sections of double-stranded DNA, was also observed in P. aeruginosa biofilms. Earlier study, revealed down-regulation of dnaB in biofilms of Streptococcus pneumonia [68]. Similar to the present report, expression of niRN and NirH was reported to be down-regulated in biofilms of P. aeruginosa [69]. Throughout the biofilm-forming process, P. aeruginosa employs iron as a signal. The two most well-studied P. aeruginosa iron acquisition systems are the lower affinity pyochelin system and the high affinity pyoverdine system. Extra cellular iron (Fe3+), which is then carried into the cell with these siderophores, is bound by pyoverdine and pyochelin [70]. The present meta-analysis of transcriptomic data revealed that genes involved in pyochelin synthesis (pchC, pchE and pchG) were up-regulated and that genes involved in pyoverdine synthesis (pvdA and pvdH) were down-regulated in biofilms of P. aeruginosa. Mutation of pvdA gene was earlier reported to abolish pyoverdine production without affecting P. aeruginosa biofilm formation [71]. This is in line with earlier findings by Banin et al, [68] which shown that pyoverdine by itself is not required for active iron uptake for biofilm formation [70]. Utilising these genes will help us control and treat biofilm-based infections in addition to providing insight into further biofilm creation and development mechanisms.
Targeting bacterial DNA-binding proteins destroyed biofilms and liberated resident bacteria, promoting their eventual host immune effector clearance or antibiotics, which are now effective at significantly lower concentrations [72]. In the present study, the fis is the only DNA binding/regulatory protein that was noticed to be up-regulated in biofilms of P. aeruginosa. Hence, fis was used as the drug target for computational based screening of drug candidate that could inhibit biofilm formation in P. aeruginosa. Computational screening and docking analysis revealed that Lys 97, Try101 and Gln100 are the fis amino acids that are involved in interaction with dexamethasone. Structural analysis of P. aeruginosa fis protein revealed that Lys97 is an essential aminoacid that is required for sequence-specific binding of fis to its target DNA site [31]. Based on the above results, it was proposed that dexamethasone could inhibit P. aeruginosa biofilm formation via targeting fis. In addition, the invitro biofilm assay revealed that dexamethasone has inhibited the P. aeruginosa biofilm formation at sub-inhibitory concentrations. Further supporting the current findings of dexamethasone's anti-biofilm activity towards P. aeruginosa biofilm is the fact that the drug previously demonstrated anti-biofilm activity against S. aureus [73].
5. Conclusions
The present study presents a robust computational way to identify key players from bacterial expression patterns by harnessing the power of meta-analysis. By comparing with other pertinent gene sets, meta-analysis has been applied for the first time in P. aeruginosa transcriptome investigations. To combat biofilm-forming pathogenic bacteria like P. aeruginosa, it is critical to comprehend the fundamental distinctions between the behaviours of planktonic and biofilm bacteria. The lack of profile samples in determining the important genes and pathways that contribute to these variations is addressed by a meta-analysis of biofilm gene expression in P. aeruginosa. A set of 83 candidate genes that were differentially expressed in the biofilm state were found by the investigation, along with fis as a potential therapeutic target for P. aeruginosa biofilm formation. These techniques were combined with meta-analysis, virtual screening, and invitro methods. Given that these genes are expected to play a significant role in biofilms, they can be used as a hallmark target list for P. aeruginosa anti-biofilm medications. While the study identifies potential therapeutic targets, the effectiveness and safety of these targets in a clinical setting are not validated. The translation of findings from in vitro studies and virtual screenings to clinical applications can be challenging and requires extensive testing.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: meta-analysis DE; Table S2: vitrual screening.
Author Contributions
Emeka PM: Conceptualization, Funding acquisition, Project administration, Methodology, Formal analysis, Writing – original draft, review & editing. Badger-Emeka L.I.: Methodology, Formal analysis, Writing – original draft, review & editing. Thirugnanasambantham K: Conceptualization, Methodology, Data curation, Validation, Formal analysis, Writing – original draft, review & editing. Alatawi A S: Methodology, Formal analysis, Writing – original draft, review & editing.
Funding
The authors extend their appreciation to the King Salman center For Disability Research for funding this work through Research Group no KSRG-2023-107.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
No human subjects are used in the present study. Hence, it’s not applicable.
Data Availability Statement
Data will be made available on request.
Acknowledgments
The authors extend their appreciation to the King Salman center For Disability Research for funding this work through Research Group no KSRG-2023-107.
Conflicts of Interest
The authors have no known competing personal or financial interests with regards to this study.
References
- Dave, A.; Samarth, A.; Karolia, R.; Sharma, S.; Karunakaran, E.; Partridge, L.; MacNeil, S.; Monk, P.N.; Garg, P.; Roy, S. Characterization of Ocular Clinical Isolates of Pseudomonas Aeruginosa from Non-Contact Lens Related Keratitis Patients from South India. Microorganisms 2020, 8, 260. [Google Scholar] [CrossRef] [PubMed]
- Mann, E.E.; Wozniak, D.J. Pseudomonas Biofilm Matrix Composition and Niche Biology. FEMS Microbiol. Rev. 2012, 36, 893–916. [Google Scholar] [CrossRef] [PubMed]
- Crouzet, M.; Le Senechal, C.; Brözel, V.S.; Costaglioli, P.; Barthe, C.; Bonneu, M.; Garbay, B.; Vilain, S. Exploring Early Steps in Biofilm Formation: Set-up of an Experimental System for Molecular Studies. BMC Microbiol. 2014, 14, 253. [Google Scholar] [CrossRef] [PubMed]
- Kimkes, T.E.P.; Heinemann, M. How Bacteria Recognise and Respond to Surface Contact. FEMS Microbiol. Rev. 2020, 44, 106–122. [Google Scholar] [CrossRef] [PubMed]
- Franklin, M.J.; Nivens, D.E.; Weadge, J.T.; Howell, P.L. Biosynthesis of the Pseudomonas Aeruginosa Extracellular Polysaccharides, Alginate, Pel, and Psl. Front. Microbiol. 2011, 2. [Google Scholar] [CrossRef]
- Ueda, A.; Wood, T.K. Connecting Quorum Sensing, c-Di-GMP, Pel Polysaccharide, and Biofilm Formation in Pseudomonas Aeruginosa through Tyrosine Phosphatase TpbA (PA3885). PLoS Pathog. 2009, 5, e1000483. [Google Scholar] [CrossRef]
- De Kievit, T.R. Quorum Sensing in Pseudomonas Aeruginosa Biofilms. Environ. Microbiol. 2009, 11, 279–288. [Google Scholar] [CrossRef]
- Chang, C.-Y. Surface Sensing for Biofilm Formation in Pseudomonas Aeruginosa. Front. Microbiol. 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Harmsen, M.; Yang, L.; Pamp, S.J.; Tolker-Nielsen, T. An Update on Pseudomonas Aeruginosa Biofilm Formation, Tolerance, and Dispersal. FEMS Immunol. Med. Microbiol. 2010, 59, 253–268. [Google Scholar] [CrossRef]
- Gupta, K.; Marques, C.N.H.; Petrova, O.E.; Sauer, K. Antimicrobial Tolerance of Pseudomonas Aeruginosa Biofilms Is Activated during an Early Developmental Stage and Requires the Two-Component Hybrid SagS. J. Bacteriol. 2013, 195, 4975–4987. [Google Scholar] [CrossRef]
- Mulcahy, L.R.; Burns, J.L.; Lory, S.; Lewis, K. Emergence of Pseudomonas Aeruginosa Strains Producing High Levels of Persister Cells in Patients with Cystic Fibrosis. J. Bacteriol. 2010, 192, 6191–6199. [Google Scholar] [CrossRef] [PubMed]
- Patel, H.; Buchad, H.; Gajjar, D. Pseudomonas Aeruginosa Persister Cell Formation upon Antibiotic Exposure in Planktonic and Biofilm State. Sci. Rep. 2022, 12, 16151. [Google Scholar] [CrossRef] [PubMed]
