
Submitted:
19 October 2024
Posted:
21 October 2024
You are already at the latest version
Abstract
This research aimed to explore the effects of Silicon (Si) on managing hydrogen peroxide levels, photosynthetic pigment content, nutrient accumulation, and the production of root and shoot dry biomass in salt-stressed sunflower (Helianthus annuus L.) plants. Conducted in a glasshouse, the experiment followed a completely randomized block design in a factorial arrangement (2 × 2) and five replicates. Pots were divided into two groups: non-salt stressed, and salt stressed (100 mM NaCl), both with Si (2 mM as stabilized sodium and potassium silicate) and without Si. The study found that Si effectively reduced hydrogen peroxide concentration and enhanced photosynthetic pigments, nutrient status, and plant growth. The results showed that Si alleviated salt stress effects in sunflower plants by significantly lowering hydrogen peroxide levels and increasing photosynthetic pigments and nutrient accumulation, leading to greater root and shoot dry biomass. These findings indicate that adding Si to the growth medium is crucial for enhancing resistance to salt-induced ionic and osmotic stress, making it a promising strategy for improving crop growth and management in saline conditions.
Keywords:
Helianthus annus
; ionic stress
; productivity
; osmotic stress
; salinity
; Silicon
1. Introduction
Soil salinity has been reported to be increasing due to the effects of climate change, causing a devastating impact on land resources (affecting 7% of them) and cultivated land (5 %) [1]. Salinity is a major environmental factor that restricts crop production in arid and semi-arid regions [2], where it impacts plant growth and yield due to osmotic effects, nutritional imbalances, oxidative damage, and/or toxicity from specific ions [3]. High salt levels in the soil lead to a high osmotic potential, restricting the uptake of water and nutrients by plants [4]. Additionally, ionic stress occurs due to the accumulation of salt reaching toxic levels in mature leaves, which hastens senescence and leads to leaf death [5]. Elevated levels of Na+ and Cl− lead to nutritional imbalances, resulting in increased production of reactive oxygen species (ROS), which negatively impact plant cells, growth, and productivity.[6]. High salinity disrupts several essential physiological and biochemical processes, such as transpiration, photosynthesis, protein synthesis, and others [7].
Various strategies are being employed to reduce the adverse effects of salinity on plant growth and productivity. Among these, silicon supplementation is regarded as a significant approach to alleviate the detrimental impacts of salinity on plant growth and yield. Silicon can enhance plant survival under different abiotic stresses, such as mineral deficiency. [8], toxicity [9], salinity [10], and drought [11]. All terrestrial plants have Si in their tissues, though the concentration varies significantly between species, ranging from 0.1% to 10% Si on a dry matter basis [12]. Overall, it was shown that both active and passive mechanisms are involved in the uptake and transport of Si in intermediate-type species like sunflower, with their roles varying based on the plant species and external Si concentrations[13].
Previous reports have indicated that Si plays a role in alleviating salt stress by reducing Na+ uptake. [15,16], ROS accumulation and transpiration rate [17,18], improvement mineral and water uptake [19,20], regulating antioxidant enzyme activity [14,20], which boosts plant growth. This positive impact of Si in reducing salinity stress, which has been previously confirmed in intermediate-Si accumulating plant species like cucumber [21] and faba bean [19]. Moreover, Si helps improve the upright position of leaf blades, which enhances light penetration and subsequently boosts photosynthesis [22,23]. Also, Si has been shown to prevent chlorophyll degradation and enlarge leaf area, resulting in greater light availability for photosynthesis [23,24]. Furthermore, enhancing plant growth and productivity in salt-affected plants through the use of Si has been documented in various plant species [11,19,23]
The primary aim of this research was to explore the interactive impact of Si on salt-stressed sunflower plants by managing ionic and osmotic stress. We demonstrate that adding silicon (Si) to the growth medium of sunflower (Helianthus annuus L.) plants can alleviate salt-induced ionic and osmotic stress, further improving dry biomass production. These findings indicate that adding Si to the growth medium significantly boost resistance to salt-induced ionic and osmotic stress, presenting a promising strategy for improving crop growth and management in saline conditions.
2. Results
2.1. Changes on Root and Shoot Dry Biomass and H2O2 Concentration After NaCl and Si Treatments
A two-way ANOVA test revealed significant (p < 0.001) interaction between NaCl and Si on RDM, SDM, and H2O2 concentration [H2O2] (Table 1). Salt stress (100 mM NaCl) evidently decreased SDM of sunflower plants by ~62% and RDM by ~47% compared to the non-salt stressed and Si-untreated plants. However, Si addition in the growth medium significantly (p <0.0001) enhanced the deleterious effects of salt stress and improved by ~27%, 68%, and ~31% the SDM, RDM, and TDM, respectively, compared to salt stressed and Si-untreated plants (Table 1). We also found that salt stress (100 mM NaCl) markedly increased [H2O2] in leaves, which also indicated the adverse effects. Si had no effect on [H2O2] under non-salt-stressed sunflower plants; however, [H2O2] was decreased by ~47% in salt-stressed sunflower under Si addition compared to salt-stressed and Si-untreated plants and showed significant (p < 0.001) differences (Table 1).
2.2. Changes on Photosynthetic Pigments Concentrations After NaCl and Si Treatments
A Two-way analysis revealed that there was significant (p < 0.001) interaction between NaCl and Si treatments on leaves photosynthetic pigments (Table 2, Table S1). Added NaCl in absence of Si decreased ChlA by 70%, ChlB by ~79%, total Chl by ~74%, and carotenoids (CA) by ~94% compared to the non-salt stressed plants without Si addition. However, under salt-stressed plants, Si supplementation significantly increased Chla by ~108%, Chlb by 125%, total Chl by 115%, and CA by ~41% compared to the salt-stressed and Si-untreated plants (Table 2).
