
Submitted:
23 October 2024
Posted:
23 October 2024
You are already at the latest version
Abstract
Various methods have been studied to speed up the wound healing process. Promoting the rapid development of blood vessels, also known as angiogenesis, is essential as this significantly accelerates skin regeneration. This study identified a polysaccharide from the red alga Osmundea pinnatifida (PSOP) that promotes angiogenesis and accelerates wound healing. Firstly, we analyzed the structural characteristics of PSOP using various techniques such as UV-visible spectrum, Fourier-transform infrared (FT-IR) spectroscopy, X-ray diffraction (X-RD), scanning electron microscopy (SEM), and high-performance liquid chromatography coupled to refractive index detector (HPLC-RID). We also evaluated the antioxidant properties of PSOP in vitro using tests for DPPH free radical scavenging capacity, iron reduction capacity (RPA), and total antioxidant capacity (TAC). Additionally, we observed a significant increase in new blood vessel formation in the chorioallantoic membrane model (CAM assay) using a fertilized hen egg, indicating the pro-angiogenic effect of PSOP.
Furthermore, we tested the wound-healing potential of PSOP hydrogel in vivo using excision wounds in the Wistar rat model. The results showed a notable acceleration of wound closure and an improvement in tissue regeneration and collagen deposition quality within 11 days, indicating the strong wound-healing activity of PSOP. Our computational study suggested that the PSOP units bind to cyclooxygenase 2 (COX-2) and vascular endothelial growth factor (VEGF) with acceptable affinities and interactions, which might explain the observed beneficial wound healing properties and the pro-angiogenic effect seen in the experimental assays. Our findings support using the polysaccharide PSOP for wound treatment due to its antioxidant and pro-angiogenic potential.
Keywords:
Osmundea pinnatifida
; Polysaccharide
; Wound healing
; Angiogenesis
; CAM assay
; molecular interactions
1. Introduction
Wounds, whether caused by surgery, burns, cuts, or severe traumatic lacerations, require effective healing. Rapid restoration of microvascular connectivity is essential to reduce ischemia and hypoxia, facilitating tissue repair and restoration processes [1]. In general, acute wound healing can take up to four weeks and involves hemostasis, inflammation, proliferation, and remodeling [2]. Furthermore, wounds are caused by a disturbance in angiogenesis, one of the leading causes of delayed wound healing. Wound healing is promoted by improved angiogenesis, which reduces excessive inflammation and provides damaged tissue with the oxygen and nutrients essential for regeneration [3]. Disturbances at any of these stages can lead to the formation of chronic wounds, such as pressure sores, diabetic ulcers, and vascular wounds. These wounds increase treatment costs and are characterized by persistent inflammation, resistance to therapies, and insensitivity of cells to healing signals, ultimately leading to a significant increase in morbidity and mortality [4], [5].
Researchers have been studying bioactive compounds from marine algae for their potential therapeutic uses in various diseases. Algal polysaccharides, in particular, have drawn significant interest due to their diverse biological activities, including anti-inflammatory, anticoagulant, antitumor, antiviral, and antioxidant properties [6], [7], [8], [9]. In addition, it has adhesive properties, is biodegradable, non-toxic, and has various physical and chemical characteristics [10]. Lately, inexpensive natural polysaccharides used in skin therapeutics have attracted attention for their effectiveness in wound healing [11] by promoting cell regeneration and collagen synthesis.
Osmundea pinnatifida, synonym Laurencia pinnatifida (Rhodobionta, Rhodophyta), is a red macroalgae, widespread (Class: Florideophyceae; Order: Ceramiales; Family: Rhodomelaceae). It’s edible for human food and dietary supplements because it contains vitamins, proteins, amino acids, fiber, and fatty acids [12]. Therefore, it represents an essential source of agar production [13], which can be investigated in the food fields. A present review highlights the biotechnological potential of these algae, including their antitumoral, antiviral, antiprotozoal, antibacterial, and antifungal activities [14]. Also, previous studies have revealed that this alga’s aqueous and hydro ethanol extract presents an antioxidant and anti-aging activity [15]. An enzymatic extract of O.pinnatifida has demonstrated important biological properties, such as antioxidant and anti-diabetic [16]. However, polysaccharide extracts from this alga remain primarily under-exploited, and their biological applications have yet to be thoroughly investigated.
The current study was designed to extract a polysaccharide from the red alga Osmundea pinnatifida, named PSOP. Firstly, the structural characteristics of the polysaccharide have been investigated. The characterization of PSOP was determined using chemical and instrumental analysis, high-performance liquid chromatography (HPLC-RID), UV-visible spectrum, FT-IR spectroscopy, X-ray diffraction (X-RD), and scanning electron microscopy (SEM). Molecular weight, total sugars, sulfates, and proteins were then determined. The in vitro antioxidant activities of PSOP were assessed by a DPPH Free radical assay, as well as total antioxidant activity and reducing power. In the second part, the pro-angiogenic potential of PSOP has been demonstrated in ovo via chicken embryo chorioallantoic membrane (CAM) and validated in vivo by accelerating the wound healing process using an excision model in Wistar rats. Additionally, the PSOP units were further explored using a computational approach, mainly by assessing binding affinities and molecular interactions by targeting cyclooxygenase 2 (COX-2) and vascular endothelial growth factor (VEGF).
2. Material and Methods
2.1. Algae Collection and Identification
Osmundea pinnatifida was collected from the Atlantic coasts of Sidi Bouzid (El Jadida, Morocco; 33°13’N-8°55’W) (Figure 1) between March and May 2023. The algae were identified at the laboratory of Marine Biotechnology and Environment, Faculty of Science of Chouaib Doukkali University. They were carefully washed, shade dried, milled, and stored in amber glass bottles at 4°C.
2.2. Extraction of the Polysaccharide-Rich Osmundea pinnatifida (PSOP)
Red algae pigments were dissolved in 95 % ethanol before the PSOP extraction, according to Lui et al. [17]. One hundred grams of dried seaweed powder were stirred for 4 hours with distilled water at 90 °C. The crude water extract was filtered, vacuum dried, and precipitated for 24 hours at 4 °C in 95 % ethanol (3v/v). The insoluble polysaccharide-rich was pelleted by centrifugation (6000 rpm, 20 min, 4 °C) using a refrigerated benchtop centrifuge (Hettich, Rotina 380R). For three days, PSOP was resuspended in distilled water and lyophilized (Christ Alpha 1-2 lyophilizer, Bioblock Scientific). The resulting extract, PSOP, was stored at 4 °C until use.
2.3. Chemical Characterization
The yield was expressed in the ratio of the dry weight of the polysaccharide extracted (g) against the dry weight of Osmundea pinnatifida (g) in percentage. Total carbohydrate content was determined using the phenol-sulfuric acid colorimetric method [18], protein concentration was determined using the method of Lowry et al. [19], and a turbidimetric method was used to quantify the sulfate content using the barium chloride/gelatin reagent [20].
2.4. Polysaccharides Spectroscopic Analysis
2.4.1. Ultraviolet Absorption Peak Detection
The polysaccharide was prepared in distilled water to a final concentration of 1 % (w/v). The solution’s UV-vis absorption spectrum was registered in the 200-800 nm wavelength range using a UV-vis spectrophotometer (JENWAY/7315, UK) [21].
2.4.2. Fourier Transform Infrared Spectrometry Analysis
The PSOP powder was applied to a Nicolet FT-IR spectrometer fitted with an attenuated reflection attachment (ATR) containing a diamond/ZnSe crystal. The spectrum was acquired with a resolution of 4 cm-1, and the measurement range was 400 to 4000 cm-1 at room temperature (25 °C). Data analysis and processing were carried out using OPUS (Bruker, Ettlingen, Germany) [22].
2.4.3. Monosaccharide Analysis by High-Performance Liquid Chromatography (HPLC-RID)
Monosaccharide compositions were analyzed using the method [23]. A 5 mg sample was dissolved in 3 mL TFA (2 mol/L) and hydrolyzed at 110 °C for 3h. The TFA was removed by washing with methanol. The hydrolyzed product was then reduced by adding 50 mg sodium tetrahydruroborate. Pyridine and acetic acid anhydride were added to the mixture at 40 °C for 2 h for acetylation. The acetylated sample was filtered and examined using high-performance liquid chromatography (LC1260 Infinity II), equipped with an Aminex HPX-87C column with H2SO4 at (0.001 N) as mobile phase, programmed with a flow rate of 0.4 mL/min and a column temperature of 60 °C. The standard sugars utilized were glucose, fructose, xylose, mannose, and glucuronic acid.
2.4.4. The Average Molecular Weight
According to Bayar et al. [24], the average molecular weight (Mw) was determined by steric exclusion chromatography (SEC).
2.4.5. Scanning Electron Microscopy
PSOP surface mapping, microstructure assessment, and morphology analysis were investigated using cryogenic scanning electron microscopy (JSM-IT500HR, JEOL).
2.4.6. X-Ray Diffraction
The XRD diffractogram of PSOP was performed using an X-ray diffractometer (D2 PHASER, Bruker, German, Berline) at room temperature. The measurement was conducted in the 2 ranges from 10 to 80 ° (2θ) with a step size of 0.02 ° and a counting time of 5 s/step.
2.5. Antioxidant Activity of Polysaccharide
2.5.1. DPPH Free Radical Scavenging Assay
DPPH radical scavenging activity was measured according to [25]. PSOP at different concentrations (1-10 mg/mL) was incubated with 0.02 % DPPH (in ethanol). DPPH radical reduction was measured at 517 nm. The standard used was butylated hydroxyanisole (BHA). DPPH radical scavenging capacity was calculated with the following equation:
where AC, AB, and AS refer to the optical densities of blank, control, and sample, respectively.
2.5.2. Total Antioxidant Activity
Total PSOP antioxidant activity was assessed according to the protocol in [26]. Absorbance was measured at 695 nm against a blank. The standard used was butylated hydroxyanisole (BHA).
2.5.3. Reducing Power Assay
The ability of PSOP to reduce iron was determined according to the method of [27]. The absorbance of the resulting solution was measured at 700 nm. Butylated hydroxyanisole (BHA) was used as the standard.
2.6. Chorioallantoic Membrane Assay
Angiogenesis activity was assessed in ovo using the chorioallantoic membrane (CAM) test with some modifications [28]. The experiment presented in the study does not require approval from the ethics committee [29].
In summary, fertilized chicken eggs from the Eurafric Poussins Company in Had Soualem, Morocco, were first cleaned with 10% betadine and then placed in an incubator at 37°C with 55-65% humidity for 3 days. The eggs were manually rotated three times a day to ensure proper development of the chicks. On the fourth day, 3 mL of albumin was removed using a sterile syringe with a 20-gauge needle, and the incubation process continued. On the eighth day, a small opening was created on the larger side of the egg, and a 1 cm² window was carefully made under a laminar flow hood. The chorioallantoic membrane was then removed without causing harm to the blood vessels. Then, the eggs were divided into 4 experimental groups (n = 6/treatment):
Group 1: eggs were treated with sterile distilled water (the control);
Group 2: eggs were treated with diclofenac DIC (50 μg/egg), which was used as a negative control;
Group 3: eggs were treated with choriogonadotropin CG (30 μg/egg), which was used as a positive control;
Group 4, 5, and 6: eggs were treated with various concentrations (25, 50, and 100 μg/egg) of PSOP;
The eggs were re-incubated for three days after sealing the windows with transparent adhesive tape. On day 12, the neovascular regions of the CAM were photographed using a stereomicroscope (Steroblue, EUROMEX).
2.7. Quantitative Analysis of Vascular Network
The photomicrographs obtained from the CAM vessels were analyzed using ImageJ software (version 1.54, National Institutes of Health, USA) and AngioTool software (version 0.6a, National Cancer Institute, USA). To quantify the topography of the vascular network, using the software ImageJ, areas of 500 mm2 were selected; the images were converted to grayscale and, in sequence, "inverted" from the plugin "edit." After that, the background was extracted from the photos (a value of 50 pixels was fixed) in the "process" plugin. The photomicrographs were saved and then opened using the AngioTool software. A typical image displaying the network topology was detected, formed by vessel area, vessel length, vessel diameter, number of junctions, and lacunarity (avascular space).
2.8. In Vivo Experimental Study
2.8.1. Animals
Twenty-four healthy adult male Wistar rats weighing 150-200 g were obtained from the Pasteur Institute of Tunis, Tunisia. The rats were kept in a ventilated room, in individual cages at room temperature (22-24 °C), with a 12-hour light and 12-hour dark cycle and free access to food and water.
2.8.2. Experimental Protocol
The excision model was used to assess the extent of wound contraction. Twenty-four rats were divided into four control and test groups:
Group 1: was treated with saline (0.9 %) and used as a control group.
Group 2: was treated with a standard drug, "Cytol Centella" cream, used as a medicated reference cream.
Group 3: was treated with PSOP hydrogel (lyophilized PSOP was mixed in 30% glycerol to obtain a final concentration of 15 mg/mL).
Group 4: was treated with glycerol (30 %).
After cleaning the rats’ skin with sterile water, it was disinfected with swabs soaked in 70 % alcohol. A light shave was performed on the back of the interscapular skin area. In brief, a circular wound of approximately 150 mm2 (1.5 cm in diameter) was made using a sharp surgical blade. All wounds were cleaned and treated every two days with sterile saline, vehicle, PSOP, and the reference drug "Cytol Centella" in a thin layer covering the wound surface until the wound was completely healed. On day 12, after wounding, all rats were anesthetized with ether and sacrificed by decapitation. Part of the re-epithelialized tissue was preserved for hydroxyproline estimation, and another part was fixed in 10 % (v/v) formalin, embedded in kerosene, and processed for histological observation.
2.8.3. Measurement of Wound Area and Burn Contraction Rate
Photographs were taken of the wound surface on days 1, 5, 7, and 11 for evaluating the healing progress. The wound surface was measured using Autodesk AutoCAD software. The rate of wound contraction can be calculated using the equation below:
2.8.4. Determination of Hydroxyproline Content
Hydroxyproline concentrations were calculated using the method of [30]. In brief, the wound biopsies were dried at 60 °C until constant weight was obtained. Then, they were hydrolyzed with 6N HCl for 3h at 120 °C. The hydrolyzed specimens were subjected to chloramines-T-oxidation and stored at room temperature for 20 min. The color developed with the presence of Ehrlich’s reagent (2.5 g of 4-dimethylamino benzaldehyde, 9.3 mL of n-propanol, and 3.9 mL of 70 % perchloric acid) at 60 °C was measured at 557 nm after cooling. The results were reported as mg/g dry weight of tissue.
2.8.5. Histological Examination
On the day one of the groups completely healed, the rats were sacrificed, and the wound site biopsies were collected and fixed immediately in formalin solution (10 %). They were then embedded in kerosene, stained with hematoxylin-eosin, and examined with a light microscope (Olympus CX41, Tokyo, Japan).
2.8.6. Statistical Analysis
The results obtained were expressed as means ± standard deviation and were analyzed with SPSS using analysis of variance (ANOVA). When the ANOVA revealed a significant effect, the Tukey test post hoc compared the values. Differences were considered statistically significant at p < 0.05.
2.9. Computational Modeling and Interactions Assay
In an attempt to rationalize the potential effect of the pro-angiogenic and wound healing of the identified O. pinnatifida monosaccharides, a computational modeling and interactions assay was realized. In this context, the binding affinity of all the identified phytochemicals with some key macromolecules involved in both pro-angiogenic and wound healing have been assessed. The crystal structures of cyclooxygenase 2 (COX-2, pdb id: 1cx2) and vascular endothelial growth factor (VEGF, pdb id: 2c7w) have been collected from RCSB databases. These macromolecules have been prepared by the deletion of crystalized water molecules and the addition of both polar hydrogen and Koleman charges… before being processed for CHARMm force field as previously described ([31], [32], [33]). The established bond categories and molecular interactions have been recorded and analyzed ([34], [35], [36]).
