
Submitted:
24 October 2024
Posted:
25 October 2024
You are already at the latest version
Abstract
The gastrointestinal (GI) tract may function as a sensory organ responding to changes in luminal content. Its ability to regulate physiological functions is, in part, determined by the bitter chemosensory capacity of the gut mucosa. We mapped the regional variation of GI tract bitter taste receptors (TAS2Rs) in order to better understand their relationship to GI function and the gut-brain axis. Mucosal biopsies from the stomach (fundus, body and antrum), duodenum (D2), proximal jejunum, terminal ileum, colon (ascending, transverse, and sigmoid), and rectum were collected from volunteers with no previous gut disorders who were undergoing routine screening by gastroscopy or colonoscopy. Nanostringtm analysis of TAS2R transcription was conducted on 300 biopsies taken from 28 volunteers. TAS2Rs showed a diverse transcription profile throughout the human GI tract. Two archetypes of gut chemosensing existed with several individuals displaying a more complete TAS2R profile in all regions of the gut. Additionally, regional variation was observed in several TAS2Rs that correlated with cytogenetic location. These data show significant inter-individual and regional variation in gut bitter chemosensory systems reflecting differing luminal sensing capacity in different gut regions and between people.
Keywords:
bitter taste receptor
; gastrointestinal tract
; TAS2R
; chemosensing
1. Introduction
The gastrointestinal (GI) tract represents a major interface between the human body and the external environment, functioning as both a vessel for the digestion and absorption of dietary nutrients as well as acting as a major chemosensory organ [1]. Gut chemosensory mechanisms detect a diverse array of compounds present in the gut lumen and elicit changes to both local and whole-body homeostasis in response to the components detected. These gut chemosensory responses are sensitive to both type and relative location of stimulus, as occurs for example when exposure of gastric tissue to taste modalities stimulate the release of the hunger promoting hormone ghrelin and increases food consumption [2,3], whereas the presence of these same tastes in the ileum can stimulate the release of the hunger suppressing hormone glucagon-like peptide-1 (GLP-1) and decrease food consumption [3,4]. These distinct response profiles suggest that characterization of not only chemosensory systems present, but also their localization is important in understanding gut chemosensing.
The gut chemosensory system is comprised of various receptors and transporters [5,6,7], with growing interest in flavor and taste modalities such as bitterness [8]. In humans, bitter taste is detected by a family of 25 bitter taste receptors (TAS2Rs) that respond to both dietary and non-dietary compounds [9]. Lingual bitter taste is a sensation that potentially signals the presence of a toxin and is more pronounced in infants and children [10,11]. Extra-lingual gut TAS2Rs are present in the gut mucosa including within the specialized chemosensing enteroendocrine cells [12], that represent less than 1% of the total gut epithelial cell population and are broadly defined by the peptide hormones they contain [13,14]. These peptide hormones include, the appetite regulating cholecystokinin (CCK), GLP-1, peptide tyrosine tyrosine (PYY) and ghrelin, the intestinal motility modulating neurotensin and serotonin, and the gut secretagogues, gastrin and secretin [15,16]. The presence of extra-lingual TAS2Rs in enteroendocrine cells infers bitter taste modalities may be gut active and affect numerous physiological processes, ranging from the regulation of ingestive behavior [3,17,18,19] and gut motility [20] to adverse GI responses to pharmaceutical products [21] and chemical absorption [22]. Additionally, single-cell sequencing of human intestinal tissue has also demonstrated the presence of specific TAS2Rs in intestinal goblet and Paneth cells, and rodent studies have shown TAS2R presence in Tuft cells [23,24], suggesting a potential role in the modulation of gut secretion, mucin homeostasis, and immunity [25,26,27].
Previous studies have detected the presence of TAS2Rs within the human gastrointestinal tract [28,29,30,31,32], and a recent review indicates likely regional variation in human TAS2Rs [32]. However, due to challenges mapping the lowly expressed TAS2R transcripts, it is still unclear the degree to which TAS2Rs vary by region of the GI tract, whether all or a subset of TAS2Rs are present in specific regions of the GI tract mucosa, and if there is conserved or differential TAS2R expression between individuals.
Here we conduct a comprehensive study of human intestinal TAS2R expression, examining biopsy samples from 28 healthy male and female adults taken from 10 regions throughout the human GI tract. We show that TAS2R transcripts are present, that specific TAS2R transcripts vary between gut regions and highlight a subset of individuals who exhibit a broader TAS2R transcriptional profile.
2. Results
2.1. Participants and Biopsy Sampling

Table 1.
Characteristics of participants and biopsy sampling (n = 28, BMI = Body Mass Index).
| Participant Number |
Age, years | Sex, M/F |
BMI, kg/m2 | Procedure Gastroscopy |
Procedure Colonoscopy |
Number of biopsy samples (total included) | |
| 1 | 24 | M | 20.1 | X | 10 | ||
| 2 | 55 | M | 28.0 | X | 10 | ||
| 3 | 49 | M | 27.0 | X | 10 | ||
| 4 | 18 | M | 28.1 | X | X | 20 | |
| 5 | 48 | F | 28.5 | X | 10 | ||
| 6 | 58 | F | 29.2 | X | 10(8) | ||
| 7 | 21 | F | 28.0 | X | 10 | ||
| 8 | 62 | F | 20.7 | X | X | 20 | |
| 9 | 35 | F | 23.1 | X | 10 | ||
| 10 | 60 | M | 29.3 | X | 10 | ||
| 11 | 21 | F | 23.3 | X | 10 | ||
| 12 | 35 | F | 23.8 | X | 10 | ||
| 13 | 44 | M | 28.4 | X | 10 | ||
| 14 | 56 | F | 24.1 | X | 10 | ||
| 15 | 56 | F | 21.3 | X | 10 | ||
| 16 | 56 | M | 24.4 | X | 10 | ||
| 17 | 57 | M | 24.7 | X | 10 | ||
| 18 | 44 | M | 24.0 | X | 10 | ||
| 19 | 61 | M | 23.3 | X | 10 | ||
| 20 | 64 | F | 26.6 | X | 10 | ||
| 21 | 65 | F | 23.5 | X | 10 | ||
| 22 | 55 | M | 25.3 | X | 10 | ||
| 23 | 44 | F | 29.8 | X | 10 | ||
| 24 | 52 | F | 24.5 | X | 10 | ||
| 25 | 30 | M | 30.0 | X | 10 | ||
| 26 | 41 | M | 29.9 | X | 10 | ||
| 27 | 63 | M | 28.2 | X | 10 | ||
| 28 | 55 | F | 27.2 | X | 10 | ||
| Average (mean, range) | 47.2, 18-65 |
25.8, 20.1-30 | |||||
| Total | 14/14 | 15 | 15 | 300 (298) | |||
* Proximal jejunum samples were excluded due to failure of QA.
2.2. TAS2R Transcription in the GI Tract
Dual biopsies taken from a given location from the same individual were pooled and RNA extracted to make 150 total RNA samples. These samples were analysed for mRNA TAS2R transcripts with 1 sample (Participant 6, Proximal jejunum) being excluded due to extremely low transcript count affecting normalisation and housekeeping procedures. Nanostring analysis demonstrated that all TAS2Rs were present in at least some individuals at all locations (Supplementary Table S3). Total RNA counts for all TAS2Rs in all locations averaged approximately 8000 per 600ng of biopsy total RNA. On average the highest transcribed TAS2R was TAS2R14 (1356 ± 51 counts) and the combined count for TAS2R sub-family of TAS2R43, 44, 45, 46 and 47 (1274 ± 39 counts) that were all detected by the same probe (Table 2). The highest single reading was 1798±218 counts for TAS2R14 was detected in the Gastric Fundus (Table 2). TAS2R1, 16, 39, 42, 60, 7 and 8 were generally lowly expressed, however, samples from some individals exhibited notely higher expression levels (Table 2).
2.3. Inter-Regional Variation of GI TAS2Rs
Mean TAS2R RNA counts across the 10 GI regions biopsied are shown in Table 2. (Table 2). For individual TAS2Rs significant (P < 0.05) regional variation was seen in all TAS2Rs except for TAS2R4 (Table 2). Notably, TAS2R40 and 38 were significantly reduced in the Gastric Fundus when compared to all other regions (P < 0.05). TAS2R41 was significantly higher in the 3 small intestinal regions of the D2 Duodenum, Proximal Jejunum and Terminal Ileum, relative to any of the gastric or large intestinal regions sampled (P < 0.05) (Table 2). TAS2R60 was significantly higher in the Terminal Ileum when compared to any other region and higher in the small intestine when compared to the gastric samples taken (P < 0.05) (Table 2). TAS2R8 and 9 were significantly higher in the Gastric Fundus when compared to all regions except the Gastric Body (P < 0.05) (Table 2).
Table 2.