2.3. Changes on Accumulation of Macronutrients and Micronutrients in Shoots and Roots After NaCl and Si Treatments
A two-way ANOVA showed changes on accumulation of macronutrients and micronutrients, and exhibited significant (p < 0.0001) interaction between NaCl and Si treatments after 30 d in sunflower plants (Figure 1 and Figure 2, Table S1). Accumulation of macronutrients in shoots and roots were lower in sunflower plants exposed to salt treatment (100 mM NaCl) (Figure 1a-f). In addition, Si application had no effect on the accumulation of N, p, K, Ca, Mg, and S in shoots and roots under non-salt-treated plants. Conversely, Si addition increased and showed a significant (p < 0.0002) difference on the accumulation of N, P, K, Ca, Mg, and S in salt-stressed plants compared to the salt-stressed and Si-untreated plants (Figure 1a-f).
The accumulation of Fe, Mn, Zn, and Cu showed a significant (p < 0.0001) interaction between NaCl and Si treatments (Figure 2a-d, Table S1). Si supplementation reveled higher micronutrients accumulation in shoots and roots of salt-stressed plants compared to Si-untreated. Meanwhile, the micronutrients accumulation increased in shoots and roots of salt-stressed sunflower plants under Si treatments compared to the salt stressed and Si-untreated plants (p < 0.0001) (Figure 2a-d).
3. Discussion
The positive impact of silicon on enhancing the growth of various salt-stressed plant species is widely acknowledged [10,19,25]. In summary, the primary function of silicon in reducing sodium toxicity in plants is largely due to its positive effects on mineral nutrition, physiology, and biochemical processes, which enhance the plant’s tolerance to abiotic stresses [23,26]. Growth characteristics were improved with Si application, especially dry biomass accumulation probably by the high photosynthetic pigments content and nutritional status (Table 1, Figs. 1 and 2). The positive effects of adding silicon, which helps mitigate the harmful impacts of high sodium toxicity on plant growth, have been previously observed in sunflowers [10,14]. Our data on plant growth indicate that sunflower plants exposed to high salinity conditions with silicon (Si) supplementation produced approximately 62% more SDB and 47% more RDB compared to those grown in high salinity without Si. These results might also be interpreted as a consequence of reduced [H2O2], which likely supports plant growth. This enhanced growth of sunflowers under high salinity can be attributed to Si improving the chlorophyll content in the leaves [23]. Furthermore, the enhancement of plant growth by silicon might be associated with the reduction of damages caused by salt stress [20]. In an initial exploration of this topic, we accepted our hypothesis and discovered that adding silicon to the growth medium serves as a sustainable method to enhance plant growth under salt stress. Silicon has been shown to protect chlorophyll from damage, leading to increased leaf area and higher dry biomass production.[36].
As previously mentioned, the hypothesis postulates that providing Si in the growth medium resulted in approximately 70% to 94% more PP per leaf compared to plants subjected to salt stress without Si treatment (Table 2). This finding suggests that Si can alleviate the harmful effects of salt stress by improving the overall photosynthetic machinery [23]. Another possible mechanism through which silicon (Si) enhances plant productivity in salt-stressed conditions may be its capacity to alter cell wall metabolism. This modification promotes cell enlargement by increasing tissue extensibility, which in turn raises the chlorophyll content in leaves [19,25,27]. Moreover, silicon has been shown to prevent chlorophyll degradation and expand leaf area, thereby enhancing the light available for the photosynthesis apparatus. [24]. These results align with our hypothesis and indicate that the enhancement of photosynthesis by silicon represents an economical strategy for plants to manage various salt stress conditions. This might be one reason for the beneficial effects of silicon application in detoxification sodium toxicity.
In this study, we found that salt stress led to elevated toxic substance levels in sunflower leaves, promoting [H2O2] production (Table 1). This effect is due to the high Na+ accumulation [19]. However, supplementing with Si helped the plants maintain lower [H2O2] levels [14]. Similar reductions have also been observed in other research studies involving Si supplementation [8,19,28]. This finding suggests that this phenomenon is a common mechanism by which silicon enhances sodium toxicity. Another possible explanation for the decrease in [H2O2] could be the increase in antioxidant enzyme activity [19,20,23]. Additionally, the other referenced studies indicate that the addition of Si helps maintain low [H2O2] and high membrane stability [29,30]. Additionally, another significant function of Si in alleviating salt stress is the reduction of ROS generation, which consequently improves plant growth [31,32]. This might explain the beneficial impact of silicon application on alleviating salt stress observed in other intermediate Si-accumulating plant species, like cucumbers [33,34] and tomato [35]. Our data directly confirm the initial hypothesis, showing that Si is crucial in boosting resistance to salt-induced osmotic stress.
Our data showed that adding Si during salt stress enhanced the accumulation of macro- and micronutrients in both the leaves and roots (Figs 1 and 2). This might explain why Si treatment is often reported to help reduce Na+ toxicity in numerous studies.[10,23,26,37]. Our observations may have important implications of Si addition enhanced mineral uptake by improving root water uptake in salt stressed plants [16,38,39,40]. Also, it has been proposed that silicon (Si) enhances salt tolerance in plants by stimulating H+-ATPase activity, which leads to increased K+ uptake.[17,26,33]. Besides, Si plays a crucial role in maintaining mineral balance in plants under salt stress, due to its ability to enhance nutrient uptake [19,21], which help to decrease the Na+ uptake [23,36]. Additionally, silicon significantly enhanced the mineral balance under salt stress conditions by promoting ionic [21,43,44] and stoichiometric homeostasis [36]. Furthermore, several studies have reported that silicon (Si) has beneficial effects in enhancing mineral nutrient uptake under high salinity conditions [26,39,41,42]. The findings of this study suggest that the enhancement on mineral nutrition through Si treatment is a prevalent method for mitigating the deleterious effects of sodium toxicity. Consistent with the observed results, this can be directly linked to an increase in dry biomass, supporting our hypothesis that Si represents an economical approach for plants to cope with salt stress.
4. Materials and Methods
The research was conducted at the School of Agricultural and Veterinary Sciences (FCAV), São Paulo State University–UNESP, located on the Jaboticabal Campus (21° 15′ 22″ S and 48°18′ 58″ W) in Brazil. The study utilized the yellow dwarf sunflower (H. annuus L. ‘Double Sungold’).
