
Submitted:
16 October 2024
Posted:
17 October 2024
You are already at the latest version
Abstract
The rise of non-albicans Candida species, exhibiting unpredictable antifungal resistance, complicates treatment and contributes to the growing threat of invasive, life-threatening infections. This study evaluated the antifungal activity of four benzo[a]phenoxazine derivatives (C34, C35, A42, and A44) against 14 Candida strains following EUCAST standards. Fluconazole interactions were analysed through fractional inhibitory concentration index (FICI) calculation and response surface analysis, based on the Bliss model. Macrophage-like J774A.1 cells were used to assess Candida killing in the presence of synergistic compounds. The MIC values against the different strains varied, with C34 showing the strongest activity, followed by C35, while A42 had the highest MIC values, indicating lower efficacy. However, A42 demonstrated the best synergy with fluconazole against fluconazole-resistant Candida strains. Cytotoxicity assays revealed that the chloropropyl group present in C35 and A42 enhanced cytocompatibility. Co-culture with macrophages showed significant yeast killing for C. albicans and C. auris when fluconazole and A42 were combined, requiring concentration 4- and 16 times lower than their MIC values, enhancing antifungal activity. Given fluconazole's fungistatic nature and the emergence of drug-resistant strains, benzo[a]phenoxazine derivatives' ability to enhance fluconazole's efficacy presents a promising strategy to address antifungal resistance in critical pathogens. These findings align with global research priorities, offering new potential avenues for developing more effective antifungal therapies.
Keywords:
Candida
; benzo[a]phenoxazines
; fluconazole
; synergy
1. Introduction
C. albicans, the most extensively studied Candida species, is a normal colonizer in healthy individuals, typically maintaining commensal interactions with its host [1,2]. However, when its equilibrium with the microbial flora or the host immune system is disrupted, the fungus shifts from a commensal to an opportunistic pathogen [3]. C. albicans causes infections that often target mucosal surfaces but can extend its reach beyond mucosal boundaries, leading to systemic or invasive infections, that are life-threatening. It is estimated that invasive candidiasis lead to more than 450 000 deaths a year [4]. Several Candida species can cause infections in humans, but over 90% of invasive diseases are associated with the same six of them [5]. While C. albicans has traditionally been linked to most of these infections, the incidence of non-albicans Candida strains has been increasing[6,7]. These include C. krusei, C. tropicalis, C. parapsilosis, C. glabrata and C. auris, such as already highlighted in the WHO fungal priority pathogens list [5,8]. The recent rise in non-albicans Candida species adds complexity to treatment, given their unpredictable susceptibility to commonly used antifungals [6,7]. This is the case of C. glabrata, developing resistance to fluconazole and echinocandins during antifungal therapy, or C. krusei, which is intrinsically resistant to fluconazole. Moreover, C. auris is becoming an increasing problem due to its resistant profile to the main classes of antifungal agents [9,10].
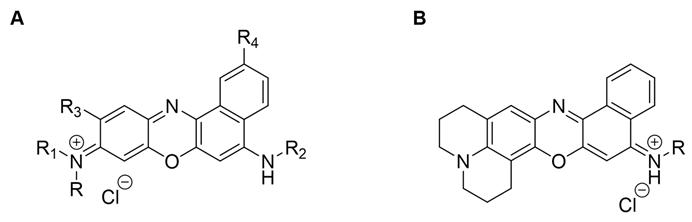
Benzo[a]phenoxazines arise as potential therapeutic approaches, due to their established antifungal activity against S. cerevisiae [11]. These compounds are Nile Blue analogues with different combinations of substituents in specific positions of the polycyclic core. Positions 2, 5 and 9 are the most common for modifications, although position 10 is also explored [12]. The conjugation of different substituents on these positions, either in terms of the type of substituent group (propyl, chloropropyl, isopenthyl, etc.) or the length of carbon chains leads to benzo[a]phenoxazine derivatives with distinct properties. Concerning the antifungal activity, previous works demonstrated that the main players were the substituents on the 5- and 9-positions [11]. In fact, the combination of propyl and dipropyl groups at the amino functions of 5- and 9-positions, respectively - a compound that was named C34 - favoured the antifungal activity against S. cerevisiae, comparing to other benzo[a]phenoxazine derivatives that differed in the length of the aliphatic chains (from 2 to 10 carbon atoms), suggesting an optimal length for the substituents on the mentioned positions [13]. C34 was also preliminary tested against different Candida spp strains, demonstrating high antifungal activity, even against multi-drug-resistant C. auris strains (submitted).
In a previous work, three compounds with different variations of C34’s structure were further tested: C35, with the same di-alkylated propyl substituent at the 9-position but with a chloropropyl group at the 5-position of the heterocycle system [11,14]; A44 and A42, where the free rotation group at 9-position in C34 and C35 was replaced by a rigid structure conferred by a julolidine moiety. Julolidine was chosen, since its cyclic structure is equivalent to the di-alkylated group of both C34 and C35 [15]. Comparing to C34, the alterations performed on C35 and A44 did not improve the antifungal activity of the compounds against S. cerevisiae. On the other hand, when both alterations were present – compound A42 - the antifungal activity was the best of the four, demonstrating that when the julolidine nucleus is present, the compound benefits from a chorine atom as a terminal of the substituent at 14-position [15]. Given the results obtained for S. cerevisiae, this work aims to explore the impact of C34’s alteration on the antifungal activity against Candida spp.
2. Results
2.1. Antifungal Activity of Benzo[a]phenoxazine Derivatives
As mentioned before, within our research group, five benzo[a]phenoxazine derivatives were preliminary tested against species from the genus Candida (submitted). C34 stood out as the compound that yielded better results, highlighting its potential as an antifungal agent. Following this, we decided to use the same approach to evaluate the activity of a set of compounds with structural variations of C34: C35, A44, and A42. C35 has a chloropropyl group at the 5-position replacing propyl, while A44 and A42 feature a rigid structure due to a julolidine moiety, chosen for its structural equivalence to C34’s di-alkylated group (Table 1). The four compounds were tested against strains of C. albicans, C. glabrata, C. parapsilosis, C. tropicalis, C. krusei, C. bracarensis, and C. auris.
After 24 and 48 hours of incubation, growth behaviour of the isolates in the presence of the compounds was analysed through calculation of growth percentage relative to control and determination of the Minimum Inhibitory Concentration (MIC).
Growth percentage was plotted against compound concentration, at both timepoints (Figure 1). Regarding C34, the growth profile was similar for all species tested, except for C. krusei which stood out as the most susceptible species following 24 h of incubation while C. tropicalis appeared to be the most resistant species (Figure 1A). These yeasts behaviour was maintained after 48 h of incubation (Figure 1B). The trend was similar for C35, with C. krusei as the most susceptible species, particularly following 48 h of incubation, followed by C. glabarata and C. bracarensis (Figure 1C,D). Similarly to C34, C. tropicalis remained the most resistant species, but C. albicans SC5314 demonstrated similar resistance (Figure 1D). For A44, Candida strains displayed a more heterogeneous distribution in terms of susceptibility, even among those from the same species. After 24 h of incubation, C. albicans 124A and C. parapsilosis 160a were among the most susceptible, while C. bracarensis 153 MT and both C. glabarata strains stood out as more resistant (Figure 1E). Following 48 h, the trend shows some variations, as C. krusei H11 and both C. albicans strains revealed themselves as more susceptible than the others. C. glabrata as C. bracarensis remained as the most resistant species to this compound (Figure 1F). As for A42, this was the compound to which all species appeared to be, overall, more resistant. C. parapsilosis 160a stood out as the most susceptible and C. bracarensis 153MT as the most resistant (Figure 1G). This pattern was altered after 48h, although C. bracarensis 153MT remained as the most resistant. The susceptibility profile of the other strains/species were highly similar, except for C. krusei, as the most susceptible (Figure 1H).
Overall, C34 and C35 were more effective than A42 and A44, showing stronger inhibitory effects, particularly against C. krusei while C. tropicalis and C. albicans were the most resistant. A44 displayed a more heterogeneous response, with C. albicans 124A and C. parapsilosis 160a being more susceptible, and C. bracarensis 153 MT and C. glabarata strains more resistant. All species appeared resistant to A42.
Figure 2 summarizes the MICs values obtained for each strains-compound combination and Table 2 the associated metrics. The MIC values obtained between replicates varied by no more than one log2 dilution.
After 24 h of incubation and considering all Candida species tested, MIC values were very similar between C34 and C35. For these compounds the values obtained ranged from 3.75 to 15 µM, with MIC geometric means of 10.1 µM and 12.9 µM, respectively. For A44, although the MIC values ranged from 7.5 µM to 30 µM, the MIC geometric mean was very close to the one of C35, with 12.3 µM. On the other hand, A42 displayed the highest MIC values, indicating that the simultaneous addition of a julolidine moiety and a chloropropyl group, did not contribute to an enhancement of the anti-Candida activity (Figure 2 and Table 2).
After 48 h of incubation, minimal deviation was found among C34 and C35 since the MIC values ranged from 7.5 µM to 30 µM in both. Nevertheless, C34 prevailed as the compound with the lowest MIC geometric mean (MIC GM), followed by C35. For both compounds containing the julolidine system (A44 and A42), the MIC values obtained achieved an upper limit of more than 30 µM, with MIC GM close to that value. In this case, it appears that the addition of a rigid structure conferred by a julolidine system decreased the activity of the compounds, comparing to their free-rotation counterparts (Figure 2 and Table 2).
Overall, our results indicated that while the species exhibited varying responses to the compounds, C. krusei was generally the most susceptible, while the most resistant species differed depending on the compound tested. Moreover, C34 was the most promising compound.
To determine the global profile of variation of all the fourteen Candida strains according to the MIC values determined for the four compounds a principal component analysis (PCA) was performed (Figure 3). Variability was explored considering the first and second PCA components, which explained a total of 74% of variability (PC1 – 48 %; PC2 – 26%).
PCA revealed a dispersed arrangement of the strain across the plot. Considering that PC1 explain the higher percentage of variability, a clear separation of both C. krusei strains (Group1) is evident, and C. auris strains together with C. bracarensis NCYC5133 and C. albicans 124A clustered closer (Group 2). Also considering PC1, the remaining strains were positioned close together but a high influence of PC2 separated the strains in two groups, one composed of C. glabrata strains and C. bracarensis 153 MT (Group 3), and the other composed of C. parapsilosis, C. tropicalis and C. albicans SC5314 (Group 4).
Group 1 included the strains with the highest susceptibility to C34 and C35 and also susceptible to A44 at both time points, while group 3 included the strains more resistant to julolidine-based compounds (A44 and A42) and also to C35 at both time points. The remaining groups include strains with a more heterogeneous profiles, but Group 2 included strains that were susceptible to C34 and A44, particularly at 24 h incubation, while Group 4 included stains that were resistant to C34 and C35, particularly at 48 h of incubation.
In accordance with the strains’ distribution, both free-rotation group compounds (C34 and C35) were clustered apart from the julolidine-based compounds (A44 and A42) at both time points and mainly according to PC2. This suggests a positive correlation among all four compounds, while underscoring the impact of varying substituents on their molecular structures. As expected, since no variability was observed with A42 after 48h incubation, this variable was neutral.
2.2. In Vitro Antifungal Activity of Fluconazole and Benzo[a]phenoxazine Derivatives
Fluconazole resistance poses a significant challenge in the management of Candida infections, requiring alternative or combination therapies [10,16,17,18]. To explore potential solutions and knowing that combining antifungal drugs can combat resistant microorganisms synergistically [19,20], we tested C34, C35, A44 and A42 in combination with fluconazole on fluconazole-resistant strains. For that, we selected four isolates of our collection: C. albicans (H65), C. glabrata (PYCC2418), C. krusei (H11) and C. auris (17-274).
