
Submitted:
16 October 2024
Posted:
17 October 2024
You are already at the latest version
Abstract
This review highlights the therapeutic potential of epigallocatechin gallate (EGCG) and forskolin in managing retinal diseases, with a focus on glaucoma, age-related macular degeneration (AMD), and diabetic retinopathy. EGCG, a potent polyphenol from green tea, exhibits significant antioxidant, anti-inflammatory, and neuroprotective effects, making it a promising candidate for reducing oxidative stress and inflammation in ocular tissues. Forskolin, a diterpene from Coleus forskohlii, increases cyclic AMP (cAMP) levels, which helps lower intraocular pressure (IOP) and provides neuroprotection. Both compounds target critical pathways involved in retinal disease progression, including oxidative stress, mitochondrial dysfunction, and inflammation, offering complementary therapeutic benefits. The review consolidates preclinical and clinical studies, highlighting the potential of EGCG and forskolin as adjunctive or alternative treatments for retinal diseases. Future research should explore the synergistic effects of these compounds, particularly in combination therapies aimed at addressing multiple pathogenic mechanisms in retinal health.
Keywords:
Epigallocatechin-gallate
; forskolin
; eye
; retinal disease
; glaucoma
; macular degeneration
; diabetic retinopathy
1. Introduction
Oxidative stress and inflammation are central to the development and progression of glaucoma and other eye diseases [1-2]. In glaucoma - a slowly progressing neurodegenerative disease affecting the optic nerve [3] - the buildup of reactive oxygen species (ROS) damages ocular tissues, particularly the trabecular meshwork and retinal ganglion cells, disrupting cellular homeostasis and contributing to increased intraocular pressure (IOP) [4]. This leads to optic nerve degeneration and vision loss. Chronic inflammation further aggravates this damage by activating pathways that degrade ocular tissue integrity [4]. In conditions like age-related macular degeneration (AMD) and diabetic retinopathy, oxidative stress also drives disease progression by damaging retinal cells and promoting harmful deposits (e.g., drusen in AMD) or impairing the blood-retinal barrier in diabetic retinopathy [5]. Inflammation accelerates cellular degeneration and contributes to retinal and optic nerve damage in these conditions [5]. Together, oxidative stress and inflammation create a cycle of tissue damage, emphasizing the need for therapeutic strategies targeting these processes to slow or prevent disease progression [4] (Figure 1).
The use of antioxidants and anti-inflammatory treatments, particularly through dietary supplements, has gained attention in managing glaucoma and other ophthalmic diseases. Antioxidants such as vitamins C and E, omega-3 fatty acids, and plant-derived compounds like flavonoids and carotenoids help neutralize ROS and reduce oxidative stress, a key factor in these diseases [6]. In glaucoma, antioxidant supplementation can protect the optic nerve and retinal ganglion cells, potentially slowing vision loss [7]. Similarly, in AMD [8] and diabetic retinopathy [9, 10], antioxidants have been shown to protect retinal cells from oxidative injury, reducing disease progression. Anti-inflammatory supplements, such as omega-3 fatty acids and curcumin, modulate inflammatory pathways that contribute to ocular damage. These supplements can reduce chronic inflammation, protecting the trabecular meshwork in glaucoma [11, 12] and preventing retinal inflammation in AMD and diabetic retinopathy [13-15]. Although more research is needed to standardize treatment protocols, antioxidants and anti-inflammatory supplements are promising adjuncts to traditional medical and surgical treatments for glaucoma and other eye diseases. Combining epigallocatechin gallate (EGCG) and forskolin in a single supplement might hold significant promise for preventing and treating various eye diseases. EGCG, a powerful antioxidant and anti-inflammatory compound found abundantly in green tea, has strong neuroprotective effects, particularly in addressing oxidative stress and inflammation in ocular pathologies [16]. Forskolin, a natural extract from Coleus forskohlii, lowers IOP by increasing cAMP levels in ocular tissues, which is critical in managing glaucoma [17]. The synergistic effects of these compounds offer a comprehensive approach to eye health, targeting oxidative stress, inflammation, and IOP reduction (Figure 2). Their combination could enhance current treatments, providing a multi-faceted defense against progressive eye diseases, making it a compelling candidate for further research and development in ocular health supplements.
This review explores the scientific foundation for the synergy between EGCG and forskolin and assesses the benefits and safety of incorporating them into vision and eye health supplements.
2. EGCG Biochemical Properties and Health Effects
2.1. Antioxidant
EGCG is well-known for its strong antioxidant effects. By neutralizing reactive oxygen species (ROS), EGCG helps mitigate oxidative stress, a key factor in the development of various organ pathologies, including ocular diseases like glaucoma, age-related macular degeneration (AMD), and diabetic retinopathy [18]. In addition to its antioxidative properties, EGCG has significant anti-inflammatory effects by inhibiting key mediators such as NF-κB and COX-2, reducing chronic inflammation in eye diseases [19].
2.2. Neuroprotection
EGCG also offers remarkable neuroprotective properties. It protects mitochondria from dysfunction [20], a crucial factor in glaucoma where mitochondrial impairment plays a pivotal role [21]. EGCG reduces neuroinflammation, enhancing communication between the nervous and immune systems, thus preserving neurological functions [22]. It also inhibits the aggregation of neurotoxic proteins like amyloid-beta and tau, known to contribute to conditions such as Alzheimer’s disease, thus protecting against neural degeneration [23].
2.3. Antitumoral
The therapeutic potential of EGCG was first identified in cancer research, where it demonstrated significant anti-cancer properties through multiple mechanisms. It inhibits cancer cell growth by inducing programmed cell death (apoptosis) and halting cell proliferation. EGCG suppresses angiogenesis, essential for tumor growth, by downregulating vascular endothelial growth factor (VEGF) and angiogenic factors [24, 25]. Additionally, EGCG modulates matrix metalloproteinases (MMPs), enzymes responsible for degrading the extracellular matrix, thereby preventing cancer cell invasion and metastasis [26]. EGCG also enhances the efficacy of cancer therapies by overcoming drug resistance, making it a potent candidate for combination treatments [27].
EGCG affects several cellular signaling pathways, such as PI3K/Akt, MAPK, and NF-κB, which are critical for cancer cell survival and proliferation [28, 29]. Furthermore, it exerts epigenetic modifications, including DNA methylation and histone modification, to activate tumor suppressor genes and inhibit oncogenes [30]. In estrogen-dependent cancers like breast cancer, EGCG modulates estrogen receptor signaling [31]. It has also been shown to inhibit androgen receptors in prostate cancer, thus reducing cancer cell proliferation and promoting apoptosis [32]. Additionally, EGCG targets lung cancer by inhibiting the cell cycle and metastasis [33], and exhibits similar anti-proliferative effects in colorectal cancer [34] and leukemia [35, 36]. Overall, EGCG’s multi-targeted approach, influencing both signaling pathways and gene expression, highlights its broad therapeutic potential in cancer management.
2.4. Metabolism and Weight Management
EGCG shares with forskolin an interesting role in metabolism and weight management, which may also benefit ocular health by reducing the risk of related eye conditions. EGCG enhances fat oxidation, converting stored fat into energy. Studies show that consuming green tea extract, rich in EGCG, can increase fat oxidation during exercise, leading to weight loss over time [37, 38]. It boosts metabolism by increasing thermogenesis, a process that burns more calories even at rest, especially when combined with caffeine, also present in green tea [39, 40]. Several studies suggest EGCG contributes to modest weight loss, particularly in combination with green tea extract, by increasing fat oxidation and metabolic rate [41, 42]. EGCG specifically targets visceral fat, a harmful fat linked to metabolic diseases such as type 2 diabetes and cardiovascular disease [38, 43].
EGCG may also reduce appetite by influencing gut hormones associated with food intake, supporting weight loss efforts [44, 45]. There is evidence that it increases feelings of fullness, leading to reduced calorie intake [46]. EGCG improves insulin sensitivity, helping regulate blood sugar levels and reducing the risk of metabolic syndrome, a condition characterized by high blood pressure, high blood sugar, excess body fat, and abnormal cholesterol levels [47-49]. EGCG may influence the expression of genes related to obesity [50]. Accordingly, EGCG may inhibit the formation of new fat cells (adipogenesis) while promoting the breakdown of existing fat cells (lipolysis) [51]. Emerging evidence suggests that EGCG combined with caffeine may positively affect the gut microbiota, which plays a role in energy balance, fat storage, and overall metabolism [52]. A healthier gut microbiome is linked to a lower risk of obesity. The anti-inflammatory properties of EGCG may help reduce chronic inflammation associated with obesity and metabolic syndrome, improving metabolic health and reducing weight gain [53].
The role of metabolic health is increasingly recognized as a crucial factor in the onset and progression of retinal diseases, especially diabetic retinopathy and age-related macular degeneration (AMD). Dysregulated glucose metabolism and obesity-related oxidative stress are known contributors to retinal damage. EGCG’s ability to enhance fat oxidation and improve insulin sensitivity may therefore have significant implications for retinal health, as these metabolic improvements can reduce the overall inflammatory and oxidative burden on the retina.
In conclusion, EGCG’s diverse biochemical properties make it a compelling therapeutic agent across multiple diseases, including metabolic syndrome, neurodegenerative conditions, cancers, and – as we will see below – ocular disorders. Its potential to mitigate oxidative stress, inflammation, and cell death highlights its value as a candidate for future treatments targeting complex health challenges.
3. EGCG Efficacy on Retinal Diseases
As metabolic dysfunction plays a central role in several retinal diseases, particularly diabetic retinopathy, the systemic effects of EGCG on glucose regulation and fat metabolism can directly influence retinal health. By improving insulin sensitivity and reducing visceral fat, EGCG may mitigate the chronic inflammation and oxidative stress that contribute to retinal cell damage in these diseases.
In ocular health, EGCG shows continued promise. By protecting retinal ganglion cells from apoptosis, it helps preserve vision in glaucoma. Its ability to modulate cellular signaling pathways supports cell survival and prevents apoptosis, while its anti-angiogenic properties benefit conditions such as diabetic retinopathy and AMD. EGCG’s metal-ion chelation abilities enhance its neuroprotective and antioxidant functions, protecting ocular tissues from oxidative damage caused by metals [54, 55].
3.1. Mitochondrial Dysfunction
Pioneering preclinical studies have demonstrated the efficacy of EGCG (shown to be more potent than Trolox) in treating oxidative eye diseases, particularly glaucoma and age-related macular degeneration (AMD) [56]. These conditions are driven by oxidative stress and mitochondrial dysfunction, which lead to the degeneration of retinal cells, including retinal ganglion cells (RGCs) and photoreceptors [57].
Mitochondrial dysfunction is a key factor in many retinal diseases. Photobiomodulation (PBM) therapy, which uses non-invasive light therapy, has gained attention for its potential to alleviate this dysfunction. PBM enhances mitochondrial function, reduces oxidative stress, and promotes cell survival. For retinal diseases, low-level lasers or light-emitting diodes (LEDs) are typically directed toward the eye in the red or near-infrared spectrum (600 to 1000 nm) for 5 to 30 minutes. PBM sessions can be conducted daily or several times per week, preferably accompanied by protective eyewear [58, 59]. Studies show that PBM promotes mitochondrial biogenesis by increasing the expression of key transcription factors like PGC-1α and NRF-1, enhancing mitochondrial respiration and improving adenosine triphosphate (ATP) production [60, 61]. This effect is crucial in high-energy-demand tissues such as the retina and optic nerve. In conjunction with PBM, EGCG enhances mitochondrial biogenesis and respiratory efficiency [62, 63], synergistically improving ATP production and reducing oxidative stress and ROS levels in ocular tissues, which amplifies the protective benefits against retinal cell apoptosis and neurodegeneration.
3.2. Oxidative Damage
In glaucoma, RGCs are particularly vulnerable to oxidative damage due to their energy demands and reliance on mitochondrial function. Consistently with its strong antioxidant power, EGCG also reduces ischemia-induced oxidative damage in hypertensive glaucoma models, preserving retinal cell function [64], even when given by the oral route [65]. In DR, oxidative stress is exacerbated by hyperglycemia, and EGCG helps neutralize ROS, protecting the retina from further damage [66], also through the activation of PKC as a neuronal survival factor [67]. Additionally, EGCG inhibits COX-2, an enzyme that in AMD promotes inflammation and oxidative damage in retinal pigment epithelial (RPE) cells [68].
