
Submitted:
24 October 2024
Posted:
26 October 2024
You are already at the latest version
Abstract
Background: Human β-defensin 2 (HBD2) is a protein important in immune system activation, produced by epithelial cells, mainly as a result of contact with microorganisms or the action of various pro-inflammatory cytokines. According to previous research, it has proven to be important in chronic urticaria, leading us to research on the HBD2 in isolated chronic angioedema. Methods: This cross-sectional study involved 102 participants (three groups): 33 patients with isolated chronic non-hereditary angioedema (AE); 33 patients with angioedema associated with chronic urticaria (CU+AE); and 35 healthy controls (CTRL). They provided saliva and serum samples to determine HBD2 levels using the Enzyme-Linked Immunosorbent Assay-ELISA. Results: Salivary HBD2 levels were significantly higher in those with CU+AE than in the CTRL (p=0.019). While salivary HBD2 values differed between those with angioedema and CTRL, the serum HBD2 values did not. Also, no correlation between the levels of HBD2 in saliva and serum was found. Conclusions: Since we found that salivary HBD2 values were significantly higher in those with CU+AE than in CTRL, this points to a possible role of the HBD2 molecule in pathogenesis of AE. Therefore, more research is needed to determine how reliabile salivary HBD2 measurement is and its significance.
Keywords:
human β-defensin 2
; angioedema
; chronic urticaria
; biomarkers
; saliva
; inflammatory factors
; mast cells
; immune response
1. Introduction
Human β-defensin 2 (HBD2) is a protein that plays an important role in activating the immune system, acting by modulating spinal pathways and the inflammatory response, thereby providing protection to the organism againstvarious pathogens. Structurally, HBD2 is a cysteine-rich cationic peptide of low molecular weight, consisting of 41 amino acids, also known as skin antimicrobial peptide 1 (SAP 1). It is produced by epithelial cells, keratinocytes, and macrophages, mainly as a result of contact with microorganisms (bacteria, viruses, fungi) or the action of various pro-inflammatory cytokines. Thus, HBD2 is secreted by the epithelial cells of the gingival mucosa, as confirmed by the expression of mRNA for HBD2 in these cells and the presence of its copies in saliva. Biopsies of the gingiva have shown that HBD2 is also secreted at very low levels in the gingival tissue of healthy individuals without signs of inflammation. Oral keratinocytes produce and secrete HBD2 primarily after stimulation by pro-inflammatory cytokines or as a result of exposure to bacterial endotoxins. Thus, just two to four hours after contact with pro-inflammatory cytokines, the synthesis of this protein significantly increases. The main function of HBD2 is antimicrobial; the peptide kills bacteria and fungi. In vitro, HBD2 promotes inflammation by recruiting CD4+ T cells and macrophages through interactions with the chemokine receptor C-C chemokine receptor 2 (CCR2) and chemokine receptor C-C 6 (CCR6) [1]. It has also been established that HBD2 induces degranulation of mast cells through interaction with the G-protein-coupled receptor MRGPRX2 [2]. Importantly, HBD2 acts as a pro-inflammatory pruritogen. In patients with skin diseases, it interacts with Toll-like receptor 4 via MRGPRX2 activation, which in turn induces histamine-independent itching [3,4]. Additionally, it has been observed that β-defensins increase vascular permeability in vivo [4].
In patients with skin diseases, HBD2 and HBD3 are primarily secreted by keratinocytes in the skin. Also, serum levels of HBD2 are significantly elevated in patients with skin diseases, including atopic dermatitis and psoriasis [5,6]. In one study, significantly elevated levels of HBD2 were measured in the serum of patients with chronic spontaneous urticaria (CSU) compared to a healthy control group [7]. Chronic spontaneous urticaria is a common inflammatory skin disease characterized by pruritic wheals and/or angioedema that persist for more than 6 weeks and are triggered by some physical or environmental stimulus (cold, heat, exercise, pressure, sunlight, vibration, water, etc. [8,9]. Additionally, urticaria and angioedema are comparable skin reactions. It is typified by a transient, localised enlargement of mucosal membranes, subcutaneous tissue, and the deep dermal layer. Although it can affect any portion of the body, the periocular area, lips, oral mucosa, vaginal areas, other mucosal regions, and other localisations are where it most frequently appears. In extreme situations, the lining of the intestines and upper respiratory tract may also be impacted. Angioedema can occur on its own, in conjunction with urticaria, or as a characteristic of anaphylaxis in disorders mediated by mast cells or bradykinin. It can also arise in conditions whose mechanisms are unknown, such as infections, uncommon diseases, or idiopathic angioedema. According to one research with HBD2 analysis by the Dunnett T3 test, CSU patients with associated angioedema had higher serum HBD2 levels than those without angioedema and the healthy control group [7]. However, there was no difference in serum HBD2 levels between CSU patients without angioedema and the healthy group. Serum HBD2 levels in CSU patients negatively correlated with the percentages of peripheral basophils. Furthermore, no significant correlation was found between HBD2 levels and disease severity (UAS7). Multiple logistic regression showed that higher HBD2 levels (> 72 pg/mL) and higher UAS (≧ 28) were significantly associated with angioedema in CSU patients, while age, sex, and vitamin deficiency affected the occurrence of angioedema.
In clinical work with chronic urticaria and non-hereditary angioedema (Figure 1), it is very important to look at the relevant disease biomarkers. According to the latest guidelines for CSU and non-hereditary angioedema treatment and diagnostic procedures, blood work should look for differential blood count(DBC), erythrocyte sedimentation rate (ESR) and/or C-reactive protein (CRP), IgG anti-TPO and total IgE values [5,6,7,8].
Mast cell activation is a key factor in the pathogenesis of CSU and chronic angioedema. This activation is well-known to involve regulators of FcεRI, IgE- independent pathways ⦏e.g., G-protein-coupled receptor X2 (MRGPRX2)⦐, tetraspanins, and the CD300 protein family [10]. In CSU patients, thyroid proteins, nuclear antigens (double-stranded DNA), and IL24 induce mast cell activation through cross-linking of FcεRI [11]. In both autoimmune subtypes of CSU (type I and IIb), there is a connection with the FcεRI receptor; however, a significant increase in serum substance P levels, which triggers mast cell activation via MRGPRX2, is also observed in CSU patients [12]. This suggests a potential contribution of the IgE-FcεRI-independent mechanism to the pathogenesis of CSU. Viral and parasitic infections are important triggers for CSU [8]. HBD molecules not only combat infections (exhibiting antimicrobial properties) but also act as mast cell secretagogues, eliciting clinical manifestations (neurogenic inflammation, pain, and itching) through a non-FcεRI cross-linking mechanism [2,4,13]. Specifically regarding CSU, Cao et al. discovered that serum levels of HBD2 were significantly elevated in CSU patients, with a significant correlation between HBD2 levels and the percentage of peripheral basophils, serum levels of translational tumor control protein (TCPA), and vitamin D levels, compared to those in a healthy control group. TCPA is a well-studied active factor in histamine release in patients with allergic diseases, including asthma and CSU [14]. In the skin of CSU patients, an oxidative tissue environment rich in cytokines (caused by the activation of various inflammatory cells in CSU) and autoimmunity (reflected by the presence of IgG antibodies to FcεRα) can lead to a conformational change in TCPA to an active dimeric form [15]. Notably, levels of the dimerized form of TCPA are increased and cause degranulation of mast cells and activation of basophils (with or without IgE sensitization) [15]. Peripheral basopenia in CSU patients indicates the activation of skin basophils [16]. A correlation has been observed between HBD2 levels and reduced basophil counts and higher TCPA levels in CSU patients, suggesting that elevated HBD2 levels may be a potential marker of basophil and mast cell activation. Kanda and Watanabe proposed that there is no positive feedback loop between histamine and HBD2 levels in patients with inflammatory skin diseases (17). Histamine synergistically increased HBD2 production in human keratinocytes. Since HBD2 can stimulate mast cell histamine release and chemotaxis via TNF-α-activated neutrophils, a paracrine loop between HBD2 and histamine levels may enhance the interaction between keratinocytes and mast cells or other inflammatory cells in the skin of CSU patients [17,18].
