
Submitted:
18 October 2024
Posted:
21 October 2024
You are already at the latest version
Abstract
Background: the red palm weevil, Rhynchophorus ferrugineus (F.) (Col.: Curculionidae: Dryophthorinae) is a pest species that infests many palm and coconut tree species across 50 countries. Larval feeding damages the palm heart and/or the palm stipe meristem, often causing the tree’s death. The damage caused by the larvae also allows phytopathogens and other insect pests to enter the tree, which can prove lethal. Methods: two active ingredients, azadirachtin and boric acid, were incorporated into an artificial diet fed to the weevils. The efficacy of these compounds was evaluated through bioassays conducted under laboratory conditions. Results: the most effective concentration for azadirachtin (95.0±5.0 %) was observed at 50.0 ppm on day 20; for boric acid (100 %), it was most efficacious at 2,500.0 ppm on day 12. Conclusions: boric acid shows significant potential in controlling the larvae of this weevil pest species. It does so in two ways - firstly, through its direct action against the red palm weevil larvae, and secondly, by its interaction with the micro-organisms associated with the larvae, which may result in more rapid mortality amongst the pest larvae themselves.
Keywords:
Red Palm Weevil
; palm pest
; larval stage
; chemical control
; effectiveness
; microbiota
1. Introduction
The red palm weevil Rhynchophorus ferrugineus (Fabricius) (Col.: Curculionidae: Dryophthorinae) is a pest species that affects numerous palm and coconut tree species in at least 50 countries. It’s distribution has been recorded over four continents: Africa (6 countries) - Djibouti, Egypt, Libya, Mauritania, Morocco, and Tunisia; The Americas: (3 countries) - Aruba, Guadeloupe, and the Netherlands Antilles; Asia (25 countries) - Bahrain, Bangladesh, Cambodia, China, India, Iran, Iraq, Israel, Japan, Kuwait, Lebanon, Malaysia, Myanmar, Oman, Pakistan, The Philippines, Qatar, Saudi Arabia, Sri Lanka, Syria, Taiwan, Thailand, United Arab Emirates, Vietnam, and Yemen; and Europe (15 countries) - Albania, Bosnia and Herzegovina, Bulgaria, Croatia, Cyprus, France, Georgia, Greece, Italy, Malta, Montenegro, Portugal, Russia, Spain, and Turkey [1]. It has also recently been found in Uruguay [2]. The biology and ecology of the red palm weevil have been reviewed in several publications (e.g., [3,4,5,6]).
The first symptoms of R. ferrugineus damage are exceedingly difficult to detect during the initial stages of infestation. Therefore, emphasis should be placed on preventive methods although this is not always possible. The most common approach is the chemical control of this pest species using insecticides [7]; however, again, this has proven difficult to implement due to the location of the larvae deep inside the stipe. Successful chemical control depends on: (i) the efficacy of the active ingredient (e.g., [8,9]); (ii) the timing of the application (e.g., [10]); (iii) the location of the palm plants – in an urban environment or in the field – due to the hazardous nature of some of the active ingredients (e.g., [11]); and (iv) the application method (e.g., [12,13,14]).
2. Materials and Methods
The insects used in these trials came from a colony kept in the Agricultural Entomology Laboratory at the University of Almeria.
The R. ferrugineus larvae were reared on an artificial diet that was optimal from a biological [3] and physiological standpoint [17]. The management of the species was conducted according to Martin and Cabello’s method [18].
A completely randomized experimental design was employed for the two trials. The method used in the two bioassays was described by Cabello et al. [8] and Barranco et al. [9]. The active ingredients evaluated (at the doses mentioned below) were incorporated into the artificial diet at the time of its preparation. The two active ingredients tested were boric acid (boric acid 99.5% wp, Panreac Quimica S.A., Castellar del Valles, Barcelona, Spain) and azadirachtin (Align 3.2% ls, Sipcam Inagra S.A., Valencia, Spain), with the following active ingredient doses per liter: 6.25, 12.5, 25 and 50 ppm of azadirachtin and 312.5, 625, 1250, 2500 and 5000 ppm of boric acid, respectively. Four replicates per treatment were performed for each bioassay. For each replicate, five second-instar larvae were taken from the colony and placed in Coulter vials (20 ml) containing a portion of the artificial diet (10-11 g) with the corresponding dose of active ingredient. The bioassays were conducted in a controlled environment cabinet at 25±2 ºC, 65±10% R.H. and a photoperiod of total darkness (0:24h; Light: Dark). Neither of the active ingredients added to the diet was used in the controls. Larval mortality was evaluated at 3, 6, 9, 12, 15 and 20 days after the start of the trials.
The number of live larvae and mortality percentage values were analyzed using generalized linear models (GZLM), having Poisson as the probability distribution and logarithm as the link function, in which dose was considered as a variable and days as a covariate. IBM SPSS version 28 software was used to carry out this analysis.
3. Results
When azadirachtin was used as the active ingredient, the number of live larvae did fall dramatically from day 15, but only at the 50 ppm dose. In contrast, when boric acid was the active ingredient, all the larvae died from day 12 at the two highest doses - 2,500 and 5,000 ppm (Table 1).