3.3. Glutamate Excitotoxicity
EGCG also plays a crucial role in preventing glutamate excitotoxicity in glaucoma by mitigating the harmful effects of excessive glutamate accumulation and the overactivation of NMDA receptors, which lead to calcium overload, oxidative stress, and mitochondrial dysfunction in retinal ganglion cells (RGCs). Glutamate excitotoxicity triggers a domino effect, where dying RGCs release more glutamate into the extracellular space, further overstimulating adjacent RGCs and exacerbating cell death through a bystander effect [69]. Additionally, EGCG helps regulate calcium homeostasis, thus preventing mitochondrial dysfunction, and enhances the clearance of excess glutamate by supporting glial cell function [70]. Through these mechanisms, EGCG interrupts the cycle of RGC degeneration, potentially halting the progression of glaucoma and protecting against vision loss [71]. Moreover, glutamate excitotoxicity is not limited to glaucoma. It plays a role in several neurodegenerative diseases, including Multiple Sclerosis (MS) [72], Alzheimer’s [73], Parkinson’s [74], and amyotrophic lateral sclerosis (ALS) [75]. Therefore, EGCG could potentially be beneficial also in the treatment of these other neurodegenerative diseases [76-79].
3.4. Inflammation
EGCG also exerts strong anti-inflammatory effects. In glaucoma, it suppresses inflammation by inhibiting key inflammatory pathways such as NF-κB and restoring Th1/Th2 cytokine balance, thus mitigating optic nerve damage [80]. In AMD, it reduces inflammation by downregulating pro-inflammatory cytokines like IL-6 and TNF-α, limiting the damage caused by choroidal neovascularization (CNV) [81]. EGCG’s ability to inhibit JNK and ERK pathways also reduces cell death in AMD [82]. In DR, EGCG inhibits the ROS/TXNIP/NLRP3 inflammasome pathway, preserving retinal cell function by reducing inflammation caused by high glucose levels [83]. This anti-inflammatory action helps to preserve retinal cell function and prevents further degeneration, especially in Müller cells, which play a critical role in maintaining retinal homeostasis.
3.5. Angiogenesis
EGCG’s antiangiogenic properties are crucial in managing conditions like AMD and DR, where abnormal blood vessel growth contributes to vision loss. Angiogenesis, driven by overexpression of VEGF, is a key factor in these diseases. EGCG inhibits VEGF production by modulating the PI3K/AKT and MAPK/ERK pathways, preventing abnormal blood vessel formation [84, 85]. It also reduces MMP-9 activity, which breaks down the extracellular matrix, further inhibiting angiogenesis in DR [86]. In AMD, EGCG reduces VEGFA expression and downregulates the HIF-1α/VEGF/VEGFR2 pathway, preventing choroidal neovascularization and limiting vision loss [81]. On the other hand, EGCG also exerts neuroprotective effects by activating endothelial nitric oxide synthase (eNOS), enhancing nitric oxide (NO) production, and improving blood perfusion. Through the phosphorylation of pathways like PI3K/Akt, EGCG stimulates eNOS, increasing NO levels and promoting vasodilation, which improves vascular function and blood flow, particularly to neural tissues such as the retina [54]. This enhanced perfusion supports better oxygen and nutrient delivery, crucial in conditions like glaucoma where impaired circulation contributes to retinal ganglion cell (RGC) degeneration. EGCG’s antioxidant properties reduce oxidative stress, protecting eNOS from uncoupling and preventing neuronal damage by scavenging reactive oxygen species (ROS) and maintaining mitochondrial health [87]. In animal models of diabetic retinopathy, EGCG has been shown to reduce blood-retinal barrier breakdown, improve retinal function, and restore the expression of tight junction proteins like claudin-1, which are critical for maintaining the integrity of the blood-retinal barrier [88]. These findings underscore the broad protective effects of EGCG in preventing both vascular and neurodegenerative complications in DR.
3.6. Fibrosis
EGCG’s antiproliferative effects are particularly useful in preventing scarring and fibrosis in conditions like glaucoma filtration surgery. EGCG inhibits myofibroblast transformation, which contributes to fibrosis, by blocking the TGF-β1/Smad signaling pathway [89]. It also prevents the migration and adhesion of RPE cells involved in the formation of epiretinal membranes in AMD, through inhibition of PDGF-beta receptor activity and downstream signaling pathways [90].
3.7. Neuroprotection
EGCG has neuroprotective properties, particularly in inhibiting apoptosis in RGCs and retinal pigment epithelial cells, which is crucial for treating glaucoma and AMD. EGCG protects RGCs from apoptosis caused by ischemia-reperfusion injury and elevated IOP by inhibiting caspase activation, a key apoptosis pathway [64, 91]. EGCG also protects RGCs from damage after optic nerve axotomy by regulating apoptosis-related genes. Axotomy increased pro-apoptotic proteins nNOS and Bax, but EGCG treatment reduced their levels, preventing excessive cell death. EGCG also boosted anti-apoptotic signaling via enhanced activation of ERK 1/2 and Akt pathways. Blocking these pathways weakened EGCG's protective effect, showing its role in promoting RGC survival by inhibiting pro-apoptotic signals and activating cell survival mechanisms [92]. Additionally, EGCG boosts NAD production by activating NMNAT2, an enzyme critical for neuron survival, which further protects RGCs from degeneration [93, 94]. In AMD, EGCG reduces UVB-induced apoptosis in retinal pigment epithelial cells by modulating the JNK and ERK pathways [82].
3.8. Autophagy
EGCG also restores autophagic activity, which is impaired in retinal diseases like AMD and glaucoma. Autophagy dysfunction in AMD leads to the accumulation of cellular debris in RPE cells, contributing to oxidative damage. EGCG restores autophagic flux by modulating the mTOR pathway, preventing apoptosis in RPE cells [95]. In glaucoma, EGCG enhances autophagic activity in Tenon’s fibroblasts, reducing fibrosis after glaucoma filtration surgery by promoting autophagosome formation and reducing p62 levels [96]. Autophagy is also disrupted in DR due to hyperglycemia. In diabetic conditions, autophagic dysfunction leads to the accumulation of cellular debris, promoting inflammation and cell death. EGCG restores autophagic activity by enhancing autophagosome formation and lysosomal function, protecting Müller cells and retinal pigment epithelial (RPE) cells from apoptosis. This mechanism is crucial in preventing retinal neurodegeneration, which is a key feature in the early stages of DR [97]. By reestablishing proper autophagy, EGCG prevents the buildup of toxic cellular components, thereby protecting retinal cells from damage.
3.9. AGEs
In addition to its antiangiogenic, antioxidant, and autophagy-regulating properties, EGCG inhibits the aldose reductase enzyme, a key player in the polyol pathway, which is activated under hyperglycemic conditions. This pathway contributes to the formation of advanced glycation end-products (AGEs), which exacerbate oxidative stress and inflammation. EGCG’s inhibition of aldose reductase reduces the accumulation of AGEs and minimizes their damaging effects on retinal cells [98]. This is particularly important as aldose reductase activity is a major contributor to microvascular complications in DR, including retinal damage.
3.10. Trabecular Meshwork
Another mechanism explaining the role of EGCG in glaucoma is linked to its effects on trabecular meshwork (TM) cells. It is known that dysfunction of TM cells increases the resistance to AH outflow, leading to IOP elevation. This dysfunction is linked to endoplasmic reticulum (ER) stress in TM cells [99]. EGCG has been shown to reduce ER stress, also including human and porcine TM cells exposed in vitro to tunicamycin, a compound that induces ER stress [100]. The results of this study showed that EGCG improved cell viability and significantly reduced the expression of key ER stress markers (ATF4, HSPA5, and DDIT3) in both human and porcine TM cells. These findings suggest that EGCG could protect TM cells from ER stress, offering a potential therapeutic approach to control IOP in glaucoma patients.
3.11. Clinical Evidence
Clinical studies support EGCG’s efficacy in improving retinal function in glaucoma patients. In a randomized, placebo-controlled trial, oral EGCG supplementation increased pattern-evoked electroretinogram (PERG) amplitudes in open-angle glaucoma patients, indicating improved retinal function [101]. Another study demonstrated that green tea extract and EGCG significantly reduced IOP in healthy volunteers [102], likely due to its relaxing effect on the trabecular meshwork [100].
Overall, EGCG’s antioxidant, anti-inflammatory, neuroprotective, and antiangiogenic properties make it a promising treatment for oxidative and inflammatory eye diseases like glaucoma, AMD, and DR (Figure 3). Its ability to be administered orally with minimal side effects, along with ongoing improvements in its delivery methods, underscores its potential as a safe and effective option for preserving vision and preventing disease progression.
4. Forskolin Biochemical Properties and Health Effects
Forskolin is a natural compound extracted from the roots of the Coleus forskohlii plant, a member of the mint family. Its primary mechanism of action involves activating adenylate cyclase, an enzyme that converts ATP to cyclic AMP (cAMP). Forskolin has been traditionally used in Ayurvedic medicine and has recently gained popularity as a dietary supplement for its various health benefits [103, 104].
4.1. Metabolism and Weight Management
Like EGCG, forskolin aids in weight loss but through different mechanisms. It stimulates cAMP production within adipose cells, promoting fat breakdown and boosting metabolism. Increased cAMP levels activate protein kinase A (PKA), which phosphorylates hormone-sensitive lipase (HSL) and other enzymes involved in lipolysis, breaking down stored fat into free fatty acids and glycerol. This contributes to fat loss and increases metabolic rate [105, 106]. Some clinical studies have shown that forskolin can reduce body fat, particularly in men [107]. In a study of overweight and obese men, forskolin reduced body fat percentage and fat mass while maintaining muscle mass. The study also suggested that forskolin might increase testosterone levels, benefiting muscle mass, energy levels, and hormonal balance, likely due to its role in cAMP elevation [108].
4.2. Cardiovascular Health
Forskolin has traditionally been used to manage cardiovascular health. It may lower blood pressure by relaxing blood vessels and improving blood flow, which can benefit those with hypertension [109]. Additionally, forskolin may enhance heart muscle contractility, useful for individuals with certain heart conditions, as cAMP plays a role in heart muscle contraction [110].
4.3. Respiratory Disorders
Regarding respiratory disorders, forskolin may help alleviate asthma by relaxing lung muscles, leading to bronchodilation, improving breathing, and reducing asthma attacks [111]. It may also reduce respiratory inflammation, benefiting people with asthma and other inflammatory conditions [112]. Furthermore, forskolin’s anti-inflammatory properties may reduce chronic inflammation, which is linked to various health issues [113]. Additionally, forskolin has antioxidant properties that help protect cells from oxidative damage [104].
4.4. Diabetes
In diabetes, forskolin may improve insulin sensitivity and regulate blood sugar levels. Some studies suggest it could aid in glucose metabolism, potentially benefiting people with type 2 diabetes [114, 115]. Preliminary research also indicates that forskolin may have anti-cancer properties, inhibiting the growth of certain cancer cells, though further research is needed to confirm its role in cancer treatment [116, 117].
4.5. Neuroprotection
Forskolin has demonstrated significant neuroprotective properties, primarily through the receptor-independent activation of adenylyl cyclase and subsequent elevation of cAMP levels. The increase in cAMP initiates downstream signaling pathways, including the activation of PKA and cAMP response element-binding protein (CREB), which is crucial for neuronal survival, differentiation, and plasticity [118, 119]. Forskolin enhances the expression of neurotrophic factors such as brain-derived neurotrophic factor (BDNF) and its receptor TrkB, promoting retinal ganglion cell (RGC) survival and neuronal resilience in neurodegenerative models [120, 121].
4.6. Antinflamatory
Forskolin’s anti-inflammatory effects, combined with its ability to mitigate oxidative stress, make it a promising candidate for neurodegenerative diseases like multiple sclerosis (MS), Alzheimer’s, and Parkinson’s disease [122-124].