Regional expression of TAS2R from human gut biopsy samples as determined by Nanostring® RNA analysis. 13 individuals gave gastroscopy samples (Gastric Fundus through to Proximal Jejunum), 13 individuals gave colonoscopy samples (Terminal Ileum through to Rectum), 2 individuals gave both gastroscopy and colonoscopy samples. (mean count ± standard deviation).
Table 2.
Regional expression of TAS2R from human gut biopsy samples as determined by Nanostring® RNA analysis. 13 individuals gave gastroscopy samples (Gastric Fundus through to Proximal Jejunum), 13 individuals gave colonoscopy samples (Terminal Ileum through to Rectum), 2 individuals gave both gastroscopy and colonoscopy samples. (mean count ± standard deviation).
| Region | Gastric Fundus |
Gastric Body |
Gastric Ant. |
D2 Duo. |
Prox. Jejunum |
Term. Ileum |
Assen. Colon |
Trans. Colon |
Sig. Colon |
Rectum |
Average |
| TAS2R1 | 23 | 14 | 20 | 16 | 29 | 11 | 9 | 7 | 5 | 9 | 14 |
| ±13a | ±8ab | ±15bc | ±9abc | ±17a | ±7bc | ±6c | ±7c | ±3c | ±5bc | ±3 | |
| TAS2R10 | 373 | 330 | 227 | 307 | 300 | 305 | 265 | 315 | 237 | 256 | 292 |
| ±68a | ±49ab | ±55c | ±56abc | ±88bc | ±69abc | ±48abc | ±73abc | ±42bc | ±48bc | ±19 | |
| TAS2R13 | 1031 | 925 | 649 | 810 | 811 | 988 | 751 | 752 | 681 | 736 | 813 |
| ±123a | ±92ab | ±68bc | ±105abc | ±147abc | ±146bc | ±92abc | ±84abc | ±72abc | ±108c | ±34 | |
| TAS2R14 | 1798 | 1553 | 1147 | 1291 | 1383 | 1527 | 1240 | 1291 | 1129 | 1206 | 1356 |
| ±218a | ±134ab | ±86c | ±146cd | ±211bcd | ±172abcd | ±143cd | ±172cd | ±110c | ±118c | ±51 | |
| TAS2R16 | 45 | 33 | 35 | 25 | 51 | 25 | 28 | 27 | 16 | 21 | 30 |
| ±26a | ±17ab | ±26bcd | ±15bcd | ±33abc | ±16d | ±18d | ±19d | ±10d | ±13cd | ±6 | |
| TAS2R19 | 718 | 668 | 511 | 533 | 554 | 691 | 514 | 555 | 497 | 505 | 575 |
| ±68a | ±48ab | ±48ab | ±60ab | ±106ab | ±78ab | ±63ab | ±66ab | ±53ab | ±59b | ±21 | |
| TAS2R20 | 1444 | 1292 | 955 | 1003 | 1052 | 1294 | 1010 | 1037 | 962 | 1001 | 1106 |
| ±110a | ±90a | ±72b | ±101b | ±149b | ±124ac | ±92bc | ±107b | ±85b | ±116b | ±35 | |
| TAS2R3 | 583 | 543 | 430 | 380 | 358 | 597 | 462 | 506 | 430 | 433 | 473 |
| ±30ab | ±26ab | ±29abc | ±21abc | ±37c | ±32a | ±26abc | ±35abc | ±34abc | ±54bc | ±12 | |
| TAS2R38 | 91 | 88 | 99 | 112 | 153 | 226 | 145 | 133 | 125 | 126 | 130 |
| ±29a | ±17b | ±16bc | ±13bc | ±20cd | ±28d | ±17cd | ±14cd | ±13bcd | ±17bc | ±7 | |
| TAS2R39 | 44 | 26 | 32 | 24 | 40 | 20 | 25 | 24 | 14 | 22 | 27 |
| ±23a | ±14ab | ±24bc | ±15bc | ±26bc | ±13c | ±16c | ±16c | ±9c | ±16bc | ±5 | |
| TAS2R4 | 615 | 598 | 469 | 503 | 520 | 637 | 480 | 545 | 439 | 515 | 532 |
| ±31 | ±26 | ±28 | ±13 | ±42 | ±28 | ±19 | ±38 | ±29 | ±77 | ±12 | |
| TAS2R40 | 68 | 71 | 86 | 100 | 122 | 124 | 98 | 112 | 98 | 88 | 97 |
| ±25a | ±14b | ±18bc | ±12bc | ±19c | ±15c | ±13bc | ±16bc | ±9bc | ±14bc | ±5 | |
| TAS2R41 | 33 | 24 | 27 | 83 | 98 | 145 | 27 | 25 | 21 | 23 | 50 |
| ±15a | ±8a | ±13a | ±9b | ±14b | ±30b | ±9a | ±10a | ±5a | ±6a | ±5 | |
| TAS2R42 | 49 | 35 | 47 | 34 | 58 | 27 | 30 | 31 | 14 | 24 | 35 |
| ±27a | ±20ab | ±36bc | ±21bc | ±38bc | ±18c | ±20c | ±20c | ±9c | ±15bc | ±7 | |
| TAS2R43* | 1567 | 1440 | 1095 | 1153 | 1082 | 1618 | 1241 | 1211 | 1132 | 1198 | 1274 |
| ±111ab | ±86ab | ±91ab | ±110ab | ±162a | ±122b | ±119ab | ±106ab | ±90ab | ±166ab | ±39 | |
| TAS2R5 | 490 | 469 | 365 | 360 | 364 | 531 | 418 | 475 | 415 | 437 | 433 |
| ±29ab | ±23abc | ±19d | ±20d | ±31d | ±37a | ±20abc | ±43bcd | ±38cd | ±48bcd | ±11 | |
| TAS2R50 | 789 | 683 | 519 | 567 | 591 | 707 | 572 | 598 | 513 | 548 | 609 |
| ±101a | ±77abc | ±56d | ±80bcd | ±123bd | ±86ac | ±80bcd | ±89bcd | ±65d | ±84d | ±27 | |
| TAS2R60 | 30 | 23 | 25 | 39 | 52 | 112 | 39 | 31 | 29 | 33 | 41 |
| ±13ab | ±9ab | ±14a | ±13bc | ±19cd | ±24d | ±18abc | ±12abc | ±9abc | ±8bc | ±5 | |
| TAS2R7 | 55 | 41 | 49 | 46 | 85 | 35 | 34 | 47 | 26 | 30 | 45 |
| ±30ab | ±24ac | ±34ac | ±20bd | ±41d | ±23c | ±23c | ±31c | ±18c | ±20ac | ±8 | |
| TAS2R8 | 54 | 37 | 47 | 34 | 52 | 44 | 30 | 42 | 23 | 33 | 40 |
| ±30a | ±22ab | ±35bc | ±22bc | ±34bc | ±29bc | ±20c | ±29c | ±15c | ±23bc | ±8 | |
| TAS2R9 | 100 | 57 | 60 | 41 | 78 | 48 | 48 | 48 | 33 | 29 | 54 |
| ±40a | ±29ab | ±43bc | ±26bc | ±52bc | ±30bc | ±31bc | ±31c | ±21bc | ±17bc | ±10 | |
| Average | 473 | 426 | 328 | 355 | 373 | 463 | 356 | 372 | 326 | 346 | |
| ±35 | ±30 | ±23 | ±25 | ±30 | ±32 | ±26 | ±27 | ±23 | ±26 |
a,b,c,d Signifies GI regions that are significantly different (P < 0.05) for a given TAS2R. * TAS2R43 includes all sub-members (TAS2R43, 44, 45, 46 and 47) of this taste receptor family. Ant. = Antrum, Duo.= Duodenum, Term. = Terminal, Trans. = Transverse, Sig. = Sigmoid.
2.4. Inter-Individual Variation in TAS2Rs
To investigate if inter-individual variation occurs in human gut epithelia TAS2Rs, particpants were compared for their TAS2R transcript profile. Several individuals appear to have altered pattern of TAS2R transcription, with 4 of 28 participants (# 5, 6, 8 ,18) having all but TAS2R1 being consistantly detected in their gut biopsies. These biopsy samples were from participants who volunteered samples from colonoscopies (2), gastroscopy (1), and from both a colonoscopy and gastroscopy (1). The presence of these specific TASR2s is consistent across biopsies from different regions and occurs in samples from both colonoscopies and gastroscopies. The low number of individuals exhibiting this variation, prevents any inference regarding sex, age or ethnicity (Supplementary Figure S2).
Where as TAS2R 16, 39, 42, 7, 8 and 9 were generally below the LOD in biospies taken from the other 24 individuals. (Figure 1).
