
Submitted:
17 October 2024
Posted:
18 October 2024
You are already at the latest version
Abstract
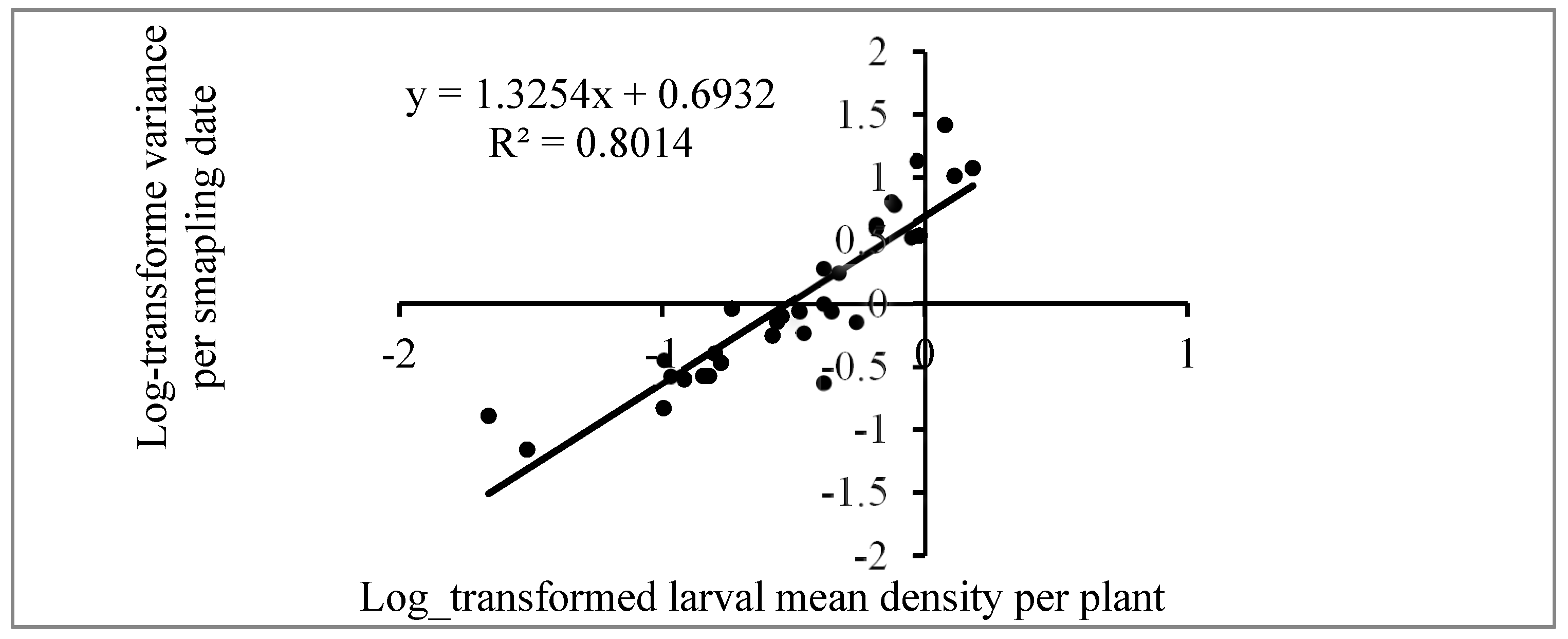
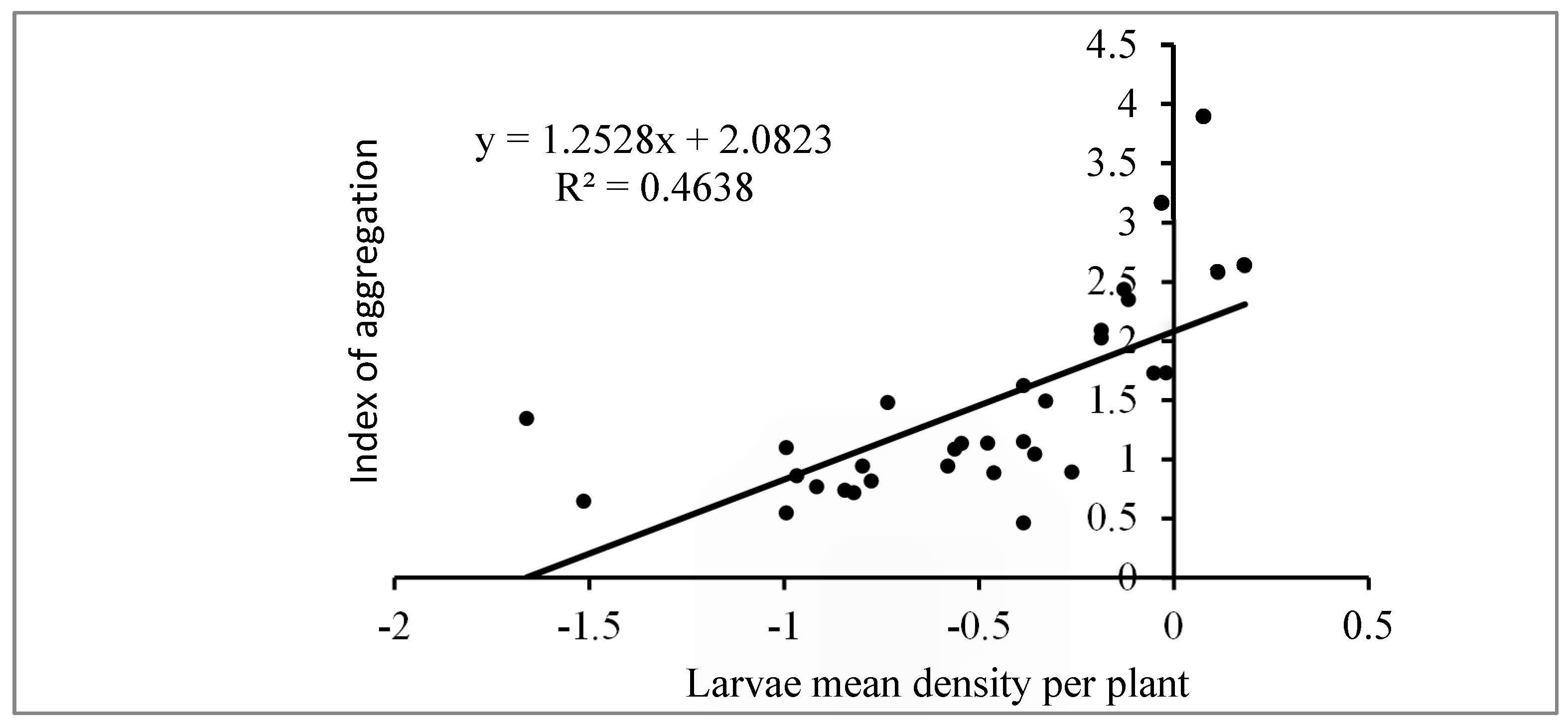
The fall armyworm (FAW), Spodoptera frugiperda, a major pest in maize production, was assessed for its temporal and spatial distribution in maize fields during both the off and rainy seasons of 2021 and 2022. In this study, 30 and 50 maize plants were randomly sampled using a "W" pattern during the dry and rainy seasons, respectively. Larval density, infestation rates, and damage severity were monitored over time. Taylor's power law and the mean crowding aggregation index were applied to evaluate the dispersion patterns of the larvae. The results indicate a higher infestation rate and larval density in agroecological zone AZE 8 (fisheries zone of South Benin) compared to AZE 6 (ferralitic soils zone) during the off-season. In the rainy season, while the percentage of damaged plants was higher in AZE 8, no significant differences in larval density between the two zones were observed. The dispersion analysis revealed moderate aggregation (aggregation index = 1.25) with a basic colony of 2.08 larvae, reflecting the aggregation oviposition behavior of FAW. This study provides valuable monitoring data on the FAW's distribution, offering insights for further research on population dynamics and developing predictive models for integrated pest management strategies.
Keywords:
1. Introduction
2. Materials and Methods
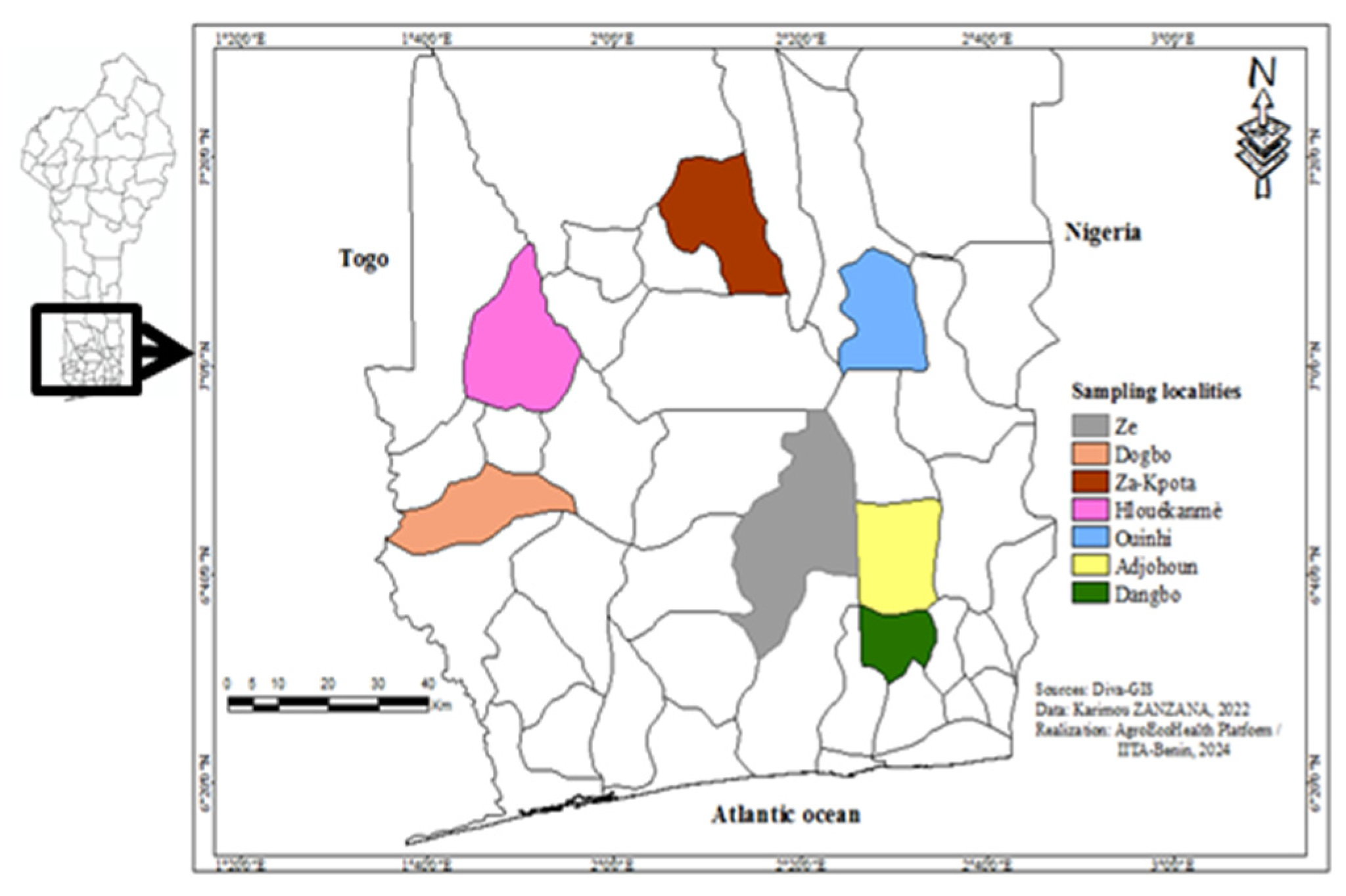
