
Submitted:
21 October 2024
Posted:
22 October 2024
You are already at the latest version
Abstract
Islatravir (ISL) is the first-in-class nucleoside reverse transcriptase translocation inhibitor (NRTtI) with novel modes of action. Data on ISL resistance is currently limited, particularly to HIV-1 non-B subtypes. This study aimed to assess prevalent nucleos(t)ide reverse transcriptase inhibitor (NRTI) resistance mutations in HIV-1 subtype C for phenotypic resistance to ISL. Prevalent single and combinations of NRTI resistance mutations were selected from a routine HIV-1 genotypic drug resistance testing database and introduced into HIV-1 subtype C-like pseudoviruses, which were then tested for ISL susceptibility. Single NRTI resistance mutations were susceptible or showed only a low level of resistance to ISL. This included thymidine analogue mutations (TAMs, i.e., M41L, D67N, K70R, T215FY, K219EQ) and non-TAMs (i.e., A62V, K65R, K70ET, L74IV, A114S, Y115F, M184V). Combinations of M184V with one or more additional NRTI resistance mutations generally displayed reduced ISL susceptibilities. This was more prominent for combinations that included M184V+TAMs, and particularly M184V+TAM-2 mutations. Combinations that included M184V+K65R did not impact significantly on ISL susceptibility. Our study suggests that ISL would be effective in treating people living with HIV (PLWH) failing tenofovir disoproxil fumarate (TDF)/lamivudine (3TC) or TDF/emtricitabine (FTC)-containing regimens but would be less effective in PLH failing zidovudine (AZT) with 3TC or FTC-containing regimens.
Keywords:
HIV
; antiretroviral therapy (ART)
; antiviral drugs
; NRTI
; islatravir (ISL)
; phenotypic
; drug resistance
; subtype C
1. Introduction
The human immunodeficiency virus (HIV) continues to be a significant threat to human health globally. HIV has infected an estimated 85.6 million individuals worldwide and the acquired immunodeficiency syndrome (AIDS) has claimed the lives of approximately 40.4 million people [1]. However, substantial advances have been made in the treatment of people living with HIV (PLWH). Combination antiretroviral therapy (cART) has proven to be effective in managing HIV infections to allow for near-normal quality and span of life in PLWH [2,3]. Antiretroviral drugs (ARVs) cannot cure an HIV infection and cART is required for life to ensure sustained viral suppression. However, optimal cART adherence can be difficult to maintain in the long-term [4], with sub-optimal adherence driving the development of antiretroviral drug resistance [5]. Alternative treatment options and novel ARVs with efficacy against resistant viral variants are required that could address these shortfalls.
Islatravir (ISL) is a novel nucleoside reverse transcriptase translocation inhibitor (NRTtI) with a unique dual mechanism of action. Apart from acting as an atypical chain terminator, it also prevents HIV Reverse Transcriptase from translocating further on the viral RNA template [6]. ISL was shown to be well tolerated and effective in several clinical trials investigating dual-drug therapies [7]. Due to its long half-life, ISL is being investigated for use in extended dosing intervals, such as weekly dosing in combination with Lenacapavir [8]. However, dose-dependent decreases in CD4 cell and total lymphocyte counts in ISL-containing regimens in some of these clinical trials [9,10] set the stage for the re-evaluation of ISL at lower doses. ISL has a high genetic barrier to the development of resistance and has shown to be effective against variants with major single nucleos(t)ide reverse transcriptase inhibitor (NRTI) drug resistance mutations (i.e., M41L, K65R, L74I, V90I) [11,12].
Since ISL is still in an early stage of clinical development, most data available on ISL drug resistance relies largely on observations in HIV-1 subtype B. However, the high genetic diversity among HIV-1 subtypes could impact on drug resistance selection pathways, and subsequently, the degree of ARV resistance imposed by resistance mutations [13]. HIV-1 subtype C predominates globally [14], and is the predominant circulating subtype among South Africa’s 7.7 million PLWH [15]. Currently, all recommended cART regimens in South Africa include at least two NRTIs, some of which (i.e., lamivudine) are often recycled between regimens [16]. To inform HIV clinicians and caregivers on the potential role of ISL in cART in the context of HIV-1 subtype C NRTI-experienced PLWH, this study evaluated the in vitro susceptibility of prevalent NRTI drug resistance mutations to ISL.
2. Materials and Methods
2.1. Antiretroviral Drugs
Islatravir was purchased from MedChemExpress (Ipswich, USA; Cat. No. HY-104012). The following reagent was obtained through BEI Resources, NIAID, NIH: Lamivudine (3TC), HRP-8146, contributed by the NIH HIV Reagent Program. ISL and 3TC were prepared in dimethyl sulfoxide (DMSO; Sigma-Aldrich, St. Louis, Missouri, United States; Cat. No. 472301) as 10 mM stocks and diluted into Dulbecco’s Modified Eagle Medium (DMEM; Thermo Fisher Scientific, Waltham, USA; Cat. No. 11995065) to a working solution of 0.5 μM and 100 μM, respectively.
2.2. Vectors
The HIV-1 gag-pol expression vectors (i.e., p8.9MJ4, p8.9NSX+) were obtained from Didier Trono (École Polytechnique Fédérale de Lausanne, Lausanne, Switzerland) and Deenan Pillay (University College London, London, United Kingdom). The vectors have been modified by introducing a PvuI endonuclease restriction site at the start (nucleotides 11 – 16) of reverse transcriptase, and an HpaI endonuclease restriction site toward the end (nucleotides 1,100 – 1,106) of reverse transcriptase. The firefly luciferase transfer vector (i.e., pCSFLW) was obtained from Nigel Temperton (Medway School of Pharmacy, Gillingham, United Kingdom). The following reagent was obtained through the NIH HIV Reagent Program, Division of AIDS, NIAID, NIH: Plasmid pHEF Expressing Vesicular Stomatitis Virus (VSV-G), ARP-4693, contributed by Dr. Lung-Ji Chang.
2.3. Laboratory-Adapted Strains
The following lab-adapted HIV-1 strains were obtained from the National Institute for Communicable Diseases (NICD; Modderfontein, Johannesburg, South Africa) as frozen culture supernatants: HIV-1 subtype C – Du151, Du179, Du422, CM9; HIV-1 subtype B – SM1, SM2, DS9, LTNP5 (Supplementary Table S1).
2.4. HEK293T Cell Culture
The HEK293T cell line was maintained in complete DMEM (Thermo Fisher Scientific, Waltham, USA; Cat. No. 11995065), containing 10% foetal bovine serum (Merck, Darmstadt, Germany; Cat. No. F0679), 25 mM D-glucose, 4 mM L-glutamine, 1 mM sodium pyruvate and 0.25 mg/ml gentamicin (Thermo Fisher Scientific, Waltham, USA; Cat. No. 15750060). The cells were cultured at 37oC under 5% CO2 in a humidified incubator and passaged every two to three days.
2.5. Selection of NRTI Drug Resistance Mutations
HIV-1 reverse transcriptase sequences (n = 7,749) from routine genotypic drug resistance testing, performed between January 2016 and December 2020, were obtained from the National Health Laboratory Services (NHLS; Charlotte Maxeke Johannesburg Academic Hospital, Johannesburg, South Africa). The twenty most prevalent NRTI drug resistance mutation combinations were identified and assessed for in vitro ISL susceptibility.
2.6. Preparation of Mutant Pseudoviruses in Reference Isolates
HIV-1-like replication defective pseudoviruses (PSVs) with the most prevalent combinations of NRTI mutations (n = 20) were prepared. In addition, PSVs containing the single mutations that constituted the combinations, as well as the A114S and A114S+M184V PSVs, were also generated. NRTI drug resistant mutations were introduced into the HIV-1 subtype C gag-pol expression vector (i.e., p8.9MJ4) through polymerase chain reaction (PCR)-based site-directed mutagenesis (SDM) using the Q5 Site-Directed Mutagenesis Kit (New England Biosciences, USA; Cat. No. E0554), according to the manufacturer’s instructions. The mutagenesis primers were designed using NEBaseChanger™ v.1.3.3. (https://nebasechangerv1.neb.com/) (Supplementary Table S3). Multiple rounds of SDM were performed to incorporate more than one mutation into a single vector.
Chemically competent DH5α bacterial cells (New England Biosciences, USA; Cat. No. C2987H) were transformed with the SDM reactions through standard heat-shock. The transformed bacteria were plated on carbenicillin-containing (100 μg/ml; Thermo Fisher Scientific, Waltham, USA; Cat. No. 10177012) Luria Bertani (LB; Thermo Fisher Scientific, Waltham, USA; Cat. No. 22700025) agar plates and incubated for approximately 16 hours at 37oC. Carbenicillin-containing (100 μg/ml) LB broth (Thermo Fisher Scientific, Waltham, USA; Cat. No. 12780052) was inoculated with bacterial colonies and incubated for approximately 16 hours at 37oC with agitation. Bacterial cells were pelleted through centrifugation at 4,500 rpm for 10 minutes at room temperature. The bacterial pellets were used to extract plasmid DNA with the QIAprep Spin Miniprep Kit (QIAGEN, Hilden, Germany; Cat. No. 27106). The eluted plasmid DNA was quantified on a NanoDrop 1000 Spectrophotometer (Thermo Fisher Scientific, Waltham, USA). The presence of NRTI drug resistance mutations in the vectors were confirmed with Sanger sequencing using the BigDye Terminator v3.1 Cycle Sequencing Kit (Thermo Fisher Scientific, Waltham, USA; Cat. No. 4337455) and ABI PRISM 3100 Genetic Analyzer (Thermo Fisher Scientific, Waltham, USA). Sequencing chromatograms were analysed using Geneious (Dotmatics, Boston, USA) and the consensus sequences submitted to the Stanford HIV Drug Resistance Database [17] to confirm the presence of the target mutation(s).
For unsuccessful SDM reactions, an alternative approach, described by Yang et al., was followed using two sets of overlapping primers [18]. For each mutation, two reactions were performed. For the first reactions, the forward mutagenesis primer and AmpR1 reverse primer were used, while the reverse mutagenesis primer and AmpF1 forward primer were used (Supplementary Table 3). The reactions were constructed as follows: 12.5 μl Q5 Hot Start High Fidelity 2x Master Mix (New England Biosciences, USA; Cat. No. M0494S), 1.25 μl forward primer (10 μM), 1.25 μl reverse primer (10 μM), 1.0 μl plasmid DNA (25 ng/μl) and 9.0 μl nuclease-free water. Thermocycling commenced as follows: initial denaturation at 98oC for 30 seconds; 25 cycles of denaturation at 98oC for 10 seconds, annealing at 50oC – 72oC (primer specific) for 30 seconds, elongation at 72oC for 10 minutes (50 sec/kb); final extension at 72oC for 2 minutes; hold at 4oC. The plasmid template DNA was digested by the addition of 0.2 μl DpnI (20 U/μl; Thermo Fisher Scientific, Waltham, USA; Cat. No. ER1702) to each reaction and incubation at 37 for 2oC hours. DH5α bacterial cells were transformed with 2.5 μl of each of the two reactions and the subsequent steps were followed as described previously. Selected mutations were also introduced into PSVs that contained a section of the reverse transcriptase from HIV-1 subtype B and C laboratory-adapted strains following the above SDM procedures.
To produce PSVs, HEK293T cells were plated in 10 ml of complete DMEM at 8 x 106 cells/ml in 10 cm Nunclon™ Delta transfection dishes (Thermo Fisher Scientific, Waltham, USA; Cat. No. 150318) and incubated overnight at 37oC under 5% CO2 in a humidified incubator. The transfection mixtures were prepared to contain the following: 1.00 μg HIV-1 gag-pol expression vector (mutant or wild-type), 0.25 μg pHEF-VSVG, 1.5 μg pCSFLW, 50 μl DMEM without additives, and 8.25 μl PEI “max” (1 mg/ml, pH 7.0; PolySciences, Warrington, USA; Cat. No. 24765-1). The transfection mixtures were incubated at room temperature for 20 minutes before adding it drop-wise to the plated cells. The plates were incubated for 48 hours at 37oC under 5% CO2 in a humidified incubator. The supernatants containing the PSVs were collected, filtered through a 0.45-micron syringe filter, aliquoted, and stored at -80oC.