Where evidence of inter-individual variation occurred (ANOVA P<0.05, TAS2R7, 8, 9, 16, 39, 42), we further investigated the observed variation by cluster analysis using multiple analytical techniques. Samples from gastroscopy and those from colonoscopy procedures were analysed independently to determine if any clustering explained the variation in transcript count number observed. Bivariate cluster plot analysis identified 2 clusters that were present for both the gastroscopy and the colonoscopy samples for TAS2R 7, 8, 9, 16, 39 and 42 and were distinct from other TAS2Rs present (Figure 2a,b). For colonscopy sites the 2 clusters explained 97.27% of the variation and for gastrscopsy sites 94.21% of point variation is explained (Figure 2a,b).
2.5. Cytogeneic and Phylogenetic Variation of GI TAS2Rs
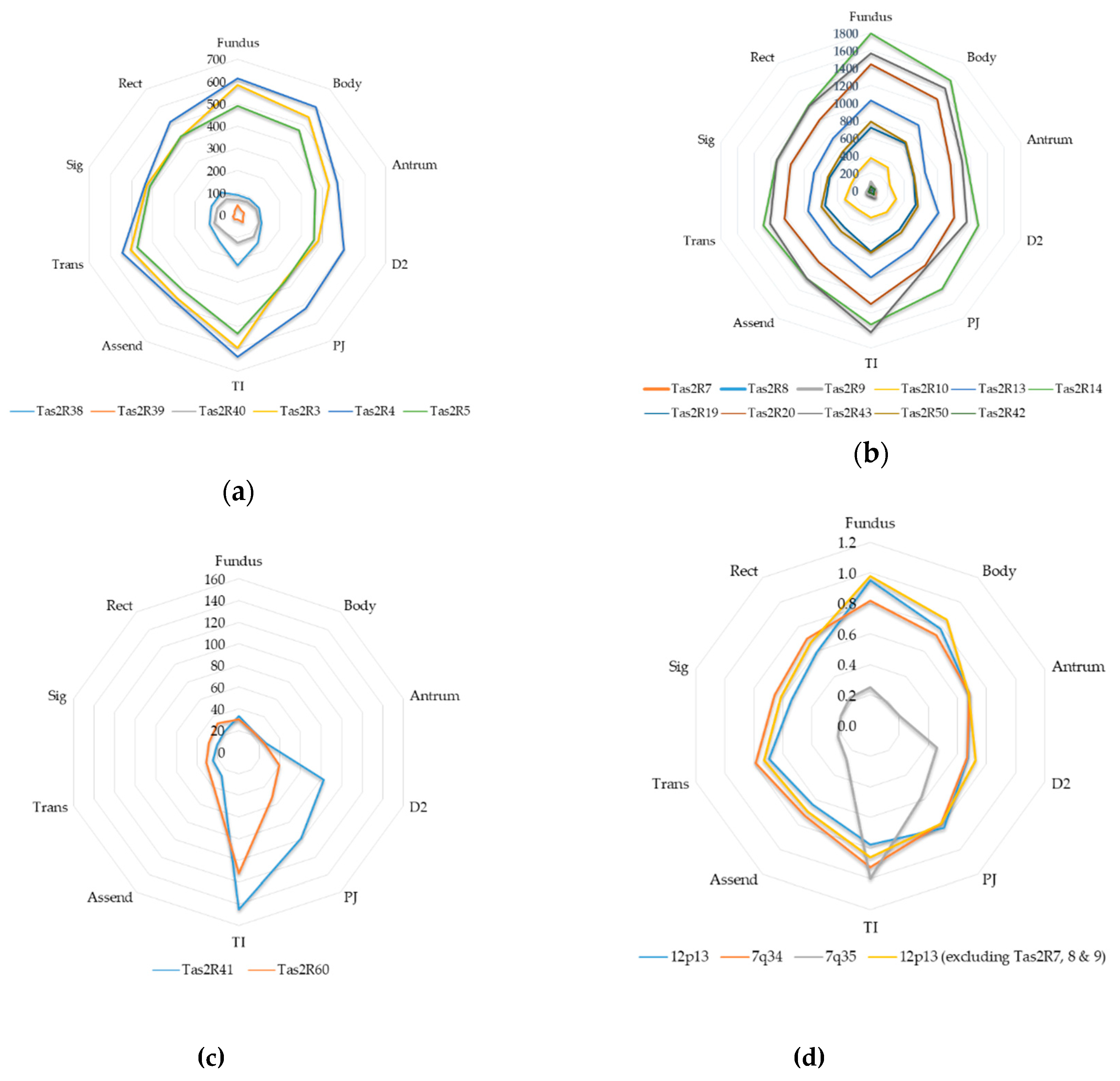
Gross patterns relating to overall TAS2R transcription and cytogenetic location were observed in specific groups of TAS2Rs. TAS2Rs present within the same chromosomal region showed a similar overall pattern of gut transcription (Figure 3). TAS2Rs located on 12p13 had higher transcription counts in the Gastric Fundus and Terminal ileum and recorded transcription numbers that ranged from being the most highly transcribed through to ones being minimally detectable in most individuals (Figure 3a). TAS2Rs present on 7q34 exhibited highest transcription numbers from Gastric Fundus, Gastric Antrum and Terminal Ileum samples. They also displayed 2 distinct groupings with TAS2R38, 39 and 40 being less transcribed when compared to TAS2R3, 4 and 5 (Figure 3b). The transcript profile for the 7q35 localized TAS2R41 and 60 is notably different from those TAS2Rs on either 12p13 or 7q34, with transcription being primarily in the small intestinal samples (Figure 3c). TAS2R1 and TAS2R16 are present within chromosomal locations 5p15 and 7q31 respectively, are remote from other TAS2Rs, and average below set LOD for all but one gut region, making assessment of cytogenetic relationship ambiguous (Table 2). Averaging and standardizing counts as a ratio to the most highly transcribed region showed that the transcription profiles are similar between 12p13 and 7q34 but very different from those of 7q35 (Figure 3d). TAS2Rs present on 12p13 were both highly and lowly transcribed and did not show consistent regional variation based on chromosomal location, however higher and lower transcript numbers did relate to their phylogenetic relationship. Of the TAS2Rs present on 12p13, the more closely related TAS2R 14, 12, 49, 48, 50 and the TAS2R43 sub-family all showed higher transcription count compared to the other 12p13 TAS2Rs of 7, 8, 9, 10 and 42. This grouping also reflected higher relative transcription in the Gastric Fundus, Gastric Body and Terminal Ileum (Table 2). TAS2R41 and 60 show close phytogenic relationships, are located on chromosome 7q35, are lowly transcribed and show significant higher transcription in the 3 small intestinal and in the terminal ileum respectively. Additionally, 4 of the 6 TAS2Rs that showed inter-individual variation (7, 8, 9 and 42) are present on 12p13 (Figure 3b) with TAS2R7, 8 and 9 having close phylogenetic relationship.
3. Discussion
Gastrointestinal tract bitter chemosensing relates to multiple aspects of gut physiology [3,33,34,35,36], it may affect gut-brain signaling and regulate whole body homeostasis [37,38]. Preceding studies have shown the presence of human TAS2Rs in biopsy samples from the stomach [39], small intestine[31] and colon [29], providing evidence for roles in intestinal function, immunity and feeding behavior. Given the broad scope of the current study and the use of a sensitive detection method that allows for the assessment of lowly transcribed receptors, the results presented here extend our knowledge of human GI TAS2R expression. We detect transcripts for most TAS2Rs in all regions of the GI tract, show novel expression patterns reflecting differential TAS2Rs expression based on GI region and chromosomal location, and highlight for the first time 2 archetypes of GI bitter sensing.
TASR2s are expressed throughout the GI tract.
In general agreement with prior work examining GI expression of human TAS2Rs, we show gastric, small intestinal and colonic expression, however several differences in TAS2R expression profile present that are worthy of discussion. Expression of TAS2R7, TAS2R10, TAS2R14, TAS2R43 and TAS2R46 was shown in the Gastric Fundus by Liszt et al. 2017 [39] using q-PCR assessment of 2 human biopsy samples. As compared to Liszt et al. 2017 [39] we show expression of TAS2R10, 14 and detect the presence of the TAS2R43 superfamily in the Gastric Fundus. Additionally, we observe variable TAS2R7 expression that is not reported in Liszt et al and may reflect the limited sample number taken during their work. Le neve et al. 2010 [31], using pooled RNA samples from 4 male donors, showed TAS2R14 but not TAS2R7 expression in human Duodenum and Jejunum samples. Our results are in line with these, showing TAS2R14 present in the small intestinal samples, whereas, TAS2R7 was not detected in samples taken from the majority of participants. Kaji et al. 2009 [30] , Latorre et al. 2016 [12] and Rozengurt et al. 2006 [29] reported the presence of TAS2Rs in the large bowel. Kaji et al. 2009 [30] showed TAS2R38 to have consistent transcription across the Ascending Colon, Sigmoidal Colon and Rectum, and showed weak banding for TAS2R1 and 4 [30]. Latorre et al. 2016 [12], observed TAS2R38 in enteroendocrine cells of the lower Colon that they confirm with antibody staining. Rozengurt et al. 2006 [29] showed 18 out of the 25 human TAS2Rs were present in a pooled sample of commercially available human Colon RNA. The data presented here is broadly in agreement with the previous published assessments of large bowel TAS2R expression. We find evidence for the presence of TAS2R4 and TAS2R38, but not TAS2R1 in samples from most participants. Of the 7 TAS2Rs that Rozengurt et al. [29] did not detect, we also do not identify TAS2R 1, 7, 8, 9, 16 and 41 in the majority of colonic samples. Conversely, we show that TAS2R14, undetected in Rozengurt’s study, was the most abundant in our data. The data presented here is also in general agreement with consensus data of human GI tract TAS2R expression reported in a recent review [32].