The selected isolates presented different degrees of susceptibility to the four compounds, with C. albicans H65 demonstrating the highest resistance to all four compounds (C34, C35, A44, A42) (Figure 4A–D), while resistance to fluconazole varied among the species with C. auris being the most resistant, and C. glabrata the least (Figure 4E).
The interactions between antimicrobial agents can be analysed by calculating FICI values after checkerboard assays. As mentioned before, interactions are interpreted as synergistic if FICI ≤0.5, indifferent if 0.5 < FICI ≤ 4, or antagonistic if FICI > 4. In this study, we investigated the MIC of fluconazole and each compound both alone and in combination against the four resistant Candida species.
The results with C34 revealed a predominantly indifferent interaction with fluconazole across three out of four species, with FICI values ranging from 0.56 to 1.00. Against C. krusei, the MIC of fluconazole decreased from 32 µg/mL to 0.125 µg/mL when combined with C34, but the resulting FICI was 1.00, indicating an indifferent interaction. For C. auris, the interaction was similar, but in C. albicans its was closer to synergism, where the combination reduced the MICs of both drugs, resulting in a FICI of 0.56. On other hand, for C. glabrata, the MIC of fluconazole increased from 16 µg/mL to 64 µg/mL when combined with C34, with FICI exceeding 4.00, suggesting an antagonistic interaction, indicating that the combination of these drugs is less effective than either drug alone (Table 3).
For fluconazole-C35 combination, the results predominantly indicated indifferent interactions, showing FICIs above 0.50. Against C. glabrata – the species for which the combination with C34 resulted in antagonistic interaction – the combination yielded a FICI of 0.56, with the MIC of fluconazole at 32 µg/mL and C35 at 7.5 µM individually, decreasing to 0.125 µg/mL and 3.75 µM, respectively, when combined. However, an exception was observed with C. krusei, where the combination of both drugs resulted in a FICI of 0.50, indicating a synergistic interaction, with the MICs of both drugs being reduced (Table 4).
The combination of fluconazole with A44 yielded a markedly different interaction profile, especially when comparing to its free rotation counterpart (C34). In this case, for C. albicans, C. krusei and C. auris the interactions were synergistic. Against C. albicans the FICI obtained was 0.31, indicating a significant reduction in MIC values when both drugs are used together. Similarly, C. krusei and C. auris showed FICIs of 0.38, highlighting strong synergistic effects. Only for C. glabrata the interaction was indifferent (FICI 0.53) (Table 5).
Remarkably, the combination of fluconazole and A42 resulted in synergistic interactions across all tested Candida species. Against C albicans, a substantial reduction in the MIC values for both drugs were observed, with a FICI of 0.26. Similarly, C. krusei and C. auris displayed FICIs of 0.31, and even C. glabrata, which showed indifferent or antagonistic interaction with the other compounds, exhibited a FICI of 0.38 with the combination with A42 (Table 6).
These results suggest that the combination of fluconazole with julolidine-based compounds, particularly A42, is more efficient than their counterparts, which may provide a highly effective option, resulting in synergy against fluconazole-resistant Candida strains.
While informative, it’s important to note that FICI analysis has some limitations such as being dependent on an inhibition endpoint - in this case, 50%. Therefore, it only considers data corresponding to the MIC, rather than all the data generated. To gain a more comprehensive understanding of these interactions, we used the Response Surface Modelling (RSM) analysis which allows the calculation and visualization of the interaction between drugs, plotting the combined effects on a three-dimensional surface. Since this approach considers all data, it is possible to have a more nuanced interpretation of the relationships between fluconazole and each compound, beyond what is captured by FICI alone.
Regarding the combination of fluconazole and C34, the FICI indicated an indifferent interaction for C. albicans, but the RSM analysis revealed a more complex interaction pattern. While the overall interaction might be indifferent, there are specific concentration ranges where the drugs interact synergistically or antagonistically, as shown by the areas of blue and red on Figure 5, respectively. For C. glabrata, which FICI showed an antagonistic interaction, the RSM analysis demonstrates that antagonism occurs at specific concentration, in accordance, but its effect is not uniform across all concentrations, and there are even regions of weak synergism. For C. krusei, both approaches indicated indifference, and for C. auris, although the overall interaction is indifferent, there are isolated concentrations where antagonism might occur (Figure 5).
When combining fluconazole with C35, the effect on C. albicans was highly similar to the combination with C34 however, despite the FICI suggesting an indifferent interaction, the actual interaction appears more complex. For C. glabrata, the indifferent interaction observed with FICI is established, although there might be minor antagonistic effects. For C. auris, the scenario is inverted: while the overall interaction remains indifferent, some drug concentration ranges begin to interact synergistically, and antagonistic effects are less prominent. Finally, the synergistic interaction observed for C. krusei is supported by this approach (Figure 6).
Fluconazole-A44 combination resulted – according to FICI – in synergistic interaction for all species, except for C. glabrata. This is confirmed by RSM analysis, especially for C. krusei and C. auris, the area of synergy is more prominent that with other compounds. For C. albicans, although there are still specific concentration ranges where the drugs interact antagonistically, the effect is weaker comparing to C34 and C35, and the overall trend is considered synergistic. The interaction of both drugs in C. glabrata is mostly indifferent, but the weak synergy, along with the absence of antagonism holds promise (Figure 7).
Finally, when fluconazole and A42 are combined, the RSM analysis reinforces the FICI results, with synergistic interaction across all Candida species tested. For all four species, the trend is strongly synergistic, even in C. glabrata, confirming that the combination of both drugs works effectively (Figure 8).
While FICI provided a summary measure of interaction, RSM analysis revealed specific concentration-dependent interaction patterns. Therefore, combining these two approaches allows for a better understanding of the complexity of drug interactions and offers a framework for optimizing drug combinations for clinical use.
Moreover, the agreement between FICI and RSM analysis results, points that the compounds containing the julolidine moiety (A44 and A42) lead to a greater synergistic interaction with fluconazole than their free rotation counterparts (C34 and C35).
2.3. Cytotoxicity Assays
Following the assessment of the in vitro interaction of these compounds with fluconazole, it became necessary to assess their cytotoxicity to determine their potential in vivo use and identify the most promising compound for further development. The MTT assay was used to test increasing concentrations of each compound to determine the concentration at which they become cytotoxic for the macrophage-like cell line J774A.1 (Figure 9).
Results showed that, for C34 and A44 the highest non-cytotoxic concentration following a 24 h incubation period was 0.47 µM of compound, with only 0.23 µM maintaining that profile following 48h. For C35, the highest non-cytotoxic concentration was 0.94 µM at 24 h, following which a noticeable cytotoxicity was observed when increasing the concentration of the compound. Remarkably, A42 was non-toxic at concentrations up to1.88 µM at 24 h and at 48 h, the maximum non-toxic concentration was 0.47 µM. These results suggest the incorporation of Cl as a terminal might enhance the cytocompatibility of these compounds with macrophage-like cells. Nevertheless, non-toxic concentrations of all compounds were below Candida MIC values.
Overall, cytotoxicity results highlighted A42 as the least cytotoxic compound. Given that the data on the antifungal interaction with fluconazole also identified this compound as the most promising against resistant Candida species, we proceeded with A42 to the next steps.
2.4. Macrophage Yeast Killing
Integrating all the results, namely FICI and RSM analysis with cytotoxicity, we proceed to evaluate whether the combination of A42 and fluconazole could enhance the antifungal activity of macrophages. Considering the fungal burden in the presence of macrophages without any drug treatment our baseline, we assessed the fungal survival in the presence of the phagocytic cells incubated with fluconazole and A42 individually and in combination, in each of the previously tested resistant Candida species (C. albicans, C. glabrata, C.krusei and C. auris). The concentrations of fluconazole used in this assay were selected for each species as the lower concentrations where the drugs might interact synergistically, as well as considering the highest non-toxic A42 concentration at 24h of incubation. Thus, for C. glabrata fluconazole was tested at 4 µg/mL (MIC 16 µg/mL), C. krusei at 8 µg/mL (MIC 32 µg/mL), C. albicans at 32 µg/mL (MIC 128 µg/mL) and C. auris at 64 µg/mL (MIC > 128 µg/mL). A42 was tested at 1.88 µM with MIC values for all species ≥ to 30 µM.
For C. glabrata and C. krusei, although the presence of fluconazole and A42 alone resulted in a reduction in yeast cell survival, the combination of fluconazole and A42 did not lead to a notable reduction in yeast survival. This indicated that in C. glabrata and C. krusei the combination did not enhance the antifungal activity of macrophages beyond the effect of individual drugs, despite the in vitro indications of synergistic interaction (Figure 10B,C).
Interestingly, C. albicans and C. auris demonstrated a different trend (Figure 10A,D). For C. albicans, the presence of fluconazole alone led to a significant reduction of yeast survival to 26% while with A42 alone, the yeast survival only reduced to 83%. Notably, with the combination of both drugs, the decrease in fungal burden was significant comparing to control and to single-drug treatment groups, leading to only 8% of yeast survival (P <0.0001) (Figure 10A). For C. auris the present of fluconazole alone reduced the yeast survival but only to 68%, however contrarily to C. albicans, the presence of A42 alone did not reduce cell survival. Nevertheless, the combination of both drugs led to significant reduction in yeast survival, to only 14% (P <0.0001) (Figure 10D).
The concentrations of drugs used in this assay were significantly lower than their respective MIC values for both fluconazole and A42 yet, for C. albicans, the fluconazole concentration employed still led to significant growth inhibition (71%) (Table 7). For the other species, the growth inhibition observed was less impressive. For A42, the non-toxic concentration was approximately 16-fold lower than the MIC values obtained for each species and did not achieve similar levels of inhibition. However, when the two compounds were combined, for C. albicans and C. auris, growth inhibition was notably enhanced. For C. albicans, the combination of compounds resulted in 92% growth inhibition, an improvement of 21% compared to the single treatment with fluconazole. For C. auris, the 86% growth inhibition observed with the combination of fluconazole and A42 was particularly remarkable, representing a 54% improvement over the inhibition achieved with fluconazole alone. C. glabrata and C. krusei did not exhibited significant improvement compared single-drug treatment.
Taking this into account, the combination of fluconazole and A42 within the non-toxic concentration limits, offers a promising strategy to enhance macrophage-mediated antifungal activity against C. albicans and C. auris. For C. glabrata and C. krusei, the lack of enhanced activity requires further studies.
3. Discussion
In the present work we evaluated four benzo[a]phenoxazine derivatives against Candida species to evaluate their potential as antimicrobials. C34 and three of its derivatives: C35, with chloropropyl group replacing a propyl at the amino of 5-position of the heterocycle system; A44 and A42, where the free rotation group at 9-position in C34 and C35 was replaced by a rigid structure conferred by a julolidine moiety. The overall range of MIC values was very similar among the four compounds, except for A42, which exhibited the highest MIC values, followed by A44. Compound C34 showed the best antifungal activity with the lowest MIC geometric mean, closely followed by C35. These same compounds had been previously tested against S. cerevisiae and C34 also displayed better activity when compared to C35 and A44 however, A42 emerged as the best compound, indicating that combining the rigid julolidine system in the chloropropyl group enhanced antimicrobial activity [11,15]. In Candida species, the results were quite different. Overall, the species tested appeared to be more resistant to the compounds than S. ceverisiae. Considering all four, the best antifungal activity belonged to C34, with A42 appearing to be the less efective compound, which was not expected.