2.1. Study Area
2.2. Sampling of Fall Armyworm Larvae in Maize Fields
| Score | Description |
| 1 | No visible leaf-feeding damage |
| 2 | Few pinholes on 1-2 older leaves |
| 3 | Several shot-hole injuries on a few leaves (<5 leaves) and small circular hole damage to leaves |
| 4 | Several shot-hole injuries on several leaves (6–8 leaves) or small lesions/pinholes, small circular lesions, and a few small elongated (rectangular-shaped) lesions of up to 1.3 cm in length present on whorl and furl leaves |
| 5 | Elongated lesions (>2.5 cm long) on 8-10 leaves, plus a few small- to mid-sized uniform to irregular-shaped holes (basement membrane consumed) eaten from the whorl and/or furl leaves |
| 6 | Several large elongated lesions are present on several whorl and furl leaves and/or several large uniform to irregular-shaped holes eaten from furl and whorl leaves |
| 7 | Many elongated lesions of all sizes present on several whorl and furl leaves plus several large uniform to irregular-shaped holes eaten from the whorl and furl leaves |
| 8 | Many elongated lesions of all sizes present on most whorl and furl leaves plus many mid to large-sized uniform to irregular-shaped holes eaten from the whorl and furl leaves |
| 9 | Whorl and furl leaves almost totally destroyed and plant dying as a result of extensive foliar damage |
2.3. Parameters
2.4. Meteorological Data
2.5. Data Analysis
2.5.1. Larval Dispersion Model Analysis in Maize Fields
2.5.2. Statistical Analysis
3. Results
3.1. Infestation of Maize by FAW in Off and Rainy Season