When analyzing the number of live larvae, model significance was found for the active ingredient azadirachtin (Omnibus test: Chi-square = 35.90, df = 5; P < 0.01) with significant effects for the dose variable (Chi-square = 17.54; df = 4; P < 0.01) and the day covariate (Chi-square = 18.36, df = 1; P < 0.01). For boric acid as the active ingredient, model significance was high (Omnibus test: Chi-square = 143.36, df = 6; P < 0.01), with effects for the dose variable (Chi-square = 112.39; df = 5; P < 0.01) and the day covariate (Chi-square = 30.97, df = 1; P < 0.01).
In the control treatments, none of the larvae died during the two bioassays (Table 1). Therefore, the mortalities found for each active ingredient and dose (Figure 1a, b) corresponded to the efficacy percentage, which was determined using the Abbott formula [19].
When analyzing the mortality rate for the active ingredient azadirachtin, significant model effects were found (Omnibus test: Chi-square = 3,858. 13, df = 5; P < 0.01) with significant effects for the dose variable (Chi-square = 1,944.32; df = 3; P < 0.01) and the day covariate (Chi-square = 3,244.65, df = 1; P < 0.01). For boric acid as the active ingredient, model significance was high (Omnibus test: Chi-square = 7,628.36, df = 6; P < 0.01), with effects for the dose variable (Chi-square = 6,166.67; df = 2; P < 0.01), and the day covariate (Chi-square = 6,563.25, df = 1; P < 0.01).
For azadirachtin, the highest larval mortality (95%) was only found at the highest dose (50.0 ppm) from day 20 of the trial. Lower azadirachtin doses were less effective on RPW (Figure 1a). In contrast, the efficacy achieved with the boric acid was more rapid and more efficient. The mortalities achieved with the two highest doses evaluated (2,500 and 5,000 ppm) were 100% from day 12 of the start of the trial (Figure 1b).
3. Discussion
The mortality results obtained for azadirachtin (Figure 1a) were better than others reported in earlier works [9,20]. The reason for this difference might be attributable to the use of different commercial products in the two assays. At the larval level, azadirachtin is a chemical that exhibits antifeeding effects that impact the insects through chemoreception (the primary effect) and by reducing their food consumption (the secondary effect). Furthermore, it affects the synthesis of vital proteins in the insect larvae’s digestive system – the more concentrated the azadirachtin dose, the greater the malfunction and mortality of the larvae [21,22]. Commercial compounds containing this active ingredient are used to control various crop pest species, including Coleoptera, Hemiptera, Diptera, Orthoptera, and Isoptera [23]. A variety of insecticides have been the subject of experimental laboratory and field trials worldwide seeking to control R. ferrugineus; of these, azadirachtin appears to show real potential for controlling this pest species [24] and other species of the genus Rhynchophorus [25] when used as the active ingredient.
In contrast, the active ingredient boric acid proved most efficacious at the highest doses used, after 12 days (Figure 1b). This is the first time, according to the existing literature, that this compound’s toxicity has been evaluated for R. ferrugineus larvae.
Boric acid has been considered an insecticide since 1948, when it began to be used to control ants, cockroaches, silverfish, wood destroying insects (including termites), and fleas [15,26,27,28,29]. As an insecticide, it is employed in dry-powder form, to which a food attractant is added. It is then sprinkled into cracks and crevices in a thin powder layer [26]. On contact, the boric acid adheres to the insects’ legs as they move through the powder. Later, the insects ingest the poison when cleaning themselves; this leads to death by starvation and dehydration after 3-10 days. However, boric acid’s mode of action on insects has not been satisfactorily explored [15].
Initial studies on the cockroach species Blattella germanica (Blattodea: Ectobiidae) reported that the insects eventually died of starvation. Through macroscopic examination of the insects’ viscera on days 1 and 2 following treatment, it was revealed that their intestines were empty and slightly enlarged. By days 3 and 4, the intestines of all the treated insects examined were empty, enlarged and filled with gas bubbles. The average food consumption during this period was less than 1 mg/insect and a 50% mortality rate was achieved after 3.8 days [30].
Habes et al. [31] investigated the same species. They concluded that boric acid exhibited insecticidal activity. According to their histological studies, they postulated that death may be due to alteration of the midgut structure. They also proved that this compound exerted a neurotoxic action on this species.
Likewise, in the ant species Linepithema humile (Hym.: Formicidae), electron-microscopic studies revealed ants that were fed on low concentrations of boric acid (0.5%) presented gross abnormalities in the microvilli and cells lining the midgut [32].
More recently, in the larvae, pupae and adults of Aedes aegypti (Dip.: Culicidae), boric acid has been reported to produce malformations as well as mortality [33].
From another perspective, some organisms and/or microbial structures, known as bacteriocytes, exist in specialized cells within insects; these are endosymbionts. Others are ectosymbionts - they are found on the body surface or on the surface of internal organs. Symbionts, which comprise mainly actinomycete fungi and bacteria, play a fundamental role in insect nutrition, as they allow many species to develop normally even though their food has limited nutritional value. Many symbiotic relationships are casual, especially for ectosymbionts, which often make up rich gut microbiota [34].
All insects (including insect pests) have symbiotic bacteria inside their body, particularly those insects that feed on restricted diets such as plant sap, vertebrate blood, or woody material. The symbionts play a prominent role in insect ecology; they aid in food digestion or provide nutrients, influence insect–plant interactions, host population, heat tolerance, and pesticide detoxification, as well as protect against natural enemies [35].