3. Results and Discussion
3.1. Chemical Characterization of PSOP
In this study, the yield of PSOP was estimated to be 32.65 %. According to Arunkuma et al. [37], crude polysaccharides isolated from L. papillosa and L. obtusa yielded around 4 % and 6 %, respectively. Polysaccharide yields vary according to species, environmental conditions, and extraction methods [38], [39]. The total sugar content of PSOP was about 88.12 % ± 1.1 %, which was higher than what was reported for algae L. pedicularioides and L. cruciata (44.8 % and 49.4 %, respectively) [40]. The results of various colorimetric tests showed that the polysaccharide analyzed contained low levels of protein (0.18 % ± 0.02 %) and low levels of sulfate (0.69 %), proving the extraction method’s precision. The polysaccharide obtained by ethanol precipitation frequently contains contaminants, such as proteins, which require purification by deproteinization [41]. However, this process can alter carbohydrate yields, modify monosaccharide composition, influence structural parameters, and affect polysaccharide bioactivity [42], [43]. Following on from the proteins discussed above, it’s worth pointing out that sulfate groups linked to polysaccharides also influence crucial biological functions, such as antitumor, anticoagulant, and immunomodulatory effects [44].
3.2. Spectroscopic Analysis and Monosaccharide Composition
Figure 2A illustrates the UV-visible spectra of PSOP. Our finding revealed a maximum absorption peak between 200 and 240 nm, which confirms that our sample is a polysaccharide [45]. A weak peak was observed at 260-280 nm, which indicates the presence of the protein trace or nucleic acid covalently linked to PSOP [46]. An absorption area identified at around 300 nm was attributed to trace pigments.
As is well known, FT-IR spectroscopy can be used to identify structural features of polymer blends, such as distinct organic groups in the polysaccharide. Figure 2B shows that PSOP exhibited absorption peaks typical of polysaccharides between 700 and 4000 cm-1. A broad hydroxyl group O-H stretching vibration peak was recorded at around 3389 cm-1 due to intra- and intermolecular hydrogen bands. Subsequently, a weak peak detected at about 2929 cm-1 was attributed to the C-H stretching vibration of sugars [47], indicating that the sample was a polysaccharide compound. Peaks appearing at about 1637 and 1412 cm-1, respectively, were attributed to carboxylate bond stretching, demonstrating that PSOP was an acidic polysaccharide [48]; this suggests the presence of uronic acids, an observation corroborated by analysis of monosaccharide composition. The hydroxyl and carboxyl groups of polysaccharides are essential to their biological properties, notably to enhance their antitumor and antioxidant effects [49]. In addition, the weak absorbance peak at 1369 cm-1 was attributed to ester sulfate S=O groups [50]. Absorption peaks between 1136 and 1047 cm-¹ were associated with C-O-C and C-O-H stretching vibrations, characteristic of the pyranose ring structure [51]. Combined, weak absorptions around 800 and 900 cm-1 originate from the stretching vibration of the C-O-S [52], indicating the existence of α and β configurations in the polysaccharide.
Figure 3A represents the HPLC-RID chromatogram of the monosaccharides detected in the PSOP. The analysis by high-performance liquid chromatography with a refractive index detector revealed the presence of the different monosaccharides, in particular, glucuronic acid, arabinose, glucose, xylose, and fructose, at retention times of 8.32, 9.39, 11.19, 12.13 and 13.64 min, respectively, by elution time of monosaccharide standards Figure 3B. Further research reveals that red algae polysaccharides are made up of a heterogeneous combination of monosaccharides. For example, the polysaccharide obtained in enzyme extracts from the alga O. pinnatifida was composed of galactose, mannose, arabinose, xylose, rhamnose, and fucose [53]. The monosaccharide composition of Laurencia dendroidal polysaccharide is mainly galactose, with smaller amounts of xylose, mannose, and glucose [54]. However, several exogenous factors such as protocol, water concentration, extraction temperature, and endogenous factors in the organisms (type of species, reproductive cycle...) can influence the polysaccharide composition [55], [56]. Indeed, the nature and proportion of monosaccharides present represent a key element affecting the activity of polysaccharides, which can confer new bioactive properties or reinforce existing ones [57].
The molecular weight (Mw) profile was analyzed by steric exclusion chromatography (SEC), revealing that PSOP had a molecular weight of 146 kDa, with a retention time of 7.25 min (Figure 4). In reality, the molecular weight of polysaccharides has a considerable influence on their biological activities. In a study by Lee and his collaborators [58], a low-molecular-weight polysaccharide (3 kDa) was obtained by controlled degradation of a high-molecular-weight polysaccharide (2238 kDa). The results showed that the low-molecular-weight polysaccharide possessed superior antioxidant activity and more effectively stimulated the immune response in specific organs subjected to oxidative stress than the higher-molecular-weight polysaccharide.
Scanning electron microscopy (SEM) is a technique that produces images of the surface of polysaccharides to analyze their structure, including shape, size, and porosity [59]. Scanning electron micrographs of PSOP are illustrated in Figure 5A. It was observed that PSOP under 10- and 20-fold magnification had a microstructure in the form of a network with many cavities and had a highly developed porosity over the entire surface area with some heterogeneity, which explains an improved ability to supply oxygen and absorb exudates; a valuable feature from a pharmacological and cosmeceutical point of view [60].
X-ray diffraction is a technique mainly used to identify polysaccharides’ crystalline or amorphous structure [61]. Figure 5B illustrates that the XRD diffractogram of PSOP, over a diffraction range of 0-80°, displays a broad diffuse band with a dominant peak at around 32° (2θ), accompanied by several other less pronounced peaks. This distribution suggests that PSOP has a semi-crystalline structure in an amorphous matrix. The degree of crystallinity of polysaccharides strongly influences their properties, such as mechanical strength, solubility, and swelling capacity [62]. For example, semi-crystalline to crystalline polysaccharides have improved water retention capacity, making them ideal for applications such as hydrogels, biofilms, nanofibers, membranes, and the medical field [63].
3.3. Antioxidant Activity of PSOP
Seaweed is increasingly seen as a natural source of antioxidant compounds [64]. Such components can be used in the cosmetics, pharmaceutical, and food industries to protect products from oxidative damage and further deterioration [65]. Three tests were used to assess the reducing and scavenging capacity of PSOP extracted from Osmundea pinnatifid algae: DPPH free radical scavenging capacity, iron reduction capacity (RPA), and total antioxidant capacity (TAC). As shown in Figure 6, the antioxidant capacity of PSOP is critical and increases with polysaccharide concentration. The data shown in Figure 6A revealed that the extract at high concentration (5 mg/mL) exhibited a free radical scavenging activity against DPPH of around 68.9 %. However, BHA shows a high activity of around 97 % at a 0.5 mg/mL concentration. The antioxidant capacity of polysaccharides may be linked to the presence of functional groups such as hydroxyls and carboxyls and to their sulfate content, which scavenges the free radical DPPH [66], [67].
The reducing power test is used to assess the antioxidant efficacy of various substances [68]. As shown in Figure 6B, the data reveal that PSOP exhibited a reducing power that correlated with increasing concentrations. A higher absorbance value indicates a more robust reducing capacity. The PSOP polysaccharide showed maximum activity at a concentration of 7 mg/mL (1.33 at OD 700 nm). Nevertheless, this activity was lower than BHA 2.9 at OD 700 nm. The reducing capacity of polysaccharides remains dependent on bioactive elements such as uronic acids, glucuronic acids, and other monosaccharide units [69].
The results of the total antioxidant capacity (TAC) of PSOP in the phosphorus-molybdenum assay are shown in Figure 6C. The data reveal that PSOP exhibited different degrees of activity, which increased with increasing concentration. However, BHA showed higher activity than PSOP at all the concentrations tested. For example, at a concentration of 1 mg/mL, BHA was able to reduce molybdenum Mo (VI) to Mo (V) with a level of 180 μmol/mL α-tocopherol compared to 49 μmol/mL α-tocopherol for PSOP at the same concentration. According to Abdelhedi et al. [70], efficiency (TAC) could be associated with the sulfate levels present in polysaccharides. In addition, Gunasekaran and collaborators have also demonstrated that sulfonic groups in polysaccharides can act as electron donors, reacting to transform free radicals into more stable compounds [71].
3.4. Angiogenesis Stimulation Assays
The chorioallantoic membrane (CAM) test is frequently used to examine different molecules’ angio-inhibitory and angio-stimulatory potential [72]. Our results have shown that PSOP action is clear and potent. It potentially caused an increase in the number of vessel branches according to the dose. Figure 7I and Table 1 show that CAM treated with different concentrations of PSOP revealed well-developed areas of neovascularization (137.22 % ± 3.7 %, 200.66 % ± 2.73 % and 250.66 % ± 3.49 % vessels for 25, 50 and 100 μg/egg, respectively) compared with the group which was not treated (set at 100 %) (p < 0.05). Diclofenac (DIC), used as an anti-angiogenic agent to validate the CAM model [73], demonstrated an ability to inhibit the process of new blood vessel formation, thereby reducing neovascularization (71.14% ± 3.97%) compared to the untreated group (p < 0.05). Choriogonadotropin (CG), used as a positive control for its pro-angiogenic role [74], significantly increased blood vessel formation (168.36% ± 2.72%) compared to the untreated group (p < 0.05). Since angiogenesis is a critical factor that promotes new blood vessel formation after chronic or ischemic wounding [75]. Angiogenesis involves phenomena such as hypoxia or inflammation, which lead to the recruitment of inflammatory cells and the release of growth factors that promote the creation of new blood vessels [76]. Among the various factors involved in angiogenesis, tumor necrosis factor-alpha (TNF-α) stands out as a pro-inflammatory cytokine. This cytokine acts on endothelial cells, producing vascular endothelial growth factor (VEGF) [77]. VEGF promotes cell proliferation, activates the expression of specific genes, stimulates nitric oxide production, and supports cell survival [78], [79]. Numerous studies have shown that polysaccharides can regulate the expression and activity of growth factors involved in angiogenesis, including VEGF [80], [81]. Therefore, our results indicate that PSOP may influence angiogenesis by activating the inflammatory process and increasing levels of VEGF and TNF-α.
3.5. Quantitative Analysis of Vascular Network
Figure 7II shows the version processed by AngioTool software. Vessel outlines are marked in yellow; vessels are marked in red, and junctions are marked in blue. The results indicate that the density of blood vessels in the PSOP-treated group is more significant than that in the control group, which reveals the pro-angiogenic nature of PSOP.
Quantitative analysis of blood vessels in Figure 8 shows that the concentrations of 50 and 100 μg/egg of PSOP extract and choriogonadotropin (CG) result in a significant increase in vascular surface area, vessel length, and diameter, as well as the number of junctions, compared to the control and the negative control diclofenac (DIC). Moreover, the analysis showed no significant difference between treatment (25 μg/egg) with PSOP extract and control. As expected, a lower lacunarity value has been identified in the PSOP-treated groups (50 and 100 μg/egg) and the positive control compared to the control and the negative control groups. The high lacunarity values signaled that empty spaces in the vascular network architecture in these groups were more frequent. According to the previous data, we deduced that PSOP extract produced remarkable results in forming new vessels. Better vascularization improves oxygen and nutrient delivery to the wound site, essential for tissue regeneration and rapid healing.
3.6. In Vivo Experimental Study
3.6.1. Morphological Evaluation
The wound contraction level was visualized and evidenced by capturing images at different time intervals (1, 5, 7, and 11 days). As shown in Figure 9, day 1 illustrates the initial state of the wounds immediately after induction and application of the various treatments. All wounds were homogeneous in appearance, with dark red coloration, well-defined margins, and visibly damaged tissue, demonstrating wound uniformity in all groups. No significant differences between the groups are discernible at this stage, which is to be expected. These observations serve as a benchmark for assessing treatment efficacy on subsequent days. On day 5 of treatment, wounds in the PSOP and Cytol Centella groups showed brown discoloration, indicating the formation of a crust resulting from the onset of blood coagulation healing. Both treatments showed a marked improvement in healing compared to the untreated groups (saline and glycerol), with a significant reduction in wound size and a less inflamed appearance. On day 7, wounds treated with saline and glycerol showed less advanced healing progression than those treated with PSOP and Cytol Centella, which delayed tissue regeneration. On the 11th day, no scarring was observed in the group treated with PSOP.
Nevertheless, an open wound and scab fragments persisted in the group treated with saline and glycerol. The morphological assessment indicates that treatment with PSOP accelerates the healing process, restoring the skin’s initial structural color. Indeed, reactive oxygen species (ROS) are often linked to delayed wound healing. When ROS are overproduced, without an effective antioxidant response, cell membranes, proteins, and DNA suffer oxidative damage, leading to prolonged inflammation and extracellular matrix degradation, which delays wound healing [82]. It is well known that polysaccharides can neutralize ROS. Their action involves reducing oxidative stress by scavenging oxidizing free radicals, thus preventing oxidative damage to cells and tissues [83]. This suggests that the antioxidant properties of PSOP help to protect cells against oxidative stress, thereby reducing wound stress and promoting faster, more effective wound healing.
3.6.2. Assessment of Wound Area
Epithelialization is an essential phase in wound healing, during which epithelial cells migrate upwards to cover and repair the wounded area [84]. In this way, the healing potential of the treatments was assessed throughout the 11-day experimental period by punctually measuring the size of the injured areas. Figure 10 shows that the groups treated with PSOP and Cytol Centella showed significantly greater healing capacity than the untreated groups (glycerol and saline). The PSOP-treated group achieved complete wound closure (100%) by day 11, indicating optimal efficiency for tissue regeneration. The Cytol Centella group also showed notable performance, with a wound reduction of 90.67% ± 0.27%, albeit lower than with PSOP.
In comparison, the glycerol and saline showed slower healing, with respective closure rates of 70.49% ± 1.09% and 66.59% ± 1.10%. These results suggest that using PSOP enables faster and more complete restoration of skin structure, illustrating its potential as a pro-healing agent. The epithelialization process is strongly influenced by wound conditions, particularly the moisture content of the wound environment. A moist environment encourages epidermal cell migration, accelerates re-epithelialization, and maintains high levels of proteinases and growth factors, all of which are essential for ideal wound healing [85]. In this sense, polysaccharides derived from algae play an important role in maintaining a moist wound-healing environment. Thanks to their hydrophilic properties, biocompatibility, and ability to retain moisture [86].
| 5 days | 7 days | 11 days | |
| Physiological serum | 32.5 ± 1.24a | 48.9 ± 2.02a | 66.59 ± 1.59a |
| Cytol Centella | 39.3 ± 1.41b | 72.54 ± 2.85b | 90.67 ± 2.08b |
| PSOP | 43.22 ± 1.92c | 80.45 ± 3.01c | 100c |
| Glycerol | 34 ± 1.74d | 54.62 ± 2.07d | 70.49 ± 1.27d |
3.6.3. Hydroxyproline and Collagen Regeneration
To confirm the regeneration of skin tissues, wound area biopsies were evaluated to estimate tissue hydroxyproline levels in all groups. As illustrated in Figure 11, the PSOP-treated group showed high hydroxyproline levels (750.81 ± 21 mg/g tissue) compared with the other groups (162.53 ± 7.71, 202.45 ± 14.8 and 626.5 ± 18.95 mg/g tissue for saline, glycerol and Cytol Centella, respectively) (p < 0.05). In reality, high levels of hydroxyproline indicate the synthesis of collagen fibers and induce the proliferation and remodeling of the wound [87]. According to Zhang et al. [88], they have shown that polysaccharides can promote the expression of interleukin-1 beta (IL-1β), which in turn stimulates VEGF production. This cascade of signals promotes angiogenesis and collagen synthesis, which is crucial for tissue regeneration and wound closure. Another factor influencing hydroxyproline production is the PI3K/Akt/mTOR signaling pathway. This pathway plays a key role in regulating cell proliferation and survival, as well as collagen synthesis [89]. Some studies have shown that polysaccharides can activate these signaling pathways, which could indirectly influence hydroxyproline production and, consequently, collagen stability and formation [90], [91]. Our experimental data and available literature evidence support the hypothesis that PSOP could modulate these pathways, thereby contributing to collagen synthesis and skin tissue healing.