Data suggests that skin inflammation caused by persistent infections (due to the activation of mast cells, basophils, macrophages, and/or T lymphocytes in the dermis of patients with CSU) can increase the production of HBD2 in CSU. According to the findings of Cao et al., higher UAS7 levels and HBD2 levels are associated with angioedema, although HBD2 levels are not significantly related to disease severity (UAS7) (7). It has also been shown that HBD2 may be involved in the pathogenesis of angioedema in CSU patients, although HBD2 does not significantly enhance itching and urticarial formation. However, the expression of HBD2 in the skin was not measured, indicating that further research is needed to clarify how HBD2 levels increase in these patients, especially those with angioedema. Comparing HBD2 protein levels and gene expression between CSU patients and a healthy control group is necessary, as well as investigating specific pathways (histaminergic, cholinergic, or others) associated with elevated HBD2 levels in CSU patients.
HBD2 leveles have been analyzed in serum and saliva. Insights regarding HBD2 levels in saliva are also important for some patients and disorders. Previous studies have shown elevated HBD2 levels in the saliva of patients with periodontal diseases compared to healthy individuals, making HBD2 a potential biomarker for detecting and preventing periodontal diseases [19,20,21]. Additionally, HBD2 may serve as a marker of inflammation with potential therapeutic effects (inflammation suppression, reduction of oxidative stress, infection fighting) [22]. Diseases for which HBD2 is believed to have therapeutic potential include viral diseases, atopic dermatitis, allergic diseases, atopic asthma, oral lichen planus, wound healing, cell damage caused by smoking, and premature birth [22]. Its primary antimicrobial function is crucial, as is its involvement in the chemotaxis of immune cells and the activation of Toll-like receptors (TLRs) on their surface, along with its strong binding ability to complement component C1, which induces mast cell degranulation through interaction with MRGPRX2 and increases vascular permeability in vivo.
Given that a recent study showed higher serum levels of HBD2 in patients with chronic CSU and associated angioedema compared to those with CSU without angioedema, this suggests a potential role for HBD2 in the pathophysiology of angioedema [7]. To date, no studies have examined the correlation between HBD2 levels in saliva and serum in patients with angioedema in relation to the severity of the clinical presentation, nor have they compared HBD2 levels in saliva and serum with coagulation factor levels. Other conditions where elevated salivary HBD2 levels have been observed include periodontal diseases, H. pylori infections, inflammatory bowel diseases, atopic dermatitis, psoriasis, and lichen sclerosus.
Overall, taking into account the numerous properties of this molecule, the current evidence suggests that HBD2 is a potential marker of inflammation, and also a molecule with a potential therapeutic effect (suppression of inflammation, antimicrobial activity, reduction of oxidative stress, etc.), which points to its potential significance for recurrent angioedema and CSU.
2. Materials and Methods
2.1. Participants
This cross-sectional study involved a total of 102 participants, involving three groups: 33 patients with isolated chronic non-hereditary angioedema (defined according to the guidelines of Zuberbier et al. from 2022), who met the inclusion and exclusion criteria; 33 patients with non-hereditary angioedema associated with chronic urticaria; and 35 healthy participants who met the exclusion criteria.
All patients were examined by an allergist/immunologist at the Outpatient Clinic for Allergology and Clinical Immunology at the Rockefeller Special Hospital for Lung Diseases. After an allergist's examination, they received a recommendation for treatment and were introduced to the research being conducted at the Rockefeller Special Hospital for Lung Diseases and were offered to participate in it. Along with verbal instructions about the methods and purpose of the research, they were reminded that participation was voluntary, anonymous, and that they could withdraw at any time. After verbal instructions, subjects were asked to read the voluntary consent and, if they agreed, sign it.
Also, all subjects were referred to a doctor of dental medicine for an examination to rule out oral disease (active periodontal disease), and those with a positive finding were included in the research.
Patients who agreed to participate and met the inclusion criteria were included in this study.
They provided a saliva sample together with the peripheral blood sample to determine HBD2 concentrations using the ELISA (Enzyme-Linked Immunosorbent Assay) method. Saliva was collected in two tubes within five minutes for each participant.
Subsequently, a peripheral blood sample (serum) was taken from the participants to determine HBD2 levels using the ELISA method, following the manufacturer's instructions, and the samples were stored at –70 °C.
The research was approved by Ethics Committee of University Hospital Center “Sestre Milosrdnice”, Zagreb, Croatia.
2.2. Saliva Sampling and Analysis for HBD2
At the initial examination, all participants were instructed to refrain from intense physical activity and psychological stress for three days prior to the saliva sampling. They were also asked to avoid eating, drinking, and brushing their teeth for 90 minutes before the sampling procedure. Total unstimulated saliva was collected from participants in the morning between 9 and 10 a.m. to minimize daily variations. A systematic review and meta-analysis [23] indicated that previous studies collecting both types of saliva (unstimulated and stimulated) showed similar results in levels of salivary biomarkers [23]. This study confirmed that unstimulated saliva is sufficient for detecting salivary biomarkers and may be relevant in future research [23].
During saliva sample collection, participants were instructed to comfortably position themselves with their heads slightly tilted forward. Immediately before saliva sampling, all participants rinsed their mouths with water to avoid contamination from other sources, then waited ten minutes before collecting the sample. They were instructed to swallow their saliva just before the collection began. Approximately 2.50 mL of saliva was collected from participants (angioedema group, angioedema with urticaria group, and control participants) in graduated tubes (Salivette) (1534.500, SARSTEDT AG & Co. KG, Nümbrecht, Germany) using the "spitting method." In this method, the participant held saliva in their mouth for 60 seconds and then spat it into the graduated tube. This process was repeated for another ten minutes. Participants did not use any materials to stimulate saliva secretion.
2.3. Serum Sampling and Analysis for HBD2
After collecting the total unstimulated saliva sample for the analysis of salivary HBD2 concentration, blood sample were taken from the participants to determine serum HBD2 concentration using the same ELISA method.
Saliva samples were delivered to the Department of Medical Laboratory Diagnostics at the "Dr. Fran Mihaljević" Clinic for Infectious Diseases, where the Salivette tubes were stored at –70 °C until analysis. To determine salivary HBD2 concentration, the frozen samples were first thawed at room temperature for 30 minutes. They were then centrifuged at 1500 x g for five minutes, and the supernatant was used for analysis. Blood samples were taken from participants in tubes without anticoagulant. After 30 minutes, these samples were centrifuged at 2500 x g for 10 minutes. Following centrifugation, the serum samples were aliquoted and frozen at –70 °C until analysis. The concentration of salivary and serum HBD2 was measured using the ELISA method with commercial reagent kit from MyBioSource (Inc., San Diego CA 92195-3308, USA). The manufacturer specifies a test sensitivity of 3.9 pg/mL and linearity from 7.8 to 500 pg/mL (samples with concentrations > 500 pg/mL were diluted and repeated in a new series). The declared coefficients of variation (CV, %) within a series were 4.5, 4.6, and 3.9 for concentrations of 93, 210, and 460 pg/mL, and between series were 5.3, 5.9, and 6.4 for concentrations of 89, 206, and 455 pg/mL. The method is based on a sandwich immunoassay principle. Antibodies specific to HBD2 are bound to the wells of a microtiter plate. Upon adding serum or saliva, HBD2 from the sample binds to the specific antibodies in the wells. After washing away the unbound fractions, a detection antibody specific to a different HBD2 epitope, conjugated with biotin, is added. After incubation, streptavidin conjugated to an enzyme (horseradish peroxidase, HRP) is added, which binds to biotin. After washing away unbound fractions, a substrate (3,3′,5,5′-Tetramethylbenzidine, TMB) is added. The intensity of the developed color is proportional to the concentration of HBD2. The ELISA method was performed using the VirClia ELISA/CLIA analyzer (Vircell, Granada, Spain).