Five seeds were evenly placed in pots (4-dm-3 polyethylene containers) and watered daily with demineralized water. The pots were filled with washed sand (3.7-dm−3) and a quarter-strength modified Hoagland nutrient solution (NS) [45] was first applied with the pH adjusted to approximately 5.8 for one week. Then, the NS was increased to half-strength for two weeks, followed by an increase to three-quarters strength until the experiment concluded. Two seven-day-old seedlings were removed, leaving three uniform seedlings.
A pot experiment was carried out in a glasshouse with natural lighting. Daytime temperatures peaked at around 33°C, while nighttime temperatures dropped to 20°C. The relative humidity ranged from 60% to 75%, and the photoperiod was set to 14 hours of light and 10 hours of darkness. The experiment followed a randomized block design with a factorial arrangement (2 × 2), featuring five replicates for each treatment. The pots were organized into two groups: non-salt stress and salt stress (100 mM NaCl), combined with either the Si-untreated and Si-treated (2 mM) plants, applied as stabilized sodium and potassium silicate (SINAKE). This resulted in four treatment combinations and a total of 40 pots.
After a week of acclimating the plants, salt treatment was initiated by adding 100 mM NaCl to the nutrient solution (NS) and continued for 30 experimental days. Si treatment was administered through the NS throughout the entire experimental period, following the method outlined by Calero et al. [10]. Thirty days after beginning the salinity treatments, the levels of photosynthetic pigments such as chlorophyll a (Chla), chlorophyll b (Chlb), total chlorophyll (Chlt), and carotenoids (CA) were measured as previously described by Lichtenthale et al. [46,47]. Leaf discs were collected from the first fully expanded leaf blade (from the top) and placed in 10 mL of 80% acetone (v/v). The mixture was then centrifuged at 4000 g at 4 °C for 10 minutes. The supernatant was separated and utilized for the chlorophyll assay. Individual samples were analyzed in triplicate.
Subsequently, leaf of sunflower seedlings was collected to determination of H2O2 concentration. Then, Alexieva et al. [48] methodology was used to estimate H2O2 concentration. Briefly, 0.2 g fresh leaves were homogenized in trichloroacetic acid (TCA) at 0.1%, and centrifuged at 11,000 × g for 15 min. The supernatant was added to the potassium phosphate buffer 100 mM (pH 7.50) and the potassium iodide solution 1.0 M. Then, the material was incubated on ice bath for 1 h, the absorbance read at 390 nm, and the H2O2 concentration was determined using a H2O2 concentration curve known as the standard curve. The H2O2 results were expressed in μMol g−1 of fresh matter (FM).
Afterward, the sunflower harvest began by cutting the plants at the soil surface and separating them into roots and shoots. The roots were washed to remove excess sand particles. Both roots and shoots were then sequentially washed in demineralized water, a 0.2% neutral detergent solution, a 0.1% HCl solution, and twice in deionized water [44]. The samples were placed in an oven (TE 394-3, Tecnal, Piracicaba, São Paulo, Brazil) and dried at 60 °C until a consistent dry mass was achieved. The root and shoot biomass were promptly measured using a digital scale (Q31711-53/Quimis—Brazil). These oven-dried samples were then ground into a fine powder. Wet digestion of the dried sunflower plant material was carried out following the method of Bataglia et al. [49]. 0.1 g of dried, ground plant material was digested using 7 mL of concentrated H2SO4 and 1.5 mL of a diacid mixture composed of nitric and perchloric acids in a 3:1 ratio. The extract was then diluted to a total volume of 30 mL in volumetric flasks to determine the concentrations of macro- (g kg−1) and micronutrients (mg kg−1). The accumulation of each nutrient was calculated by multiplying its concentration by the corresponding dry biomass [10].
All data were subjected to normality distribution checks using the Kolmogorov-Smirnov test and Levene test (p < 0.05) for unequal variance. After confirming normality and unequal variance, a two-way ANOVA was performed to examine the main effects of two levels of NaCl (Na) and two levels of Si (Si), and their interaction (Na × Si). When the F test indicated significant differences among treatments, mean comparisons were carried out using the Scott-Knott test (p < 0.05). All analyses were conducted using the statistical software R v. 4.4.[50].
5. Conclusions
In conclusion, our study assumes the hypothesis that Si supplementation is an economic and alternative efficient to ameliorate the growth of salt-stressed sunflower plants. The decreasing H2O2 concentration and increasing photosynthetic pigments and nutritional status is an important mechanism of Si to improving the growth of salt-stressed sunflower seedlings. Our findings suggested that Si-supplemented plants play an important role in detoxifying sodium toxicity by regulating ionic and osmotic stress, which lead to increase growth production.
Supplementary Materials
The following supporting information can be downloaded at website of this paper posted on Preprints.org, Table S1: Results of the two-way ANOVA of the effects of NaCl, Si, and their interaction on shoot and root dry biomass, H2O2 concentration, photosynthetic pigments, and shoot and root macros- and micronutrient accumulations (n = 5).
Author Contributions
Conceptualization, A.C.H. and D.A.C.; methodology, R.M.P.; validation, A.C.H., D.A.C. and R.M.P.; formal analysis, A.C.H.; investigation, A.C.H., D.A.C.; K.P.C.; D.O.V. and G.S.S.J.; resources, R.M.P.; data curation, A.C.H.; writing—original draft preparation, A.C.H.; writing—review and editing, A.C.H., R.M.P.; Y.P.D. K.P.C.; D.O.V.; visualization, X.X.; supervision, A.C.H.; project administration, R.M.P.; funding acquisition, A.C.H. All authors have read and agreed to the published version of the manuscript.
Funding
Work in the CamposLab was funded by the Brazilian Federal Agency for the Support and Evaluation of Graduate Education (CAPES – Grant no. 88887.975003/2024-00, XXXXX and Fundação de Amparo a Pesquisa do Estado de Mato Grosso (FAPEMAT – Grant no. 000258/2023).
Data Availability Statement
All relevant data are within the paper.