3.6.4. Histological Evaluation
Re-epithelialization corresponds to the restoration of the epithelium after a wound, thanks to the proliferation and migration of epithelial cells on the wound surface, creating a protective cover over regenerating tissue. In addition, granulation tissue formation, neovascularization, and collagen deposition are fundamental mechanisms identified throughout the cutaneous wound healing process [92]. To assess the level of wound healing, skin tissue from treated and untreated groups was stained with hematoxylin-eosin and examined at the end of the experiment. The evaluation focused on re-epithelialization, collagen and fibroblast formation, neovascularization, and the deposition of inflammatory cells. As illustrated in Figure 12, microscopic photographs of the untreated groups (saline and glycerol) reveal the persistence of an invasive inflammatory infiltrate with necrosis of the epidermis and dermis, and we also observed low collagen synthesis, a consequence of incomplete repair and re-epithelialization. Nevertheless, the wound tissue sections of the PSOP and Cytol Centella treated groups demonstrated denser epithelization, neo-vascularization, as well as the disappearance of inflammatory cells, which confirms our earlier results regarding the determination of hydroxyproline in wound biopsies.
Histological sections from the PSOP-treated group showed complete healing, supported by accelerated cell proliferation, protection against oxidative damage, and reduced inflammation. The results also confirmed the PSOP’s pro-angiogenic potential, which seems capable of stimulating the expression of VEGF, thereby promoting the proliferation of endothelial cells and fibroblasts.
3.7. Computational Findings
The PSOP-identified monosaccharides showed different affinities to two targeted receptors (Table 2). Nevertheless, they all had negative binding affinities, which might support their biological effects. The predicted binding affinities ranged between −5.6 and −6.4 kcal/mol for COX-2 and −4.1 and −4.6 kcal/mol for VEGF. Such variations have been reported to be the result of 3D chemical structures of the ligands ([34], [33], [93]). The identified monosaccharides established acceptable molecular interactions with both targeted receptors, including four to six conventional H-bonds (Table 3). The established molecular interactions concerned several key residues. For instance, fructose while complexed with COX-2, fructose interacted twice with ASN39 and once with each of CYS47, GLU465, CYS41, and GLY45 (Table 3 and Figure 13). While complexed with VEGF, glucose interacted twice with LEU39 and once with THR36, LEU35, and LYS45. PSOP units were also deeply embedded in the pocket region of the targeted receptors and showed a vicinity of up to 2.072 Å only.
Deep embedding (<2.5 Å), as those of the current study, was previously reported to enhance the bioactivities such as antiproliferative, anti-inflammatory, toxicity alleviation, and antimicrobial effects ([32], [93], [34], [35]). The binding affinities, the deep embedding, and the revealed molecular interactions of PSOP building blocks indicate that the pro-angiogenic and wound-healing potential effects of PSOP chemicals are thermodynamically possible. These predicted bioactivities had already been confirmed by the in vitro study in rats. Our results support the beneficial effects of natural-derived compounds, phytotherapy, and medicinal plants, including algae ([94], [93], [35]).
4. Conclusions
This is an unprecedented report on the antioxidant activity, healing, and pro-angiogenic properties of PSOP extracted from the red alga Osmundea pinnatifida. The PSOP is genuinely and potentially interesting due to the antioxidant activities confirmed by various tests (DPPH, TAC and RPA). These activities include eliminating the DPPH free radical, reducing iron (III), and converting molybdenum Mo (VI) to Mo (V). The findings of our study show that PSOP has a pro-angiogenic effect when tested using the CAM assay. This effect was further confirmed by observing an accelerated wound healing process in Wistar rats after 11 days, involving angiogenesis, fibrotic tissue formation, remodeling, and reduced healing time using the circular excision model. Our combined experimental and computational results suggest that PSOP could be exploited as a natural bioactive material to improve the healing of chronic wounds, such as diabetic ulcers, burns, or even to promote vascularization during skin grafting procedures, etc.
Author Contributions
Conceptualization: AH, IY and IBA; Methodology: ZB, ME, AF, HBS and SE; Software: ZB, MK and RB; Validation: AH, IBA and SH; Supply of reagents and materials: IBA, SH and SE; Writing – original draft preparation, ZB; Writing – review & editing: JMP, HK and RB; Visualization: IBA, RB, JMP and HK. All authors have read and accepted the published version of the manuscript.
Funding
This work was supported by the Tuniso-Moroccan project (code 20PRD16).
Ethical Statements
All animal protocols complied with European Union legislation (Directive 2010/63/EU) on protecting animals used for scientific purposes and were approved by the local ethics committee.
Acknowledgments
The authors would like to express their deep gratitude to Professor Samira Etahiri for his expertise in the identification of algae.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Han, X.; Ju, L.S.; Irudayaraj, J. Oxygenated Wound Dressings for Hypoxia Mitigation and Enhanced Wound Healing. Mol. Pharmaceutics 2023, 20, 3338–3355. [Google Scholar] [CrossRef] [PubMed]
- Raziyeva, K.; Kim, Y.; Zharkinbekov, Z.; Kassymbek, K.; Jimi, S.; Saparov, A. Immunology of Acute and Chronic Wound Healing. Biomolecules 2021, 11, 700. [Google Scholar] [CrossRef] [PubMed]
- Bhat, G.P.; Maurizio, A.; Motta, A.; Podini, P.; Diprima, S.; Malpighi, C.; Brambilla, I.; Martins, L.; Badaloni, A.; Boselli, D.; et al. Structured Wound Angiogenesis Instructs Mesenchymal Barrier Compartments in the Regenerating Nerve. Neuron 2024, 112, 209–229.e11. [Google Scholar] [CrossRef] [PubMed]
- Mota, F.A.R.; Passos, M.L.C.; Santos, J.L.M.; Saraiva, M.L.M.F.S. Comparative Analysis of Electrochemical and Optical Sensors for Detection of Chronic Wounds Biomarkers: A Review. Biosensors and Bioelectronics 2024, 251, 116095. [Google Scholar] [CrossRef] [PubMed]
- Rathna, R.P.; Kulandhaivel, M. Advancements in Wound Healing: Integrating Biomolecules, Drug Delivery Carriers, and Targeted Therapeutics for Enhanced Tissue Repair. Arch Microbiol 2024, 206, 199. [Google Scholar] [CrossRef]
- Boujhoud, Z.; Feki, A.; Eleroui, M.; Lakhram, M.; Kraiem, M.; Dghim, A.; Zeroual, A.; Youlyouz Marfak, I.; Essayagh, S.; Hilali, S.; et al. The Anti-Angiogenic, Anti-Inflammatory and Anticoagulant Potential of a Polysaccharide Extracted from the Brown Alga Cystoseira Humilis. European Polymer Journal 2024, 220, 113461. [Google Scholar] [CrossRef]
- Peipei, L.; Qinghong, Z.; Yin, C.; Pengfei, H.; Junjie, Z. Structure and Anticoagulant Activity of a Galactoarabinan Sulfate Polysaccharide and Its Oligosaccharide from the Green Algae, Codium Fragile. International Journal of Biological Macromolecules 2024, 279, 135255. [Google Scholar] [CrossRef]
- Torres, M.D.; Flórez-Fernández, N.; Domínguez, H. Chondrus Crispus Treated with Ultrasound as a Polysaccharides Source with Improved Antitumoral Potential. Carbohydrate Polymers 2021, 273, 118588. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, S.; Yang, M.; Ding, J.; Huang, Y.; Zhu, Y.; Zhou, M.; Yan, B. Antiviral Activity of a Polysaccharide from Sargassum Fusiforme against Respiratory Syncytial Virus. International Journal of Biological Macromolecules 2024, 279, 135267. [Google Scholar] [CrossRef]
- Luo, M.; Zhang, X.; Wu, J.; Zhao, J. Modifications of Polysaccharide-Based Biomaterials under Structure-Property Relationship for Biomedical Applications. Carbohydrate Polymers 2021, 266, 118097. [Google Scholar] [CrossRef]
- Kumar, M.; Kumar, D.; Garg, Y.; Mahmood, S.; Chopra, S.; Bhatia, A. Marine-Derived Polysaccharides and Their Therapeutic Potential in Wound Healing Application - A Review. International Journal of Biological Macromolecules 2023, 253, 127331. [Google Scholar] [CrossRef] [PubMed]
- Biancacci, C.; Abell, R.; McDougall, G.J.; Day, J.G.; Stanley, M.S. Annual Compositional Variation in Wild Osmundea Pinnatifida (Hudson) Stackhouse from the West Coast of Scotland. J Appl Phycol 2022, 34, 1661–1675. [Google Scholar] [CrossRef]
- Rodrigues, D.; Freitas, A.C.; Pereira, L.; Rocha-Santos, T.A.P.; Vasconcelos, M.W.; Roriz, M.; Rodríguez-Alcalá, L.M.; Gomes, A.M.P.; Duarte, A.C. Chemical Composition of Red, Brown and Green Macroalgae from Buarcos Bay in Central West Coast of Portugal. Food Chemistry 2015, 183, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Silva, P.; Pereira, L. Concise Review of Osmundea Pinnatifida (Hudson) Stackhouse. J Appl Phycol 2020, 32, 2761–2771. [Google Scholar] [CrossRef]
- Moreira, B.R.; Vega, J.; Sisa, A.D.A.; Bernal, J.S.B.; Abdala-Díaz, R.T.; Maraschin, M.; Figueroa, F.L.; Bonomi-Barufi, J. Antioxidant and Anti-Photoaging Properties of Red Marine Macroalgae: Screening of Bioactive Molecules for Cosmeceutical Applications. Algal Research 2022, 68, 102893. [Google Scholar] [CrossRef]
- Rodrigues, D.; Sousa, S.; Silva, A.; Amorim, M.; Pereira, L.; Rocha-Santos, T.A.P.; Gomes, A.M.P.; Duarte, A.C.; Freitas, A.C. Impact of Enzyme- and Ultrasound-Assisted Extraction Methods on Biological Properties of Red, Brown, and Green Seaweeds from the Central West Coast of Portugal. J. Agric. Food Chem. 2015, 63, 3177–3188. [Google Scholar] [CrossRef]
- Liu, G.; Xu, S.; Chen, L. Chemical Composition and Bioactivities of a Water-Soluble Polysaccharide from the Endodermis of Shaddock. International Journal of Biological Macromolecules 2012, 51, 763–766. [Google Scholar] [CrossRef]
- Ortega-Calvo, J.J.; Mazuelos, C.; Hermosin, B.; Saiz-Jimenez, C. Chemical Composition ofSpirulina and Eukaryotic Algae Food Products Marketed in Spain. J Appl Phycol 1993, 5, 425–435. [Google Scholar] [CrossRef]
- Lowry, OliverH. ; Rosebrough, NiraJ.; Farr, A.L.; Randall, RoseJ. PROTEIN MEASUREMENT WITH THE FOLIN PHENOL REAGENT. Journal of Biological Chemistry 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Llyod, A.G.; Tudball, N.; Dodgson, K.S. Infrared Studies on Sulphate Esters III. O-Sulphate Esters of Alcohols, Amino Alcohols and Hydroxylated Amino Acids. Biochimica et Biophysica Acta 1961, 52, 413–419. [Google Scholar] [CrossRef]
- He, R.; Zhao, Y.; Zhao, R.; Sun, P. Antioxidant and Antitumor Activities in Vitro of Polysaccharides from E. Sipunculoides. International Journal of Biological Macromolecules 2015, 78, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.-J.; Zhang, J.-G.; Sun, Y.-H.; Qu, J.; Li, L.; Prasad, C.; Wei, Z.-J. Physicochemical Properties and Antioxidant Activities of Polysaccharides Sequentially Extracted from Peony Seed Dreg. International Journal of Biological Macromolecules 2016, 91, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.-H.; Xie, M.-Y.; Nie, S.-P.; Shen, M.-Y.; Wang, Y.-X.; Li, C. Isolation, Chemical Composition and Antioxidant Activities of a Water-Soluble Polysaccharide from Cyclocarya Paliurus (Batal.) Iljinskaja. Food Chemistry 2010, 119, 1626–1632. [Google Scholar] [CrossRef]
- Bayar, N.; Kriaa, M.; Kammoun, R. Extraction and Characterization of Three Polysaccharides Extracted from Opuntia Ficus Indica Cladodes. International Journal of Biological Macromolecules 2016, 92, 441–450. [Google Scholar] [CrossRef] [PubMed]
- Bersuder, P.; Hole, M.; Smith, G. Antioxidants from a Heated Histidine-Glucose Model System. I: Investigation of the Antioxidant Role of Histidine and Isolation of Antioxidants by High-Performance Liquid Chromatography. J Amer Oil Chem Soc 1998, 75, 181–187. [Google Scholar] [CrossRef]
- Prieto, P.; Pineda, M.; Aguilar, M. Spectrophotometric Quantitation of Antioxidant Capacity through the Formation of a Phosphomolybdenum Complex: Specific Application to the Determination of Vitamin E. Analytical Biochemistry 1999, 269, 337–341. [Google Scholar] [CrossRef]
- Yıldırım, A.; Mavi, A.; Kara, A.A. Determination of Antioxidant and Antimicrobial Activities of Rumex Crispus L. Extracts. J. Agric. Food Chem. 2001, 49, 4083–4089. [Google Scholar] [CrossRef]
- Namvar, F.; Mohamad, R.; Baharara, J.; Zafar-Balanejad, S.; Fargahi, F.; Rahman, H.S. Antioxidant, Antiproliferative, and Antiangiogenesis Effects of Polyphenol-Rich Seaweed ( Sargassum Muticum ). BioMed Research International 2013, 2013, 1–9. [Google Scholar] [CrossRef]
- Institutional Animal Care and Use Committee Guidebook; Office of Laboratory Animal Welfare, 2002.