Combining two drugs in the treatment of infection offers some potential advantages, including the reduction of the dosage of each compound, thereby decreasing their toxicity and broadening the spectrum of cellular targets, which may overpass resistance [19,20]. For example, Caballero et al. (2021) combined isavuconazole and echinocandins against C. auris, suggesting a promising avenue for further exploration [21]. Taking this into account, we selected four fluconazole-resistant Candida to test the combination of this well-known drug with these four compounds. Surprisingly, A42 emerged as the compound with the best performance when in combination, leading to synergistic interactions in all tested species, equally supported by the results obtained with the calculation of the FICI and response surface analysis based on the Bliss model. Recent studies have focused on the interactions of fluconazole with new compounds against Candida species. For example, Kane and co-workers (2024) tested bisphosphonates against different Candida species isolates, along with their interaction with fluconazole, from where a promising combination emerged [22]. Similarly, Albuquerque et al. (2024) explored molecular hybrids of aza-bicyclic 2-isoxazoline-acylhydrazone as anti- Candida agents, revealing one hybrid that – when combined with fluconazole - led to synergistic effects on a fluconazole-resistant Candida strain [23]. These findings underscore the importance of developing new strategies, either through developing novel compounds or exploring their combinations, to improve antifungal activity and, therefore, treatment effectiveness.
To consider the applicability of the benzo[a]phenoxazine derivatives under study, it was necessary to evaluate their cytotoxicity. Results showed that the presence of the chloropropyl group in C35 and A42 appears to be essential for the cytocompatibility of these types of compounds, compared with C34 and A44. Moreover, its combination with the julolidine system further improved the cytocompatibility, highlighting compound A42 as the best to combine with mammalian cells.
Co-culturing of fungi with macrophages in the presence of promising compounds is a common method to evaluate how the compounds influence fungal vulnerability to host immune defenses. In our group, this approach has been used to assess the ability of macrophages to kill Candida cells in the presence of selected phytocompounds, either in their free form or encapsulated [24], with the latter demonstrating superior activity. More recently, Gutierrez-Gongora et al. (2024) [25] used this approach to test mussel extracts in co-cultures of C. neoformans and macrophages, proving their efficacy in reducing fungal survival, both on their own and in combination with fluconazole. In the present work, results obtained with the macrophage yeast killing showed a significant decrease in yeast survival, namely for C. albicans and C. auris, when fluconazole and A42 were used in combination, when compared to their single forms. It is worth noting that the concentration of drugs responsible for these results were 4- and 16-fold lower, respectively, than the MIC value for the species and remained capable of potentiating the antifungal activity of both compounds. Fluconazole is known for its fungistatic nature, paving the way for the development of resistance among isolates [10,26,27]. Efflux pumps, particularly mediated by ATP-binding cassette (ABC) transporters (Cdr1p and Cdr2p) and the major facilitator superfamily (MFS) transporter (Mdr1p) are often associated with azole resistance in C. albicans [28]. Interestingly, Nile Red, also belonging to the benzophenoxazine family, has been reported as a fluorescence substrate of these transporters [29], suggesting that these types of compounds could potentially inhibit them in fluconazole-resistant strains, thereby enhancing the activity of fluconazole, by increasing its intracellular accumulation [26].
Our findings demonstrate that combining fluconazole with benzo[a]phenoxazine derivatives may be effective against fluconazole-resistant Candida species, C. albicans and C. auris, suggesting that these compounds could be a promising strategy for overcoming antifungal resistance. These are the two Candida species among the four ‘critical threat’ pathogens ranked on the WHO report [8], and the research on new and effective antifungal agents, along with possible synergies are aligned with the identified research and development priorities.
4. Materials and Methods
4.1. Fungal Strains and Cell Lines
Fourteen isolates of C. albicans, C. parapsilosis, C. tropicalis, C glabrata, C. krusei, C. bracarensis and C. auris were used in this work. These strains belong to the collection of Biology Department of the University of Minho and were obtained from different sources, identified and conserved in 30% glycerol at -80ºC. When needed, yeast cells were routinely grown at 30 ºC on YPD agar medium (Yeast extract 1%, Peptone, 1%, Glucose 2% and Agar 2%).
For cell culture assays, the murine macrophage-like cell line J774A.1 was used. The cell line was cultured in Dulbecco’s modified Eagle’s medium (DMEM; Gibco) high glucose supplemented with 10% heat-inactivated foetal bovine serum (FBS; Gibco), 2 mM glutamine, 1 mM sodium pyruvate, and 25 mM HEPES buffer, in tissue culture flasks (Nunc) in a humified atmosphere with 5% (v/v) CO2 at 37 °C (Binder CB150).
4.2. Drugs and Medium
Four benzo[a]phenoxazine derivatives were synthetized in the Center of Chemistry of the University of Minho with a purity higher than 95%, as previously described [11,13,15]. Stock solutions were prepared in dimethylsulphoxide (DMSO) to a final concentration of 12 mM. Stock solutions of fluconazole were prepared in DMSO at 25.6 mg/mL. For the susceptibility assays, RPMI 1640 medium (with L-glutamine and a pH indicator but without bicarbonate) was supplemented with glucose to a final concentration of 2% and buffered to pH 7.0 with 0.165 M 3-(N-morpholino)propanesulfonic acid (MOPS) (Sigma-Aldrich). Serial dilutions of all benzo[a]phenoxazine derivatives and fluconazole were performed in supplemented RPMI 1640 medium and added to 96-well plates, to obtain the intended final concentrations.
4.3. Minimum Inhibitory Concentration Assays
In vitro susceptibility of all isolates to the four benzo[a]phenoxazine derivatives was evaluated according to European Committee on Antimicrobial Susceptibility testing (EUCAST) protocols for yeasts (Eucast Definitive Document E.Def 7.3.2).
Susceptibility to benzo[a]phenoxazine derivatives was evaluated by testing concentrations ranging from 0.47 to 30 μM, while for fluconazole the concentration varied between 0.125 to 128 μg/mL. Following incubation on YPD agar medium for 24 hours at 30 ºC, isolates were suspended in sterile water adjusted to a cellular density adjusted of 1-3 x 106 cell/mL. Working concentrations of 0.5-1.5 x 105 cell/mL were used. Each microplate included both sterility and growth controls. After 24 and 48 hours of incubation at 30 ºC, absorbance was measured at 530 nm using a microplate reader. The Minimum Inhibitory Concentration (MIC) of each compound was determined as the lowest drug concentration with growth inhibition of ≥ 50%, in comparison with the growth control.
4.4. Checkerboard Assays – Microplate Preparation
Interaction between fluconazole with each of the benzo[a]phenozaxine derivatives (C34, C35, A44 or A42) followed the same guidelines as the Minimum Inhibitory Concentration (MIC) Assays, adjusted to a broth dilution checkerboard approach [30,31], and following the drug dilution scheme recommended by EUCAST. For that, 96 well flat bottom plates were used. Final concentrations ranged from 0.47 to 30 μM for each one of the benzo[a]phenoxazine derivatives and 0.125 to 128 μg/mL for fluconazole. For microplate preparation, 50 μL of each concentration of C34, C35, A44 or A42 was added into wells 1 to 11 of each column, while 50 μL of each concentration of fluconazole was added into wells A to G of each line. The wells of column 12 and line H contained each compound and fluconazole alone, respectively. Well H12 served as the growth control, containing RPMI with DMSO, but lacking any antifungal drug. The yeast cell suspensions were prepared in sterile water, as previously described, and each well was inoculated with 100 μL of the suspension. For each combination, an uninoculated blank plate, with sterile water instead of cells, was included. Microplates were incubated at 30 ºC and after 24 h of incubation, the absorbance was read at 530 nm. The percentage of growth for each well was calculated by comparison with the growth in the drug-free wells, using a 50% endpoint of growth inhibition, both for drugs tested alone and in combination.
4.4.1. Analysis of Results
MICs that exceeded the highest concentration tested (off-scale MICs) were adjusted to the next highest concentration. To assess drug interaction, the Fractional Inhibitory Concentration Index (FICI) and the Response Surface Modelling (RSM) were calculated and analysed.
4.4.1.1. Fractional Inhibitory Concentration index (FICI)
In all wells corresponding to a MIC, the FICI was calculated as follows:
FICI = FIC A + FIC B = (CA / MIC A) + (CB/ MIC B),
where CA and CB are the concentrations of drugs A and B when used in combination, and MICA and MICB are the concentrations of drugs A and B when used alone. For interpretation, it was considered that: if FICI ≤0.5, the interaction was synergistic; if 0.5 < FICI ≤ 4, the interaction was indifferent; if FICI > 4, the interaction was antagonistic [32].
4.4.1.2. Response Surface Modelling (RSM)
Response Surface Modelling is an approach that allows the use of all data instead of a pre-defined endpoint and has been used for testing drug interactions in yeasts, namely belonging to the genus Candida [30,31,33]. Briefly, experimental data is analysed by expressing the growth percentages for each well and fitting dose-response curves for each drug using a Hill equation. Combined with the Bliss independence model – used to predict drug effects under the assumption of no interaction – allows for the prediction of a theoretical response. Comparing the observed experimental results with the theoretical predictions enables the evaluation of drug interactions and the identification of synergistic or antagonistic effects. Therefore, the data regarding the combinations of fluconazole and the compounds was processed using the Combenefit software, allowing visualization of interaction [34].
4.5. Cytotoxicity Assays
In vitro studies were performed using the murine macrophage-like cell line J774A.1 routinely cultured as previously described [35]. Macrophages were seeded on 96-well tissue culture plates (Nunc) at 1 x 104 cells/well and allowed to adhere overnight at 37 ºC in a 5% CO2 atmosphere. Different concentrations of free compounds were incubated with the cells and cell viability was assessed via the 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide (MTT) assay [36] after 24 and 48 hours of incubation. Results were expressed relative to untreated cells (100% cell viability) [37].
4.6. Macrophage Yeast Killing
To evaluate the impact of free benzo[a]phenoxazine derivatives and fluconazole on macrophage-mediated killing of Candida species a modified protocol was used [38]. Briefly, 1 x 104 macrophages per well were seeded in 96-well plates (Nunc) and incubated overnight at 37 ºC with 5% CO2. Chosen concentrations of free compounds and fluconazole were added. Candida cells (5 x 104 cells/well) were then added and incubated for 24 h at 37 ºC with 5% CO2. After incubation, macrophages were lysed, with 10% saponin solution, and serial dilutions of the lysates were plated on YPD agar. After 24 h of incubation, colony-forming units (CFUs) were enumerated.
4.7. Data Analysis
Data were analysed with ANOVA followed by Šídák’s or Dunnett’s T3 multiple comparisons test, using GraphPad Prism (9.0 version, GraphPad Software, USA). Unless otherwise stated, results shown are from at least three independent experiments with three replicates each, except for in vitro interactions with fluconazole (FICI, RSM, and macrophage yeast killing), where results shown are from two independent experiments, with two to three replicates each. Statistically significant was considered at p-value < 0.05. Principal component analysis (PCA) was performed using Orange data mining suite software (version 3.37.0), to assess the strain variability of Candida species, following the MIC assays. The analysis, visualization and quantification of drug combinations regarding the interaction between fluconazole and each compound were performed with the Combenefit software [34].
Author Contributions
Conceptualization, Methodology and Validation, P. S., M.J.S, M. S. T. G., and M. I. P. Investigation, M. I. P., B. G., P. P. S., and A. C. B. Formal Analysis, and Data Curation, M. I. P., P. P. S., and A.C.B. Writing - Original Draft Preparation, M. I. P. Writing - Review and Editing, P. S., M.J.S, and M. S. T. G. Visualization, M. I. P., P. P. S., and A.C.B. Supervision, P. S., M.J.S, and M. S. T. G. Resources, and Funding Acquisition, P. S., M.J.S, and M. S. T. G. Project Administration, P.S. All authors read and approved the final manuscript.
Funding
This work was supported by FEDER (European Fund for Regional Development)-COMPETEQREN-EU in the framework of the Strategic Funding UIDB/04050/2020 (https://doi.org/10.54499/UIDB/04050/2020) and UID/QUI/00686/2020. FCT is also acknowledged for the Ph.D. scholarships UI/BD/150872/2021 (https://doi.org/10.54499/UI/BD/150872/2021) (M. I. P.), 2020.08235.BD (P. P. S.), SFRH/BD/133513/2017 and COVID/BD/152169/2021 (A. C. B.).