Acknowledgments
The authors are grateful to the São Paulo State University -UNESP, School of Agricultural and Veterinarian Sciences-FCAV, for providing the necessary facilities for this study. We would also like to thank GENPLANT research for their technical support of the project. We wish to thank the Brazilian Federal Agency for the Support and Evaluation of Graduate Education (CAPES) for providing the fellowship for the first author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Abbas, T.; Balal, R.; Shahid, M.; Pervez, M.; Ayyub, C.; Aqueel, M.A.; Javaid, M. Silicon-Induced Alleviation of NaCl Toxicity in Okra (Abelmoschus Esculentus) Is Associated with Enhanced Photosynthesis, Osmoprotectants and Antioxidant Metabolism. Acta Physiol. Plant. 2015, 37, 1–15. [Google Scholar] [CrossRef]
- Siddiqui, M.N.; Mostofa, M.G.; Akter, M.M.; Srivastava, A.K.; Sayed, M.A.; Hasan, M.S.; Tran, L.-S.P. Impact of Salt-Induced Toxicity on Growth and Yield-Potential of Local Wheat Cultivars: Oxidative Stress and Ion Toxicity Are among the Major Determinants of Salt-Tolerant Capacity. Chemosphere 2017, 187, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Munns, R.; James, R.; Gilliham, M.; Flowers, T.; Colmer, T. Tissue Tolerance: An Essential but Elusive Trait for Salt-Tolerant Crops. Funct. Plant Biol. 2016, 43, 1103–1113. [Google Scholar] [CrossRef]
- Munns, R.; Gilliham, M. Salinity Tolerance of Crops - What Is the Cost ? Tansley Insight Salinity Tolerance of Crops – What Is the Cost ? New Phytol. 2015, 208, 668–673. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of Salinity Tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef]
- Kim, Y.-H.; Khan, A.L.; Waqas, M.; Lee, I.-J. Silicon Regulates Antioxidant Activities of Crop Plants under Abiotic-Induced Oxidative Stress: A Review. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef]
- Kaya, M.D.; Akdoğan, G.; Kulan, E.G.; Dağhan, H.; Sari, A. Salinity Tolerance Classification of Sunflower (Helianthus Annuus L.) and Safflower (Carthamus Tinctorius L.) by Cluster and Principal Component Analysis. Appl. Ecol. Environ. Res. 2019, 17, 3849–3857. [Google Scholar] [CrossRef]
- Oliveira, R.L.L. de; Prado, R. de M.; Felisberto, G.; Checchio, M.V.; Gratão, P.L. Silicon Mitigates Manganese Deficiency Stress by Regulating the Physiology and Activity of Antioxidant Enzymes in Sorghum Plants. J. Soil Sci. Plant Nutr. 2019, 1–11. [Google Scholar] [CrossRef]
- Santos Silva, B.; de MELLO PRADO, R.; Calero Hurtado, A.; APARECIDA DE ANDRADE, R.; PEREIRA DA SILVA, G. Ammonia Toxicity Affect Cations Uptake and Growth in Papaya Plants Inclusive with Silicon Addition. Acta Biológica Colomb. 2020, 25, 345–353. [Google Scholar] [CrossRef]
- Calero Hurtado, A.; Chiconato, D.A.; Prado, R. de M.; Sousa Junior, G. da S.; Felisberto, G. Silicon Attenuates Sodium Toxicity by Improving Nutritional Efficiency in Sorghum and Sunflower Plants. Plant Physiol. Biochem. 2019, 142, 224–233. [Google Scholar] [CrossRef]
- Alsaeedi, A.; El-Ramady, H.; Alshaal, T.; El-Garawany, M.; Elhawat, N.; Al-Otaibi, A. Silica Nanoparticles Boost Growth and Productivity of Cucumber under Water Deficit and Salinity Stresses by Balancing Nutrients Uptake. Plant Physiol. Biochem. 2019, 139, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Deshmukh, R.; Sonah, H.; Belanger, R. New Evidence Defining the Evolutionary Path of Aquaporins Regulating Silicon Uptake in Land Plants. J. Exp. Bot. 2020. [Google Scholar] [CrossRef]
- Liang, Y.; Sun, W.; Zhu, Y.G.; Christie, P. Mechanisms of Silicon-Mediated Alleviation of Abiotic Stresses in Higher Plants: A Review. Environ. Pollut. 2007, 147, 422–428. [Google Scholar] [CrossRef]
- Conceição, S.; Neto, C.; Marques, E.; Barbosa, A.; Galvão, J.; de Oliveira, T.; Okumura, R.; Martins, J.; Costa, T.; Gomes-Filho, E. Silicon Modulates the Activity of Antioxidant Enzymes and Nitrogen Compounds in Sunflower Plants under Salt Stress. Arch. Agron. Soil Sci. 2019, 65, 1237–1247. [Google Scholar] [CrossRef]
- Bosnic, P.; Bosnic, D.; Jasnic, J.; Nikolic, M. Silicon Mediates Sodium Transport and Partitioning in Maize under Moderate Salt Stress. Environ. Exp. Bot. 2018, 155, 681–687. [Google Scholar] [CrossRef]
- Flam-Shepherd, R.; Huynh, W.Q.; Coskun, D.; Hamam, A.M.; Britto, D.T.; Kronzucker, H.J. Membrane Fluxes, Bypass Flows, and Sodium Stress in Rice: The Influence of Silicon. J. Exp. Bot. 2018, 69, 1679–1692. [Google Scholar] [CrossRef]
- Liang, Y.; Zhang, W.; Chen, Q.; Liu, Y.; Ding, R. Effect of Exogenous Silicon (Si) on H+-ATPase Activity, Phospholipids and Fluidity of Plasma Membrane in Leaves of Salt-Stressed Barley (Hordeum Vulgare L.). Environ. Exp. Bot. 2006, 57, 212–219. [Google Scholar] [CrossRef]
