Submitted:
21 October 2024
Posted:
23 October 2024
You are already at the latest version
Abstract
Microplastics (MP), defined as plastic particles less than 5 mm, pose a significant global environmental threat, particularly in aquatic ecosystems, due to their persistence and potential harmful effects on wildlife and human health. They can absorb persistent organic pollutants (POP) like polycyclic aromatic hydrocarbons (PAH) and polychlorinated biphenyls (PCB), raising concerns about their impact on biota. To elucidate this, the present study employed attenuated total reflectance Fourier transform infrared spectroscopy (ATR-FTIR) to analyse the characteristics of MP sourced from commercial cosmetics. It also utilised nematodes, which are widely accepted indicators of environmental change. Indeed, despite the known risks of MP in aquatic habitats, knowledge of their effects on free-living and beneficial nematodes is limited. This study investigated the toxicity of MP on Caenorhabditis elegans and two entomopathogenic nematode species, Steinernema feltiae (the enviroCORE strain SB12[1]) and Steinernema carpocapsae (a commercial strain from e-NEMA) in laboratory bioassays. Nematodes were exposed to various concentrations of MP and other pollutants in controlled settings over 72 to 96 hours. Additionally, high-throughput 18S rDNA sequencing was used to analyse nematode biodiversity in sediments from the River Barrow (RB) in SE Ireland. Results revealed that MP adversely affected nematode survival and community structure, highlighting the ecological risks posed by MP pollution in river sediments and emphasizing the need for further research into the health of benthic ecosystems in Ireland.
Keywords:
microplastics
; entomopathogenic nematodes
; Caenorhabditis elegans
; polycyclic aromatic hydrocarbons
; pesticides
; ATR-FTIR
1. Introduction
Microplastics are becoming persistent pollutants that are of growing concern [1], due to their potential toxicity to aquatic biota and human health, as well as their association with pathogenic bacteria, and affects on the food web [2,3]. Microplastics have become ubiquitous worldwide, in the ocean, on land, and even in polar glaciers where human activity is minimal, and are regarded as a rising threat to ecological and environmental sustainability [4]. Occurrence of MP in water is an emerging worldwide environmental issue, with an estimated annual release of approximately 0.7 to 1.8 million tonnes of MP into the environment being reported in Europe [5]. Microplastic particles have been reported in seafood, processed food, and beverages [6]. The large surface area and small volume (particle size is usually less than 5 mm) of MP can lead to the adsorption and transfer of various environmental pollutants such as PAH, pesticides, polybrominated diphenyl ethers, pharmaceutical and personal care products, and heavy metals including chromium (CrVI), nickel (NiII), copper (CuII), zinc (Zn), cadmium (Cd), lead (Pb), and titanium (Ti), from environmental media to aquatic food webs. Furthermore, pollutants carried in MP may have an impact on aquatic organisms when discharged into the aquatic environment [7,11]. Plasticizers have been identified within MP, which also demonstrate a significant ability to adsorb toxic chemical compounds, including polycyclic aromatic hydrocarbons (PAH) and dichlorodiphenyltrichloroethane (DDT). These substances can be transferred to organisms and impact human health through the food chain [12]. In the past few years, research on MP in aquatic environments has received considerable attention, with aquatic MP pollution being listed as one of the top ten environmental challenges to be solved globally, partly due to the negative impact of MP particles on a range of aquatic organisms, such as phytoplankton, zooplankton, and fish [4,13]. To date, MP have been found in honey, sugar, beer, milk, table salt, bottled water, tap water, seafood, and rice, among other foods and beverages [14,18].
Microplastics contaminate a wide range of environmental compartments, which includes oceans, rivers, lakes, soils, and atmosphere, as well as sources of drinking water [19,20]. Furthermore, rivers play a fundamental role in the transportation of plastic particles into lakes, seas, and oceans [21]. Microplastics have the potential to affect soil fauna via a variety of mechanisms, including ingestion by soil living organisms, release of toxic plastic additives, and serving as vectors for transport of pathogenic bacteria [20,22]. Additionally, pesticides have the potential to be transported by MP, which accumulate in soil or sediment and serve as a pesticide sink [23,24]. Moreover, freshwater sediments act as a sink for MP and source for toxic chemicals, and as a result, benthic organisms can be exposed to possibly significant amounts of MP [25,26]. Small and lightweight MP accumulate downstream in rivers, however MP are also often detected in riverbed sediment, showing persistence over time [27].
Polycyclic aromatic hydrocarbons (PAH) are a large class of aromatic compounds produced through inadequate combustion or pyrolysis activities, such as automobile exhaust, cigarette smoking, industrial processes, consumption of grilled and smoked food, fossil fuel, and forest fires. Consequently, PAH have been found in high quantities in air, water, and soil [28,29]. Pesticides are chemical mixtures that are used to prevent, remove, repel, or neutralize organisms that cause issues in agriculture, such as microorganisms (bacteria, fungi, and viruses), insects, animals, and weeds [30]. Pesticides have adverse implications in ecosystems that vary depending on the pollutant concentration, quantity, and exposure time. Pesticides pose a significant hazard due to their persistence in the environment and tendency to accumulate in organisms [30], leading to contamination of water and soil, even when used at low rates, and with poor biodegradability [31]. For example, 1,3- dichloropropene is frequently used because of its effectiveness in controlling pathogenic nematodes and weeds [32]. Herbicides are synthetic chemicals that aid in the management and elimination of unwanted weeds [30]. atrazine is a commonly used, highly effective, and low-cost pesticide for controlling broadleaf and grassy weeds. It is a major pollutant of soil and aquatic systems, and it has been shown to disrupt aquatic flora and animal reproduction, which can have a negative impact on community structure [33,34].
Nematodes are the most abundant and diverse group of metazoans found in sediment and soil, and they play a critical role in the food webs of benthic and soil environments [35]. Nematodes are non-segmented roundworms that live in almost every ecosystem on the planet and have considerable potential as environmental indicators. They are used as bioindicators in assessments of anthropogenic pollution because of their ecological significance, widespread prevalence, and exceptionally high individual densities [36]. Nematodes have been used to study gene expression in relation to environmental challenges, as well as in laboratory ecotoxicity assays, and in situ monitoring of the ecological impact of a variety of enviromental problems [36,37].
Entomopathogenic nematodes (EPN) from the families Steinernematidae (genus Steinernema) and Heterorhabditidae (genus Heterorhabditis) of the Phylum Nematoda are soil-dwelling, lethal insect parasites which can be an alternative to chemical insecticides, and are easily mass produced in vivo and in vitro [38,39]. Their symbiotic bacteria are released into their insect host hemocoel by nematode free-living infective juveniles (IJ), which penetrate insect hosts through natural body openings and pores in the cuticle. The subsequent resulting bacterial septicemia kills the host quickly, and its cadaver is changed to a nutritious broth for the nematodes, mediated by the nematode symbiotic bacteria. By creating bacteriocins, antimicrobials, and other antibiotics, the bacteria protect the cadaver resource from rivals. Infective juveniles feed on the bacterial broth andmature into adults, which can give rise to multiple generations until resources are depleted. Then, new IJ emerge from the cadaver and disperse in the soil, looking for new hosts to infect [40,42].
Caenorhabditis elegans is a model organism that has been widely used to study genotoxic reactions to environmental contaminants from molecular to organismal levels [43,44]. Because of its small size, rapid life cycle (3-4 days), short average lifetime (2-3 weeks), and ease of rearing, this transparent, free-living nematode offers numerous benefits for genetic investigations [45,46]. Caenorhabditis elegans has shown a low-dose response to several chemicals after being used in assessing the toxicity of a range of environmental pollutants, including zinc oxide (ZnO) nanoparticles, which are commonly found in toothpaste, beauty products, sunscreens, and textiles, as well as bisphenol A (BPA), which is widely used in the manufacture of plastic products [46,48]. Brenner (1974) emphasized the advantages of biological testing with C. elegans. Caenorhabditis elegans displays rapid growth on agar plates or in liquid culture at temperatures ranging from 15 to 25°C, using Escherichia coli as a food source. Its fast growth is evidenced by a half-day doubling time and a generation period lasting only 3 to 5 days. Additionally, advantages of using C. elegans include the possibility of self- or cross-fertilization [45]. Caenorhabditis elegans is one of the species that has been used to study the toxicity of MP [19,49]. In a number of previous studies, MP have been shown to have negative physical and biological impacts on C. elegans [19,44,46,47,48,49,50].
However, there is limited research on the impact of MP on other nematode indicators, such as EPN, apart from the present study involving testing C. elegans and EPN as sentinels of MP pollution. For assessing sediment quality, nematodes have been used as an effective alternative to the existing conventional methods for this task, such as chemical analyses [25]. Both morphological and molecular (DNA-based) methods have been employed in the identification of nematodes in order to facilitate an effective assessment of biodiversity [51,52]. In the 1970s, Zullini (1976) studied the relationship between nematode community structure and sediment pollution (anthropogenic contamination with heavy metals and organic pollutants) and discovered that nematode community structure of river sediments was related to pollution and site structure [53,54].
Hence, the aims of the current study were (1) to improve understanding of MP properties and their interactions with organic pollutants, (2) to determine the toxicity effects of MP particles using three nematode species (C. elegans, Steinernema feltiae (SB12(1) and Steinernema carpocapsae) as sentinel organisms, and (3) to observe possible effects of MP on nematode communities from Irish river sediments via high-throughput sequencing of nematode 18S rDNA for taxa identification.
2. Materials and Methods
2.1. Effects of Contaminated MP on Nematodes
2.1.1. Microplastics’ Samples Preparation
Isolation of polyethylene MP from a commercial exfoliating product was adapted from Napper et al., (2015) and Lei et al., (2017) by selecting two most commonly used commercial facial scrubs, including (i) Clean And Clear Exfoliating Daily Wash, and (ii) Clean And Clear Blackhead Clearing Daily Scrub [55,56]. Each product content was diluted to approximately 1L of boiling water to extract MP for further analysis. The mixture was then subjected to vacuum filtration using a suction pump attached to a buchner funnel with Whatman filter paper N°4. Filter papers retaining MP were dried at room temperature overnight and then dried in an oven at 60°C for 5 minutes. Once dried, the particles were transferred into labelled 1.5 cm centrifuge tubes. The recovered MP were transferred on slides analysed under a stereoscope (Optika microscopes) at a magnification of 40X. The total magnification was calculated by multiplying the objective lens magnification (4) by the ocular lens magnification (10), resulting in a total magnification of X40. Furthermore, MP particles were characterised using attenuated total reflectance Fourier transform infrared (ATR-FTIR) spectroscopy. Spectra were collected from 4000 cm-1 to 500 cm-1 using a Perkin Elmer Spectrum 65 FT-IR Spectrometer. A background scan was performed between each sample after cleaning the ATR diamond crystal with 70% 2-propanol. Absorption bands were recorded and compared to the reference spectra. Commercial clear polyethylene (PE) particles 1000μm (http://www.cospheric.com/) were used as reference to identify the recovered MP.
2.1.2. Nematode Cultures
Caenorhabditis elegans (sourced from cgc.umn.edu) and two species of entomopathogenic nematodes (EPN), including Steinernema feltiae (SB12(1) enviroCORE strain) and Steinernema carpocapsae (a commercial e-NEMA strain) were chosen as sentinel organisms to evaluate MP toxicity. Rearing techniques of EPN and C. elegans were adapted from [45,57,58,59].
2.1.2.1. Culturing and Maintenance of Caenorhabditis elegans
Nematode growth medium (NGM) was used to grow C. elegans (see Table 1; [45]). To make 1 L nematode growth (NG) agar, 3 g sodium chloride (NaCl), 17 g bacteriological agar (Nº3) and 2.5 g bacteriological peptone were placed into a 1 L Duran bottle holding 975 mL deionised water. The mixture was autoclaved for 20 minutes at 121°C. The mixture was allowed to cool to 60 °C. The NGM Duran bottle was filled with 25 mL KH2PO4 pH 6.0, 1 mL of 1 M MgSO4, 1 mL of 1 M CaCl2 and 1 mL of 5 mg/mL cholesterol in ethanol. The solution of NGM was then aseptically poured into Petri plates. Prior to use, the plates were left to dry at room temperature for 2-3 days in a laminar airflow, for detection of any contaminants.
2.1.2.2. Seeding of NGM Plates
Escherichia coli OP50 (E. coli OP50) (supplied with the nematode from the Caenorhabditis Genetics Center, University of Minnesota) was used as the food source for the growth of C. elegans. Aseptically, approximately 20 µL of E. coli OP50 liquid culture was added to 50 mL Luria broth (LB) and incubated at 37 °C overnight in a shaking incubator. Escherichia coli OP50 was aseptically seeded onto NG plates in a laminar flow. After allowing the plates to dry, square-shaped piece of agar containing C. elegans were placed in the centre of the seeded plates (Figure 1). The plates were sealed with parafilm (to prevent the agar from drying out) and incubated at 21°C for nematode growth before being used as working stocks for toxicity tests and other experiments. The plates were checked after 24 hours to ensure that there were viable growing nematodes.
2.1.2.2.1. Buffer preparation
To prepare a 500 mL potassium phosphate buffer (KH2PO4/K2HPO4), 34 g KH2PO4 were placed in a 250 mL Duran bottle containing 250 mL deionised water. Subsequently, 45.6 g K2HPO4 were placed in a separate 250 mL Duran bottle with 200 mL deionised water. To raise the pH from 4.0 to 6.0, 60 mL K2HPO4 solution was added to KH2PO4. After the liquid reached pH 6.0, filter sterlisation was applied to remove any particles.
2.1.2.2.2. Chemical preparation
To make 1 M calcium chloride (CaCl2), 5.545 g (CaCl2) were added in a 50 mL Duran bottle holding 50 mL deionised water. To make 1 M magnesium sulphate (MgSO4), 6.015 g (MgSO4) were transferred in a new 50 mL Duran bottle containing 50 mL deionised water. These solutions were then autoclaved. Afterward, 50 mg cholesterol were transferred to a 50 mL Duran bottle with the addition of 95% ethanol to make 5 mg/mL cholesterol in ethanol. This mixture was not autoclaved.
2.1.2.2.3. Culturing and maintenance of entomopathogenic nematodes
Two species of EPN, Steinernema feltiae (the SB12(1) enviroCORE strain) and Steinernema carpocapsae (a commercial e-NEMA strain) were cultured, maintained, and kept at 9 ᵒC in the laboratory. Galleria mellonella was sourced commercially from Live Foods Direct (Sheffield, UK) and was used as a host to culture EPN. Two sheets of Whatman filter paper were used to line the lids of standard size Petri dishes with a diameter of 100 mm and a height of 15 mm. The filter paper was moistened with approximately 1-1.5 mL of a dense IJ suspension. Following this, five late instar larvae of G. mellonella were carefully placed on the filter paper and the base of the Petri dish was added as a lid to close the dish. To keep the filter paper moist, the dish was loosely wrapped with parafilm. The plates were subsequently incubated for 3-7 days in darkness at a temperature of 21ᵒC until mortality of G. mellonella occurred. Nematode IJ were recovered using the White trap method [60] after insect mortality had occurred. An inverted Petri dish (60 X 15 mm) was placed in a clear comtainer (150 X 30 mm). Filter paper was used to cover the dish and create a raised platform. Enough sterile deionised water was then placed in the clear container to completely submerge the filter paper's edges. The infected insects were placed on the platform and incubated for up to 14 days at room temperature. Nematodes were harvested after emergence by decanting the water into an empty clear dish. The infected host's transparent dish was refilled with sterile deionised water and incubated for another 1-3 days. Incubating for an additional 1-3 days ensures maximal emergence of nematodes from infected insects, thereby enhancing experimental yield. Nematodes were then maintained at 9°C until needed, with no more than two weeks between emergence and experimentation (Figure 2).
2.1.3. Caenorhabditis elegans Age Synchronisation Using Alkaline Hypochlorite Solution (Bleaching)
The bleaching technique is used for age synchronising C. elegans cultures at the first larval stage (L1). Age synchronising in the rearing of C. elegans ensures that all the nematodes in a culture reach L1 stage at the same time, promoting consistency in experimental timing and comparisons. Caenorhabditis elegans can be synchronized by isolating newly hatched larvae or by treating gravid adults with bleach (by killing everything but embryos) and isolating eggs, which are resistant to bleach treatment. After treatment with alkaline hypochlorite solution (Table 2), embryos are incubated in liquid media without food, which allows hatching but prevents further development [57,58,59,60].
To age synchronise C. elegans the nematodes were cultured on NGM plates until adult stage. The nematodes were allowed to grow two to five days to recover enough embryos. Therefore, seeded NGM plates were washed out to recover all the nematodes using Ringer’s solution. The nematodes in the Ringer’s solution were transferred into 15 mL Falcon conical centrifuge tubes and centrifuged for 2 minutes at 1500 rpm, at room temperature. The supernatant was discarded, and this step was repeated three times to ensure that most of the worms were transferred in the Ringer' s solution. Most of the Ringer’s solution was aspirated without disturbing the nematode pellet. Subsequently, 15 mL of the 20% alkaline hypochlorite solution was added to each tube. The tubes were gently inverted and agitated for five minutes and centrifuged for 1 minute at 1500 rpm. Following that, 20% alkaline hypochlorite solution was aspirated without disturbing the nematode pellet. About 15 mL Ringer’s solution was added to the tube. The tube was furthermore centrifuged for 1 minute and this step was repeated three to four times since the treatment may still be active. The embryos were transferred onto new unseeded NGM plates (without food - E. coli OP50) to allow the eggs to hatch (Figure 3). The culture plates were incubated for 3 days prior MP toxicity tests.