These multifaceted actions suggest that forskolin not only preserves neuronal integrity but also enhances the brain’s repair mechanisms, offering potential benefits in both acute and chronic neurodegenerative conditions.
5. Forskolin Efficacy on Retinal Diseases
Forskolin has shown promising therapeutic potential in the treatment of glaucoma and other retinal diseases, primarily through its effects on intraocular pressure (IOP) and neuroprotection.
5.1. IOP
In glaucoma, forskolin may be beneficial by reducing intraocular pressure (IOP), potentially preventing optic nerve damage and vision loss [125]. Elevated cAMP levels promote two primary effects: they stimulate the active resorption of aqueous humor (AH) from the posterior chamber into the stroma [126], and cause relaxation of the ciliary muscle along with the trabecular meshwork [127]. This enhances AH outflow and effectively lowers intraocular pressure (IOP), a critical factor in glaucoma management [128, 129]. Additionally, forskolin has neuroprotective effects by enhancing cAMP levels, which support neuronal survival and function, providing a safeguard against neurodegeneration in retinal ganglion cells in glaucoma [130]. In fact, forskolin induces on one hand the synthesis and the expression of BDNF by astrocytes [131] and endothelial cells [132] lining RGCs. On the other hand, it enhances the translocation of the cognate receptor TrkB to the cell membrane of RGCs [133].
5.2. Neuroprotection
Although topical administration of forskolin is well-studied in preclinical models, oral delivery may offer broader therapeutic benefits [125]. Numerous experimental studies suggest forskolin as a neuroprotective agent due to its ability to reduce intraocular pressure (IOP) in both animal models and humans [126, 134]. Its protective effects on retinal ganglion cells (RGCs) in glaucoma models are also well-documented [118], with indirect neuroprotection arising from its IOP-lowering capacity. In a double-blind, randomized clinical trial, patients with primary open-angle glaucoma treated with 1% forskolin eye drops (administered three times daily for 4 weeks) exhibited significant IOP reduction [135], possibly due to forskolin's ability to reduce aqueous humor accumulation [136]. In another clinical study, a dietary supplement containing forskolin lowered IOP and improved pattern electroretinogram amplitude in glaucoma patients, suggesting enhanced RGC function or survival [137]. Forskolin’s neuroprotective effects are partially attributed to its role in boosting neurotrophin activity. Meyer-Franke et al. demonstrated that forskolin, when combined with brain-derived neurotrophic factor (BDNF), ciliary-derived neurotrophic factor (CTNF), and insulin-like growth factor-1 (IGF-1), increased RGC survival in vitro [118]. Similarly, in a feline model, forskolin augmented RGC survival when used alongside BDNF and CTNF following axonal injury [138]. Animal studies also support these findings. Dietary supplementation with forskolin, homotaurine, spearmint, and B vitamins protected RGCs from degeneration in rodent models of optic nerve injury [139] and glaucoma [140]. This combination decreased inflammation and apoptotic markers, preserving visual function without altering IOP in glaucomatous models [140].
Moreover, forskolin has also shown promising therapeutic potential in the treatment of glaucoma and other retinal diseases, primarily through its effects on oxidative stress, inflammation, blood perfusion and angiogenesis, and glucose metabolism.
5.3. Oxidative Stress and Inflammation
Forskolin’s antioxidant properties further protect ocular tissues from oxidative stress-induced damage, a common pathway in diseases like AMD and diabetic retinopathy [104]. Forskolin also helps reduce inflammation, which is a contributing factor in retinal diseases [141]. By lowering the levels of inflammatory factors such as ICAM-1 and TNF-α, and decreasing the number of adherent leukocytes in retinal microvasculature, forskolin mitigates retinal inflammation, especially in diabetic conditions [113, 142, 143]. This reduction in inflammation protects retinal cells from further damage, particularly in retinal ganglion cells (RGCs), where forskolin’s neuroprotective effects play a crucial role [144].
5.4. Blood Perfusion and Angiogenesis
Forskolin promotes vasodilation by increasing cyclic AMP (cAMP) and cyclic GMP (cGMP) levels, improving blood flow and supporting the delivery of oxygen and nutrients to the retina [145]. This is particularly beneficial in conditions like AMD, where blood supply to the retina is often impaired. Additionally, forskolin inhibits angiogenesis, which is the abnormal formation of new blood vessels that contributes to vision loss in proliferative diabetic retinopathy and neovascular AMD [146]. In a preclinical experimental setting, it was investigated whether forskolin, a protein kinase A (PKA) agonist, affects toll-like receptor 4 (TLR4) signaling and retinal endothelial cell (REC) permeability in high glucose conditions [147]. The results showed that forskolin restored the levels of tight junction proteins (ZO-1 and occludin) and improved REC permeability. Both forskolin and TLR4 inhibition reduced the high glucose-induced increase in REC permeability, though their actions were not synergistic. Forskolin influenced both MyD88-dependent and -independent TLR4 signaling pathways independently of Epac1. These findings suggest that targeting PKA or TLR4 could offer new therapeutic approaches for retinal vascular conditions. This may help reduce fluid accumulation and retinal edema. Diabetic retinopathy can involve the formation of new, abnormal blood vessels (neovascularization). If forskolin can influence pathways related to angiogenesis (the formation of new blood vessels), it could potentially play a role in preventing or reducing this process. Indeed, a study involving a cancer model showed that forskolin's activation of the cAMP/PKA pathway could have an inhibitory effect on vascular endothelial growth factor (VEGF) production, a key driver of retinal angiogenesis in pathologies like diabetic retinopathy [148].
5.5. Glucose Metabolism
In diabetic retinopathy, forskolin plays a role in modulating glucose metabolism, which is crucial for preventing retinal damage. Studies have shown that forskolin can reduce retinal glucose concentrations by lowering the expression of glucose transporter 1, a protein that mediates glucose uptake into cells. This leads to a decrease in retinal glucose levels and reduces inflammation associated with diabetic conditions [149]. Forskolin’s ability to restore tight junction proteins and improve retinal endothelial cell permeability further supports retinal health by preventing fluid leakage and retinal edema, common issues in diabetic retinopathy.
Improving glucose metabolism is critical in the prevention and treatment of diabetic retinopathy, as prolonged hyperglycemia accelerates retinal damage through multiple pathways. Forskolin’s ability to regulate glucose levels and enhance retinal endothelial cell function offers a dual benefit of addressing both systemic metabolic dysregulation and localized retinal health. This metabolic regulation may provide a therapeutic advantage in managing the progression of retinal diseases.
Collectively, these biochemical properties position forskolin as a valuable molecule for developing treatments aimed at reducing IOP, protecting against oxidative and inflammatory damage, and preserving neuronal health in various eye diseases (Figure 4).
6. Predicted Cooperative Effects of EGCG and Forskolin
Although no experimental data have been published so far, the predicted cooperative effects of epigallocatechin gallate (EGCG) and forskolin in the treatment of glaucoma and other ophthalmic diseases stem from their complementary biochemical properties and mechanisms of action. EGCG, known for its potent antioxidant and anti-inflammatory properties, effectively reduces oxidative stress and inflammation, which are key contributors to the pathogenesis of several retinal diseases, including glaucoma, diabetic retinopathy, and age-related macular degeneration (AMD). In addition to reducing oxidative damage, EGCG’s neuroprotective capabilities are crucial in preserving retinal ganglion cells (RGCs), which helps prevent optic nerve damage and vision loss in glaucoma.
Forskolin, on the other hand, acts primarily by activating adenylate cyclase, an enzyme that increases cyclic AMP (cAMP) levels. This leads to enhanced aqueous humor outflow and a consequent reduction in intraocular pressure (IOP), a key therapeutic target in glaucoma management. By lowering IOP, forskolin directly addresses one of the main risk factors for optic nerve damage and disease progression in glaucoma.
When combined, EGCG and forskolin offer a multifaceted approach to ocular treatment. EGCG’s neuroprotective and anti-inflammatory actions help enhance the overall health of ocular tissues, protecting against retinal degeneration and inflammation, while forskolin’s IOP-lowering effect specifically targets glaucoma’s root cause. Furthermore, both compounds exhibit anti-inflammatory and anti-angiogenic properties, which can be particularly beneficial in managing diabetic retinopathy and AMD, where abnormal blood vessel growth and inflammation exacerbate retinal damage.
The synergistic effects of EGCG and forskolin, therefore, hold the potential to not only slow disease progression but also improve overall ocular health by addressing multiple pathogenic mechanisms. Through their combined antioxidant, anti-inflammatory, neuroprotective, and IOP-lowering actions, these compounds can offer a comprehensive therapeutic strategy for retinal diseases.
Additionally, considering the well-established link between metabolic dysfunction and retinal diseases, the systemic effects of both EGCG and forskolin—especially their roles in glucose metabolism and weight management—add another layer of therapeutic potential. By improving metabolic health, these compounds could reduce the overall oxidative and inflammatory burden on the retina, potentially slowing the progression of conditions such as diabetic retinopathy and AMD. This predicted cooperative interaction between EGCG and forskolin suggests a promising avenue for future research and development of comprehensive treatment strategies for glaucoma and other ophthalmic diseases, leveraging the strengths of both compounds to achieve superior clinical outcomes.
7. Formulation Issues
Despite their efficacy, the oral bioavailability of both EGCG (epigallocatechin gallate) and forskolin faces significant challenges due to their poor solubility, limited permeability, and instability in the gastrointestinal tract [150]. EGCG, being a polyphenolic compound, suffers from poor aqueous solubility and is further hindered by its large molecular size and hydrophilicity, which restrict its absorption. It is also highly susceptible to degradation in the gut environment [151]. Encapsulation techniques, nanoparticles, and co-administration with substances like piperine or phospholipids have been explored to improve EGCG’s bioavailability [152, 153]. Similarly, forskolin, a labdane diterpene, shares the problem of limited solubility and permeability, exacerbated by its hydrophobic nature and susceptibility to enzymatic breakdown [151]. To improve the bioavailability of these compounds, several advanced formulation strategies have been explored [154]. Nanoparticle-based delivery systems can enhance their solubility and stability, promoting better absorption. Self-emulsifying drug delivery systems (SEDDS) can improve both solubility and permeability by increasing lipophilicity and facilitating intestinal transport. Complexation with suitable carriers is another strategy that can boost both solubility and stability, potentially improving the systemic exposure of these compounds [154]. Lipid nanoparticle encapsulation also enhances ocular retention, making it a promising topical delivery method for retinal diseases [155].
Taken together, these methods could significantly enhance the bioavailability of EGCG and forskolin, and co-administration may offer synergistic therapeutic effects in treating various conditions.
In summary, the bioavailability of EGCG and forskolin is hindered by their poor solubility, permeability, and stability. Nanoparticles, SEDDS, and complexation are promising strategies to improve their oral bioavailability and might allow their co-administration, potentially offering synergistic benefits.
8. Conclusions
EGCG and forskolin show considerable promise as therapeutic agents for retinal diseases due to their combined antioxidant, anti-inflammatory, neuroprotective, and IOP-lowering properties. The available preclinical evidence, particularly in animal models and in vitro studies, supports their potential to mitigate the progression of conditions such as glaucoma, AMD, and DR. However, despite the strong foundation of preclinical data, there is a notable lack of robust clinical studies confirming these effects in humans. This gap in clinical evidence limits the translation of these findings into practical treatments.
The existing literature highlights several bioavailability challenges, particularly regarding the oral administration of both compounds, which may reduce their effectiveness in clinical settings. Advanced formulation strategies, such as encapsulation and nanoparticle-based delivery, should be explored further to improve systemic exposure and therapeutic efficacy.
Given the significant body of preclinical research, the field would benefit from well-designed clinical trials that explore the safety and efficacy of EGCG and forskolin, both individually and in combination. Such trials could validate their therapeutic potential in humans and clarify their roles as adjuncts or alternatives to conventional treatments for retinal diseases. The promising synergistic effects of these two compounds, targeting multiple pathways involved in retinal disease progression, provide a compelling rationale for their combined use in future clinical studies. These trials would be crucial in determining whether the substantial benefits observed in preclinical models can be replicated in human patients.