Data Availability
Data Availability:All original data generated during the study are included in the article in the form of figures and tables. For any further inquiries, please contact the corresponding author.
Acknowledgments
We would like to acknowledge the contribution of Bruna Oliveira, Carolina Osório and Ticiana Fernandes for their vital support during the macrophage yeast killing assays.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Chow EWL, Pang LM, Wang Y (2021) From jekyll to hyde: The yeast–hyphal transition of Candida albicans. Pathogens 10:859. [CrossRef]
- Rosenbach A, Dignard D, Pierce J V., et al. (2010) Adaptations of Candida albicans for growth in the mammalian intestinal tract. Eukaryot Cell 9:1075–1086. [CrossRef]
- da Silva Dantas A, Lee KK, Raziunaite I, et al. (2016) Cell biology of Candida albicans–host interactions. Curr Opin Microbiol 34:111–118. [CrossRef]
- Denning DW (2024) Global incidence and mortality of severe fungal disease. Lancet Infect Dis 24:e428–e438. [CrossRef]
- Gómez-Gaviria M, Ramírez-Sotelo U, Mora-Montes HM (2023) Non-albicans Candida Species: Immune Response, Evasion Mechanisms, and New Plant-Derived Alternative Therapies. Journal of Fungi 9:. [CrossRef]
- Logan C, Martin-Loeches I, Bicanic T (2020) Invasive candidiasis in critical care: challenges and future directions. Intensive Care Med 46:2001–2014. [CrossRef]
- Friedman DZP, Schwartz IS (2019) Emerging fungal infections: New patients, new patterns, and new pathogens. Journal of Fungi 5:. [CrossRef]
- 2022; 8. World Health Organization (2022) WHO fungal priority pathogens list to guide research, development and public health action.
- Gómez-Gaviria M, Mora-Montes HM (2020) Current aspects in the biology, pathogeny, and treatment of Candida krusei, a neglected fungal pathogen. Infect Drug Resist 13:1673–1689. [CrossRef]
- Pristov KE, Ghannoum MA (2019) Resistance of Candida to azoles and echinocandins worldwide. Clinical Microbiology and Infection 25:792–798. [CrossRef]
- Leitão MIPS, Raju BR, Cerqueira NMFSA, et al. (2020) Benzo[a]phenoxazinium chlorides: Synthesis, antifungal activity, in silico studies and evaluation as fluorescent probes. Bioorg Chem 98:. [CrossRef]
- Martinez V, Henary M (2016) Nile Red and Nile Blue: Applications and syntheses of structural analogues. Chem Eur J 22:13764–13782. [CrossRef]
- Leitão MIPS, Raju BR, Naik S, et al. (2016) Synthesis and photophysical studies of new benzo[a]phenoxazinium chlorides as potential antifungal agents. Tetrahedron Lett 57:3936–3941. [CrossRef]
- Frade VHJ, Sousa MJ, Moura JCVP, Gonçalves MST (2008) Synthesis of naphtho [2,3-a]phenoxazinium chlorides: Structure-activity relationships of these heterocycles and benzo[a]phenoxazinium chlorides as new antimicrobials. Bioorg Med Chem 16:3274–3282. [CrossRef]
- Raju BR, Leitão MIPS, Sousa MJ, et al. (2020) New NIR dyes based on quinolizino[1,9-hi]phenoxazin-6-iminium chlorides: synthesis, photophysics and antifungal activity. Dyes and Pigments 173:. [CrossRef]
- Farmakiotis D, Kontoyiannis DP (2017) Epidemiology of antifungal resistance in human pathogenic yeasts: current viewpoint and practical recommendations for management. Int J Antimicrob Agents 50:318–324. [CrossRef]
- Arastehfar A, Lass-Flörl C, Garcia-Rubio R, et al. (2020) The quiet and underappreciated rise of drug-resistant invasive fungal pathogens. Journal of Fungi 6:1–34. [CrossRef]
- Arendrup MC, Patterson TF (2017) Multidrug-resistant Candida: Epidemiology, molecular mechanisms, and treatment. Journal of Infectious Diseases 216:S445–S451. [CrossRef]
- Fioriti S, Brescini L, Pallotta F, et al. (2022) Antifungal Combinations against Candida Species: From Bench to Bedside. Journal of Fungi 8:. [CrossRef]
- Carmo A, Rocha M, Pereirinha P, et al. (2023) Antifungals: From Pharmacokinetics to Clinical Practice. Antibiotics 12:. [CrossRef]
- Caballero U, Kim S, Eraso E, et al. (2021) In vitro synergistic interactions of isavuconazole and echinocandins against Candida auris. Antibiotics 10:. [CrossRef]
- Kane A, Dinh H, Campbell L, et al. (2024) Spectrum of activity and mechanisms of azole–bisphosphonate synergy in pathogenic Candida. Microbiol Spectr 12:. [CrossRef]
- de Albuquerque IKP, de Santana DL, de Assis Graciano dos Santos F, et al. (2024) Novel aza-bicyclic 2-isoxazoline acylhydrazone hybrids and their synergistic potential with fluconazole against a drug-resistant Candida albicans strain. Brazilian Journal of Microbiology 55:1811–1816. [CrossRef]
- Miranda-Cadena K, Dias M, Costa-Barbosa A, et al. (2021) Development and Characterization of Monoolein-Based Liposomes of Carvacrol, Cinnamaldehyde, Citral, or Thymol with Anti-Candida Activities. Antimicrobial Agents Chemotherapy 65:e01628-20.
- Gutierrez-Gongora D, Woods M, Prosser RS, Geddes-McAlister J (2024) Natural compounds from freshwater mussels disrupt fungal virulence determinants and influence fluconazole susceptibility in the presence of macrophages in Cryptococcus neoformans. Microbiol Spectr 12:e02841-23. [CrossRef]
- Lu H, Shrivastava M, Whiteway M, Jiang Y (2021) Candida albicans targets that potentially synergize with fluconazole. Crit Rev Microbiol 47:323–337. [CrossRef]
- Berman J, Krysan DJ (2020) Drug resistance and tolerance in fungi. Nat Rev Microbiol 18:319–331. [CrossRef]
- Nishimoto AT, Sharma C, Rogers PD (2020) Molecular and genetic basis of azole antifungal resistance in the opportunistic pathogenic fungus Candida albicans. Journal of Antimicrobial Chemotherapy 75:257–270. [CrossRef]
- Ivnitski-Steele I, Holmes AR, Lamping E, et al. (2009) Identification of Nile red as a fluorescent substrate of the Candida albicans ATP-binding cassette transporters Cdr1p and Cdr2p and the major facilitator superfamily transporter Mdr1p. Anal Biochem 394:87–91. [CrossRef]
- Bidaud AL, Djenontin E, Botterel F, et al. (2020) Colistin interacts synergistically with echinocandins against Candida auris. Int J Antimicrob Agents 55:. [CrossRef]
- Bidaud AL, Botterel F, Chowdhary A, Dannaouia E (2019) In Vitro Antifungal Combination of Flucytosine with Amphotericin B, Voriconazole, or Micafungin against Candida auris Shows No Antagonism. Antimicrob Agents Chemother 63:. [CrossRef]
- Bidaud AL, Schwarz P, Herbreteau G, Dannaoui E (2021) Techniques for the assessment of in vitro and in vivo antifungal combinations. Journal of Fungi 7:1–16. [CrossRef]
- Schwarz P, Nikolskiy I, Bidaud AL, et al. (2022) In Vitro Synergy of Isavuconazole Combined With Colistin Against Common Candida Species. Front Cell Infect Microbiol 12:. [CrossRef]
- Di Veroli GY, Fornari C, Wang D, et al. (2016) Combenefit: An interactive platform for the analysis and visualization of drug combinations. Bioinformatics 32:2866–2868. [CrossRef]
- Sabino R, Sampaio P, Carneiro C, et al. (2011) Isolates from hospital environments are the most virulent of the Candida parapsilosis complex. BMC Microbiol 11:. [CrossRef]
- Madesh M, Balasubramanian KA (1997) A microtiter plate assay for superoxide using MTT reduction method. Indian journal of biochemistry and biophysics 34:535–539.
- Carneiro C, Correia A, Collins T, et al. (2015) DODAB:monoolein liposomes containing Candida albicans cell wall surface proteins: A novel adjuvant and delivery system. European Journal of Pharmaceutics and Biopharmaceutics 89:190–200. [CrossRef]
- McKenzie CGJ, Koser U, Lewis LE, et al. (2010) Contribution of Candida albicans cell wall components to recognition by and escape from murine macrophages. Infect Immun 78:1650–1658. [CrossRef]
Figure 1.
Effect of C34, C35, A44, and A42 concentrations on the growth of fourteen strains of seven species of Candida after 24h hours (left) and 48h (right) of incubation. A and B – C34; C and D – C35; E and F – A44; G and H – A42.
Figure 1.
Effect of C34, C35, A44, and A42 concentrations on the growth of fourteen strains of seven species of Candida after 24h hours (left) and 48h (right) of incubation. A and B – C34; C and D – C35; E and F – A44; G and H – A42.
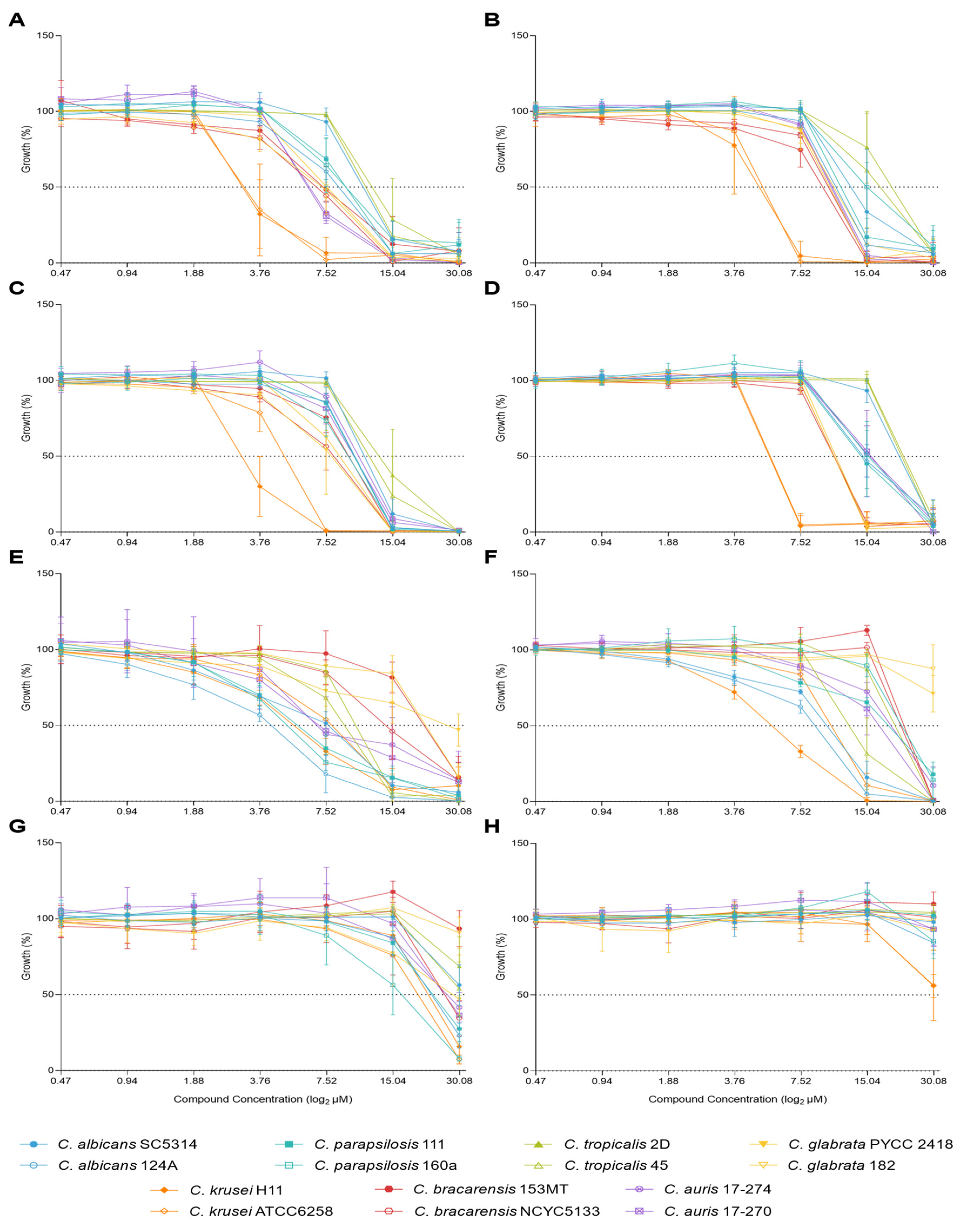
Figure 2.