2.1.4. Toxicity Bioassay of EPNs and C. elegans Using Microplastic Pollutants
Nematodes suspensions were transferred in sterile Falcon tubes to obtain the required concentration for the assays. The Falcon tubes were left upright for 30 minutes to ensure complete settlement of nematodes at the bottom of the tubes. The supernatant was slowly decanted using a sterile glass pipette to obtain approximately 10 IJ/100 µl suspension. Subsequently, 96 well plates were used to conduct the toxicity assays using a fresh batch of EPNs and C. elegans. To investigate the effects of MP on EPN and C. elegans, each well contained 100 µl of sterilised deionised water with approximately 10 juveniles of EPN and C. elegans at L1 stage, and approximately 0.01 g MP particles. The particles had been treated with Ringer’s solution, which was used as an untreated control, and atrazine (100 ppb), 1,3 – dichloropropene (100 ppb), naphthalene (100ppb), and fluorene (100 ppb), for 12 months prior to toxicity bioassays. Afterward, 100 µl of PAH and the pesticides mentioned above were added in each well (see Table 3). Nematodes were then counted under a stereo microscope (Optica SZM-2) at magnifications ranging from 7X (minimum) to 45X (maximum). The total magnification was calculated by multiplying the objective lens magnification (0.7) and (4.5) by the ocular lens magnification (10). The nematode mortality was assessed after 72 and 96 hours exposure (Figure 4).
2.1.5. Statistical Analysis
ANOVA was carried out to identify significant effects on nematode mortality by the induced MP treatments. The statistical analysis was conducted using Microsoft Office Excel. The level of significance was set at P ≤ 0.05.
2.2. Effects of MP on Nematode Communities in Irish River Sediments
2.2.1. Sediment Sampling and Processing
River sediment samples were collected in triplicate from four locations along the RB from a 10 cm depth using a Dutch auger (Table 4). The selected sites were chosen for analysis due to the presence of MP contamination as reported in prior research conducted by Murphy et al., (2022). The collected samples were stored at -20 ᵒC prior to analysis. Subsequently, 25 g sediment sub-samples were transferred in sterile Falcon tubes containing 25 mL deionised water and centrifuged for 2 minutes at 3500 rpm. The supernatant was discarded, and the remaining materials were placed in Petri dishes and dried overnight at room temperature. Furthermore, the dried sediment soil was thoroughly homogenized using a mortar and pestle. Afterward, total DNA was extracted from 0.25 g of the dried sediment sub-samples using the Qiagen DNeasy® PowerSoil® Pro kit following the manufacturer’s protocols. Prior to exporting to a sequencing company (Novogene, UK), the concentration and quality of the DNA were evaluated using QubitTM 4 Fluorometer InvitrogenTM (Thermo Fisher Scientific). DNA purity was then assessed using 1.5% agarose gels (Figure 8). The DNA samples were stored at -80 ᵒC until being sent for sequencing. The primer pair MN18F and 22R [62] was used on the Illumina paired-end platform, and the nematode 18S V4V5 rRNA region was sequenced.
2.2.2. 18S rDNA Gene Amplification and DNA Sequencing
The sediment DNA samples were sent to Novogene Ltd. (Beijing) for subsequent Polymerase Chain Reaction (PCR), library preparation, and DNA sequencing. Specifically, a PCR primer set MN18F (forward) (5' - CGCGAATRGCTCATTACAACAGC - 3') and 22R (reverse) (5' - GCCTGCTGCCTTCCTTGGA - 3'), adapted from [62], was provided by the author to the company and used to amplify the V4V5 region of the 18S gene in nematodes. Following PCR amplification, libraries were produced by Novogene using the NEBNext® UltraTM DNA Library Prep Kit for Illumina, following the manufacturer's guidelines, and indexed. Library quantification was performed by the company using Qubit and qPCR, and sequencing was done on the Illumina platform, generating 250 bp paired-end reads.
2.2.3. Analysis of the DNA Sequencing Data
The author conducted the extraction of sediment DNA samples, which were subsequently submitted to Novogene for sequencing. Novogene provided the DNA sequence data, as well as diversity indices. The author further worked on the diversity indices, OTU table, and heatmap in Microsoft Excel using the data provided by Novogene for statistical analysis purposes. The alpha diversity indices of nematodes in river sediment, including observed operational taxonomic units (OTU) number, Chao1, Simpson, and Shannon values, were analysed by the author using the QIIME software on Illumina MiSeq sequencing data by the sequencing company Novogene Ltd. (Beijing). The paired-end reads were assigned to samples based on their unique barcodes and truncated by cutting off the barcode and primer sequences. For Alpha diversity, the number of OTU was used to evaluate the diversity of the river sediment nematode communities. Paired-end reads were merged using the fast and accurate tool FLASH (V1.2.7), specifically designed to combine reads with overlapping sequences into raw tags. Quality filtering on the raw tags were performed under specific filtering conditions to obtain the high-quality clean tags according to the Qiime (V1.7.0) quality-controlled process. Sequence analysis was performed by Uparse software (Uparse v7.0.1090) using all the effective tags and clustered into operational taxonomic units (OTU) at ≥ 97% similarity, in order to analyse the species diversity in each sample. For each representative sequence, Qiime (Version 1.7.0) in Mothur method was used to perform the assignment of taxonomy against the SSU rRNA database of SILVA138 database. The species annotation was done at each taxonomic rank (kingdom, phylum, class, order, family, genus, species) with a threshold of 0.8~1. A principal component analysis (PCA) was performed to investigate the clustering of samples based on OTU. The more similar the composition of communities among the samples are, the closer the distance of their corresponding data points on the PCA graph are. Analysis of Similarity (Anosim) and permutational MANOVA (ADONIS) in R software were also used to evaluate the significance in nematode communities between sediment samples from the four selected locations. Heatmaps were generated using the FactoMineR package in R software. Boxplots were formed to analyze the difference of Alpha and Beta diversity indices across nematode community groups from the selected locations. T-test, wilcox and Tukey tests were used to look at the significance of MP impacts on nematode communities.
The maturity indices (MI) based on the nematode colonizer-persister (c-p) scale [63] was calculated to assess the health of river sediments by submitting the sediment nematode data obtained from18S rDNA sequencing to NINJA (Nematode INdicator Joint Analysis; [64], provided at https://shiny.wur.nl/ninja/.
3. Results
3.1. Effects of Contaminated MP on Nematodes
3.1.1. Interactions Between Microplastics and Organic Pollutants
In this study, product 1, Clean And Clear Exfoliating Daily Wash, was selected for the toxicity bioassay due to the size of its MP (Figure 5), which ranged from approximately 29 to 300 µm, compared to the MP of product 2, measuring beween 51.5 µm to 1 mm. Prior to toxicity assessments, the extracted MP were immersed in various treatments for 12 months, including Ringer’s solution (untreated control), atrazine (100 ppb), 1,3 – dichloropropene (100 ppb), naphthalene (100 ppb), and fluorene (100 ppb).
The absorption of contaminants by MP primarily depends on factors such as particle size hydrophobicity [65,66]. Figure 6a and Table 5 present a comparison between the absorption bands obtained from the recorded data and those found in polymer reference spectra. ATR-FTIR analysis of MPs exposed to atrazine for a 12 month period showed clear evidence of atrazine being retained on the MP surface. Figure 6b shows that, in addition to the 4 peaks associated with the original MP (Figure 6a), 12 additional peaks appear that correlate with peaks on the atrazine reference spectrum (Figure 6c). These peaks are at 3300 cm-1, 3150 cm-1, 1618 cm-1, 1544 cm-1, 1401 cm-1, 1343 cm-1, 1304 cm-1,1264 cm-1, 1165 cm-1, 1128 cm-1, 1056 cm-1, 991 cm-1, 809 cm-1, and 803 cm-1 . For MP exposed to 1,3-dichloropropene (Figure 6d,e) and naphthalene (Figure 6f,g) the same type of correlation was not observed, however a number of additional peaks appear on MP spectra after exposure (Figure 6d,f) which are not present for the original MP (Figure 6a). The additional peaks that appear in Figure 6d,f do not appear in the spectra for MP exposed to atrazine or fluorene so they cannot be ascribed to degradation of the polymer. This suggests that there may be some interaction between the MPs and these substances, but this interaction is not clear, and dichloropropene and naphthalene do not seem to be retained on the MP to the same extent as atrazine.
3.1.2. Toxicity Bioassays of MP on Nematodes
The results were recorded as mean percentage mortality of nematodes in each treatment. Investigating nematode exposure to MP particles in different treatments has demonstrated that chemicals adhere to MP and are subsequently transferred into nematodes. Acute exposure to contaminated MP particles resulted in a time and dose dependent increase in nematodes mortality. All three nematode species were readily affected by MP treatments, however the mean mortality percentange was significantly different among all the treatments (Figure 7). After a 72 hours exposure, results showed high mortality in S. carpocapsae IJ and C. elegans J3 in all the treatments containing MP, compared to mortality in S. feltiae SB 12(1) IJ. However, S. carpocapsae IJ were the most sensitive to all the treatments during both 72 and 96-hour time exposures. In addition, S. carpocapsae IJ were significantly affected by MP treated with all the treatments compared to S. feltiae SB 12(1) IJ and C. elegans J3 (P < 0.05) (Figure 7). Steinernema feltiae SB 12(1) IJ were the least affected in all the treatments. There was only a significant difference between the mortality of S. feltiae SB 12(1) IJ exposed to the combination treatments of atrazine with no MP and the mortality in the treatment with MP (P < 0.05), which confirms that MP particles are causing some impacts on nematodes. Mortality of the three nematode species was significantly higher in the combination treatments of 1,3 – dichloropropene with no MP and with MP (F = 2.861, P = 0.002724) in comparison to the treatments with atrazine, naphthalene, and fluorene.
A single factor one-way ANOVA analysis of the data showed significant mortality variation among the three nematode species. The analysis of variance (ANOVA) was conducted on the three nematode species and various treatments, with results presented in Table 6. Notably, Table 6 presents a significant effect of increasing exposure time to naphthalene on the three nematode species. Specifically, following a 96-hour exposure to naphthalene-treated MP, there was a statistically significant difference in nematode mortality (one-way ANOVA: F = 1.954, P = 0.008807).
When comparing the mortality rates of S. carpocapsae IJ, C. elegans J3, and S. feltiae SB 12(1) IJ to those of the untreated control (nematode medium with MP), it was observed that the mortality rates were 26%, 21%, and 23%, respectively. The mortality of S. carpocapsae IJ and C. elegans J3 increased significantly to 60%, and 41%, respectively, specifically in MP treated with 1,3–dichloropropene, at 96 hours exposure. However, S. feltiae SB 12(1) IJ mortality was lower than that of S. carpocapsae IJ and for C. elegans J3 reaching 38%. Furthermore, the mortality rates of S. carpocapsae IJ, C. elegans J3 and S. feltiae SB 12(1) IJ, after 96 hours of exposure to MP treated with atrazine, were 46%, 41% and 35%, respectively, compared to 32%, 29% and 25% respectively, exposed to MP that were not treated (control). The mortality rates of S. carpocapsae IJ, C. elegans J3 and S. feltiae SB 12(1) IJ were also affected in MP treated with naphthalene at 96 hours, reaching 40%, 35% and 30%, respectively, compared to the control treatment containing MP and showing mortality of 34%, 27% and 23%, respectively. Moreover, the mortality rates of S. carpocapsae IJ, C. elegans J3, and S. feltiae SB 12(1) IJ were impacted by exposure to MP treated with fluorene, resulting in rates of 41%, 37%, and 33% respectively. These rates were notably higher than the control rates of 30%, 26%, and 22%, respectively.
Significant differences in mortality of all species were observed among certain treatments containing MP. For example, after a 96-hour exposure, S. carpocapsae IJ mortalities caused by atrazine and 1,3-dichloropropene were significantly (P < 0.05) higher compared to those caused by naphthalene and fluorene in the same nematode. Overall, there was a significant impact of MP on the three nematode species among the treatments. Microplastics treated with 1,3 – dichloropropene were found to be the most toxic treatment in this bioassay as most nematodes were affected compared to the rest of treatments (Figure 7a-h).
3.2. Characteristics of DNA Sequences from Sediment Nematodes
The electrophoresis of sediment extracted DNA samples are illustrated in Figure 8. After Novogene Ltd. (Beijing) optimised the sequencing results of sediment nematode DNA fragments on the Illumina paired-end platform, effective DNA sequences encoding vital biological functions within an organism were generated from 20 sediment samples. A total of 2,499,108 clean reads, defined as high-quality sequencing reads that are free from errors, artifacts, or contamination, were obtained through analysis provided by the sequencing company Novogene Ltd. (Beijing). The mean number of reads obtained for DHC samples was 119,975 ± 4251.099, for GI samples was 125,182 ± 4426.391, for MCR samples was 129,313 ± 5852.505, and for MLF samples was 125,352 ± 3313.495.
Figure 8.
(a) Agarose (1.5%) gel electrophoresis of river sediment DNA at 100 volts for 40 minutes. Note: M1 indicates Trans 15K plus marker, lane 1 to 20 represent samples; (b) PCR products gel electrophoresis of samples processed by Novogene Ltd. (Beijing) sequencing company. Note: M1 indicates Trans 100bp ladder. PCR amplicons amplified in different samples; lane 1-5: DHC samples, lane 6-10: MLF samples, lane 11-15: MCR sample; and lane 16-20: and lane 11-20: GI samples.
Figure 8.
(a) Agarose (1.5%) gel electrophoresis of river sediment DNA at 100 volts for 40 minutes. Note: M1 indicates Trans 15K plus marker, lane 1 to 20 represent samples; (b) PCR products gel electrophoresis of samples processed by Novogene Ltd. (Beijing) sequencing company. Note: M1 indicates Trans 100bp ladder. PCR amplicons amplified in different samples; lane 1-5: DHC samples, lane 6-10: MLF samples, lane 11-15: MCR sample; and lane 16-20: and lane 11-20: GI samples.
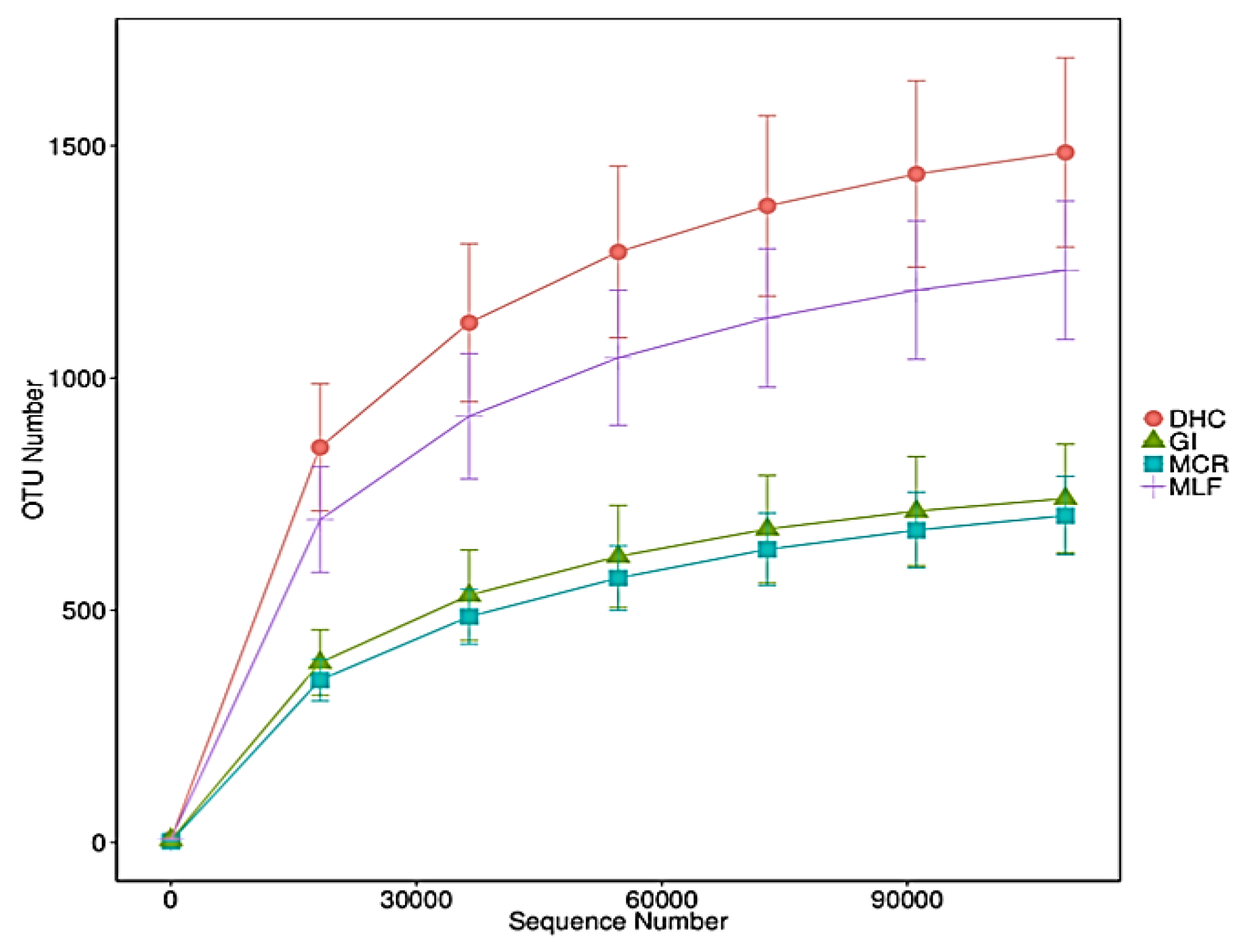
Figure 9.
The Rarefaction curve of the number of operational taxonomic units (OTUs) of a sediment nematode community. Operational taxonomic units were delineated at 97% sequence similarity: Dolmen Hotel Carlow, GI: Great Island, MCR: Macmurroughs, MLF: Milford. The X-axis represents the sequence number, while the Y-axis depicts the observed number of OTUs.
Figure 9.
The Rarefaction curve of the number of operational taxonomic units (OTUs) of a sediment nematode community. Operational taxonomic units were delineated at 97% sequence similarity: Dolmen Hotel Carlow, GI: Great Island, MCR: Macmurroughs, MLF: Milford. The X-axis represents the sequence number, while the Y-axis depicts the observed number of OTUs.