Gastrointestinal TAS2Rs exhibit regional variation.
TAS2R transcription showed significant variation along the length of the human GI tract. Generally, TAS2Rs exhibit two gross patterns of transcription with the highest transcript being in the Gastric Fundus, Gastric Body and Terminal Ileum for TAS2R3, 4, 5, 13, 14, 19, 20, 43 and 50, whereas other TAS2Rs were elevated in the small or large intestinal regions (TAS2R38, 40, 41 and 60). Overall, the Gastric Fundus and the Terminal Ileum recorded the highest TAS2R counts, suggesting enhanced chemosensing capacity in these gut regions that may reflect likely exposure to dietary and microbiome derived compounds respectively. Interestingly, TAS2R38 was most highly transcribed in the terminal Ileum, is a known receptor for amine moieties containing molecules (such as acetythiourea and caprolactum) [9] and its allelic haplotype is implicated in respiratory response to bacterial infection [40] . This, combined with the TAS2R38’s specificity towards bacterial quorum sensing compounds [41], suggests TAS2R38 may have a potential role in bacterial overgrowth sensing in the terminal Ileum. A degree of SI regionality, particularly in the TI, was also observed for TAS2R41 and 60. Small intestinal enteroendocrine cells contain appetite suppressing and insulin sensitizing hormones, such as CCK, GLP-1 and PYY [42], with the activation gastrointestinal of TAS2Rs implicated in the reduction of food intake and enhanced glucose control [3,38]. The delivery of a TAS2R41 or TAS2R60 specific agonist could potentially activate only small intestinal TAS2Rs, triggering the release of appetite suppressing peptide hormones without the unwanted activation of gastric or colonic enteroendocrine cells [43,44]. To date, TASR2R41 has only two known agonist, the antibiotic chloramphenicol and the artificial sweetener sucralose [45,46], neither of which are specific to TAS2R41 or likely to be the endogenous or environmental ligands, TAS2R60 is an currently an orphan receptor.
Gastrointestinal TAS2Rs exhibit inter-individual variation.
We introduce the concept of gut super tasters as evidenced by a more complete repertoire of TAS2Rs throughout their GI tract. This complement of TAS2Rs is present in all GI samples taken from these individuals and suggests a conserved regulatory mechanism that is unlikely to be directly affected by exposure to specific dietary/bacterial compound(s). This conclusion is further supported as one participant, who volunteered samples from both a gastroscopy and colonoscopy, exhibited an enhanced TAS2R repertoire along the entire length of the GI tract. Importantly, an expanded GI TAS2R repertoire may confer a broader chemosensory capacity to these individuals that may modulate their GI responses to specific compounds, be they dietary, bacterial or endogenous in nature. For example, TAS2R16 was detected only in the gut super tasters and is the sole receptor for the detection of natural compounds arbutin and amygdalin D [9], which are present in the leaf of bearberry plants and stone fruit kernels respectively. The presence of GI TAS2R16 may allow for a more pronounced gastrointestinal response to these plant toxins and hence may afford additional protection from accidental poisoning when consuming the associated food. Additionally Salicin, the willow bark derived anti-inflammatory agent [47] , is also a TAS2R16 specific agonist that may trigger GI side effects in some people [48]. It is possible that these reported side effects are, in part, mediated by the actions of GI TAS2R16 in these individuals, and that side effects for other drugs are in part dictated by the presence of specific TAS2Rs. We believe this is the first evidence suggesting the existence of gut super tasters and has ramifications for interventions targeting GI TAS2Rs, and for the understanding of gastrointestinal side effect to pharmaceuticals and bitter bioactives. Unlike the oral super tasters, “gut super tasters” are not defined based on their specific genetic polymorphism, but on a general increase in the spectrum of TAS2Rs present with the gut. It is possible that along with a general increase in the spectrum of TAS2Rs present, there may be genetic polyphorphisms and/or changes in the absolute number of enteroendocrine cells. Indeed, recent studies report changes in both enteroendocrine cell number in response to diet and highlight the role of haplotype variation in oral bitter taste perception [49,50].
Expression levels vs chromosome location and gene relatedness
Human TAS2Rs are located on 3 cytogentic locations, 5p15, 7q31-35 and 12p13 that contain a single gene, a cluster of 9 genes and a cluster of 15 genes respectively. As previously reported for rodent TAS2Rs [51], TAS2Rs from the same chromosomal location exhibited similar patterns of transcription along the human GI tract suggesting conserved cis-regulatory elements between closely located genes. Although relative GI patterns of transcription were conserved within a given chromosomal location, absolute transcription number was not. It is possible that there is copy number variation, deletions of regulatory factors that affects the gene transcription, or the presence of gene choice elements such as those that effectively turn on or off transcription of adjacent odorant receptor genes [52]. TAS2R41 and 60 both reside on chromosomal region 7q35, show close phylogenetic relationship and are the only TAS2R genes that have significantly elevated small intestine transcript count. Again, this suggests that in addition to genetic and locational similarities, they may also have conserved regulatory elements and their transcription maybe regulated together.
Strengths and limitations
There are several limitations to the current work, including the characterization of transcript rather than protein expression, the inability to localize TAS2R expression within a specific cell type, and the assessment of transcript without additional haplotype analysis. These limitations were considered during the study design and reflect decisions made to increase participant recruitment rates by not assessing genetic information, and to ensure optimal detection of lowly expressed TAS2Rs by extracting RNA from the entire biopsy sample. Further to this, the characterization of TAS2R proteins was considered impractical due to the current lack of effective TAS2R antibodies [53]. Strengths of this study include the relatively large number of participants, the strict criteria for participant inclusion and the choice of the highly sensitive NanoString detection technology that allows for detailed resolution of lowly transcribed genes without pooling clinical samples [54].
4. Materials and Methods
Figure 4.
Schematic overview of methods: Participants undergoing routine Gastroscopy and/or Colonoscopy were recruited and volunteered 10 biopsies per procedure taken in biopsy pairs. Dual biopsies were processed together with total RNA extracted. TAS2R analysis of total RNA samples was conducted using Nanosting™ direct RNA counting and analyzed for gastrointestinal expression, regional variation, and inter-individual variation.
Figure 4.
Schematic overview of methods: Participants undergoing routine Gastroscopy and/or Colonoscopy were recruited and volunteered 10 biopsies per procedure taken in biopsy pairs. Dual biopsies were processed together with total RNA extracted. TAS2R analysis of total RNA samples was conducted using Nanosting™ direct RNA counting and analyzed for gastrointestinal expression, regional variation, and inter-individual variation.
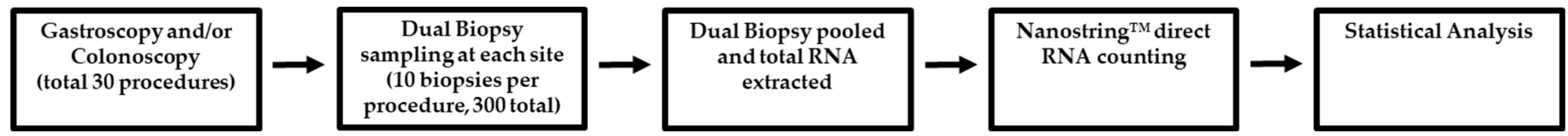
4.1. Ethics and Recruitment
Ethical approval was obtained from the New Zealand Health and Disability Ethics (HEDC) committee prior to the commencement of the study (NTY/11/08/077). This research was conducted in alignment with the Declaration of Helsinki and all participants gave informed consent before taking part in this study, Participants were recruited from routine endoscopy lists at regional hospitals of the Waitemata District Health Board (Auckland, New Zealand). Participants were approached prior to undergoing either a colonoscopy or a gastroscopy, a medical history was taken, suitability for the trial was determined and informed consent was obtained. Inclusion criteria for this study was: male or female, 18-65 years of age, normal gross gastrointestinal tract anatomy, generally healthy as ascertained by self-report, undergoing screening gastroscopy or colonoscopy, and provision of written informed consent. Exclusion criteria for this study was: obese as defined by BMI >30kg/m2, previous GI tract resection (including gastric banding), diagnosed diabetes mellitus (type 1 or 2), diagnosed medical conditions known to affect gut function including but not limited to malignant or inflammatory diseases of the gut such as: inflammatory bowel disease (Crohns, colitis), ischemic bowel, or coeliac disease. Current or recent use of the following medications: non-steroidal anti-inflammatory drugs (NSAIDs), proton-pump inhibitors, glucocorticoids, immunomodulators, anticoagulants, weight loss medication, and alcohol or drug dependency. Extreme food restriction and/or excessive physical activity as seen in anorexia nervosa, bulimia or similar eating disorders that alter body composition in a short space of time, and/or purging (vomiting/using laxatives). Unwilling/unable to comply with study protocol. Participating in another clinical intervention trial.