Minimum Inhibitory Concentrations (MICs) of the selected benzo[a]phenoxazines against Candida species after 24 hours (left) and 48 hours (right) of growth, summarized in heat map format. The colour code to the right indicates the range of MIC values obtained.
Figure 2.
Minimum Inhibitory Concentrations (MICs) of the selected benzo[a]phenoxazines against Candida species after 24 hours (left) and 48 hours (right) of growth, summarized in heat map format. The colour code to the right indicates the range of MIC values obtained.
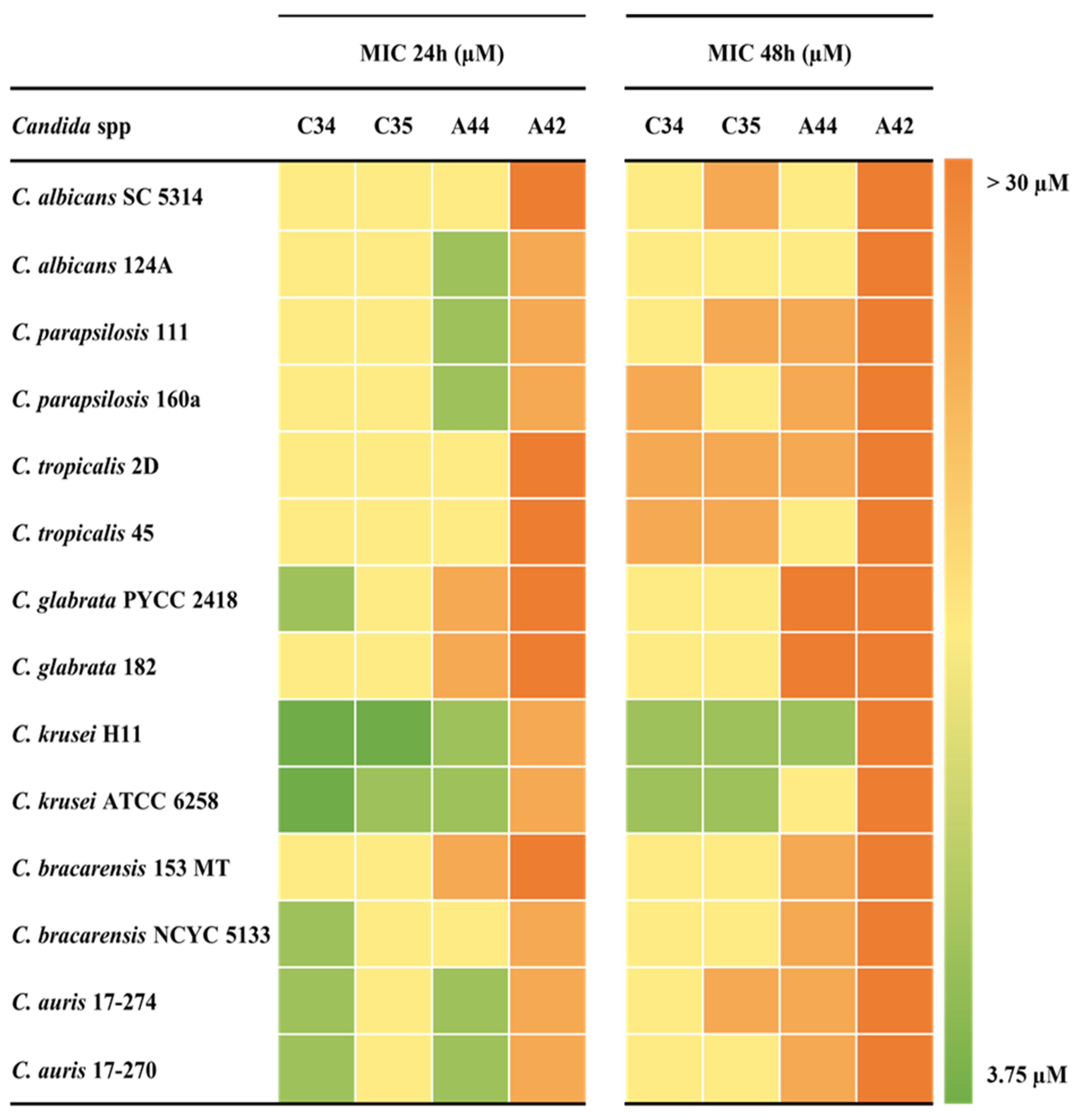
Figure 3.
Principal component analysis (PCA) of MICs data from fourteen strains of Candida spp. A -Scores: 14 strains segregation. B – Loadings: benzo[a]phenoxazine derivatives and timepoints tested.
Figure 3.
Principal component analysis (PCA) of MICs data from fourteen strains of Candida spp. A -Scores: 14 strains segregation. B – Loadings: benzo[a]phenoxazine derivatives and timepoints tested.
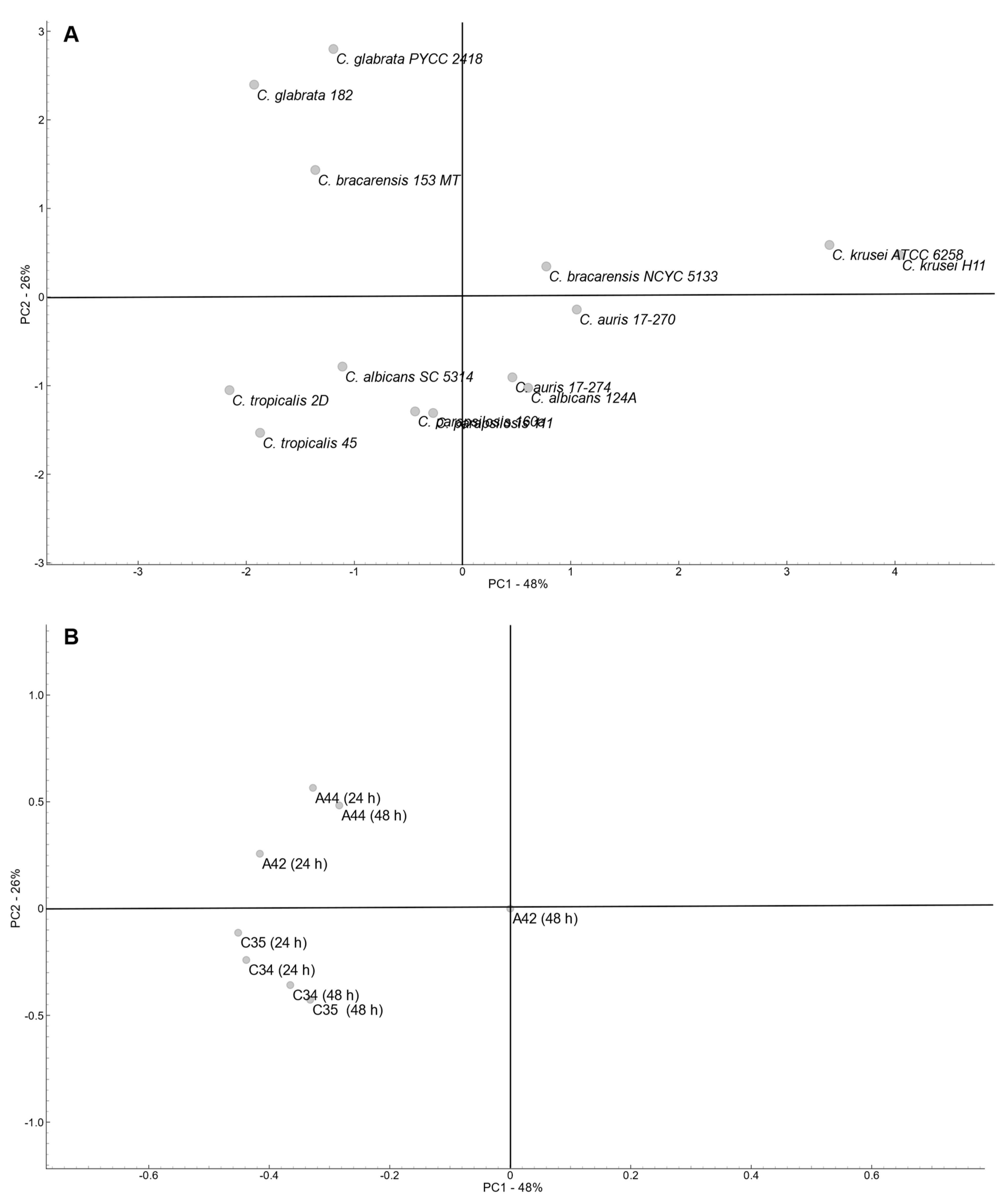
Figure 4.
Effect of the concentration of the selected compounds and fluconazole on the growth of four Candida species (C. albicans, C. glabrata, C. krusei and C. auris). A - C34; B - C35; C - A44; D - A42; E - Fluconazole.
Figure 4.
Effect of the concentration of the selected compounds and fluconazole on the growth of four Candida species (C. albicans, C. glabrata, C. krusei and C. auris). A - C34; B - C35; C - A44; D - A42; E - Fluconazole.