- Pattnaik, S.; Barik, S.; Muralitharan, G.; Busi, S. Ferulic Acid Encapsulated Chitosan-tripolyphosphate Nanoparticles Attenuate Quorum Sensing Regulated Virulence and Biofilm Formation in Pseudomonas Aeruginosa PAO1. IET Nanobiotechnology 2018, 12, 1056–1061. [Google Scholar] [CrossRef] [PubMed]
- Ben Hur, D.; Kapach, G.; Wani, N.A.; Kiper, E.; Ashkenazi, M.; Smollan, G.; Keller, N.; Efrati, O.; Shai, Y. Antimicrobial Peptides against Multidrug-Resistant Pseudomonas Aeruginosa Biofilm from Cystic Fibrosis Patients. J. Med. Chem. 2022, 65, 9050–9062. [Google Scholar] [CrossRef]
- Saeki, E.K.; Martins, H.M.; Camargo, L.C. de; Anversa, L.; Tavares, E.R.; Yamada-Ogatta, S.F.; Lioni, L.M.Y.; Kobayashi, R.K.T.; Nakazato, G. Effect of Biogenic Silver Nanoparticles on the Quorum-Sensing System of Pseudomonas Aeruginosa PAO1 and PA14. Microorganisms 2022, 10, 1755. [Google Scholar] [CrossRef]
- Brindhadevi, K.; LewisOscar, F.; Mylonakis, E.; Shanmugam, S.; Verma, T.N.; Pugazhendhi, A. Biofilm and Quorum Sensing Mediated Pathogenicity in Pseudomonas Aeruginosa. Process Biochem. 2020, 96, 49–57. [Google Scholar] [CrossRef]
- Secor, P.R.; James, G.A.; Fleckman, P.; Olerud, J.E.; McInnerney, K.; Stewart, P.S. Staphylococcus Aureus Biofilm and Planktonic Cultures Differentially Impact Gene Expression, Mapk Phosphorylation, and Cytokine Production in Human Keratinocytes. BMC Microbiol. 2011, 11, 143. [Google Scholar] [CrossRef]
- Vallet, I.; Diggle, S.P.; Stacey, R.E.; Cámara, M.; Ventre, I.; Lory, S.; Lazdunski, A.; Williams, P.; Filloux, A. Biofilm Formation in Pseudomonas Aeruginosa: Fimbrial Cup Gene Clusters Are Controlled by the Transcriptional Regulator MvaT. J. Bacteriol. 2004, 186, 2880–2890. [Google Scholar] [CrossRef]
- Li, H.; Li, X.; Wang, Z.; Fu, Y.; Ai, Q.; Dong, Y.; Yu, J. Autoinducer-2 Regulates Pseudomonas Aeruginosa PAO1 Biofilm Formation and Virulence Production in a Dose-Dependent Manner. BMC Microbiol. 2015, 15, 192. [Google Scholar] [CrossRef]
- Dolatshah, L.; Tabatabaei, M. A Phenotypic and Molecular Investigation of Biofilm Formation in Clinical Samples of Pseudomonas Aeruginosa. Mol. Biol. Res. Commun. 2021, 10, 157–163. [Google Scholar] [CrossRef]
- Toyofuku, M.; Roschitzki, B.; Riedel, K.; Eberl, L. Identification of Proteins Associated with the Pseudomonas Aeruginosa Biofilm Extracellular Matrix. J. Proteome Res. 2012, 11, 4906–4915. [Google Scholar] [CrossRef] [PubMed]
- Park, A.J.; Murphy, K.; Krieger, J.R.; Brewer, D.; Taylor, P.; Habash, M.; Khursigara, C.M. A Temporal Examination of the Planktonic and Biofilm Proteome of Whole Cell Pseudomonas Aeruginosa PAO1 Using Quantitative Mass Spectrometry. Mol. Cell. Proteomics 2014, 13, 1095–1105. [Google Scholar] [CrossRef] [PubMed]
- Kaleta, M.F.; Petrova, O.E.; Zampaloni, C.; Garcia-Alcalde, F.; Parker, M.; Sauer, K. A Previously Uncharacterized Gene, PA2146, Contributes to Biofilm Formation and Drug Tolerance across the ɣ-Proteobacteria. npj Biofilms Microbiomes 2022, 8, 54. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, D.; Natarajan, J. Integrated Meta-Analysis and Machine Learning Approach Identifies Acyl-CoA Thioesterase with Other Novel Genes Responsible for Biofilm Development in Staphylococcus Aureus. Infect. Genet. Evol. 2021, 88, 104702. [Google Scholar] [CrossRef]
- Zhou, G.; Soufan, O.; Ewald, J.; Hancock, R.E.W.; Basu, N.; Xia, J. NetworkAnalyst 3.0: A Visual Analytics Platform for Comprehensive Gene Expression Profiling and Meta-Analysis. Nucleic Acids Res. 2019, 47, W234–W241. [Google Scholar] [CrossRef]
- Choi, J.K.; Yu, U.; Kim, S.; Yoo, O.J. Combining Multiple Microarray Studies and Modeling Interstudy Variation. Bioinformatics 2003, 19, i84–i90. [Google Scholar] [CrossRef]
- Zhang, Y.; Jenkins, D.F.; Manimaran, S.; Johnson, W.E. Alternative Empirical Bayes Models for Adjusting for Batch Effects in Genomic Studies. BMC Bioinformatics 2018, 19, 262. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and Integrative Analysis of Large Gene Lists Using DAVID Bioinformatics Resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Sherman, B.T.; Hao, M.; Qiu, J.; Jiao, X.; Baseler, M.W.; Lane, H.C.; Imamichi, T.; Chang, W. DAVID: A Web Server for Functional Enrichment Analysis and Functional Annotation of Gene Lists (2021 Update). Nucleic Acids Res. 2022, 50, W216–W221. [Google Scholar] [CrossRef]
- Li, J.; Miao, B.; Wang, S.; Dong, W.; Xu, H.; Si, C.; Wang, W.; Duan, S.; Lou, J.; Bao, Z.; et al. Hiplot: A Comprehensive and Easy-to-Use Web Service for Boosting Publication-Ready Biomedical Data Visualization. Brief. Bioinform. 2022, 23. [Google Scholar] [CrossRef]
- Zhou, J.; Gao, Z.; Zhang, H.; Dong, Y. Crystal Structure of the Nucleoid-Associated Protein Fis (PA4853) from Pseudomonas Aeruginosa. Acta Crystallogr. Sect. F Struct. Biol. Commun. 2020, 76, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A Visualization System for Exploratory Research and Analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef]
- Gan, J.; Liu, J.; Liu, Y.; Chen, S.; Dai, W.; Xiao, Z.-X.; Cao, Y. DrugRep: An Automatic Virtual Screening Server for Drug Repurposing. Acta Pharmacol. Sin. 2023, 44, 888–896. [Google Scholar] [CrossRef] [PubMed]
- Kuriata, A.; Gierut, A.M.; Oleniecki, T.; Ciemny, M.P.; Kolinski, A.; Kurcinski, M.; Kmiecik, S. CABS-Flex 2.0: A Web Server for Fast Simulations of Flexibility of Protein Structures. Nucleic Acids Res. 2018, 46, W338–W343. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute Performance Standards for Antimicrobial Disk Susceptibility Tests: Approved Standard - Eleventh Edition; 2012; Vol. 32; ISBN 1562384856.
- Pobiega, K.; Kraśniewska, K.; Derewiaka, D.; Gniewosz, M. Comparison of the Antimicrobial Activity of Propolis Extracts Obtained by Means of Various Extraction Methods. J. Food Sci. Technol. 2019, 56, 5386–5395. [Google Scholar] [CrossRef] [PubMed]
- Elshikh, M.; Ahmed, S.; Funston, S.; Dunlop, P.; McGaw, M.; Marchant, R.; Banat, I.M. Resazurin-Based 96-Well Plate Microdilution Method for the Determination of Minimum Inhibitory Concentration of Biosurfactants. Biotechnol. Lett. 2016, 38, 1015–1019. [Google Scholar] [CrossRef]
- Kim, Y.-M.; Son, H.; Park, S.-C.; Lee, J.-K.; Jang, M.-K.; Lee, J.R. Anti-Biofilm Effects of Rationally Designed Peptides against Planktonic Cells and Pre-Formed Biofilm of Pseudomonas Aeruginosa. Antibiotics 2023, 12, 349. [Google Scholar] [CrossRef]
- Roehm, N.W.; Rodgers, G.H.; Hatfield, S.M.; Glasebrook, A.L. An Improved Colorimetric Assay for Cell Proliferation and Viability Utilizing the Tetrazolium Salt XTT. J. Immunol. Methods 1991, 142, 257–265. [Google Scholar] [CrossRef]
- Akshaya, B.S.; Premraj, K.; Iswarya, C.; Muthusamy, S.; Ibrahim, H.-I.M.; Khalil, H.E.; Ashokkumar, V.; Vickram, S.; Senthil Kumar, V.; Palanisamy, S.; et al. Cinnamaldehyde Inhibits Enterococcus Faecalis Biofilm Formation and Promotes Clearance of Its Colonization by Modulation of Phagocytes in Vitro. Microb. Pathog. 2023, 181, 106157. [Google Scholar] [CrossRef]
- Harikrishnan, P.; Arayambath, B.; Jayaraman, V.K.; Ekambaram, K.; Ahmed, E.A.; Senthilkumar, P.; Ibrahim, H.I.M.; Sundaresan, A.; Thirugnanasambantham, K. Thidiazuron, a Phenyl-Urea Cytokinin, Inhibits Ergosterol Synthesis and Attenuates Biofilm Formation of Candida Albicans. World J. Microbiol. Biotechnol. 2022, 38, 1–14. [Google Scholar] [CrossRef]
- Emeka, P.M.; Badger-Emeka, L.I.; Ibrahim, H.I.M.; Thirugnanasambantham, K.; Hussen, J. Inhibitory Potential of Mangiferin on Glucansucrase Producing Streptococcus Mutans Biofilm in Dental Plaque. Appl. Sci. 2020, 10, 1–17. [Google Scholar] [CrossRef]
- Tuon, F.F.; Dantas, L.R.; Suss, P.H.; Tasca Ribeiro, V.S. Pathogenesis of the Pseudomonas Aeruginosa Biofilm: A Review. Pathogens 2022, 11, 300. [Google Scholar] [CrossRef] [PubMed]