The syntrophic relationship within ectosymbiosis provides an important ecological evolutionary advantage (e.g., [35,36,37]). However, knowledge regarding several aspects of these associations is yet to be fully unraveled [35].
It has already been documented that palm weevils (including R. ferrugineus) host a diverse array of protozoan, fungal, viral, and bacterial species [38,39]. This is due to them having to feed on material, such as palm stipe, that has limited nutritional value (as mentioned before), thus the ectosymbionts provide them with a much-needed microbiota.
The intestinal microbiota of R. ferrugineus is composed of facultative and obligate anaerobic bacteria that metabolize through fermentation. It is assumed that these bacteria manage the fermentation of the palm tissue in the tunnels where the larvae develop and might play a key role in providing nutrition to the insect [40,41]. The contribution of microbial ectosymbionts to the physiology, reproduction, and detoxification of secondary plant metabolites in palm weevils, and in R. ferrugineus, is not yet fully understood [39]. Nevertheless, we do know how important this gut microbiota is in R. ferrugineus development [42,43].
Boric acid possesses insecticidal properties (as shown above). It is also an effective biocide, exhibiting bactericidal, fungicidal, and virucidal properties [44,45]. Furthermore, boric acid is approved for use in several countries to protect wooden structures, timber, and related products (e.g., [26,46,47]) whereas in other areas, such as in the E.U., an exclusion criterion applies that is subject to derogation [48]. Indeed, efforts are underway for boric acid to be re-evaluated and reapproved.
The results presented into the mortality-effectiveness of boric acid (Figure 1b) could be due to the compound’s direct toxic effect on the larvae; this cannot be ruled out. On the other hand, it may also be because the larvae starve as a result of an alteration in the midgut, as has been demonstrated in other insect species (see above) along with the elimination of their intestinal microbiota. In this regard, boric acid has been shown to induce dysbiosis in termite and cockroach species. Apart from a general decrease in microbial diversity, the relative abundance of some symbionts essential for nutrition in these insects decreased in response to higher boric acid concentrations [49,50].
Moreover, boric acid has been shown to have a synergistic effect when entomopathogens are applied, as occurs in other insect species [51,52,53,54,55].
The findings of this study are very promising and important for increasing our knowledge into the effects of boric acid (and its derivatives) as an insecticide and/or synergist in the control of this destructive species. However, we must still fully investigate any possible undesirable effects that this compound might have on the palm trees themselves. In general, the importance of boron is undeniable. It is an essential micronutrient for cultivated plants since both its deficiency and overabundance (toxicity) in the soil can negatively affect plant growth and development [56], especially for the date palm [57].
5. Conclusions
-The active ingredient azadirachtin demonstrated good potential in controlling the pest species, with an efficacy of 95.0 ± 5.0 % at a dose of 50.0 ppm on day 20.
-In contrast, the active ingredient boric acid showed even greater potential, showing 100 % efficacies at doses of 2,500 and 5,000 ppm at day 12.
Author Contributions
Conceptualization, T.C.; methodology, T.C.; software, M.G.; validation, T.C. and M.G.; formal analysis, M.G.; investigation, T.C.; resources, T.C.; data curation, T.C. and M.G.; writing—original draft preparation, T.C.; writing—review and editing, M.G.; visualization, T.C. and M.G.; supervision, T.C.; project administration, T.C.; funding acquisition, T.C. All authors have read and agreed to the published version of the manuscript
Funding
This research was funded by National Institute of Agrarian and Food Research and Technology (INIA), Spain, grant number SC97-117-C2.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors on request.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- EPPO. Rhynchophorus ferrugienus. EPPO Global Database. European and Mediterranean Plant Protection Organization. Available online: https://gd.eppo.int/taxon/RHYCFE/distribution (accessed on 23 September 2024).
- Resolucion, no. Resolucion no. 1.079/022 DGSA aprueba plan de contingencia para Rhynchophorus ferrugineus. Ministerio de Ganadería, Agricultura y Pesca, Gobierno de Uruguay. 2022, p. 2. Available online: https://www.gub.uy/ministerio-ganaderia-agricultura-pesca/institucional/normativa/resolucion-n-1079022-dgsa-aprueba-plan-contingencia-para-rhynchophorus (accessed on 23 September 2024).