- Edwards, C.A.; O’Brien, W.D. Modified Assay for Determination of Hydroxyproline in a Tissue Hydrolyzate. Clinica Chimica Acta 1980, 104, 161–167. [Google Scholar] [CrossRef]
- Badraoui, R.; Allouche, M.; El Ouaer, D.; Siddiqui, A.J.; Ishak, S.; Hedfi, A.; Beyrem, H.; Pacioglu, O.; Rudayni, H.A.; Boufahja, F. Ecotoxicity of Chrysene and Phenanthrene on Meiobenthic Nematodes with a Case Study of Terschellingia Longicaudata: Taxonomics, Toxicokinetics, and Molecular Interactions Modelling. Environmental Pollution 2023, 316, 120459. [Google Scholar] [CrossRef]
- Chira, A.; Kadmi, Y.; Badraoui, R.; Aouadi, K.; Alhawday, F.; Boudaya, M.; Jamoussi, K.; Kallel, C.; El Feki, A.; Kadri, A.; et al. GC-MS/MS Analysis and Wound Repair Potential of Urtica Dioica Essential Oil: In Silico Modeling and In Vivo Study in Rats. Curr Pharm Biotechnol 2024. [Google Scholar] [CrossRef] [PubMed]
- Ishak, S.; Allouche, M.; Alotaibi, G.S.; Alwthery, N.S.; Al-Subaie, R.A.; Al-Hoshani, N.; Plavan, O.-A.; Selamoglu, Z.; Özdemir, S.; Plavan, G.; et al. Experimental and Computational Assessment of Antiparkinson Medication Effects on Meiofauna: Case Study of Benserazide and Trihexyphenidyl. Marine Pollution Bulletin 2024, 205, 116668. [Google Scholar] [CrossRef] [PubMed]
- Rahmouni, F.; Hamdaoui, L.; Saoudi, M.; Badraoui, R.; Rebai, T. Antioxidant and Antiproliferative Effects of Teucrium Polium Extract: Computational and in Vivo Study in Rats. Toxicology Mechanisms and Methods 2024, 34, 495–506. [Google Scholar] [CrossRef] [PubMed]
- Kraiem, M.; Ben Hamouda, S.; Eleroui, M.; Ajala, M.; Feki, A.; Dghim, A.; Boujhoud, Z.; Bouhamed, M.; Badraoui, R.; Pujo, J.M.; et al. Anti-Inflammatory and Immunomodulatory Properties of a Crude Polysaccharide Derived from Green Seaweed Halimeda Tuna: Computational and Experimental Evidences. Marine Drugs 2024, 22, 85. [Google Scholar] [CrossRef] [PubMed]
- Mhadhbi, N.; Dgachi, S.; Belgacem, S.; Ahmed, A.B.; Henry, N.; Loiseau, T.; Nasr, S.; Badraoui, R.; Naϊli, H. Design, Theoretical Study, Druggability, Pharmacokinetics and Properties Evolution of a New Organo-Bromocadmate Compound as Prospective Anticancer Agent. Journal of Molecular Structure 2023, 1274, 134439. [Google Scholar] [CrossRef]
- Arunkumar, K.; Sreena, K.S.; Moosa, M.; Mohan, G.; Raja, R. Cytotoxic Characterization of Optically Negative Codium Fragile Polysaccharide against HeLa and MCF Cell Lines. Bioactive Carbohydrates and Dietary Fibre 2023, 29, 100341. [Google Scholar] [CrossRef]
- Kravchenko, A.O.; Byankina Barabanova, A.O.; Glazunov, V.P.; Yakovleva, I.M.; Yermak, I.M. Seasonal Variations in a Polysaccharide Composition of Far Eastern Red Seaweed Ahnfeltiopsis Flabelliformis (Phyllophoraceae). J Appl Phycol 2018, 30, 535–545. [Google Scholar] [CrossRef]
- Yaich, H.; Garna, H.; Besbes, S.; Paquot, M.; Blecker, C.; Attia, H. Effect of Extraction Conditions on the Yield and Purity of Ulvan Extracted from Ulva Lactuca. Food Hydrocolloids 2013, 31, 375–382. [Google Scholar] [CrossRef]
- Siddhanta, A.; Goswami, A.M.; Munisamy, S.; Mody, K.H.; Ramavat, B.K.; Mairh, O.P. Sulphated Galactans of Marine Red Alga Laurencia Spp. (Rhodomelaceae, Rhodophyta) from the West Coast of India. Indian Journal of Marine Sciences 2002, 31, 305–309. [Google Scholar]
- Hu, W.; Yu, A.; Bi, H.; Gong, Y.; Wang, H.; Kuang, H.; Wang, M. Recent Advances in Artemisia Argyi Levl. et Vant. Polysaccharides: Extractions, Purifications, Structural Characteristics, Pharmacological Activities, and Existing and Potential Applications. International Journal of Biological Macromolecules 2024, 279, 135250. [Google Scholar] [CrossRef]
- Yi, Y.; Xu, W.; Wang, H.-X.; Huang, F.; Wang, L.-M. Natural Polysaccharides Experience Physiochemical and Functional Changes during Preparation: A Review. Carbohydrate Polymers 2020, 234, 115896. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.; Chen, F.; Yang, W.; Huang, H. Preparation, Deproteinization and Comparison of Bioactive Polysaccharides. Trends in Food Science & Technology 2021, 109, 564–568. [Google Scholar] [CrossRef]
- Wang, Z.; Xie, J.; Shen, M.; Nie, S.; Xie, M. Sulfated Modification of Polysaccharides: Synthesis, Characterization and Bioactivities. Trends in Food Science & Technology 2018, 74, 147–157. [Google Scholar] [CrossRef]
- Jose, G.M.; Raghavankutty, M.; Kurup, G.M. Sulfated Polysaccharides from Padina Tetrastromatica Induce Apoptosis in HeLa Cells through ROS Triggered Mitochondrial Pathway. Process Biochemistry 2018, 68, 197–204. [Google Scholar] [CrossRef]
- Yin, Z.; Zhang, W.; Zhang, J.; Kang, W. Isolation, Purification, Structural Analysis and Coagulatory Activity of Water-Soluble Polysaccharides from Ligustrum Lucidum Ait Flowers. Chemistry Central Journal 2017, 11, 98. [Google Scholar] [CrossRef]
- Eljoudi, S.; Feki, A.; Bkhairia, I.; Barkia, A.; Ben Amara, I.; Nasri, M.; Hajji, M. New Polysaccharides Extracted from Malcolmia Triloba: Structure Characterization, Biological Properties and Application to Beef Meat Preservation. Journal of Food Composition and Analysis 2022, 107, 104380. [Google Scholar] [CrossRef]
- Yuan, Q.; Lin, S.; Fu, Y.; Nie, X.-R.; Liu, W.; Su, Y.; Han, Q.-H.; Zhao, L.; Zhang, Q.; Lin, D.-R.; et al. Effects of Extraction Methods on the Physicochemical Characteristics and Biological Activities of Polysaccharides from Okra (Abelmoschus Esculentus). International Journal of Biological Macromolecules 2019, 127, 178–186. [Google Scholar] [CrossRef]
- Ramamoorthy, S.; Gnanakan, A.; S. Lakshmana, S.; Meivelu, M.; Jeganathan, A. Structural Characterization and Anticancer Activity of Extracellular Polysaccharides from Ascidian Symbiotic Bacterium Bacillus Thuringiensis. Carbohydrate Polymers 2018, 190, 113–120. [Google Scholar] [CrossRef]
- Pei, Y.; Yang, S.; Xiao, Z.; Zhou, C.; Hong, P.; Qian, Z.-J. Structural Characterization of Sulfated Polysaccharide Isolated From Red Algae (Gelidium Crinale) and Antioxidant and Anti-Inflammatory Effects in Macrophage Cells. Front. Bioeng. Biotechnol. 2021, 9. [Google Scholar] [CrossRef]
- Mateos-Aparicio, I.; Martera, G.; Goñi, I.; Villanueva-Suárez, M.-J.; Redondo-Cuenca, A. Chemical Structure and Molecular Weight Influence the in Vitro Fermentability of Polysaccharide Extracts from the Edible Seaweeds Himathalia Elongata and Gigartina Pistillata. Food Hydrocolloids 2018, 83, 348–354. [Google Scholar] [CrossRef]
- Jaballi, I.; Sallem, I.; Feki, A.; Cherif, B.; Kallel, C.; Boudawara, O.; Jamoussi, K.; Mellouli, L.; Nasri, M.; Amara, I.B. Polysaccharide from a Tunisian Red Seaweed Chondrus Canaliculatus: Structural Characteristics, Antioxidant Activity and in Vivo Hemato-Nephroprotective Properties on Maneb Induced Toxicity. International Journal of Biological Macromolecules 2019, 123, 1267–1277. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, D.; Costa-Pinto, A.R.; Sousa, S.; Vasconcelos, M.W.; Pintado, M.M.; Pereira, L.; Rocha-Santos, T.A.P.; Costa, J.P. da; Silva, A.M.S.; Duarte, A.C.; et al. Sargassum Muticum and Osmundea Pinnatifida Enzymatic Extracts: Chemical, Structural, and Cytotoxic Characterization. Marine Drugs 2019, 17, 209. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, L.G.; da Silva, A.C.R.; Noseda, M.D.; Fuly, A.L.; de Carvalho, M.M.; Fujii, M.T.; Sanchez, E.F.; Carneiro, J.; Duarte, M.E.R. Chemical Structure and Snake Antivenom Properties of Sulfated Agarans Obtained from Laurencia Dendroidea (Ceramiales, Rhodophyta). Carbohydrate Polymers 2019, 218, 136–144. [Google Scholar] [CrossRef] [PubMed]
- Hans, N.; Pattnaik, F.; Malik, A.; Naik, S. Comparison of Different Green Extraction Techniques and Their Influence on Chemical Characteristics of Sulfated Polysaccharide (Fucoidan) from Padina Tetrastromatica and Turbinaria Conoides. Algal Research 2023, 74, 103199. [Google Scholar] [CrossRef]
- Konstantin, B.; Anastasia, P.; Nikolay, I.; Daria, P. Seasonal Variations in the Chemical Composition of Arctic Brown Macroalgae. Algal Research 2023, 72, 103112. [Google Scholar] [CrossRef]
- Wang, Z.; Zheng, Y.; Lai, Z.; Hu, X.; Wang, L.; Wang, X.; Li, Z.; Gao, M.; Yang, Y.; Wang, Q.; et al. Effect of Monosaccharide Composition and Proportion on the Bioactivity of Polysaccharides: A Review. International Journal of Biological Macromolecules 2024, 254, 127955. [Google Scholar] [CrossRef]
- Lee, Q.; Xue, Z.; Luo, Y.; Lin, Y.; Lai, M.; Xu, H.; Liu, B.; Zheng, M.; Lv, F.; Zeng, F. Low Molecular Weight Polysaccharide of Tremella Fuciformis Exhibits Stronger Antioxidant and Immunomodulatory Activities than High Molecular Weight Polysaccharide. International Journal of Biological Macromolecules 2024, 136097. [Google Scholar] [CrossRef]
- Wang, Y.; Li, Y.; Ma, X.; Ren, H.; Fan, W.; Leng, F.; Yang, M.; Wang, X. Extraction, Purification, and Bioactivities Analyses of Polysaccharides from Glycyrrhiza Uralensis. Industrial Crops and Products 2018, 122, 596–608. [Google Scholar] [CrossRef]
- Khan, B.M.; Qiu, H.-M.; Xu, S.-Y.; Liu, Y.; Cheong, K.-L. Physicochemical Characterization and Antioxidant Activity of Sulphated Polysaccharides Derived from Porphyra Haitanensis. International Journal of Biological Macromolecules 2020, 145, 1155–1161. [Google Scholar] [CrossRef]
- Teng, C.; Li, S.; Xu, L.; Ma, K.; Lu, Y.; Feng, J.; Chai, Z.; Hu, X.; Zhou, W.; Li, Y. Structural Characterization, Physicochemical Properties and Hypoglycemic Activity of Sulfated Polysaccharides from Porphyra Yezoensis. Food Bioscience 2024, 62, 105163. [Google Scholar] [CrossRef]
- Naghdi, S.; Rezaei, M.; Tabarsa, M.; Abdollahi, M. Ultrasonic-Assisted Enzymatic Extraction of Sulfated Polysaccharide from Skipjack Tuna by-Products. Ultrasonics Sonochemistry 2023, 95, 106385. [Google Scholar] [CrossRef] [PubMed]
- Seidi, F.; Yazdi, M.K.; Jouyandeh, M.; Habibzadeh, S.; Munir, M.T.; Vahabi, H.; Bagheri, B.; Rabiee, N.; Zarrintaj, P.; Saeb, M.R. Crystalline Polysaccharides: A Review. Carbohydrate Polymers 2022, 275, 118624. [Google Scholar] [CrossRef] [PubMed]
- Marinho, G.S.; Sørensen, A.-D.M.; Safafar, H.; Pedersen, A.H.; Holdt, S.L. Antioxidant Content and Activity of the Seaweed Saccharina Latissima: A Seasonal Perspective. J Appl Phycol 2019, 31, 1343–1354. [Google Scholar] [CrossRef]
- Hermund, D. Antioxidant Properties of Seaweed-Derived Substances. In Bioactive Seaweeds for Food Applications; 2018; pp. 201–221 ISBN 978-0-12-813312-5.