The presence of blood in saliva (hemolysis) was tested by visual reading, and saliva samples containing blood were excluded from the study [24]. Hemolytic serum samples were also excluded from the study.
3. Results
The results of the salivary HBD2 measurement in patients with angioedema compared to the control group (i.e., across three groups) are presented in Figure 2. The analysis of the obtained HBD2 values indicates that the groups of participants statistically significantly differed in terms of salivary HBD2 levels, with a small effect size (p = 0.019; ε² = 0.079). When comparing the groups pairwise, salivary HBD2 levels were significantly higher in those with angioedema accompanied by urticaria (CU+AE) than in the control group (CTRL) (p = 0.019).
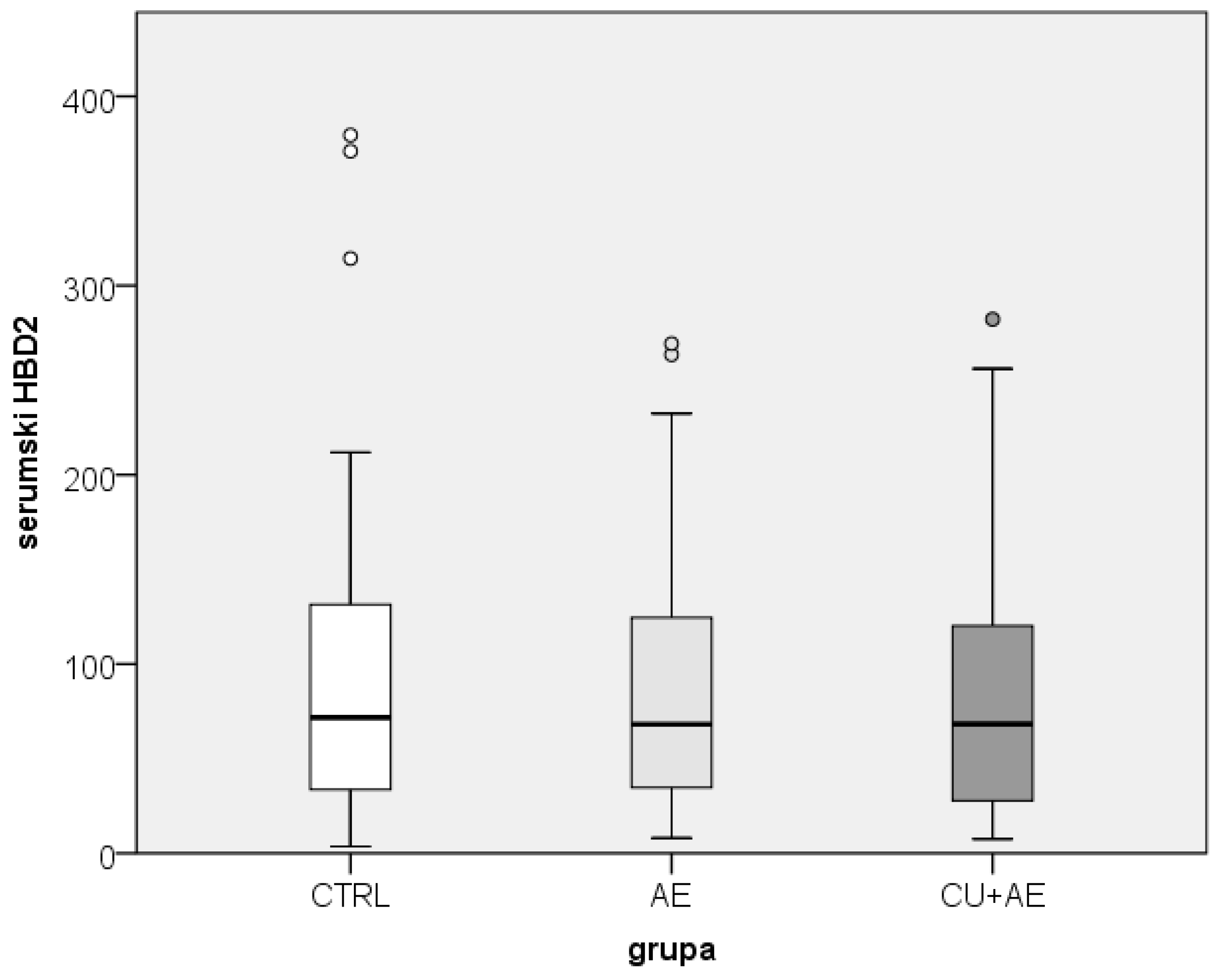
While the salivary HBD2 values differed among groups of patients with angioedema and healthy participants, the serum HBD2 values did not differ between them (Figure 3).
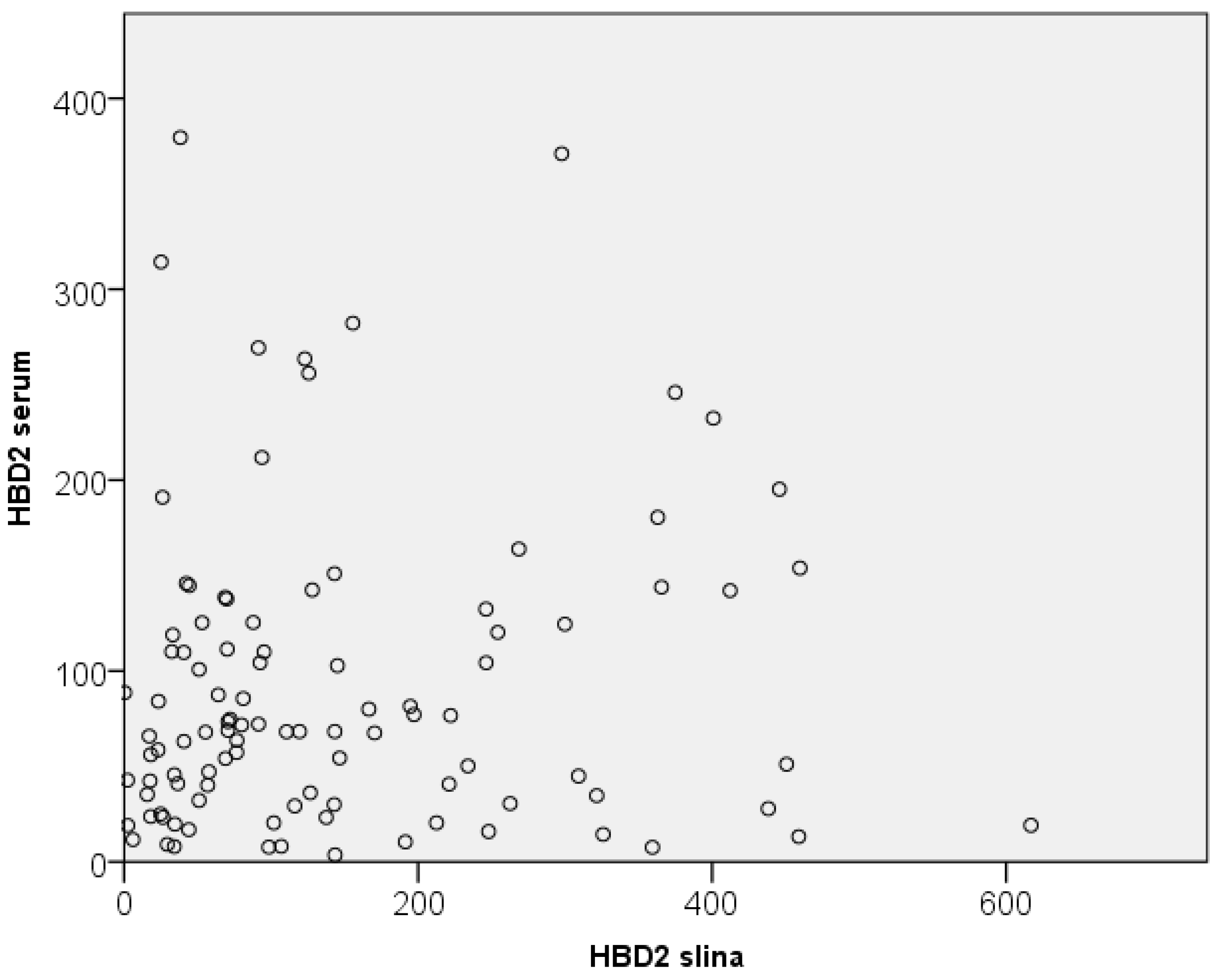
The HBD2 values in saliva were also compared with those in serum of the participants. A comparative overview of the range of serum and salivary HBD2 values is presented in Figure 4.