3.2.1. Sediment Nematode Community Composition
At the class level, the relative richness of Enoplea was 32% and of Chromadorea 68%. However, Enoplea richness was only higher than that of Chromadorea at the DHC location. In general, Chromadorea was the dominant class accounting for 22.60%, 35.70%, 55.10%, 46.10%, 23.00%, 53.80%, 24.50%, 37.30%, 89.40%, 80.40%, 57.10%, 97.00%, 90.30%, 91.50%, 82.80%, 89.90%, 98.10%, 92.50%, 98.60%, and 94.60% of the nematode community at DHC1, DHC2, DHC3, DHC4, DHC5, MLF1, MLF2, MLF3, MLF4, MLF5, MCR1, MCR2, MCR3, MCR4, MCR5, GI1, GI2, GI3, GI4, and GI5, respectively (Figure 10). At the order level, Chromadorida (21.4%), Desmodorida (6.3%), Monhysterida (36.5%), Dorylaimida (14.6%), and Triplonchida (13.7%) were the dominant orders, accounting for 92.4% of the total nematode sequences obtained (Figure 10a). Of the nine most frequently occuring nematode orders, significant differences were found in the abundances of Monhysterida, Dorylaimida, and Chromadorida compared to Triplonchida and Desmodorida (Figure 10b).
Relative abundance of the 10 most prevalent nematode genera in the RB sediment samples obtained by 18S rDNA gene sequencing were summarized in Figure 11a. The dominant genera group in the river sediment nematode community of DHC was Triplonchida and Mermithida, both with a relative abundance of 28%, followed by Monhysterida (21%). Chromadorida was less abundant in the MCR (9%), but mostly dominated in the MLF (28%), followed by Monhysterida (25%). Desmodorida was mainly identified in the MCR (6%) and GI 17%. Araeolaimida was not present in the other two locations, with lower abundance observed in the MLF (2%) and MCR (11%). However, the main nematode group in the sediment nematode communities of MCR and GI were Monhysterida, with a relative abundance of 57% and 44%. On the other hand, Chromadorida was the most dominant in the GI (34%) and MLF (28%) samples, followed by Dorylaimida which was in abundance in the MLF (20%) and DHC (34%) samples. Desmodorida was in a higher abundance in the GI (17%) samples, and a lower number was observed in the DHC (1%) samples. Monhysterida and Araeolaimida were the most dominant nematode orders in the MCR compared to the rest of the locations (Figure 11a). The heatmap shows that the orders Monhysterida and Araeolaimida were the most dominant in the sediment samples from MCR compared to sediment samples from the other three locations, followed by Dorylaimida, Triplonchida, and Tylenchida which were in higher abundance in DHC and MLF samples (Figure 11b).
At the genus level, there was a high diversity of nematodes within the DHC, MLF, and MCR samples, and a much lower diversity was observed in the GI samples. Monhysterida OTU were identified in all the samples from the four locations representing an average of 21 ± 11%, 25 ± 12%, 57 ± 22%, and 44 ± 26% of the total genera from DHC, MLF, MCR, and GI samples, respectively (Figure 12a). In order to study the differences in nematode communities, the distribution of OTU was measured. The analysis revealed that 63 OTU were shared among all the samples. In addition, 16 unique OTU were found in GI, 21 in MCR, 27 in MLF, and 65 unique OTU only appeared in DHC location (Figure 12b).
3.2.2. Nematode Alpha and Beta Diversity
The alpha diversity of the nematode communities was determined using Chao1, Simpson, ACE, Goods coverage and Shannon’s diversity metrics (Table 7). Both observed OTU numbers and Chao1 showed a high level of species richness in the DHC location, followed by the MLF, and MCR samples. However, a low level of species richness was observed in the GI samples. Likewise, both Shannon and Simpson indices showed a high level of species richness and evenness in the MCR and DHC locations compared to the MLF and GI samples.
The impacts of MP were assessed on nematode composition at the OTU level, which represents the highest taxonomic resolution. Significant variations in nematode community diversity among the four locations were identified when comparing alpha diversity indices, specifically observed species, Chao1, Simpson’s diversity indices, and Shannon as illustrated in Figure 13a-d.
Beta diversity shown in Figure 14 also confirmed significant differences in nematode communities among the four locations. The two principal component axes explained 55.17% of the total variation in the nematode community, between which the PC1 axis explained 38.55% while the PC2 axis explained 16.62% of this variation (Figure 14a). Nematode communities from three locations (GI, MCR and MLF) were clustered together, separate from that in DHC. In addition, some DHC and MLF samples were clustered together. The nematode community structures in samples from each location were significantly different among them. There were significant differences as sediment samples from DHC, for example, were well dispersed away from each other, but two samples from this location were clustered with four samples from the MLF location. Principal component analysis (PCA) demonstrated samples separation at the OTU level sampled from the four locations. Furthermore, analysis of similarity (Anosim) and permutational MANOVA indicated that significant differences existed among the nematode communities in the twenty river sediment samples across the four locations (Figure 14 b to g), indicating that MP pollutants in the sediments have a significant impact on nematode communities.
3.2.3. Trophic Guilds and c-p Scale Association Across Locations
Nematode communities were significantly affected by the presence of MP particles in the river sediment. The occurrence of nematde trophic groups differed significantly among the four locations (P < 0.05). Coloniser-persister (c-p) classification is based on life cycle properties. Nematodes of c-p-1 are regarded as enrichment opportunists; they have short life cycles and are often found in disturbed environments. In contrast, nematodes of c-p-5 have long life cycles and tend to inhabit stable, mature ecosystems. Coloniser-persister of herbivores are called p-p [67]. In the present study, bacterivores were the most diverse and abundant trophic group across locations, whereas omnivores were least abundant. Plant parasites (p-p) were abundant across all the locations (see Table 8).
Sediment samples from the four locations presented a significant difference in nematode trophic group abundance. Sediment samples from DHC - site A (38.37%) had the highest abundance of herbivores followed by those from MLF - site B (23.97%). The lowest number of herbivores was observed in MCR - site C (13.29%) and GI - site D (4.9%) samples. The abundance of bacterivores was significantly (P = 0.01) higher in MCR - site C (70.47%) and GI - site D (44%), followed by DHC - site A (33.4%). The lowest number was observed in MLF - site B (27.3%). The abundance of predators was the highest in sediment samples from GI - site D (50.9%) and MLF - site B (30.8%), followed by DHC - site A (21.8%). The lowest number was observed in sediment samples from MCR - site C (15.1%). Moreover, omnivores are shown to decrease in abundance in line with the most polluted locations DHC - site A (6.5), MCR - site C (1.1) and GI - site D (0.1), compared to MLF - site B (17.9) which had the highest in abundance (Figure 15).
Nematodes were also classified into five c-p groups (1-5) according to their colonizer-persister life strategy. Throughout the locations, c-p2 nematodes contributed the major proportion. However, c-p4 nematodes were the least occurring and only observed in site A and B, but not in site C and D. Majority of nematode groups were classified under c-p2 and c-p3 categories, with c-p2 predominately represented. Sediment samples from MCR - site C (81.3%) and DHC - site A (55.4%) had the highest abundances of c-p2 nematodes, followed by GI - site D (46.3%). The lowest abundance was observed in samples from MLF - site B (35.6%). However, sediment samples from GI - site D (53.6%) and MLF - site B (40.4%) had the highest abundance of c-p3, followed by DHC - site A (35.3%). The lowest abundace was observed in samples from MCR - site C (17.4%). Lastly, sediment samples from MLF -site B (23.6%) had the highest abundance of c-p4 and , sediment samples from DHC - site A (10.5%) had the lowest. No c-p4 nematodes were observed in sediment samples from MCR - site C (0%) and GI - site D (0%) (Figure 16).
The maturity indices at site B for MLF (2.77 ± 0.51) exhibited a slightly greater value in comparison to site A for DHC (2.29 ± 0.44), site C for MCR (2.2 ± 0.16), and site D for GI (2.54 ± 0.27) as depicted in Figure 17. There was no significant difference in maturity indices among the four locations (P= 0.154). However, the maturity indices of the four locations were less than 3, indicating low sediment food web maturity indices and persistence of MP pollutants among the four locations [51]. In contrast, there were significant differences among the Sigma maturity indices (ƩMI) between the four locations (P < 0.05) (Figure 18). The sigma maturity indices recorded at each site were as follows: site B for MLF (3.13 ± 0.27), site A for DHC (3.10 ± 0.51), site C for MCR (2.43 ± 0.33), and site D for GI (2.61 ± 0.26).
The analysis of the ecological triangle model depicted in Figure 19 demonstrated that sediment samples from site C - MCR and site A - DHC were closer and placed toward the 100% stressed environment, indicating a stressed state, likely influenced by various factors including temperature, pH, dissolved oxygen levels, heavy metals, pesticides, organic pollutants, and nutrient levels. The sediment samples from site B - MLF and site D – GI, on the other hand, were found to be more stable, despite lower nutrient levels, as denoted as indicated by their positioning on the c-p triangle. Overall, results suggested that the river sediment maintained relative stability and exhibited low levels of stress, implying good sediment quality suitable for nematodes. However, the presence of MP in the sediment could potentially perturb the nematode community, leading to alterations in species composition or abundance (Table 7). Notably, the GI samples contained a higher number of MP as reported in a previous study by [68].While the positioning of the sediment samples at the nematode c-p triangle at the bottom by the stability and stress axis may signal good sediment health, the presence of MP could still adversely impact the nematode community within these sediment samples.
The Food Web analysis depicted in Figure 20 was used for the evaluation of sediment health in the River Barrow (RB). The analysis displayed in Figure 20a revealed indications of poor sediment health as evidenced by the alignment of all data points along the lower end of the structure index axis. This observed pattern may be attributed to various factors including decreased oxygen levels, elevated pollutant concentrations, or degradation of habitat. Additionally, the alignment of the nematode food web analysis data points along the lower end of the graph suggested the presence of significant stressors impacting the health of sediment and the nematode community within the river sediment. It is important that further investigation and ongoing monitoring of these factors be conducted in order to gain a comprehensive understanding and effectively address potential threats to the ecosystem.
4. Discussion
To the best of our knowledge, this is the first study to evaluate possible toxic effects of MP pollutants in EPN. Previous studies [50,69,70] in relation to MP toxicity have used C. elegans mostly and the freshwater amphipod Gammarus duebeni as sentinel organisms, while focusing more on the impacts of MP upon ingestion, reproduction, oxidative stress, with less emphasis on assessing mortality. In this study, it was observed that contaminated MP were able to uptake toxicants such as atrazine, resulting in the mortality of nematodes. Similar studies in relation to MP as carriers of toxicants, have also shown that MP act as vectors of various environmental pollutants such as polycyclic aromatic hydrocarbons (PAH), or hydrophobic persistent organic pollutants (POP), pesticides, and heavy metals, thus affecting test organisms, including nematodes [11,49]. Furthermore, [43,46] have also found that MP cause mortality of C. elegans, as well as inhibiting their growth and reproduction [43,46]. This study is the first to evaluate the effects of MP on the three nematode species tested. The findings can now be compared to other studies that have investigated the impact of MP on nematodes. Previous research also examined how MP affect C. elegans, concluding that MP are causing significant issues in this particular nematode species [19,70]. By comparing the results of this study to these previous findings, a more comprehensive understanding of the impact of MP on nematodes can be achieved.
This study demonstrated that increasing MP exposure time from 72 to 96 hours increased the mortality of nematodes exposed to the various treatments. In the case of naphthalene, these increases were from 26 to 30% (S. feltiae SB 12(1) IJ), 29 to 40% (S. carpocapsae IJ) and 26 to 35% (C. elegans J3). Nematode mortality was shown to increase prominently in the presence of MP treated with atrazine and 1,3 – dichloropropene than that in the presence of MP treated with naphthalene and fluorene. [19,46] have also reported the impacts of MP on C. elegans. When comparing the three species, the results of this paper clearly showed that S. carpocapsae IJ and C. elegans J3 responded differently to all the treatments containing MP compared to S. feltiae SB 12(1) IJ. In addition, the S. carpocapsae IJ and C. elegans J3 mean percentage mortality levels were significantly higher (P < 0.05) compared to that in S. feltiae SB 12(1) IJ. From the three species used C. elegans is usually the most sensitive test organism [19]. However, C. elegans J3 mortality was lower compared to that in S. carpocapsae IJ. This presents a possible justification for the utility of S. carpocapsae IJ as sentinel organisms in future toxicology assays, and research based evidence suggests that S. carpocapsae can also be used as a sentinel organism alongside C. elegans. This study demonstrated that the toxicity of MP on the three nematode species differed depending on the treatments and exposure time. The cause of mortality in nematodes could be as a result of leaching of MP additives as this was also oberved by [19]. Similar findings indicated that toxic chemicals absorbed by MP may potentially have very high impact on C. elegans tissues inducing toxic effects [44,49,50].
This study is also the first to fill the gap of knowledge on MP pollution and its environmental impact on the RB sediment nematodes communities. The majority of similar studies focused on the impacts of MP on nematode (namely C. elegans) growth, reproduction, and survival [19,71], as well as MP ingestion by benthic organisms such as freshwater amphipods Gammarus duebeni [72]. Studies [20,25] showed that nematodes found in soil and river sediment can act as bioindicators of MP environmental pollution. Recently, the authors of this study confirmed the presence of MP in RB sediment samples [68]. A similar study by [69] made similar observation confirming plastic microfibres widely present in freshwater sediments.
The higher number of reads obtained in the MCR samples are likely due to the abundance of Monhysterida in this particular location (57 ± 22 %) (Figure 10a). The average read length was 301 bp which was larger than 99% of Good`s coverage, as defined in the scikit-bio documentation (http://scikit-bio.org/docs/latest/generated/skbio.diversity.alpha.goods_coverage.html#skbio.diversity.alpha.goods_coverage) for the 18 S rRNA gene region, with an average GC content of 46%. According to rarefaction curves, a common method for assessing the biodiversity of samples, OTU saturation was achieved at approximately 60,000 reads per sample.The dilution curves of the sediment samples tended to be flat at all four locations, while the overall OTU coverage of the samples appeared to have reached saturation, suggesting that the sequencing depth adequately reflected the structure of the nematode community (Figure 9).
The four locations selected in this study revealed significant differences in nematode communities. For example, the DHC location was sandy/gravel, the MLF location was gravel, the MCR location was muddy/fine grain size, and the GI location was muddy/fine grains, which were the notable differences across the study locations. The smaller the grain size, the less diverse a nematode assemblage is [73]. For instance, compared to the sediment samples from the other three locations, the GI location's sediment samples had higher abundances of only three nematode orders. The three main orders observed from the GI sediment samples were Monhysterida, followed by Chromadorida, and Dorylamida. Monhesterida were among the most numerous groups of nematodes in the sediment samples according to the data of this study. According to [19], MP and their associated toxicants have the potential to adversely impact soil nematodes. As many of the harmful chemicals introduced into waters bind to settling particles, soft sediments are frequently very polluted [74]. This could potentially explain the difference in nematode feeding types and communities observed across the four locations.[75] similarly observed that sediment grain size and organic matter content have strong correlations with nematode communities.
In this study, it was observed that the coarser the sediments the more diverse the communities. This was particularly observed in DHC and MLF samples compared to MCR and GI samples, as the diversity and abundance of sediment nematodes significantly decreased in MCR and GI samples compared to DHC and MLF. It has been also shown via assessment of MP in Irish river sediment, that GI was the most polluted location with MP compared to the rest of the locations used in this current work [68]. There is a clear evidence that MP have significantly affected the nematode communities as the number of nematode orders in the MCR and GI were lower compared to that in the DHC and MLF samples (Figure 12b). Similar findings were observed by [76].
The increasing MP pollutants have been considered as an emerging threat to biodiversity and ecosystem functioning while exerting significant negative effects on the abundance of nematodes [20,77]. Furthermore, the authors’ recent paper [68] revealed that finer sediments act as a sink for MP deposition, which clearly explained the decline in the species abundance and nematode feeding types in the MCR and GI locations. These two particular sites were found to have fine sediments with more deposited MP compared to sediment samples from DHC and MLF locations. [77] confirmed that the presence of various MP in the sediments can reduce nematode feeding types and alter the sediment pH. Despite all this evidence however, the impacts of MP pollution on nematode communities of sediments in the RB in other counties in Ireland remains largely unknown.
The nematodes community structure is indicative of the condition or health of the soil/sediment in which nematodes live, as they respond rapidly to disturbance and enrichment [51]. The impact of MP contamination in the river sediment that caused the disruption of nematode communities was evaluated using a number of indices. For free-living nematodes, the maturity and sigma maturity indices were used to assess the environmental disruption caused by MP. Nematodes were assigned to c-p values according to Bongers (1990). The most resistant nematode taxa to MP pollution were found to be those in class c-p2. [64] also reported similar results, however, it was regarding heavy metals rather than MP. Among the nematode feeding type composition, there was more variation in sites A - DHC and D - MLF compared to sites C - MCR and D - GI. Bacterivores were the most dominant, followed by predators and herbivores. The abundance of bacterivores was high in sites C (MCR) and D (GI). A plausible explanation is that the presence of environmental factors, including organic/inorganic carbon, nitrogen, phosphorus, and other minerals in the environment may modify the direct negative impact of MP. [2,20] also made similar observations. On the other hand, omnivores were the lowest in abundance. Sites C - MCR and D - GI had the highest level of MP compared to the two other locations. The disturbance in nematode feeding types could potentially be caused by the presence of MP. Therefore, these indices can be used as guidelines by the EU Water Framework Directive (WFD) for monitoring MP pollution in the RB sediments. Lower maturity and sigma maturity indices in river sediment have also been observed throughout the four locations, particularly in locations with a high number of MP, including MCR and GI, indicating a disturbed sediment nematode community by MP. The findings reported here support the hypothesis that perturbation generally leads to a lower maturity indices [63]. The disturbance caused by MP likely disrupts the natural habitat and food sources of sediment nematodes, ultimately impacting their development and maturity levels. Thus, the findings from this study provide further evidence linking perturbation, specifically from MP, to lower maturity indices in sediment nematode communities.