- Hamzaoui, A.; Ghariani, M.; Sellem, I.; Hamdi, M.; Feki, A.; Jaballi, I.; Nasri, M.; Amara, I.B. Extraction, Characterization and Biological Properties of Polysaccharide Derived from Green Seaweed “Chaetomorpha Linum” and Its Potential Application in Tunisian Beef Sausages. International Journal of Biological Macromolecules 2020, 148, 1156–1168. [Google Scholar] [CrossRef]
- Ma, X.-T.; Sun, X.-Y.; Yu, K.; Gui, B.-S.; Gui, Q.; Ouyang, J.-M. Effect of Content of Sulfate Groups in Seaweed Polysaccharides on Antioxidant Activity and Repair Effect of Subcellular Organelles in Injured HK-2 Cells. Oxidative Medicine and Cellular Longevity 2017, 2017, e2542950. [Google Scholar] [CrossRef]
- Khemakhem, I.; Abdelhedi, O.; Trigui, I.; Ayadi, M.A.; Bouaziz, M. Structural, Antioxidant and Antibacterial Activities of Polysaccharides Extracted from Olive Leaves. International Journal of Biological Macromolecules 2018, 106, 425–432. [Google Scholar] [CrossRef]
- Wang, J.; Hu, S.; Nie, S.; Yu, Q.; Xie, M. Reviews on Mechanisms of In Vitro Antioxidant Activity of Polysaccharides. Oxidative Medicine and Cellular Longevity 2015, 2016, e5692852. [Google Scholar] [CrossRef]
- Abdelhedi, O.; Jridi, M.; Najjaa, H.; Zouari, N.; Sebai, H.; Nasri, M. Sulfated Polysaccharides from the Viscera of Mustelus Shark: Characterization and Antioxidant, Anticoagulant and Anti-Proliferative Activities. Bioactive Carbohydrates and Dietary Fibre 2024, 31, 100399. [Google Scholar] [CrossRef]
- Gunasekaran, S.; Govindan, S.; Ramani, P. Sulfated Modification, Characterization and Bioactivities of an Acidic Polysaccharide Fraction from an Edible Mushroom Pleurotus Eous (Berk.) Sacc. Heliyon 2021, 7. [Google Scholar] [CrossRef]
- Ribatti, D.; Nico, B.; Vacca, A.; Roncali, L.; Burri, P.H.; Djonov, V. Chorioallantoic Membrane Capillary Bed: A Useful Target for Studying Angiogenesis and Anti-Angiogenesis in Vivo. Anat. Rec. 2001, 264, 317–324. [Google Scholar] [CrossRef]
- Muhammad, O.; Lama, R. The Effect of Diclofenac Sodium on Blood Vessel Formation. Medbiotech Journal 2018, 02. [Google Scholar] [CrossRef]
- Berndt, S.; Blacher, S.; Perrier d’Hauterive, S.; Thiry, M.; Tsampalas, M.; Cruz, A.; Péqueux, C.; Lorquet, S.; Munaut, C.; Noël, A.; et al. Chorionic Gonadotropin Stimulation of Angiogenesis and Pericyte Recruitment. The Journal of Clinical Endocrinology & Metabolism 2009, 94, 4567–4574. [Google Scholar] [CrossRef]
- Han, G.; Ceilley, R. Chronic Wound Healing: A Review of Current Management and Treatments. Adv Ther 2017, 34, 599–610. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Cazares, D.; Chavez-Dominguez, R.; Carlos-Reyes, A.; Lopez-Camarillo, C.; Hernadez de la Cruz, O.N.; Lopez-Gonzalez, J.S. Contribution of Angiogenesis to Inflammation and Cancer. Front. Oncol. 2019, 9. [Google Scholar] [CrossRef]
- Yu, R.; Zhong, J.; Zhou, Q.; Ren, W.; Liu, Z.; Bian, Y. Kaempferol Prevents Angiogenesis of Rat Intestinal Microvascular Endothelial Cells Induced by LPS and TNF-α via Inhibiting VEGF/Akt/P38 Signaling Pathways and Maintaining Gut-Vascular Barrier Integrity. Chemico-Biological Interactions 2022, 366, 110135. [Google Scholar] [CrossRef]
- Mauroux, A.; Gofflo, S.; Breugnot, J.; Malbouyres, M.; Atlas, Y.; Ardidie-Robouant, C.; Marchand, L.; Monnot, C.; Germain, S.; Bordes, S.; et al. Angiogenesis and Full Thickness Wound Repair in a Cell Sheet-Based Vascularized Skin Substitute. Acta Biomaterialia 2024, 187, 123–137. [Google Scholar] [CrossRef]
- Jozkowicz, A.; Cooke, J.P.; Guevara, I.; Huk, I.; Funovics, P.; Pachinger, O.; Weidinger, F.; Dulak, J. Genetic Augmentation of Nitric Oxide Synthase Increases the Vascular Generation of VEGF. Cardiovascular Research 2001, 51, 773–783. [Google Scholar] [CrossRef]
- Keykhaee, M.; Sorouri, F.; Rahimifard, M.; Baeeri, M.; Forumadi, A.; Firoozpour, L.; Khoobi, M. Polysaccharide-Based Hydrogel Enriched by Epidermal Growth Factor Peptide Fragment for Improving the Wound Healing Process. Heliyon 2023, 9, e22749. [Google Scholar] [CrossRef]
- Malektaj, H.; Nour, S.; Imani, R.; Siadati, M.H. Angiogenesis Induction as a Key Step in Cardiac Tissue Regeneration: From Angiogenic Agents to Biomaterials. International Journal of Pharmaceutics 2023, 643, 123233. [Google Scholar] [CrossRef]
- Dong, Y.; Wang, Z. ROS-Scavenging Materials for Skin Wound Healing: Advancements and Applications. Front. Bioeng. Biotechnol. 2023, 11. [Google Scholar] [CrossRef]
- Liu, Y.; Zhu, Y.; Zheng, Q.; Lu, H.; Huang, H.; Zhang, J.; Fang, Z.; Lin, L.; Ma, P. Recent Progress in the Efficacy of Algal Saccharides on Skin Repair. Algal Research 2024, 78, 103403. [Google Scholar] [CrossRef]
- Tan, S.T.; Dosan, R. Lessons From Epithelialization: The Reason Behind Moist Wound Environment. [CrossRef]
- Rezvani Ghomi, E.; Khalili, S.; Nouri Khorasani, S.; Esmaeely Neisiany, R.; Ramakrishna, S. Wound Dressings: Current Advances and Future Directions. Journal of Applied Polymer Science 2019, 136, 47738. [Google Scholar] [CrossRef]
- Andryukov, B.G.; Besednova, N.N.; Kuznetsova, T.A.; Zaporozhets, T.S.; Ermakova, S.P.; Zvyagintseva, T.N.; Chingizova, E.A.; Gazha, A.K.; Smolina, T.P. Sulfated Polysaccharides from Marine Algae as a Basis of Modern Biotechnologies for Creating Wound Dressings: Current Achievements and Future Prospects. Biomedicines 2020, 8, 301. [Google Scholar] [CrossRef] [PubMed]
- Karna, E.; Szoka, L.; Huynh, T.Y.L.; Palka, J.A. Proline-Dependent Regulation of Collagen Metabolism. Cell. Mol. Life Sci. 2020, 77, 1911–1918. [Google Scholar] [CrossRef]
- Zhang, H.; Chen, J.; Cen, Y. Burn Wound Healing Potential of a Polysaccharide from Sanguisorba Officinalis L. in Mice. International Journal of Biological Macromolecules 2018, 112, 862–867. [Google Scholar] [CrossRef]
- Bao, L.; Cai, X.; Zhang, M.; Xiao, Y.; Jin, J.; Qin, T.; Li, Y. Bovine Collagen Oligopeptides Accelerate Wound Healing by Promoting Fibroblast Migration via PI3K/Akt/mTOR Signaling Pathway. Journal of Functional Foods 2022, 90, 104981. [Google Scholar] [CrossRef]
- Chen, Z.-Y.; Chen, S.-H.; Chen, C.-H.; Chou, P.-Y.; Yang, C.-C.; Lin, F.-H. Polysaccharide Extracted from Bletilla Striata Promotes Proliferation and Migration of Human Tenocytes. Polymers 2020, 12, 2567. [Google Scholar] [CrossRef]
- Nie, X.; Li, J.; Cheng, Y.; Rangsinth, P.; Wu, X.; Zheng, C.; Shiu, P.H.-T.; Li, R.; Xu, N.; He, Y.; et al. Characterization of a Polysaccharide from Amauroderma Rugosum and Its Proangiogenic Activities in Vitro and in Vivo. International Journal of Biological Macromolecules 2024, 271, 132533. [Google Scholar] [CrossRef]
- Chouikhi, A.; Ktari, N.; Bardaa, S.; Hzami, A.; Ben Slima, S.; Trabelsi, I.; Asehraou, A.; Ben Salah, R. A Novel Probiotic Strain, Lactiplantibacillus Plantarum LC38, Isolated from Tunisian Camel Milk Promoting Wound Healing in Wistar Diabetic Rats. Arch Microbiol 2021, 204, 24. [Google Scholar] [CrossRef]
- Ben Saad, H.; Frikha, D.; Bouallegue, A.; Badraoui, R.; Mellouli, M.; Kallel, H.; Pujo, J.M.; Ben Amara, I. Mitigation of Hepatic Impairment with Polysaccharides from Red Alga Albidum Corallinum Supplementation through Promoting the Lipid Profile and Liver Homeostasis in Tebuconazole-Exposed Rats. Pharmaceuticals 2023, 16, 1305. [Google Scholar] [CrossRef]
- Bédoui, I.; Nasr, H.B.; Ksouda, K.; Ayadi, W.; Louati, N.; Chamkha, M.; Choura, S.; Gargouri, J.; Hammami, S.; Affes, H.; et al. Phytochemical Composition, Bioavailability and Pharmacokinetics of Scorzonera Undulata Methanolic Extracts: Antioxidant, Anticancer, and Apoptotic Effects on MCF7 Cells. Pharmacognosy Magazine 2024, 20, 218–229. [Google Scholar] [CrossRef]
Figure 1.
The location of the seaweed collection site uses ArcGIS version 10.7 (Sidi Bouzid coastal zone, El Jadida, Morocco).
Figure 1.
The location of the seaweed collection site uses ArcGIS version 10.7 (Sidi Bouzid coastal zone, El Jadida, Morocco).
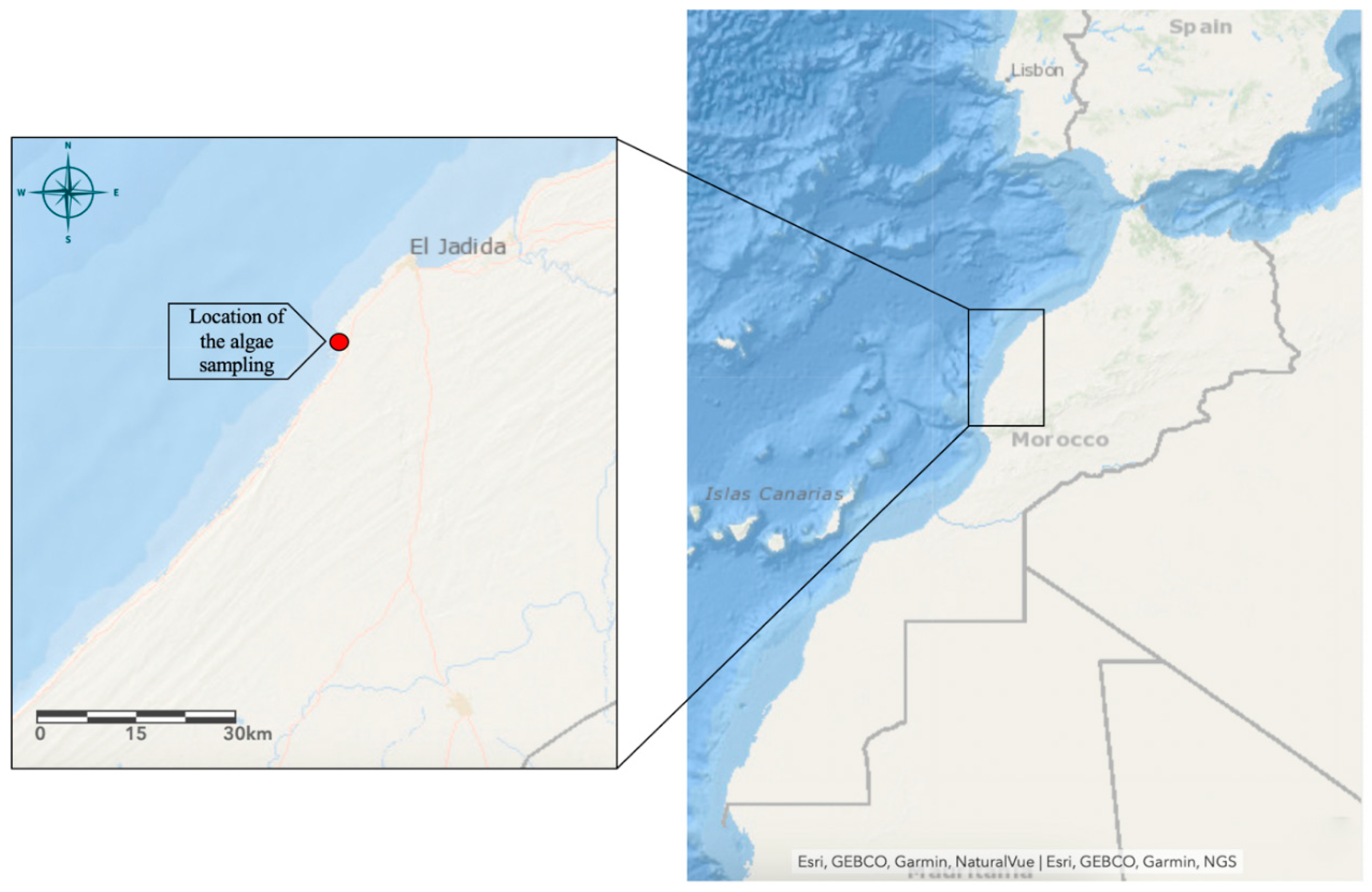
Figure 2.
Preliminary characterization of PSOP. (A) UV–vis absorption spectrum of the PSOP; (B) Infra-red spectrum of the PSOP.
Figure 2.
Preliminary characterization of PSOP. (A) UV–vis absorption spectrum of the PSOP; (B) Infra-red spectrum of the PSOP.
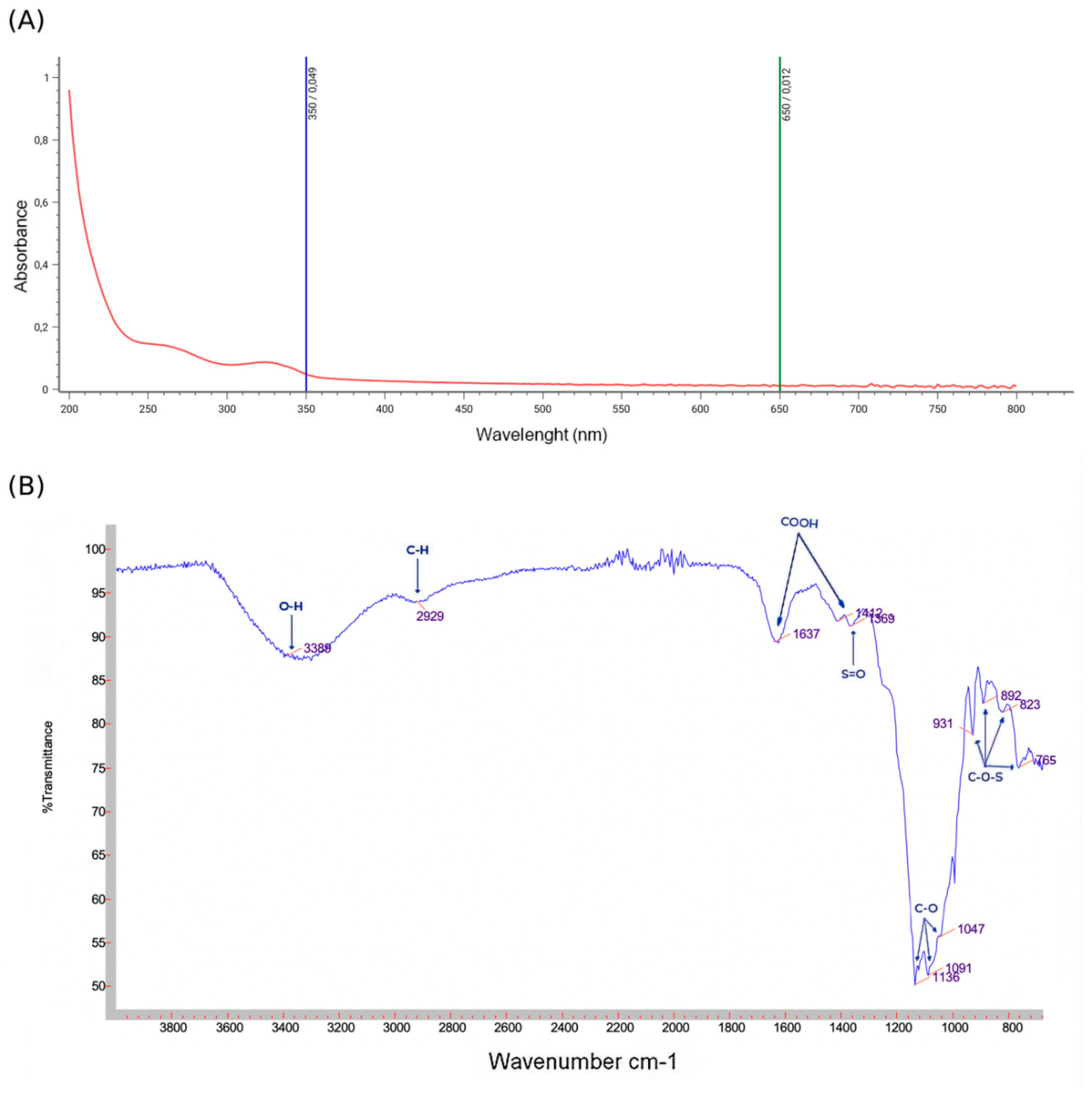
Figure 3.
Preliminary characterization of PSOP. (A) Monosaccharide composition analysis by HPLC-RID; (B) standards used for HPLC. GlcA: Acid Glucuronic; Ara: Arabinose; Man: Mannose; Glc: Glucose; Xyl: Xylose and Fru: Fructose.
Figure 3.
Preliminary characterization of PSOP. (A) Monosaccharide composition analysis by HPLC-RID; (B) standards used for HPLC. GlcA: Acid Glucuronic; Ara: Arabinose; Man: Mannose; Glc: Glucose; Xyl: Xylose and Fru: Fructose.