- Kumar, S.; Milstein, Y.; Brami, Y.; Elbaum, M.; Elbaum, R. Mechanism of Silica Deposition in Sorghum Silica Cells. New Phytol. 2017, 213, 791–798. [Google Scholar] [CrossRef]
- Ahmad, P.; Abass, M.; Alam, P.; Nasser, M.; Wijaya, L.; Ali, S.; Ashraf, M. Silicon (Si) Supplementation Alleviates NaCl Toxicity in Mung Bean [Vigna Radiata (L.) Wilczek] through the Modifications of Physio-Biochemical Attributes and Key Antioxidant Enzymes. J. Plant Growth Regul. 2019, 38, 70–82. [Google Scholar] [CrossRef]
- Calero Hurtado, A.; Chiconato, D.A.; Prado, R. de M.; Sousa Junior, G. da S.; Gratão, P.L.; Felisberto, G.; Olivera Viciedo, D.; Mathias dos Santos, D.M. Different Methods of Silicon Application Attenuate Salt Stress in Sorghum and Sunflower by Modifying the Antioxidative Defense Mechanism. Ecotoxicol. Environ. Saf. 2020, 203, 110964. [Google Scholar] [CrossRef]
- Alsaeedi, A.; El-Ramady, H.; Alshaal, T.; El-Garawani, M.; Elhawat, N.; Al-Otaibi, A. Exogenous Nanosilica Improves Germination and Growth of Cucumber by Maintaining K+/Na+ Ratio under Elevated Na+ Stress. Plant Physiol. Biochem. 2018, 125, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.X.; Xia, Y.C.; Liu, L.C.; Yin, J.L.; Ma, D.F. Beneficial Effects of Silicon on Salt Tolerance in Plants. J. Plant Nutr. Fertil. 2019, 25, 498–509. [Google Scholar] [CrossRef]
- Alzahrani, Y.; Kuşvuran, A.; Alharby, H.F.; Kuşvuran, S.; Rady, M.M. The Defensive Role of Silicon in Wheat against Stress Conditions Induced by Drought, Salinity or Cadmium. Ecotoxicol. Environ. Saf. 2018, 154, 187–196. [Google Scholar] [CrossRef]
- Agarie, S.; Agata, W.; Kubota, H.; Kaufmann, P.B. Physiological Role of Silicon in Photosynthesis and Dry Matter Production in Rice Plants. Crop Sci. 1992. [Google Scholar]
- Etesami, H.; Jeong, B.R. Silicon (Si): Review and Future Prospects on the Action Mechanisms in Alleviating Biotic and Abiotic Stresses in Plants. Ecotoxicol. Environ. Saf. 2018, 147, 881–896. [Google Scholar] [CrossRef] [PubMed]
- Rios, J.J.; Martínez-Ballesta, M.C.; Ruiz, J.M.; Blasco, B.B.; Carvajal, M.; Martinez-Ballesta, M.C.; Ruiz, J.M.; Blasco, B.B.; Carvajal, M. Silicon-Mediated Improvement in Plant Salinity Tolerance: The Role of Aquaporins. Front. Plant Sci. 2017, 8, 1–10. [Google Scholar] [CrossRef]
- Liu, B.; Soundararajan, P.; Manivannan, A. Mechanisms of Silicon-Mediated Amelioration of Salt Stress in Plants. Plants 2019, 8, 307. [Google Scholar] [CrossRef]
- Fialová, I.; Šimková, L.; Vaculíková, M.; Luxová, M. Effect of Si on the Antioxidative Defense of Young Maize Roots under NaCl Stress. Silicon 2018, 10, 2911–2914. [Google Scholar] [CrossRef]
- Zhu, Z.; Wei, G.; Li, J.; Qian, Q.; Yu, J. Silicon Alleviates Salt Stress and Increases Antioxidant Enzymes Activity in Leaves of Salt-Stressed Cucumber (Cucumis Sativus L.). Plant Sci. 2004, 167, 527–533. [Google Scholar] [CrossRef]
- Gomathi, R.; Rakkiyapan, P. Comparative Lipid Peroxidation, Leaf Membrane Thermostability, and Antioxidant System in Four Sugarcane Genotypes Differing in Salt Tolerance. Int. J. Plant Physiol. Biochem. 2011, 3, 67–74. [Google Scholar]
- Coskun, D.; Britto, D.T.; Huynh, W.Q.; Kronzucker, H.J. The Role of Silicon in Higher Plants under Salinity and Drought Stress. Front. Plant Sci. 2016, 7, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Coskun, D.; Deshmukh, R.; Sonah, H.; Menzies, J.G.; Reynolds, O.; Ma, J.F.; Kronzucker, H.J.; Bélanger, R.R. The Controversies of Silicon’s Role in Plant Biology. New Phytol. 2019, 221, 67–85. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Zhang, W.; Chen, Q.; Ding, R. Effects of Silicon on H+-ATPase and H+-PPase Activity, Fatty Acid Composition and Fluidity of Tonoplast Vesicles from Roots of Salt-Stressed Barley (Hordeum Vulgare L.). Environ. Exp. Bot. 2005, 53, 29–37. [Google Scholar] [CrossRef]
- Zhu, Y.; Xu, X.; Hu, Y.; Han, W.; Yin, J.; Li, H.; Gong, H. Silicon Improves Salt Tolerance by Increasing Root Water Uptake in Cucumis Sativus L. Plant Cell Rep. 2015, 34, 1629–1646. [Google Scholar] [CrossRef]
- Eraslan, F.; Inal, A.; Pilbeam, D.J.; Gunes, A. Interactive Effects of Salicylic Acid and Silicon on Oxidative Damage and Antioxidant Activity in Spinach (Spinacia Oleracea L. Cv. Matador) Grown under Boron Toxicity and Salinity. Plant Growth Regul. 2008, 55, 207–219. [Google Scholar] [CrossRef]
- Calero Hurtado, A.; Aparecida Chiconato, D.; de Mello Prado, R.; da Silveira Sousa Junior, G.; Olivera Viciedo, D.; de Cássia Piccolo, M. Silicon Application Induces Changes C:N:P Stoichiometry and Enhances Stoichiometric Homeostasis of Sorghum and Sunflower Plants under Salt Stress. Saudi J. Biol. Sci. 2020. In press. [Google Scholar] [CrossRef]