3.2. Density of FAW Larvae During the Dry and Rainy Season
3.3. Percentage of Damaged Plants During the Off and Rainy Seasons
3.4. Plant Damage Severity in 2021 and 2022 During the Dry and Rainy Season
3.5. Dispersion Pattern of FAW Larvae
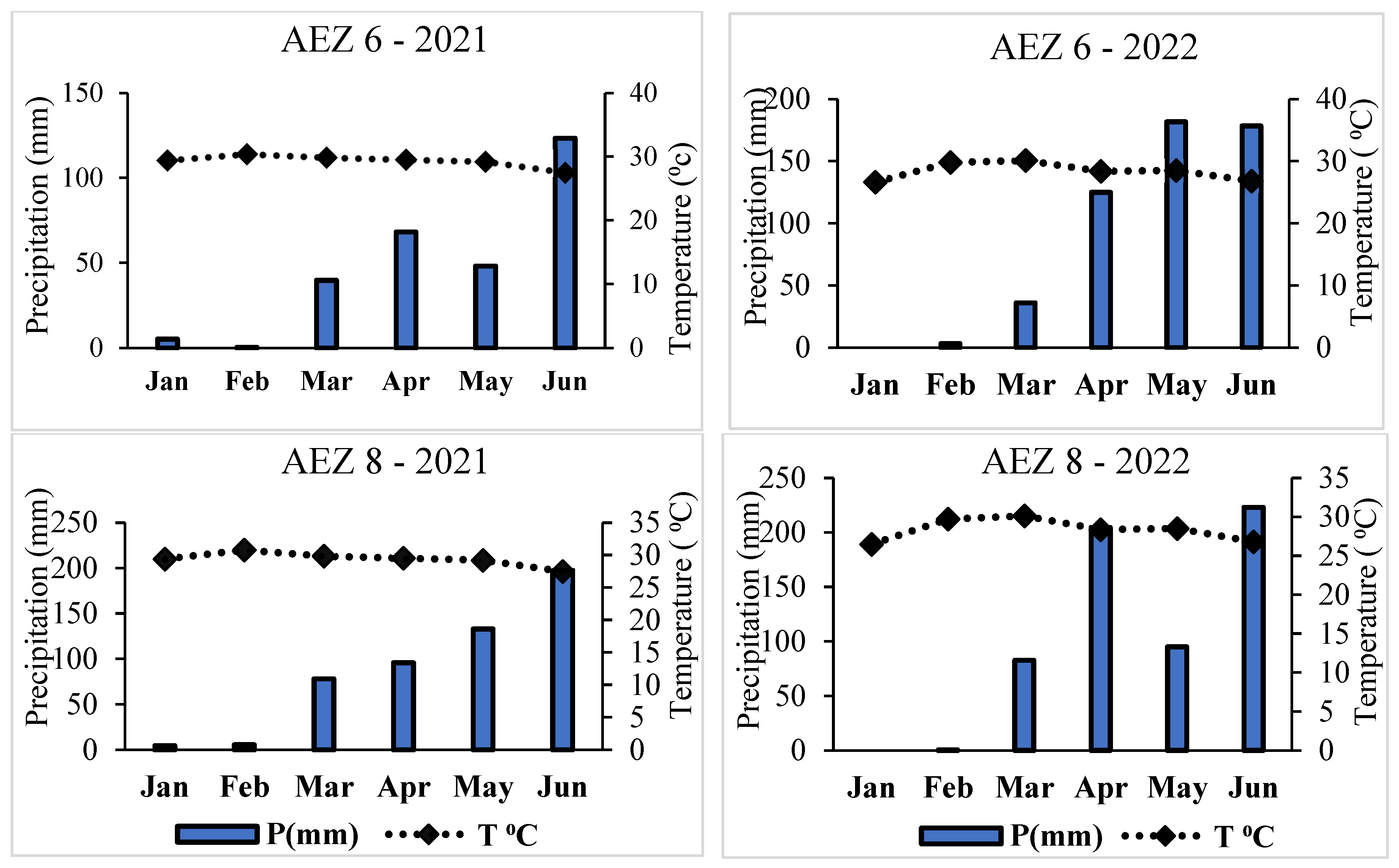
3.6. Temperature and Precipitation During the Survey
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Goergen, G.; Kumar, P.L.; Sankung, S.B.; Togola, A.; Tamò, M. First Report of Outbreaks of the Fall Armyworm Spodoptera frugiperda (J E Smith) (Lepidoptera, Noctuidae), a New Alien Invasive Pest in West and Central Africa. PLoS One 2016, 11, 0165632. [Google Scholar] [CrossRef] [PubMed]
- CABI. Spodoptera frugiperda (Fall Armyworm). Available online: https://www.cabi.org/isc/datasheet/29810 (accessed on 14 July 2024).
- Tao, W.C.; Zhang, X.Y.; Zhang, Y.; Deng, X.Y.; Zhang, H.L.; Zhang, Z.H.; Li, Q.; Jiang, C.X. Effects of the Host Plants of the Maize-Based Intercropping Systems on the Growth, Development and Preference of Fall Armyworm, Spodoptera Frugiperda (Lepidoptera: Noctuidae). Insects 2024, 15, 26. [Google Scholar] [CrossRef]
- Prasanna, B.M.; Huesing, J.E.; Eddy, R.; Peschke, V.M. Fall armyworm in Africa: A Guide For Intergrated Pest Management; 1st ed.; CDMX: CIMMYT: Mexico, 2018. [Google Scholar]
- 5. Rwomushana, I.; Bateman, M.; Beale, T.; Beseh, P.; Cameron, K.; Chiluba, M.; Clottey, V.; Davis, T.; Day, R.; Early, R.; et al. Fall Armyworm: Impacts and Implications for Africa. CABI Evidence Note Update, CABI: Oxfordshire, UK.; 2018;