PSV titration was performed by preparing eight, two-fold serial dilutions of supernatant in 50 μl of complete DMEM in Nunc™ Edge™ 96-well culture plates (Thermo Fisher Scientific, Waltham, USA; Cat. No. 167542). After the addition of HEK293T cells (2 x 104 cells/50 μl), the plates were incubated for 48 hours at 37oC under 5% CO2 in a humidified incubator. The expression of firefly luciferase, indicative of PSV infection, was assessed using the Bright-Glo™ Luciferase Substrate (Promega, Madison, USA; Cat. No. E263B). For this, 100 μl of substrate was added to the wells of the 96-well culture plate and incubated for two minutes at room temperature in the dark. The well contents were mixed and transferred to the corresponding wells of a white 96-well plate. Bioluminescence was quantified on the GloMax® Explorer Multimode Microplate Reader (Promega, Madison, USA) in relative light units (RLU). PSV dilutions that produced a bioluminescence of 1 x 106 RLU were used in subsequent in vitro phenotypic assays as a standardized PSV input.
2.7. Preparation of Mutant Pseudoviruses in Laboratory-Adapted Strains
Viral RNA was extracted from viral culture supernatants using the QIAamp Viral RNA Mini Kit (QIAGEN, Hilden, Germany; Cat. No. 52906) according to the manufacturer’s instructions. The viral RNA was reverse transcribed using the SuperScript™ III First-Strand Synthesis System for RT-PCR (Thermo Fisher Scientific, Waltham, USA; Cat. No. 18080-51) in conjunction with the Outer Reverse Primer (10 μM; 5’- GCTTGGATGCACACTAAATTTTCC-3’) according to the manufacturer’s instructions.
The first-round PCR reactions were performed using the Expand™ High Fidelity PCR System (Merck, Darmstadt, Germany; Cat. No. EHIFI-RO) according to the manufacturer’s instructions and contained the following: 5 μl PCR Buffer (10x), 5 μl Outer Forward Primer (10 μM, 5’-CCATGGCTATTTTTTGCACTGC-3’), 5 μl Outer Reverse Primer (10 μM), 1 μl PCR Grade Nucleotide Mix (10 mM of each dNTP), 0.75 μl Enzyme Mix (3.5 U/μl), 30.75 μl water, and 2.5 μl cDNA reaction. Thermocycling commenced as follows: initial denaturation at 94oC for 2 minutes; 10 cycles of denaturation at 94oC for 15 seconds, annealing at 55.5oC for 30 seconds, elongation at 72oC for 1 minute 10 seconds (45 sec/kb); 20 cycles of denaturation at 94oC for 15 seconds, annealing at 55.5oC for 30 seconds, elongation at 72oC for 1 minute 10 seconds (+5 seconds per successive cycle); final extension at 72oC for 7 minutes; hold at 4oC.
The second-round PCR reactions were performed using the Platinum™ SuperFi II Green PCR Master Mix (Thermo Fisher Scientific, Waltham, USA; Cat. No. 12369010) with primers that introduced PvuI and HpaI restriction sites at the 5’- and 3’-end of the amplicon, respectively. The reactions contained the following: 25 μl Platinum™ SuperFi II Green PCR Master Mix (2x), 2.5 μl Inner Forward Primer (10 μM; 5’-CGATCGAAACTGTACCAGTAAAATTAAAGC-3’), 2.5 μl Inner Reverse Primer (10 μM; 5’- GTTAACTGTTTTACATCATTAGTGTGGG-3’), 10 μl SuperFi II GC Enhancer (5x), 9 μl water and 1.0 μl first-round PCR reaction. Thermocycling commenced as follows: initial denaturation at 98oC for 30 seconds; 35 cycles of denaturation at 98oC for 10 seconds, annealing at 56.4oC for 30 seconds, elongation at 72oC for 1 minute 10 seconds (45 sec/kb); final extension at 72oC for 5 minutes; hold at 4oC. The PCR reactions were analysed on a 1% agarose gel in Tris-Acetate-EDTA (TAE) buffer (Thermo Fisher Scientific, Waltham, USA; Cat. No. B49) containing 0.35 μg/ml ethidium bromide (Bio-Rad, Johannesburg, South Africa; Cat. No. 1610433) to confirm the expected size of the amplicons (i.e., 1,096 bp). The second-round PCR amplicon spanned from amino acid 4 (HXB2 nucleotide number 2,560) to amino acid 368 (HXB2 nucleotide number 3,656) in HIV-1 Reverse Transcriptase. The amplicons were gel-purified with the Zymoclean™ Gel DNA Recovery Kit (Zymo Research, Orange, USA; Cat. No. D4001), the eluted DNA quantified on a NanoDrop 1000 Spectrophotometer (Thermo Fisher Scientific, Waltham, USA), and sequenced. The absence of ARV drug resistance mutations in the wild-type strains was confirmed with Sanger sequencing and submission to the Stanford HIV Drug Resistance Database [17].
The purified amplicons were cloned into the PCR-XL-2-TOPO vector using the TOPO™ XL-2 Complete PCR Cloning Kit (Thermo Fisher Scientific, Waltham, USA; Cat. No. K8050-10) according to the manufacturer’s instructions. Bacterial transformation, culture and plasmid DNA extraction were performed as described previously. The plasmid DNA from the resulting clones were sequenced as described previously to confirm the identity of their inserts. After sequence confirmation, the TOPO clones were subjected to double digests: 0.5 μl PvuI (20 U/μl; New England Biosciences, USA; Cat. No. R3150S), 0.5 μl HpaI (5 U/ μl; New England Biosciences, USA; Cat. No. R0105S), 1 μl T4 polynucleotide kinase (10 U/μl; New England Biosciences, USA; Cat. No. M0201S), 7 μl rCutSmart™ Buffer (10x), 1 μg plasmid DNA, and nuclease-free water to 70 μl final volume. Reactions were incubated at 37oC for 1 hour. The HIV-1 gag-pol expression vectors were also subjected to double-digests, similar to the TOPO clones, but the reactions contained 1.0 μl recombinant Shrimp Alkaline Phosphatase (1 U/μl; New England Biosciences, USA; Cat. No. M0371S) instead of T4 polynucleotide kinase. Reactions were incubated at 37oC for 1.5 hours, followed by 65oC for 5 minutes to deactivate the phosphatase.
The inserts from the TOPO clones were then sub-cloned into the HIV-1 gag-pol expression vectors (i.e., p8.9NSX+ for subtype B strains, p8.9MJ4 for subtype C strains) using the Quick Ligation™ Kit (New England Biosciences, USA; Cat. No. M2200S). The reactions contained the following: 10 μl Quick Ligase Reaction Buffer (2x), 17 ng insert, 183.5 ng vector backbone, 1 μl Quick Ligase Enzyme, and nuclease-free water to 20 μl final volume. Reactions were incubated at room temperature for 5 minutes and 5 μl of each reaction was used for the transformation of DH5α bacterial cells as described previously. The plasmid DNA from the resulting clones were sequenced to confirm the identity of their inserts. Consensus sequences were submitted to the Stanford HIV Drug Resistance Database [17] to confirm the absence of ARV drug resistance mutations. NRTI drug resistance mutations of interest were introduced into the clones by SDM (Supplementary Table S2), and PSVs were produced and tittered, as previously described.
2.8. In Vitro Phenotypic ISL Susceptibility Testing
An in vitro single-cycle phenotypic assay was used to assess the susceptibility of the PSVs to ISL [19,20,21], with some modification. Briefly, eleven, duplicate, 3-fold serial dilutions of ISL in 50 μl complete DMEM were prepared in the wells of Nunc™ Edge™ 96-well culture plates (Thermo Fisher Scientific, Waltham, USA; Cat. No. 167542) with concentrations ranging from 500 nM to 0.008 nM. Two wells received complete DMEM only and functioned as the no-drug control. 3TC was included as a positive inhibition control over concentrations ranging from 100 μM to 0.002 μM. HEK293T cells were prepared in complete DMEM at 4 x 105 cells/ml, PSVs were added to the cells at half the standardised dilution, and 50 μl of the cell/virus mixture was added to the wells of the plates. The plates were incubated for 48 hours at 37oC under 5% CO2 in a humidified incubator, after which, the expression of firefly luciferase in the wells of the plates were assessed and quantified as previously described. All PSVs were assessed in three or more independent assays.
The percent viral activities were calculated for each PSV over the ISL concentration range that was tested, relative to the no-drug control. The inhibitory concentration-50s (IC50) were calculated for each PSV using the FORECAST function in Microsoft Excel (Redmond, USA). The degree of ISL susceptibility was expressed as fold-change (FC) in IC50, relative to the wild-type PSV IC50. The assay’s FC technical cut-off (TCO) for ISL was determined at the 99th percentile of the wild-type PSV IC50 values, determined over multiple repeat assays. The degrees of ISL resistance were set at increasing increments of the TCO value.
2.9. Statistics
FC values of PSVs with single or combination mutations were compared to that of the wildtype PSV using parametric, unpaired, two-tailed t-tests. Differences in IC50 values between mutant subtype B and C PSV were compared using a one-way analysis of variance (ANOVA).
3. Results
3.1. Sequence database
3.1.1. Regimens
Most sequences were obtained from patients failing a ritonavir-boosted protease inhibitor-based (PI/r) second-line regimen (70.89%, n = 5,493/7,749), while 12.67% (n = 982/7,748) and 0.84% (n = 65/7,7498) were obtained from patients failing a non-nucleoside reverse transcriptase inhibitor (NNRTI) or integrase strand-transfer inhibitor (INSTI)-based regimens, respectively. For 0.74% (n = 57/7,748) of sequences, the information on the ARVs in the regimens were incomplete, while for 14.87% (n = 1,152/7,748) of sequences, no treatment information was provided. Details of current and prior treatment are shown in Supplementary Table S3.
3.1.2. NRTI resistance mutations
Among the 7,749 sequences, NRTI drug resistance mutations were observed in 71.2% (n = 5521) of the sequences. The 20 most prevalent combinations (Supplementary Table S4) and individual single NRTI drug resistance mutations that constitute the combinations (Supplementary Table S5) were identified. The M184V mutation was the most prevalent mutation, both as a single mutation and in combination with other NRTI drug resistance mutations. The M184V mutation was observed in each of the 20 most prevalent mutation combinations. The thymidine analogue mutations (TAMs; M41L, D67N, K70R, T215FY, K219QE) were more prevalent in combination with other mutations than in isolation. This was also observed for the non-TAM mutations (i.e., K70ET, L74IV, Y115F), and for K65R. Sequences containing TAM-2 pathway mutation combinations (i.e., D67N/K70R/T215F/K219QE) were more prevalent (4.0%, n = 244/5,521) than sequences containing TAM-1 pathway mutation combinations (i.e., M41L/L210W/T215Y) (1.9%, n = 109/5,521). However, all TAM-1 pathway mutation combinations lacked the L210W mutation, and only the M41L+T215F or M41L+T215Y combinations were observed. We also observed the TAM-2 pathway mutation combination where only D67N+K07R was present. The remaining TAM-2 pathway mutation combinations contained at least three TAM-2 mutations. The A114S mutation, shown to reduce ISL susceptibility, either in isolation or in combination with the M184V mutation [12], was not observed in any of the sequences.
3.2. In Vitro Phenotypic ISL Susceptibility Testing
3.2.1. TCO for ISL and Assay-Defined Classifications
To establish a meaningful threshold for the interpretation of FC values of the HIV-1 subtype C mutant PSVs, a TCO was derived from the ISL IC50 values obtained from multiple independent in vitro phenotypic assays (n = 41) of the wild-type HIV-1 subtype C p8.9MJ4 PSV against ISL. The TCO was established at the 99th percentile (18.26 nM) of the average IC50 (8.3 ± 5.0 nM S.D.), which translated to a TCO of 2.2 FC (Figure 1A, 1B). Thus, PSVs with a FC > 2.2 were considered to have a reduced susceptibility to ISL. Classifications of the degrees of reduced susceptibility or resistance were set at increments of the TCO as follows: FC < 2.2 = Susceptible, 2.2 ≤ FC ˂ 4.4 = Potential Low-
Level Resistance, 4.4 ≤ FC ˂ 6.6 = Low-Level Resistance, 6.6 ≤ FC ˂ 8.8 = Intermediate-Level Resistance, FC ≥ 8.8 = High-Level Resistance. Similarly, for the interpretation of the FC values of the HIV-1 subtype B mutant PSVs, a TCO was established at the 99th percentile (15.29 nM) of the average IC50 (5.9 ± 5.5 nM S.D.) of the wild-type HIV-1 subtype B p8.9NSX+, from multiple independent in vitro phenotypic assays (n = 13). The TCO was determined to be at a FC cutoff of 1.9 FC (Figure 1C, 1D). Classifications of the degrees of reduced susceptibility or resistance were set at increments of the TCO as follows: FC < 1.9 = Susceptible, 1.9 ≤ FC ˂ 3.8 = Potential Low-Level Resistance, 3.8 ≤ FC ˂ 5.7 = Low-Level Resistance, 5.7 ≤ FC ˂ 7.6 = Intermediate-Level Resistance, FC ≥ 7.6 = High-Level Resistance.