- Hojjati, F.; Roointan, A.; Gholaminejad, A.; Eshraghi, Y.; Gheisari, Y. Identification of Key Genes and Biological Regulatory Mechanisms in Diabetic Nephropathy: Meta-Analysis of Gene Expression Datasets. Nefrología 2023, 43, 575–586. [Google Scholar] [CrossRef]
- Ramasamy, A.; Mondry, A.; Holmes, C.C.; Altman, D.G. Key Issues in Conducting a Meta-Analysis of Gene Expression Microarray Datasets. PLoS Med. 2008, 5, e184. [Google Scholar] [CrossRef]
- Cho, H.; Kim, H.; Na, D.; Kim, S.Y.; Jo, D.; Lee, D. Meta-Analysis Method for Discovering Reliable Biomarkers by Integrating Statistical and Biological Approaches: An Application to Liver Toxicity. Biochem. Biophys. Res. Commun. 2016, 471, 274–281. [Google Scholar] [CrossRef] [PubMed]
- Whiteley, M.; Bangera, M.G.; Bumgarner, R.E.; Parsek, M.R.; Teitzel, G.M.; Lory, S.; Greenberg, E.P. Gene Expression in Pseudomonas Aeruginosa Biofilms. Nature 2001, 413, 860–864. [Google Scholar] [CrossRef]
- Schembri, M.A.; Kjærgaard, K.; Klemm, P. Global Gene Expression in Escherichia Coli Biofilms. Mol. Microbiol. 2003, 48, 253–267. [Google Scholar] [CrossRef]
- Resch, A.; Rosenstein, R.; Nerz, C.; Götz, F. Differential Gene Expression Profiling of Staphylococcus Aureus Cultivated under Biofilm and Planktonic Conditions. Appl. Environ. Microbiol. 2005, 71, 2663–2676. [Google Scholar] [CrossRef]
- Post, D.M.; Held, J.M.; Ketterer, M.R.; Phillips, N.J.; Sahu, A.; Apicella, M.A.; Gibson, B.W. Comparative Analyses of Proteins from Haemophilus Influenzae Biofilm and Planktonic Populations Using Metabolic Labeling and Mass Spectrometry. BMC Microbiol. 2014, 14, 329. [Google Scholar] [CrossRef]
- Bottagisio, M.; Barbacini, P.; Bidossi, A.; Torretta, E.; DeLancey-Pulcini, E.; Gelfi, C.; James, G.A.; Lovati, A.B.; Capitanio, D. Phenotypic Modulation of Biofilm Formation in a Staphylococcus Epidermidis Orthopedic Clinical Isolate Grown Under Different Mechanical Stimuli: Contribution From a Combined Proteomic Study. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef]
- Zemke, A.C.; D’Amico, E.J.; Torres, A.M.; Carreno-Florez, G.P.; Keeley, P.; DuPont, M.; Kasturiarachi, N.; Bomberger, J.M. Bacterial Respiratory Inhibition Triggers Dispersal of Pseudomonas Aeruginosa Biofilms. Appl. Environ. Microbiol. 2023, 89. [Google Scholar] [CrossRef] [PubMed]
- Ueda, A.; Attila, C.; Whiteley, M.; Wood, T.K. Uracil Influences Quorum Sensing and Biofilm Formation in Pseudomonas Aeruginosa and Fluorouracil Is an Antagonist. Microb. Biotechnol. 2009, 2, 62–74. [Google Scholar] [CrossRef] [PubMed]
- Naughton, S.; Parker, D.; Seemann, T.; Thomas, T.; Turnbull, L.; Rose, B.; Bye, P.; Cordwell, S.; Whitchurch, C.; Manos, J. Pseudomonas Aeruginosa AES-1 Exhibits Increased Virulence Gene Expression during Chronic Infection of Cystic Fibrosis Lung. PLoS One 2011, 6, e24526. [Google Scholar] [CrossRef]
- Taylor, P.K.; Zhang, L.; Mah, T.-F. Loss of the Two-Component System TctD-TctE in Pseudomonas Aeruginosa Affects Biofilm Formation and Aminoglycoside Susceptibility in Response to Citric Acid. mSphere 2019, 4. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.; Zhou, C.; Qin, J.; Jiang, Y.; Li, D.; Tang, X.; Luo, J.; Kong, J.; Wang, K. Molecular Mechanisms of DNase Inhibition of Early Biofilm Formation Pseudomonas Aeruginosa or Staphylococcus Aureus: A Transcriptome Analysis. Biofilm 2024, 7, 100174. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wilks, J.C.; Danhorn, T.; Ramos, I.; Croal, L.; Newman, D.K. Phenazine-1-Carboxylic Acid Promotes Bacterial Biofilm Development via Ferrous Iron Acquisition. J. Bacteriol. 2011, 193, 3606–3617. [Google Scholar] [CrossRef]
- Li, K.; Xu, C.; Jin, Y.; Sun, Z.; Liu, C.; Shi, J.; Chen, G.; Chen, R.; Jin, S.; Wu, W. SuhB Is a Regulator of Multiple Virulence Genes and Essential for Pathogenesis of Pseudomonas Aeruginosa. MBio 2013, 4. [Google Scholar] [CrossRef]
- Shi, J.; Jin, Y.; Bian, T.; Li, K.; Sun, Z.; Cheng, Z.; Jin, S.; Wu, W. <scp>SuhB</Scp> Is a Novel Ribosome Associated Protein That Regulates Expression of <scp>MexXY</Scp> by Modulating Ribosome Stalling in <scp> P </Scp> Seudomonas Aeruginosa. Mol. Microbiol. 2015, 98, 370–383. [Google Scholar] [CrossRef]
- Schinner, S.; Engelhardt, F.; Preusse, M.; Thöming, J.G.; Tomasch, J.; Häussler, S. Genetic Determinants of Pseudomonas Aeruginosa Fitness during Biofilm Growth. Biofilm 2020, 2, 100023. [Google Scholar] [CrossRef]
- Moor, H.; Teppo, A.; Lahesaare, A.; Kivisaar, M.; Teras, R. Fis Overexpression Enhances Pseudomonas Putida Biofilm Formation by Regulating the Ratio of LapA and LapF. Microbiology 2014, 160, 2681–2693. [Google Scholar] [CrossRef]
- Long, Y.; Fu, W.; Wang, S.; Deng, X.; Jin, Y.; Bai, F.; Cheng, Z.; Wu, W. Fis Contributes to Resistance of Pseudomonas Aeruginosa to Ciprofloxacin by Regulating Pyocin Synthesis. J. Bacteriol. 2020, 202. [Google Scholar] [CrossRef] [PubMed]
- Holden, E.R.; Yasir, M.; Turner, A.K.; Wain, J.; Charles, I.G.; Webber, M.A. Massively Parallel Transposon Mutagenesis Identifies Temporally Essential Genes for Biofilm Formation in Escherichia Coli. Microb. Genomics 2021, 7. [Google Scholar] [CrossRef] [PubMed]
- Gélinas, M.; Museau, L.; Milot, A.; Beauregard, P.B. The de Novo Purine Biosynthesis Pathway Is the Only Commonly Regulated Cellular Pathway during Biofilm Formation in TSB-Based Medium in Staphylococcus Aureus and Enterococcus Faecalis. Microbiol. Spectr. 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Fang, X.; Wood, T.K.; Huang, Z.J. A Systems-Level Approach for Investigating Pseudomonas Aeruginosa Biofilm Formation. PLoS One 2013, 8, e57050. [Google Scholar] [CrossRef] [PubMed]
- Mueller, R.S.; McDougald, D.; Cusumano, D.; Sodhi, N.; Kjelleberg, S.; Azam, F.; Bartlett, D.H. Vibrio Cholerae Strains Possess Multiple Strategies for Abiotic and Biotic Surface Colonization. J. Bacteriol. 2007, 189, 5348–5360. [Google Scholar] [CrossRef]
- Li, H.; Du, X.; Chen, C.; Qi, J.; Wang, Y. Integrating Transcriptomics and Metabolomics Analysis on Kojic Acid Combating Acinetobacter Baumannii Biofilm and Its Potential Roles. Microbiol. Res. 2022, 254, 126911. [Google Scholar] [CrossRef]
- Yadav, M.K.; Kwon, S.K.; Cho, C.G.; Park, S.; Chae, S.; Song, J. Gene Expression Profile of Early in Vitro Biofilms of Streptococcus Pneumoniae. Microbiol. Immunol. 2012, 56, 621–629. [Google Scholar] [CrossRef]
- Thöming, J.G.; Tomasch, J.; Preusse, M.; Koska, M.; Grahl, N.; Pohl, S.; Willger, S.D.; Kaever, V.; Müsken, M.; Häussler, S. Parallel Evolutionary Paths to Produce More than One Pseudomonas Aeruginosa Biofilm Phenotype. npj Biofilms Microbiomes 2020, 6, 2. [Google Scholar] [CrossRef]
- Banin, E.; Vasil, M.L.; Greenberg, E.P. Iron and Pseudomonas Aeruginosa Biofilm Formation. Proc. Natl. Acad. Sci. 2005, 102, 11076–11081. [Google Scholar] [CrossRef]
- Kang, D.; Kirienko, N. V. High-Throughput Genetic Screen Reveals That Early Attachment and Biofilm Formation Are Necessary for Full Pyoverdine Production by Pseudomonas Aeruginosa. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef]
- Novotny, L.A.; Goodman, S.D.; Bakaletz, L.O. Targeting a Bacterial DNABII Protein with a Chimeric Peptide Immunogen or Humanised Monoclonal Antibody to Prevent or Treat Recalcitrant Biofilm-Mediated Infections. eBioMedicine 2020, 59, 102867. [Google Scholar] [CrossRef] [PubMed]
- Saleh, M.M.; Yousef, N.; Shafik, S.M.; Abbas, H.A. Attenuating the Virulence of the Resistant Superbug Staphylococcus Aureus Bacteria Isolated from Neonatal Sepsis by Ascorbic Acid, Dexamethasone, and Sodium Bicarbonate. BMC Microbiol. 2022, 22, 268. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Data pre-processing and processing; (a) Comparison and contrast between the biofilm and planktonic samples using PCA plots of batch effect removal; (b) Density plots of batch effect elimination against log2 of read counts display the relative distribution of various counts within each group.