Figure 3.
Comparison of serum HBD2 between groups. HBD2-human beta defensin 2; CTRL- healthy controls; AE-angioedema; CU+AE-angioedema associated with urticaria.
Figure 3.
Comparison of serum HBD2 between groups. HBD2-human beta defensin 2; CTRL- healthy controls; AE-angioedema; CU+AE-angioedema associated with urticaria.
Additional statistical analysis (scatter plot analysis and Spearman correlation) showed no correlation between the levels of HBD2 in saliva and serum (they did not correlate linearly), indicating that salivary HBD2 is not a good indicator of serum HBD2 values (Figure 5). Additionally, no correlation was observed between serum and salivary HBD2 values and other serum biomarkers.
4. Discussion
Based on the numerous roles of HBD2 and the recently determined higher values of HBD2 in patients with CSU, in this study, we aimed to investigate its levels in patients with chronic angioedema. As is known, β-defensins are antimicrobial peptides that contribute to the resistance of epithelial (mucosal/skin) surfaces to microbial colonization [25]. They have been confirmed in all mammalian species and are predominantly produced by epithelial cells lining various organs such as the skin epidermis, bronchial mucosa, oral mucosa, and the genitourinary tract. In humans, β-defensins like HBD2 induce the activation and degranulation of mast cells, leading to the release of histamine, prostaglandin D2 and other mediators [26]. It is also essential to mention genetic factors, as β-defensins are encoded by genes that produce antimicrobial peptides found in white blood cells such as macrophages, granulocytes, and NK cells [27,28]. Defensins can enhance the innate immune system and strengthen the adaptive immune system through the chemotaxis of immune cells (T-cells, monocytes, dendritic cells, and mast cells) to the site of infection [29].
HBD2 is a cationic peptide of low molecular weight, rich in cysteine (composed of 41 amino acids), produced by epithelial cells, keratinocytes and macrophages, primarily upon contact with microorganisms (bacteria, viruses, fungi) or the action of various pro-inflammatory cytokines [30,31,32]. It is also crucial for numerous immune functions of the body. Regarding its role, HBD2 is essential in activating the immune system, modulating signaling pathways and the inflammatory response, and protecting the body from various microorganisms (primarily bacteria and fungi). In addition to its primary antimicrobial function, HBD2 may also be involved in the chemotaxis of immune cells and the activation of Toll-like receptors (TLR) on their surface. It also binds strongly to the C1 component of the complement system. Moreover, HBD2 promotes inflammation (in vitro) by recruiting CD4+ T cells and macrophages through interaction with chemokine receptors CCR2 and CCR6 [4,6]. HBD2 also induces mast cell degranulation via interaction with the G-protein coupled mast cell receptor X2 (Mas-related G-protein coupled receptor member X2, MRGPRX2) and increases vascular permeability (in vivo). Thus, interactions of HBD2 with the mast cell receptor MRGPRX2 induce mast cell degranulation [2]. MRGPRX2 is a protein predominantly found on skin mast cells, sensory neurons and keratinocytes, and its activation on mast cells leads to IgE-independent manifestations resembling type 1 hypersensitivity (pseudo-allergic reactions) [33].
The role of HBD2 has been demonstrated in several dermatological mucosal and allergic conditions. Regarding the production of defensins in the skin, human defensins HBD2 and HBD3 are primarily produced in keratinocytes but have also been detected in serum. In patients with skin diseases, significantly higher serum levels of HBD2 have been observed, including atopic dermatitis and psoriasis [5,6]. It has been noted that in patients with skin diseases, HBD2 acts as a pro-inflammatory pruritogen, with the interaction of HBD2 with Toll-like receptor 4 promoting the activation of MRGPRX2, which induces itch independent of histamine [3,4]. In addition to these diseases, HBD2 is also essential in CSU, as skin inflammations with urticaria may be triggered or exacerbated by persistent infections (due to the activation of mast cells, basophils, macrophages, and/or T cells), which can increase HBD2 production in the dermis. It is also essential to highlight that CSU is often accompanied by angioedema, which may be linked to HBD2, as higher serum levels of HBD2 have been demonstrated in patients with CSU and accompanying angioedema compared to those with CSU without angioedema [7].