4.2. Biopsy Tissue Sampling
Table 3.
Location of Biopsy Sampling.
| Sample Number | Procedure | Sample Name | Location Notes |
| 1 | Gastroscopy | Gastric Fundus |
|
| 2 | Gastroscopy | Gastric Body |
Approximately midpoint along the greater curvature |
| 3 | Gastroscopy | Gastric Antrum |
|
| 4 | Gastroscopy | D2 Duodenum |
|
| 5 | Gastroscopy | Proximal Jejunum |
Or as close to the Duodenojejunal Flexure as possible |
| 6 | Colonoscopy | Terminal Ileum |
Proximal to the ileocecal sphincter |
| 7 | Colonoscopy | Ascending Colon |
|
| 8 | Colonoscopy | Transverse Colon |
|
| 9 | Colonoscopy | Sigmoid Colon |
|
| 10 | Colonoscopy | Rectum |
Dual 2-3 mm intestinal biopsy samples were taken from gastric fundus, gastric body, gastric antrum, duodenum (2nd part (D2) and either the proximal jejunum or 4th part (D4) of the duodenum (referred to as proximal jejunum sample), for participants undergoing gastroscopy, and rectum, sigmoidal colon, transverse colon, ascending colon and terminal ileum for participants undergoing colonoscopy (Supplementary Figure S1). Samples were rinsed in RNase free water (Life Technologies, Carlsbad, CA, USA, cat # 10977-023), and immediately immersed in RNAlater® (Life Technologies, cat # AM7021). Samples were kept at 4oC for 24h then frozen at -80oC until analysis, as described in the manufactures protocol.
4.3. NanoString Bitter Taste Receptor Analysis
Dual biopsy samples were processed together using a TissueLyser II (Qiagen, Limburg, Netherland, cat #: 85300), total RNA was extracted using QIAzol Lysis Reagent (Qiagen, Cat #: 79306) and RNA extraction kit (Qiagen, cat #: 217004) as described in the manufactures’ protocol. Samples were analysed for mRNA transcription of human TAS2Rs by NanoString® (NanoString® Technologies, Inc. Seattle, WA). RNA Probes were designed by NanoString® Technologies Inc (Supplementary Figure S2). NanoString® experiments were conducted by New Zealand Genomics Ltd (Dunedin, New Zealand) and analysed at The New Zealand Institute of Plant and Food Research Ltd (Auckland, New Zealand) as described in the manufacture’s protocol (see Supplementary Table S1 for probe design). Sequence similarity of 5 TAS2Rs (Ta2R43, 44, 45, 46 and 47) within the probe target areas made the design of specific probes impractical and 1 probe (TAS2R43) detected all of the members of this super family with greater than 93% efficiency. Briefly, RNA was quantified using a NanoDrop 2000 UV-Vis spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA). 600ng per sample was added to the Nanostring® probe hybridisation reaction and incubated for 18h at 64oC. Hybridized probes were then purified using the nCounter® Prep Station (NanoString Technologies, Inc.). Samples were washed to remove excess capture and reporter probes then immobilized on a streptavidin-coated cartridge to remove transcript-specific ternary complexes. Data collection was carried out on the nCounter® Digital Analyser (NanoString® Technologies, Inc.) counting individual fluorescent barcodes to quantify target RNA molecules present in each sample. Analysis was conducted as per the manufactures’ recommendations. Briefly, raw Count data from the nCounter was normalized relative to a standard curve constructed using spike in exogenous control samples, then subsequently to a standard curve generated using the geometric mean of 6 reference genes B2M (NM_004048.2), G6PD (NM_000402.2) GAPDH (NM_002046.3), YWHAZ (NM_003406.2), TUBA1A (NM_006009.2) POLR2A (NM_000937.2). The background hybridization signal was determined using the spike in negative controls. All non-background RNAs underwent a background correction by subtraction of the mean counts of the negative controls +2 standard deviations. Final RNA counts that were below 50 were classified as being minimally detectable.
4.4. Statistical Analysis
For statistical analysis, normalized mRNA count data (as described above) were log transformed. Analysis of variance testing (ANOVA) was performed to determine if significant differences in RNA count existed between gut sites and between participants. When significant differences did occur, post-hoc analysis was conducted using Tukeys HSD to determine where these differences occurred.
For the investigation into the clustering of participants the gut sites were split into two groups, the upper Gastroscopy sites and lower Colonoscopy gut sites. Similar to all other analyses the response variable (gene expression) underwent log transformation. This analysis excluded participant 6 due sample BS31’s failure at QA.
Multiple methods for cluster analysis were conducted to determine presence and number of clusters for TAS2R RNA count. For cluster analysis the following procedures were employed: 1. ANOVA analysis to test whether there is any evidence of difference between participants. 2. Hierarchical clustering dendrograms using ward.d2 clustering criteria (24). 3. A plot of within group sum of squares by number of clusters by K-means partitioning. 4. NbClust (25) package within R, for both ward.d2 and k means with 26 criteria. For each method the function applies 26 criteria for determining the number of clusters, it then takes the best number of clusters as the one with the majority of the criteria suggesting that it is the best partition. 5. Genes identified from previous plot of expression vs participant, any with significant ANOVA p-values and a consensus of two clusters were further investigated to see how participants were grouped. 6. Several genes were found to have the same grouping of participants in their level of transcription. With the participants who are in both upper and lower gut-sites being present in the same groups for both regions. The selected genes were then combined, and a principal component analysis performed (PCA). From the PCA we produced a cluster-plot for the participants.
All the analyses other than those specifically noted were conducted in R: A language and environment for statistical computing. A P value of < 0.05 was designated as significant.
5. Conclusions
In this present study we characterize the GI profile of TAS2Rs in the human gut and determine the inter-regional and inter-individual variation of the TAS2Rs present. The data shows that although there is a generally conserved pattern of transcription both between regions and between individuals, specific TAS2Rs show regional variation and inter-individual variation. In the future this may allow for gastrointestinal therapies targeting specifically located TAS2Rs and for those targeted to individual TAS2R expression profiles.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, E.W. and J.I.; methodology, E.W. R.W. S.D.P and J.I.; formal analysis, K.R.; investigation, E.W R.W. and A.M; writing—original draft preparation, E.W.; writing—review and editing R.W. K.R. A.M. S.D.P. and J.I.
Funding
This research was funded by The New Zealand Ministry for Business and Innovation, grant number C11X1004.