Figure 1.
Data pre-processing and processing; (a) Comparison and contrast between the biofilm and planktonic samples using PCA plots of batch effect removal; (b) Density plots of batch effect elimination against log2 of read counts display the relative distribution of various counts within each group.
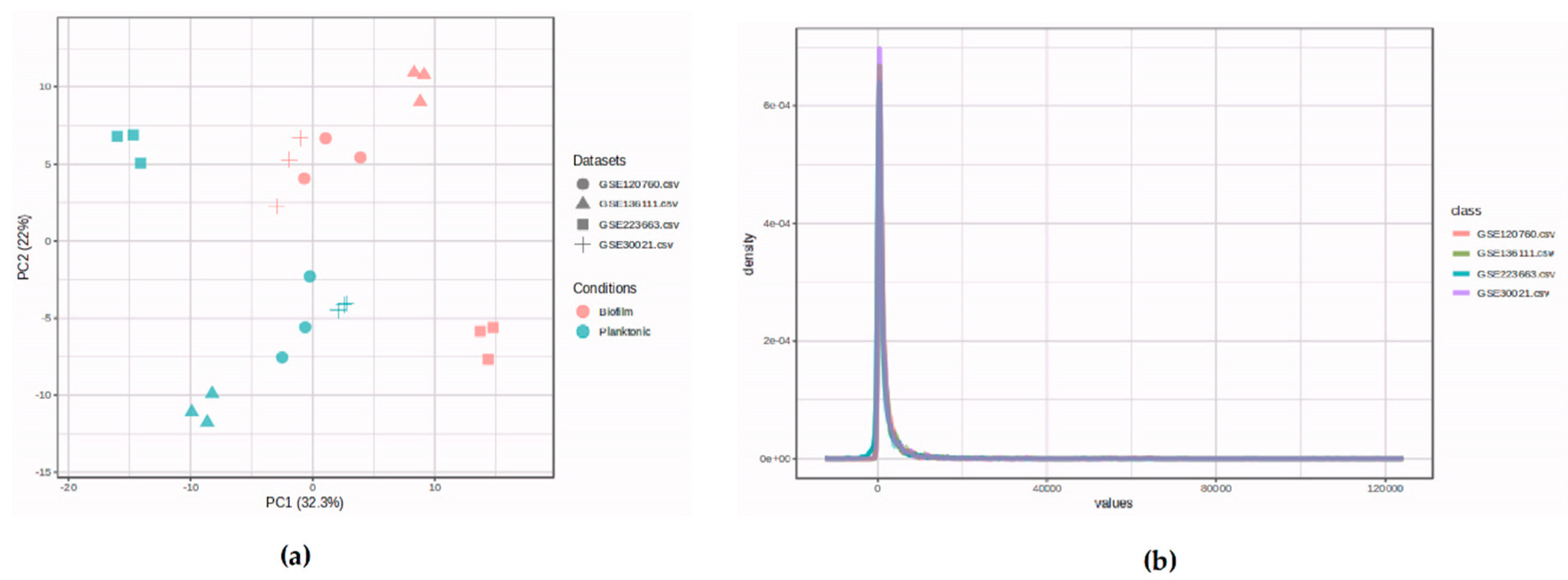
Figure 2.
Analysis of GEOs. (a) The top 50 DEGs' heatmap based on adjusted p-value; (b) Volcano plot of genes that were selected based on meta-analysis using random effect size.
Figure 2.
Analysis of GEOs. (a) The top 50 DEGs' heatmap based on adjusted p-value; (b) Volcano plot of genes that were selected based on meta-analysis using random effect size.
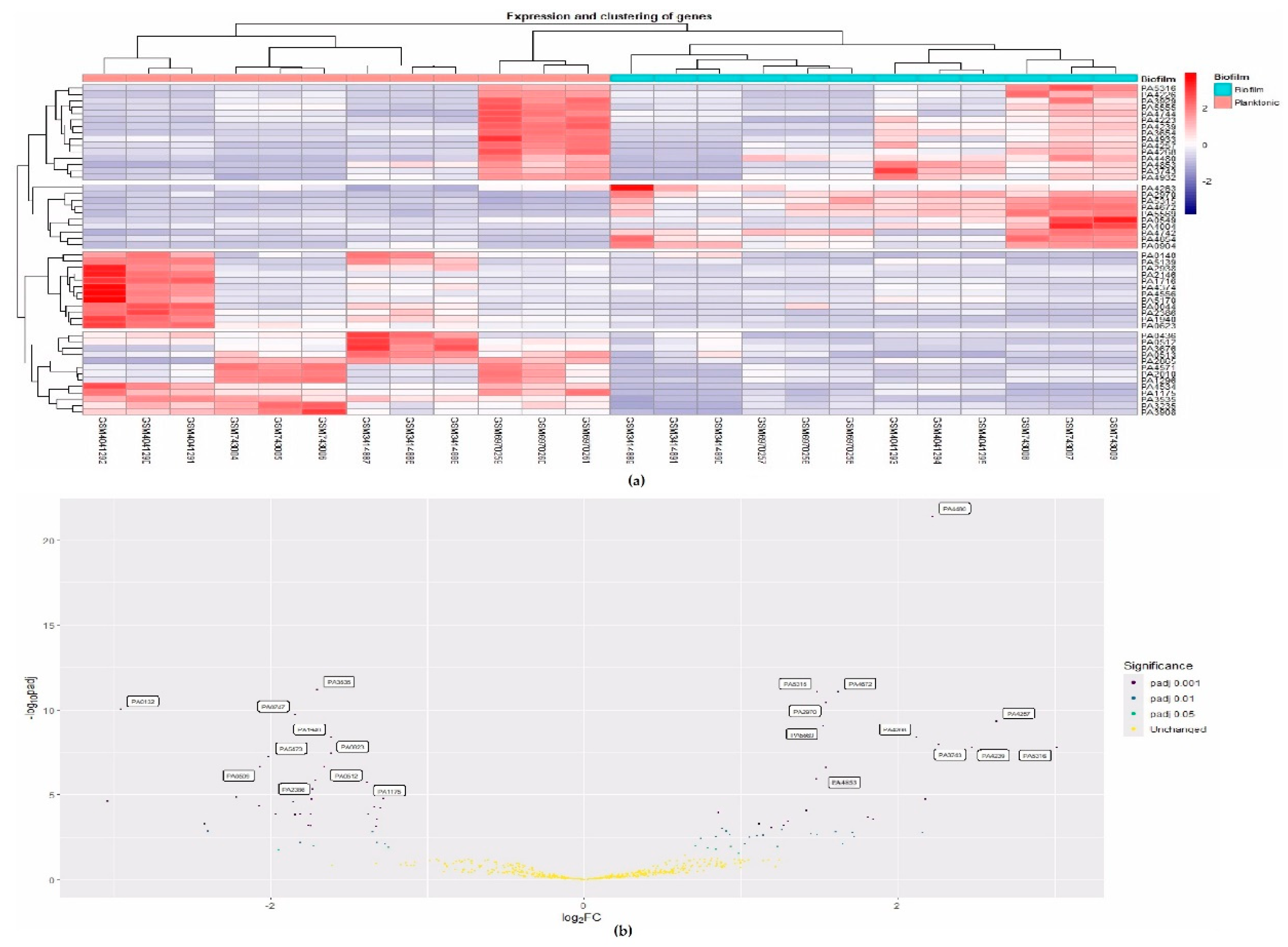
Figure 3.
Functional enrichment analysis of DEGs up-regulated in biofilm samples. (a) Enriched gene GO terms under molecular function; (b) Enrichment GO terms under biological process; (c) Enriched GO terms under cellular component; (d) Enriched KEGG pathways.
Figure 3.