3.2.2. ISL Susceptibility of Single NRTI Drug Resistance Mutations
PSVs (n = 16) containing single NRTI resistance mutations were tested in vitro for phenotypic susceptibility to ISL. The majority of the PSVs (75%, n = 12) with single NRTI resistance mutations (i.e., M41L, K65R, D67N, K70E/R/T, L74I, A114S, Y115F, T215Y, K219E/Q) were fully susceptible to ISL (FC < 2.2; Figure 2). Three PSVs with single NRTI resistance mutations showed a potential low-level resistance to ISL: A62V – FC = 3.2 ± 1.3 FC, L74V – FC = 2.5 ± 1.5 FC, T215Y – FC = 2.9 ± 0.2 FC). The PSV containing the M184V mutation was on the cusp of potential- and low-level resistance (FC = 4.4 ± 1.1 FC). It is important to note that the classifications of ISL susceptibility for several of the PSVs with single NRTI resistance mutants (i.e., M41L, L74V, A114S, Y115F, M184V, T215Y, K219E) spanned over two or three classifications due to the variability in the assay. However, none of the mutations caused intermediate or high-level resistance to ISL.
A two-sample t-test was performed on the FC values to ascertain whether there were any statistically significant differences in ISL susceptibility between PSVs with single NRTI resistance mutants and the HIV-1 subtype C wild-type PSV (Supplementary Table S6). No statistically significant differences (p > 0.05) were observed for PSVs with the M41L, A62V, D67N, K70R, L74IV or K219EQ mutations. However, statistically significant lower FC values were observed for PSVs that contained K65R (0.4 ± 0.2 FC, p = 0.009), K70E (FC = 0.3 ± 0.1, p < 0.001), K70T (FC = 1.5 ± 0.3 FC, p = 0.033) or T215F (FC = 0.4 ± 0.03, p < 0.001) and may be indicative of hyper-susceptibility to ISL. Only four PSVs had FC values significantly higher than the wild-type PSV: A114S (FC = 2.1 ± 0.3, p = 0.031), Y115F (FC = 2.2 ± 0.5, p = 0.029), M184V (Fc = 4.4 ± 1.1, p = 0.008), and T215Y (FC = 2.9 ± 0.2, p = 0.004). However, although their changes in FC were significantly higher than the wild-type PSV, these mutations did not dramatically affect ISL susceptibility.
Published phenotypic data on ISL resistance were only available for a few of the NRTI resistance mutations (i.e., M41L, K65R, L74I/V, A114S, M184V, and T215Y) (Supplementary Table S6). However, for the PSV with the L74V single mutant, we observed a FC value of 2.5 (± 1.5), which was 12.5-fold higher than the reported value in the literature [23] (i.e., FC = 0.2). Since the PSV with this mutation also showed a high level of variation in IC50 in our in vitro assay (Figure 2, Supplementary Table S6), we generated and tested strain-derived PSVs with this mutation to ascertain whether the observations were strain and/or subtype (i.e., B or C) related. All the strain-derived mutant PSVs that contained L74V were susceptible to ISL, with ISL FC values well below their corresponding wild-type references, irrespective of subtype (Figure 3; Supplementary Table S7).
An inter-subtype analysis with a one-way ANOVA, utilizing the Brown-Forsythe and Welch parameters, comparing the IC50 values of the strain-derived mutant PSVs was performed (Figure 4A, 4B). The ANOVA specifically scrutinized the IC50 values (i.e., not FC values) since no PSV could be used as a representative (or reference) for both subtypes. The analysis revealed a significant difference (p = 0.017) in IC50 only between the subtype B DS9-L74V PSV (2.76 ± 1.27 nM) and wild-type subtype C MJ4 PSV (8.32 ± 4.99 nM) (Figure 4B). An intra-subtype analysis of subtype B strain-derived mutant PSVs with Welch’s t-test revealed that the DS9-L74V PSV was significantly more susceptible (FC = 0.35 ± 0.16, p = 0.009) to ISL than the wild-type NSX PSV (Supplementary Table S7). No significant differences were observed for the other subtype B strain-derived mutant PSVs. All subtype C strain-derived mutant PSVs had FC values lower than that of the wild-type MJ4 PSV. This difference was significant for the Du151-L74V PSV (FC = 0.58 ± 0.06, p < 0.001), Du422-L74V PSV (FC = 0.36 ± 0.26, p 0.024) and CM9-L74V PSV (FC = 0.65 ± 0.10, p = 0.005).
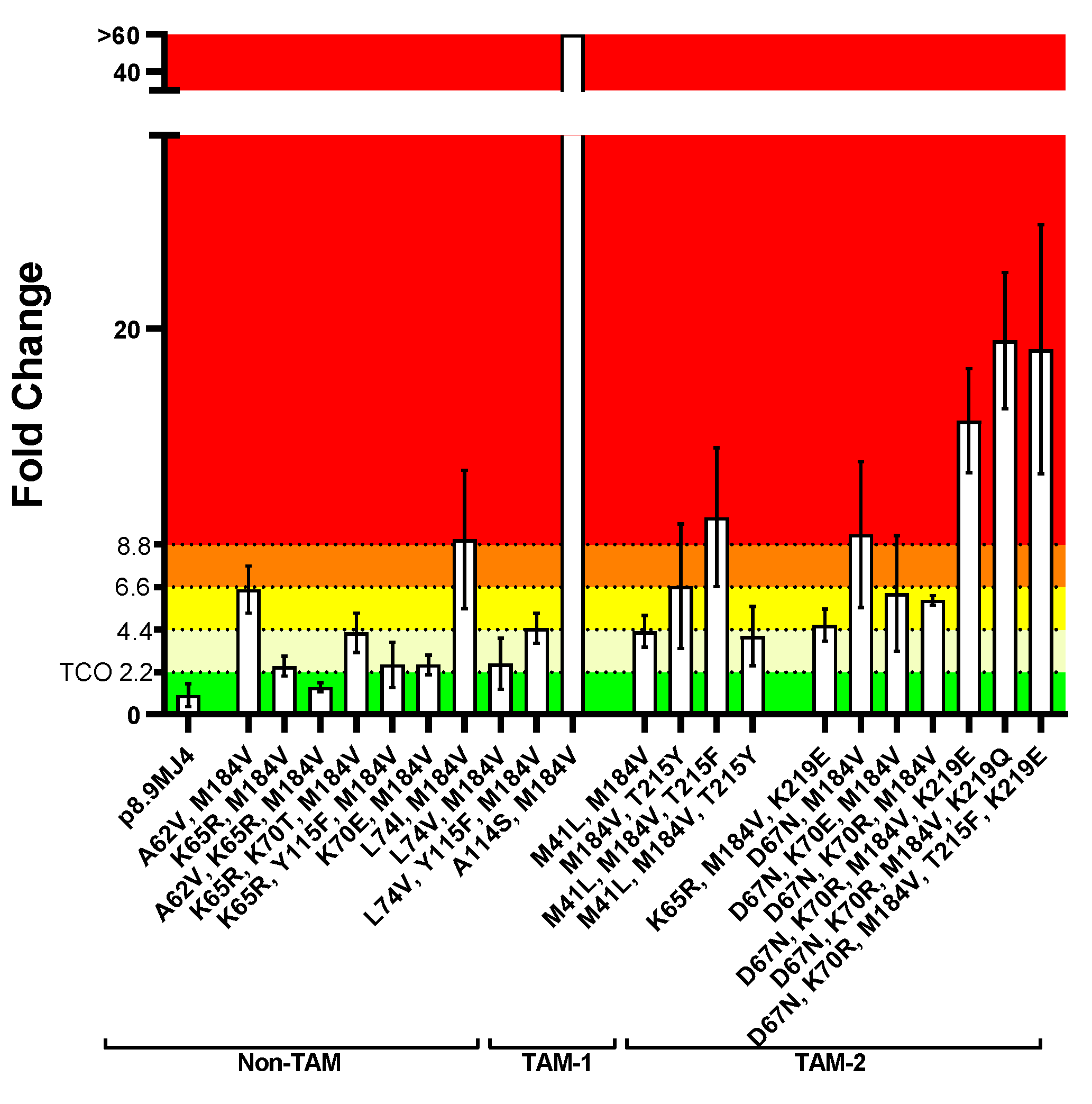
3.2.3. ISL Susceptibility of Combined NRTI Drug Resistance Mutations
Overall, PSVs with combinations of two or more NRTI resistance mutations were less susceptible to ISL (Figure 5) than those with single NRTI resistance mutations (Figure 2). Regarding PSVs containing non-TAMs only, one combination (i.e., K65R/A62V/M184V) showed full susceptibility to ISL (FC = 1.4 ± 0.2; Figure 5, Supplementary Table S8). Other PSVs containing combinations with K65R or K70E showed potential low-level resistance to ISL. The PSV with A62V/M184V was on the cusp of low- to intermediate-level ISL resistance (FC = 6.5 ± 1.0). Although PSVs with combinations containing L74V (i.e., L74V/M184V, L74V/Y115F/M184V) showed low-level ISL resistance (FC 2.6 – 4.5), the PSV with L74I/M184V showed high-level resistance (FC = 9.1 ± 2.9). This was unexpected, as the PSV with the single L74I or M814V mutations were susceptible or showed a low level of resistance (L74I: FC = 0.8 ± 0.1 FC, M184V: FC = 4.4 ± 1.1; Figure 2). The PSV with the A114S/M184V combination showed an extremely high level of resistance (FC > 60) to ISL. This was another unexpected result since the PSV with the single A114S mutation was susceptible to ISL (2.1 ± 0.3 FC), and the PSV with the single M184V mutation had a potential low level of resistance (Figure 2).
The PSVs that contained combinations with TAM-1 mutations showed a potential low- to high-level ISL resistance. The combinations of M41L/M184V (FC = 4.3 ± 0.7) and M41L/M184V/T215Y (FC = 4.1 ± 1.3) showed potential low-level resistance to ISL, while the combination of M41L/M184/T215F (FC = 10.2 ± 3.1) showed a high level of resistance. The M184V/T215Y combination showed an intermediate level of ISL resistance (FC = 6.7 ± 2.6). The PSVs that contained combinations with TAM-2 mutations generally showed higher levels of ISL resistance. Three combinations (i.e., K65R/M184V/K219E, D67N/K70E/M184V, D67N/K70R/M184V) showed low-level ISL resistance (FC 4.6 – 6.3).
Interestingly, the addition of K70E or K70R seems to sensitize the D67N/M184V combination to ISL as they were more susceptible (FC 5.9 – 6.3) than the D67N/M184V combination (FC = 9.3 ± 3.1), which showed an intermediate level of resistance. When three or more TAM-2 mutations were present, a high level of resistance to ISL was observed: D67N/ K70R/M184V/K219E (FC = 15.2 ± 2.2), D67N/ K70R/M184V/K219Q (FC = 19.4 ± 2.9), D67N/K70R/M184V/T215F/K219E (FC = 18.9 ± 5.3).
A parametric, unpaired, two-tailed t-test was performed to assess statistical significance in FC differences to the wild-type MJ4 PSV (Supplementary Table S8). The majority of PSVs with combinations of NRTI resistance mutations showed a significant difference (p-value < 0.03) in FC. No significant differences were observed for the following combinations: L74I/M184V (p = 0.060), M184V/T215Y (p = 0.094), K65R/Y115F/M184V (p = 0.144), D67N/M184V (p = 0.062), D67N/K70E/M184V (p = 0.092), and L74V/M184V (p = 0.162). Apart from the L74V/M184V and K65R/Y115F/M184V combinations, with FC values close to the TCO (i.e., FC = 2.6 ± 1.1), the remaining combinations had much higher average FC values and the lack of statistical significance is as a result of the large standard deviations observed with these combinations.