Analysis of river sediment samples from the four locations along the RB shows that MP do affect the nematode communities, causing a decline in nematode diversities, feeding type, and maturity index. Therefore, based on the findings of this study, it can be inferred that MP pollution of river sediment in the four locations along the RB is evident. This statement is supported by the observed decrease in nematode diversities, changes in feeding type composition, and lower maturity indices in sediment nematode communities. The disruption caused by MP contamination likely alters the natural habitat and food sources of sediment-dwelling nematodes, ultimately influencing their development and maturity levels. The results from this research provide further evidence of the relationship between disturbance, particularly from MP, and the reduced maturity indices in sediment nematode communities. This in turn indicates that the health and condition of the river sediment in the four locations are indeed affected by MP pollution, as suggested by the observed changes in nematode communities. Nematode communities are at risk as a result of the accumulation of MP contaminants in river sediments. Nematode communities were mostly affected by the presence of MP in samples from the MCR and GI locations. A decrease in the number of nematode feeding types, and maturity indices was also observed in the MCR and GI locations, potentially associated with the high number of MP. Furthermore, sediment MP contamination can modify the composition of feeding types in nematode communities, both directly and indirectly, due to differences in the sensitivity of various nematode species [78,79]. The low sediment web maturity indices observed can be attributed to the ongoing rise in MP within the RB sediments [68]. Therefore, it is important for ecologists to use these indices as a means to improve the sediment quality of the RB by reducing the use of non-biodegradable plastics and thereby mitigating the environmental effects of MP. Stakeholders such as policymakers, industry, government agencies, and the general public can also play a role in reducing non-biodegradable inputs and mitigating the environmental impacts of MP. Implementing regulations on the use of plastics, promoting recyclable and biodegradable materials, and educating the public on the importance of reducing plastic use and pollution can all contribute to improving the sediment quality of the RB. Collaborative efforts between environmental activists, policymakers, industry, and the public are essential in addressing the issue of MP contamination and promoting environmental sustainability. The impact of this research is significant, as it highlighted the negative effects of MP on beneficial nematodes and nematode biodiversity in river sediments, ultimately affecting the overall ecological health of aquatic ecosystems. The findings of this research can be implemented in several ways: (1) Environmental policy and regulation: The data on the impacts of MP on nematode communities can inform policymakers and regulators about the potential ecological risks posed by MP in aquatic environments. This information can be used to develop strategies to reduce MP pollution and protect benthic ecosystems. (2) Eco-friendly product development: The identification of commercial cosmetic products as a potential source of MP underscored the importance of developing eco-friendly alternatives that do not contain harmful pollutants. Relevant industry can use this information to reformulate their products and reduce their environmental impact. (3) Further research: The study highlighted the need for more research on the pathways of nanoparticles in organisms and the effects of MP on different types of organisms. Future studies can build upon these findings to deepen understanding of the ecological implications of MP pollution. By addressing the issue of MP pollution and its effects on benthic organisms, such as nematodes, steps can be taken to protect and preserve aquatic ecosystems for future generations. This study will contribute to the limited work on the impacts of MP pollutants in the RB sediments.
5. Conclusions
This paper presents for the first time the toxic effects of MP pollutants on EPN. Attenuated total reflectance Fourier transform infrared spectroscopy showed that atrazine accumulated on the surface of polyethylene MP. There was some evidence of interaction between these MP and both 1,3–dichloropropene and naphthalene, while fluorene did not appear to be retained on the MP surface. The present study showed that S. carpocapsae IJ and C. elegans J3, when exposed in vitro to MP for 72 to 96 hours, exhibited significant effects in all treatments. The mortality percentages were notably higher compared to S. feltiae (SB12(1) IJ. Furthermore, continuous exposure of nematodes to MP led in increasing nematodes mortality. Therefore, experiments using the three species of nematodes as test organisms demonstrated that MP treated with a range of toxicants affected the survival percentages of the nematodes. The results demonstrated the toxicity of MP among the three species, confirming the hypothesis that MP particles act as vector of contaminants for nematodes.
This work explored the capacity of MP to absorb and transfer chemical pollutants into nematodes, providing experimental insights and considerations for studying EPN as model organisms. This suggests that EPN can serve as excellent models for studying the transfer of chemical pollutants via MP in aquatic ecosystems. Microplastics have the ability of absorbing toxic substances. Therefore, limiting the release of MP may reduce their ability to damage organisms and ecosystems. Thus, these findings raise a great concern regarding the long-term exposure of nematode species to MP and highlight the urgent need of more studies. Subsequent research may consider using S. carpocapsae as a sentinel organism in toxicological studies due to the extensive use of C. elegans in such assays. This is justified by the significant findings observed in this study, where both S. carpocapsae and C. elegans showed notable responses to exposure to MP. The mortality rates of both nematode species were higher compared to S. feltiae, indicating the potential toxicity of MP to nematodes. Given the prevalence of research focusing on C. elegans in toxicological assessments, incorporating S. carpocapsae as a sentinel organism could provide a more comprehensive understanding of the impacts of MP on nematode communities. This approach would also broaden the scope of research on the effects of MP on various nematode species, ultimately contributing to a more thorough assessment of the ecological implications of MP pollution.
This study additionally examines the distribution of sediment-dwelling nematode communities within the RB. The abundance and diversity of sediment nematodes were significantly affected by the presence of MP. In conclusion, MP have negative impacts on nematode communities. This work gave an insight into the status on the community structure of nematodes in river sediment from the DHC, MLF, MCR, and GI locations. The differences in sediment types and MP occurrence can shape the meiofaunal nematode communities. This study revealed that the highest nematode richness and abundance were observed in the DHC location, followed by MLF, while the lowest nematode richness and abundance were found in the MCR and GI locations. Monhysterida was the predominant order across all the samples, particularly in the MCR samples. The differences in nematode communities in the various sediments indicate significant MP impacts on the nematode community composition.
The composition of the nematode communities in the sediments, the trophic groups recorded and the maturity index can be used in ecological monitoring of the RB. The accumulation of MP in river sediments, as demonstrated by [68] poses a threat to nematode communities. Therefore, it could be useful to assess the preliminary ecosystem functions of the RB sediment in order to mitigate the potential impact of MP accumulation on nematode communities. This assessment can be conducted by analysing the composition of nematode communities, trophic groups, and maturity index as indicators for ecological monitoring of the RB. Preliminary ecosystem functions refer to the initial assessment of the ecological processes and functions in a particular ecosystem, such as the distribution and diversity of sediment-dwelling nematode communities. By analysing the composition of nematode communities, trophic groups, and maturity index as indicators for ecological monitoring, one can gain valuable insights into the status of the ecosystem and potential impacts of MP accumulation on nematode communities. This preliminary assessment can help in understanding the baseline conditions of the ecosystem and in recognising any changes that may occur due to factors such as MP accumulation. This information can then be used to develop strategies to mitigate the negative impact of MP on nematode communities, such as implementing measures to reduce MP pollution and restore the ecosystem balance.
The findings from this work should facilitate further research concerning MP pollutants in the RB at different counties in Ireland. This will provide better indications of the stability of the sediments that benthic nematode communities inhabit and provide better insights into how such communities may respond to anthropogenic disturbance which includes MP. Finally, based on the findings from this study, sediment nematodes can serve as bioindicators of MP environmental pollution.
Author Contributions
Conceptualization, L.M., J.C., K.G. and T.K.D.; methodology, L.M., J.C., and T.K.D.; software, L.M.; validation, L.M., J.C., K.G. and T.K.D.; formal analysis, L.M., J.C., K.G. and T.D.K.; investigation, L.M., J.C., K.G., and T.K.D.; writing—original draft preparation, L.M.; writing—review and editing, L.M. and T.K.D.; visualization, L.M., J.C., K.G., and T.K.D; supervision, J.C., K.G., and T.K.D. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Irish Research Council, grant number GOIPG/2019/2786.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in this study are incorporated within the article.
Acknowledgments
This study was funded by the Irish Research Council (IRC) under the Government of Ireland Postgraduate Scholarship programme, project ID: GOIPG/2019/2786.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- He, D.; Luo, Y.; Lu, S.; Liu, M.; Song, Y.; Lei, L. Microplastics in soils: Analytical methods, pollution characteristics and ecological risks. TrAC - Trends Anal. Chem. 2018, 109, 163–172. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, W.; Zhang, Z.; Grossart, H.P.; Gadd, G.M. Microplastics provide new microbial niches in aquatic environments. Appl. Microbiol. Biotechnol. 2020, 104, 6501–6511. [Google Scholar] [CrossRef] [PubMed]
- Talbot, R.; Chang, H. Microplastics in freshwater: A global review of factors affecting spatial and temporal variations. Environ. Pollut. 2022, 292, 118393. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Li, W.; Yang, W.; Li, W.; Yang, J.; Yang, F. Ecological effects of microplastics on microorganism characteristics in sediments of the Daihai Lakeshore, China. J. Freshw. Ecol. 2021, 37, 21–37. [Google Scholar] [CrossRef]
- European Commission. Directorate-General for Environment. EU action against microplastics, 2023. [Google Scholar]
- Wright, S.L.; Kelly, F.J. Plastic and Human Health: A Micro Issue? Environ. Sci. Technol. 2017, 51, 6634–6647. [Google Scholar] [CrossRef]
- Borges-Ramírez, M.M.; Escalona-Segura, G.; Huerta-Lwanga, E.; Iñigo-Elias, E.; Osten, J.R. von Organochlorine pesticides, polycyclic aromatic hydrocarbons, metals and metalloids in microplastics found in regurgitated pellets of black vulture from Campeche, Mexico. Sci. Total Environ. 2021, 801, 149674. [Google Scholar] [CrossRef]
- Liu, G.; Zhu, Z.; Yang, Y.; Sun, Y.; Yu, F.; Ma, J. Sorption behavior and mechanism of hydrophilic organic chemicals to virgin and aged microplastics in freshwater and seawater. Environ. Pollut. 2019, 246, 26–33. [Google Scholar] [CrossRef]
- Singla, M.; Díaz, J.; Broto-Puig, F.; Borrós, S. Sorption and release process of polybrominated diphenyl ethers (PBDEs) from different composition microplastics in aqueous medium: Solubility parameter approach. Environ. Pollut. 2020, 262. [Google Scholar] [CrossRef]
- Yazdani Foshtomi, M.; Oryan, S.; Taheri, M.; Darvish Bastami, K.; Zahed, M.A. Composition and abundance of microplastics in surface sediments and their interaction with sedimentary heavy metals, PAHs and TPH (total petroleum hydrocarbons). Mar. Pollut. Bull. 2019, 149, 110655. [Google Scholar] [CrossRef]
- Du, S.; Zhu, R.; Cai, Y.; Xu, N.; Yap, P.S.; Zhang, Y.; He, Y.; Zhang, Y. Environmental fate and impacts of microplastics in aquatic ecosystems: A review. RSC Adv. 2021, 11, 15762–15784. [Google Scholar] [CrossRef]
- Zhang, B.; Chen, L.; Chao, J.; Yang, X.; Wang, Q. Research Progress of Microplastics in Freshwater Sediments in China. Environ. Sci. Pollut. Res. 2020, 27, 31046–31060. [Google Scholar] [CrossRef] [PubMed]
- Ogonowski, M.; Motiei, A.; Ininbergs, K.; Hell, E.; Gerdes, Z.; Udekwu, K.I.; Bacsik, Z.; Gorokhova, E. Evidence for selective bacterial community structuring on microplastics. Environ. Microbiol. 2018, 20, 2796–2808. [Google Scholar] [CrossRef] [PubMed]
- Kosuth, M.; Mason, S.A.; Wattenberg, E.V. Anthropogenic contamination of tap water, beer, and sea salt. PLoS One 2018, 13, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Kunz, A.; Shim, W.J.; Walther, B.A. Microplastic contamination of table salts from Taiwan, including a global review. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Li; Shi, Y.; Yang, L.; Xiao, L.; Kehoe, D.K.; Gun’ko, Y.K.; Boland, J.J.; Wang, J.J. Microplastic release from the degradation of polypropylene feeding bottles during infant formula preparation. Nat. Food 2020, 1, 746–754. [CrossRef]
- Dessì, C.; Okoffo, E.D.; O’Brien, J.W.; Gallen, M.; Samanipour, S.; Kaserzon, S.; Rauert, C.; Wang, X.; Thomas, K.V. Plastics contamination of store-bought rice. J. Hazard. Mater. 2021, 416, 125778. [Google Scholar] [CrossRef]
- Thiele, C.J.; Hudson, M.D.; Russell, A.E.; Saluveer, M.; Sidaoui-Haddad, G. Microplastics in fish and fishmeal: an emerging environmental challenge? Sci. Rep. 2021, 11, 1–12. [Google Scholar] [CrossRef]
- Kim, S.W.; Waldman, W.R.; Kim, T.Y.; Rillig, M.C. Effects of Different Microplastics on Nematodes in the Soil Environment: Tracking the Extractable Additives Using an Ecotoxicological Approach. Environ. Sci. Technol. 2020, 54, 13868–13878. [Google Scholar] [CrossRef]
- Yang, B.; Li, P.; Entemake, W.; Guo, Z.; Xue, S. Concentration-Dependent Impacts of Microplastics on Soil Nematode Community in Bulk Soils of Maize: Evidence From a Pot Experiment. Front. Environ. Sci. 2022, 10, 1–11. [Google Scholar] [CrossRef]
- Bellasi, A.; Binda, G.; Pozzi, A.; Galafassi, S.; Volta, P.; Bettinetti, R. Microplastic contamination in freshwater environments: A review, focusing on interactions with sediments and benthic organisms. Environ. - MDPI 2020, 7. [Google Scholar] [CrossRef]
- Murphy, L.; Germaine, K.; Dowling, D.N.; Kakouli-Duarte, T.; Cleary, J. Association of Potential Human Pathogens with Microplastics in Freshwater Systems. 2020, 112–120. [Google Scholar] [CrossRef]
- Rodrigues, M.O.; Abrantes, N.; Gonçalves, F.J.M.; Nogueira, H.; Marques, J.C.; Gonçalves, A.M.M. Spatial and temporal distribution of microplastics in water and sediments of a freshwater system (Antuã River, Portugal). Sci. Total Environ. 2018, 633, 1549–1559. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Seijo, A.; Santos, B.; Ferreira Da Silva, E.; Cachada, A.; Pereira, R. Low-density polyethylene microplastics as a source and carriers of agrochemicals to soil and earthworms. Environ. Chem. 2019, 16, 8–17. [Google Scholar] [CrossRef]
- Schenk, J.; Höss, S.; Brinke, M.; Kleinbölting, N.; Brüchner-Hüttemann, H.; Traunspurger, W. Nematodes as bioindicators of polluted sediments using metabarcoding and microscopic taxonomy. Environ. Int. 2020, 143, 105922. [Google Scholar] [CrossRef]
- Höss, S.; Rauchschwalbe, M.T.; Fueser, H.; Traunspurger, W. Food availability is crucial for effects of 1-μm polystyrene beads on the nematode Caenorhabditis elegans in freshwater sediments. Chemosphere 2022, 298. [Google Scholar] [CrossRef]
- Drummond, J.D.; Schneidewind, U.; Li, A.; Hoellein, T.J.; Krause, S.; Packman, A.I. Microplastic accumulation in riverbed sediment via hyporheic exchange from headwaters to mainstems. Sci. Adv. 2022, 8. [Google Scholar] [CrossRef]
- da Silva Junior, F.C.; Felipe, M.B.M.C.; de Castro, D.E.F.; Araújo, S.C. da S.; Sisenando, H.C.N.; Batistuzzo de Medeiros, S.R. A look beyond the priority: A systematic review of the genotoxic, mutagenic, and carcinogenic endpoints of non-priority PAHs. Environ. Pollut. 2021, 278. [Google Scholar] [CrossRef]
- Honda, M.; Suzuki, N. Toxicities of polycyclic aromatic hydrocarbons for aquatic animals. Int. J. Environ. Res. Public Health 2020, 17, 1363. [Google Scholar] [CrossRef]
- de Souza, R.M.; Seibert, D.; Quesada, H.B.; de Jesus Bassetti, F.; Fagundes-Klen, M.R.; Bergamasco, R. Occurrence, impacts and general aspects of pesticides in surface water: A review. Process Saf. Environ. Prot. 2020, 135, 22–37. [Google Scholar] [CrossRef]
- Li, H.; Wang, F.; Li, J.; Deng, S.; Zhang, S. Adsorption of three pesticides on polyethylene microplastics in aqueous solutions: Kinetics, isotherms, thermodynamics, and molecular dynamics simulation. Chemosphere 2021, 264, 128556. [Google Scholar] [CrossRef]
- Cheng, H.; Zhang, D.; Huang, B.; Song, Z.; Ren, L.; Hao, B.; Liu, J.; Zhu, J.; Fang, W.; Yan, D.; et al. Organic fertilizer improves soil fertility and restores the bacterial community after 1,3-dichloropropene fumigation. Sci. Total Environ. 2020, 738, 140345. [Google Scholar] [CrossRef] [PubMed]
- Graymore, M.; Stagnitti, F.; Allinson, G. Impacts of atrazine in aquatic ecosystems. Environ. Int. 2001, 26, 483–495. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Kumar, V.; Chauhan, A.; Datta, S.; Wani, A.B.; Singh, N.; Singh, J. Toxicity, degradation and analysis of the herbicide atrazine. Environ. Chem. Lett. 2018, 16, 211–237. [Google Scholar] [CrossRef]
- Yeates, G.W.; Ferris, H.; Moens, T.; Van Der Putten, W.H. The role of nematodes in ecosystems; 2009; ISBN 9781845933852. [Google Scholar]
- Wilson; Kakouli-Duarte, M.T. Ematodes as environmental indicators; 2009; ISBN 9781845933852. [Google Scholar]
- Haight. MUDRY and J. PASTERNAK Toxicity of seven heavy metals on panagrellus Silusiae: The efficacy of the free-living nematode an an in vivo toxicological bioassay. 2017, 91, 399–404.
- Gulcu, B.; Cimen, H.; Raja, R.K.; Hazir, S. Entomopathogenic nematodes and their mutualistic bacteria: Their ecology and application as microbial control agents. Biopestic. Int. 2017, 13, 79–112. [Google Scholar] [CrossRef]
- Kary, N.E.; Sanatipour, Z.; Mohammadi, D.; Dillon, A.B. Combination effects of entomopathogenic nematodes, Heterorhabditis bacteriophora and Steinernema feltiae, with Abamectin on developmental stages of Phthorimaea operculella (Lepidoptera, Gelechiidae). Crop Prot. 2021, 143. [Google Scholar] [CrossRef]
- Akhurst Antibiotic Activity of Xenorhabdus spp., Bacteria Symbiotically Associated with Insect Pathogenic Nematodes of the Families Heterorhabditidae and Steinernematidae. 1982.