Functional enrichment analysis of DEGs up-regulated in biofilm samples. (a) Enriched gene GO terms under molecular function; (b) Enrichment GO terms under biological process; (c) Enriched GO terms under cellular component; (d) Enriched KEGG pathways.
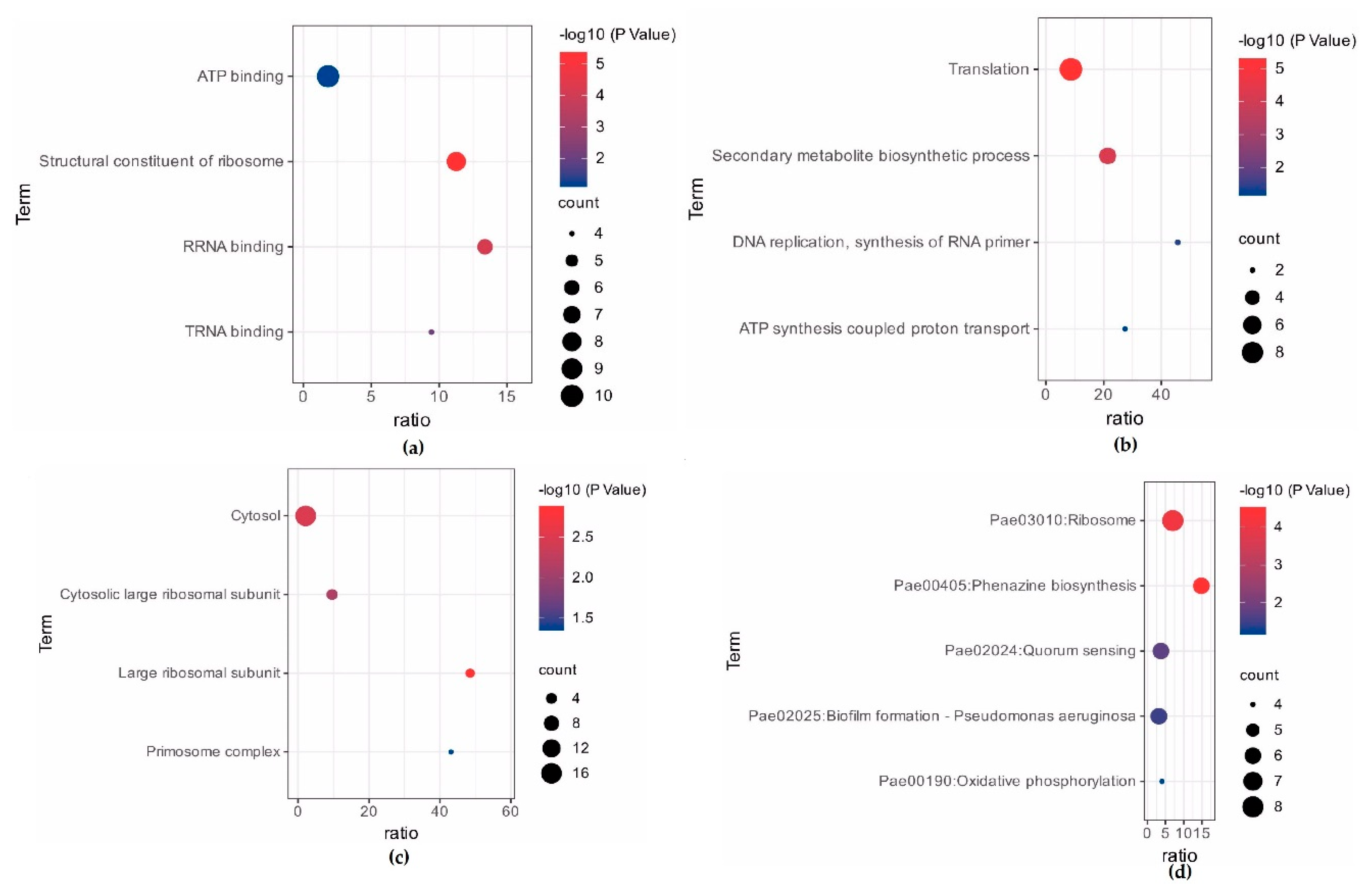
Figure 4.
Functional enrichment analysis of DEGs down-regulated in biofilm samples.
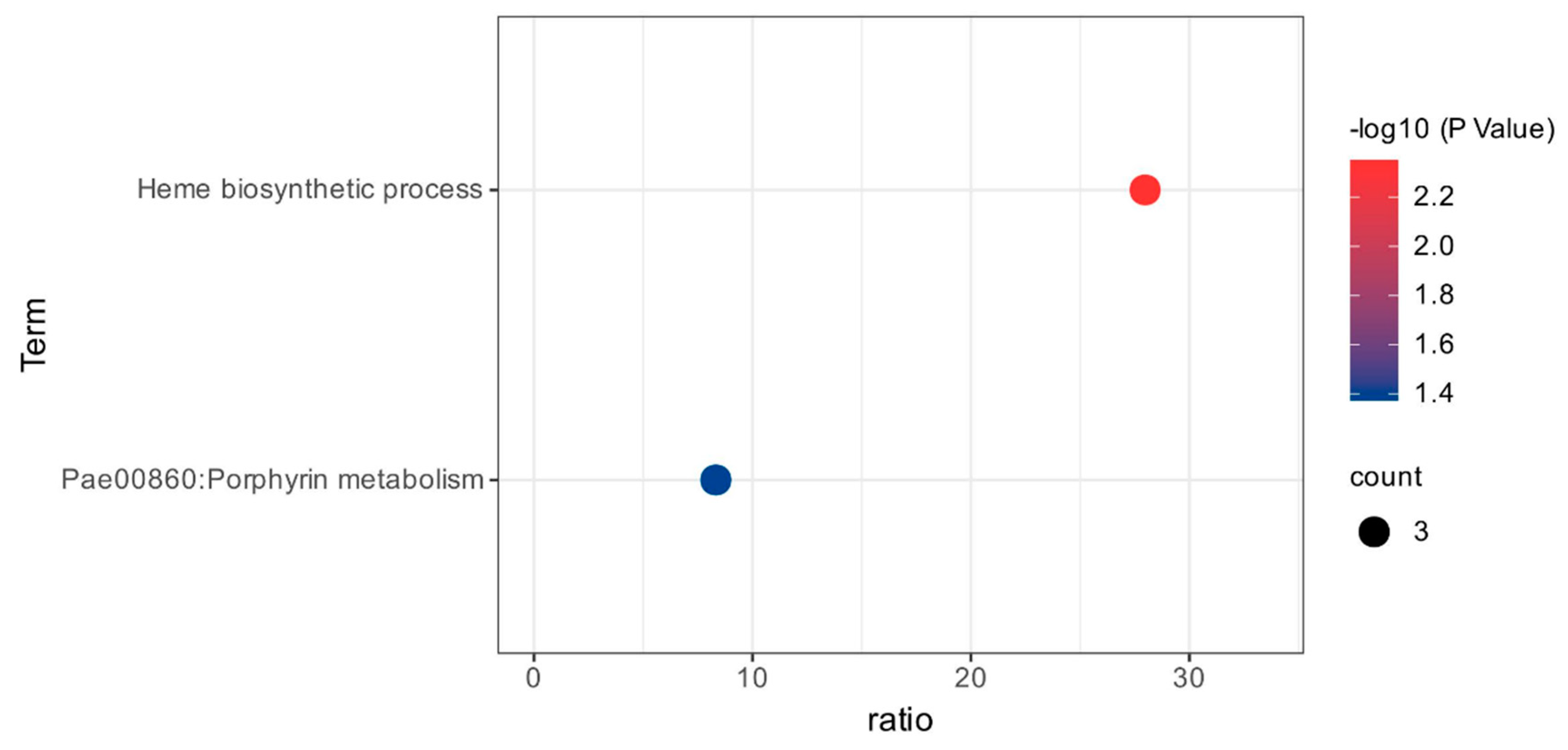
Figure 5.
Ribosome pathway from KEGG database. Red colour filed denotes genes that are up-regulated in P. aeruginosa biofilm formation (pae03010).
Figure 5.
Ribosome pathway from KEGG database. Red colour filed denotes genes that are up-regulated in P. aeruginosa biofilm formation (pae03010).