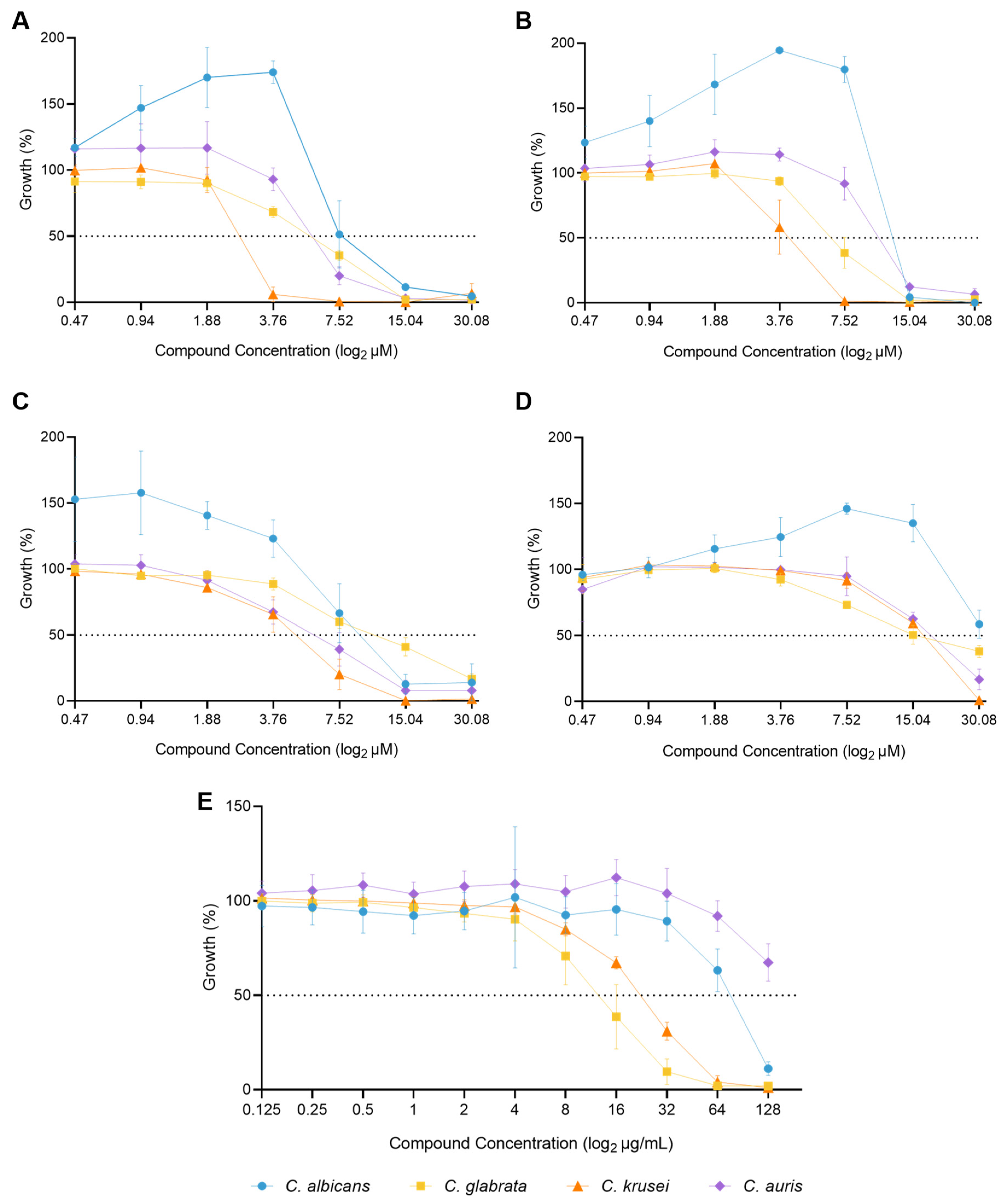
Figure 5.
Combination of C34 and fluconazole against C. albicans, C. glabrata, C. krusei and C. auris, analysed by response surface modelling based on the Bliss model. Synergy is mapped on the experimental response surface.
Figure 5.
Combination of C34 and fluconazole against C. albicans, C. glabrata, C. krusei and C. auris, analysed by response surface modelling based on the Bliss model. Synergy is mapped on the experimental response surface.
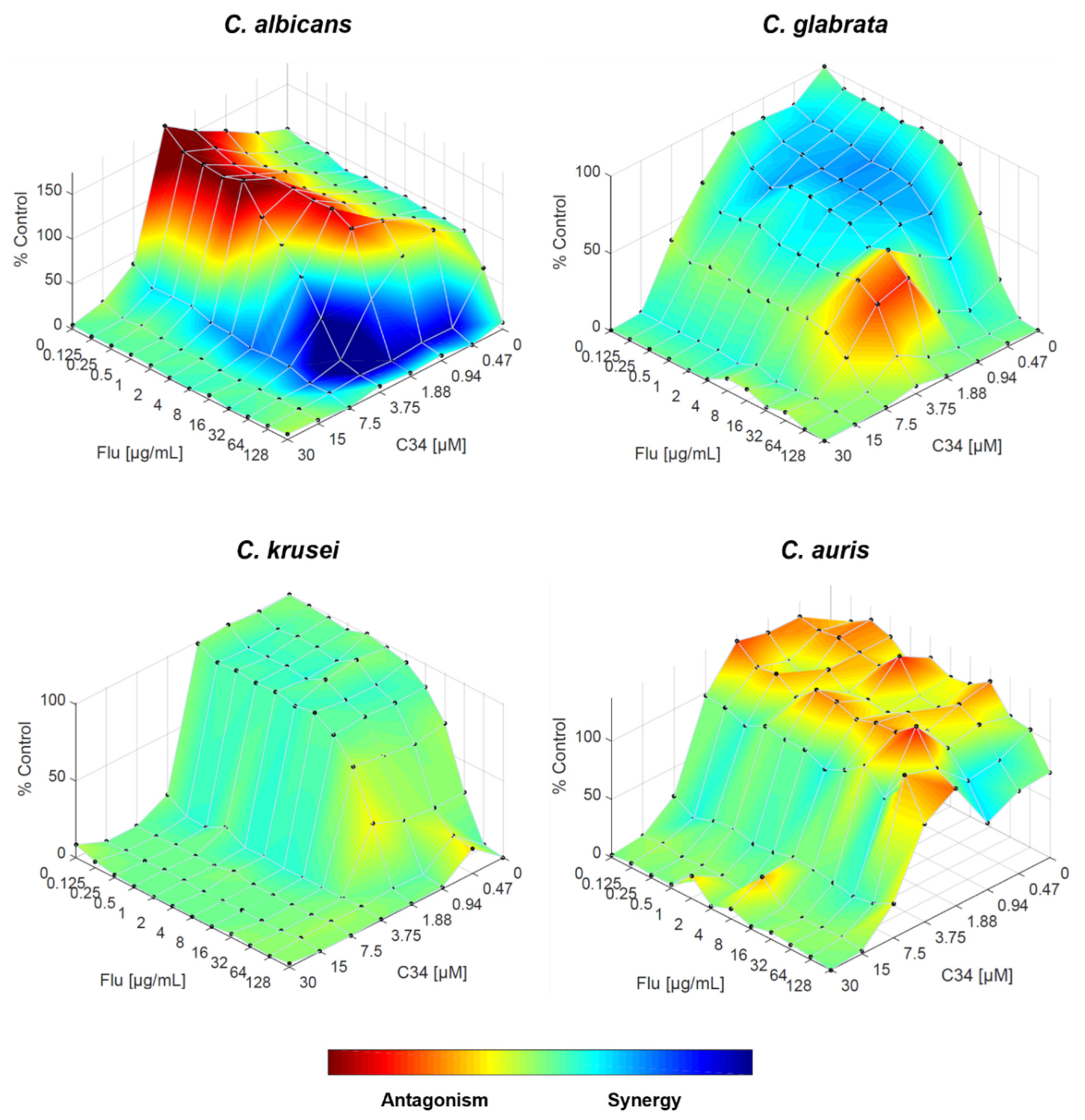
Figure 6.
Combination of C35 and fluconazole against C. albicans, C. glabrata, C. krusei and C. auris, analysed by response surface modelling based on the Bliss model. Synergy is mapped on the experimental response surface.
Figure 6.
Combination of C35 and fluconazole against C. albicans, C. glabrata, C. krusei and C. auris, analysed by response surface modelling based on the Bliss model. Synergy is mapped on the experimental response surface.
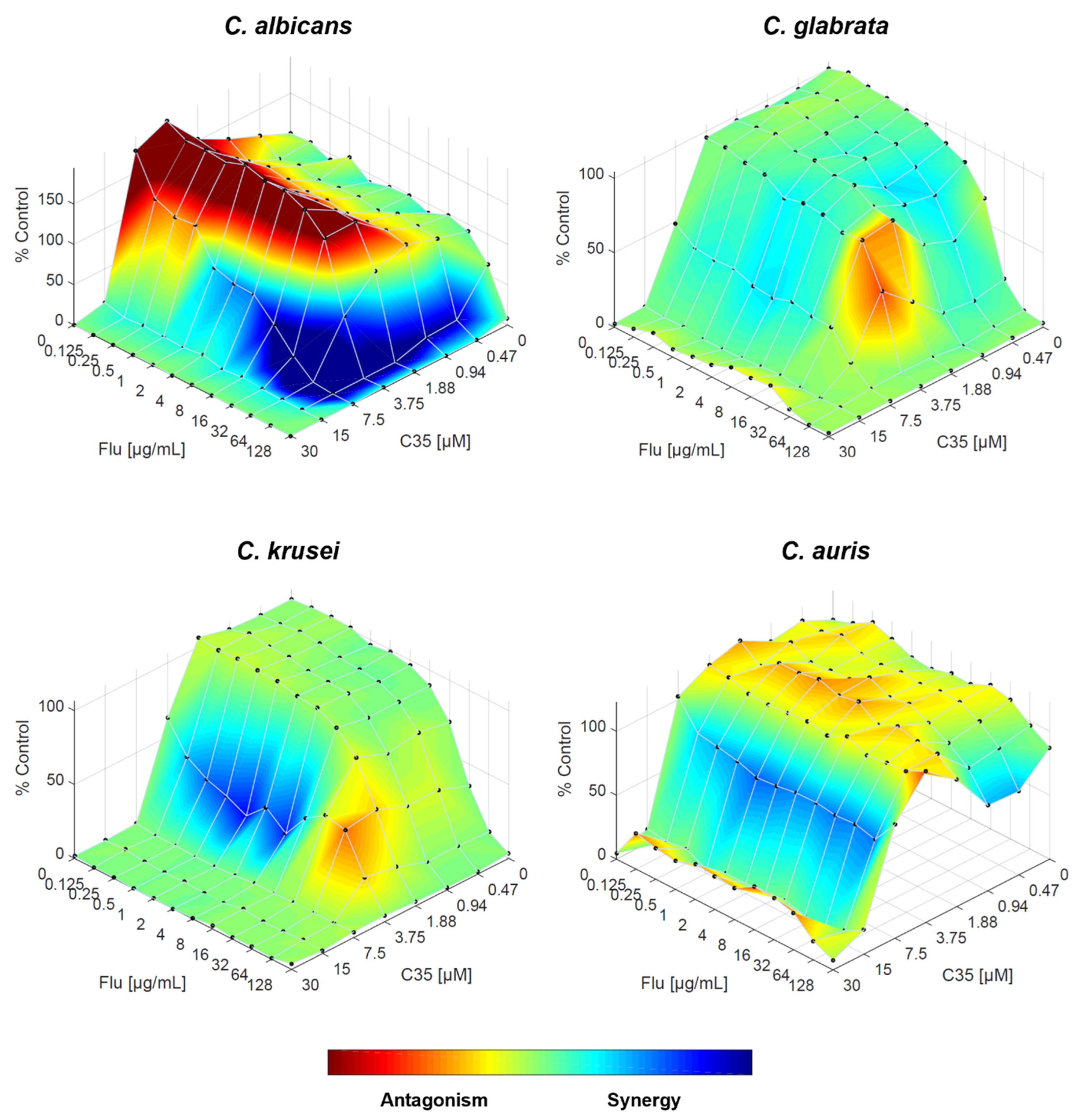
Figure 7.
Combination of A44 and fluconazole against C. albicans, C. glabrata, C. krusei and C. auris, analysed by response surface modelling based on the Bliss model. Synergy is mapped on the experimental response surface.
Figure 7.
Combination of A44 and fluconazole against C. albicans, C. glabrata, C. krusei and C. auris, analysed by response surface modelling based on the Bliss model. Synergy is mapped on the experimental response surface.