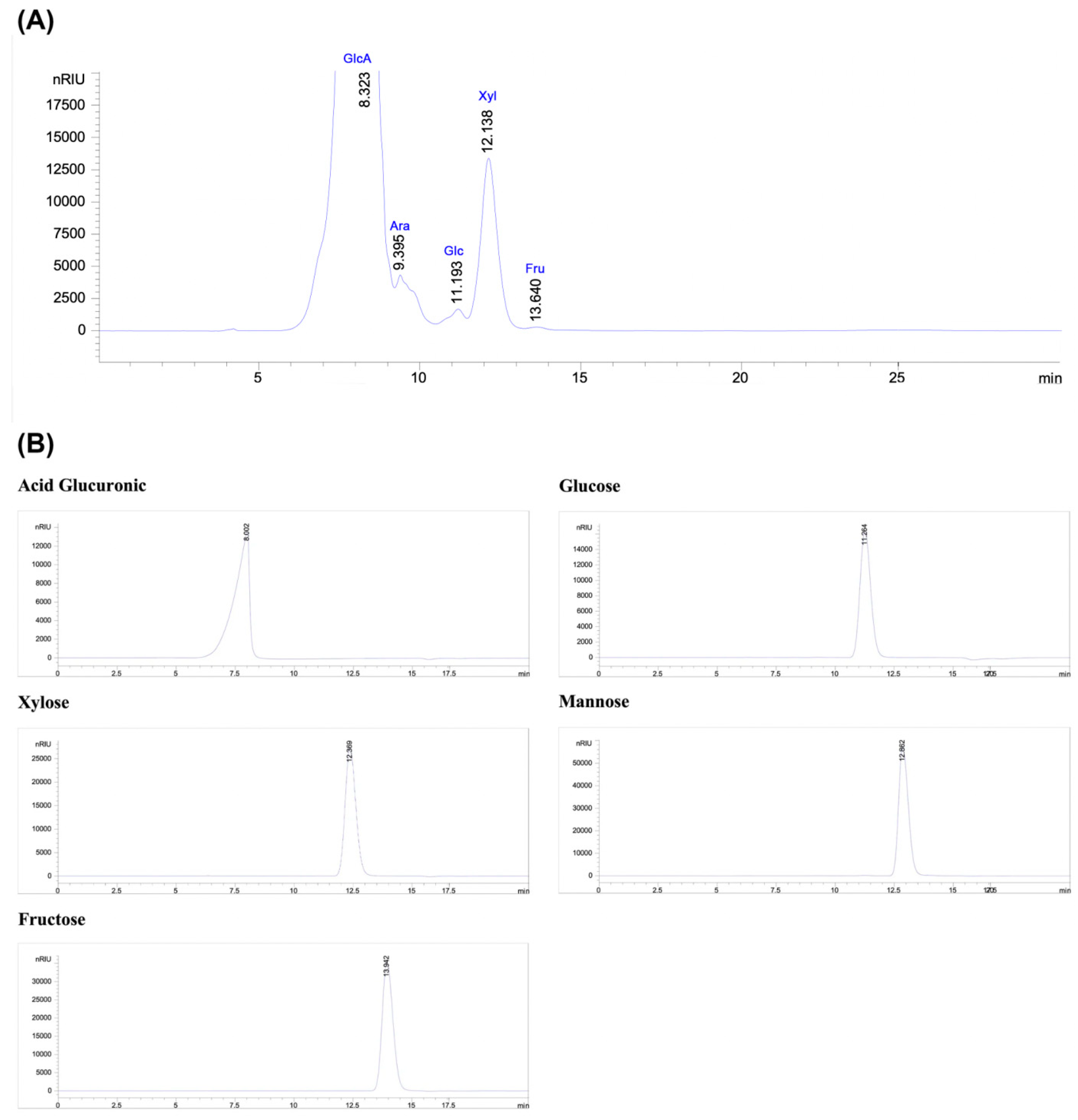
Figure 4.
The average molecular weight of PSOP by steric exclusion chromatography (SEC).
Figure 5.
PSOP Structure characterization. (A) SEM microstructure analysis of PSOP; (B) X-ray diffraction pattern.
Figure 5.
PSOP Structure characterization. (A) SEM microstructure analysis of PSOP; (B) X-ray diffraction pattern.
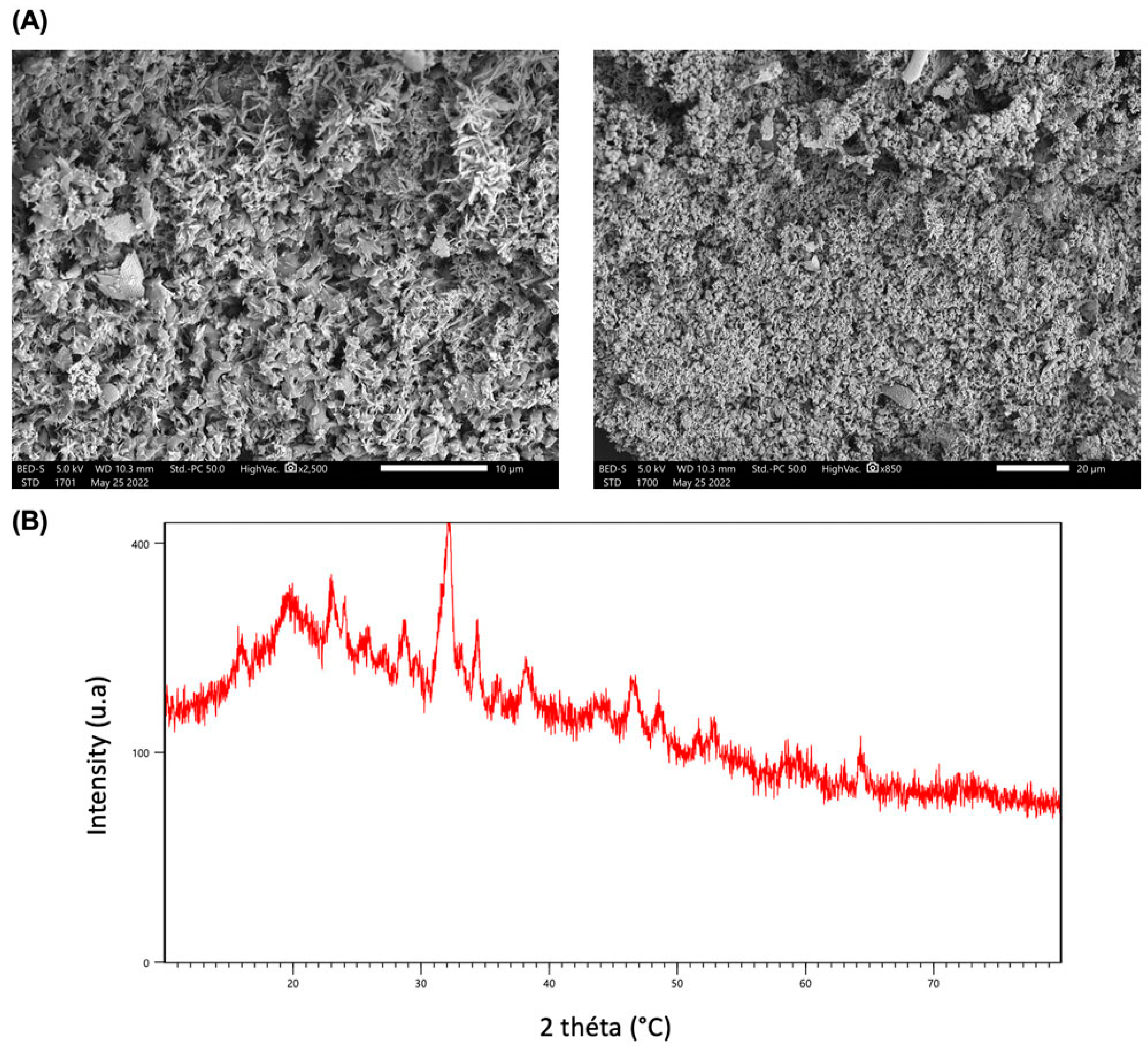
Figure 6.
Antioxidant potentials of PSOP. (A) DPPH radical-scavenging activity; (B) reducing power capacity; and (C) total antioxidant capacity.
Figure 6.
Antioxidant potentials of PSOP. (A) DPPH radical-scavenging activity; (B) reducing power capacity; and (C) total antioxidant capacity.
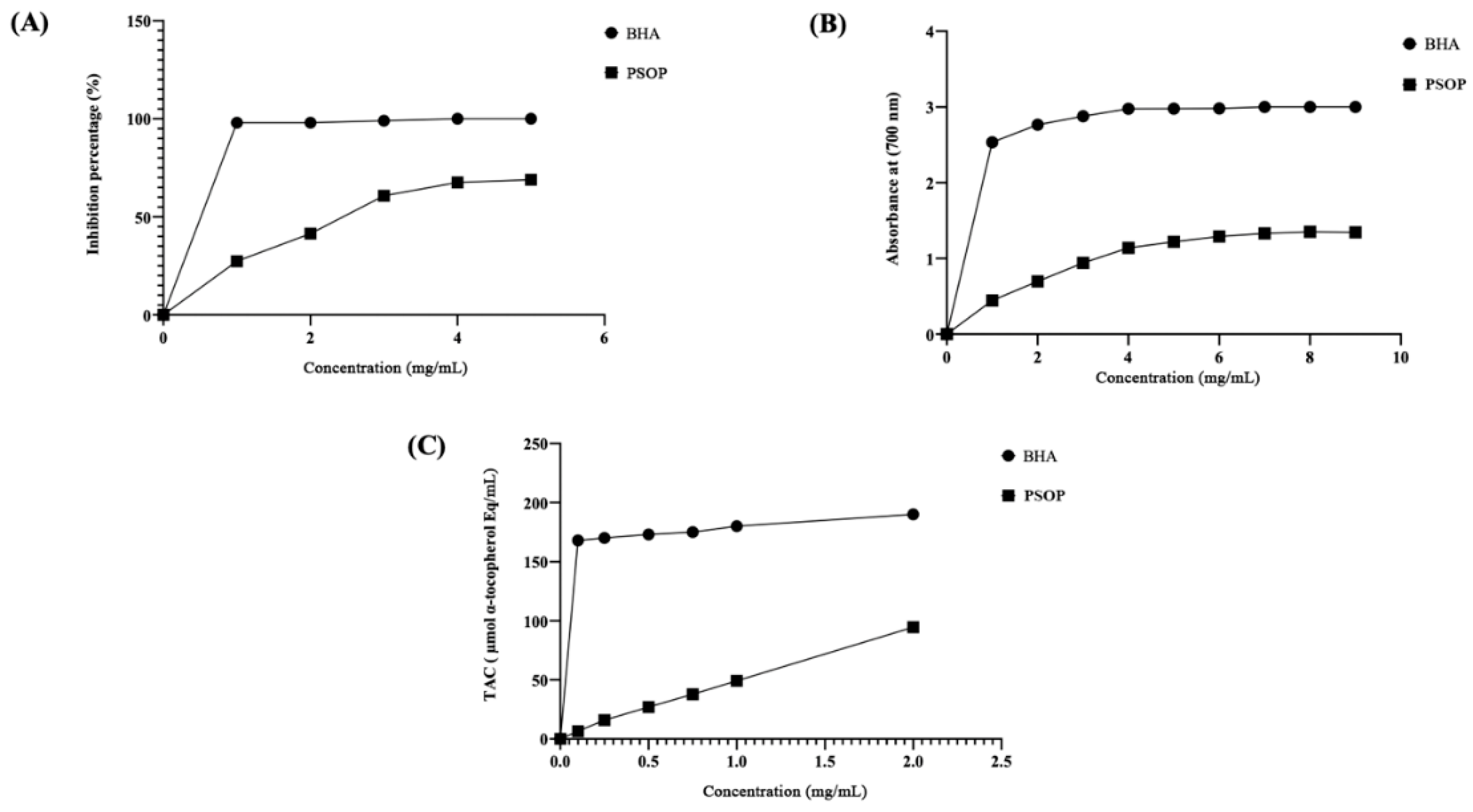
Figure 7.
PSOP stimulated neovascularization of the chorioallantoic membrane in 12-day-old fertilized eggs. (A) distilled water (control); (B) Diclofenac (50 μg/egg); (C) Choriogonadotropin (30 μg/egg); (D) PSOP (25 μg/egg); (E) PSOP (50 μg/egg) and (F) PSOP (100 μg/egg). (I): Original CAM images; (II): Resulting images after processing with AngioTool v 0.6a. The ring fields were photographed (magnification, ×10).
Figure 7.
PSOP stimulated neovascularization of the chorioallantoic membrane in 12-day-old fertilized eggs. (A) distilled water (control); (B) Diclofenac (50 μg/egg); (C) Choriogonadotropin (30 μg/egg); (D) PSOP (25 μg/egg); (E) PSOP (50 μg/egg) and (F) PSOP (100 μg/egg). (I): Original CAM images; (II): Resulting images after processing with AngioTool v 0.6a. The ring fields were photographed (magnification, ×10).
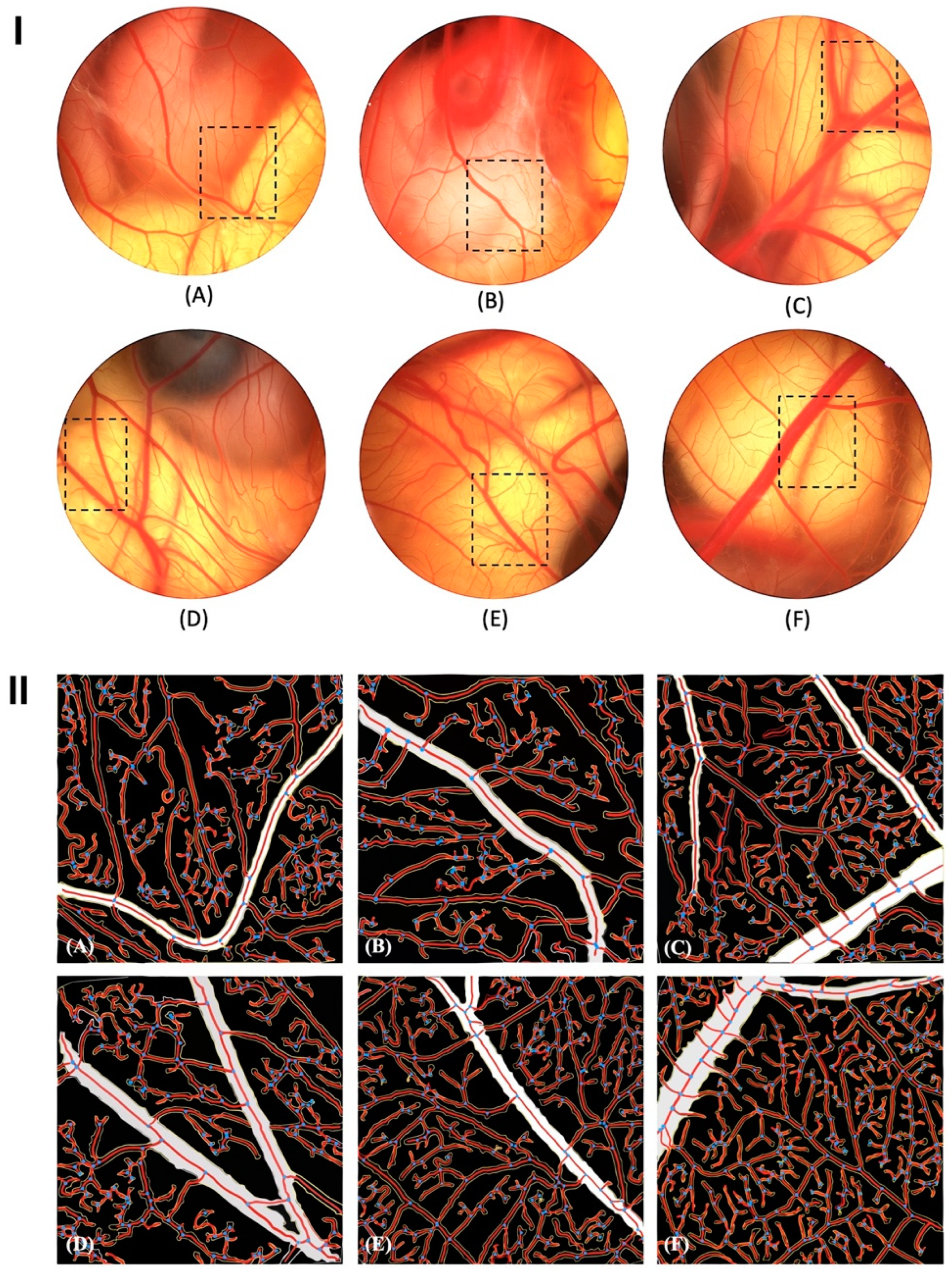
Figure 8.