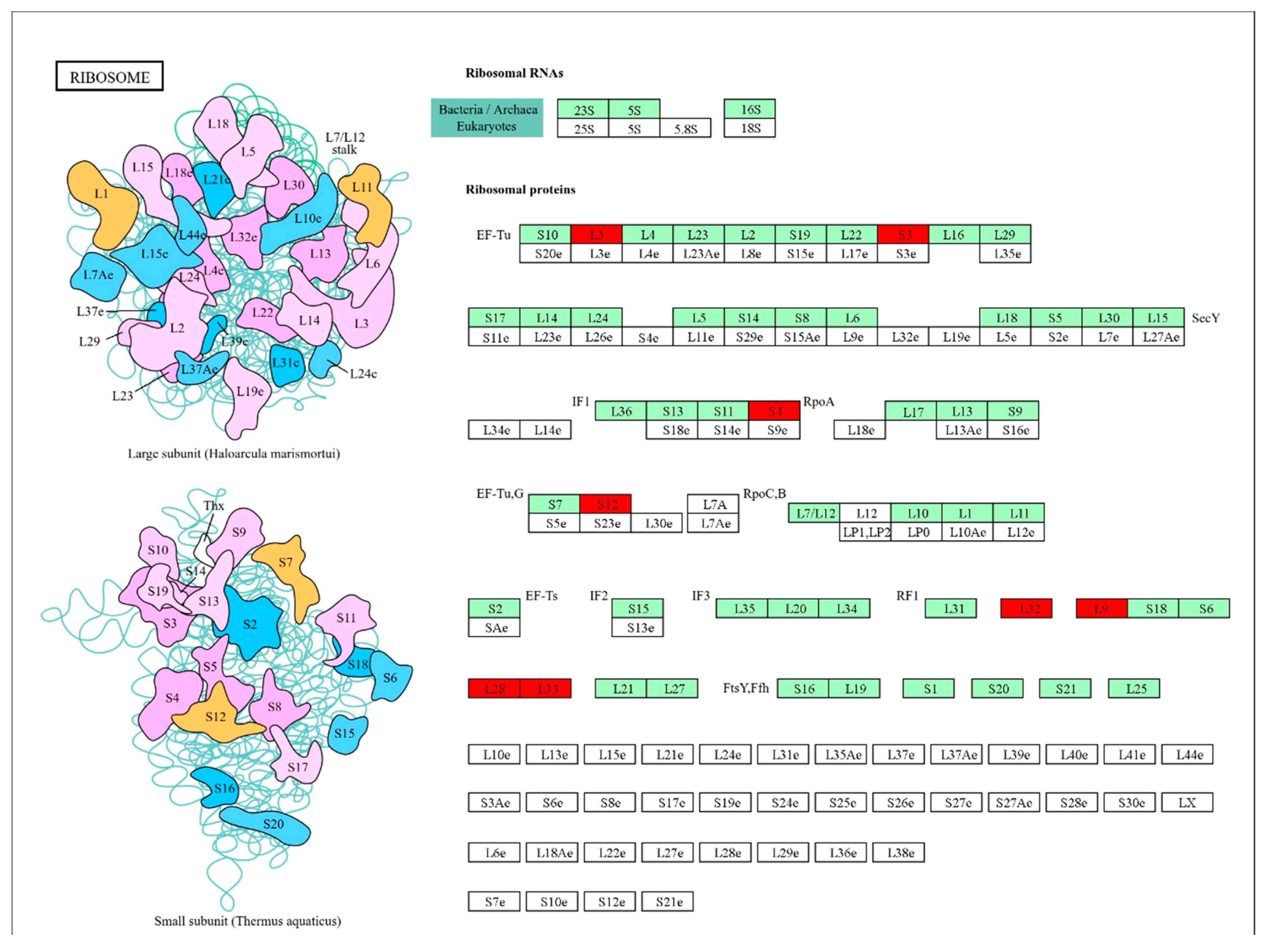
Figure 6.
Quorum sensing pathway from KEGG database. Red colour filed denotes genes that are up-regulated in P. aeruginosa biofilm formation (pae02024).
Figure 6.
Quorum sensing pathway from KEGG database. Red colour filed denotes genes that are up-regulated in P. aeruginosa biofilm formation (pae02024).
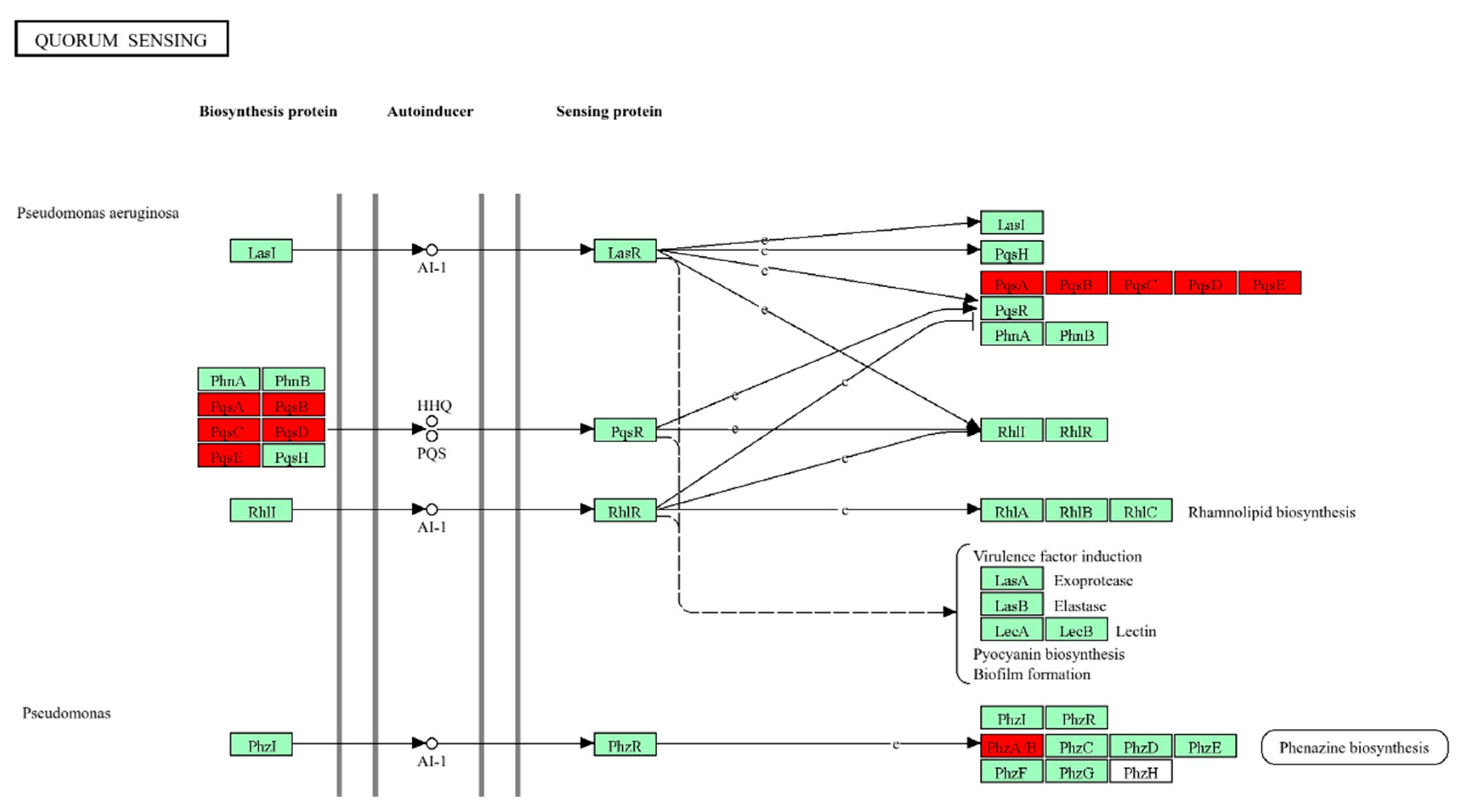
Figure 7.
Biofilm formation pathway from KEGG database. Red colour filed denotes genes that are up regulated in P. aeruginosa biofilm formation (map05111).
Figure 7.
Biofilm formation pathway from KEGG database. Red colour filed denotes genes that are up regulated in P. aeruginosa biofilm formation (map05111).