In conclusion, while the preclinical evidence is strong, the need for clinical validation is pressing. The published reports should serve as a call to action for initiating clinical trials that assess the real-world impact of these supplements, potentially optimizing their use through combination therapies for retinal health.
Funding
This research received no external funding.
Data Availability Statement
No new data were created or analyzed in this study.
Acknowledgments
Critical proofreading of the manuscript was done with the aid of ChatGPT.
Conflicts of Interest
The author declares no conflicts of interest.
References
- Ahmad, A. , Ahsan, H. Biomarkers of inflammation and oxidative stress in ophthalmic disorders. J Immunoassay Immunochem. 2020, 41, 257–271, Epub 2020 Feb 11. [Google Scholar] [CrossRef] [PubMed]
- Böhm EW, Buonfiglio F, Voigt AM, Bachmann P, Safi T, Pfeiffer N, Gericke, A. Oxidative stress in the eye and its role in the pathophysiology of ocular diseases. Redox Biol. 2023, 68, 102967, Epub 2023 Nov 18. [CrossRef] [PubMed]
- Gupta N, Ang LC, Noël de Tilly L, Bidaisee L, Yücel YH. Human glaucoma and neural degeneration in intracranial optic nerve, lateral geniculate nucleus, and visual cortex. Br J Ophthalmol. 2006, 90, 674–8, Epub 2006 Feb 7. [CrossRef] [PubMed]
- Baudouin C, Kolko M, Melik-Parsadaniantz, S. , Messmer EM. Inflammation in Glaucoma: From the back to the front of the eye, and beyond. Prog Retin Eye Res. 2021, 83, 100916, Epub 2020 Oct 17. [Google Scholar] [CrossRef] [PubMed]
- Jadeja RN, Martin PM. Oxidative Stress and Inflammation in Retinal Degeneration. Antioxidants (Basel). 2021, 10, 790. [Google Scholar] [CrossRef] [PubMed]
- Maiuolo J, Bulotta RM, Oppedisano F, Bosco F, Scarano F, Nucera S, Guarnieri L, Ruga S, Macri R, Caminiti R, Musolino V, Gliozzi M, Carresi C, Cardamone A, Coppoletta A, Nicita M, Carnevali A, Scorcia V, Mollace, V. Potential Properties of Natural Nutraceuticals and Antioxidants in Age-Related Eye Disorders. Life (Basel). 2022, 13, 77. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Lawler T, Liu Y, Christensen K, Vajaranant TS, Mares, J. Dietary Antioxidants, Macular Pigment, and Glaucomatous Neurodegeneration: A Review of the Evidence. Nutrients. 2019, 11, 1002. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Kushwah N, Bora K, Maurya M, Pavlovich MC, Chen, J. Oxidative Stress and Antioxidants in Age-Related Macular Degeneration. Antioxidants (Basel). 2023, 12, 1379. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Williams M, Hogg RE, Chakravarthy, U. Antioxidants and diabetic retinopathy. Curr Diab Rep. 2013, 13, 481–7. [Google Scholar] [CrossRef] [PubMed]
- She C, Shang F, Cui M, Yang, X. , Liu, N. Association between dietary antioxidants and risk for diabetic retinopathy in a Chinese population. Eye (Lond). 2021, 35, 1977–1984, Epub 2020 Oct 2. [Google Scholar] [CrossRef] [PubMed]
- Lin, C. , Wu, X. Curcumin Protects Trabecular Meshwork Cells From Oxidative Stress. Invest Ophthalmol Vis Sci. 2016, 57, 4327–32. [Google Scholar] [CrossRef] [PubMed]
- Tourtas T, Birke MT, Kruse FE, Welge-Lüssen UC, Birke, K. Preventive effects of omega-3 and omega-6 Fatty acids on peroxide mediated oxidative stress responses in primary human trabecular meshwork cells. PLoS One, 2012; 7, e31340, Epub 2012 Feb 3. [CrossRef] [PubMed]
- Lawrenson JG, Evans JR. Omega 3 fatty acids for preventing or slowing the progression of age-related macular degeneration. Cochrane Database Syst Rev. 2015, 2015, CD010015. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Fathima S, Prokopiou E, Georgiou, T. Omega-3 Polyunsaturated Fatty Acids and Their Anti-Oxidant, Anti-Inflammatory and Neuroprotective Effects in Diabetic Retinopathy: A Narrative Review. Front Biosci (Landmark Ed). 2023, 28, 153. [Google Scholar] [CrossRef] [PubMed]
- Allegrini D, Raimondi R, Borgia A, Sorrentino T, Montesano G, Tsoutsanis P, Cancian G, Verma Y, De Rosa FP, Romano MR. Curcumin in Retinal Diseases: A Comprehensive Review from Bench to Bedside. Int J Mol Sci. 2022, 23, 3557. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Li J, Du L, He JN, Chu KO, Guo CL, Wong MOM, Pang CP, Chu WK. Anti-inflammatory Effects of GTE in Eye Diseases. Front Nutr. 2021, 8, 753955. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Wagh VD, Patil PN, Surana SJ, Wagh KV. Forskolin: upcoming antiglaucoma molecule. J Postgrad Med. 2012, 58, 199–202. [Google Scholar] [CrossRef] [PubMed]
- Mokra, D. , Joskova, M., Mokry, J. Therapeutic Effects of Green Tea Polyphenol (‒)-Epigallocatechin-3-Gallate (EGCG) in Relation to Molecular Pathways Controlling Inflammation, Oxidative Stress, and Apoptosis. Int J Mol Sci. 2022, 24, 340. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Kim SR, Seong KJ, Kim WJ, Jung JY. Epigallocatechin Gallate Protects against Hypoxia-Induced Inflammation in Microglia via NF-κB Suppression and Nrf-2/HO-1 Activation. Int J Mol Sci. 2022, 23, 4004. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Oliveira MR, Nabavi SF, Daglia M, Rastrelli L, Nabavi SM. Epigallocatechin gallate and mitochondria. A story of life and death. Pharmacol Res. 2016, 104, 70–85, Epub 2015 Dec 29. [Google Scholar] [CrossRef] [PubMed]
- Catalani E, Brunetti K, Del Quondam, S. , Cervia, D. Targeting Mitochondrial Dysfunction and Oxidative Stress to Prevent the Neurodegeneration of Retinal Ganglion Cells. Antioxidants (Basel). 2023, 12, 2011. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Payne A, Nahashon S, Taka E, Adinew GM, Soliman KFA. Epigallocatechin-3-Gallate (EGCG): New Therapeutic Perspectives for Neuroprotection, Aging, and Neuroinflammation for the Modern Age. Biomolecules. 2022, 12, 371. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Afzal O, Dalhat MH, Altamimi ASA, Rasool R, Alzarea SI, Almalki WH, Murtaza BN, Iftikhar S, Nadeem S, Nadeem MS, Kazmi, I. Green Tea Catechins Attenuate Neurodegenerative Diseases and Cognitive Deficits. Molecules. 2022, 27, 7604. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Rodriguez SK, Guo W, Liu L, Band MA, Paulson EK, Meydani, M. Green tea catechin, epigallocatechin-3-gallate, inhibits vascular endothelial growth factor angiogenic signaling by disrupting the formation of a receptor complex. Int J Cancer. 2006, 118, 1635–44. [Google Scholar] [CrossRef] [PubMed]
- Leong H, Mathur PS, Greene GL. Green tea catechins inhibit angiogenesis through suppression of STAT3 activation. Breast Cancer Res Treat. 2009, 117, 505–15, Epub 2008 Sep 28. [Google Scholar] [CrossRef] [PubMed]
- Tanabe H, Suzuki T, Ohishi T, Isemura, M. , Nakamura, Y., Unno, K. Effects of Epigallocatechin-3-Gallate on Matrix Metalloproteinases in Terms of Its Anticancer Activity. Molecules. 2023, 28, 525. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Aggarwal V, Tuli HS, Tania M, Srivastava S, Ritzer EE, Pandey A, Aggarwal D, Barwal TS, Jain A, Kaur G, Sak K, Varol M, Bishayee, A. Molecular mechanisms of action of epigallocatechin gallate in cancer: Recent trends and advancement. Semin Cancer Biol. 2022, 80, 256–275, Epub 2020 May 24. [Google Scholar] [CrossRef] [PubMed]
- Singh BN, Shankar, S. , Srivastava RK. Green tea catechin, epigallocatechin-3-gallate (EGCG): mechanisms, perspectives and clinical applications. Biochem Pharmacol. 2011, 82, 1807–21, Epub 2011 Jul 30. [Google Scholar] [CrossRef] [PubMed]
- Talib WH, Awajan D, Alqudah A, Alsawwaf R, Althunibat R, Abu AlRoos M, Al Safadi A, Abu Asab S, Hadi RW, Al Kury LT. Targeting Cancer Hallmarks with Epigallocatechin Gallate (EGCG): Mechanistic Basis and Therapeutic Targets. Molecules. 2024, 29, 1373. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Li F, Qasim S, Li D, Dou QP. Updated review on green tea polyphenol epigallocatechin-3-gallate as a cancer epigenetic regulator. Semin Cancer Biol. 2022, 83, 335–352, Epub 2021 Jan 13. [Google Scholar] [CrossRef] [PubMed]
- Marín V, Burgos V, Pérez R, Maria DA, Pardi, P. , Paz, C. The Potential Role of Epigallocatechin-3-Gallate (EGCG) in Breast Cancer Treatment. Int J Mol Sci. 2023, 24, 10737. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Kumar M, Verma S, Rawat S, Dhatwalia SK. Exploring integrative approaches: EGCG’s potential in combating prostate cancer. WCRJ, 2744. [CrossRef]
- Sehgal A, Bhat AM, Dogra D, Rawat, S. , Dhatwalia SK. EGCG: The antioxidant powerhouse in lung cancer management and chemotherapy enhancement. Advances in Redox Research, 2023, 9, 100085. [CrossRef]
- Luo KW, Xia J, Cheng BH, Gao HC, Fu LW, Luo XL. Tea polyphenol EGCG inhibited colorectal-cancer-cell proliferation and migration via downregulation of STAT3. Gastroenterol Rep (Oxf). 2020, 9, 59–70. [CrossRef] [PubMed] [PubMed Central]
- Wang J, Xie Y, Feng Y, Zhang L, Huang X, Shen X, Luo, X. (-)-Epigallocatechin gallate induces apoptosis in B lymphoma cells via caspase-dependent pathway and Bcl-2 family protein modulation. Int J Oncol. 2015, 46, 1507–15, Epub 2015 Feb 3. [Google Scholar] [CrossRef] [PubMed]
- Della Via FI, Shiraishi RN, Santos I, Ferro KP, Salazar-Terreros MJ, Franchi Junior GC, Rego EM, Saad STO, Torello CO. (-)-Epigallocatechin-3-gallate induces apoptosis and differentiation in leukaemia by targeting reactive oxygen species and PIN1. Sci Rep. 2021, 11, 9103. [CrossRef] [PubMed] [PubMed Central]
- Hodgson AB, Randell RK, Jeukendrup AE. The effect of green tea extract on fat oxidation at rest and during exercise: evidence of efficacy and proposed mechanisms. Adv Nutr. 2013, 4, 129–40. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Li F, Gao C, Yan P, Zhang M, Wang Y, Hu Y, Wu X, Wang, X. , Sheng, J. EGCG Reduces Obesity and White Adipose Tissue Gain Partly Through AMPK Activation in Mice. Front Pharmacol. 2018, 9, 1366. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Gosselin, C. , Haman, F. Effects of green tea extracts on non-shivering thermogenesis during mild cold exposure in young men. Br J Nutr. 2013, 110, 282–8, Epub 2012 Dec 14. [Google Scholar] [CrossRef] [PubMed]