Specifically regarding CSU, the pathogenesis in these patients involves significant mast cell activation through FcεRI receptor regulators and IgE-independent pathways (such as the MRGPRX2 mentioned above, tetraspanins, and the CD300 family of proteins) [8]. It is known that mast cell activation can be triggered by thyroid proteins, nuclear antigens (double-stranded DNA), and IL-24, which occurs via cross-linking of FcεRI that induces mast cell activation [9]. In autoimmune CSU, two autoimmune subtypes/endotypes (type I and IIb autoimmunity) have been observed, involving FcεRI receptor activity. However, an increase in serum levels of substance P has also been noted in patients with CSU, which triggers mast cell activation via MRGPRX2, suggesting the involvement of an IgE-FcεRI-independent mechanism in the pathogenesis [10]. Since infections can be possible causes and/or triggers for CSU, the role of HBD molecules is also possible in this process. In addition to their antimicrobial activities, HBD molecules act as mast cell secretagogenes, inducing neurogenic inflammation, pain, and itching (through a non-FcεRI cross-linking mechanism) [2,4,8,11]. In patients with CSU, research has revealed significantly elevated serum levels of HBD2, which correlate with the percentage of peripheral basophils, and with the level of a serum protein called translationally controlled tumor protein (TCTP) and vitamin D, compared to a healthy control group [7]. Additionally, in patients with allergic diseases (including asthma and CSU), the TCTP mentioned above is a crucial active factor in histamine release [14]. In the skin of patients with CSU, an oxidative environment rich in cytokines is present (due to the activation of various inflammatory skin cells), and autoimmune processes are underway (with IgG directed against the FcεRα receptor), which can induce conformational changes in TCTP into its active dimeric form [15]. Furthermore, increased levels of the dimerized form of TCTP have been observed in patients with CSU, leading to mast cell degranulation and basophil activation, independent of sensitization (with or without IgE sensitization). Typically, TCTP causes basophil histamine release dependent on IgE [15]. A recent study on patients with CSU measured significantly higher serum levels of HBD2 compared to healthy individuals [7]. Additionally, measuring HBD2 levels in CSU patients with associated angioedema (using Dunnett's T3 test) revealed higher levels compared to those without angioedema and healthy individuals, although there was no difference in HBD2 levels between CSU patients without angioedema and the healthy group. A negative correlation was observed between serum HBD2 levels and the percentage of peripheral basophils in patients with CSU, although no significant correlation was found between HBD2 levels and disease severity (UAS7). However, statistical analysis (multiple logistic regression) demonstrated that higher HBD2 levels (>72 pg/ml) and more severe disease (UAS ≥28) were significantly associated with angioedema in CSU patients (although no association was found between angioedema and age, gender, or vitamin deficiency) [7]. Since basophil counts are proven biomarkers of CSU severity (peripheral basopenia in these patients confirms skin basophil activation), it is essential to note the observed correlation between HBD2 levels and lower basophil counts as well as higher TCTP levels. Therefore, increased HBD2 levels may be a potential biomarker of basophil and mast cell activation [16,34]. When accompanying angioedema occurs with CSU, it is essential to address it, as it may be associated with HBD2. Most angioedemas in CSU patients are histaminergic, mediated by mast cells, and associated with itching [35]. Clinically, patients with CSU and angioedema exhibit more significant disease activity and longer disease duration compared to those without angioedema [36,37]. However, since CSU severity (measured by the UAS7 questionnaire) only assesses the presence of urticaria and itching without considering the presence of angioedema, it is essential to evaluate angioedema and its activity when assessing the condition and quality of life in CSU patients. Cao and colleagues found that greater CSU severity (higher UAS7) and HBD2 levels were associated with angioedema, while there was no significant correlation between HBD2 levels and CSU severity (UAS7) [7]. Thus, in CSU patients, HBD2 may be potentially involved in the pathogenesis of their associated angioedema (while HBD2 was not strongly linked to itching and urticaria formation). It is also reported that in patients with inflammatory skin diseases, there is no positive feedback loop between histamine and HBD2 levels [17]. According to research results, histamine synergistically increases HBD2 production in human keratinocytes when combined with TNF-α or IFN-γ expression. Since HBD2 can stimulate mast cells to release histamine and attract TNF-α-activated neutrophils, it is possible that in the skin of CSU patients, a paracrine loop between HBD2 and histamine levels enhances the interaction between keratinocytes and mast cells or other inflammatory cells [17,18]. Other diseases where elevated HBD2 levels have been observed, and potential therapeutic implications may arise include periodontal diseases, H. pylori infections, inflammatory bowel diseases, atopic dermatitis, psoriasis, and lichen sclerosis. Among the conditions where HBD2 may have a therapeutic effect are viral diseases, allergic conditions such as allergic asthma and atopic dermatitis, oral lichen planus, wound healing, cell damage from smoking, and preterm birth [22]. Additionally, oral production of HBD2 has been established, with epithelial cells of the gingival mucosa secreting it (confirmed by mRNA expression for HBD2 in these cells and its presence in saliva) [38,39]. Oral epithelial cells primarily produce and secrete HBD2 in response to pro-inflammatory cytokine stimulation or bacterial endotoxin activity after contact with pro-inflammatory cytokines (which happens rapidly, within just two to four hours), significantly increasing the synthesis of this protein [40]. Moreover, studies have shown that the level of HBD2 in the saliva of patients with periodontal disease is significantly higher than that of healthy people. This suggests that HBD2 can be used as a potential biomarker for detecting and preventing periodontal disease [19,20,21]. Due to insufficient data on HBD2 expression in the skin of patients with allergic mucocutaneous diseases such as CSU and chronic urticaria, further research is required to clarify the causes/aetiology of increased HBD2 levels in these patients. This is particularly important for CSU, especially in patients with angioedema. Future research should include a comparison of HBD2 levels and the expression of related genes between CSU patients and healthy control groups, as well as an investigation into the potential connection between the pathogenic pathways of angioedema/urticaria (whether histaminergic, cholinergic, or otherwise) and elevated HBD2 levels in CSU patients. Considering this molecule's numerous properties, current evidence suggests that HBD2 is a potential marker of inflammation and may also have therapeutic potential (anti-inflammatory, antimicrobial effects, reduction of oxidative stress, and more). This indicates its potential significance for recurrent angioedema and urticaria [22]. Our results for salivary HBD2 determination in patients with angioedema compared to the control group (i.e., across the three groups) and analysis of the obtained HBD2 levels show that the participant groups differed significantly in salivary HBD2 levels, although with a small effect size (p = 0.019). Comparing the groups in pairs revealed that salivary HBD2 was significantly higher in those with angioedema and urticaria compared to the control group (p = 0.019). Furthermore, salivary HBD2 values were positively correlated with age (p = 0.021), indicating an association between HBD2 levels and age. Several studies have explored the value of HBD2 in allergic skin diseases, though only one has investigated serum HBD2 levels in patients with CSU associated with angioedema. However, no studies have examined serum or salivary HBD2 in patients with isolated angioedema [7,41,42,43,44,45,46]. This indicates that our results are the first obtained from investigating HBD2 levels in patients with isolated angioedema. In a related study, Cao et al., in their 2021 study, demonstrated significantly elevated serum HBD2 levels in a group of CSU patients with associated angioedema compared to a healthy group and a group without angioedema [7]. Our results do not align with their findings, as they did not observe higher values. While our study found differences in salivary HBD2 levels between patients with angioedema and healthy participants, serum HBD2 levels did not differ. Importantly, we also compared salivary HBD2 levels with serum levels in the participants. Additional statistical analysis found no correlation between HBD2 levels in saliva and serum (no linear correlation), suggesting that salivary HBD2 may not be a good indicator of serum HBD2 levels in our participants. Furthermore, no correlation was observed between serum or salivary HBD2 levels and other serum biomarkers. Therefore, more research is needed to determine how reliable salivary HBD2 measurement is and its significance.
5. Conclusions
Since salivary HBD2 values were statistically significantly different between the groups and are much higher in those with chronic angioedema with urticaria than in controls, it indicates that HBD2 may be involved in the patogenesis of chronic angioedema. Salivary HBD2 is not a good indicator of serum HBD2 values. Therefore, more research is needed to determine how reliable salivary HBD2 measurement is and its significance.