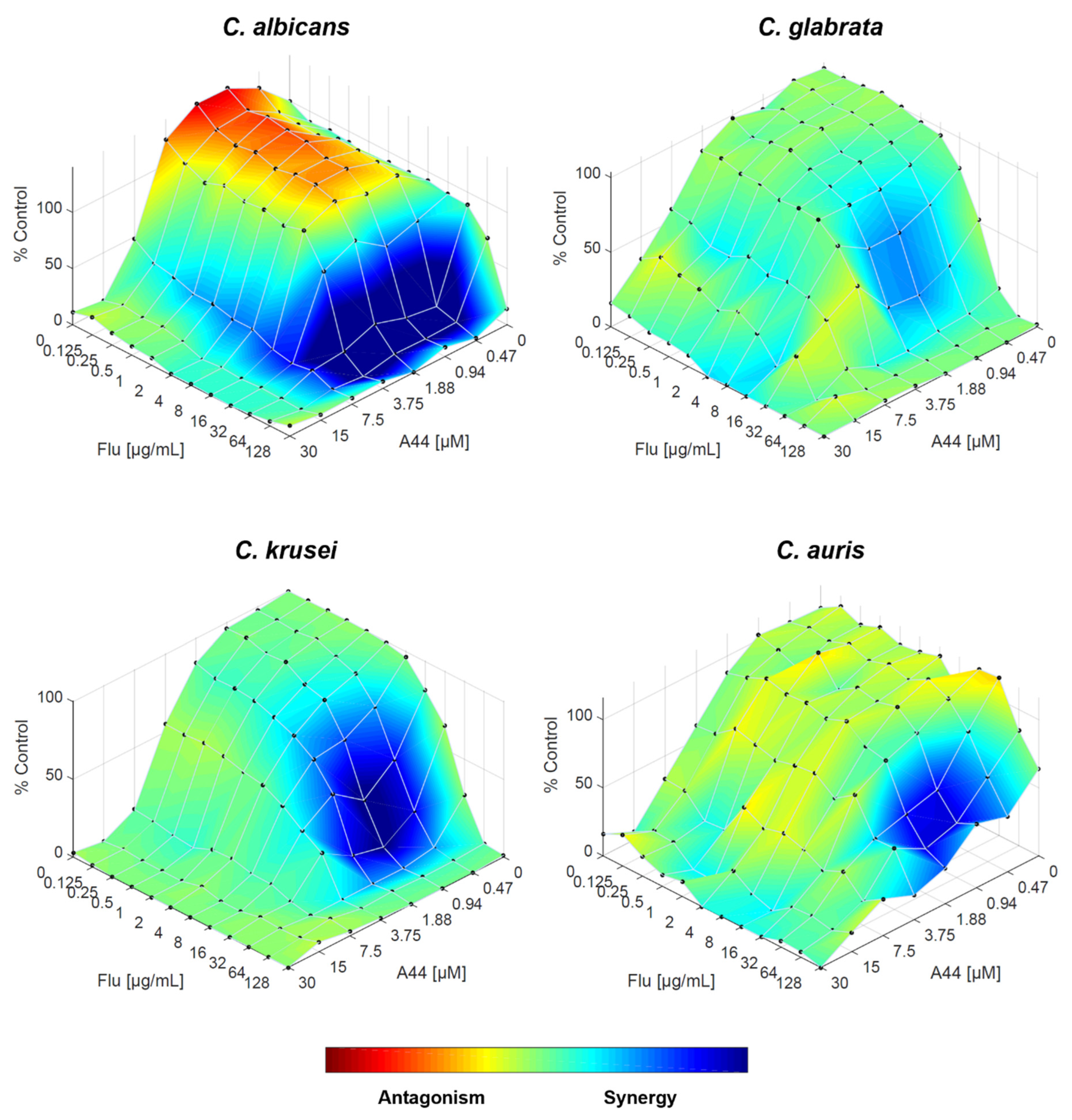
Figure 8.
Combination of A42 and fluconazole against C. albicans, C. glabrata, C. krusei and C. auris, analysed by response surface modelling based on the Bliss model. Synergy is mapped on the experimental response surface.
Figure 8.
Combination of A42 and fluconazole against C. albicans, C. glabrata, C. krusei and C. auris, analysed by response surface modelling based on the Bliss model. Synergy is mapped on the experimental response surface.
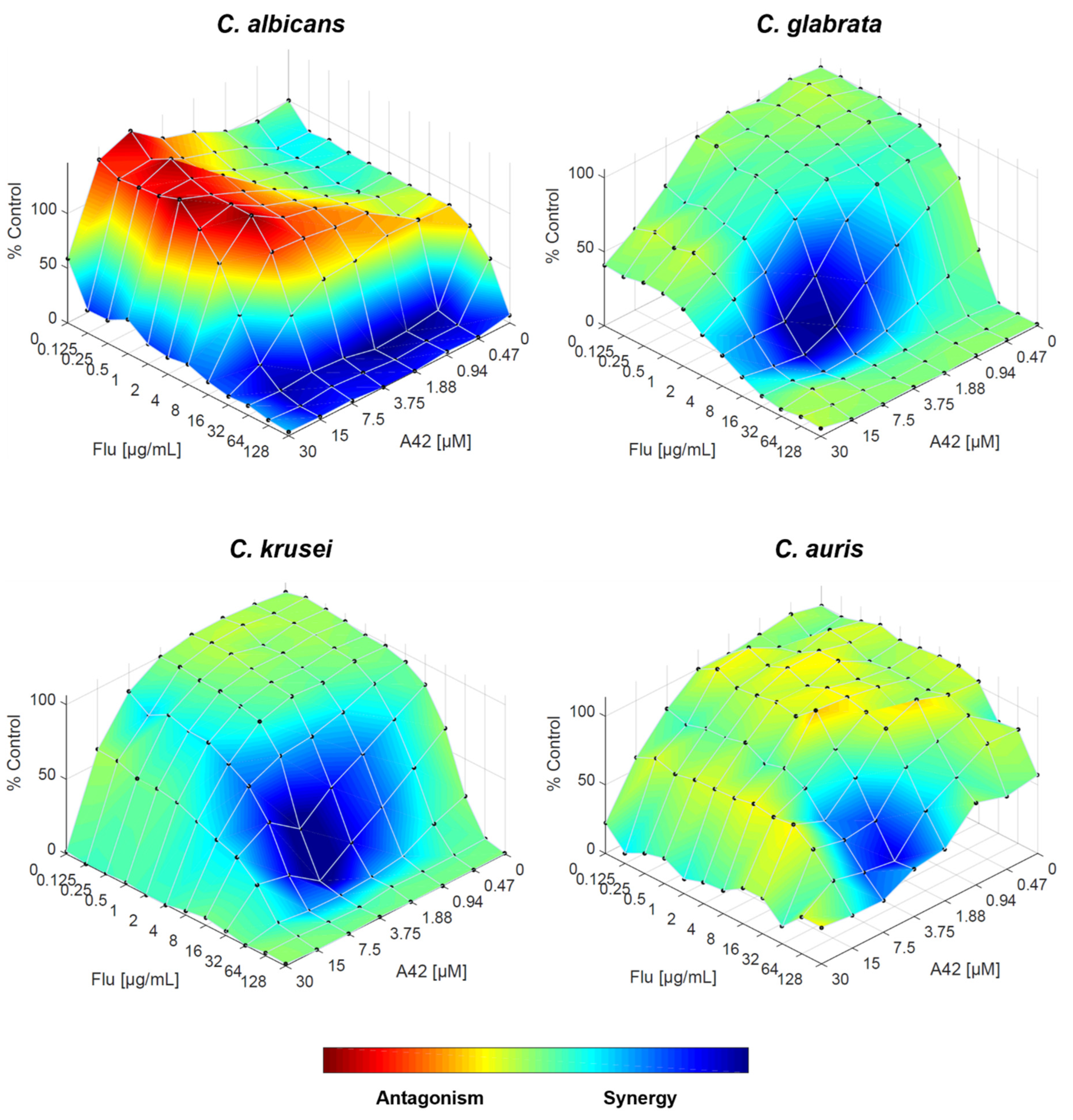
Figure 9.
Effect of increasing concentrations of each benzo[a]phenoxazine (C34, C35, A44 and A42) on the viability of J774A.1 cells. Viability was assessed using the MTT assay after 24 h (black bars) and 48 h (white bars) of incubation. Results indicate the mean ± SD of three independent assays. Significant differences between the two time points of incubation, 24 h and 48 h, are represented by: *P <0.05, **P <0.01, ***P <0.001, ****P <0.0001.
Figure 9.
Effect of increasing concentrations of each benzo[a]phenoxazine (C34, C35, A44 and A42) on the viability of J774A.1 cells. Viability was assessed using the MTT assay after 24 h (black bars) and 48 h (white bars) of incubation. Results indicate the mean ± SD of three independent assays. Significant differences between the two time points of incubation, 24 h and 48 h, are represented by: *P <0.05, **P <0.01, ***P <0.001, ****P <0.0001.
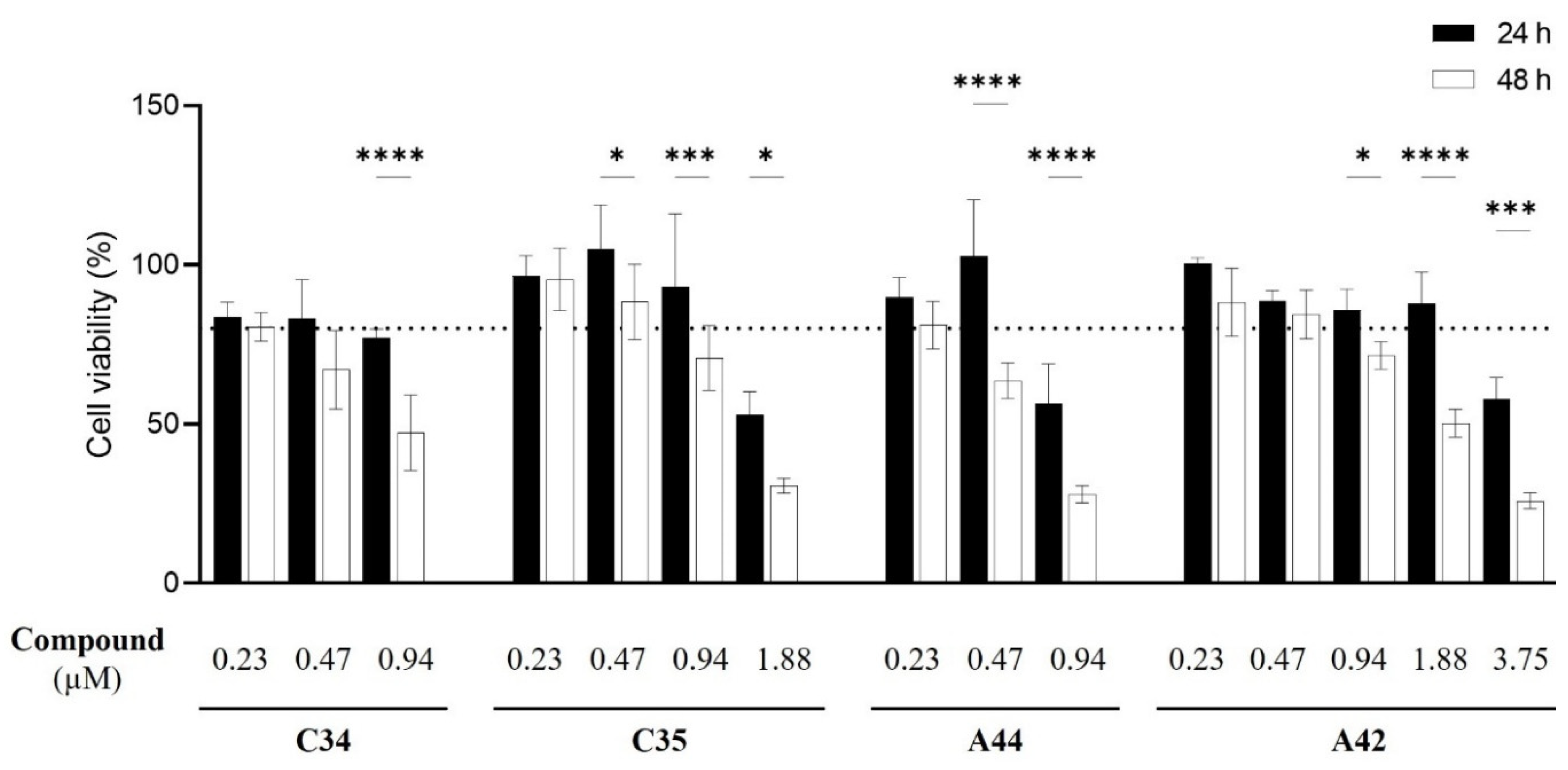
Figure 10.