- Khoshgoftarmanesh, A.H.; Khodarahmi, S.; Haghighi, M. Effect of Silicon Nutrition on Lipid Peroxidation and Antioxidant Response of Cucumber Plants Exposed to Salinity Stress. Arch. Agron. Soil Sci. 2014, 60, 639–653. [Google Scholar] [CrossRef]
- Gurmani, A.; Bano, A.; Ullah, N.; Khan, H.; Jahangir, M.; Flowers, T.J. Exogenous Abscisic Acid (ABA) and Silicon (Si) Promote Salinity Tolerance by Reducing Sodium (Na+) Transport and Bypass Flow in Rice (“Oryza Sativa” Indica). Aust. J. Crop Sci. 2013, 7, 1219–1226. [Google Scholar]
- Liu, P.; Yin, L.; Wang, S.; Zhang, M.; Deng, X.; Zhang, S.; Tanaka, K. Enhanced Root Hydraulic Conductance by Aquaporin Regulation Accounts for Silicon Alleviated Salt-Induced Osmotic Stress in Sorghum Bicolor L. Environ. Exp. Bot. 2015, 111, 42–51. [Google Scholar] [CrossRef]
- Li, H.; Zhu, Y.; Hu, Y.; Han, W.; Gong, H. Beneficial Effects of Silicon in Alleviating Salinity Stress of Tomato Seedlings Grown under Sand Culture. Acta Physiol. Plant. 2015, 37, 1–9. [Google Scholar] [CrossRef]
- Yin, L.; Wang, S.; Tanaka, K.; Fujihara, S.; Itai, A.; Den, X.; Zhang, S. Silicon-Mediated Changes in Polyamines Participate in Silicon-Induced Salt Tolerance in Sorghum Bicolor L. Plant Cell Environ. 2016, 39, 245–258. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.F. Characterization of the System and Molecular Mapping of the Silicon Transporter Gene in Rice. Plant Physiol. 2004, 136, 3284–3289. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.X.; Ma, Y.P.; Liu, Y.L. Effects of Silicon (Si) on Growth, Quality and Ionic Homeostasis of Aloe under Salt Stress. South African J. Bot. 2015, 98, 26–36. [Google Scholar] [CrossRef]
- Hurtado, A.C.; Chiconato, D.A.; de Mello Prado, R.; Sousa Junior, G. da S.; Viciedo, D.O.; Díaz, Y.P.; Peña Calzada, K.; Gratão, P.L. Silicon Alleviates Sodium Toxicity in Sorghum and Sunflower Plants by Enhancing Ionic Homeostasis in Roots and Shoots and Increasing Dry Matter Accumulation. Silicon 2020, 12, In. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. The Water-Culture Method for Growing Plants without Soil. Circ. Calif. Agric. Exp. Stn. 1950, 347, 1–32. [Google Scholar]
- Lichtenthaler, H.K. Chlorophylls and Carotenoids: Pigments of Photosynthetic Biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Buschmann, C.; Knapp, M. How to Correctly Determine the Different Chlorophyll Fluorescence Parameters and the Chlorophyll Fluorescence Decrease Ratio RFd of Leaves with the PAM Fluorometer. Photosynthetica 2005, 43, 379–393. [Google Scholar] [CrossRef]
- Alexieva, V.; Sergiev, I.; Mapelli, S.; Karanov, E. The Effect of Drought and Ultraviolet Radiation on Growth and Stress Markers in Pea and Wheat. Plant. Cell Environ. 2001, 24, 1337–1344. [Google Scholar] [CrossRef]
- Bataglia, O.C.; Teixeira, J.P.F.; Furlani, P.R.; Furlani, A.M.C.; Gallo, J.R. Métodos de Análise Química de Plantas; 1st ed.; Instituto Agronômico de Campinas: Campinas, 1983. [Google Scholar]
- Team, R.C. A Language and Environment for Statistical Computing. R Found. Stat. Comput. 2023.
Figure 1.
Shoot and root N accumulation (a), shoot and root P accumulation (b), shoot and root K accumulation (c), shoot and root Ca accumulation (d), shoot and root Mg accumulation (e), and shoot and root S accumulation (f), grown under NaCl (0 and 100 mM NaCl) treatments combined with the absence and presence (2 mM) of Si. Data are means ± standard deviation (SD) (n = 5). Different normal small letters (e.g., a, b) or italic small letters (e.g., a, b, c) indicate significant differences between Si treatments under non-salt stress and salt stress conditions, respectively; and different uppercase letters (e.g., A, B) show significant differences between non-salt and salt treatments at the same Si treatment (e.g., 0 mM or 2 mM), according to Scott–Knott test (p < 0.05). F values and significance (**p < 0.01) from a two-way ANOVA). Si × NaCl, NaCl–Si interaction.
Figure 1.
Shoot and root N accumulation (a), shoot and root P accumulation (b), shoot and root K accumulation (c), shoot and root Ca accumulation (d), shoot and root Mg accumulation (e), and shoot and root S accumulation (f), grown under NaCl (0 and 100 mM NaCl) treatments combined with the absence and presence (2 mM) of Si. Data are means ± standard deviation (SD) (n = 5). Different normal small letters (e.g., a, b) or italic small letters (e.g., a, b, c) indicate significant differences between Si treatments under non-salt stress and salt stress conditions, respectively; and different uppercase letters (e.g., A, B) show significant differences between non-salt and salt treatments at the same Si treatment (e.g., 0 mM or 2 mM), according to Scott–Knott test (p < 0.05). F values and significance (**p < 0.01) from a two-way ANOVA). Si × NaCl, NaCl–Si interaction.