- Katada S, Yanagimoto A, Matsui Y, Hibi M, Osaki N, Kobayashi, S. , Katsuragi, Y. Effect of tea catechins with caffeine on energy expenditure in middle-aged men and women: a randomized, double-blind, placebo-controlled, crossover trial. Eur J Nutr. 2020, 59, 1163–1170, Epub 2019 May 6. [Google Scholar] [CrossRef] [PubMed]
- Bose M, Lambert JD, Ju J, Reuhl KR, Shapses SA, Yang CS. The major green tea polyphenol, (-)-epigallocatechin-3-gallate, inhibits obesity, metabolic syndrome, and fatty liver disease in high-fat-fed mice. J Nutr. 2008, 138, 1677–83. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- James, A. , Wang, K., Wang, Y. Therapeutic Activity of Green Tea Epigallocatechin-3-Gallate on Metabolic Diseases and Non-Alcoholic Fatty Liver Diseases: The Current Updates. Nutrients. 2023, 15, 3022. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Choi C, Song HD, Son Y, Cho YK, Ahn SY, Jung YS, Yoon YC, Kwon SW, Lee YH. Epigallocatechin-3-Gallate Reduces Visceral Adiposity Partly through the Regulation of Beclin1-Dependent Autophagy in White Adipose Tissues. Nutrients. 2020, 12, 3072. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Song WY, Aihara Y, Hashimoto T, Kanazawa, K. , Mizuno, M. (-)-Epigallocatechin-3-gallate induces secretion of anorexigenic gut hormones. J Clin Biochem Nutr. 2015, 57, 164–9, Epub 2015 Sep 1. [Google Scholar] [CrossRef] [PubMed]
- Li H, Kek HC, Lim J, Gelling RW, Han, W. Green tea (-)-epigallocatechin-3-gallate counteracts daytime overeating induced by high-fat diet in mice. Mol Nutr Food Res. 2016, 60, 2565–2575, Epub 2016 Sep 1. [Google Scholar] [CrossRef] [PubMed]
- Fernandes RC, Araújo VA, Giglio BM, Marini ACB, Mota JF, Teixeira KS, Monteiro PA, Lira FS, Pimentel GD. Acute Epigallocatechin 3 Gallate (EGCG) Supplementation Delays Gastric Emptying in Healthy Women: A Randomized, Double-Blind, Placebo-Controlled Crossover Study. Nutrients. 2018, 10, 1122. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Ma SB, Zhang R, Miao S, Gao B, Lu Y, Hui S, Li L, Shi XP, Wen AD. Epigallocatechin-3-gallate ameliorates insulin resistance in hepatocytes. Mol Med Rep. 2017, 15, 3803–3809, Epub 2017 Apr 7. [Google Scholar] [CrossRef] [PubMed]
- Wu X, Yang M, He Y, Wang F, Kong Y, Ling TJ, Zhang, J. EGCG-derived polymeric oxidation products enhance insulin sensitivity in db/db mice. Redox Biol. 2022, 51, 102259, Epub 2022 Feb 9. [Google Scholar] [CrossRef] [PubMed]
- Legeay S, Rodier M, Fillon L, Faure, S. , Clere, N. Epigallocatechin Gallate: A Review of Its Beneficial Properties to Prevent Metabolic Syndrome. Nutrients. 2015, 7, 5443–68. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Sae-Tan S, Grove KA, Kennett MJ, Lambert JD. (-)-Epigallocatechin-3-gallate increases the expression of genes related to fat oxidation in the skeletal muscle of high fat-fed mice. Food Funct. 2011, 2, 111–6, Epub 2011 Jan 4. [Google Scholar] [CrossRef] [PubMed]
- Söhle J, Knott A, Holtzmann U, Siegner R, Grönniger E, Schepky A, Gallinat S, Wenck H, Stäb F, Winnefeld, M. White Tea extract induces lipolytic activity and inhibits adipogenesis in human subcutaneous (pre)-adipocytes. Nutr Metab (Lond). 2009, 6, 20. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Zhu MZ, Zhou F, Ouyang J, Wang QY, Li YL, Wu JL, Huang JA, Liu ZH. Combined use of epigallocatechin-3-gallate (EGCG) and caffeine in low doses exhibits marked anti-obesity synergy through regulation of gut microbiota and bile acid metabolism. Food Funct. 2021, 12, 4105–4116. [Google Scholar] [CrossRef] [PubMed]
- Nainu F, Frediansyah A, Mamada SS, Permana AD, Salampe M, Chandran D, Emran TB, Simal-Gandara, J. Natural products targeting inflammation-related metabolic disorders: A comprehensive review. Heliyon. 2023, 9, e16919. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Ng TK, Chu KO, Wang CC, Pang CP. Green Tea Catechins as Therapeutic Antioxidants for Glaucoma Treatment. Antioxidants (Basel). 2023, 12, 1320. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Boroughani M, Tahmasbi Z, Heidari MM, Johari M, Hashempur MH, Heydari, M. Potential therapeutic effects of green tea (Camellia sinensis) in eye diseases, a review. Heliyon. 2024, 10, e28829. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Zhang, B. , Osborne NN. Oxidative-induced retinal degeneration is attenuated by epigallocatechin gallate. Brain Res. 2006, 1124, 176–87, Epub 2006 Nov 3. [Google Scholar] [CrossRef] [PubMed]
- Eells, JT. Mitochondrial Dysfunction in the Aging Retina. Biology (Basel). 2019, 8, 31. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Geneva, II. Photobiomodulation for the treatment of retinal diseases: a review. Int J Ophthalmol. 2016, 9, 145–52. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Siqueira, RC. Photobiomodulation Using Light-Emitting Diode (LED) for Treatment of Retinal Diseases. Clin Ophthalmol. 2024, 18, 215–225. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Zhang CX, Lou Y, Chi J, Bao XL, Fan, B. , Li GY. Considerations for the Use of Photobiomodulation in the Treatment of Retinal Diseases. Biomolecules. 2022, 12, 1811. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Valter K, Tedford SE, Eells JT, Tedford CE. Photobiomodulation use in ophthalmology - an overview of translational research from bench to bedside. Front Ophthalmol (Lausanne). 2024, 4, 1388602. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Sommer AP, Zhu, D. Green tea and red light--a powerful duo in skin rejuvenation. Photomed Laser Surg. 2009, 27, 969–71. [Google Scholar] [CrossRef] [PubMed]
- Sommer AP, Bieschke J, Friedrich RP, Zhu D, Wanker EE, Fecht HJ, Mereles D, Hunstein, W. 670 nm laser light and EGCG complementarily reduce amyloid-β aggregates in human neuroblastoma cells: basis for treatment of Alzheimer's disease? Photomed Laser Surg. 2012, 30, 54–60, Epub 2011 Oct 26. [CrossRef] [PubMed]
- Zhang B, Safa R, Rusciano D, Osborne NN. Epigallocatechin gallate, an active ingredient from green tea, attenuates damaging influences to the retina caused by ischemia/reperfusion. Brain Res. 2007, 1159, 40–53, Epub 2007 May 26. [CrossRef] [PubMed]
- Zhang B, Rusciano D, Osborne NN. Orally administered epigallocatechin gallate attenuates retinal neuronal death in vivo and light-induced apoptosis in vitro. Brain Res. 2008, 1198, 141–52, Epub 2007 Dec 15. [Google Scholar] [CrossRef] [PubMed]
- Du J, Wang Y, Tu Y, Guo Y, Sun X, Xu X, Liu X, Wang L, Qin, X. , Zhu, M., Song, E. A prodrug of epigallocatechin-3-gallate alleviates high glucose-induced pro-angiogenic factor production by inhibiting the ROS/TXNIP/NLRP3 inflammasome axis in retinal Müller cells. Exp Eye Res. 2020, 196, 108065, Epub 2020 May 12. [Google Scholar] [CrossRef] [PubMed]
- Mandel SA, Avramovich-Tirosh Y, Reznichenko L, Zheng H, Weinreb O, Amit T, Youdim MB. Multifunctional activities of green tea catechins in neuroprotection. Modulation of cell survival genes, iron-dependent oxidative stress and PKC signaling pathway. Neurosignals. [CrossRef] [PubMed]
- Chan CM, Huang JH, Lin HH, Chiang HS, Chen BH, Hong JY, Hung CF. Protective effects of (-)-epigallocatechin gallate on UVA-induced damage in ARPE19 cells. Mol Vis. 2008, 14, 2528–34. [Google Scholar] [PubMed] [PubMed Central]
- Boccuni, I. , Fairless, R. Retinal Glutamate Neurotransmission: From Physiology to Pathophysiological Mechanisms of Retinal Ganglion Cell Degeneration. Life (Basel). 2022, 12, 638. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Chen F, Jiang L, Shen C, Wan H, Xu L, Wang N, Jonas JB. Neuroprotective effect of epigallocatechin-3-gallate against N-methyl-D-aspartate-induced excitotoxicity in the adult rat retina. Acta Ophthalmol. 2012, 90, e609–e615, Epub 2012 Sep 13. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L. , Chen, F., Wang, N. Neuroprotective Effects of Epigallocatechin-3-gallate against N-methyl-D-aspartate Induced Excitotoxicity in Rat Retina. Investigative Ophthalmology & Visual Science. 2012, 53, 6574. [Google Scholar]
- Ellwardt, E. , Zipp, F. Molecular mechanisms linking neuroinflammation and neurodegeneration in MS. Exp Neurol. 2014; 262 Pt A:8-17, Epub 2014 Feb 14. [Google Scholar] [CrossRef] [PubMed]
- Wang, R. , Reddy PH. Role of Glutamate and NMDA Receptors in Alzheimer's Disease. J Alzheimers Dis. 2017, 57, 1041–1048. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Iovino L, Tremblay ME, Civiero, L. Glutamate-induced excitotoxicity in Parkinson's disease: The role of glial cells. J Pharmacol Sci. 2020, 144, 151–164, Epub 2020 Aug 1. [Google Scholar] [CrossRef] [PubMed]
- Spalloni A, Nutini M, Longone, P. Role of the N-methyl-d-aspartate receptors complex in amyotrophic lateral sclerosis. Biochim Biophys Acta. 2013, 1832, 312–22, Epub 2012 Nov 29. [Google Scholar] [CrossRef] [PubMed]
- Xu Q, Langley M, Kanthasamy AG, Reddy MB. Epigallocatechin Gallate Has a Neurorescue Effect in a Mouse Model of Parkinson Disease. J Nutr. 2017, 147, 1926–1931, Epub 2017 Aug 23. [Google Scholar] [CrossRef] [PubMed]
- Tang S, Zhang Y, Botchway BOA, Wang, X., Huang, M., Liu, X. Epigallocatechin-3-Gallate Inhibits Oxidative Stress Through the Keap1/Nrf2 Signaling Pathway to Improve Alzheimer Disease. Mol Neurobiol. 2024; Epub ahead of print. [CrossRef] [PubMed]
- Koh SH, Lee SM, Kim HY, Lee KY, Lee YJ, Kim HT, Kim J, Kim MH, Hwang MS, Song C, Yang KW, Lee KW, Kim SH, Kim OH. The effect of epigallocatechin gallate on suppressing disease progression of ALS model mice. Neurosci Lett. 2006, 395, 103–7, Epub 2005 Dec 13. [Google Scholar] [CrossRef] [PubMed]
- Schuldesz AC, Tudor R, Nandarge PS, Elagez A, Cornea A, Ion R, Bratosin, F. , Prodan, M., Simu, M. The Effects of Epigallocatechin-3-Gallate Nutritional Supplementation in the Management of Multiple Sclerosis: A Systematic Review of Clinical Trials. Nutrients. 2024, 16, 2723. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Zhang WH, Chen, Y. , Gao LM, Cao YN. Neuroprotective role of epigallocatechin-3-gallate in acute glaucoma via the nuclear factor-κB signalling pathway. Exp Ther Med. 2021, 22, 1235, Epub 2021 Aug 31. [Google Scholar] [CrossRef] [PubMed]