Acknowledgments
The staff, nurses, and gastroenterologists from Waitakere hospital surgical department. Dr Kevin Sutton for obtaining funding for this research, and Dr Dominic Lomiwes and Dr Roger Harker for reviewing the manuscript.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Abbreviations
| TAS2R | Bitter Taste Receptor |
| LOD | Limit of Detection |
| LD | linear dichroism |
References
- Psichas, A.; Reimann, F.; Gribble, F.M. Gut chemosensing mechanisms. J. Clin. Investig. 2015, 125, 908–917. [CrossRef]
- Al Massadi, O.; Pardo, M.; Roca-Rivada, A.; Castelao, C.; Casanueva, F.F.; Camino, L.M.S. Macronutrients act directly on the stomach to regulate gastric ghrelin release. J. Endocrinol. Investig. 2010, 33, 599–602. [CrossRef]
- Janssen, S.; Laermans, J.; Verhulst, P.-J.; Thijs, T.; Tack, J.; Depoortere, I. Bitter taste receptors and α-gustducin regulate the secretion of ghrelin with functional effects on food intake and gastric emptying. Proc. Natl. Acad. Sci. 2011, 108, 2094–2099. [CrossRef]
- Feng, X.; Zhong, S.; Yang, J.; Wang, Y.; Liu, J. Effects on Glucagon-Like Peptide-1 Secretion by Distal Ileal Administration of Nutrients. Obes. Surg. 2013, 23, 1774–1782. [CrossRef]
- Díez-Sampedro, A.; Hirayama, B.A.; Osswald, C.; Gorboulev, V.; Baumgarten, K.; Volk, C.; Wright, E.M.; Koepsell, H. A glucose sensor hiding in a family of transporters. Proc. Natl. Acad. Sci. USA 2003, 100, 11753–11758. [CrossRef]
- Jang, H.-J.; Kokrashvili, Z.; Theodorakis, M.J.; Carlson, O.D.; Kim, B.-J.; Zhou, J.; Kim, H.H.; Xu, X.; Chan, S.L.; Juhaszova, M.; et al. Gut-expressed gustducin and taste receptors regulate secretion of glucagon-like peptide-1. Proc. Natl. Acad. Sci. USA 2007, 104, 15069–15074. [CrossRef]
- Tolhurst, G.; Heffron, H.; Lam, Y.S.; Parker, H.E.; Habib, A.M.; Diakogiannaki, E.; Cameron, J.; Grosse, J.; Reimann, F.; Gribble, F.M. Short-Chain Fatty Acids Stimulate Glucagon-Like Peptide-1 Secretion via the G-Protein-Coupled Receptor FFAR2. Diabetes 2012, 61, 364–371. [CrossRef]
- Avau, B.; Depoortere, I. The bitter truth about bitter taste receptors: beyond sensing bitter in the oral cavity. Acta Physiol. 2015, 216, 407–420. [CrossRef]
- Meyerhof, W.; Batram, C.; Kuhn, C.; Brockhoff, A.; Chudoba, E.; Bufe, B.; Appendino, G.; Behrens, M. The Molecular Receptive Ranges of Human TAS2R Bitter Taste Receptors. Chem. Senses 2010, 35, 157–170. [CrossRef]
- Beauchamp, G.K.; Mennella, J.A. Flavor Perception in Human Infants: Development and Functional Significance. Digestion 2011, 83, 1–6. [CrossRef]
- Rozin, P.; A Vollmecke, T. Food Likes and Dislikes. Annu. Rev. Nutr. 1986, 6, 433–456. [CrossRef]
- Latorre, R.; Huynh, J.; Mazzoni, M.; Gupta, A.; Bonora, E.; Clavenzani, P.; Chang, L.; Mayer, E.A.; De Giorgio, R.; Sternini, C. Expression of the Bitter Taste Receptor, T2R38, in Enteroendocrine Cells of the Colonic Mucosa of Overweight/Obese vs. Lean Subjects. PLOS ONE 2016, 11, e0147468. [CrossRef]
- Gunawardene, A.R.; Corfe, B.M.; Staton, C.A. Classification and functions of enteroendocrine cells of the lower gastrointestinal tract. Int. J. Exp. Pathol. 2011, 92, 219–231. [CrossRef]
- Sternini, C., L. Anselmi, and E. Rozengurt, Enteroendocrine cells: a site of 'taste' in gastrointestinal chemosensing. Curr Opin Endocrinol Diabetes Obes, 2008. 15(1): p. 73-8.
- Moran, G.W., et al., Enteroendocrine cells: neglected players in gastrointestinal disorders? Therap Adv Gastroenterol, 2008. 1(1): p. 51-60.
- Posovszky, C.; Wabitsch, M. Regulation of Appetite, Satiation, and Body Weight by Enteroendocrine Cells. Part 1: Characteristics of Enteroendocrine Cells and Their Capability of Weight Regulation. Horm. Res. Paediatr. 2015, 83, 1–10. [CrossRef]
- Walker, E.; Lo, K.; Tham, S.; Pahl, M.; Lomiwes, D.; Cooney, J.; Wohlers, M.; Gopal, P. New Zealand Bitter Hops Extract Reduces Hunger During a 24 h Water Only Fast. Nutrients 2019, 11, 2754. [CrossRef]
- Walker, E.G.; Lo, K.R.; Pahl, M.C.; Shin, H.S.; Lang, C.; Wohlers, M.W.; Poppitt, S.D.; Sutton, K.H.; Ingram, J.R. An extract of hops (Humulus lupulus L.) modulates gut peptide hormone secretion and reduces energy intake in healthy-weight men: a randomized, crossover clinical trial. Am. J. Clin. Nutr. 2022, 115, 925–940. [CrossRef]
- Walker, E.; Lo, K.; Gopal, P. Gastrointestinal delivery of bitter hop extract reduces appetite and food cravings in healthy adult women undergoing acute fasting. Obes. Pillars 2024, 11, 100117. [CrossRef]
- Sakai, H.; Sato, K.; Kai, Y.; Chiba, Y.; Narita, M. Denatonium and 6-n-Propyl-2-thiouracil, Agonists of Bitter Taste Receptor, Inhibit Contraction of Various Types of Smooth Muscles in the Rat and Mouse. Biol. Pharm. Bull. 2016, 39, 33–41. [CrossRef]
- Clark, A.A.; Liggett, S.B.; Munger, S.D. Extraoral bitter taste receptors as mediators of off-target drug effects. FASEB J. 2012, 26, 4827–4831. [CrossRef]
- Jeon, T.-I.; Seo, Y.-K.; Osborne, T.F. Gut bitter taste receptor signalling induces ABCB1 through a mechanism involving CCK. Biochem. J. 2011, 438, 33–37. [CrossRef]
- Liszt, K.I.; Wang, Q.; Farhadipour, M.; Segers, A.; Thijs, T.; Nys, L.; Deleus, E.; Van der Schueren, B.; Gerner, C.; Neuditschko, B.; et al. Human intestinal bitter taste receptors regulate innate immune responses and metabolic regulators in obesity. J. Clin. Investig. 2022, 132. [CrossRef]
- Lei, H.; Yu, D.; Xue, Y.-B.; Li, Y.-H.; Gong, S.-M.; Peng, Y.-Y.; Liu, K.-F.; Buratto, D.; Yang, Y.; Zhang, S.-S.; et al. Tuft cells utilize taste signaling molecules to respond to the pathobiont microbe Ruminococcus gnavus in the proximal colon. Front. Immunol. 2023, 14, 1259521. [CrossRef]
- Schneider, C.; O’leary, C.E.; Locksley, R.M. Regulation of immune responses by tuft cells. Nat. Rev. Immunol. 2019, 19, 584–593. [CrossRef]
- Gustafsson, J.K.; Johansson, M.E.V. The role of goblet cells and mucus in intestinal homeostasis. Nat. Rev. Gastroenterol. Hepatol. 2022, 19, 785–803. [CrossRef]
- Gassler, N. Paneth cells in intestinal physiology and pathophysiology. World J. Gastrointest. Pathophysiol. 2017, 8, 150–160. [CrossRef]
- Wu, S.V.; Rozengurt, N.; Yang, M.; Young, S.H.; Sinnett-Smith, J.; Rozengurt, E. Expression of bitter taste receptors of the T2R family in the gastrointestinal tract and enteroendocrine STC-1 cells. Proc. Natl. Acad. Sci. USA 2002, 99, 2392–2397. [CrossRef]
- Rozengurt, N.; Wu, S.V.; Chen, M.C.; Huang, C.; Sternini, C.; Rozengurt, E. Colocalization of the α-subunit of gustducin with PYY and GLP-1 in L cells of human colon. Am. J. Physiol. Liver Physiol. 2006, 291, G792–G802. [CrossRef]
- Kaji, I.; Karaki, S.-I.; Fukami, Y.; Terasaki, M.; Kuwahara, A. Secretory effects of a luminal bitter tastant and expressions of bitter taste receptors, T2Rs, in the human and rat large intestine. Am. J. Physiol. Liver Physiol. 2009, 296, G971–G981. [CrossRef]
- Le Nevé, B.; Foltz, M.; Daniel, H.; Gouka, R. The steroid glycoside H.g.-12 from Hoodia gordonii activates the human bitter receptor TAS2R14 and induces CCK release from HuTu-80 cells. Am. J. Physiol. Liver Physiol. 2010, 299, G1368–G1375. [CrossRef]
- Descamps-Solà, M.; Vilalta, A.; Jalsevac, F.; Blay, M.T.; Rodríguez-Gallego, E.; Pinent, M.; Beltrán-Debón, R.; Terra, X.; Ardévol, A. Bitter taste receptors along the gastrointestinal tract: comparison between humans and rodents. Front. Nutr. 2023, 10, 1215889. [CrossRef]
- Nolden, A.A.; McGeary, J.E.; Hayes, J.E. Differential bitterness in capsaicin, piperine, and ethanol associates with polymorphisms in multiple bitter taste receptor genes. Physiol. Behav. 2016, 156, 117–127. [CrossRef]
- Kauer, J., et al., Adult picky eating. Phenomenology, taste sensitivity, and psychological correlates. Appetite, 2015. 90: p. 219-28.