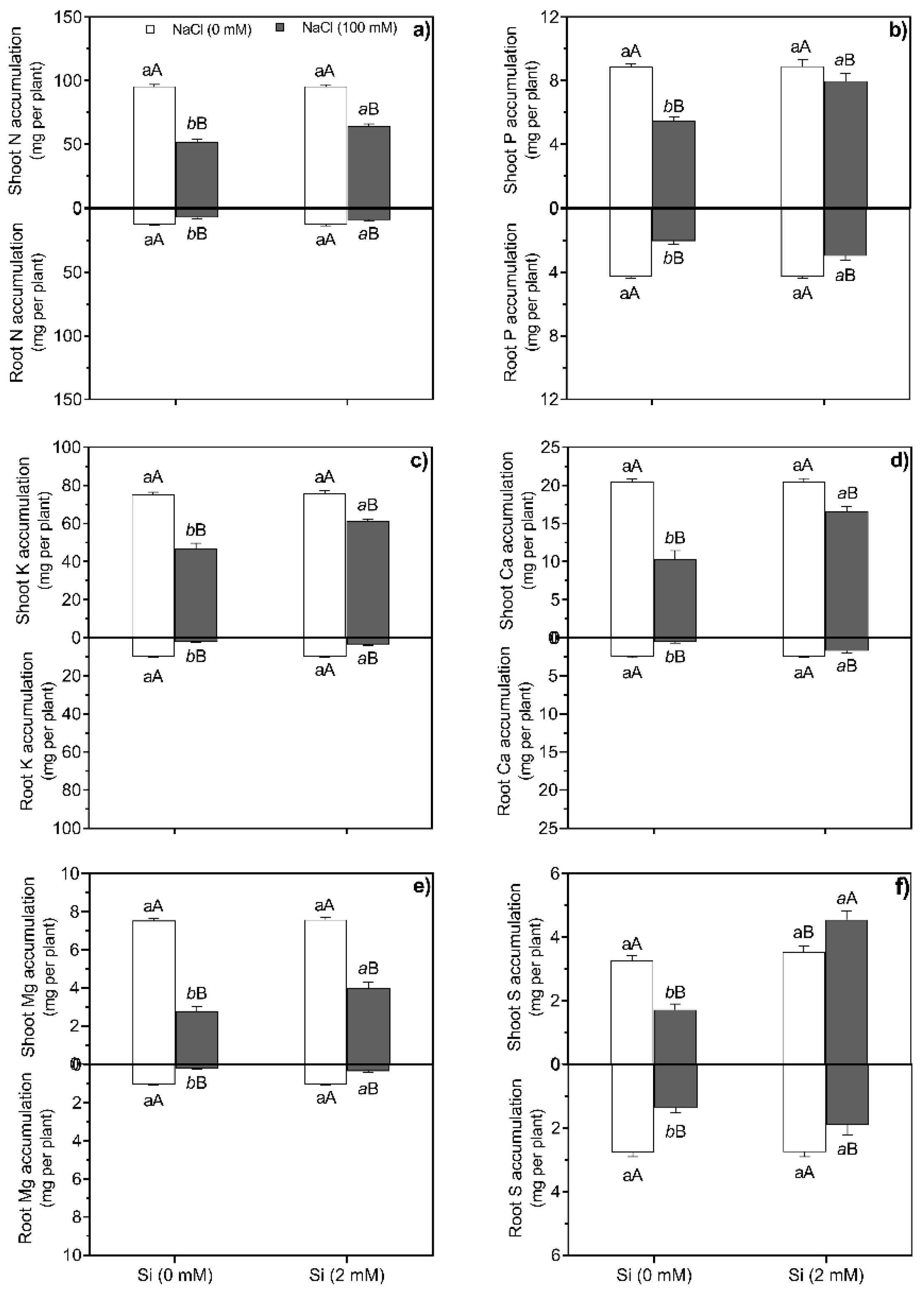
Figure 2.
Accumulation of micronutrients in shoots and roots of sunflower plants. Shoot and root Fe accumulation (a), shoot and root Mn accumulation (b), shoot and root Zn accumulation (c), shoot and root Cu accumulation (d), grown under non-salt and salt stress (100 mM NaCl) treatments combined with the absence and presence (2 mM) of Si. Data are means ± standard deviation (SD) (n = 5). Different normal small letters (e.g., a,b) in the same column indicate significant differences among Si treatments under non-salt stress; different italic small letters (e.g., a,b) shown significant differences among Si treatments under salt stress conditions and different uppercase letters (e.g., A, B) indicate significant differences between non-salt and salt treatments at the same level of Si, according to the Scott–Knott test (p < 0.05). (F values from ANOVA: **p < 0.01). NaCl × Si, NaCl–Si interaction.
Figure 2.
Accumulation of micronutrients in shoots and roots of sunflower plants. Shoot and root Fe accumulation (a), shoot and root Mn accumulation (b), shoot and root Zn accumulation (c), shoot and root Cu accumulation (d), grown under non-salt and salt stress (100 mM NaCl) treatments combined with the absence and presence (2 mM) of Si. Data are means ± standard deviation (SD) (n = 5). Different normal small letters (e.g., a,b) in the same column indicate significant differences among Si treatments under non-salt stress; different italic small letters (e.g., a,b) shown significant differences among Si treatments under salt stress conditions and different uppercase letters (e.g., A, B) indicate significant differences between non-salt and salt treatments at the same level of Si, according to the Scott–Knott test (p < 0.05). (F values from ANOVA: **p < 0.01). NaCl × Si, NaCl–Si interaction.