- Xu J, Tu Y, Wang Y, Xu X, Sun X, Xie L, Zhao Q, Guo Y, Gu Y, Du J, Du S, Zhu, M. , Song, E. Prodrug of epigallocatechin-3-gallate alleviates choroidal neovascularization via down-regulating HIF-1α/VEGF/VEGFR2 pathway and M1 type macrophage/microglia polarization. Biomed Pharmacother. 2020, 121, Epub 2019 Nov 25. 109606. [Google Scholar] [CrossRef] [PubMed]
- Cao G, Chen M, Song Q, Liu Y, Xie L, Han Y, Liu Z, Ji, Y. , Jiang, Q. EGCG protects against UVB-induced apoptosis via oxidative stress and the JNK1/c-Jun pathway in ARPE19 cells. Mol Med Rep. 2012, 5, 54–9, Epub 2011 Sep 9. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L. , Zhang ZK, Liang, S. Epigallocatechin-3-gallate protects retinal vascular endothelial cells from high glucose stress in vitro via the MAPK/ERK-VEGF pathway. Genet Mol Res. 2016; 15. [Google Scholar] [CrossRef] [PubMed]
- Blasiak J, Chojnacki J, Szczepanska J, Fila M, Chojnacki C, Kaarniranta, K. , Pawlowska, E. Epigallocatechin-3-Gallate, an Active Green Tea Component to Support Anti-VEGFA Therapy in Wet Age-Related Macular Degeneration. Nutrients. 2023, 15, 3358. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Lee HS, Jun JH, Jung EH, Koo BA, Kim YS. Epigalloccatechin-3-gallate inhibits ocular neovascularization and vascular permeability in human retinal pigment epithelial and human retinal microvascular endothelial cells via suppression of MMP-9 and VEGF activation. Molecules. 2014, 19, 12150–72. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Shankar S, Chen Q, Srivastava RK. Inhibition of PI3K/AKT and MEK/ERK pathways act synergistically to enhance antiangiogenic effects of EGCG through activation of FOXO transcription factor. J Mol Signal. 2008, 3, 7. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Zhang, Z. , Zhang, D. (-)-Epigallocatechin-3-Gallate Inhibits eNOS Uncoupling and Alleviates High Glucose-Induced Dysfunction and Apoptosis of Human Umbilical Vein Endothelial Cells by PI3K/AKT/eNOS Pathway. Diabetes Metab Syndr Obes. Erratum in: Diabetes Metab Syndr Obes. 2020, 13, 2751. 10.2147/DMSO.S274564. 2020, 13, 2495–2504. [Google Scholar] [CrossRef] [PubMed]
- Silva KC, Rosales MA, Hamassaki DE, Saito KC, Faria AM, Ribeiro PA, Faria JB, Faria JM. Green tea is neuroprotective in diabetic retinopathy. Invest Ophthalmol Vis Sci. 2013, 54, 1325–36. [Google Scholar] [CrossRef] [PubMed]
- Lin HL, Qin YJ, Zhang YL, Zhang YQ, Chen YL, Niu YY, Pang CP, Chu WK, Zhang HY. Epigallocatechin-3-gallate (EGCG) inhibits myofibroblast transformation of human Tenon's fibroblasts. Exp Eye Res. 2020, 197, 108119, Epub 2020 Jun 27. [Google Scholar] [CrossRef] [PubMed]
- Chan CM, Huang JH, Chiang HS, Wu WB, Lin HH, Hong JY, Hung CF. Effects of (-)-epigallocatechin gallate on RPE cell migration and adhesion. Mol Vis. 2010, 16, 586–95. [Google Scholar] [PubMed] [PubMed Central]
- Shen C, Chen L, Jiang L, Lai TY. Neuroprotective effect of epigallocatechin-3-gallate in a mouse model of chronic glaucoma. Neurosci Lett. 2015, 600, 132–6, Epub 2015 Jun 5. [Google Scholar] [CrossRef] [PubMed]
- Peng PH, Chiou LF, Chao HM, Lin S, Chen CF, Liu JH, Ko ML. Effects of epigallocatechin-3-gallate on rat retinal ganglion cells after optic nerve axotomy. Exp Eye Res. 2010, 90, 528–34, Epub 2010 Jan 28. [Google Scholar] [CrossRef] [PubMed]
- Fang F, Zhuang P, Feng X, Liu P, Liu D, Huang H, Li L, Chen W, Liu L, Sun Y, Jiang H, Ye J, Hu, Y. NMNAT2 is downregulated in glaucomatous RGCs, and RGC-specific gene therapy rescues neurodegeneration and visual function. Mol Ther. 2022, 30, 1421–1431, Epub 2022 Jan 31. [Google Scholar] [CrossRef] [PubMed]
- Tribble JR, Jöe M, Varricchio C, Otmani A, Canovai A, Habchi B, Daskalakis E, Chaleckis R, Loreto A, Gilley J, Wheelock CE, Jóhannesson G, Wong RCB, Coleman MP, Brancale A, Williams PA. NMNAT2 is a druggable target to drive neuronal NAD production. Nat Commun. Erratum in: Nat Commun. 2024, 15, 8143. 10.1038/s41467-024-52439-7. 2024, 15, 6256. [Google Scholar] [CrossRef] [PubMed]
- Li CP, Yao J, Tao ZF, Li XM, Jiang, Q. , Yan, B. Epigallocatechin-gallate (EGCG) regulates autophagy in human retinal pigment epithelial cells: a potential role for reducing UVB light-induced retinal damage. Biochem Biophys Res Commun. 2013, 438, 739–45, Epub 2013 Jul 31. [Google Scholar] [CrossRef] [PubMed]
- Zhang YL, Zhang YQ, Lin HL, Qin YJ, Zeng J, Chen YL, Niu YY, Pang CP, Chu WK, Zhang HY. Epigallocatechin-3-gallate increases autophagic activity attenuating TGF-β1-induced transformation of human Tenon's fibroblasts. Exp Eye Res. 2021, 204, 108447, Epub 2021 Jan 16. [Google Scholar] [CrossRef] [PubMed]
- Wang L, Sun X, Zhu M, Du J, Xu J, Qin X, Xu X, Song, E. Epigallocatechin-3-gallate stimulates autophagy and reduces apoptosis levels in retinal Müller cells under high-glucose conditions. Exp Cell Res. 2019, 380, 149–158, Epub 2019 Apr 15. [Google Scholar] [CrossRef] [PubMed]
- Sampath, C. , Sang, S. , Ahmedna, M. In vitro and in vivo inhibition of aldose reductase and advanced glycation end products by phloretin, epigallocatechin 3-gallate and [6]-gingerol. Biomed Pharmacother. 2016, 84, 502–513, Epub 2016 Sep 28. [Google Scholar] [CrossRef] [PubMed]
- Kasetti RB, Maddineni P, Millar JC, Clark AF, Zode GS. Increased synthesis and deposition of extracellular matrix proteins leads to endoplasmic reticulum stress in the trabecular meshwork. Sci Rep. 2017, 7, 14951. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Zhou L, He JN, Du L, Ho BM, Ng DS, Chan PP, Tham CC, Pang CP, Chu WK. Epigallocatechin-3-Gallate Protects Trabecular Meshwork Cells from Endoplasmic Reticulum Stress. Oxid Med Cell Longev. 2022, 2022, 7435754. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Falsini B, Marangoni D, Salgarello T, Stifano G, Montrone L, Di Landro S, Guccione L, Balestrazzi E, Colotto, A. Effect of epigallocatechin-gallate on inner retinal function in ocular hypertension and glaucoma: a short-term study by pattern electroretinogram. Graefes Arch Clin Exp Ophthalmol. 2009, 247, 1223–33, Epub 2009 Mar 17. [Google Scholar] [CrossRef] [PubMed]
- Gasiunas, K. , Galgauskas, S. Green tea-a new perspective of glaucoma prevention. Int J Ophthalmol. 2022, 15, 747–752. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Salehi B, Staniak M, Czopek K, Stępień A, Dua K, Wadhwa R, Kumar Chellappan D, Sytar O, Brestic M, Ganesh Bhat, N., et al. The Therapeutic Potential of the Labdane Diterpenoid Forskolin. Applied Sciences. 2019; 9, 4089. [CrossRef]
- Pullaiah, T. (2022). Pharmacology of Coleus forskohlii and Forskolin. In: Forskolin. Springer, Singapore. [CrossRef]
- Schimmel, RJ. Stimulation of cAMP accumulation and lipolysis in hamster adipocytes with forskolin. Am J Physiol. C: Pt 1), 1984; 246(1 Pt 1):C63-8. [Google Scholar] [CrossRef] [PubMed]
- Whitehead A, Krause FN, Moran A, MacCannell ADV, Scragg JL, McNally BD, Boateng E, Murfitt SA, Virtue S, Wright J, Garnham J, Davies GR, Dodgson J, Schneider JE, Murray AJ, Church C, Vidal-Puig A, Witte KK, Griffin JL, Roberts LD. Brown and beige adipose tissue regulate systemic metabolism through a metabolite interorgan signaling axis. Nat Commun. 2021, 12, 1905. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Loftus HL, Astell KJ, Mathai ML, Su XQ. Coleus forskohlii Extract Supplementation in Conjunction with a Hypocaloric Diet Reduces the Risk Factors of Metabolic Syndrome in Overweight and Obese Subjects: A Randomized Controlled Trial. Nutrients. 2015, 7, 9508–22. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Godard MP, Johnson BA, Richmond SR. Body composition and hormonal adaptations associated with forskolin consumption in overweight and obese men. Obes Res. 2005, 13, 1335–43. [Google Scholar] [CrossRef] [PubMed]
- Schlepper M, Thormann J, Mitrovic, V. Cardiovascular effects of forskolin and phosphodiesterase-III inhibitors. Basic Res Cardiol. 1989; 84, Suppl 1, 197–212. [CrossRef] [PubMed]
- Baumann G, Felix S, Sattelberger, U. , Klein, G. Cardiovascular effects of forskolin (HL 362) in patients with idiopathic congestive cardiomyopathy--a comparative study with dobutamine and sodium nitroprusside. J Cardiovasc Pharmacol. 1990, 16, 93–100. [Google Scholar] [CrossRef] [PubMed]
- González-Sánchez R, Trujillo X, Trujillo-Hernández B, Vásquez C, Huerta M, Elizalde, A. Forskolin versus sodium cromoglycate for prevention of asthma attacks: a single-blinded clinical trial. J Int Med Res. 2006, 34, 200–7. [Google Scholar] [CrossRef] [PubMed]
- Ma C, Zou L, Xia Y, Tu Y, Xue D, Yang Y, Liu D, Liu, Y. , Wu, H., Dan, H., You, P. Extracts of Coleus forskohlii relieves cough and asthma symptoms via modulating inflammation and the extracellular matrix. J Cell Biochem. 2019, 120, 9648–9655, Epub 2018 Dec 5. [Google Scholar] [CrossRef] [PubMed]
- Tran QTN, Wong WSF, Chai CLL. Labdane diterpenoids as potential anti-inflammatory agents. Pharmacol Res. 2017, 124, 43–63, Epub 2017 Jul 25. [Google Scholar] [CrossRef] [PubMed]
- Ríos-Silva M, Trujillo X, Trujillo-Hernández B, Sánchez-Pastor E, Urzúa Z, Mancilla E, Huerta, M. Effect of chronic administration of forskolin on glycemia and oxidative stress in rats with and without experimental diabetes. Int J Med Sci. 2014, 11, 448–52. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Chen JY, Peng SY, Cheng YH, Lee IT, Yu YH. Effect of Forskolin on Body Weight, Glucose Metabolism and Adipocyte Size of Diet-Induced Obesity in Mice. Animals (Basel). 2021, 11, 645. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Salzillo A, Ragone A, Spina A, Naviglio, S. , Sapio, L. Forskolin affects proliferation, migration and Paclitaxel-mediated cytotoxicity in non-small-cell lung cancer cell lines via adenylyl cyclase/cAMP axis. Eur J Cell Biol. 2023, 102, 151292, Epub 2023 Jan 27. [Google Scholar] [CrossRef] [PubMed]
- Sapio L, Gallo M, Illiano M, Chiosi E, Naviglio D, Spina A, Naviglio, S. The Natural cAMP Elevating Compound Forskolin in Cancer Therapy: Is It Time? J Cell Physiol. 2017, 232, 922–927, Epub 2016 Nov 20. [Google Scholar] [CrossRef] [PubMed]