- Martin, M.M.; Cabello, T. Biologia y ecologia del curculionido rojo de la palmera, Rhynchophorus ferrugineus (Olivier, 1790) (Col.: Dryophthoridae); University of Almeria: Almeria, Spain, 2005; pp. 1–202. [Google Scholar]
- Cabello, T. Population biology and dynamics of the red palm weevil. In Rhynchophorus ferrugineus (Col.: Dryophothoridae) in Spain. First International Conference on the Red Palm Weevil; Valencia Fundacion, Agroalimed, Ed.; Generalitat Valenciana, Conselleria de Agricultura y Medio Ambiente: Valencia, Spain, 2006; pp. 19–34. [Google Scholar]
- Dembilio, O.; Jaques, J.A. Biology and management of Red Palm Weevil. In Sustainable pest management in date palm: current status and emerging challenges; Waqas, W., Faleiro, J.R., Miller, T.A., Eds.; Springer International: Cham, Switzerland, 2015; pp. 13–36. [Google Scholar] [CrossRef]
- Rochat, D.; Dembilio, O; Jaques, J.A.; Suma, P.; Pergola, A.; Hamidi, R.; Kontodimas, D.; Soroker, V. Rhynchophorus ferrugineus: taxonomy, distribution, biology, and life cycle. In: Handbook of major palm pests; Audsley, N., Soroker, V., Colazza, S., Eds.; John Willey & Sons Ltd.: Oxford, UK, 2017; pp. 69–104. [Google Scholar] [CrossRef]
- Faleiro, J.R. A review of the issues and management of the red palm weevil Rhynchophorus ferrugineus (Col.: Rhynchophoridae) in coconut and date palm during the last one hundred years. Int. J. Trop. Insect Sc. 2006, 26, 135–154. [Google Scholar] [CrossRef]
- Cabello, T.; de la Pena, J.; Barranco, P.; Belda, J. Laboratory evaluation of imidacloprid and oxamyl against Rhynchophorus ferrugineus. Tests of Agrochemicals and Cultivar. Annals of Applied Biology 18 Supplement 130, 6–7.
- Barranco, P.; de la Peña, J.; Martin, M.M.; Cabello, T. Efficacy of chemical control of the new palm pest Rhynchophorus ferrugineus (Curculionidae: Coleoptera). Bol. San. Veg. Plagas 1998, 24, 301–306. [Google Scholar]
- Solano-Rojas, Y.; Gamez, M.; Lopez, I.; Garay, J.; Varga, Z.; Cabello, T. Conservation strategy for palm groves: Optimal chemical control model for red palm weevil Rhynchophorus ferrugineus. Agronomy 2021, 11, 1–13. [Google Scholar] [CrossRef]
- Bracchetti, L.; Cocci, P.; Palermo, F.A. Multiple aspects of the fight against the red palm weevil in an urban area: study case, San Benedetto del Tronto (Central Italy). Insects 2023, 14, 502. [Google Scholar] [CrossRef]
- Hernandez, D.; Folk, F.; Sanchez, A.; Fernandez, R. Control of red palm weevil (Rhynchophorus ferrugineus) using trunk injections and foliar sprays. Bol. San. Veg. Plagas 2003, 29, 563–573. [Google Scholar]
- Dembilio, O.; Riba, J.M.; Gamon, M.; Jacas, J.A. Mobility and efficacy of abamectin and imidacloprid against Rhynchophorus ferrugineus in Phoenix canariensis by different application methods. Pest Manag. Sci. 2015, 71, 1091–1098. [Google Scholar] [CrossRef]
- FAO. Red Palm Weevil: Guidelines on management practices; Food and Agriculture Organization of the United Nations: Rome, Italy, 2020; pp. 1–86. [Google Scholar] [CrossRef]
- See, A.S; Salleh, A.B.; Bakar, F. A, Yusof, N.A.; Abdulamir, A.S.; Heng, L.Y. Risk and health effect of boric acid. Am. J. Appl. Sci. 2010, 7, 620–627. [Google Scholar] [CrossRef]
- Boone, C.; Bond, C.; Stone, D. Boric Acid General Fact Sheet; National Pesticide Information Center, Oregon State University Extension Services. Available online: http://npic.orst.edu/factsheets/boricgen.pdf] (accessed on 29 June 2024).
- Alarcon, F.J.; Martinez, T.F.; Barranco, P.; Cabello, T.; Diaz, M.; Moyano, F.J. Digestive proteases during development of larvae of red palm weevil, Rhynchophorus ferrugineus (Col.: Curculionidae). Insect Biochem. Molec. 2002, 32, 265–274. [Google Scholar] [CrossRef]
- Martin, M.M.; Cabello, T. Rearing management of red palm weevil, Rhynchophorus ferrugineus (Col.: Dryophthoridae), in artificial diet and effects on biology and adult biometry. Bol. San. Veg. Plagas 2006, 32, 631–641. [Google Scholar]
- Andujar, A.; Barranco, P.; Belda, J.E.; Cabello, T.; Carreño, R. Analisis de eficacia de productos fitosanitarios. Phytoma España 1997, 92, 32–42. [Google Scholar]
- Merghem, A.; Mohamed, A.A. Impact of neem extracts, Azadirachta indica induced against red palm weevil, Rhynchophorus ferrugineus attacking date palm orchards in Egypt. Egypt. Acad. J. Biolog. Sci. 2017, 9, 109–117. [Google Scholar] [CrossRef]
- Dhra, G.; Ahmad, M.; Kumar, J.; Patanjali, P.K. Mode of action of azadirachtin: a natural insecticide. Int. Res. J. Biol. Sci. 2018, 7, 41–46. [Google Scholar]
- Kilani-Morakchi, S.; Morakchi-Goudjil, H.; Sifi, K. Azadirachtin-based insecticide: overview, risk assessments, and future directions. Front. Agron. 2021, 3. [Google Scholar] [CrossRef]
- Morgan, E.D. Azadirachtin, a scientific goldmine. Bioorgan. Med. Chem. 2009, 17, 4096–4105. [Google Scholar] [CrossRef]
- Naveed, H.; Andoh, V.; Islam, W.; Chen, L.; Chen, K. Sustainable pest management in date palm ecosystems: unveiling the ecological dynamics of red palm weevil (Col.: Curculionidae) Infestations. Insects 2023, 14, 859. [Google Scholar] [CrossRef]
- Gabr, B.; Lemmons, J.M.; El-Bokl, M.M. Potential of neem oil extract against palmetto weevil larvae, Rhynchophorus cruentatus (Col.: Curculionidae) and its impact on some detoxification enzymes. J. Entomol. Acarol. Res. 2022, 54, 10470. [Google Scholar] [CrossRef]
- EPA. R.E.D. Fact Boric acid. United States Environmental Protection Agency, EPA-738-F-93-006. Available online: https://nepis.epa.gov/Exe/ZyPDF.cgi/2000E7FQ.PDF?Dockey=2000E7FQ.PDF] (accessed on 2 October 2024).