- Toepfer, S.; Fallet, P.; Kajuga, J.; Bazagwira, D.; Mukundwa, I.P.; Szalai, M.; Turlings, T.C.J. Streamlining Leaf Damage Rating Scales for the Fall Armyworm on Maize. J Pest Sci 2021, 94, 1075–1089. [Google Scholar] [CrossRef]
- Montezano, D.G.; Specht, A.; Sosa-Gómez, D.R.; Roque-Specht, V.F.; Sousa-Silva, J.C.; Paula-Moraes, S. V.; Peterson, J.A.; Hunt, T.E. Host Plants of Spodoptera frugiperda (Lepidoptera: Noctuidae) in the Americas. African Entomology 2018, 26, 286–300. [Google Scholar] [CrossRef]
- Hafeez, M.; Li, X.W.; Zhang, J.M.; Zhang, Z.J.; Huang, J.; Wang, L.K.; Khan, M.M.; Shah, S.; Fernández-Grandon, G.M.; Lu, Y. Bin Role of Digestive Protease Enzymes and Related Genes in Host Plant Adaptation of a Polyphagous Pest, Spodoptera frugiperda. Insect Sci 2021, 28, 611–626. [Google Scholar] [CrossRef]
- Hafeez, M.; Li, X.; Chen, L.; Ullah, F.; Huang, J.; Zhang, Z.; Zhang, J.; Siddiqui, J.A.; Zhou, S.X.; Ren, X.Y.; et al. Molecular Characterization and Functional Analysis of Cytochrome P450-Mediated Detoxification CYP302A1 Gene Involved in Host Plant Adaptation in Spodoptera frugiperda. Front Plant Sci 2023, 13, 1–11. [Google Scholar] [CrossRef]
- Tambo, J.A.; Kansiime, M.K.; Rwomushana, I.; Mugambi, I.; Nunda, W.; Mloza Banda, C.; Nyamutukwa, S.; Makale, F.; Day, R. Impact of Fall Armyworm Invasion on Household Income and Food Security in Zimbabwe. Food Energy Secur 2021, 10, 299–312. [Google Scholar] [CrossRef]
- FAO. The Global Action for Fall Armyworm Control: Action Framework 2020–2022: Working Together to Tame the Global Threat. 2020.