Effect of treatments on the density and morphology of the vascular network in the chorioallantoic membrane (CAM). (A) Vessel area, (B) Vessel length, (C) Vessel diameter, (D) Number of junctions, and (E) Lacunarity. Data are presented as percentages, and the control group is set to 100 %. CTRL: Control; DIC: Diclofenac; CG: Choriogonadotropin; PSOP1: 25 μg/egg; PSOP2: 50 μg/egg; PSOP3: 100 μg/egg. The data represents the mean ± SEM of six samples in each group. a,b,c,d, indicate significant differences between different groups (p < 0.05).
Figure 8.
Effect of treatments on the density and morphology of the vascular network in the chorioallantoic membrane (CAM). (A) Vessel area, (B) Vessel length, (C) Vessel diameter, (D) Number of junctions, and (E) Lacunarity. Data are presented as percentages, and the control group is set to 100 %. CTRL: Control; DIC: Diclofenac; CG: Choriogonadotropin; PSOP1: 25 μg/egg; PSOP2: 50 μg/egg; PSOP3: 100 μg/egg. The data represents the mean ± SEM of six samples in each group. a,b,c,d, indicate significant differences between different groups (p < 0.05).
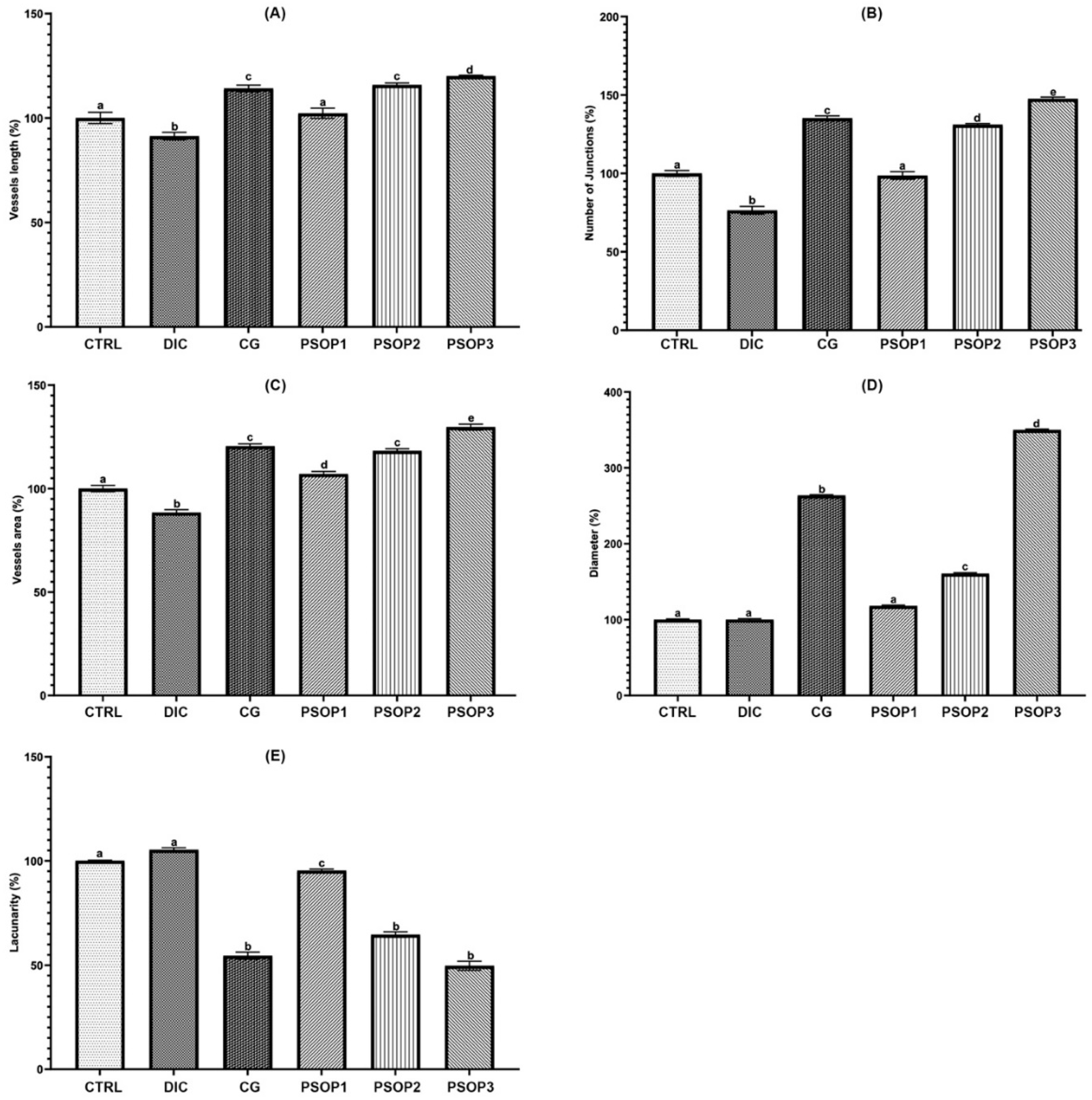
Figure 9.
Representative photographs of the macroscopic appearance of 1.5 cm2 wounds excised on rats at days 1, 5, 7, and 11 of the group treated with physiological serum, glycerol, Cytol Centella, and PSOP hydrogel.
Figure 9.
Representative photographs of the macroscopic appearance of 1.5 cm2 wounds excised on rats at days 1, 5, 7, and 11 of the group treated with physiological serum, glycerol, Cytol Centella, and PSOP hydrogel.
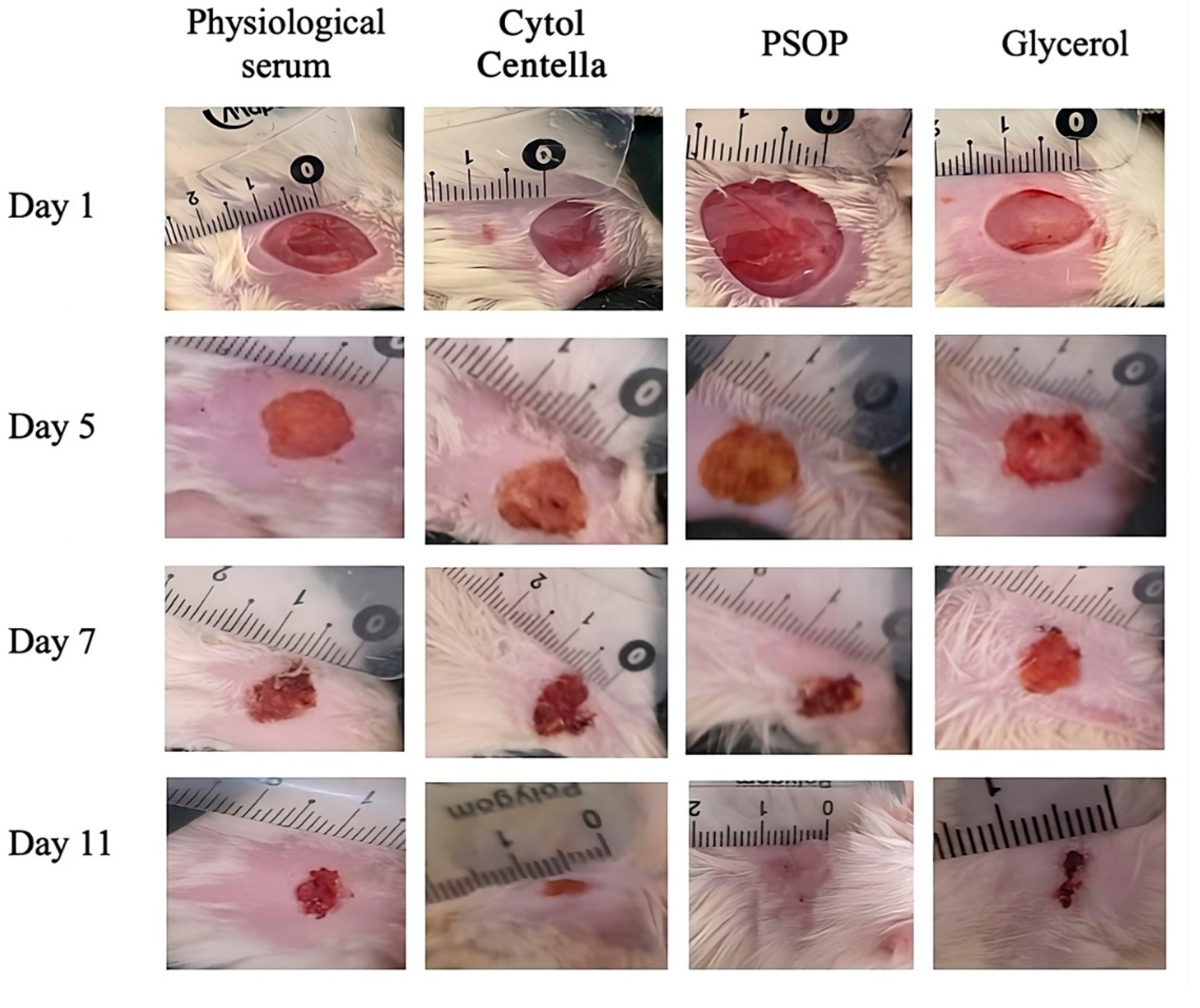
Figure 10.
Percentage of wound area contraction of different groups of rats. Each value represents the mean ± SEM of results from 6 rats. a,b,c, d In the same column indicate significant differences (p < 0.05).
Figure 10.
Percentage of wound area contraction of different groups of rats. Each value represents the mean ± SEM of results from 6 rats. a,b,c, d In the same column indicate significant differences (p < 0.05).
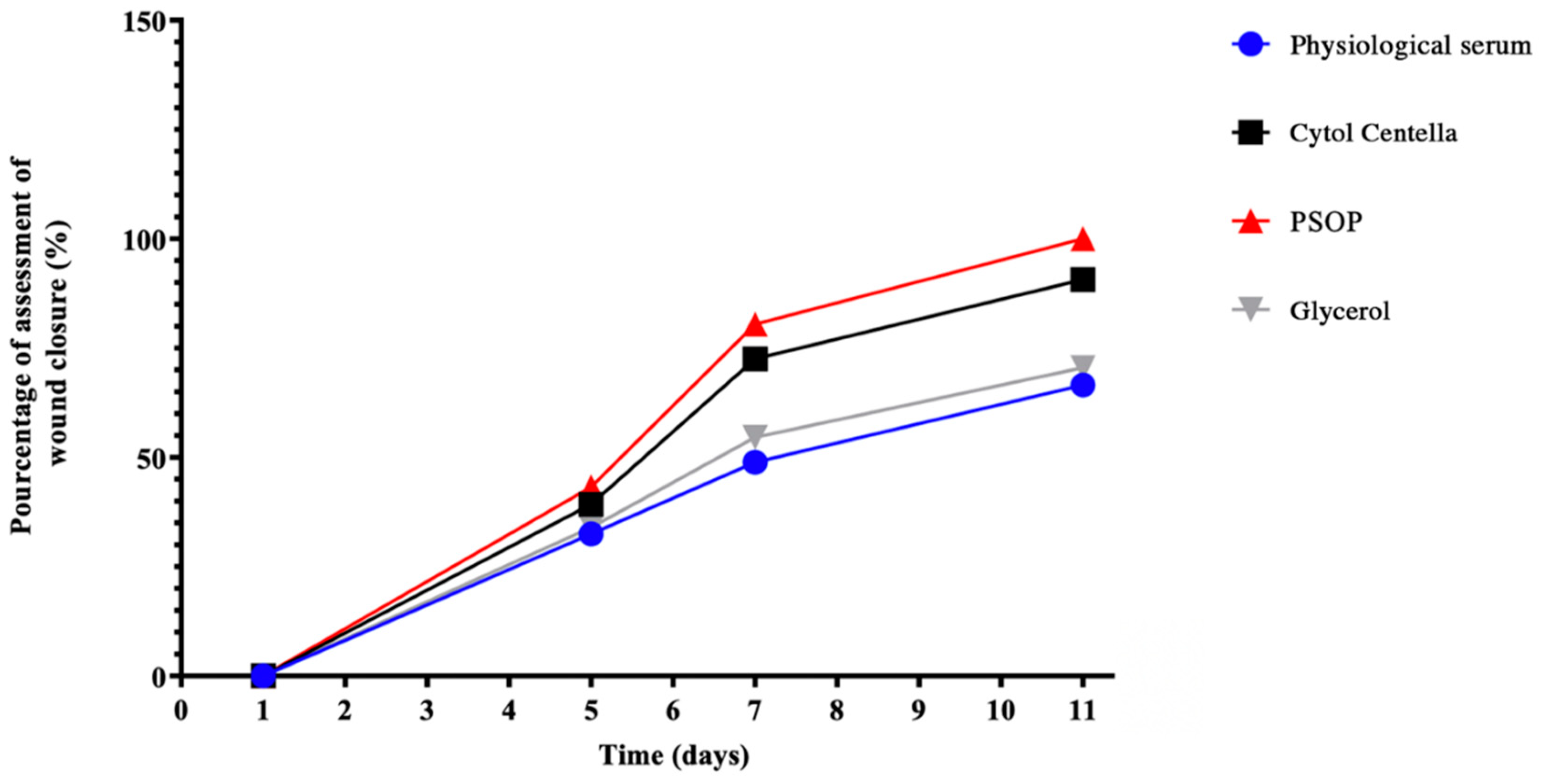
Figure 11.
Hydroxyproline concentrations in biopsies from experimental rat wounds. Different letters on each bar indicate significant differences between treatments (p < 0.05).
Figure 11.
Hydroxyproline concentrations in biopsies from experimental rat wounds. Different letters on each bar indicate significant differences between treatments (p < 0.05).