- Woods, W. G. An introduction to boron: history, sources, uses and chemistry. Environ. Health Perspect. 1994, 102, 5–11. [Google Scholar]
- Cox, C. Boric acid and borates. J. Pesticide Reform 2004, 24, 10–15. [Google Scholar]
- Bernard, C.E.; Harrass, M.C.; Manning, M.J. Boric acid and inorganic borate pesticides. In Hayes’ handbook of pesticide toxicology, 3rd ed.; Krieger, R., Ed.; Elsevier: Amsterdam, NL, 2010; pp. 2033–2053. [Google Scholar] [CrossRef]
- Cochran, D.G. Toxic effects of boric acid on the german cockroach. Experientia 1995, 51, 561–63. [Google Scholar] [CrossRef]
- Habes, D.; Morakchi, S.; Aribi, N.; Farine, J.P.; Soltani, N. Boric acid toxicity to the german cockroach, blattella germanica: alterations in midgut structure, and acetylcholinesterase and glutathione s-transferase activity. Pestic. Biochem. Phys. 2006, 84, 17–24. [Google Scholar] [CrossRef]
- Klotz, J.; Amrhein, C.; McDaniel, S.; Rust, M.; Reierson, D. Assimilation and toxicity of boron in the Argentine ant (Hym.: Formicidae). J. Entomol. Sci. 2002, 37, 193–99. [Google Scholar] [CrossRef]
- Sharawi, S.E. Mortality and malformation effects of boric acid against larval stage of Aedes aegypti (Culicidae: Diptera). bioRxiv 5562. [Google Scholar] [CrossRef]
- Thompson, S.N.N.; Simpson, S.J.J. Nutrition. In Encyclopedia of Insects; Resh, V.H., Carde, R.T., Eds.; Academic Press: San Diego, CA, USA, 2003; pp. 807–813. [Google Scholar]
- Kashkouli, M.; Mehrabadi, M.; Fathipour, Y. The symbionts. In Microbial approaches for insect pest management; Omkar, *!!! REPLACE !!!*, Ed.; Springer Nature: Singapore, 2021; pp. 219–269. [Google Scholar]
- Gupta, A.; Nair, S. Dynamics of insect–microbiome interaction influence host and microbial symbiont. Front. Microbiol. 2020, 11, 1357. [Google Scholar] [CrossRef]
- Drishnan, N.; Csiszar, V.; Mori, T.F.; Garay, J. Genesis of ectosymbiotic features based on commensalistic syntrophy. Sci. Rep. 2024, 14, 1366. [Google Scholar] [CrossRef]
- Lefevre, C; Charles, H.; Vallier, A.; Delobel, B.; Farrell, B.; Heddi, A. Endosymbionte phylogenesis in the Dryophthoridae weevils: evidence for bacterial replacement. Mol. Biol. Evol. 2004, 21, 965–973. [Google Scholar] [CrossRef]
- Hoddle, M.S.; Antony, B.; El-Shafie, H.A.F.; Chamorro, M.L.; Milosavljevic, I.; Lohr, B.; Faleiro, J.R. Taxonomy, biology, symbionts, omics, and management of Rhynchophorus palm weevils (Col.: Curculionidae: Dryophthorinae). Annu. Rev. Entomol. 2024, 69, 455–479. [Google Scholar] [CrossRef]
- Tagliavia, M.; Messina, E.; Manachini, B.; Cappello, S.; Quatrini, P. The gut microbiota of larvae of Rhynchophorus ferrugineus (Col.: Curculionidae). BMC Microbiol. 2014, 14, 136. [Google Scholar] [CrossRef]
- Habineza, P.; Muhammad, A.; Ji, T.; Xiao, R.; Yin, X.; Hou, Y.; Shi, Z. The promoting effect of gut microbiota on growth and development of red palm weevil, Rhynchophorus ferrugineus (Col.: Dryophthoridae) by modulating its nutritional metabolism. Front. Microbiol. 2019, 10, 1212. [Google Scholar] [CrossRef]
- Butera, G.; Ferraro, C.; Colazza, S.; Alonzo, G.; Quatrini, P. The culturable bacterial community of frass produced by larvae of Rhynchophorus ferrugineus (Col.: Curculionidae) in the Canary Island date palm. Lett. Appl. Microbiol. 2012, 54, 530–536. [Google Scholar] [CrossRef]
- Habineza, P.; Muhammad, A.; Ji, T.; Xiao, R.; Yin, X.; Hou, Y.; Shi, Z. The promoting effect of gut microbiota on growth and development of red palm weevil, Rhynchophorus ferrugineus (Col.: Dryophthoridae) by modulating its nutritional metabolism. Front. Microbiol. 2019, 10, 1212. [Google Scholar] [CrossRef]
- McDonnell, G.E. Antisepsis, disinfection, and sterilization: type, action, and resistance, 2nd ed.ASM Press: Washington, DC, USA, 2017; pp. 1–432. [Google Scholar]
- Celikezen, F.C.; Sahin, I.H. Investigation of antimicrobial effects of some boron compounds. Bitlis Eren Univ. J. Sci. 2023, 12, 591–595. [Google Scholar] [CrossRef]
- Boric acid and precursor to boric acid: environmentier II assessment. CAS Registry number: 10043-35-3, 11113-50-1, 13460-51-0, 12008-41-2, 26038-87-9, 1303-96-4. Government of Australia. Available online: https://www.industrialchemicals.gov.au/sites/default/files/Boric%20acid%20and%20precursors%20to%20boric%20acid_%20Environment%20tier%20II%20assessment.pdf (accessed on 5 October 2014).