- Han, S.-P.; Zhou, Y.-Y.; Wang, D.; Qin, Q.-J.; & P.S.; He, Y.-Z. Impact of Host Plants on Biological Characteristics and Vg/VgR Expression of Spodoptera frugiperda. J Pest Sci 2022, 96, 1569–1577.
- Koffi, D.; Kyerematen, R.; Eziah, V.Y.; Osei-Mensah, Y.O.; Afreh-Nuamah, K.; Aboagye, E.; Osae, M.; Meagher, R.L. Assessment of Impacts of Fall Armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae) on Maize Production in Ghana. J Integr Pest Manag 2020, 11, 1–7. [Google Scholar] [CrossRef]
- Suby, S.B.; Soujanya, P.L.; Yadava, P.; Patil, J.; Subaharan, K.; Prasad, G.S.; Babu, K.S.; Jat, S.L.; Yathish, K.R.; Vadassery, J.; et al. Invasion of Fall Armyworm (Spodoptera frugiperda) in India: Nature, Distribution, Management and Potential Impact. Curr Sci 2020, 119, 44–51. [Google Scholar] [CrossRef]
- Kogan, M.; Herzog, D.C. Sampling Methods in Soybean Entomology; Springer Science & Business Media.: New York, 1980. [Google Scholar]
- Rahmathulla, V.K.; Sathyanarayana, K.; Angadi, BS. Influence of Abiotic Factors on Population Dynamics of Major Insect Pests of Mulberry. Pakistan Journal of Biological Sciences 2015, 18, 215–223. [Google Scholar] [CrossRef]
- Cammell, M.E.; Knight, J.D. Effects of Climatic Change on the Population Dynamics of Crop Pests. Adv Ecol Res 1992, 22, 117–162. [Google Scholar]
- Hamby, K.A.; E. Bellamy, D.; Chiu, J.C.; Lee, J.C.; Walton, V.M.; Wiman, N.G.; York, R.M.; Biondi, A. Biotic and Abiotic Factors Impacting Development, Behavior, Phenology, and Reproductive Biology of Drosophila Suzukii. J Pest Sci 2016, 89, 605–619. [Google Scholar] [CrossRef]
- Omoregie, M.E.; Enobakhare, D.A.; Omoregie, A.O. Population Dynamics of the Fall Armyworm, Spodoptera frugiperda J. E. Smith (Lepidoptera: Noctuidae) on Early and Late Season Maize. Animal Research International 2023, 20, 4734–4740. [Google Scholar]
- Schowalter, T.D. Population Systems. In Insect Ecology: An Ecosystem Approach, 3rd ed.; Schowalter, T.D; Academic Press, Cambridge, Massachusetts, United States, 2011, 129–156.
- Nboyine, J.A.; Kusi, F.; Abudulai, M.; Badii, B.K.; Zakaria, M.; Adu, G.B.; Haruna, A.; Seidu, A.; Osei, V.; Alhassan, S.; et al. A New Pest, Spodoptera frugiperda (J.E. Smith), in Tropical Africa: Its Seasonal Dynamics and Damage in Maize Fields in Northern Ghana. Crop Protection 2020, 127, 104960. [Google Scholar] [CrossRef]
- Afshari, A.; Soleiman-Negadian, E.; Shishebor, P. Population Density and Spatial Distribution of Aphis Gossypii Glover (Homoptera: Aphididae) on Cotton in Gorgan. J. Agric. Sci. Technol 2009, 11, 27–38. [Google Scholar]
- Kuno, E. Sampling and Analysis of Insect Populations. 1991, 285–304.
- Melo, E.P.; Fernandes, M.G.; Degrande, P.E.; Cessa, R.M.A.; Salomão, J.L.; Nogueira, R.F. Distribuição Espacial de Plantas Infestadas Por Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae) Na Cultura Do Milho. Neotrop Entomol 2006, 35, 689–697. [Google Scholar] [CrossRef]
- Báez, M.S.; J.E., I.; Villanueva, F. Distribución Espacial y Tamaño de Muestra de Los Gusanos: Cogollero Spodoptera frugiperda (Smith) y Elotero Heliothis Zea (Boddie) En Cultivo de Maíz. Folia Entomológica Mexicana 1980, 45, 58–59.