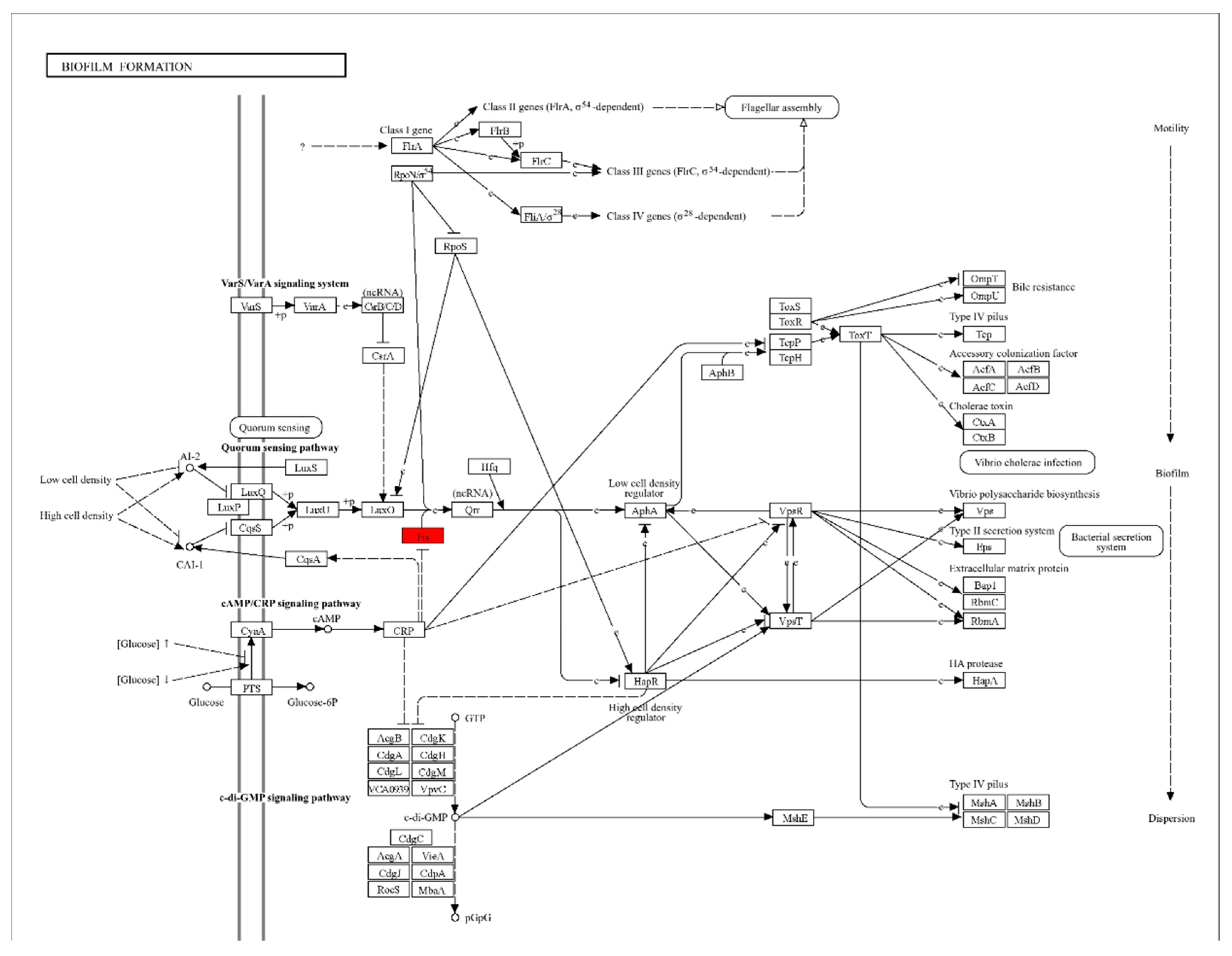
Figure 8.
Computational analysis of dexamethasone- fis interaction. (a). Interaction of dexamethasone to the active site of P. aeruginosa factors for inversion stimulation (fis); (b) RMSF of the dexamethasone as well as native fis protein.
Figure 8.
Computational analysis of dexamethasone- fis interaction. (a). Interaction of dexamethasone to the active site of P. aeruginosa factors for inversion stimulation (fis); (b) RMSF of the dexamethasone as well as native fis protein.

Figure 9.
Anti-microbial potential of dexamethasone against P. aeruginosa investigated. (a) agar well disc diffusion method; (b) Zone of inhibition (mm) of dexamethasone and Ciprofloxacin (20µg/well) against P. aeruginosa. The means ± standard error from three replicates were used to express the values, and *P≤0.05 indicated that the results were significant.
Figure 9.
Anti-microbial potential of dexamethasone against P. aeruginosa investigated. (a) agar well disc diffusion method; (b) Zone of inhibition (mm) of dexamethasone and Ciprofloxacin (20µg/well) against P. aeruginosa. The means ± standard error from three replicates were used to express the values, and *P≤0.05 indicated that the results were significant.
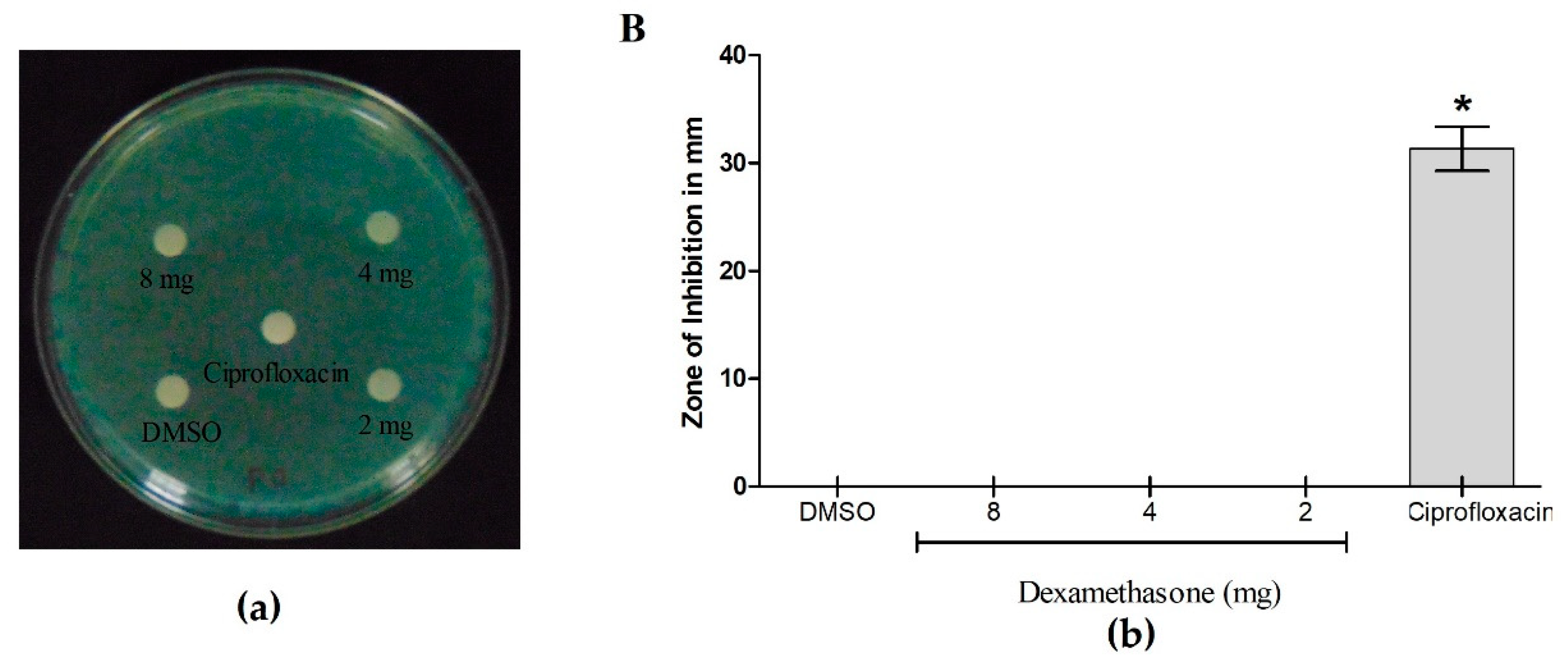
Figure 10.
Effect of dexamethasone on P. aeruginosa biofilm formation. (a) Quantitative assessment of the P. aeruginosa biofilm using crystal violet staining; (b) Quantitative assessment of the P. aeruginosa biofilm using XTT reduction assay; (c) The P. aeruginosa biofilm was evaluated using acryline orange staining. The vehicle control utilized was DMSO, and the results were reported as the means ± standard error of three replicates. Findings were deemed noteworthy when *P < 0.05.
Figure 10.
Effect of dexamethasone on P. aeruginosa biofilm formation. (a) Quantitative assessment of the P. aeruginosa biofilm using crystal violet staining; (b) Quantitative assessment of the P. aeruginosa biofilm using XTT reduction assay; (c) The P. aeruginosa biofilm was evaluated using acryline orange staining. The vehicle control utilized was DMSO, and the results were reported as the means ± standard error of three replicates. Findings were deemed noteworthy when *P < 0.05.