- Boric acid. Government of Canada, Canada. Available online: https://www.canada.ca/en/health-canada/services/chemicals-product-safety/boric-acid.html (accessed on 5 October 2014).
- Boucard, P.; Denize, C. Potential for substitution of substances used in wood preservative (PT8). Ineris-207016-2757679-v1.0. Available online: https://www.ineris.fr/en/potential-substitution-substances-used-wood-preservatives-pt8-issues-related-future-approval (accessed on 5 October 2014).
- Jiang, M.; Dong, F.Y.; Pan, X.Y.; Zhang, Y.N.; Zhang, F. Boric acid was orally toxic to different instars of Blattella germanica (Blattodea: Blattellidae) and caused dysbiosis of the gut microbiota. Pestic. Biochem. Physiol. 2021, 172, 104756. [Google Scholar] [CrossRef] [PubMed]
- Ashbrook, A.R.; Schwarz, M.; Schal, C.; Mikaelyan, A. Lethal disruption of the symbiotic gut community in Eastern subterranean termite caused by boric acid. BioRxiv. 2024, 2024.06.26.600876. [Google Scholar] [CrossRef]
- Govindarajan, R.; Jayaraj, S.; Narayanan, K. Mortality of the tobacco caterpillar, Spodoptera litura, when treated with Bacillus thuringiensis combinations with boric acid and insecticides. Phytoparasitica 1976, 4, 193–1966. [Google Scholar] [CrossRef]
- Cisneros, J.; Perez, J.A.; Penagos, D.I.; Ruiz, J.V.; Goulson, D.; Caballero, P.; Cave, R.D.; Williams, T. Formulation of nucleopolyhedrovirus with boric acid for control of Spodoptera frugiperda (Lep.: Noctuidae) in maize. Biol. Control 2002, 23, 87–95. [Google Scholar] [CrossRef]
- Biswal, G.; Singh, D.; Dhal, N.K. Synergistic effect of Bacillus subtilis and boric acid on management of bacterial with disease of potato caused by Rlastonia solanacearum in coastal plans of Odisha under field condition. Indian Phytopathology 2018, 71, 431–434. [Google Scholar] [CrossRef]
- Chao, Y.; Wang, M.; Dai, W.; Dong, F.; Wang, K.; Zhang, F. (Synergism between hydramethylnon and Metarhizium anisopliae and their influence on the gut microbiome of Blattella germanica. Insects 2020, 11, 538. [Google Scholar] [CrossRef]
- Yang, R.; Zhang, M.; Schal, C.; Jiang, M.; Cai, T.; Zhang, F. Boric acid enhances Metarhizium anisopliae virulence in Blattella germanica by disrupting the gut and altering its microbial community. Biol. Control. 2021, 152, 104430. [Google Scholar] [CrossRef]
- Thakur, S.; Sinha, A.; Bag, A.G. Boron – a critical element for fruit nutrition. Commun. Soil Sci. Plan. 2023, 54, 2899–2914. [Google Scholar] [CrossRef]
- Pillay, A.E.; Williams, J.R.; El-Mardi, M.O.; Hassan, S.M.; Al-Hamdi, A. Boron and the alternate-bearing phenomenon in the date palm (Phoenix dactylifera). J. Arid Environ. 2005, 62, 199–207. [Google Scholar] [CrossRef]
Figure 1.
Efficacy percentage of the two active ingredients: (a) azadirachtin and (b) boric acid, at different doses (ppm), as a function of the number of days post application (DAT), when incorporated in an artificial diet and offered to II-instar Rhynchophorus ferrugineus larvae under laboratory conditions (25 ºC and 0:24 hours of light/dark).
Figure 1.
Efficacy percentage of the two active ingredients: (a) azadirachtin and (b) boric acid, at different doses (ppm), as a function of the number of days post application (DAT), when incorporated in an artificial diet and offered to II-instar Rhynchophorus ferrugineus larvae under laboratory conditions (25 ºC and 0:24 hours of light/dark).