Author Contributions
Author Contributions: Conceptualization, M.Š., L.L.-M. A.S.-M., and M.A.; methodology and visualization, M.Š., L.L.-M., A.S.-M., M.A.; M.H.; T.Š.; and A.T.K.; formal analysis, investigation, software and validation, M.Š., L.L.-M.; M.H.; T.Š.; and A.T.K.; resources, M.Š; data curation, M.Š., L.L.-M A.S.-M., A.G..; N.-B.P., and L.N.-B.; writing—original draft preparation and writing—review and editing, M.Š., L.L.-M., A.S.-M., M.A.; N.-B. P., and L.N.-B.; supervision and funding acquisition L.L.-M.; project administration, M.Š. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and approved Ethics Committee of University Hospital Center “Sestre Milosrdnice”, Zagreb, Croatia, in December 2022; number of protocol: 251-29-11-21-08, for studies involving humans.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study. Written informed consent has been obtained from the patient(s) to publish this paper.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation
References
- Jin, G.; Kawsar, H.I.; Hirsch, S.A.; Zeng, C.; Jia, X.; Feng, Z.; Ghosh, SK.; Zheng, Q.Y.; Zhou, A.; McIntyre, T.M.; Weinberg, A. An antimicrobial peptide regulates tumor-associated macrophage trafficking via the chemokine receptor CCR2, a model for tumorogenesis. PLos One. 2010, 5, e10993. [Google Scholar] [CrossRef]
- Subramanian, H.; Gupta, K.; Lee, D.; Bayir, A.K.; Ahn, H.; Ali, H. ß-defensins activate human mast cells via mas-related gene X2. J Immunol. 2013, 191, 345–52. [Google Scholar] [CrossRef] [PubMed]
- J., *!!! REPLACE !!!*; Luo, J.; Mack, MR.; Yang, P.; Zhang, F.; Wang, G.; Gong, X.; Cai, T.; Mei, Z.; Kim, BS.; Yin, S.; Hu, H. The antimicrobial peptide human ß-defensin 2 promotes itch trought Toll-like receptor 4 signaling in mice. J Allergy Clin Immunol 2017, 140, 885-e6. [Google Scholar] [CrossRef]
- Zhang, L.; McNeil, B.D. ß-defensins are proinflammatory pruritogens that activate Mrgprs. J Allergy Clin Immunol 2019, 143, 1960–1962-e5. [Google Scholar] [CrossRef] [PubMed]
- Kanda, N.; Watanabe, S. Increased serum human ß-defensin-2 levels in atopic dermatitis: relationship to IL-22 and oncostatin M. Immunobiology. 2012, 2017, 436–45. [Google Scholar] [CrossRef] [PubMed]
- Jin, T.; Sun, Z.; Chen, X.; Wang, Y.; Li, R.; Ji, S.; Zhao, Y. Serum human ß-defensin-2 is a possible biomarker for monitoring response to JAK inhibitor in psoriasis patient. Dermatology. 2017, 233, 164–169. [Google Scholar] [CrossRef]
- Tra Cao, T.B.; Cha, H.Y.; Yang, E.M.; Choi, B.Y.; Park, H.S.; Ye, Y.M. Serum human ß-defensin 2 is increased in angioedema accompanying chronic spontaneous urticaria. Int Arch Allergy Immunol. 2021, 182, 1066–1071. [Google Scholar] [CrossRef]
- Zuberbier, T.; Abdul Latiff, A.H.; Abuzakouk, M.; Aquilina, S.; Asero, R.; Baker, D.; Ballmer-Weber, B.; Bangert, C.; Ben-Shoshan, M.; Bernstein, J.A.; et al. The international EAACI/GA²LEN/EuroGuiDerm/APAAACI guideline for the definition, classification, diagnosis, and management of urticaria. Allergy. 2022, 77, 734–766. [Google Scholar] [CrossRef]
- Pozdarec, I.; Lugović-Mihić, L.; Artuković, M.; Stipuć-Marković, A.; Kuna, M.; Ferček, I. Chronic inducible urticaria: classification and prominent features of physical and non-physical types. Acta Dermatovenerol Alp Pannonica adria. 2020, 29, 141–148. [Google Scholar] [CrossRef]
- Bulfone-Paus, S.; Nilsson, G.; Draber, P.; Blank, U.; Levi-Schaffer, F. Positive and negative signals in mast cell activation. Trends Immunol. 2017, 38, 657–667. [Google Scholar] [CrossRef]
- Maurer, M.; Eyerich, K.; Eyerich, S.; Ferrer, M.; Gutermuth, J.; Hartmann, K.; Jakob, T.; Kapp, A. , Kolkhir, P.; Larens-Linnemann, D.; et al. Urticaria: Colegium internationale allergologicum (CIA) update 2020. Int Arch Allergy Immunol 2020, 181, 321–333. [Google Scholar] [CrossRef] [PubMed]
- Vena, G.A.; Cassano, N.; Di Leo, E.; Calogiuri, G.F.; Nettis, E. Focus on the role of substance P in chronic urticaria. Clin Mol Allergy. 2018, 16, 24. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, H.; Gopta, K.; Ali, H. Roles of Mas-related G protein-coupled receptor X2 on mast cell-mediated host defense, pseudoallergic drug reactions, and chronic inflammatory diseases. J Allergy Clin Immunol. 2016, 138, 700–710. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, Y.; Kasakura, K.; Kawakami, T. Histamine releasimg factor, a new therapeutic target in allergic diseases. Cells. 2019, 8, 1515. [Google Scholar] [CrossRef]
- Ulambayar, B.; Lee, H.; Yang, E.M.; Park, H.S.; Lee, K.; Ye, Y.M. Dimerized, not monomeric, translationally controlled tumor protein induces basophil activation and mast cell degranulation in chronic urticaria. Immune Netw. 2019, 19–e20. [Google Scholar] [CrossRef]
- Sánchez-Borges, M.; Caballero-Fonseca, F.; Capriles-Hulett, A.; González-Aveledo, L.; Maurer, M. Factors linked to disease severity and time to remission in patients with chronic spontaneus urticaria. J Eur Acad Dermatol Venereol. 2017, 31, 964–971. [Google Scholar] [CrossRef]
- Kanda, N.; Watanabe, S. Histamine enhances the production of human ß-defensin-2 in human keratinocytes. Am J Physiolαα Cell Physiol 2007, 293, C1916-23. [Google Scholar] [CrossRef]
- Niyonsaba, F.; Ogawa, H.; Nagaoka, I. Human ß-defensin-2 functions as a chemotactic agent for tumour necrosis factor-α-treated human neutrophils. Immunology. 2004, 111, 273–281. [Google Scholar] [CrossRef]
- Öztürk, A.; Kurt-Bayrakdar, S.; Avci, B. Comparison of gingival crevicular fluid and serum human beta-defensin-2 levels between periodontal health and disease. Oral Dis. 2021, 27, 993–1000. [Google Scholar] [CrossRef]
- Ribeiro, A.E.R.A.; Lourenco, A.G.; Motta, A.C.F.; Komesu, M.C. Influence of peridontal conditions on levels of human beta defensins 1 and 2 in saliva. J Microbiol Exp. 2018, 6, 00186. [Google Scholar] [CrossRef]
- Güncü, G.N.; Yilmaz, D.; Könönen, E.; Gürsoy, U.K. Salivary antimicrobial peptides in early detection of periodontitis. Front Cell Infect Microbiol. 2015, 5, 99. [Google Scholar] [CrossRef]
- Cieślik, M.; Bagińska, N.; Górski, A.; Jończyk-Matysiak, E. Human β-defensin 2 and its postulated role in modulation of the immune response. Cells. 2021, 10, 2991. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Agra, M.; González-Serrano, J.; de Pedro, M:, Virto; Caponio, V.C.A.; Prieto-Ibáñez, E.; Hernández, G.; Pintor-López, M.R. Salivary biomarkers in burning mouth syndrome: A systematic review and meta-analysis. Oral Dis 2023, 229, 2600–2613. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.H.; Kho, H.S. Blood contamination in salivary diagnostics: Current methods and their limitations. Clin Chem Lab Med. 2019, 57, 1115–1124. [Google Scholar] [CrossRef] [PubMed]