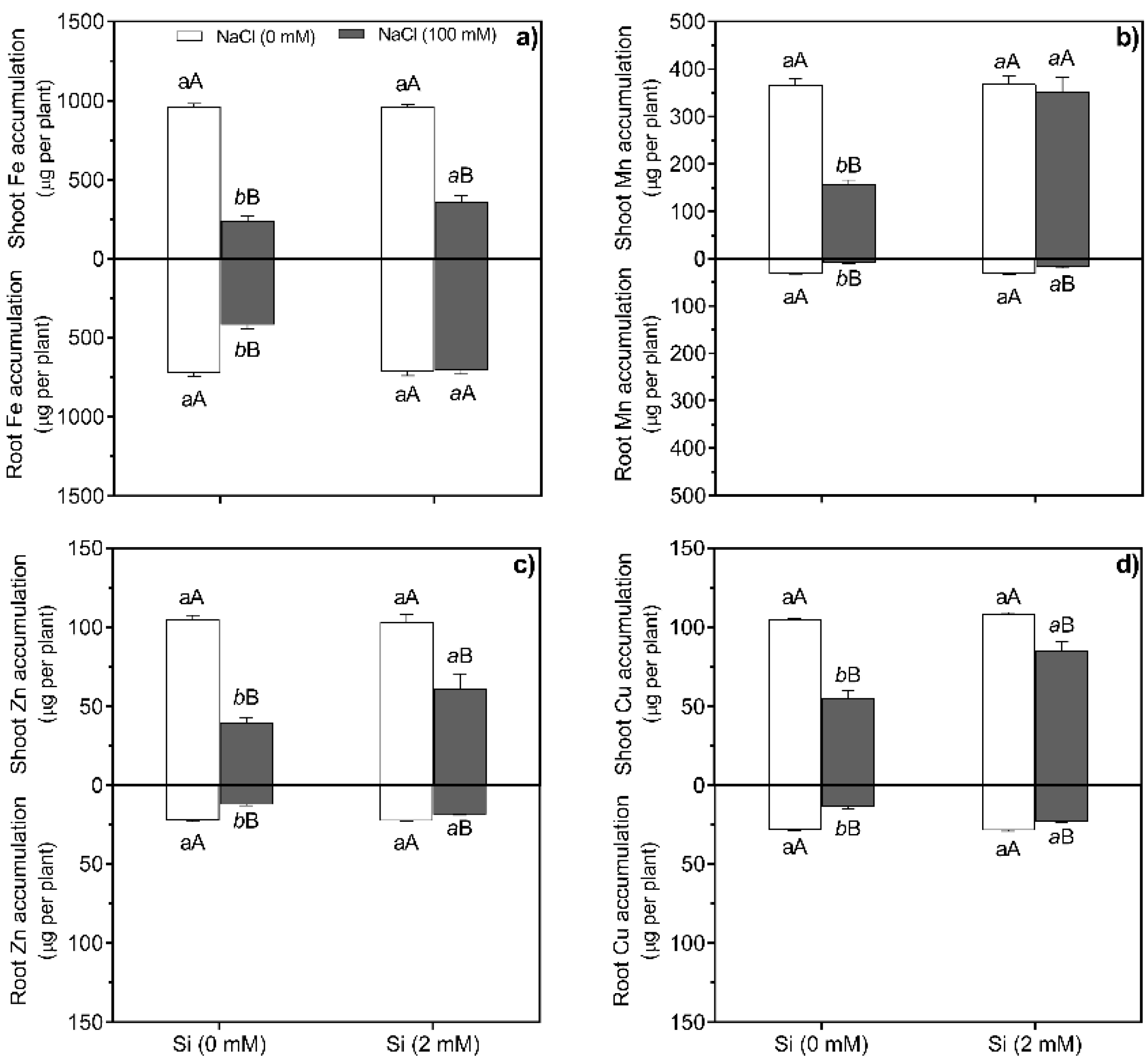

Table 1.
Effect of absence or presence (2 mM) of Si on H2O2 concentration, root dry biomass (RDB) and shoot dry biomass (SDB) of sunflower plants grown in non and salt stress (100 mM NaCl) conditions.
Table 1.
Effect of absence or presence (2 mM) of Si on H2O2 concentration, root dry biomass (RDB) and shoot dry biomass (SDB) of sunflower plants grown in non and salt stress (100 mM NaCl) conditions.
| Treatments | H2O2 concentration | RDM | SDM | |
| NaCl | Si | |||
| mM | µMol g−1 FM | g plant−1 | ||
| 0 | 0 | 6.97 ± 0.27 aB | 0.95 ± 0.01aA | 2.46 ± 0.07bA |
| 2 | 6.79 ± 0.14 aB | 0.97 ± 0.03aA | 2.48 ± 0.06bA | |
| 100 | 0 | 22.15 ± 0.51 aA | 0.29 ± 0.03bB | 1.35 ± 0.05bB |
| 2 | 11.78 ± 0.34 bA | 0.46 ± 0.03aB | 1.73 ± 0.07aB | |
Notes: Data are means ± standard deviation (SD) (n = 5). NaCl (0 mM); NaCl (100 mM); Si (0 mM); Si (2 mM). Different normal small letters (e.g., a, b) or italic small letters (e.g., a, b, c) in the same column indicate significant differences between Si treatments under non-salt stress and salt stress conditions, respectively; and different uppercase letters (e.g., A, B) indicate significant differences between non-salt and salt treatments at the same Si treatment (e.g., 0 or 2 mM), according to Scott–Knott test (p < 0.05). (F values and significance (**p < 0.01) from a two-way ANOVA). Si × NaCl, NaCl–Si interaction. FM, fresh matter.
Table 2.
Effect of absence or presence (2 mM) of Si on photosynthetic pigments (Chla, Chlb, Chlt, and CA) concentrations in sunflower plants grown under non and salt stress (100 mM NaCl) conditions.
Table 2.
Effect of absence or presence (2 mM) of Si on photosynthetic pigments (Chla, Chlb, Chlt, and CA) concentrations in sunflower plants grown under non and salt stress (100 mM NaCl) conditions.
| Treatments | Photosynthetic pigments | ||||
|---|---|---|---|---|---|
| NaCl | Si | Chla | Chlb | Total Chl | CA |
| mM | µg cm−2 | ||||
| 0 | 0 | 0.40 ± 0.01 aA | 0.38 ± 0.01 aA | 0.78 ± 0.01 aA | 3.04 ± 0.01 aA |
| 2 | 0.41 ± 0.01 aA | 0.39 ± 0.01 aA | 0.80 ± 0.02 aA | 3.06 ± 0.02 aA | |
| 100 | 0 | 0.12 ± 0.01 bB | 0.08 ± 0.01 bB | 0.20 ± 0.02 bB | 1.57 ± 0.03 bB |
| 2 | 0.25 ± 0.01 aB | 0.18 ± 0.01 aB | 0.43 ± 0.01 aB | 2.21 ± 0.02 aB | |
Notes: Data are means ± standard deviation (SD) (n = 5). Chla, chlorophyll a; Chlb, chlorophyll b; Chlt, chlorophyll (a + b); CA, carotenoids. Different normal small letters (e.g., a, b) or italic small letters (e.g., a, b, c) in the same column indicate significant differences between Si treatments under non-salt stress and salt stress conditions, respectively; and different uppercase letters (e.g., A, B) indicate significant differences between non-salt and salt treatments at the same Si treatment (e.g., 0 mM or 2 mM), according to Scott–Knott test (p < 0.05). (F values and significance (**: p < 0.01) from a two-way ANOVA). Si × NaCl, NaCl–Si interaction.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