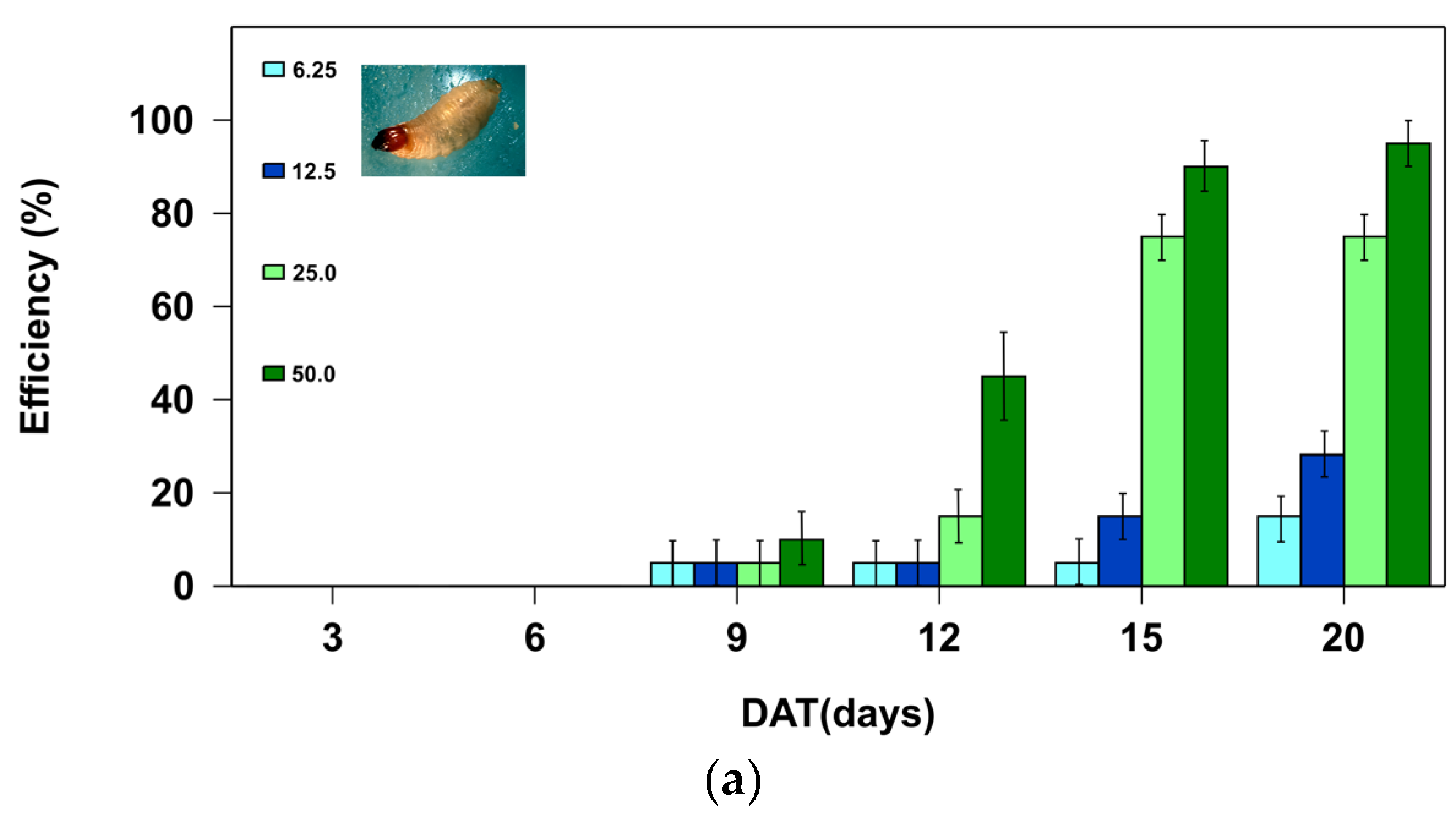
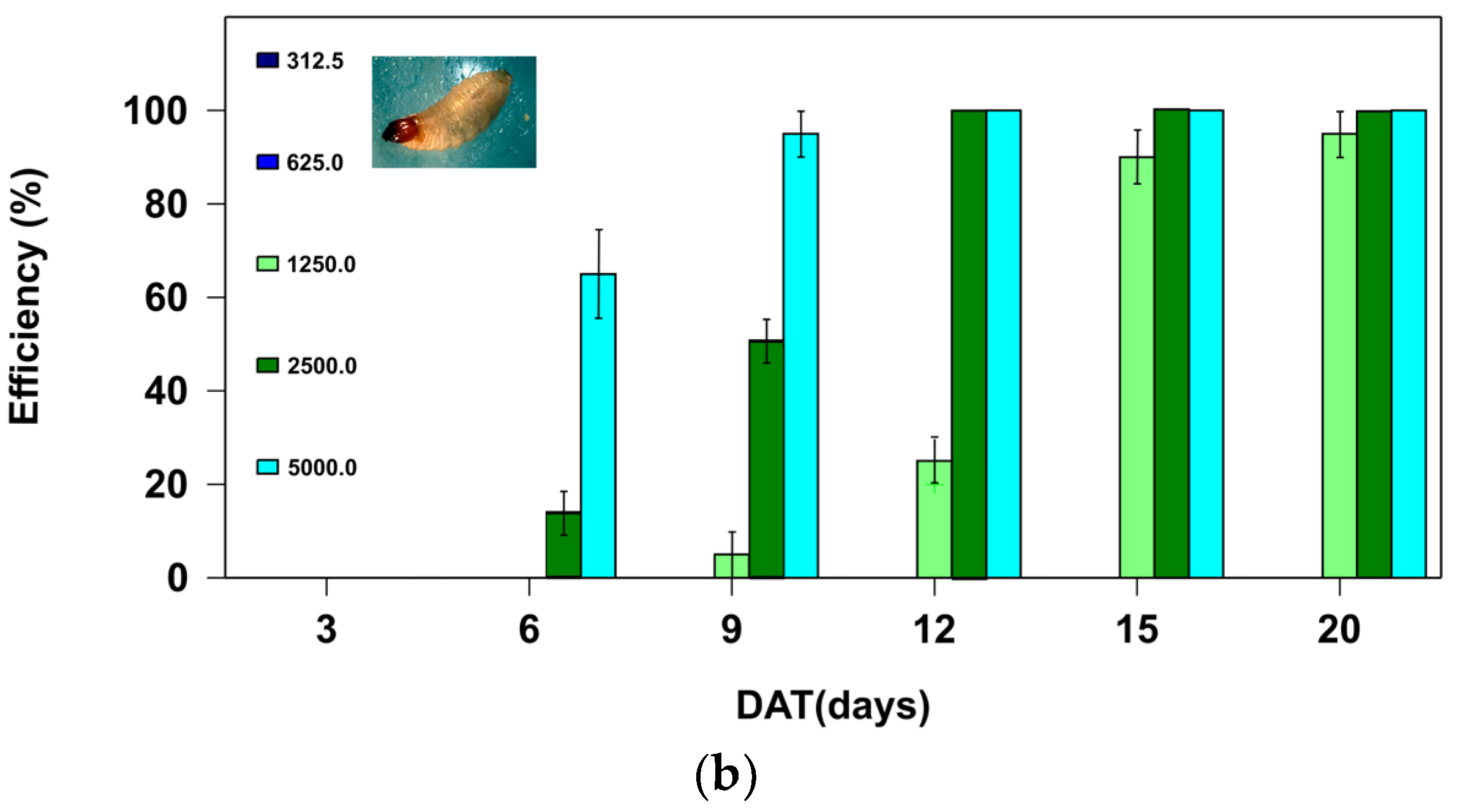

Table 1.
Average number (±SE) of live Rhynchophorus ferrugineus larvae (II-instar) as a function of time, when fed an artificial diet with two active ingredients at different doses (ppm) under laboratory conditions (25 ºC and 0:24 hours of Light/Dark).
Table 1.
Average number (±SE) of live Rhynchophorus ferrugineus larvae (II-instar) as a function of time, when fed an artificial diet with two active ingredients at different doses (ppm) under laboratory conditions (25 ºC and 0:24 hours of Light/Dark).
| Active Ingredient | Dose (ppm) | Day after treatments (DAT) | |||||
|---|---|---|---|---|---|---|---|
| 3 | 6 | 9 | 12 | 15 | 20 | ||
| Azadirachtin | 0 (control) |
5.0 (0.0) |
5.0 (0.0) |
5.0 (0.0) |
5.0 (0.0) |
5.0 (0.0) |
5.0 (0.0) |
| 6.3 | 5.0 (0.0) |