- Bode, H.B. Entomopathogenic bacteria as a source of secondary metabolites. Curr. Opin. Chem. Biol. 2009, 13, 224–230. [Google Scholar] [CrossRef]
- Labaude, S.; Griffin, C.T. Transmission success of entomopathogenic nematodes used in pest control. Insects 2018, 9, 72. [Google Scholar] [CrossRef]
- Hu, J.; Li, X.; Lei, L.; Cao, C.; Wang, D.; He, D. The Toxicity of (Nano)Microplastics on C. elegans and Its Mechanisms. Handb. Environ. Chem. 2020, 95, 259–278. [Google Scholar] [CrossRef]
- Ficociello, G.; Gerardi, V.; Uccelletti, D.; Setini, A. Molecular and cellular responses to short exposure to bisphenols A, F, and S and eluates of microplastics in C. elegans. Environ. Sci. Pollut. Res. 2021, 28, 805–818. [Google Scholar] [CrossRef]
- Brenner, S. The Genetics of Caenorhabditis elegans. Genetics 1974, 77, 71–94. [Google Scholar] [CrossRef] [PubMed]
- Shang, X.; Lu, J.; Feng, C.; Ying, Y.; He, Y.; Fang, S.; Lin, Y.; Dahlgren, R.; Ju, J. Microplastic (1 and 5 μm) exposure disturbs lifespan and intestine function in the nematode Caenorhabditis elegans. Sci. Total Environ. 2020, 705, 135837. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.W.; Li, S.W.; Hsiu-Chuan Liao, V. Chronic ZnO-NPs exposure at environmentally relevant concentrations results in metabolic and locomotive toxicities in Caenorhabditis elegans. Environ. Pollut. 2017, 220, 1456–1464. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.; Yang, J.; Li, H.; Cui, C.; Yu, Y.; Liu, Y.; Lin, K. The chronic toxicity of bisphenol A to Caenorhabditis elegans after long-term exposure at environmentally relevant concentrations. Chemosphere 2016, 154, 546–551. [Google Scholar] [CrossRef]
- Jewett, E.; Arnott, G.; Connolly, L.; Vasudevan, N.; Kevei, E. Microplastics and Their Impact on Reproduction—Can we Learn From the C. elegans Model? Front. Toxicol. 2022, 4, 1–21. [Google Scholar] [CrossRef]
- Mueller, M.T.; Fueser, H.; Höss, S.; Traunspurger, W. Species-specific effects of long-term microplastic exposure on the population growth of nematodes, with a focus on microplastic ingestion. Ecol. Indic. 2020, 118, 106698. [Google Scholar] [CrossRef]
- Du Preez, G.; Daneel, M.; De Goede, R.; Du Toit, M.J.; Ferris, H.; Fourie, H.; Geisen, S.; Kakouli-Duarte, T.; Korthals, G.; Sánchez-Moreno, S.; et al. Nematode-based indices in soil ecology: Application, utility, and future directions. Soil Biol. Biochem. 2022, 169. [Google Scholar] [CrossRef]
- Lu, Q.; Liu, T.; Wang, N.; Dou, Z.; Wang, K.; Zuo, Y. A review of soil nematodes as biological indicators for the assessment of soil health. Front. Agric. Sci. Eng. 2020, 7, 275–281. [Google Scholar] [CrossRef]
- Zullini, A. Nematodes as indicators of river pollution. Nematol. Mediterr. 1976, 4, 13–22. [Google Scholar]
- Wu, H.C.; Chen, P.C.; Tsay, T.T. Assessment of nematode community structure as a bioindicator in river monitoring. Environ. Pollut. 2010, 158, 1741–1747. [Google Scholar] [CrossRef]
- Napper, I.E.; Bakir, A.; Rowland, S.J.; Thompson, R.C. Characterisation, quantity and sorptive properties of microplastics extracted from cosmetics. Mar. Pollut. Bull. 2015, 99, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Lei, K.; Qiao, F.; Liu, Q.; Wei, Z.; Qi, H.; Cui, S.; Yue, X.; Deng, Y.; An, L. Microplastics releasing from personal care and cosmetic products in China. Mar. Pollut. Bull. 2017, 123, 122–126. [Google Scholar] [CrossRef] [PubMed]
- Corsi, A.K.; Wightman, B.; Chalfie, M. A Transparent window into biology: A primer on Caenorhabditis elegans. WormBook 2015, 1–31. [Google Scholar] [CrossRef] [PubMed]
- McMullen, J.G.; Stock, S.P. In vivo and In vitro Rearing of Entomopathogenic Nematodes (Steinernematidae and Heterorhabditidae). 2014, 1–7. [CrossRef]
- Stiernagle, T. Maintenance of C. elegans. WormBook 2006, 1–11. [Google Scholar] [CrossRef]
- White. A Method for Obtaining Infective Nematode Larvae from Cultures. 1927, 66, 302–303.
- Porta-de-la-Riva, M.; Fontrodona, L.; Villanueva, A.; Cerón, J. Basic Caenorhabditis elegans methods: synchronization and observation. J. Vis. Exp. 2012, 64, 4019. [Google Scholar] [CrossRef]
- Bhadury, P.; Austen, M.C.; Bilton, D.T.; Lambshead, P.J.D.; Rogers, A.D.; Smerdon, G.R. Development and evaluation of a DNA-barcoding approach for the rapid identification of nematodes. Mar. Ecol. Prog. Ser. 2006, 320, 1–9. [Google Scholar] [CrossRef]
- Bongers, T. The maturity index: an ecological measure of environmental disturbance based on nematode species composition. Oecologia 1990, 83, 14–19. [Google Scholar] [CrossRef]
- Sieriebriennikov, B.; Ferris, H.; de Goede, R.G.M. NINJA: An automated calculation system for nematode-based biological monitoring. Eur. J. Soil Biol. 2014, 61, 90–93. [Google Scholar] [CrossRef]
- Yu, F.; Yang, C.; Zhu, Z.; Bai, X.; Ma, J. Adsorption behavior of organic pollutants and metals on micro/nanoplastics in the aquatic environment. Sci. Total Environ. 2019, 694, 133643. [Google Scholar] [CrossRef]
- Menéndez-Pedriza, A.; Jaumot, J. Interaction of environmental pollutants with microplastics: A critical review of sorption factors, bioaccumulation and ecotoxicological effects. Toxics 2020, 8, 40. [Google Scholar] [CrossRef] [PubMed]
- Bongers, T.; Bongers, M. Functional diversity of nematodes. Appl. Soil Ecol. 1998, 10, 239–251. [Google Scholar] [CrossRef]
- Murphy, L.; Germaine, K.; Kakouli-Duarte, T.; Cleary, J. Assessment of microplastics in Irish river sediment. Heliyon 2022, 8, e09853. [Google Scholar] [CrossRef]
- Mateos-Cárdenas, A.; O’Halloran, J.; van Pelt, F.N.A.M.; Jansen, M.A.K. Beyond plastic microbeads – Short-term feeding of cellulose and polyester microfibers to the freshwater amphipod Gammarus duebeni. Sci. Total Environ. 2021, 753, 141859. [Google Scholar] [CrossRef]
- Chen, H.; Hua, X.; Li, H.; Wang, C.; Dang, Y.; Ding, P.; Yu, Y. Transgenerational neurotoxicity of polystyrene microplastics induced by oxidative stress in Caenorhabditis elegans. Chemosphere 2021, 272, 129642. [Google Scholar] [CrossRef]
- Kim, S.W.; Kim, D.; Jeong, S.W.; An, Y.J. Size-dependent effects of polystyrene plastic particles on the nematode Caenorhabditis elegans as related to soil physicochemical properties. Environ. Pollut. 2019, 258, 113740. [Google Scholar] [CrossRef]
- Mateos-Cárdenas, Alexandra, R.J.; Jansen, John O’Halloran, F.N.A.M. van P. and M.A.K.J. Impacts of Microplastics in the Irish Freshwater Environment. Available online: https://www.epa.ie/publications/research/epa-research-2030-reports/Research_Report_377.pdf.
- Tytgat, B.; Nguyen, D.T.; Nguyen, T.X.P.; Pham, T.M.; Long, P.K.; Vanreusel, A.; Derycke, S. Monitoring of marine nematode communities through 18S rRNA metabarcoding as a sensitive alternative to morphology. Ecol. Indic. 2019, 107, 105554. [Google Scholar] [CrossRef]
- Höss, S.; Claus, E.; Von der Ohe, P.C.; Brinke, M.; Güde, H.; Heininger, P.; Traunspurger, W. Nematode species at risk - A metric to assess pollution in soft sediments of freshwaters. Environ. Int. 2011, 37, 940–949. [Google Scholar] [CrossRef]
- Stark, J.S.; Mohammad, M.; McMinn, A.; Ingels, J. Diversity, Abundance, Spatial Variation, and Human Impacts in Marine Meiobenthic Nematode and Copepod Communities at Casey Station, East Antarctica. Front. Mar. Sci. 2020, 7. [Google Scholar] [CrossRef]
- Steyaert, M.; Garner, N.; Van Gansbeke, D.; Vincx, M. Nematode communities from the North Sea: Environmental controls on species diversity and vertical distribution within the sediment. J. Mar. Biol. Assoc. United Kingdom 1999, 79, 253–264. [Google Scholar] [CrossRef]
- Boots, B.; Russell, C.W.; Green, D.S. Effects of Microplastics in Soil Ecosystems: Above and below Ground. Environ. Sci. Technol. 2019, 53, 11496–11506. [Google Scholar] [CrossRef] [PubMed]
- Heininger, P.; Höss, S.; Claus, E.; Pelzer, J.; Traunspurger, W. Nematode communities in contaminated river sediments. Environ. Pollut. 2007, 146, 64–76. [Google Scholar] [CrossRef] [PubMed]
- Ji, Z.; Huang, Y.; Feng, Y.; Johansen, A.; Xue, J.; Tremblay, L.A.; Li, Z. Effects of pristine microplastics and nanoplastics on soil invertebrates: A systematic review and meta-analysis of available data. Sci. Total Environ. 2021, 788, 147784. [Google Scholar] [CrossRef]
Figure 1.
Seeded NGM plates.
Figure 2.
Rearing of entomopathogenic nematodes.

Figure 3.
Caenorhabditis. elegans age synchronization. (a) Various C. elegans stages on an agar culture plate; the substrate was pre-inoculated with the nematode food, Escherichia coli OP50; (b) Hatched eggs on unseeded NGM plates; (c) Age synchronised N2 C. elegans J3 at 20 °C after 3 days.
Figure 3.
Caenorhabditis. elegans age synchronization. (a) Various C. elegans stages on an agar culture plate; the substrate was pre-inoculated with the nematode food, Escherichia coli OP50; (b) Hatched eggs on unseeded NGM plates; (c) Age synchronised N2 C. elegans J3 at 20 °C after 3 days.

Figure 4.
The assessment of nematode mortality was done using a stereoscope. The tests were done in 96 well plates containing various treatments and 10 nematodes in each well.
Figure 4.
The assessment of nematode mortality was done using a stereoscope. The tests were done in 96 well plates containing various treatments and 10 nematodes in each well.
Figure 5.
(a) Microplastics extracted from cosmetic products; (b) Stereoscope images of the extracted MP.
Figure 5.
(a) Microplastics extracted from cosmetic products; (b) Stereoscope images of the extracted MP.

Figure 6.
(a) ATR-FTIR spectrum of commercial clear polyethylene; ATR-FTIR spectra of MP after immersion in: (b) 100ppb atrazine; (c) atrazine reference spectrum; (d) 100 ppb 1,3–dichloropropene; (e) 1,3–dichloropropene reference spectrum; (f) 100 ppb naphthalene; (g) naphthalene reference spectrum; (h) 100 ppb fluorene; (i) fluorene reference spectrum, showing MP absorption of toxic chemicals. The blue circles on the spectra represent original peaks before MP exposure to treatments.
Figure 6.
(a) ATR-FTIR spectrum of commercial clear polyethylene; ATR-FTIR spectra of MP after immersion in: (b) 100ppb atrazine; (c) atrazine reference spectrum; (d) 100 ppb 1,3–dichloropropene; (e) 1,3–dichloropropene reference spectrum; (f) 100 ppb naphthalene; (g) naphthalene reference spectrum; (h) 100 ppb fluorene; (i) fluorene reference spectrum, showing MP absorption of toxic chemicals. The blue circles on the spectra represent original peaks before MP exposure to treatments.
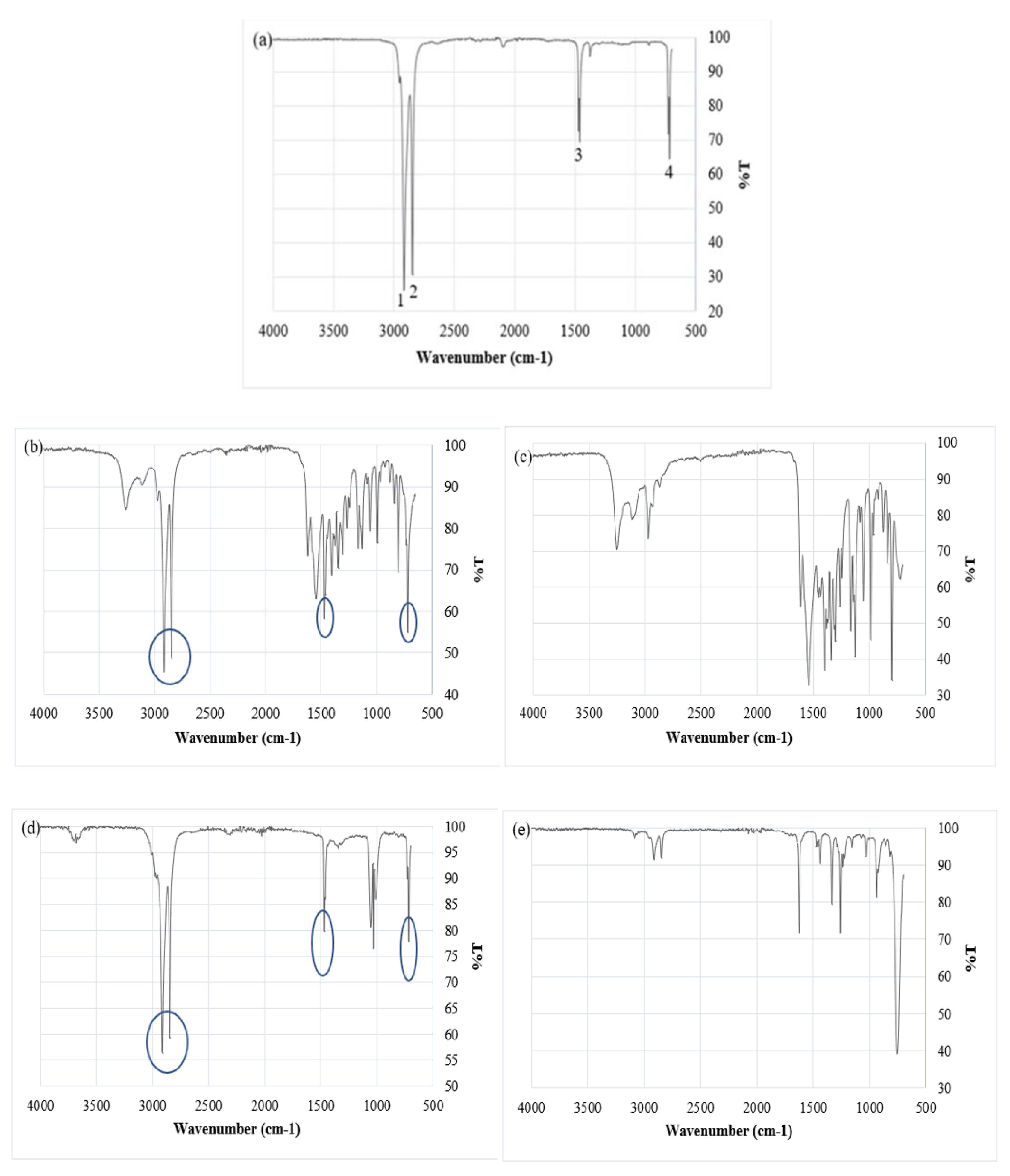
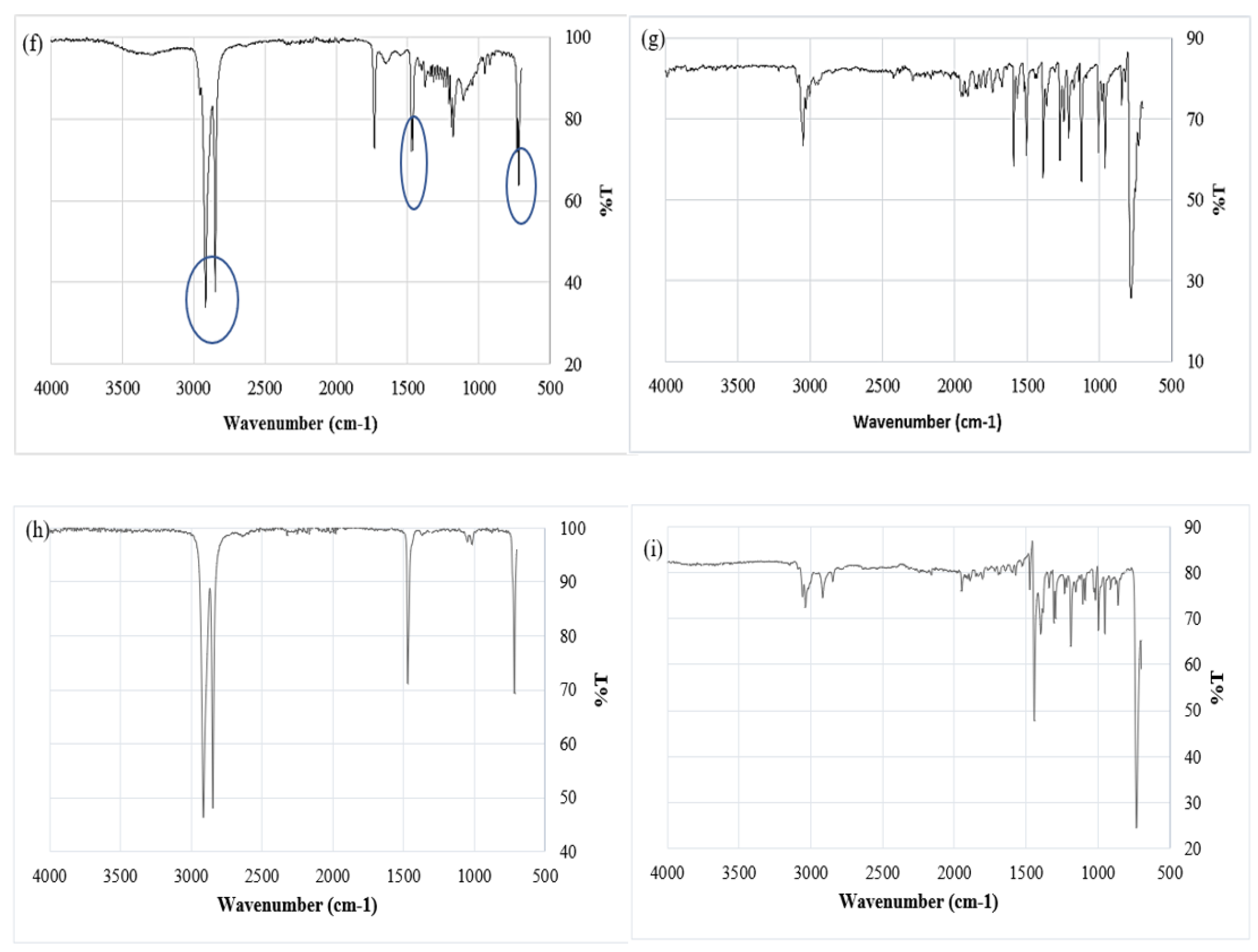
Figure 7.