- Štrajtenberger, M.; Stipić-Marković, A.; Barac, E.; Artuković, M.; Lugović-Mihić, L. Human β -defensin-2 a connection between infections and allergic skin diseases. Acta Dermatovenerol Alp Pannonica Adriat. 2024, 33, 135–139. [Google Scholar] [CrossRef] [PubMed]
- White, S.H.; Wimley, W.C.; Selsted, M.E. Structure, function, and membrane integration of defensins. Curr Opin Struct Biol. 1995, 5, 521–7. [Google Scholar] [CrossRef]
- Ganz, T. Defensins: antimicrobial peptides of innate immunity. Nat Rev Immunol. 2003, 3, 710–20. [Google Scholar] [CrossRef]
- Hellgren, O.; Sheldon, B.C. Locus-specific protocol for nine different innate immune genes (antimicrobial peptides: β-defensins) across passerine bird species reveals within-species coding variation and a case of trans-species polymorphisms. Mol Ecol Resour. 2011, 11, 686–692. [Google Scholar] [CrossRef]
- van Dijk, A.; Veldhuizen, E.J.; Haagsman, H.P. Avian defensins. Vet Immunol Immunopathol. 2008, 124, 1–18. [Google Scholar] [CrossRef]
- Deptula, J.; Tokarz-Deptula, B.; Deptula, W. Defensins in humans and animals. Postepy hig. Med. Dosw. 2019, 73, 152–158. [Google Scholar] [CrossRef]
- Harder, J.; Bartels, J.; Christophers, E.; Schröder, J.M. A peptide antibiotic from human skin. Nature. 1997, 387, 861. [Google Scholar] [CrossRef]
- Schröder, J.M.; Harder, J. Human beta-defensin-2. Int J Biochem. Cell Biol. 1999, 31, 645–51. [Google Scholar] [CrossRef]
- Akin, C.; Elhosni, M.; Khokar, D.S. Mast cells and mast cell disorders. In: Rich RR, Fleicher TA, Schroeder HW, Weyand CM, Corry DB, Puck J (eds.). Clinical immunology: principles and practice. Elsevier. 2023, p, 563. [CrossRef]
- Kuna, M.; Štefanović, M.; Ladika Davidović, B.; Mandušić, N.; Birkić Belanović, I.; Lugović-Mihić, L. Chronic urticaria biomarkers IL-6, ESR and CRP in correlation with disease severity and patient quality of life-a pilot study. Biomedicines. 2023, 11, 2232. [Google Scholar] [CrossRef] [PubMed]
- Huston, D.P.; Sabato, V. Decoding the enigma of urticaria and angioedema. J Allergy Clin Immunol Pract. 2018, 6, 1171–1175. [Google Scholar] [CrossRef] [PubMed]
- Sussman, G.; Abuzakouk, M.; Bérard, F.; Canonica, W.; Oude Elberink, H.; Giménez-Arnau, A.; Grattan, C.; Hollis, K.; Hunteer, S.; Knulst, A.; et al. Angioedema in chronic spontaneous urticaria is underdiagnosed and has a substantial impact: Analyses from ASSURE-CSU. Allergy. 2018, 73, 1724–1734. [Google Scholar] [CrossRef]
- Puxeddu, I.; Petrelli, F.; Angelotti, F.; Croia, C.; Migliorini, P. Biomarkers in chronic spontaneous urticaria: current targets and clinical implications. J Asthma Allergy. 2019, 12, 285–295. [Google Scholar] [CrossRef]
- Mathews, M.; Jia, H.P.; Guthmiller, J.M.; Losh, G.; Graham, S.; Johnson, G.K.; Tack, B.K.; McCray, P.B.Jr. Production of beta-defensin antimicrobial peptides by the oral mucosa and salivary glands. Infect. Immun. 1999, 67, 2740–2745. [Google Scholar] [CrossRef]
- Dale, B.A.; Fredericks, L.P. Antimicrobial peptide sin the oral environment. Expression and function in health and disease. Curr. Issues Mol. Biol. 2005, 7, 119–133. [Google Scholar] [CrossRef]
- Krisanaprakornkit, S.; Kimball, J.R.; Weinberg, A.; Darveau, R.P.; Bainbridge, B.W.; Dale, B.A. Inducible expression of human beta-defensin 2 by Fusobacterium nucleatum ino ral epithelial cells: Multiple signaling pathways and role of commensal bacteria in innate immunity and the epithelial barrier. Infect. Immun. 2000, 68, 2907–2915. [Google Scholar] [CrossRef]
- Jansen, P.A; Rodijk-Olthuis, D.; Hollox, E.J.; Kamsteeg, M.; Tjabringa, G.S.; de Jongh, G.J.; van Vlijmen-Willems, I.M.; Bergboer, J.G.; van Rossum, M.M.; de Jong, E.M.; et al. Beta-defensin-2 protein is a serum biomarker for disease activity in psoriasis and reaches biologically relevant concentrations in lesional skin. PLoS One. 2009, 4–e4725. [Google Scholar] [CrossRef]
- Yu, L.; Li, L. Potential biomarkers of atopic dermatitis. Front Med (Lausanne). 2022, 9, 1028694. [Google Scholar] [CrossRef] [PubMed]
- Park, C.H.; Min, S.Y.; Yu, H.W.; Kim, K.; Kim, S.; Lee, H.J.; Kim, J.H. Effects of apigenin on RBL-2H3, RAW264.7, and HaCaT cells: anti-allergic, anti-inflammatory, and skin-protective activities. Int J Mol Sci 2020, 24, 4620. [Google Scholar] [CrossRef] [PubMed]
- Hata, T.R.; Gallo, R.L. Antimicrobial peptides, skin infections, and atopic dermatitis. Semin Cutan Med Surg. 2008, 27, 144–150. [Google Scholar] [CrossRef]
- Clausen, M.L.; Jungersted, J.M.; Andersen, P.S.; Slotved, H.C.; Krogfelt, K.A.; Agner, T. Human β-defensin-2 as a marker for disease severity and skin barrier properties in atopic dermatitis. Br J Dermatol. 2013, 169, 587–93. [Google Scholar] [CrossRef] [PubMed]
- Ong, P.Y.; Ohtake, T.; Brandt, C.; Strickland, I.; Boguniewicz, M.; Ganz, T.; Gallo, R.L.; Leung, D.Y. Endogenous antimicrobial peptides and skin infections in atopic dermatitis. N Engl J Med. 2002, 347, 1151–60. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Mu, Z.L.; Chen, X.; Wen, G.D.; Zhao, Y.; Zhang, J.Z. Atopic dermatitis-like graft-versus-host disease and lichen planus-like graft-versus-host disease: alterations in skin barrier function and related molecules. Chin Med J (Engl). 2017, 130, 1459–1466. [Google Scholar] [CrossRef]
- de Jongh, G.J.; Zeeuwen, P.L.; Kucharekova, M.; Pfundt, R.; van der Valk, P.G.; Blokx, W.; Dogan, A.; Hiemstra, P.S.; van de Kerkhof, P.C.; Schalkwijk, J. High expression levels of keratinocyte antimicrobial proteins in psoriasis compared with atopic dermatitis. J Invest Dermatol. 2005, 125, 1163–73. [Google Scholar] [CrossRef]
Figure 1.
Isolated chronic angioedema.
Figure 2.
Comparison of salivary HBD2 between groups. (Circles indicate outlier values. Groups connected by a horizontal line are statistically significantly different.) HBD2-human beta defensin 2; CTRL- healthy controls; AE-angioedema; CU+AE-angioedema associated with urticaria.
Figure 2.
Comparison of salivary HBD2 between groups. (Circles indicate outlier values. Groups connected by a horizontal line are statistically significantly different.) HBD2-human beta defensin 2; CTRL- healthy controls; AE-angioedema; CU+AE-angioedema associated with urticaria.