4. Discussion
Considering the integral role of NRTIs in all cART regimens, the development and exploration of novel NRTIs that are capable of effectively addressing prevailing NRTI-resistant viral variants is needed. ISL, a novel NRTtI with a unique mechanism of action, has an extended half-life in vivo and a characteristic resistance profile. Data on ISL phenotypic resistance is limited, particularly for HIV-1 non-B subtypes. To gain insight into the potential value of ISL as a treatment option in NRTI-experienced PLWH with HIV-1 subtype C, this study assessed the in vitro phenotypic drug susceptibility of HIV-1-like subtype C PSVs with common NRTI resistance mutations to ISL. Prevalent NRTI resistance mutations were identified from a routine genotypic drug resistance testing database, and PSVs containing the NRTI resistance mutations were generated and tested for susceptibility to ISL in vitro. Phenotypic data for most PSVs with single or combinations of mutations were in agreement with the limited published phenotypic data available. No single NRTI resistant mutation caused intermediate or high-level resistance to ISL. The highly prevalent M184V mutation conferred only a low level of resistance to ISL, whereas the K65R mutation was found to cause potential hyper-susceptibility to ISL. PSVs with combinations of NRTI resistance mutations, all containing the M184V mutation, were generally less susceptible to ISL than those with single NRTI resistance mutations. Combinations with the K65R, L74V, and K70E non-TAMs were either susceptible, or had a potential low-level resistance to ISL. Combinations with TAM-1, and particularly TAM-2 mutations, showed low- to high-level resistance to ISL. Both the type and number of TAMs impacted on the degree of ISL resistance.
The genotypic drug resistance testing database that was employed for this study included sequences from routine drug resistance testing performed during 2016 – 2020. The South African National Treatment Guidelines recommended cART initiation on TDF with either FTC or 3TC, and efavirenz (EFV) [24]. PLWH failing this first-line regimen were switched to a protease inhibitor (PI)-based second-line regimen containing either AZT or abacavir (ABC) with 3TC and either ritonavir(r)-boosted lopinavir (LPV/r) or atazanavir (ATV/r). In May 2019, the guidelines changed to replace EFV in the first-line regimen with dolutegravir (DTG), an INSTI [25]. The majority of sequences were obtained from PLWH managed under the 2015 guidelines, with the minority managed under to the 2019 guidelines. Routine genotypic drug resistance testing for PLWH failing cART was only recommended for those failing a PI-based second-line regimen. As such, the majority of sequences that were scrutinized in this study were from PLWH who failed a PI-based regimen that contained AZT or ABC, and 3TC as NRTIs in their current regimens, and with d4T, FTC or TDF in their previous regimens. This was reflected in their drug resistance profiles as we observed TAM-1 (i.e., M41L, T215Y) and TAM-2 (i.e., D67N, K70R, T215F, K219EQ) mutations, typically associated with sub-optimal use of AZT or d4T [26], as well as the L74IV and Y115F mutations which are typically associated with sub-optimal use of ABC [27]. The M184V mutation was most prevalent and is typically selected for with sub-optimal use of either 3TC or FTC [28,29]. The presence of this mutation among the sequences is likely as a result of sub-optimal use of 3TC/FTC (XTC) during first-line cART and sub-optimal use of 3TC during second-line cART. The presence of the K65R mutation is most likely as a consequence of sub-optimal use of TDF [30,31,32] during first-line cART, but could also be as a result of sub-optimal use of ABC during second-line cART [27]. Mutations at K70 (i.e., E/R/T) were also observed, likely as a consequence of sub-optimal use of ABC or TDF [27,32]. The A62V mutation, usually observed in combination with other NRTI resistance mutations [33], was also observed in two combinations (i.e., A62V/M184V, K65R/A62V/M184V) among the sequences.
A South African National Survey showed the M184V and K65R NRTI resistance mutations to be highly prevalent (78.2% and 57.5%, respectively) among PLWH who were failing a TDF-based regimen [34]. This highly prevalent M184V NRTI resistance mutation typically decreases XTC susceptibility by 100- to 1 000-fold in in vitro settings [28,29]. On the contrary, the M184V mutation is approximately three-fold more susceptible to AZT [35] and causes hyper-susceptibility to TDF [36]. Our data showed that this mutation conferred potential low level resistance to ISL (FC = 4.4 ± 1.1), which is in agreement with the literature where M184V showed a modest decrease in ISL susceptibility by 5-fold [37]. An in vivo animal study performed on rhesus macaques showed no viral breakthroughs when infected with an M184V-containing simian immunodeficiency virus (SIV) in the presence of ISL [38]. This implies that ISL would still be efficacious in the treatment of PLWH with M184V-containing viral variants, which is much needed due to the mutation’s high prevalence. When appearing alone, the K65R mutation has been reported cause resistance to XTC, ABC, and TDF (8.4- and 8.8-fold) [39], but increases susceptibility to AZT [35,40]. Our data showed that K65R alone was significantly more susceptible to ISL (i.e., 0.4 ± 0.2 FC) than the wild-type reference, possibly suggesting its hyper-susceptible nature. This is in agreement with a previous study performed in an HIV-1 subtype B setting [37]. Both the K65R and M184V mutations cause a deficit in viral replication [36]. An in vitro passage study showed that the single K65R mutant rapidly reverted to wild-type in the presence of ISL, whereas no reversion was noted for M184V [41]. This points towards a sustained efficacy for ISL on K65R-containing viral variants.
Separately and in combination, the K65R and M184V mutations are two of the most common NRTI mutations to develop in those infected with HIV-1 subtype C, who are failing on a first-line NNRTI-backbone regimen containing TDF and 3TC [42,43]. This was also observed in our routine genotypic drug resistance testing database, which showed that the K65R/M184V combination was the second highest in terms of prevalence (i.e., 2.35%). While K65R and M184V are antagonistic mutations and cause a viral fitness deficit [44], the Stanford HIV Drug Resistance Algorithm [17] (https://hivdb.stanford.edu/) predicts the combination to confer high-level resistance to ABC and XTC, and intermediate resistance to TDF. Our data showed that this combination conferred potential low-level resistance to ISL. The K65R mutation potentially sensitised the M184V mutant to ISL as it led to a 1.8-fold reduction in FC. This increase in ISL susceptibility of K65R+M184V has also been observed in previous studies [37,45]. Interestingly, such an increase in susceptibility with K65R+M184V has also been observed with AZT [44]. In addition, we observed that combinations that contained other NRTI mutations (i.e., A62V, K70T, Y115F, or K219E), in addition to K65R+M184V, were susceptible to, or conferred only a potential low-level resistance to ISL. The presence of other NRTI resistance mutations in combination with K65R+M184V did not cause an additional decrease in ISL susceptibility. This finding is significant, especially in the context of subtype C, as we observed that 13.11% (n = 724) of the sequences from PLWH failing second-line therapy, that contained NRTI resistance mutations, contained the K65R+M184V mutations in combination with other mutations.
Although AZT or ABC were occasionally included in first-line treatment, AZT was one of the main NRTIs used in PI-based second-line treatment. AZT has a low barrier to the development of drug resistance and often selects for TAMs [46,47,48]. TAMs occur in two distinguishable, yet overlapping patterns [26,49,50]: Type 1 TAMs include M41L, L210W, and T215Y; Type 2 TAMs include D67N, K70R, T215F, and K219E/Q. As 18.2% (n = 4,275) of patients in the routine genotypic resistance database received AZT in their current regimens, several Type-1 (M41L, T215Y) and Type-2 (D67N, K70R, T215F, K219E/Q) TAMs were observed. The TAMs were observed either alone, or in combination with other NRTI drug resistance mutations. Individually, most of the PSVs with single TAMs were susceptible to ISL, although the T215Y mutant conferred potential low-level resistance to ISL. According to the Stanford HIV Drug Resistance Algorithm (https://hivdb.stanford.edu/), T215Y is predicted to be susceptible to XTC, whilst conferring potential low-level resistance (~1.5-fold) to ABC and TDF. The remaining TAMs are predicted to be susceptible to XTC, ABC, and TDF, with the exception of T215F, which is predicted to confer potential low-level resistance to ABC and TDF. Limited data is available on the susceptibility of single TAM-containing variants on ISL susceptibility. A previous study reported a FC ≤ 2.5 for three single TAMs (i.e., M41L, L74I, T215Y) [12], which is in agreement with our findings for these single mutations.
Although individual TAMs did not affect ISL susceptibility in our study, the combination of TAMs with other NRTI drug resistance mutations led to a decrease in ISL susceptibility in most cases. This was most evident for combinations with TAM-2 mutations. The combination of TAM-1 mutations (i.e., M41L+M184V+T215Y) did not decrease ISL susceptibility. However, M41L+M184V, with the addition of the T215F TAM-2 mutation, caused a significant decrease in ISL susceptibility. Furthermore, the addition of other TAM-2 mutations (i.e., K70R/K219EQ, K70R/T215F/K219Q) to D67N/M184V significantly decreased ISL susceptibility. On the contrary, the addition of K70E (a non-TAM) or K70R (a TAM-2) to D67N/M184V increased ISL susceptivity, relative to that of D67N/M184V. Brenner et. al [51] showed that the combination of three TAM-1 mutations (M41L+L210W+T215Y) conferred low-level resistance to ISL (2.1 FC). However, in combination with M184V, it conferred high-level resistance to ISL (40 FC) [51]. In addition, Diamond et. al [12] showed that four TAM-2 mutations together (D67N+K70R+T215F+K219Q), in the absence of M184V, conferred only low-level resistance to ISL (3.8 FC).
The A62V mutation has been observed mostly in conjunction with the K65R mutation, partially correcting the viral fitness deficit caused by the latter [52]. However, in this study, the A62V+K65R+M184V mutation was found to be susceptible to ISL, most likely due to the hyper-susceptible nature of K65R+M184V. Contrasting this, the absence of the K65R mutant had a notable effect, as the A62V+M184V combination conferred a low to intermediate level of resistance to ISL. Although this may be a point of concern, the prevalence of the A62V+M184V combination was observed in only 1.01% of sequences with NRTI drug resistance mutations in the genotypic drug resistance database.
We observed three variants at position 70 (i.e., K70E, K70R, K70T) with FC values below the TCO. Two of these (i.e., K70E, K70T) showed a significant hyper-susceptibility to ISL. When in combination with D67N+M184V, K70E and K70R seemed to sensitise the D67N/M184V mutation to ISL. However, the addition of more TAM-2 mutations to D67N/K70R/M184V significantly reduced ISL susceptibility.
The L74V mutation showed a potentially low level of resistance to ISL (FC = 2.5 ± 1.5 FC) in our study, which was higher than reported by Grobler et al. (2022) in subtype B (FC = 0.21 ± 0.07 FC) [37]. Further evaluation of the L74V mutation in both subtypes B and C PSVs in this study did show potential hyper-susceptibility in one of the four HIV-1 subtype B PSVs (i.e., DS9-L74V). However, this was not observed in the remaining subtype B or C PSVs, and although unexplored, suggests that hyper-susceptibility may be strain-specific for this NRTI resistan mutation. An additional mutation at position 74 (i.e., L74I) was observed at a low frequency among our sequences and did not affect ISL susceptibility by itself. However, in combination with M184V, it significantly reduced ISL susceptibility.
The A114S NRTI resistance mutation was initially identified in a selection study, which showed that it incurs a high level of resistance to ISL (24-fold) [41]. Similarly, another in vitro dose-escalation study reported the A114S+M184V combination to have an FC of 37.9 to ISL, as well as confer very high levels of resistance to XTC (3TC: FC > 368, FTC: FC > 990) [12]. Interestingly, both studies reported the hyper-susceptible nature of the dual mutation against TDF (50-fold [41]/0.03 FC [12])and AZT (0.4 FC) [12]. Although the A114S mutation alone did not decrease susceptibility to ISL in our study (i.e., 2.1 ± 0.3 FC), a significant reduction in ISL susceptibility (i.e., FC > 60) was observed for the A114S+M184V combination. However, this mutation is uncommon, as the Stanford HIV Drug Resistance Database [53,54] (https://hivdb.stanford.edu/) depicted its frequency as < 0.01%. In addition, the A114S+M184V mutant combination was shown to cause a deficit in viral fitness, leading to a significantly decreased infectivity when compared to wild-type [41]. Therefore, although the A114S+M184V combination is of concern as it severely impacts on ISL susceptibility, its low prevalence in PLWH and its detrimental impact on the virus fitness is reassuring.