Comparison of mortality of S. feltiae SB 12(1) IJ, S. carpocapsae IJ and C. elegans J3 exposed to a mixture of MP in various treatments after 72 hours (a-d), 96 hours (e-h).
Figure 7.
Comparison of mortality of S. feltiae SB 12(1) IJ, S. carpocapsae IJ and C. elegans J3 exposed to a mixture of MP in various treatments after 72 hours (a-d), 96 hours (e-h).
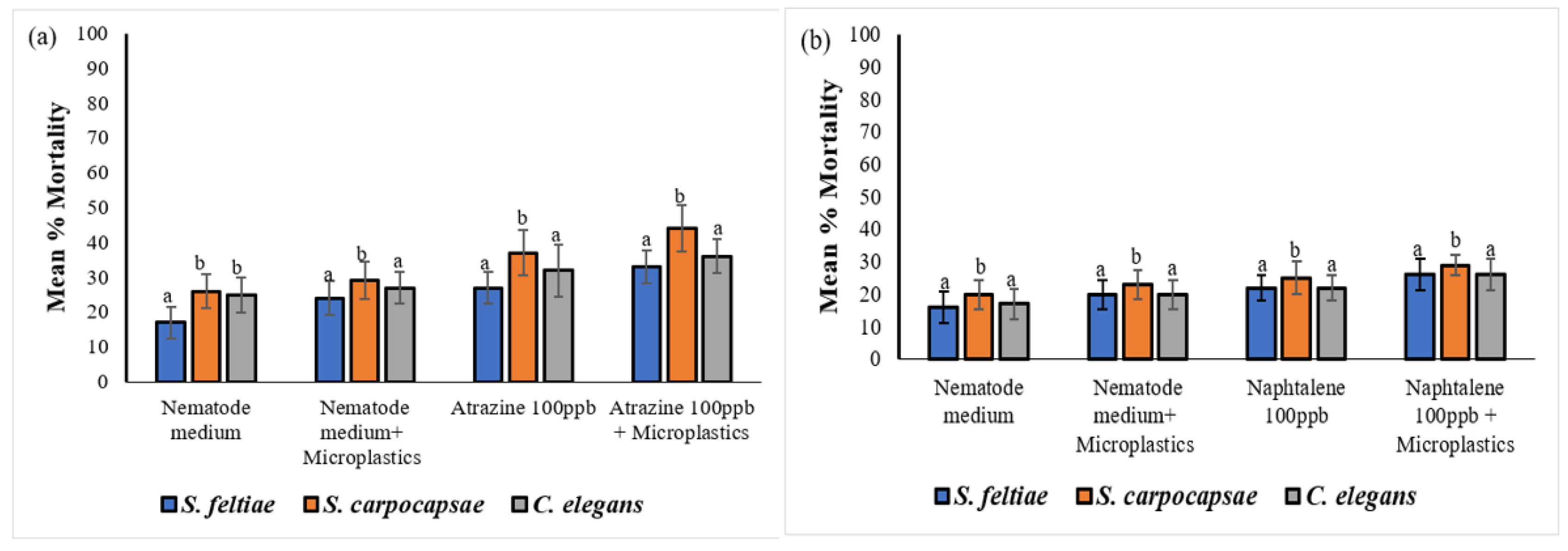
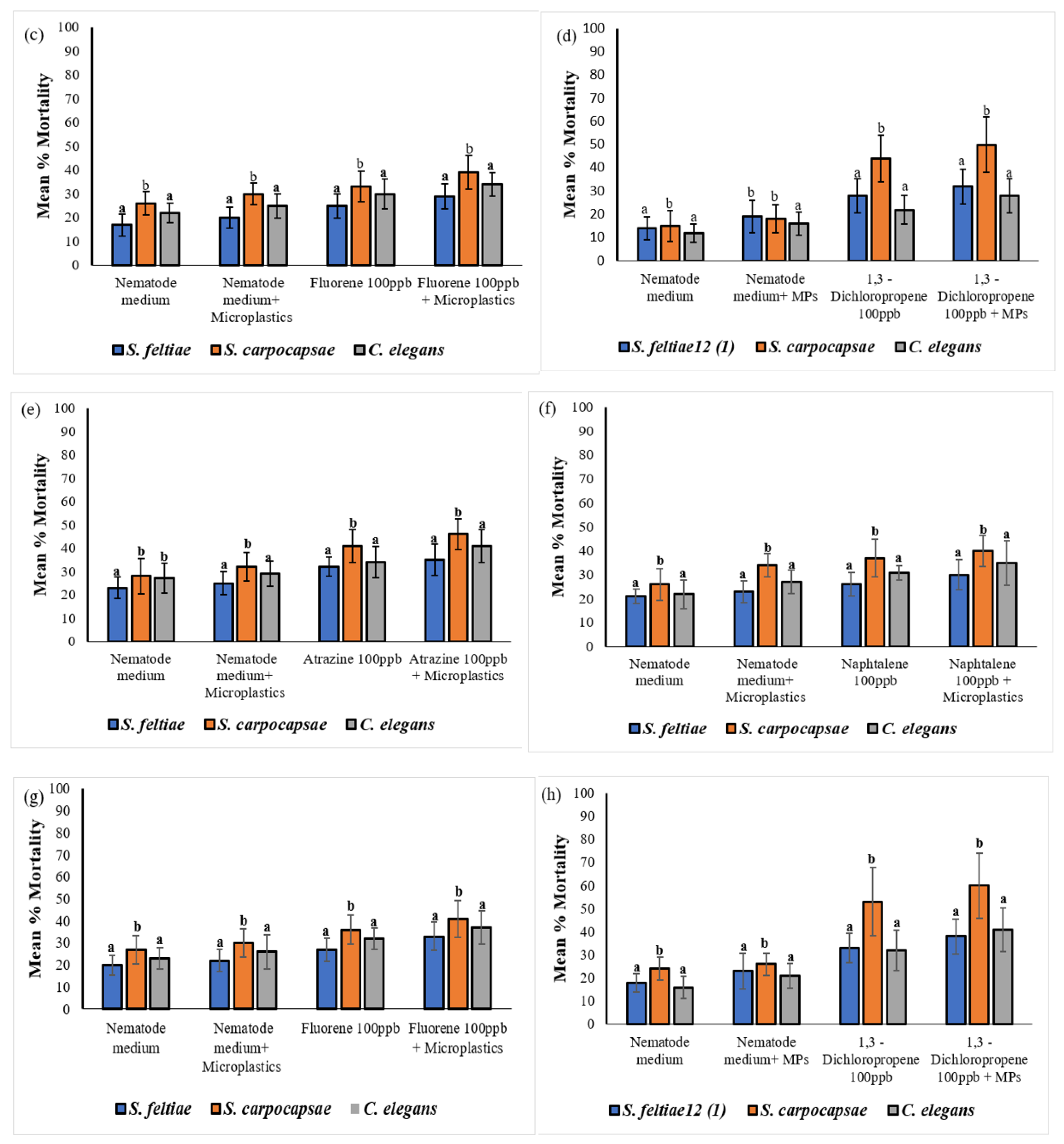
Figure 10.
River sediment nematode abundances. (a) Relative abundance of sediment nematode classes in samples from the locations of the study ; (b) The relative distribution of sequences in the nematode dataset within different nematode orders for all sediment samples.
Figure 10.
River sediment nematode abundances. (a) Relative abundance of sediment nematode classes in samples from the locations of the study ; (b) The relative distribution of sequences in the nematode dataset within different nematode orders for all sediment samples.
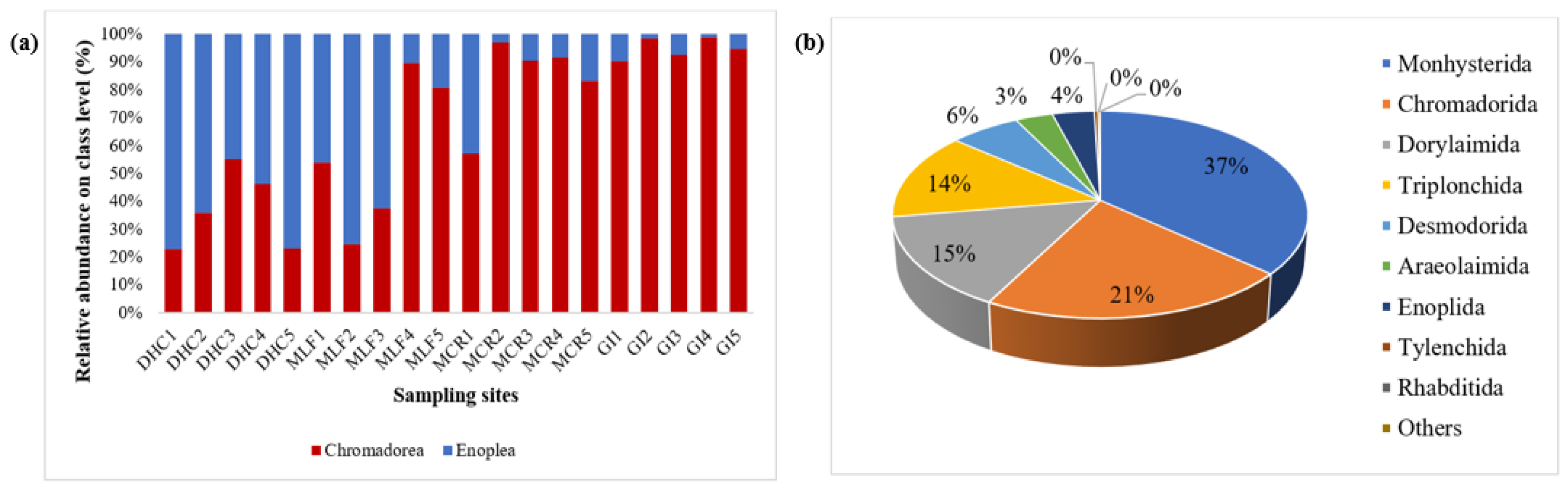
Figure 11.
River sediment nematode community composition. (a) The most frequently occurring nematode genera in the study sites and (b) Heatmap representing the abundances of the nine most prevalent nematode orders among the four locations. Location group name DHC: Dolmen Hotel – Carlow; GI: Great Island – Wexford; MCR: Macmurroughs – Wexford; and MLF: Milford – Carlow The colour-coded indicates the heatmap scale from 1 to –1.
Figure 11.
River sediment nematode community composition. (a) The most frequently occurring nematode genera in the study sites and (b) Heatmap representing the abundances of the nine most prevalent nematode orders among the four locations. Location group name DHC: Dolmen Hotel – Carlow; GI: Great Island – Wexford; MCR: Macmurroughs – Wexford; and MLF: Milford – Carlow The colour-coded indicates the heatmap scale from 1 to –1.
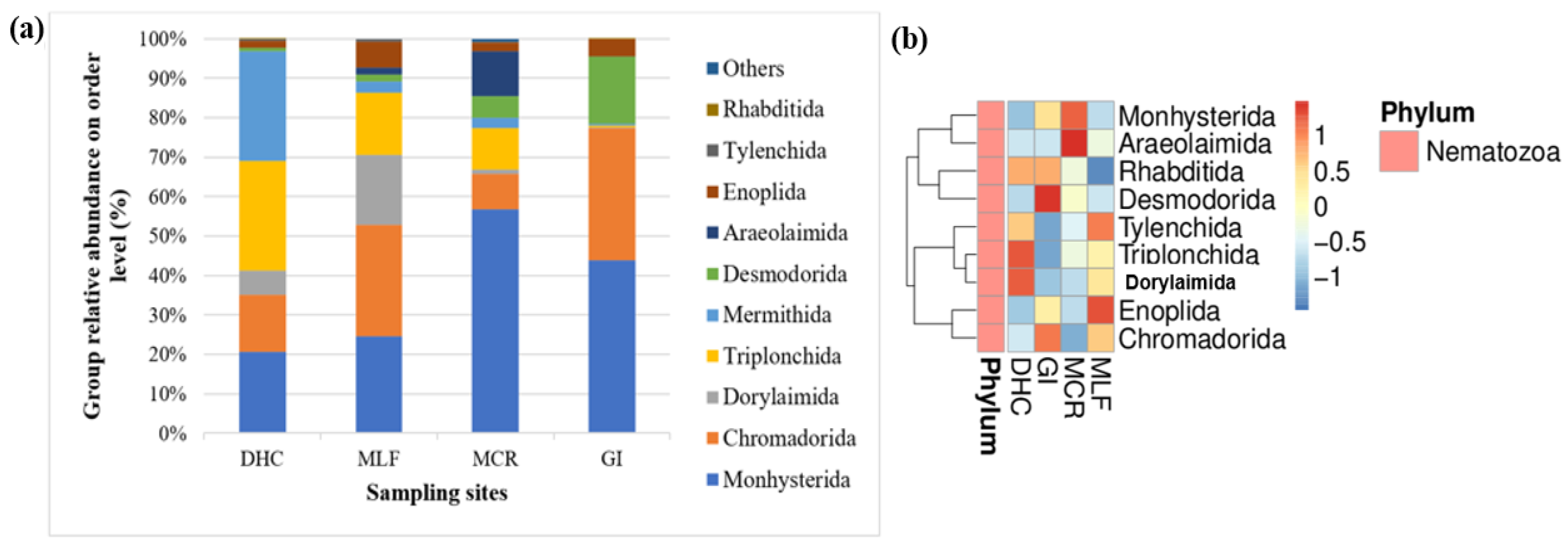
Figure 12.
Relative abundance of the various orders in the nematode communities at the various study sites. (a) The relative distribution of sequences in the nematode dataset within different nematode orders for each of the 20 analysed sediment samples and (b) Venn diagram displaying the overlap of shared OTUs among the four locations (at 97% similarity).
Figure 12.
Relative abundance of the various orders in the nematode communities at the various study sites. (a) The relative distribution of sequences in the nematode dataset within different nematode orders for each of the 20 analysed sediment samples and (b) Venn diagram displaying the overlap of shared OTUs among the four locations (at 97% similarity).
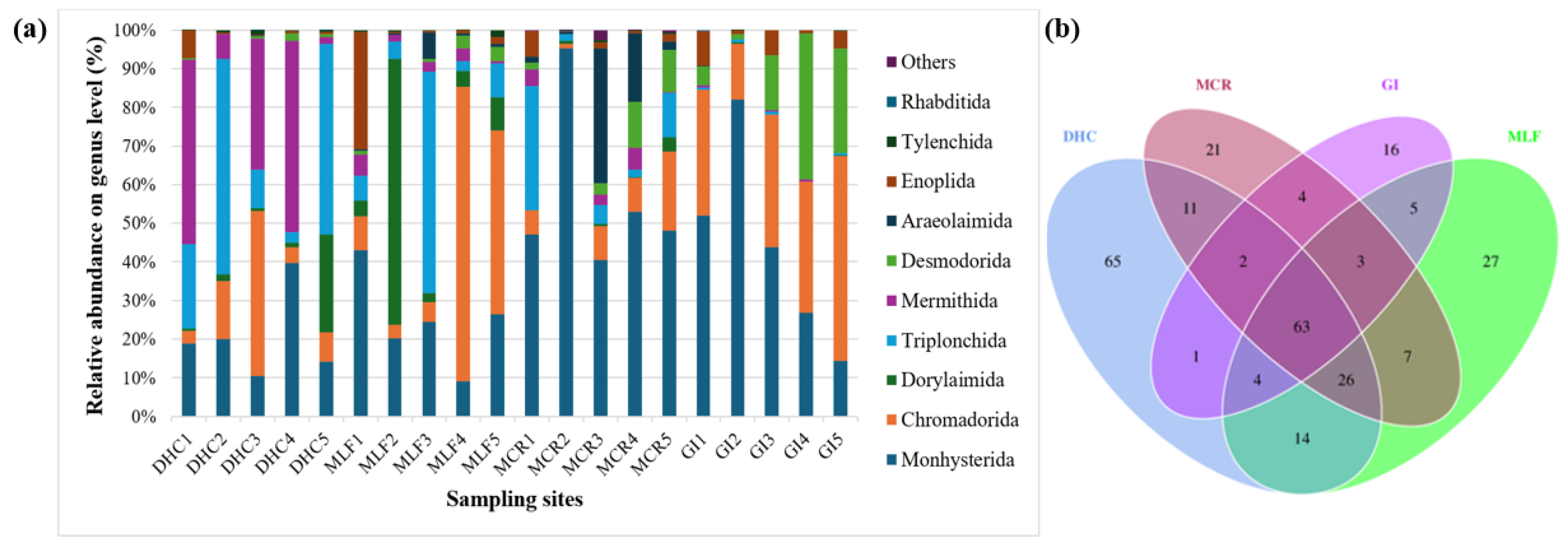
Figure 13.