- Meyer-Franke A, Kaplan MR, Pfrieger FW, Barres BA. Characterization of the signaling interactions that promote the survival and growth of developing retinal ganglion cells in culture. Neuron. 1995, 15, 805–19. [Google Scholar] [CrossRef] [PubMed]
- Riaz SS, Theofilopoulos S, Jauniaux E, Stern GM, Bradford HF. The differentiation potential of human fetal neuronal progenitor cells in vitro. Brain Res Dev Brain Res. 2004, 153, 39–51. [Google Scholar] [CrossRef] [PubMed]
- Heo H, Yoo M, Han D, Cho Y, Joung, I. , Kwon YK. Upregulation of TrkB by forskolin facilitated survival of MSC and functional recovery of memory deficient model rats. Biochem Biophys Res Commun. 2013, 431, 796–801, Epub 2013 Jan 9. [Google Scholar] [CrossRef] [PubMed]
- Lamas NJ, Johnson-Kerner B, Roybon L, Kim YA, Garcia-Diaz A, Wichterle H, Henderson CE. Neurotrophic requirements of human motor neurons defined using amplified and purified stem cell-derived cultures. PLoS One. Erratum in: PLoS One. 2015, 10, e0119195. 10.1371/journal.pone.0119195. 2014, 9, e110324. [Google Scholar] [CrossRef] [PubMed]
- Karthikeyan B, Harini L, Krishnakumar V, Kannan VR, Sundar K, Kathiresan, T. Insights on the involvement of (-)-epigallocatechin gallate in ER stress-mediated apoptosis in age-related macular degeneration. Apoptosis. 2017, 22, 72–85. [Google Scholar] [CrossRef] [PubMed]
- Owona BA, Zug C, Schluesener HJ, Zhang ZY. Protective Effects of Forskolin on Behavioral Deficits and Neuropathological Changes in a Mouse Model of Cerebral Amyloidosis. J Neuropathol Exp Neurol. 2016, 75, 618–27, Epub 2016 May 31. [Google Scholar] [CrossRef] [PubMed]
- Alharbi M, Alshammari A, Kaur G, Kalra S, Mehan S, Suri M, Chhabra S, Kumar N, Alanazi WA, Alshanwani AR, Al-Ghamdi AH, Narula AS, Kalfin, R. Effect of Natural Adenylcyclase/cAMP/CREB Signalling Activator Forskolin against Intra-Striatal 6-OHDA-Lesioned Parkinson's Rats: Preventing Mitochondrial, Motor and Histopathological Defects. Molecules. 2022, 27, 7951. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Vetrugno M, Uva MG, Russo V, Iester M, Ciancaglini M, Brusini P, Centofanti M, Rossetti LM. Oral administration of forskolin and rutin contributes to intraocular pressure control in primary open angle glaucoma patients under maximum tolerated medical therapy. J Ocul Pharmacol Ther. 2012, 28, 536–41, Epub 2012 Jun 25. [Google Scholar] [CrossRef] [PubMed]
- Caprioli J, Sears M, Bausher L, Gregory, D. , Mead, A. Forskolin lowers intraocular pressure by reducing aqueous inflow. Invest Ophthalmol Vis Sci. 1984, 25, 268–77. [Google Scholar] [PubMed]
- Shen X, Koga T, Park BC, SundarRaj, N. , Yue BYJT. Rho GTPase and cAMP/protein kinase A signaling mediates myocilin-induced alterations in cultured human trabecular meshwork cells. J Biol Chem. 2008, 283, 603–612, Epub 2007 Nov 5. [Google Scholar] [CrossRef] [PubMed]
- Kapoor T, Mehan S, Suri M, Sharma N, Kumar N, Narula AS, Alshammari A, Alasmari AF, Alharbi M, Assiri MA, Kalfin, R. Forskolin, an Adenylcyclase/cAMP/CREB Signaling Activator Restoring Myelin-Associated Oligodendrocyte Destruction in Experimental Ethidium Bromide Model of Multiple Sclerosis. Cells. 2022, 11, 2771. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Wagh V, Kalpana, Inamdar B, Samanta, M. 2009. The effect of forskolin ophthalmic inserts on intraocular pressure in rabbit eyes. Int. J. Pharmacy and Pharmaceutical Sci.
- Shim MS, Kim KY, Ju WK. Role of cyclic AMP in the eye with glaucoma. BMB Rep. 2017, 50, 60–70. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Russo R, Adornetto A, Cavaliere F, Varano GP, Rusciano D, Morrone LA, Corasaniti MT, Bagetta, G. , Nucci, C. Intravitreal injection of forskolin, homotaurine, and L-carnosine affords neuroprotection to retinal ganglion cells following retinal ischemic injury. Mol Vis. 2015, 21, 718–29. [Google Scholar] [PubMed] [PubMed Central]
- Juric DM, Loncar, D., Carman-Krzan, M. Noradrenergic stimulation of BDNF synthesis in astrocytes: mediation via alpha1- and beta1/beta2-adrenergic receptors. Neurochem Int. 2008; 52, 297–306, Epub 2007 Jul 4. [CrossRef] [PubMed]
- Nakahashi T, Fujimura H, Altar CA, Li J, Kambayashi J, Tandon NN, Sun, B. Vascular endothelial cells synthesize and secrete brain-derived neurotrophic factor. FEBS Lett. 2000, 470, 113–7. [Google Scholar] [CrossRef] [PubMed]
- Meyer-Franke A, Wilkinson GA, Kruttgen A, Hu M, Munro E, Hanson MG Jr, Reichardt LF, Barres BA. Depolarization and cAMP elevation rapidly recruit TrkB to the plasma membrane of CNS neurons. Neuron. 1998, 21, 681–93. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Caprioli, J. , Sears, M. Forskolin lowers intraocular pressure in rabbits, monkeys, and man. Lancet. 1983, 1, 958–60. [Google Scholar] [CrossRef] [PubMed]
- Majeed M, Nagabhushanam K, Natarajan S, Vaidyanathan P, Kumar SK. A Double-blind, Randomized Clinical Trial to Evaluate the Efficacy and Safety of Forskolin Eye Drops 1% in the Treatment of Open Angle Glaucoma – A Comparative Study. J Clin Trials. 2014; 4, 184. [CrossRef]
- Rusciano D, Pezzino S, Mutolo MG, Giannotti R, Librando A, Pescosolido, N. Neuroprotection in Glaucoma: Old and New Promising Treatments. Adv Pharmacol Sci. 2017, 2017, 4320408, Epub 2017 Oct 17. [Google Scholar] [CrossRef] [PubMed]
- Sisto D, Lavermicocca N, Errico D, Rusciano D (2013) Oral Administration of Forskolin and Rutin Contributes to Reduce Intraocular Pressure and Improve PERG (Pattern Electroretinogram) Amplitude in Glaucomatous Patients. JSM Biotechnol Bioeng 2, 1036.
- Watanabe M, Tokita Y, Kato M, Fukuda, Y. Intravitreal injections of neurotrophic factors and forskolin enhance survival and axonal regeneration of axotomized beta ganglion cells in cat retina. Neuroscience. 2003; 116, 733–742. [CrossRef] [PubMed]
- Locri F, Cammalleri M, Dal Monte M, Rusciano D, Bagnoli, P. Protective Efficacy of a Dietary Supplement Based on Forskolin, Homotaurine, Spearmint Extract, and Group B Vitamins in a Mouse Model of Optic Nerve Injury. Nutrients. 2019, 11, 2931. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Cammalleri M, Dal Monte M, Amato R, Bagnoli P, Rusciano, D. A Dietary Combination of Forskolin with Homotaurine, Spearmint and B Vitamins Protects Injured Retinal Ganglion Cells in a Rodent Model of Hypertensive Glaucoma. Nutrients. 2020, 12, 1189. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Tavares LP, Negreiros-Lima GL, Lima KM, E Silva PMR, Pinho V, Teixeira MM, Sousa LP. Blame the signaling: Role of cAMP for the resolution of inflammation. Pharmacol Res. 2020, 159, 105030, Epub 2020 Jun 17. [Google Scholar] [CrossRef] [PubMed]
- Suresh V, Deepika G, Bantal V, Beedu SR, Rupula, K. Evaluation of Anti-inflammatory and Anti-nociceptive Potentials of Andrographolide and Forskolin: In vivo Studies. Journal of Biologically Active Products from Nature. 2018; 8, 326–334. [CrossRef]
- Chen, Y. , Eklund KK. Insights image for Forskolin attenuates the NLRP3 inflammasome activation and IL-1β secretion in human macrophages. Pediatr Res. 2019; 86, 785. [Google Scholar] [CrossRef]
- Liu L, Jiang Y, Steinle JJ. Forskolin eye drops improve retinal damage from ischemia/reperfusion. Mol Vis. P: ;27, 365-369. PMCID, 2021; 27, 365–369. [PubMed]
- Neto, M. , Lunardi, C., Rodrigues, G. and Bendhack, L. (2011) Vasodilatation induced by forskolin involves cyclic GMP production. Journal of Biophysical Chemistry, 2, 373-379. [CrossRef]
- Nedvetsky PI, Zhao X, Mathivet T, Aspalter IM, Stanchi F, Metzger RJ, Mostov KE, Gerhardt, H. cAMP-dependent protein kinase A (PKA) regulates angiogenesis by modulating tip cell behavior in a Notch-independent manner. Development. 2016, 143, 3582–3590. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Liu L, Jiang Y, Steinle JJ. Forskolin regulates retinal endothelial cell permeability through TLR4 actions in vitro. Mol Cell Biochem. 2021; 476, 4487–4492. [CrossRef]
- Xue Y, Sun R, Zheng W, Yang, L. , An, R. Forskolin promotes vasculogenic mimicry and invasion via Notch-1-activated epithelial-to-mesenchymal transition in syncytiolization of trophoblast cells in choriocarcinoma. Int J Oncol. 2020, 56, 1129–1139, Epub 2020 Feb 20. [Google Scholar] [CrossRef] [PubMed]
- You ZP, Xiong B, Zhang YL, Shi L and Shi, K. Forskolin attenuates retinal inflammation in diabetic mice. Mol Med Rep. 2018; 17, 2321–2326. [CrossRef]
- Gupta, S. , Kesarla, R., Omri, A. Formulation strategies to improve the bioavailability of poorly absorbed drugs with special emphasis on self-emulsifying systems. ISRN Pharm. 2013, 2013, 848043. [Google Scholar] [CrossRef]
- Chen G, Kang W, Li W, Chen, S. , Gao, Y. Oral delivery of protein and peptide drugs: from non-specific formulation approaches to intestinal cell targeting strategies. Theranostics. 2022, 12, 1419–1439. [Google Scholar] [CrossRef] [PubMed]
- Shi M, Shi YL, Li XM, Yang R, Cai ZY, Li QS, Ma SC, Ye JH, Lu JL, Liang YR, Zheng XQ. Food-grade Encapsulation Systems for (-)-Epigallocatechin Gallate. Molecules. 2018, 23, 445. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Zagury, Y. , Kazir, M., Livney YD. Improved antioxidant activity, bioaccessibility and bioavailability of EGCG by delivery in β-lactoglobulin particles. Journal of Functional Foods. 2019, 52, 121–130. [Google Scholar] [CrossRef]
- Ameta RK, Soni, K. , Bhattarai, A. Recent Advances in Improving the Bioavailability of Hydrophobic/Lipophilic Drugs and Their Delivery via Self-Emulsifying Formulations. Colloids and Interfaces. 2023, 7, 16. [Google Scholar] [CrossRef]
- Fangueiro JF, Andreani T, Fernandes L, Garcia ML, Egea MA, Silva AM, Souto EB. Physicochemical characterization of epigallocatechin gallate lipid nanoparticles (EGCG-LNs) for ocular instillation. Colloids Surf B Biointerfaces. 2014, 123, 452–60, Epub 2014 Sep 28. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Main causes and effects of oxidative damage and inflammation leading to the development of the main retinal diseases.
Figure 1.
Main causes and effects of oxidative damage and inflammation leading to the development of the main retinal diseases.