5.0 (0.0) |
4.8 (0.3) |
4.8 (0.3) |
4.8 (0.3) |
4.3 (0.3) |
|
| 12.5 | 5.0 (0.0) |
5.0 (0.0) |
4.8 (0.3) |
4.8 (0.3) |
4.8 (0.3) |
4.3 (0.3) |
|
| 25.0 | 5.0 (0.0) |
5.0 (0.0) |
4.8 (0.3) |
4.8 (0.3) |
4.3 (0.3) |
4.0 (0.0) |
|
| 50.0 | 5.0 (0.0) |
5.0 (0.0) |
4.5 (0.3) |
2.8 (0.5) |
0.5 (0.3) |
0.3 (0.3) |
|
| Boric Acid | 0 (control) |
5.0 (0.0) |
5.0 (0.0) |
5.0 (0.0) |
5.0 (0.0) |
5.0 (0.0) |
5.0 (0.0) |
| 312.5 | 5.0 (0.0) |
5.0 (0.0) |
5.0 (0.0) |
5.0 (0.0) |
5.0 (0.0) |
5.0 (0.0) |
|
| 625.0 | 5.0 (0.0) |
5.0 (0.0) |
5.0 (0.0) |
5.0 (0.0) |
5.0 (0.0) |
5.0 (0.0) |
|
| 1250.0 | 5.0 (0.0) |
5.0 (0.0) |
4.8 (0.3) |
3.8 (0.3) |
0.5 (0.3) |
0.3 (0.3) |
|
| 2500.0 | 5.0 (0.0) |
4.3 (0.3) |
2.3 (0.5) |
0 | 0 | 0 | |
| 5000.0 | 5.0 (0.0) |
1.8 (0.5) |
0.3 (0.3) |
0 | 0 | 0 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