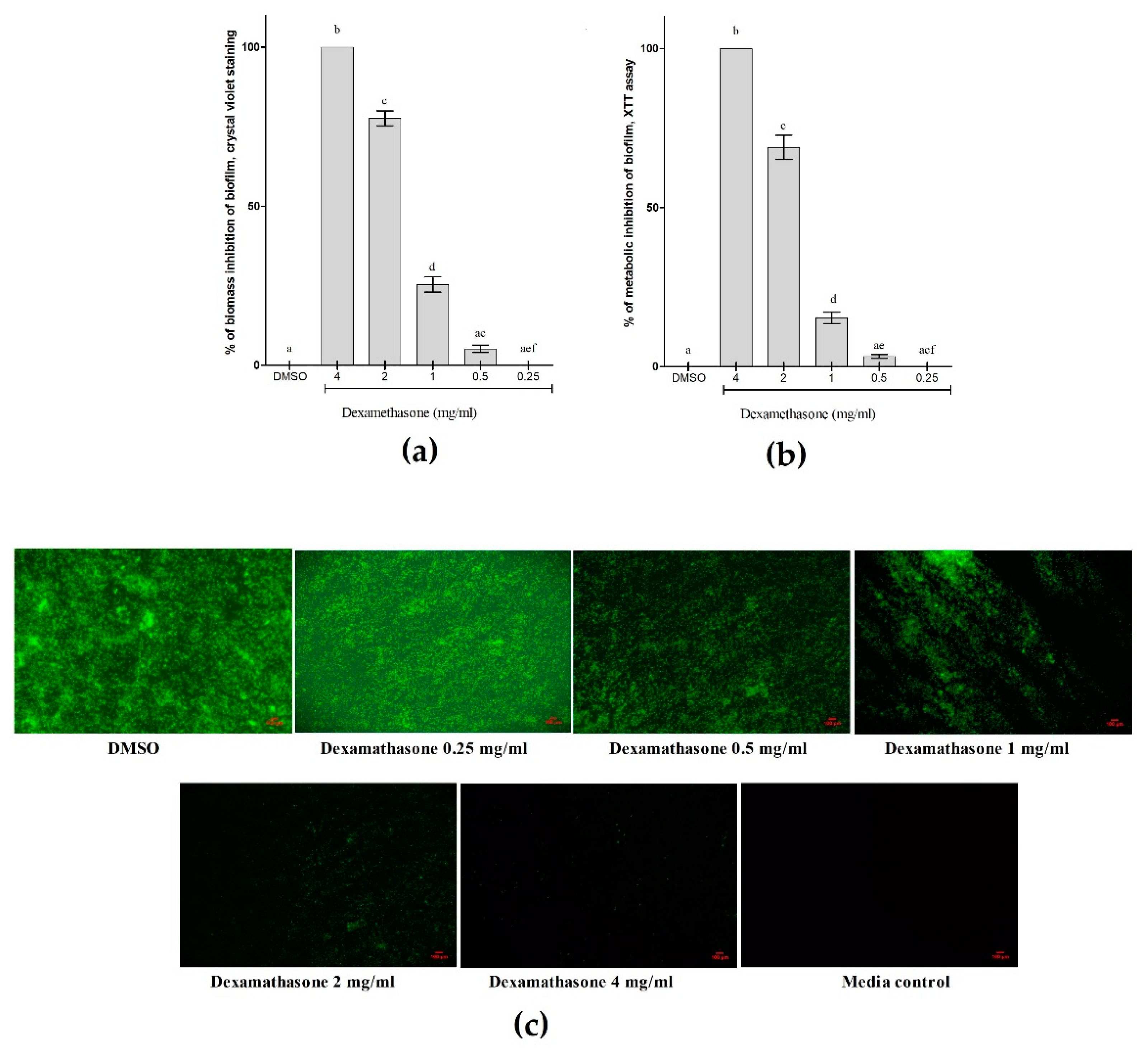

Table 1.
RNA-Seq/microarray samples used for differential expression analysis.
| S.No. | GEO series ID | Total Number of Samples (Control: Test) | Planktonic (Control) | Biofilm (Test) | Study Platform |
| 1 | GSE30021 | 6 (3:3) | GSM743004, GSM743005, GSM743006 | GSM743007, GSM743008, GSM743009 |
GPL84 [Pae_G1a] Affymetrix Pseudomonas aeruginosa Array |
| 2 | GSE120760 | 6 (3:3) | GSM3414886, GSM3414887, GSM3414888 | GSM3414889, GSM3414891, GSM3414890 | GPL84 [Pae_G1a] Affymetrix Pseudomonas aeruginosa Array |
| 3 | GSE136111 | 6 (3:3) | GSM4041290, GSM4041291, GSM4041292 | GSM4041293, GSM4041294, GSM4041295 | Illumina NextSeq 500 (Pseudomonas aeruginosa) |
| 4 | GSE223663 | 6 (3:3) | GSM6970259, GSM6970260, GSM6970261 |
GSM6970256, GSM6970257, GSM6970258 |
Illumina HiSeq 2500 (Pseudomonas aeruginosa) |
Table 2.
DEGs identified based on meta-analysis.
| S.No. | Expression | Significance | No. of genes |
| 1 | Down-regulated | padj 0.001 | 30 |
| 2 | Down-regulated | padj 0.01 | 5 |
| 3 | Down-regulated | padj 0.05 | 3 |
| 4 | Unchanged | Unchanged | 447 |
| 5 | Up-regulated | padj 0.001 | 21 |
| 6 | Up-regulated | padj 0.01 | 17 |
| 7 | Up-regulated | padj 0.05 | 7 |
Table 3.
List of top 25 up-regulated genes in biofilm.
| Locus Tag | Gene symbol | Gene name | Combined effect size | Padj value |
| PA5316 | rpmB | 50S ribosomal protein L28 | 3.01 | 1.57E-08 |
| PA4257 | rpsC | 30S ribosomal protein S3 | 2.63 | 4.86E-10 |
| PA4239 | rpsD | 30S ribosomal protein S4 | 2.47 | 1.58E-08 |
| PA3743 | trmD | tRNA (guanine-N1)-methyltransferase | 2.26 | 1.19E-08 |
| PA4480 | mreC | rod shape-determining protein MreC | 2.22 | 4.06E-22 |
| PA4933 | - | hypothetical protein | 2.17 | 1.86E-05 |
| PA0549 | - | hypothetical protein | 2.15 | 0.0017 |
| PA4268 | rpsL | 30S ribosomal protein S12 | 2.12 | 4.32E-09 |
| PA3929 | cioB | cyanide insensitive terminal oxidase | 1.85 | 0.0003 |
| PA3654 | pyrH | uridylate kinase | 1.81 | 0.0002 |
| PA4004 | - | conserved hypothetical protein | 1.72 | 0.0028 |
| PA4226 | pchE | dihydroaeruginoic acid synthetase | 1.71 | 0.0017 |
| PA4223 | pchH | probable ATP-binding component of ABC transporter | 1.65 | 0.0022 |
| PA4672 | pth | peptidyl-tRNA hydrolase | 1.62 | 5.52E-14 |
| PA4853 | fis | DNA-binding protein Fis | 1.60 | 0.0016 |
| PA2970 | rpmF | 50S ribosomal protein L32 | 1.54 | 3.67E-11 |
| PA4854 | purH | phosphoribosylaminoimidazolecarboxamide formyltransferase | 1.54 | 2.52E-07 |
| PA5569 | rnpA | ribonuclease P protein component | 1.53 | 8.64E-10 |
| PA5555 | atpG | ATP synthase gamma chain | 1.48 | 0.0023 |
| PA5315 | rpmG | 50S ribosomal protein L33 | 1.48 | 8.82E-12 |
| PA4932 | rplI | 50S ribosomal protein L9 | 1.48 | 1.22E-06 |
| PA4744 | infB | translation initiation factor IF-2 | 1.45 | 0.0020 |
| PA0904 | lysC | aspartate kinase alpha and beta chain | 1.42 | 8.08E-05 |
| PA4263 | rplC | 50S ribosomal protein L3 | 1.30 | 0.0004 |
| PA4742 | truB | tRNA pseudouridine 55 synthase | 1.27 | 0.0006 |
Table 4.
List of top 25 down-regulated genes in biofilm.
| Locus Tag | Gene symbol | Gene name | Combined effect size | Padj value |
| PA3234 | yjcG | sodium:solute symporter | -3.04 | 2.34E-06 |
| PA0132 | bauA | Beta-alanine:pyruvate transaminase | -2.95 | 9.56E-11 |
| PA4931 | dnaB | replicative DNA helicase | -2.42 | 0.0005 |
| PA0459 | clpC | ClpA/B protease ATP binding subunit | -2.40 | 0.0013 |
| PA2413 | pvdH | L-2,4-diaminobutyrate:2-ketoglutarate 4-aminotransferase, PvdH | -2.22 | 1.40E-05 |
| PA3568 | ymmS | acetyl-coa synthetase | -2.07 | 4.76E-05 |
| PA0509 | nirN | Dihydro-Heme d1 Dehydrogenase | -2.06 | 2.42E-07 |
| PA5473 | - | conserved hypothetical protein | -2.01 | 5.86E-08 |
| PA2586 | gacA | response regulator GacA | -1.97 | 0.0001 |
| PA5153 | - | amino acid (lysine/arginine/ornithine/histidine/octopine) ABC transporter periplasmic binding protein | -1.95 | 0.0177 |
| PA3091 | - | hypothetical protein | -1.86 | 2.54E-05 |
| PA0747 | pauC | aldehyde dehydrogenase | -1.84 | 1.91E-10 |
| PA0301 | spuE | polyamine transport protein | -1.84 | 0.00014 |
| PA5139 | - | hypothetical protein | -1.81 | 0.0070 |
| PA1716 | pscC | Type III secretion outer membrane protein PscC precursor | -1.81 | 0.0001 |
| PA1296 | - | 2-hydroxyacid dehydrogenase | -1.75 | 0.0006 |
| PA2146 | - | conserved hypothetical protein | -1.74 | 0.0007 |
| PA0044 | exoT | exoenzyme T | -1.74 | 0.0001 |
| PA4571 | - | cytochrome c | -1.73 | 1.81E-05 |
| PA2938 | - | transporter | -1.73 | 4.82E-06 |
| PA5170 | arcD | arginine/ornithine antiporter | -1.71 | 0.0104 |
| PA2386 | pvdA | L-ornithine N5-oxygenase | -1.71 | 1.45E-06 |
| PA3535 | eprS | serine protease | -1.70 | 6.74E-12 |
| PA0512 | nirH | siroheme decarboxylase subunit | -1.66 | 2.30E-07 |
| PA1940 | - | hypothetical protein | -1.61 | 4.33E-09 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