This study has its limitations. The different classifications of phenotypic resistance were placed at increments of the TCO and are not linked to clinical outcomes. These classifications merely served as a means to contextualize the degrees of ISL resistance in the assay that was employed. Additionally, the average FC value of at least three independent screens for the resistance classifications of each mutant PSV was utilized. For some PSVs, the phenotypic responses were either on the cusp of two neighbouring classifications or the standard deviations overlapped two or three neighbouring classifications. However, despite this, this study’s results were mostly in agreement with the limited literature on phenotypic ISL resistance currently available. We also observed large variations in FC values in the repeat assays for some of the single and combinations of NRTI resistance mutations. This may also be strain-specific, since we did not observe similar variations in the strain-derived PSVs compared to the MJ4-L74V PSV. However, these variations (up to ±1.7 FC) were greater than the assay variation for the wild-type MJ4 PSV (i.e., ±0.6 FC), suggesting that there may be some other mechanisms in place that impact on the interaction between ISL and certain mutated amino acids in the active site of Reverse Transcriptase. We argue that this is a plausible hypothesis, considering the unique mechanisms of action of ISL. Lastly, since all the NRTI mutation combinations observed among the sequences in our database contained M184V, TAMs in the absence of M184V were not investigated in this study. However, the result of our study reflects on a “real world” setting and its implication on ISL as a treatment option following failure on AZT/XTC-containing regimens.
ARV resistance and cross-resistance is the Achillies heel of cART. As we have shown in this study, regimen choices may have an impact on the efficacy of novel ARVs, even those with novel mechanisms of action. This study implies that failure on a TDF/XTC-containing cART would possibly have a minimal impact on ISL efficacy in PLWH. However, due to frequent selection of TAMs in conjunction with the M184V NRTI resistance mutation, PLWH failing AZT/XTC-containing cART would most likely not benefit from ISL. With the shift from NNRTI-based first-line regimens (e.g. TDF/3TC/EFV) to INSTI-based regimens (e.g. TDF/3TC/DTG) [55], ISL would remain efficacious since prevalent NRTI resistance mutations, selected for by TDF (i.e., K65R) and 3TC (M184V), would have a low impact on ISL susceptibility. Despite the effectiveness of PI-based second-line regimens that contain AZT/FTC, the selection of TAMs in the presence of M184V could negate the efficacy of ISL. The Nucleosides and Darunavir/Dolutegravir in Africa (NADIA) Trial demonstrated that TDF/3TC was non-inferior to AZT/3TC in second-line cART [56]. The data from this trial supports maintaining TDF and 3TC at the time of a switch to second-line treatment. Due to the absence of AZT in such regimens, ISL could potentially be used effectively as an alternative switching option, or in subsequent third-line cART. However, the full treatment histories of PLWH should be taken into consideration prior to switching to ISL-containing regimens.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: Reference sequences of laboratory-adapted strains; Table S2: Sequences of primers used for site-directed mutagenesis to synthesize single and combination mutation profiles; Table S3: NRTI exposure of PLWH from which sequences were obtained; Table S4: The prevalence of the top 20 combination NRTI mutations in patients with NRTI drug-resistance mutations; Table S5: The prevalence of the individual mutations that constitute the top 20 combination NRTI mutations in patients with NRTI drug-resistance mutations; Table S6: Average IC50- and FC values of single NRTI resistant mutants in subtype C; Table S7: Intra-subtype comparison of fold-change values of the L74V mutant in laboratory-adapted HIV-1 PSVs in response to ISL; Table S8: Average IC50- and FC values of combination NRTI-resistant mutants in subtype C. References [57,58,59,60,61,62,63] were cited in the Supplementary Materials.
Author Contributions
Conceptualization, A.E.B.; resources, A.E.B., K.S., M.P.; writing—original draft preparation, H.B.; writing—review and editing, A.E.B., K.S., M.P.; visualization, H.B. and A.E.B.; supervision, A.E.B., K.S.; project administration, A.E.B.; funding acquisition, A.E.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Poliomyelitis Research Foundation (PRF), grant number 21/66.
Institutional Review Board Statement
Ethical review and approval were waived for this study due to this study not involving humans or animals. Ethics waiver obtained from the Human Research Ethics Committee (University of the Witwatersrand): reference no. R19/49, protocol no. W-CBP-220503-01.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Materials, further inquiries can be directed to the corresponding authors.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- WHO Fact Sheets: HIV and AIDS Available online:. Available online: https://www.who.int/news-room/fact-sheets/detail/hiv-aids (accessed on 18 September 2024).
- Lee, L.M.; Karon, J.M.; Selik, R.; Neal, J.J.; Fleming, P.L. Survival after AIDS Diagnosis in Adolescents and Adults during the Treatment Era, United States, 1984-1997. JAMA 2001, 285, 1308–1315. [Google Scholar] [CrossRef] [PubMed]
- Nieuwkerk, P.T.; Gisolf, E.H.; Reijers, M.H.; Lange, J.M.; Danner, S.A.; Sprangers, M.A. ; NATIVE Study Group; PROMETHEUS Study Group; ADAM Study Group Long-Term Quality of Life Outcomes in Three Antiretroviral Treatment Strategies for HIV-1 Infection. AIDS 2001, 15, 1985–1991. [Google Scholar] [CrossRef]
- Mate, K.K.V.; Engler, K.; Lessard, D.; Lebouché, B. Barriers to Adherence to Antiretroviral Therapy: Identifying Priority Areas for People with HIV and Healthcare Professionals. Int J STD AIDS 2023, 34, 677–686. [Google Scholar] [CrossRef]
- Kheswa, J.G. Exploring the Factors and Effects of Non-Adherence to Antiretroviral Treatment by People Living with HIV/AIDS. Indo-Pacific Journal of Phenomenology 2017, 17, 1–11. [Google Scholar] [CrossRef]
- Michailidis E; Marchand B; Kodama EN; Singh K; Matsuoka M; Kirby KA; Ryan EM; Sawani AM; Nagy E; Ashida N; et al. Mechanism of Inhibition of HIV-1 Reverse Transcriptase by 4’-Ethynyl-2-Fluoro-2’-Deoxyadenosine Triphosphate, a Translocation-Defective Reverse Transcriptase Inhibitor. The Journal of biological chemistry 2009, 284. [Google Scholar] [CrossRef]
- Matthews, R.P.; Rudd, D.J.; Fillgrove, K.L.; Zhang, S.; Tomek, C.; Stoch, S.A.; Iwamoto, M. A Phase 1 Study to Evaluate the Drug Interaction Between Islatravir (MK-8591) and Doravirine in Adults Without HIV. Clinical Drug Investigation 2021, 41, 629. [Google Scholar] [CrossRef]
- Amy Colson; Gordon Crofoot; Peter J. Ruane; Moti Ramgopal; Alexandra W. Dretler; Ronald G. Nahass; Gary Sinclair; Mezgebe Berhe; Chris Deaton; Angela S. Liu; et al. Efficacy and Safety of Weekly Islatravir Plus Lenacapavir in PWH at 24 Weeks: A Phase II Study - CROI Conference.; Denver, Colorado, USA, 2024; Vol. Abstract 208. 7 March.
- Correll T; Molina JM; Klopfer S; Grandhi A; Lahoulou R; Zhou Y-P; Eves K; Squires K O46 Total Lymphocyte and CD4+ T-Cell Count Changes in Participants Receiving Islatravir (0. 25, 0.75 and 2.25 Mg QD) and Doravirine +/- Lamivudine: Post-Hoc Analysis from a Phase 2b Dose- Ranging Study (P011). Journal of the International AIDS Society 2022, 25, e26009. [Google Scholar] [CrossRef]
- Mills AM; Rizzardini G; Ramgopal MN; Osiyemi OO; Bogner JR; Hagins DP; Paredes R; Reynes J; Rockstroh JK; Carr A; et al. Switch to Fixed-Dose Doravirine (100 Mg) with Islatravir (0·75 Mg) Once Daily in Virologically Suppressed Adults with HIV-1 on Bictegravir, Emtricitabine, and Tenofovir Alafenamide: 48-Week Results of a Phase 3, Randomised, Controlled, Double-Blind, Non-Inferiority Trial. The lancet. HIV 2024, 11. [Google Scholar] [CrossRef]
- Diamond, T.L.; Goh, S.L.; Ngo, W.; Rodriguez, S.; Xu, M.; Klein, D.J.; Grobler, J.A.; Asante-Appiah, E. No Antagonism or Cross-Resistance and a High Barrier to the Emergence of Resistance in Vitro for the Combination of Islatravir and Lenacapavir. Antimicrobial Agents and Chemotherapy 2024, 68, e00334–24. [Google Scholar] [CrossRef]
- Diamond, T.L.; Ngo, W.; Xu, M.; Goh, S.L.; Rodriguez, S.; Lai, M.-T.; Asante-Appiah, E.; Grobler, J.A. Islatravir Has a High Barrier to Resistance and Exhibits a Differentiated Resistance Profile from Approved Nucleoside Reverse Transcriptase Inhibitors (NRTIs). Antimicrob Agents Chemother 2022, 66, e0013322. [Google Scholar] [CrossRef]
- Singh, K.; Flores, J.A.; Kirby, K.A.; Neogi, U.; Sonnerborg, A.; Hachiya, A.; Das, K.; Arnold, E.; McArthur, C.; Parniak, M.; et al. Drug Resistance in Non-B Subtype HIV-1: Impact of HIV-1 Reverse Transcriptase Inhibitors. Viruses 2014, 6, 3535. [Google Scholar] [CrossRef]
- Hemelaar, J.; Elangovan, R.; Yun, J.; Dickson-Tetteh, L.; Fleminger, I.; Kirtley, S.; Williams, B.; Gouws-Williams, E.; Ghys, P.D.; Abimiku, A.G.; et al. Global and Regional Molecular Epidemiology of HIV-1, 1990–2015: A Systematic Review, Global Survey, and Trend Analysis. The Lancet Infectious Diseases 2019, 19, 143–155. [Google Scholar] [CrossRef]
- UNAIDS Country Facts Sheet: South Africa (2023). 2024.
- South Africa National Department of Health 2023 ART Clinical Guidelines for the Management of HIV in Adults, Pregnancy and Breastfeeding, Adolescents, Children, Infants and Neonates. 2023.
- Liu TF; Shafer RW Web Resources for HIV Type 1 Genotypic-Resistance Test Interpretation. Clinical infectious diseases : an official publication of the Infectious Diseases Society of America 2006, 42. [CrossRef]
- Yang, Z.; Chen, Z.; Zhang, Y. A Simple and Economical Site-Directed Mutagenesis Method for Large Plasmids by Direct Transformation of Two Overlapping PCR Fragments. Biotechniques 2022, 73, 239–245. [Google Scholar] [CrossRef]
- Gupta, R.K.; Kohli, A.; McCormick, A.L.; Towers, G.J.; Pillay, D.; Parry, C.M. Full-Length HIV-1 Gag Determines Protease Inhibitor Susceptibility within in Vitro Assays. AIDS 2010, 24, 1651–1655. [Google Scholar] [CrossRef]
- Naldini, L.; Blömer, U.; Gallay, P.; Ory, D.; Mulligan, R.; Gage, F.H.; Verma, I.M.; Trono, D. In Vivo Gene Delivery and Stable Transduction of Nondividing Cells by a Lentiviral Vector. Science 1996, 272, 263–267. [Google Scholar] [CrossRef]
- Parry, C.M.; Kohli, A.; Boinett, C.J.; Towers, G.J.; McCormick, A.L.; Pillay, D. Gag Determinants of Fitness and Drug Susceptibility in Protease Inhibitor-Resistant Human Immunodeficiency Virus Type 1. J Virol 2009, 83, 9094–9101. [Google Scholar] [CrossRef]
- Diamond, T.L.; Ngo, W.; Xu, M.; Goh, S.L.; Rodriguez, S.; Lai, M.-T.; Asante-Appiah, E.; Grobler, J.A. Islatravir Has a High Barrier to Resistance and Exhibits a Differentiated Resistance Profile from Approved Nucleoside Reverse Transcriptase Inhibitors (NRTIs). Antimicrob Agents Chemother 66. [CrossRef]
- Grobler, J.A.; Huang, Q.; Hazuda, D.J.; Lai, M. Efficacy of MK-8591 against Diverse HIV-1 Subtypes and NRTI-Resistant Clinical Isolates. In Proceedings of the Presented at HIV Drug Therapy, Glasgow, UK; October 28 2018. [Google Scholar]
- SANDOH National Department of Health, SA. 2015 National Consolidated for the Prevention of Mother-to-Child Transmission of HIV (PMTCT) and the Management of HIV in Children, Adolescents and Adulta 2015.