- Green, E.; Jacobson, A.; Haase, L.; Murphy, C. Neural correlates of taste and pleasantness evaluation in the metabolic syndrome. Brain Res. 2015, 1620, 57–71. [CrossRef]
- A Mennella, J. Ontogeny of taste preferences: basic biology and implications for health. Am. J. Clin. Nutr. 2014, 99, 704S–711S. [CrossRef]
- Dotson, C.D.; Zhang, L.; Xu, H.; Shin, Y.-K.; Vigues, S.; Ott, S.H.; Elson, A.E.T.; Choi, H.J.; Shaw, H.; Egan, J.M.; et al. Bitter Taste Receptors Influence Glucose Homeostasis. PLOS ONE 2008, 3, e3974. [CrossRef]
- Kok, B.P., et al., Intestinal bitter taste receptor activation alters hormone secretion and imparts metabolic benefits. Mol Metab, 2018. 16: p. 76-87.
- Liszt, K.I.; Ley, J.P.; Lieder, B.; Behrens, M.; Stöger, V.; Reiner, A.; Hochkogler, C.M.; Köck, E.; Marchiori, A.; Hans, J.; et al. Caffeine induces gastric acid secretion via bitter taste signaling in gastric parietal cells. Proc. Natl. Acad. Sci. USA 2017, 114, E6260–E6269. [CrossRef]
- Lee, R.J.; Xiong, G.; Kofonow, J.M.; Chen, B.; Lysenko, A.; Jiang, P.; Abraham, V.; Doghramji, L.; Adappa, N.D.; Palmer, J.N.; et al. T2R38 taste receptor polymorphisms underlie susceptibility to upper respiratory infection. J. Clin. Investig. 2012, 122, 4145–4159. [CrossRef]
- Verbeurgt, C.; Veithen, A.; Carlot, S.; Tarabichi, M.; Dumont, J.E.; Hassid, S.; Chatelain, P. The human bitter taste receptor T2R38 is broadly tuned for bacterial compounds. PLOS ONE 2017, 12, e0181302. [CrossRef]
- Egerod, K.L.; Engelstoft, M.S.; Grunddal, K.V.; Nøhr, M.K.; Secher, A.; Sakata, I.; Pedersen, J.; Windeløv, J.A.; Füchtbauer, E.-M.; Olsen, J.; et al. A Major Lineage of Enteroendocrine Cells Coexpress CCK, Secretin, GIP, GLP-1, PYY, and Neurotensin but Not Somatostatin. Endocrinology 2012, 153, 5782–5795. [CrossRef]
- Harada, Y.; Koseki, J.; Sekine, H.; Fujitsuka, N.; Kobayashi, H. Role of Bitter Taste Receptors in Regulating Gastric Accommodation in Guinea Pigs. J. Pharmacol. Exp. Ther. 2019, 369, 466–472. [CrossRef]
- Kuwahara, A. Involvement of the Gut Chemosensory System in the Regulation of Colonic Anion Secretion. BioMed Res. Int. 2015, 2015, 1–9. [CrossRef]
- Lossow, K.; Hübner, S.; Roudnitzky, N.; Slack, J.P.; Pollastro, F.; Behrens, M.; Meyerhof, W. Comprehensive Analysis of Mouse Bitter Taste Receptors Reveals Different Molecular Receptive Ranges for Orthologous Receptors in Mice and Humans. J. Biol. Chem. 2016, 291, 15358–15377. [CrossRef]
- Thalmann, S.; Behrens, M.; Meyerhof, W. Major haplotypes of the human bitter taste receptor TAS2R41 encode functional receptors for chloramphenicol. Biochem. Biophys. Res. Commun. 2013, 435, 267–273. [CrossRef]
- Fiebich, B.L.; Appel, K. Anti-inflammatory effects of willow bark extract. Clin. Pharmacol. Ther. 2003, 74, 96–96. [CrossRef]
- Saller, R., J. Melzer, and M. Felder, Pain Relief with a Proprietary Extract of Willow Bark in Rheumatology. An Open Trial. Swiss Journal of Integrative Medicine, 2008. 20(3): p. 156-162.
- Widmayer, P.; Goldschmid, H.; Henkel, H.; Kã¼Per, M.; Kã¶Nigsrainer, A.; Breer, H.; Küper, M. High fat feeding affects the number of GPR120 cells and enteroendocrine cells in the mouse stomach. Front. Physiol. 2015, 6, 53–53. [CrossRef]
- Roudnitzky, N.; Behrens, M.; Engel, A.; Kohl, S.; Thalmann, S.; Hübner, S.; Lossow, K.; Wooding, S.P.; Meyerhof, W. Receptor Polymorphism and Genomic Structure Interact to Shape Bitter Taste Perception. PLOS Genet. 2015, 11, e1005530. [CrossRef]
- Prandi, S.; Voigt, A.; Meyerhof, W.; Behrens, M. Expression profiling of Tas2r genes reveals a complex pattern along the mouse GI tract and the presence of Tas2r131 in a subset of intestinal Paneth cells. Cell. Mol. Life Sci. 2018, 75, 49–65. [CrossRef]
- Khan, M.; Vaes, E.; Mombaerts, P. Regulation of the Probability of Mouse Odorant Receptor Gene Choice. Cell 2011, 147, 907–921. [CrossRef]
- Behrens, M.; Born, S.; Redel, U.; Voigt, N.; Schuh, V.; Raguse, J.-D.; Meyerhof, W. Immunohistochemical Detection of TAS2R38 Protein in Human Taste Cells. PLOS ONE 2012, 7, e40304. [CrossRef]
- Veldman-Jones, M.H., et al., Evaluating Robustness and Sensitivity of the NanoString Technologies nCounter Platform to Enable Multiplexed Gene Expression Analysis of Clinical Samples. Cancer Res, 2015. 75(13): p. 2587-93.
Figure 1.
Inter-individual variation in specific TAS2R RNA transcript count. TAS2R RNA transcript counts of 298 gut biopsy (analysed as biopsy pairs) samples taken from 28 individuals were determined by Nanostring analysis. Four individuals (5, 6, 8 and 18) showed altered patterns of TAS2R transcription with notable presence of TAS2R16, 39, 42, 7, 8 and 9. Participant 5 volunteered samples from a gastroscopy, participants 6 and 18 that volunteered samples from colonoscopies with the Proximal Jejunum sample from participant 6 being excluded from analysis due to failure of QA processes, and participant 8 volunteered samples from both a colonoscopy and gastroscopy. Within these individuals, TAS2R16, 39, 42, 7, 8 and 9 were consistently detected across all biopsy samples.
Figure 1.
Inter-individual variation in specific TAS2R RNA transcript count. TAS2R RNA transcript counts of 298 gut biopsy (analysed as biopsy pairs) samples taken from 28 individuals were determined by Nanostring analysis. Four individuals (5, 6, 8 and 18) showed altered patterns of TAS2R transcription with notable presence of TAS2R16, 39, 42, 7, 8 and 9. Participant 5 volunteered samples from a gastroscopy, participants 6 and 18 that volunteered samples from colonoscopies with the Proximal Jejunum sample from participant 6 being excluded from analysis due to failure of QA processes, and participant 8 volunteered samples from both a colonoscopy and gastroscopy. Within these individuals, TAS2R16, 39, 42, 7, 8 and 9 were consistently detected across all biopsy samples.