Alpha diversities of the nematode communities associated with the river sediment samples from DHC, MLF, MCR, and GI; observed species, Chao1, Simpson , and Shannon are displayed in a, b, c, and d respectively.
Figure 13.
Alpha diversities of the nematode communities associated with the river sediment samples from DHC, MLF, MCR, and GI; observed species, Chao1, Simpson , and Shannon are displayed in a, b, c, and d respectively.
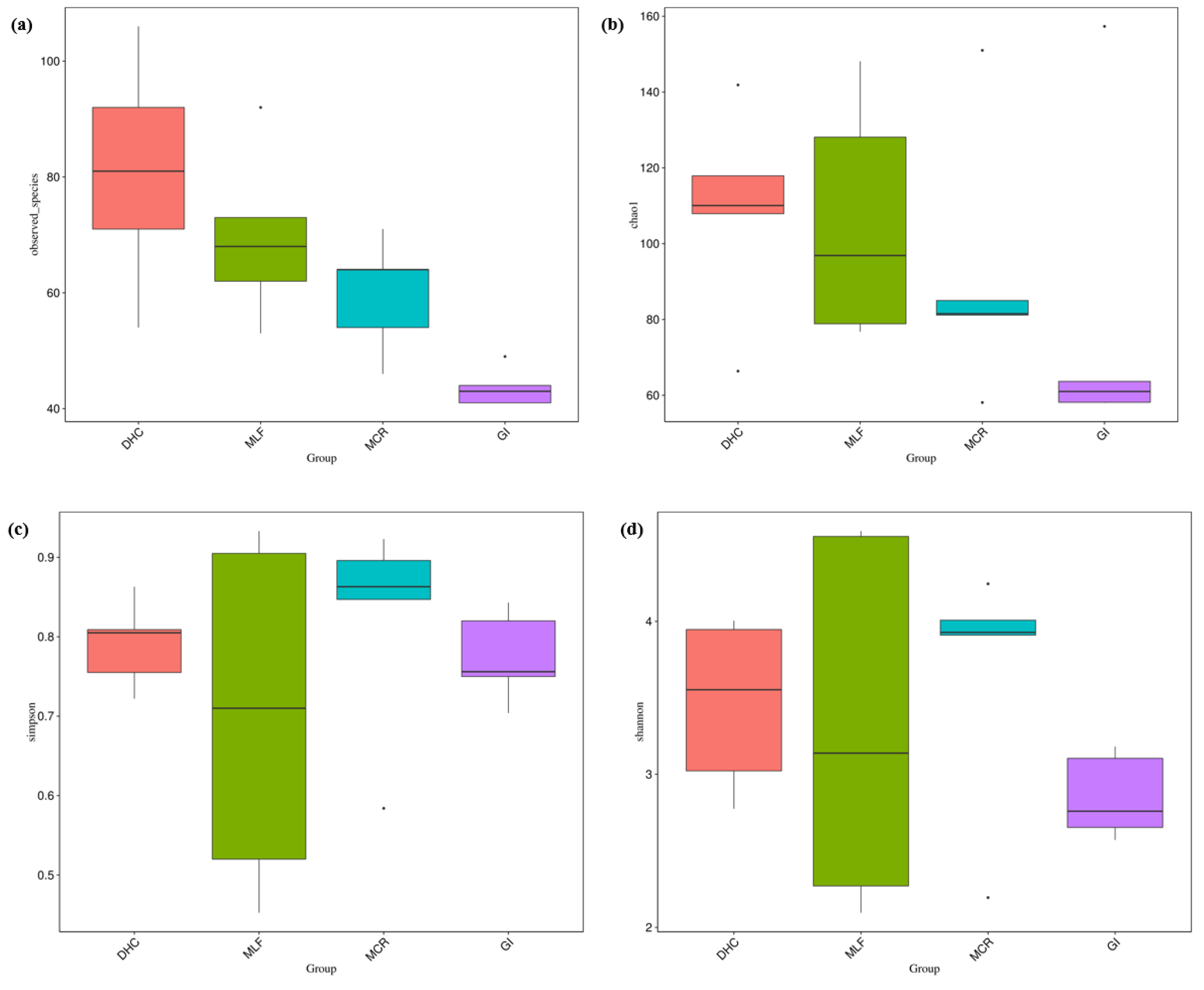
Figure 14.
Beta diversities of the nematode communities associated with the river sediment (a) Principal coordinate analysis (PCoA) of sediment nematode communities based on Bray–Curtis distances; PCoA plots are based on Bray-Curtis distances at the OTU level (97% sequence similarity). Red are Dolmen Hotel – Carlow samples, Green are Milford samples, Blue are Macmurroughs samples, and cyan are great island samples. (b) to (g) Anosim results of samples .
Figure 14.
Beta diversities of the nematode communities associated with the river sediment (a) Principal coordinate analysis (PCoA) of sediment nematode communities based on Bray–Curtis distances; PCoA plots are based on Bray-Curtis distances at the OTU level (97% sequence similarity). Red are Dolmen Hotel – Carlow samples, Green are Milford samples, Blue are Macmurroughs samples, and cyan are great island samples. (b) to (g) Anosim results of samples .
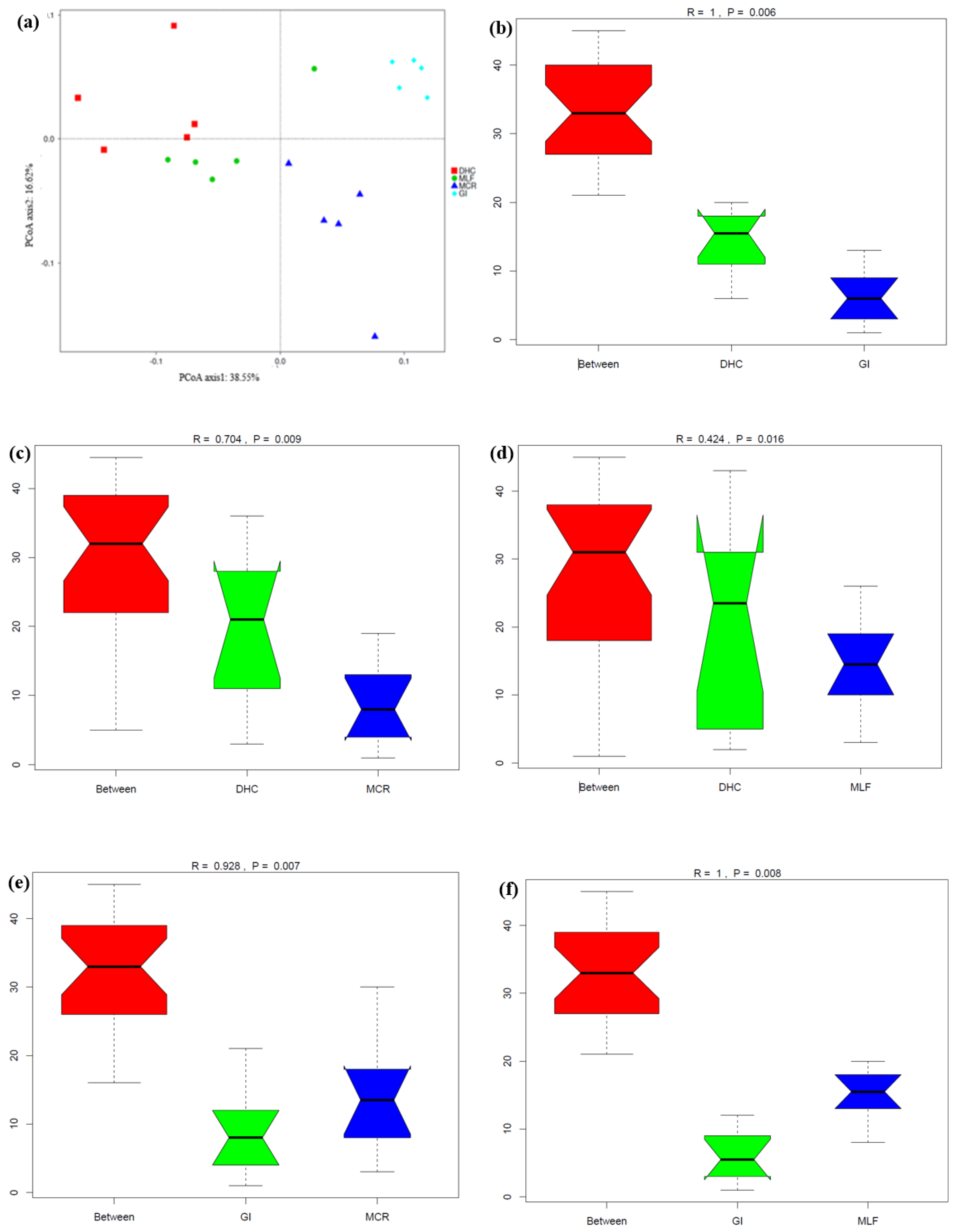
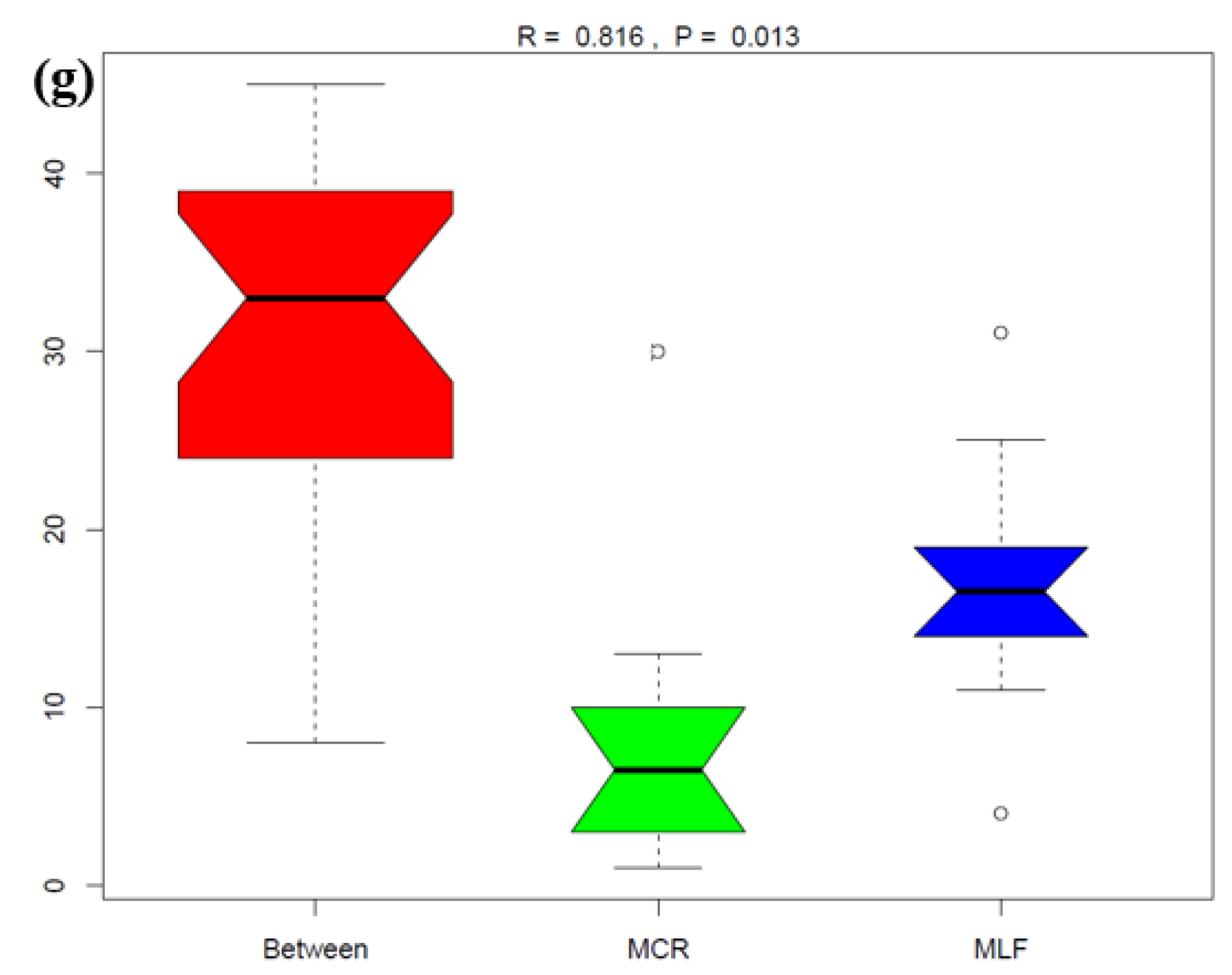
Figure 15.
Nematode abundance of various feeding (trophic) groups (guilds) across locations. Feeding type composition of nematode assemblage in the River Barrow sediment samples. Site A (DHC), site B (MLF), site C (MCR), and site D (GI). The percentage was determined by summing the abundances of all samples collected at each site and subsequently presenting the total cumulative abundance of all samples within each respective site.
Figure 15.
Nematode abundance of various feeding (trophic) groups (guilds) across locations. Feeding type composition of nematode assemblage in the River Barrow sediment samples. Site A (DHC), site B (MLF), site C (MCR), and site D (GI). The percentage was determined by summing the abundances of all samples collected at each site and subsequently presenting the total cumulative abundance of all samples within each respective site.
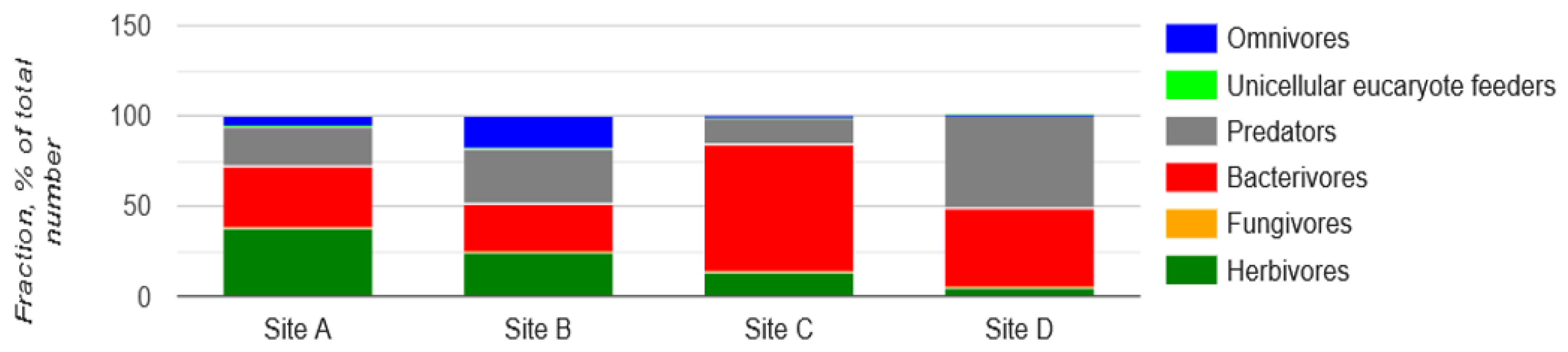
Figure 16.
Relative abundance (%) of nematode assemblage C–P groups at four different locations. The percentage was determined by adding up the abundances of all samples at each site and presenting the total cumulative abundance of all samples at each site.
Figure 16.
Relative abundance (%) of nematode assemblage C–P groups at four different locations. The percentage was determined by adding up the abundances of all samples at each site and presenting the total cumulative abundance of all samples at each site.
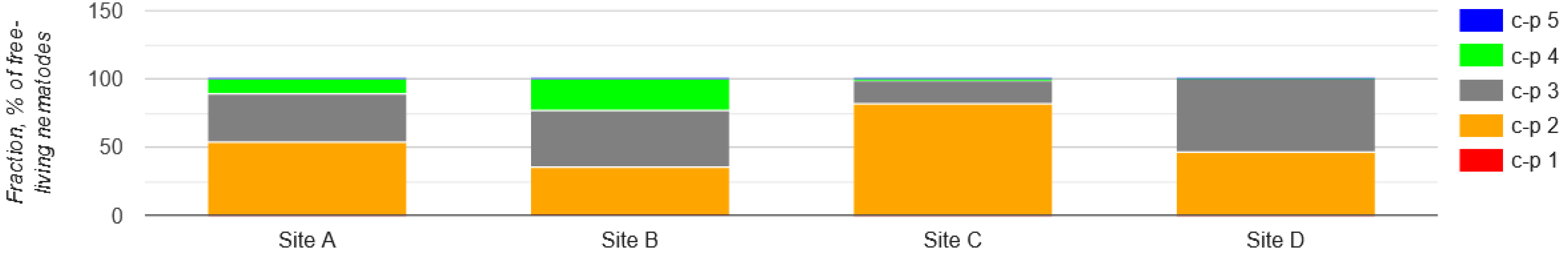
Figure 17.
Comparison of the mean (± SD) maturity index values of the four locations for nematodes.