- SANDOH National Department of Health, SA. 2019 ART Clinical Guidelines for the Management of HIV in Adults, Pregnancy, Adolescents, Children, Infants and Neonates. 2019.
- Marcelin AG; Delaugerre C; Wirden M; Viegas P; Simon A; Katlama C; Calvez V Thymidine Analogue Reverse Transcriptase Inhibitors Resistance Mutations Profiles and Association to Other Nucleoside Reverse Transcriptase Inhibitors Resistance Mutations Observed in the Context of Virological Failure. Journal of medical virology 2004, 72. [CrossRef]
- Tisdale, M.; Alnadaf, T.; Cousens, D. Combination of Mutations in Human Immunodeficiency Virus Type 1 Reverse Transcriptase Required for Resistance to the Carbocyclic Nucleoside 1592U89. Antimicrob Agents Chemother 1997, 41, 1094–1098. [Google Scholar] [CrossRef]
- Boucher, C.A.; Cammack, N.; Schipper, P.; Schuurman, R.; Rouse, P.; Wainberg, M.A.; Cameron, J.M. High-Level Resistance to (-) Enantiomeric 2’-Deoxy-3’-Thiacytidine in Vitro Is Due to One Amino Acid Substitution in the Catalytic Site of Human Immunodeficiency Virus Type 1 Reverse Transcriptase. Antimicrob Agents Chemother 1993, 37, 2231–2234. [Google Scholar] [CrossRef]
- Tisdale, M.; Kemp, S.D.; Parry, N.R.; Larder, B.A. Rapid in Vitro Selection of Human Immunodeficiency Virus Type 1 Resistant to 3’-Thiacytidine Inhibitors Due to a Mutation in the YMDD Region of Reverse Transcriptase. Proceedings of the National Academy of Sciences of the United States of America 1993, 90, 5653. [Google Scholar] [CrossRef]
- Brenner, B.G.; Oliveira, M.; Doualla-Bell, F.; Moisi, D.D.; Ntemgwa, M.; Frankel, F.; Essex, M.; Wainberg, M.A. HIV-1 Subtype C Viruses Rapidly Develop K65R Resistance to Tenofovir in Cell Culture. AIDS 2006, 20, F9–13. [Google Scholar] [CrossRef]
- Brenner, B.G.; Coutsinos, D. The K65R Mutation in HIV-1 Reverse Transcriptase: Genetic Barriers, Resistance Profile and Clinical Implications. HIV therapy 2009, 3, 583. [Google Scholar] [CrossRef]
- Margot, N.A.; Isaacson, E.; McGowan, I.; Cheng, A.K.; Schooley, R.T.; Miller, M.D. Genotypic and Phenotypic Analyses of HIV-1 in Antiretroviral-Experienced Patients Treated with Tenofovir DF. AIDS 2002, 16, 1227–1235. [Google Scholar] [CrossRef]
- Maldonado, J.O.; Mansky, L.M. The HIV-1 Reverse Transcriptase A62V Mutation Influences Replication Fidelity and Viral Fitness in the Context of Multi-Drug-Resistant Mutations. Viruses 2018, 10, 376. [Google Scholar] [CrossRef]
- Steegen, K.; Bronze, M.; Papathanasopoulos, M.A.; van Zyl, G.; Goedhals, D.; Variava, E.; MacLeod, W.; Sanne, I.; Stevens, W.S.; Carmona, S. HIV-1 Antiretroviral Drug Resistance Patterns in Patients Failing NNRTI-Based Treatment: Results from a National Survey in South Africa. Journal of Antimicrobial Chemotherapy 2017, 72, 210–219. [Google Scholar] [CrossRef]
- Melikian Gl; Rhee SY; Taylor J; Fessel WJ; Kaufman D; W, T. ; Pv, T.-C.; A, Z.; Gk, R.; R, K.; et al. Standardized Comparison of the Relative Impacts of HIV-1 Reverse Transcriptase (RT) Mutations on Nucleoside RT Inhibitor Susceptibility. Antimicrobial agents and chemotherapy 2012, 56. [Google Scholar] [CrossRef]
- Weber, J.; Chakraborty, B.; Weberova, J.; Miller, M.D.; Quiñones-Mateu, M.E. Diminished Replicative Fitness of Primary Human Immunodeficiency Virus Type 1 Isolates Harboring the K65R Mutation. Journal of Clinical Microbiology 2005, 43, 1395. [Google Scholar] [CrossRef]
- J. A. Grobler; Q. Huang; D.J. Hazuda; M.-T. Lai Efficacy of MK-8591 against Diverse HIV-1 Subtypes and NRTI-Resistant Clinical Isolates. Journal of the International AIDS Society 2022, 25, e26009. [Google Scholar] [CrossRef]
- Murphey-Corb M; Rajakumar P; Michael H; Nyaundi J; Didier PJ; Ab, R. ; H, M.; Sg, S.; Ma, P. Response of Simian Immunodeficiency Virus to the Novel Nucleoside Reverse Transcriptase Inhibitor 4’-Ethynyl-2-Fluoro-2’-Deoxyadenosine in Vitro and in Vivo. Antimicrobial agents and chemotherapy 2012, 56. [Google Scholar] [CrossRef]
- Borroto-Esoda, K.; Parkin, N.; Miller, M.D. A Comparison of the Phenotypic Susceptibility Profiles of Emtricitabine and Lamivudine. Antivir Chem Chemother 2007, 18, 297–300. [Google Scholar] [CrossRef]
- Whitcomb JM; Parkin NT; Chappey C; Hellmann NS; Petropoulos CJ Broad Nucleoside Reverse-Transcriptase Inhibitor Cross-Resistance in Human Immunodeficiency Virus Type 1 Clinical Isolates. The Journal of infectious diseases 2003, 188. [CrossRef]
- Cilento, M.E.; Reeve, A.B.; Michailidis, E.; Ilina, T.V.; Nagy, E.; Mitsuya, H.; Parniak, M.A.; Tedbury, P.R.; Sarafianos, S.G. Development of Human Immunodeficiency Virus Type 1 Resistance to 4′-Ethynyl-2-Fluoro-2′-Deoxyadenosine Starting with Wild-Type or Nucleoside Reverse Transcriptase Inhibitor-Resistant Strains. Antimicrobial Agents and Chemotherapy 2021, 65. [Google Scholar] [CrossRef]
- Invernizzi, C.F.; Coutsinos, D.; Oliveira, M.; Moisi, D.; Brenner, B.G.; Wainberg, M.A. Signature Nucleotide Polymorphisms at Positions 64 and 65 in Reverse Transcriptase Favor the Selection of the K65R Resistance Mutation in HIV-1 Subtype C. J Infect Dis 2009, 200, 1202–1206. [Google Scholar] [CrossRef]
- TenoRes Study Group Global Epidemiology of Drug Resistance after Failure of WHO Recommended First-Line Regimens for Adult HIV-1 Infection: A Multicentre Retrospective Cohort Study. The Lancet. Infectious Diseases 2016, 16, 565. [CrossRef]
- White KL; Margot NA; Wrin T; Petropoulos CJ; Miller MD; Naeger LK Molecular Mechanisms of Resistance to Human Immunodeficiency Virus Type 1 with Reverse Transcriptase Mutations K65R and K65R+M184V and Their Effects on Enzyme Function and Viral Replication Capacity. Antimicrobial agents and chemotherapy 2002, 46. [CrossRef]
- Cilento ME; Wen X; Reeve AB; Ukah OB; Snyder AA; Carrillo CM; Smith CP; Edwards K; Wahoski CC; Kitzler DR; et al. HIV-1 Resistance to Islatravir/Tenofovir Combination Therapy in Wild-Type or NRTI-Resistant Strains of Diverse HIV-1 Subtypes. Viruses 2023, 15. [Google Scholar] [CrossRef]
- Harrigan, P.R.; Kinghorn, I.; Bloor, S.; Kemp, S.D.; Nájera, I.; Kohli, A.; Larder, B.A. Significance of Amino Acid Variation at Human Immunodeficiency Virus Type 1 Reverse Transcriptase Residue 210 for Zidovudine Susceptibility. J Virol 1996, 70, 5930–5934. [Google Scholar] [CrossRef]
- Kellam, P.; Boucher, C.A.; Larder, B.A. Fifth Mutation in Human Immunodeficiency Virus Type 1 Reverse Transcriptase Contributes to the Development of High-Level Resistance to Zidovudine. Proceedings of the National Academy of Sciences of the United States of America 1992, 89, 1934. [Google Scholar] [CrossRef]
- Larder, B.A.; Kemp, S.D. Multiple Mutations in HIV-1 Reverse Transcriptase Confer High-Level Resistance to Zidovudine (AZT). Science 1989, 246, 1155–1158. [Google Scholar] [CrossRef] [PubMed]
- Cozzi-Lepri, A.; Ruiz, L.; Loveday, C.; Phillips, A.N.; Clotet, B.; Reiss, P.; Ledergerber, B.; Holkmann, C.; Staszewski, S.; Lundgren, J.D.; et al. Thymidine Analogue Mutation Profiles: Factors Associated with Acquiring Specific Profiles and Their Impact on the Virological Response to Therapy. Antiviral Therapy 2005, 10, 791–802. [Google Scholar] [CrossRef] [PubMed]
- Flandre, P.; Descamps, D.; Joly, V.; Meiffrédy, V.; Tamalet, C.; Izopet, J.; Aboulker, J.-P.; Brun-Vézinet, F. Predictive Factors and Selection of Thymidine Analogue Mutations by Nucleoside Reverse Transcriptase Inhibitors According to Initial Regimen Received. Antiviral Therapy 2003, 8, 65–72. [Google Scholar] [CrossRef]
- Brenner BG; Oliveira M; Ibanescu RI; Routy JP; Thomas R Doravirine Responses to HIV-1 Viruses Bearing Mutations to NRTIs and NNRTIs under in Vitro Selective Drug Pressure. The Journal of antimicrobial chemotherapy 2023, 78. [CrossRef]
- Svarovskaia, E.S.; Feng, J.Y.; Margot, N.A.; Myrick, F.; Goodman, D.; Ly, J.K.; White, K.L.; Kutty, N.; Wang, R.; Borroto-Esoda, K.; et al. The A62V and S68G Mutations in HIV-1 Reverse Transcriptase Partially Restore the Replication Defect Associated with the K65R Mutation. J Acquir Immune Defic Syndr 2008, 48, 428–436. [Google Scholar] [CrossRef]
- Rhee, S.; Gonzales, M.J.; Kantor, R.; Betts, B.J.; Ravela, J.; Shafer, R.W. Human Immunodeficiency Virus Reverse Transcriptase and Protease Sequence Database. Nucleic Acids Research 2003, 31, 298–303. [Google Scholar] [CrossRef]
- Shafer, R.W. Rationale and Uses of a Public HIV Drug-Resistance Database. J INFECT DIS 2006, 194, S51–S58. [Google Scholar] [CrossRef]
- Iacobucci, G. HIV: Dolutegravir Should Be Preferred Treatment Option in All Populations, Says WHO. BMJ 2019, 366, l4831. [Google Scholar] [CrossRef]
- Paton NI; Musaazi J; Kityo C; Walimbwa S; Hoppe A; Balyegisawa A; Asienzo J; Kaimal A; Mirembe G; Lugemwa A; et al. Efficacy and Safety of Dolutegravir or Darunavir in Combination with Lamivudine plus Either Zidovudine or Tenofovir for Second-Line Treatment of HIV Infection (NADIA): Week 96 Results from a Prospective, Multicentre, Open-Label, Factorial, Randomised, Non-Inferiority Trial. The Lancet. HIV 2022, 9. [Google Scholar] [CrossRef]
- Obasa, A.E.; Engelbrecht, S.; Jacobs, G.B. Near Full-Length HIV-1 Subtype B Sequences from the Early South African Epidemic, Detecting a BD Unique Recombinant Form (URF) from a Sample in 1985. Sci Rep 2019, 9, 6227. [Google Scholar] [CrossRef]
- Mikhail, M.; Wang, B.; Lemey, P.; Beckholdt, B.; Vandamme, A.-M.; Gill, M.J.; Saksena, N.K. Full-Length HIV Type 1 Genome Analysis Showing Evidence for HIV Type 1 Transmission from a Nonprogressor to Two Recipients Who Progressed to AIDS. AIDS Res Hum Retroviruses 2005, 21, 575–579. [Google Scholar] [CrossRef]
- Ledwaba, J.; Sayed, Y.; Pillay, V.; Morris, L.; Hunt, G. Low Frequency of Protease Inhibitor Resistance Mutations and Insertions in HIV-1 Subtype C Protease Inhibitor-Naïve Sequences. AIDS Res Hum Retroviruses 2019, 35, 673–678. [Google Scholar] [CrossRef]
- Maldarelli, F.; Kearney, M.; Palmer, S.; Stephens, R.; Mican, J.; Polis, M.A.; Davey, R.T.; Kovacs, J.; Shao, W.; Rock-Kress, D.; et al. HIV Populations Are Large and Accumulate High Genetic Diversity in a Nonlinear Fashion. J Virol 2013, 87, 10313–10323. [Google Scholar] [CrossRef]
- Papathanasopoulos, M.A.; Cilliers, T.; Morris, L.; Mokili, J.L.; Dowling, W.; Birx, D.L.; McCutchan, F.E. Full-Length Genome Analysis of HIV-1 Subtype C Utilizing CXCR4 and Intersubtype Recombinants Isolated in South Africa. AIDS Res Hum Retroviruses 2002, 18, 879–886. [Google Scholar] [CrossRef]
- Williamson, C.; Morris, L.; Maughan, M.F.; Ping, L.-H.; Dryga, S.A.; Thomas, R.; Reap, E.A.; Cilliers, T.; van Harmelen, J.; Pascual, A.; et al. Characterization and Selection of HIV-1 Subtype C Isolates for Use in Vaccine Development. AIDS Research and Human Retroviruses 2003, 19, 133–144. [Google Scholar] [CrossRef]
- Van Harmelen, J.; Williamson, C.; Kim, B.; Morris, L.; Carr, J.; Abdool Karim, S.S.; McCutchan, F. Characterization of Full-Length HIV Type 1 Subtype C Sequences from South Africa. AIDS Research and Human Retroviruses 2001, 17, 1527–1531. [Google Scholar] [CrossRef]
Figure 1.