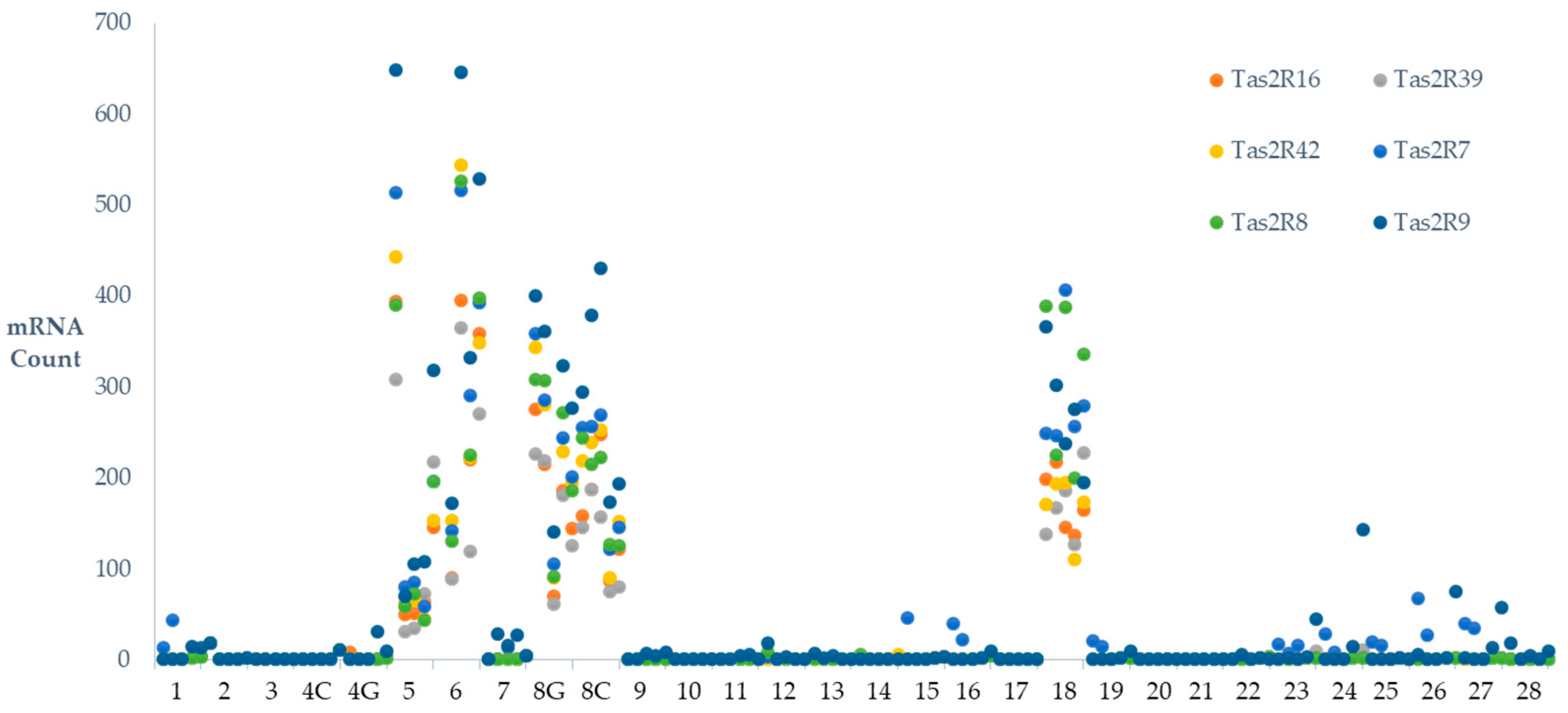
Figure 2.
Cluster analysis of Inter-individual variation in TAS2R RNA transcript count. For TAS2Rs where evidence of individual variation existed (ANOVA P<0.05, TAS2R7, 8, 9, 16, 39, 42) a cluster analysis of TAS2R transcription was conducted. Gut biopsies from 27 individuals (participant 6 was excluded from analysis due to an excluded data point for their proximal jejunum measure) were analyzed for clusters that explained point variability observed in the data. As most individuals only provided samples from one procedure, data was split into biopsies from individuals and by gastroscopy/colonoscopy. Individuals (designated by number) were found to cluster into 2 groups for both the gastroscopy and colonoscopy biopsy sites. When compared to the other participants sampled, individual 5 and 8 showed a different clustering for gastroscopy biopsy sites, and individual 8 and 18 for the colonoscopy biopsy sites. These clusters explained 94.21 and 97.27% of variations for gastroscopy and colonoscopy sites respectively.
Figure 2.
Cluster analysis of Inter-individual variation in TAS2R RNA transcript count. For TAS2Rs where evidence of individual variation existed (ANOVA P<0.05, TAS2R7, 8, 9, 16, 39, 42) a cluster analysis of TAS2R transcription was conducted. Gut biopsies from 27 individuals (participant 6 was excluded from analysis due to an excluded data point for their proximal jejunum measure) were analyzed for clusters that explained point variability observed in the data. As most individuals only provided samples from one procedure, data was split into biopsies from individuals and by gastroscopy/colonoscopy. Individuals (designated by number) were found to cluster into 2 groups for both the gastroscopy and colonoscopy biopsy sites. When compared to the other participants sampled, individual 5 and 8 showed a different clustering for gastroscopy biopsy sites, and individual 8 and 18 for the colonoscopy biopsy sites. These clusters explained 94.21 and 97.27% of variations for gastroscopy and colonoscopy sites respectively.
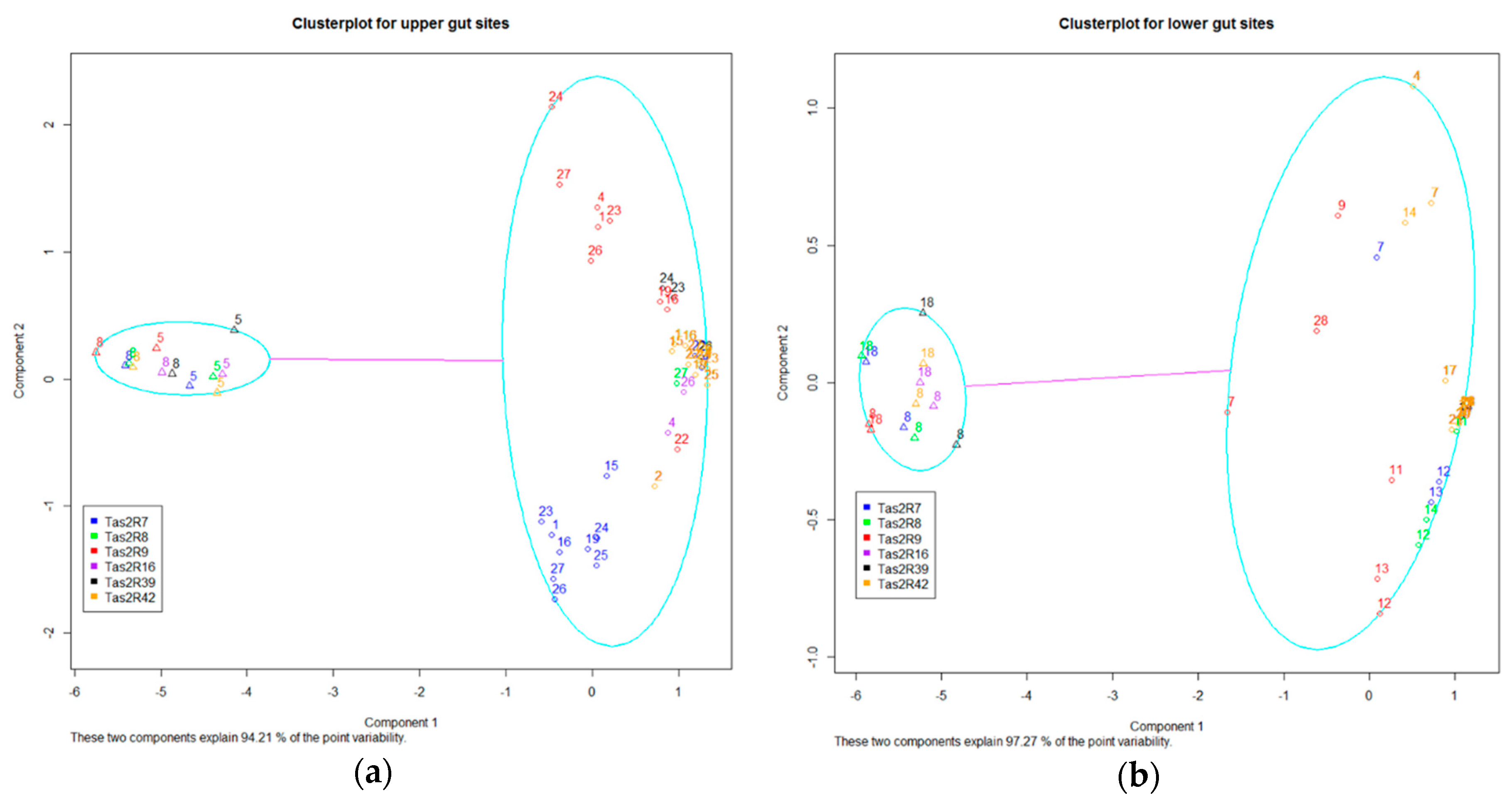
Figure 3.
Bitter Taste receptor (TAS2R) transcription cytogenetic location: TAS2R transcripts from 10 gut regions were compared to cytogenetic location. (a) TAS2Rs located on 12p13 (b) TAS2Rs located on 7q34 (c) TAS2Rs located on 7q35 (d) Consensus transcription ratio profile of all TAS2Rs found on 12p13, 7q34 and 7q35. Transcript profiles are conserved throughout the gut by cytogenetic location, but not for absolute transcript count. TAS2Rs on 7q35 have a notably different profile of expression to those of either 12p13 or 7q34.
Figure 3.
Bitter Taste receptor (TAS2R) transcription cytogenetic location: TAS2R transcripts from 10 gut regions were compared to cytogenetic location. (a) TAS2Rs located on 12p13 (b) TAS2Rs located on 7q34 (c) TAS2Rs located on 7q35 (d) Consensus transcription ratio profile of all TAS2Rs found on 12p13, 7q34 and 7q35. Transcript profiles are conserved throughout the gut by cytogenetic location, but not for absolute transcript count. TAS2Rs on 7q35 have a notably different profile of expression to those of either 12p13 or 7q34.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