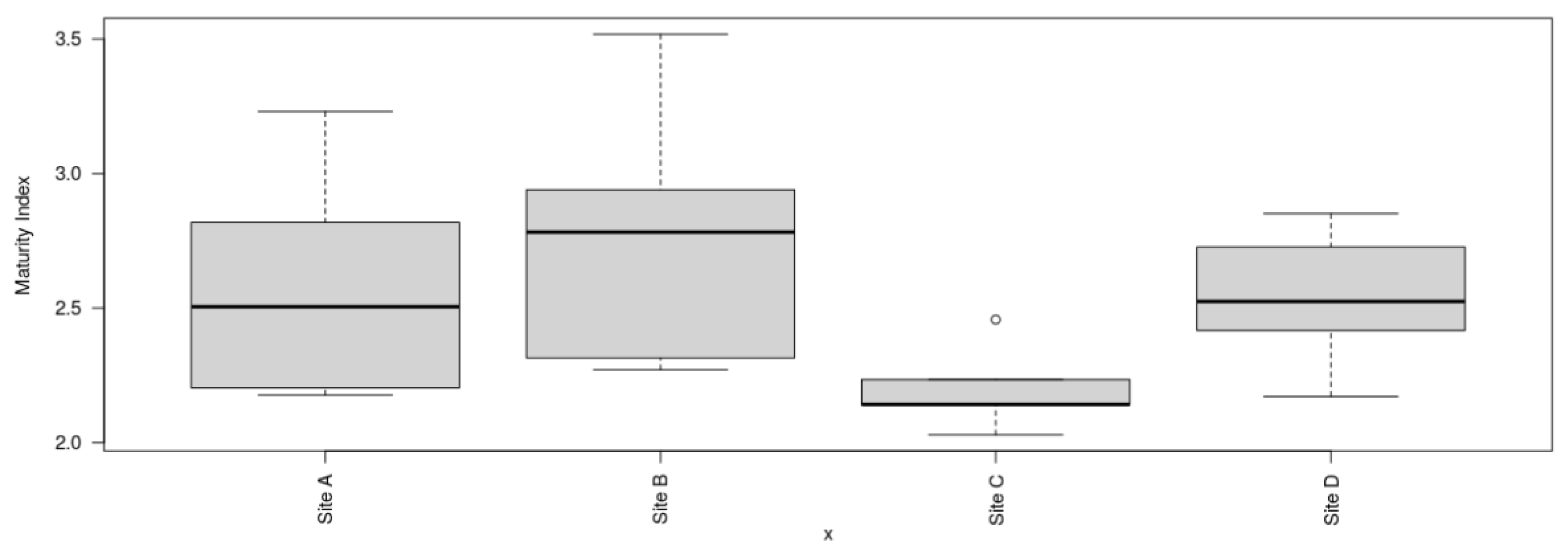
Figure 18.
Comparison of the mean sigma maturity index values of the four locations for nematodes.
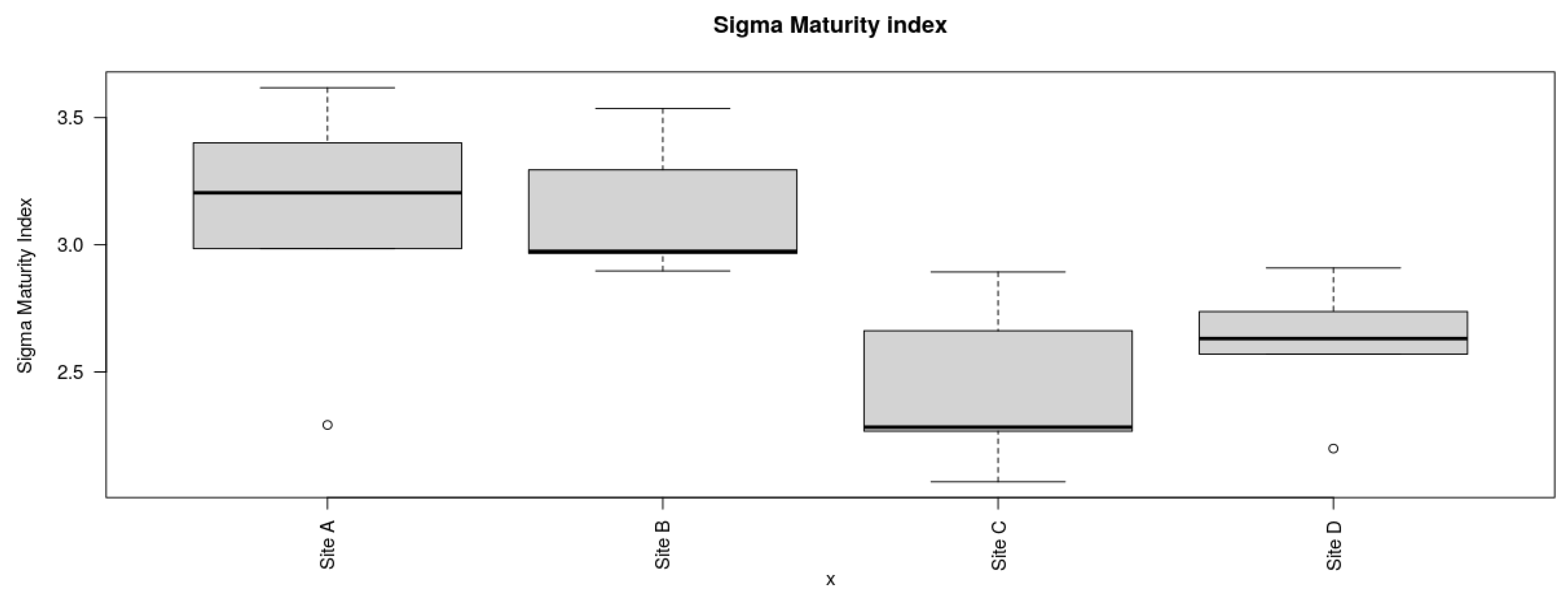
Figure 19.
Colonizer-persister triangle depicting river sediment status regarding nematodes c-p groups in the River Barrow.
Figure 19.
Colonizer-persister triangle depicting river sediment status regarding nematodes c-p groups in the River Barrow.
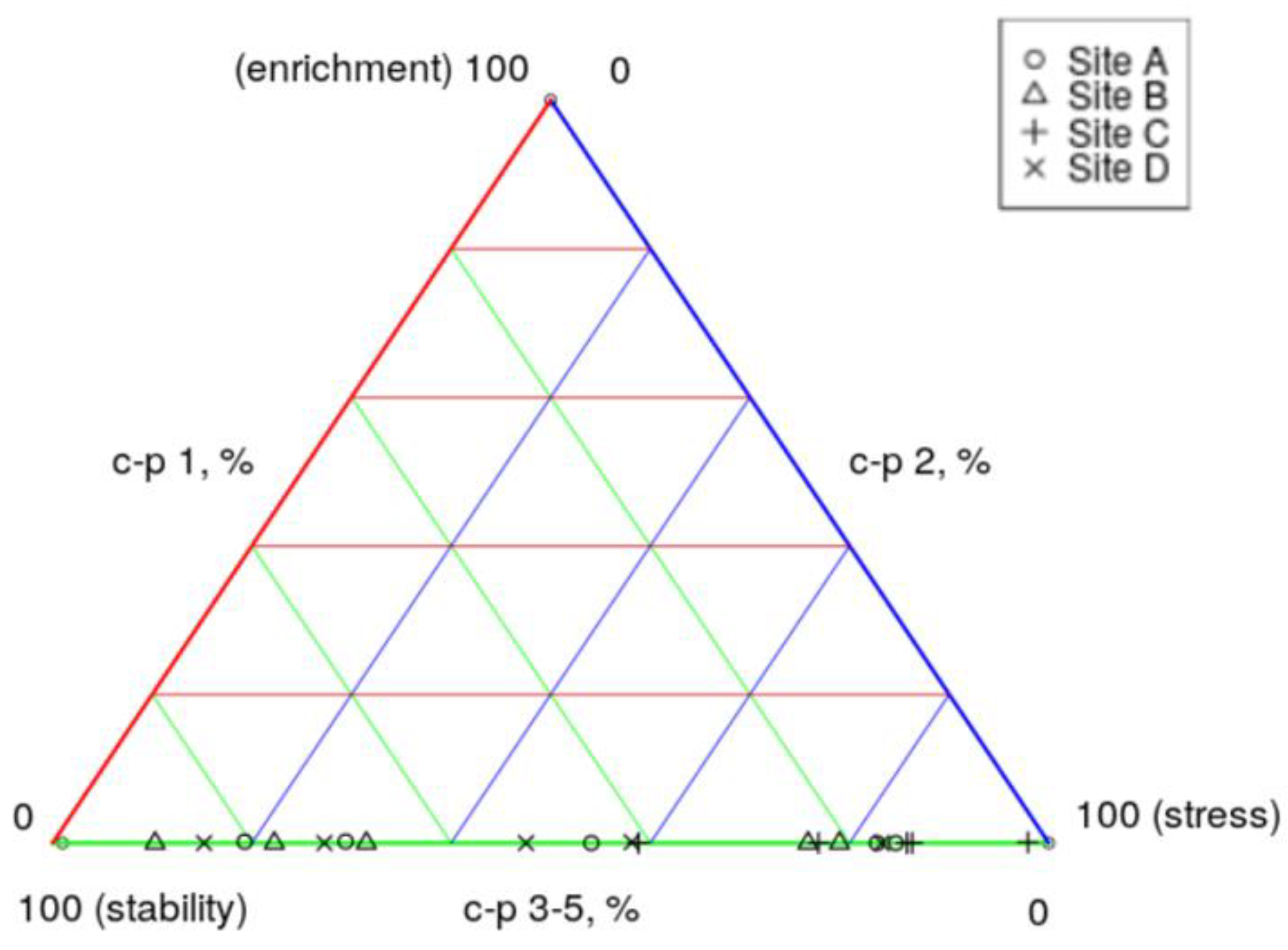
Figure 20.
(a) Food web analysis of nematode communities at different River Barrow locations sampled and (b) Food web analysis interpretation scheme.
Figure 20.
(a) Food web analysis of nematode communities at different River Barrow locations sampled and (b) Food web analysis interpretation scheme.
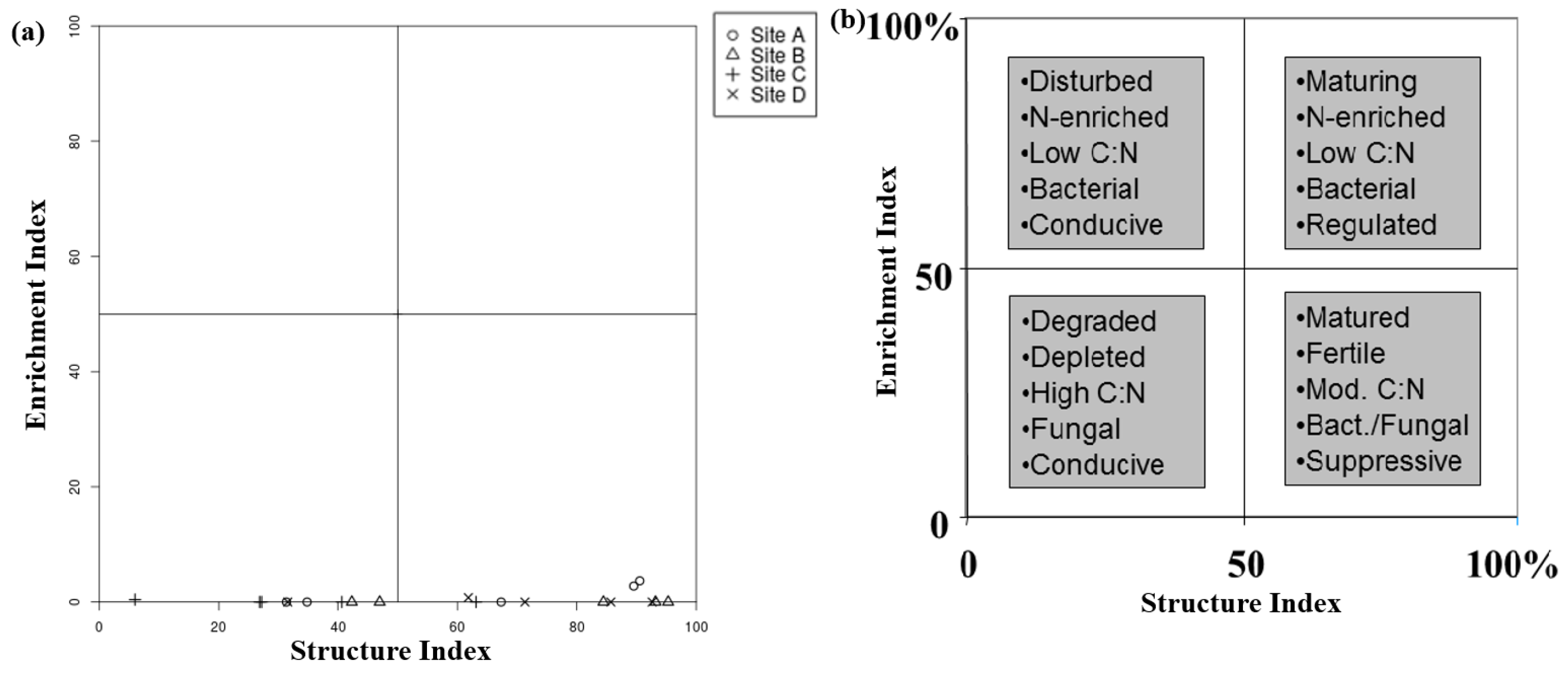

Table 1.
Nematode growth medium (NGM).
| Reagent | Amount to be added (1 L) |
|---|---|
| Bacteriological agar (Nº3) | 17.0 g |
| Bacteriological peptone | 1.25 g |
| Calcium chloride (CaCl2) | 1.0 mL |
| Sodium chloride (NaCl) | 3.0 g |
| 5 mg/mL Cholesterol in ethanol | 1.0 mL |
| Potassium phosphate buffer (KH2PO4/K2HPO4) pH 6.0 | mL |
Table 2.
Alkaline hypochlorite solution (20%) (15 mL).
| Reagent | Amount to be added (15 mL) |
|---|---|
| Sodium hydroxide (NaOH) | 3.75 mL |
| Bleach | 3.0 mL |
| Double deionised water | 8.25 mL |
Table 3.
Experimental design. The toxicity bioassays were run in 96 well plates.
 |
Table 4.
DNA concentration from Irish river sediment samples, and sampling coordinates (August 2021).
Table 4.
DNA concentration from Irish river sediment samples, and sampling coordinates (August 2021).
| Sampling coordinates | Sampling sites | MP Samples | Ds DNA (ng/µL) |
|---|---|---|---|
| 52°49'58.5"N 6°55'30.0"W | Dolmen Hotel - Carlow | 1 DHC | 56 |
| 2 DHC | 59 | ||
| 3 DHC | 51 | ||
| 4 DHC | 55 | ||
| 5 DHC | 57 | ||
| 52°47'04.7"N 6°57'60.0"W | Milford - Carlow | 1 MLF | 53 |
| 2 MLF | 59 | ||
| 3 MLF | 57 | ||
| 4 MLF | 39.4 | ||
| 5 MLF | 55 | ||
| 52°25'13.0"N 6°56'04.9"W | Macmurroughs - Wexford | 1MCR | 39 |
| 2 MCR | 31.7 | ||
| 3 MCR | 29.3 | ||
| 4 MCR | 40.9 | ||
| 5 MCR | 28.3 | ||
| 52°16'58.9"N 6°59'52.3"W | Great Island - Wexford | 1 GI | 46.9 |
| 2 GI | 42.3 | ||
| 3 GI | 45.4 | ||
| 4 GI | 42.6 | ||
| 5 GI | 49.4 |
Table 5.
ATR-FTIR peak data and assignments for polyethylene.
| Polymer | Absorption bands ((cm-1) | Assignment |
| Polyethylene (PE) | 2915 | C-H stretch |
| 2848 | C-H stretch | |
| 1472 | CH2 bend | |
| 717 | CH2 rock |
Table 6.
F and P-Value of nematode mortality in various treatments.
| Treatments | Exposure time (hours) | F | P-Value |
|---|---|---|---|
| Nematode medium, atrazine and MP | 96 | 0.964 | 0.527931 |
| Nematode medium, naphthalene and MP | 72 | 0.938 | 0.563178 |
| Nematode medium, naphthalene and MP | 96 | 1.954 | 0.008807 |
| Nematode medium, fluorene and MP | 72 | 1.645 | 0.039446 |
| Nematode medium, fluorene and MP | 96 | 0.639 | 0.913678 |
| Nematode medium, 1,3-dichloropropene and MP | 72 | 0.718 | 0.843136 |
| Nematode medium, 1,3-dichloropropene and MP | 96 | 0.948 | 0.548654 |
Table 7.
Alpha diversity indices of nematode communities from river sediment samples (n = 5). The data shows the averages and ± standard deviations.
Table 7.
Alpha diversity indices of nematode communities from river sediment samples (n = 5). The data shows the averages and ± standard deviations.
| Sample name | Observed number of species | Shannon | Simpson | Chao1 | ACE | Goods coverage |
|---|---|---|---|---|---|---|
| DHC | 80.8 ± 19.84 | 3.46 ± 0.55 | 0.7908 ± 0.054 | 108.82 ± 27.295 | 114.798 ± 34.818 | 0.980 ± 0.006 |
| MLF | 69.6 ± 14.57 | 3.33 ± 1.202 | 0.7042 ± 0.218 | 105.741 ± 31.369 | 107.27 ± 21.354 | 0.9818 ± 0.004 |
| MCR | 59.8 ± 9.81 | 3.66 ± 0.828 | 0.8226 ± 0.137 | 91.347 ± 35.021 | 81.037 ± 16.566 | 0.9874 ± 0.004 |
| GI | 43.6 ± 3.29 | 2.85 ± 0.274 | 0.7746 ± 0.056 | 79.628 ± 43.50 | 72.519 ± 15.127 | 0.9874 ± 0.003 |
Table 8.
Feeding types and colonizer-persistent class assignments of nematode taxa in river sediment.
Table 8.
Feeding types and colonizer-persistent class assignments of nematode taxa in river sediment.
| Family | c-p class | p-p class | Feeding type |
|---|---|---|---|
| Trichodoridae | 0 | 4 | Herbivores – ectoparasites |
| Tylenchidae | 0 | 2 | Herbivores – epidermal/root hair feeders |
| Monhysteridae | 2 | 0 | Bacterivores |
| Plectidae | 2 | 0 | Bacterivores |
| Rhabditidae | 1 | 0 | Bacterivores |
| Achromadoridae | 3 | 0 | Predators |
| Cyatholaimidae | 3 | 0 | Predators |
| Dorylaimidae | 4 | 0 | Omnivores |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