Variation in IC50 and FC of ISL against wild-type PSV. (A) Multiple independent in vitro assays (n = 41) were conducted to determine the IC50 of ISL against the wild-type HIV-1 subtype C virus (i.e., p8.9MJ4). The average IC50 = 8.3 nM ± 5.0 nM (median IC50 = 7.3 nM). (B) Each IC50 value was divided by the average IC50 value to determine the FC. The average FC was therefore 1.0 ± 0.6 FC (median FC = 0.87). TCO: obtained from the 99th percentile of the IC50 values. (C) Multiple independent in vitro assays (n = 13) were conducted to determine the IC50 value of ISL against the wild-type HIV-1 subtype B virus (i.e., p8.9NSX). The average IC50 concentration was shown to be 7.9 nM (median IC50 = 5.9 nM), with a standard deviation of ± 5.5 nM. (D) Each IC50 value was divided by the average IC50 value to obtain an average FC value of 1 (median FC = 0.74). The standard deviation of the FC value was shown to be ± 0.7.
Figure 1.
Variation in IC50 and FC of ISL against wild-type PSV. (A) Multiple independent in vitro assays (n = 41) were conducted to determine the IC50 of ISL against the wild-type HIV-1 subtype C virus (i.e., p8.9MJ4). The average IC50 = 8.3 nM ± 5.0 nM (median IC50 = 7.3 nM). (B) Each IC50 value was divided by the average IC50 value to determine the FC. The average FC was therefore 1.0 ± 0.6 FC (median FC = 0.87). TCO: obtained from the 99th percentile of the IC50 values. (C) Multiple independent in vitro assays (n = 13) were conducted to determine the IC50 value of ISL against the wild-type HIV-1 subtype B virus (i.e., p8.9NSX). The average IC50 concentration was shown to be 7.9 nM (median IC50 = 5.9 nM), with a standard deviation of ± 5.5 nM. (D) Each IC50 value was divided by the average IC50 value to obtain an average FC value of 1 (median FC = 0.74). The standard deviation of the FC value was shown to be ± 0.7.
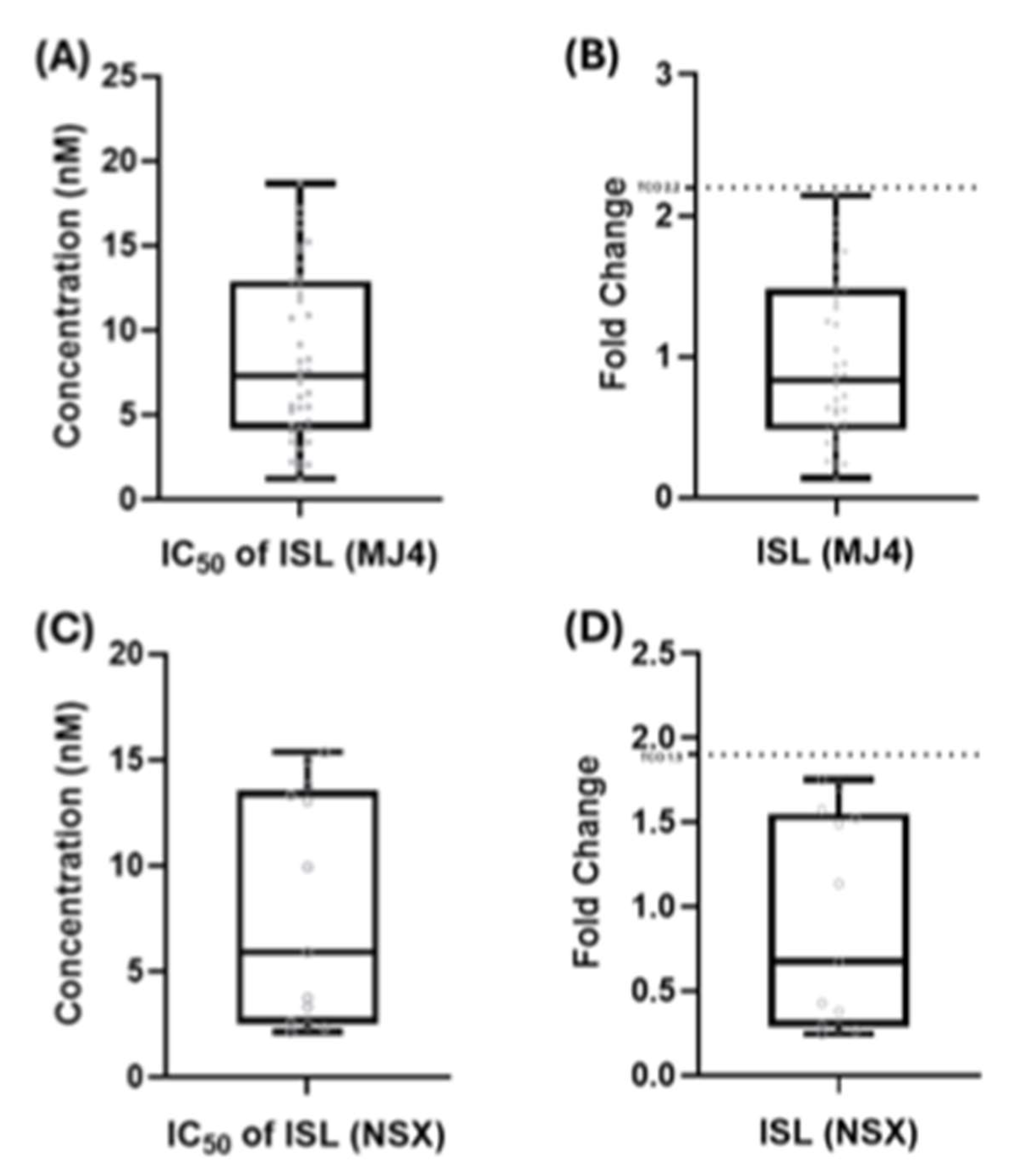
Figure 2.
Fold change in IC50 of single mutants in subtype C PSVs compared to the wild-type PSV. Following the in vitro phenotypic activity assays, the IC50 value for each mutant was compared against the IC50 of the MJ4 wild-type PSV, allowing the determination of fold changes. The 99th percentile of variation in the wild-type IC50 value, calculated to be 2.2 (TCO), served as the threshold for categorizing mutants as either susceptible or having a decreased susceptibility to ISL. Among the single mutants, M41L, K65R, D67N, K70E/R/T, L74I, A114S, Y115F, T215F, and K219E/Q demonstrated susceptibility to ISL. In contrast, A62V, L74V, and T215Y exhibited potential low-level resistance, and M184V exhibited low-level resistance. Susceptible (■), potential low-level resistance (■), low-level resistance (■), intermediate resistance (■), high-level resistance (■).
Figure 2.
Fold change in IC50 of single mutants in subtype C PSVs compared to the wild-type PSV. Following the in vitro phenotypic activity assays, the IC50 value for each mutant was compared against the IC50 of the MJ4 wild-type PSV, allowing the determination of fold changes. The 99th percentile of variation in the wild-type IC50 value, calculated to be 2.2 (TCO), served as the threshold for categorizing mutants as either susceptible or having a decreased susceptibility to ISL. Among the single mutants, M41L, K65R, D67N, K70E/R/T, L74I, A114S, Y115F, T215F, and K219E/Q demonstrated susceptibility to ISL. In contrast, A62V, L74V, and T215Y exhibited potential low-level resistance, and M184V exhibited low-level resistance. Susceptible (■), potential low-level resistance (■), low-level resistance (■), intermediate resistance (■), high-level resistance (■).
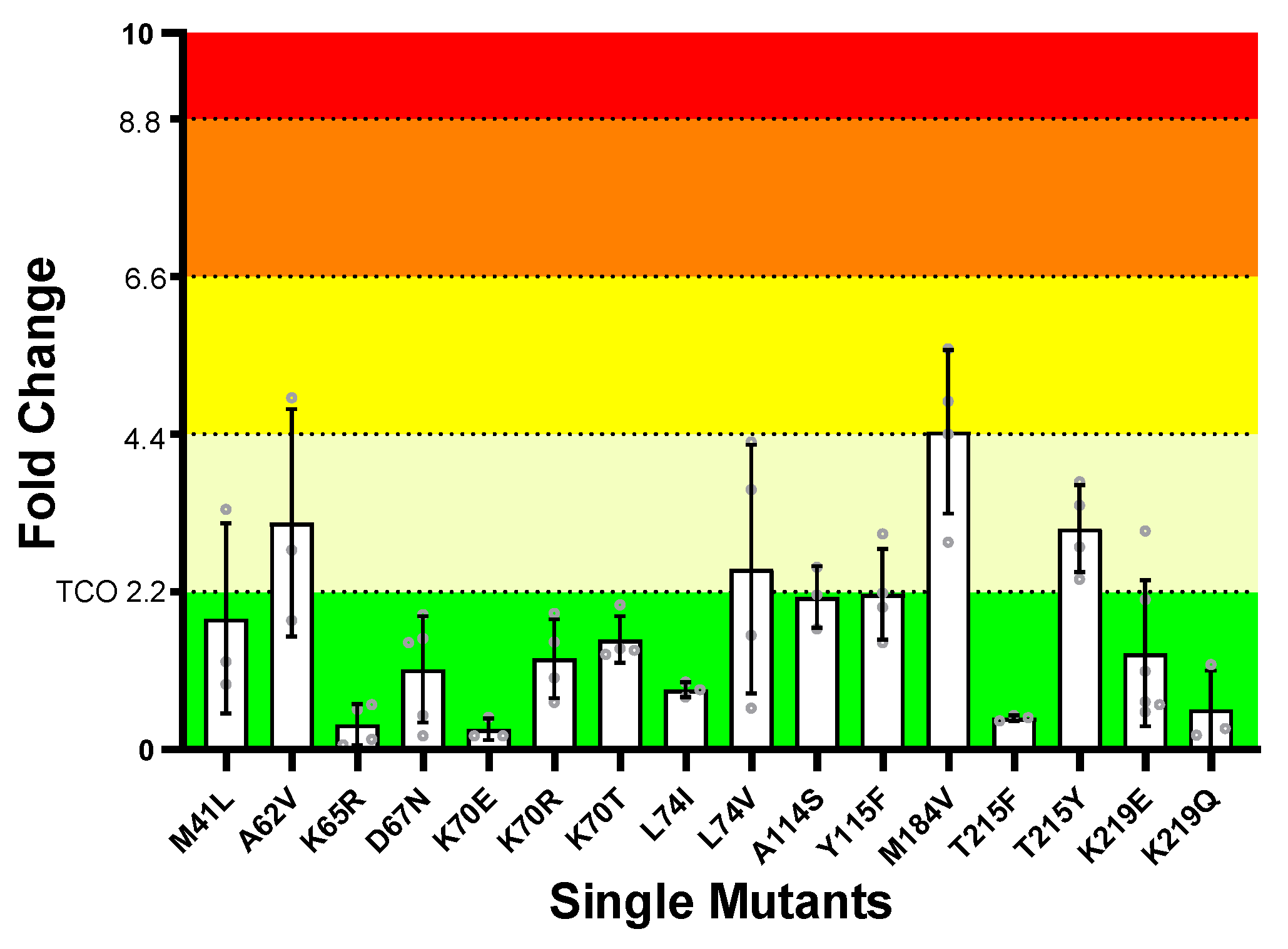
Figure 3.