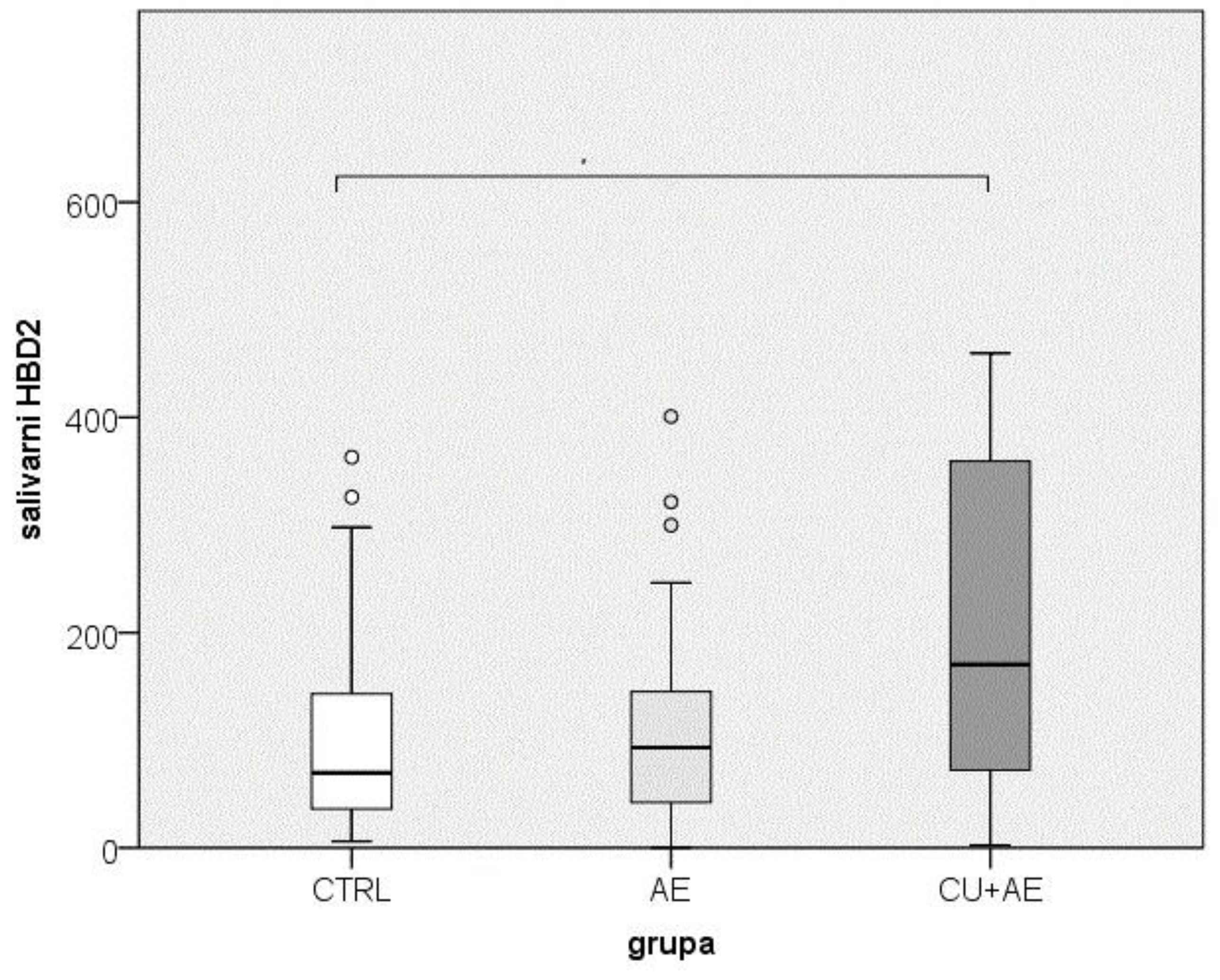
Figure 4.
Comparative display of the range of values for serum and salivary HBD2.
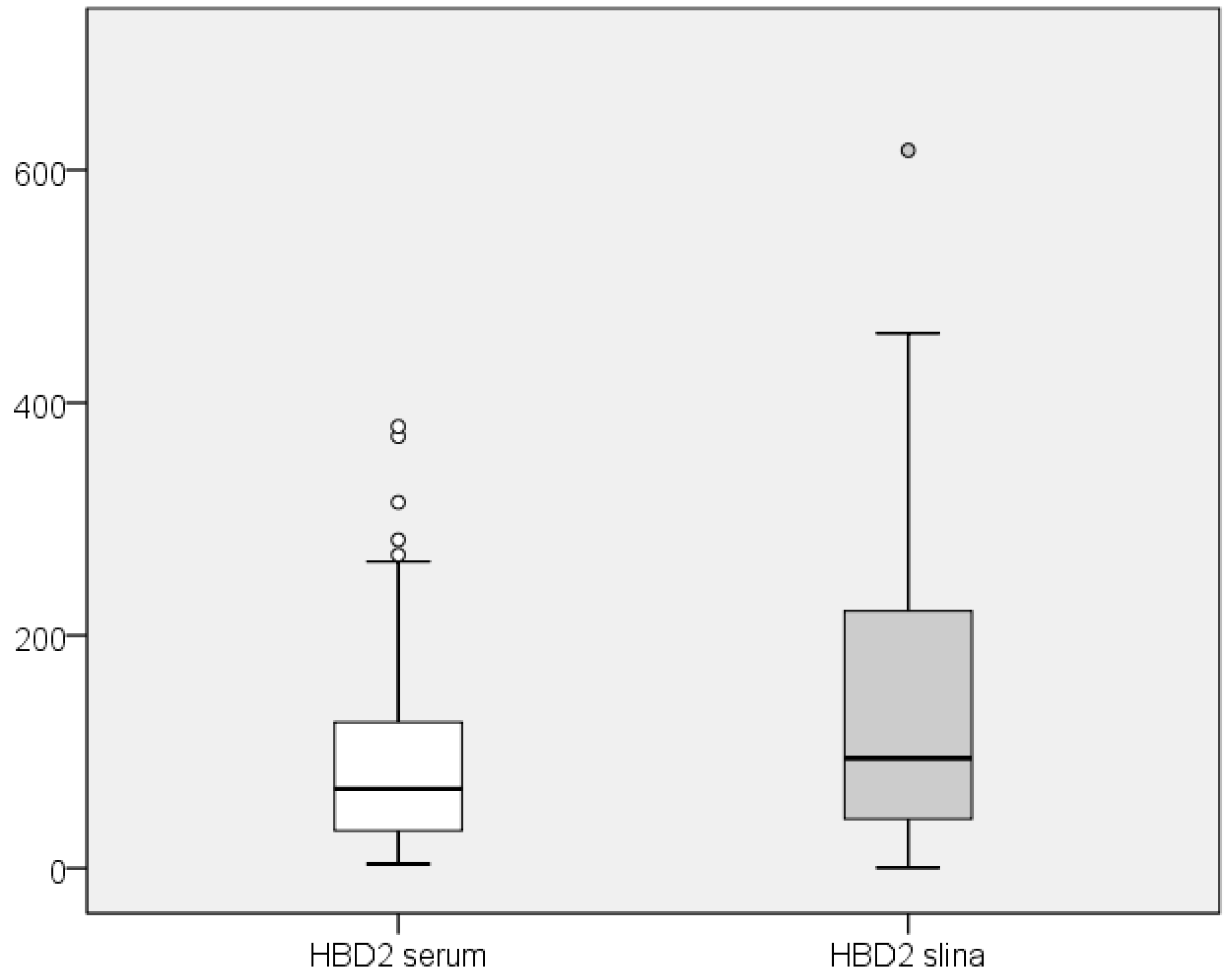
Figure 5.
Relationship between serum and salivary HBD2 values. HBD2-human beta defensin 2
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