- Hernández-Mendoza, J.L.; López-Barbosa, E.C.; Garza-González, E.; Mayek-Pérez, N. Spatial Distribution of Spodoptera frugiperda (Lepidoptera: Noctuidae) in Maize Landraces Grown in Colima, Mexico. Int J Trop Insect Sci 2008, 28, 126–129. [Google Scholar] [CrossRef]
- He, Y.; Wang, K.; Du, G.; Zhang, Q.; Li, B.; Zhao, L.; He, P.; Chen, B. Temporal and Spatial Distribution Patterns of Spodoptera frugiperda in Mountain Maize Fields in China. Insects 2022, 13, 938. [Google Scholar] [CrossRef]
- Elliott, N.C.; Kieckhefer, R.W. and; Walgenbach, D.D. Binomial Sequential Sampling Methods for Cereal Aphids in Small Grains. Journal of Economic Entomology 1990, 83, 1381–1387. [Google Scholar] [CrossRef]
- Taylor, L.R.; Woiwod, I.P. &; Perry, J.N. The Negative Binomial as a Dynamic Ecological Model for Aggregation, and the Density Dependence of k. J Anim Ecol 1979, 48, 289–304. [Google Scholar]
- Taylor, L.R. Assessing and Interpreting the Spatial Distributions of Insect Populations. Annual review of entomology. Vol. 29 1984, 321–358. [Google Scholar] [CrossRef]
- Caniço, A.; Mexia, A.; Santos, L. Seasonal Dynamics of the Alien Invasive Insect Pest. Journal insects 2020, 11, 1–12. [Google Scholar]
- Anandhi, S.; Saminathan, V.R.; Yasodha, P.; Roseleen, S.S.J.; Sharavanan, P.T.; Rajanbabu, V. Correlation of Fall Armyworm Spodoptera frugiperda (J.E. Smith) with Weather Parameters in Maize Ecosystem. Int J Curr Microbiol Appl Sci 2020, 9, 1213–1218. [Google Scholar] [CrossRef]
- Ramasamy, M.; Das, B.; Ramesh, R. Predicting Climate Change Impacts on Potential Worldwide Distribution of Fall Armyworm Based on CMIP6 Projections. J Pest Sci 2021, 95, 841–854. [Google Scholar] [CrossRef]
- Davis, F.M.; Ng, S.S.; Williams, W.P. Visual Rating Scales for Screening Whorl-Stage Corn for Resistance to Fall Armyworm. Mississippi Agricultural & Forestry Experiment Station, Bulletin Technique 186, Université d’état de Mississippi,USA. 1992.
- Atachi, P.; Dannon, E.A.; Arodokoun, Y.D.; Tamò, M. Distribution and Sampling of Maruca Vitrata (Fabricius) (Lep., Pyralidae) Larvae on Lonchocarpus Sericeus (Poir) H.B. and K. Journal of Applied Entomology 2002, 126, 188–193. [Google Scholar] [CrossRef]
- Iwao, S. A New Regression Method for Analyzing the Aggregation Pattern of Animal Populations. Water (Basel) 1968, 10, 1–20. [Google Scholar] [CrossRef]
- Ruesink, W.G. Introduction to Sampling Theory. Probability and Statistics for Economists 2017, 299–350. [Google Scholar]
- Taylor, L.R. Aggregation, Variance and the Mean. Nature 1961, 189, 732–735. [Google Scholar] [CrossRef]
- Southwood, T.R.E. Ecological Methods. London,UK: Chapman & Hall, 1978, 524.
- Lloyd, M. Mean Crowding. J Anim Ecol, 36 1967, 1-30.
- Zahner, P.; J.; B.; Delucchi, V.; Graf, B. Distribution and Sampling of Winter Populations of Panonychus Ulmi Koch (Acarina: Tetranychidae) on Apple Tree. Oecol. Appl. 1985, 6, 99–110.
- Deutsch, C.A.; Tewksbury, J.J.; Tigchelaar, M.; Battisti, D.S.; Merrill, S.C.; Huey, R.B.; Naylor, R.L. Climate Change Increase in Crop Losses to Insect Pests in a Warming Climate. Science (1979) 2018, 361, 31. [Google Scholar]
- Vicente, J.R.; Vaz, A.S.Q.; Isabel, A.; Buchadas, A.R.; Guisan, A.; Kueffer, C.; Marchante, E.; Marchante, H.; Cabral, J.A.; Nesper, M.; et al. Alien Plant Species: Environmental Risks in Agricultural and Agro-Forest Landscapes Under Climate Change. In Climate Change-Resilient Agriculture and Agroforestry: Ecosystem Services and Sustainability; 2019, 215–234.
- Tepa-Yotto, G.T.; Tonnang, H.E.Z.; Goergen, G.; Subramanian, S.; Kimathi, E.; Abdel-Rahman, E.M.; Flø, D.; Thunes, K.H.; Fiaboe, K.K.M.; Niassy, S.; et al. Global Habitat Suitability of Spodoptera frugiperda (Je Smith) (Lepidoptera, Noctuidae): Key Parasitoids Considered for Its Biological Control. Insects 2021, 12, 273. [Google Scholar] [CrossRef]
- Sharanabasappa; Kalleshwaraswamy, C.M.; Maruthi, M.S.; Pavithra, H.B. Biology of Invasive Fall Army Worm Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae) on Maize . Indian Journal of Entomology 2018, 80, 540.