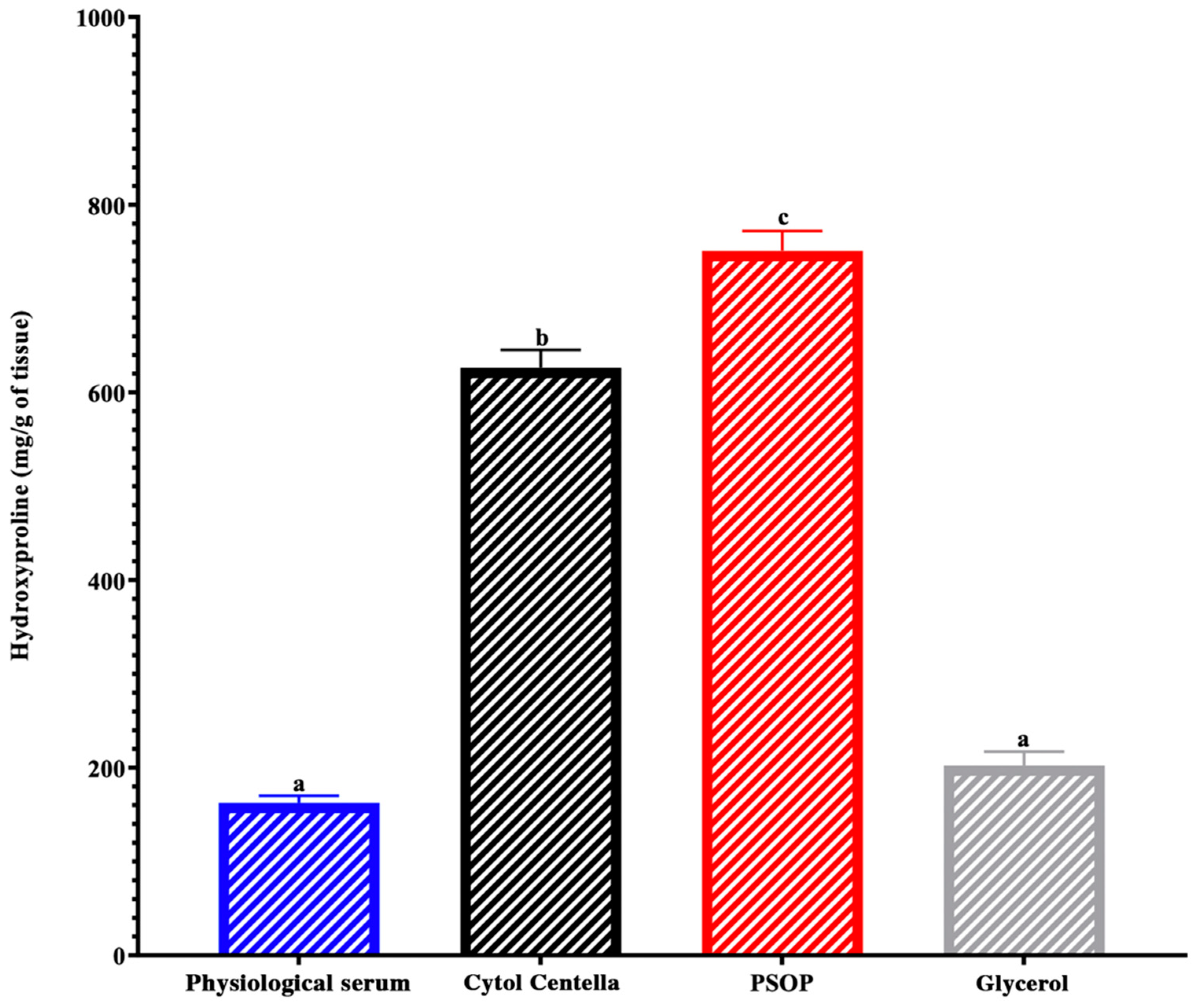
Figure 12.
Representative microphotographs of epidermal and dermal wound structure on day 11 in rats treated with saline, glycerol, Cytol Centella, and PSOP hydrogel. Tissues were visualized at 50 and 100× magnification. Arrows indicate the following: E: epiderm and D: derm.  : Ulceration
: Ulceration  : Inflammatory infiltrate
: Inflammatory infiltrate  : Fibers of collagen
: Fibers of collagen
: Ulceration : Inflammatory infiltrate : Fibers of collagen
Figure 12.
Representative microphotographs of epidermal and dermal wound structure on day 11 in rats treated with saline, glycerol, Cytol Centella, and PSOP hydrogel. Tissues were visualized at 50 and 100× magnification. Arrows indicate the following: E: epiderm and D: derm. : Ulceration : Inflammatory infiltrate : Fibers of collagen
: Ulceration : Inflammatory infiltrate : Fibers of collagen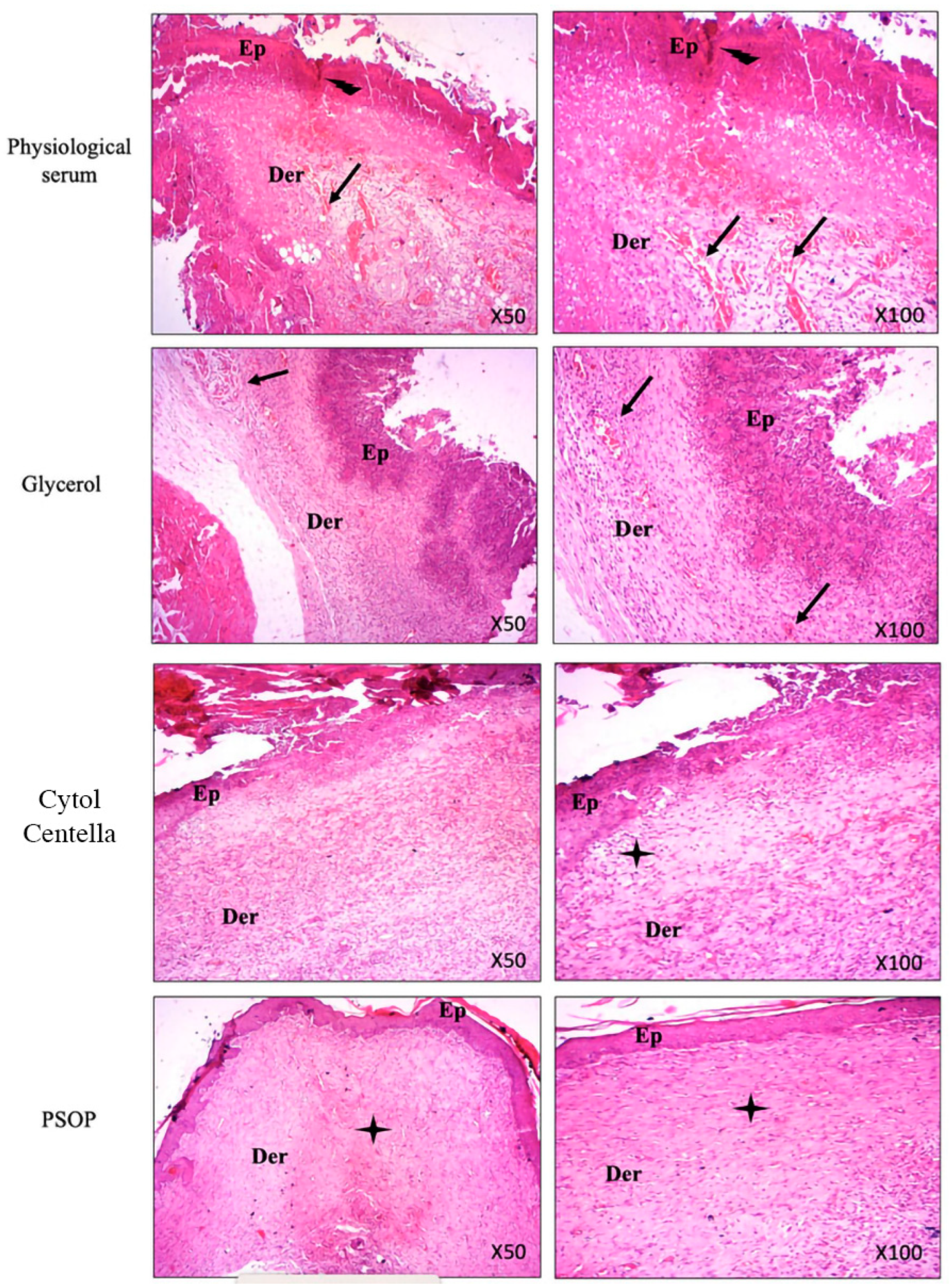
Figure 13.
3D illustration of the pocket regions (A and B) and resulted diagrams of interactions (A’ and B’) of the O. pinnatifida identified monosaccharides with the two targeted receptors: 1cx-2 and 2c7w for cyclooxygenase 2 (COX-2, A) and vascular endothelial growth factor (VEGF, B), respectively.
Figure 13.
3D illustration of the pocket regions (A and B) and resulted diagrams of interactions (A’ and B’) of the O. pinnatifida identified monosaccharides with the two targeted receptors: 1cx-2 and 2c7w for cyclooxygenase 2 (COX-2, A) and vascular endothelial growth factor (VEGF, B), respectively.
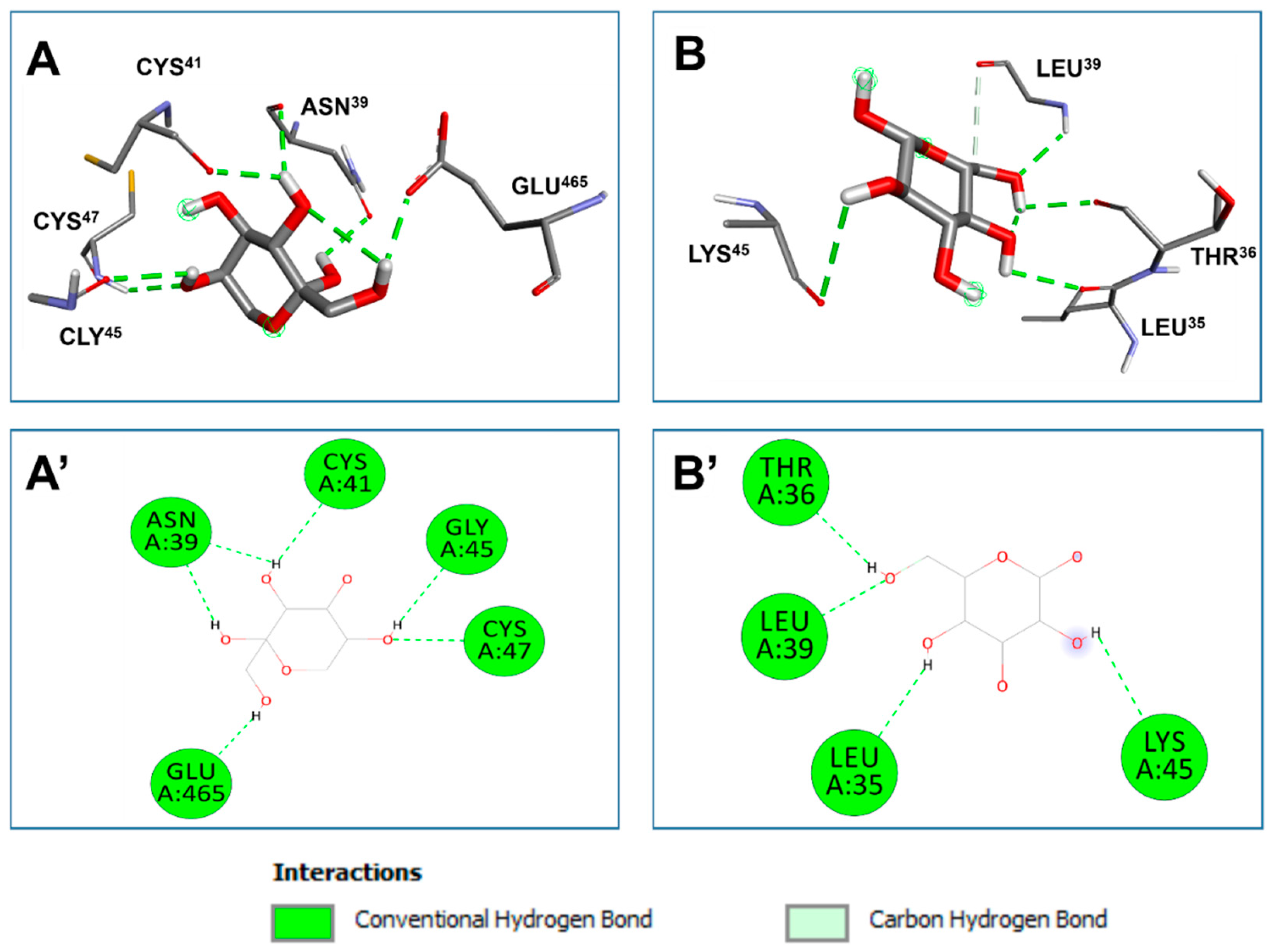

Table 1.
Percentage of blood vessels in CAM assay.
| Groups | Control | Diclofenac (50 μg/egg) |
Choriogonadotropin (30 μg/egg) |
PSOP (25 μg/egg) |
PSOP (50 μg/egg) |
PSOP (100 μg/egg) |
|---|---|---|---|---|---|---|
| Vessel Number (%) | 100a | 71.14 ± 3.97b | 168.36 ± 2.72c | 137.22 ± 3.7d | 200.66 ± 2.73e | 250.66 ± 3.49f |
Data are presented as percentages, and the control group is set to 100 %. Values are expressed as means ± SD for 6 eggs in each group. a, b, c, d, e, f Indicate significant differences between different groups (p < 0.05).
Table 2.
Binding affinity of root mean square deviation of the O. pinnatifida identified monosaccharides with the two targeted receptors: 1cx2 and 2c7w for cyclooxygenase 2 (COX-2) and vascular endothelial growth factor (VEGF), respectively.
Table 2.
Binding affinity of root mean square deviation of the O. pinnatifida identified monosaccharides with the two targeted receptors: 1cx2 and 2c7w for cyclooxygenase 2 (COX-2) and vascular endothelial growth factor (VEGF), respectively.
| Compound No. | COX-2 (1cx2) |
VEGF (2c7w) |
|---|---|---|
| Binding Affinity (kcal × mol−1) | ||
| Arabinose | −5.7 | −4.5 |
| Fructose | −6.4 | −4.5 |
| Glucose | −6.3 | −4.6 |
| Glucoronic acid | −5.6 | −4.4 |
| Xylose | −6.1 | −4.1 |
| RMSD (lower – upper) | ||
| Arabinose | 0.0 - 30.13 | 0.0 - 31.90 |
| Fructose | 0.0 - 30.82 | 0.0 - 9.08 |
| Glucose | 0.0 - 22.70 | 0.0 - 9.14 |
| Glucoronic acid | 0.0 - 41.78 | 0.0 - 14.83 |
| Xylose | 0.0 - 9.86 | 0.0 - 30.91 |
Table 3.
Number of conventional H-bonds, closest interacting residues and distance to closest interacting residue (Å) of the O. pinnatifida identified monosaccharides with the two targeted receptors: 1cx2 and 2c7w, for cyclooxygenase 2 (COX-2) and vascular endothelial growth factor (VEGF), respectively.
Table 3.
Number of conventional H-bonds, closest interacting residues and distance to closest interacting residue (Å) of the O. pinnatifida identified monosaccharides with the two targeted receptors: 1cx2 and 2c7w, for cyclooxygenase 2 (COX-2) and vascular endothelial growth factor (VEGF), respectively.
| Saccharide | No. Conventional H-Bonds |
Closest Interacting Residues |
||
|---|---|---|---|---|
| Interacting Residues | Closest residue (Distance, Å) | No. closest interacting residues | ||
| Cyclooxygenase 2 (COX-2) | ||||
| Fructose | 6 | CYS47, ASN39, GLU465, ASN39, CYS41, GLY45 | GLU465:OE1 (2.072) |
5 |
| Vascular Endothelial growth factor (VEGF) | ||||
| Glucose | 4 | LEU39, THR36, LEU35, LYS45, LEU39 | LEU39:HN (2.247) |
4 |
| Bold residues: amino acids interacting with conventional H-bond | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