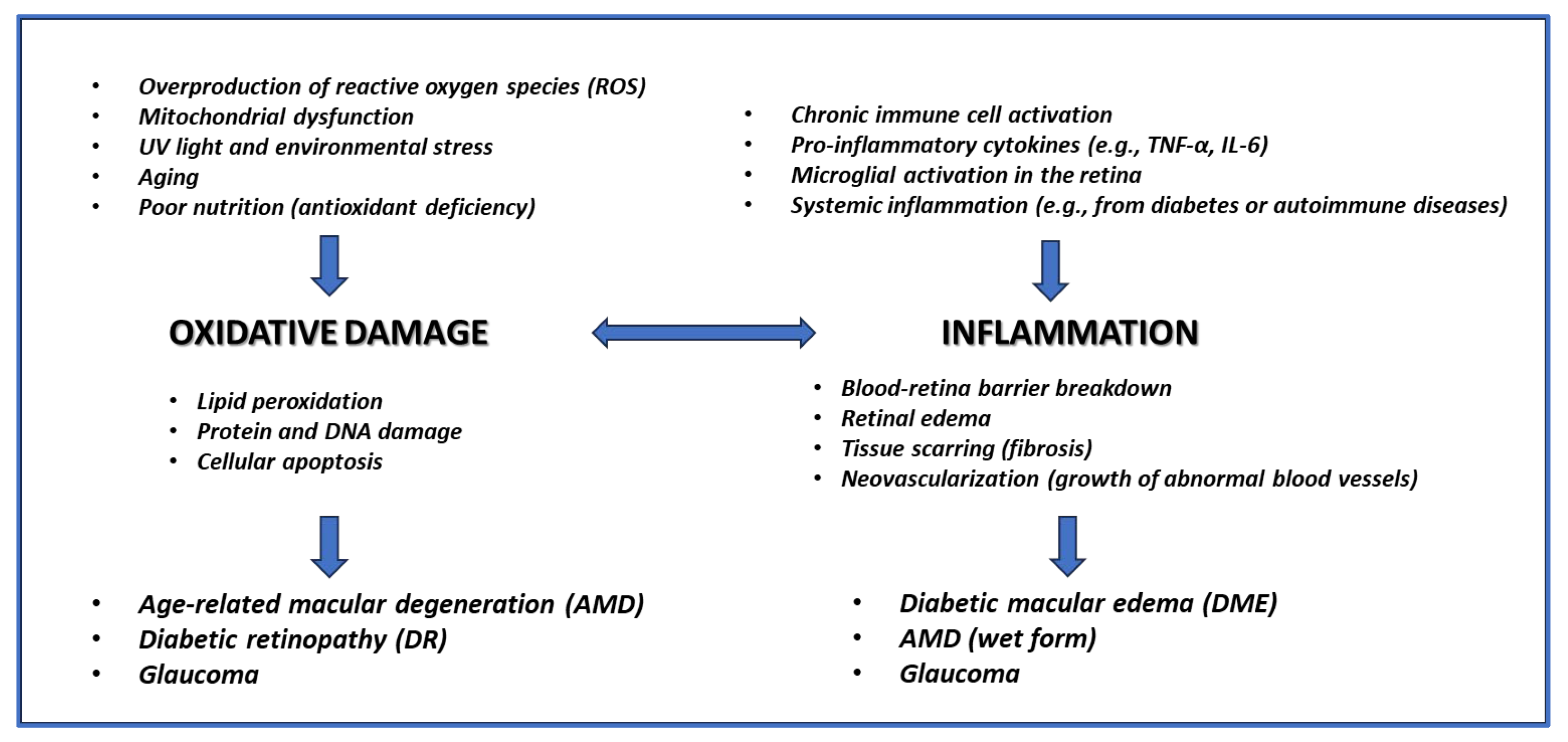
Figure 2.
Chemical structure of EGCG and Forskolin.
Figure 3.
Pleiotropic effects of EGCG (the main catechin present in green tea) relevant to ophthalmic diseases. Related activities are highlighted with the same color: yellow for neuroprotection, pink for vasoactive effects, green for antioxidant activity, light blue for cell survival, light grey for IOP control, dark grey for antifibrotic effect.
Figure 3.
Pleiotropic effects of EGCG (the main catechin present in green tea) relevant to ophthalmic diseases. Related activities are highlighted with the same color: yellow for neuroprotection, pink for vasoactive effects, green for antioxidant activity, light blue for cell survival, light grey for IOP control, dark grey for antifibrotic effect.
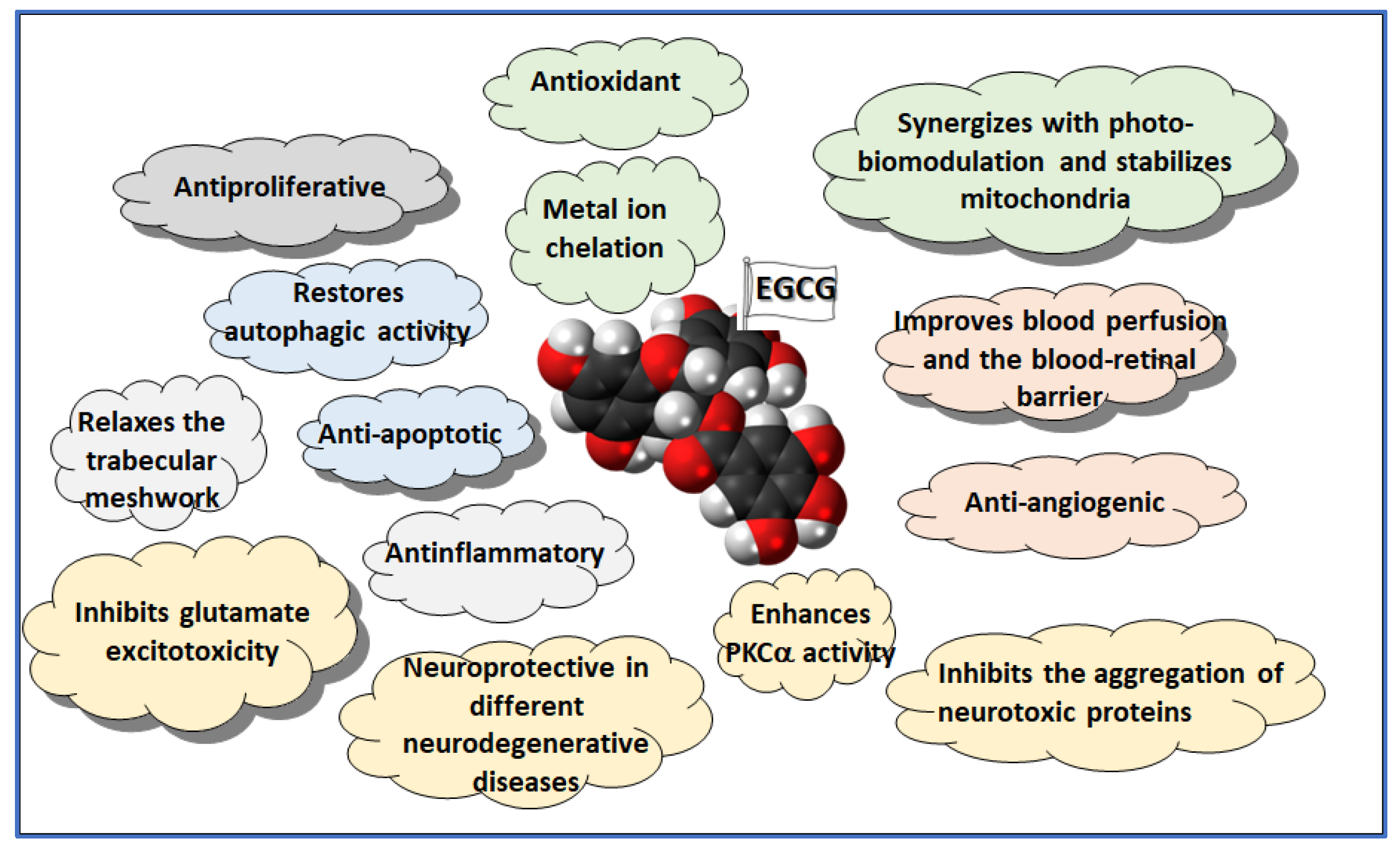
Figure 4.
Multiple effects of forskolin (a receptor-independent activator of adenyl-cyclase) relevant to ophthalmic diseases.
Figure 4.
Multiple effects of forskolin (a receptor-independent activator of adenyl-cyclase) relevant to ophthalmic diseases.
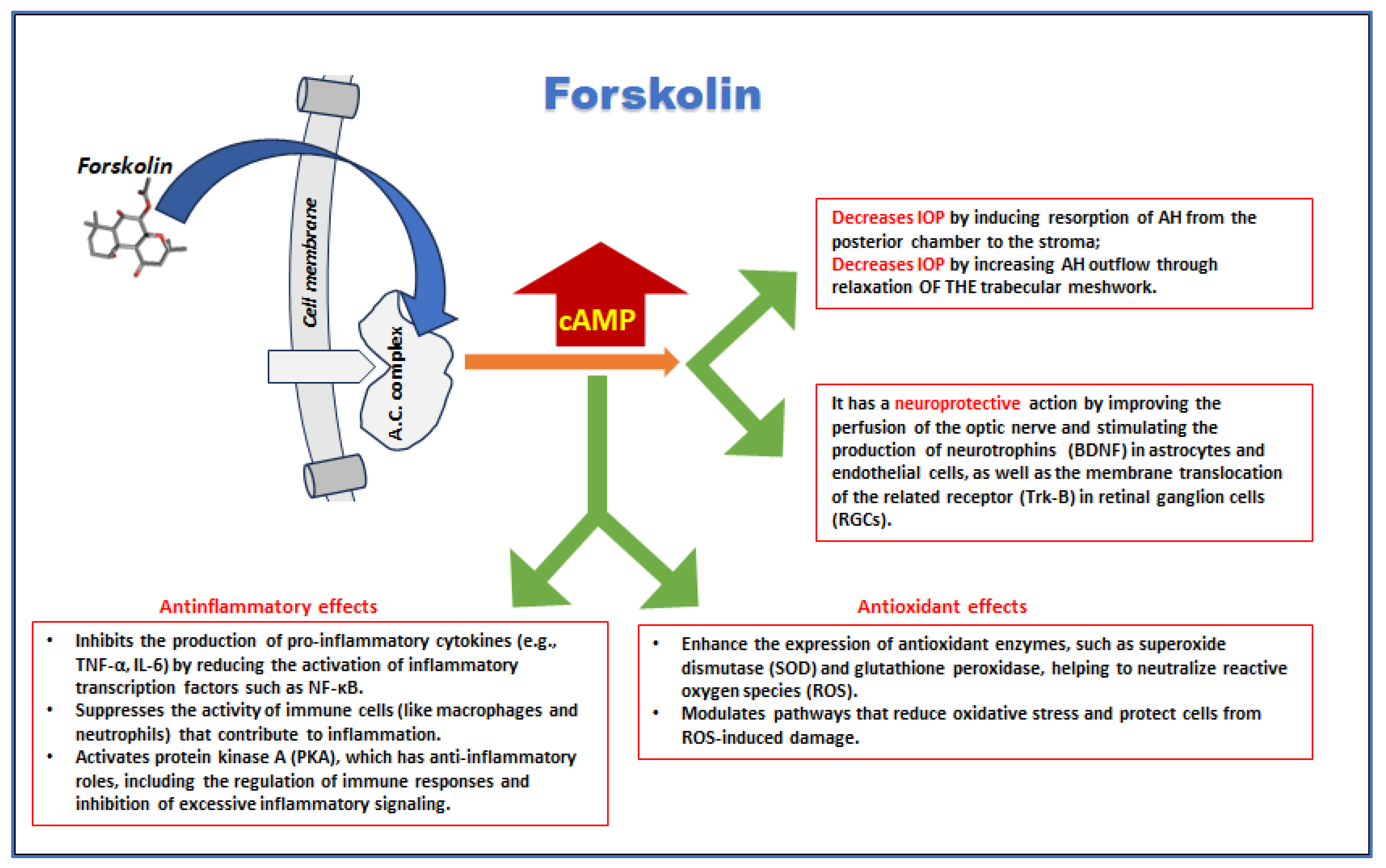
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