- Bakry, M.M.S.; Abdel-Baky, N.F. Population Density of the Fall Armyworm, Spodoptera frugiperda (Smith) (Lepidoptera: Noctuidae) and Its Response to Some Ecological Phenomena in Maize Crop. Brazilian Journal of Biology 2023, 83, 1–17. [Google Scholar] [CrossRef]
- Murúa, G.; Molina-Ochoa, J.; Coviella, C. Population Dynamics of the Fall Armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae) and Its Parasitoids in Northwestern Argentina. Florida Entomologist 2006, 89, 175–182. [Google Scholar] [CrossRef]
- Mitchell, E.R.; McNeil, J.N.; Westbrook, J.K.; Silvain, J.F.; Lalanne-Cassou, B.; Chalfant, R.B.; Pair, S.D.; Waddill, V.H.; Sotomayor-Rios, A.; Proshold, F.I. Seasonal Periodicity of Fall Armyworm, (Lepidoptera: Noctuidae) in the Caribbean Basin and Northward to Canada. J Entomol Sci 1991, 26, 39–50. [Google Scholar] [CrossRef]
- Pannuti, L.E.R.; Baldin, E.L.L.; Hunt, T.E.; Paula-Moraes, S. V. On-Plant Larval Movement and Feeding Behavior of Fall Armyworm (Lepidoptera: Noctuidae) on Reproductive Corn Stages. Environ Entomol 2016, 45, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Sparks, A.N. A Review of the Biology of the Fall Armyworm. Fla Entomol 1979, 62, 82–87. [Google Scholar] [CrossRef]
- Vilarinho, E.C.; Fernandes, O.A.; Hunt, T.E.; Caixeta, D.F. Movement of “Spodoptera frugiperda” Adults (Lepidoptera: Noctuidae) in Maize in Brazil. 2011, 94, 480–488.
- Winsou, J.K.; Tepa-Yotto, G.T.; Thunes, K.H.; Meadow, R.; Tamò, M.; Sæthre, M.G. Seasonal Variations of Spodoptera frugiperda Host Plant Diversity and Parasitoid Complex in Southern and Central Benin. Insects 2022, 13, 491. [Google Scholar] [CrossRef]
- Early, R.; González-Moreno, P.; Murphy, S.T.; Day, R. Forecasting the Global Extent of Invasion of the Cereal Pest Spodoptera Frugiperda, the Fall Armyworm. NeoBiota 2018, 50, 25–50. [Google Scholar] [CrossRef]
- Mitchell, F.L. Natural Control and Spatial Distribution of Fall Armyworm ( Spodoptera Frugiperda ) Within Louisiana Corn Fields; Louisiana State University and Agricultural & Mechanical College 1985.
- Farias, P.R.S.; Barbosa, J.C.; Busoli, A.C. Distribuição Espacial Da Lagarta-Do-Cartucho, Spodoptera Frugiperda (J.E. Smith) (Lepidoptera: Noctuidae), Na Cultura Do Milho. Neotrop Entomol 2001, 30, 681–689. [Google Scholar] [CrossRef]
- Sétamou, M.; Schulthess, F.; Poehling, H.M.; Borgemeister, C. Spatial Distribution of and Sampling Plans for Mussidia nigrivenella (Lepidoptera: Pyralidae) on Cultivated and Wild Host Plants in Benin. Environ Entomol 2000, 29, 1216–1225. [Google Scholar] [CrossRef]
- Moradi Vajargah, M.; Golizadeh, A.; Dastjerdi, H.R.; Zalucki, M.P.; Hassanpour, M.; Naseri, B. Population Density and Spatial Distribution Pattern of Hypera postica (Coleoptera: Curculionidae) in Ardabil, Iran. Not Bot Horti Agrobot Cluj Napoca 2011, 39, 42–48. [Google Scholar] [CrossRef]
- Guimapi, R.A.; Niassy, S.; Mudereri, B.T.; Abdel-Rahman, E.M.; Tepa-Yotto, G.T.; Subramanian, S.; Mohamed, S.A.; Thunes, K.H.; Kimathi, E.; Agboka, K.M.; et al. Harnessing Data Science to Improve Integrated Management of Invasive Pest Species across Africa: An Application to Fall Armyworm (Spodoptera frugiperda) (J.E. Smith) (Lepidoptera: Noctuidae). Glob Ecol Conserv 2022, 35, e02056. [Google Scholar] [CrossRef]