Results of macrophage killing assay using the cellular line J774A.1 and four fluconazole-resistant Candida species. A - C. albicans; B - C. glabrata; C - C. krusei; D - C. auris. These graphics compare the yeast survival after incubation with macrophages, macrophages with fluconazole or A42 (alone) and macrophages with the combination of both drugs to evaluate the synergistic effect. Results indicate the mean ± SD of two independent assays. Significant differences with Candida cells incubated with macrophages are represented by *P <0.05, **P <0.01, ***P <0.001, ****P <0.0001. Significant differences between the single drug effects and the combination are represented by #P <0.05, ##P <0.01, ###P <0.001, ####P <0.0001.
Figure 10.
Results of macrophage killing assay using the cellular line J774A.1 and four fluconazole-resistant Candida species. A - C. albicans; B - C. glabrata; C - C. krusei; D - C. auris. These graphics compare the yeast survival after incubation with macrophages, macrophages with fluconazole or A42 (alone) and macrophages with the combination of both drugs to evaluate the synergistic effect. Results indicate the mean ± SD of two independent assays. Significant differences with Candida cells incubated with macrophages are represented by *P <0.05, **P <0.01, ***P <0.001, ****P <0.0001. Significant differences between the single drug effects and the combination are represented by #P <0.05, ##P <0.01, ###P <0.001, ####P <0.0001.
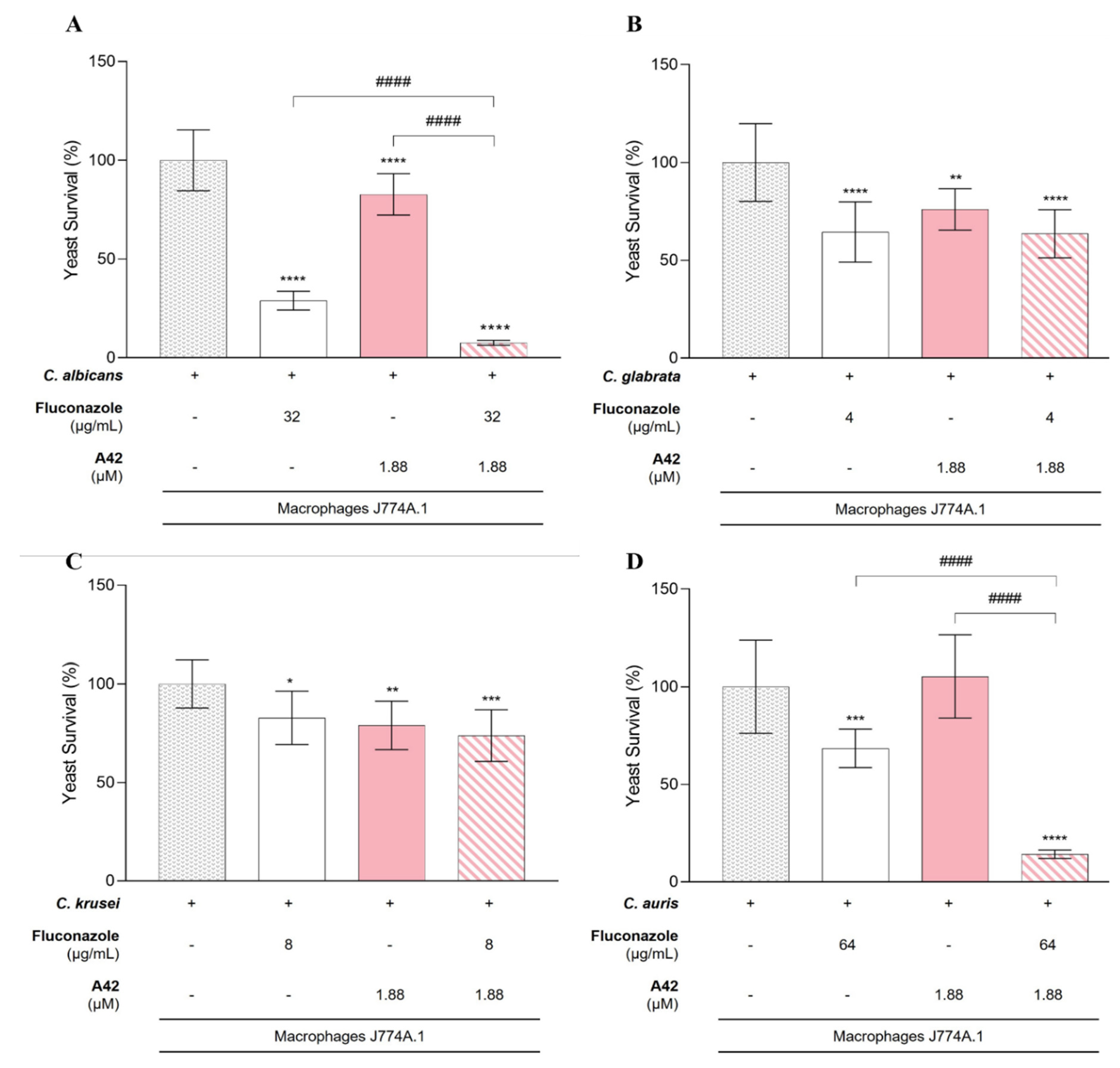

Table 1.
Benzo[a]phenoxazine derivatives used in this study, molecular structures, and MICs against S. cerevisiae PYCC 4072. Common positions for synthetic modifications with free rotation substituent group (A) and in the presence of a rigid structure, namely julolidine (B) is shown.
Table 1.
Benzo[a]phenoxazine derivatives used in this study, molecular structures, and MICs against S. cerevisiae PYCC 4072. Common positions for synthetic modifications with free rotation substituent group (A) and in the presence of a rigid structure, namely julolidine (B) is shown.
 | ||||||||
| Core Structure | Compound | R | R1 | R2 | R3 | R4 | MIC (µM) | Ref |
| A | C34 | (CH2)2CH3 | (CH2)2CH3 | (CH2)2CH3 | H | H | 1.56 |
As 7 [13] |
| A | C35 | (CH2)2CH3 | (CH2)2CH3 | (CH2)3Cl | H | H | 6.25 | As C35 / 6 [11] |
| B | A44 | (CH2)2CH3 | - | - | - | - | 6.25 | As 4c [15] |
| B | A42 | (CH2)3Cl | - | - | - | - | 0.78 | As 4d [15] |
Table 2.
MIC metrics of the selected benzo[a]phenoxazines against Candida species after 24 h and 48 h of growth.
Table 2.
MIC metrics of the selected benzo[a]phenoxazines against Candida species after 24 h and 48 h of growth.
| MIC 24h (µM) | MIC 48h (µM) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Metrics | C34 | C35 | A44 | A42 | C34 | C35 | A44 | A42 | ||
| Mode | 15 | 15 | 7.5 | 30 | 15 | 15 | 30 | >30 | ||
| Range MIC | 3.75 – 15 | 3.75 – 15 | 7.5 – 30 | 30 - >30 | 7.5 – 30 | 7.5 – 30 | 7.5 - >30 | >30 | ||
| MIC GM | 10.1 | 12.9 | 12.3 | 40.4 | 15.8 | 17.4 | 24.6 | 60.0 | ||
GM – Geometric Mean. For GM calculations, off-scale MICs were converted to the next highest concentration.
Table 3.
Interaction of fluconazole with C34 against C. albicans, C. glabrata, C. krusei and C. auris.
Table 3.
Interaction of fluconazole with C34 against C. albicans, C. glabrata, C. krusei and C. auris.
| MIC of Drugs Alone | MIC of the Drugs in Combination | Lowest FICI for the Combination | Interpretation | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Species | Flu (µg/mL) | C34 (µM) |
Flu (µg/mL) | C34 (µM) |
||||||
| C. albicans | 128 | 7.5 | 64 | 0.47 | 0.56 | IND | ||||
| C. glabrata | 16 | 7.5 | 64 | 1.88 | 4.25 | ANT | ||||
| C. krusei | 32 | 3.75 | 0.125 | 3.75 | 1 | IND | ||||
| C. auris | >128 | 7.5 | 0.125 | 7.5 | 1 | IND | ||||
Table 4.
Interaction of fluconazole with C35 against C. albicans, C. glabrata, C. krusei and C. auris.
Table 4.
Interaction of fluconazole with C35 against C. albicans, C. glabrata, C. krusei and C. auris.
| MIC of Drugs Alone | MIC of the Drugs in Combination | Lowest FICI for the Combination | Interpretation | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Species | Flu (µg/mL) | C35 (µM) |
Flu (µg/mL) | C35 (µM) |
||||||
| C. albicans | 128 | 15 | 64 | 0.47 | 0.53 | IND | ||||
| C. glabrata | 16 | 7.5 | 8 | 0.47 | 0.56 | IND | ||||
| C. krusei | 32 | 7.5 | 0.125 | 3.75 | 0.50 | SYN | ||||
| C. auris | >128 | 15 | 0.125 | 15 | 1 | IND | ||||
Table 5.
Interaction of fluconazole with A44 against C. albicans, C. glabrata, C. krusei and C. auris.
Table 5.
Interaction of fluconazole with A44 against C. albicans, C. glabrata, C. krusei and C. auris.
| MIC of Drugs Alone | MIC of the Drugs in Combination | Lowest FICI for the Combination | Interpretation | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Species | Flu (µg/mL) | A44 (µM) |
Flu (µg/mL) | A44 (µM) |
||||||
| C. albicans | 128 | 7.5 | 32 | 0.47 | 0.31 | SYN | ||||
| C. glabrata | 16 | 15 | 8 | 0.47 | 0.53 | IND | ||||
| C. krusei | 32 | 7.5 | 8 | 0.94 | 0.38 | SYN | ||||
| C. auris | >128 | 15 | 64 | 1.88 | 0.38 | SYN | ||||
Table 6.
Interaction of fluconazole with A42 against C. albicans, C. glabrata, C. krusei and C. auris.
Table 6.
Interaction of fluconazole with A42 against C. albicans, C. glabrata, C. krusei and C. auris.
| MIC of Drugs Alone | MIC of the Drugs in Combination | Lowest FICI for the Combination | Interpretation | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Species | Flu (µg/mL) | A42 (µM) |
Flu (µg/mL) | A42 (µM) |
||||||
| C. albicans | 128 | >30 | 32 | 0.47 | 0.26 | SYN | ||||
| C. glabrata | 16 | 30 | 4 | 3.75 | 0.38 | SYN | ||||
| C. krusei | 32 | 30 | 8 | 1.88 | 0.31 | SYN | ||||
| C. auris | >128 | 30 | 64 | 3.75 | 0.31 | SYN | ||||
Table 7.
Summary of the antifungal activity of fluconazole, A42 and their combination in the presence of macrophages against four fluconazole-resistant Candida species.
Table 7.
Summary of the antifungal activity of fluconazole, A42 and their combination in the presence of macrophages against four fluconazole-resistant Candida species.
| Fluconazole (µg/mL) |
A42 (µM) |
Growth Inhibition (%) | Combination Improvement over Fluconazole (%) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Species | MIC | Tested | MIC | Tested | Flu | A42 | Combination | ||||
| C. albicans | 128 | 32 | >30 | 1.88 | 71% | 17% | 92% | 21% | |||
| C. glabrata | 16 | 4 | 30 | 1.88 | 36% | 24% | 36% | 0% | |||
| C. krusei | 32 | 8 | 30 | 1.88 | 17% | 21% | 26% | 9% | |||
| C. auris | >128 | 64 | 30 | 1.88 | 32% | 0% | 86% | 54% | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