Single mutant L74V in wild-type HIV-1 subtype B and C laboratory-adapted strains. A phenotypic activity assay was conducted to determine whether ISL had similar potencies against the single mutant L74V in different wild-type strains and subtypes of HIV-1. IC50 values are expressed as fold change differences to the IC50 of the relevant control subtype. The TCO for each subtype is indicated on the graph by a dotted line.
Figure 3.
Single mutant L74V in wild-type HIV-1 subtype B and C laboratory-adapted strains. A phenotypic activity assay was conducted to determine whether ISL had similar potencies against the single mutant L74V in different wild-type strains and subtypes of HIV-1. IC50 values are expressed as fold change differences to the IC50 of the relevant control subtype. The TCO for each subtype is indicated on the graph by a dotted line.
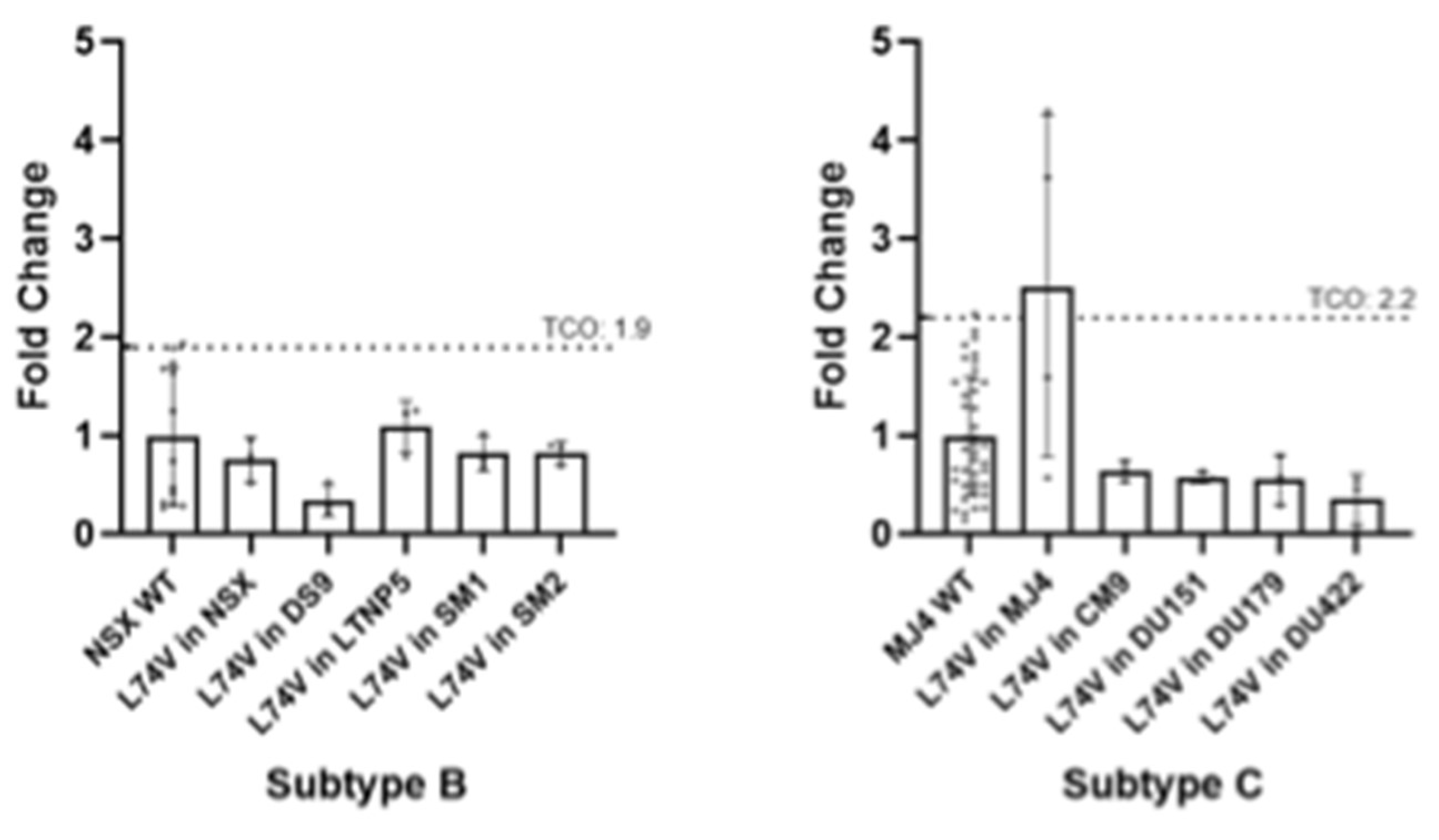
Figure 4.
IC50 values of L74V-in laboratory-adapted PSVs and inter-subtype one-way ANOVA test. (A) IC50 values of the L74V mutation in laboratory-adapted strains show that the LTNP5 PSV has the highest IC50 (0.0087 µM ± 0.0011 µM). DS9 had the lowest IC50 of 0.0028 µM ± 0.0013 µM. (B) Statistical analysis was performed using parametric parameters, assuming no equal standard deviation. The grid shows the p-values of the inter-subtype comparisons in IC50 values. A significant difference was observed between the IC50 values of the wild-type MJ4 PSV and the L74V-DS9 PSV (p-value 0.017).
Figure 4.
IC50 values of L74V-in laboratory-adapted PSVs and inter-subtype one-way ANOVA test. (A) IC50 values of the L74V mutation in laboratory-adapted strains show that the LTNP5 PSV has the highest IC50 (0.0087 µM ± 0.0011 µM). DS9 had the lowest IC50 of 0.0028 µM ± 0.0013 µM. (B) Statistical analysis was performed using parametric parameters, assuming no equal standard deviation. The grid shows the p-values of the inter-subtype comparisons in IC50 values. A significant difference was observed between the IC50 values of the wild-type MJ4 PSV and the L74V-DS9 PSV (p-value 0.017).
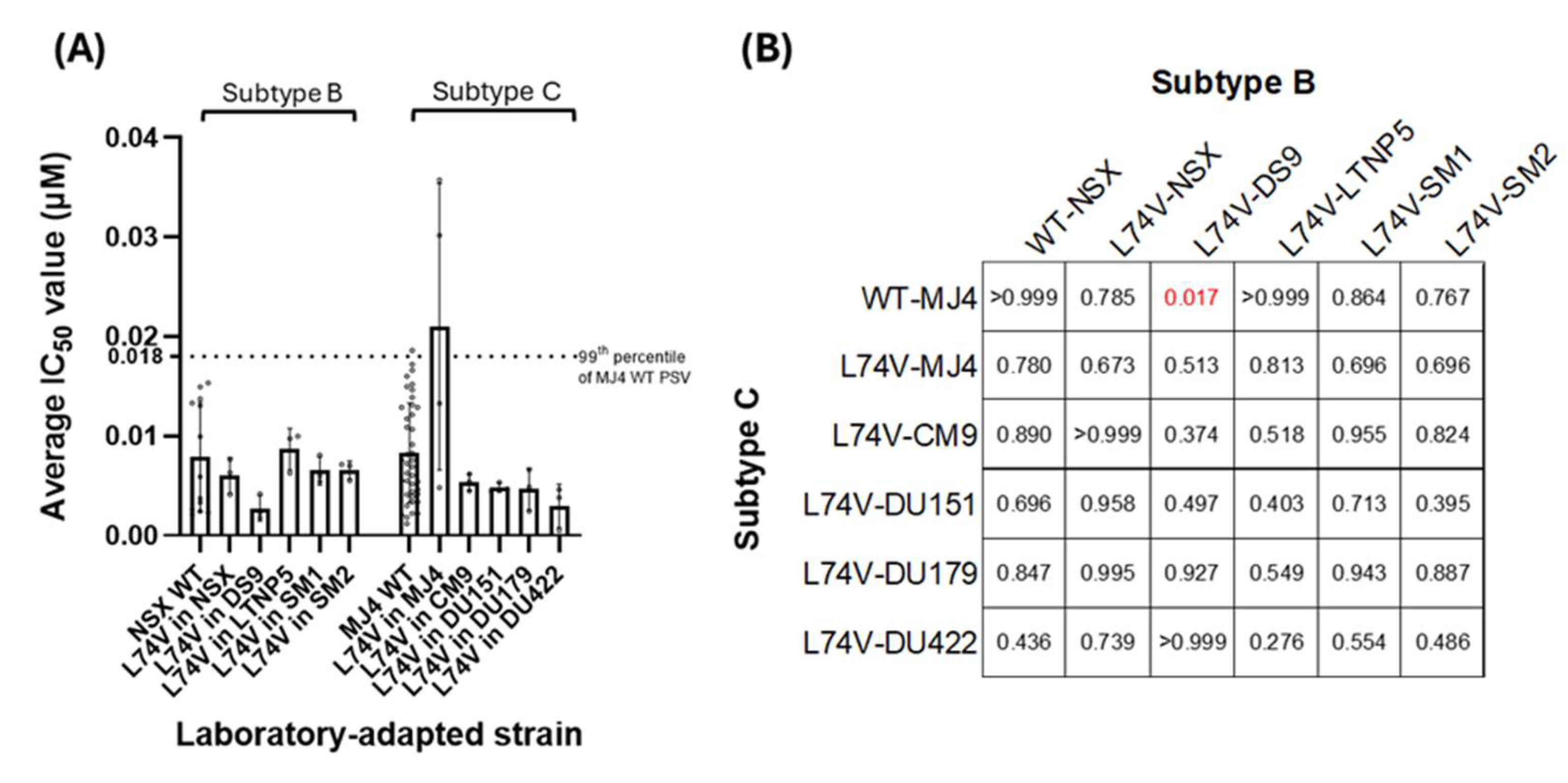
Figure 5.
Fold change in IC50 of mutation combinations in subtype C PSVs compared to the wild-type MJ4 PSV. Following the in vitro phenotypic activity assays, the IC50 value for each mutant was compared against the IC50 of the MJ4 wild-type PSV, allowing for the determination of fold changes. The 99th percentile of variation in the wild-type IC50 value, calculated to be 2.2 (TCO), served as the threshold for categorizing mutants as either susceptible or resistant to ISL. It was observed that the combination of NRTI mutations generally increased resistance to ISL. The A114S/M184V mutation combination showed a very high level of resistance to ISL. Its IC50 value was greater than the highest ISL concentration tested, and consequently, its FC value was > 60. Susceptible (■), potential low-level resistance (■), low-level resistance (■), intermediate resistance (■), high-level resistance (■).
Figure 5.
Fold change in IC50 of mutation combinations in subtype C PSVs compared to the wild-type MJ4 PSV. Following the in vitro phenotypic activity assays, the IC50 value for each mutant was compared against the IC50 of the MJ4 wild-type PSV, allowing for the determination of fold changes. The 99th percentile of variation in the wild-type IC50 value, calculated to be 2.2 (TCO), served as the threshold for categorizing mutants as either susceptible or resistant to ISL. It was observed that the combination of NRTI mutations generally increased resistance to ISL. The A114S/M184V mutation combination showed a very high level of resistance to ISL. Its IC50 value was greater than the highest ISL concentration tested, and consequently, its FC value was > 60. Susceptible (■), potential low-level resistance (■), low-level resistance (■), intermediate resistance (■), high-level resistance (■).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