- Sedaratian-Jahromi, A. Effects of Entomopathogens on Insect Predators and Parasitoids. In Microbes for Sustainable lnsect Pest Management; 2021, 2, 83–231. [Google Scholar]

| Season | Agroecological zone | Infestation rate (%) (Mean ± SE) | F | P | |
| Year 2021 | Year 2022 | ||||
| Off-season | AEZ 6 | 36.94 ± 2.61bB | 29.97 ± 2.25aC | 11.69 | 0.0001 |
| AEZ 8 | 36.50 ± 2.00bB | 29.87 ± 2.27aC | |||
| Rainy season | AEZ 6 | 42.40 ± 2.90aB | 41.44 ± 2.32aB | 3.80 | 0.01 |
| AEZ 8 | 47.96 ± 2.87abA | 50.94 ± 1.90bA | |||
| F | 3.54 | 21.49 | |||
| P | 0.01 | 0.0001 | |||
| Season | Agroecological zone | Sampling week | Larval density (Mean ± SE) | |
| Year 2021 | Year 2022 | |||
| Off-season | AEZ 6 | 1 | 0.88 ± 0.08abc | 0.96 ± 0.09abc |
| 2 | 0.58 ± 0.05d | 0.68 ± 0.05cde | ||
| 3 | 0.71 ± 0.06cd | 0.67 ± 0.06cde | ||
| 4 | - | 0.56 ± 0.05ed | ||
| 5 | - | 0.43 ± 0.04e | ||
| Overall mean | 0.73 ± 0.03B | 0.66 ± 0.02B | ||
| AEZ 8 | 1 | 1.11 ± 0.09a | 1.07 ± 0.11ab | |
| 2 | 0.82 ± 0.07bc | 1.20 ± 0.09a | ||
| 3 | 0.98 ± 0.70ab | 0.95 ± 0.07abc | ||
| 4 | - | 0.87 ± 0.08bc | ||
| 5 | - | 0.82 ± 0.07bcd | ||
| Overall mean | 0.98 ± 0.04A | 0.99 ± 0.03A | ||
| F = 7.15; P < 0.0001 | F = 10.42; P < 0.0001 | |||
| Rainy season | AEZ 6 | 1 | 0.44 ± 0.03b | 0.18 ± 0.03d |
| 2 | 0.62 ± 0.04a | 0.42 ± 0.03c | ||
| 3 | 0.69 ± 0.04a | 0.36 ± 0.03c | ||
| 4 | - | 0.38 ± 0.03c | ||
| 5 | - | 0.77 ± 0.04a | ||
| Overall mean | 0.59 ± 0.02C | 0.43 ± 0.01C | ||
| AEZ 8 | 1 | 0.48 ± 0.04b | 0.37 ± 0.03c | |
| 2 | 0.56 ± 0.04a | 0.21 ± 0.02d | ||
| 3 | 0.63 ± 0.04a | 0.39 ± 0.03c | ||
| 4 | - | 0.45 ± 0.03c | ||
| 5 | - | 0.67 ± 0.03b | ||
| Overall mean | 0.56 ± 0.02C | 0.42 ± 0.01C | ||
| F=6.38; P < 0.0001 | F = 35.88; P < 0.0001 | |||
| Season | Agroecological zones | Percentage of damaged plants (Mean ± SE) |
F | P | |
| Year 2021 | Year 2022 | 8.09 |
0.0005 |
||
| Off-season | AEZ 6 | 58.33 ± 1.51bB | 51.53 ± 2.31aC | ||
| AEZ 8 | 55.72 ± 1.82abB | 53.73 ± 2.35aC | |||
| Rainy season | AEZ 6 | 80.92 ± 1.94cA | 63.44 ± 2.06aB | 16.61 | 0.0001 |
| AEZ 8 | 82.22 ± 2.36cA | 72.05 ± 1.86bA | |||
| F | 66.42 | 19.91 | |||
| P | 0.0001 | 0.0001 | |||
| Season | Agroecological zone | Sampling week | Plant Damage Score Per AEZ (Scale 1–9) (Mean ± SE) | |
| Year 2021 | Year 2022 | |||
| Off-season | AEZ 6 | 1 | 3.36 ± 0.10b | 2.58 ± 0.07b |
| 2 | 3.17 ± 0.09bc | 2.25 ± 0.06c | ||
| 3 | 3.30 ± 0.08bc | 2.30 ± 0.06c | ||
| 4 | - | 2.73 ± 0.07ab | ||
| 5 | - | 2.24 ± 0.07c | ||
| Overall mean | 3.28 ± 0.05A | 2.42 ± 0.03BC | ||
| AEZ 8 | 1 | 2.95 ± 0.08c | 2.32 ± 0.07c | |
| 2 | 3.13 ± 0.08bc | 2.63 ± 0.07ab | ||
| 3 | 3.82 ± 0.15a | 2.63 ± 0.06ab | ||
| 4 | - | 2.89 ± 0.07a | ||
| 5 | - | 2.84 ± 0.07a | ||
| Overall mean | 3.30 ± 0.06A | 2.66 ± 0.03A | ||
| F = 8.60; P < 0.0001 | F = 12.91; P < 0.0001 | |||
| Rainy season | AEZ 6 | 1 | 2.03 ± 0.05b | 1.55 ± 0.04f |
| 2 | 2.10 ± 0.05b | 2.56 ± 0.07b | ||
| 3 | 2.32 ± 0.05a | 2.70 ± 0.08b | ||
| 4 | - | 1.98 ± 0.05e | ||
| 5 | - | 3.48 ± 0.08a | ||
| Overall mean | 2.16 ± 0.02B | 2.46 ± 0.03B | ||
| AEZ 8 | 1 | 2.00 ± 0.05b | 1.84 ± 0.04e | |
| 2 | 2.10 ± 0.05b | 1.79 ± 0.05e | ||
| 3 | 2.32 ± 0.05a | 2.45 ± 0.07c | ||
| 4 | - | 2.23 ± 0.05d | ||
| 5 | - | 3.41 ± 0.08a | ||
| Overall mean | 2.14 ± 0.03B | 2.35 ± 0.02C | ||
| F = 197.99; P < 0.0001 | F = 13.53; P < 0.0001 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).



